Embed Size (px)
Citation preview
ll
I
I
- I

•
------=--------- :s--·-
----~
~0 CH3-c,0_
Acetate
. ~CoASH acetyl CoA + A TP ~ynthetase
AMP+ PPi
~0 CH3 --c·-scoA
Acetyl ;:;oA
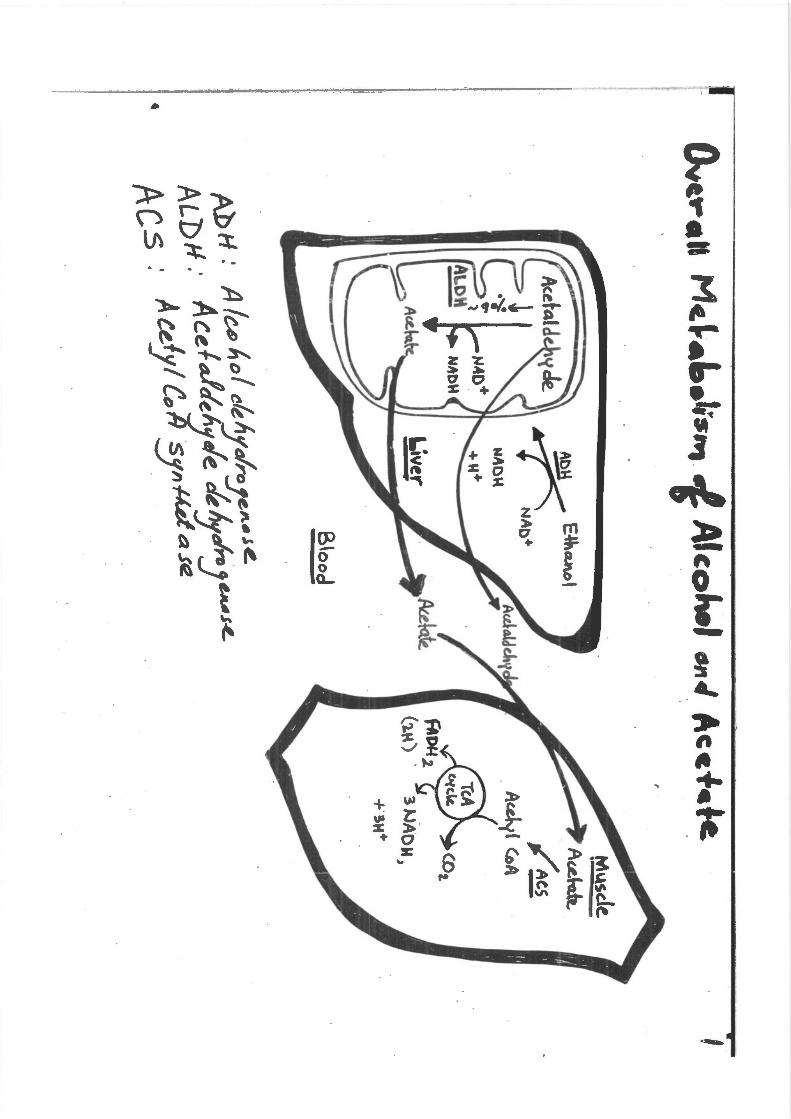
Fig. 25.4. The activation of acetate to acetyl Co A
NADPH NADP+, H+
Cytochrome Cytochrome P450 reductase P450
Fig. 25.5. General structure of cytochrome P450 enzymes.·0 2 binds to the P450fe·-he.me in the active site and is activated to a reactive form by accepting electrons. The electrons are donated by the cytochrome P450 reductase, which contains an FAD plus an FMN or Fe-S center to facilitate the transfer of single electrons from NADPH to 0 2• The P450 enzymes involved in steroidogenesis have a somewhat different structure. For CYP2El, RH is ethanol (CH3CH20H) and ROH is &cetaldehyde (CH3COH).
---
\
I \
\
I
w\
\ \
I
'
AcetaiFdeh~:o+ ALDH
NADH + H+
.------. 0 . . ~ : _ CH3C,
0_: : _____ .,:
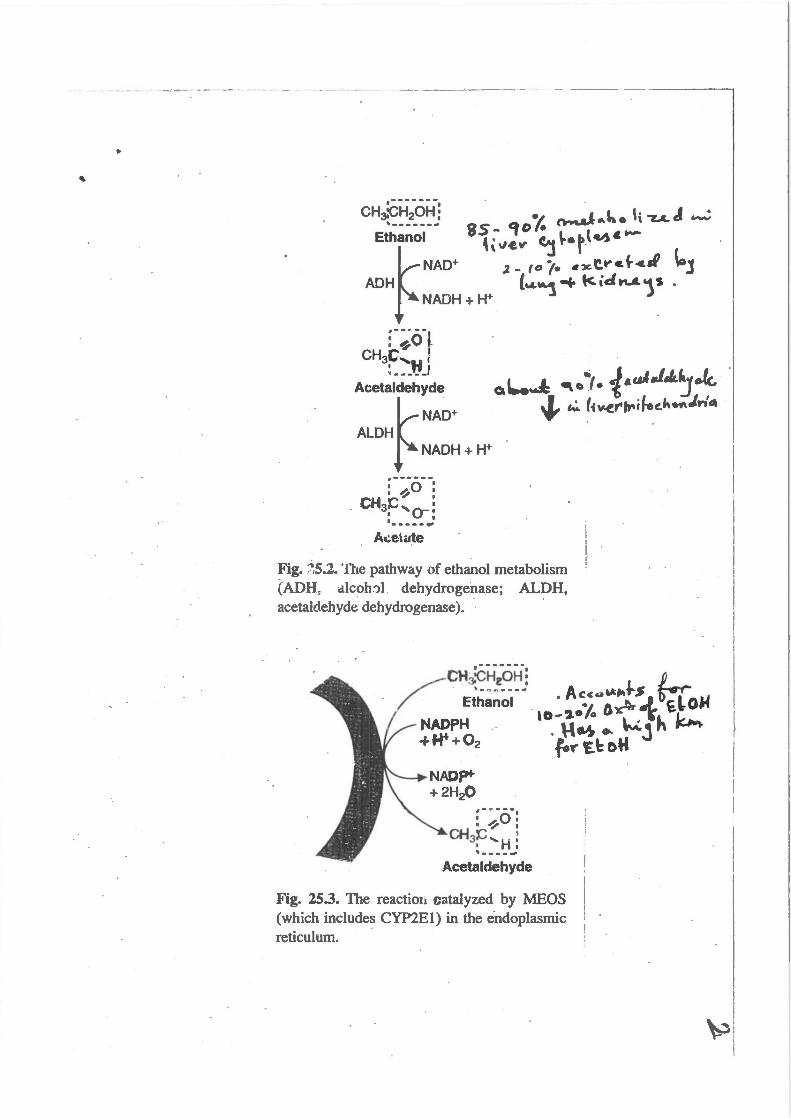
Fig. ::':5.2. The pathway of ethanol metabolism (ADH: alcoh::>l dehydrogenase; ALDH, acetaldehyde dehydrogenase). - ·
-------• •
Acetaldehyde
Fig. 25.3. The reactioli cntalyzed by MEOS (which includes CYP2El) in the e-ndoplasmic reticulum.
-- ----~
I I
~~ . I

if .. ..
I , . _:____ LJ·
Metabolism of Ethanol
Ethanol is a dietary fuel that is metabolized to acetate principally in the liver, with the generation of NADH. The principal route for metabolism of ethanol is through hepatic alcohol dehydrogenases, which oxidize ethanol to acetaldehyde in the cytosol (Fig. 25.1). Acetaldehyde is further oxidized by acetaldehyde dehydrogentises to acetate, principal/yin mitochondria. Acetaldehyde, which is toxic, also may enter the blood. NADH produced by these reactions is used for adenosine triphosphate (ATP) generation rnro.~gh oxidative phosphorylation. Most of the acetate enters the blood and is taken up by s.~eletal muscles and other tissues1 where it is activated to acetyl CoA and is oxidized in the TCA cycle.
·Approximately 10 to 20% of ingested ethanol is oxidized through a microsomal oxidizing system (MEOS) , comprising cytochrome P450 enzyme~ in the endoplasmic reticulum (especially CYP2El). CYP2El has a high Kmfor ethanol and is inducible ·by ethl:tnol. Therefore, the proportion of ethai'IDl metabolized through this route is greater at high ethanol concentrations, and greater after chronic consumption of ethanoL

- )) i .s ~ .. d .. u l·i """' -/; f-C:, / L( e-u s e :
/ rc. e. J W ~., J e ~~.. r... C. 1--. -" ,._ - ~ = _o .r~ =- 'jfo L~,
~ u .J !J ey~ .:. /,wr :=. rJ 7 ~ J z. I•"J . ~ .. -- ~ tnl4'1t: tV "d, r
- L.ve.-J!J~Je'"" ,.,~lilU: tt.~{/ke,r .. ~ 16 ir
- 13 ~.;_ u.s e N 12. D t!" i J keo $"~ I d:::t
- 7o I:..J ,.,..,.,.. ~ N 1~ K.t /J :: 130"tJ"o Kt:A/
,, . s~t!JUW - C.,..e.J 1171 ~ m.ual ,.J s,...tl
• , c .. b~,.,.,_lA.nf.'·~ ~ ,.J .;J.() ,.. rt ( cfJ
''.JJ,.,. v;,.r~ ~:rcar~·s.e vr· J Arf' --) ..21• ¥$a
cP ~ o2~.#c...
- {?,_ sl- #b ~rt,f.,..,~ ra~"y mu&tft .. ~ (,5)/-1. moJua/L aarbJe-> h ,4. ,.,11u. s ...... a. go"/.~ J'" ~of:t' VJ t.tf.,"l;~el_b 6,.,,;. + rl:.C.
- ])u..-t~ pr•lo':J~d ~ ,·1/".f , tJ.h·/iz.4
/,· ••• ~ F k· '?J •II fiSSul! {a~~~~ lrtti..Tr/,e) · &? . i~,u(
1/ f, 5 /J fYI~.S -f 1: e. I" lf..P b• 4 tP1 by fY' • rl2
/;~It fOO ~ f/ m I! f

CL Ll c ON~OC £ NtES/5
• C fucose 5y"t/ust's Is R~r~~~r~ f .. .Surviv«l
- iM~NtJj.S i C,/~.~co..VJ~ · . (.JJlllcl•fc _ ~ (.tJ ~r"''",fc -"( - p,.,..;.. •"~
.:- =::. ~ -
. f!!",;,• 6th{,i ~ (.ZJ 01Ct~loac#.laft. ,._. u• fO::I!..i•n• '; - - · ~ -£ I.)_~'J_t!!1, --7- l.~ ~iiJsc;/J•$fl'•lcS 4:- ,:-,, c/oJC.
~
1
C,af,cf•se. ~ c,f,,.se
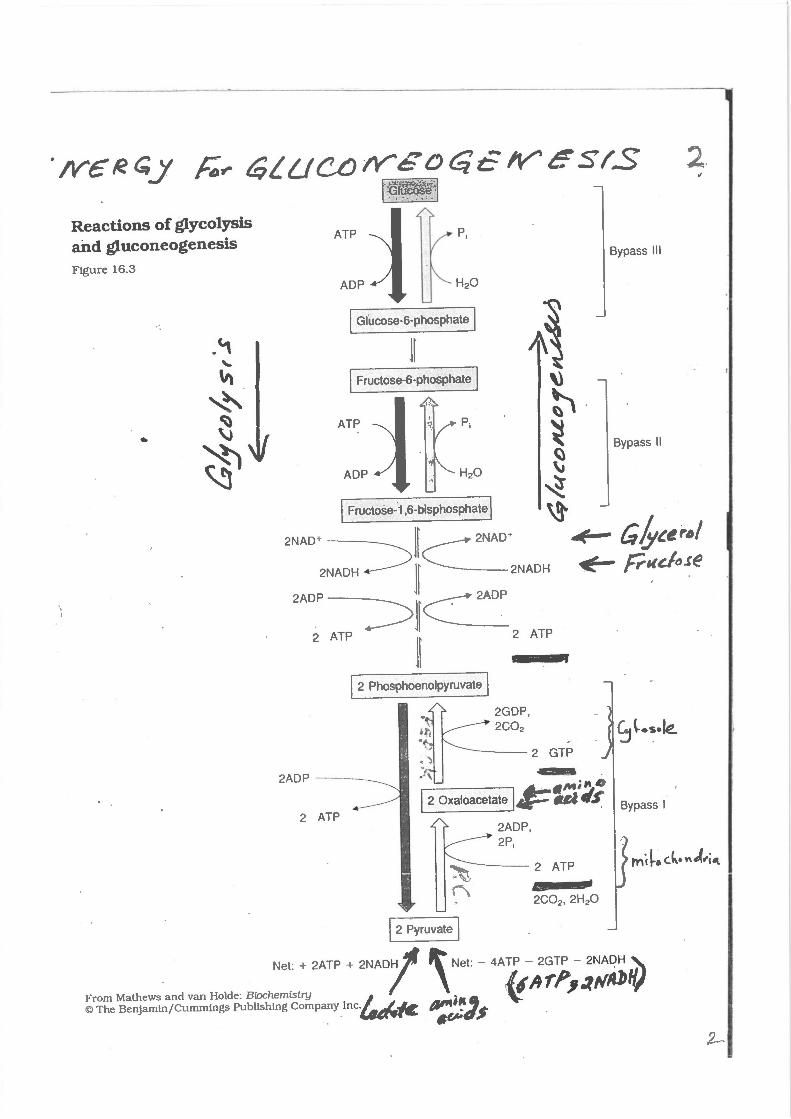
Reactions of glycolysis arid gluconeogenesis Fig ure 16 .3
ATP P,
ADP
I Gi"ucose-6-phosphate I Jr
I Fructose-6-phosphate I
ATP
ADP
2NAD + Jr 2NAD+
2~j[c=2NADH 2ADP~JrC::
2 ATP Jr 2 ATP
12 Phosphoenolpyruvate I
2ADP ---
2 ATP
2GDP, 2C0 2
---2 GTP
~
I ~ami"o
~ Oxaloacetate 14"'- tt.d iiS' 2ADP , 2P;
---2 ATP
[ 2 Pyruvate I Net: + 2ATP + 2NAD1
From Mathews and van Holde: Biochemis try © The Benjamin /Cummings Publishing Company I nc~{c.
Bypass Ill
Bypass II
Bypass I
""' ~·
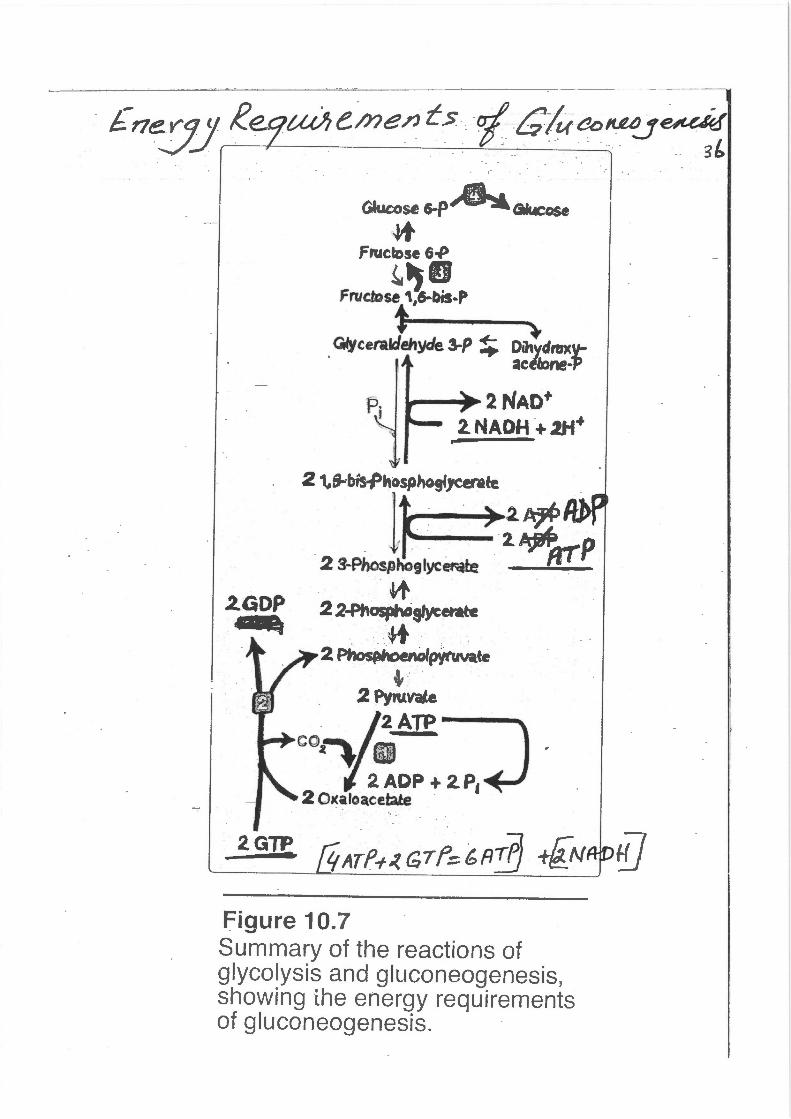
· £ne:yy Re Uthe.me?Jt> ufG!uaoKMJ~ . . . .
~...-..j~ 2 NADt 2.NADH +»i+
2.GTP
Figure 10.7 Summary of the reactions of glycolysis and gluconeogenesis, showing ihe energy requirements of gluconeogenesis.
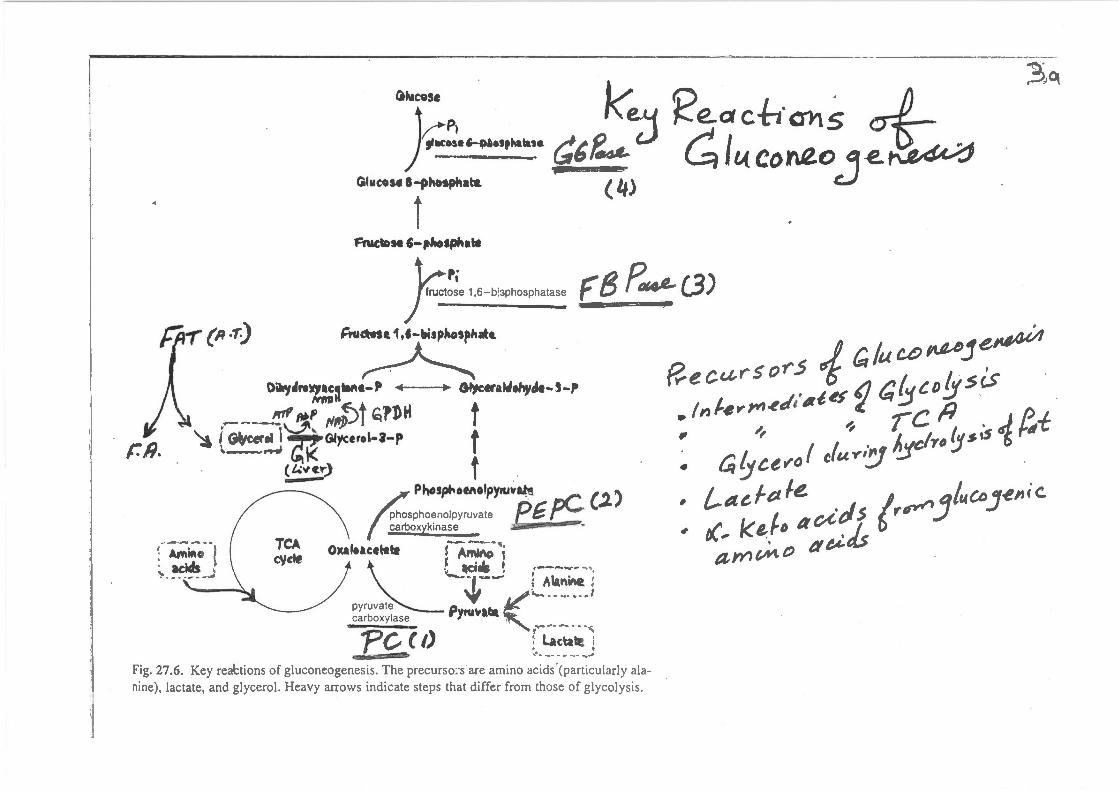
r:a.
---~;0\
C.hiC9Se 1<: ~~P· e..y 'f-...eact1·al1s · ... - .......... G6~ u Gi«con.e.o -e.~
Glo«o .. l-phosphala ( 4} 'j I
F'NCIDH '-pltosphatf
kr; ) nu~ose 1,6-b::;phosphatase F 8 P fJI4L C3)
;n.~a.se.1,1-bisphospllatt.
~ DiWJ.I,.~~W\e-P ,. .,. Glycetal.tehyde-5-P
~---~'!"~' /VIt})~t ~1DH f ~ ( G(ve«.t l ~Giycerel-a-p f
..~_ . ..,.,~ ~K (Uvt~) f ____. .
~ Phos~oMolpyiU~~ (.l)
sphoenolpyruvale .fs G ~ boxyk1nase "' •
T~ O)C~Iuuttte lA~-,, eye "' ~~ ~i4 ! ,------ ~'. ,._ __ -~--- l Alanine ~
~ (""'-----.--1 pyruvate '"" carboxylase Pytu\lltla :- -.- _____ . _ .
PC (I) f. ~eta~ i ~ ...... -. -.- ._J
f?.e cat" s o-r .5 i r;, /u U) P'Pje~ :{nt~ .. ,..,~.t/atd ') a{_jco!:Jsis . ~, ~ rc P · • cq '.J ce '~"" I cf u ..-i~ A_:jclro 'J s ,5 ~ ~-I: • L-adet le. · ' tC~ keJD ao-itl.s (r~ J/uCDJ~"ic.
ame#to t¥ei-Js
Fig. 27.6. Key reabions of gluconeogenesis. The precurso::s·are amino acids.(particularly alanine), lactate, and glycerol. Heavy arrows indicate steps that differ from those of glycolysis.
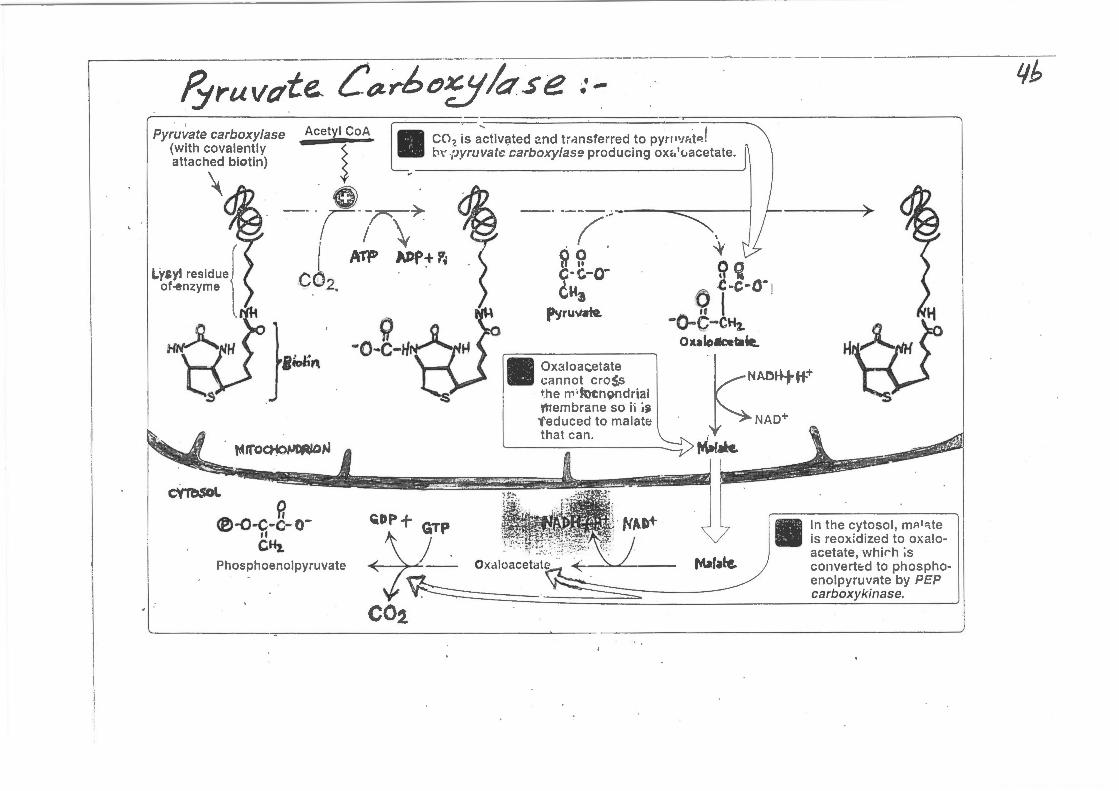
fyruvctte.. C~-rhc5k.se ··•
Pyruvate carboxylase (with covalently attached biotin)
\
Acetyl CoA
i •
.... i CO ?. iS actiV~ted :?.nd tr.-3nSferred to pytP'JI1t~ ! b\' pyruvate; carboxylase producing ox<.'uacetate.
·o . -- · r- ·-r-*
( / \ ----~.;-
lysyl residue { of~nzyme
I AW APf>:t ~ .co2.
~~ c-c-o-CH3
~ru\1•
\
'*' Qc;a t-c-o-1
0{ 10 -()..C-CH:z. Ouloketa~ }~~" Oxaloac.etate
cannot cro~ ~he n,;~n~mdrial V'JTembrane so h i~ 'feduced to malate that can.
. ~ NAO»ftt+
t'AfrOCHQ~ON
CVTb$0!.. p,
e-o-c-c-o-11
CHJ. Phosphoenolpyruvate
<altP + L\)TP '"'"7' 7 "== Oxal~ace;;;~ , C01 - .
~NAD+ M.t•
• Mal ale.
In the cytosol, m;::~ 1 1te is reox:dized to oxaloacetate, whirh ;s convertE:d to phosphoenolpyruvr~te by PEP carboxykinase.
qb
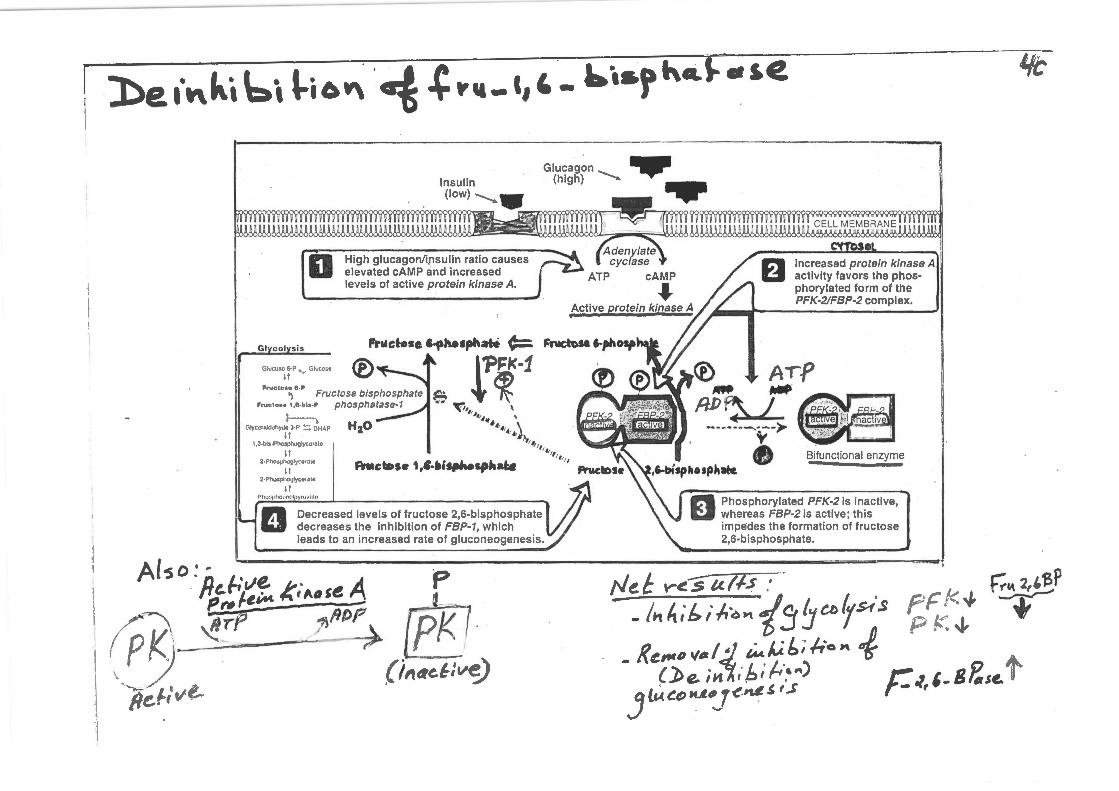
be l"kt bt l-i6W\ ·~ .(! "" _ c,' _ ~···r "•'" • s.e
Glucagon~ (high} lllllP ..
0 High glucagon/insulin ratio causes elevated cAMP and increased levels of active protein kinase A.
Ft&~dose '-.al\osph• ¢:::.
Glucu,o1 ~·P ' ·· Gluco;o ® ~PFI< •1
~hn~ ~ ~ . e ~
Fructoae 1,G·bls-P ~111 \
!----, '"'·~ ' G!yccrnlclohy\Jc 3.f' ~:~ Ot-tAP Hz.O 1'4111~ . \
'I ~,;
1.3-bls-Ph,.,;phogl~·ccrolc I >,, ~,,1 ~ t ,,,,, ,,,
l·r""""~"t'"•••" Fnaetose 1,~·11i.,W.ecphatc '' 2· Ph~Js!'IIIO\Jirce• ~to
It
II Decreased levels of fructose 2,6-bisphosphate decreases the inhibition of FBP-1, which leads to an increased rate of gluconeogenesis.
Al-so·~ ·fie. f,· ,;e ". ~ e,_leV>< ~1/l.t>St!. A 1:>
cAMP fJ • Active protein kinase A '
Increased protein kinase A activity favors the phosphorylated form of the PFK-2/FBP-2 complex.
ATf .. ,.. ft/)~ -------·-->-\'
• Bifunctional enzyme
ft Phosphorylated PFK-2 is inactive, U whereas FBP-2 is active ; this
impedes the format ion of fructose 2,6-bisphosphate.
tJ ~ t ~~Siill-.s . - /.,lt;b; /.<:. ... i j ';CLJ frsr's pF!< .J,
pP.::, ~
l.ft
t;:~ 2,,~p y
P 1/ , ~:P p~t>P
,_. !'.) ' 'o-/
[f~l .{ittt~ei:ve)
.. (( ~'" IJ\/IJt / :l ~. /J. b ; /-r"o "' aft ( 1> e in '1t ,· .b' 1-,. • ") o t )t"'~""-'"J<~~ t'.s F- .t, &-Br.sc. flef, vt,.,
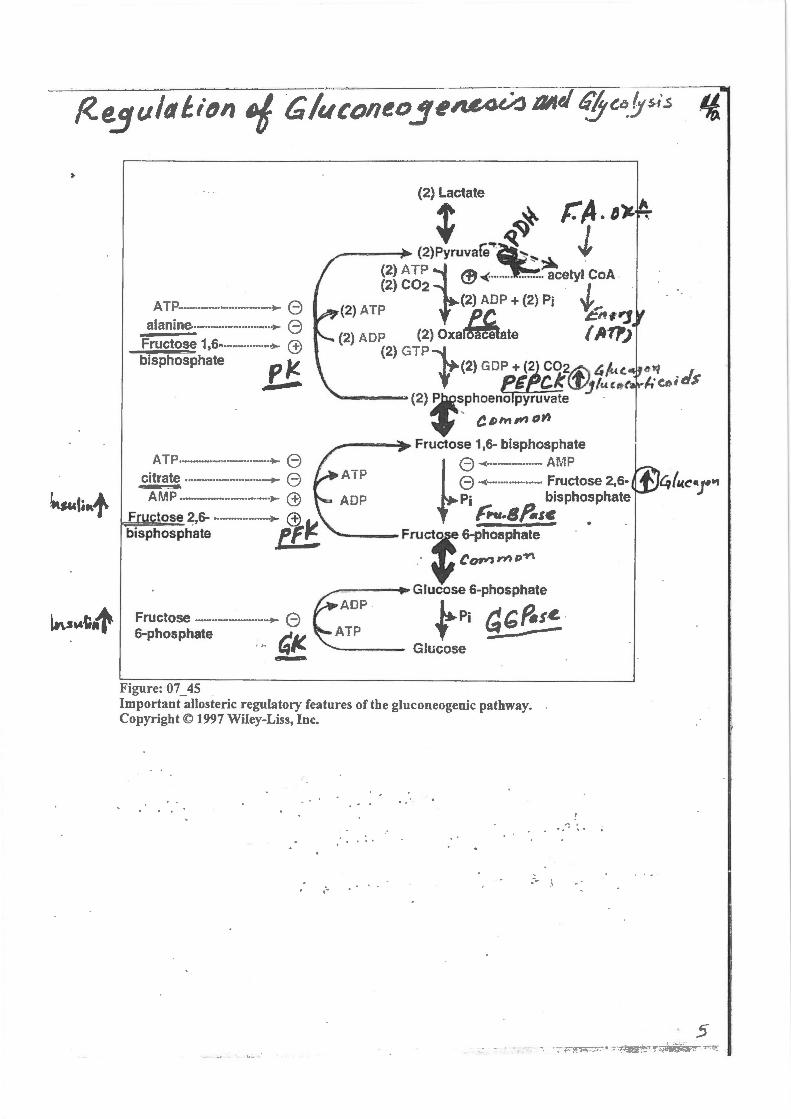
ATP ................................. ~ 8 alanine ............................ ~ e
Fructose 1,6 ................... ~ ffi bisphosphate p J!.
.....,....
ATP ................................. ~ B citr~te ............................... ~ B Afi.~P ................................ ~ ffi
F tose 2,6- .................... >- ffi bisphosphata eF!;
(2) Lactate + ~ r.A.I~. ---~ (2)Pyruvar.~( J
(2) ATP } ' ·~ (2) C0
2 @ ..(···-··· ...... _. acetyl CoA .
(2) ATP (2} ADP + (2) Pi ~,. . PC. C"l.,
(2) ADP (2) Oxa~ate { ~11') {2) GTP f
(2) GOP + (2 CO~ &/«c. G. CkiJijtu.c,;
"---=-- (2) ~~phoeno pyruvate ·
+ a D"" , ",
~---~Fructose 1,6- bisphosphate
l 8 ~·· · ····- ··· · ····-· AMP
t~ ;::.~;.~~~~~~~;.~; . ~ or'2 17) ~>""'
Frul6-phosphate
· ~Giucose6-phosphate AOP ~
~;~~~~~;;;;·-·-.. ····-J, ATP f Pi ~(;;~!!_ 1.4~ - Glucose -
Figure: 07 45 Important-allosteric regulatory features of the gluconeogenic pathway. Copyright© 1997 Wiley-Liss, Inc.
I
.. ..
· 5
..
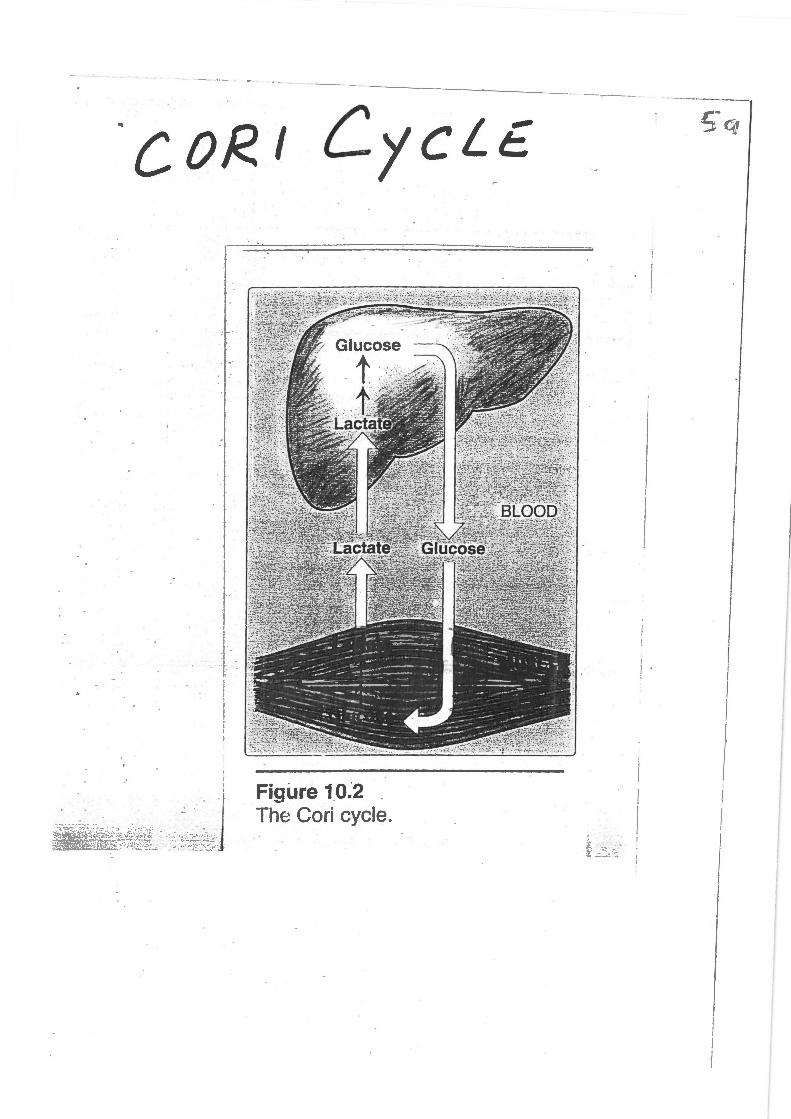
COR I CycLE
I
t ( .
I
~lfi~i~i.~~~,if' .. •· z~•;sA~J Figure 10.2 . The Cori cycle.
i I . I
I I I
• ..
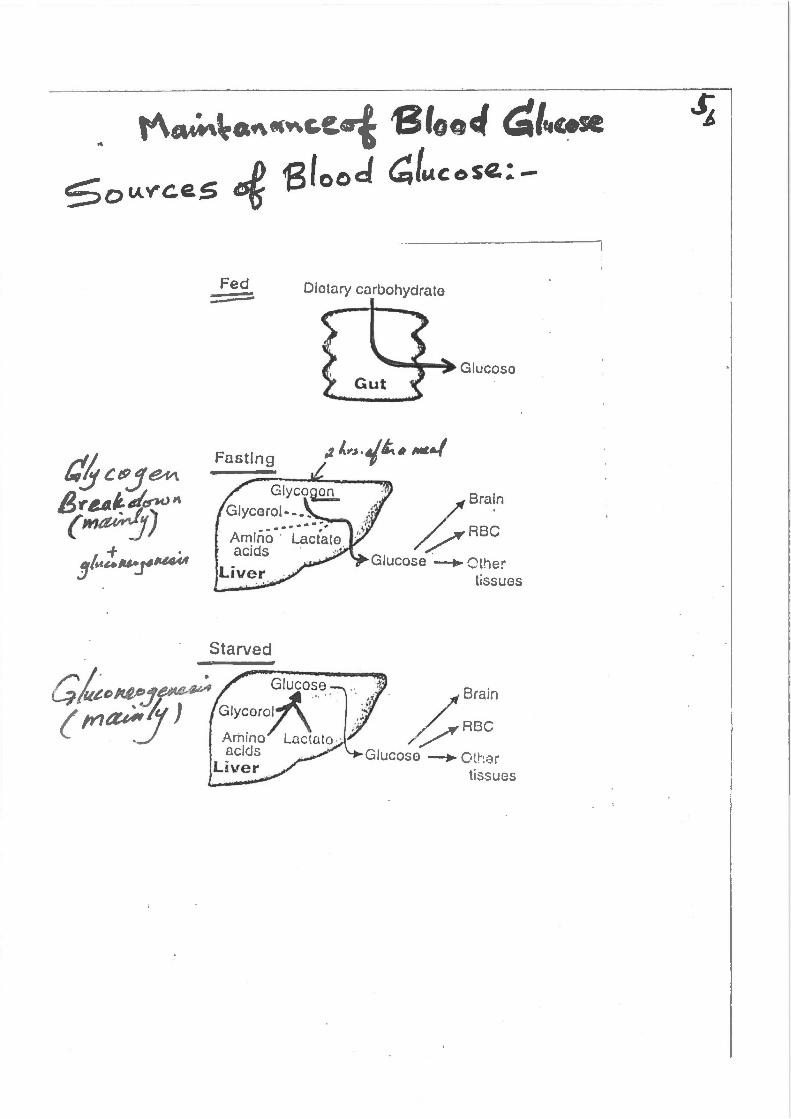
Fed ~
Starved
Itt~~
Blood ~{l.lcose:-
Dietary carbohydrate
Glucose
~::~n Glucose ~ C'the:
tissues
Gie~>~.J,.-:. Glucose
( !YI(UIP!J) , Glycaro.t\ Amrno Lactato . acids -
Liver
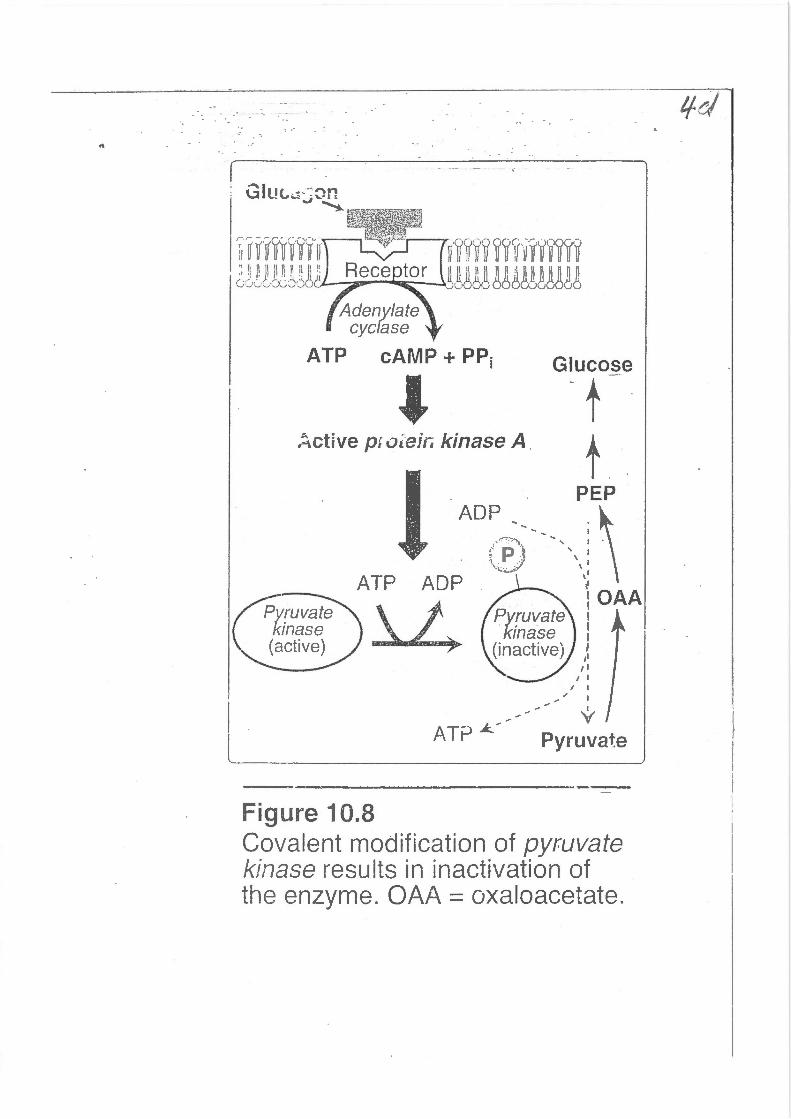
1
Glut...,::;~
1 ':.i. ~~v .... ~YI~T!.ll" ~¥!rli1rv·~~~~·~ c:::-.·0 GXC)•jOO.----:.~~~~--l.."'li"·flfl() !8~M!!8~!
A deny/ate cyclase
ATP cAMP+ PPi
' :..ctive pi oi.··ejr; kinase A .
ATP ADP
~
ATP k-
Figure 10.8
Glucose
t t
I I
- / I I y,
Pyruva~e
Covalent modification of pyruvate kinase results in inactivation of the enzyme. OAA = oxaloacetate.
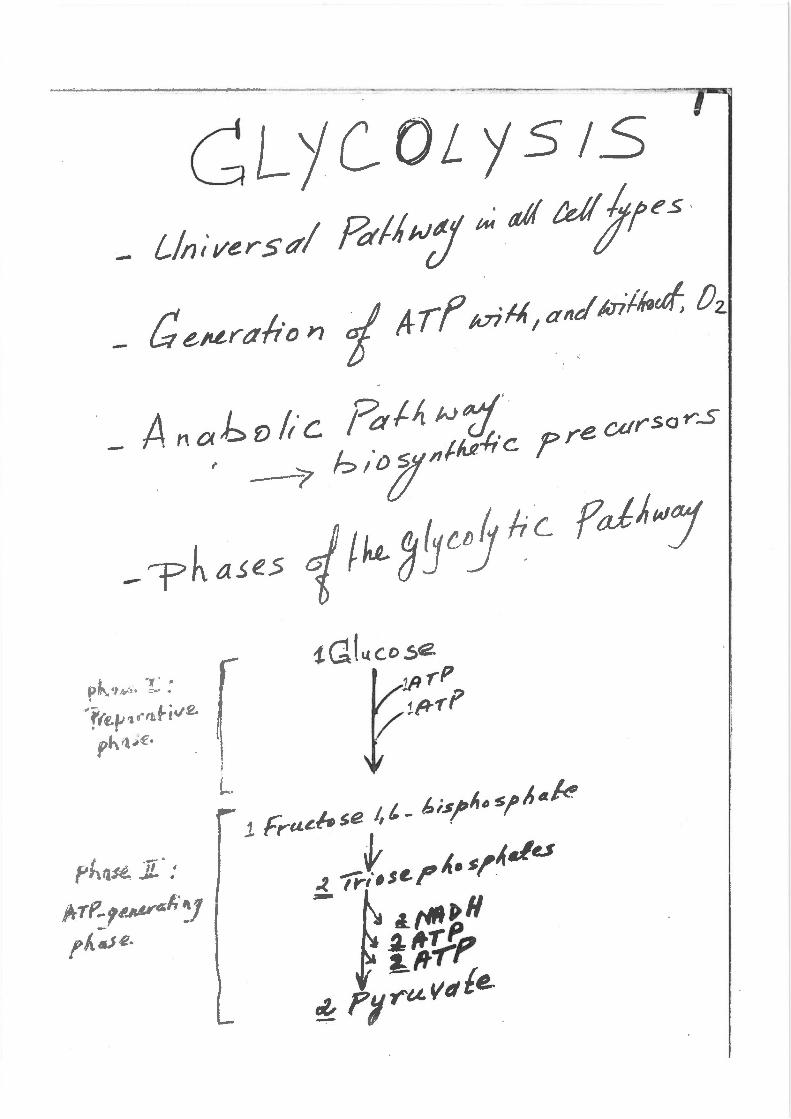
CLjC0Lj.51.S - Lfn;versttl ~/iwJ ~ atf tel! jtte.s - CurL.rtth·on 1 Aff tvi~1 ancllrrilbtt/; Oz.
- An o)~ v fr·c ;?af..~ N~· ·· ' · tLA'-/ic p~e cursor'S
. --'7 f? I 0 ~ "'""-" · I
- r-ph a~e51 /4 3~c..0 he fcdhwJ
-~~~;:·.-~~;v& r 1
ph <tA"· I I
1. GtL.tco se 111.,-P !f"~f

GLj( OL.Y S.!S .'-
.IJcct.Lr .s .;,. pi/ ,4,q,. c~!l.s
Jlo 0..- re. '14. lf'l.tM. ~f! f {IJ--r ~ fy tDl:fs/£~ t~nauJ,,·c. fumtAt fCJ.It' ~
6 r~ +- P- ~tl'P -rt.P,· ~ l ArP + 2· t r:/qf.e
" -r!S..s lJL,! Hat A a r~·e. 5Dr 6 (4tt~>$L
• flrtUie .. ~crl' 8/.tJJ Cells .. Co~AHJ /t.tts ~ rdiitc
. "it/III:J "'"""•" ~--~r~.&•t;~lc. ,._J 1.-~Aifre m tq de _/I_J; €Y:S
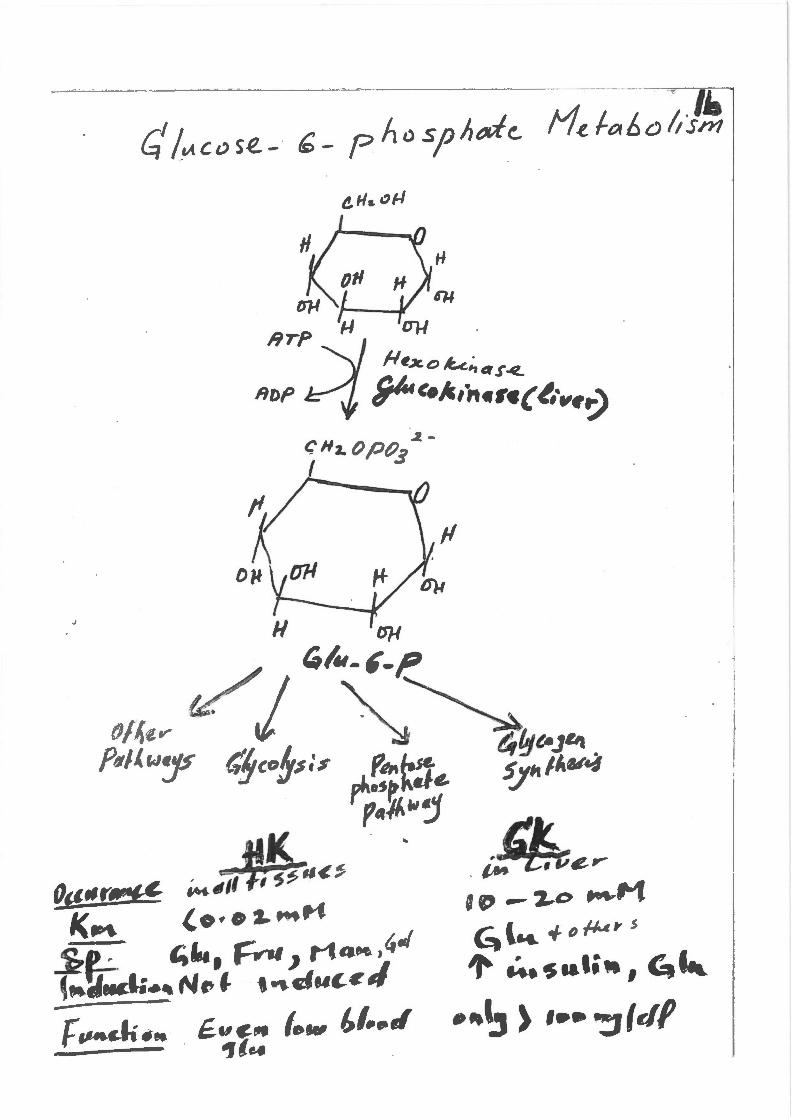
HTP
J
. ~4r:e~ ~ @:' --~o ~M c;.l .... +o~" s
,. U.,saali•, G.
.,,.~ ) ••• ":JIJf
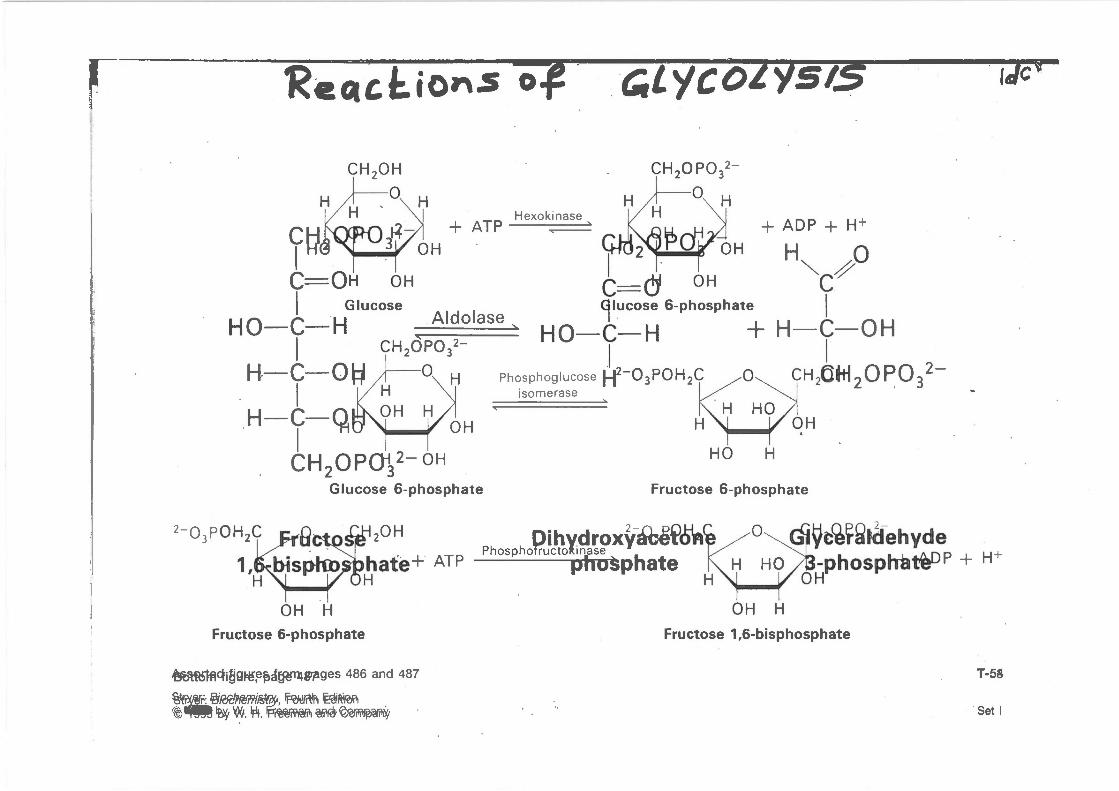
,-~ ~.
'RecuJ:.ions o~ · . ~Lycol'/SIS · tdcr-
r~ C==OH OH
Glucose I HO-C-H
I H-C-0~
. I H-C-Q
Aldolase
I CH OPQ-J 2- OH 2 3
Glucose 6-phosphate
Hexoki nase ...--
isomerase 0
HO
+ ADP + H+
H"'- ~0 c~
H
I H-C-OH
I CH20ki20 P032-
Fructose 6-phosphate
20H PhosphoPruiPto~~a~~xy~eHSA1
phosphate H
~t~~~t(fehyde -phosphil~P + H+
OH OH H
Fructose 6-phosphate
~cli~9~~~a!j~!11UP1!9eS 486 and 487
%¥lif.!fi_ %R!C!Pf(mi~, ~ 12ei~fefrll ~-- iB'y VJI.J. 'Pt. rp-~ ~~
OH H Fructose 1 ,6-bisphosphate
T-58
. Set I
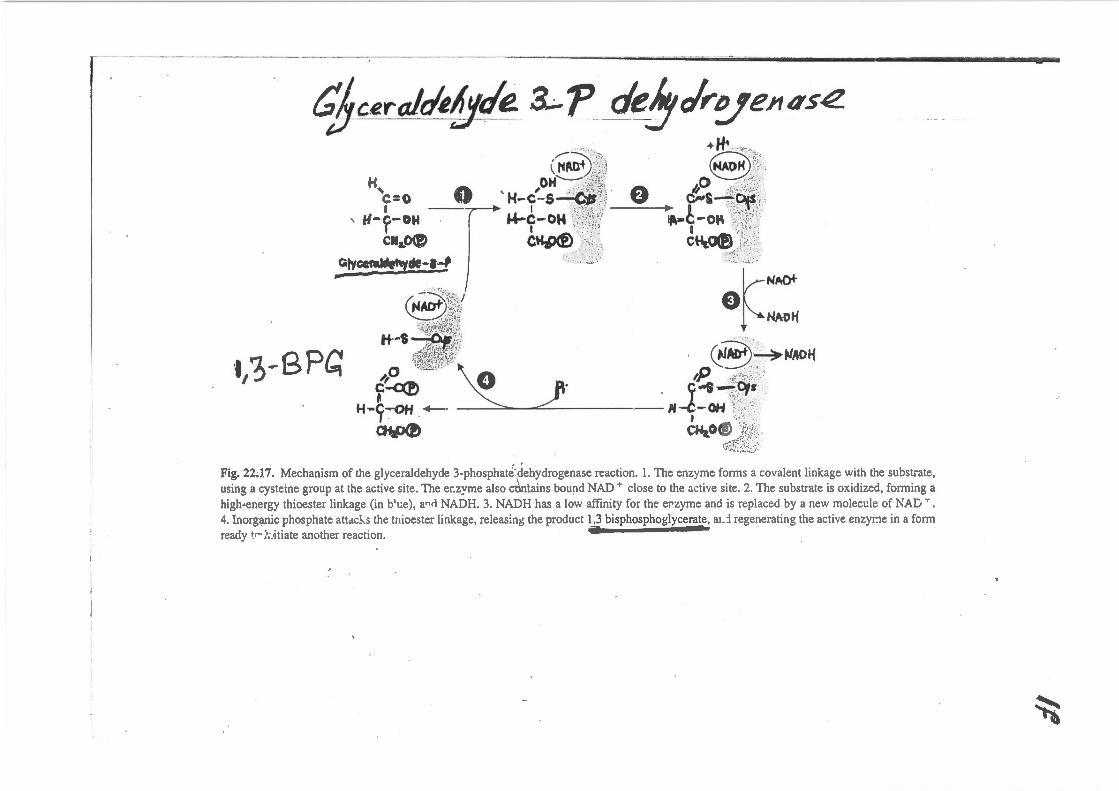
qj_ceralrA!Jtlt#: &.p de!Jclr~e;~as~ .. ,_,. •H• "' . ., ... _
--~-'::.:l. s ·t· .. ( ~#tnt).~i0 NAOH k';•. H OH"--: ~t'i~~· 0 "' ···"',,. 'c=o 0 'H-c-s4 ii\ 0 &-s_;;f'";;t,:?;\::"
I --r I ·:;r~\;{:\;J I i:i,:~/" ' H- c- OH · ...,_C- OH ~::/'~'!J:f-1 IA•C-Ot\ .,;r,j~{0'~
I I 't•·:t:··"~'' I ·.<'•:;.:.:*'1
CH 1\18\ ! cu ~ \::.:;.1 1'14\ ,·~:· .. \ ,_...,~ ~ :~·r:r~: .. ~. C\-lr.~ .~:~-::~;· "1'1~-~~ ''i:i;'l~:l c:!l);;;t: III'O'
~-:-....:r;'L(., / or .·t/3-BPC:
.A~ l)~'l' IU.I' ~~ ... :,;
~('' to:> •. ~i:.<51 NADti ~ ,;(f,..-~··t" ¥; '-¥,;-- ·~f•Nti~ ........ ,~ .. ;.,. -.. ,
IT ,!!;)~~) . 0"3:~~,40~ 0 ··-·'··~ .n "'"· ~ ......t"Y''t\ 0 2 tr ··."'_ .. ~.7_ ·.;:···.·· c~ . . ;..s~·~, . ~ ~ .. ,.,,\'. .
H-C--oH ~· ,._t-OH '{:·_?: .. ··~· 1 · · 1 ··;.d ~ ·. ~ cu,o® ·.:· .. ,. . ,·,~~~:~L~~-
Fig. 22J.7. Mechanism of the glyceraldehyde 3-phosphat~'.dehydrogenase reaction. 1. The enzyme forms a covalent linkage with the substrate, using a cysteine group at the active site. The er.zyme also ccintains bound NAD + close to the active site. 2. The substrate is oxidized, forming a high-energy thioester linkage (in b'l;e), a.,rl NADH. 3. NADH has a low affinity for the erzyme and is replaced by a new molect!le of NAG T .
4. Inorganic phosphate attdc!-.s the tnioester linkage, releasing the product 1,3 bisphosphogiycerate, ad regenerating the active enzyr:-1e in a form ready •r ;•;..itiate another reaction.
~
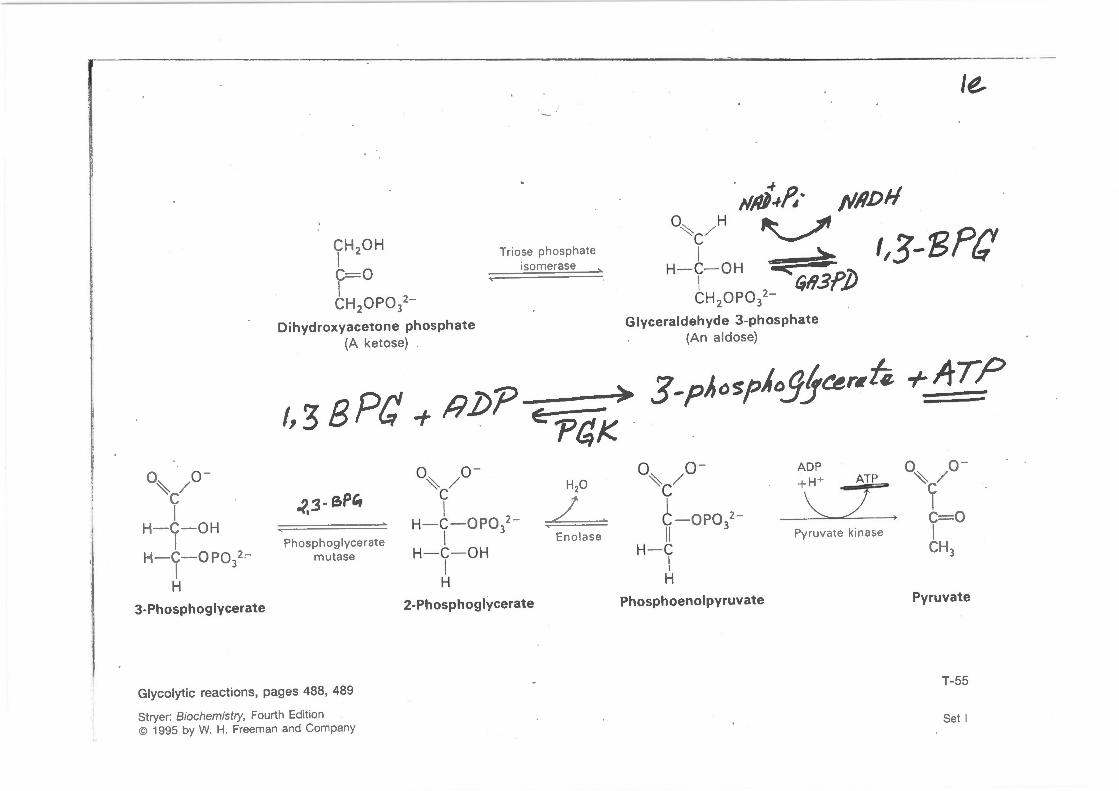
CH 20H I C= O I CH OPO 2-2 3
Oihydroxyacetone phosphate (A ketose)
~
Triose phosphate isomerase
0~ / H ~c
-+ 1{/1)-1-~ ..
~ I ~
/e,
fti/IDH
t,3-8?C H- C- OH '!iic;: •
tH OPO 2- ~ll3f]J 2 3
Glyceraldehyde 3 -phosphate (A n aldose)
> I, 3 B Pf} + !1.0? Eo 'PrJ,K.
3-p/,ospioJ!Je.er~h +ATP
o~ / o~c
I H- C- OH
I H- C- OPO 2·-
1 3
H
3-Phosphoglycerate
-?.3- e.P'r
Phosphoglycerate mutase
Glycolytic reactions, pages 488, 489
Stryer: Biochemistry, Fourth Edition © 1995 by W. H. Freeman and Company
o o-~/ H20 c
I / H- C- OPO 2-I 3 Enolase
H- C- OH I H
2-Phosphoglycerate
o~ / o- ADP o~ / o-+ H+ ATP ~c -y- ~c
t - OPO 2 -~ I
> ~ C= O II
3 Pyruvate kinase I
H- C CH 3
I H
Phosphoenolpyruvate Pyruvate
T-55
Set I
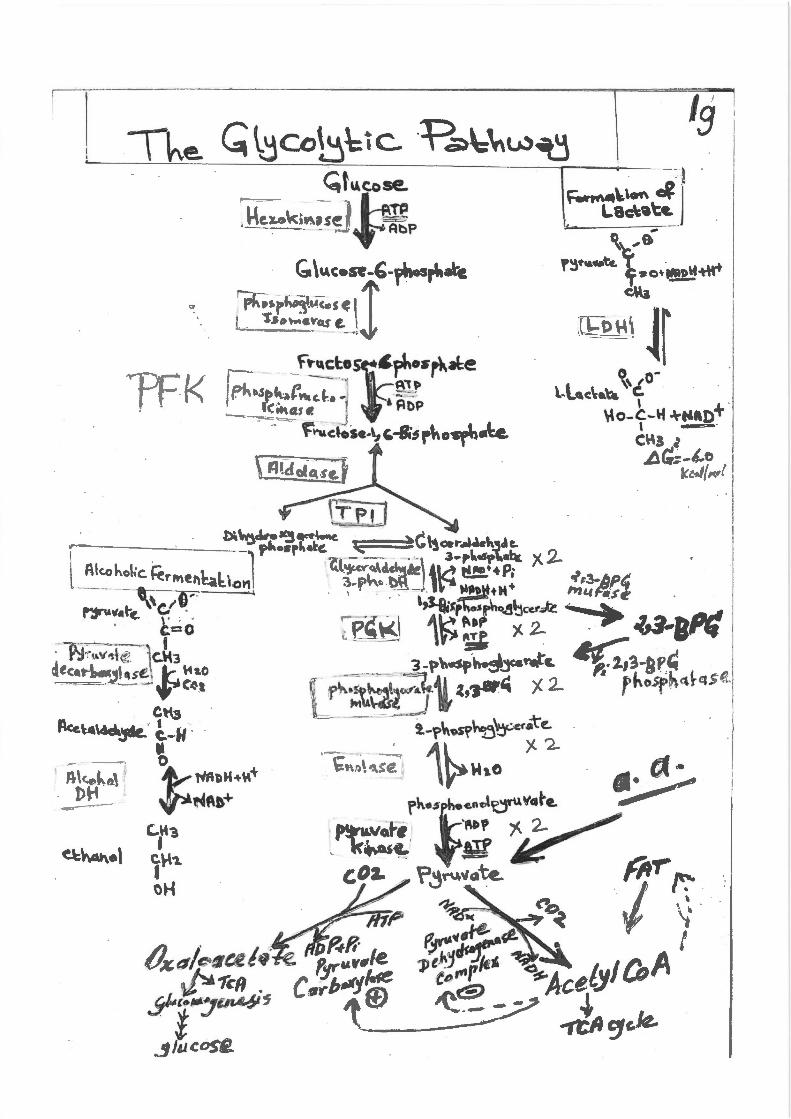
,{'441~~«19 ~1'c/l
.9-r:~.,..c.6is ~/tlCOS£
I
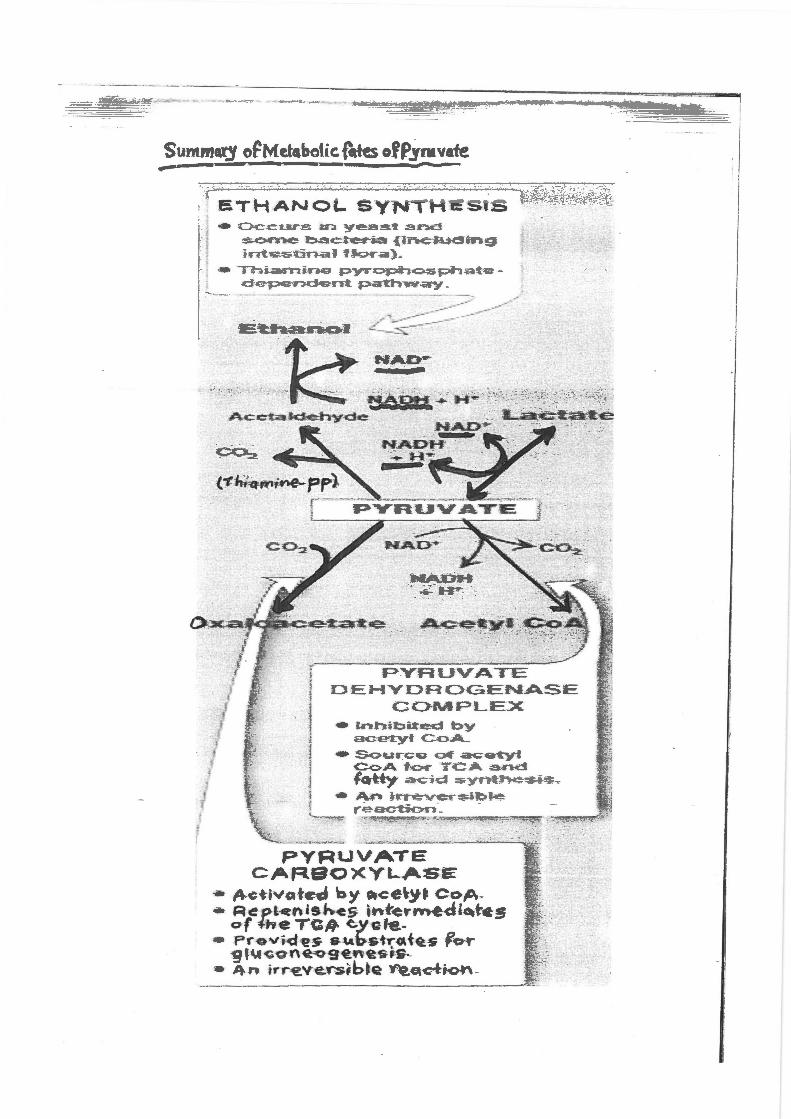
Summatj c~Metabotic. ~tes ~fP_yruvate .,..-{~~-------------·---· --~-----. :.. .
·, ETHANOt.. SYNTHEsis ":·/·:-- c~e:Ut~ }.lF.l y~s~ ar-x:! ~ ~e-t:-c::r-G:~ n.ne r.~ei:ng in"t.'letsti.....-aJ 'f~.a) .• 1 j
11 - .,-;::-:oi.amino P'YT"Ophc;;;;..s.J:*l.ui:g- t 1 ~ d~nd~<nt. p.a~IT!Nay _ 1:
1----~ ... .lri..o.~ .;,:::.?---~:::--j -·:~ : ,, . . '·~1:: . . . ~- '"'·' • :,~ ":~-. ~ ;-...,. ~,· :;.~~= . ;-4-..~::Q~~~~g,~ ·~,·~7~--~_..:,~t';;;-<>-
~ -. -- - - ::.:- "'-""' ....... ""
I-'·~ •. -"'
PYRUVATE D ·EHVDROGENASE
COI\oi.PLEX .._ Jo-hi:Q~~ bY
ac-etyl ~:::>A-~ S<:>up;;:o Q'f a<;:oct:yi
C.OA t~ TCA and fo-tty- ~j9 ~Yf"_l--"th~~-~ j-~~$i~l>¢:
PYRUVATE CAFlSOXY~S.E
• ,4-ctivatetl by e~tce~y.t CoP.-* R~tcnis~s 1-h-te-rrne-di-~t-es
o-f 4tie TG-JI<. eyGf'e.~ • :Pr-ov-i<f'e-5' •~b<str«t~-s fo-r
'9 rct.R<>«tn~~~~~ • 4.n kr'€Ve.r.slbhi~ ~~dio~ -

Lad-n.fe. vO ~>ro clu ad a . L. ~ .t, · 1 j ',.,. /.A Mer4 o, .r ~ ~/,.1 -· . 0 meet ~ ,_..,'4.,'j o.~~M,.'iljJ '
!. Ce.!ls Vt-1-~ low . ~£)" dl'Mfllld
J.. 7D c@;e tc;t' lA ~c,u.~t:l-enef[J · drunaAds ~ "!Jer-ot.~.$.!J ezt!re:L?._y
/YJIAJ3.de 1.~ ~"" ld 14¢~.-.' .s-lo 10-/P . I
3 · t!J f ,tfl:-U·CA _, •. ., [, (') pJ),·, ~~~ ~~~-4. ,I
1"() s,u.1'"v(V- , "f (/ · . . w/,J f!liXC!L 6 ~lSI~,., S . 1!.4_,._1
., I
L . r'"d~k. >s~rl (•·'<- 1-~-.P
· ,,J; C. A {;i, JfJ S 1-s ! -~ pll 0 · :J. ( 7- ··.s-,. 7- vs-J
is ~tn~s! ~,.,DY~ e.~ 1 ,mdabtib:c
· ao.:d() sis . ~ • w. ~lr.--.,dut-f,'-."') 4 f.,d/c. auc:/
. cl ~c.- -reo. s,Ld t.dt. /, · ~ a-A· 11 )1 q '~- ~
t1 tf1 f /!. tffM n'J ,r>J (!_a I.A S e. v.i ~, (J:J. f. Jn.e J Q-&- . met·cfll /iV~ n~. -t!. f "'~ /r'SJf'l r"e&tLfli_!] fr---:
Ce ll at' !e. i C;y--c,uJerl'!} -~-s ~~ :-• /mf!~·f'e_gj /J,_ /_rJUt~/ov-1
eJ · fYl_J" ~rcf;P.f tn/.,.--c./;n,
• R. ~ sr; ret. io t;_J F~ !u re.. e J - P t-d.., o h4 r1 &m /:. o It's I¥)

.. {.L..-. ~~""1J !I.e J ~~WJo-rrA.#.Je.. . 3.b
• ~ i reef tP- j.;._£; -/i o ......
fjp:tft:d-i~- pAo.>? .o.y /~
---
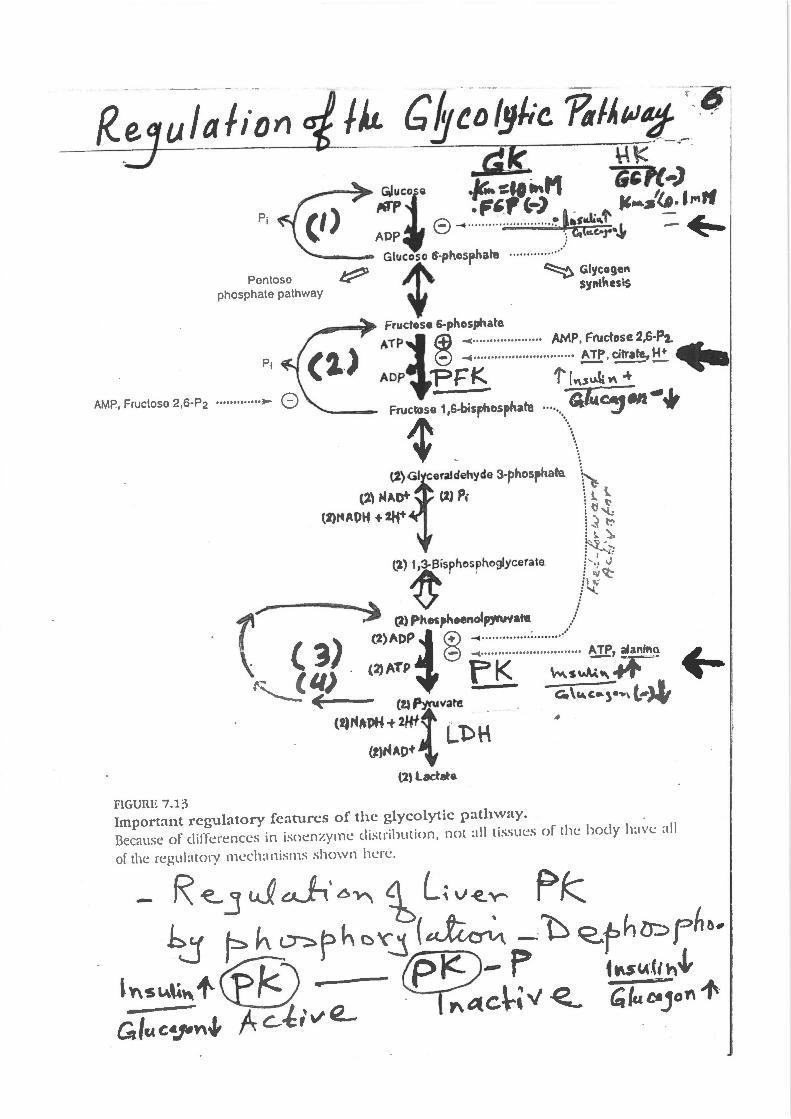
fiGUilE 7.1;) Important regulatory features of the glycolytic pathway. Because of dillerences in isoenzyme distribution, not all tissues or the body h~lVC all
of the regubtoty mech:tnisms shown he t-c.
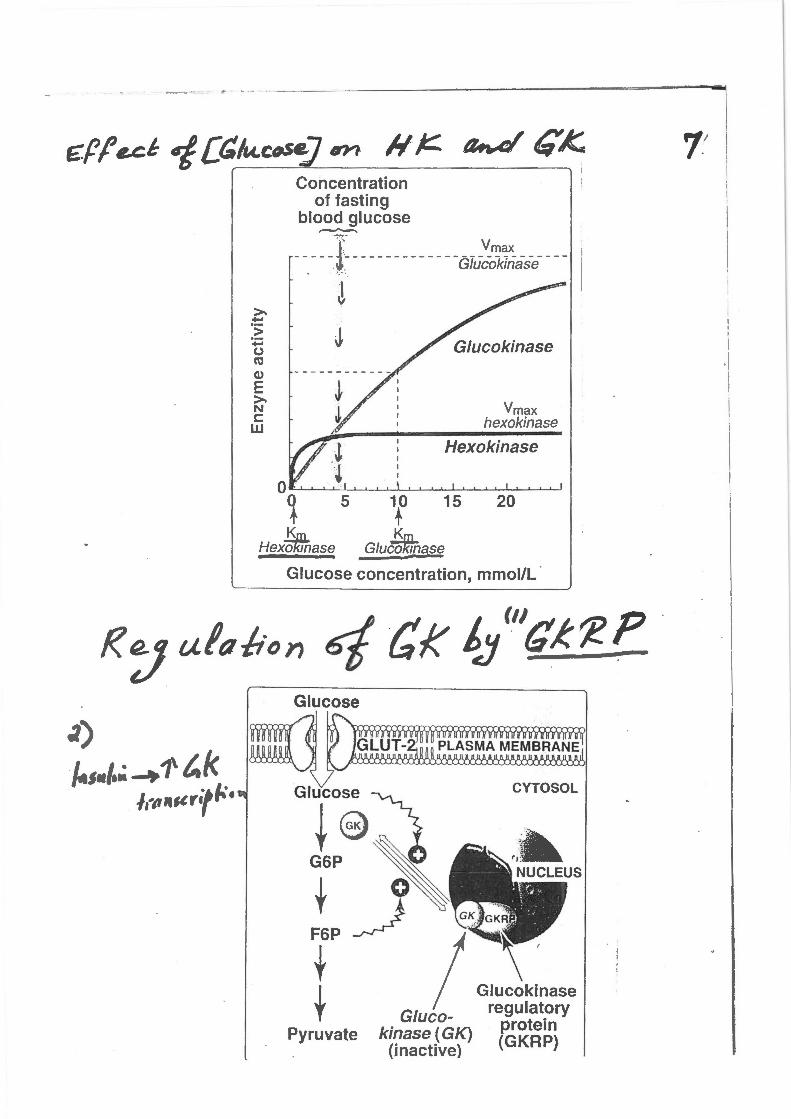
~)
~~ .. , • .; ~ 1\ t. k I til ,.,cr•f ,_. •
>--> -(.) ro Q)
E >N c w
Concentration of fasting
blood glucose ,_.,._..
-~--~1~----------- --Glu~o~tiase ___ _
J J
\0
Vmax hexokinase
Hexokinase
15 20
.!S.m. K Hexokinase Glucomnase -
Glucose concentration, mmoi/L ·
Glucose
kinase (GK) (inactive)
CYTOSOL
Glucokinase regulatory
protein (GKRP)
I
·I i I
' I
i ' i l
l 1.
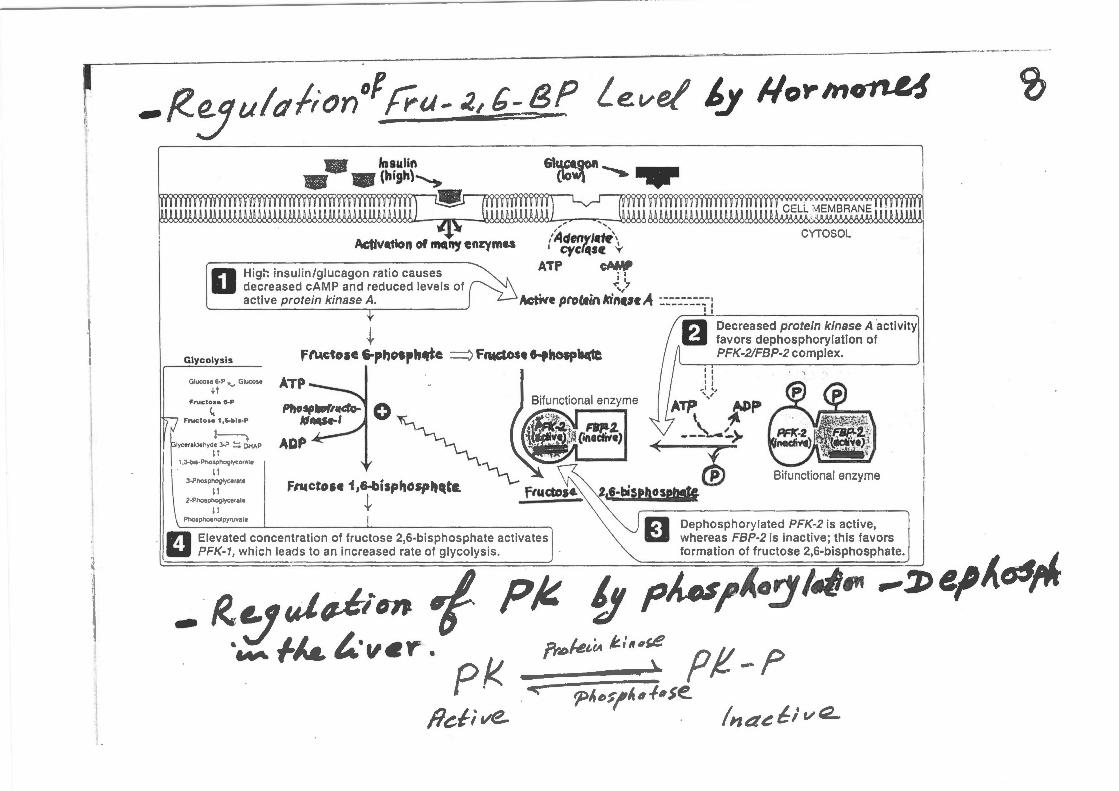
-~~uldfion°f !Yu- ;., G:g Level ~.Y #o-rm~ 1lmllf lnaulltl\ -"i'l'~n-,.. ~
~~~~WW\1 n~~mwww~mw~~WW~~~
_, ,, M~>MM~&&J!fJ6M6<X'IIU<~~>MM.I&'\66~
ActM,tlon or.,..._ en:rvme.s /Adenyl•~\ CYTOSOL
··:~ -f ' cycf~ts« v
decreased cAMP and reduced levels of ..-,,7 0 Higt': insulin/glucagon ratio causes ATP c~
active protein kinase A. Active pro CAin lrin«n ..4 :.-_-_-_-_-_-_-.-: ~------------------------~
+ Glycolysis F"--ctoae 6-phosptt~e :::::) Frwc:tost f"fhOipb4(1e
Glucose 6·P '-" Glucose ATP +t
FNetose 8-P
<.. Fruc l o a• 1 ,6-b ls · P
lyceraldehyd! 3·P ~ ~HAP APP If
1 ,J·b,s·Phosphoglycornlo
l l 3·Phospnoglycerate
lf 2-PhOsphoglycerate
F"'erose 1 ,&-bispttosp.,qtc.
+ ll PhOsphoenolpyruvate
.:JI Elevated concentration of fructose 2,6-bisphosphate activates
1:'1 Decreased protein kinase A ·activity U favors dephosphorylation of
PFK-2/FBP-2 complex.
Bifunctional enzyme
1!:'1 Dephosphorylated PFK-2 is active,
~
W PFK-1 , which leads to an increased rate of glycolysis .
- ((. '=' ""~~,. ·~ fk t..·v•r·.
1':.11 whereas FBP-2 is inactive; this favors formation of fructose 2,6-bisphosphate.
P~ !f pA-J~J.•!]I.I'" -::r>e,lfttJ$,It
pi< llefive..
fr.&>l~e-i.A it./""¢ \...
~ rp~ t>~l'" IJ "" se. /, lv~
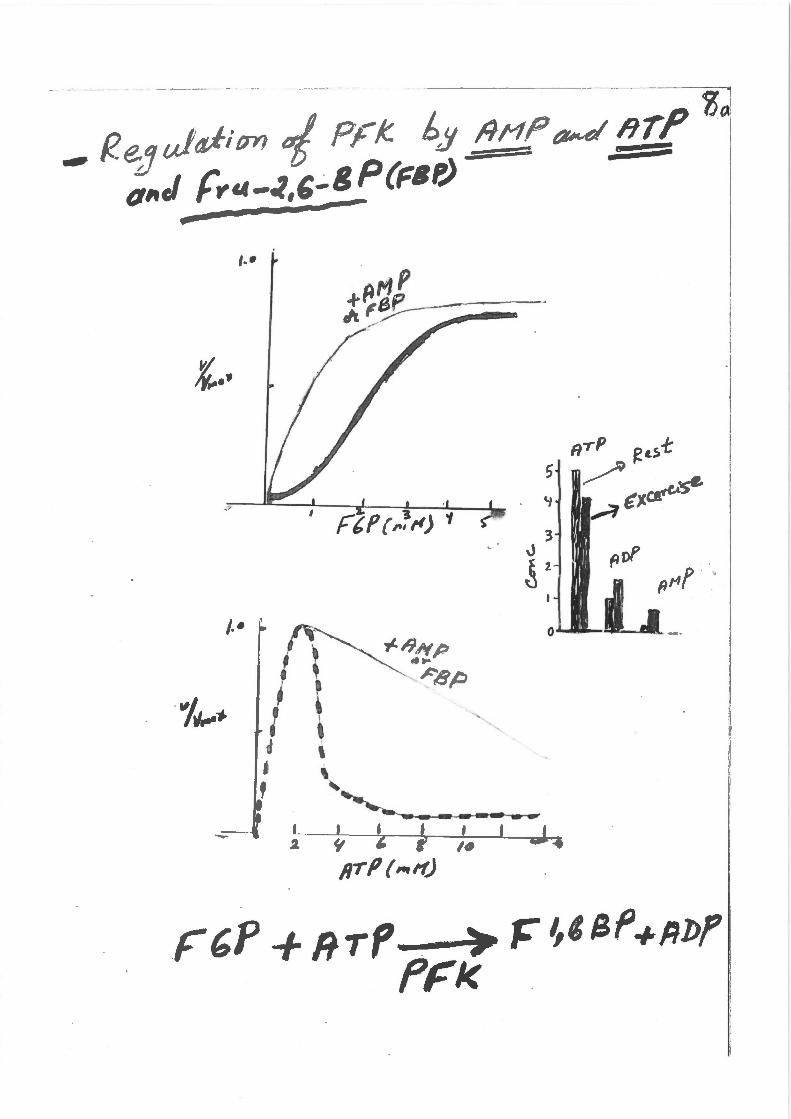
, ..
J.• ' ~'
i ; l ~I I t 1-J,..• 1 '
I , ; ~
' ' I I t f I 2 ~ ' t ltJ I d
1rrP r ,.,If)
F'P + RTP 0 . )o ~~~Sf +Rl>f PFI< . I
I ! I

-
- ~!uDr'ide. ~/LA' b-r'l-s Gn~ldr.se
f' l"' .or,.dt:Xt#!d MdeY' ~f,;6; / ~ b4t!f~y/J e ntJ(O se ro ,PTf2venl :J>enk/ earie$
.4-t'.sentC::. ~lsne-~
{tJ fi.,fa v..fl,.f arst!ni~ (./ir .sene~ fL) pre.J/a.nf nd ArP ~el' 11111-1;::, ,4 ?_ro du cl·i <!:>.,.. ~:1-!1 ~ ~ t:t lys;s (CeJmp~le JA9;{/, r;- ~ 4lc. Sub$/rJe r ~~~3Pl>)
{?r,' VA~,I aY"S~nlG {_ aYseni ~~ , , • /~r':' s./.e~. f::.<e c.~~~~.x. !Ari/1.. - ~~~ "iti;.~"'e;J f
~f..,;~' /-,'1\J . 7>bP.c~plex_ • tiC~ /<~~.!}/,.!-nl'a/1! ~A.Jtf':J~"tl.$
.. 6rentc.l..le/-~ tt-U:, «- {<'ct.o
«t:A.'el ck!J.y dro .' ~"' ~~ se .. ~s~ 1VJ.e,c.IP6/oq;c~ d'V.,~t.u·lt~tA~,S'f~k"(/..
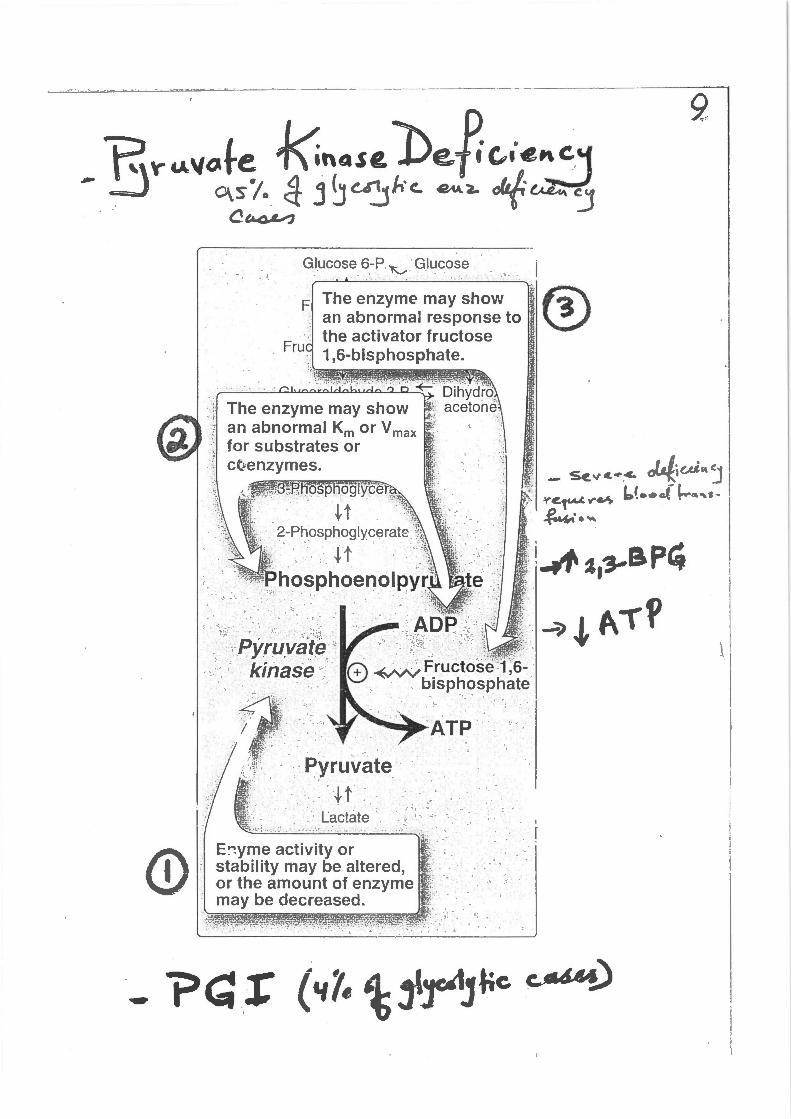
--------·-~--------~
Glucose 6-.P 'K,_.. ·Glucose -. -"" . . . . . . .. •. -~ .: .. ·: ....
, ····
·, .;·<: ·~r.;,)Q~i~ ':: : .. · kinase · .. -
. .
·Pyruvate · r· .,. t
;!"!iii i' 1 · -. · ' : tactate · · · q:ti~.,v l .-. :~:_·-:;'. '· ,_~_: , ~"--'.;':i~-::'~·;.'-.-~\. ;;: . ,_;-.i i ·.-.- --~ " -~"'. ...
E~yme activity or stability may be altered or the amount of enzy~e may be decreased.
. ,; '.
. . . ... . ~~:~
·. _.-,,
~
• - se" ._~ . .c. J.4,; W• J I .y~,tM .... ~ b~ •• ~f ~~""., _ I ~· .....
;
- ~ .... ~ ~,3--apc;
-"> J. A -rf









![ppt [??? ??????]...םייניב םוכיס –ATP היגרנא רורחשל תמרוג תוינגרוא תובוכרת קוריפ ATP תריציל ןחרזו ADP םיביגמ הז](https://img.pdfslide.net/doc/110x75/5e56a833e8dc9911e062c4dd/ppt-aatp-oe-.jpg)