Embed Size (px)
Citation preview

fnt. J . Cancer: 7, 101-11 1 (1971)
STUDIES ON MURINE SARCOMA VIRUS:
BY IMMUNOFLUORESCENCE 11. DETECTION OF GROUP-SPECIFIC ANTIGENS
Jean-Claude CHUAT, Franqoise LASQUELLEC, Anne-Marie L’HIRONDEL
and Michel BOIRON Laboratoire d’Hkmatologie exptrimentale, Institut de Recherches sur les Leuckmies,
H6pital St-Louis, Paris loe, France
The group-specific ( G S ) antigen of murine tumor viruses was demonstrated by immunofluorescence in mouse cells recently infected by mouse sarcoma virus, strain Moloney (MSV-M), with the sesum of rats carrying long-transplanted MSV-M tumors. GS antigen was detected 15 h post-infection and was also present in various mouse and rat cell lines chronically infected with murine tumor viruses. The antigen was strictly localized in the cytoplasm of infected cells and was also found in mouse and rat cells chronically infected by members of the two major subgroups of murine tumor viruses. Further, the sera employed were shown to contain exclusively GS antibodies and the tumors used for immunization were found by several techniques to be free of virus envelope ( V ) tintigens after a given number of passages in vivo. V antigens were visible only at the cell membrane and the time course of appearance of both GS and Vantigens in recently infected cells was parallel. In contrast, GS antigen was not observed in two hamster tumor lines transformed by MSV-M.
An antigen specific to the murine tumor virus group has been demonstrated by the immuno- diffusion (ID) and complement-fixation (CF) techniques (Geering et al., 1966; Huebner et al., 1966) and has been shown to be an internal com- ponent of the virus particle (Schafer et al., 1969). These studies have brought to light many simi- larities in the structure and properties of avian and murine tumor viruses (Huebner, 1967). How- ever, the application of immunofluorescence (IF) to the characterization and localization in infected cells of the murine group-specific (GS) antigen has not been described.
The present study was initiated to see if the results obtained independently by Payne et al. (1966) and Kelloff and Vogt (1966) for avian tumor viruses could be transposed into the murine virus group. In addition to IF, the rat immune
sera employed in this investigation had to be examined by various techniques and the relevant information is given in the present paper.
MATERIAL AND METHODS
Virus
MSV, Moloney isolate (MSV-M) preparations were derived from mouse tumor tissue or from the growth medium of 78 Al. The various strains of murine leukemia virus (MULV), Moloney (MLV), Rauscher (RLV), Gross passage A (GLV) were prepared from the viremic plasma and/or spleen cell-free extracts of infected mice or from the growth medium of the respective cell line. The virus was brought to the desired concentra- tion by spinning at 56,003xg for 90 min in the Spinco model L ultracentrifuge. The viral pellet
Received: June 5, 1970. (and in revised form September 29, 1970).
101

CHUAT ET AL.
was suspended in 0.05 M sucrose for infection or otherwise in Dulbecco’s phosphate-buffered saline (PBS). A sample of GLV, kindly made available to us by Dr. L. Gross (Veterans Administration Hospital, Bronx, N.Y., USA), was also utilized.
Sera
a ) Rat sera. Newborn W-CF (random bred) and W-FU (inbred) rats were inoculated intra- muscularly with MSV-M and the tumors which subsequently developed were serially transplanted by subcutaneous inoculation on the back in adult rats of the respective strain. Two of these tumor lines, designated as MSV-WCF-3 and MSV- WFU-I, were used in the present experiments. The cell dosage a t each tumor generation was so adjusted as to be compatible with long survival and the animals were exsanguinated when mori- bund. Individual serum samples were collected at each tumor generation up to the 15th. In addition, Dr. R. J. Huebner (N.I.H., Bethesda, Md., USA) generously provided a sample of his own rat sera (MSV 20) and Dr. Schafer (Max- Planck-Institut fur Virusforschung, Tubingen, Germany) kindly supplied a rabbit immune serum to tween-ether-t reated R LV (RTE/2).
b ) Mouse sera. (BALB/c x C57BL) F1 hybrid mice were immunized with syngeneically prepared (BALB/c) MSV-M or MLV, or with X-irradiated (6,000 R) cells from a transplanted C57/BL MLV lymphoma. (C3H xC57BL) F1 mice were irn-
munized with GLV grown in C3H or with for- malin-treated (GLV) lymphoma cells.
Cells
A list of the cell lines used in the present study can be found in Table I with the corresponding reference. The lines MSV-WCF-3 and MSV- WFU-1 were derived by trypsinization from the two rat tumor lines described above under Sera, when these were at their 10th and 15th passages in vivo, respectively. Each cell line consists of two cell types, viz. elongated and round cells. The round cells grow in clusters attached to a cell sheet formed by elongated cells. All the cells were in Eagle’s MEM supplemented with 10% calf serum, arginine and antibiotics (penicillin, streptomycin).
Immunoji’uorescence
The cells to be tested were either grown as monolayers on coverslips in Leighton tubes, washed and air-dried, o r collected by trypsiniza- tion from Falcon 250 ml tissue-culture flasks (Falcon Plastics, Los Angeles, Calif., USA), washed, suspended in PBS and layered over coverslips until the PBS had evaporated. Both established cell lines and recently infected cells were used. In the latter case, the virus brought to the desired concentration in serum-free MEM was layered over the cell monolayer, stirred for 3 h at 37” C with a Rocker Platform (Bellco Glass, Vineland, N.J., USA). After this, the cells were
TABLE I
SUMMARY OF THE CULTURE CELL LINES USED IN THIS STUDY
Cell line Host species and strain Virus strain Origin ’ Reference
7 8 A l . . . Rat W/CF MSV-M I, embryo cells Bernard ef a/. (1 967) MSV- WCF-3 I 1 Chuat (present article) MSV-FU-1 . W/FU 11 ERThV-C GLV I , thymocytes Ioachim (1969) JLS-VS . . Mouse, BALB/c RLV I , thyniocytes-I-spleen Wright and Lasfargues
JLS-VII . , M LV I , bone marrow cells Wright ef a/. (1967) CL-I . . . Non-infected I , embryo cells Hartley and Rowe (1966)
(clone P 25) Hamster MSV-M I I Huebner ef u / . (1966)
cells ( I 965)
HT- I
HT-4 . . . MSV-M Perk el a/. (1969)
’ I : normal cells infected in virro, 11: tunlor cells grown in vi lro. See “ Material and Methods ”.
102

ANTIGENS OF M U K l N t SAKCOMAS
washed and maintenance medium with 5 % calf serum was added. Cells were taken at the end of incubation and at regular intervals thereafter. Once dry, the cells on coverslips were either left unfixed or fixed in acetone at room temperature for 10 min. The coverslips were either processed immediately or stored a t -20" C in screw-capped vials until needed.
Immunofluorescence reactions were performed with the indirect technique, using a Bellco Fluorescence Kit (Bellco Glass, Vineland, N.J., USA). The coverslips were dipped in PBS, drained and reacted with properly diluted immune or control serum. After incubation a t 37" C for 45 rnin and three washings in PBS, commercial fluorescein-conjugated (anti IgG) globulin was added. Conjugated globulin was purchased from Hyland Laboratories, Los Angeles, Calif. (anti- mouse and anti-hamster-IgG) and from Micro- biological Associates, Bethesda, Md (anti-rat and anti-rabbit IgG). After a 1 h incubation a t 37" C and three additional washings, the coverslips were dipped in distilled water, immediately mounted in buffered glycerol and examined in UV light in an Ortholux microscope (Leitz, Wetzlar, Germany) equipped with a BG 12/3 mm primary filter and an HBO-200 mercury lamp.
Imrnunodifusion
GS antigen preparations were obtained in either of two ways: ( I ) by disruption of virus-producing cells grown in tissue culture (78 A l , JLS-V5, ER Th/V-A) and (2) by detergent and/or ether treatment of virus suspensions. The procedures followed in the present study are summarized as follows :
Disruption of virus-producing cells: 2 x lo7 cells, washed three times by slow centrifugation in PBS, disrupted by three cycles of freezing and thawing (dry ice +methanol) or sonication. Centrifugation a t 5,000 xg /15 min, 10,000 x g / 5 min, 100,000 x g / 9 0 min-supernatant from each run. Concentration 5 x by polyvinylpyrroli- done ( t 4 " C) through a dialysis tubing.
Disruption of virus suspensions: Tween-ether: 10 or 100xvirus concentrate from culture fluid or viremic plasmattween 80 ( 1 mg/ml) in cold PBS (30 min at 0" C)+half volume peroxide-free ether (shaken 2 h at 20" C). Low-speed centrifu- gation, H,O-phase removed, bubbled with nitro- gen, 100,OOOxg/90 min. Ether alone was occa-
sionally employed. Sodium dodecylsulfate ( S D S ) : Virus concentrate+SDS dropwise to 0.5 % final (5 rnin at 20" C ) + a drop saturated KCL (0" C). Low-speed centrifugation, excess detergent re- moved, 100,000 xg/90 min.
Micro-immunodiffusion tests were performed either with 2% Noble Agar in distilled water, using a commercial punch set (Gelman Instru- ment Co., Ann Arbor, Mich., USA) or Immuno- plate pattern C (Hyland Laboratories, Los Angeles, Calif., USA). In both cases; the slides were incubated for 24 h a t room temperature in a humidified chamber. A buffer containing cad- mium (Crowle, 1961 ), as suggested by Fink et al. (1969), was occasionally utilized.
Other techniques
a) Serological techniques detecting viral envelope andlor virus-specific cell surface antigens. For in vivo neutralization tests, undiluted MSV was incubated for 1 h at 37" C with a n equal amount of undiluted serum and 0.2 ml of the mixture was inoculated into each of five to 10 newborn mice. In absorption tests, properly diluted serum was mixed with varying amounts of infected o r control cells or with pelletized virus. After 30 rnin a t 37" C, followed by 18 h a t 1 4 " C, cells or virus were removed by centrifugation (10,000 x g ) o r ultracentrifugation (56,000 xg) respectively. The techniques for cytotoxic (Fenyo et al., 1968) membrane immunofluorescence (MIF) (Klein and Klein, 1964a; Chuat et al., 1969) and mouse anti- body production (MAP) (Klein and Klein, 19646) tests have been described. For the latter (A/Sn X C57BL) F1 hybrids were used, as the sera were tested on an A/Sn transplantable lymphoma.
b) Radioactive labelling and sucrose density gradients. For labelling, cultured cells of MSV- WFU-1 and of 78 A1 were grown for 48 h in 400 ml of medium containing 2 mCi of 3 H - ~ r i - dine (specific activity: 18 Ci/mmole). The culture fluids were then clarified by low-speed centrifu- gation and concentrated 100 x by spinning a t 56,000 x g and suspending in 0.4 ml of 5 % sucrose in Tris 0.001 5 M. This was layered over 4.6 ml of a preformed linear gradient of 15-60% sucrose and run at 130,000 x g for 15 h in an SW 65 Spinco rotor. Three-drop fractions were collected from the bottom of the punctured tubes and radio- activity was measured in each fraction by a scin- tillation counter.
103
ANTIGENS OF MURINE SARCOMAS
A 4 FIGURE I
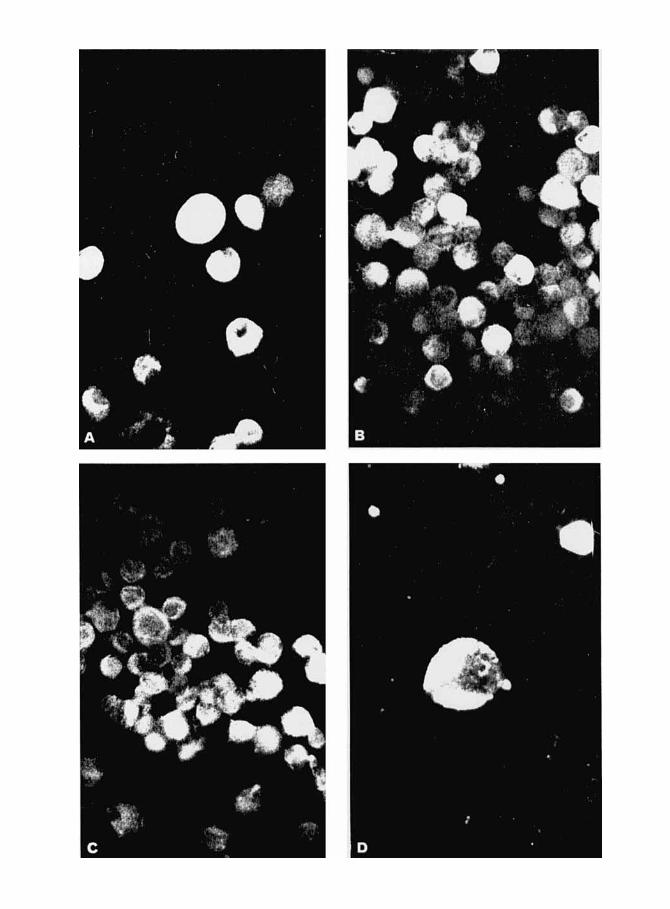
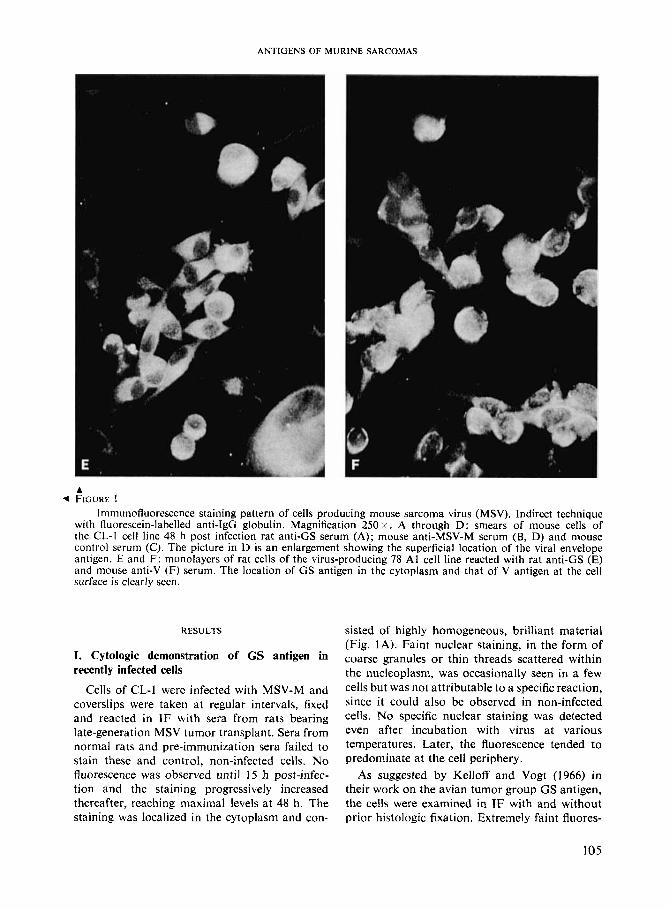
Immunofluorescence staining pattern of cells producing mouse sarcoma virus (MSV). Indirect technique with fluorescein-labelled anti-IgG globulin. Magnification 250 x . A through D: smears of mouse cells of the CL-I cell line 48 h post infection rat anti-GS serum (A); mouse anti-MSV-M serum (B, D) and mouse control serum (C) . The picture in D is an enlargement showing the superficial location of the viral envelope antigen. E and F: monolayers of rat cells of the virus-producing 78 A1 cell line reacted with rat anti-GS (E) and mouse anti-V (F) serum. The location of GS antigen in the cytoplasm and that of V antigen at the cell surface is clearly seen.
RESULTS
I. Cytologic demonstration of GS antigen in recently infected cells
Cells of CL-1 were infected with MSV-M and coverslips were taken at regular intervals, fixed and reacted in IF with Sera from rats bearing late-generation MSV tumor transplant. Sera from normal rats and pre-immunization Sera failed to stain these and control, non-infected cells. No fluorescence was observed until 15 h post-infec- As suggested by Kelloff and Vogt (1966) in tion and the staining progressively increased their work on the avian tumor group GS antigen, thereafter, reaching maximal levels a t 48 h. The the cells were examined in IF with and without staining was localized in the cytoplasm and con- prior histologic fixation. Extremely faint fluores-
sisted of highly homogeneous, brilliant material (Fig. 1A). Faint nuclear staining, in the form of coarse granules o r thin threads scattered within the nucleoplasrn, was occasionally seen in a few Cells but Was not attributable to a specific reaction, since it could also be observed in non-infected Cells. NO Specific nuclear Staining was detected even after incubation with virus a t various temperatures. Later, the fluorescence tended to Predominate a t the cell Periphery.
105

CHUAT ET AL.
cence or none at all was seen in the absence of fixation, suggesting that the material detected was restricted to the cytoplasm.
11. Demonstration of the group reactivity of rat immune sera
To confirm the cytoplasmic localization of GS antigen, and to determine whether the rat sera detect only GS antigen or both GS and type- specific (V) antigens, we resorted to (1) comparison of the fluorescence obtained with the serum of tumor-bearing rats and pooled immune serum prepared in mice with virus or lymphoma cells, and (2) further characterization of the rat and mouse sera employed in neutralization, ID, MIF, cytotoxicity and absorption tests. In this respect, it must be emphasized that the term V antigens is used for convenience, and that distinction between viral envelope antigens and cell surface type-specific antigens, which requires the use of electron microscopy with visually labelled antibody (Aoki et al., 1970), has not been attempted in the present study.
The distinction between GS and V antigens by immunofluorescence with recently infected cells
Pooled serum from mice immunized with MSV-M was reacted with fixed, MSV-infected, CL-I cells at various times post infection, as described above. Despite several washings, faint staining with a very uniform distribution was present at the end of the absorption period. However, no difficulty was encountered in recog- nizing the synthesis of progeny V antigen, which started at 15 h. The staining was very bright and affected only a given proportion of the cells, thus permitting sharp distinction with the uniform and dim background staining due to residual virus (Fig. 1B and D). As the proportion of stained cells in various experiments exactly paralleled that observed with rat immune serum, much attention was given to the distinction between the aspects of the two antigens. Three major criteria were retained, viz. (1) extremely faint fluorescence or none at all was observed with anti-GS-serum without prior histologic fixation of the cells, whereas no such requirement was apparent with anti-V-serum; (2) the morphologic aspect of the staining obtained with anti-V-serum was markedly different, consisting of bright patches located exclusively at the cell surface (Fig. lD), as was obvious when focus was varied between extreme
positions; (3) additional distinction between the two specificities was provided by absorption with intact virus or with virus-producing, living cells (78 Al, JLS-VS). For this, both anti-GS and anti-V sera were so diluted as to give comparable staining intensity, absorbed and titered again. Whereas the reactivity of rat antiserum remained unaffected, there was a sharp decrease of fluores- cence with absorbed mouse anti-serum with a parallel decrease in cytotoxic or MIF tests.
Irnmunodiffusion reactions
Representative reactions are shown in Figure 2. No reaction was ever detected between mouse anti-V serum and any of the material tested. In contrast, it can be seen that rat anti-sera reacted after degradation of the virus particles by ether, tween-ether or SDS treatment, whereas the same serum gave no reaction if the virus preparations were made of intact particles. The single apparent exception was MSV-M harvested from 78 A1 cells after maintaining the cultures without medium change for 10 days. In that case, a weak reaction with seemingly undegraded virus was observed. An antigen cross-reacting with degraded virion antigen was found in the cytoplasmic fraction of disrupted virus-producing cell lines (78 Al, JLS- V5, JLS-VI 1). In addition, the seru n was also reacted with degraded GLV from two sources: one was a generous gift of Dr. Gross and the other was sedimented from growth fluid of the ERTh V-G GLV-producing rat cell line of Dr. Ioachim. As can be seen in Figure 2 , a reaction was observed only with degraded virus and the same antigen was detected in the dis- rupted cells of this line.
Taken all together, the need for detergent degradation of the particles and the cross-reaction between MSV-M and GLV indicate that sera of rats carrying MSV-M induced tumor transplants contain antibody to an inner, group-specific component of the virus particle which is also present in the cytoplasm of virus-producing cul- tures. The reaction seen with undegraded MSV-M obtained from 10-day-old growth fluid suggests that spontaneous degradation of fragilized virions had taken place during incubation in agar.
Treatment of the agar plates after incubation with a cadmium-containing buffer, as suggested by Fink er al. (1969), did not alter the reactivity pattern observed. More precisely, this treatment
106

ANTIGENS OF MURINE SARCOMAS
did not in our hands lead to a visible reaction between rat serum and undegraded MSV-M.
Finally, the sera kindly provided by Drs. Hueb- ner and Schafer gave reactions of identity with ours.
Reactivity to V antigens of antisera fvom rats hearing transplanted M S V tumors
The serum samples collected at the various successive tumor generations of MSV-CF-3 and MSV-FU-1 were tested in parallel t o see if they contained antibodies to V antigens. The finding that apparently normal rat sera were frequently cytotoxic for the mouse lymphoma target cells Y A C precluded the use of the cytotoxic test and left us with the virus-neutralizing and membrane immunofluorescence reactions. Results appearing in Table I1 (A and B) show that neutralizing antibodies were no longer produced after several passages of both tumors in vivo. The Table similarly shows that V antibodies detected by MIF tended to decrease with passing tumor generations. The results obtained with mouse anti-V serum are shown for comparison (Table 11, C) .
In addition, the two MSV-M tumors described in this paper grown in vitro were examined in the
TABLE 11
I N V l V O NEUTRALIZATION ' A N D MEMBRANE IMMUNOFLUORESCENCE TESTS WITH RAT ANTI-GS
A N D MOUSE ANTI-MSV-M SERA
Donor species Immunization No' with Fluore'- tumor/ cence total index and strain with
A. W/CF rat MSV-W-CF 3, generation No.
3 0/4 NT 15 515
control 818 B. W/FU rat MSV-FU-1,
generation No. 2 417 0.44 4 NT 0.38 6 8/8 0.10
10 616 0.10 1 1 NT 0.22 1 3 818 0.11 15 616 0.09
control 717 - C. BALB/C x
C57BL/6 F1 MSV-M 019 0.90 control 919 -
Each mouse inoculated intramuscularly with 0.2 ml of a 1: l MSV-M immune or control serum mixture after 30 min at 37" C.
Performed on living 78 A l cells. Fluorescence index (FI): % negative cells with control serum minus % negative cells with test serum, divided by the former figure. Reaction considered as positive if IFZ0.3.
NT: not tested.
FIGURE 2 Immunodiffusion reactions of a serum from an MSV tumor-bearing rat with various antigen preparations. R = Serum from a W/FU rat immunized with the 13th tumor generation of MSV/FU-I tumor. ( I ) MSV
from mouse tumors; (2) same after TE; (3) cytoplasmic fraction of 78 A1 cells (W/CF rat cell line, permanent producer of MSV-M); (4) GLV from mouse plasma after TE; (5) MSV from 78 A1 cell culture fluid after TE; (6) same except after SDS; (7) same except after spontaneous degradation; (8) same from a 4-day culture (non degraded); (9) GLV from mouse plasma.
Abbreviations: MSV-M = murine sarcoma virus, Moloney isolate; GLV: Gross leukemia virus; TE: Tween-ether treatment; SDS: Sodium dodecylsulfate.
107
CHUAT ET A t .
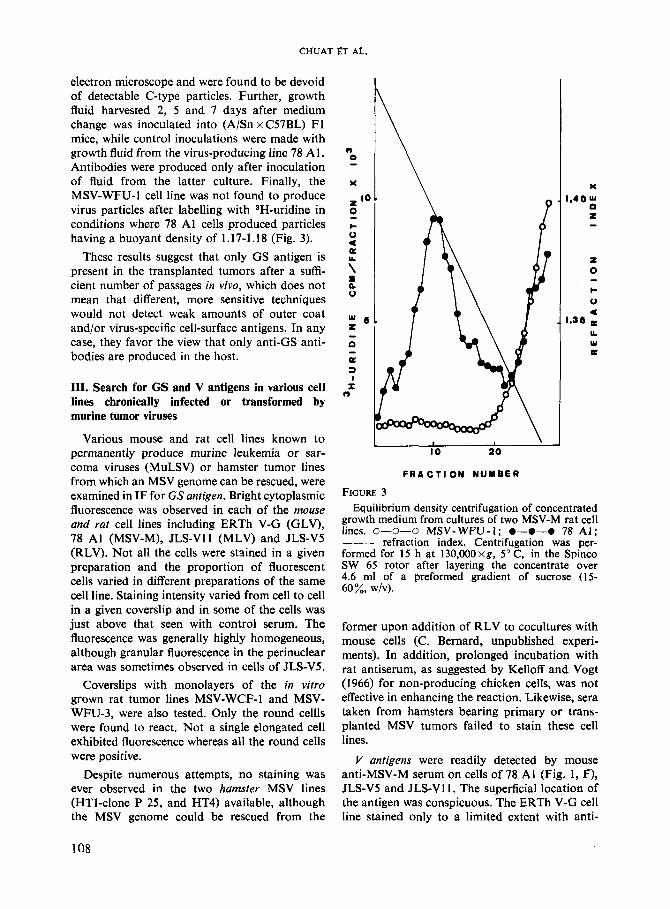
electron microscope and were found to be devoid of detectable C-type particles. Further, growth fluid harvested 2, 5 and 7 dsys after medium change was inoculated into (A/Sn x C57BL) F1 mice, while control inoculations were made with growth fluid from the virus-producing line 78 Al. Antibodies were produced only after inoculation of fluid from the latter culture. Finally, the MSV-WFU-1 cell line was not found to produce virus particles after labelling with SH-uridine in conditions where 78 A1 cells produced particles having a buoyant density of 1.17-1.18 (Fig. 3).
These results suggest that only GS antigen is present in the transplanted tumors after a suffi- cient number of passages in vivo, which does not mean that different, more sensitive techniques would not detect weak amounts of outer coat and/or virus-specific cell-surface antigens. In any case, they favor the view that only anti-GS anti- bodies are produced in the host.
111. Search for GS and V antigens in various cell lines chronically infected or transformed by murine tumor viruses
Various mouse and rat cell lines known to permanently produce murine leukemia or sar- coma viruses (MuLSV) or hamster tumor lines from which an MSV genome can be rescued, were examined in IF for GS antigen. Bright cytoplasmic fluorescence was observed in each of the mouse and rat cell lines including ERTh V-G (GLV), 78 A1 (MSV-M), JLS-V11 (MLV) and JLS-VS (RLV). Not all the cells were stained in a given preparation and the proportion of fluorescent cells varied in different preparations of the same cell line. Staining intensity varied from cell to cell in a given coverslip and in some of the cells was just above that seen with control serum. The fluorescence was generally highly homogeneous, although granular fluorescence in the perinuclear area was sometimes observed in cells of JLS-VS.
Coverslips with monolayers of the in vitro grown rat tumor lines MSV-WCF-1 and MSV- WFU-3, were also tested. Only the round cellls were found to react. Not a single elongated cell exhibited fluorescence whereas all the round cells were positive.
Despite numerous attempts, no staining was ever observed in the two hamster MSV lines (HT1-clone P 25, and HT4) available, although the MSV genome could be rescued from the
I \ 10 20
F R A C T I O N NUMBER
FIGURE 3 Equilibrium density centrifugation of concentrated
growth medium from cultures of two MSV-M rat cell lines. 0-0-0 MSV-WFU-I; 0-0-0 78 A l ; ___ refraction index. Centrifugation was per- formed for 15 h at 130,OOOxg, 5” C, in the Spinco SW 65 rotor after layering the concentrate over 4.6 ml of a preformed gradient of sucrose (15- 60%, wlv).
former upon addition of RLV to cocultures with mouse cells (C. Bernard, unpublished experi- ments). In addition, prolonged incubation with rat antiserum, as suggested by Kelloff and Vogt (1966) for non-producing chicken cells, was not effective in enhancing the reaction. Likewise, sera taken from hamsters bearing primary or trans- planted MSV tumors failed to stain these cell lines.
V antigens were readily detected by mouse anti-MSV-M serum on cells of 78 A1 (Fig. 1, F), JLS-VS and JLS-V1 1 . The superficial location of the antigen was conspicuous. The ERTh V-G cell line stained only to a limited extent with anti-
108

ANTIGENS OF MURINE SARCOMAS
MSV and anti-GLV mouse serum whereas the latter was positive with 78 A1 (MSV-M) cells. This confirms previous unpublished data with cytotoxic tests where potent anti-GLV sera gave a weak but discernible reaction with YAC (MLV) lymphoma cells. Consequently, clear-cut distinc- tion between the two subgroups of murine tumor viruses, viz. Gross (G) and Friend-Moloney- Rauscher (FMR) was not possible. Finally, the two rat tumor lines MSV-CF-3 and MSV-FU-1 were negative when reacted with mouse anti-V sera.
DISCUSSION
Earlier investigations of murine leukemia virus (RLV, FLV, MLV) antigens by IF were per- formed with antiserum produced by immuniza- tion of heterologous species (rabbits or monkeys) with presumably intact particles. Fink and Malm- gren (1963) were pioneering to detect antigens in the tissues of leukemic animals and Osato et al. (1964) and Yoshida et al. (1966) used IF to study murine leukemia virus replication in vitro. The evidence concerning the nuclear or cytoplas- mic localization of the antigens was conflicting and the specificity (group or type) was not assessed. The use of syngeneically obtained mouse antiserum was restricted to the study of mem- brane antigens on lymphoma cells (Klein and Klein, 1964a).
In the present studies, and provided the tumors used for immunizing the animals had undergone several passages in vivo, the antigen detected in IF with rat serum appeared to be restricted to the cytoplasm, in the sense that, (1) we failed to observe any specific nuclear staining, even early after infection and in varied conditions of tem- perature, as recommended by Payne et al. (1966) for avian tumor viruses; (2) the antigen was seen only after histologic fixation of the cells. In addition, the reactivity of the rat antiserum was not impaired by prior absorption with virus or virus-containing material. The antigen was present in all the chronically infected mouse and rat cell lines tested, including cells producing GLV, which in itself suggests that the antigen was actually GS antigen. However, GS seems to be absent from all the cells of two hamster MSV lines (HT-I and HT-4).
In contrast, the antigen detected with mouse anti-viral Serum was found to be strictly located at the cell surface. In fact, the antigen was visible
even without prior fixation and was no longer stainable by virus-absorbed mouse antiserum. However, the type or rather subgroup specificity of the antigen could not be assessed, as'there was a cross-reaction between anti-GLV and anti-MLV mouse antisera.
As reported for the avian GS antigen with rabbit antiserum to split products of avian myelo- blastosis virus (AMV), the rabbit antiserum to split RLV kindly furnished by Dr. Schafer gave reactions identical to those of tumor-bearing rat sera.
On the whole, and including absence of nuclear staining, these observations are in accordance with those of Kelloff and Vogt (1966), who used the serum of hamsters bearing tumors induced by the Schmidt-Ruppin strain of Rous Sarcoma virus (RSV-SR) for detecting avian GS antigen, while chicken antiserum to intact virus was used to demonstrate viral envelope, type-specific antigens.
GS antigen was visible as early as 15 h post infection in mouse cells.
Owing to the fact that the time course of appearance of both GS and V antigens in recently infected cells was strictly parallel, much care was taken to assess the group-specificity of the reaction observed in I F with the serum of rats carrying transplants of MSV-M induced tumors. In addition to IF, the group-reactivity was further strengthened by other relevant im- munological techniques.
The reasons for believing that GS antigen was effectively detected are direct and indirect. Direct arguments are the cytoplasmic location of the antigen in IF and its presence in cells infected by members of the two major subgroups of murine tumor viruses, viz. G and FMR. In fact, the antigen reacted with a cytoplasmic antigen in cells producing GLV. In addition, the sera used in IF were able to detect in ID tests an antigen present in disrupted infected cells and in degraded virus particles, and a cross-reaction between the two virus subgroups was observed. The antigen was not sedimented by forces known to pelletize intact virions. Moreover, absorption tests showed that the reaction was not abolished after incuba- tion with living, virus-producing cells or intact virus particles in conditions where anti-viral anti- bodies were absorbed out. Indirect arguments can be derived from the absence of antibodies to
109

CHUAT ET AL.
the virus envelope or to cell surface virus-specific antigens in the sera employed. In effect, the sera of rats carrying serial transplants of two different MSV tumors were found to contain neutralizing antibody during the very first tumor generations only. Antibodies to envelope antigens present on the surface of MSV-producing rat cells (78 Al) were found to persist somewhat longer but ulti- mately to disappear.
Further, the two tumor lines used for immuniza- tion were grown in tissue culture. The cells of these cultures were not found by electron micro- scopic examination to produce virus particles and were not stained in I F tests with mouse anti- viral antibodies. Likewise, the growth fluid of the cultures failed to evoke cytotoxic antibodies upon inoculation in mice, whereas that of 78 A1 cul- tures did. The same was true of short-term cultures of the tumors. Finally, and in contrast to 78 A1 cells, no evidence of virus release from the tumor cells could be obtained after labelling with 3H-uridine.
Huebner et al. (1966) also noted that neutrali- zing antibodies tend to disappear from the serum of MSV tumor-bearing rats wjth passing tumor generations. This is the murine counterpart of hamster carrying RSV-SR tumors (Sarma et al., 1964) and the present experiments show that these transplanted rat tumors are probably non- producing tumors. The same may not be true of rats carrying transplanted lymphomas induced by murine leukemia viruses, as their serum con- tains neutralizing or cytotoxic antibodies (Hartley et al., 1965; Geering et al., 1966). In fact, murine leukemia viruses are known to fully replicate in rat tissues.
ACKNOWLEDGEMENT
We wish to thank Dr. Jacqueline Lasneret who kindly performed the electron microscope exami- nations described in this paper. We also acknow- ledge the skilful technical assistance of Mrne Mar- tine Mauchauffk.
ETUDE DU VIRUS DU SARCOME MURIN: 11. DETECTION DES ANTIGPNES SPECIFIQUES DE GROUPE PAR IMM UNOFLUORESCENCE
L'antigtne spe'cifque de groupe (gs) des tumeurs mirrines a C t t mis en Cvidence par immunofluorescence dans des cellules de souris ricemment inferties par Ie virus du sarcome murin, souche Moloney ( VSM-M), au moyen de strum de rats porteurs de tumeurs a VSM-M plusieurs fois transplanttes. L'antigtne gs a t t t dCcelC I5 heures aprPs l'infection; il Ctait e'galement pre'sent dans diverses Iignies cellulaires de rats er de souris chroniquement infertees par les virus des tumeurs murines. L'antigtne Ptait strictement localisk et n'apparaissait que dans le cytoplasme des cellules infectCes; il a aussi e'tt de'celi dans les cellules de rat et de souris chroniquement infectees par des virus appartenant aux deux principaux sous-groupes des virus des tumeurs murines. En outre, on a constatt que les se'rums utilists contenaient exclusivement les anticorps gs; diverses techniques ont montri que les titmeurs employees pour I'immunisation ne contenaient plus d'antigtnes ( V ) a enveloppe virale aprts un certain nombre de passages in vivo. Les antigtnes V n'itaient visibles que sur la membrane cellulaire et le temps ntcessaire a ['apparition des antigenes gs et V dans les celiules ricemment infecte'es e'tait du m6me ordre. Par contre, f'antigzne gs n'a pas Pt& observe' dans deux souches tumorales de hamster transformkes par le VSM-M.
REFERENCES
AOKI, T., BOYSE, E. A,, OLD, L. J . , DE HARVEN, E., HAMMERLING, v., and WOOD, H. A., G (Gross) and H-2 cell surface antigens: Location on Gross leukemia cells by electron microscopy with visually labelled antibody. Proc. nut. Acad. Sci. (Wash.), (1967).
BERNARD, C., BOIRON, M., and LASNERET, J., Trans- formation et infection chronique de cekles em- bryonnaires de rat par le virus du sarcome de Moloney. C. R. Acud. Sci. (Paris), 264, 2170-2173
65, 569-576 (I 970).
110

ANTIGENS OF MURINE SARCOMAS
CHUAT, J. C., BERMAN, L., GUNVBN, P., and KLEIN, E., Studies on murine sarcoma virus: antigenic charac- terization of murine sarcoma virus induced tumor cells. Int. J . Cancer, 4, 465-479 (1969).
CROWLE, A. J., Immunodiffusion, pp. 170-171. Aca- demic Press, New York (1961).
FENYO, E. M., KLEIN, G., and SWIECH, K., Selection of an immuno-resistant Moloney lymphoma sub- line with decreased concentration of tumor-specific surface antigens. J . nut. Cancer Inst., 40, 69-89 (1968).
FINK, M. A. and MALMGREN, R. A., Fluorescent antibody studies pf the viral antigen in a murine leukemia (Rauscher). J . nut. Cancer Inst., 31, 1 I 1 1- 1121 (1963).
FINK, M.A.,SIBAL,L.R., WIVEL,N.A., COWLES, C.A., and O’CONNOR, T. E., Some characteristics of an isolated group antigen common to most strains of murine leukemia virus. Virology, 37,605-614 (1969).
GEERING, G., OLD, L. J., and BOYSE, E. A., Antigens of leukemia induced by naturally occurring murine leukemia virus: Their relation to the antigen of Gross virus and other murine leukemia viruses. J . exp. Med., 124, 753-772 (1966).
HARTLEY, J. W., and ROWE, W. P., Production of altered foci in tissue culture by defective Moloney sarcoma virus particles. Proc. nut. Acad. Sci.
HARTLEY, J. W., RoWE, W. P., CAPPS, W. I . , and HUEBNER, R. J., Complement-fixation and tissue culture assays for moiise leukemia viruses. Proc.
HUEBNER, R. J., The murine leukemia sarcoma virus complex. Proc. nut. Acad. Sci. (Wash.), 58, 835- 842 (1967).
HUEBNER, R. J., HARTLEY, J. W., ROWE, W. P. LANE, W. T., and CAPPS, W. I . , Rescue of the defective genome of Moloney sarcoma virus from a non infectious hamster tumor and the production of pseudotype sarcoma viruses with varioas murine leukemia viruses. Proc. nut. Acad. Sci. (Wash.),
IOACHIM, H. L., Divergence in tumor induction of thymus cells transformed in vitro by Gross leukemia virus. J. nut. Cancer Insr., 42, 101-113 (1969).
(Wash.), 55, 780-786 (1966).
nut. Acad. Sci. (Wash.), 53, 931-938 (1965).
56, 1164-1169 (1966).
KELLOFF, G., and VOGT, P. K., Localization of avian tumor virus group-specific antigen in cell and virus Virology, 29, 377-384 (1966).
KLEIN, E., and KLEIN, G., Antigenic properties of lymphomas induced by the Moloney agent. J. nut. Cancer Inst., 32, 547-568 (1964~).
KLEIN, E., and KLEIN, G., Mouse antibody production test for the assay of the Moloney virus. Nature (Lond.), 204, 339-342 (1964h).
OSATO, T., MIRAND, E. A., and GRACE, J. T., Propa- nation and immunofluorescent investigations of Friend virus in tissue culture. Nature ( L h d . ) , 201, 52-54 (1964).
PAYNE, F. E., SOLOMON, J. J., and PURCHASE, H. G., Immunofluorescent studies of group-specific antigen of the avian sarcoma-leukosis viruses. Proc. nut. Acad. Sci. (Wash), 55, 341-349 (1966).
PERK, K., VIOLA, M. V., SMITH, K. L., WIVEL, N. A., and MOLONEY, J. B., Biologic studies on hamster tumors induced by the murine sarcoma virus (Moloney). Cancer Res., 29, 1089-1 102 (1969).
SARMA, P. S., HUEBNER, R. J., and ARMSTRONG, D., A simplified tissue culture tube neutralization test for Rous sarcoma virus. Proc. SOC. exp. Biol.
SCHAFER, W., ANDERER, F. A., BAUER, H., and PETER, L., Studies on mouse leukemia viruses. 1. Isolation and characterization of a group-specific antigen. Virology, 38, 387-394 (1969).
WRIGHT, B. S., and LASFARGUES, J. C., Long-term propagation of the Rauscher murine leukemia virus in tissue culture. J. nut. Cancer Inst., 35, 319-327 (1965).
WRIGHT, B. S., O’BRIEN, P. A., SHIBLEY, G. P., MAYYASI, S. A., and LASFARGUES, J. C., Infection of an established mouse bone marrow cell line (JLS-V9) with Rauscher and Moloney murine leukemia viruses. Cancer Res., 27, 1672-1677 (1967).
YOSHIDA, K., SMITH, K. L., and PINKEL, D., Studies of murine leukemia viruses. I. Detection of Moloney and Rauscher viruses by indirect immunofluores- cence. Proc. SOC. exp. Biol. ( N . Y.j , 121, 72-81 (1966).
( N . Y.), 115, 481-486 (1964).
111





























