Embed Size (px)
Citation preview

Ultrastructural localisation and functionalimplications of Corticotropin releasing factor,
Urocortin and their receptors in cerebellarneuronal development

© 2003 by J.D. Swinny
All rights reserved. No part of this book may be reproduced or transmitted in any form or by
any means without written permission of the author and the publisher holding the copyright
of the published articles.
Cover and page design: P. van der Sijde, Groningen, The Netherlands
Printed by: Ponsen & Looijen B.V., Wageningen, The Netherlands
This thesis was fundet by BCN

RIJKSUNIVERSITEIT GRONINGEN
Ultrastructural localisation and functional
implications of Corticotropin releasing factor,
Urocortin and their receptors in
cerebellar neuronal development
Proefschrift
ter verkrijging van het doctoraat in deMedische Wetenschappen
aan de Rijksuniversiteit Groningenop gezag van de
Rector Magnificus, dr. F. Zwarts,in het openbaar te verdedigen op
woensdag, 8 oktober, 2003om 14.15 uur
door
Jerome Dominic Swinny
geboren op 6 oktober 1969te Dundee, Zuid Afrika

Promotor: Prof. dr. A. Gramsbergen
Co-promotor: dr. J.J.L. van der Want
Beoordelingscommissie: Prof. dr. H.W.G.M. Boddeke
Prof. dr. C.I. De Zeeuw
Prof. dr. P.G. Luiten

Paranimfen: Florence Postollec
Virginia Vadillo-Rodriguez

Dedicated to my parents

CONTENTS
Chapter 1 General Introduction 9
Chapter 2 The expression of Corticotropin releasing factor in the 35
developing rat cerebellum: a light and electron microscopic
study
J. D. Swinny, D. Kalicharan , J. IJkema-Paassen, J.J.L. van der Want,
A. Gramsbergen
Chapter 3 The localisation of urocortin in the adult rat cerebellum: 55
A light and electron microscopic study
J. D. Swinny, D. Kalicharan, A. Gramsbergen , J.J.L. van der Want.
Neuroscience (2002) ;114(4):891-903.
Chapter 4 The postnatal developmental expression pattern of 73
urocortin in the rat olivocerebellar system
J.D. Swinny, D. Kalicharan, F. Shi, A. Gramsbergen ,
J.J.L. van der Want.
Journal of Comparative Neurology: submitted
Chapter 5 Corticotropin releasing Factor receptor types 1 and 2 are 91
differentially expressed in pre- and postsynaptic elements
in the postnatal developing rat cerebellum
J. D. Swinny, D. Kalicharan, E.H. Blaauw, J. IJkema-Paassen,
F. Shi, A. Gramsbergen, J.J.L. van der Want.
European Journal of Neuroscience: (2003);18:549-562.
Chapter 6 Corticotropin releasing factor and Urocortin modulate 115
Purkinje cell dendritic outgrowth and differentiation in vitro
J. D. Swinny, J. IJkema-Paassen, F. Metzger, A. Gramsbergen,
J.J.L. van der Want
Submitted: Mol. Cell. Neuroscience
Chapter 7 General discussion and conclusions 135
Summary 143
Samenvatting (Summary in Dutch) 144
Acknowledgements 146
Curriculum Vitae 148


CHAPTER 1
General Introduction

10
Chapter 1
1.1 General introduction
The development of the nervous system is governed by the intrinsic genetic blueprint
of neurons and, importantly, epigenetic cues, acting e.g. via a wide variety of chemical
signals, which either promote or repress growth. This results in the development of
highly specific patterns of connectivity, which are usually maintained throughout
life, but are subject to plastic changes based on adaptations to changing environmental
needs. The establishment of causal relationships between the temporal expression of
neurochemical substrates such as neuropeptides, and well-defined developmental
events are at the forefront of elucidating the process of normal development, and in
case of pathologies, potential therapies.
Contemporary theory holds that the role of the cerebellum is that of a coordinating
centre, using sensory inputs from the periphery to fine-tune our movement and postural
control (Doyon et al., 2002). Apart from motor coordination, the cerebellum is also
thought to be involved in motor learning and higher cognitive function (Allen et al.,
1997; Bloedel and Bracha, 1997). The cerebellum, in relation to other brain structures,
enjoys a protracted developmental profile, providing a convenient vantage point to
view ongoing developmental processes. This accessibility of the developing
cerebellum provides a highly amenable model to study the roles of neuropeptides
expressed during normal development.
One of the peptides growing in stature as a modulator of neuronal development is
corticotropin releasing factor (CRF) (Vale et al., 1981). Apart from that, CRF is well
established as the major physiological regulator of the hypothalamic-pituitary-adrenal
(HPA) axis and serves to coordinate the mammalian endocrine, autonomic and
behavioural responses to stress (Koob and Heinrichs, 1999). There are, however,
extrahypothalamic sources of CRF, most prominent being the collection of brainstem
nuclei collectively termed the pre-cerebellar nuclei. They express CRF in their axons
that form the major afferent systems to the cerebellum, namely the mossy fibres and
climbing fibres (Palkovitz et al., 1987). Recently discovered urocortin (Vaughan et
al., 1995) has a more restricted expression profile and is believed to be crucial in
sensori-motor integration (Bittencourt et al., 1999).
Both CRF and urocortin couple to two G-protein receptors, namely CRF-R1 and
CRF-R2. In vivo, at adult age, activation of CRF-R1 has been shown to induce
locomotor activity (Contarino et al., 2000), while the activation of CRF-R2 has an
inhibitory effect (Valdez et al., 2002). Crucially, in vitro, CRF has been shown to be

11
Introduction
necessary for the induction of long term depression (LTD) (Miyata et al., 1999), a
type of synaptic plasticity and viewed as the cellular basis of learning (Ito, 1984).
This alludes to a direct role for CRF-like peptides on motor coordination and motor
learning in adulthood. However, the onset of expression of CRF (Bishop and King,
1999) urocortin and their receptors (Chang et al., 1993) occurs embryonically, prior
to any synapses being formed, suggesting an initial role for CRF-like peptides in the
development of the cerebellar circuitry. During the first three postnatal weeks,
cerebellar neurons undergo considerable structural changes. This period coincides
with the maturation of the “CRF system”. These structural changes attract considerable
interest since morphological changes are considered integral in higher brain function
such as learning and memory. To define the potential roles of CRF-like peptides in
dendritic growth and synapse formation, ultrastructural data on their localisation
within the developing cerebellar circuitry is essential. In this thesis, these aspects are
investigated in an attempt to establish causal relationships between the presence of
CRF-like peptides and well-defined cellular and behavioural processes that occur
during cerebellar development.
Figure 1. An overview of the cerebellar cortex showing the three layers and five differenttypes of neurons. A section of a single folium in both the sagittal and transverse planes
illustrates the general organisation of the cerebellar cortex (Kandel et al., 2000).

12
Chapter 1
1.2 Cerebellar function
Most of our mundane activities, such as walking, cycling or writing are carried out
with ease and precision. Many parts of the brain have to interact to ensure such
coordinated motor activity. Parameters of this coordination are adjusted during early
years of development and years of motor learning during adulthood. The cerebellum
plays a specific role in producing smooth motor acts and can be considered the brain’s
engine of agility. Indeed, damage to this structure results in imprecise and unsure
movements, and even impaired thoughts about movement (see Muller et al., 1998).
In view of the extremely regular anatomical organisation of the cerebellar cortex
(see figure 1) (Eccles et al., 1967), it is thought that the cerebellum may perform a
general computational role, which is similar for all its target systems (Marr, 1969;
Albus, 1971).
1.2.1 Role in motor coordination
The cerebellum is well known to be essential in coordination among the joints of one
limb, between limbs, between the eye and hand in reaching, pointing or tracking
tasks, or between the eye and extremeties during gait (see figure 2 for afferent and
efferent pathways). The cerebellum influences motor output by evaluating differences
between intended and actual motor acts. Furthermore, it is able to coordinate the
operation of motor centres in the cortex and the brain stem whilst a movement is
ongoing as well as during repetitions of the same movement.
Figure 2. The three functional regions of the cerebellum, namely the vermis, the hemispheres
and flocculonodular lobe have different inputs and outputs (Kandel et al., 2000).

13
Introduction
The role of the cerebellum in the control of human movements was first described in
great detail by the English neurologist Gordon Holmes (Holmes, 1917). Holmes’s
clinical studies on first world war casualties with gunshot injuries restricted to the
cerebellum showed that these patients had lost the ability to move properly. Their
movements were highly ataxic and they had problems with body posture and balance,
and particularly striking were abnormalities in muscle tone. Another dominant finding
was their disability to perform coordinated movements like the finger-to-nose test,
normally performed at ease with eyes shut, but almost impossible for cerebellar
patients even under visual control.
Behavioural evidence from patients with cerebellar lesions indicate that the cerebellum
effects smooth coordinated motor acts by predicting and adjusting for the forces
acting on a limb during complex movement. Goodkin et al. (1993) reported of a
stroke patient who had that part of his cerebellum destroyed that controlled his right
hand. Simple movements like flexing and extending his wrist was unimpaired, when
compared with his left hand. In contrast, compound movements which necessitated
the coordination of multiple joints, like reaching to a visual target and precision
pinch of a seen object were impaired. In a further study of patients with cerebellar
lesions videotaped as they made reaching movements towards a small ball suspended
in front of themselves, Bastian et al. (1996) showed that, in comparison to healthy
subjects, these individuals first reached beyond the target and looped back. In contrast,
the hands of the healthy subjects went directly to the ball. By computing the rotational
forces of each joint, the researchers were able to deduce that the cerebellar patients
were having trouble balancing opposing forces of the multiple joints involved, namely
the wrist, elbow and shoulder, concluding that the cerebellum effects smooth motor
acts by controlling the interaction torques across multiple joints.
In relation to the balancing of forces across multiple joints, the cerebellum is also
thought to be responsible for coordinating the timing of joint movements. Timmann
et al. (1999) investigated the role of the cerebellum in the timing of finger opening as
subjects threw a ball overarm. Patients with cerebellar lesions threw more slowly
than controls, were markedly less accurate, had more variable hand trajectories, and
showed increased variability in the timing, amplitude, and velocity of finger opening,
suggesting that the cerebellum is crucial for accurate timing in complex multi-joint
movements.

14
Chapter 1
1.2.2 Motor learning at a behavioural level
Various bodies of data suggest that the cerebellar cortical circuits are used in learning
motor skills. There is little doubt that the cerebellum is needed for the most simple
types of motor learning such as associative learning. Converging lines of evidence
from lesioning, recording, stimulation, reversible inactivation and brain-imaging
studies indicate that the cerebellum is essential for eyeblink conditioning, a type of
discrete sensory–motor learning (Hesslow and Yeo, 1998). In brief, the animal
associates a tone with an air puff, so that eventually the tone by itself is sufficient to
elicit the blinking of the eye. Selective lesions (electrolytic or chemical) of the
cerebellum prevent the acquisition and retention of conditioned eyeblink responses.
Electrophysiological studies indicate that cells in specific regions of the cerebellum
undergo learning-induced changes during eyeblink conditioning. For example,
Purkinje cells in the cortex (particularly in lobule VI) decrease their activity while
cells in the interpositus nucleus increase their activity, which is consistent with
inhibitory projections from the cortex to the interpositus nucleus. The involvement
of the cerebellum in eyeblink conditioning is also supported by studies that show
that electrical stimulation of the two major afferents to the cerebellum – the mossy
fibers from the pontine nuclei and the climbing fibers from the inferior olive – can
substitute for the peripheral conditioned stimulus and unconditioned stimulus,
respectively.
Another example of learning that the cerebellum plays an important role in is the
vestibulo-ocular reflex, a coordinated response that maintains the eyes on a fixed
target when the head is rotated (Ito, 1984). Lesioning the vestibulocerebellum
obliterates the ability of experimental animals to effect this form of adaptation. The
cerebellum is also involved in more complex motor acts like the control of limb
movement or hand-eye coordination that are acquired through trial and error. Once
the behaviour becomes adapted or learnt, it is performed automatically.
1.2.3 Motor learning at a cellular level
Great efforts are dedicated to correlating behavioural learning at the cellular level
and vice versa. The most fashionable hypothesis is that of Marr-Albus-Ito. In 1969
in his seminal “A Theory of Cerebellar Cortex,” David Marr, proposed that plasticity,
termed long term depression (LTD), at synapses between parallel and Purkinje cells
could be the basis for motor learning (see figure 3). This theory has been further
expanded (Albus 1971; Ito, 1984) and holds that the climbing fibre input to Purkinje
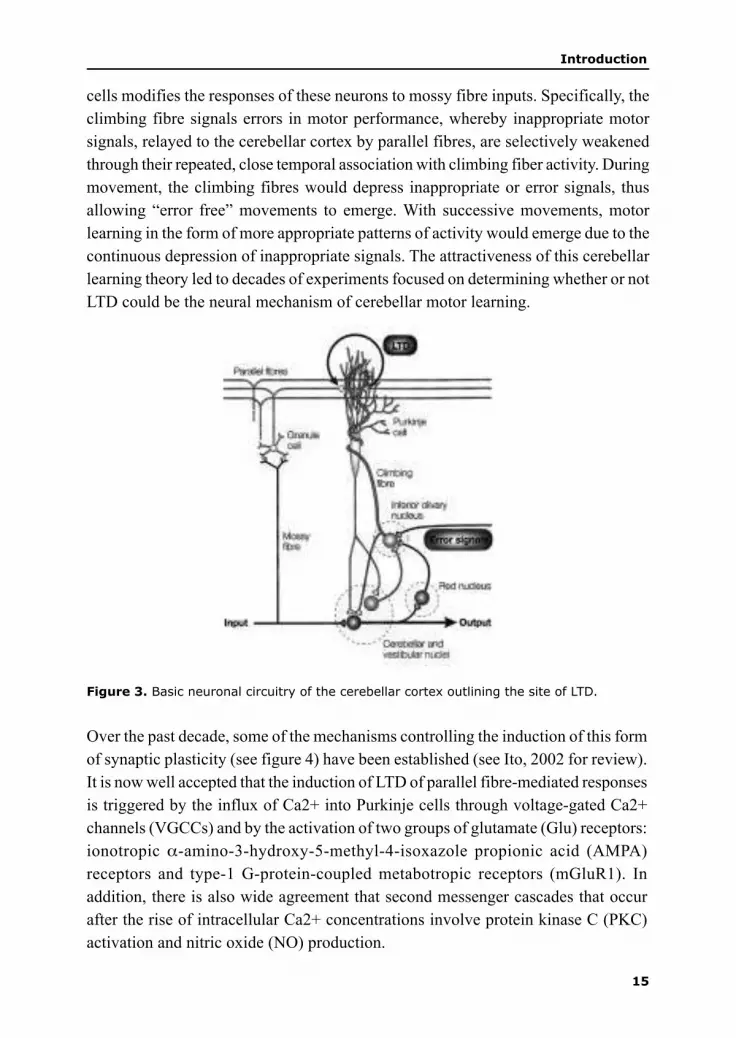
15
Introduction
cells modifies the responses of these neurons to mossy fibre inputs. Specifically, the
climbing fibre signals errors in motor performance, whereby inappropriate motor
signals, relayed to the cerebellar cortex by parallel fibres, are selectively weakened
through their repeated, close temporal association with climbing fiber activity. During
movement, the climbing fibres would depress inappropriate or error signals, thus
allowing “error free” movements to emerge. With successive movements, motor
learning in the form of more appropriate patterns of activity would emerge due to the
continuous depression of inappropriate signals. The attractiveness of this cerebellar
learning theory led to decades of experiments focused on determining whether or not
LTD could be the neural mechanism of cerebellar motor learning.
Figure 3. Basic neuronal circuitry of the cerebellar cortex outlining the site of LTD.
Over the past decade, some of the mechanisms controlling the induction of this form
of synaptic plasticity (see figure 4) have been established (see Ito, 2002 for review).
It is now well accepted that the induction of LTD of parallel fibre-mediated responses
is triggered by the influx of Ca2+ into Purkinje cells through voltage-gated Ca2+
channels (VGCCs) and by the activation of two groups of glutamate (Glu) receptors:
ionotropic α-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid (AMPA)
receptors and type-1 G-protein-coupled metabotropic receptors (mGluR1). In
addition, there is also wide agreement that second messenger cascades that occur
after the rise of intracellular Ca2+ concentrations involve protein kinase C (PKC)
activation and nitric oxide (NO) production.

16
Chapter 1
Whilst most of the current LTD studies have focused on the roles of conventional
neurotransmitters like glutamate, there has been little emphasis on the possible roles
of the plethora of neuropeptides contained in climbing fibres. This route might prove
lucrative in further gaining a composite understanding of climbing fibre action in
motor learning. In an effort to further understand climbing fibre-derived neuropeptide
Figure 4. (for colorinformation: see page 149) Model of signal transduction during LTD.Four categories of molecules are placed in sequential steps, from first messengers to receptors/channels to second messengers and kinases/phosphatases (the phosphorylation–dephosphorylation (PD) system). Blue outlines and arrows indicate candidate modulators oflong-term depression (LTD). Red outlines and arrows indicate candidate mediators. Greencircles indicate candidate coincidence detectors (C1, C2, C3 and C4). δ2R, δ2 receptor; AA,arachidonic acid; AMPAR, α-amino-3-hydroxy-5-methyl-4- isoxazole propionic acid receptor;CRF, corticotropin-releasing factor; CRFR1, CRF type 1 receptor; d, membrane depolarization;DAG, diacylglycerol; GC, guanylyl cyclase; Glu, glutamate; IGF1, insulin-like growth factor1; IGF1R, IGF1 receptor; InsP3R, inositol-1,4,5-trisphophate receptor; MAPK, mitogen-activated protein kinase; mGluR1, type 1 metabotropic glutamate receptor; NO, nitric oxide;PKC, protein kinase C; PKG, protein kinase G; PP2A, protein phosphatase 2A; PTK, proteintyrosine kinase; SG, slow-EPSP (excitatory postsynaptic potential) generator associated withmGluR1 (Ito, 2002).

17
Introduction
function, Miyata et al. (1999) showed that CRF is crucial for the induction of LTD.
With the identification of newer members of the CRF family, like urocortin, and the
characterisation of the receptors through which these peptides act, their precise
localisation within the cerebellar circuitry will provide a crucial stepping stone to
the eventual understanding of their roles in cerebellar function.
Much energy has been invested in attempting to show that LTD is not merely an
artefactual phenomenon. The holy grail of cerebellar functional studies would be to
link the occurrence of LTD, that is learning at a cellular level with learning at a
behavioural level. The behavioural and cellular assessment of mice, mutant for the
key role players in LTD have gone some way in solidifying the LTD hypothesis. In
an elegant study, Ichise et al. (2000) showed that using an mGluR1 knockout mouse,
LTD was abolished. Motor coordination in these mice was impaired strongly
suggesting that LTD underlies motor learning in the cerebellum. Furthermore, De
Zeeuw et al. (1998) have also shown that inhibition of PKC abolishes LTD and motor
learning.
Emerging evidence shows that LTD is not the only type of plasticity resident in the
cerebellum (Hansel et al., 2001). In vitro studies have demonstrated long-term
plasticity at multiple sites in both the cerebellar cortex and deep nuclei. Known sites
include all Purkinje cell inputs: parallel fibre, inhibitory interneuron, and climbing
fibre synapses. In addition to LTD, LTP (long-term potentiation) also had been
observed at the synapse between Purkinje cells and deep nuclear neurons. Finally,
deep nuclear neurons and granule cells exhibit activity-dependent, non-synaptic
changes in intrinsic excitability. Hence, cellular mechanisms of plasticity must be
linked to circuit mechanisms of behaviour to understand learning and memory (Carey
and Lisberger, 2002).
1.3 Cerebellar development
In rodents, there are strong causal relationships between certain developmental stages
of cerebellar and motor behavior. Rats are able to stand only from PD 8 and are also
capable of a walking a few steps (Geisler et al., 1996). This “atactic” pattern of
walking continues until PD 14. From PD 15, in the course of one to two days, this
pattern of immature walking shifts into the fluent and adult-like walking pattern
(Westerga et al., 1990). Recordings of EMG patterns in the hind leg and back muscles
showed that from that age, regular bursts occur in hind leg muscles during the stance
and the swing phase respectively (Westerga and Gramsbergen, 1993,1994). Ablation
of one of the cerebellar hemispheres at PD 5 or 10 does not influence the immature

18
Chapter 1
walking pattern, but the shift into the fluent walking pattern does not occur
(Gramsbergen, 1993) and this indicates that the cerebellum is indeed involved in the
feed-forward programming of posture and movement which is essential for the adult
walking patterns.
The initial phase of neuronal development is governed by intrinsic patterns under the
influence of genetic imprints. However, the secondary phase of development, governed
largely by activity dependent mechanisms, and acting via electrical signalling and
trophic factors is crucial for the further developments of neurites and accounts for
the most active period of neural development and circuitry formation (Wong and
Ghosh, 2002). The temporal and spatial identification of the neurochemical substrates
expressed during development will help to elucidate their roles in the establishment
of the cerebellar circuitry and hence motor activity. This understanding will also
help in the approach to possible therapies of pathologies arising during development
since the cerebellum is especially prone to such perturbation due to its late
development. The studies undertaken in this thesis will be restricted to the rodent.
Hence, unless otherwise specified, the discussion will pertain to the development of
the rodent cerebellum.
1.3.1 Intrinsic development
Birth and migration of cerebellar neurons
In comparison to other brain structures, the cerebellum undergoes a protracted and
belated developmental profile, resulting in a large component of development
occurring postnatally. In humans, the development of the cerebellum (and particularly
the proliferation of future granule cells as indicated by the increase in DNA-P) starts
in the last trimester of pregnancy and continues into the first year of life (Dobbing
and Sand, 1973). In rodents, the cerebellar cortex almost totally develops during the
first three postnatal week (Smart and Dobbing, 1971).
Chick-Quail chimera experiments show that both the mesencephalon and
metencephalon contribute to the developing cerebellum The junction between the
mes- and metencephalon, the isthmus, has been shown to be an important organising
centre (Hatten and Heintz, 1995). This is evidenced by the number of important
secreted and regulated genes expressed in this region. The reciprocal repression of
one of the mouse homologues of the Drosophila gene orthodenticle, Otx2 and Gbx2,
a homologue of the Drosophila gene unplugged forms the isthmus organisor, which
in turn uses Fgf8 (fibroblast growth factor 8) and Engrailed (En1) to pattern the

19
Introduction
prospective mid-/hindbrain region. Cells from both the mesencephalon and the
metencephalon give rise to cerebellar tissues. Mutations in several of these genes
have marked impact upon cerebellar development with the best documented examples
being Wingless 1 (Wnt1) and En1. Knockouts of these genes largely or totally eliminate
the cerebellum (Goldowitz and Hamre, 1998; Wang and Zoghbi, 2001).
Cells in the cerebellum arise from two different germinal matrices, namely the
ventricular epithelium and the rhombic lip (see Goldowitz and Hamre, 1998 for
review). Deep cerebellar nuclei, Purkinje cells, Golgi, stellate and basket cells all
arise from the ventricular neuroepithelium. The first cells to leave the ventricular
zone are the deep cerebellar nuclei and vestibular nuclei at approximately embryonic
day (E) 10-12. Round about E 13, precursor Purkinje cells are born. Shortly after
their final mitosis, they migrate during E 14-17 along radial glial fibres, over the
already formed deep cerebellar nuclei into the cerebellar anlage. It is also at this
stage that Purkinje cells begin to express the calcium binding protein, calbindin.
This successful migration of Purkinje cells is dependent upon the Reelin pathway.
Mutations in the Reln gene or in components of its signaling pathway lead to various
cerebellar defects (Rice and Curran, 1999).
The formation of a secondary germinal matrix from the rhombic lip – the external
granular layer (EGL) – occurs at about the time when nuclear and Purkinje cells
have stopped dividing. As these EGL cells migrate over the cerebellar surface in a
subpial position, the Golgi neurons are born from the diminishing ventricular zone.
Postnatally, in rats, the EGL will seed the internal granular layer with an abundance
of granule cells. Also generated at this time are the stellate and basket cells that
colonise the molecular layer.
In contrast to Purkinje cells that migrate radially in an outward fashion, granule cell
neuroblasts migrate in an inward, radial manner with the presumed aid of radial glial
fibres (more properly known as the Bergmann fibres of the Golgi epithelial cells),
through the molecular layer and past the developing Purkinje cells. Perturbations of
Purkinje cells have devastating effects on granule cell numbers. However, the converse
is not true. This is evidenced by the X-irradiation which destroys granule cells with
no consequences on Purkinje cell number. Granule cells are, however, pivotal for
establishing the precise spatial domains of the cerebellum by their ability to read
migratory signals in extra-cerebellar territories. When they are incapable of doing
this, they travel into ectopic regions, pulling Purkinje cells in this aberrant journey.

20
Chapter 1
1.3.2 Activity-dependent development of the cerebellum
During the first postnatal week, Purkinje cells rearrange from a disordered multi-
tiered layer to their characteristic monolayer, with their apical regions oriented towards
the pial surface. This is also the stage at which swellings in their apical regions of
their somata, the apical caps, become pronounced. The apical caps are the regions
from which the primary dendrites grow out. The two major afferent systems, the
climbing fibres, which originate from inferior olivary neurons and the parallel fibres,
the axons of granule cells heavily influence the growth and differentiation of Purkinje
cell dendrites through activity dependent mechanisms and by chemoattractive and
repulsive means.
By late embryogenesis, multiple inferior olivary axons make numerous synaptic
contacts on somatic spines of Purkinje cells (Morara et al., 2001). From the end of
the first postnatal week, climbing fibres undergo a complex process of regression of
multiple innervation (Crepel et al., 1981) and translocation to the developing Purkinje
cell dendrites ensues (Scelfo et al., 2003). It was initially believed that competition
for the limited space on the Purkinje cell soma and its dendrites results in elimination
of supernumerary climbing fibres (Hume and Purves , 1981; Purves and Hume 1981).
What is clear is that glutamatergic signalling via parallel fibres is crucial in the
regression of multiple climbing fibres. Within a strict time frame (postnatal days 15-
16), N-methyl-D-aspartate (NMDA) receptor signalling between parallel fibres and
Purkinje cells is essential for the reduction of climbing fibres (Kakizawa et al., 2000).
Also, in mGlur1 mutants, multiple climbing fibres persist into adulthood (Ichise et
al., 2000).
As mentioned above, granule cells have little influence on Purkinje cell number.
However, they do play a crucial role in their postnatal development. In cases where
the granule cell population has been disrupted, Purkinje cells fail to form a monolayer,
and show altered morphologies, ranging from misoriented to stunted dendritic trees
(Rakic and Sidman, 1973; Ross et al., 1990). The settling of granule cells below, and
the stacking of parallel fibres above Purkinje cells are thought to, in a mechanical
fashion contribute to the formation of the Purkinje cell monolayer and the outgrowth
and branching of their dendrites (Altman and Bayer, 1997).
There appears to be a reciprocal trophic interaction between climbing fibres, parallel
fibres and Purkinje cells with either element capable of exerting remarkable actions
on the other. Following de-afferentation of climbing fibres by lesioning the inferior
olive with 3-Acetyl pyridine, Purkinje cells undergo hyperspiny transformation and

21
Introduction
become hyperinnervated by parallel fibres (Sotelo and Arsenio-Nunes, 1976). This
phenomenon probably reflects a degree of competition between the climbing fibres
and parallel fibres for the synaptic domains of the Purkinje cell dendritic tree. This is
highlighted by the contrasting effects which these two afferent systems have on
Purkinje cell spine formation. Whilst climbing fibres have an inhibitory effect (Sotelo
et al., 1975), parallel fibres are known to induce spine formation (Baptista et al.,
1994). Furthermore, the climbing fibre-deprived Purkinje cell is able to induce
sprouting from nearby intact climbing fibres and a new arbour is able to fully restore
synaptic connectivity (Rossi et al., 1991). Following Purkinje cell deletion, climbing
fibres become heavily atrophic and reduced in size (Rossi et al., 1993). Climbing
fibres are also capable of innervating Purkinje cells of embryonic cerebellum, grafted
onto an adult unlesioned cerebellum (Armengol et al., 1989; Rossi et al., 1995). In
this case, collaterals of intact climbing fibre arbours elongate through the pial surface
and enter the graft to innervate the Purkinje cells. This intimate reciprocal regulation
between axon and target are under the influence of trophic signals from Purkinje
cells and the conventional neurotransmitters and the host of neuropeptides secreted
by the climbing and parallel fibres.
1.3.2.1 Trophic factors in the cerebellum
Climbing fibres and mossy fibres release aspartate or glutamate which in turn
depolarise their target neurons, the Purkinje cells and granule cells respectively. Co-
localised with these neurotransmitters are a diverse group of neuropeptides such as
calcitonin gene related peptide (CGRP), enkephalin, cholycystokinin, B50,
neuropeptide Y, insulin-growth factor (Kwong et al., 2000). The functional
significance of these peptides in cerebellar afferents remains largely unexplored.
The unequivocal significance of conventional trophic factors in cerebellar
development such as neurtrophin-3 and brain-derived neurotrophic factor (Neveu
and Arenas, 1996; Schwartz et al., 1997; Bates et al., 1999) has been detailed. The
potential of CRF, which is expressed in climbing and mossy fibres, as a possible
trophic factor, facilitating the activity-dependent phase of cerebellar development
remains wholly unexplored.
CRF has been shown to augment the effects of neurotransmitters in the cerebellum.
Indeed, the excitatory effects of glutamate were accentuated with the inhibitory effects
of GABA being attenuated (Bishop, 1990). Crucially, CRF is essential for the induction
of plasticity at the parallel fibre-Purkinje cell synapse (Miyata et al., 1999).
Interestingly, cerebellar CRF (Bishop and King, 1999) and the receptors to which it

22
Chapter 1
binds (Chang et al., 1993) are expressed at early stages of cerebellar development,
prior to any functional synapses being formed. The fact that CRF augments the effects
of glutamate (Bishop, 1990), a neurotransmitter directly implicated in activity
dependent dendritic development (Metzger et al., 1998) suggests that climbing fibre
released CRF might function first as a trophic agent with a shift to that of a
neuromodulator once the functional circuitry has been established. The comparison
of the spatial and temporal localisation of CRF, newer members like urocortin and
their receptors throughout cerebellar ontogeny has not yet been elucidated. The relation
between their expression profiles and well-defined stages of cerebellar and motor
development are crucial for the understanding of the way the “CRF system” influences
motor function. Furthermore, due to the ubiquitous nature of CRF-like peptides,
functional information gleamed from studies into the cerebellum could serve as a
template for future studies in other brain centres where perturbations of the “CRF
system” are strongly implicated in diverse pathologies like Alzheimer’s disease,
epilepsy and major depression.
1.4 Corticotropin releasing factor
The characterisation of a 41 amino acid peptide, termed corticotropin releasing factor
(Vale et al., 1981) proved a fundamental breakthrough in understanding the stress
response. Since then, three new members belonging to this family of neuropeptides
have been characterised, these being urocortin (Vaughan et al., 1995), urocortin II/
stresscopin-related peptide (Lewis et al., 2001) and stresscopin/urocortin III (Reyes
et al., 2001). CRF peptides bind, with varying affinities, to two receptors, termed
CRF-R1 and CRF-R2. Efforts are now concentrated on delineating the distinct roles
each member might subserve in light of their distinct distributional patterns and that
of the receptors.
1.4.1 CRF receptors
cDNAs encoding CRF-R1 and CRF-R2 have been cloned from vertebrates as
evolutionary distant as fish and humans. CRF-R1 and CRF-R2 belong to the class B
subfamily of seven-transmembrane receptors that signal by coupling to G-proteins.
There is one functional variant and several non-functional variants of the CRF-R1, a
415–420-amino-acid polypeptide (Chang et al., 1993; Chen et al., 1993; Vita et al.,
1993). The CRF-R2, a 397–438-amino-acid protein, has three functional splice
variants, CRF-R2A–C (Lovenburg et al., 1995; Perrin et al., 1995). Although their
N-terminal sequences and tissue distribution differ, these splice variants show no

23
Introduction
major pharmacological differences. The CRF-R1 and CRF-R2 are remarkably
homologous (~70% amino acid identity). Both CRF-R1 and CRF-R2 receptors signal
via cAMP as a second messenger.
Figure 5. Schematic overview of the different CRF peptides, their binding affinities to thetwo CRF receptors and their physiological effects. As indicated by the solid arrows, both CRFand UCN bind with relative equal affinities to CRF receptor 1. However, compared to CRF,UCN, UCN II and UCN III bind with greater affinities to CRF receptor two.
1.4.1.2 Localisation of CRF-like peptides in the brain
CRF-mRNA and protein are abundantly distributed in the CNS with major sites of
expression in the parvoventricular nucleus of the hypothalamus, cerebral cortex,
cerebellum and the amygdalar–hippocampal complex, an area important for stress
adaptation, learning and memory. In the periphery, CRF is expressed in the adrenal
gland, testis, placenta, gut, spleen, thymus and skin (Swanson et al., 1983).
In general, there is very little neuroanatomical overlap in the distribution of urocortin
(UCN) and CRF in the brain, suggesting differential functional roles for these peptides.
Although there is some conflict in the literature with respect to the cellular localization
of UCN expression in the rat brain, all studies agree that UCN-immunoreactive (UCN-

24
Chapter 1
ir) perikarya and mRNA expression are most prominent in the Edinger–Westphal
nucleus (EWN) and the lateral superior olive. Other brain regions reported to contain
moderate levels of urocortin mRNA include the cerebellum, hippocampus, neocortex,
olfactory system, basal ganglia, amygdala, and the supraoptic (SON), ventromedial
(VMH), and paraventricular (PVN) nuclei of the hypothalamus (Bittencourt et al.,
1999).
The distribution of urocortin within the human CNS is less well investigated, but
there is evidence of both urocortin mRNA and urocortin-ir in the cerebral cortex,
hypothalamus, medulla, pons, and cerebellum of the human brain, with no significant
regional differences in concentration. Similar to the rat, UCN-ir has also been found
within the human anterior pituitary, particularly in cells that co-express growth
UROCORTIN
CRF
Figure 6. Comparison of the rat brain distribution of CRF and urocortin immunoreactive cellsin the rodent brain. (Smagin et al., 2001). All abbreviations are taken from Paxinos andWatson’s (1982) brain atlas.

25
Introduction
hormone or prolactin (Takahashi et al., 1998; Iino et al., 1999).
UCN II mRNA is predominantly localised subcortically. Major sites of expression
include stress-related cell groups such as the paraventricular, supraoptic, and arcuate
nuclei of the hypothalamus, and the locus coeruleus of the rostral pons. Motor nuclei
of the brainstem (trigeminal, facial, hypoglossal), as well of the spinal ventral horn,
are also identified as sites of UCN II mRNA expression (Lewis et al., 2001).
The main sites of UCN III mRNA expression in the central nervous system include
the hypothalamus, brainstem, and lateral septum bed nucleus of stria terminalis . The
pituitary, cerebellum, and cerebral cortex showed little or no detectable mRNA
expression. In the periphery, UCN III mRNA is expressed in small intestine and
skin, with no detectable expression in heart, aortic vessel, liver, or lung (Reyes et al.,
2001).
1.4.2 Localisation of CRF receptors
In general, there is very little overlap in the distribution of the CRF-R1, CRF-R2a,
and CRF-R2b subtypes in the rat, suggesting distinctive functional roles. The
distribution of CRF-R1 mRNA in mouse is fundamentally similar to that in rat, with
expression predominating in the cerebral cortex, sensory relay nuclei, and in the
cerebellum and its major afferents. CRF-R2 mRNA shows comparable expression in
rat and mouse brain, being distinct from, and more restricted than that of CRF-R1.
Major neuronal sites of CRF-R2 expression included aspects of the olfactory bulb,
lateral septal nucleus, bed nucleus of the stria terminalis, ventromedial hypothalamic
nucleus, medial and posterior cortical nuclei of the amygdala, ventral hippocampus,
Figure 7.Corticotropin-releasing factor receptor (CRF-R) distributions in rodent brain.Schematic drawing of a sagittal section through the rat brain shows the distribution andrelative density of cells expressing CRF-R1 and CRF-R2 mRNAs. The CRF-R2 transcript showsa more restricted distribution that is largely non-overlapping with that of CRF-R1 (Van Pett et
al., 2000).

26
Chapter 1
mesencephalic raphe nuclei, and novel localisations in the nucleus of the solitary
tract and area postrema. Several sites of expression in the limbic forebrain overlap
partially with ones of androgen receptor expression. Rat and mouse pituitary display
CRF-R1 mRNA signal continuously over the intermediate lobe and over a subset of
cells in the anterior lobe, whereas CRF-R2 transcripts show expression mainly in the
posterior lobe (Van Pett et al., 2000).
1.4.3 Behavioral effects of CRF peptides and their receptors
Behavioural studies on the roles of CRF peptides are almost exclusively restricted to
their stress or anxiety inducing effects. Mice null for the CRF-R1 displayed increased
exploratory activity and markedly reduced anxiety-related behavior. The resultant
disruption to the hypothalamic-pituitary-adrenal (HPA) axis also resulted in the
receptor’s failure to elicit the characteristic hormonal response to stress (Timpl et al.,
1998; Smith et al., 1998). CRF-R2 deficient mice showed enhanced anxious behavior
in relation to CRF-R1 mice. The enhanced anxiogenic effect in CRF-R2 deficient
mice was not due to abberrations in the HPA axis, but rather reflected impaired
responses in specific brain regions involved in emotional and autonomic function
(Bale et al., 2000; Kishimoto et al., 2000).
Intracerebroventricular administration of CRF and UCN affects performance in tests
of learning and memory. Under difficult learning conditions (massed trials),
pretreatment with CRF or UCN facilitated the acquisition of spatial navigation in the
Morris water maze. Under less difficult learning conditions (spaced trials), both
peptides impaired water maze performance. With post-training treatment, the peptides
were equipotent in facilitating the consolidation of passive avoidance learning. The
performance-enhancing effects of UCN in both water maze and passive avoidance
paradigms were reversed by pretreatment with a broad CRF receptor antagonist, or
antalarmin, a potent, nonpeptide, CRF-R1 selective receptor antagonist. These findings
are consistent with a hypothesis that CRF receptor agonists affect performance in
tests of learning and memory by increasing arousal.
Chronic treatment with CRA1000, a specific nonpeptidic CRF-R1 selective antagonist
significantly decreased locomotor activity in the dark phase of the diurnal cycle
suggesting that CRF-R1 receptors are involved in the regulation of locomotor activity.
In contrast, UCN II has been shown to induce mild locomotor inhibition. Importantly,
no studies have been undertaken to determine the effects of CRF or UCN on brain
development or motor coordination.

27
Introduction
1.4.4 CRF in the cerebellum
1.4.4.1 Cerebellar CRF distribution
Palkovitz et al. (1987) showed that CRF is localised in the axons of several pre-
cerebellar nuclei. These axons form the two main afferent systems to the cerebellum,
namely the mossy fibre and climbing fibre systems resulting in a prominent CRF
presence in the cerebellum. Since then, the distribution profile of CRF in various
species has been documented.
In the rat cerebellum, van den Dungen et al. (1988) showed that CRF is localised
within climbing and mossy fibres from PD 8 onwards. However, in the mouse, Bishop
and King (1999) have shown that CRF is already expressed at embryonic stages in
cerebellum. The presence of CRF this early could be indicative of it subserving a
different role in this time period as opposed to its purported neuromodulatory function
in the adult structure. The above hypothesis has recently been enhanced by Ha et al.
(2000) who have shown that CRF induces proliferation of cerebellar astrocytes in a
dose dependent manner. Furthermore, CRF has been shown to induce neurite
outgrowth in locus coeruleus derived neurons (Cibelli et al., 2001).
At early postnatal stages, CRF is diffusely distributed throughout the cerebellum
without any strict lobular gradient evident. However, in adulthood, most
immunoreactivity is resident in afferents projecting to the central and posterior lobules,
especially vermal lobules IX and X and the flocculus and paraflocculus. This anterior-
posterior gradient appears to be species specific. In the mouse (Overbeck and King,
1999), most CRF labeled profiles occur in lobules VIII, IX & X and in the rabbit
(Errico and Barmack, 1993), in lobules VIII & IX. In the opossum (Cummings et al.,
1994), a far more even distribution had been described. By implication of the
heterogeneous lobular distribution at adulthood, not all climbing and mossy fibres
contain CRF, suggestive of a rather discrete role in specified areas of the cerebellum.
The influence of CRF on the myriad of developmental processes involved in the
establishment of the functional circuitry, such as axonal-target development,
recognition and synapse formation (Oberdick et al., 1998; Sotelo and Chedotal, 1997;
Sotelo, 1999) and ongoing plasticity of olivocerebellar axis (Strata and Rossi, 1998)
remains to be elucidated. Of particular importance are the signalling factors involved
in the initiation of Purkinje cell dendritic outgrowth and its further development
which results in a structure composed of functionally distinct zones, resulting in
climbing fibres being restricted to the proximal dendritic regions and parallel fibres,
the more distal dendritic compartment.

28
Chapter 1
1.4.4.2 Cerebellar UCN distribution
The localisation of UCN in the cerebellum is less clear. Bittencourt et al. (1999)
reported scant UCN immunoreactivity in the rat cerebellar cortex with the deep nuclei
receiving a sparse to moderate projection. The most prominent UCN-ir projection
to the cerebellar cortex was seen consistently in the granular layer of the flocculus.
The remainder of the cortex contained only widely scattered fibres. Using
radioimmunoassay, Takahashi et al., (1998), showed a prominent expression of UCN
in the human cerebellum. It must be mentioned that none of these studies were
dedicated to exploring the cerebellar expression of UCN. Hence, a comprehensive
evaluation of the distribution of UCN in the cerebellum, during development and
adulthood is mandated.
1.4.4.3 Cerebellar CRF receptor distribution
In the rat, the cerebellum is the most prominent seat of CRF-R1 expression in the
brain. This receptor is first expressed at embryonic day 17. Van Pett et al., (2000)
showed that in adulthood, CRF-R1 is localised over the Purkinje and granule layer
of cerebellar cortex. Each of the deep cerebellar nuclei were strongly and uniformly
positive. Similar labelling intensity was observed in a number of major pre- and
post-cerebellar nuclei, including the red nucleus, lateral nucleus, reticular nucleus
and basilar pontine nucleus. The inferior olive displayed scant evidence of CRF-R1
expression. Bishop et al. (2000) showed that in the mouse cerebellum, CRF-R1 is
distributed over the somata most of Purkinje cells as well as their primary dendrites.
Glial processes in the molecular layer were also immunoreactive. In the granule cell
layer, scattered immunoreactive puncta are present.
The expression of CRF-R2 in the cerebellum is less definitive. Van Pett et al. (2000)
reported a clear absence of CRF-R2 expression in both mouse and rat cerebella. In
contrast, Bishop et al. (2000) reported of immunoreactive puncta throughout the
molecular layer in all lobules. Furthermore, molecular layer interneurons, namely
basket and/or stellate cells were also immunopositive. In the Purkinje cell layer, the
immunolabeling was confined to the basal pole of the Purkinje cell including the
initial axonal segment. In the granule cell layer, labelling was evident in cell bodies
and the initial axonal segments Golgi cells.
All the studies on the cerebellar localisation of CRF receptors have been at the light
microscopical level in the adult mouse. Whilst this provides a good overview of the

29
Introduction
various cells expressing receptor protein, the sub-cellular compartmentation is difficult
to discern. This is crucial since in terms of receptor studies, data on whether they are
pre- or postsynaptically localised or both are of paramount importance if an integrative
picture of their role in the multiple synaptic loci of the cerebellar circuitry is to be
understood. Hence, the ultrastructural distribution in the rat during development is
warranted.
1.5 Scope of the thesis
In the first part of the thesis, complementary immunocytochemical techniques will
be used to map the distributions of CRF, UCN and CRF receptors in the developing
rat cerebellum (chapters 2 to 5). We will use light microscopy to detail their lobular
distributional pattern. This is particularly important in the cerebellum since different
lobules receive inputs from and project to different brain regions. Lobules IX, X, the
flocculus and paraflocculus for example are mainly concerned with vestibular
function. Hence, a precise lobular localisation will provide insight into the potential
physiological roles of the peptide. However, data on the ultrastructural localisation
of the CRF-like peptides and their receptors are crucial if we are to understand the
locus of their action. This is important with respect to the Purkinje cells especially,
since functionally heterologous zones exist on this cell type due to its diverse inputs
it receives. Indeed, climbing fibres form excitatory synapses on the proximal stubby
dendritic spines whereas parallel fibres synapse on the distal thinner spines.
Furthermore, basket and stellate cells form inhibitory synapses on the somata and
dendritic shafts of Purkinje cells respectively. Using pre- and postembedding immuno-
electron microscopy, we will detail the expression of CRF, UCN and CRF receptors
at various developmental stages at the sub-cellular level. The intention is to build a
composite of the potential roles of these proteins in the developing cerebellum.
In chapter 6, emphasis will be placed on the development of the Purkinje cell dendrite
and the possible roles of CRF peptides. The unique morphology of the Purkinje cell
dendritic tree, namely its elaborate arborisation is essential for it’s correct functioning
as an integrator of the sensory signals conveyed via the parallel fibres. The outgrowth
of primary dendrites and their further branching all occurs during the critical second
and third postnatal weeks in the rat, a period when CRF expression becomes fully
manifest, suggesting a causative role for these agents in shaping the dendritic tree.
Using organotypic slice cultures of the postnatal cerebellum, we tested the hypotheses
built upon the localisation data. By applying synthetic CRF, UCN, antagonist against
CRF receptors and secondary messenger pathways to cerebellar slice cultures at crucial

30
Chapter 1
stages of development, we will delineate the contributions of these peptides to Purkinje
cell development.
REFERENCES
Allen, G. Buxton, R.B. Wong, E.C. Courchesne, E. (1997) Attentional activation of the cerebellum
independent of motor involvement. Science. 275, 1940-3.
Albus, J.S. (1971) A theory of cerebellar function. Math Biosc 10: 25-61.
Altman, J. Bayer, S.A. (Eds.) (1997) Development of the cerebellar system. In relation to its evolution,
structure and function. CRC Press.
Armengol, J.A. Sotelo, C. Angaut, P. Alvarado-Mallart, R.M. (1989) Organization of Host Afferents
to Cerebellar Grafts Implanted into Kainate Lesioned Cerebellum in Adult Rats. Eur J Neurosci. 1, 75-
93.
Bishop, G.A. (1990) Neuromodulatory effects of corticotropin releasing factor on cerebellar Purkinje
cells: an in vivo study in the cat. Neuroscience. 39, 251-7.
Bishop, G.A. King, J.S. (1999) Corticotropin releasing factor in the embryonic mouse cerebellum.
Exp Neurol. 160, 489-99.
Bishop, G.A. Seelandt, C.M. King, J.S. (2000) Cellular localization of corticotropin releasing factor
receptors in the adult mouse cerebellum. Neuroscience. 101, 1083-92.
Baptista, C.A. Hatten, M.E. Blazeski, R. Mason, C.A. (1994) Cell-cell interactions influence survival
and differentiation of purified Purkinje cells in vitro. Neuron. 12, 243-60.
Bastian, A.J. Martin, T.A. Keating, J.G. Thach, W.T. (1996) Cerebellar ataxia: abnormal control of
interaction torques across multiple joints. J Neurophysiol. 76, 492-509.
Bates, B. Rios, M. Trumpp, A. Chen, C. Fan, G. Bishop, J.M. Jaenisch, R. (1999) Neurotrophin-3 is
required for proper cerebellar development. Nat Neurosci. 2, 115-7.
Bittencourt, J.C. Vaughan, J. Arias, C. Rissman, R.A. Vale, W.W. Sawchenko, P.E. (1999) Urocortin
expression in rat brain: evidence against a pervasive relationship of urocortin-containing projections
with targets bearing type 2 CRF receptors. J Comp Neurol. 415, 285-312.
Bloedel, J.R. Bracha, V. (1997) Duality of cerebellar motor and cognitive functions. Int Rev Neurobiol.
41, 613-34.
Carey, M. Lisberger, S. (2002) Embarrassed, but not depressed: eye opening lessons for cerebellar
learning. Neuron. 35, 223-6.
Chang, C.P. Pearse, R.V. O’Connell, S. Rosenfeld, M.G. (1993) Identification of a seven transmembrane
helix receptor for corticotropin-releasing factor and sauvagine in mammalian brain. Neuron. 11, 1187-
95.
Chen, R. Lewis, K.A. Perrin, M.H. Vale, W.W. (1993) Expression cloning of a human corticotropin-
releasing-factor receptor. Proc. Natl. Acad. Sci. U S A. 90, 8967-71.
Cibelli, G. Corsi, P. Diana, G. Vitiello, F. Thiel, G. (2001) Corticotropin-releasing factor triggers
neurite outgrowth of a catecholaminergic immortalised neuron via cAMP and MAP kinase signalling
pathways. Eur. J. Neurosci.13, 1339-48.
Contarino, A. Dellu, F. Koob, G.F. Smith, G.W. Lee, K.F. Vale, W.W. Gold, L.H. (2000) Dissociation
of locomotor activation and suppression of food intake induced by CRF in CRFR1-deficient mice.
Endocrinology. 141, 2698-702.
Crepel, F. Delhaye-Bouchaud, N. Dupont, J.L. (1981) Fate of the multiple innervation of cerebellar
Purkinje cells by climbing fibers in immature control, x-irradiated and hypothyroid rats. Brain Res.

31
Introduction
227, 59-71.
Cummings, S.L. Young, W.S III. King, J.S. (1994) Early development of cerebellar afferent systems
that contain corticotropin-releasing factor. J Comp Neurol. 350, 534-49.
De Zeeuw, C.I. Hansel, C. Bian, F. Koekkoek, S.K. van Alphen, A.M. Linden, D.J. Oberdick, J. (1998)
Expression of a protein kinase C inhibitor in Purkinje cells blocks cerebellar LTD and adaptation of
the vestibulo-ocular reflex. Neuron. 20, 495-508.
Doyon, J. Song, A.W. Karni, A. Lalonde, F. Adams, M.M. Ungerleider, L.G. (2002) Experience-
dependent changes in cerebellar contributions to motor sequence learning. Proc Natl Acad Sci. 99,
1017-22.
Dobbing, J. Sands, J. (1973) Quantitative growth and development of human brain. Arch Dis Child.
48, 757-67.
Eccles, J.C. (1967) Circuits in the cerebellar control of movement. Proc Natl Acad Sci. 58, 336-43.
Errico, P. Barmack, N.H. (1993) Origins of cerebellar mossy and climbing fibers immunoreactive for
corticotropin-releasing factor in the rabbit. J Comp Neurol. 336, 307-20.
Geisler, H.C. Westerga, J. Gramsbergen, A. (1996)The function of the long back muscles during postural
development in the rat. Behav Brain Res. 80, 211-5.
Goldowitz, D. Hamre, K. (1998) The cells and molecules that make a cerebellum. Trends Neurosci.
21, 375-82.
Goodkin, H.P. Keating, J.G. Martin, T.A. Thach, W.T. (1993) Preserved simple and impaired compound
movement after infarction in the territory of the superior cerebellar artery. Can J Neurol. 20 Suppl
3:S93-104.
Gramsbergen, A. (1993) Consequences of cerebellar lesions at early and later ages: clinical relevance
of animal experiments. Early Hum Dev. 34, 79-87.
Ha, B.K. Bishop, G.A. King, J.S. Burry, R.W. (2000) Corticotropin releasing factor induces proliferation
of cerebellar astrocytes. J Neurosci Res. 62, 789-98.
Hallonet, M. E. Teillet, M. A. Le Douarin, N. M. (1990) A new approach to the development of the
cerebellum provided by the quail-chick marker system. Development. 108,19-31.
Hallonet, M. E. Le Douarin, N. M. (1993) Tracing neuroepithelial cells of the mesencephalic and
metencephalic alar plates during cerebellar ontogeny in quail-chick chimaeras. Eur. J. Neurosci. 5,
1145-1155.
Hansel, C. Linden, D.J. D’Angelo, E. (2001) Beyond parallel fiber LTD: the diversity of synaptic and
non-synaptic plasticity in the cerebellum. Nat Neurosci. 4, 467-75.
Hatten, M. E. Heintz, N. (1995) Mechanisms of neural patterning and specification in the developing
cerebellum. Annu. Rev. Neurosci. 18, 385-408.
Hesslow, G. Yeo, C. (1998) Cerebellum and learning: a complex problem. Science. 280, 1817-9.
Holmes, J. (1917) The symptoms of acute cerebellar injuries due to gunshot injuries. Brain. 40, 461-
535.
Hume, R.I. Purves, D. (1981) Geometry of neonatal neurones and the regulation of synapse elimination.
Nature. 293, 469-471.
Ichise, T. Kano, M. Hashimoto, K. Yanagihara, D. Nakao, K. Shigemoto, R. Katsuki, M. Aiba, A.
(2000) mGluR1 in cerebellar Purkinje cells essential for long-term depression, synapse elimination,
and motor coordination. Science. 288, 1832-5.
Iino, K. Sasano, H. Oki, Y. Andoh, N. Shin, R.W. Kitamoto, T. Takahashi, K. Suzuki, H. Tezuka, F.
Yoshimi, T. Nagura, H. (1999) Urocortin expression in the human central nervous system. Clin

32
Chapter 1
Endocrinol. 50, 107-14.
Ito, M. (1984) The Cerebellum and Neural Control. New York: Raven.
Ito, M. (2002) The molecular organization of cerebellar long-term depression. Nat Rev Neurosci. 3,
896-902.
Kakizawa, S. Yamasaki, M. Watanabe, M. Kano, M. (2000) Critical period for activity-dependent
synapse elimination in developing cerebellum. J Neurosci. 20, 4954-61.
Kandel, E.R. Schwartz, J.H. Jessell, T.M. (2000) Principles of neural science. McGraw-Hill.
Koob, G.F. Heinrichs, S.C. (1999) A role for corticotropin releasing factor and urocortin in behavioral
responses to stressors. Brain Res. 848, 141-52.
Kwong, W.H. Chan, W.Y. Lee, K.K. Fan, M. Yew, D.T. (2000) Neurotransmitters, neuropeptides and
calcium binding proteins in developing human cerebellum: a review. Histochem J. 32, 521-34.
Lewis, K. Li, C. Perrin, M.H. Blount, A. Kunitake, K. Donaldson, C. Vaughan, J. Reyes, T.M. Gulyas,
J. Fischer, W. Bilezikjian, L. Rivier, J. Sawchenko, P.E. Vale, W.W. (2001).Identification of urocortin
III, an additional member of the corticotropin-releasing factor (CRF) family with high affinity for the
CRF2 receptor. Proc Natl Acad Sci. 98, 7570-5.
Lovenberg, T.W. Liaw, C.W. Grigoriadis, D.E. Clevenger, W. Chalmers, D.T. De Souza, E.B. Oltersdorf,
T. (1995) Cloning and characterization of a functionally distinct corticotropin-releasing factor receptor
subtype from rat brain. Proc. Natl. Acad. Sci. U S A. 92, 836-40.
Marr, D. (1969) A theory of cerebellar cortex. J Physiol. 202, 437-70.
Marin, F. Puelles, L. (1994) Patterning of the embryonic avian midbrain after experimental inversions:
a polarizing activity from the isthmus. Dev. Biol. 163, 19–37.
Metzger, F. Wiese, S. Sendtner, M. (1998) Effect of glutamate on dendritic growth in embryonic rat
motoneurons. J Neurosci. 18, 1735-42.
Miyata, M. Okada, D. Hashimoto, K. Kano, M. Ito, M. (1999) Corticotropin-releasing factor plays a
permissive role in cerebellar long-term depression. Neuron. 22, 763-75.
Morara, S. van der Want, J.J. de Weerd, H. Provini, L. Rosina, A. (2001) Ultrastructural analysis of
climbing fiber-Purkinje cell synaptogenesis in the rat cerebellum. Neuroscience. 108, 655-71.
Muller, R.A. Courchesne, E. Allen, G. (1998) The cerebellum: so much more. Science. 282, 879-80.
Neveu, I. Arenas, E. (1996) Neurotrophins promote the survival and development of neurons in the
cerebellum of hypothyroid rats in vivo. J Cell Biol. 133, 631-46.
Overbeck, T.L. King, J.S. (1999) Developmental expression of corticotropin releasing factor in the
postnatal murine cerebellum. Brain Res Dev Brain Res. 115, 145-59.
Palkovits, M. Leranth, C. Gorcs, T. Young, W.S. (1987) Corticotropin-releasing factor in the
olivocerebellar tract of rats: demonstration by light- and electron-microscopic immunohistochemistry
and in situ hybridization histochemistry. Proc Natl Acad Sci. 84, 3911-5.
Perrin, M. Donaldson, C. Chen, R. Blount, A. Berggren, T. Bilezikjian, L. Sawchenko, P. Vale, W.
(1995) Identification of a second corticotropin-releasing factor receptor gene and characterization of a
cDNA expressed in heart. Proc. Natl. Acad. Sci. U S A. 92, 2969-73.
Purves, D. Hume, R.I. (1981) The relation of postsynaptic geometry to the number of presynaptic
axons that innervate autonomic ganglion cells. J Neurosci. 1, 441-452,.
Rakic, P. Sidman, R.L. (1973) Organization of cerebellar cortex secondary to deficit of granule cells
in weaver mutant mice. J Comp Neurol. 152, 133-61.
Reyes, T.M. Lewis, K. Perrin, M.H. Kunitake, K.S. Vaughan, J. Arias, C.A. Hogenesch, J.B. Gulyas,
J. Rivier, J. Vale, W.W. Sawchenko, P.E. (2001) Urocortin II: a member of the corticotropin-releasing

33
Introduction
factor (CRF) neuropeptide family that is selectively bound by type 2 CRF receptors. Proc Natl Acad
Sci. 98, 2843-8.
Rice, D.S. Curran, T. (1999) Mutant mice with scrambled brains: understanding the signalling pathways
that control cell positioning in the CNS. Genes Dev. 13, 2758-73.
Ross, M.E. Fletcher, C. Mason, C.A. Hatten, M.E. Heintz, N. (1990) Meander tail reveals a discrete
developmental unit in the mouse cerebellum. Proc Natl Acad Sci. 87, 4189-92.
Rossi, F. Wiklund, L. van der Want, J.J. Strata. P. (1991) Reinnervation of cerebellar Purkinje cells by
climbing fibres surviving a subtotal lesion of the inferior olive in the adult rat. I. Development of new
collateral branches and terminal plexuses. J Comp Neurol. 308, 513-35.
Rossi, F. Borsello, T. Vaudano, E. Strata, P. (1993) Regressive modifications of climbing fibres following
Purkinje cell degeneration in the cerebellar cortex of the adult rat. Neuroscience. 53, 759-78.
Rossi, F. Jankovski, A. Sotelo, C. (1995) Differential regenerative response of Purkinje cell and inferior
olivary axons confronted with embryonic grafts: environmental cues versus intrinsic neuronal
determinants. J Comp Neurol. 359, 663-77.
Scelfo, B. Strata, P. Knopfel, T. (2003) Sodium imaging of climbing fiber innervation fields in
developing mouse Purkinje cells. J Neurophysiol.
Schwartz, P.M. Borghesani, P.R. Levy, R.L. Pomeroy, S.L. Segal, R.A. (1997) Abnormal cerebellar
development and foliation in BDNF-/- mice reveals a role for neurotrophins in CNS patterning. Neuron.
19, 269-81.
Smagin, G.N. Heinrichs, S.C. Dunn, A.J. (2001) The role of CRH in behavioural responses to stress.
Peptides. 22, 713-24.
Smith, G.W. Aubry, J.M. Dellu, F. Contarino, A. Bilezikjian, L.M. Gold, L.H. Chen, R. Marchuk, Y.
Hauser, C. Bentley, C.A. Sawchenko, P.E. Koob, G.F. Vale, W. Lee, K.F. (1998) Corticotropin releasing
factor receptor 1-deficient mice display decreased anxiety, impaired stress response, and aberrant
neuroendocrine development. Neuron. 20, 1093-102.
Sotelo, C. Hillman, D.E. Zamora, A.J. Llinas, R. (1975) Climbing fiber deafferentation: its action on
Purkinje cell dendritic spines. Brain Res. 98, 574-81.
Sotelo, C. Arsenio-Nunes, M.L. (1976) Development of Purkinje cells in absence of climbing fibers.
Brain Res. 111, 289-95.
Swanson, L.W. Sawchenko, P.E. Rivier, J. Vale, W.W. (1983) Organization of ovine corticotropin-
releasing factor immunoreactive cells and fibers in the rat brain: an immunohistochemical study.
Neuroendocrinology. 36, 165-86.
Takahashi, K. Totsune, K. Sone, M. Murakami, O. Satoh, F. Arihara, Z. Sasano, H. Iino, K. Mouri, T.
(1998) Regional distribution of urocortin-like immunoreactivity and expression of urocortin mRNA
in the human brain. Peptides. 19, 643-7.
Timmann, D. Watts, S. Hore, J. (1999) Failure of cerebellar patients to time finger opening precisely
causes ball high-low inaccuracy in overarm throws. J Neurophysiol. 82, 103-14.
Timpl, P. Spanagel, R. Sillaber, I. Kresse, A. Reul, J.M. Stalla, G.K. Blanquet, V. Steckler, T. Holsboer,
F. Wurst, W. (1998) Impaired stress response and reduced anxiety in mice lacking a functional
corticotropin-releasing hormone receptor 1. Nat Genet. 19, 162-6.
Takahashi, K. Totsune, K. Sone, M. Murakami, O. Satoh, F. Arihara, Z. Sasano, H. Iino, K. Mouri, T.
(1998) Regional distribution of urocortin-like immunoreactivity and expression of urocortin mRNA
in the human brain. Peptides. 19, 643-7.
Thomas, K.R. Capecchi, M.R. (1990) Targeted disruption of the murine int-1 proto-oncogene resulting
in severe abnormalities in midbrain and cerebellar development. Nature. 346, 847–850.

34
Chapter 1
Van Pett, K. Viau, V. Bittencourt, J.C. Chan, R.K. Li, H.Y. Arias, C. Prins, G.S. Perrin, M. Vale, W.
Sawchenko, P.E. (2000) Distribution of mRNAs encoding CRF receptors in brain and pituitary of rat
and mouse. J Comp Neurol. 428, 191-212.
Valdez, G.R. Inoue, K. Koob, G.F. Rivier, J. Vale, W. Zorrilla, E.P. (2002) Human urocortin II: mild
locomotor suppressive and delayed anxiolytic-like effects of a novel corticotropin-releasing factor
related peptide. Brain Res. 943, 142-50.
Vale, W. Spiess, J. Rivier, C. Rivier, J. (1981) Characterization of a 41-residue ovine hypothalamic
peptide that stimulates secretion of corticotropin and beta-endorphin. Science. 213, 1394-7.
van den Dungen, H.M. Groenewegen, H.J. Tilders, F.J. Schoemaker, J. (1988) Immunoreactive
corticotropin releasing factor in adult and developing rat cerebellum: its presence in climbing and
mossy fibres. J Chem Neuroanat. 1, 339-49.
Van Pett, K. Viau, V. Bittencourt, J.C. Chan, R.K. Li, H.Y. Arias, C. Prins, G.S. Perrin, M. Vale, W.
Sawchenko, P.E. (2000) Distribution of mRNAs encoding CRF receptors in brain and pituitary of rat
and mouse. J. Comp. Neurol. 428, 191-212.
Vaughan, J. Donaldson, C. Bittencourt, J. Perrin, M.H. Lewis, K. Sutton, S. Chan, R. Turnbull, A.V.
Lovejoy, D. Rivier, C. et al. (1995) Urocortin, a mammalian neuropeptide related to fish urotensin I
and to corticotropin-releasing factor. Nature. 378, 233-4.
Vita, N. Laurent, P. Lefort, S. Chalon, P. Lelias, J.M. Kaghad, M. Le Fur, G. Caput, D. Ferrara, P.
(1993) Primary structure and functional expression of mouse pituitary and human brain corticotrophin
releasing factor receptors. FEBS. Lett. 335, 1-5.
Wang, V.Y. Zoghbi, H.Y. (2001) Genetic regulation of cerebellar development. Nat Rev Neurosci. 2,
484-91.
Westerga, J. Gramsbergen, A. (1994) Development of the EMG of the soleus muscle in the rat. Brain
Res Dev Brain Res. 80, 233-43.
Wong, R.O. Ghosh, A. (2002) Activity-dependent regulation of dendritic growth and patterning. Nat
Rev Neurosci. 3, 803-12.
Wurst, W. Auerbach, A.B. Joyner, A.L. (1994) Multiple developmental defects in Engrailed-1 mutant
mice: an early mid-hindbrain deletion and patterning defects in forelimbs and sternum. Development.
120, 2065-75.

35
CHAPTER 2
The expression of Corticotropin releasing factor in thedeveloping rat cerebellum:
a light and electron microscopic study
J. D. Swinny1, D. Kalicharan1 , J. IJkema-Paassen2, J.J.L. van der Want1,
A. Gramsbergen2
1 Laboratory for Cell Biology and Electron Microscopy, & Graduate School of
Behavioural and Cognitive Neuroscience, University of Groningen,
The Netherlands;2 Department of Medical Physiology, University of Groningen, The Netherlands.

Chapter 2
ABSTRACT
In the cerebellum, climbing fibers containing corticotropin releasing factor (CRF)
are crucial for the induction of long term depression at the Purkinje cell-parallel
fiber synapse, a type of synaptic plasticity proposed as the cellular basis of learning.
, CRF expression commences at late embryonic stages, prior to the formation
of any functional synapses. Prevailing thought leans towards the supposition of CRF,
initially playing a role in establishing the cerebellar circuitry, with a shift in adulthood
to that of a neuromodulator. In the present study, we detail the spatial and temporal
distributional profile of CRF to further understand the possible roles of this peptide
in the developing rat cerebellum. The study showed that the expression of CRF was
not evenly distributed rather exhibiting inter - and intra-lobular heterogeneity, being
localised in variable amounts in mossy fiber rosettes and climbing fiber terminal
profiles from as early as postnatal day 3. In the posterior regions especially lobules
IX and X, CRF labelling was more prominent in comparison to the central (V-VIII)
and anterior lobules (I-IV). This disparate lobular distribution persisted through to
adulthood. No cerebellar neurons were conclusively found to contain CRF.
Furthermore, between postnatal days 6 to 9, in the posterior lobules, CRF labelling
at the level of the Purkinje cell clearly showed the characteristic climbing fiber terminal
arrangement related to the capuchon stage whereas in the anterior lobules, labelling
is confined to the perisomatic profiles. These differences might signify that
translocation of the climbing fibers from the Purkinje cell soma to its dendrite is not
uniform for all climbing fibers with CRF possibly playing a facilitative role in this
process. CRF immunoreactivity was also zonally arranged in sagittal bands across
the molecular layer, these appearing as early as postnatal day 9 and persisting through
to adulthood indicating that not all climbing fibers are immunoreactive. Since not all
climbing fibers contain CRF, the process of long term depression and hence learning
could possibly not be uniform across the cerebellum but be limited to certain zones.
The above study lends credence to the supposition that CRF serves, at least, a dual
role in the cerebellum, facilitating postnatal development as well as acting as a
neuromodulator once functional synapses have formed.

37
the expression of corticotropin
INTRODUCTION
The processes involved in the development of cerebellar circuitry eventually manifest
in a structure pivotal for the fine-tuning of motor coordination (see Welsh et al. 1995;
ckelgren 1998) and higher cognitive functions (Muller et al. 1998). An
understanding of these events could provide a convenient paradigm for neuronal
learning as well as that of brain behaviour relationships. Attention has recently shifted
towards factors purportedly involved in development and plasticity in the cerebellum
(Sotelo and Chedotal, 1997; Chen and Tonegawa, 1997), one of these being
corticotropin-releasing factor (CRF) (King et al. 1997)
The conventional view of CRF has always been that of an agent involved in the
hypothalamus-pituitary-adrenal response to stress (Vale et al. 1981) as well as other
central nervous system and immune disorders (De Souza, 1995). Recently, CRF has
been found to play an important neuromodulatory role in the adult cerebellum. Miyato
(1999) have shown this neuropeptide to be crucial in the induction of long term
depression (LTD), a form of synaptic plasticity proposed as the cellular basis of
learning in the cerebellum (Ito, 1982). Also, Bishop and King (1992) have shown
that in the adult cerebellum, CRF augments synaptic efficacy at the mossy fiber –
climbing fiber – Purkinje cell synapses.
In contrast to its more established role in the adult cerebellum, the function of CRF
in the postnatal cerebellum remains enigmatic. Van den Dungen et al. (1988) showed
from postnatal day (PD) 8 in the rat, CRF is found in the two main afferent
systems viz. mossy fibers and climbing fibers. However, Bishop and King (1999)
have shown that in the mouse, CRF immunoreactivity is present in the cerebellum
from birth, long before these afferent systems have formed their adult connections.
Up to PD15 there is also extensive immunoreactivity in the external granular layer.
This layer consists largely of migrating neurons. The presence of CRF this early
could be indicative of it subserving a different role in this time period as opposed to
its purported neuromodulatory function in the adult structure. The above hypothesis
has recently been enhanced by Ha et al. (2000) who have shown that CRF induces
proliferation of cerebellar astrocytes in a dose dependent manner. The influence of
CRF on the myriad of developmental processes involved in the establishment of
functional circuitry such as axonal target recognition and synapse formation (Oberdick
1998; Sotelo and Chedotal, 1997; Sotelo, 1999) and ongoing plasticity of
olivocerebellar axis (Strata and Rossi, 1998) remains to be elucidated.
Labelling studies in the mouse have shown that at early stages, CRF is evenly
distributed throughout the cerebellum with a more discrete lobular localisation in the
adult structure (Overbeck and King, 1999; Yamano and Tohyama, 1994). This lobular

Chapter 2
localisation of CRF appears to be species specific, e.g. in the mouse (Overbeck and
King, 1999), most CRF labelled profiles occur in lobules VIII, IX & X and in the
rabbit (Errico and Barmack, 1993), in lobules VIII & IX. In the opossum (Cummings
et al., 1994), a far more even distribution had been described. By implication of the
heterogeneous lobular distribution at adulthood, not all climbing and mossy fibers
contain CRF suggestive of a rather discrete role in specified areas of the cerebellum.
This raises the question of the role that CRF plays in the development of cerebellar
circuitry and whether a specific relationship might exist with its expression and motor
development in the early postnatal period. Also, since CRF is important for LTD and
that CRF expression develops differently in distinct parts of the cerebellum, is the
phenomenon of LTD uniform throughout this structure?
address these issues, as a first step in the present study, we mapped the topographical
distribution of CRF immunoreactivity in the developing rat cerebellum with a view
to eventually relating its localisation to cerebellar development and motor behaviour.
Experimental procedures
Immunocytochemistry
Black-hooded Lister rats were studied at the following postnatal days:
3,6,9,12,15,20,25,30 and 40. The day of birth was considered PD 0. An average of
six animals were used at each age. Ethical approval to conduct the study was obtained
from the Ethics Committee on Animal Experimentation, University of Groningen.
forts were made to minimise the number of animals used and their suffering.
Animals were anaesthetised with ether and perfused transcardially with first a solution
containing 2% PVP, 0.4% NaNO3 in 0.1 M phosphate buffer (pH 7.4), followed by
4% paraformaldehyde, 0.2% picric acid in 0.1M phosphate buffered saline (PBS)
(pH 7.4). The brains were removed and stored in the same paraformaldehyde/picric
acid/PBS solution.
For light microscopy, brains were stored overnight in 30% sucrose to cryoprotect the
The tissue was frozen and 20mm thick cryosections were cut in the sagittal
wo animals per age group were also sectioned in the transverse plane.
Immunoreactivity was visualised by the avidin-biotin-peroxidase complex method,
performed as described previously by Yamano and Tohyama, 1994. Briefly, free-
floating sections were immersed for two hours in a pre-incubation medium containing
1% normal goat serum, 1% bovine serum albumin, 0.1% glycine, 0.1% lysine, 0.1%
cold water fish gelatine and 0.05% triton in 0.1M TRIS buffered saline (TBS) (pH
The sections were then incubated with rabbit anti-CRF antibodies (Peninsula
Laboratories, San Carlos, CA), reactive for human and rat (diluted 1:1000 with pre-

39
the expression of corticotropin
incubation medium) overnight at room temperature. After washing with TBS, the
sections were incubated with biotinylated goat anti-rabbit IgG (diluted 1:200 in pre-
incubation medium) for two hours at room temperature. After further washing, the
sections were incubated with avidin-biotin-complex (VectastainR ABC kit) for one
hour at room temperature. Immunoreactivity was visualised by incubation with 5mg
3,3 diaminobenzidine and 0.03% hydrogen peroxide in 10ml of 0.01 M PBS (pH
7.4) for 5 to 10 minutes at room temperature. The reaction was stopped by washing
the sections in TBS. The sections were mounted on gelatine-coated slides, counter
stained with cresyl violet for 2 minutes, dehydrated and coverslipped with Permount.
Evaluation of sections
For light microscopical evaluation of the labelling pattern, sagittal and transverse
sections were studied. At each age, images of sections were projected onto paper
using a projection microscope and these were plotted by hand. From these plots, a
composite sketch was drawn summarising the labelling pattern for that age group.
Selected sections were photographed on a Zeiss microscope and digitally adjusted to
enhance the contrast.
For electron microscopy 50mm sections were cut on a vibratome and processed as
for light microscopy with immunoreactivity being enhanced according to the gold
substituted silver peroxidase method (van den Pol, 1985). Sections were then
osmicated in 1% OsO4,
1.5% potassium hexacyanoferrate in 0.1M cacodylate buffer
(pH 7.6) for 15 minutes, dehydrated in a graded series of ethanols and embedded in
Epon. Ultrathin sections were cut on an LKB Ultratome, counterstained with uranyl
acetate and lead citrate and inspected with a Philips CM 100 transmission microscope.
Sections were processed as above with the omission of the primary antibody.
TS
The light microscopical analysis of sagittal sections used in this study showed a
distinct lobular distribution of CRF which changed with age, hence emphasis will be
placed thereon. The nomenclature of Altman and Bayer, (1997) was adopted
Postnatal day 3
At this stage, only the five major lobes of the cerebellum can be distinguished viz.
the anterobasal lobe (lobules I-III), the anterodorsal lobe (lobules IV-V), the central

Chapter 2
Figure 1. Schematic representation of mid-sagittal sections through the vermis of the ratcerebellum indicating CRF labelling at different postnatal ages. (A) Lobular nomenclatureadopted according to Altman and Bayer (1997). (B) At PD 3 CRF labelling was evenly distributedand appeared in the form of dot-like puncta (arrow) and isolated fiber-like profiles in thewhite matter (arrowhead). (C) At PD 6 the posterior lobules showed the most labelling in theform of fibers (arrowhead) as well as rosettes (arrow) in the internal granular layer as well asfibers around the Purkinje cell soma. On occasion, isolated fibers penetrated into the molecular
er. The least amount of labelling was to be found in the anterior lobules (especially I andII) and it was located primarily in the region of the Purkinje cell body. The internal granular
er was almost empty. (D) At PD 12 there was an overall increase in the intensity oflabelling throughout all lobules with many more CRF positive climbing fibers having penetratedinto the molecular layer of lobules IX and X. Also, clearly distinguishable climbing fibersbecame evident in the anterior lobules. (E and F) PD 15 to 40 showed the final maturation ofthe labelled profiles. The most intense labelling was found in the apical regions of lobules IXand X closely followed by the central lobules (V, VI, VIII) and finally the anterior lobules (I-IV).

41
the expression of corticotropin
lobe (lobules VI-VIII), the posterior lobe (lobule IX) and the inferior lobe (lobule X)
(fig 1 B). On the whole, CRF labelling was extremely weak, taking the appearance
of dot-like puncta and tiny individual fibers (fig 2 A and B). Although weak and
ficult to visualise and conclude any precise lobular specificity, these profiles tended
to appear in the posterior lobe and inferior lobe and to a lesser extent in the central
The dot-like puncta occurred mainly in the formative internal granular layer
with the fiber-like profiles at the level of the Purkinje cells as well as in the external
granular layer. It was impossible to distinguish whether these were immature mossy
or climbing fibers. Also in evidence were isolated fibers entering the cerebellum
presumably from the inferior cerebellar peduncle.
Postnatal day 6
At this stage, the adult-like lobular architecture of the cerebellum could be clearly
distinguished (fig 1 C). Coupled with the rapid maturation of cerebellar morphology
was a striking increase in the intensity of the labelled profiles. The labelling was
Figure 2. Nomarski interference photomicrographs of CRF labelling at early stages.(A and B) At PD 3 isolated profiles (solid arrow heads) and puncta (arrows) appeared laterally(A) as well as more medially (B) in the developing lobules. (C) At PD 6 in lobule II, labelling
as concentrated at the level of the Purkinje cell soma (open block arrows) with labelling inthe internal granular layer extremely weak. (D) In lobule X at this stage, fiber labelling wasin the internal granular layer (open arrow head), the Purkinje cell layer as well as the molecular
er (block arrow).

Chapter 2
found almost exclusively in the developing main cerebellar afferents viz. mossy fibers
and climbing fibers. However, at these early stages, there was also extensive labelling
in the external granular layer.
Lobules X and IXa,b displayed fairly dense labelling. However, it was not evenly
distributed throughout the lobule, predominating in the lateral regions and especially
at the lobular apex (fig 2 D). The most abundant labelling occurred in the internal
granular layer in the form of mossy fiber rosettes as well as axons of climbing fibers.
ficult to notice any discernable difference in the intensity of labelling between
mossy fibers and climbing fibers. There was little or no labelling in the folia. Between
PD 6 and 9, some labelled fibers, presumably climbing fibers at the level of the
Purkinje cells were between the capuchon stage and that of pericellular nesting.
The distributional pattern was similar in lobules V, VI, VII and VIII and IXc, but
with much less intensity as well as frequency in terms of labelled profiles. Here, the
labelling was mainly in the internal granular layer. Mossy fiber rosettes as well as
climbing fibers could on occasion be clearly distinguished. The climbing fibers
Figure 3. Photomicrographs of CRF labelling at PD 12. (A) shows that in lobule II, labellingas in the form of puncta primarily at the Purkinje cell layer and that it was not evenly
distributed throughout the lobule appearing weak in the folial regions. (B) Shows that isolatedfibers had penetrated into the molecular layer (arrows). (C) In contrast, lobule X showed afar more even distribution of labelled profiles. These being in the internal granular layer,Purkinje cell layer and molecular layer. (D) Shows that certain climbing fiber profiles (arrowhead) were clearly at a different stage of development compared with those in lobule II,
having penetrated higher into the molecular layer.

43
the expression of corticotropin
sporadically showed pericellular nests on the somata of Purkinje cells. The appearance
of the labelling in the entire anterobasal lobe (lobules I, II and III) as well as the
remaining lobule of the anterodorsal lobe (IV) was in stark contrast to the rest of the
cerebellum taking on the form of numerous densely labelled puncta or pericellular
nests surrounding the Purkinje cell soma without any axonal processes being labelled
Figure 4. Photomicrographs comparing CRF labelling in lobules II and X at PD 15 to 40. (A)In lobule II at PD 15, the pattern of labelling resembled that of the other lobules, however,labelled profiles were still far less numerous than in lobule X at the corresponding stage. (B)An enlargement shows that labelling was concentrated at the apex of lobule IIa. (C) Showsthat lobule X was far more extensively labelled, however, there also existed regions of weakerlabelling especially in the folium (arrow heads). (D) Shows the full extent of labelled profilesespecially in the apex. (E) Shows the maturity of climbing fibers in lobule X and (F) showsmossy fiber rosettes (arrows) as well as fibers in the internal granular layer at PD 24.

Chapter 2
The internal granular layer was almost completely absent of any mossy or
climbing fibers. This pattern of labelling began to taper off at PD 12 when fiber-like
profiles appeared.
Electron microscopic data showed that labelled profiles, presumably climbing fibers,
had at this stage made synapses on the soma of Purkinje cells (fig 5 C) as well as on
the proximal dendrites (fig 5 D). It was difficult to notice any discernable differences
between synapses made in the various lobules.
Postnatal days 9 and 12
At these stages, there was an overall increase in the intensity of the labelled profiles
(fig 1 D). In those lobules with an already intense labelling (X and IX), there was a
more even distribution throughout these lobules (fig 3 C). One also observed single
translocations of climbing fiber profiles from the soma of Purkinje cell to their
dendrites in the molecular layer (fig 3D). Evidence for the apparent increased number
of synapses formed in the molecular layer was apparent at the electron microscopic
level (fig 6 B).
Figure 5. (A) Low-power photomicrograph and (B) magnified region of CRF labelling in theflocculus and paraflocculus regions. (C) Photomicrograph of a transverse section at PD 18showing the tendency for labelled profiles to be arranged in sagittal bands. (D) Photomicrographof a control section showing no labelling.

45
the expression of corticotropin
The anterior and central lobules still lagged far behind the posterior and inferior
lobules in terms of extent and intensity of labelled profiles. From PD 12 onwards,
the punctate profiles in lobules I-III were on the wane giving rise to isolated mossy
fiber and climbing fiber profiles (fig 3 A and B). The climbing fibers appeared to be
at an earlier stage of development than those in lobules IX and X having not yet
translocated from the Purkinje cell soma to its dendrites. The mossy fibers as well
appeared far less numerous and labelled less intense than those in lobules IX and X.
PD 9 was the first stage to observe a zonal arrangement of labelled profiles in the
transverse plane. This occurred at the molecular layer as well as in the internal granular
. Fig 5 C shows this arrangement more clearly at PD 18. The above distributional
pattern of labelling is followed through to adulthood.
Figure 6. Electron micrographs of CRF labelled profiles PD 6. (A) Survey of CRF labelling inlobule II and (B) in lobule X. Calibration bars = 0.5µm. Labelling was found primarily aroundthe Purkinje cell somata at the level of the nucleus (Pc nuc). In lobule X, CRF containingprofiles made synaptic contacts which resembled those in lobule II. These varicose profilescontained spherical vesicles and occasional mitochondria. (C) These synaptic contacts wereat the level of the Purkinje cell soma (peri-somatic) (arrow heads) or (D) on the stubbydendrites of the Purkinje cell (arrows). Calibration bars = 0.25µm.

Chapter 2
Postnatal day 15
At this stage the distributional pattern was characterised by a rapid advancement in
the number and intensity of labelled profiles in those areas (anterior and central
lobules) that up till now were slower to develop (fig 1 D). The intensity as well as the
extent was greatly increased but still did not approach that of lobules IX and X. In
lobules IX and X, the appearance of synapses formed on the Purkinje cell soma was
infrequent with most climbing fibers traversing the soma (fig 7 A) to form synapses
in the molecular layer.
Figure 7. Electron micrographs of CRF labelled profiles at PD 12 in lobule X. (A) Survey atthe level of the Purkinje cell soma. Calibration bar = 2µm. (B) Enlargement showing a CRFprofile making a triple synapse with a Purkinje cell spine (arrow heads). Calibration bar =
0.25µm.
Figure 8. Electron micrographs of CRF labelled profiles at PD 15 & 24. (A) At PD 15, alabelled CRF fiber traversing the soma of a Purkinje cell (arrow heads). At this stage, somaticsynapses have not been encountered. The soma contains a large substantive cisternaecharacteristic for a Purkinje cell. (B) Shows at PD 24, a labelled plexus (arrows) appositionedbetween fibers and the Purkinje cell soma. No synapses were found at the level. Calibrationbars = 1.5µm.

47
the expression of corticotropin
Only lobules II b, III and VI c still exhibited labelling of a weak intensity and an
uneven distribution. However, even in these lobules, there was an overall increase as
well as more labelled profiles that were at the level of the Purkinje cell layer or had
penetrated into the molecular layer. The remaining lobules of the anterior (I, IIa, IV)
and central lobes (V, VI a,b, VII,VIII) all showed dense though still uneven labelling
in both the internal granular layer and molecular layer.
Postnatal days 24-40
At adult stages, we observed the greatest degree of labelling, occurring in the posterior
and inferior lobules, although lobule IX c showed slightly less but still more than
anterior and central lobules (fig 1 F; fig 4 A and B). Even in adulthood, labelling was
not even within these lobules, predominating in the apex and appearing weak in the
folial and lateral regions of the lobules (fig 4 C and D). Also, labelling was more
abundant in the vermal lobes (in comparison to the hemispheric regions), appearing
also in the flocculus and paraflocculus (fig 5 A and B). At the electron microscopic
level, these regions had profiles, which contained either less labelled product or were
completely empty. The labelling was contained in climbing fibers (fig 4 E) and mossy
fiber rosettes (fig 4D and 8) as well as plexus at the level of the Purkinje cell soma
In the anterior lobules, labelling also was inhomogeneous predominating in apical
regions (fig 4 A and B). However, these lobules showed greater areas of weak or no
labelling.
Figure 9. Electron micrographs of CRF labelling in mossy fiber profiles. (A) At PD 12, mossyfibers containing CRF were numerous. These contained numerous spherical vesicles,mitochondria, and made multiple synapses (arrow heads). (B) By PD 24, mossy fiber rosetteshad increased in size. Calibration bars = 0.25µm.

Chapter 2
DISCUSSION
CRF distribution
The present study shows that in the rat cerebellum, the distribution of CRF labelling
is highly dependent upon the age as well as the region under view. The posterior
aspects of the cerebellum (especially lobules IX and X) showed intense labelling in
the two main afferent systems i.e. mossy fibers and climbing fibers from as early as
PD 6 with this pattern persisting into adulthood with a striking increase in intensity
and frequency of labelled profiles at later stages. However, in anterior lobules between
the stages of PD 6-12, labelling appeared weak and predominantly around the Purkinje
cell with a subsequent change later to more intense labelling in the mossy fibers and
climbing fibers. These findings add to those of Palkovits et al. (1987) and van den
Dungen et al. (1988) who failed to elucidate any lobular distribution of CRF. Another
important finding was that not all climbing fibers were CRF-positive with variations
occurring within as well as between lobules. The same was noticeable for the mossy
fiber system.
The posterior lobules IXa & b and X which displayed the most intense labelling and
moreover, which appeared earliest, essentially receive mossy fiber input from the
vestibular system as well as climbing fibers from the dorsal cap of the inferior olive
oogd and Glickstein, 1998). The early and overwhelming presence of CRF in the
vestibulo-cerebellar system, the phylogenetically oldest part of the cerebellum, could
possibly signify a crucial role for this peptide in the maturation and subsequent
functioning of this system, more so than in any other function involving the
cerebellum. In contrast to other lobules, climbing fibers and mossy fiber rosettes
appeared here to be equally CRF-positive making it difficult to discern any dominance
between the two systems. Although heavily labelled, even within these lobules,
labelled profiles predominated within certain areas especially the apex (as opposed
to the folial and lateral regions). This rather discrete localisation of CRF in only
certain regions of the cerebellum is perhaps suggestive of the role of this neuropeptide
being confined to these specific areas.
Other areas of the cerebellum lagged behind in terms of the temporal as well as the
quantitative expression of CRF. In lobules V, VI a & b as well as VIII, the CRF
distribution developed slightly slower than IX and X and never reached their level of
. In the anterior lobules, the labelling first appeared predominantly around
the Purkinje cell soma in the form of punctate profiles. From PD 12 onwards, the
labelled profiles took on the appearance of climbing fibers and mossy fibers, not
dissimilar from those in other lobules, although far less frequent and intense.

49
the expression of corticotropin
Importantly, up to this stage, the internal granular layer appeared almost devoid of
any labelled profiles in these anterior lobules. Somatotopical maps indicate a large
degree of overlap in terms of afferent input to these areas from precerebellar nuclei,
indicating perhaps a more generic role for CRF in these areas. The fact that CRF
occurs predominantly in the vermis is suggestive of it being involved more in
spinocerebellar function such as the control of axial muscles and posture. The fact
that CRF was never evenly distributed throughout the lobule but located either apically
or laterally might be in keeping with the functional compartmentation of the
cerebellum (Voogd and Glickstein, 1998). Since the source of CRF is from the
precerebellar nuclei, the heterogeneous distribution of CRF within the cerebellum
indicated in this study could be a marker for the differential functional development
of these nuclei. It raises the question whether this disparate distribution is suggestive
of this neuropeptide serving different functions in different parts of the cerebellum.
Alternatively, it could merely be an indication of the extent to which these precerebellar
nuclei are represented in the cerebellar cortex.
CRF during maturation
The present data show that CRF is present in the rat cerebellum much earlier than PD
8 as previously reported by van den Dungen et al. (1988). They also reported the
disappearance of the pericellular nest stage of labelling around the Purkinje cells
from PD 16 to 18 onwards, which is in contrast to our study where we found that
depending on the region underview, this type of labelling had disappeared as early as
PD 6 in the posterior lobules and around PD 12 in the anterior lobules. However, our
study is in concordance with their findings that not all climbing fibers contained
CRF and not all CRF positive fibers were equally immunoreactive. Our findings
also tend to contrast somewhat with those of Overbeck and King, (1999) who found
a far more uniform lobular distribution of CRF positive profiles up till PD 10, after
which a differential lobular distribution became more apparent. Our data suggest
that this differential lobular distribution occurs much sooner, as early as PD 6. Since
this study was performed in a mouse, there appears to be a clear species difference in
terms of the ontogeny of CRF-containing profiles.
Bishop and King, (1999) have shown that CRF is expressed in the embryonic murine
cerebellum. Given the overlap in its distribution patterns in the two species, it is
most likely that it is also expressed in the embryonic rodent cerebellum, though
further studies need to corroborate this supposition.
CRF is known to augment the activity of neurotransmitters at the Purkinje cell synapse
(Bishop and King, 1992). However, these studies were restricted to adult material. It

Chapter 2
could therefore be inferred that this neuropeptide has a vastly different role in postnatal
or developing tissue since functional synapses have not yet formed. King and
Overbeck, (1999) have proposed that CRF plays a developmental role in the
cerebellum with a subsequent shift to that of a neuromodulatory function once the
functional circuitry has been established. This hypothesis has been enhanced by the
recent publication of Ha et al. (2000) showing that CRF caused proliferation of
cerebellar astrocytes in vitro. However, the establishment of the cerebellar circuitry
remains subject to a myriad of processes (Strata et al., 1997; Vogel, 1998). We have
shown that CRF from a very early age is not ubiquitous, with some afferents to
certain regions containing the neuropeptide (posterior regions) and others not (anterior
regions). This is in keeping with the findings of Chedotal et al. (1997) that suggest
the anterior and posterior regions of the cerebellum are under separate controlling
developmental factors despite the consequential rather homogenous morphology.
Chedotal and Sotelo, (1993) have also shown that a sub-population of climbing fibers
portray a rather unique developmental profile, synapsing rather precociously at PD 0
on the distal dendrites of Purkinje cells and later retracting to form pericellular nests
at around PD 6. They termed this early stage of climbing fiber synaptogenesis, the
“creeper stage”. These afferents projected exclusively to vermal lobule X. The early
expression of CRF in climbing fibers could be a useful marker to study climbing
fiber synaptogenesis in other regions of the cerebellum.
For CRF to play a pivotal role in the maturation of the climbing fiber and mossy
fiber systems, differences in labelled and non-labelled profiles would be expected.
, ultrastructural analysis failed to elucidate any discernable morphological
ferences in the maturation processes between CRF containing profiles and those
lacking it. Whatever differences were apparent were between afferents containing
CRF in different parts of the cerebellum and we have concentrated on the two extremes
to highlight the contrasting conditions i.e. lobule II and lobule X. From an early
stage (PD 6) there were differences in the labelling picture between CRF containing
climbing fibers in lobules II and X. It raises the question whether the disparity in the
extent of labelling in these two regions is due to differences in the maturity of the
climbing fiber and mossy fiber rosettes in the two regions, differential maturation of
the respective precerebellar nuclei or merely the time-onset on CRF production in
these nuclei. CRF labelling appeared different in lobule II when compared with that
of lobule X. In lobule II, CRF was concentrated primarily in puncta around the soma
of the Purkinje cell without any labelling of axonal processes whereas in lobule X,
label was clearly visible in the axon as well as in terminals synapsing on the Purkinje
cell soma. Ultrastructural analysis clearly showed climbing fibers having made

51
the expression of corticotropin
synapses on the soma of Purkinje cells as well as their proximal dendrites. The
translocation of climbing fiber synapses from the Purkinje cell soma to their proximal
dendrites appeared to occur at disparate rates in the different lobules suggestive of
the maturation of this afferent system occurring at different rates in different parts of
the cerebellum. The temporal differences in the expression of CRF mRNA in the
precerebellar nuclei probably impacts on the development of those axons containing
it. How CRF fits into the chemo-affinity hypothesis espoused by Alcantara et al.
(2000) and Vogel, (1998) remains to be elucidated. Comparative electrophysiological
recordings in different regions could go some way to confirm whether the early
expression of CRF in posterior lobules represents the precocious establishment of
functional neuronal networks or merely an anatomic maturation.
CRF as a neuro-modulator
In vitro studies by Bishop and King, (1992) have shown that CRF augments the
activity of neurotransmitters at the synapses of climbing fibers and mossy fibers on
Purkinje cells. Our study shows that not all climbing fibers contain CRF. This raises
the question of how important CRF and the CRF-receptor system is to the functioning
of climbing fibers (Bloedel and Bracha, 1998) and whether the dynamics of this
ferent system differs between lobules as well as the impact of CRF positive and
negative climbing fiber on motor co-ordination (Ebner, 1998; Welsh et al., 1995).
It would also be prudent to correlate the distribution of CRF with that of its receptor.
, it is known that two CRF receptors are expressed in the cerebellum
(Lovenberg et al. 1995; Chang et al 1993; Potter et al. 1994). Bishop et al. (2000)
have shown that in the adult mouse, receptor type one is present on the somas and
primary dendrites of Purkinje cells of all the lobules of the cerebellar cortex as well
as in radial glia of certain vermal lobules. Receptor type two is also distributed
throughout all lobules being localised to the basal pole of the Purkinje cell, certain
basket and stellate cells as well as granule cells. However, Van Pett et al. (2000)
have shown that there is a degree of disparity in terms of CRF receptor mRNA
expression between the rat and mouse cerebellum. Notwithstanding, this universal
distribution of the receptors could indicate that the reach of CRF in the cerebellum is
far and wide though its impact on cerebellar function is discretely regulated to perhaps
certain time points in certain regions. Importantly, studies of knock-out mice for the
various CRF receptors (Kishimoto et al., 2000; Bale et al., 2000) show that these
animals have various motor deficits.

Chapter 2
CRF is principally involved in the body’s stress response via the HPA axis. The fact
, as shown by this and other studies, is extensively expressed in the cerebellum
from a very early age through to adulthood implies a pivotal role of CRF in cerebellar
processes. However, a robust theory incorporating CRF activity with that of current
thinking on cerebellar function such as the Marr-Albus-Ito theory remains elusive,
resulting in a degree of conjecture regarding the primary role of CRF in this structure.
Miyata et al. (1999), using adult cerebellar slices have shown that CRF contained in
climbing fibers is crucial for the induction of LTD between the parallel fiber-Purkinje
cell synapse. However, a host of other factors have been found to be important for
cerebellar LTD (Daniel et al., 1998; Ichise et al., 2000) and Hansel and Linden,
(2000) have shown that LTD is possible at climbing fiber – Purkinje cell synapses as
well. In both studies, there is no mention of the region of the cerebellum where the
recordings were done raising the question whether these phenomena are equally
inducible in every cerebellar lobule or does learning within the cerebellum take place
via other mechanisms in CRF-poor regions. Kenyon, (1997) has also postulated that
another site of long term memory storage could also be the synapses of parallel
fibers onto stellate/basket cell interneurons. Since we have found CRF labelling in
climbing fibers to exhibit a temporal and spatial heterogeneity, the questions arise as
to how soon LTD can manifest itself and whether LTD is limited to certain regions of
the cerebellum. Also, is LTD the predominant mechanism underlying memory storage
in the cerebellum? It could be suggestive of learning and memory being fairly disparate
processes in different regions of the cerebellum.
Conclusions
This study shows that there is a prominent CRF cerebellar expression throughout the
lifetime of the rat indicative of a pivotal role in the functioning of this structure. The
study also shows that CRF is not evenly distributed throughout the cerebellum but
predominates in the posterior aspects, especially vermal lobules IX and X suggesting
that the role of CRF could be more dominant in these regions. Further studies in
progress to manipulate the expression of this neuropeptide will shed some light on
its function in the cerebellum.
REFERENCES
Alcantara S., Ruiz M., De Castro F., Soriano E., Sotelo C. (2000) Netrin 1 acts as an attractive or as arepulsive cue for distinct migrating neurons during the development of the cerebellar system.Development 127(7):1359-72.

53
the expression of corticotropin
Altman J., Bayer S.A. (Eds.) (1997) Development of the cerebellar system. In relation to its evolution,structure and function. CRC Press.
.L., Contarino A., Smith G.W., Chan R., Gold L.H., Sawchenko P.E., Koob G.F., Vale W.W.,Lee K.F. (2000) Mice deficient for corticotropin-releasing hormone receptor-2 display anxiety-likebehaviour and are hypersensitive to stress. Nat Genet. 24(4),410-4.
Bishop G.A., Seelandt C.M., King J.S. (2000) Cellular localization of corticotropin releasing factorreceptors in the adult mouse cerebellum. Neuroscience. 101(4),1083-92.
Bishop G.A., King J.S. (1999) Corticotropin releasing factor in the embryonic mouse cerebellum. ExpNeurol. 160(2),489-99.
Bishop G.A., King J.S. (1992) Differential modulation of Purkinje cell activity by enkephalin andcorticotropin releasing factor. Neuropeptides. 22(3), 167-74.
Bloedel J.R., Bracha V. (1998) Current concepts of climbing fiber function. Anat Rec. 253(4), 118-26.
Chang C.P., Pearse R.V., O’Connell S, Rosenfeld M.G. (1993) Identification of a seven transmembranehelix receptor for corticotropin-releasing factor and sauvagine in mammalian brain. Neuron. 11(6),187-95.
Chedotal A., Bloch-Gallego E., Sotelo C. (1997) The embryonic cerebellum contains topographiccues that guide developing inferior olivary axons. Development. 124(4), 861-70.
Chedotal A., Sotelo C. (1993) The ‘creeper stage’ in cerebellar climbing fiber synaptogenesis precedesthe ‘pericellular nest’—ultrastructural evidence with parvalbumin immunocytochemistry. Brain ResDev Brain Res; 76(2): 207-20.
Chen C., Tonegawa S.. (1997) Molecular genetic analysis of synaptic plasticity, activity-dependentneural development, learning, and memory in the mammalian brain. Annu Rev Neurosci; 20:157-84
Cummings S.L., Young W.S III., King J.S. (1994) Early development of cerebellar afferent systemsthat contain corticotropin-releasing factor. J Comp Neurol; 350(4): 534-49.
Daniel H., Levenes C., Crepel F. (1998) Cellular mechanisms of cerebellar LTD. Trends Neurosci.21(9), 401-7.
De Souza E.B. (1995) Corticotropin-releasing factor receptors: physiology, pharmacology, biochemistryand role in central nervous system and immune disorders. Psychoneuroendocrinology. 20(8), 789-
Ebner T.J. (1998) A role for the cerebellum in the control of limb movement velocity. Curr OpinNeurobiol. 8(6),762-9.
Errico P., Barmack N.H. (1993) Origins of cerebellar mossy and climbing fibers immunoreactive forcorticotropin-releasing factor in the rabbit. J Comp Neurol. 336(2), 307-20.
Ha B.K., Bishop G.A., King J.S., Burry R.W. (2000) Corticotropin releasing factor induces proliferationof cerebellar astrocytes. J Neurosci Res. 62(6), 789-98.
Hansel C., Linden D.J. (2000) Long-term depression of the cerebellar climbing fiber—Purkinje neuronsynapse. Neuron. 26(2),473-82.
Ichise T., Kano M., Hashimoto K., Yanagihara D., Nakao K., Shigemoto R., Katsuki M., Aiba A.(2000) mGluR1 in cerebellar Purkinje cells essential for long-term depression, synapse elimination,and motor coordination. Science. 288(5472), 1832-5.
Ito M. (1989) Long-term depression. Annu Rev Neurosci; 12:85-102.
Kenyon G.T. (1997) A model of long-term memory storage in the cerebellar cortex: a possible role forplasticity at parallel fiber synapses onto stellate/basket interneurons. Proc Natl Acad Sci U S A. 94(25),14200-5.
King J.S., Madtes P., Bishop G.A., Overbeck T.L.(1997) The distribution of corticotropin-releasingfactor (CRF), CRF binding sites and CRF1 receptor mRNA in the mouse cerebellum. Prog Brain
14; 55-66.

Chapter 2
Kishimoto T., Radulovic J., Radulovic M., Lin C.R., Schrick C., Hooshmand F., Hermanson O.,Rosenfeld M.G., Spiess J. (2000) Deletion of crhr2 reveals an anxiolytic role for corticotropin-releasinghormone receptor-2. Nat Genet. 24(4),415-9.
Lovenberg T.W., Liaw C.W., Grigoriadis D.E., Clevenger W., Chalmers D.T., De Souza E.B., Oltersdorf(1995) Cloning and characterization of a functionally distinct corticotropin-releasing factor receptor
subtype from rat brain. Proc Natl Acad Sci. 92(3), 836-40.
Miyata M., Okada D., Hashimoto K., Kano M., Ito M. (1999) Corticotropin-releasing factor plays apermissive role in cerebellar long-term depression. Neuron. 22(4),763-75.
Muller R.A., Courchesne E., Allen G. (1998) The cerebellum: so much more. Science. 282, 879-80.
Oberdick J., Baader SL., Schilling K. (1998) From zebra stripes to postal zones: deciphering patternsof gene expression in the cerebellum. Trends Neurosci; 21(9): 383-90.
Overbeck T.L., King J.S. (1999) Developmental expression of corticotropin releasing factor in thepostnatal murine cerebellum. Brain Res Dev Brain Res. 115(2),145-59.
Palkovits M., Leranth C., Gorcs T., Young W.S. (1987) Corticotropin-releasing factor in theolivocerebellar tract of rats: demonstration by light- and electron-microscopic immunohistochemistryand in situ hybridization histochemistry. Proc Natl Acad Sci U S A. 84(11),3911-5.
Potter E., Sutton S., Donaldson C., Chen R., Perrin M., Lewis K., Sawchenko P.E., Vale W. (1994)Distribution of corticotropin-releasing factor receptor mRNA expression in the rat brain and pituitary.Proc Natl Acad Sci. 91(19), 8777-81.
Sotelo C. (1999) From Cajal’s chemotaxis to the molecular biology of axon guidance. Brain Res Bull.50(5-6), 395-6.
Sotelo C., Chedotal A. (1997) Development of the olivocerebellar projection. Perspect DevNeurobiol.5(1), 57-67.
Strata P., Rossi F. (1998) Plasticity of the olivocerebellar pathway. Trends Neurosci. 21(9), 407-13.
Strata P., Tempia F., Zagrebelsky M., Rossi F. (1997) Reciprocal trophic interactions between climbingfibres and Purkinje cells in the rat cerebellum. Prog Brain Res. 114, 263-82.
., Spiess J., Rivier C., Rivier J. (1981) Characterization of a 41-residue ovine hypothalamicpeptide that stimulates secretion of corticotropin and beta-endorphin. Science; 213,1394-7.
van den Dungen H.M., Groenewegen H.J., Tilders F.J., Schoemaker J. (1988) Immunoreactivecorticotropin releasing factor in adult and developing rat cerebellum: its presence in climbing andmossy fibres. J Chem Neuroanat. 1(6),339-49.
van den Pol AN. (1985) Silver-intensified gold and peroxidase as dual ultrastructural immunolabelsfor pre- and postsynaptic neurotransmitters. Science; 228(4697):332-5.
an Pett K., Viau V., Bittencourt J.C., Chan R.K., Li H.Y., Arias C., Prins G.S., Perrin M., Vale W.,Sawchenko P.E. (2000) Distribution of mRNAs encoding CRF receptors in brain and pituitary of ratand mouse. J Comp Neurol. 428(2), 191-212.
ogel M.W. (1998) Images in neuroscience. Brain development, II. Establishing synaptic connections.Am J Psychiatry. 155(5),581.
oogd J., Glickstein M. (1998) The anatomy of the cerebellum. Trends Neurosci. 21(9), 370-5.
elsh J.P., Lang E.J., Sugihara I., Llinas R. (1995) Dynamic organization of motor control within theolivocerebellar system. Nature. 374, 453-7.
ickelgren I. (1998) The cerebellum: the brain’s engine of agility. Science. 281,1588-90.
amano M., Tohyama M. (1994) Distribution of corticotropin-releasing factor and calcitonin gene-
related peptide in the developing mouse cerebellum. Neurosci Res. 19(4),387-96.

55
CHAPTER 3
The localisation of urocortin in the adult rat cerebellum:A light and electron microscopic study
J. D. Swinny1, D. Kalicharan1, A. Gramsbergen2 , J.J.L. van der Want1
1 Laboratory for Cell Biology and Electron Microscopy, and2 Department of Medical Physiology, Graduate School of Behavioural and
Cognitive Neuroscience, University of Groningen, The Netherlands
Neuroscience (2002); 114(4):891-903.

56
Chapter 3
ABSTRACT
Light and electron microscopic immunocytochemistry was used to identify the cellular
and subcellular localisation of urocortin in the adult rat cerebellum. Urocortin
immunoreactivity (UCN-ir) was visualised throughout the cerebellum, yet
predominated in the posterior vermal lobules, especially lobules IX and X, the
flocculus, paraflocculus and deep cerebellar nuclei. Cortical immunoreactivity was
most evident in the Purkinje cell layer and molecular layer. Reaction product, though
sparse, was found in the somata of Purkinje cells, primarily in the region of the Golgi
apparatus. Purkinje cell dendritic UCN-ir was compartmentalised, with it being
prevalent in proximal regions especially where climbing fibres synapsed, yet absent
in distal regions where parallel fibres synapsed. In the Purkinje cell layer, the labelling
was also contained in axonal terminals, synapsing directly on Purkinje cell somata.
These were identified as axon terminals of basket cells based on their morphology.
Terminals of stellate cells in the upper molecular layer also expressed the peptide.
Whilst somata of inferior olivary neurons showed intense immunoreactivity, axonal
labelling was indistinct, with only the terminals of climbing fibres containing reaction
product. UCN-ir in the mossy fibre-parallel fibre system was restricted to mossy
fibre rosettes of mainly posterior lobules and the varicose terminals of parallel fibres.
Furthermore, labelling also was prevalent in glial perikarya and their sheaths. The
current study shows, firstly, that urocortin enjoys a close ligand-receptor symmetry
in the cerebellum, probably to a greater degree than CRF since CRF itself is found
exclusively in the two major cerebellar afferent systems. Its congregation in excitatory
and inhibitory axonal terminals suggests a significant degree of participation in the
synaptic milieu, perhaps in the capacity as a neurotransmitter or effecting the release
of co-localised neurotransmitters. Finally, its unique distribution in the Purkinje cell
dendrite might serve as an anatomical marker of discrete populations of dendritic
spines.

57
localisation of urocortin
INTRODUCTION
Corticotropin releasing factor (CRF) is principally involved in the regulation of the
body’s endocrine, autonomic and behavioural response to stress (Koob and Heinrichs,
1999). It plays an important role in the cerebellum, based on its presence and
distribution in the embryo (Bishop and King, 1999) and in the adult (Palkovits et al.,
1987). CRF-containing projections are particularly prominent in the posterior lobules,
especially lobules VIII to X (Overbeck and King, 1999). At a physiological level, Ha
et al., (2000) have shown that CRF causes proliferation of cerebellar astrocytes in
vitro. Also, Bishop (1990) has shown that CRF augments the activity of excitatory
neurotransmitters at the climbing fibre-Purkinje cell synapse and Miyata et al. (1999)
have shown that CRF contained in climbing fibres is crucial for the induction of long
term depression (LTD), a form of synaptic plasticity proposed as the cellular basis of
learning in the cerebellum (Ito. 1984). Taken together, these data suggest that CRF
has in the cerebellum, a dual role first in development and later in synaptic plasticity.
The widespread distribution of both CRF receptor 1 (CRF-R1) and CRF receptor 2
(CRF-R2) in the cerebellum (Bishop et al., 2000), coupled with the disparate lobular
CRF localisation suggests that other members of the CRF-like peptides could be
involved in cerebellar functions. Urocortin is a relatively recent addition to the growing
family of CRF-like neuropeptides shown to mediate their effects via both CRF
receptors. Urocortin appears to couple preferentially with CRF-R2 leading to
speculation that it is the natural ligand of the type two receptor (Vaughan et al.,
1995). The localisation of urocortin suggests a role in motor control and sensorimotor
integration. The urocortin distribution in the brain is rather restricted, in contrast to
CRF, with expression occurring predominantly in the Edinger-Westphal nucleus and
the lateral superior olive. Minor sites of expression include the inferior olive, pontine
grey, tegmental reticular nucleus, lateral reticular nucleus, and in the red nucleus and
nucleus prepositus hypoglossi (for a comprehensive evaluation in rat brain, see
Bittencourt et al., 1999). At a physiological level, centrally administered urocortin
inhibits food intake to a greater extent than CRF but induces anxiety-like behaviour
and locomotor activity to a lesser degree (Spina et al., 1996).
There are conflicting data regarding the precise localisation of urocortin in the
cerebellum. Bittencourt et al. (1999) reported weak immunoreactive projections to
the molecular, Purkinje cell and granular cell layers of the flocculus, paraflocculus
and lingula as well to the deep nuclei of rat cerebellum. Morin et al. (1999), in
comparing the distribution of CRF and urocortin in the rat brain made mention only

58
Chapter 3
of CRF “cells” in the cerebellum. Kozicz et al. (1998) also failed to report urocortin
cerebellar immunoreactivity in a study of the rat. Using radioimmunoassay, Takahashi
et al. (1998), showed a prominent quantity of this peptide in the human cerebellum.
Species and methodological differences could account for the incongruencies in the
above studies. With a view to elucidating the possible role of urocortin in cerebellar
function we provide detailed information on its laminar, lobular and cellular
distribution.
Experimental procedures
Immunocytochemistry
A total of eight black-hooded Lister rats were studied at postnatal day (PD) 40. The
day of birth was considered PD 0. Approval to conduct the study was obtained from
the Ethics Committee on Animal Experimentation, University of Groningen. All efforts
were made to minimise the number of animals used and their suffering. Animals
were deeply anaesthetised with sodium pentobarbital (NembutalR, i.p., 50mg/kg)
and perfused transcardially first with a solution containing 2% polyvinyl pyrrolidone
(molecular weight 30 000), 0.4% NaNO3 in 0.1 M phosphate buffer (PB) (pH 7.4) for
one minute, and then with 4% paraformaldehyde and 0.2% picric acid in 0.1M PB
(pH 7.4) for ten minutes. The brains were removed and stored overnight in the fixative
solution at 4o C.
For light microscopy, brains were stored overnight at 4oC in 30% sucrose for
cryoprotection and cryo-sectioned (30µm thick) in either the sagittal or transverse
plane. Three animals were evaluated in either plane. Immunoreactivity was visualised
by the avidin-biotin-peroxidase complex (ABC) method. Briefly, free-floating sections
were immersed for two hours in a pre-incubation medium containing 1% normal
rabbit serum and 1% bovine serum albumin in 0.1M phosphate buffered saline (PBS)
(pH 7.4). The sections were then incubated overnight at 4o C with affinity purified
goat anti-urocortin IgG (Santa Cruz Biotechnologies, Santa Cruz, CA), diluted 1:100
with PBS. After washing with PBS, the sections were incubated with biotinylated
rabbit anti-goat IgG, diluted 1:200 in PBS, for two hours at room temperature. After
further washing, the sections were incubated with ABC (Vectastain Elite, Vector
Labs, Burlingame, CA) for one hour at room temperature. Immunoreactivity was
visualised by incubation with 5mg 3,3 diaminobenzidine HCl (DAB) and 0.03%
hydrogen peroxide in 10 ml of 0.1 M PBS (pH 7.4) for 5 to 10 minutes at room
temperature. The reaction was stopped by washing the sections in cold PBS. In some
sections, immunostaining was further enhanced according to the gold substituted

59
localisation of urocortin
silver peroxidase (GSSP) method (van den Pol and Gorcs, 1985).
Controls
Sections were processed as above with the omission of the primary antibody or with
the addition of a blocking peptide (Santa Cruz Biotechnologies, Santa Cruz, CA),
diluted 1:100 with PBS.
The sections were mounted on gelatine-coated slides. Some of the sections were
counter stained with cresyl violet for 2 minutes, dehydrated and coverslipped with
DepexR mounting medium.
Evaluation of sections
Images of sections were projected onto paper using a projection microscope and
these were plotted by hand. From these plots, a composite sketch was drawn
summarising the labelling pattern. Selected sections were photographed on an
Olympus Digital camera, mounted on an Olympus B50 optical microscope. These
images were adjusted to enhance the contrast and brightness and saved at 300 dpi.
Transmission electron microscopy
Single immunolabelling:
From the cerebella of two animals, 50 µm thick sections were cut on a Vibratome in
the sagittal and transverse planes and processed as for light microscopy except, that
Triton X-100, 0.025% was added to the primary antibody incubation.
Immunoreactivity was enhanced according to the gold substituted silver peroxidase
method (van den Pol and Gorcs, 1985). Sections were then osmicated in 1% OsO4,
1.5% potassium hexacyanoferrate in 0.1M cacodylate buffer (pH 7.6) for 15 minutes,
dehydrated in a graded series of ethanols and embedded in Epon. Semithin sections
(1 µm) were cut on an LKB Ultratome, stained with toluidine blue and used for
orientation purposes. Ultrathin sections were then cut, counterstained with uranyl
acetate and lead citrate, and examined with a Philips CM 100 transmission electron
microscope.
Double immunolabelling:
To prove conclusively that climbing fibre terminals contained urocortin, double
immunolabelling with urocortin and CRF (a positive marker for climbing fibres)
was performed, according to a modified protocol of Yi et al., (2001). Briefly, sections
were placed in phosphate buffer (PB) containing 0.1% sodium borohydride to
inactivate residual aldehyde groups in the tissue sections. To improve reagent
penetration, the sections were then treated with PB containing 0.05% Triton X-100.

60
Chapter 3
To prevent non-specific binding of the immunoreagents, sections were incubated in
blocking solution, which was PBS, pH 7.4, containing 5% normal goat serum (NGS),
5% BSA, and 0.1% CWFS gelatin. After blocking, sections were incubated in a
mixture of rabbit anti-CRF (1:1000) and goat anti-urocortin (1:100) primary antibodies
diluted with PBS containing 0.2% acetylated bovine serum albumin (BSA-c), (Aurion,
Wageningen, The Netherlands) (PBS/BSA-c, pH 7.4). After washes with PBS/BSA-
c, sections were incubated overnight in the first secondary antibody, a conjugate
which was ultrasmall gold-conjugated F(ab’)2 (0.8nm) fragments of rabbit anti-goat
IgG diluted 1:100 with PBS/BSA-c. After washes, silver enhancement was performed
according the manufacturer’s instructions. After the first silver enhancement, sections
were washed with PB and then incubated with the second secondary antibody, namely
biotinylated goat anti-rabbit IgG, diluted 1:200 in PBS. Immunoreactivity was
enhanced according to the GSSP procedure (van den Pol and Gorcs, 1985) and the
sections processed further for transmission electron microscopy, in the same manner
as for single immunolabelling.
Antiserum
The antiserum used in this study was raised against a peptide mapping at the carboxy
terminus of urocortin of rat origin (identical to corresponding mouse and human
sequences). It reacts with urocortin of mouse, rat and human and does not cross-
react with corticotropin-releasing factor.
RESULTS
Regional distribution
Note that due to variations in the intensity of lobular labelling, all micrographs are of
regions from either lobule IX or X, unless stated to the contrary.
Urocortin immunoreactivity (UCN-ir) could be visualised in all vermal lobules, the
hemispheres as well as the flocculus and paraflocculus of the cerebellar cortex.
Labelling appeared to be constant medio laterally, from vermis to hemispheres.
However, a distinct staining gradient was noted in the sagittal plane (Fig. 1). The
posterior lobules (especially lobules IX and X), the flocculus, and the paraflocculus
exhibited the most abundant labelling, closely followed by lobules VII, VIII and
then lobule VI. The anterior lobules appeared to contain appreciably less
immunoreactivity (Fig. 2). UCN-ir was also present in the white matter, deep cerebellar
nuclei (fastigial, interposed and dentate), and the lateral vestibular nuclei (Fig. 3).

61
localisation of urocortin
Figure 1. Schematic representation of UCN-ir in the mid-sagittal and transverse planes ofthe cerebellum, concentrating on the labelling in Purkinje cells. (A) shows the vermal lobularimmunoreactivity pattern in the sagittal plane with the labelling predominating in posteriorlobules, especially lobules IX and X. Labelling is mainly in the Purkinje cell dendrites (arrow)as well as some somata (arrowhead). (B) shows the labelling pattern of the anterior lobulesin the transverse plane with the UCN-ir being somewhat weaker. However intenseimmunoreactivity could be visualised in the deep cerebellar nuclei as well as the lateralvestibular nucleus. (C) shows the labelling pattern of the posterior lobules in the transverseplane with UCN-ir being particularly prominent in the Purkinje cell dendrites in the vermis. CI & II, crus one and two; DN, dentate nucleus; FL, flocculus; PFd, dorsal region of theparaflocculus; PFv, ventral region of the paraflocculus; FN, fastigial nucleus; IN, interposednucleus; LS, lobulus simplex; Vn, lateral vestibular nucleus.

62
Chapter 3
Figure 2. Photomicrographs of UCN-ir showing differences in intensity between anterior andposterior lobules. (A) in lobule X, the labelling pattern is intense and concentrated in thePurkinje cell dendrites (arrowheads), with weak localisation in the regions of their somata.(B) an overview of lobule II indicating immunoreactivity of relatively weak intensity, and (C),a higher magnification of the framed area showing the labelling being concentrated aroundthe Purkinje cell somata and puncta in the molecular layer (arrowheads). Note that to observethe relatively weak immunoreactivity in lobule II, the DAB reaction product had to be enhancedaccording to the GSSP method.

63
localisation of urocortin
Cellular localisation
Labelled profiles were evident in all layers of the cerebellar cortex, predominantly in
the molecular and Purkinje cell layers. Some Purkinje cells contained sparse reaction
product in their perikarya, associated with small vesicles in the region of the Golgi
apparatus. Axonal labelling was mainly at terminals synapsing on the deep cerebellar
nuclei. Dendritic labelling appeared in the main shafts as well as in the spines of the
proximal dendritic domain synapsing with climbing fibres. While the somatic and
axonal reaction product was conspicuously associated with spherical vesicles, this
was not the case in dendrites, where labelling often occurred in large clumps that are
most likely related to tubular elements of endoplasmic reticulum (Fig. 4).
In the Purkinje cell layer, varicosities synapsing directly on somata of Purkinje cells
showed UCN-ir. These were characteristic of basket cell terminals. In the upper
molecular layer, terminals of stellate cells synapsing directly on Purkinje cell dendritic
shafts also contained UCN-ir (Fig. 5).
UCN-ir was evident in the mossy fibre-parallel fibre system. Mossy fibre rosettes,
especially those in the posterior lobules, exhibited reaction product. In the molecular
Figure 3. Photomicrographs of UCN-ir in the deep cerebellar nuclei and lateral vestibularnucleus. (A) an overview of the extent and intensity of labelling in the transverse plane. (B)an overview of the labelling of the fastigial nucleus in the sagittal plane. (C) a highermagnification of a selected area showing the labelling around individual neurons of the dentatenucleus. (D) is a higher magnification of (B) showing labelled puncta that represent terminalvaricosities of Purkinje cell axons. IV, fourth ventricle.

64
Chapter 3
Figure 4. Micrographs showing UCN-ir in the Purkinje cell. (A) a photomicrograph showingUCN-ir in the proximity of Purkinje cell somata and proximal dendrites. Arrowheads point tolabelled puncta which could be either Purkinje cell spines, parallel fibre varicosities or climbingfibre varicosities. (B) moderate UCN-ir in a Purkinje cell soma is associated with Golgi-likevesicles (arrowheads). In (C), a labelled Purkinje cell dendritic spine (arrowhead) is shownmaking an asymmetric synapse with an unlabelled terminal varicosity. In the vicinity, threeunlabelled dendritic spines (asterisks) can be observed. G, Golgi apparatus; Pc, Purkinje cell

65
localisation of urocortin
Figure 5. Basket and stellate cell UCN-ir. (A) shows a basket cell with prominent somaticlabelling (arrowheads), located in the vicinity of the Golgi apparatus. (B) a detail of a Purkinjecell soma showing labelling in a profile characteristic of a basket cell terminal. (C) Showsreaction product in the upper molecular layer. In the centre, is a stellate cell terminal containingreaction product (arrow heads), making direct synaptic contact on the shaft of a Purkinje celldendrite. Reaction product is also contained within the glial sheath, the dendritic shaft itselfand within parallel fibres in the vicinity. Note that this micrograph is of material sectioned inthe transverse plane. Hence parallel fibres are viewed in the longitudinal plane. Bc, basketcell; Bt, basket cell terminal; Gs, glial sheath; Pd, Purkinje cell dendrite; Pf, parallel fibres;
Ps, Purkinje cell soma; St, stellate cell terminal.

66
Chapter 3
layer, labelling was evident in the axons of granule cells, that is, parallel fibres.
Whilst some of this reaction product was observed in the shafts, most of it was in the
synaptic varicosities of the parallel fibres. The palisades of Bergmann’s glia were
also labelled (Fig. 6).
At the light microscopic level, no labelled olivo-cerebellar axonal profiles were
observed in either the white matter or cortical layers of the cerebellum. Although, in
the inferior olive, dense cytoplasmic labelling was evident. However, at the electron
microscopic level, profiles in close association with Purkinje cell dendrites and
resembling climbing fibre axonal terminals in the molecular layer contained reaction
product. The results of the double immunolabelling with antibodies against CRF and
urocortin proved these to be climbing fibres since in the molecular layer, CRF is
found exclusively in climbing fibres. CRF immunoreactivity is exhibited in the form
Figure 6. UCN-ir in the mossyfibre-parallel fibre system andglial elements. (A) a survey of alabelled mossy fibre rosetteshowing clustered reactionproduct (arrowheads). (B) anoverview of parallel fibre labellingin the shafts (arrows) and smallervesicular labelling in pre-synapticvaricosities and terminals(arrowheads). The dendriticspines which make synapticcontact with urocortin positiveparallel fibres all are free ofreaction product. However, anumber of other Purkinje celldendritic profiles exhibit denselabelling (open arrowheads).Note also that the micrograph isof a section, cut in the transverseplane. Therefore, parallel fibresare viewed in the longitudinalplane. (C) shows a region, cut inthe sagittal plane, of the uppermolecular layer. Reaction product(arrowheads) is present inparallel fibre varicosities makingsynaptic contact on Purkinje celldendritic spines. Note that,similar to (B), spines postsynapticto parallel fibre terminals are freeof reaction product. (D) anoverview of a basket cell with theinsert being a magnified regionof the boxed area, showing alabelled glial sheath as it bordersthe perikaryon. The labelling is inthe form of large clumps thatalmost seem continuous.

67
localisation of urocortin
Figure 7. UCN-ir in olivo-cerebellarprojection neurons. (A) a photo-micrograph of the principle anddorsal accessory olive showingprominent somatic labelling. (B) anelectron micrograph of urocortinsingle immunolabelling, according tothe GSSP method, showing UCN-irin a varicose terminal, makingsynaptic contacts (arrowheads) ontwo spines of a proximal Purkinjecell dendrite. The terminal shows allthe morphological characteristics ofa climbing fibre. (C) shows urocortinand CRF double immunolabellingwithin a climbing fibre terminal.UCN-ir is exhibited in the form ofuniform, enhanced ultrasmall(0.8nm) immunogold particles(arrowheads), whereas CRFimmunoreactivity appears asclumpy, irregular silver deposits,characteristic of the GSSP method(arrows). Whilst CRF immuno-reactivity was confined to climbingfibres UCN-ir was evident in climbingfibres and Purkinje cell dendriticspines. Dao, dorsal accessory olive;Po, principal olive.
of clumpy, peroxidase reaction product and UCN-ir is represented by uniform,
ultrasmall immunogold particles (Fig. 7).
DISCUSSION
The present study is the first to comprehensively detail the distribution of urocortin
in the adult rat cerebellum. Using light and electron microscopic immuno-
cytochemistry, we were able to pinpoint the localisation of this neuropeptide both at
cellular and sub-cellular levels. The present study shows that urocortin is extensively
expressed throughout the adult rat cerebellum, with highest amounts occurring in
vermal lobules IX and X, the flocculus, and the paraflocculus. This is in contrast to
a previous study (Bittencourt et al., 1999) that showed only labelled fibres in the
flocculus, paraflocculus and lingula. The main reason for this discrepancy could be
the different antisera used. Another reason is that in other regions of the cerebellum
where UCN-ir was somewhat weaker, most of the labelling tended to occur in axon
terminals, that are more readily visualised at the electron microscopic level. Our
study is, however, consonant with the previous finding of prominent labelling in the

68
Chapter 3
cerebellar nuclei and vestibular nuclei (Bittencourt et al., 1999). Additional sites
found to be immunoreactive were the synaptic terminals of the inhibitory basket and
stellate cells and the Bergmann’s fibres.
Antiserum specificity
One of the most crucial elements of immunohistochemical studies is the specificity
of the antiserum that ensures a cogent relationship between the reaction product and
the antigen under examination. Despite the relatively pervasive labelling (in
comparison to CRF) encountered with the antiserum used in the study, we are
convinced of its specificity in reliably binding to urocortin for the following reasons.
Firstly, labelling was not evenly distributed across the cerebellum but consistently
exhibited a distinct gradient in the posterior to anterior plane. Also, labelling tended
to be concentrated in specific axonal terminals, namely those of basket, stellate,
granule and Purkinje cells. At the electron microscopic level, the UCN-ir was present
within these terminals without any aspecific labelling on other profiles. Finally, when
somatic labelling occurred, this was never nuclear, but always cytoplasmic, and
occurred in close proximity to the Golgi apparatus.
Urocortin and CRF receptor localisation
Although purely an immunocytochemical approach, this study suggests an intimate
interaction between urocortin and the CRF receptors, obviating, at least in the
cerebellum, the constant issue of ligand-receptors mismatches that has beset
investigators since the initial CRF studies (Chalmers et al., 1995, Potter et al., 1994).
Urocortin is known to mediate its effects by coupling to both known CRF receptors
(Vaughan et al., 1995). In the mouse, Bishop et al. (2000) showed that both receptors
are localised on Purkinje cells (their somata, dendrites and initial axonal segments)
with CRF-R1 alone being further localised in the radial glia as well as in puncta in
the granular layer. CRF-R2 is also localised in basket and Golgi cells and the initial
axonal segments of Purkinje cells. Some of the most prominent labelling found in
this study occurred in the form of axon terminals synapsing on Purkinje cell somata.
These are unlikely to be climbing fibre terminals since climbing fibres do not make
synaptic contacts on Purkinje cell somata at such late adult stages. Based on their
morphology, they are most likely to be the inhibitory terminals of basket cells since
these neurons do indeed form axo-somatic synapses with Purkinje cells in adulthood
(Palay and Chan-Palay, 1974). Also, the basket cell somata were found to exhibit
prominent somatic labelling. However, the possibility of these being the recurrent
collaterals of Purkinje cell axons cannot be entirely excluded due to the close structural
resemblance of these terminals and those of basket cell axon terminals (Altman and

69
localisation of urocortin
Bayer, 1997). Importantly, however, urocortin-containing varicosities synapsing on
the somata of Purkinje cells are to be expected since Bishop et al. (2000) has shown
that, at least in the mouse, CRF-R2 is located in this region. Stellate cells also exhibited
labelling at their synapses on Purkinje dendrites, though to a lesser degree. Another
prominent site of labelling was the synaptic varicosities of parallel fibres which
form excitatory synapses on Purkinje cell spines. It is rather perplexing why a single
neuropeptide would be localised at both excitatory and inhibitory synapses. However,
since CRF receptors are localised to postsynaptic sites (Purkinje cell somata in the
case of basket cell axons and Purkinje dendrites in the case of parallel fibres), this
receptor system could play an integral part of neuromodulation in the cerebellum.
The fact that the reaction product at these synapses was localised in vesicles in
appreciable amounts suggests some kind of neurotransmitter capacity (Schwartz et
al., 2000).
Another major site of UCN-ir was the Purkinje cell itself. The perikarya were weakly
labelled in comparison to axons and dendrites. This compartment probably serves
merely as the site of synthesis and packaging of the peptide, which may be rapidly
transported to the neurites. Labelling in the dendrites was localised in the main shafts
and in some spines. Spines which formed synapses with terminals containing reaction
product, i.e. parallel fibre-dendritic synapses, did not contain any labelling. However,
spines forming synapses with climbing fibre did indeed contain labelling. The
labelling pattern observed in this study suggests a compartmentation of urocortin in
the Purkinje cell dendrite. Even at the light microscopic level (Fig. 4 A), UCN-ir
never reached the distal domain of the dendritic tree. Rather, it predominated in the
proximal domain known to be contacted by climbing fibres. This is in keeping with
current evidence that individual Purkinje cell dendrites are compartmentalised into
two different synaptic domains due to their contiguous innervation by climbing
fibres and parallel fibres (Morando et al., 2001; Bravin et al., 1999; Strata et al.,
1997). The fact that in axonal elements, UCN-ir is associated with vesicles, whereas
dendritic and glial reaction product is not, enhances the supposition that the peptide
may have different functions. Studies investigating urocortin distribution in Purkinje
cell dendrites after afferent perturbation such as climbing fibre lesioning by 3-acetyl
pyridine (Rossi et al., 1991a, b; Desclin and Colin, 1980) or in hypogranular mice
mutant (Crepel and Mariani, 1976; Mariani et al., 1977; Crepel et al., 1980) will
shed more light on the way its targeting to the dendritic environment is controlled.
A point of contention was the labelling of climbing fibres. Bittencourt et al. (1999)
reported reaction product only in projections to the inferior olive and not the cell

70
Chapter 3
bodies themselves. However, the present study shows substantial amounts of reaction
product within cell bodies of inferior olivary neurons. Furthermore, no
immunoreactivity was readily visible in their axonal shafts at the light microscopic
level in any part of the cerebellum. At the electron microscopic level, however, axonal
terminals in the molecular layer fitting the profile of climbing fibres did express
reaction product. Moreover, double labelling experiments which co-localised CRF
and urocortin proved conclusively that urocortin is contained in the terminals and
not in the axons of climbing fibres. Hence, it appears that synthesised urocortin in
inferior olivary neurons is rapidly transported to the axonal terminals, precluding the
localisation of significant amounts in the fibre tracts. If this is the case, then it implies
that the inferior olive processes urocortin differently to CRF, since CRF is clearly
localised in fibre tracts (Palkovits et al., 1987). The functional significance of this
disparate handling of two different peptides which react essentially on the same
receptors system remains to be elucidated.
Urocortin in cerebellar function
The processes involved in the development of the cerebellar circuitry eventually
manifest in a structure pivotal for the fine-tuning of motor coordination (see Welsh
et al., 1995; Wickelgren, 1998) and perhaps higher cognitive functions (Muller et al.,
1998). An understanding of these events could provide a convenient paradigm for
neuronal learning and brain behaviour relationships. The emerging picture of how
urocortin, CRF and their receptors are distributed in the cerebellum encourages one
to attempt to delineate the roles played by these two peptides in cerebellar function.
Urocortin is localised in the targets of both the mossy fibre and the climbing fibre
systems, granule cell axons, Purkinje cells, inhibitory interneurons, and the deep
cerebellar nuclei. More specifically, the presynaptic location of urocortin in excitatory
and inhibitory axonal terminals and its post-synaptic location in the shafts and spines
of Purkinje cell dendrites, suggests an integrative role in the neuronal circuit dynamics
of the cerebellar cortex.
The observations that vestibulocerebellar folia, i.e. lobules IX and X, the flocculus,
and paraflocculus, appeared to be more heavily labelled than the rest of the cortex,
suggests that urocortin might be more intimately involved in the vestibular aspects
of cerebellar function. Its prominence in this region of the cerebellum is consonant
with that of CRF, suggesting that in the adult cerebellum, CRF-like peptides mainly
function in vestibular control. Also, one of the most prominent sites of urocortin
localisation in the brain is the Edinger-Westphal nucleus. In the cat, this site provides
projections to the anterior and posterior cerebellar cortices and the fastigial and

71
localisation of urocortin
interposed nuclei, with the flocculus being the main terminal field (Roste and
Dietrichs, 1988). However, both CRF receptors are, by and large, evenly distributed
throughout the mouse cerebellum (Bishop et al., 2000), intimating that other, yet
undiscovered CRF-like peptides might be responsible for the activation of CRF
receptors resident in these regions.
In conclusion, the present study shows that urocortin has a specific and prominent
expression in the cerebellum, adding to the growing body of evidence that this peptide
is involved in motor control. The data shown here on the urocortin distribution may
also serve as a guide in prospective physiological studies.
REFERENCES
Bishop, G.A., Seelandt, C.M., King, J.S., 2000. Cellular localization of corticotropin releasing factor receptors in
the adult mouse cerebellum. Neuroscience. 101(4), 1083-92.
Bishop, G.A., King J.S., 1999. Corticotropin releasing factor in the embryonic mouse cerebellum. Exp Neurol.
160(2),489-99.
Bishop, G.A., 1990. Neuromodulatory effects of corticotropin releasing factor on cerebellar Purkinje cells: an in
vivo study in the cat. Neuroscience. 39(1), 251-7.
Bittencourt, J.C., Vaughan, J., Arias, C., Rissman, R.A., Vale, W.W., Sawchenko, P.E., 1999. Urocortin expression
in rat brain: evidence against a pervasive relationship of urocortin-containing projections with targets bearing type
2 CRF receptors. J Comp Neurol. 415(3), 285-312.
Bravin, M., Morando, L., Vercelli, A., Rossi, F., Strata, P., 1999. Control of spine formation by electrical activity
in the adult rat cerebellum. Proc Natl Acad Sci U S A. 96(4), 1704-9.
Chalmers, D.T., Lovenberg, T.W., De Souza, E.B., 1995. Localisation of novel corticotropin-releasing factor
receptor (CRF2) mRNA expression to specific sub-cortical nuclei in rat brain: comparison with CRF1 receptor
mRNA expression. J Neurosci.15,6340-6350.
Chen, Y., Brunson, K.L., Muller, M.B., Cariaga, W., Baram, T.Z., 2000. Immunocytochemical distribution of
corticotropin-releasing hormone receptor type-1 (CRF(1))-like immunoreactivity in the mouse brain: light
microscopy analysis using an antibody directed against the C-terminus. J Comp Neurol. 420(3), 305-23.
Crepel, F., Delhaye-Bouchaud, N., Guastavino, J.M., Sampaio, I., 1980. Multiple innervation of cerebellar Purkinje
cells by climbing fibres in staggerer mutant mouse. Nature. 283(5746),483-4.
Crepel, F., Mariani, J., 1976. Multiple innervation of Purkinje cells by climbing fibres in the cerebellum of the
Weaver Mutant Mouse. J Neurobiol. 7(6),579-82.
Desclin, J.C., Colin, F., 1980. The olivocerebellar system. II. Some ultrastructural correlates of inferior olive
destruction in the rat. Brain Res.187(1), 29-46.
Ha, B.K., Bishop, G.A., King, J.S., Burry, R.W., 2000. Corticotropin releasing factor induces proliferation of
cerebellar astrocytes. J Neurosci Res. 62(6), 789-98.
Ito, M., 1984. The cerebellum and Neural Control. Raven Press. New York.
Koob, G.F., Heinrichs, S.C., 1999. A role for corticotropin releasing factor and urocortin in behavioural responses
to stressors. Brain Res. 848(1-2), 141-52.
Kozicz, T., Yanaihara, H., Arimura, A., 1998. Distribution of urocortin-like immunoreactivity in the central nervous
system of the rat. J Comp Neurol. 391(1),1-10.
Mariani, J., Crepel, F., Mikoshiba, K., Changeux, J.P., Sotelo, C., 1977. Anatomical, physiological and biochemical
studies of the cerebellum from Reeler mutant mouse. Philos Trans R Soc Lond B Biol Sci 281(978), 1-28.

72
Chapter 3
Miyata, M., Okada, D., Hashimoto, K., Kano, M., Ito, M., 1999. Corticotropin-releasing factor plays a permissive
role in cerebellar long-term depression. Neuron. 22(4), 763-75.
Morando, L., Cesa, R., Rasetti, R., Harvey, R., Strata, P., 2001. Role of glutamate delta -2 receptors in activity-
dependent competition between heterologous afferent fibres. Proc Natl Acad Sci U S. 98(17), 9954-9.
Morin, S.M., Ling, N., Liu, X.J., Kahl, S.D., Gehlert, D.R., 1999. Differential distribution of urocortin- and
corticotropin-releasing factor-like immunoreactivities in the rat brain. Neuroscience. 92(1), 281-91.
Muller, R.A., Courchesne, E., Allen, G., 1998. The cerebellum: so much more. Science. 282, 879-80.
Overbeck, T.L., King, J.S., 1999. Developmental expression of corticotropin releasing factor in the postnatal
murine cerebellum. Brain Res Dev Brain Res. 115(2), 145-59.
Palay, S.L., Chan-Palay, V., (Eds) 1974. Cerebellar Cortex. Cytology and Organisation. Berlin: Springer.
Palkovits, M., Leranth, C., Gorcs, T., Young, W.S., 1987. Corticotropin-releasing factor in the olivocerebellar tract
of rats: demonstration by light- and electron-microscopic immunohistochemistry and in situ hybridisation
histochemistry. Proc Natl Acad Sci U S A. 84(11), 3911-5.
Potter, E., Sutton, S., Donaldson, C., Chen, R., Perrin, M., Lewis, K., Sawchenko, P.E., Vale, W., 1994. Distribution
of corticotropin-releasing factor receptor mRNA expression in the rat brain and pituitary. Proc Natl Acad Sci USA.
91,8777-8781.
Rossi, F., Wiklund, L., van der Want, J.J., Strata, P., 1991. Reinnervation of cerebellar Purkinje cells by climbing
fibres surviving a subtotal lesion of the inferior olive in the adult rat. I. Development of new collateral branches
and terminal plexuses. J Comp Neurol. 308(4), 513-35.
Rossi, F., van der Want, J.J., Wiklund, L., Strata, P., 1991. Reinnervation of cerebellar Purkinje cells by climbing
fibres surviving a subtotal lesion of the inferior olive in the adult rat. II. Synaptic organization on reinnervated
Purkinje cells. J Comp Neurol. 308(4), 536-54.
Roste, G.K., Dietrichs, E., 1988. Cerebellar cortical and nuclear afferents from the Edinger-Westphal nucleus in
the cat. Anat Embryol (Berl). 178(1), 59-65.
Schwartz, J.H., 2000. Neurotransmitters. In: Kandel, E.R., Schwarz, J.H., Jessell, T.M. (Eds), Principles of Neural
Science. McGraw-Hill, pp280-297.
Spina, M., Merlo-Pich, E., Chan, R.K., Basso, A.M., Rivier, J., Vale, W., Koob, G.F., 1996. Appetite-suppressing
effects of urocortin, a CRF-related neuropeptide. Science. 273(5281), 561-4.
Strata, P., Tempia, F., Zagrebelsky, M., Rossi, F., 1997. Reciprocal trophic interactions between climbing fibres
and Purkinje cells in the rat cerebellum. Prog Brain Res. 14, 263-82.
Takahashi, K., Totsune, K., Sone, M., Murakami, O., Satoh, F., Arihara, Z., Sasano, H., Iino, K., Mouri, T., 1998.
Regional distribution of urocortin-like immunoreactivity and expression of urocortin mRNA in the human brain.
Peptides. 19(4), 643-7.
Vaughan, J., Donaldson, C., Bittencourt, J., Perrin, M.H., Lewis, K., Sutton, S., Chan, R., Turnbull, A.V., Lovejoy,
D., Rivier, C., et al. 1995. Urocortin, a mammalian neuropeptide related to fish urotensin I and to corticotropin-
releasing factor. Nature. 378(6554), 233-4.
van den Pol, A.N., Gorcs, T., 1986. Synaptic relationships between neurons containing vasopressin, gastrin-releasing
peptide, vasoactive intestinal polypeptide, and glutamate decarboxylase immunoreactivity in the suprachiasmatic
nucleus: dual ultrastructural immunocytochemistry with gold-substituted silver peroxidase. J Comp Neurol. 252(4),
507-21.
Welsh, J.P., Lang, E.J., Sugihara, I., Llinas, R., 1995. Dynamic organization of motor control within the
olivocerebellar system. Nature. 374, 453-7.
Wickelgren, I., 1998. The cerebellum: the brain’s engine of agility. Science. 281,1588-90.
Yamano M., Tohyama M. (1994) Distribution of corticotropin-releasing factor and calcitonin gene-related peptide
in the developing mouse cerebellum. Neurosci Res. 19(4),387-96.

73
CHAPTER 4
The postnatal developmental expression pattern of urocortinin the rat olivocerebellar system
Jerome D. Swinny1, Dharamdajal Kalicharan1, Fuxin Shi3 ,
Albert Gramsbergen2 , Johannes J.L. van der Want1
1Laboratory for Cell Biology and Electron Microscopy, and2Department of Medical Physiology, Graduate School of Behavioural and
Cognitive Neurosciences,3Department of Membrane Cell Biology, University of Groningen,
The Netherlands.
Journal of Comparative Neurology: submitted

74
Chapter 4
ABSTRACT
Urocortin belongs to the family of corticotropin releasing factor (CRF)-like peptides
that play an important role in sensorimotor coordination. CRF induces locomotor
activity whilst urocortin has an inhibitory effect. Here, we document the regional
and subcellular localisation of urocortin in the developing rat cerebellum to compare
it with CRF. Urocortin immunoreactivity (UCN-ir) was evident within the white
matter and cerebellar cortex of all vermal lobes during the first postnatal week. Cortical
immunoreactivity was in the form of weak, scattered immunoreactive puncta that
encircled Purkinje cell somata. Towards the end of the first postnatal week, Purkinje
cells expressed UCN-ir, particularly in lobules I,II, IX and X. By postnatal day 12,
UCN-ir had increased appreciably in all lobules. In Purkinje cells, labelling was
spread throughout their somata and proximal dendrites. By postnatal day 15, labelling
in lobules I-IV appeared to wane, yet still prevailed in the central and posterior lobules.
This anterior to posterior gradient persisted through to adulthood. The study shows
that urocortin and CRF have similar regional distribution profiles during development
suggesting synergistic roles within the vestibulocerebellum. The onset of the adult
distributional pattern of urocortin at the stage when rats are capable of fluent walking
patterns further strengthens the correlation between CRF like peptides and postural
control. An important difference between urocortin and CRF is the localisation of
urocortin, and not CRF, within Purkinje cells implying that urocortin probably has
an additional role in modulating the signals eminating from the cerebellar cortex to
the deep cerebellar nuclei.

75
postnatal developmental expression
INTRODUCTION
The processes involved in the development of cerebellar circuitry eventually manifest
in a structure pivotal for the integration of sensory information and locomotor activity,
providing for coordinated motor activity (Ito, 1984) and, its is thought, higher cognitive
function (Wickelgren, 1998). Studying the underlying structural or morphological
development in relation to behavioural development possibly may elucidate causal
relationships and provide a convenient paradigm for brain behaviour relationships.
The protracted development of the cerebellum, in relation to other brain structures
provides a convenient model to study such events.
Corticotropin-releasing factor (CRF) (Vale et al., 1981) is the first member of a family
of peptides shown to modulate the body’s response to stress (Heinrich and Koob,
2000). Considerable evidence shows that coupled to its stress response, CRF is also
capable of inducing locomotor activity (Contarino et al., 2000; Terawaki et al., 2002;
Zorrilla et al., 2002). The precise neural correlates for this effect on motor activity is
unknown (Zorrilla et al., 2002). CRF is prominently expressed in the cerebellum
(Palkovitz et al., 1987). The principle sites of expression are mossy fibres and climbing
fibres (Palkovitz et al., 1987). During ontogeny, labelling studies have shown that at
early stages, CRF is evenly distributed throughout the cerebellum. From the stage of
postnatal day (PD) 15 onwards, a more discrete lobular localisation and parasagittal
banding pattern emerges (Overbeck and King, 1999; Yamano and Tohyama, 1994;
van den Dungen et al., 1988). This stage of development corresponds to the time
when fluent and adult-like walking patterns emerge (Westerga and Gramsbergen,
1990).
In vitro studies show that the excitatory effect of CRF becomes progressively stronger
from PD 9 (Bishop, 2002). At this stage, the rat is able to stand on its fours and to
perform a few steps (Westerga and Gramsbergen, 1990). Evidence indicates that the
stage of cerebellar development is the limiting factor for the walking pattern
(Gramsbergen et al., 1998) in which extremity movements and postural control are
closely coupled (Gramsbergen, 1999). Furthermore, CRF is essential for the induction
of long-term depression in the cerebellum (Miyata et al., 1999), a phenomenon
proposed as the cellular basis of learning (Ito, 1984). Taken together, the above
suggests a causal relationship between the developmental expression of CRF and
coordinated motor activity.
Recently, we have shown that urocortin (Vaughan et al., 1995), a relatively new
addition to the burgeoning class of CRF-like peptides is expressed in the adult rat
cerebellum (Swinny et al., 2002). Urocortin was localised not only within climbing
and mossy fibres, but most intriguingly, also within cerebellar neurons (Swinny et

76
Chapter 4
al., 2002). Urocortin has been shown to induce mild locomotor suppression. The
presence of urocortin in the cerebellum and in other specific regions of the brain
(Bittencourt et al., 1999) as well as its direct effects on locomotion (Valdez et al.,
2002) strongly suggests an important role in motor control and sensorimotor
integration. Up till now, there are no data on the developmental expression of urocortin
in the cerebellum. In the present study, we provide data on the temporal and spatial
distribution of urocortin immunoreactive profiles in the cerebellum during the first
three postnatal weeks.
MATERIAL AND METHODS
Immunocytochemistry
Light microscopy was used to map the regional distribution of urocortin
immunoreactivity and immunoelectron microscopy was used to positively identify
the immunoreactive profiles.
Black-hooded Lister rats were studied between the ages of PD 3 to PD 25. The day
of birth was considered PD 0. Approval to conduct the study was obtained from the
Ethics Committee on Animal Experimentation, University of Groningen. All efforts
were made to minimise the number of animals used and their suffering. Animals
were deeply anaesthetised with sodium pentobarbital (NembutalR, i.p., 50mg/kg)
and perfused transcardially, first with a solution containing 2% polyvinyl pyrrolidone
(molecular weight 30 000), 0.4% NaNO3 in 0.1 M phosphate buffer (PB) (pH 7.4) for
one minute, and then with 4% paraformaldehyde and 0.2% picric acid in 0.1M PB
(pH 7.4) for ten minutes. The brains were removed and stored overnight in the fixative
solution at 4o C.
Light microscopy
Brains were stored overnight at 4oC in 30% sucrose for cryoprotection and cryo-
sectioned (30µm thick) in either the sagittal or transverse plane. Two animals were
evaluated in either plane. Immunoreactivity was visualised by the avidin-biotin-
peroxidase complex (ABC) method. Briefly, free-floating sections were immersed
for two hours in a pre-incubation medium containing 1% normal rabbit serum and
1% bovine serum albumin in 0.1M phosphate buffered saline (PBS) (pH 7.4). The
sections were then incubated overnight at 4o C with affinity purified rabbit anti-
urocortin IgG (Sigma), diluted 1:2000 with PBS. After washing with PBS, the sections
were incubated with biotinylated goat anti-rabbit IgG, diluted 1:200 in PBS, for two
hours at room temperature. After further washing, the sections were incubated with
ABC (Vectastain Elite, Vector Labs, Burlingame, CA) for one hour at room

77
postnatal developmental expression
temperature. Immunoreactivity was visualised by incubation with 5 mg 3,3 diamino-
benzidine HCl (DAB) and 0.03% hydrogen peroxide in 10 ml of 0.1 M PBS (pH 7.4)
for 5 to 10 minutes at room temperature. The reaction was stopped by washing the
sections in cold PBS.
Selected sections were photographed with an Olympus Digital camera, mounted on
an Olympus B50 optical microscope. These images were adjusted to enhance the
contrast and saved at 300 dpi.
Transmission electron microscopy
Fifty micrometer sections were cut on a Vibratome in the sagittal and transverse
planes. Immunoreactivity was visualised by the avidin-biotin-peroxidase complex
(ABC) method. Briefly, free-floating sections were immersed for two hours in a pre-
incubation medium containing 1% normal rabbit serum and 1% bovine serum albumin
in 0.1M phosphate buffered saline (PBS) (pH 7.4) and processed as for light
microscopy except that 0.025% Triton X-100 was added to the primary antibody
incubation. Immunoreactivity was enhanced according to the gold substituted silver
peroxidase method (van den Pol and Gorcs, 1986). Sections were then osmicated in
1% OsO4,
1.5% potassium hexacyanoferrate in 0.1M cacodylate buffer (pH 7.6) for
15 minutes, dehydrated in a graded series of ethanols and embedded in Epon. Semithin
sections (1 µm) were cut on an LKB Ultratome, stained with toluidine blue and used
for orientation purposes. Serial ultrathin sections were cut, counterstained with uranyl
acetate and lead citrate and examined with a Philips CM 100 transmission electron
microscope.
Antiserum
The antiserum used in this study was raised against a peptide mapping at the carboxy
terminus of urocortin of rat origin (identical to corresponding mouse and human
sequences). It reacts with urocortin of mouse, rat and human and does not cross-
react with CRF.
Controls
Western blotting was used to check the specificity of the urocortin antiserum used as
well as any cross-reactivity with CRF. Briefly, samples were prepared from fresh PD
8 and PD 15 rat cerebellums, lysed in sample buffer (3ml sample buffer/mg
cerebellum; 5%SDS, 5% a-mercaptoethanol, 8M urea, 6.25mM Tris-HCl, PH 6.8,
and 0.01% bromophenol blue). Rat urocortin (Sigma) was used as a positive control.
Rat CRF (Sigma) was used to assess the cross reactivity of the antiserum with CRF.
Five or ten nanogram urocortin and CRF, and 5 microliter cerebellum samples were

78
Chapter 4
separated on 12.5% SDS-PAGE (Bio-Rad, Hercules, CA), blotted on a pure
nitrocellulose membrane (Bio-Rad, Hercules, CA) and probed with rabbit anti-
urocortin (1:500; Sigma). The CRF blot was visualised by a horseradish peroxidase-
conjugated rabbit anti-goat antibody (1:5000, Sigma) or anti-rabbit antibody (1:5000,
Nenox), and processed using ECL ( Amersham Pharmacia Biotech).
Figure 1. Western blot analysis of cerebellarhomogenates of PDs 8 and 15 showing thespecificity the antiserum. (A) at both stages,the urocortin antiserum recognised a singleprotein, in the range of 6.5 to 16.5 kDa. (B)shows that the antiserum was specific forurocortin and did not cross-react with CRF.
RESULTS
Controls
Western blotting
The antiserum was highly specific, recognising a single protein in the range of 6.5 to
16.5 kDa from homogenates of both PD 8 and PD 15. This is in accordance with
previous observations of the molecular weight of urocortin (Vaughan et al., 1995).
The immunoblots also revealed that the urocortin antiserum was not cross-reactive
with CRF (figs 1 A and B).
Light microscopy
Cerebellum (sagittal plane)
During the first postnatal week, urocortin immunoreactivity (UCN-ir) was present
throughout the cerebellar cortex and the deep cerebellar nuclei. At this stage, labelling
was most intense in lobules I, II, IX, X, followed by lobules IV, V, VIII and then
lobules VI and VII. In the cerebellar cortex, labelling, in the form of punctate elements,
was most abundant in the external granular layer (EGL) and the still multi-layered
Purkinje cell layer. Labelled puncta in the Purkinje cell layer generally encircled
Purkinje cell somata, with the cell bodies themselves not appearing to be
immunoreactive (fig. 2 A-C). From the second postnatal week, urocortin
immunoreactive punctate elements in the cerebellar white matter distributed fascicles

79
postnatal developmental expression
Figure 2. Light microscopical surveys of UCN-ir, viewed in the sagittal plane from PDs 3-12.(A) and (B) show that during the first postnatal week, the immunoreactivity was not restrictedto any particular lobule and was resident mainly in the Purkinje cell layer and in the externalgranular layer. (C) a magnified region of the boxed area in (B) showing immunoreactivepunctate elements around a Purkinje cell soma (arrowheads). (D) at PD 9, strong labellingwas evident in fascicle bundles in the white matter of anterior lobules (arrowheads) and inthe Purkinje cell layers of lobules I and II (arrows). (E) in posterior lobules, similar labellingwas evident in the white matter (arrowheads) and particularly in the Purkinje cell layer oflobule IX b and the base of lobule IX (arrows). (F) a magnified view of the boxed area in (E)showing reaction product diffusely spread throughout the soma of a Purkinje cell without anysignificant dendritic labelling at this stage. (G) shows that PD 12, immunoreactivity wasgenerally evenly distributed throughout all lobules, having increased appreciably in thoselobules which had earlier exhibited weak labelling. (H) is a magnified view of the boxed areain (G) showing that at PD 12, immunoreactivity within Purkinje cells had spread into theprimary dendritic shafts (I) an overview of the molecular layer in lobule IX showingimmunoreactive interneuron (arrowheads). Scale bars: A, B, G, 100 µm; C, F, H, 12 µm; D-E, 50 µm; I, 25 µm.

80
Chapter 4
particularly to lobules I and II of the anterior lobe, but also, to a lesser extent, lobules
IX and X of the posterior lobe. Reaction product in the Purkinje cell layer was most
prominent in lobule I and lobule IX b and to a lesser extent in lobule X (fig 2 D-F).
Between PDs 9-12, immunoreactivity in the Purkinje cell layer increased considerably
in all vermal lobules. Reaction product, especially in the posterior and central lobules,
appeared to be spread across the entire somata of Purkinje cells and into their proximal
dendrites. Between the stages of PDs 6- 12, labelling in interneurons in the molecular
layer, most likely stellate and basket cells, was evident (figs 2 G-I).
Figure 3. Light microscopical surveys of UCN-ir, viewed in the sagittal plane from PDs 15 and25. (A) shows that at PD 15, immunoreactivity in lobule II had decreased whilst still remainingintense in (B) the central lobules (lobule VI shown) and (C) the posterior lobules (lobule Xshown). (D) a magnified view of lobule IX at PD 15 showing that reaction product had spreadinto the primary dendrites of Purkinje cells (arrowheads). At PD 25, the anterior lobules suchas lobule II (E) showed weak or indistinct labelling in the Purkinje cell layer in comparison totheir central and posterior counterparts, (F) and (G) respectively. (H) a survey of lobule IX atPD 25 showing a climbing fibre profile making contact with a Purkinje cell with (I) a magnifiedview of the boxed area in (H). Scale bars; A-C, E-G, 50 µm; D, H, 25 µm; I, 12 µm.

81
postnatal developmental expression
At PD 15, urocortin immunoreactivity remained prominent in Purkinje cells of the
central and posterior lobules whilst immunoreactivity within anterior lobules appeared
less conspicuous. Dendritic labelling was most prominent in the posterior lobules,
with Purkinje cells in the anterior lobules rarely exhibiting any dendritic reaction
product. Dendritic labelling was always restricted to the proximal dendritic shafts
and was not present in the tiny branchlets (figs 3 A-D). From the stages of PDs 15-
25, immunoreactivity appeared to decrease especially in the anterior lobules yet
persisting in the central and posterior lobules (figs E-G). Immunoreactive profiles
resembling characteristic climbing fibres were noticeable from PD 15 onwards,
especially in the posterior lobules (figs 3 H-I).
Cerebellum (transverse plane)
Transverse sections were examined to assess whether urocortin immunoreactivity
exhibited any parasagittal banding pattern. At all developmental stages examined,
no such pattern or gradient in immunoreactivity intensity in the medio-lateral plane
was evident (fig 4).
Figure 4. Light microscopical surveys of urocortin immunoreactivity viewed in the transverseplane. (A) shows at PD 12, immunoreactivity in lobules I and III being consistent in themediolateral plane with (B) showing a similar distribution in posterior lobules. (C) at PD 25,immunoreactivity in lobule I and III was much weaker compared with earlier stages. However,in the posterior lobules (D) immunoreactivity remained consistently intense from vermalregions to hemispheres. Scale bars 200 µm

82
Chapter 4
Inferior Olive
At PD 3, strong immunoreactivity was evident in the inferior olivary complex.
Immunoreactive neurons included those of the principal olive, dorsal and medial
accessory olives including the dorsal cap. This same pattern of labelling persisted
throughout ontogeny.
Electron microscopy
Purkinje cell layer
The ultrastructural data showed that urocortin immunoreactivity was present in the
cytoplasm of Purkinje cell somata. Close inspection revealed that in the apical cap of
Purkinje cell somata, labelling was often associated with endoplasmic reticulum and
Figure 5. Overviews of urocortinimmunoreactivity at different stageswithin the inferior olive. (A) at PD 3intense immunoreactivity was evidentin the dorsal accessory olive, theprincipal olive and medial accessoryolive. This pattern of expressionremained consistent during the secondpostnatal week (B) and in youngadulthood (C). DC, dorsal cap of Kooy;DOA, dorsal accessory olive; MAO,medial accessory olive; PO, principalolive. Scale bars; 100 µm

83
postnatal developmental expression
Figure 6. Electronmicrographs showing the distribution of UCN-ir in Purkinje cell profiles atdifferent developmental stages. (A) an overview at PD 6 of the still multi-layered Purkinjecell layer showing reaction product (arrowheads) in the somata of two Purkinje cells. (B)shows the distribution of immunoreactivity within the soma of a Purkinje cell at PD 12.Reaction product had increased appreciably, with it being closely associated with theendoplasmic reticulum (arrowheads). (C) shows a soma of a Purkinje cell at PD 15, withreaction product at the endoplasmic reticulum (arrowhead) and also in a multivesicular body(arrow). (D) at PD 25, reaction product was convincingly demonstrated in the Purkinje celldendrite (arrowhead) with (E) showing reaction product selectively localised within glia apposedto a Purkinje cell dendrite (arrowheads). Pc, Purkinje cell; Pd, Purkinje cell dendrite. Scalebars: A, 5 µm; B-E, 1 µm.

84
Chapter 4
multi-vesicular bodies. Labelling was present in Purkinje cells of lobules IX and X
from as early as PD 6. Most of the reaction product between the stages of PDs 6 to 12
was confined to their somata. Between the stages of PDs 12-15, urocortin
immunoreactivity was convincingly demonstrated in the primary dendritic shafts of
Purkinje cells. At the young adulthood stage (PD 25 onwards), dendritic labelling
was restricted to the proximal shafts and isolated spines, that were often contacted
by climbing fibres. Purkinje cell spines with synapses of parallel fibres did not exhibit
immunoreactivity (fig 6).
Figure 7. Electron micrographs showing urocortin immunoreactivity with the external granularlayer and molecular layer. (A) shows a reaction product contained within a mixture of celltypes in the external granular layer at PD 3 (arrowheads). (B) shows immunoreactive granulecells (arrowheads) at PD 3, identified by their electron dense nuclei. (C) shows reactionproduct (arrowheads) within an interneuron located in the lower molecular layer at PD 6. Thecell is deemed to be a basket cell due to its location and its darker nucleus (in comparison tostellate cells). (D) shows an immunoreactive neuron (arrowheads), imaged from the uppermolecular layer. Its location and pale nucleus suggests that it is a stellate cell. Bc, basket cell;Gc, granule cell; Sc, stellate cell. Scale bars: A, 5 µm; B-D, 1 µm.

85
postnatal developmental expression
External granular layer and molecular layer
At PD 3, urocortin immunoreactivity was readily visualised in a mixture of
differentiating neurons that are present in the external granular layer (fig 7 A).
Differentiating granule cells, identified by their spherical and electron dense nuclei,
were found to be immunoreactive (fig 7 B). Most of the immunoreactivity was found
in their somata, but was more intense in the axons, the developing parallel fibres.
Granule cells were thus immunoreactive prior to the onset of polar extension that
immediately precedes the formation of parallel fibres. This resulted in most of the
labelled product, at later stages, being present in parallel fibres, whereas minimal
amounts were located in the somatic regions, even when the cell bodies descended
Figure 8. Electron micrographs showing urocortin immunoreactivity within cerebellar afferents.(A) an overview of the Purkinje cell layer showing shows labelled terminals (arrowheads)forming synaptic contacts on somatic spines of a Purkinje cell at PD 6, characteristic ofclimbing fibres at this stage with (B) a magnified region of the boxed area in (A). (C) showsreaction product at PD 25 in a climbing fibre terminal, identified by its contact on a proximalspine of a Purkinje cell dendrite and the dense spherical vesicles contained within. (D) asurvey of the upper molecular layer showing immunoreactive parallel fibres at PD 9. Cf,climbing fibre; Pc, Purkinje cell; Pd, Purkinje cell dendrite; Sp, Purkinje cell spine. Scale bar:A and D, 1 µm; B and C, 200 nm.

86
Chapter 4
through the molecular layer. From PD 6 onwards, UCN-ir was visualised in basket
and stellate inhibitory interneurons of the molecular layer. Basket cells, identified by
their spherical and darker nuclei (in comparison to stellate cells) and their position,
juxtaposed between parallel fibres and immediately adjacent to the Purkinje cells,
exhibited prominent, scattered somatic labelling. Immunoreactive stellate cells were
identified by their pale nuclei and position in the upper molecular layer (figs 7 C-D).
Afferent systems
Cell bodies of inferior olivary neurons contained significant amounts of reaction
product in the cytoplasm from as early as PD 3 (see fig 4). Labelled terminal
varicosities forming characteristic climbing fibre synaptic contacts on spines of
Purkinje cell somata were evident at PD 6. At later stages (PD 15 onwards), labelled
profiles, resembling climbing fibre terminals, were found to make synaptic contact
with the proximal spines of Purkinje cell dendrites. Labelling in the terminals consist
of dense precipitates (figs 8 A-C). At PD 9, labelled parallel fibre varicosities made
synaptic contact on Purkinje cell dendritic spines (fig 8 D).
DISCUSSION
The current study provides evidence of prominent urocortin expression in the
developing cerebellum. Expression is most intense in lobules I and II of the anterior
lobe and lobules IX and X of the posterior lobe, closely followed the central lobules.
By PD 12, differences in labelling intensity within lobules are difficult to discern. A
plateau in immunoreactive intensity is reached at PD 15, after which, urocortin
expression appears to taper off, most markedly in the anterior lobe. Furthermore, by
serial sectioning of specimens prepared for immunoelectron microscopy, we could
confirm that at early stages during development, urocortin is localised within Purkinje
cells and also in profiles such as glial sheaths, climbing fibre and basket cell terminals
which are closely apposed to Purkinje cell somata and are also immunoreactive
(Swinny et al., 2002). This is an important confirmation since CRF is not expressed
by any cerebellar neurons.
The developmental distribution profiles of urocortin and CRF, unsurprisingly, show
strong similarities. Both peptides show an initial widespread distribution throughout
the cerebellum during the first two postnatal weeks (Overbeck and King, 1999;
Yamano and Tohyama, 1994). At later stages, immunoreactivity of both peptides
appears to diminish in the anterior lobules of the cerebellum, yet persisting in central
and posterior lobules. Since CRF receptors are also expressed during development
(Chang et al., 1993), CRF and urocortin would appear to be functional already during

87
postnatal developmental expression
the early stages of cerebellar morphological development and synaptogenesis. In
other brain centres, CRF has been shown to modulate neurite outgrowth (Cibelli et
al., 2001). Taken together, it strongly implies that urocortin and CRF assume
important, though yet to be defined roles in the establishment of the cerebellar
circuitry.
There are also some striking differences between the expression profiles of urocortin
and CRF. CRF is evident in the entire fibre tracts of inferior olivary neurons. However,
urocortin appears to be selectively localised in only their terminals once they reach
the molecular layer and are termed climbing fibres. The widespread and intense
urocortin immunoreactivity within the neurons of the inferior olive, the source of
climbing fibres, confirms the presence of this peptide in the climbing fibre terminals.
Also, using immunoelectron microscopy, we were able to identify immunoreactive
profiles as climbing fibres. At later stages, only isolated climbing fibres showed
urocortin immunoreactivity in their fibre tracts (see figure 3 H) indicating that
urocortin is not only transiently expressed but remains so into adulthood. The
differential processing by the inferior olive of CRF and urocortin, possibly indicates
that these two peptides subserve varying roles at the climbing fibre-Purkinje cell
synapse.
Furthermore, at later stages, whilst CRF and urocortin show similar lobular
distribution patterns, a parasagittal banding pattern of CRF positive climbing fibres
emerges between the second and third postnatal weeks suggesting that these profiles
innervate specific compartments of Purkinje cells (Yamano and Tohyama, 1994). In
this and in a previous study (Swinny et al., 2002), we observed that there was no
indication of a parasagittal banding pattern with respect to urocortin immunoreactive
profiles, only a lobular gradient could be observed. This might indicate that urocortin
serves a more global role within the cerebellar circuitry in comparison to CRF, that
seems more restricted.
A crucial difference between urocortin and CRF is the expression of urocortin in
Purkinje cells and cerebellar interneurons. This implies that urocortin is not only
involved with the processing of sensory information conveyed within cerebellar
afferents, but also with signals leaving the cerebellar cortex via Purkinje cells. A
tantalising hypothesis for the CRF system in the cerebellar circuitry would be that
CRF and urocortin are neuromodulatory in the input loop via the cerebellar afferents
with urocortin being part of the output loop via the projection of Purkinje cell axons
to the deep cerebellar nuclei. Most labelling studies show that CRF and urocortin are
predominantly localised in the posterior vermal lobules, the flocculus and
paraflocculus, the regions often termed the vestibulocerebellum. This strongly
suggests that the cerebellar CRF system could be instrumental in the vestibular and

88
Chapter 4
postural component of motor control. This is borne out of the strong correlation
between urocortin and CRF reaching their adult-like expression patterns at about PD
15 and the ability of the animal at this stage, to effect fluent and coordinated walking
patterns (Westerga and Gramsbergen, 1990). Earlier neurophysiological studies
indicate that this fluent walking pattern is dependent upon feed forward control of
posture (Gramsbergen et al., 1998). Behavioral studies on the vestibular ocular reflex
or other tests to assess postural control mechanisms using recently generated urocortin
mutants (Vetter et al., 2002; Wang et al., 2002) will further illuminate the role of
urocortin in cerebellar function.
REFERENCES
Bishop GA. 2002. Development of a corticotropin-releasing factor-mediated effect on the firing rate
of Purkinje cells in the postnatal mouse cerebellum. Exp Neurol 178:165-74.
Bittencourt JC, Vaughan J, Arias C, Rissman RA, Vale WW, Sawchenko PE. 1999. Urocortin expression
in rat brain: evidence against a pervasive relationship of urocortin-containing projections with targets
bearing type 2 CRF receptors. J Comp Neurol. 415: 285-312.
Chang CP, Pearse RV, O’Connell S, Rosenfeld MG. 1993. Identification of a seven transmembrane
helix receptor for corticotropin-releasing factor and sauvagine in mammalian brain. Neuron 11: 1187-
95.
Cibelli G, Corsi P, Diana G, Vitiello F, Thiel G. 2001. Corticotropin-releasing factor triggers neurite
outgrowth of a catecholaminergic immortalised neuron via cAMP and MAP kinase signalling pathways.
Eur J Neurosci 13: 1339-48.
Contarino A, Dellu F, Koob GF, Smith GW, Lee KF, Vale WW, Gold LH. 2000. Dissociation of
locomotor activation and suppression of food intake induced by CRF in CRFR1-deficient mice.
Endocrinology 141:2698-702.
Gramsbergen A. 1998. Posture and locomotion in the rat: independent or interdependent development?
Neurosci Biobehav Rev 22:547-53.
Gramsbergen A, Geisler HC, Taekema H, van Eykern LA. 1999. The activation of back muscles during
locomotion in the developing rat. Brain Res Dev Brain Res 112:217-28.
Ito M. 1984. The Cerebellum and Neural Control. New York: Raven.
Koob GF, Heinrichs SC. 1999. A role for corticotropin releasing factor and urocortin in behavioural
responses to stressors. Brain Res 27: 41-52.
Miyata M, Okada D, Hashimoto K, Kano M, Ito M. 1999. Corticotropin-releasing factor plays a
permissive role in cerebellar long-term depression. Neuron 22: 763-75.
Overbeck TL, King JS. 1999. Developmental expression of corticotropin releasing factor in the postnatal
murine cerebellum. Brain Res Dev Brain Res 115: 145-59.
Palkovits M, Leranth C, Gorcs T, Young WS. 1987 Corticotropin-releasing factor in the olivocerebellar
tract of rats: demonstration by light- and electron-microscopic immunohistochemistry and in situ
hybridization histochemistry. Proc Natl Acad Sci U S A 84: 3911-5.
Swinny JD, Kalicharan D, Gramsbergen A, van der Want JJL. 2002. The localisation of urocortin in
the adult rat cerebellum: a light and electron microscopic study. Neuroscience 114, 891-903.
Terawaki K, Koike K, Yuzurihara M, Kurauchi K, Ishige A, Sasaki H, Murakami K, Inoue M. 2001.

89
postnatal developmental expression
An inhibitory effect of cytokine-induced neutrophil chemoattractant on corticotropin-releasing factor-
induced increase in locomotor activity. Brain Res 917:133-7.
Valdez GR, Inoue K, Koob GF, Rivier J, Vale W, Zorrilla EP. 2002. Human urocortin II: mild locomotor
suppressive and delayed anxiolytic-like effects of a novel corticotropin-releasing factor related peptide.
Brain Res 943:142-50.
Vale W, Spiess J, Rivier C, Rivier J. 1981. Characterization of a 41-residue ovine hypothalamic peptide
that stimulates secretion of corticotropin and beta-endorphin. Science 213: 1394-7.
Van den Dungen HM, Groenewegen HJ, Tilders FJ, Schoemaker J. 1988. Immunoreactive corticotropin
releasing factor in adult and developing rat cerebellum: its presence in climbing and mossy fibres. J
Chem Neuroanat. 1 : 339-49.
Van den Pol AN, Gorcs T. 1986. Synaptic relationships between neurons containing vasopressin, gastrin-
releasing peptide, vasoactive intestinal polypeptide, and glutamate decarboxylase immunoreactivity
in the suprachiasmatic nucleus: dual ultrastructural immunocytochemistry with gold-substituted silver
peroxidase. J Comp Neurol 252: 507-21.
Vaughan J, Donaldson C, Bittencourt J, Perrin MH, Lewis K, Sutton S, Chan R, Turnbull AV, Lovejoy
D, Rivier C, et al. 1995. Urocortin, a mammalian neuropeptide related to fish urotensin I and to
corticotropin-releasing factor. Nature 378: 233-4.
Vetter DE, Li C, Zhao L, Contarino A, Liberman MC, Smith GW, Marchuk Y, Koob GF, Heinemann
SF, Vale W, Lee KF. 2002. Urocortin-deficient mice show hearing impairment and increased anxiety-
like behavior. Nat Genet 31: 363-9.
Wang X, Su H, Copenhagen LD, Vaishnav S, Pieri F, Shope CD, Brownell WE, De Biasi M, Paylor R,
Bradley A. 2002. Urocortin-deficient mice display normal stress-induced anxiety behavior and
autonomic control but an impaired acoustic startle response. Mol Cell Biol 22:6605-10.
Westerga J, Gramsbergen A. 1990. The development of locomotion in the rat. Brain Res Dev Brain
Res 57:163-74.
Wickelgren I. 1998. The cerebellum: the brain’s engine of agility. Science 281: 1588-90.
Yamano M, Tohyama M. 1994. Distribution of corticotropin-releasing factor and calcitonin gene-
related peptide in the developing mouse cerebellum. Neurosci Res 19: 387-96.
Zorrilla EP, Valdez GR, Nozulak J, Koob GF, Markou A. 2002. Effects of antalarmin, a CRF type 1
receptor antagonist, on anxiety-like behavior and motor activation in the rat. Brain Res 952:188-99.

90

91
Chapter 5
Corticotropin releasing factor receptor types 1 and 2 aredifferentially expressed in pre- and postsynaptic elements in
the postnatal developing rat cerebellum
J. D. Swinny1, D. Kalicharan1, E.H. Blaauw1, J. IJkema-Paassen2, F. Shi3 ,
A. Gramsbergen2 , J.J.L. van der Want1
1Laboratory for Cell Biology and Electron Microscopy,2 Department of Medical Physiology, Graduate School of Behavioural and
Cognitive, Neuroscience, &3Department of Membrane Cell Biology, University of Groningen,
The Netherlands
European Journal of Neuroscience: (2003);18(3):549-562

92
Chapter 5
ABSTRACT
Corticotropin releasing factor (CRF)-like proteins act via two G-protein coupled
receptors (CRF-R1 and CRF-R2) playing important neuromodulatory roles in stress
responses and synaptic plasticity. Cerebellar expression of CRF-like ligands have
been well documented, but not their receptor localisation. This is the first combination
light microscopy ultrastructural study to localize CRF receptors immunohistologically
(im) in the developing rat cerebellum. Both CRF-R1 and R2 were expressed in
climbing fibres from early stages (postnatal day 3) to the adult but CRF-R2im was
only prominent in throughout the molecular layer in the posterior cerebellar lobules.
CRF-R1im was concentrated in apical regions of Purkinje cell (PC) somata and later
in primary dendrites exhibiting a diffuse cytoplasmic appearance. In PC’s, CRF-
R1im was never membrane bound postsynaptically in dendritic spines while CRF-
R2im was found on plasmic membranes of PC from postnatal day 15 onwards. We
conclude that the localisation of these receptors in cerebellar afferents implies their
control of the presynaptic release of CRF-like ligands, impacting on the sensory
information being transmitted from afferents. Furthermore, the fact that CRF-R2 is
membrane bound at synapses, while CRF-R1 is not, suggests that ligands couple to
CRF-R2 via synaptic transmission and to CRF-R1 via volume transmission. Finally,
the distinct expression-profiles of receptors along structural domains of PC’s suggest
that the role for these receptors is to modulate afferent inputs.

93
corticotropin releasing factor receptor types 1 and 2
INTRODUCTION
Corticotropin releasing factor (CRF)-like peptides modulate neurite growth (Cibelli
et al., 2000) and synaptic plasticity in the forms of long term depression (Miyata et
al., 1999) and long term potentiation (Wang et al., 2000). Currently, this family of
stress-related peptides consists of CRF (Vale et al., 1981), urocortin (Vaughan et al.,
1995), stresscopin related peptide/urocortin II (Lewis et al., 2001) and stresscopin/
urocortin III (Reyes et al., 2001). CRF in the cerebellum is restricted to the two
major afferent systems, namely the mossy fibres and climbing fibres (Palkovitz et
al., 1987). We have recently demonstrated that urocortin is localised within the
afferents and within cerebellar neurons in the adult rat cerebellum (Swinny et al.,
2002). This disparate localisation of CRF and urocortin suggests contrasting roles
for these peptides in cerebellar function.
CRF-like peptides mediate their effects via two receptors, namely CRF receptor type
one (CRF-R1) (Chang et al., 1993; Chen et al., 1993; Vita et al., 1993) and CRF
receptor type two (CRF-R2) (Lovenburg et al., 1995; Perrin et al., 1995). Both
receptors belong to the family of G-protein coupled receptors and act by activating
adenylate cyclase (Bilizekjian and Vale, 1983; Giguere and Labrie, 1983; Aguilera
et al., 1983). In the rat brain, the most prominent seat of expression of CRF-R1 is the
cerebellum, occurring predominantly in Purkinje cells, from embryonic day 17 through
to adulthood (Chang et al., 1993). In adulthood, Radulovic et al. (1998), van Pett et
al. (2000), Chen et al. (2000) and Bishop et al. (2000) consistently observed,
immunoreactivity in all cerebellar cortical layers and the deep cerebellar nuclei.
However, reports on the immunocytochemical localisation of CRF-R2 are
contradictory. Van Pett et al. (2000) reported no cerebellar CRF-R2 immunoreactivity
whereas Bishop et al. (2000) showed CRF-R2 immunoreactivity in the cerebellar
cortical layers.
The above light microscopical studies suggest that the Purkinje cell is one of the
main sites of CRF receptor expression. However, they do not address such issues as
to whether the receptors are localised in pre- or postsynaptic elements, or both. An
ultrastructural examination at the electron microscopic level is imperative since the
Purkinje cell is composed of functionally distinct regions due to the heterologous
synaptic inputs it receives on different locations (see Voogd and Glickstein, 1998 for
a review). Inhibitory interneurons, like basket cells and stellate cells synapse on the
Purkinje cell somata and dendritic shafts respectively. Climbing fibres and parallel
fibres, provide excitatory input at the proximal stubby dendritic spines and distal,
thinner dendritic spines respectively (Palay and Chan-Palay, 1974; Sotelo, 1978).
This synaptic patterning and morphological development largely occurs within the

94
Chapter 5
first three postnatal weeks in the rat (Altman and Bayer, 1997).
A comprehensive investigation of the developmental expression of CRF-R1 and CRF-
R2 in different regions of the cerebellum, especially the Purkinje cell layer will help
in the understanding of the roles of CRF-like peptides serve, first in motor development
and subsequently in motor learning.
MATERIAL AND METHODS
Black-hooded Lister rats were studied from postnatal day (PD) 3 to 25. The day of
birth was considered postnatal day (PD) 0. Twelve animals in total were used. Approval
to conduct the study was obtained from the Ethics Committee on Animal
Experimentation, University of Groningen. All efforts were made to minimise the
number of animals used and their suffering. Animals were deeply anaesthetised with
sodium pentobarbital (NembutalR, i.p., 50mg/kg).
Western blotting
Levels of CRF receptors in rat cerebellum and the specificity of the binding of the
antisera used, were evaluated by Western immunoblot (Shi et al., 2001; Towbin et
al., 1979). Cerebella samples at different ages were prepared from fresh rat cerebellum
lysed in sample buffer (3ml/mg, 5%SDS, 5% a-mercaptoethanol, 8M urea, 6.25mM
Tris-HCl, pH 6.8, and 0.01% bromophenol blue). Twenty microliter cerebellum
samples were separated on 12.5% SDS-PAGE (Bio-Rad, Hercules, CA), blotted on a
pure nitrocellulose membrane (Bio-Rad, Hercules, CA) and probed with goat anti-
CRF-R1 or CRF-R2 (1:500; Santa Cruz). The CRF-R1 blot was followed by
horseradish peroxidase-conjugated rabbit anti-goat antibody (1:5000, Sigma,
Stinheim, Germany), and processed using ECL (Amersham Pharmacia Biotech)
according to the manufacturer’s instruction. The CRF-R2 blot was followed by
alkaline phosphatase-conjugated rabbit anti-goat antibody (1:3000, Sigma) and colour
processed by nitro blue tetrazolium and 5-bromo-4-chloro-3-indolyl (Sigma).
Immunocytochemistry
Following anaesthesia, animals were perfused transcardially first with 10 ml of a
solution containing 2% polyvinyl pyrrolidone (molecular weight 30000), 0.4% NaNO3
in 0.1 M phosphate buffer (PB) (pH 7.4), and then with 100ml of 4%
paraformaldehyde and 0.2% picric acid in 0.1M PB (pH 7.4). The brains were removed
and stored overnight in the fixative solution at 4o C.
For light microscopy, brains were stored overnight at 4oC in 30% sucrose for

95
corticotropin releasing factor receptor types 1 and 2
cryoprotection and cryo-sectioned (30µm thick) in the sagittal plane. Immuno-
reactivity was visualised by fluorescence. Briefly, free-floating sections were
immersed for two hours in a pre-incubation medium containing 1% normal rabbit
serum and 1% bovine serum albumin in 0.1M phosphate buffered saline (PBS) (pH
7.4). The sections were incubated overnight at 4o C with affinity purified goat anti-
CRF-R1 IgG or goat anti-CRF-R2 IgG (Santa Cruz Biotechnologies, Santa Cruz,
CA), diluted 1:400 in PBS. After washing with PBS, the sections were incubated
with Alexa Fluor® 488 rabbit anti-goat IgG, diluted 1:200 in PBS, for two hours at
room temperature. After further washing, the sections were mounted in Dako anti-
fading mounting medium (Dako Corporation, Ca).
Imaging
All sections were evaluated. However, selected sections were photographed on an
Olympus Digital camera, mounted on an Olympus B50 fluorescence microscope.
When necessary, these images were adjusted to enhance the contrast and brightness
and saved at 300 dpi.
Electron microscopy
Pre-embedding immunocytochemistry
Animals were perfusion fixed as for light microscopy. Fifty micrometer sections
were cut on a Vibratome in the sagittal plane. Immunoreactivity was visualised by
the avidin-biotin-peroxidase complex (ABC) method. Briefly, free-floating sections
were immersed for two hours in a pre-incubation medium containing 1% normal
rabbit serum, 0.025% Triton X-100 and 1% bovine serum albumin in 0.1M phosphate
buffered saline (PBS) (pH 7.4). The sections were incubated overnight at 4o C with
either affinity purified goat anti-CRF-R1 IgG or goat anti-CRF-R2 IgG (Santa Cruz
Biotechnologies, Santa Cruz, CA), diluted 1:400 in PBS. After washing with PBS,
the sections were incubated with biotinylated rabbit anti-goat IgG, diluted 1:200 in
PBS, for two hours at room temperature. After further washing, the sections were
incubated with ABC (Vectastain Elite, Vector Labs, Burlingame, CA) for one hour at
room temperature. Immunoreactivity was visualised by incubation with 5 mg 3,3
diaminobenzidine HCl (DAB) and 0.03% hydrogen peroxide in 10 ml of 0.1 M PBS
(pH 7.4) for 5 to 10 minutes at room temperature. The reaction was stopped by
washing the sections in cold PBS. Immunoreactivity was enhanced according to the
gold substituted silver peroxidase method (van den Pol and Gorcs, 1986). Sections
were osmicated in 1% OsO4,
1.5% potassium hexacyanoferrate in 0.1M cacodylate
buffer (pH 7.6) for 15 minutes, dehydrated in a graded series of ethanols and embedded
in Epon. Semithin sections (1 µm) were cut on an LKB Ultratome, stained with

96
Chapter 5
toluidine blue and used for orientation purposes. Ultrathin sections were cut,
counterstained with uranyl acetate and lead citrate and examined with a Philips CM
100 transmission electron microscope.
Post-embedding immunocytochemistry
Postembedding immunogold labelling was performed according to a modified
protocol of Petralia et al. (1998). Briefly, animals were fixed as for light microscopy.
The cerebella were removed, postfixed, washed, and 100 µm sagittal sections were
cut with a Vibratome. In the last two steps, tissue was kept in phosphate buffer (0.1
M with 4% glucose). Tissue was cryoprotected in a series of 10, 20, and 30% glycerol
(last step overnight) in 0.1 M phosphate buffer, and was plunge-frozen in liquid
propane in a Leica EM CPC. Frozen tissue was immersed in 1.5% uranyl acetate in
methanol at -90°C in a Leica AFS freeze-substitution instrument, infiltrated with
Lowicryl HM 20 resin at -45°C, and polymerized with UV light (-45 to 0°C). Thin
sections were cut on a Leica Reichert Ultracut S ultramicrotome, placed on nickel
grids (Electron Microscopy Sciences, Fort Washington), incubated in 0.1% sodium
borohydride plus 50 mM glycine in Tris-buffered saline/0.1% Triton X-100 (TBST)
for 10 min, and in blocking serum in TBST for 10 min (10% normal goat serum
(NGS). Sections on the grids were incubated in primary antibody (goat anti-CRF-R1
or R2, diluted 1.25 in PBS) overnight at 4°C. After washing in PBS, the grids were
incubated in 1:50 immunogold (0.8 nm rabbit anti-goat immunogold; Amersham:
Arlington Heights). Following washing, the immunogold particles were further
enhanced for 10 minutes in the dark according to a modified protocol of Yi et al.
(2001). Concentrations of primary antibodies were selected to minimise background
immunogold labelling. Such background artefactual staining was examined in control
sections, (processed with the omission of the primary antibodies) and, within the
experimental sections. Immunogold labelled sections were considered acceptable if
they showed little or no labelling inside the mitochondria and nucleus.
Controls
To assess antibody specificity, both primary antibodies were separately pre-incubated
with their specific blocking peptides (Santa Cruz Biotechnologies, Santa Cruz, CA).
Cross-reactivity between the two antisera was tested by applying the blocking peptide
for CRF-R2 in the pre-incubation medium with anti-CRF-R1 and vice versa. To
assess cross-reactivity with CRF or urocortin, the receptor antibodies were pre-
incubated with either synthetic CRF or urocortin peptides. Sections were incubated
with the individual cocktails overnight, and processed further for western blotting or
light microscopy.

97
corticotropin releasing factor receptor types 1 and 2
Antisera
The CRF-R1 antiserum used in this study was raised against a peptide mapping
within an internal region of CRF-R1 of human origin (sc 12381), specific for CRF-
RI of mouse, rat and human origin and does not cross-react with CRF-R2. The CRF-
R2 antiserum used in this study was raised against a peptide mapping at the amino
terminus of CRF-R2 of mouse origin and is specific for CRF-R2 and does not cross-
react with CRF-R1.
RESULTS
Western blotting
The specificity of the anti-CRF-R1 and CRF-R2 antisera were assessed by
immunoblotting rat cerebellar homogenates. Since only single immunoreactive bands
were evident for both antisera, only the regions of the blots containing the
immunoreactive bands are shown. For CRF-R1, the antiserum recognised a protein
in the 40-45 kDa range, which is in accordance with previous observations of the
molecular weight of rat cerebellar CRF-R1 protein (Chang et al., 1993; Grigoriadis
and DeSouza, 1989; Vita et al., 1993). Weak indistinct bands were evident in
immunoblots from homogenates of PD 1 to PD 4, with signal intensity increasing
from PD 8 onwards through to PD 12. A slight tapering off was observed in adult
homogenates (fig 1 A). Preadsorption of the primary antiserum with a blocking peptide
consisting of the immunogenic epitope (100 µg/ml) completely abolished the CRF-
R1 band (data not shown). The specificity of CRF-R1 immunoreactivity was also
demonstrated by the absence of labelling in cerebellar sections that were stained
using antiserum pretreated with the antigenic peptide (data not shown). For CRF-
Figure 1. Western blot analysis of postnatal cerebellar homogenates of PDs 1,4,8,12 and25, showing the specificity and degree of binding of CRF receptor antisera. (A) CRF-R1antiserum recognised a single protein of approximately 40kDa. Weak banding was observedbetween the stages of PD 1 to PD 4. Expression became more intense between the stages ofPD 4 to PD 8 with PD 12 showing the most intense banding pattern. (B) CRF-R2 antiserumconsistently recognised a single protein of approximately 50kDa. In contrast to CRF-R1,banding was more intense and was expressed as early as PD 1 through to young adulthood.However, most intense immunoblotting was observed between the stages of PD 4 through toPD 12.
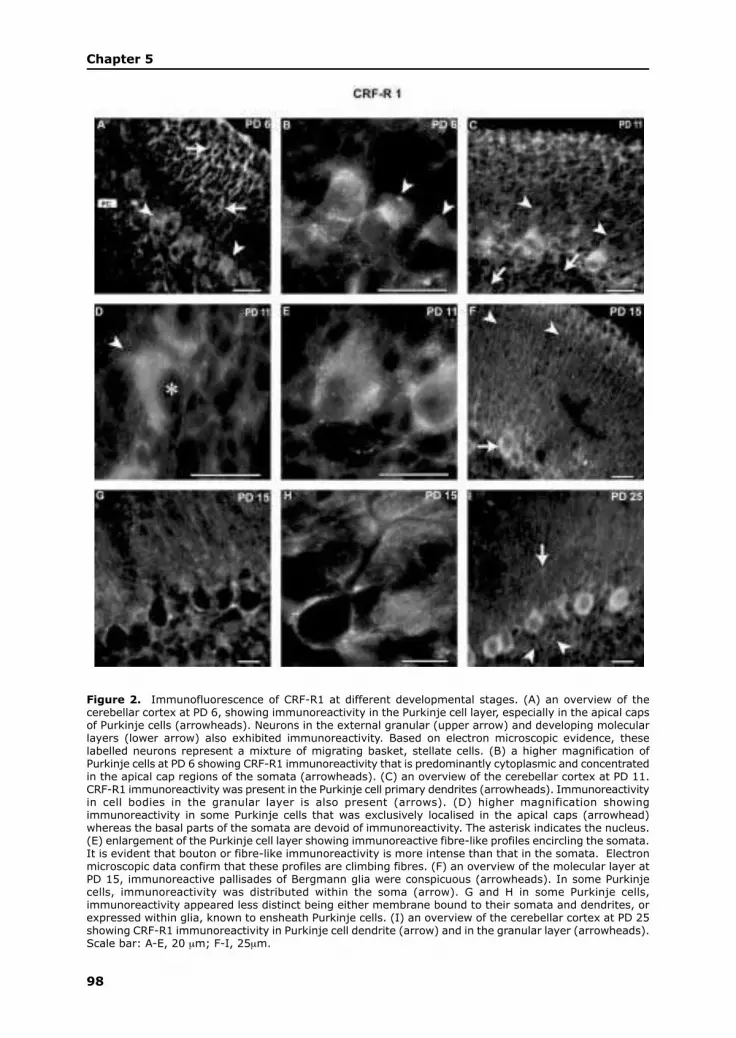
98
Chapter 5
Figure 2. Immunofluorescence of CRF-R1 at different developmental stages. (A) an overview of thecerebellar cortex at PD 6, showing immunoreactivity in the Purkinje cell layer, especially in the apical capsof Purkinje cells (arrowheads). Neurons in the external granular (upper arrow) and developing molecularlayers (lower arrow) also exhibited immunoreactivity. Based on electron microscopic evidence, theselabelled neurons represent a mixture of migrating basket, stellate cells. (B) a higher magnification ofPurkinje cells at PD 6 showing CRF-R1 immunoreactivity that is predominantly cytoplasmic and concentratedin the apical cap regions of the somata (arrowheads). (C) an overview of the cerebellar cortex at PD 11.CRF-R1 immunoreactivity was present in the Purkinje cell primary dendrites (arrowheads). Immunoreactivityin cell bodies in the granular layer is also present (arrows). (D) higher magnification showingimmunoreactivity in some Purkinje cells that was exclusively localised in the apical caps (arrowhead)whereas the basal parts of the somata are devoid of immunoreactivity. The asterisk indicates the nucleus.(E) enlargement of the Purkinje cell layer showing immunoreactive fibre-like profiles encircling the somata.It is evident that bouton or fibre-like immunoreactivity is more intense than that in the somata. Electronmicroscopic data confirm that these profiles are climbing fibres. (F) an overview of the molecular layer atPD 15, immunoreactive pallisades of Bergmann glia were conspicuous (arrowheads). In some Purkinjecells, immunoreactivity was distributed within the soma (arrow). G and H in some Purkinje cells,immunoreactivity appeared less distinct being either membrane bound to their somata and dendrites, orexpressed within glia, known to ensheath Purkinje cells. (I) an overview of the cerebellar cortex at PD 25showing CRF-R1 immunoreactivity in Purkinje cell dendrite (arrow) and in the granular layer (arrowheads).Scale bar: A-E, 20 µm; F-I, 25µm.

99
corticotropin releasing factor receptor types 1 and 2
Figure 3. CRF-R2 immunofluorescence in the cerebellar cortex at different postnatal stages. (A) an overviewfrom lobule III showing immunoreactivity predominating in Purkinje cell somata (arrowheads). The molecularand granular layers show indistinct labelling. (B) an overview of lobule IX showing strong immunostainingin Purkinje cell somata (arrows) and primary dendrites (arrowhead). (C) a detail of the Purkinje cell layer.The immunoreactivity is localised in the cytoplasm and extends into the initial axonal segments (arrowheads).(D) an overview and (E), a higher magnification of the Purkinje cell layer at PD 11, showing thatimmunoreactivity within Purkinje cells is more restricted in comparison with earlier stages. Reactivity isrestricted to cytoplasmic membranes, especially in the basal regions and initial axonal segments (arrowhead).(F) at PD 11, immunoreactive fibre-like profile (arrowheads), extending from the granular layer into themolecular layer were evident. Such bouton or fibre-like profiles were more intense than similar profilesexpressing CRF-R2 immunoreactivity. These are probably a combination of climbing fibres, Purkinje cellaxons or recurrent axonal collaterals based on electron microscopic data (see figure 7 B). (G to I) from PD15 through to adulthood, Purkinje cell immunoreactivity is clearly condensed and membrane bound totheir somata with the cytoplasm being relatively free of labelling. Scale bars: A-F, 20µm; G-I, 25 µm

100
Chapter 5
R2, the antiserum recognised a protein in the 40~50 kDa range, also in close
approximation of previous data (Lovenberg et al., 1995; Perrin et al., 1995; Vita et
al., 1993) (fig 1 B). More intense banding, in comparison to CRF-R1, was evident
from homogenates taken at PD 1. Banding appeared to be equally intense between
the stages of PDs 4 to 12 with a slight decrease at PD 25. Like CRF-R1, preadsorption
of the primary antiserum with a blocking peptide consisting of the immunogenic
epitope (100 g/ml) completely abolished the CRF-R2 band (data not shown). To
assess cross-reactivity between the two receptor antisera, pre-incubation with the
alternate blocking peptides did not reduce immunoreactivity (data not shown).
Similarly, pre-incubation of the two antisera with CRF or urocortin peptides resulted
in no discernable decrease in the intensity of immunoreactivity, confirming that the
antisera are not cross-reactive with these peptides.
Light microscopy
CRF-R1
PD 3 was the first stage to be examined, showing rather weak labelling. However,
from PD 6 onwards, immunoreactivity increased appreciably in all layers of the
cerebellar cortex, in fibre tracts in the cerebellar white matter and in the deep cerebellar
nuclei. The reaction product was most abundant in Purkinje cells, the molecular
layer and the external granular layer. In Purkinje cells, immunoreactivity was first
concentrated in the apical caps and basal region of Purkinje cell cytoplasm. There
were no apparent regional differences between Purkinje cell immunoreactivity.
Interneurons in the molecular layer also exhibited immunoreactivity (figs 2 A and
B). Electron microscopic evidence showed conclusively that both basket and stellate
cells are immunoreactive (see figure 6).
Between the stages of PDs 9 to 12, labelling in Purkinje cells was preferentially
localised in the apical cap of somata and in the proximal regions of the primary
dendrites. This pattern of labelling was not uniform in all Purkinje cells, with some
cells exhibiting labelling in the basal parts of the somata (figs 2 C and D). Also
evident at this stage were varicose fibre-like immunoreactive profiles at the level of
the Purkinje cell somata (fig 2 E). It was apparent that for both CRF-R1 and CRF-
R2, fibre-like or bouton immunoreactivity was more intense than that of the labelling
in cell bodies. At the electron microscopical level, these profiles were shown to be
climbing fibres (see figure 5). At PD 15, Purkinje cell dendritic labelling had become
more pronounced. Labelling within the somata of Purkinje cells was diffuse, not
restricted to any specific regions as in the earlier stages. Immunoreactivity within
the palisades of Bergmann glia was fully evident especially in the central and posterior
lobules (fig 2F). A subset of Purkinje cells showed immunoreactivity only around
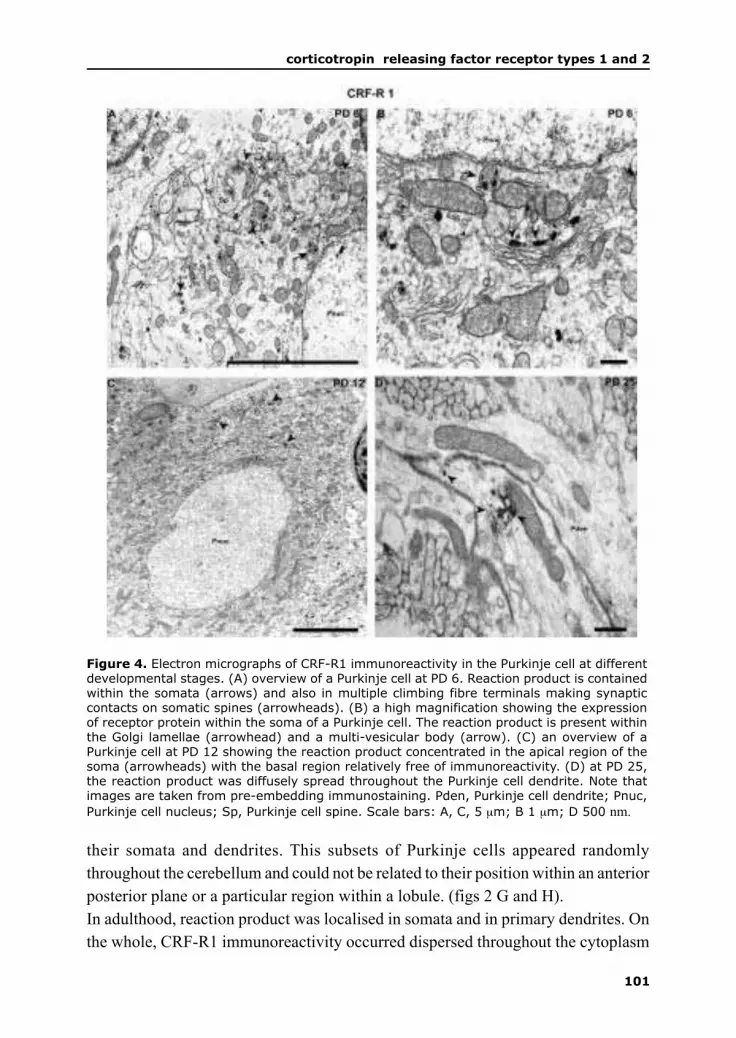
101
corticotropin releasing factor receptor types 1 and 2
their somata and dendrites. This subsets of Purkinje cells appeared randomly
throughout the cerebellum and could not be related to their position within an anterior
posterior plane or a particular region within a lobule. (figs 2 G and H).
In adulthood, reaction product was localised in somata and in primary dendrites. On
the whole, CRF-R1 immunoreactivity occurred dispersed throughout the cytoplasm
Figure 4. Electron micrographs of CRF-R1 immunoreactivity in the Purkinje cell at differentdevelopmental stages. (A) overview of a Purkinje cell at PD 6. Reaction product is containedwithin the somata (arrows) and also in multiple climbing fibre terminals making synapticcontacts on somatic spines (arrowheads). (B) a high magnification showing the expressionof receptor protein within the soma of a Purkinje cell. The reaction product is present withinthe Golgi lamellae (arrowhead) and a multi-vesicular body (arrow). (C) an overview of aPurkinje cell at PD 12 showing the reaction product concentrated in the apical region of thesoma (arrowheads) with the basal region relatively free of immunoreactivity. (D) at PD 25,the reaction product was diffusely spread throughout the Purkinje cell dendrite. Note thatimages are taken from pre-embedding immunostaining. Pden, Purkinje cell dendrite; Pnuc,
Purkinje cell nucleus; Sp, Purkinje cell spine. Scale bars: A, C, 5 µm; B 1 µm; D 500 nm.

102
Chapter 5
was not bound to any cell membranes (fig 2 I). Furthermore, there was no sagittal
banding pattern with respect to CRF-R1 immunoreactivity (data not shown).
Figure 5. Electron micrographs of climbing fibres showing dense and selective CRF-R1immunoreactivity. (A) shows an immunoreactive climbing fibre terminal making synapticcontacts on spines of Purkinje cell dendritic spine at PD 15. (B) shows similar profiles at PD25. Note that the dendritic spines are always free of reaction product whilst the dendriticshaft contains reaction product (arrowheads). Images are taken of pre-embeddingimmunostaining. Cf, climbing fibre; Pden, Purkinje cell dendrite; Sp, Purkinje cell spine.Scale bars, 1 µm

103
corticotropin releasing factor receptor types 1 and 2
CRF-R2
In the first postnatal week, CRF-R2 immunoreactivity was weak and rather indistinct.
At PD 6, immunoreactivity was evident in Purkinje cells and in the granular layer. In
the anterior lobules, labelling was sparse (fig 3 A). However, in the posterior lobules,
labelling in the molecular layer was more intense. Purkinje cell immunoreactivity
Figure 6. Electron micrographs of CRF-R1 immunoreactivity within interneurons in themolecular layer. (A) an overview of a stellate cell at PD 3 identified by its pale nucleus (incomparison to basket cells) showing reaction product within its soma (arrowheads). (B) adetail of an interneuron in the upper molecular layer expressing prominent somaticimmunoreactivity. (C), shows the dendrite of the soma in (B) containing prominent reactionproduct. (D) shows an interneuron in the molecular layer containing membrane boundimmunogold particles (arrowheads) with the insert showing gold particles membrane boundto dendritic profiles also in the molecular layer. Note that figures A to C are of pre-embeddingimmunocytochemistry and figure D of post-embedding immunocytochemistry. Scale bars:A,C 1 µm; B, D, 2 µm

104
Chapter 5
was prominently expressed in apical caps. It was difficult to positively ascertain
whether immunoreactivity in the basal regions was present within the initial axonal
segments of Purkinje cells or in climbing fibres since immunoelectron microscopy
revealed reaction product in both profiles. In certain Purkinje cells, labelling was
evident in the primary dendrites (fig 3 B and C). Between the stages of PDs 9 to 12,
Purkinje cell immunoreactivity was localised mainly in the basal regions of their
somata and was apposed to the cell membranes (fig 3 D and E). Immunoreactive
fibre-like profiles originating from the granular layer, encircling Purkinje cell somata
and encroaching on their proximal primary dendrites were observed at this stage.
These are presumably climbing fibres, primary Purkinje cell axons or recurrent axonal
collaterals (fig 3 F). CRF-R2 immunoreactive boutons were in greater abundance in
comparison to CRF-R1 (see fig. 2 E). Between PDs 15 to 25, Purkinje cell
immunoreactivity increased, mainly localised on somatic membranes and near the
initial axonal segment. Some Purkinje cells exhibited immunoreactivity in the
proximal dendritic regions (figs 3 G to I).
Electron microscopy
CRF-R1
Early CRF-R1 labelling was observed in Purkinje cell somata and in climbing fibre
terminals making synaptic contact with somatic spines of Purkinje cells. In Purkinje
cells, the reaction product was localised either within the trans Golgi network, in
small vesicles and in multivesicular bodies (figs 4 A and B). Importantly, somatic
spines of Purkinje cells were always free of label, whereas the climbing fibre terminals
making synaptic contact with spines expressed strong CRF-R1 immunoreactivity.
From PDs 6 to 11 onwards, Purkinje cell labelling increased in density and was
concentrated within the apical caps showing a clear gradient between the apical and
basal regions, evident at PD 12 (fig 4 C). Dendritic labelling became pronounced
from PD 15 onwards, with reaction product in the primary dendrites and their branches.
Dendritic spines were always free of label. Indeed, using pre- and postembedding
techniques, during the entire period of ontogeny, CRF-R1im in the Purkinje cell was
never found on or close to synaptic membrane specialisations (fig 4 D).
Climbing fibre terminals show intense immunoreactivity from PD 3 onwards. At this
stage, they made synaptic contacts on somatic spines on Purkinje cells (see fig 4 A).
Climbing fibre immunoreactivity persisted through to adulthood, when the
translocation of the somatic terminals to the stubby spines of Purkinje cell dendrites
had been established. Postembedding labelling showed that the reaction product was
localised in the cytoplasm and was not related to membranes (data not shown) (fig 5
A and B).

105
corticotropin releasing factor receptor types 1 and 2
Figure 7. Electron micrographs of CRF-R2 immunoreactivity in Purkinje cells at different postnatal stages.(A) an overview of the soma of a Purkinje cell at PD 6 showing immunogold particles closely associatedwith rough endoplasmic reticulum (arrowheads). (B) shows part of a soma of a Purkinje cell at PD 12.Evident are numerous immunoreactive climbing fibre terminals making synaptic contacts on somatic spines.(C) shows a region of a soma of a Purkinje cell at PD 15. Immunogold particles are cytoplasmicallylocalised and membrane bound (arrowheads). (D) shows a Purkinje cell soma at PD 25 with membranebound immunogold particles (arrowheads). (E) is a magnified view of the rectangular boxed area in (D)showing an immunogold particle in the perisynaptic region of a basket cell-Purkinje cell synapse (arrow-heads). (F) is a magnified view of the square boxed area in (D) showing an immunogold particle that isclosely attached to the presynaptic membrane specialisation. Bt, basket cell terminal; Cf, climbing fibre;Pnuc, Purkinje cell nucleus; Sp, Purkinje cell somatic spine. Scale bars: A-D, 2µm; E and F, 200nm.

106
Chapter 5
At early developmental stages, the external granular layer and the molecular layer
exhibited immunoreactivity in various cellular elements. From PD 6 differentiating
neurons resembling basket or stellate cells expressed CRF-R1im. Reaction product
was mainly localised in somata and dendrites and not in terminals. The pattern of
somatic and dendritic expression persisted into adulthood, with postembedding
labelling showing reaction product often membrane bound in the somata and dendrites
(fig 6).
Figure 8. Electron micrographs of CRF-R2 at Purkinje cell-parallel fibre contacts at PDs 12and 25. (A) an overview showing parallel fibres making synaptic contact with immunoreactivedendritic spines at PD 12. Note the reaction product contained within the dendritic branch aswell. (B) a magnified view of the boxed area in (A), showing scattered reaction productwithin the spine and in the region of the postsynaptic density (arrowheads). (C) an overviewof parallel fibre-Purkinje cell contacts at PD 25 showing immunoreactivity selectively localisedin parallel fibre terminals with the insert being a magnified view of the boxed area. (D) amagnified view of a similar contact showing that were either in the lumen of the parallel fibreterminal or (E), membrane bound in the perisynaptic region (arrowhead). Figures A to C areof pre-embedding immunostaining with D and E of post-embedding immunostaining. Pden,Purkinje cell dendrite; Pf, parallel fibre; Sp, Purkinje cell spine. Scale bars: A, C, 1 µm; B, D,E 200 nm

107
corticotropin releasing factor receptor types 1 and 2
CRF-R2
Within the first postnatal week, pre- and postembedding labelling showed that
Purkinje cell immunoreactivity was purely cytoplasmic, particularly in the region of
the rough endoplasmic reticulum. Membrane bound reaction product at this early
stage was not evident (fig 7 A). At PD 12, the reaction product was evident in somata,
while climbing fibre terminals making synaptic contact on somatic spines were also
immunoreactive (fig 7 B). Using postembedding labelling, PD 15 was shown to be
the first stage at which reaction product was localised bound to plasma membranes
(fig 7 C). At later stages, immunogold particles were found in close proximity to
terminals contacting Purkinje cell somata. These were either basket cell terminals or
recurrent axonal collaterals of Purkinje cells. The reaction product was always
perisynaptic. Membrane bound reaction product was also associated with
vacuolisation, presumably due to endocytotic receptor internalisation (figs 7 D – F).
In the molecular layer, CRF-R2im was present in Purkinje cell dendritic spines
contacted by parallel fibres. Labelling in spines was only expressed from PD 9 through
Figure 9. Electron micrographs ofCRF-R2 immunoreactivity inclimbing fibre profiles. (A) showstwo characteristic climbing fibreprofiles making synaptic contactson spines of Purkinje cells at PD 15with (B) showing similarimmunoreactive profiles at PD 25.Cf, climbing fibre; Sp, Purkinje celldendritic spine. Scale bars: 1µm.

108
Chapter 5
to PD 15. During this period, parallel fibre terminals were always free of
immunoreactivity (figs 8 A and B). Labelling in parallel fibre terminals was evident
at later stages. In the presynaptic terminal, immunogold particles were visualised,
close to the synaptic membrane specialisation (figs 8 C – E).
CRF-R2im was expressed in climbing fibres from early stages (see fig 7 B) through
to adulthood. It could be documented using pre- and postembedding labelling that
reaction product was often associated with vesicles and was rarely bound to
presynaptic membranes (fig 9).
DISCUSSION
The current study is the first to provide a detailed account of the localisation of CRF-
R1 and CRF-R2 at different developmental stages at the subcellular level. We have
largely concentrated on the developing Purkinje cells since they develop their dendritic
tree and afferent input during the first three postnatal weeks (Sotelo, 1978) and are
also considered the organising centre of the cerebellum (Hatten, 1999). The present
study addresses probing questions posed by Bishop et al. (2000) that can only be
addressed by using immunoelectron microscopy. Firstly, are these receptors localised
presynaptically or postsynaptically? Secondly, do cerebellar afferents express CRF
receptors? Such data is crucial for an integrative analysis of light microscopic
observations on CRF and urocortin localisation, an approach essential in the
elucidation of the role of the CRF system in cerebellar development and in motor
learning (Miyata et al., 1999).
Technical considerations
We have used three different immunocytochemical techniques to provide
complementary information on the ultrastructural localisation of CRF receptors. We
used antisera which, based on our controls, appear to offer a high degree of specificity
for the CRF receptors. Light microscopy was used to confirm the earlier findings
(Bishop et al., 2000) and to extend those findings to the developing cerebellum. Two
independent methods of immunocytochemistry at the electron microscopic level were
used to assess consistency of our results and the precise subcellular distribution of
the receptors. Pre-embedding immunocytochemistry using a peroxidase reaction is a
good indicator of the immunoreactivity of a profile (Lopez-Bendito et al., 2001).
However, the diffuse nature of the reaction product precludes the precise localisation
of receptors, especially G-protein coupled receptors which are expected to be
predominantly membrane bound. We have used postembedding immuno-
cytochemistry using immunogold particles since this is regarded as the method to

109
corticotropin releasing factor receptor types 1 and 2
provide the most accurate localisation (Lopez-Bendito et al., 2001). However, the
trade-off, compared to the peroxidase method, is a decreased intensity on labelling
and this is evident in this study.
CRF receptors in cerebellar afferents
From PDs 3-4, CRF-R1 and CRF-R2 are expressed in climbing fibres and mossy
fibre rosettes while only parallel fibres express CRF-R2. CRF and urocortin bind
with equal affinity to CRF-R1, however, urocortin exhibits a 40-fold greater affinity
for CRF-R2, suggesting that urocortin is the natural ligand for CRF-R2 (Vaughan et
al., 1995). In a previous study, we showed that both CRF and urocortin are expressed
in climbing fibres and mossy fibre rosettes whereas only urocortin is present in parallel
fibres (Swinny et al., 2002). Hence, CRF and urocortin are capable of coupling to
CRF-R1 and CRF-R2 respectively presynaptically. The presynaptic localisation of
these receptors is in accordance with physiological data from other brain regions
(Lawrence et al., 2002; Lewis et al., 2002). The reason and consequences for this
presynaptic interaction is unclear. However, presynaptic localisation of receptors,
acting as autoreceptors (Merighi, 2002), is not uncommon for neuropeptides. For
example, both CGRP and its receptors are localised in climbing fibres (A. Rosina,
personal communication). The presynaptic coupling to receptors, usually in concert
with other factors such as receptor activity modifying proteins (Born et al., 2002)
usually serves to control the release of the neuropeptide and/or other co-stored
modulators (Malcangio and Bowery, 1999; Khakh and Henderson, 2000). The
presynaptic localisation of CRF receptors suggests that they might play a role in the
release of CRF-like ligands.
In climbing fibres, CRF and urocortin probably adopt the roles of neuromodulators,
as opposed to the roles of classic neurotransmitters. While receptors of classical
neurotransmitters are generally concentrated directly beneath or in close proximity
of the postsynaptic membrane (Merighi, 2002) CRF-R1 or CRF-R2 were never
visualised on postsynaptic sites of climbing fibre contacts, namely the proximal stubby
spines. CRF-R1 was consistently visualised only in primary dendrites and their
branches. This potential neuromodulator role of CRF, as opposed to a neurotransmitter
function, is validated by studies of Bishop, (1990) who showed that CRF itself had
no excitatory action on Purkinje cell activity. Instead, CRF caused Purkinje cell
excitation by potentiating the effects of endogenous neurotransmitters like glutamate.
Cross-talk between different receptors is not uncommon (Bloch et al., 1999; Satake
et al., 2000). The interaction between the climbing fibre CRF and glutamatergic
systems would appear to be crucial since both CRF receptors (Miyata et al., 1999)
and metabotropic glutamate receptors (mGluR-1) (Ichise T et al., 2000) are essential

110
Chapter 5
for LTD. mGlu-R1 are also necessary for the elimination of supernumery climbing
fiber (Ichise et al., 2000), processes deemed crucial in the correct wiring and
functioning of the cerebellar circuitry.
CRF receptors in Purkinje neuron
The Purkinje cell showed significant CRF-R1 and CRF-R2 immunoreactivity from
early developmental stages through to adulthood. CRF-R1 immunoreactivity showed
a more dynamic developmental profile than CRF-R2, occurring at early stages in the
Purkinje cell somata, but shifting first into the apical caps, the precursor regions for
dendritic outgrowth (Altman and Bayer, 1997) and later into the primary dendrites
and dendritic branches. Also, CRF-R1 was not evidently bound to membranes,
however, it was always occurring in the cytoplasm of somata or dendrites. These
observations are rather unexpected since CRF-R1 belongs to the family of G-protein
coupled receptors which are usually membrane bound. However, this cytoplasmic
localisation is consonant with the report by Radulovic et al. (1998), who showed that
Purkinje neurons are the only cell type in the brain without membrane bound reaction
product. The scarcity of membrane bound CRF-R1 expression could be a reflection
of the degree of ligand-receptor coupling and subsequent cytoplasmic internalisation
that this receptor undergoes, so as to limit the effects of the ligands binding to it
(Bloch et al., 1999).
In contrast, CRF-R2 immunoreactivity resided predominantly in Purkinje cell somata,
particularly in the basal regions and axon hillocks. Only during the developmental
stages of PD 9 to 15 was reaction product evident in dendritic spines contacted by
parallel fibres. This is a critical period of Purkinje cell dendritic development thought
to be heavily influenced by parallel fibre activity (Altman and Bayer, 1997). It is
speculative whether urocortin contained in parallel fibres (Swinny et al., 2002) acts
directly on CRF-R2 contained in the spines during this time, perhaps playing a
collaborative role with other agents implicated synapse formation and dendritic
maturation. An additional piece of evidence for CRF-R2 being more active at the
synaptic level, in comparison to CRF-R1, is that it appears to be membrane bound,
especially in the Purkinje cell somata. The presence of CRF-R2, membrane bound
on the Purkinje cell soma, a region contacted by basket cell terminals suggests that
CRF-R2 could be coupled postsynaptically to urocortin released from basket cell
terminals or recurrent axonal collaterals of Purkinje cells.
The translocation of CRF-R1 into the developing dendrite infers an active role of the
receptor in the early outgrowth of Purkinje cells dendrites. Dendritic development is
governed by intrinsic patterns (Threadgill et al., 1997) and extrinsic cues (Whitford
et al., 2002). CRF, by coupling to CRF-R1 has been shown to mediate neurite

111
corticotropin releasing factor receptor types 1 and 2
outgrowth in catecholaminergic immortalized neuron (Cibelli et al., 2001). Further
investigations are needed to determine whether CRF, or particularly urocortin are
implicated in the early intrinsic pattern of Purkinje cell dendritic development. Since
CRF-R1 was mainly in the dendritic shafts and CRF-R2 in dendritic spines, these
two receptors, in concert, could play significant roles in modulating the development
of the functionally different domains of Purkinje cell dendrites.
In conclusion, the presynaptic expression of CRF receptors, together with co-localised
CRF and urocortin could play a role in differentially modulating the transmission of
sensory information contained within the cerebellar afferent systems. Secondly, in
light of the established developmental role of CRF receptors, their localisation within
different regions of the Purkinje cell dendritic tree alludes to their participation in
the intrinsic program regulating the outgrowth and maturation of Purkinje cell
dendrites. Functional studies are underway to test the above hypotheses.
REFERENCES
Altman, J. & Bayer, S.A. (1997) Development of the cerebellar system. In relation to its evolution,
structure and function. CRC Press.
Aguilera, G. Harwood, J.P. Wilson, J.X. Morell, J. Brown, J.H. Catt, K.J. (1983) Mechanisms of
action of corticotropin-releasing factor and other regulators of corticotropin release in rat pituitary
cells. J. Biol. Chem. 258, 8039-45.
Bilezikjian, L.M. Vale, W.W. (1983) Stimulation of adenosine 3',5'-monophosphate production by
growth hormone-releasing factor and its inhibition by somatostatin in anterior pituitary cells in vitro.
Endocrinology. 113, 1726-31.
Bishop, G.A. (1990) Neuromodulatory effects of corticotropin releasing factor on cerebellar Purkinje
cells: an in vivo study in the cat. Neuroscience. 39, 251-7.
Bishop, G.A. Seelandt, C.M. King, J.S. (2000) Cellular localization of corticotropin releasing factor
receptors in the adult mouse cerebellum. Neuroscience. 101, 1083-92.
Bloch, B. Dumartin, B. Bernard, V. (1999) In vivo regulation of intraneuronal trafficking of G protein-
coupled receptors for neurotransmitters. Trends Pharmacol Sci. 20, 315-9.
Born, W. Muff, R. Fischer, J.A. (2002) Functional interaction of G protein-coupled receptors of the
adrenomedullin peptide family with accessory receptor-activity-modifying proteins (RAMP). Microsc.
Res. Tech. 57, 14-22.
Chang, C.P. Pearse, R.V. 2nd. O’Connell, S. Rosenfeld, M.G. (1993) Identification of a seven
transmembrane helix receptor for corticotropin-releasing factor and sauvagine in mammalian brain.
Neuron. 11, 1187-95.
Chen, R. Lewis, K.A. Perrin, M.H. Vale, W.W. (1993) Expression cloning of a human corticotropin-
releasing-factor receptor. Proc. Natl. Acad. Sci. U S A. 90, 8967-71.
Chen, Y. Brunson, K.L. Muller, M.B. Cariaga, W. Baram, T.Z. (2000) Immunocytochemical distribution
of corticotropin-releasing hormone receptor type-1 (CRF(1))-like immunoreactivity in the mouse brain:
light microscopy analysis using an antibody directed against the C-terminus. J Comp Neurol May
8;420(3):305-23.

112
Chapter 5
Cibelli, G. Corsi, P. Diana, G. Vitiello, F. Thiel, G. (2001) Corticotropin-releasing factor triggers
neurite outgrowth of a catecholaminergic immortalised neuron via cAMP and MAP kinase signalling
pathways. Eur. J. Neurosci.13, 1339-48.
Giguere, V. Labrie, F. (1983) Additive effects of epinephrine and corticotropin-releasing factor (CRF)
on adrenocorticotropin release in rat anterior pituitary cells. Biochem. Biophys. Res. Commun. 110,
456-62.
Grigoriadis, D.E. De Souza, E.B. (1989) Heterogeneity between brain and pituitary corticotropin-
releasing factor receptors is due to differential glycosylation. Endocrinology. 25, 1877-88.
Hatten, M.E. (1999) Central nervous system neuronal migration. Annu. Rev. Neurosci. 22, 511-39.
Ichise, T. Kano, M. Hashimoto, K. Yanagihara, D. Nakao, K. Shigemoto, R. Katsuki, M. Aiba, A.
(2000) mGluR1 in cerebellar Purkinje cells essential for long-term depression, synapse elimination,
and motor coordination. Science. 288, 1832-5.
Khakh, B.S. Henderson, G. (2000) Modulation of fast synaptic transmission by presynaptic ligand-
gated cation channels. J. Auton. Nerv. Syst. 81, 110-21.
Lawrence, A.J. Krstew, E.V. Dautzenberg, F.M. Ruhmann, A. (2002) The highly selective CRF(2)
receptor antagonist K41498 binds to presynaptic CRF(2) receptors in rat brain. Br. J. Pharmacol. 136,
896-904.
Lewis, K. Li, C. Perrin, M.H. Blount, A. Kunitake, K. Donaldson, C. Vaughan, J. Reyes, T.M. Gulyas,
J. Fischer, W. Bilezikjian, L. Rivier, J. Sawchenko, P.E. Vale, W.W. (2001) Identification of urocortin
III, an additional member of the corticotropin-releasing factor (CRF) family with high affinity for the
CRF2 receptor. Proc. Natl. Acad. Sci. U S A. 98, 7570-5.
Lewis, M.W. Hermann, G.E. Rogers, R.C. Travagli, R.A. (2002) In vitro and in vivo analysis of the
effects of corticotropin releasing factor on rat dorsal vagal complex. J. Physiol. 543, 135-46.
Lopez-Bendito, G. Shigemoto, R. Lujan, R. Juiz, J.M. (2001) Developmental changes in the localisation
of the mGluR1alpha subtype of metabotropic glutamate receptors in Purkinje cells. Neuroscience.
105, 413-29.
Lovenberg, T.W. Liaw, C.W. Grigoriadis, D.E. Clevenger, W. Chalmers, D.T. De Souza, E.B. Oltersdorf,
T. (1995) Cloning and characterization of a functionally distinct corticotropin-releasing factor receptor
subtype from rat brain. Proc. Natl. Acad. Sci. U S A. 92, 836-40.
Malcangio, M. Bowery, N.G. (1999) Peptide autoreceptors: does an autoreceptor for substance P exist?
Trends. Pharmacol. Sci. 20, 405-7.
Merighi, A. (2002) Costorage and coexistence of neuropeptides in the mammalian CNS. Prog.
Neurobiol. 66, 161-90.
Miyata, M. Okada, D. Hashimoto, K. Kano, M. Ito, M. (1999) Corticotropin-releasing factor plays a
permissive role in cerebellar long-term depression. Neuron. 22, 763-75.
Palay, S. Chan-Palay, V. (1974) Cerebellar cortex: Cytology and organisation. Springer, New York.
Palkovits, M. Leranth, C. Gorcs, T. Young, W.S. 1987 Corticotropin-releasing factor in the
olivocerebellar tract of rats: demonstration by light- and electron-microscopic
immunohistochemistry and in situ hybridization histochemistry. Proc. Natl. Acad. Sci. U S A. 84,
3911-5.
Petralia, R.S. Zhao, H.M. Wang, Y.X. Wenthold, R.J. (1998) Variations in the tangential distribution of
postsynaptic glutamate receptors in Purkinje cell parallel and climbing fiber synapses during
development. Neuropharmacology. 37, 1321-34.
Perrin, M.H. Donaldson, C.J. Chen, R. Lewis, K.A. Vale, W.W. (1993) Cloning and functional expression
of a rat brain corticotropin releasing factor (CRF) receptor. Endocrinology. 133, 3058-61.

113
corticotropin releasing factor receptor types 1 and 2
Perrin, M. Donaldson, C. Chen, R. Blount, A. Berggren, T. Bilezikjian, L. Sawchenko, P. Vale, W.
(1995) Identification of a second corticotropin-releasing factor receptor gene and characterization of
a cDNA expressed in heart. Proc. Natl. Acad. Sci. U S A. 92, 2969-73.
Radulovic, J. Sydow, S. Spiess, J. (1998) Characterization of native corticotropin-releasing factor
receptor type 1 (CRFR1) in the rat and mouse central nervous system. J. Neurosci. Res. 54, 507-21.
Reyes, T.M. Lewis, K. Perrin, M.H. Kunitake, K.S. Vaughan, J. Arias, C.A. Hogenesch, J.B. Gulyas,
J. Rivier, J. Vale, W.W. Sawchenko, P.E. (2001) Urocortin II: a member of the corticotropin-releasing
factor (CRF) neuropeptide family that is selectively bound by type 2 CRF receptors. Proc. Natl. Acad.
Sci. U S A. 98, 2843-8.
Satake, S. Saitow, F. Yamada, J. Konishi, S. (2000) Synaptic activation of AMPA receptors inhibits
GABA release from cerebellar interneurons. Nat. Neurosci. 3, 551-8.
Shi, F. Nomden, A. Oberle, V. Engberts, J.B. Hoekstra, D. (2001) Efficient cationic lipid-mediated
delivery of antisense oligonucleotides into eukaryotic cells: down-regulation of the corticotropin-
releasing factor receptor. Nucleic Acids Res. 29, 2079-87.
Sotelo, C. (1978) Purkinje cell ontogeny: formation and maintenance of spines. Prog. Brain. Res. 48,
149-70.
Swinny, J. Kalicharan, D. Gramsbergen, A. van der Want, J. (2002) The localisation of urocortin in
the adult rat cerebellum: a light and electron microscopic study. Neuroscience. 14, 891-903.
Threadgill, R. Bobb, K. Ghosh, A. (1997) Regulation of dendritic growth and remodeling by Rho,
Rac, and Cdc42. Neuron. 19, 625-34.
Towbin, H. Staehelin, T. Gordon, J. (1979) Electrophoretic transfer of proteins from polyacrylamide
gels to nitrocellulose sheets: procedure and some applications. Proc. Natl. Acad. Sci. 76, 4350-4.
Vale, W. Spiess, J. Rivier, C. Rivier, J. (1981) Characterization of a 41-residue ovine hypothalamic
peptide that stimulates secretion of corticotropin and beta-endorphin. Science. 213, 1394-7.
Van den Pol, A.N. Gorcs, T. (1986) Synaptic relationships between neurons containing vasopressin,
gastrin-releasing peptide, vasoactive intestinal polypeptide, and glutamate decarboxylase
immunoreactivity in the suprachiasmatic nucleus: dual ultrastructural immunocytochemistry with gold-
substituted silver peroxidase. J. Comp. Neurol. 252, 507-21.
Van Pett, K. Viau, V. Bittencourt, J.C. Chan, R.K. Li, H.Y. Arias, C. Prins, G.S. Perrin, M. Vale, W.
Sawchenko, P.E. (2000) Distribution of mRNAs encoding CRF receptors in brain and pituitary of rat
and mouse. J. Comp. Neurol. 428, 191-212.
Vaughan, J. Donaldson, C. Bittencourt, J. Perrin, M.H. Lewis, K. Sutton, S. Chan, R. Turnbull, A.V.
Lovejoy, D. Rivier, C. et al. (1995) Urocortin, a mammalian neuropeptide related to fish urotensin I
and to corticotropin-releasing factor. Nature. 378, 287-92.
Vita, N. Laurent, P. Lefort, S. Chalon, P. Lelias, J.M. Kaghad, M. Le Fur, G. Caput, D. Ferrara, P.
(1993) Primary structure and functional expression of mouse pituitary and human brain corticotrophin
releasing factor receptors. FEBS. Lett. 335, 1-5.
Voogd, J. Glickstein, M. (1998) The anatomy of the cerebellum. Trends Neurosci. 21, 370-5.
Wang, H.L. Tsai, L.Y. Lee, E.H. (2000) Corticotropin-releasing factor produces a protein synthesis—
dependent long-lasting potentiation in dentate gyrus neurons. J. Neurophysiol. 83, 343-9.
Whitford, K.L. Dijkhuizen, P. Polleux, F. Ghosh, A. (2002) Molecular control of cortical dendrite
development. Annu Rev Neurosci. 25, 127-49.
Yi H, Leunissen, J. Shi, G. Gutekunst, C. Hersch, S. (2001) A novel procedure for pre-embedding
double immunogold-silver labelling at the ultrastructural level. J. Histochem. Cytochem. 49, 279¯284.

114

115
CHAPTER 6
Corticotropin releasing factor and Urocortin modulatePurkinje cell dendritic outgrowth and differentiation in vitro
J. D. Swinny1, F. Metzger2 , J. IJkema-Paassen2, A. Gramsbergen2,
J.J.L. van der Want1
1Laboratory for Cell Biology and Electron Microscopy,2 Department of Medical Physiology, Graduate School of Behavioural and
Cognitive, Neuroscience, University of Groningen,
The Netherlands
Submitted: Mol. Cell. Neuroscience

116
Chapter 6
ABSTRACT
The precise outgrowth and arborisation of dendrites is crucial for their function as
integrators of signals relayed from axons and hence, the functioning of the brain.
Proper dendritic differentiation is particularly resonant for Purkinje cells since the
intrinsic activity of this cell-type is governed by functionally distinct regions of its
dendritic tree. The initial phase of dendritic development is governed by intrinsic
patterns. However, activity dependent mechanisms, under the influence of electrical
signalling and trophic factors account for the most active period of dendritogenesis.
A yet unexplored trophic modulator of Purkinje cell dendritic development is
corticotropin releasing factor (CRF) and family member, urocortin, both localised in
climbing fibres. Here we use organotypic cerebellar slice cultures to investigate the
roles of CRF and urocortin on Purkinje cell dendritic development. Intermittent
exposure (12 hours per day for 10 days in vitro) of CRF and urocortin induced
significantly more dendritic outgrowth (45 % and 70 % respectively) and elongation
(25 % and 15 % respectively) compared with untreated cells. Conversely, constant
exposure to CRF and urocortin significantly inhibited dendritic outgrowth. Using
specific antagonists against the CRF receptors, it could be demonstrated that both
receptors are involved in the induction of dendritic outgrowth whereas CRF receptor
1 modulates CRF-induced elongation of dendrites. Furthermore, the trophic effects
of CRF and urocortin are mediated by the protein kinase A and the mitogen activating
protein kinase pathways. The study shows unequivocally that CRF and urocortin are
important regulators of dendritic development. However, their effects are dependent
upon their degree of expression that in turn, influences the functional status of their
receptors. The current data offer insights, and possible therapeutic possibilities for
growing number of pathologies that show perturbations between CRF peptide
expression and their receptors.

117
corticotropin releasing factor and Urocortin
INTRODUCTION
Dendrites play a crucial role in the integration and computation of signals relayed
from axons and consequently, the activity of neurons (Magee, 2000; Reyes, 2001).
The diversity of dendritic arbor types within the CNS underscores the significance
of dendritic morphology to the specialised functioning of neurons (Vetter et al., 2001).
Since the spatial patterning of dendritic arborisation is integral to neuronal function,
and hence proper brain functioning, an understanding of the processes and factors
involved in the precise growth and differentiation of dendrites is crucial. This will
also facilitate more directed interventions of malformations and pathologies of a
developmental origin such as spinocerebellar ataxia (Yang et al., 2000; Jeong et al.,
2000).
The precise orientation and differentiation of the most elaborate dendritic trees in the
brain, namely those of Purkinje cells is of particular importance. This is highlighted
by the fact that the functionally distinct regions of its dendritic tree (Bravin et al.,
1999) serve specific roles in the intrinsic activity of Purkinje cells (Womack and
Khodakhah, 2002). Also, reciprocal trophic interactions between the Purkinje cell
dendrites and their afferents, climbing fibres and parallel fibres (Strata et al., 1997)
ensure that the afferents selectively innervate distinct regions on the dendritic tree.
Indeed, parallel fibres which convey sensory information from the mossy fibre system
form synapses exclusively on the distal thorny dendritic spines while axons of the
inferior olivary complex, namely climbing fibres, which convey signals from higher
brain centres, selectively innervate the more proximal stubby dendritic spines
(Sugihara et al., 2000). Since the Purkinje cell is considered the organising centre of
the cerebellar cortex, the formation and differentiation of Purkinje cell dendrites is
central for the integration and computation of its diverse inputs, allowing the
cerebellum to function as the centre for motor coordination.
During development, intrinsic programs contribute to the initial acquisition of a
polarised form, with distinct axons and dendrites (Dotti et al., 1988; Zhang et al.,
2002) and also allowing for the distinctive morphologies of different neuronal types
(Cannon et al., 1999). This holds true for Purkinje cells as well (Soha and Herrup,
1995). However, activity dependent mechanisms, particularly the signalling from
afferents accounts for the most active period of dendritic growth (Zhang and Poo,
2001; Wong and Ghosh, 2002). The importance of activity-dependent mechanisms
resonates most soundly for Purkinje cells, since the greatest amount of dendritic
growth occurs during the most active period of synaptogenesis, which in the rat,
occurs from the second postnatal week onwards. In relation to electrical signalling
also trophic factors play a dynamic role in either inducing or repressing dendritic

118
Chapter 6
growth (McAllister, 2001). A yet unexplored possible trophic agent of Purkinje cell
development is corticotropin releasing factor (CRF) (Vale et al., 1981).
In the cerebellum, CRF is selectively localised in climbing and mossy fibres (Palkovitz
et al., 1987). In adulthood, CRF released from climbing fibres is crucial for the
induction of long term depression at the parallel fibre-Purkinje cell synapse, a type
of synaptic plasticity considered to be the cellular correlate of motor learning.
Interestingly, cerebellar CRF (Bishop and King, 1999) and the receptors to which it
binds (Chan et al., 1993) are expressed already at early stages of cerebellar
development, prior to any functional synapses being formed. CRF augments the effects
of glutamate (Bishop, 1990), a neurotransmitter directly implicated in activity
dependent dendritic development (Metzger et al., 1998), suggesting that climbing
fibre released CRF might first function as a trophic agent. We have recently shown
that a newer member of the CRF family of peptides, urocortin (Vaughan et al., 1995)
is also expressed in the rat cerebellum (Swinny et al., 2002). The major sites of
expression were the Purkinje cells, and climbing and parallel fibres. Our earlier
observations show that urocortin is also expressed in the developing rat cerebellum
suggesting that both CRF and urocortin could play a trophic role in Purkinje cell
development. Both CRF and urocortin act via two G-protein coupled receptors, namely
CRF receptor 1 (CRF-R1) and CRF receptor 2 (CRF-R2) with urocortin having a
greater binding affinity for CRF-R2. In the Purkinje cell, CRF-R1 is localised on the
dendrites whilst CRF-R2 is restricted to the somata (Bishop et al., 2000; Bishop and
King, 2002; Swinny et al.,2003). Since CRF and urocortin couple to the two CRF
receptors with different binding affinities (Vaughan et al., 1995), these two peptides
might subserve different trophic roles. To explore this postulate, we have used
organotypic slice cultures of rat cerebellum to study the effects of CRF and urocortin
on Purkinje cell dendritic differentiation in vitro.
METHODS
Materials
Human and rat CRF, rat urocortin and alpha-helical CRF (a non-selective CRF receptor
antagonist) were obtained from Sigma (Zwijndrecht, The Netherlands). KT 5720 (a
selective PKA antagonist) and PD98059 (a selective MAPK antagonist) were obtained
from Calbiochem (Breda, The Netherlands). Antalarmin was a generous gift of KC
Rice and H Habib (NIH). Chemicals were dissolved in a 50:50 mixture of DMSO and
ethanol. Minimal essential medium (MEM), Hank’s balanced salt solution (HBSS),
glutamax I and horse serum were purchased from Invitrogen (Breda, The Netherlands).

119
corticotropin releasing factor and Urocortin
Organotypic slice cultures of rat cerebellum
Approval to conduct the study was obtained from the Ethics Committee on Animal
Experimentation, University of Groningen. All efforts were made to minimise the
number of animals used and their suffering. The preparation of cerebellar slices was
performed according to a protocol modified from Metzger et al. (2000). In brief, 8-
day-old Black-hooded Lister rat pups were decapitated and the brain aseptically
removed. The cerebellum was rapidly dissected in ice-cold preparation medium (MEM
containing 2 mM glutamax I, pH 7.3) and the meninges were carefully removed.
Sagittal slices of 400 µm were cut using a McIllwain tissue cutter, separated with
fine forceps and transferred onto humidified transparent membranes (Millicell-CM,
Millipore). They were cultured on a liquid layer of MEM containing HBSS (25%),
horse serum (25%), glutamax I (2 mM) and NaHCO3 (5 mM), HEPES (10 mM),
pH 7.3, in a humidified atmosphere with 5% CO2 at 37 °C.
CRF receptor desensitization is time-dependent, with ~ 50% down-regulation
occurring within 3 hours in Y79 cells (Dautzenberg et al., 2002). We therefore explored
the effects of intermittent exposure or constant exposure of CRF and urocortin on
Purkinje cell dendritic development. Thus, slices were either exposed to
pharmacological agents for 12 hours per day for 10 days in vitro (DIV) or continuously
for 10 DIV.
Immunocytochemistry
Slice cultures were fixed after 10 DIV in phosphate buffer (100 mM) containing 4%
paraformaldehyde. All further steps were performed in 100 mM phosphate buffer
(PB), pH 7.3. The slices were permeabilised and non-specific binding sites blocked
with Triton X-100 (0.3%) and normal goat serum (2%). Monoclonal antibody against
calbindin D-28K (Sigma) was applied (1: 1000) and the slices incubated at 4 °C
overnight. After washing, the slices were incubated with goat-antimouse Alexa Fluor
633 (Molecular Probes), (1: 500) for 2 hours. The slices were washed again and
mounted with Mowiol (Calbiochem). Slices were investigated by fluorescence
microscopy.
Quantitative morphological analysis of Purkinje cell dendritic trees
Purkinje cells with little overlap with neighbouring cells were selected in order to
fully quantify the entire dendritic trees as described (Metzger and Kapfhammer, 2000;
Schrenk et al., 2002). Purkinje cells were photographed with a high-resolution CCD
camera at a 200x magnification. The images were analysed using a morphometric

120
Chapter 6
program supplied by analySIS Soft Imaging System (Munster, Germany). The
following parameters were determined from micrographs according to the definitions
of Metzger and Kapfhammer (2000) and Schrenk et al. (2002): (i) the number of
primary dendrites; (ii) the number of dendritic branching points; (iii) the length of the
longest dendrite (i.e. distance between the cell body and the most distal dendritic
ending); (iv) the total dendritic tree area by connecting the ends of all terminal dendritic
tips of a single Purkinje cell including the soma with straight lines. Additionally, the
branching density, the area and the number of branching points per dendrite were
calculated for each Purkinje cell from the obtained raw data.
Calculations and statistics
Individual experiments were performed with 5-6 cerebellar slices per sample and
repeated 2-3 times using matched controls, and the obtained data were pooled. From
each sample, 15-40 Purkinje cells were evaluated. Results are expressed in the text
and graphically as mean ± standard error of the mean (SEM). The statistical
significance of differences in parameters was assessed by parametric one-way analysis
of variance (ANOVA) followed by Dunnett’s multiple comparison test using SPSS
statistical software (Chicago).
RESULTS
Intermittent exposure of slices to CRF and urocortin stimulate Purkinje cell
dendritic outgrowth and elongation
Since CRF receptors are rapidly down-regulated (Hauger et al., 2000) by G-protein
receptor kinase 3 (Dautzenberg et al., 2001; 2002), we investigated the effects of
intermittent and continuous exposure of CRF and urocortin, and hence, different
degrees of CRF receptor desensitisation, on Purkinje cell dendritic development. We
therefore treated cerebellar slice cultures for each 12 hours per day with CRF or
urocortin. Generally, treatment with CRF, urocortin or any antagonist used in this
study had no discernable effect on Purkinje cell survival (data not shown). Intermittent
exposure of cerebellar slices to CRF and urocortin had overall stimulatory effects on
Purkinje cell dendritic growth and development. At 4 DIV, cells from control slices
exhibited numerous immature or unbranched dendritic processes. In comparison, CRF
and urocortin treated cells had longer primary dendrites that branched more extensively.
At 10 DIV, both control and treated cells showed the characteristic morphology of
Purkinje cells. However, cells treated with CRF and urocortin had, on average more
primary dendrites, which were longer and branched more (figure 1). The effects of
picomolar (pM), nanomolar (nM) and micromolar (µM) concentrations of CRF and

121
corticotropin releasing factor and Urocortin
Figure 1. Comparative morphologies of calbindin-immunostained Purkinje cells from slicestreated with the CRF and urocortin over 4 and 10 DIV daily for 12 hours. (A) a cell fromcontrol experiments at 4 DIV exhibiting relatively short unbranched primary dendrites. (Band C) in comparison, CRF and urocortin (1 nM) treated cells had longer primary dendriteswith apparently more branches (D) shows the characteristic morphology of a Purkinje cellafter 10 DIV. (E and F) show cells treated with CRF and urocortin respectively at a concentra-tion of 1 nM. In both treatment groups, more primary dendrites are evident with these beingmore elongated compared to controls. Scale bar = 50 µm.
Figure 2. Quantitative morphological analysis of Purkinje cell dendritic trees showing themeans and SEM following treatment of cerebellar slices with different concentrations of CRF
and urocortin for 12 hours per day for 10 DIV.*** P < 0.01 versus respective controls.

122
Chapter 6
urocortin on Purkinje cell dendritic growth and differentiation were quantified. Both
CRF and urocortin caused a concentration dependent increase in the number of primary
dendrites with 1 nM concentration resulting in the most significant amount of primary
dendritic outgrowth (fig. 2 A). At 10 DIV, urocortin, at 1 nM concentration was the
most potent in inducing dendritic outgrowth (mean of 5.3 vs 3.1 primary dendrites
for control). One nM concentrations of CRF and urocortin also resulted in the most
significant increases of branching points per Purkinje cell (fig. 2 B). However, there
were no significant differences between branching points per primary dendrite
suggesting that both factors had no effect on the induction of new branching (fig. 2
C). Compared to controls, CRF and urocortin treated cells had significantly longer
dendrites indicating that CRF at 1 nM was most potent in inducing dendritic elongation
(mean length 174 vs 139 µm in controls) (fig 2 D). Furthermore, CRF (1pM and
1nM) caused a statistical significant increase in total dendritic area (fig 2 E) whereas
cells treated with urocortin (1 nM and 1 µM) showed significantly lower areas per
dendrite (fig. 2 F).
An apparent discrepancy occurred between the effects of CRF and urocortin on the
dendritic branching and area (revealing the branching density of the dendritic tree)
when calculated per Purkinje cell or single dendrite (see fig. 2). We therefore performed
a linear correlation analysis to correlate the branching density either per cell or per
single dendrite. The analysis per Purkinje cell revealed very poor correlations under
control conditions as well as after CRF or urocortin treatment (fig. 3A, table 1) whereas
the calculation per single dendrite greatly improved this correlation (fig. 3B, table 1).
This supports an idea that dendritic branching is a stoichiometric process driven by
factors influencing dendritic lengthening, in this case, CRF. The correlation analyses
further supported the calculation per dendrite as relevant because the apparent number
of primary dendrites (y-intersects in fig. 3) only in this analysis revealed values
qualitatively fitting to our observations since we observed Purkinje cells always bearing
one to seven primary dendrites in our cultures (table 1). Using this analysis we observed
a highly significant increase in the branching density of 55 % after CRF treatment as
compared to controls whereas urocortin did not reveal any change in branching density
but showed a tendency to induce more primary dendrites although this difference did
not reach significance (table 1). These data indicate that CRF and urocortin might
exert different effects on Purkinje cell dendritic development with CRF being more
active in already-formed dendrites, influencing their further differentiation while
urocortin initiates dendritic outgrowth. For clarity, in the rest of the manuscript we
therefore only show data calculated per single dendrite.

123
corticotropin releasing factor and Urocortin
Table 1: Linear regression analysis of morphological parameters. Correlations betweendendritic area and branching points (see fig. 3) were calculated per cell or per primarydendrite. Correlations were fitted to a straight line equation revealing a slope (branchingdensity) and the y-intercept (primary dendrites emerging from the cell body where dendritearea is zero). ** p<0.01 as assessed by unpaired t-test. R indicates quality of fit.
Treatment Slope Y-intercept R
Evaluation per cellControl 0.85 ± 0.18 16.0 ± 4.3 0.366CRF 1 nM 1.07 ± 0.26 30.1 ± 8.0 0.494Urocortin 1 nM 1.01 ± 0.39 30.1 ± 9.6 0.319
Evaluation per dendriteControl 1.35 ± 0.13 1.6 ± 1.1 0.663CRF 1 nM 2.09 ± 0.21** 0.3 ± 1.6 0.802Urocortin 1 nM 1.45 ± 0.23 4.5 ± 1.4 0.636
Figure 3 (for colorinfor-mation: see page 149)Linear regression analysis ofthe correlation betweendendritic area and branch-ing. Data for the dendritictree area and the numberof branching points wereeither calculated either perPurkinje cell (A) or per sin-gle dendrite (B) and theobtained data fitted to astraight line equation(dashed lines) revealingslope (branching density)and y-intercept (primarydendrites/cell). For numericdata, see table 1.

124
Chapter 6
In order to gauge how soon the stimulatory effects of CRF and urocortin are manifested,
we quantified the morphological parameters at 4 DIV. Cells treated with CRF and
urocortin had on average more primary dendrites although these differences were not
significantly different to control values (fig. 4 A). Application of both CRF and
urocortin resulted in cells having significantly longer primary dendrites (129 and 114
respectively vs 79 µm in controls) (fig. 4 B). At 4 DIV, CRF-treated cells had
significantly more branching points per dendrite (fig. 4 C) and a greater area per
dendrite (fig. 4 D). Compared to the 10 DIV cultures, these data suggest that acute
exposure of CRF and urocortin in vitro has a more pronounced effect on dendritic
elongation whilst long-term exposure is required to maintain dendrites.
Constant exposure of CRF and urocortin to cerebellar slices inhibits Purkinje
cell dendritic development
Instead of exposing slices to CRF or urocortin for 12 hours per day, slices were
continuously exposed to the peptides at 1 µM concentration for 10 DIV in order to
mimic a state of continuous and complete CRF receptor down-regulation. The constant
exposure of CRF and urocortin resulted in Purkinje cells showing significantly less
growth than cells from control slices. Cells had shorter primary dendrites and less
Figure 4. Comparison of quantitative data for short-term vs. long-term treatments of slicecultures with CRF or urocortin. Cerebellar slices were treated with CRF or urocortin (each 1nM) for 12 hours per day for 4 or 10 DIV and Purkinje cell dendrites morphologically ana-lysed. * P < 0.05; ** P < 0.01; *** P < 0.001 versus respective control.

125
corticotropin releasing factor and Urocortin
Figure 5. Fluorescence micrographs of Purkinje cells from a control slice and slices treatedwith 1 µM of CRF and urocortin continuously for 10 DIV. Purkinje cells from treated sliceshave relatively shorter primary dendrites and less dendritic branching particularly in urocortintreated slices. Scale bar = 50 µm.
Figure 6. Quantitative morphological analysis of Purkinje cell dendritic trees showing themeans and SEM following treatment of cerebellar slices with CRF and urocortin 1 µM continu-ously for 10 DIV. Continuous exposure of cerebellar slices to CRF and urocortin, had, ingeneral, inhibitory effects on Purkinje cell growth. * P < 0.05; *** P < 0.001 versus control.
dendritic branching, with cells from urocortin effecting the most significant amount
of inhibition (fig. 5).

126
Chapter 6
The continuous exposure of slices to 1µM urocortin had a potent inhibitory effect on
all morphological parameters evaluated. The number of primary dendrites growing
out was reduced by urocortin (2.0 vs 3.4 for control; fig. 6 A). Dendrites were
significantly shorter when treated with CRF and urocortin (127 and 107 µm
respectively vs 146 µm for control (fig. 6 B). Compared to control values, 1 µM
concentrations of CRF and urocortin also significantly decreased the total number of
dendritic branches per primary dendrite (fig. 6 C) with urocortin significantly
decreasing the area per dendrite (fig. 6 D). These data indicate that CRF receptors
were down regulated by continuous peptide application and that CRF receptor
signalling is necessary for proper dendrite outgrowth in cerebellar slice cultures.
Blockade of CRF receptors and secondary messenger cascades
CRF receptor antagonists
The specificity of the CRF and urocortin effects on Purkinje cell dendrite outgrowth
was tested using receptor-specific blockers. a-helical-CRF (a-h-CRF), which inhibits
both known CRF receptors (CRF-R1 and CRF-R2) ( Menzaghi et al., 1994) was used
as a non-specific CRF receptor antagonist. Antalarmin (Webster et al., 1994) was
used as a selective CRF-R1 antagonist.
A-h-CRF alone did not have any significant effects on Purkinje cell dendritic growth
and differentiation when compared with cells from control slices (fig. 7 A). Cells
from slices treated with CRF and urocortin (1 nM) in the presence of a-h-CRF had
less primary dendrites, which were shorter and branched less, when compared with
cells from slices treated with CRF and urocortin alone (figs. 7 B and C. compare with
fig. 1). In the presence of a-h-CRF, the CRF and urocortin induced outgrowth of
primary dendrites was attenuated (3.4 and 3.1 for CRF and urocortin respectively).
Although a-h-CRF significantly reduced the total number of branch points per cell
(data not shown), there was no significant reduction in branching points per dendrite
(fig. 8 B). A-h-CRF significantly reduced the dendritic length of CRF-treated cells
(174 µm reduced to 150 µm). However, the area per dendrite was unaffected by a-h-
CRF (fig. 8 C).
The CRF-R1-specific antagonist antalarmin alone did not alter normal Purkinje cell
dendritic development in vitro when compared with cells from untreated slices (fig. 7
D. see fig 1A). In contrast, the CRF and urocortin-induced dendritic lengthening and
branching effects were markedly inhibited by antalarmin (fig 7 E and F). The addition
of antalarmin to CRF and urocortin treated slice cultures resulted in cells having

127
corticotropin releasing factor and Urocortin
Figure 7. Comparative morphologies of Purkinje cells from slices treated with the CRF-R1and CRF-R2 antagonist a-h-CRF alone, antalarmin, the specific CRF-R1 antagonist alone orwith CRF and urocortin in the presence of a-h-CRF and antalarmin for 12 hours per day for12 DIV. (A) cells from slices treated with a-h-CRF resembled cells from untreated slices (seefig. 1 A). (B and C) however, the presence of a-h-CRF in CRF and urocortin treated slicesattenuated the stimulatory effects of CRF and urocortin (see also fig. 1 B and C). (D-F)antalarmin significantly reduced the dendritic length of CRF and urocortin treated cells. Scalebar = 50 µm.
significantly less primary dendrites (2.7 and 3.9 respectively) compared with those
treated CRF and urocortin alone (fig. 8 A). Antalarmin significantly reduced the total
number of dendritic branches of CRF and urocortin-treated cells (65.7 and 68.5
respectively reduced to 38 and 40 respectively). However, the branching points per
dendrite of CRF and urocortin treated cells were unaffected by antalarmin treatment
(fig. 8 B). The mean dendritic length of CRF-treated cells (174 µm to 151 µm) as
well as the mean total dendritic area (32607 µm2 to 18530 µm2) was significantly
reduced by antalarmin. However, the area per primary dendrite was unaffected by
antalarmin alone (fig. 8 C) although it abolished the urocortin effect.
CRF receptor secondary messenger cascades
Both CRF receptors couple to G proteins resulting in cAMP stimulation and protein
kinase A (PKA) activation (Chang et al., 1993; De Souza, 1995). However, CRF and
urocortin have been shown to activate diverse signalling cascades, such as the mitogen

128
Chapter 6
activating protein kinase (MAPK) pathways (Rossant et al., 1999; Blank et al., 2003).
We used the specific PKA inhibitor, KT 5720 (Kase et al., 1987) and the specific
MAPK inhibitor, PD 98059 (Alessi et al., 1995) to assess the roles of the PKA and
MAPK pathways in the CRF and urocortin-induced dendritic differentiation. The
culturing of slices in the presence of KT 5720 and PD 98059 alone did not significantly
alter the normal growth of Purkinje cells when compared with cells from untreated
slices. In the presence of KT 5720, the stimulatory effects of CRF 1 nM and urocortin
1 nM on Purkinje cell dendritic outgrowth was significantly reduced (4.5 and 5.3
respectively reduced to 3.4 and 4.0 mean number of primary dendrites per cell
respectively; fig.8A). PD 98059 was more potent in inhibiting the CRF and urocortin
Figure 8. Quantitative morphological analysis of Purkinje cell dendritic trees showing themeans and SEM following treatment of cerebellar slices with a-h-CRF (CRF-R1 and 2 antago-nist), antalarmin (CRF-R1 antagonist), KT 5720 (PKA antagonist) and PD 98059 (MAPK an-tagonist) (1 µM) alone or with CRF and urocortin (1 nM) for 12 hours per day for 10 DIV. Thedata represents the means and SEM of parameters assessed. (A) a-h-CRF significantly at-tenuated the urocortin-induced outgrowth of primary dendrites, to a greater degree thanantalarmin. PD 98059 was most potent in reversing the CRF and urocortin-induced dendriticoutgrowth. (B and C) following treatment with the antagonists, the parameters of branchingpoints per dendrite and area per dendrite were comparable to control values. ** P < 0.01;*** P< 0.001.

129
corticotropin releasing factor and Urocortin
induced dendritic outgrowth (reduced to 2.6 and 2.9 mean number of primary dendrites
per cell respectively). Whilst the total number of dendritic branches per cell, dendritic
length and total dendritic areas of CRF and urocortin treated cells were all significantly
reduced to levels of untreated controls, the branching points per primary dendrite and
area per dendrite were not significantly altered by the treatment of slices with CRF
and urocortin concomitantly with KT 5720 and PD 98059 (figs. 8 B and C). In fact,
the inhibitory effect of urocortin on the dendritic area per primary dendrite was
abolished in the presence of either K/ 5720 or PD 98059 (fig.8C).
DISCUSSION
In the current study, we have used a well-established model of Purkinje cell dendritic
development (Metzger and Kapfhammer, 2000; Schrenk et al., 2002; Gundlfinger et
al., 2003) to show that CRF and urocortin dynamically effect dendritic outgrowth and
elongation. The constant exposure of cerebellar slices to CRF and urocortin, which,
one could reasonably expect to lead to sustained receptor desensitisation (Hauger et
al., 2000), resulted in grossly under-developed dendritic trees. Conversely, intermittent
exposure of these peptides stimulated Purkinje cell dendritic outgrowth and elongation,
indicating that the expression of CRF-like peptides and the functional status of the
CRF receptor system are crucial for normal dendritic development. CRF and urocortin
most likely modulate different phases of the dendritic developmental process.
Induction of dendritic outgrowth
Based on the quantitative morphological assessment, urocortin is a potent inducer of
dendritic outgrowth, more so than CRF. Indeed, urocortin caused a 70% increase in
number of primary dendrites compared with control values, and 17% increase when
compared with cells from slices treated with CRF alone (see figure 2). The effects of
urocortin appear to predominate in the initial phases of dendritic development, namely
the initial outgrowth, since its effects on parameters concerned with later dendritic
differentiation, such as elongation and branching were always less than that of CRF.
Both CRF and urocortin are expressed in climbing fibres (Swinny et al., 2002) which
form synapses on Purkinje cell somata as early as embryonic day 19 (Morara et al.,
2001) and then translocate to the proximal dendritic regions (Scelfo et al., 2003).
CRF-like peptides act via two G-protein coupled receptors with urocortin showing
selectivity for CRF-R2. We have recently shown that, postnatally, CRF-R2 is restricted
to the Purkinje cell somata, while CRF-R1 enjoys a more dynamic ontogenic
distributional profile (Swinny et al., 2003). Indeed, at early postnatal stages, CRF-R1
protein was first localised in the cell body, but infused into the developing dendritic

130
Chapter 6
shafts. Based on the localisation data of CRF receptors (Bishop et al., 2000; King and
Bishop, 2002; Swinny et al., 2003) and the current study, a delicate interplay probably
ensues between climbing fibre co-localised CRF and urocortin. At the initial stage of
climbing fibre synaptogenesis on Purkinje cell somata, the effects of urocortin, via
CRF-R2 probably predominate, initiating dendritic outgrowth.
Dendritic elongation
The current study shows unequivocally that CRF and urocortin elongate primary
dendrites. CRF had the most influence on dendritic length, with cells from CRF-
treated slices having dendrites 25% longer than untreated cells and 9% longer than
urocortin treated cells. Furthermore, the dendritic lengthening effects of CRF appear
to be mediated by CRF-R1. The similar degree of inhibition of dendrite elongation by
the CRF-R1 and CRF-R2 receptor antagonist, a-h-CRF and the specific CRF-R1
antagonist, antalarmin, implies that CRF-R1 primarily mediates this process. This is
also consonant with localisation data that show CRF-R1 primarily distributed in the
growing dendritic shafts, closely trailing the upward migration of climbing fibres
(King and Bishop, 2002; Swinny et al., 2003). Furthermore, both CRF and urocortin
significantly increased total dendritic branching per cell. However, only CRF
significantly increased the number of branch points per primary dendrite showing
that the enhanced total dendritic branching effect of urocortin is a consequence of the
additional primary dendrites and not directly attributable to specific branching effects,
as is the case with CRF. Hence, after the phase of dendritic outgrowth, which is
probably heavily influenced by urocortin acting via CRF-R2, the effects of CRF
predominate, in concert with the upwardly-migrating CRF-R1, resulting in the
elongation and branching of the newly established dendrites.
Secondary messenger cascades
Metzger and Kapfhammer, (2000) have shown that protein kinase C (PKC) is critical
in dendritic development, with the activation of this pathway resulting in the inhibition
of normal dendritic development. In an attempt to understand the signal transduction
pathways involved in the CRF or urocortin-induced dendritic outgrowth, we used
specific antagonists against the two main secondary messenger pathways implicated,
namely the cAMP-PKA and MAPK pathways. Blockade of the PKA pathway caused
a 5 and 26% decrease in the CRF and urocortin induced dendritic outgrowth
respectively, whilst inhibition of the MAPK pathway resulted in a 42 and 45% decrease
respectively, suggesting that signals mediating the stimulatory effects of CRF and
urocortin on dendritic outgrowth are predominantly transduced via the MAPK pathway.
This is in accordance with an earlier study by Cibelli et al. (2001) who also showed

131
corticotropin releasing factor and Urocortin
the importance of PKA and MAPK in mediating the stimulator effects of CRF on
dendritic outgrowth in locus coeruleus-derived neurons.
The precise signal transduction pathway involved in CRF and urocortin-induced
dendritic elongation is somewhat ambiguous, based on the data emanating from this
study. PKA and MAPK antagonism resulted in similar inhibition of the CRF-induced
dendritic elongation. However, blockade of the MAPK showed stronger inhibition in
the urocortin-treated cells compared with PKA blockade. Notwithstanding, the PKA
pathway is reputed to be crucial in dendritic organisation. Microtubule–associated
protein (MAP) 2 is involved in microtubule assembly and the stabilisation of dendrites
(Kaech et al., 1996). MAP 2 is also one of the major PKA-anchoring proteins in
dendrites (Vallee et al., 1981). Not surprisingly, Harada et al. (2002) have shown that
MAP 2 and PKA signal transduction are essential for proper dendritic elongation.
Since G-protein coupled receptor signalling does not consist of solitary, linear
pathways (Belcheva and Coscia, 2002), and the fact that urocortin is not selective for
only CRF-R2 (Vaughan et al., 1995), the results of the current study are probably due
to “noise” from cross-talk between the two CRF receptors and the different
intracellular signalling cascades.
In conclusion, the study shows that intermittent exposure of Purkinje cells to CRF
and urocortin has a strong trophic action, stimulating dendritic outgrowth and
elongation. In converse, prolonged exposure results in underdeveloped dendritic trees.
This overt inhibition of dendritic structure, and probably synaptic remodelling, might
render an explanation, and potential therapeutic modalities, for pathologies where
early life stressful events or sustained stressful stimuli are know to manifest in
conditions like anxiety or major depression (De Souza, 1995), in which perturbations
of the CRF peptide/receptor system has been implicated.
REFERENCES
Alessi, D.R., Cuenda, A., Cohen, P., Dudley, D.T., Saltiel, A.R., 1995. PD 098059 is a specific inhibitor
of the activation of mitogen-activated protein kinase kinase in vitro and in vivo. J. Biol. Chem. 270,
27489-94.
Belcheva, M.M., Coscia, C.J., 2002. Diversity of G protein-coupled receptor signaling pathways to
ERK/MAP kinase. Neurosignals. 11, 34-44.
Bishop, G.A., 1990. Neuromodulatory effects of corticotropin releasing factor on cerebellar Purkinje
cells: an in vivo study in the cat. Neuroscience. 39, 251-7.
Bishop, G.A., King, J.S., 1999. Corticotropin releasing factor in the embryonic mouse cerebellum. Exp.
Neurol. 160, 489-99.
Bishop, G.A., Seelandt, C.M., King, J.S., 2000. Cellular localization of corticotropin releasing factor

132
Chapter 6
receptors in the adult mouse cerebellum. Neuroscience. 101, 1083-92.
Blank, T., Nijholt, I., Grammatopoulos, D.K., Randeva, H.S., Hillhouse, E.W., Spiess, J., 2003.
Corticotropin-releasing factor receptors couple to multiple G-proteins to activate diverse intracellular
signalling pathways in mouse hippocampus: role in neuronal excitability and associative learning. J.
Neurosci. 23, 700-7.
Cannon, R.C., Wheal, H.V., Turner, D.A., 1999. Dendrites of classes of hippocampal neurons differ in
structural complexity and branching patterns. J. Comp. Neurol. 413, 619-33.
Chang, C.P., Pearse, R.V., O’Connell, S., Rosenfeld, M.G., 1993. Identification of a seven transmembrane
helix receptor for corticotropin-releasing factor and sauvagine in mammalian brain. Neuron. 11, 1187-
95.
Cibelli, G., Corsi, P., Diana, G., Vitiello, F., Thiel, G., 2001. Corticotropin-releasing factor triggers
neurite outgrowth of a catecholaminergic immortalized neuron via cAMP and MAP kinase signalling
pathways. Eur. J. Neurosci. 13. 1339-48.
Dautzenberg, F.M., Braun, S., Hauger, R.L., 2001. GRK3 mediates desensitization of CRF1 receptors: a
potential mechanism regulating stress adaptation. Am. J. Physiol. Regul. Integr. Comp. Physiol. 280,
R935-46
Dautzenberg, F.M., Wille, S., Braun, S., Hauger, R.L., 2002. GRK3 regulation during CRF- and urocortin-
induced CRF1 receptor desensitization. Biochem. Biophys. Res. Commun. 298, 303-8.
De Souza, E.B., 1995. Corticotropin-releasing factor receptors: physiology, pharmacology, biochemistry
and role in central nervous system and immune disorders. Psychoneuroendocrinology. 20, 789-819.
Dotti, C.G., Sullivan, C.A., Banker, G.A., 1988. The establishment of polarity by hippocampal neurons
in culture. J. Neurosci. 8, 1454-68.
Gundlfinger, A., Kapfhammer, J.P., Kruse, F., Leitges, M., Metzger, F., 2003. Different regulation of
Purkinje cell dendritic development in cerebellar slice cultures by protein kinase Cα and -β. J. Neurbiol,
in press.
Harada, A., Teng, J., Takei, Y., Oguchi, K., Hirokawa, N., 2002. MAP2 is required for dendrite elongation,
PKA anchoring in dendrites, and proper PKA signal transduction. J. Cell Biol. 158, 541-9.
Hauger, R.L., Smith, R.D., Braun, S., Dautzenberg, F.M., Catt, K.J., 2000. Rapid agonist-induced
phosphorylation of the human CRF receptor, type 1: a potential mechanism for homologous
desensitization. Biochem. Biophys. Res. Commun. 268, 572-6.
Jeong, Y.G., Hyun, B.H., Hawkes, R., 2000. Abnormalities in cerebellar Purkinje cells in the novel
ataxic mutant mouse, pogo. Brain. Res. Dev. Brain. Res. 125, 61-7.
Kaech, S., Ludin, B., Matus, A., 1996. Cytoskeletal plasticity in cells expressing neuronal microtubule-
associated proteins. Neuron. 17, 1189-99.
Kase, H., Iwahashi, K., Nakanishi, S., Matsuda, Y., Yamada, K., Takahashi, M., Murakata, C., Sato, A.,
Kaneko, M., 1987. K-252 compounds, novel and potent inhibitors of protein kinase C and cyclic
nucleotide-dependent protein kinases. Biochem. Biophys. Res. Commun. 142, 436-40
King, J.S., Bishop, G.A., 2002. The distribution and cellular localization of CRF-R1 in the vermis of the
postnatal mouse cerebellum. Exp. Neurol. 178, 175-85.
Magee, J.C., 2000. Dendritic integration of excitatory synaptic input. Nat. Rev. Neurosci. 1, 181-90.
McAllister, A.K., 2001. Neurotrophins and neuronal differentiation in the central nervous system. Cell,
Mol, Life, Sci. 58, 1054-60.
Menzaghi, F., Howard, R.L., Heinrichs, S.C., Vale, W., Rivier, J., Koob, G.F., 1994. Characterization of
a novel and potent corticotropin-releasing factor antagonist in rats. J. Pharmacol. Exp. Ther. 269, 564-

133
corticotropin releasing factor and Urocortin
72.
Metzger, F., Wiese, S., Sendtner, M., 1998. Effect of glutamate on dendritic growth in embryonic rat
motoneurons. J. Neurosci. 18, 1735-42.
Metzger, F., Kapfhammer, J.P., 2000. Protein kinase C activity modulates dendritic differentiation of rat
Purkinje cells in cerebellar slice cultures. Eur. J. Neurosci. 12, 1993-2005.
Morara, S., van der Want, J.J., de Weerd, H., Provini, L., Rosina, A., 2001. Ultrastructural analysis of
climbing fiber-Purkinje cell synaptogenesis in the rat cerebellum. Neuroscience.108, 655-71.
Newport, DJ., Wilcox, M.M., Stowe, Z.N.. 2002. Maternal depression: a child’s first adverse life event.
Semin. Clin. Neuropsychiatry. 7,113-9.
Palkovits, M., Leranth, C., Gorcs, T., Young, W.S., 1987. Corticotropin-releasing factor in the
olivocerebellar tract of rats: demonstration by light- and electron-microscopic immunohistochemistry
and in situ hybridization histochemistry. Proc. Natl. Acad. Sci. 84, 3911-5.
Reyes, A., 2001. Influence of dendritic conductances on the input-output properties of neurons. Annu.
Rev. Neurosci. 24, 653-75.
Rossant, C.J., Pinnock, R.D., Hughes, J., Hall, M.D., McNulty, S., 1999. Corticotropin-releasing factor
type 1 and type 2alpha receptors regulate phosphorylation of calcium/cyclic adenosine 3',5'-
monophosphate response element-binding protein and activation of p42/p44 mitogen-activated protein
kinase. Endocrinology. 140, 1525-36.
Scelfo, B., Strata, P., Knopfel, T., 2003. Sodium imaging of climbing fiber innervation fields in developing
mouse Purkinje cells. J Neurophysiol.
Schrenk, K., Kapfhammer, J.P., Metzger, F., 2002. Altered dendritic development of cerebellar Purkinje
cells in slice cultures from protein kinase Cgamma-deficient mice. Neuroscience. 110, 675-89.
Swinny, J.D., Kalicharan, D., Gramsbergen, A., van der Want, J.J., 2002. The localisation of urocortin in
the adult rat cerebellum: a light and electron microscopic study. Neuroscience. 114, 891-903.
Swinny, J. D., Kalicharan, D., Blaauw, E.H., IJkema-Paassen, J., Shi, F., Gramsbergen, A., van der
Want, J.J.L. (2003) European Journal of Neuroscience. 18(3), 549-562.
Vale, W., Spiess, J., Rivier, C., Rivier, J., 1981. Characterization of a 41-residue ovine hypothalamic
peptide that stimulates secretion of corticotropin and beta-endorphin. Science. 213, 1394-7.
Vallee, R.B., DiBartolomeis, M.J., Theurkauf, W.E., 1981. A protein kinase bound to the projection
portion of MAP 2 (microtubule-associated protein 2). J. Cell Biol. 90, 568-76.
Vaughan, J., Donaldson, C., Bittencourt, J., Perrin, M.H., Lewis, K., Sutton, S., Chan, R., Turnbull,
A.V., Lovejoy, D., Rivier, C., et al.,1995. Urocortin, a mammalian neuropeptide related to fish urotensin
I and to corticotropin-releasing factor. Nature. 378, 233-4.
Vetter, P., Roth, A., Hausser, M., 2001. Propagation of action potentials in dendrites depends on dendritic
morphology. J. Neurophysiol. 85, 926-37.
Webster, E.L., Lewis, D.B., Torpy, D.J., Zachman, E.K., Rice, K.C., Chrousos, G.P., 1996. In vivo and
in vitro characterization of antalarmin, a nonpeptide corticotropin-releasing hormone (CRH) receptor
antagonist: suppression of pituitary ACTH release and peripheral inflammation. Endocrinology. 137,
5747-50.
Weinstock M., 2001. Alterations induced by gestational stress in brain morphology and behaviour of the
offspring. Prog. Neurobiol. 65,427-51.
Wong, R.O.. Ghosh, A.. 2002. Activity-dependent regulation of dendritic growth and patterning. Nat.
Rev. Neurosci. 3, 803-12.
Yang, Q., Hashizume, Y., Yoshida, M., Wang, Y., Goto, Y., Mitsuma, N., Ishikawa, K., Mizusawa, H.,
2000. Morphological Purkinje cell changes in spinocerebellar ataxia type 6. Acta. Neuropathol. (Berl).

134
Chapter 6
100, 371-6.
Zhang, L.I., Poo, M.M., 2001. Electrical activity and development of neural circuits. Nat. Neurosci.
4,1207-14.
Zhang, Z., Casey, D.M., Julien, J.P., Xu, Z., 2002. Normal dendritic arborization in spinal motoneurons
requires neurofilament subunit L. J. Comp. Neurol. 450, 144-52.

135
Chapter 7
General discussion and conclusions

136
Chapter 7
General discussion and conclusions
The cerebellum is crucial for the coordination of motor activity as well the learning
of skilled motor acts. This influence of the cerebellum on motor coordination begins
during ontogeny with cerebellar development not only closely tailoring the
development of motor coordination, but also impacts on the execution of smooth
motor acts in later life. Hence, an understanding of the factors governing the
development of the cerebellar circuitry is intrinsic to the understanding of normal
motor development, and in case of pathologies, possible therapeutic avenues.
Corticotropin releasing factor (CRF) was recently elevated to a status of prominence
in cerebellar studies when it was shown that CRF contained in one of the main
cerebellar afferent systems, namely the climbing fibres is essential for cerebellar
long term depression (LTD) (Miyata et al., 1999), a type of synaptic plasticity proposed
as the cellular basis of motor learning (Ito, 1984). Intriguingly, mRNA studies showed
that CRF and one of its receptors, CRF receptor 1 (CRF-R1) is expressed from late
embryonic stages, through to adulthood (Chang et al., 1993). An added fascination is
the recent identification of a newer member of the CRF family of peptides, namely
urocortin (Vaughan et al., 1995) as well as an additional receptor, CRF-R2 (Lovenberg
et al., 1995). The early onset of expression, prior to the formation of any functional
synapses suggests that initially, CRF might subserve a developmental role in the
cerebellar circuitry, with a shift to that of a neuromodulator once the cerebellar circuitry
has been established.
To explore this tantalising hypothesis, a precise understanding of the temporal and
spatial expression of CRF, urocortin and their receptors in the developing cerebellum
was mandatory. The morphological development of the rat cerebellum is well
documented. Furthermore, different stages of the formation of the cerebellar circuitry
can be equated with distinct stages of development at the behavioural level. Hence,
by detailing the ultrastructural expression of CRF peptides at timed intervals during
the early postnatal developmental stages, a strong correlation could be observed
between these peptides and ongoing developmental processes. The hypotheses that
evolved from the localisation studies were then tested in vitro in organotypic slice
cultures so as to cement the roles of CRF and urocortin as key players during cerebellar
development.
In chapter 2, it was shown that CRF is selectively expressed within the two major
afferent systems, the climbing fibres and mossy fibres and not by cerebellar neurons.
An early diffuse expression at postnatal day (PD) 3 throughout the different cerebellar
lobules, was followed by a more restricted distributional pattern from PD onwards.
Indeed, from early adulthood, CRF expression predominated in the posterior vermal

137
general discussion and conclusions
lobules, especially lobules IX and X and also the flocculus and paraflocculus. Taken
together, this further alludes to a generic role for CRF at early postnatal stages with
a shift to that of a more discrete functional modulator in certain cerebellar regions.
Intriguingly, CRF containing climbing fibres projecting to different cerebellar lobules
appeared to exhibit disparate stages of development and varying immunoreactive
intensities. Indeed, labelled climbing fibres in lobule two, at the stages of PD 6-9,
were mainly at the level of the Purkinje cell soma, whereas those climbing fibres in
lobule IX or X has already traversed the Purkinje cell layer and entered the molecular
layer. It is unclear whether the differences in the temporal expression of CRF in
fibres projecting to different parts of the cerebellum are a consequence of the different
birth-dates of Purkinje cells in the various regions or are indicators of the rates of
functional maturity of these regions. If CRF directly impacts on cerebellum
development and later function, it’s thus apparent that such a role is temporally
continguous in different regions of the cerebellum and might be related to the
functional maturity of the different pre-cerebellar systems.
The predominance of CRF immunoreactivity in the vestibular cerebellum
overwhelmingly indicts it as a crucial entity in vestibular function. As mentioned
above, CRF labelled climbing fibres in lobules IX, X, the flocculus and paraflocculus
are the first to translocate from the Purkinje cell somata to their primary dendrites.
The translocation between the stages of PDs 6-9 coincides remarkably with the stage
of development of the vestibular and postural system at which the rat is able to stand
on its fours and to perform a few steps (Westerga and Gramsbergen, 1990). In the
next days, rats are able to walk for longer periods, however, without clear cut
qualitative changes in the immature locomotor patterns. Between the stages of PDs
15-18, the diffuse expression of CRF becomes more restricted, manifesting in
transverse zones of immuno-positive climbing fibres and zones not showing
immunoreactivity. At this stage, the reduced immunoreactivity parallels with the onset
of fluent motor patterns that are generally initiated between PDs 15-16. Evidence
indicates that the stage of cerebellar development is the limiting factor for the fluent
walking pattern to emerge (Gramsbergen, 1998) in which extremity movements and
postural control are closely coupled (Gramsbergen, 1999). Whilst the presence of
CRF in the flocculus and paraflocculus provides unambiguous evidence for the role
of CRF in vestibular function, close inspection of the projections from CRF zones in
the vermis and hemispheres will provide further insights into other functional roles
of CRF.
A perplexing issue to arise from the CRF localisation study was the apparent lack of
CRF immunoreactivity in certain regions of the cerebellum while the CRF receptor
mRNA expression occurs throughout the cerebellum. This suggested that CRF

138
Chapter 7
receptors in CRF “free” regions of the cerebellum might be coupled to by another
member of the CRF family of peptides. Urocortin, a newer member appeared to be a
likely candidate since it binds to both CRF receptors (Vaughan et al., 1995).
Since the literature was littered with conflictual reports of urocortin expression in
the cerebellum, in chapter 3, as a first step, we detailed its expression in the adult
cerebellum and in chapter 4, the postnatal distributional profile. There was a high
degree of overlap in terms of regional developmental expression of urocortin compared
with CRF. The expression of urocortin in the developing cerebellum is not evenly
distributed throughout different lobules. Immunoreactivity was evident within the
first postnatal week in the Purkinje cell layers of the anterior lobules I-III and posterior
lobules IX-X. By PD 12, it was difficult to discern differences in intensity of urocortin
immunoreactivity between different lobules. Following a plateau in immunoreactivity
at PD 15, immunoreactivity abated, particularly in the anterior lobules. Urocortin
was shown to present in some unique profiles. Like CRF, urocortin was also contained
with climbing and mossy fibres. However, in contrast, urocortin was also expressed
by cerebellar neurons. In particular, Purkinje cells, granule cells and inhibitory
interneurons of the molecular layer.
While both CRF and urocortin were shown to be co-localised in climbing fibres,
they exhibited different immunoreactive profiles. CRF immunoreactivity could be
visualised in climbing fibre tracts as well as terminals. However, urocortin, for the
most part, was contained almost exclusively in the terminals and not in the fibre
tracts. This difference in the processing of climbing fibre-derived CRF and urocortin
by the inferior olive is the first intimation of diverging roles for these two peptides.
The congregation of urocortin in axonal terminals, not unlike conventional
neurotransmitters like glutamate, suggests a prominent role in the synaptic
environment. It is known that CRF acts as a neuromodulator since it causes Purkinje
cell excitability by augmenting glutamate activity. (Bishop, 1990). It remains to be
elucidated whether urocortin directly or indirectly influences Purkinje cell activity.
It has been shown that CRF is crucial for the induction of LTD (Miyata et al., 1999),
hence only the early phase or protein synthesis independent phase of this phenomenon.
In comparison to CRF, urocortin has been shown to cause cyclic AMP binding protein
(CREB) phosphorylation (Rossant et al., 1999) that generally is an indicator of gene
expression, and subsequent protein synthesis. A fertile area of future studies could
be the possible role of urocortin in LTD, and if so, whether its role extends to different
phases of this process, perhaps the protein synthesis segment.
A fundamental finding was that Purkinje cells express urocortin. These cells provide
the sole output of the cerebellar cortex. This certainly extends the role of the “CRF
system” since it was commonly believed that this system was only implicated in

139
general discussion and conclusions
modulating sensory information entering the cerebellum. More specifically, both CRF
and urocortin might be involved in modulating signals entering the cerebellar cortex,
via climbing and mossy fibres, while only urocortin is implicated with those exiting
the cerebellar cortex, via Purkinje cell axons. The presence of urocortin in posterior
lobules at later stages, particularly in lobules IX and X reinforces the roles of CRF
peptides in vestibular and postural control. An interesting future line of work would
be to explore the integrity of the vestibulo-occulo reflex (VOR) (Ito, 1984) in
urocortin-deficient mice (Vetter et al., 2002). The principal site of urocortin expression,
the Edinger-Westphal nucleus projects to the flocculus as well as providing pre-
ganglionic innervation to ciliary muscles (Hultborn et al., 1978; Roste and Dietrichs,
1988). The concurrent localisation of urocortin in strategic centres of this circuit
suggests that it could be an important role player.
To fully interpret the emerging picture of CRF and urocortin localisation in the circuitry
of the cerebellum, data was needed on the localisation of the receptors through which
they transduce their signals. All previous studies on CRF receptor localisation had
concentrated mostly on outlining their mRNA distributional profiles. Data showing
the site of protein expression at the subcellular level was needed to understand their
roles in the context of the synaptically and functionally diverse cerebellar circuitry.
CRF and urocortin act by coupling to two G-protein coupled receptors (Chang et al.,
1993; Lovenberg et al., 1995). While both CRF and urocortin couple with relatively
equal affinity to CRF-R1, urocortin bind to CRF-R2 with much greater affinity
(Vaughan et al., 1995). In chapter 5, we addressed the issue of CRF receptor
localisation. The precise localisation of the receptors within the cerebellar circuitry
would further help to elucidate the relative roles of CRF and urocortin in the
cerebellum. This is of particular importance with respect to the Purkinje cell, since
this cell-type receives functionally diverse inputs on its different regions.
A novel finding was that CRF receptors occur in both pre- and postsynaptic elements.
Indeed, both CRF-R1 and 2 was localised in climbing fibre profiles as well as in
Purkinje cells. Receptor protein was also localised in inhibitory interneurons in the
molecular layer and in Bergman glia.
None of the previous localisation studies, all at the light microscopical level, have
shown that either CRF-R1 or R2 are localised in climbing fibres. However, this was
the first study to examine receptor localisation at the electron microscopic level. In
other regions, recent physiological evidence certainly appears to validate the
presynaptic localisation of CRF receptors. Specific physiological studies in the
cerebellum are lacking, hence the functional significance of presynaptic receptors in
climbing fibres is yet unclear. Nevertheless, presynaptic receptors generally serve to
control the synaptic release of their ligands. Hence, presynaptic CRF-R1 and 2 might

140
Chapter 7
selectively control the release of CRF and urocortin respectively, either during activity
dependent phases of development or in adulthood. CRF is essential for the induction
of cerebellar LTD. Following plasticity like LTD, it has been directly established
that there is decreased presynaptic function (Zakharenko et al., 2002) with the
dynamics of synaptic vesicle fusion also altered. Hence, presynaptic CRF receptors
might come into play during phases of LTD, limiting the release of CRF and urocortin
and perhaps, through receptor cross-talk, other agents involved in LTD like glutamate
via the presynaptic metabotropic glutamate receptors.
The remit of the CRF receptor localisation study was to establish structure-function
paradigms. It was apparent that CRF-R1 and CRF-R2 are differentially expressed in
the developing Purkinje cell, suggesting divergent roles for the two receptors. CRF-
R2 was predominantly localised on the Purkinje cell soma, first diffusely at early
stages but becoming membrane bound from PD 15 onwards. Based on the evidence
above, CRF-R2 conveys signals from axons synapsing on Purkinje cell somata. At
early developmental stages, climbing fibres make synaptic contact on somatic spines
of Purkinje cells. If CRF peptides are involved in the activity dependent phase of
development, the localisation data suggest that climbing fibre-released urocortin by
coupling to somatically bound CRF-R2 might be instrumental in initiating dendritic
outgrowth. Evidence from subsequent in vitro studies certainly support this notion.
At later stages of development, once climbing fibres have translocated to Purkinje
cell dendrites, the Purkinje cell soma is generally innervated by GABAergic basket
cell terminals or recurrent axon collateral of Purkinje cells (Altman and Bayer, 1997).
The localisation of CRF-R2 in this region is not unexpected since we have shown
that both basket and Purkinje cells contain urocortin, suggesting that the locus of
action of CRF-R2 is on postsynaptic membranes. At first glance, it might appear
perplexing why an apparently excitatory peptide like urocortin might be contained
in inhibitory terminals. However, Chavas and Marty, (2003) have shown the
GABAergic terminals in the cerebellum being capable of producing excitatory as
well as inhibitory effects. The biochemical makeup of these “mixed” synapses are
not yet fully known, but urocortin could very well likely play a role in buffering the
inhibitory effects of these synapses. The postsynaptic membrane activity of CRF-R2
is further validated by its localisation at parallel fibre-Purkinje cell synapses.
In comparison to CRF-R2, CRF-R1 undergoes a more dynamic postnatal distribution.
At early stages, labelling was localised in the somata but shifted into the dendrites as
these developed. CRF-R1 begins to be expressed on Purkinje cell from embryonic
day 17, prior to synaptogenesis. During development, CRF-R1 might act as a
signalling molecule to guide CRF and urocortin containing climbing fibres in finding
their contacts, since climbing fibres first synapse on Purkinje cell somata, and then

141
general discussion and conclusions
translocate to the dendrites. Once synaptogenesis is initiated, CRF and urocortin
might act as trophic agents with the peptides and the receptor being instrumental in
Purkinje cell dendritic growth and differentiation.
In chapter 6, we explore the hypothesis that CRF and urocortin actively contribute to
Purkinje cell dendritic growth and differentiation using organotypic slice cultures.
Using cerebellar slice cultures of rats from PD 8, we showed that both CRF and
urocortin have a potent effect on Purkinje cell dendritic differentiation. Intermittent
exposure of the slices to CRF and urocortin (12 hours per day for 10 days in vitro)
enhanced dendritic outgrowth and elongation whereas continuous exposure of the
slices to the peptides inhibited growth.
Urocortin was shown to be more potent in inducing dendritic outgrowth, with cells
from slices exposed to it intermittently having the most number of primary dendrites,
on average, 5.3. In contrast, CRF was shown to be more potent than urocortin in
causing dendritic elongation. This finding vindicates earlier suppositions derived
from CRF receptor localisation data that showed that CRF-R2 is mainly expressed in
the Purkinje cell soma and CRF-R1 in the Purkinje cell dendritic shaft. Furthermore,
in line with previous studies on the binding affinities of CRF and urocortin, the use
of receptor antagonists showed that CRF mediated its effects predominantly via CRF-
R1 and urocortin, apparently, via both receptors. Finally, blockade of the PKA and
MAPK pathways implicated these secondary messenger pathways in mediating the
trophic effects of CRF and urocortin. (see for figures 1 and 2: page 150)
REFERENCES
Bishop, G.A. (1990) Neuromodulatory effects of corticotropin releasing factor on cerebellar Purkinje
cells: an in vivo study in the cat. Neuroscience. 39, 251-7.
Chang, C.P. Pearse, R.V. O’Connell, S. Rosenfeld, M.G. (1993) Identification of a seven transmembrane
helix receptor for corticotropin-releasing factor and sauvagine in mammalian brain. Neuron. 11, 1187-
95.
Chavas, J. Marty, A. (2003) Coexistence of excitatory and inhibitory GABA synapses in the cerebellar
interneuron network. J Neurosci. 23, 2019-31.
Gramsbergen, A. (1998) Posture and locomotion in the rat: independent or interdependent development?
Neurosci Biobehav Rev. 22, 547-53.
Gramsbergen, A. Geisler, H.C. Taekema, H. van Eykern, L.A. (1999) The activation of back muscles
during locomotion in the developing rat. Brain Res Dev Brain Res. 112, 217-28.
Hultborn, H. Mori, K. Tsukahara, N. (1978) Cerebellar influence on parasympathetic neurones
innervating intra-ocular muscles. Brain Res. 159, 269-78.
Lovenberg, T.W. Liaw, C.W. Grigoriadis, D.E. Clevenger, W. Chalmers, D.T. De Souza, E.B. Oltersdorf,
T. (1995) Cloning and characterization of a functionally distinct corticotropin-releasing factor receptor
subtype from rat brain. Proc. Natl. Acad. Sci. U S A. 92, 836-40.

142
Chapter 7
Ito, M. (1984) The Cerebellum and Neural Control. New York: Raven.
Miyata, M. Okada, D. Hashimoto, K. Kano, M. Ito, M. (1999) Corticotropin-releasing factor plays a
permissive role in cerebellar long-term depression. Neuron. 22, 763-75.
Rossant, C.J. Pinnock, R.D. Hughes, J. Hall, M.D. McNulty, S. (1999) Corticotropin-releasing factor
type 1 and type 2alpha receptors regulate phosphorylation of calcium/cyclic adenosine 3',5'-
monophosphate response element-binding protein and activation of p42/p44 mitogen-activated protein
kinase. Endocrinology. 140, 1525-36.
Roste, G.K. Dietrichs, E. (1988) Cerebellar cortical and nuclear afferents from the Edinger-Westphal
nucleus in the cat. Anat Embryol. 178, 59-65.
Vaughan, J. Donaldson, C. Bittencourt, J. Perrin, M.H. Lewis, K. Sutton, S. Chan, R. Turnbull, A.V.
Lovejoy, D. Rivier, C. et al. (1995) Urocortin, a mammalian neuropeptide related to fish urotensin I
and to corticotropin-releasing factor. Nature. 378, 233-4.
Vetter, D.E. Li, C. Zhao, L. Contarino, A. Liberman, M.C. Smith, G.W. Marchuk, Y. Koob, G.F.
Heinemann, S.F. Vale, W. Lee, K.F. (2002) Urocortin-deficient mice show hearing impairment and
increased anxiety-like behavior. Nat Genet. 31, 363-9.
Westerga, J. Gramsbergen, A. (1990) The development of locomotion in the rat. Brain Res Dev Brain
Res. 57, 163-74.
Zakharenko, S.S. Zablow, L. Siegelbaum, S.A. (2002) Altered presynaptic vesicle release and cycling
during mGluR-dependent LTD. Neuron. 35, 1099-110.

143
SUMMARY
The cerebellar circuitry plays a central role in motor coordination and motor learning.
What’s more, the development of motor coordination is dependent upon the
development of the cerebellum and its circuitry. Thus, an understanding of the factors
influencing cerebellar development is integral to elucidating how normal motor
development ensues and allows for directed interventions in cases of pathologies.
A plethora of neuropeptides are expressed within the cerebellum and in pre-cerebellar
centres, which supply the cerebellar afferents. In this thesis, we investigate the role
of corticotropin releasing factor (CRF) and urocortin in the developing rat cerebellum
in an attempt to idendtify causal relations between their spatial and temporal
expression, well-defined stages of cerebellar development and motor development.
Both CRF and urocortin initially are distributed diffusely throughout the cerebellum
and at later stages expression predominates in the posterior cerebellum, or the
vestibulo-cerebellum. An important difference at later stages is that whilst CRF is
exclusively expressed in the cerebellar afferents, urocortin is localised in the afferents
as well as Purkinje neurons, the sole output of the cerebellar cortex. The receptors of
CRF and urocortin are expressed in climbing fibres and their contacts, the Purkinje
cells, suggesting that the receptors modulate the release of CRF and urocortin from
climbing fibres as well as the effects of these peptides on Purkinje cells once they are
released. Finally, using organotypic slice cultures, we have shown that CRF and
urocortin have direct and differential trophic actions on Purkinje cells, inducing the
outgrowth and differentiation of Purkinje cell dendrites.

144
SAMENVATTING
De kleine hersenen spelen een belangrijke rol bij het coordineren van motorische
activiteit en bij het aanleren van complexe bewegingen zoals lopen, slikken, spreken,
fietsen en autorijden. Het leren van een goede coordinatie van motorische bewegingen
is afhankelijk van de ontwikkeling van de kleine hersenen en de aanleg van neuronale
circuits. Daarom is het van belang om de factoren, die bijdragen aan de ontwikkeling
van de kleine hersenen te bestuderen. Hierdoor kunnen we een beter begrip krijgen
welke circuits de normale motorische activiteit reguleren en controleren en mogelijk
ook hoe afwijkingen in de motoriek verklaard zouden kunnen worden.
Dit proefschrift gaat over Corticotropin Releasing Factor (CRF) en de rol van
aanverwante neuropeptiden in de kleine hersenen, het cerebellum. CRF speelt een
belangrijke rol bij stress. Bij stressvolle situaties komt CRF vrij in een deel van de
hersenen -de hypothalamus- en veroorzaakt daar de zgn. stressreaktie met als
kenmerken o.a. verhoogde bloeddruk, versnelde hartslag en toename van zweten.
CRF in het cerebellum speelt waarschijnlijk geen rol bij de stress response. Eerder
onderzoek, in het cerebellum, heeft aannemelijk gemaakt dat CRF op volwassen leeftijd
noodzakelijk is om specifieke, fysiologische veranderingen in de synaptische aktiviteit
tussen zenuwcellen, de zgn. synaptische plasticiteit, te kunnen laten verlopen. De
ontdekking van het optreden van verschillende vormen van synaptische plasticiteit,
bij excitatoire en inhibitoire zenuwverbindingen, heeft duidelijk gemaakt hoe
zenuwcellen synaptische input integreren. Langdurige veranderingen in synaptische
effectiviteit kunnen stimulerend zijn, dit staat bekend als “Long Term Potentiation”
(LTP) of remmend zijn, “Long Term Depression” (LTD). Het type synaptische
plasticiteit dat in het cerebellum veel aandacht heeft gekregen is de LTD. LTD kan
optreden als twee zenuwvezels, de parallelvezel en de klimvezel, tegelijk aktief zijn
ten opzichte van een derde zenuwcel, in dit geval de Purkinje cel. Het effect van de
simultane aktiviteit leidt er toe dat de synaptische signaaloverdracht tussen
parallelvezels en Purkinje cellen vermindert, deze verandering wordt beschouwd als
de cellulaire basis van leren en geheugen van motorische activiteit.
In het cerebellum zelf zijn geen zenuwcellen die CRF produceren, maar er zijn CRF-
producerende zenuwcellen, die vanuit de hersenstam met hun vezels in het cerebellum
synaptische contacten vormen. Deze vezels heten klimvezels en maken contact met
Purkinje cellen. De klimvezel-Purkinje cel synaps is uniek, in het volwassen zoogdier
cerebellum ontvangt iedere Purkinje cel slechts één klimvezel. De klimvezel vormt
ongeveer 1500 synaptische contacten op de dendrieten van één Purkinje cel. Purkinje
cellen hebben een sterk vertakte dendrietboom, de grootste van alle zenuwcellen. Die

145
dendrietboom ontwikkelt zich bij zoogdieren pas na de geboorte. De groei van de
dendrietboom en de vertakkingen wordt in belangrijke mate gestuurd door
neuropeptiden, die door de klimvezels worden afgegeven.
Tijdens de postnatale ontwikkeling komen CRF en het nauw verwante urocortin in
wisselende hoeveelheden voor. In dit proefschrift wordt aangetoond dat de plaats en
het moment van de aanwezigheid van CRF en urocortin gekoppeld is aan specifieke
ontwikkelingsstadia en de daarmee samenhangende motorische ontwikkeling. Zowel
CRF als urocortin zijn aanvankelijk diffuus verspreid in de kleine hersenen. Op latere
leeftijden komen ze voornamelijk voor in het achterste deel van de kleine hersenen,
het vestibulocerebellum, dat een belangrijke rol speelt bij het evenwicht. Een opvallend
verschil tussen CRF en urocortin is, dat CRF uitsluitend in klimvezels voorkomt,
terwijl urocortin aanwezig is zowel in klimvezels als in Purkinje cellen. Behalve de
localisatie van CRF en urocortin is ook onderzocht waar de receptoren van CRF en
urocortin gelocaliseerd zijn. Het blijkt dat de respectievelijke receptoren op een
karakteristieke manier verdeeld zijn. Zij komen tot expressie bij de synaptische
contacten van klimvezels met Purkinje cellen, maar ook in de klimvezels zelf. Dit
zou kunnen betekenen, dat de receptoren de afgifte van CRF en urocortin van
klimvezels kunnen reguleren, in de vorm van presynaptische modulatie van de Purkinje
cellen en de synaptische overdracht.
Bij het onderzoek zijn ook kweken van hersenplakjes gebruikt om aan te tonen dat
CRF en urocortin een direct effect hebben op Purkinje cellen. Het blijkt dat korte,
onderbroken expositie de uitgroei van dendrieten stimuleert. Bij constante blootstelling
van CRF en urocortin vindt een remming in de ontwikkeling van de Purkinje cel
dendrietboom plaats. Er worden veel minder vertakkingen gevormd en er ontstaat
een “kale boom” terwijl bij afwisselende blootstelling aan CRF en urocortin een
sterke groei optreedt van de Purkinje cel dendrietboom, met meer vertakkingen. Deze
kweekmethode laat ook duidelijk zien dat CRF en urocortin niet hetzelfde effect
hebben op de uitgroei van de Purkinje cel dendrieten, maar op specifieke onderdelen
als gevolg van de verschillende signalen. Of deze situaties zich ook bij de normale
ontwikkeling van de kleine hersenen voordoen, is nog niet bekend, maar met de
resultaten van dit onderzoek hebben inzicht gegeven in de betekenis van CRF en
urocortin tijdens de ontwikkeling van de kleine hersenen. Verder onderzoek zal zich
moeten richten op de andere factoren die groei-en ontwikkelings-processen in het
cerebellum beïnvloeden. Kennis omtrent de determinanten van deze processen is
van groot belang voor de behandeling van verworven en aangeboren motorische
handicaps.

146
ACKNOWLEDGEMENTS
Without any experience in neuroscience or hardly having peered into a microscope,
my two supervisors, Albert Gramsbergen and Han van der Want gave me the benefit
of the doubt, and a position in their research groups. Sheer folly or desperation?
Firstly, to both of you, I’m grateful that your loyalty was to supervising and mentoring
the individual first and then the project. Secondly, you were brave enough to allow
me great latitude in the execution of the project whilst still keeping me on the straight
and narrow, and for that I thank you.
Han, my direct supervisor, your interest and concern for me was never confined to
the aspects of my project. Being in a foreign country, it was extremely comforting to
know that if ever I needed help, there was somebody who I could count on. The
manner in which you manage the department always made it a pleasurable work
environment.
Albert, my promotor, I am truly appreciative of your ability to make me see the bigger
picture. Your infectious enthusiasm and continued wonderment at my little discoveries
was constant fuel and sustenance during the “winters” of the project.
To the members of the lab for electron microscopy where I was based. I thank you
firstly for the way you made me feel completely welcome and were constantly
concerned with my welfare. To the research technicians viz. Bert Blaauw, Nel Blom,
Freark Dijk, Ruby Kalicharan, Geert Kors, Ietse Stokroos and Henk de Weerd, having
to learn everything from scratch, you showed remarkable patience in teaching me the
various techniques of electron microscopy. You were never too busy and no question
was too trivial. A special note of thanks is extended to Ruby Kalicharan who worked
most extensively on my project. Ruby, zonder jou kom ik nergens!! Your utmost
dedication to the project ensured that no matter what obstacles were in our path, they
would be surmounted. Your constant concern and support for my welfare will not be
forgotten.
The photographers, Bert Hellinga, Dick Huizinga and Peter van der Syde, thank you
for assuring that I constantly had access to top-quality images. Bert, thank you for
teaching me the finer tricks of Photoshop! Peter, thank for your time and effort in
preparing this thesis.
Greetje Noppert, you have been a constant source of support from the very first emails
before I had even arrived in Groningen. It was always reassuring to know that I could
rely on your professionalism and efficiency.
Robert Liem, Janneke Mijnster and Natasha Gounko, thank for all the helpful

147
discussions and continued interest in my work.
To the colleagues the department of Medical Physiology. Although it not based full-
time on the 8th floor, the friendly and supportive atmosphere made me feel at home.
A special word of thanks is extended to Jos. Thank you for the training and advice,
the help with the experiments of the project, especially with the preparation of “royal
quality” cerebellar slice cultures. Most importantly, I value your pleasant demeanor
and enthusiasm on work-related issues or otherwise.
Friedrich Metzger, thank you for showing the kind of interest you did in my project,
teaching me new techniques and for patiently putting up with all my unending queries.
The best is yet to come!
Nieske Brower and Knut Biber, thank you for all the constructive criticisms and helpful
assistance, especially with the in situ hybridization experiments.
Erik Boddeke, thanks for all the support and advice.
Thank you to Fuxin Shi, Department of Membrane Cell Biology for help with western
blotting.
Cesar Hulstaert, international students coordinator and Gerry Hoogenberg, the manager
of the research group, thank you for your continued help and support, especially in
the difficult first months of my arrival and for taking care of all my administrative
tangles.
I am grateful to the BCN for the financial support throughout my PhD studies and
also for the printing of this thesis. A special note of thanks to Diana Koopmans and
Rob Visser.
To my friends outside the lab, with special mention to my paranymphs Florence and
Virginia, and Anne, Danny, Luc, Norel. I consider myself fortunate to have crossed
your paths. We have massaged one another’s aches, celebrated our small achievements,
and all along the way, we never failed to see the funny side of life. I hope this is only
the beginning.
Last but not least, recognition and gratitude is also deserving of those who enabled
me to get into a position to actually undertake this course of study. I thank my parents
for their unstinting support and encouragement throughout my life. I also express my
most sincerest gratitude to my siblings for their continued support during this period,
especially my brother Andrew, crime-fighter extraordinair!

148
CURRICULUM VITAE
The author of this thesis was born in Dundee, South Africa on 6 October, 1969. He
completed his secondary schooling in 1987 at Bechet Secondary School, Durban. In
1988, he undertook undergraduate studies in pharmacy at the University of Natal,
graduating in 1992. After doing his pharmacist’s internship in 1993, he undertook a
Masters in Medical Science (Pharmacology) at the University of Durban-Westville
(UDW), graduating in 1996. After lecturing in the department of Pharmacy at (UDW)
from 1997-1999, he was awarded an Ubbo Emmius Scholarship to undertake doctoral
studies at the Departments of Cell Biology (Lab for Electron Microscopy) and Medical
Physiology, University of Groningen.

149
Chapter 1, Figure 4. (see page 16)
Chapter 6, figure 3. (see page 123)

150
Figure 1. Schematized summary of CRF and urocortin expressionin the adult rat cerebellum. CRF is expressed exclusively in thecerebellar afferent systems whereas urocortin is localized in theafferents as well as in cerebellar neurons.Note that in inferior olivary axons, CRF is located throughoutthe fibre tract, whereas urocortin is selectively localized in theterminals of climbing fibres.
Figure 2. Schematized summary of CRF receptor expression in the postnatal rat cerebellum.Both receptors are expressed in climbing fibre terminals and in Purkinje cells. Initial Purkinjecell CRF-R1 immunoreactivity is localized somatically but then infuses into the growingdendrites. CRF-R2 immunoreactivity remains in the Purkinje cell soma, diffusely spread atearly stages, but bound to plasmic membranes at later stages.
Figures of Chapter 7

151

152





























