Embed Size (px)
DESCRIPTION
第八章 糖代谢 第一节 糖酵解( Glycolysis ) 第二节 丙酮酸的去路 第三节 三羧酸循环( TCA ) 第四节 蔗糖和多糖的分解 第五节 乙醛酸循环途径 第六节 磷酸戊糖途径 第七节 糖类的合成 第八节 血糖水平的调节. 第一节 糖酵解 ( Glycolysis ). 概念:糖酵解是描述葡萄糖通过 果糖二磷酸酯 的途径降解为丙酮酸并提供 ATP 的一系列反应。 它是首先被阐明的代谢途径,由于 G.Emlden, O.Meyerhof, J.K.Parnas 三人的贡献最大,又称 EMP 途径。. 一、化学历程(三个阶段 10 个反应). - PowerPoint PPT Presentation
Citation preview

第八章 糖代谢第一节 糖酵解( Glycolysis )第二节 丙酮酸的去路第三节 三羧酸循环( TCA)第四节 蔗糖和多糖的分解第五节 乙醛酸循环途径第六节 磷酸戊糖途径第七节 糖类的合成第八节 血糖水平的调节

第一节 糖酵解( Glycolysis ) 概念:糖酵解是描述葡萄糖通过果糖二磷酸酯的途径降解为丙酮酸并提供 ATP的一系列反应。它是首先被阐明的代谢途径,由于 G.Emlden, O.Meyerhof, J.K.Parnas 三人的贡献最大,又称 EMP 途径。
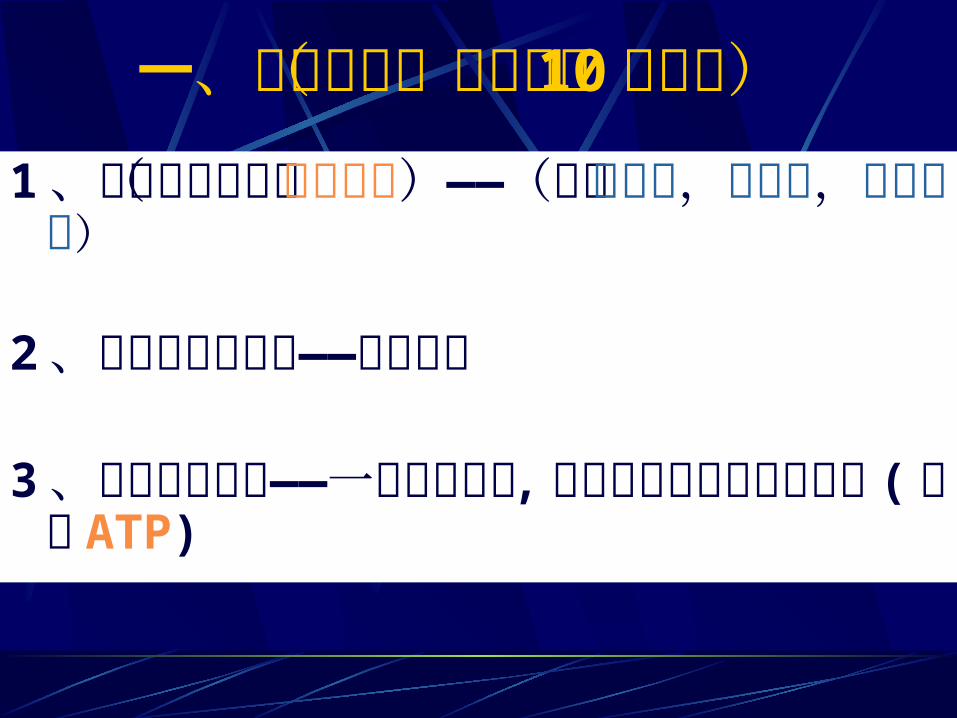
一、化学历程(三个阶段 10个反应) 1 、己糖的磷酸化(活化过程)——(已糖磷酸化,异构化,再磷酸化)2 、磷酸己糖的裂解——二步反应3、丙酮酸的生成——一步脱氢反应 ,二步底物水平磷酸化反应 (产生 ATP)
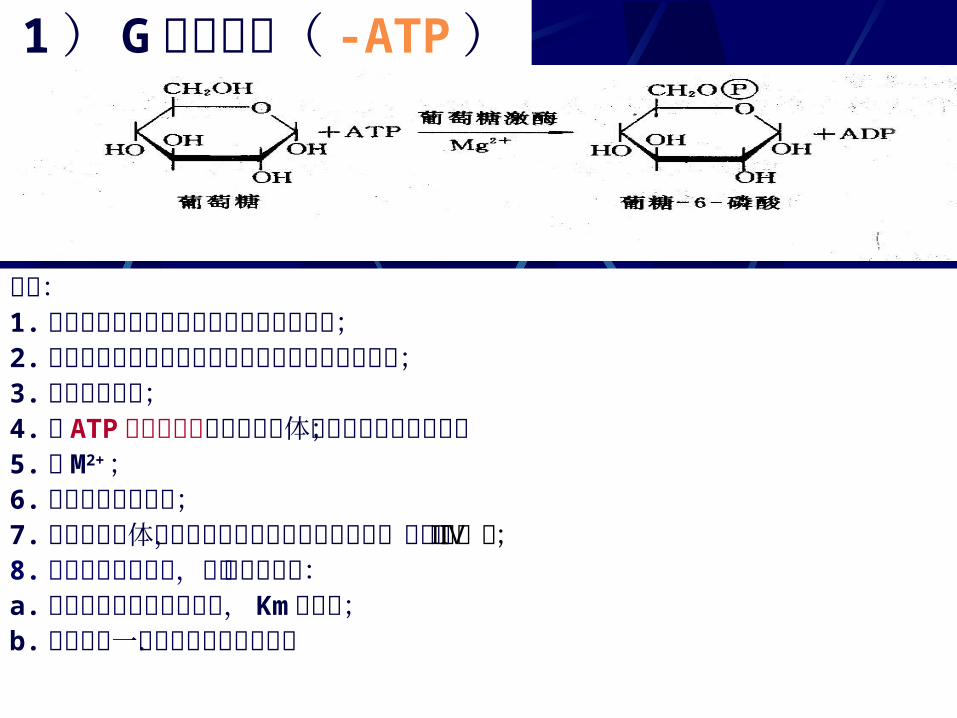
1 ) G的磷酸化( -ATP )
要点:1.葡萄糖进入细胞后首先的反应的是磷酸化;2.磷酸化后葡萄糖即不能自由通过细胞膜而逸出细胞;3.己糖激酶催化;4.由 ATP 的磷酸基团转移给接受体的反应都由激酶催化;5.需 M2+;6.基本上是不可逆的;7.哺乳类动物体内已发现有四种己糖激酶同功酶,分别为Ⅰ至Ⅳ型;8. 肝细胞中存在的是,称为葡萄糖激酶:a.它对葡萄糖的亲和力很低, Km 值很高;b.此酶的另一个特点是受激素控制.
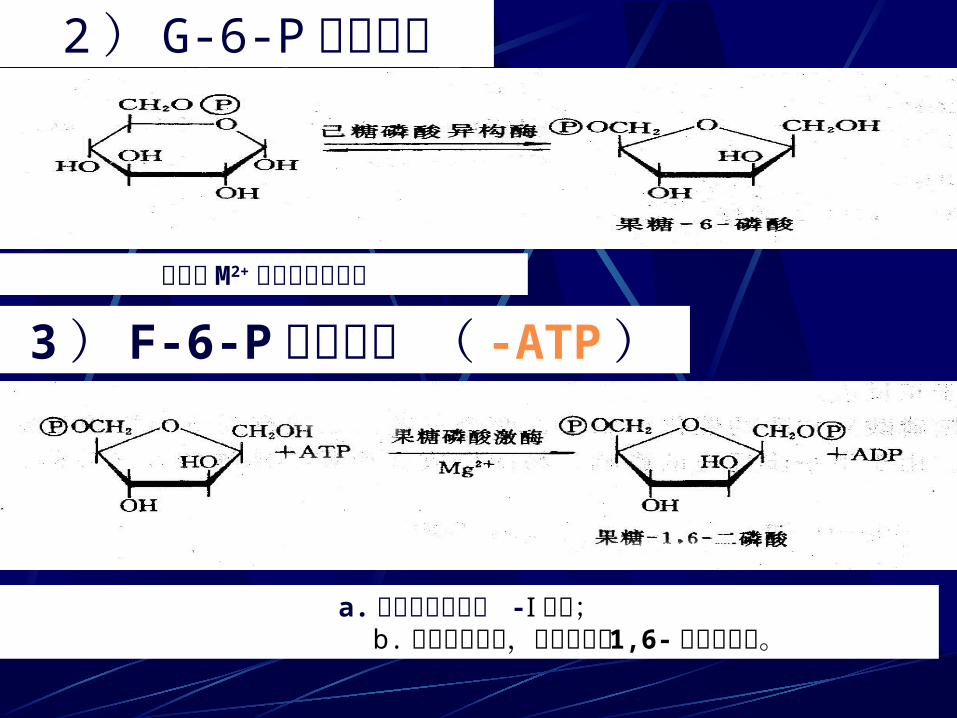
2 ) G-6-P 的异构化
3 ) F-6-P 再磷酸化 ( -ATP )是需要 M2+ 参与的可逆反应
a. 由磷酸果糖激酶 -Ⅰ 催化; b. 是非平衡反应,倾向于生成 1,6- 双磷酸果糖。

4 ) 1,6- 二磷酸果糖的裂解
( 1)此步反应是可逆的( 2)由醛缩酶( alkolase)催化,而且有利于已糖的合成,所以称为醛缩酶
( 3)最终产生 2分子丙糖,即磷酸二羟丙酮和 3-磷酸甘油醛。
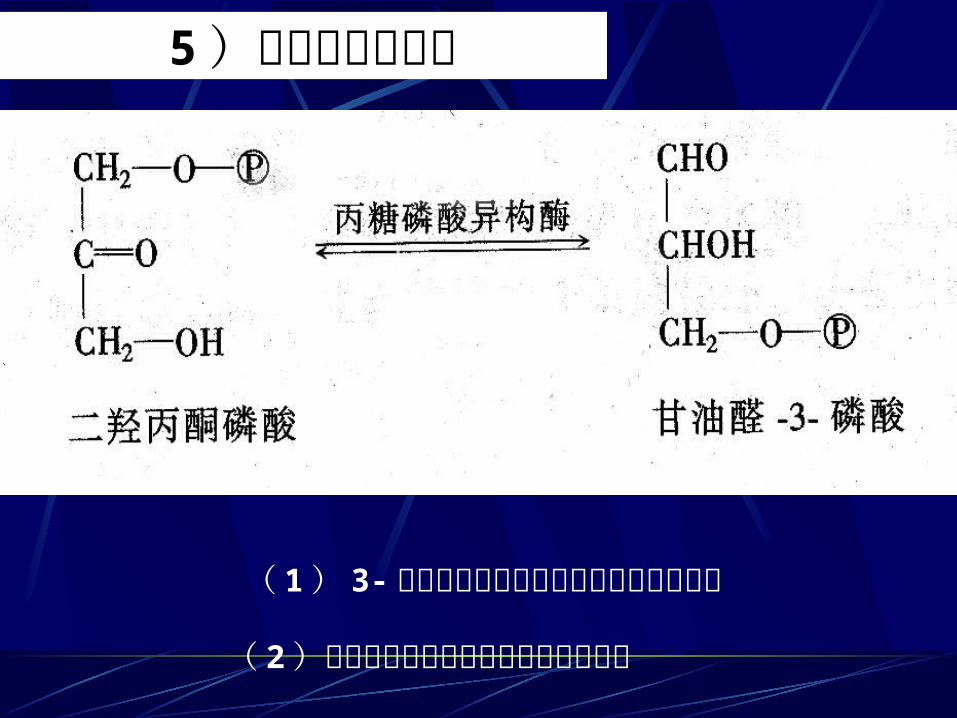
5 )磷酸丙糖的互变
( 1) 3-磷酸甘油醛和磷酸二羟丙酮是同分异构( 2)在磷酸丙糖异构酶催化下可互相转变
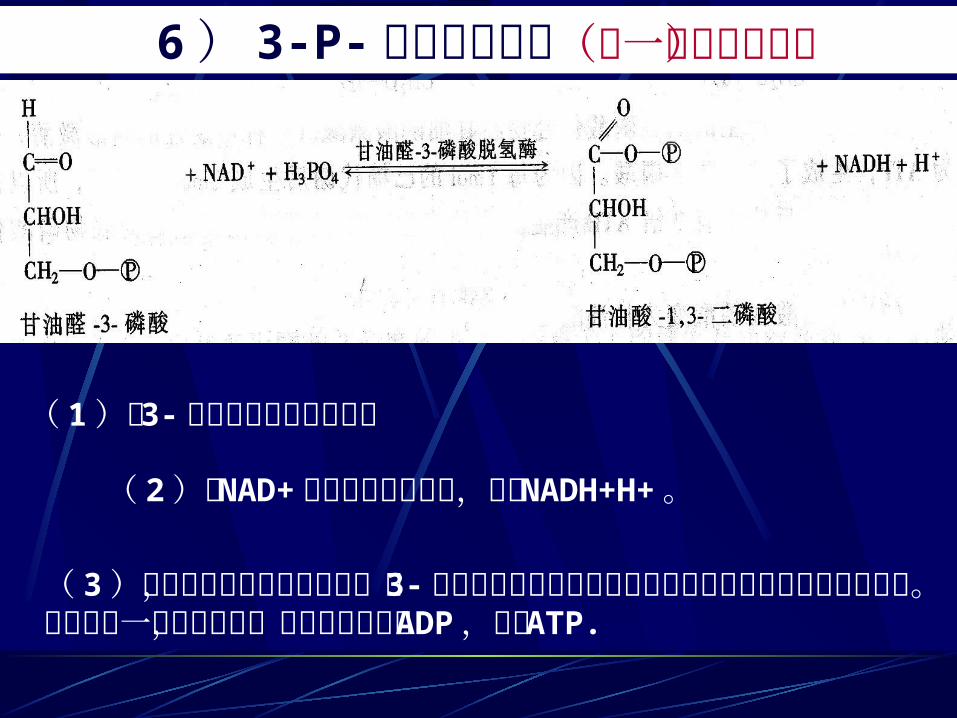
6 ) 3-P- 甘油醛的氧化(唯一的氧化反应)
( 1)由 3-磷酸甘油醛脱氢酶催化 ( 2)以 NAD+为辅酶接受和电子,生成 NADH+H+。
( 3)参加反应的还有无机磷酸,当 3-磷酸甘油醛的醛基氧化脱氢成羧基即与磷酸形成混合酸酐。该酸酐含一高能磷酸键,可将能量转移至 ADP,生成 ATP.
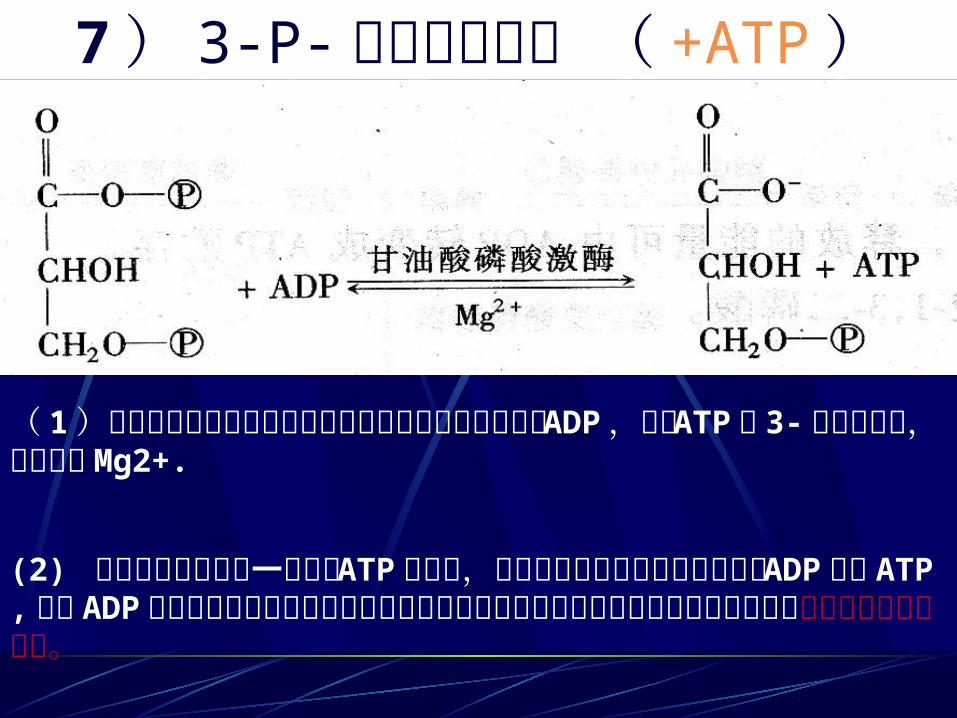
7 ) 3-P- 甘油酸的生成 ( +ATP )
( 1)磷酸甘油酸激酶催化混合酸酐上的磷酸从羧基转移到 ADP,形成 ATP和 3-磷酸甘油酸,反应需要Mg2+.
(2) 这是酵解过程中第一次产生 ATP 的反应,将底物的高能磷酸基直接转移给 ADP 生成 ATP, 这种 ADP 或其他核苷二磷酸的磷酸化作用与底物的脱氢作用直接相偶联的反应过程称为底物水平磷酸化作用。
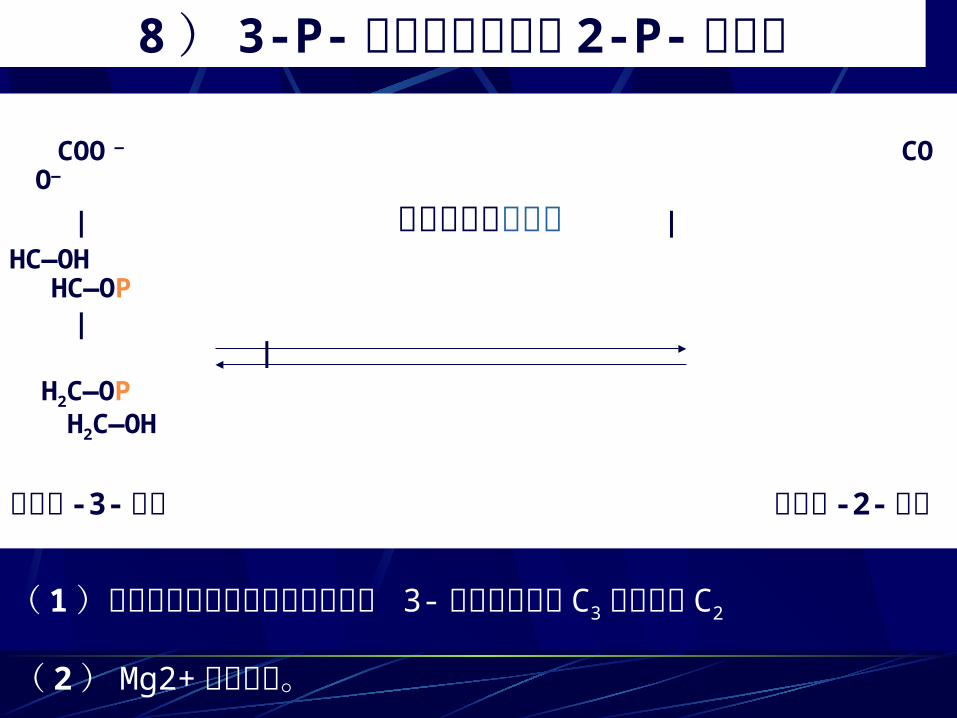
8 ) 3-P- 甘油酸异构化为 2-P- 甘油酸 COO — COO—
| 磷酸甘油酸变位酶 |HC—OH HC—OP | | H2C—OP H2C—OH
甘油酸 -3- 磷酸 甘油酸 -2- 磷酸( 1)磷酸甘油酸变位酶催化磷酸基从 3- 磷酸甘油酸的 C3 位转移到C2
( 2) Mg2+ 是必需的。
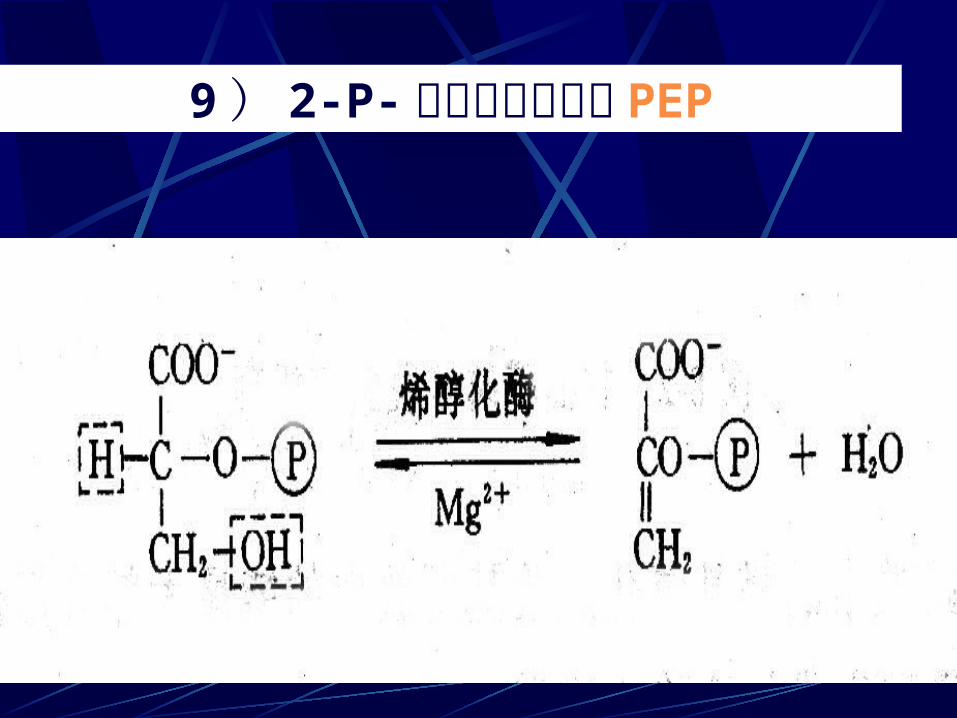
9 ) 2-P- 甘油酸脱水形成 PEP

10 )丙酮酸的生成( +ATP )
( 1)由丙酮酸激酶催化的。 ( 2)丙酮酸激酶的作用需要 K+ 和 Mg2+ 参与。
( 3)在胞内这个反应是不可逆的。 ( 4)糖酵解途径中第二次底物水平磷酸化。


二、 EMP 的讨论(以 G 为底物) 1 、细胞定位:细胞质
总反应式: C6H12O6+2NAD++2ADP→2C3H4O3+2NADH+2H++2ATP2 、 EMP 反应中大部分可逆,只有已糖激酶( 1 ),磷酸已糖激酶( 3 ),丙酮酸激酶( 10 )催化的反应不可逆,这三个反应称限速反应,三种酶称限速酶。3 、 EMP 中 ATP 的计量 反应 ATP 的变化1 ) G→G-6-P -13 ) F-6-P →FDP -17 ) 1,3- 二磷酸甘油酸 →
3- 磷酸甘油酸 +210 ) PEP → 丙酮酸 +2 净变化 +2
产能效率: 31%

4 、 EMP 的生理意义 (1) 在缺氧情况下快速释放能量,使机体仍能进行生命活动,这对肌肉收缩更为重要.(2) 成熟的红细胞没有线粒体,完全依赖糖酵解提供能量.(3)神经、白细胞、骨髓等代谢极为活跃,即使不缺氧也常由糖酵解提供部分能量.(4) 酵解过程的中间产物可为机体其它生物合成提供碳架。
三、糖酵解的调节糖酵解途径的中三个不可逆反应分别为:己糖激酶磷酸果糖激酶 糖酵解途径的调节酶丙酮酸激酶

1 、己糖激酶对糖酵解的调节(1) 己糖激酶为变构酶,受其产物葡萄糖 -6- 磷酸的强烈抑制;葡萄糖激酶分子内不存在葡萄糖 -6- 磷酸的变构部位,故不受葡萄糖 -6- 磷酸的影响。(2) 长链脂酰 CoA 对其有变构抑制作用,着在饥饿时减少肝和其他组织摄取葡萄糖有一定意义.(3) 胰岛素可诱导葡萄糖激酶基因的转录,促进酶的合成.

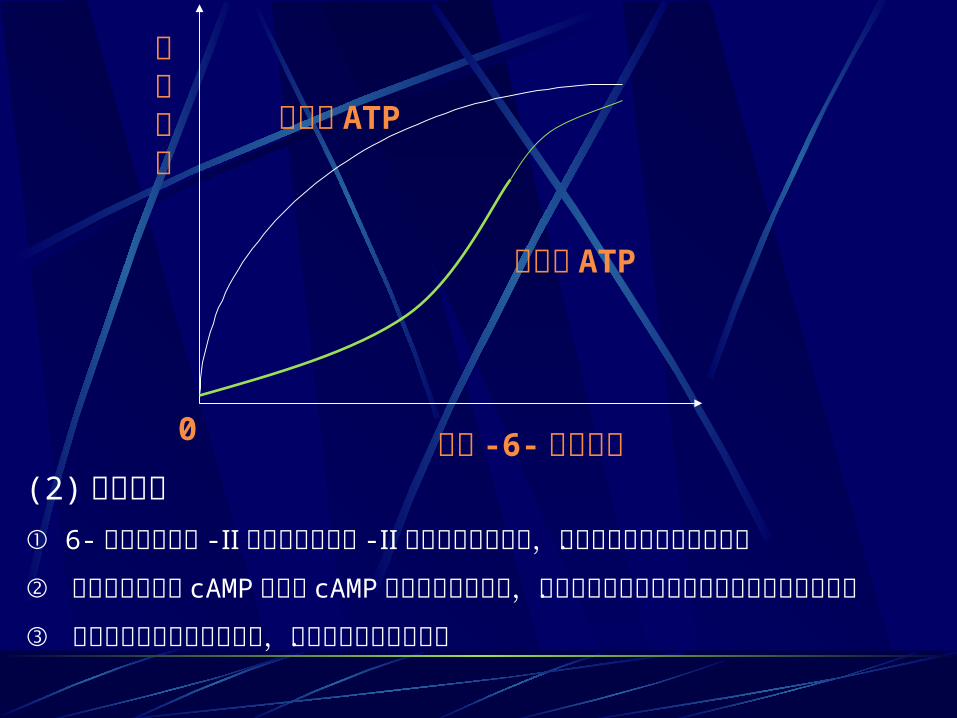
2 、磷酸果糖激酶 -Ⅰ 是控制糖酵解的关键酶(限速酶)(1) 变构调节① 6- 磷酸果糖激酶 -Ⅰ 是一四聚体,受多种变构调节剂的影响.② ATP 和柠檬酸是 6- 磷酸果糖激酶 -Ⅰ 的变构抑制剂.③ 6- 磷酸果糖激酶 -Ⅰ 的变构激活剂有 AMP , ADP , 1,6- 二磷酸果糖和 2,6-二磷酸果糖.④ 1,6- 二磷酸果糖是 6- 磷酸果糖激酶 -Ⅰ 的反应产物,这中产物正反馈作用是比较少见的,它有利于糖的分解.
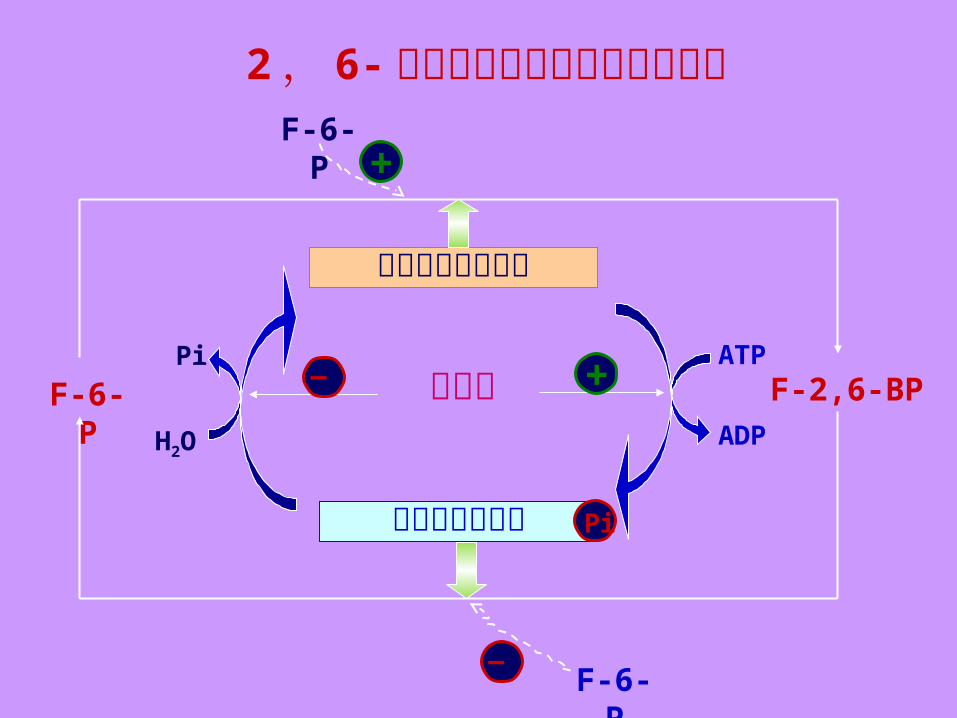
⑤ 果糖 -2 , 6- 二磷酸是 6- 磷酸果糖激酶 -Ⅰ 最强的的变构激活剂. ⑥ 果糖 -2 , 6- 二磷酸由 6- 磷酸果糖激酶 -Ⅱ 催化 6- 磷酸果糖 C2 磷酸化而成.
⑦ 6- 磷酸果糖激酶 -Ⅱ 实际上是一种双功能酶,在酶蛋白中具有两个分开的催化中心,故同时具有 6- 磷酸果糖激酶 -Ⅱ 和果糖双磷酸酶 -Ⅱ 两种活性.
高浓度 ATP
低浓度 ATP
0 果糖 -6-磷酸浓度
反应速度
(2) 共价修饰① 6- 磷酸果糖激酶 -Ⅱ 或果糖双磷酸酶 -Ⅱ 还可在激素作用下,以共价修饰方式进行调节.
② 胰高血糖素通过 cAMP 及依赖 cAMP 的蛋白激酶磷酸化,磷酸化后其激酶活性减弱而磷酸酶活性升高. ③ 磷蛋白磷酸酶将其去磷酸后,酶活性的变化则相反.
2 , 6- 二磷酸果糖合成和降解的调控
磷酸化的前后酶
去磷酸化的前后酶
F-6-P 低血糖
Pi
+— F-2,6-BPATP
ADPH2O
Pi
+F-6-P
F-6-P—

3 、丙酮酸激酶对糖酵解的调节 (1) 果糖 -1,6- 二磷酸是丙酮酸激酶的变构激活剂,而长链脂肪酸、乙酰 CoA 、 ATP 则有抑制作用.(2) 在肝内丙氨酸也有变构抑制作用.(3) 共价修饰方式调节
① 依赖 cAMP 的蛋白激酶和依赖 Ca2+ 、钙调蛋白的蛋白激酶均可使其磷酸化而失活. ② 胰高血糖素可通过 cAMP 抑制丙酮酸激酶的活性.
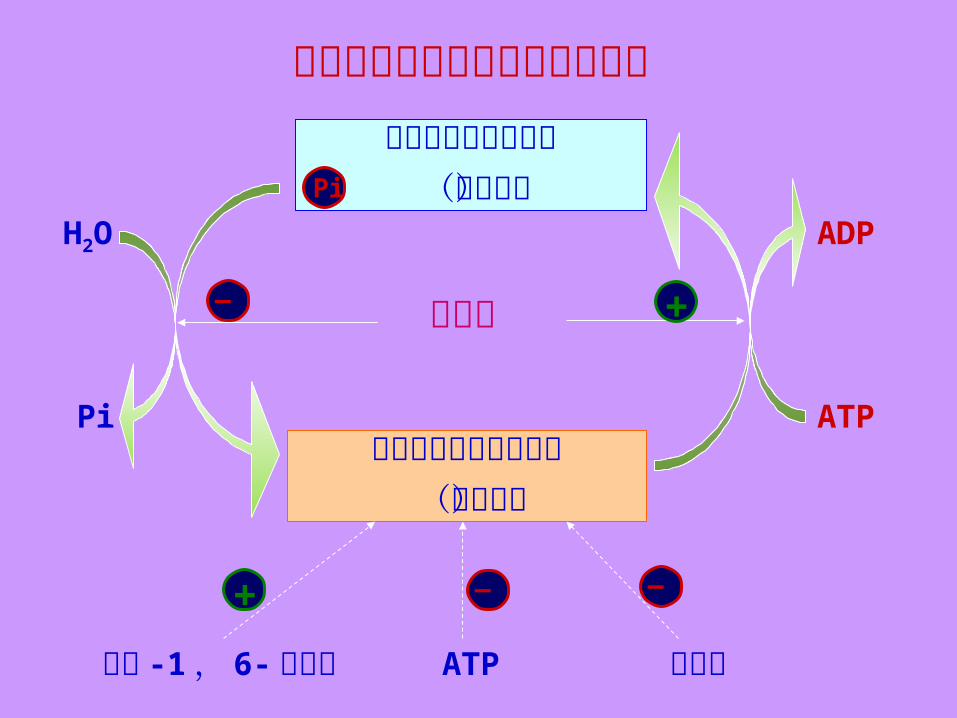
丙酮酸激酶催化活性控制关系图磷酸化的丙酮酸激酶
(低活性)
去磷酸化的丙酮酸激酶(高活性)
H2O
Pi ATP
ADP
果糖 -1, 6-二磷酸 ATP 丙氨酸— —+
低血糖Pi
+—

(1) 当能量消耗多,细胞内 ATP/AMP 比值降低时, 6- 磷酸果糖激酶 -Ⅰ 和丙酮酸激酶均被激活,加速葡糖的分解.反之 , 细胞内 ATP 的储备丰富时,通过糖酵解分解的葡萄糖就少.(2) 正常进食时,肝亦仅氧化少量葡萄糖,主要由氧化脂肪获得能量.(3) 进食后,胰高血糖素分泌减少,胰岛素分泌增加,果糖 -2 ,6- 二磷酸的合成增加,加速糖循糖酵解途径分解,主要是生成乙酰 CoA 以合成脂肪酸.(4) 饥饿时,胰高血糖素分泌增加,抑制了果糖 -2 , 6- 二磷酸的合成和丙酮酸激酶的活性,即抑制糖酵解,这样才能有效地进行糖异生,维持血糖的水平.
4 、共同调节
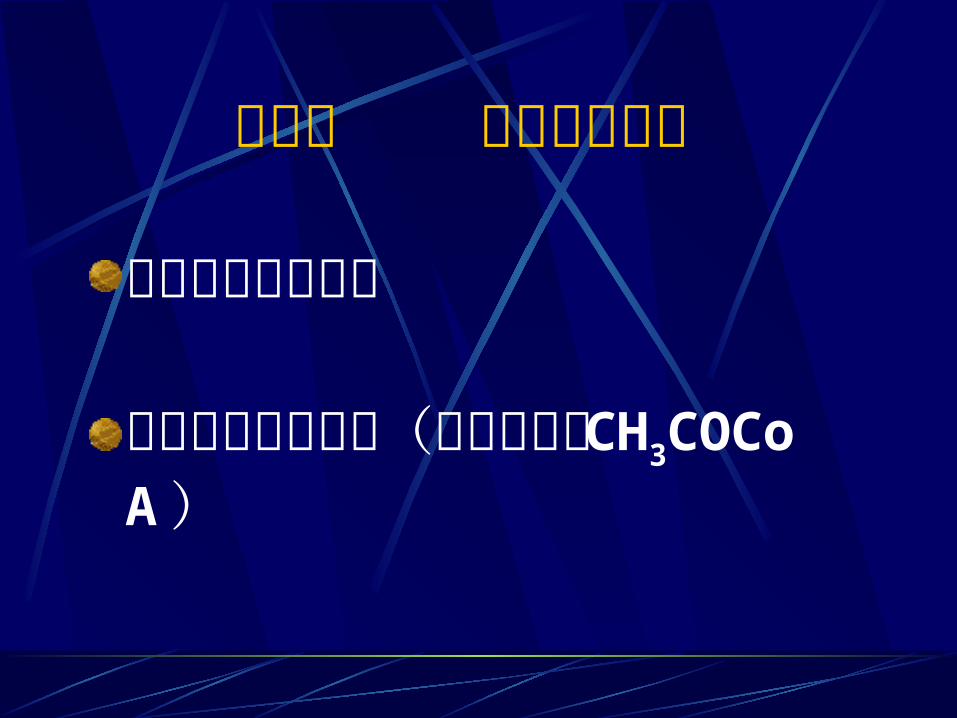
第二节 丙酮酸的去路 丙酮酸的无氧降解丙酮酸的有氧降解(氧化脱羧成CH3COCoA )

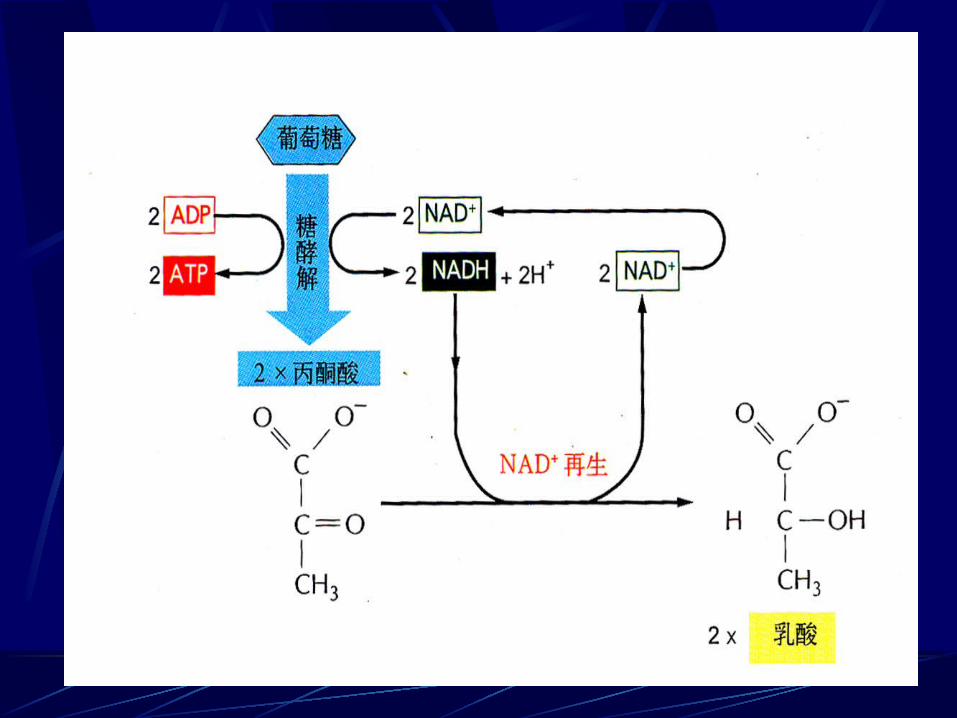
一、丙酮酸的无氧降解 1 、生成乳酸
在无氧条件下,把糖酵解中生成的 NADH 中的H 交给丙酮酸生成乳酸的过程称为乳酸发酵。
• 剧烈活动的肌肉细胞、胡萝卜根、玉米、豌豆和马铃薯在无氧条件下。•利用乳酸发酵还可以制造酸牛奶、泡菜等。
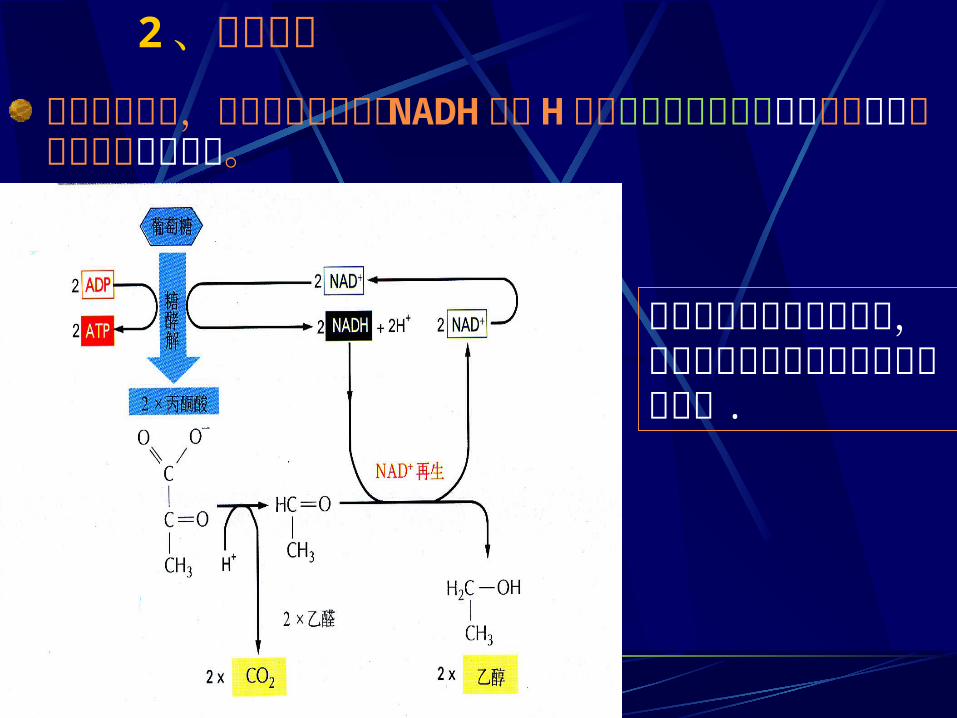
2、乙醇发酵在无氧条件下,把糖酵解中生成的 NADH 中的 H 交给丙酮酸脱羧产物乙醛生成乙醇的过程称为乙醇发酵。
在大多数植物和微生物中,在有些可以厌氧生长的生物如酵母中 .
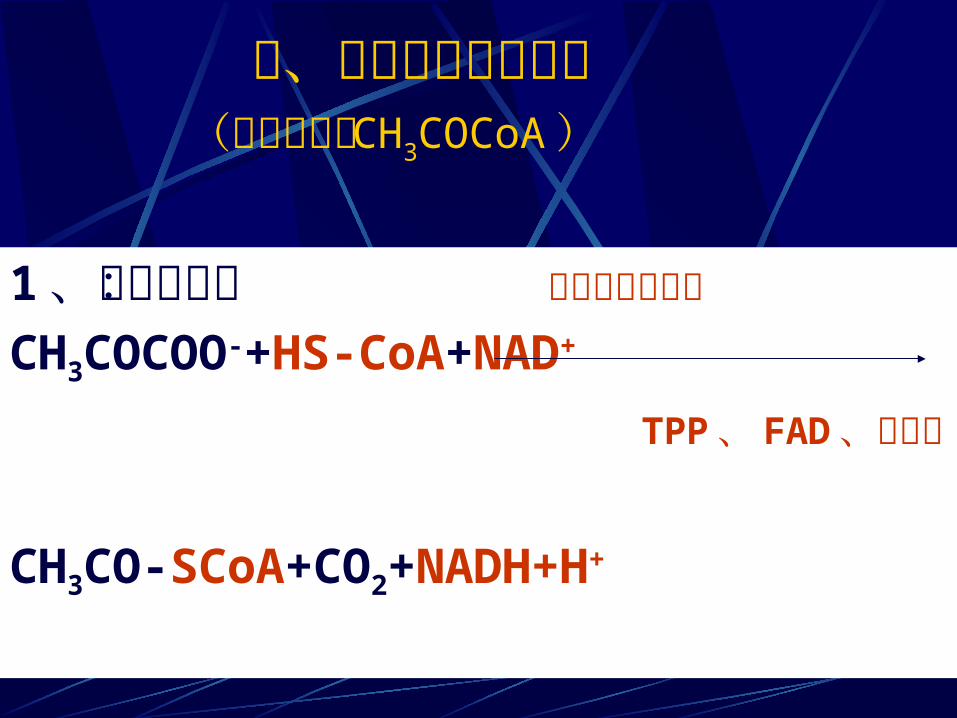
二、丙酮酸的有氧降解(氧化脱羧成 CH3COCoA )
1 、总反应式: 丙酮酸脱氢酶系CH3COCOO-+HS-CoA+NAD+ TPP 、 FAD 、硫辛酸
CH3CO-SCoA+CO2+NADH+H+
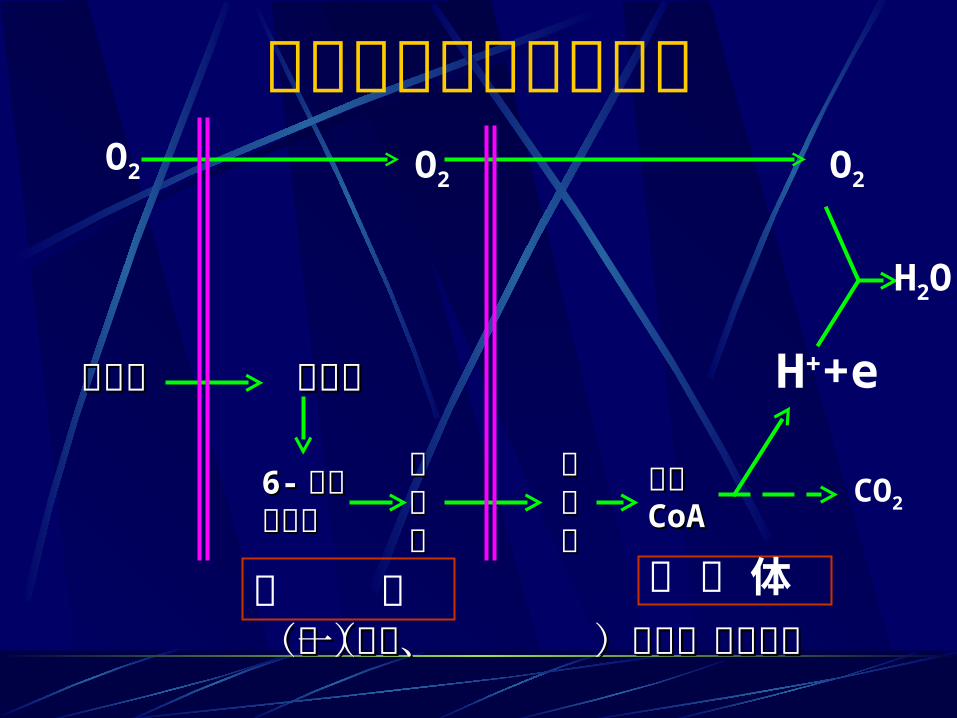
葡萄糖有氧氧化的概况O2
O2 O2
H2O
H++e
CO2
乙酰乙酰CoACoA丙丙酮酮酸酸
丙丙酮酮酸酸6-6- 磷酸磷酸葡萄糖葡萄糖
葡萄糖葡萄糖葡萄糖葡萄糖
线 粒 体胞 液(第一阶段) (第二、三阶段)(第一阶段) (第二、三阶段)
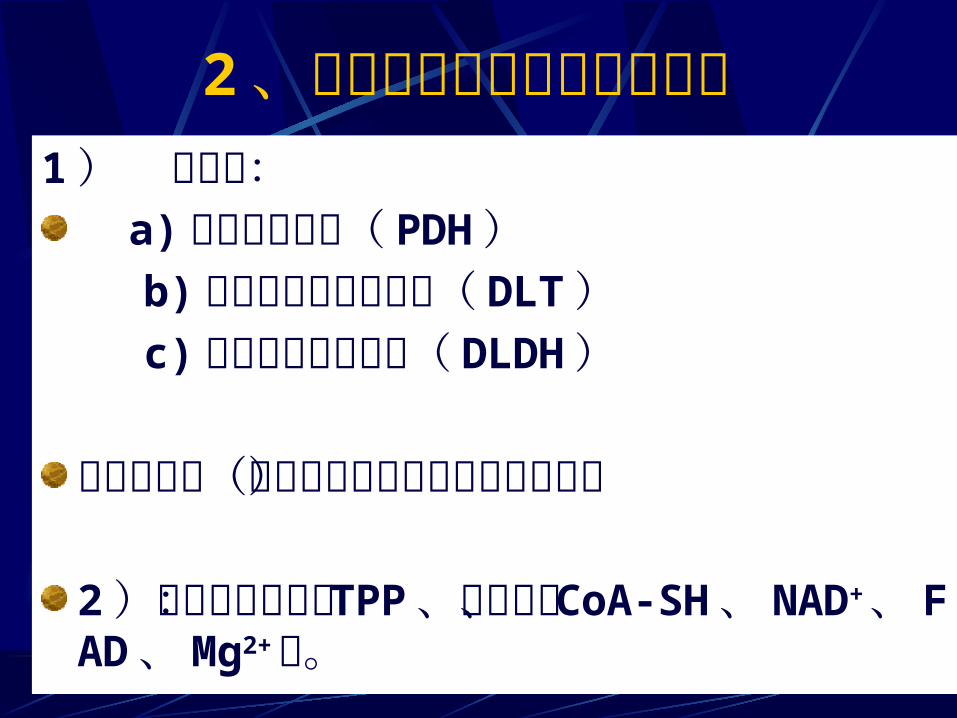
2、丙酮酸脱氢酶系组成与装配1 ) 三种酶: a) 丙酮酸脱氢酶( PDH )
b) 二氢硫辛酸转乙酰酶( DLT ) c) 二氢硫辛酸脱氢酶( DLDH ) 若干调控酶(对丙酮酸氧化脱羧起调控作用)2)若干辅助因子: TPP 、硫辛酸、 CoA-SH 、NAD+、 FAD 、 Mg2+等。
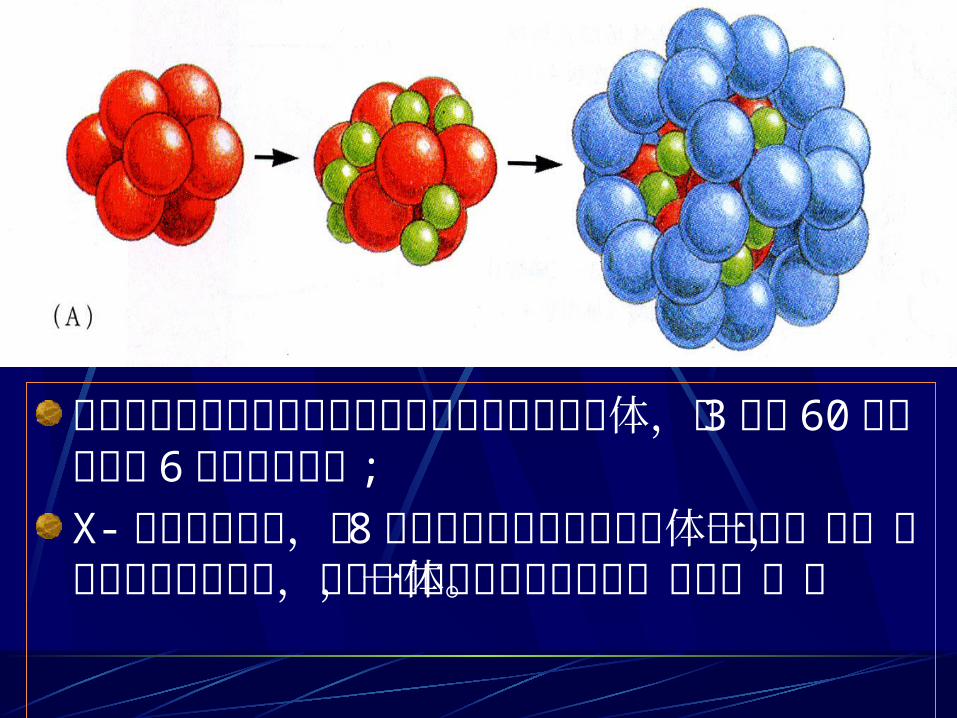
大肠杆菌中的丙酮酸脱氢酶复合物为圆球状多面体,由 3种酶 60 条多肽链和 6种辅因子组成 ;X- 射线研究表明,有 8个硫辛酸转乙酰酶的三聚体组合在一起,形成中空的方型结构,其他两种酶与这个核心结合,成为一体。

3、丙酮酸氧化脱羧的机理
丙酮酸脱羧形成羟乙基-TPP-E1;由二氢硫辛酰胺转乙酰酶(E2)催化使羟乙基-TPP-E1上的羟乙基被氧化成乙酰基,同时转移给硫辛酰胺,形成乙酰硫辛酰胺 -E2;二氢硫辛酰胺转乙酰酶(E2)还催化乙酰硫辛酰胺的乙酰基转移给CoA生成乙酰 CoA ,离开酶复合体,同时氧化过程中的2个电子使硫辛酰胺上的二硫键还原为 2个巯基;二氢硫辛酰胺脱氢酶(E3)使还原的二氢硫辛酰胺脱氢重新生成硫辛酰胺,以进行下一轮反应.同时将氢传递给FAD,生成 FADH2;在二氢硫辛酰胺脱氢酶(E3)催化下,将 FADH2上的H转移给 NAD+,形成 NADH+H+.

4、砷化物对硫辛酰胺的毒害作用5、丙酮酸脱氢酶系的调控( 1)产物控制 即由 NADH和乙酰 -CoA控制。这两种产物表现的抑制作用是和酶的作用底物 NAD+ 和 CoA竞争酶的活性部位,是竞争性抑制。乙酰 -CoA抑制 E2 , NADH抑制 E3 。如果 [NADH]/[NAD+]和 [乙酰 -CoA]/[CoA]的比值高, E2 则处于与乙酰基结合的形式,这时不可能接受在 E1 上与 TPP结合着的羟乙基基团,使 E1 上的 TPP停留在与羟乙基结合的状态,从而抑制了丙酮酸脱羧作用的进行。( 2)磷酸化和去磷酸化的调控E1 的磷酸化和去磷酸化是使丙酮酸脱氢酶复合体失活和激活的重要方式。在处于丙酮酸脱氢酶复合体核心位置的 E2 分子上结合着两种特殊的酶,一种称为激酶,另一种称为磷酸酶。激酶使丙酮酸脱氢酶组分磷酸化,磷酸酶则是脱去丙酮酸脱氢酶的磷酸基团,从而使之活化。 Ca2+通过激活磷酸酶的作用,也使丙酮酸脱氢酶活化。

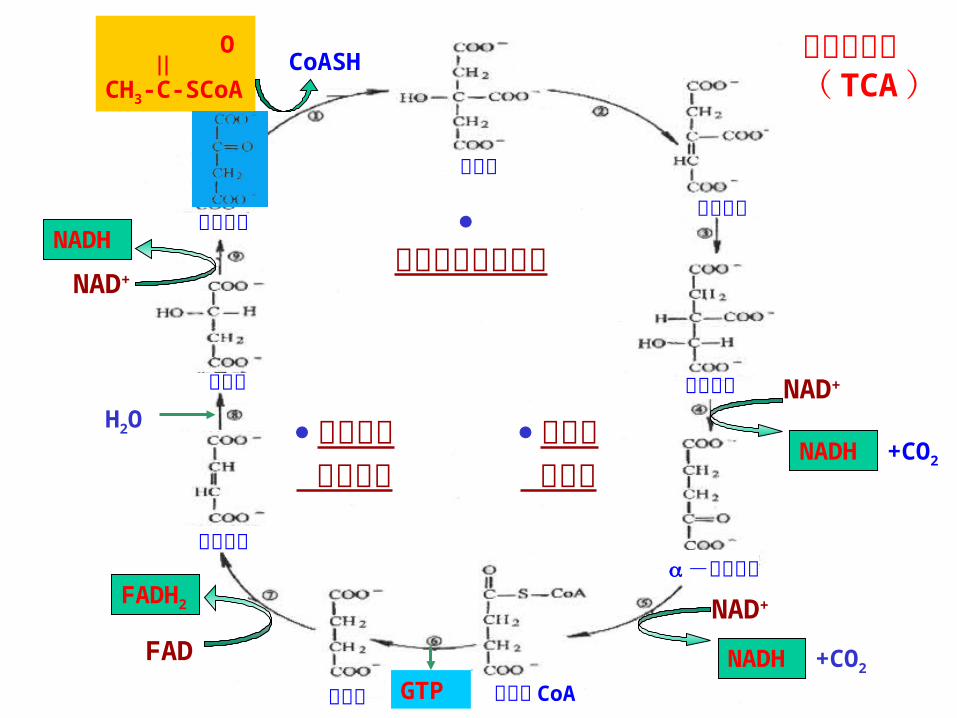
第三节 三羧酸循环( tricarboxylic acid cycle, TCA )
TCA 循环:指从乙酰 CoA与 OAA 缩合成柠檬酸,再经一系列氧化、脱羧,重新产生OAA 的循环过程,乙酰基则在循环中氧化成 CO2放出。(脱羧) Krebs 循环 1937年由德国科学家 Hans Krebs 提出, 1953年获诺贝尔奖。
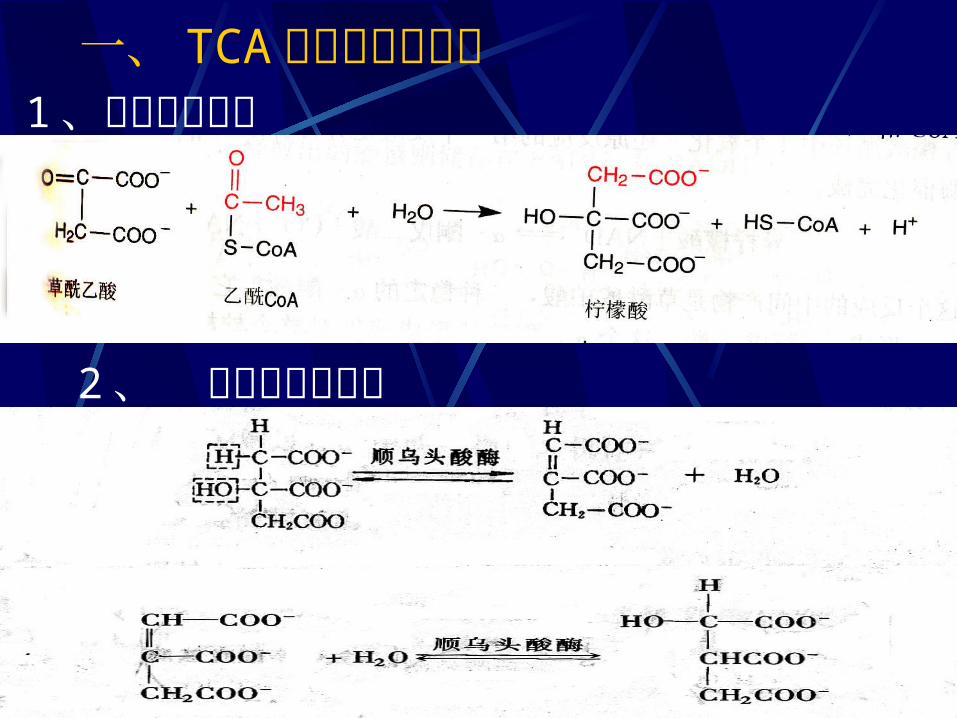
一、 TCA 循环的化学历程 1 、柠檬酸的合成
2 、 异柠檬酸的形成
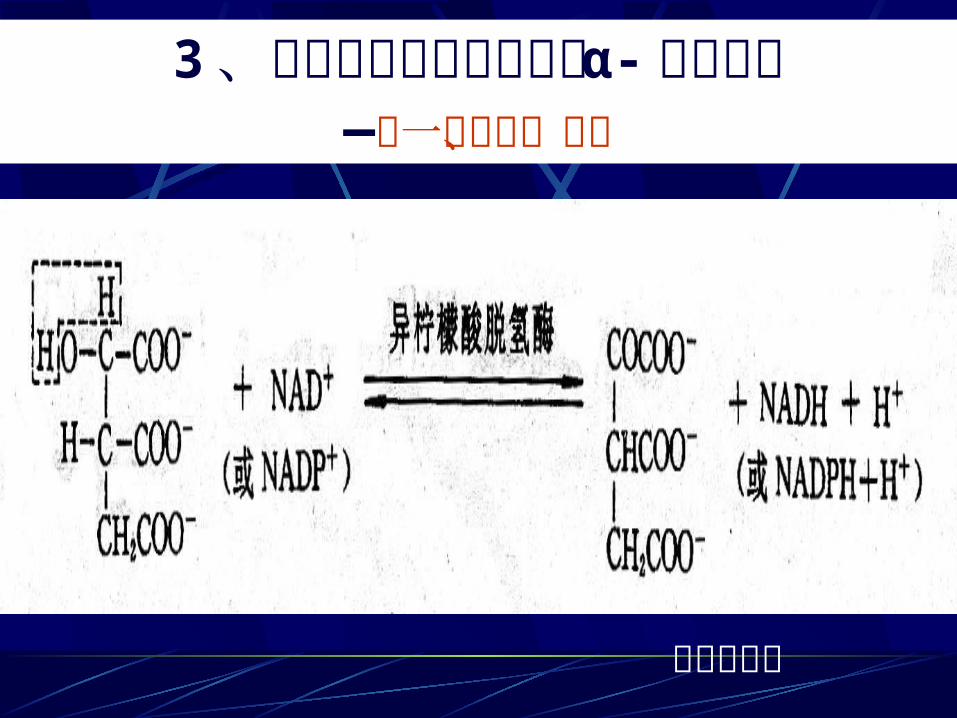
3、异柠檬酸氧化脱羧生成 α- 酮戊二酸—第一次脱氢、脱羧
草酰琥珀酸
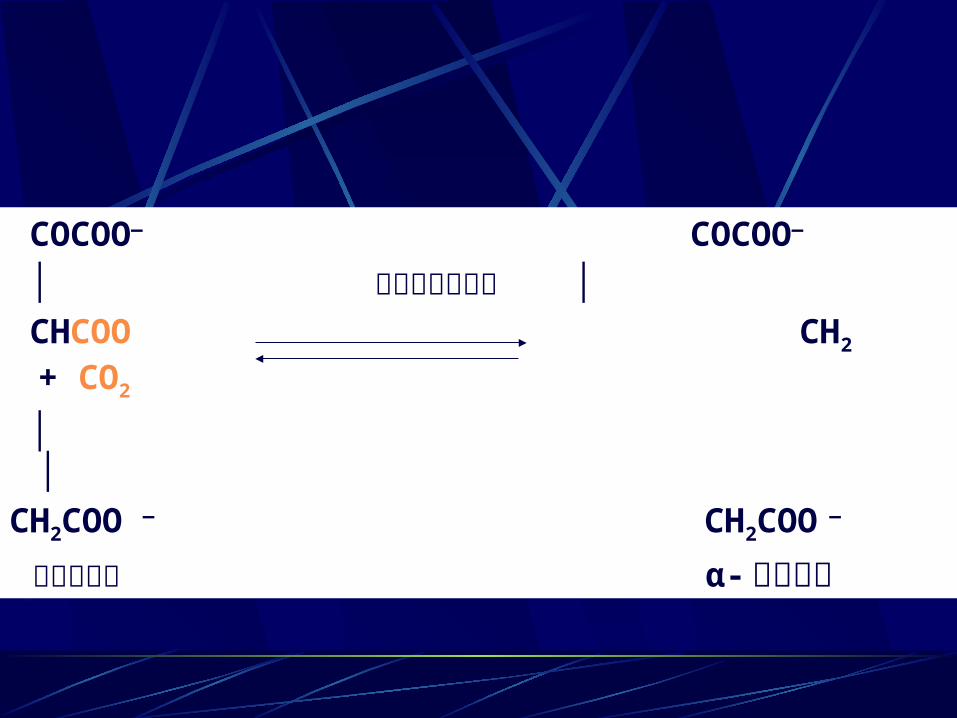
COCOO— COCOO—
│ 异 异柠檬酸脱氢酶 │ CHCOO CH2 + CO2
│ │CH2COO — CH2COO —
草酰琥珀酸 α- 酮戊二酸
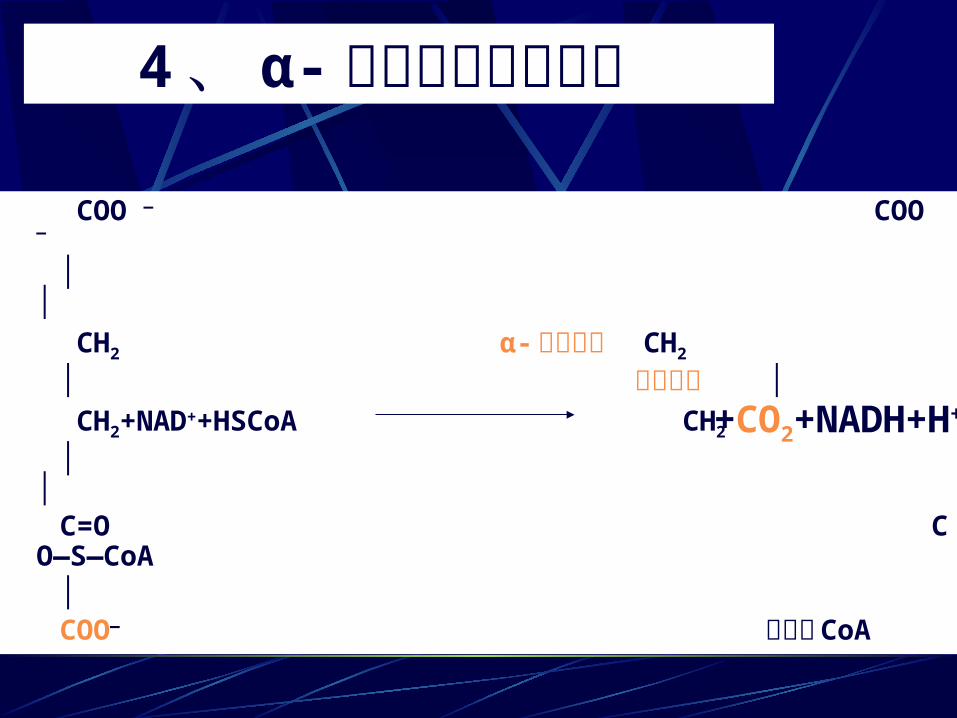
4、 α- 酮戊二酸氧化脱羧 COO — COO —
│ │ CH2 α- 酮戊二酸 CH2 │ 脱氢酶系 │ CH2+NAD++HSCoA CH2
│ │ C=O CO—S—CoA │ COO— 琥珀酰 CoA
+CO2+NADH+H+
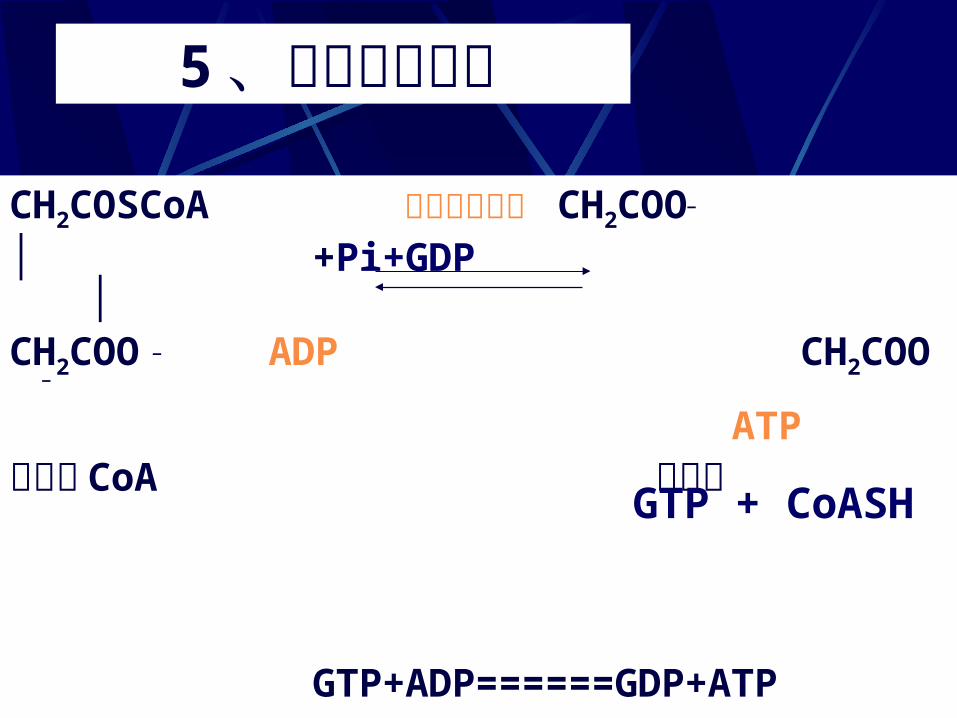
5、琥珀酸的生成 CH2COSCoA 琥珀酸硫激酶 CH2COO—
│ +Pi+GDP │ CH2COO — ADP CH2COO —
ATP琥珀酰 CoA 琥珀酸
GTP+ADP======GDP+ATP
GTP + CoASH
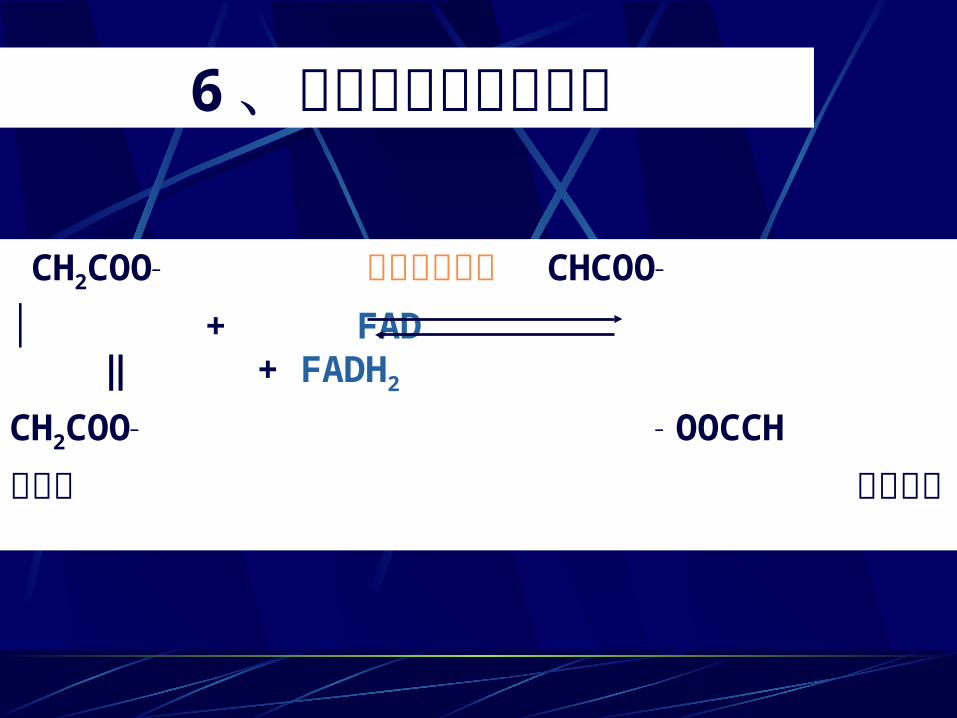
6、琥珀酸生成延胡索酸 CH2COO— 琥珀酸脱氢酶 CHCOO—
│ + FAD ‖ + FADH2
CH2COO— — OOCCH 琥珀酸 延胡索酸

7 、苹果酸的生成 COO— COO—
│ │ CH 苹果酸酶 HO—C—H ‖ + H2O │ HC H —C—H │ │ COO— COO—
延胡索酸 苹果酸
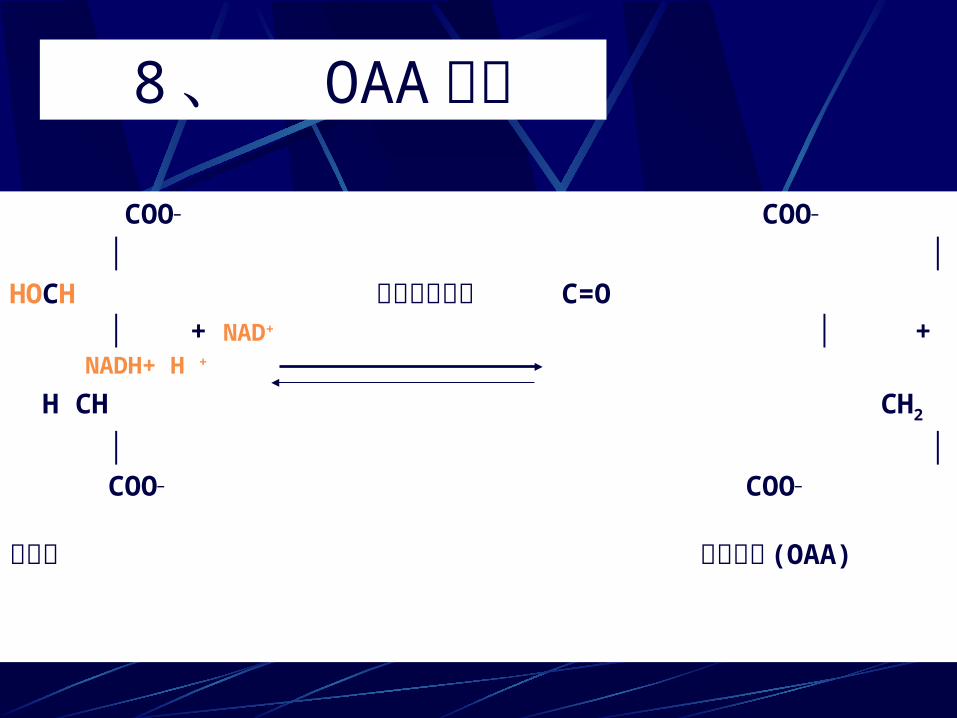
8、 OAA再生 COO— COO—
│ │HOCH 苹果酸脱氢酶 C=O │ + NAD+ │ + NADH+ H +
H CH CH2
│ │ COO— COO—
苹果酸 草酰乙酸 (OAA)
OCH3-C-SCoA
CoASH
NADH +CO2
FADH2
H2O
NADH
+CO2NADHGTP
三羧酸循环 ( TCA )
草酰乙酸 再生阶段
柠檬酸的生成阶段
氧化脱 羧阶段
柠檬酸
异柠檬酸
顺乌头酸
-酮戊二酸
琥珀酸 琥珀酰 CoA
延胡索酸
苹果酸
草酰乙酸
NAD+
NAD+
FADNAD+

总 结 1 、 TCA循环以乙酰 CoA 为底物,循环进行一周,消耗一分子乙酰 CoA ,经过二次脱羧,四次脱氢,一次底物水平磷酸化被完全分解;2 、总反应:CH3COSCOA+3NAD++FAD+ADP+Pi+2H2O
→2CO2+3NADH+3H + +FADH2+ATP
3 、细胞定位:线粒体基质中,整个循环单向进行。
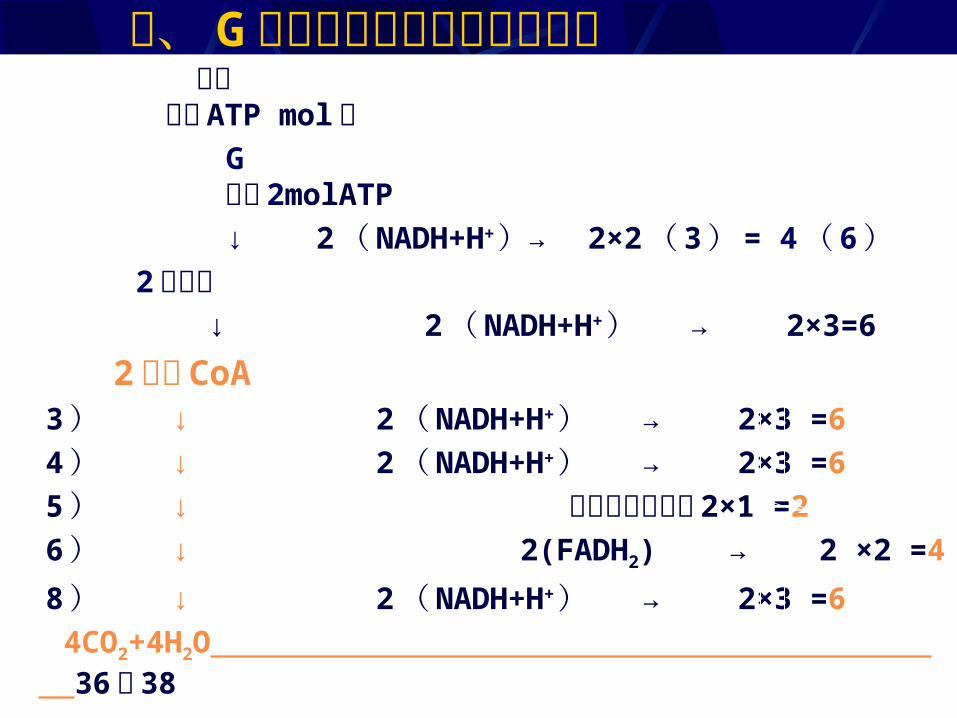
过程 产生 ATP mol数 G 净得 2molATP ↓ 2 ( NADH+H+ )→ 2×2 ( 3 ) = 4 ( 6 ) 2 丙酮酸 ↓ 2 ( NADH+H+ ) → 2×3=6 2 乙酰 CoA 3 ) ↓ 2 ( NADH+H+ ) → 2×3 =6 4 ) ↓ 2 ( NADH+H+ ) → 2×3 =6 5 ) ↓ 底物水平磷酸化 2×1 =2 6 ) ↓ 2(FADH2) → 2 ×2 =4 8 ) ↓ 2 ( NADH+H+ ) → 2×3 =6 4CO2+4H2O 36 或 38
二、 G 有氧分解的化学和能量计量

还原当量与 ATP 的转运通过线粒体内膜的物质转运还原当量的转运α- 磷酸甘油穿梭:脑、心肌、骨骼肌苹果酸 -天冬氨酸穿梭:脑、心肌、肝、红肌
ATP 、 ADP 、 Pi 的转运

线粒体外 NADH的氧化磷酸化作用
磷酸甘油穿梭系统 苹果酸—天冬氨酸穿梭系统
酵解(细胞质)
氧化磷酸化 (线粒体)
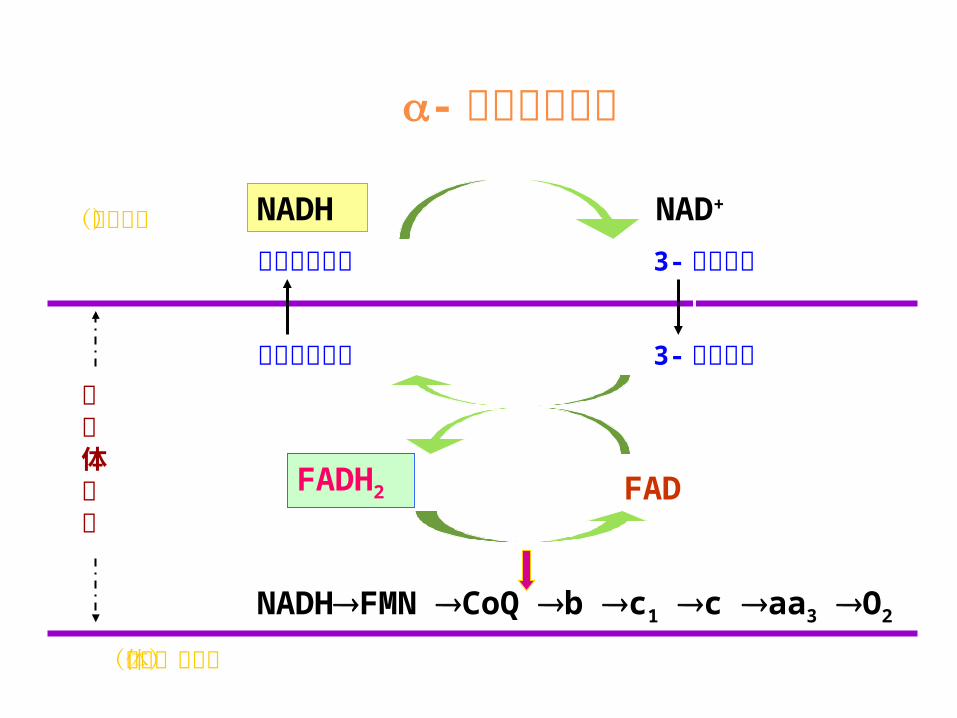
- 磷酸甘油穿梭
(线粒体基质)
磷酸二羟丙酮 3-磷酸甘油磷酸二羟丙酮 3-磷酸甘油
FADFADH2
NADHFMNCoQbc1caa3O2
NADH NAD+
线粒体内膜
(细胞液)
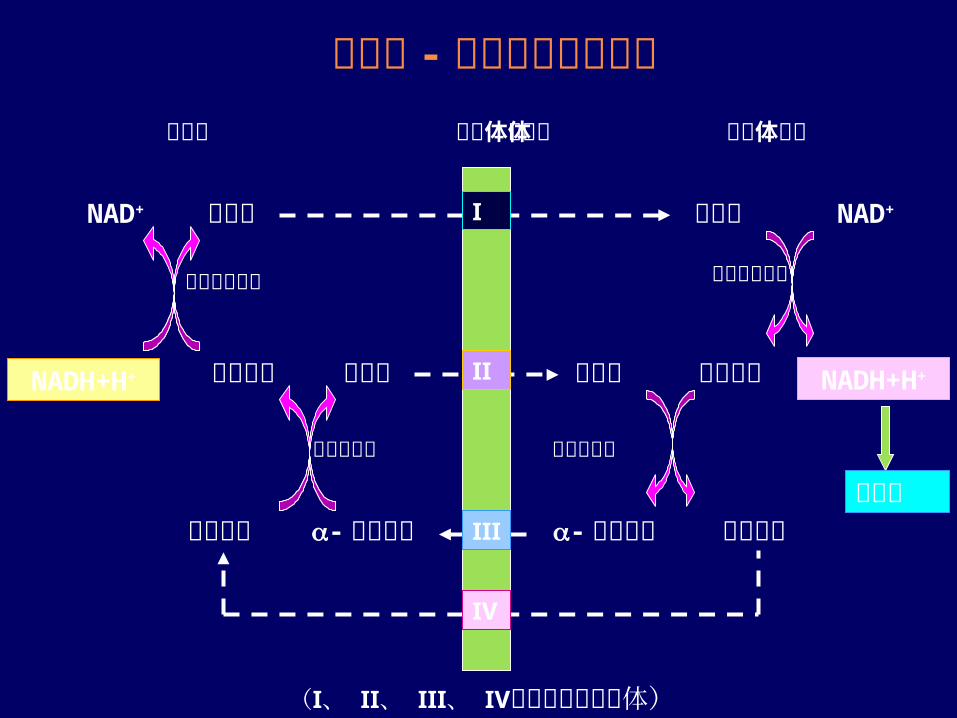
苹果酸 -草酰乙酸穿梭作用细胞液 线粒体内膜体
天冬氨酸-酮戊二酸
苹果酸
草酰乙酸 谷氨酸
-酮戊二酸天冬氨酸
苹果酸
谷氨酸NADH+H+
NAD+
草酰乙酸
NAD+
线粒体基质
苹果酸脱氢酶NADH+H+
Ⅳ
Ⅰ
Ⅱ
Ⅲ
苹果酸脱氢酶
谷草转氨酶 谷草转氨酶
(Ⅰ、 Ⅱ、 Ⅲ、 Ⅳ为膜上的转运载体)
呼吸链
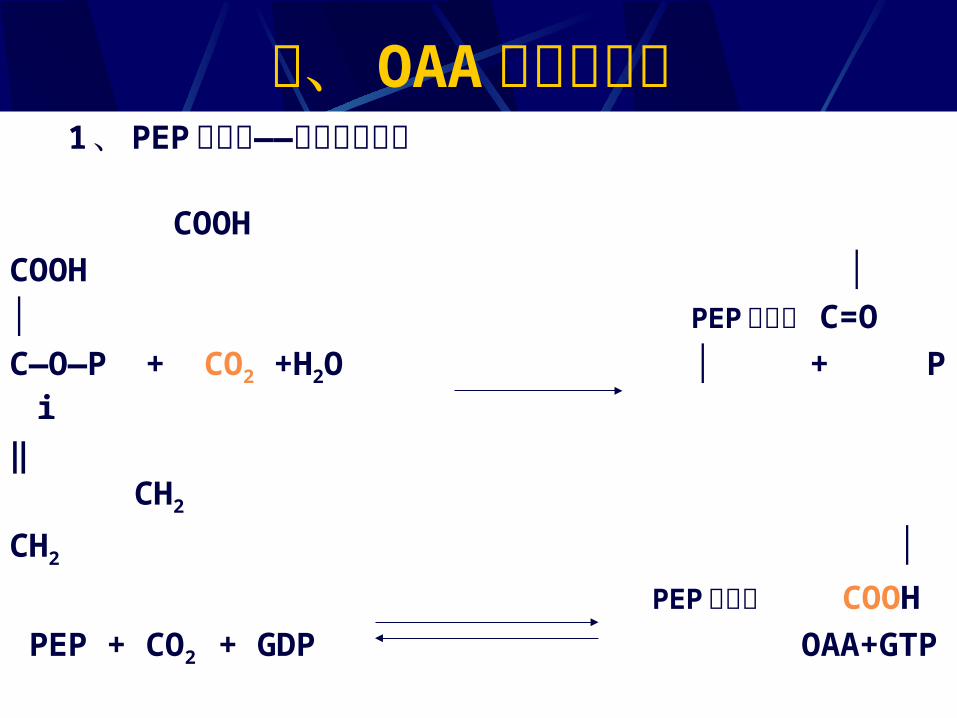
三、 OAA 的回补反应 1 、 PEP 的羧化——植物和细菌中 COOHCOOH ││ PEP 羧化酶 C=OC—O—P + CO2 +H2O │ + Pi‖ CH2
CH2 │
PEP 羧激酶 COOH PEP + CO2 + GDP OAA+GTP
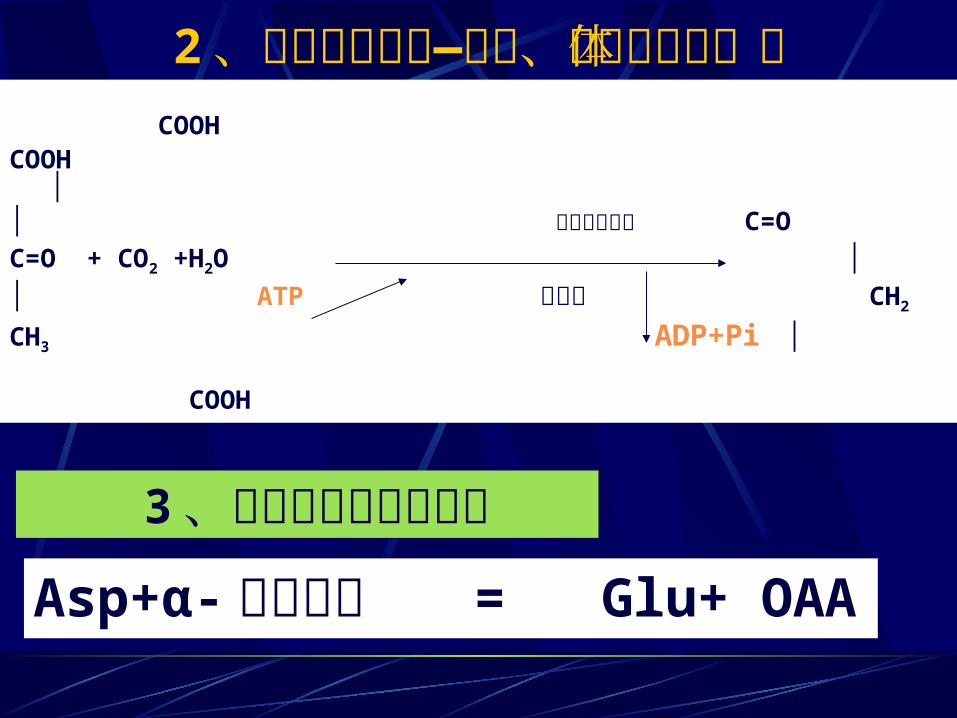
2 、丙酮酸的羧化—动物、酵母胞线粒体中 COOHCOOH │ │ 丙酮酸羧化酶 C=O C=O + CO2 +H2O ││ ATP 生物素 CH2
CH3 ADP+Pi │ COOH
3 、通过氨基酸转氨作用Asp+α- 酮戊二酸 = Glu+ OAA

四、 TCA循环的生物学意义1 、 TCA环是有机体获得生命活动所需能量的最主要途径。1 ) G 有氧分解中,每个 G通过 TCA环可得 24 个 ATP ,远远超过 EMP 或 G 无氧降解所产生ATP 的数目。2 )脂肪、氨基酸等有机物作为呼吸底物分解,彻底氧化时所产生的能量也主要通过 TCA循环。

2 、 TCA循环是物质代谢的枢纽1 ) TCA 是糖、脂肪、氨基酸等彻底分解的共同途径。2 ) TCA 中产生的 OAA 、 α- 酮戊二酸、柠檬酸、琥珀酰 CoA 和延胡索酸又是合成糖、氨基酸、脂肪酸、卟啉等的原料,因此能将各种有机物代谢联系起来,起到物质代谢的枢纽作用。3 、是发酵产物重新氧化的途径。

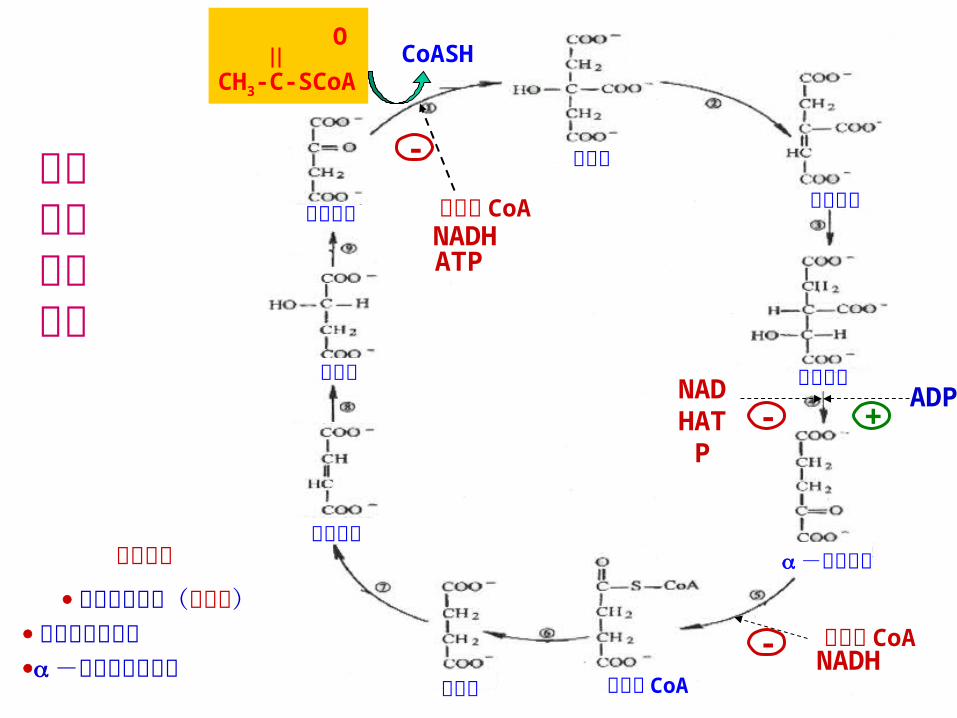
五、三羧酸循环的调控 调节三羧酸循环速度的关键酶:柠檬酸合酶;异柠檬酸脱氢酶;α- 酮戊二酸脱氢酶。
三羧酸循环中的酶的活性主要靠底物提供的情况推动,并受其生成产物的抑制,同时还受到变构效应物的调节。
OCH3-C-SCoA
CoASH三羧酸循环的调节
柠檬酸
异柠檬酸
顺乌头酸
-酮戊二酸
琥珀酸 琥珀酰 CoA
延胡索酸 调节位点 柠檬酸合成酶(限速酶)
异柠檬酸脱氢酶 -酮戊二酸脱氢酶
ADP+
NADHATP
-
琥珀酰 CoANADH
-
琥珀酰 CoANADHATP
-
苹果酸
草酰乙酸

柠檬酸合酶——柠檬酸和琥珀酰 CoA 分别是柠檬酸合酶的底物草酰乙酸和乙酰 CoA 的竞争性抑制剂,二者浓度的增加,抑制柠檬酸合酶的活性,另外,该酶还受到 NADH 的抑制。异柠檬酸脱氢酶(别构酶)——ADP 是别构激活剂,异柠檬酸、 NAD+等对酶的活性也有促进作用。NADH则抑制它的活性。 α- 酮戊二酸脱氢酶复合体——产物琥珀酰辅酶 A和 NADH抑制,也受高能荷抑制。当细胞内 ATP水平高时,可使循环速度减慢。
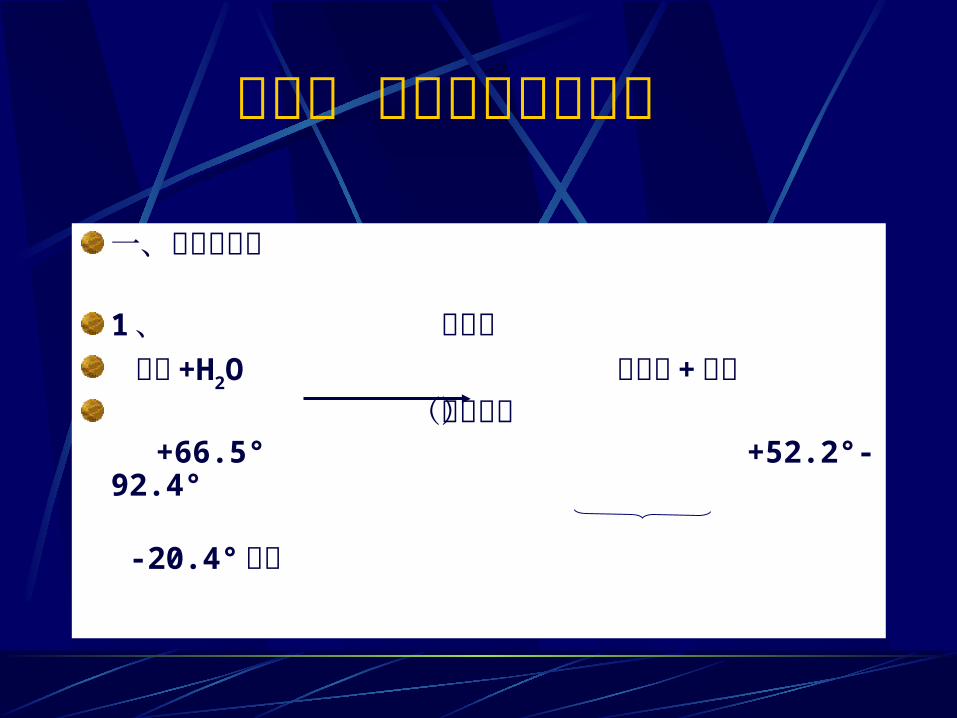
第四节 蔗糖和多糖的分解 一、蔗糖的水解1 、 蔗糖酶 蔗糖 +H2O 葡萄糖 + 果糖 (转化酶)
+66.5° +52.2°-92.4° -20.4°左右
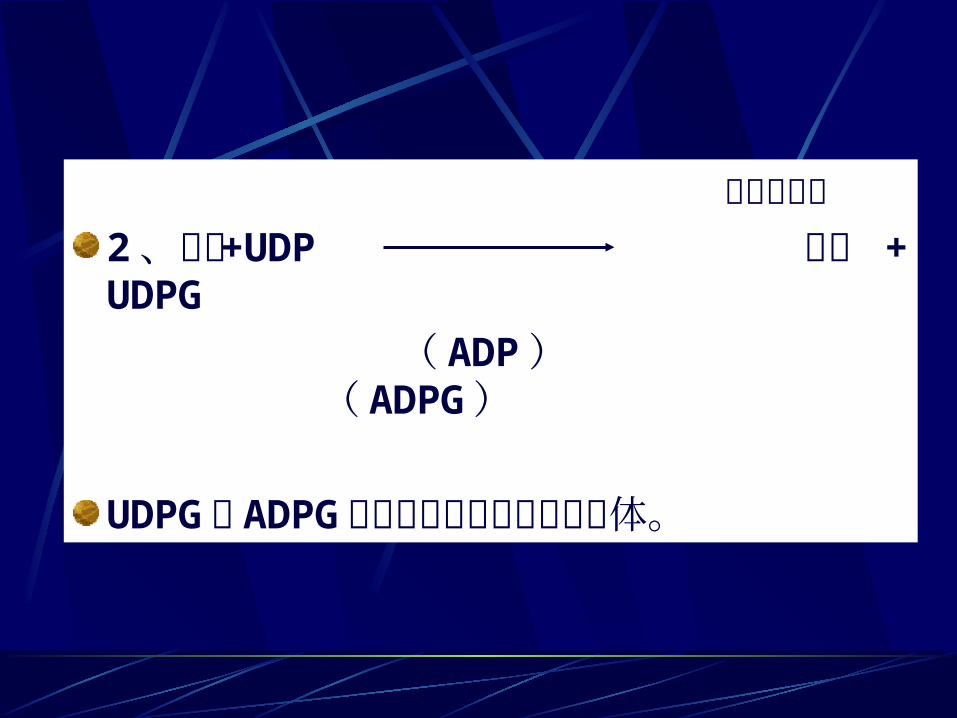
蔗糖合成酶2 、蔗糖 +UDP 果糖 +UDPG
( ADP ) ( ADPG )UDPG 和 ADPG常作为淀粉合成的糖基供体。

常见双糖的酶促降解
麦芽糖 +H2O 2 葡萄糖麦芽糖酶
乳糖 +H2O葡萄糖 +半乳糖--
β-半乳糖苷酶

淀粉(糖原)进入 EMP 的反应 ATP\/ ADP ↑( 磷酸己糖旁路或称磷酸戊糖途经 )
葡萄糖→葡萄糖 -6- 磷酸 = 果糖 -6- 磷酸→二磷酸果糖 ↓ ↑葡萄糖磷酸变位酶 P 酶 ↓淀粉 → 葡萄糖 -1- 磷酸 ↓(糖原) ↓ ( 糖醛酸途经 )

二、淀粉的降解 1 、淀粉的水解α-淀粉酶 (内切酶 )β-淀粉酶 (外切酶 )脱枝酶麦芽糖酶
2、淀粉的磷酸解淀粉磷酸化酶( P酶)
α-淀粉酶
β-淀粉酶
淀粉磷酸化酶 脱支酶淀粉 +nH3PO4 nG-1-p+少量葡萄糖

1 、淀粉的水解1 ) α-淀粉酶(液化酶)a )作用方式:可以任意水解淀粉分子内部 的 α-1,4- 糖苷键。b)终产物: 麦芽糖、葡萄糖(直链) 麦芽糖、葡萄糖、 α-极限糊精(枝链)c )性质:耐高温, t=40℃活性最大, 70℃仍稳定但不耐酸,当 pH≤3.3 即失活。

2 ) β-淀粉酶(外淀粉酶)a )作用方式 从淀粉非还原端顺次切下两分子葡萄糖(一个麦芽糖)也仅作用于 α-1,4 糖苷键。b)产物 β-麦芽糖(直链); β-麦芽糖 +β-极限糊精(枝链)。c) 性质 不耐高温( 70℃下酶蛋白变性而失活),但对酸度稳定 , pH=3.3 仍稳定。

3 ) 脱枝酶( R酶、异淀粉酶) 作用方式:水解 α-淀粉酶和 β-淀粉酶作用后留下的极限糊精中的 α-1,6- 糖苷键。 4 ) 麦芽糖酶( α- 葡萄糖苷酶) 作用方式:水解 α-1,4- 糖苷键
2 、淀粉的磷酸解淀粉磷酸化酶( P酶)
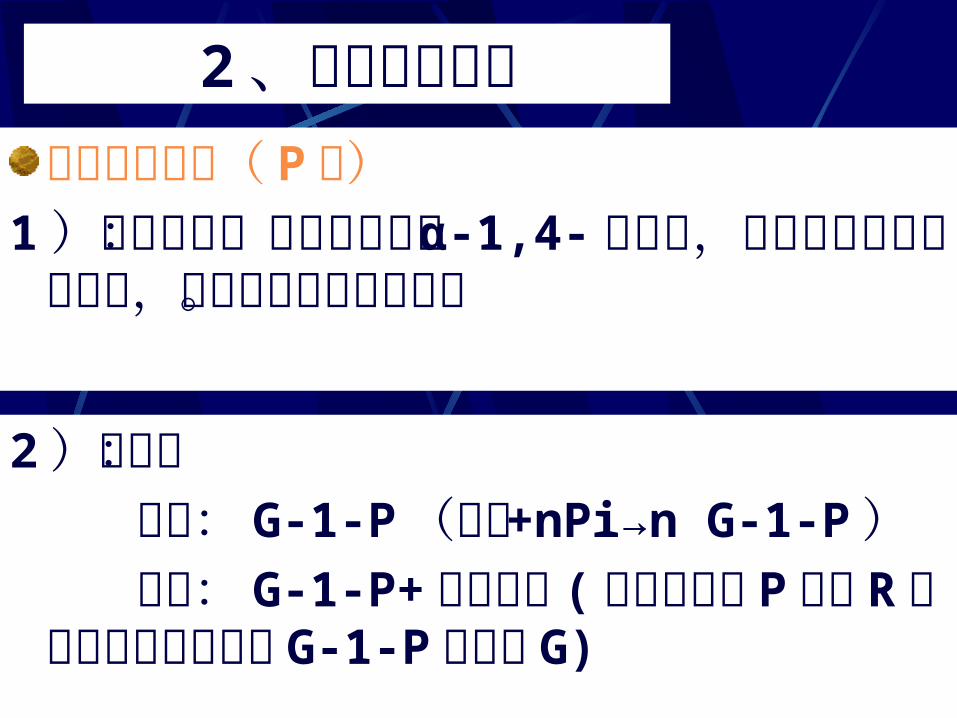
1 )作用方式:作用于淀粉的 α-1,4- 糖苷键,从直链的非还原端开始,逐个进行磷酸解反应。2)产物: 直链: G-1-P (淀粉 +nPi→n G-1-P ) 枝链: G-1-P+极限糊精 (极限糊精在 P酶和 R酶作用下继续降解为 G-1-P 和少量G))
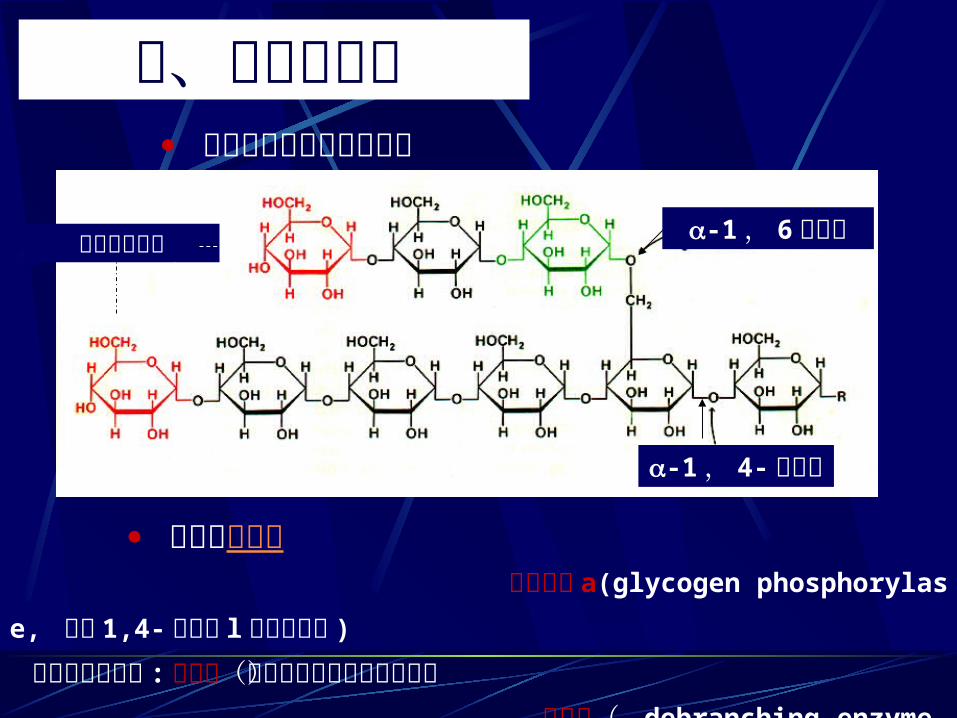
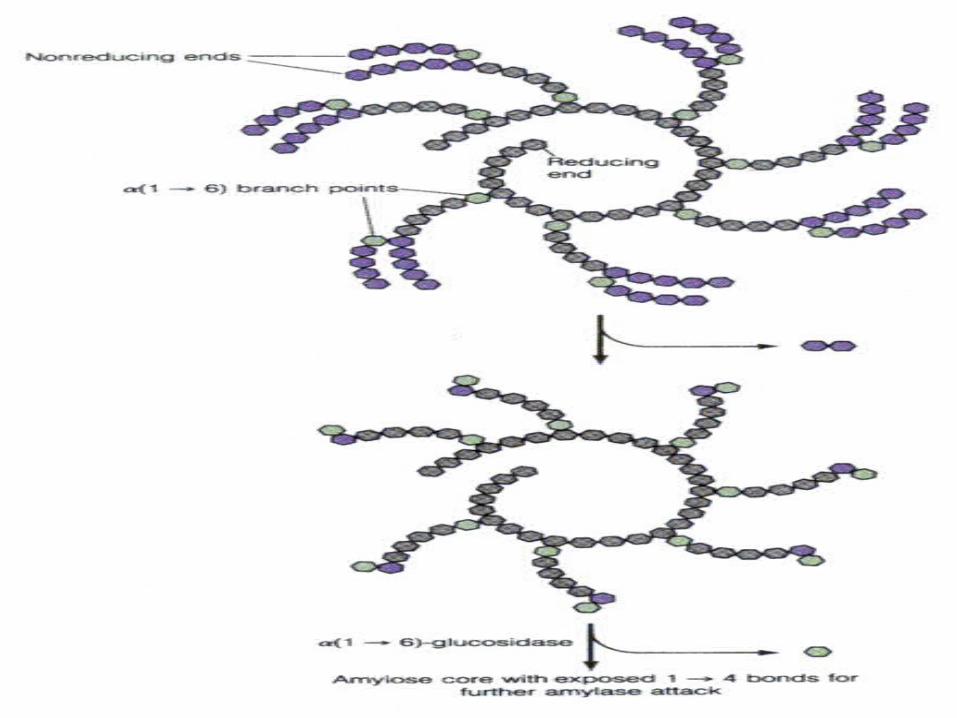
三、糖原的降解 糖原的结构及其连接方式
磷酸化酶 a(glycogenphosphorylase, 催化 1,4- 糖苷键 l磷酸解断裂 )
三种酶协同作用 :转移酶(催化寡聚葡萄糖片段转移) 脱枝酶( debranchingenzyme,催化 1,6-糖苷键水解断裂)
糖原的磷酸解
-1, 4-糖苷键
-1, 6糖苷键非还原性末端
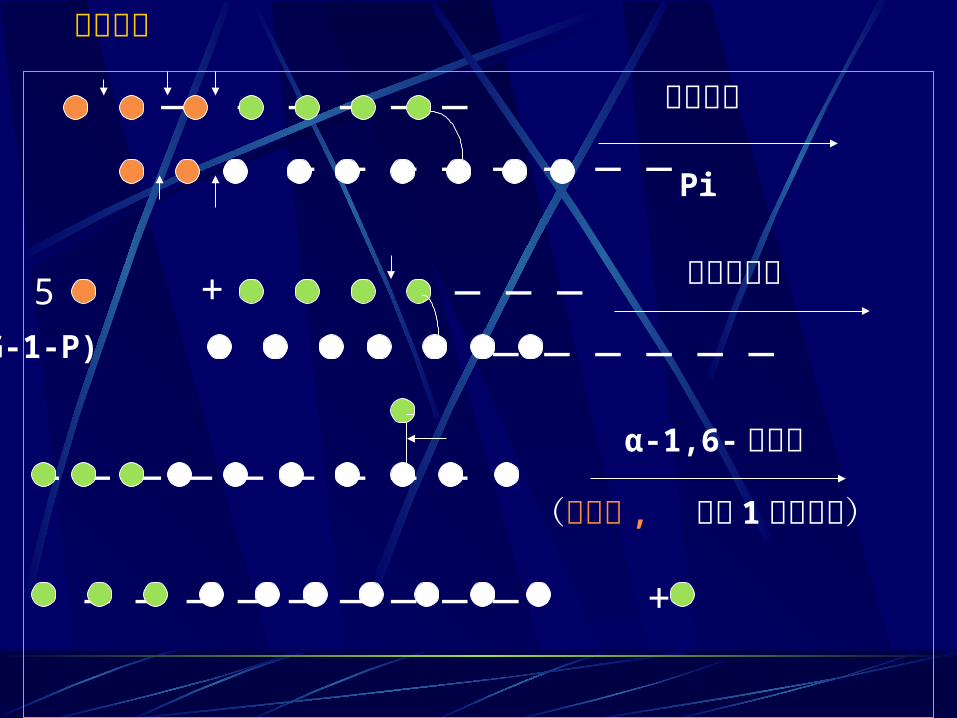
— — — — — — — — — — — — — —
5 + — — — — — — — — —
— — — — — — — — —
— — — — — — — — — +
磷酸化酶Pi
糖基转移酶
α-1,6- 糖苷酶
非还原端
(脱枝酶 ,释放 1个葡萄糖)
(G-1-P)
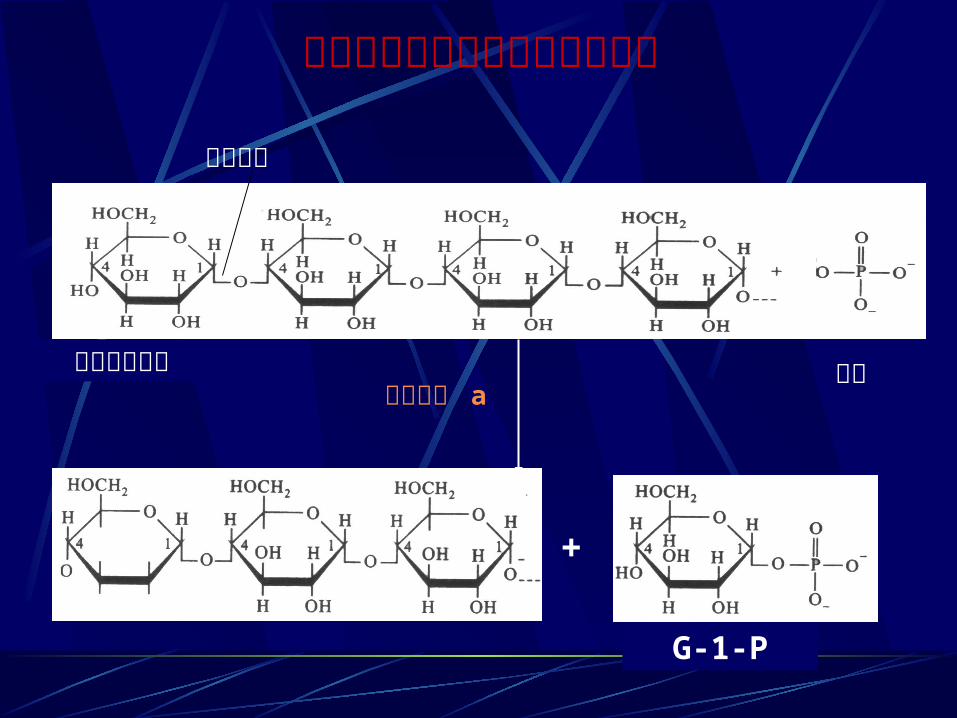
糖原磷酸化酶的作用位点及产物
G-1-P
磷酸化酶 a非还原性末端 磷酸
+
断键部位

其它糖进入单糖分解的途径半乳糖
半乳糖 -1-P
UDP-半乳糖UDP-葡萄糖
葡萄糖 -1-磷酸
糖原或淀粉
葡萄糖
葡萄糖 -6-磷酸
果糖
葡萄糖
果糖 -6-磷酸 果糖 -1 、 6-磷酸
磷酸二羟丙酮
3-磷酸甘油
甘油
3-磷酸甘油醛
进入糖酵解甘露糖
甘露糖 -6-磷酸
ATP
ADP
ATP ADP
ATP
ADP
ATP
ADP
ATP
ADP
ATP
ADP
NADH+H+
NAD+
Pi
UTP
PPi

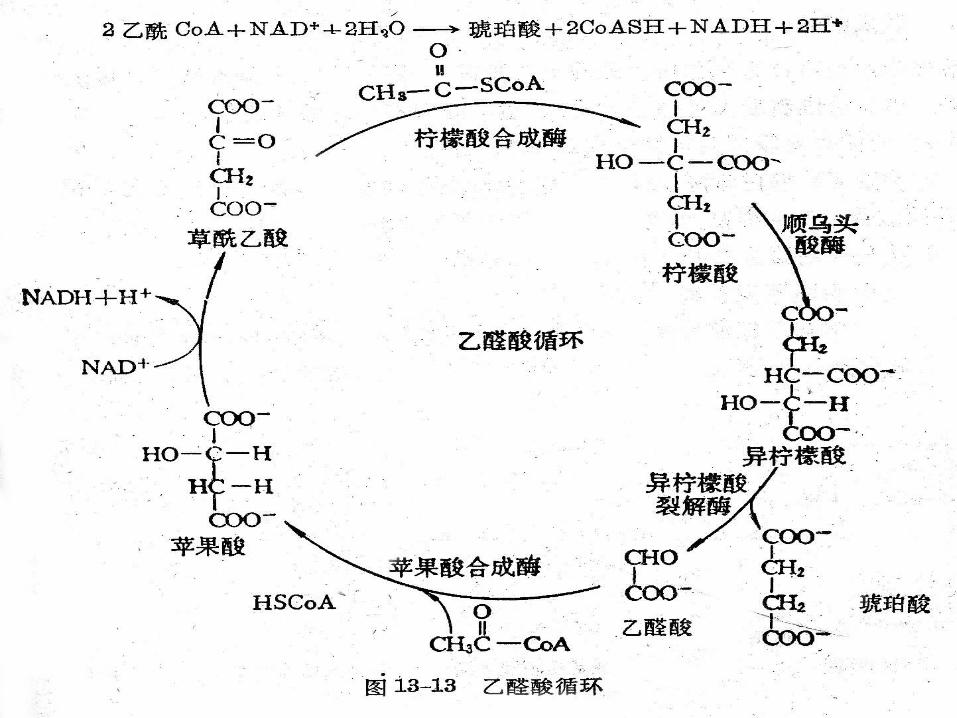
第五节 乙醛酸循环途径 该循环存在于植物细胞的乙醛酸循环体,动物细胞不存在乙醛酸体,故不存在乙醛酸循环。 一、乙醛酸循环的化学过程 乙醛酸循环共有五个反应,其中包括二个关键性酶:异柠檬酸裂解酶和苹果酸合成酶。
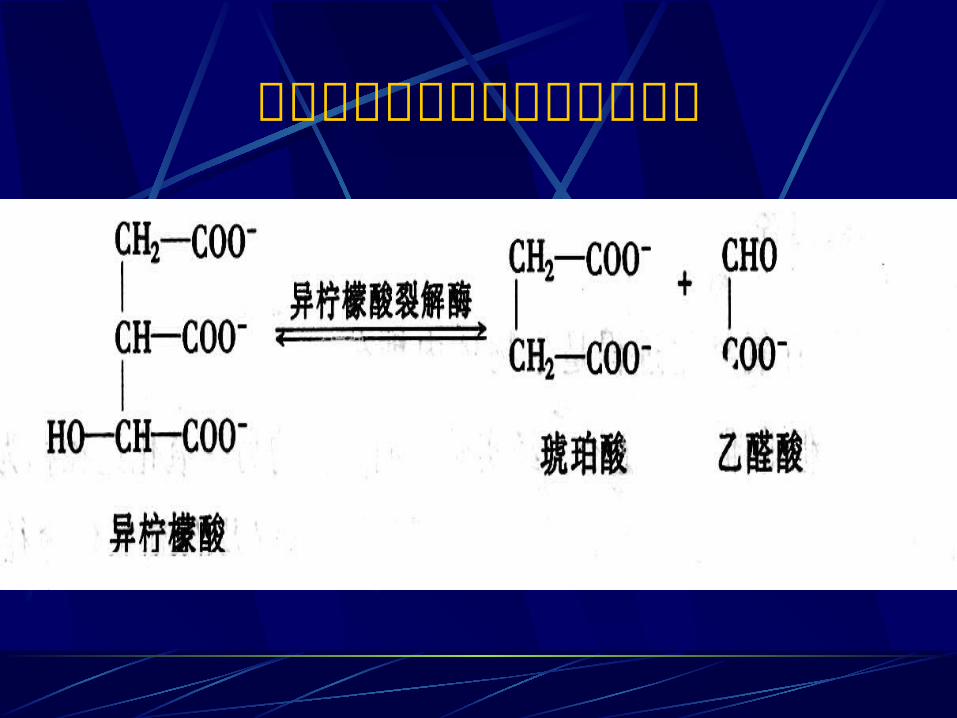
异柠檬酸裂解为琥珀酸和乙醛酸
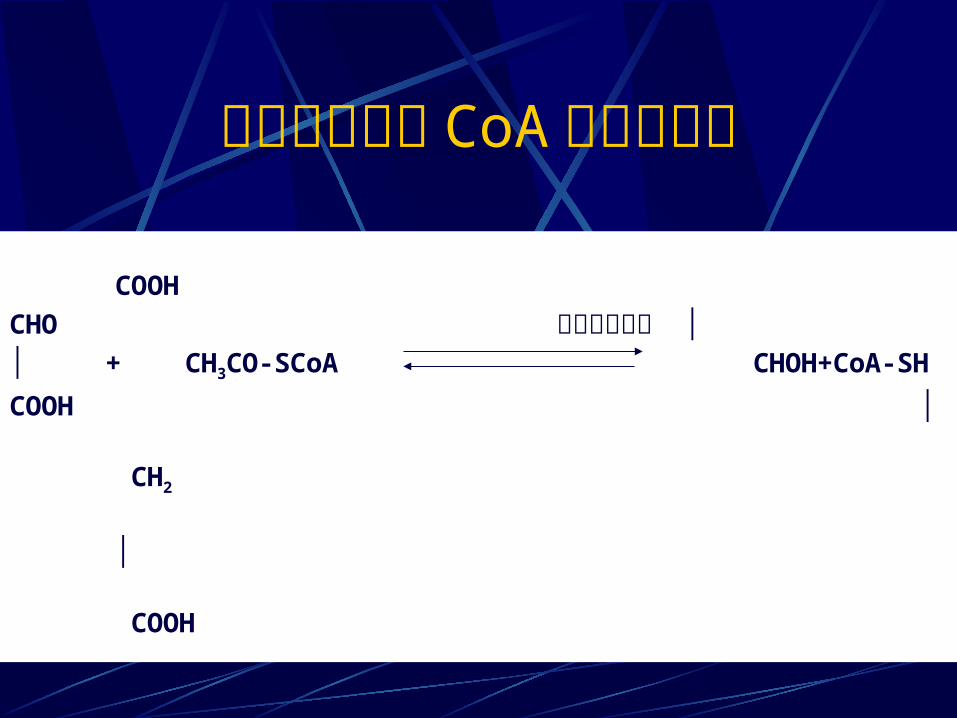
乙醛酸和乙酰 CoA 合成苹果酸 COOHCHO 苹果酸合成酶 ││ + CH3CO-SCoA CHOH+CoA-SHCOOH │ CH2
│ COOH

二、与 TCA循环的比较

1 、 TCA循环的综合效果是 1摩尔 CH3COSCoA经过 TCA环完全氧化成 CO2+H2O 。
2 、乙醛酸循环的综合效果是 2摩尔 CH3COSCoA→ 变成四碳二羧酸(琥珀酸),也就是说乙酰 CoA经过乙醛酸循环,乙酰基不但保存下来,而且有净合成。

乙醛酸循环的总反应如下: 2CH3CO-SCoA+2H2O+NAD+→琥珀酸 +2CoASH+NADH+H+
联系:由乙醛酸循环合成的琥珀酸可进入 TCA环作为 TCA环中间产物的补充方式。

三、乙醛酸循环的生物学意义1 、可以以二碳物为起始物合成 TCA环中的二羧酸和三羧酸,作为三羧酸循环上化合物的补充—回补反应(不是主要的)。2 、某些以乙酸、乙醇为营养物质的微生物可利用乙酸、乙醇作为唯一碳源通过乙醛酸循环合成机体所需的能源和碳源;植物体可利用脂肪降解的产物乙酰 CoA通过乙醛酸循环转变成糖。
第六节 磷酸戊糖途径 磷酸戊糖途径( PPP )又称磷酸已糖旁路( HMS ),该途径特点是 G 从 G-6-P出发经脱氢生成 6-P- 葡萄糖酸→ CO2+ 磷酸戊糖。

一、磷酸戊糖途径的发现(二个实验) G→3-P- 甘油醛 ==========1,3- 二磷酸甘油酸→丙酮酸 3-P- 甘油醛脱氢酶( -SH ) ↓碘乙酸或氟化物 失活 (EMP 中断 ) 按推理, EMP被中断, G 则不被消耗;然而,可 G仍被消耗,证明G 还有其他代谢途径。
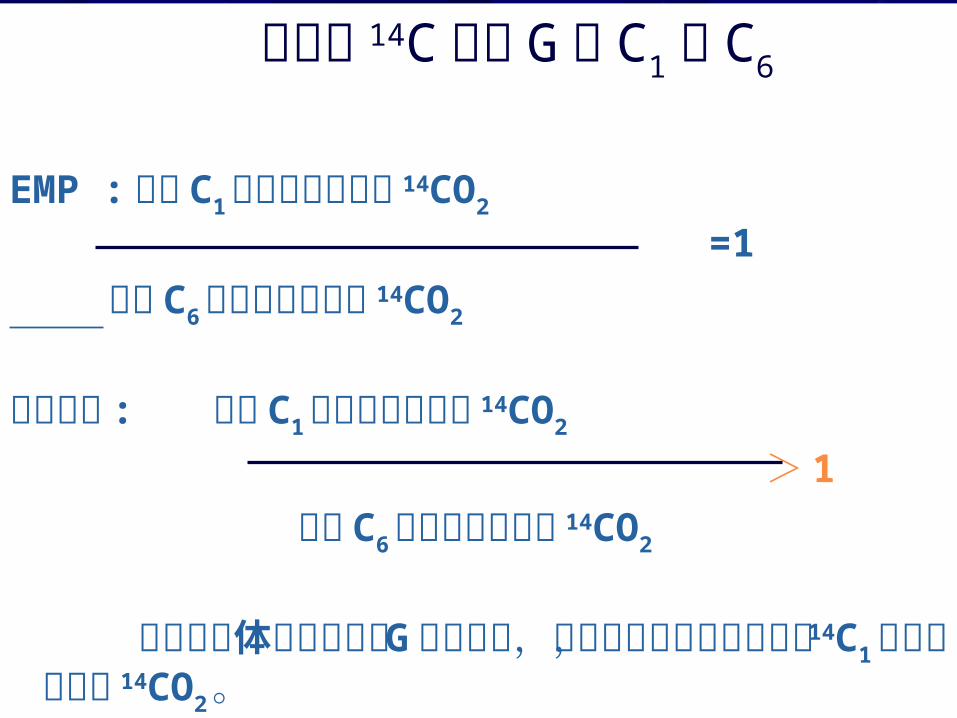
同位素 14C标记 G 的 C1 和 C6
EMP :标记 C1葡萄糖所释放的 14CO2 =1 标记 C6葡萄糖所释放的 14CO2
实验表明 : 标记 C1葡萄糖所释放的 14CO2 > 1 标记 C6葡萄糖所释放的 14CO2
说明生物体还有其他的 G氧化途径,且在其它的氧化途径中, 14C1更容易氧化成 14CO2。
二、磷酸戊糖途径的化学历程1 、磷酸葡萄糖氧化脱羧1 ) 6- 磷酸葡萄糖脱氢( NADP+ 为 H 受体)2 ) 6- 磷酸葡萄糖酸的生成3 ) 5- 磷酸核酮糖的生成( NADP+ 为 H 受体)2 、磷酸已糖的再生1 )磷酸戊糖的相互转化2 )转酮反应3 )转醛反应4 )转酮反应
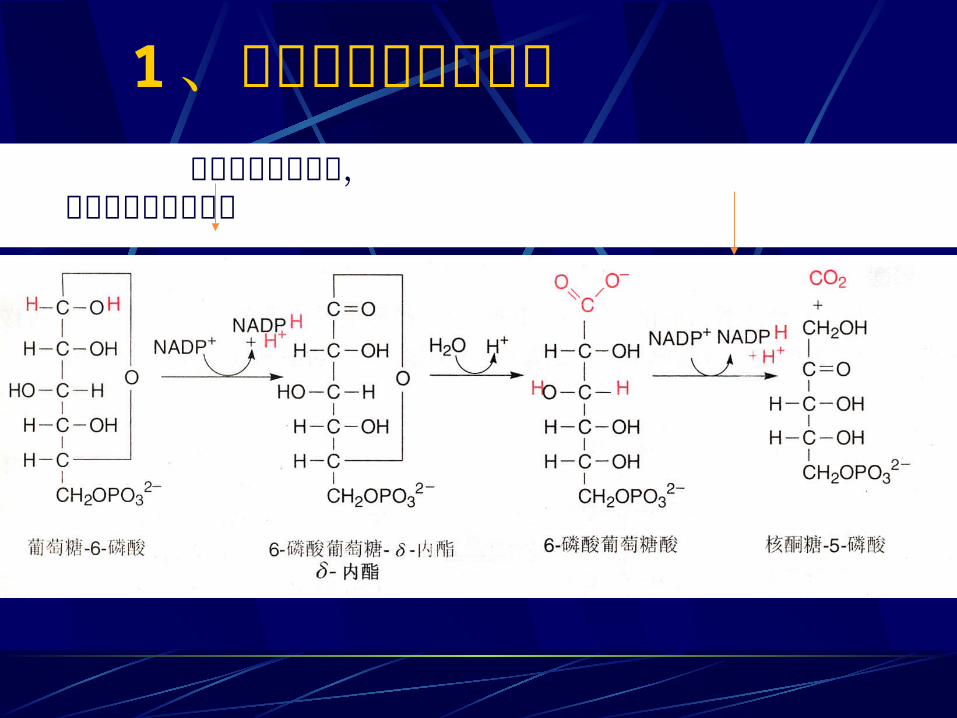
1 、磷酸葡萄糖氧化脱羧 磷酸葡萄糖脱氢酶, 磷酸葡萄糖酸脱氢酶
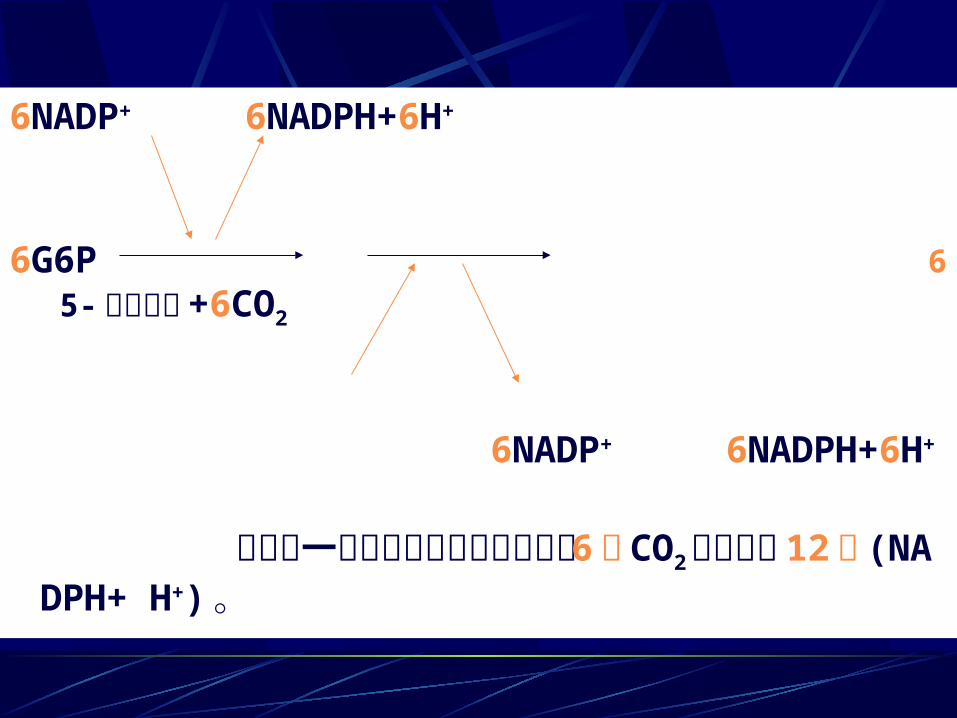
6NADP+ 6NADPH+6H+
6G6P 6 5- 磷酸核酮 +6CO2
6NADP+ 6NADPH+6H+
相当于一个葡萄糖被彻底氧化放出 6 个 CO2 同时生成 12 个 (NADPH+ H+) 。
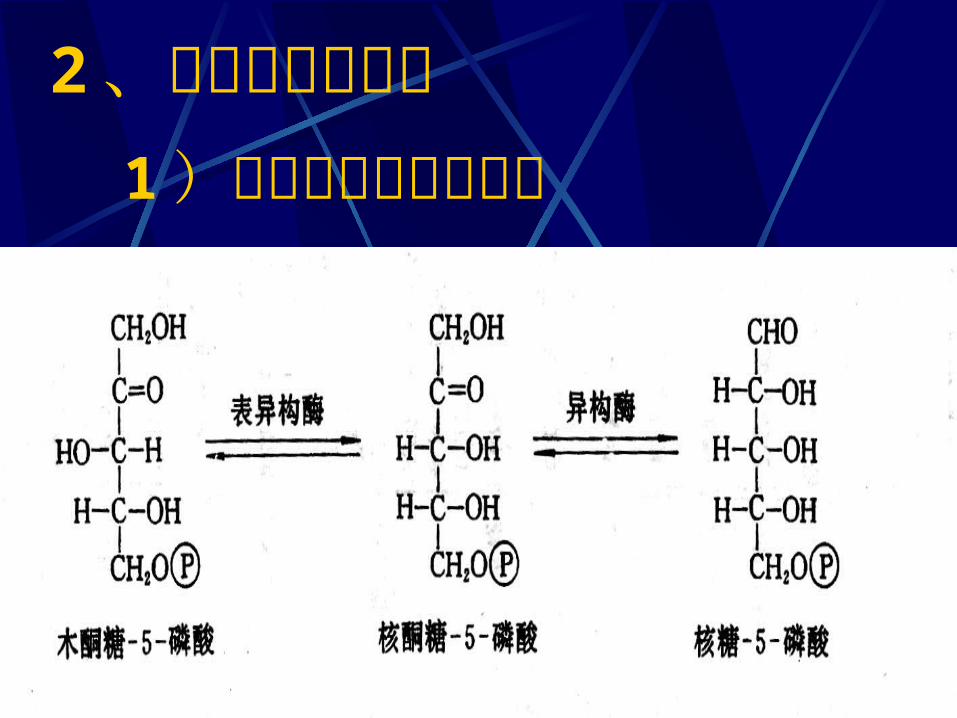
1 )磷酸戊糖的相互转化2 、磷酸已糖的再生
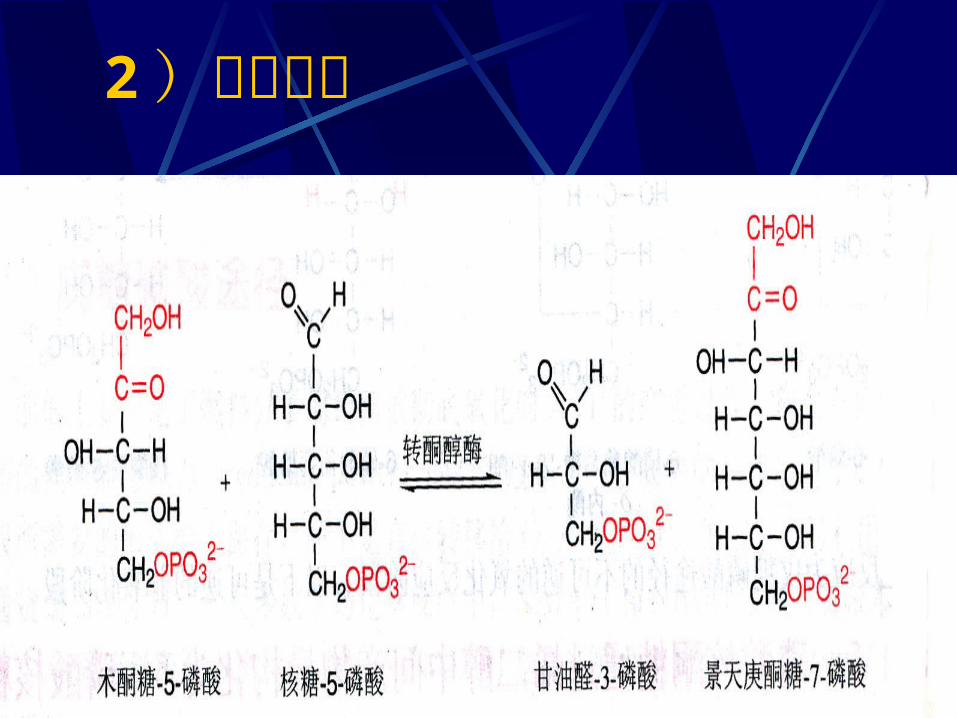
2 )转酮反应
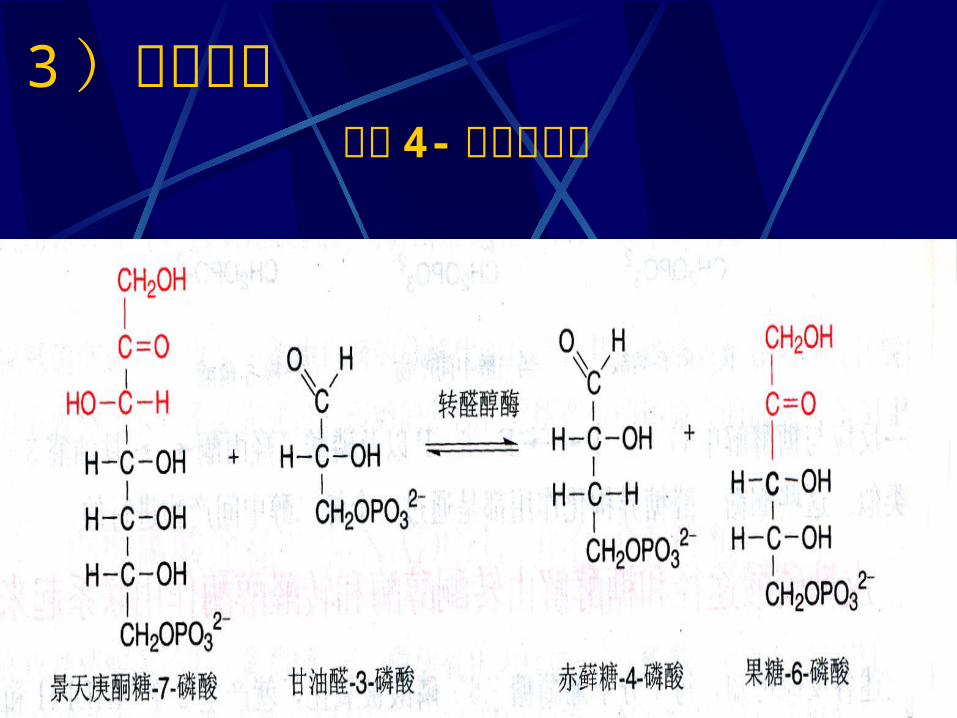
3 )转醛反应 生成 4- 磷酸赤藓糖
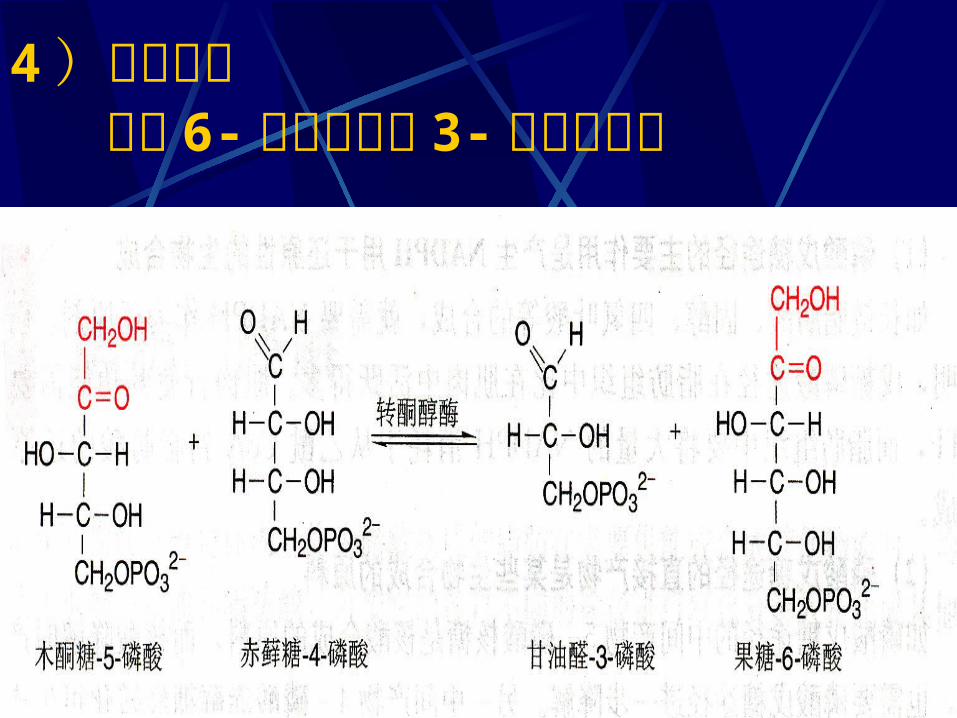
4 )转酮反应 生成 6- 磷酸果糖及 3- 磷酸甘油醛
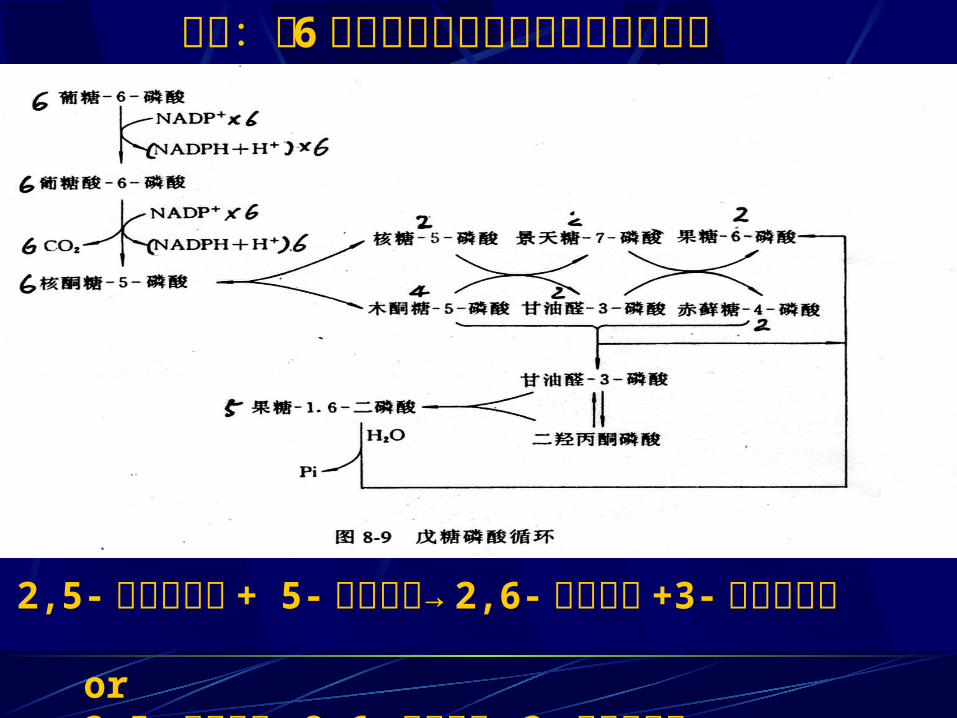
小结:以 6 分子葡萄糖为例说明磷酸戊糖途径
2,5-磷酸木酮糖 + 5-磷酸核糖→ 2,6-磷酸果糖 +3-磷酸甘油醛or3,5-磷酸核糖→ 2,6-磷酸果糖 +3-磷酸甘油醛

三、 PPP 途径的特点 1 ) G 直接脱氢和脱羧,不经过 TCA 或 EMP 途径2 )脱氢酶的辅酶为 NADP+ 而不是 NAD+
四、 PPP 途径的细胞定位细胞质中动物的肝脏中、骨髓、脂肪组织、肾上腺皮质、红细胞等组织中。

五、 PPP 途径的生物学意义 1 ) PPP 中产生的 NADPH ,供给组织中合成代谢的需要, NADPH 是合成脂肪酸、胆固醇、四氢叶酸等的供 H 体,也是 NO3
- 、 NO3- 还原, NH3 同化等反应所需的供 H 体——细胞的各种合成反应提供主要还原力。
2 ) PPP 途径中的中间物是各种生物合成的原料3 )在植物中,已糖再生阶段的一系列中间产物与酶同光合作用中卡尔文循环的大多数中间产物和酶相同,因此 PPP 可与光合作用联系起来,实现某些单糖间的互变。4 ) NADPH+H+ 有时也可经呼吸链氧化供能。

六、戊糖磷酸途径的调节葡萄糖 -6- 磷酸脱氢酶为戊糖磷酸途径的限速酶其活性受 NADP+/NADPH比例的调节 在戊糖磷酸途径的氧化阶段,葡萄糖 -6- 磷酸脱氢酶活性最低,是途径的限速酶, NADPH 竞争性地抑制葡萄糖 -6- 磷酸脱氢酶活性,非氧化阶段无调控酶,反应速度受控于底物浓度。

第七节 糖类的合成 一、葡萄糖的合成(光合作用、葡萄糖异生)二、葡萄糖核苷酸的合成三、蔗糖的生物合成四、淀粉的合成五、糖原的合成

一、糖异生反应 糖异生概念:非糖物质 ( 乳酸 , 甘油 ,某些氨基酸 )转变成糖的过程 (gluconeogenesis) 。部位:肝、肾糖异生前体:凡是能生成丙酮酸的物质均可以转变成葡萄糖。乳酸、三羧酸循环的中间物,转变成丙酮酸、 α- 酮戊二酸、草酰乙酸的氨基酸,脂肪水解产生的甘油等。

--- 糖酵解中三个不可逆反应须由其他反应完成
– PA PEP
– F-1,6-BP F-6-P
– G-6-P G
( 一 ) 糖异生反应历程

糖异生途径:丙酮酸 葡萄糖
A. A. 丙酮酸羧化支路丙酮酸羧化支路B. 1,6-B. 1,6- 二磷酸果糖转变成二磷酸果糖转变成 6-6- 磷酸果糖磷酸果糖C. 6-C. 6- 磷酸葡萄糖水解为葡萄糖磷酸葡萄糖水解为葡萄糖

糖异生主要途径和关键反应
非糖物质转化成糖代谢的中间产物后,在相应酶催化下 ,绕过糖酵解途径的三个不可逆反应 , 利用糖酵解途径其它酶生成葡萄糖的途径称为糖异生。
糖原(或淀粉)1-磷酸葡萄糖
6-磷酸果糖
1, 6-二磷酸果糖3-磷酸甘油醛磷酸二羟丙酮
2 磷酸烯醇丙酮酸
2 丙酮酸
葡萄糖己糖激酶
果糖激酶二磷酸果糖磷酸酯酶
丙酮酸激酶丙酮酸羧化酶
6-磷酸葡萄糖磷酸酯酶6-磷酸葡萄糖
PEP羧激酶2草酰乙酸

糖异生途径(反应 1、 2)
CHCH22
C OC OCOOHCOOH
丙酮酸丙酮酸
COO-COO-C OC OCHCH22
COOHCOOH
COOCOO- - OO
C-O P-OC-O P-O--
CHCH2 2 OO--COCO22 ATP ADPATP ADP ++ PiPi
草酰乙酸草酰乙酸 磷酸烯醇磷酸烯醇型丙酮酸型丙酮酸
丙酮酸丙酮酸羧化酶羧化酶磷酸烯醇型磷酸烯醇型丙酮酸羧激丙酮酸羧激酶酶
GTP GDPGTP GDP COCO2 2
(1)(1) (2)(2)
生物素生物素

线粒体
丙酮酸
丙酮酸
草酰乙酸
苹果酸苹果酸
草酰乙酸
磷酸烯醇式丙酮酸
TCANADH+H+
NAD+NAD+
NADH+H+
GTP
CO2+GDP
丙酮酸羧化酶
PEP羧激酶
天冬氨酸GOT
Asp

糖异生途径(反应 3)
6-6- 磷酸果糖磷酸果糖
1,6-1,6- 二磷酸果糖二磷酸果糖
ATATPP
ADPADP
6-6- 磷酸果磷酸果糖激酶糖激酶 -1-1果糖双磷果糖双磷酸酶酸酶 -1-1PiPi
(3)(3)

二磷酸果糖磷酸酯酶+H2O +Pi
1,6-二磷酸果糖
PPO
H2COH2CO
HO
OH
HOHH
H
HH2CO
OH
6-磷酸果糖
PO H2CO
HO
OH
HH
H

糖异生途径(反应 4)
葡萄糖葡萄糖
6-6- 磷酸葡磷酸葡萄糖萄糖
ATATPP
ADPADP
6-6- 磷酸葡磷酸葡萄糖酶萄糖酶PiPi
(3)(3) 己糖激酶己糖激酶(葡萄糖激酶)(葡萄糖激酶)

+H2O +Pi
6-磷酸葡萄糖磷酸酯酶P
6-磷酸葡萄糖
H
葡萄糖

葡萄糖葡萄糖 6-6- 磷酸磷酸葡萄糖葡萄糖 6-6- 磷酸磷酸果糖果糖 1,6-1,6- 二磷二磷酸果糖酸果糖磷酸二磷酸二羟丙酮羟丙酮 3-3- 磷酸磷酸甘油醛甘油醛
磷酸烯醇磷酸烯醇型丙酮酸型丙酮酸
丙酮酸丙酮酸三羧酸三羧酸循环循环
乙酰乙酰 CoACoA苹果酸苹果酸 苹果酸苹果酸
草酰乙酸草酰乙酸草酰乙酸草酰乙酸线 粒 体线 粒 体
丙酮酸丙酮酸
(1)(1)(2)(2)
(3)(3)(4)(4)
糖 异 生 途 径糖 异 生 途 径NADNADHH+++NADH+NADH

糖糖酵酵解解和和葡葡萄萄糖糖异异生生的的关关系系
A
B
C1
C2
(胞液)(线粒体)
葡萄糖
丙酮酸草酰乙酸天冬氨酸
磷酸二羟丙酮3-P-甘油醛
-酮戊二酸乳酸
谷氨酸 丙氨酸
TCA循环乙酰 CoA
PEP
G-6-P
F-6-P
F-1.6-P
丙酮酸草酰乙酸谷氨酸-酮戊二酸
天冬氨酸
3-P-甘油 甘油
AG-6-P磷酸酯酶(葡萄糖 -6- 磷酸酶)
BF-1.6-P磷酸酯酶(果糖二磷酸酶 )
C1 丙酮酸羧化酶C2PEP羧激酶
回 顾

糖异生耗能2 丙酮酸 葡萄糖
– 2 丙酮酸 2PEP: 2 ATP×2=4
– 2 3-P- 甘油 酸 21,3-BP- 甘油酸: 1×2=2
– 共计 6 分子 ATP

糖异生的生理意义
3 、调节酸碱平衡 , 有利于乳酸的利用
1 、维持血糖水平恒定。糖异生的主要原料为乳酸、氨基酸及甘油。饥饿时主要为氨基酸和甘油。
2 、补充糖原储备– 直接途径: G UDPG 糖原– 间接途径(三碳途径): G 三碳化合物 糖原
( 丙酮酸、乳酸 )

底物循环( substrate cycle)作用物的互变反应分别由不同的酶催化其单
向反应,这种互变循环称之为底物循环。当两种酶活性相等时,则不能将代谢向前推进,称之为无效循环。

乳酸循环
葡萄糖葡萄糖葡萄糖 葡萄糖 葡萄糖葡萄糖
乳酸乳酸 乳酸乳酸乳酸乳酸丙酮酸丙酮酸 丙酮酸丙酮酸NADHNADH
NADNAD++NADHNADHNADNAD++
糖糖异异生生途途径径
糖糖酵酵解解途途径径
肝 血液 肌肉肝 血液 肌肉 CoriCori 循环循环

乳酸循环的形成是由于肝和肌肉组织中酶的特点所致。
生理意义:避免损失乳酸;防止乳酸堆积造成酸中毒。
乳酸循环是耗能的过程, 2分子乳酸异生为葡萄糖需消耗 6分子 ATP。

( 二 ) 、糖异生作用的调控1 、果糖 -6- 磷酸和果糖 -1 , 6- 二磷酸的相互转化的调节
调节酶:果糖二磷酸酶正调节物:柠檬酸、 ATP 是激活剂负调节物: Pi、 ADP 、 AMP 和果糖 -2 , 6- 二磷酸
2 、磷酸烯醇式丙酮酸和丙酮酸的转化的调节调节酶:丙酮酸羧化酶负调节物: ADP ,正调节物乙酰 CoA

二、葡萄糖核苷酸的合成

常用于蔗糖和淀粉合成的葡萄糖的活化体是 UDPG (尿二苷葡萄糖) ADPG (腺二苷葡萄糖) 合成反应为: G-1-P+UTP==UDPG+PPi UDPG焦磷酸化酶 PPi+H2O==2Pi 焦磷酸化酶 G-1-P+ATP==ADPG+PPi ADPG焦磷酸化酶

植物细胞中 蔗糖合成活化体—— UDPG淀粉合成活化体—— ADPG (主) UDPG (少)纤维素合成活化体—— GDPG 和 UD
PG

三、蔗糖的生物合成 1 、 蔗糖合成酶途径 蔗糖合成酶
UDPG + 果糖 ======== 蔗糖 + UDP
2 、磷酸蔗糖合成酶途径 磷酸蔗糖合成酶 UDPG + F-6-P ============ 磷酸蔗糖 + UDP
蔗糖磷酸酯酶 磷酸蔗糖 +H2O ————— 蔗糖 +Pi

四、淀粉的合成
ADPG 转葡萄糖苷酶 ADPG+“引物” ADP+ Gn+1 ( Gn) ( n≥2 )
UDPG 转葡萄糖苷酶
UDPG+ “引物” UDP+ Gn+1 ( Gn ) ( n≥2 )
1、直链淀粉的合成——淀粉合成酶的作用 由淀粉合成酶催化,需引物( Gn ) ,ADPG供糖基,形成 α-1,4糖苷键。

直链淀粉的合成
A
ADPG引物( Gn )+
+
直链淀粉 (Gn+1)
A
ADP

2 、枝链淀粉的合成 1 )淀粉合成酶
催化形成 α-1,4 糖苷键,直链部分交替作用共同完成
2 ) Q 酶(分支酶) 既能催化 α-1,4 糖苷键的断裂(非还原端切下 6-
7 个 G 的寡糖片段); 又能催化 α-1,6 糖苷键的形成(该段移至直链淀粉中的严格 G 上,形成 α-1,6 糖苷键——分枝点); 分枝部分交替作用共同完成.

在 Q酶作用下的支链淀粉的合成
+
Q酶( 1)
Q酶( 2)
B
A
A
A
B
B
nm
m
m
n
n

五、糖原的合成•由葡萄糖合成糖原的过程称糖原生成作用.•包括三种酶的催化作用: UDP- 葡萄糖焦磷酸化酶,糖原合酶,糖原分支酶. 糖原生物合成过程与植物支链淀粉合成过程相似,但参与合成的引物、酶、糖基供体等是不相同的。 引物:结合有一个寡糖链的多肽 酶:糖原合成酶,分支酶 糖基供体: UDPG

1 、 UDP-葡萄糖焦磷酸化酶(UDP-glucosepytophosphorylase)
—— 催化单糖基的活化形成糖核苷二磷酸,为各种聚糖形成时,提供糖基和能量。动物细胞中糖元合成时需 UDPG ;植物细胞中蔗糖合成时需 UDPG ,淀粉合成时需 ADPG ,纤维素合成时需 GDPG 和 UDPG 。2、糖原合成酶( glycogen synthase ) —— 催化 -1,4- 糖苷键合成3.糖原分支酶 ( glycogenbranchingenzyme) —— 催化 -1,6- 糖苷键合成

UDPG 的结构
G UDP

糖核苷酸的生成
+
+PPi
1-磷酸葡萄糖 UTP UDPG

糖原合成酶反应
UDPG
UDP
糖原( n个 G分子)
糖原( n+1)

糖原新分支的形成
糖原核心 糖原核心
糖原核心 糖原核心
非还原性末端
-1, 4糖苷键
-1, 6糖苷键
糖原分支酶

第八节 血糖水平的调节 血液中的葡萄糖含量称为血糖。按真糖法测定,正常空腹血糖浓度为 3.89-6.11mmol/L( 70 ~100mg%)。 一、血糖的来源与去路

二、血糖水平的调节 (一)组织器官 1 .肝脏;
2 .肌肉等外周组织。(二)激素1 .降低血糖浓度的激素—胰岛素;2 .升高血糖浓度的激素—胰高血糖素、肾上腺素、糖皮质激素、生长激素、甲状腺激素。
(三)神经系统

三、糖原分解和合成的调控 糖原的分解和合成都是根据肌体的需要由一系列的调控机制进行调控,其限速酶分别为磷酸化酶和糖原合成酶。它们的活性是受磷酸化或去磷酸化的共价修饰的调节及变构效应的调节。二种酶磷酸化及去磷酸化的方式相似,但其效果相反。 糖原合成酶
a(有活性 )
糖原磷酸化酶 b(无活性 )
OH
OH ATP
ADPH2O
Pi
糖原合成酶 b(无活
性 )糖原磷酸化酶 a(有活性 )
P
P

( 一 ) 胰岛素1. 是体内唯一降低血糖的激素,也是唯一同时促进糖原、脂肪、蛋白质合成的激素.2. 机制(1) 促进肌、脂肪组织等的细胞膜葡萄糖载体将葡萄糖转运入细胞.(2) 通过增强磷酸二酯酶的活性,降低 cAMP 水平,从而使糖原合酶的活性增强,磷酸化酶活性降低,加速糖原合成,抑制糖原分解.(3) 通过激活丙酮酸脱氢酶磷酸酶而使丙酮酸脱氢酶激活,加速丙酮酸氧化成 乙酰 CoA ,从而加快糖的有氧氧化.(4) 抑制肝内糖异生:通过抑制磷酸烯醇式丙酮酸羧激酶的合成以及促进氨基酸进入肌组织并合成蛋白质,减少肝糖异生的原料.(5) 通过抑制脂肪组织内的激素敏感性脂肪酶,可减缓脂肪动员的速率.

(二 )胰高血糖素1. 是体内主要升高血糖的激素.2. 机制(1) 经肝细胞膜受体激活依赖 cAMP 的蛋白激酶,从而抑制糖原合酶和激活磷酸化酶,迅速使肝糖原分解,血糖升高. (2) 通过抑制 6- 磷酸果糖激酶 -2 ,激活果糖二磷酸酶 -2 ,从而减少 2,6- 二磷酸果糖的合成,后者是 6- 磷酸果糖激酶 -1 的最强的变构激活剂,又是果糖二磷酸酶 -1 的抑制剂.于是糖酵解被抑制,糖异生则加速. (3) 促进磷酸烯醇式丙酮酸羧激酶的合成;抑制肝 L 型丙酮酸激酶;加速肝摄取血中的氨基酸,从而增强糖异生.(4) 通过激活脂肪组织内激素敏感性脂肪酶,加速脂肪动员 .

(三 )糖皮质激素1. 糖皮质激素可引起血糖升高,肝糖原增加.2. 机制(1) 促进肌蛋白质分解,分解产生的氨基酸转移到肝进行糖异生.磷酸烯醇式丙酮酸激酶的合成常增强. (2) 抑制肝外组织摄取和利用葡萄糖,抑制点为丙酮酸的氧化脱羧. (3) 在糖皮质激素存在时,其他促进脂肪动员的激素才能发挥最大的效果 .(四 )糖皮质激素1. 强有力的升高血糖的激素 .2. 机制:通过肝和肌的细胞膜受体、 cAMP 、蛋白激酶级联激活磷酸化酶,加速糖原分解.在肝内,糖原分解为葡萄糖.3. 在肌内则经糖酵解生成乳酸,并通过乳酸循环间接升高血糖水平.

激素通过 cAMP-蛋白激酶调节代谢示意图
ATP cAMP+PPi
内在蛋白质的磷酸化作用
改变细胞的生理过程
细胞膜
细胞膜
cR
蛋白激酶 (无活性)c + R cAMP
蛋白激酶(有活性)
受体 环化酶激素
G蛋白
非磷酸化蛋白激酶ATPADP
磷酸化蛋白激酶

cAMP 激活蛋白激酶的作用机理

激素对肝糖原合成与分解的调控
意义:由于酶的共价修饰反应是酶促反应,只要有少量信号分子(如激素)存在,即可通过加速这种酶促反应,而使大量的另一种酶发生化学修饰,从而获得放大效应。这种调节方式快速、效率极高。
肾上腺素或胰高血糖素1 、腺苷酸环化酶(无活性) 腺苷酸环化酶(活性)
2 、 ATP
cAMPR、 cAMP3 、蛋白激酶(无活性) 蛋白激酶(活性)
4 、磷酸化酶激酶(无活性) 磷酸化酶激酶(活性)5、磷酸化酶 b(无活性) 磷酸化酶 a(活性)
6、糖原
6-磷酸葡萄糖1-磷酸葡萄糖
葡萄糖血液
肾上腺素或胰高血糖素1
3
2102
104
106
108
葡萄糖
ATPADP
ATPADP
4
5
6