Embed Size (px)
Citation preview

/3-VLDL Metabolism by Pigeon MacrophagesEvidence for Two Binding Sites with Different Potentials for
Promoting Cholesterol Accumulation
Steven J. Adelman and Richard W. St. Clair
Previous studies from our laboratory (J Upld Res 1988;29:643-656) have shownthat thioglycolate-eliclted peritoneal macrophages from White Carneau and ShowRacer pigeons, like mammalian macrophages, have on their surfaces specificreceptors for acetyiated low density llpoproteln (acLDL) and ^-migrating very lowdensity lipoprotelns (0-VLDL). The binding kinetics of 0-VLDL were complex, how-ever, suggesting more than one binding site. The purpose of the present study wasto further characterize these 0-VLDL binding sites. Scatchard analysis of 13Sl-0-VLDLblndlng curves indicated at least two classes of binding sites. The first binds pigeon/3-VLDL and LDL with high affinity (Kd approximately 7 jig/ml), Is down-regulated bycholesterol loading, requires calcium, and Is destroyed by the proteolytlc enzyme,pronase. This pigeon 0-VLDL receptor Is specific for pigeon /3-VLDL and LDL anddoes not recognize HDL, acLDL, methyl LDL, cynomolgus monkey LDL, or rabbit0-VLDL. Like the mammalian macrophage /3-VLDL receptor, the "pigeon /3-VLDLreceptor" has many of the characteristics of an LDL receptor. The second class ofbinding sites Is relatively nonspecific, recognizing both pigeon and rabbit 0-VLDL,LDL, acLDL, methyl LDL, and HDL Binding to this site Is not altered by Incubation ofmacrophages with pronase or by cholesterol loading. This binding site has lowaffinity for /3-VLDL (Kd approximately 100 f*glm\), but high capacity. We have calledthis the "llpoproteln binding site," a term used by others to describe similarllpoproteln binding characteristics on a variety of cells. Not only does binding to thissite promote the internallzation and degradation of lipoprotelns, but It may alsofacilitate the Independent uptake of cholesterol. This conclusion Is based on theobservation that more cholesterol accumulates In cells incubated with rabbit0-VLDL, which binds only to the llpoproteln binding site, than can be accounted forby 0-VLDL uptake and degradation. Since the llpoproteln binding site recognizes avariety of normal, as well as abnormal, lipoprotelns, It would not require thegeneration of abnormal llpoprotein products, as must occur with the scavengerreceptor, to promote the accumulation of cholesteryl esters in macrophages ofatherosclerotic lesions. This, coupled with the fact that the lipoproteln binding site isnot down-regulated by cholesterol loading, suggests that It could provide analternative mechanism to the scavenger receptor pathway for the formation of foamcells. (Arteriosclerosis 9:673-683, September/October 1989)
W hite Carneau (WC) pigeons are susceptible tonaturally occurring and cholesterol-aggravated aor-
tic atherosclerosis, while Show Racer (SR) pigeons areresistant, even though both breeds have the same con-centrations of plasma lipoproteins and other risk factors.1
As a result, comparative studies with these breeds ofpigeons provides a unique opportunity to understand thebiochemical and cellular factors that influence susceptibil-ity to atherosclerosis at the level of the arterial wall.
From The Arteriosclerosis Research Center, Department ofPathology, Bowman Gray School of Medicine, Wake ForestUniversity, Wlnston-Salem, North Carolina.
Steven J. Adelman is now at the division of Metabolic Disor-ders, Wyetrt-Ayerst Research, Philadelphia, Pennsylvania.
This work was supported by SCOR Grant HL-14164 from theNational Heart, Lung, and Blood Institute. Dr. Adelman wassupported by National Research Service Award InstitutionalGrant HL-07115.
Address for reprints: Dr. Richard W. St Clair, Department ofPathology, Bowman Gray School of Medicine, Wake ForestUniversity, Winston-Salem, NC 27103.
Received January 12, 1989; revision accepted May 3, 1989.
Atherosclerotic lesions of humans and a variety ofexperimental animals are characterized by the presenceof cholesteryl ester-rich foam cells, many of which arederived from circulating monocyte/macrophages.2-5 Thesecholesteryl ester-rich cells appear early in lesion forma-tion and are present in varying amounts throughout thestages of progression of the atherosclerotic plaque. Sincethe macrophage appears to play an important role in theprogression, and possibly regression, of atheroscleroticlesions, understanding the mechanisms responsible forcholesterol accumulation in macrophages is essential toour understanding of the etiology of this disease. We haverecently reported that peritoneal macrophages from bothWC and SR pigeons have binding sites that are similar to/3-migrating very low density lipoprotein (/3-VLDL) recep-tors and acetylated LDL (scavenger) receptors on mam-malian macrophages.6 Lipoproteins bind to these siteswith high affinity and specificity. In addition, lipoproteinuptake and metabolism by these two pathways results inthe stimulation of cholesterol esterification and cholesterylester accumulation in pigeon macrophages.
673
by guest on July 21, 2018http://atvb.ahajournals.org/
Dow
nloaded from

674 ARTERIOSCLEROSIS V O L 9, No 5, SEPTEMBER/OCTOBER 1989
Both WC and SR pigeons form cholesteryl ester-rich/3-VLDL in response to cholesterol feeding.7 In contrast tomammalian 0-VLDL, no apoprotein E (apo E) can bedetected in pigeon /3-VLDL or in any of the other pigeonlipoproteins.7 This is of interest since a number of recentstudies have demonstrated that apo E mediates theinteraction of /3-VLDL with /3-VLDL receptors on mamma-lian macrophages,89 which are now believed to be mod-ified forms of the low density lipoprotein (LDL) recep-tor.10'11 Thus, the ligand(s) responsible for the binding ofpigeon /3-VLDL to pigeon macrophages must be funda-mentally different from those responsible for binding tomammalian macrophages. In addition to binding to ahigh-affinity site on pigeon macrophages, Scatchard analy-sis of 4°C binding data suggests that pigeon 0-VLDL canalso interact with a second, lower-affinity site.6
Our previous study showed that rabbit /3-VLDL, a lipopro-tein rich in apo E, was a poor competitor for the binding ofpigeon /3-VLDL to pigeon macrophages.6 This is consistentwith the conclusion that apo E does not mediate the high-affinity binding of 0-VLDL to pigeon macrophages. Rabbit/J-VLDL was also a poor competitor for the binding of acetyllow density lipoprotein (acLDL) to the scavenger receptor onthese cells. Despite poor competition of rabbit /8-VLDL forbinding to these two high-affinity binding sites, preliminarystudies demonstrated that rabbit 0-VLDL was capable ofstimulating substantial cholesterol esterificatJon and cho-lesteryl ester accumulation in pigeon macrophages. Basedon these observations, we set out to further characterize themetabolism of rabbit 0-VLDL by pigeon macrophages. Theresults of these studies indicate that pigeon macrophagespossess a low-affinity binding site, which recognizes avariety of lipoproteins. This is similar to the lipoproteinbinding site first described by Bachorik et al.1213 on hepa-tocyte membranes. The results indicate that rabbit /3-VLDLwhich binds to this site, stimulates the accumulation ofcholesterol by a process that can only be partially explainedby intemalization and degradation of the intact /3-VLDL
MethodsMaterials
Sodium 125l-iodide (IMS 300, carrier-free, in NaOHsolution, pH 7 to 11) and 114C-oleic acid (CFA 243) werepurchased from Amersham Corporation, Arlington Heights,Illinois. Silica gel-coated glass plates for thin-layer chrc-matography (TLC) were purchased from American Scien-tific Products, McGraw Park, Illinois. Stigmasterol andoleic acid were purchased from Applied Science Labora-tories, Incorporated, State College, Pennsylvania. Pro-nase, fibrinogen, and bovine serum albumin (BSA) werefrom Sigma Chemical Company, St. Louis, Missouri.Eagle's minimal essential medium (MEM), fetal bovineserum (FBS), and calf serum were obtained from Hazel-ton Research Products, Denver, Pennsylvania. Tissueculture dishes were purchased from Coming Glass Works,Corning, New York. All other tissue culture supplies wereobtained from KC Biological, Lenexa, Kansas. Fluid thio-glycolate medium was purchased from Difco Laborato-ries, Detroit, Michigan and was prepared as described bythe manufacturer. Gel electrophoresis supplies were from
Pharmacia, Piscataway, New Jersey. All other chemicalswere reagent grade and were purchased from FisherScientific Company, Raleigh, North Carolina.
LipoproteinsAnimals were fasted overnight before blood was
obtained for lipoprotein isolation. Rabbit /3-VLDL wasobtained from New Zealand White rabbits (Franklin, WakeForest, NC) who were fed a pelleted rabbit chow diet6
containing 0.5% cholesterol and 10% lard for at least1 month. Pigeon /3-VLDL and hypercholesterolemic LDLwere from pigeons fed a cholesterol-rich (0.5% choles-terol, 10% lard) diet for at least 3 months. Normal pigeonLDL and high density lipoprotein (HDL) were from pigeonsfed a pelleted grain diet. Monkey LDL was obtained fromMacaca fasdcularis monkeys fed Purina monkey chow towhich cholesterol was added.14 Lipoproteins were iso-lated from the pooled plasma of approximately 12 pigeons,six monkeys, and three rabbits. /3-VLDL was isolated fromthe plasma of hypercholesterolemic pigeons and rabbitsby ultracentrifugation (d<1.006 g/ml) as described.15 LDLand HDL fractions were obtained by a combination ofultracentrifugation and agarose column chromatographyon 4% agarose (Bio-Gel A-15m, Bio-Rad Laboratories,Richmond, CA).16 AcLDL was prepared from hyperiipemicmonkey and pigeon LDL.17 Lipoproteins were iodinatedby the iodine monochloride method.18 Less than 10% ofthe 12SI in the lipoprotein preparations was soluble inchloroform and methanol (2:1), and more than 97% wasprecipitable in 10% trichloroacetic acid (TCA). All lipopro-teins were stored in a sterile state and were sterilizedagain just before use with a 0.45-^m Millipore filter(Millipore Corporation, Bedford, MA), and they were usedwithin 3 weeks of isolation. The chemical composition ofthe lipoproteins was determined as described previously.7
The specific activities of the iodinated lipoproteins rangedfrom 200 to 600 cpm/ng protein. Unless otherwise indi-cated, all lipoprotein concentrations were added to cells inculture on the basis of lipoprotein protein.
Cell CultureWC and SR pigeons (6 to 12 months of age) were
obtained from our breeding colony and were fed a cholesterol-free pelleted grain diet Elicited peritoneal macrophageswere obtained as described by using thioglycolate.18 Exper-iments in which cells were incubated at 37°C were carriedout in a CO2 incubator with bicarbonate-buffered MEMcontaining 2.5 mg/ml lipoprotein-deficient serum (LPDS).19
Experiments at 4°C were carried out in the same mediumbuffered with 4-(1 -hydroxyethyl)-1 -piperazineethanesulfonicadd (HEPES) and incubated in air.
Determination of 128l-labeled UpoprotelnBinding and Metabolism
Macrophages were incubated with 12sl-pigeon /3-VLDLor 12Sl-rabbit /3-VLDL at the protein concentrations indi-cated in the legends of the figures and tables. Cell surfacebinding was determined after 3 hours at 4°C, while lipo-protein uptake and degradation were measured after5 hours at 37°C, unless indicated otherwise. Preliminaryexperiments showed that equilibrium of binding at 4°C for all
by guest on July 21, 2018http://atvb.ahajournals.org/
Dow
nloaded from

RECEPTORS ON PIGEON MACROPHAGES Adelman and St. Clair 675
Table 1. Composition of Pigeon and Rabbit 0-Very Low Density LJpoproteln
Source
Rabbit 0-VLDL
WC /3-VLDL(N=6)
SR0-VLDL(N=3)
Plasma chol(mg/dl)
2249±215
1872±376
1361±189
Prot
5.1 ±0.31
5.6+0.42
6.2±0.38
PL
12.9+0.70
15.4±0.43
15.4±0.82
% composition
TG
4.5+0.88
5.9±1.01
8.8±2.62
UC
10.8±0.40
13.0±0.25
11.8±0.95
CE
66.7±1.33
60.1 ±1.01
57.8±2.34
Chol/Prot
10.2±0.57
8.80±0.73
7.40±0.13
0-VLDL were isolated from the pooled plasma from two to four cholesterol-fed rabbits and three to five chdesterol-fed pigeons for eachpreparation. The results are the means±SEM for the indicated number of /3-VLDL preparations analyzed for each spedes.
Prot=protein, PL=phospholipld, TG=trlglyceride, UC=unestertfled cholesterol, CE=cnc4esteryl ester, Chol/Prot=total cholesterol/protein, /3-VLDL=beta-migrating very low density llpoprotelns.
lipoproteins was reached by 2 hours. After incubationsat 37°C, an aliquot of culture medium was taken for thedetermination of lipoprotein degradation by measure-ment of TCA-soluble, noniodide ^ l . 2 0 The cell layer waswashed five times with phosphate-buffered saline (PBS)containing 2 mg/ml BSA, then three times with PBS. Thecells were digested with 0.5 N NaOH for a minimum of30 minutes, and an aliquot was counted in a Tracor 1185gamma counter to determine 125I radioactivity. A secondaliquot was taken from each dish for protein determin-ation.21 The results were corrected for degraded lipopro-tein in control dishes incubated without cells. Specificbinding or metabolism was defined as the differencebetween the amount of 125l-lipoprotein that was bound,internalized, or degraded in the absence or presence ofunlabeled homologous lipoprotein.
Cholesterol Mass and EsterlflcatlonCells for experiments to measure cholesterol mass or
esterificatjon were incubated as indicated in the tablesand figures. The cell monolayers were washed as indi-cated above and were scraped from the dishes into PBS.Cells were disrupted by sonication, and aliquots weretaken for protein determination and for llpid extraction bythe method of Bligh and Dyer.22 The total and freecholesterol mass were determined by gas-liquid chroma-tography as described previously14 by the method ofIshikawa et al.23 with stigmasterol as an internal standard.Cholesterol estertfication was determined by the incorpo-ration of 14C-oleate into cholesteryl esters after incubationof the cells with lipoproteins in the presence of a 14C-ole-ate/albumin substrate (6:1 oleate/aJbumin molar ratio,2 jiCi/dish, 11 247 dpm/nmol)24 for the times indicated inthe tables and figures. 3H-cholesterol was added during lipidextraction as an internal standard. Upids were separated byTLC on silica gel plates in a solvent system consisting of ahexane/ethyl ether/glacial acetic add, 146:50:4 (vol/vol/vol), and the cholesterol and cholesteryl ester fractionswere identified by comparison to authentic cholesterol andcholesteryl oleate standards. 3H and 14C were counted in aBeckman LS 230 liquid scintillation counter.
Sodium Dodecyl Sulfate-GradlentGel Electrophoresls
Apoproteins from isolated lipoproteins were separatedby electrophoresis as previously described,7 except thatPharmacia pre-poured gradient gels (4% to 30%) were
used after an initial equilibration of the gels with sodiumdodecyl surfate (SDS) by pre-running them for 2 hours at100 volts. We used the buffers described by Laemmli,25
which contained 1% SDS. Lipoproteins were dialyzedagainst deionized water and then delipidated in ether/ethanol.26 The samples were resolubilized in 3% SDS and5% /3-mercaptoethanol at 100°C for 2 minutes. The sam-ples were allowed to cool, and a solution containingsucrose and bromphenol blue (final concentrations of8% sucrose and 0.002% bromphenol blue) was added toeach sample. Aliquots of 40 /xg of protein were applied toeach lane, and the separations were carried out at 15°C.
ResultsCharacteristics of Pigeon and Rabbit p-VeryLow Density LJpoproteln
The composition of representative samples of /3-VLDLfrom cholesterol-fed pigeons and rabbits is shown inTable 1. The /3-VLDLs from both species were rich incholesteryl esters. Cholesteryl esters accounted for58% to 60% of the mass of pigeon /J-VLDL and 67% of themass of rabbit /3-VLDL. The percentages of total protein,phospholipid, and unesterified cholesterol were similar forboth rabbit and pigeon 0-VLDL On the average, rabbit0-VLDL had a greater cholesterol/protein ratio than pigeon/3-VLDL. The percent cholesterol ester and the cholesterol/protein ratio were positively correlated with plasma cho-lesterol concentrations for both pigeons and rabbits (datanot shown).
The apoproteins of rabbit and WC pigeon /3-VLDL arecompared in Figure 1. The major apoproteins of rabbit/3-VLDL are apo B (B-100 and B-48) and apo E. The majorapoproteins of pigeon £-VLDL correspond in size to apoB-100 and apo A-l, with several unidentified proteins alsopresent. In marked contrast to rabbit 0-VLDL, pigeon/3-VLDL contained no detectable apo E as demonstratedby this intentionally overloaded gel. The lack of apo E is aconsistent finding for all pigeon lipoproteins, whetherisolated from fed or fasted, normal or hypercholester-olemic, WC or SR pigeons.7
Binding and Metabolism of Rabbit p-Very LowDensity LJpoproteln by Pigeon Macrophages
0-VLDL from cholesterol-fed rabbits binds to a receptoron mammalian macrophages that recognizes apo E.89
by guest on July 21, 2018http://atvb.ahajournals.org/
Dow
nloaded from

676 ARTERIOSCLEROSIS VOL 9, No 5, SEPTEMBER/OCTOBER 1989
apoB
94,000
67,000
43,000
30,000
20,100
14,400
apoAII.C
M.W.
Stds.
RabbitBVLDL
PigeonBVLDL
Figure 1. Apoproteins of ^-migrating very low density llpopro-telns (0-VLDL) from cholesterol-fed rabbits and White Carneaupigeons. Apoproteins (apo) from the d< 1.006 g/ml (/3-VLDL)fraction were separated by sodium dodecyi sulfate (SDS) gradi-ent gel (4% to 30%) electrophoresls. Forty micrograms of proteinwas applied to each lane of the gel. The molecular weightstandards were composed of (Pharmacia LMW standards) phos-phoryiase b, 94 000; albumin, 67 000; ovalbumin, 43 000; car-bonic anhydrase, 30 000; trypsin Inhibitor, 20 100; and lactalbu-mln, 14 400. The location of the mammalian apolipoprotelns,apo B, apo E, apo A-l, apo A-ll, and apo C are shown.
Since pigeon /3-VLDL lacks apo E, yet is taken up by aspecific receptor-mediated process by pigeon macrc-phages,6 we determined whether apo E-rich rabbit /3-VLDLwould bind and be metabolized by pigeon macrophagesand whether the presence of apo E would enhance itsbinding relative to pigeon 0-VLDL Figure 2 shows surfacebinding at 4°C of rabbit and pigeon /3-VLDL to the samebatch of pigeon macrophages. As seen in this repre-sentative experiment, rabbit /3-VLDL bound to WC and SRmacrophages with saturable kinetics. Scatchard analysis
500
400
300
200
100
WC Macrophages
Rabbit fl-VLDL" ° Total binding
• Non-specific binding• Specific binding
Pigeon fl-VLDL" Specific binding
500
400
300
200
100
Pound (nflffnol i
SR Macrophages
&25 25 50 100
0-VLDL Concentration (eg/ml)
Rgure 2. Concentration dependence of binding of 126l-labeledrabbtt and pigeon ^-migrating very low density lipoproteins(/3-VLDL) by White Cameau (WC) and Show Racer (SR) macro-phages. Cells were incubated for 3 hours at 4°C with theIndicated concentrations of 125l-labeled 0-VLDL isolated fromcholesterol-fed WC pigeons or rabbits. Specific binding wascalculated as the difference between binding in the absence orpresence of 1 mg/ml unlabeled homologous 0-VLDL Total,specific, and non-specific binding are shown for rabbit /3-VLDL,while only specific binding is shown for pigeon /3-VLDL Theresults are the means of triplicate determinations at each pointThe inserts show the Scatchard plots for the specific binding datafor rabbit and pigeon 0-VLDL
of the specific binding data (Figure 2 inserts) suggestedthat rabbit /3-VLDL bound to a single class of sites ofrelatively low affinity, but high capacity. The apparentdissociation constant (Kd), calculated from four individualexperiments similar to those described in Fig-ure 2, was 100 fuQlm\, with a range of 62 to 123 /xg/ml. Adirect comparison in the same experiment with rabbit andpigeon ^VLDL demonstrated that pigeon 0-VLDL boundto at least two classes of binding sites on pigeon macro-phages. The initial high-affinity site, estimated from thefirst four points on the Scatchard curve, gave an apparentKd of 7±4 fjuQlm\ (N=4). The low-affinity site, estimatedfrom the final three points on the Scatchard curve, hadsimilar binding kinetic to the low-affinity site that boundrabbit 0-VLDL The results with pigeon /3-VLDL are con-sistent with our previously published studies demonstrat-ing the curvilinear nature of the Scatchard plot.6
Even though rabbit /3-VLDL bound to pigeon macro-phages with much lower affinity than pigeon /3-VLDL, itwas efficiently internalized and degraded by saturableprocesses in both WC and SR macrophages (Figure 3).
by guest on July 21, 2018http://atvb.ahajournals.org/
Dow
nloaded from
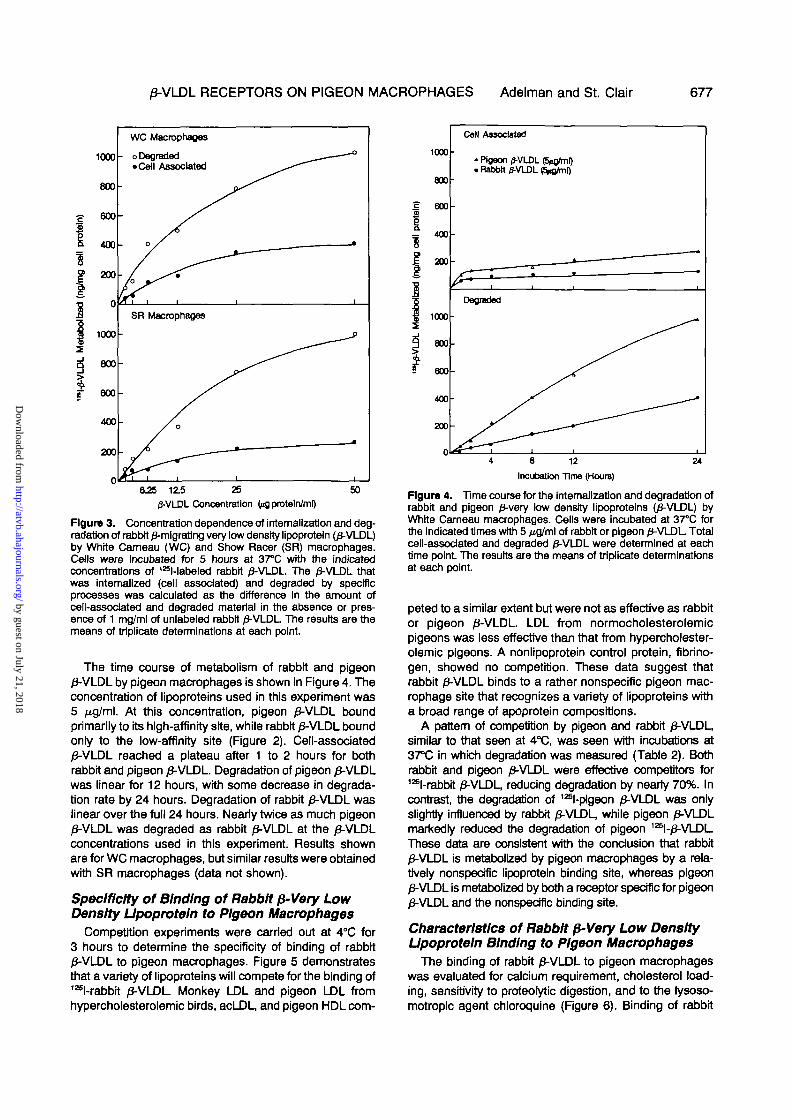
0-VLDL RECEPTORS ON PIGEON MACROPHAGES Adelman and St. Clair 677
WC Macrophages
025 12.5 260-VLDL Concentration (?g proteln/ml)
50
Figure 3. Concentration dependence of intemallzation and deg-radation of rabbit /3-mlgrating very low density lipoprotein (£-VLDL)by White Cameau (WC) and Show Racer (SR) macrophages.Ceils were incubated for 5 hours at 3TC with the indicatedconcentrations of 12Sl-labeled rabbit /3-VLDL. The /3-VLDL thatwas internalized (cell associated) and degraded by specificprocesses was calculated as the difference In the amount ofcell-associated and degraded material in the absence or pres-ence of 1 mg/ml of unlabeled rabbit /3-VLDL The results are themeans of triplicate determinations at each point.
The time course of metabolism of rabbit and pigeon0-VLDL by pigeon macrophages is shown in Figure 4. Theconcentration of lipoproteins used in this experiment was5 Mg/ml. At this concentration, pigeon /3-VLDL boundprimarily to its high-affinity site, while rabbit /3-VLDL boundonly to the low-affinity site (Figure 2). Cell-associated/3-VLDL reached a plateau after 1 to 2 hours for bothrabbit and pigeon /3-VLDL. Degradation of pigeon 0-VLDLwas linear for 12 hours, with some decrease in degrada-tion rate by 24 hours. Degradation of rabbit /J-VLDL waslinear over the full 24 hours. Nearly twice as much pigeon0-VLDL was degraded as rabbit 0-VLDL at the /3-VLDLconcentrations used in this experiment. Results shownare for WC macrophages, but similar results were obtainedwith SR macrophages (data not shown).
Specificity of Binding of Rabbit p-Very LowDensity Upoproteln to Pigeon Macrophages
Competition experiments were carried out at 4°C for3 hours to determine the specificity of binding of rabbit0-VLDL to pigeon macrophages. Figure 5 demonstratesthat a variety of lipoproteins will compete for the binding of125l-rabbrt /3-VLDL Monkey LDL and pigeon LDL fromhypercholesterolemic birds, acLDL, and pigeon HDL com-
Cell Associated
»Pigeon 0-VLDL (5rt>frnl). Rabbtt 0-VLDL ^ m l )
8 12
Incubation Time (Hours)
24
Figure 4. Time course for the intemallzatJon and degradation ofrabbit and pigeon 0-very low density lipoproteins (/J-VLDL) byWhite Carneau macrophages. Cells were incubated at 37°C forthe indicated times with 5 jig/ml of rabbit or pigeon 0-VLDL Totalcell-associated and degraded 0-VLDL were determined at eachtime point The results are the means of triplicate determinationsat each point.
peted to a similar extent but were not as effective as rabbitor pigeon /3-VLDL. LDL from normocholesterolemicpigeons was less effective than that from hypercholester-olemic pigeons. A nonlipoprotein control protein, fibrino-gen, showed no competition. These data suggest thatrabbit 0-VLDL binds to a rather nonspecific pigeon mac-rophage site that recognizes a variety of lipoproteins witha broad range of apoprotein compositions.
A pattern of competition by pigeon and rabbit /3-VLDL,similar to that seen at 4°C, was seen with incubations at37°C in which degradation was measured (Table 2). Bothrabbit and pigeon /3-VLDL were effective competitors for126l-rabbit /J-VLDL, reducing degradation by nearly 70%. Incontrast, the degradation of 126l-pigeon /3-VLDL was onlyslightty influenced by rabbit 0-VLDL, while pigeon 0-VLDLmarkedly reduced the degradation of pigeon 12S1-/3-VLDLThese data are consistent with the conclusion that rabbit/3-VLDL is metabolized by pigeon macrophages by a rela-tively nonspecific lipoprotein binding site, whereas pigeon/3-VLDL is metabolized by both a receptor specific for pigeon0-VLDL and the nonspecific binding site.
Characteristics of Rabbtt p-Very Low DensityUpoproteln Binding to Pigeon Macrophages
The binding of rabbit 0-VLDL to pigeon macrophageswas evaluated for calcium requirement, cholesterol load-ing, sensitivity to proteolytic digestion, and to the lysoso-motropic agent chloroquine (Figure 6). Binding of rabbit
by guest on July 21, 2018http://atvb.ahajournals.org/
Dow
nloaded from

678 ARTERIOSCLEROSIS VOL 9, No 5, SEPTEMBER/OCTOBER 1989
"t-Rabblt-0-VLDL (3.16 ufl/ml), 3 hr.F 4*C
Mlbrinogen
pigeon NLDL
LDLacetyl LDLpigeon HLDLpigeon HDL
V rabbit 0-VLDLx pigeon /5-VLDL
Bound3 hr, 4°C
Degraded5 hr., 37 °C
40 80 120 160 200 240 280 320
Competitor Concentration
Figure 5. Competition for binding of 125Mabeled rabbit 0-verylow density lipoprotelns (J3-VLDL). 1S5l-labeted rabbit 0-VLDL wasadded to each dish of White Carneau macrophages at a concen-tration of 3.16 >ig/ml In the presence of the indicated unlabeledcompetitors over a concentration range of haif log units from0.316 to 316 /^ /ml . Binding of 128Mabeled rabbit /3-VLDL In theabsence of competitor was 66.5±5.2 ng/mg cell protein(means±SD, N=6). Each point represents the percent change inbinding of 12Sl-labeled rabbit /3-VLDL from the value withoutadded competitor. Duplicate dishes were used for each concen-tration of the various competitors. NLDL and HLDL refer to LDLIsolated from normal or cholesterol-fed pigeons, respectively.
Table 2. Competition for Degradation of 125MabeledRabbit and Pigeon 0-Very Low Density Upoproteln InWhite Carneau Pigeon Peritoneal Macrophages
Competitor
None
Pigeon /3-VLDL(100 jig/ml)
Rabbit 0-VLDL(lOOMfl/ml)
12BNabeled pigeon/3-VLDL
288±34(100%)
61 ±8(21%)
229±36(80%)
12»l-labeled rabbit0-VLDL
208+20(100%)
73+16(35%)64±6(31%)
Values are given as nanograms of 12Bl-0-VLDL degraded/milligram of protein and are the means+SD for quadruplicateIncubations. The values in parentheses represent the percent ofdegradation occurring in the presence of the competitor relativeto no competitor.
125l-tebeled rabbit or pigeon /3-VLDL (5 Mg/ml) were incubatedwith White Carneau pigeon peritoneal macrophages for 5 hoursat 37°C in the presence or absence of the indicated unlabeled(competitor) /3-VLDL, and degradation was determined.
0-VLDL=p-mlgrating very low density lipoprotein.
Calcium Pronase Choi. ChloroquineLoading
•ng/mgCell Protein
Figure 6. The effects of calcium, pronase, chloroquine, andcholesterol loading on the rabbit 0-migratlng very low densitylipoprotein (/3-VLDL) receptor pathway in White Cameau pigeonmacrophages. The amount of 125l-labeled rabbit 0-VLDL bound ordegraded Is shown within each bar, and the percent change bythe bar height. Cells were incubated with 5 ng/ml of 12Sl-labeledrabbit /3-VLDL In the presence or absence of 100 /xg/ml ofunlabeled rabbit 0-VLDL Binding and specific degradation wereas described in Methods. To assess calcium dependence, celtswere cooled to 4°C for 1 hour and washed three times wtth coldcalcium-free phosphate-buffered saline (PBS) containing 100 /iMEDTA with or without 2 mM calcium. The 12BI-Iabeled rabbit0-VLDL was added, and the cells were Incubated at 4°C for3 hours. For pronase sensitivity, cell monolayers were washedthree times wtth PBS at 37°C, serum-free Eagle's MinimumEssential Medium (MEM) wtth or without 3 figlm\ pronase wasadded, and the cells were incubated at 37°C. After 20 minutes,the cells were chilled on Ice for 1 hour and washed three timeswtth cold PBS. Fresh MEM (4°C) containing 2.5 mg/ml lipoprotein-deficient serum (LPDS) plus '^-labeled rabbit /3-VLDL, with orwithout unlabeled rabbit 0-VLDL, was added, and specific bind-ing was determined after 3 hours. The effect of cholesterolloading on rabbit /3-VLDL binding was determined after pre-incubation of cells wtth pigeon /3-VLDL (50 HQ proteln/ml) for48 hours. The cells were washed thrice with PBS and Incubatedwith MEM containing 2.5 mg/ml of LPDS for 2 hours at 37°C toallow internalization of surface-bound /3-VLDL The cells werechilled at 4°C for 1 hour, washed thrice with PBS, and incubatedwith 12Sl-labeled rabbit /3-VLDL with or without unlabeled rabbit/3-VLDL at 4°C for 3 hours to determine specific binding. Cellularcholesterol content was determined in parallel dishes as describedIn Methods. Total cholesterol content increased from 22 /ig/mgcell protein before loading to 114 /ig/mg cell protein after loading.Sensitivity of degradation to chloroquine was determined afterpretreatment of cells for 1 hour at 37°C wtth 50 /xM chloroquine.12SHabeled rabbit 0-VLDL with or without unlabeled rabbit 0-VLDLwas added to the dishes in the presence of 50 /JA chloroquineand incubated for 5 hours at 37°C. Degradation was determinedas described In Methods. The results are the means of triplicatedeterminations.
0-VLDL was diminished approximately 60% in the absenceof caJcium, suggesting some requirement for divaJentcations. Degradation of rabbit /3-VLDL appeared to belargely lysosomaJ, as seen by the 73% reduction indegradation when incubated in the presence of 50 ^Mchloroquine. Both of these results are similar to that seenpreviously wtth pigeon /3-VLDL binding to pigeon macro-phages.0 In contrast, the binding of rabbit /3-VLDL was
neither influenced by pretreatment of the cells with pro-nase nor was it inhibited by cholesterol loading.
Ability of Rabbit p-Very Low DensityUpoproteln to Deliver Cholesterolto Pigeon Macrophages
To determine whether the metabolism of rabbit /3-VLDLby pigeon macrophages would result in cholesterol deliv-
by guest on July 21, 2018http://atvb.ahajournals.org/
Dow
nloaded from
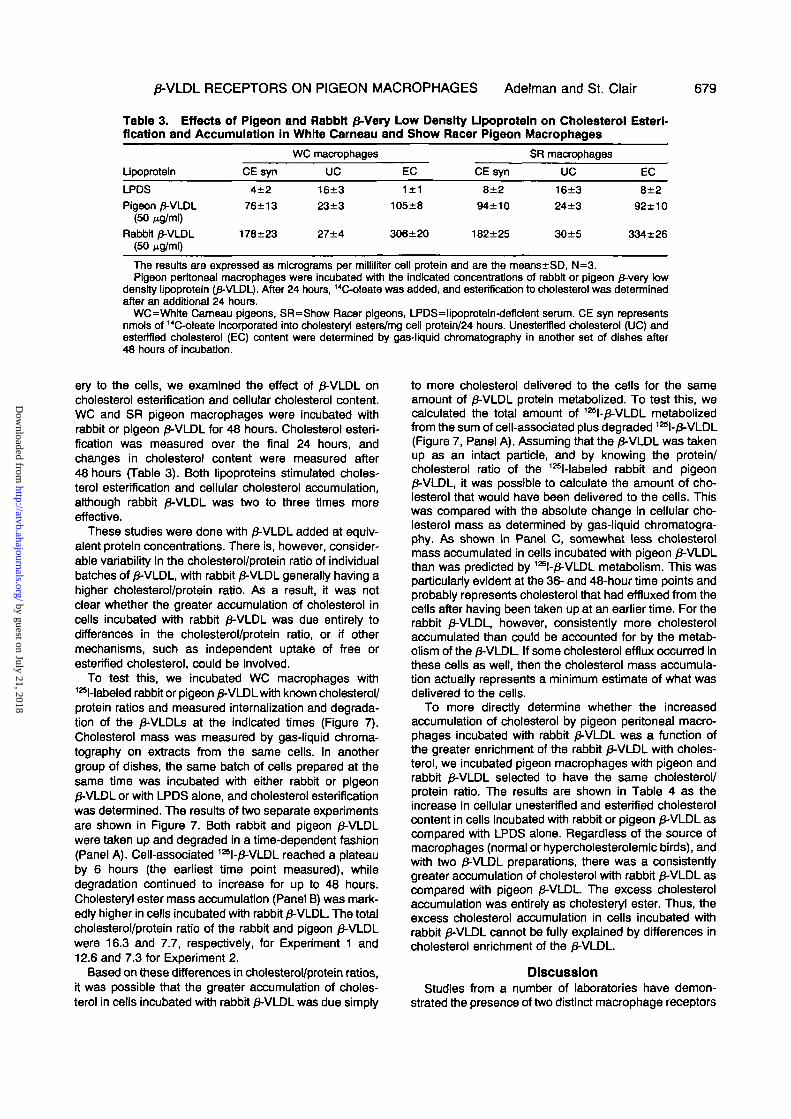
0-VLDL RECEPTORS ON PIGEON MACROPHAGES Adelman and St. Clair 679
Table 3. Effects of Pigeon and Rabbit 0-Very Low Density Upoproteln on Cholesterol Esterl-fication and Accumulation in White Cameau and Show Racer Pigeon Macrophages
Upoproteln
LPDSPigeon /3-VLDL
(50ftg/ml)Rabbit /3-VLDL
CE syn
4±276±13
178±23
WC macrophages
UC
16±323±3
27±4
EC
105±8
306±20
CE syn
8±294±10
182±25
SR macrophages
UC
16±324±3
30±5
EC
8±292±10
334±26
The results are expressed as micrograms per milliliter cell protein and are trie means±SD, N=3.Pigeon peritoneal macrophages were incubated with the indicated concentrations of rabbit or pigeon £-very low
density lipoproteln O-VLDL). After 24 hours, 14C-oleate was added, and esterification to cholesterol was determinedafter an additional 24 hours.
WC=White Cameau pigeons, SR=Show Racer pigeons, LPDS=lipoproteln-deflclent serum. CE syn representsnmols of 14C-oleate Incorporated into cholesteryl esters/mg cell protein/24 hours. Unesterlfied cholesterol (UC) andestertfled cholesterol (EC) content were determined by gas-liquid chromatography in another set of dishes after48 hours of incubation.
ery to the cells, we examined the effect of 0-VLDL oncholesterol esterification and cellular cholesterol content.WC and SR pigeon macrophages were incubated withrabbit or pigeon /3-VLDL for 48 hours. Cholesterol esteri-fication was measured over the final 24 hours, andchanges in cholesterol content were measured after48 hours (Table 3). Both lipoproteins stimulated choles-terol esterification and cellular cholesterol accumulation,although rabbit /3-VLDL was two to three times moreeffective.
These studies were done with /J-VLDL added at equiv-alent protein concentrations. There is, however, consider-able variability in the cholesterol/protein ratio of individualbatches of /3-VLDL, with rabbit /3-VLDL generally having ahigher cholesterol/protein ratio. As a result, it was notclear whether the greater accumulation of cholesterol incells incubated with rabbit /3-VLDL was due entirely todifferences in the cholesterol/protein ratio, or if othermechanisms, such as independent uptake of free oresterified cholesterol, could be involved.
To test this, we incubated WC macrophages with125l-labeled rabbit or pigeon 0-VLDL with known cholesterol/protein ratios and measured internalization and degrada-tion of the 0-VLDLs at the indicated times (Figure 7).Cholesterol mass was measured by gas-liquid chroma-tography on extracts from the same cells. In anothergroup of dishes, the same batch of cells prepared at thesame time was incubated with either rabbit or pigeon/3-VLDL or with LPDS alone, and cholesterol esterificationwas determined. The results of two separate experimentsare shown in Figure 7. Both rabbit and pigeon /3-VLDLwere taken up and degraded in a time-dependent fashion(Panel A). Cell-associated 1ssl-/3-VLDL reached a plateauby 6 hours (the earliest time point measured), whiledegradation continued to increase for up to 48 hours.Cholesteryl ester mass accumulation (Panel B) was mark-edly higher in cells incubated with rabbit 0-VLDL The totalcholesterol/protein ratio of the rabbit and pigeon /3-VLDLwere 16.3 and 7.7, respectively, for Experiment 1 and12.6 and 7.3 for Experiment 2.
Based on these differences in cholesterol/protein ratios,it was possible that the greater accumulation of choles-terol in cells incubated with rabbit /3-VLDL was due simply
to more cholesterol delivered to the cells for the sameamount of /J-VLDL protein metabolized. To test this, wecalculated the total amount of 12Sl-£-VLDL metabolizedfrom the sum of cell-associated plus degraded 12Sl-/3-VLDL(Figure 7, Panel A). Assuming that the /3-VLDL was takenup as an intact particle, and by knowing the protein/cholesterol ratio of the 125l-labeled rabbit and pigeon/3-VLDL, it was possible to calculate the amount of cho-lesterol that would have been delivered to the cells. Thiswas compared with the absolute change in cellular cho-lesterol mass as determined by gas-liquid chromatogra-phy. As shown in Panel C, somewhat less cholesterolmass accumulated in cells incubated with pigeon /3-VLDLthan was predicted by 126l-0-VLDL metabolism. This wasparticularly evident at the 36- and 48-hour time points andprobably represents cholesterol that had effluxed from thecells after having been taken up at an earlier time. For therabbit 0-VLDL, however, consistently more cholesterolaccumulated than could be accounted for by the metab-olism of the 0-VLDL If some cholesterol efflux occurred inthese cells as well, then the cholesterol mass accumula-tion actually represents a minimum estimate of what wasdelivered to the cells.
To more directly determine whether the increasedaccumulation of cholesterol by pigeon peritoneal macro-phages incubated with rabbit /3-VLDL was a function ofthe greater enrichment of the rabbit /3-VLDL with choles-terol, we incubated pigeon macrophages with pigeon andrabbit 0-VLDL selected to have the same cholesterol/protein ratio. The results are shown in Table 4 as theincrease in cellular unesterified and esterified cholesterolcontent in cells incubated with rabbit or pigeon /3-VLDL ascompared with LPDS alone. Regardless of the source ofmacrophages (normal or hypercholesterolemic birds), andwith two 0-VLDL preparations, there was a consistentlygreater accumulation of cholesterol with rabbit 0-VLDL ascompared with pigeon /3-VLDL The excess cholesterolaccumulation was entirely as cholesteryl ester. Thus, theexcess cholesterol accumulation in cells incubated withrabbit /3-VLDL cannot be fully explained by differences incholesterol enrichment of the 0-VLDL.
DiscussionStudies from a number of laboratories have demon-
strated the presence of two distinct macrophage receptors
by guest on July 21, 2018http://atvb.ahajournals.org/
Dow
nloaded from

680 ARTERIOSCLEROSIS VOL 9, No 5, SEPTEMBER/OCTOBER 1989
Experiment 1 Experiment 2A. Metabolism
6 12 24' 38 ' 6 24
B. Cholesterol Accumulation
n - 3
- /
—roil As
—f48
8 12 24 38 48
C. Choi, from /3VLDL Metabolism vs. Choi. Accumulation
8 12 24 36Incubation Time (hour*)
D. Cholesterol Esterlflcatlon160
•o=plgeon/3VLDL g c• D= rabbit /3VLDL £ S
FC = FreeChol. * £ 120
EC = Estertfied Choi. | g
8 48Incubation Time (hour*)
Figure 7. Relationship of ^-migrating very low density lipo-protein (/J-VLDL) metabolism to cholesterol accumulation andestertflcation in White Carneau pigeon macrophages incubatedwith rabbit or pigeon 0-VLDL. Macrophages were pre-incubated with lipoproteln-deficient serum (LPDS) (2.5 mgprotein/ml) for 48 hours. In Experiment 1, cells were incubatedat 37°C in medium containing LPDS and 12Sl-labeled rabbit orpigeon 0-VLDL (40 \i% proteln/ml) for the indicated times. InExperiment 2, the /3-VLDLs were added at 50 /ig proteln/ml./3-VLOL metabolism and cholesterol mass accumulation weredetermined on the same dishes of cells. Another set of dishesusing the same cells was prepared at the same time formeasurement of cholesterol esteriflcation. The mass of choles-terol (unesterified plus esterifled) that accumulated was com-pared with the amount of cholesterol that was calculated to bedelivered by 0-VLDL metabolism (cell-associated plus degrad-ed). Cholesterol delivered by 0-VLDL metabolism was deter-mined from the total 12Sl-0-VLDL protein metabolized times thecholesterol/protein ratio of the j8-VLDL The results are themeans of duplicate or triplicate dishes at each time point
that mediate the uptake of lipoproteins, the scavengerreceptor, and the 0-VLDL receptor. The scavenger recep-tor has been the most extensively studied (for a review,see reference 27). This receptor has been shown torecognize a variety of "abnormal" lipoproteins and to bepresent on monocyte/macrophages and peritonea) mac-rophages from virtually every species tested,27 includingpigeons.8 The /3-VLDL receptor, although initially thoughtto be distinct from the LDL receptor,27.28 is now believed tobe a form of the LDL receptor.1011 Nevertheless, sincethere are some differences in the binding characteristicsof 0-VLDL to this receptor relative to the LDL receptorfound on skin fibroblasts, and since we have no directevidence in the pigeon that the /3-VLDL receptor is a formof the LDL receptor, we will continue to refer to it as the/3-VLDL receptor.
We have recently reported that pigeon peritoneal mac-rophages metabolize /3-VLDL by what appears to be areceptor-mediated mechanism.8 Several of the character-istics of /3-VLDL binding and metabolism by pigeon peri-toneal macrophages are different from the mammalian0-VLDL receptor. As shown previously and extended inthe present report, it is clear that there are at least twoclasses of binding sites on pigeon peritoneal macro-phages for pigeon 0-VLDL. This is most apparent from thecurvilinear Scatchard plot. If the Scatchard curve is dividedinto its high- and low-affinity components (Figure 2), thenthe apparent Kd for binding and the B max can beestimated to be approximately 7 /ig/ml and 200 ng/mg cellprotein for the high-affinity site and 100 ̂ g/ml and about600 ng/mg cell protein for the low-affinity site. Binding tothe high-affinity site (hereafter referred to as the pigeon/3-VLDL receptor), is specific for pigeon /3-VLDL and LDLBased on competition studies, the high-affinity pigeon/3-VLDL receptor does not bind acLDL, methyl LDL,mammalian LDL, HDL, or rabbit /3-VLDL. Uptake by thepigeon 0-VLDL receptor requires calcium and is down-regulated by cholesterol loading. The receptor appears tobe a protein, as seen by its susceptibility to pronasedigestion. The 0-VLDL taken up by this receptor is deliv-ered to the lysosomes for degradation, since lysosomo-tropic agents, such as chloroquine, effectively inhibitdegradation. Uptake by the pigeon 0-VLDL receptor iseffective in delivering cholesterol to macrophages and instimulating cholesterol esterification. Thus, the pigeon/3-VLDL receptor resembles the mammalian /3-VLDL recep-tor in many of its characteristics.
There is, however, at least one major difference betweenthe pigeon and mammalian /3-VLDL receptor. Binding of0-VLDL to the mammalian 0-VLDL receptor is mediatedprimarily by apo E,88 and although LDL will bind, it bindswith an affinity that is approximately 100-fold less than/3-VLDL Pigeon /3-VLDL, like all pigeon lipoproteins, doesnot contain apo E.7 As a result, apo E cannot be mediatingthe binding to the pigeon /3-VLDL receptor. Furthermore,rabbit /3-VLDL, a lipoprotein rich in apo E (Figure 1), doesnot compete for binding of pigeon /3-VLDL to the pigeon0-VLDL receptor. These results clearly indicate that apo Eis not mediating binding of pigeon 0-VLDL to the /3-VLDLreceptor, although it does not exclude the possibility thatother pigeon apolipoproteins (Figure 1) might participate
by guest on July 21, 2018http://atvb.ahajournals.org/
Dow
nloaded from

0-VLDL RECEPTORS ON PIGEON MACROPHAGES Adelman and St. Clair 681
Table 4. Cholesterol Accumulation by White Cameau Pigeon Peritoneal Macrophages Incubatedwith Pigeon or Rabbit 0-Very Low Density LJpoprotein with Equivalent Cholesterol/Protein Ratio
Increase in chol content over controls
Cholesterol/protein ratio
Experiment 1
Pigeon 0-VLDL
Rabbit 0-VLDL
Experiment 2
Pigeon 0-VLDL
Rabbit p-VLDL
8.3
8.3
9.910.0
Unesterified
(MQ chol/mg cell
5.9
9.2
0
3.4
Estertfled
protein)
40.2
72.8
12.0
74.0
The data for control cells are the means of quadruplicate incubations, while the data for pigeon and rabbit /3-VLDLare the means of duplicate Incubations.
White Cameau pigeon peritoneal macrophages were incubated with medium containing llpoprotein-deflclentserum (LPDS) (2.5 mg protein/ml) for 48 hours. Cells were then incubated for 24 hours at 37°C in medium containingLPDS and rabbit or pigeon /3-very low density lipoprotein (0-VLDL) at 40 /ig protein/ml. The /3-VLDLs were selectedto have equivalent cholesterol/protein ratios within the same experiment Cells were analyzed for cholesterol massby gas-liquid chromatography. Control cells were incubated In LPDS medium for 48 hours and then harvested forcholesterol mass. For Experiment 1, control cells contained 42.4±3.51 (mean±SD) and 7.5±2.85 NG of unestertfiedand estertfied cholesterol, respectively, per milligram cell protein. For Experiment 2, these control values were46.7±2.87 and 71.4±14.6 /*g per milligram cell protein, respectively. The macrophages from Experiment 2 werefrom cholesterol-fed birds, which is why the control cells were enriched in cholesteryl esters.
in binding. Cynomolgus monkey LDL, whose principalapoprotein is apo B, does not compete for binding to thepigeon /3-VLDL receptor.6 Thus, the pigeon /3-VLDL recep-tor, even though resembling the mammalian /3-VLDLreceptor in a number of ways, does not recognize eithermammalian apo B or apo E.
Although rabbit /3-VLDL does not bind to the pigeon/3-VLDL receptor, Scatchard analysis indicates that it doesbind to a single class of sites on pigeon peritonea)macrophages with low affinity (apparent Kd approximately100 /xg/ml), but high capacity (B max approximately600 ng/mg cell protein). Evidence that pigeon and rabbitp-VLDL are binding to the same low-affinity site is basedon their similar binding affinity and the ability of pigeon0-VLDL to compete for binding with rabbit 0-VLDL Thelow-affinity site appears to recognize not only rabbit andpigeon /3-VLDL, but virtually all other lipoproteins tested,including LDL, HDL, and acLDL. Further evidence that thelow-affinity site is distinct from the high-affinity pigeon/3-VLDL receptor is the fact that binding of rabbit /3-VLDLto the low-affinity site was not affected by pronase diges-tion, nor was binding decreased by cholesterol loading.
The characteristics of the low-affinity site are similar towhat has been referred to by others by a variety of namesincluding the "EDTA-resistant receptor"29 and "the lipopro-tein binding site."30 This lipoprotein binding site has beendescribed for a number of cells and tissues including,hepatocytes,30 liver membranes,13'31 and macrophages.32-33
Even pigeon smooth muscle cells and skin fibroblasts inculture that do not express LDL receptors bind significantamounts of lipoproteins to a site with similar characteristics.34
In pigeon smooth muscle cells and fibroblasts, binding to thelipoprotein binding site does not result in intemalizatJon ofthe lipoprotein.34 In contrast in mammalian hepatocytes,intemalizatJon and degradation of LDL and HDL occurs afterbinding to the lipoprotein binding site,30 as it does with rabbit0-VLDL in human monocyte macrophages.35 Likewise, inpigeon macrophages, rabbit /3-VLDL, which appears to bind
only to the lipoprotein binding site, is efficiently internalizedand degraded. Degradation occurs in the lysosomes, asseen from the inhibition of degradation by chloroquine.
Given the role of macrophages in the pathogenesis ofatherosclerosis, it is important to know whether the lipo-protein binding site can play a role in cholesterol accumu-lation by these cells. As seen from the data in Figure 7 andTables 3 and 4, rabbit 0-VLDL was consistently moreeffective than pigeon /3-VLDL in promoting cholesterolaccumulation by pigeon peritoneal macrophages. Someof this difference could be accounted for by a greatercholesterol/protein ratio of the rabbit /3-VLDL used insome of the experiments. However, even when the p-VLDLwas selected to have the same cholesterol/protein ratio,or when the difference in this ratio was accounted for inthe calculation of cholesterol delivery to cells, rabbit0-VLDL consistently caused greater accumulation of cho-lesterol in cells. This difference could not be explained byuptake of the intact 0-VLDL particles and their degrada-tion. In arriving at this conclusion, we have assumed thatall of the cholesterol that entered the cells during the48 hours of incubation remained in the cell. This isprobably not true, however, particularly for the later timepoints, since the LPDS in the culture medium, and per-haps also the /J-VLDL itself, may facilitate the efflux ofsome of the cholesterol that entered the cells at an earliertime. As a result, the data on cholesterol accumulationmust be viewed as a minimum estimate of the amount ofcholesterol entering on /3-VLDL. Cells incubated withpigeon 0-VLDL at a concentration of 50 /xg/ml accumu-lated cholesterol at a rate equivalent to, or slightly lessthan, could be accounted for by /3-VLDL metabolism. Onthe other hand, cells incubated with rabbit 0-VLDL con-sistently accumulated from 30% to 50% more cholesterolthan could be accounted for by 0-VLDL metabolism. Onepossible explanation for this effect is that binding to thelipoprotein binding site on pigeon peritoneal macrophagesnot only results in the internalization and degradation of
by guest on July 21, 2018http://atvb.ahajournals.org/
Dow
nloaded from

682 ARTERIOSCLEROSIS V O L 9, No 5, SEPTEMBER/OCTOBER 1989
0-VLDL, but also facilitates the uptake of cholesterolindependent of the degradation of the entire 0-VLDLparticle. Such a phenomenon has been described for theuptake of HDL cholesterol in the liver and steroidogenictissues, in rat hepatocytes and adrenal cells in culture, inhuman and ratfibroblasts,38-4Ofor LDLin skin fibroblasts,14
and for arterial smooth muscle cells.1441 In steroidogenictissue, the process of independent uptake of cholesterolappears secondary to binding to the so-called HDLreceptor.40 Curiously, the HDL receptor has many of thesame characteristics as the lipoprotein binding site, andthey may, in fact, be one and the same.42 The specificnature of this binding site is uncertain, however. Someresearchers have even suggested that HDL may bind tohydrophobic lipid domains on the cell surface rather thanto a specific protein receptor.43
Regardless of the nature of the Interaction of /3-VLDLwith pigeon macrophages, a number of possible mecha-nisms exist for the independent uptake of cholesterol.Surface transfer of unesterified cholesterol between lipo-proteins and cells, and the esterification and storage ofthis cholesterol as cholesteryl esters, has been describedin rat hepatoma cells in culture4449 and in arterial smoothmuscle cells incubated with high concentrations of humanLDL.41 Alternatively, independent uptake of cholesterylesters could occur. Although such a phenomenon hasbeen described in a variety of cells and tissues,3739 themechanism is unclear. Potential mechanisms includefacilitated transfer of cholesteryl esters by transfer pro-teins secreted by macrophages4847 or uptake and retro-endocytosis of /3-VLDL. The latter mechanism has beensuggested for HDL metabolism by macrophages.46
The greater ability of rabbit /3-VLDL to deliver cholesterolto cells, as compared to pigeon /3-VLDL, cannot be explainedby differences In the cholesterol/protein ratio of these lipo-proteins. This may suggest that more subtle differences inthe surface conformation of rabbit and pigeon /3-VLDL areimportant in mediating independent cholesterol transfer intomacrophages. A more likely explanation, however, is thatthe difference is related to the particular binding site on themacrophage that mediates 0-VLDL uptake. The majority ofpigeon /3-VLDL (unless very high concentrations are used)appears to be taken up and degraded by the high-affinitypigeon /3-VLDL receptor. This process is tightty regulated bycellular cholesterol concentrations and apparently does notpromote the independent uptake of cholesterol. Rabbit0-VLDL is not recognized by this regulated receptor butinstead is taken up by the lipoprotein binding site, as ispigeon 0-VLDL when present at high concentrations. Thelipoprotein binding site promotes the uptake and degrada-tion of the intact /3-VLDL, as well as perhaps the indepen-dent uptake of cholesterol. Thus, the relative proportion ofthe /3-VLDL taken up by the two binding sites on pigeonmacrophages will likely determine the extent to which cho-lesterol accumulation occurs.
The results of this and previous studies6 have shown thatpigeon peritoneal macrophages have at least three mecha-nisms for uptake of abnormal lipoproteins. These include:the scavenger lipoprotein receptor, the pigeon 0-VLDL recep-tor, and the lipoprotein binding site. The pigeon /3-VLDLreceptor is readily down-regulated by cholesterol loading. As
a result, in the atherosclerotic lesion, one would expect thelipoprotein binding site and the scavenger receptor to pre-dominate. Given the high capacity of the lipoprotein bindingsite, and its ability to recognize a variety of normal andabnormal lipoproteins, it could play an important role in theformation of macrophage foam ceBs in atherosclerotic lesions.Such a mechanism should be particularly efficient in areas ofthe arterial wall where microdomains of very high concen-trations of plasma lipoproteins are found. Although macro-phages from both WC and SR pigeons express pigeon/3-VLDL receptors, it remains to be seen whether differencesin susceptibility to atherosclerosis between the two breeds ofpigeons can be related to the function of the lipoproteinbinding site. The demonstration that rabbit /3-VLDL onlybinds to the low-affinity lipoprotein binding site on pigeonmacrophages provides a useful new tool to further explorethe function of this binding site in the pigeon model.
AcknowledgmentsWe acknowledge the excellent technical assistance of Molly
Leight and Patricia Hester and the assistance of Patricia Brooksand Dorian Seman In the preparation of this manuscript.
References1. St Clair RW. Metabolic changes in the arterial wall associated
with atherosclerosis In the pigeon. Fed Proc 1983:422480-24852. Ross R. The pathogenesis of atherosclerosis—an update. N
Engl J Med 1986:314:488-5003. Poole JCF, Florey HW. Changes In the endothelium of the
aorta and the behavior of macrophages in experimentalatheroma of rabbits. J Pathol Bacteriol 1958:75:245-252
4. Schaffner T, Taylor K, Bantuccl EJ, et al. Arterial foamcells with distinctive immunomorphologlc and histochemicaJfeatures of macrophages. Am J Pathol 1980;100:57-80
5. Fowler S, Shlo H, Haley NJ. Characterization of llpld-ladenaortic cells from cholesterol-fed rabbits. IV. Investigation ofmacrophage-llke properties of aortic ceil populations. LabInvest 1979:41:372-378
6. Adelman SJ, St Clair RW. Lipoprotein metaboflsm by mac-rophages from atherosclerosis-susceptible White Cameau andresistant Show Racer pigeons. J LJpid Res 1988;29:643-656
7. Barakat HA, St Clair RW. Characterization of plasmalipoproteins of grain-fed and cholesterol-fed White Cameauand Show Racer pigeons. J Upld Res 1985;26:1252-1268
8. Innerartty TL, Arnold KS, Welsgraber KH, Mahley RW.Apollpoproteln E Is the determinant that mediates the recep-tor uptake of 0-very low density lipoproteins by mousemacrophages. Arteriosclerosis 1986:6:114-122
9. Bates SR, Coughlln BA, Mazzone T, Borensztajn J, GetzGS. Apoproteln E mediates the interaction of 0-VLDL withmacrophages. J Upld Res 1987;28:787-797
10. Koo C, Wernette-Hammond ME, Innerartty Tl_ Uptake ofcanine 0-very low density lipoproteins by mouse peritonealmacrophages is mediated by a low density lipoprotein recep-tor. J Btol Chem 1986;261:11194-11201
11. Ellsworth JL, Kraemer FB, Cooper AD. Transport of /3-verylow density lipoproteins and chylomicron remnants by mac-rophages is mediated by the low density lipoprotein receptorpathway. J Biol Chem 1987;262:2316-2325
12. Bachorik PS, Livingston JN, Cooke J, Kwtterovlch PO.The binding of low density lipoprotein by liver membranes Inthe pig. Biochem Blophys Res Commun 1976:69:927-935
13. Bachorik PS, Kwtterovlch PO, Cooke JC. Isolation of aporcine liver plasma membrane fraction that binds low den-sity lipoproteins. Biochemistry 1978;17:5287-5299
14. St Clair RW, Mrtschelen JJ, Leight M. Metabolism by cellsIn culture of low-density lipoproteins of abnormal composi-
by guest on July 21, 2018http://atvb.ahajournals.org/
Dow
nloaded from

/3-VLDL RECEPTORS ON PIGEON MACROPHAGES Adelman and St. Clair 683
tion from non-human primates with diet-Induced hypercho-lesterolemla. Biochem Biophys Acta 1980;618:63-79
15. St. Clalr RW, Randolph RK, Joldnen MP, Clarkson TB,Barakat HA. Relationship of plasma lipoprotelns and themonocyte-macrophage system to atherosclerosis severity Incholesterol-fed pigeons. Arteriosclerosis 1986;6:614-626
16. Rudel LL, Leo JA, Morris MD, FaHs JM. Characterization ofplasma lipoproteins separated and purified by agarose-column chromatography. Blochem J 1974; 139:89-95
17. Basu SK, Goldstein JL, Anderson RGW, Brown MS.Degradation of cationlzed low density lipoprotein and regu-lation of cholesterol metabolism in homozygous familialhypercholesterolemia fibroblasts. Proc Nati Acad Sci USA1976;73:3178-3182
18. McFariane AS. Metabolism of plasma proteins. In: MunroHN, Allison JB, eds. Mammalian protein metabolism, vol. 1.New York: Academic Press, 1964:297-341
19. S t Clalr RW, Smith BP, Wood LL Stimulation of choles-terol esterifteation in Rhesus monkey arterial smooth musclecells. Ore Res 1977;40:166-173
20. Goldstein JL, Brown MS. Binding and degradation of lowdensity lipoprotelns by cultured human fibroblasts. Compar-ison of cells from a normal subject and from a patient withhomozygous familial hypercholesterolemia J Biol Chem1974;249:5153-5162
21. Lowry OH, Rosebrough NJ, Fan- AL, Randall RJ. Proteinmeasurement with the Folln phenol reagent. J Biol Chem1951:193:265-275
22. Bllgh EG, Dyer WJ. A rapid method of total lipkl extractionand purification. Can J Blochem Physlol 1959;37:911-917
23. Ishlkawa TT, MacGee J, Morrison JA, Qlueck CJ. Quan-titative analysis of cholesterol in 5 to 20 jd of plasma. J UpldRes 1974:15:286-291
24. Smith BP, St. Clalr RW, Lewis JC. Cholesterol estertflca-tion and cholesteryl ester accumulation In cultured pigeonand monkey arterial smooth muscle cells. Exp Mol Pathol1979:30:190-208
25. Laemmll UK. Cleavage of structural proteins during the assem-bly of the head of bacteriophage T4. Nature 1970:227:680-685
26. Scanu A. Forms of human serum high density lipoproteinprotein. J Upld Res 1966;7:295-306
27. Brown MS, Goldstein JL Lipoprotein metabolism in themacrophage: Implications for cholesterol deposition in ath-erosclerosis. Annu Rev Biochem 1983:52:223-261
28. Mahley RW. Development of accelerated atherosclerosis.Concepts derived from cell biology and animal model stud-ies. Arch Pathol Lab Med 1983:107:393-399
29. Kite T, Brown MS, WatanatM Y, Goldstein JL Deficiencyof low density lipoprotein receptors In liver and adrenal glandof the WHHL rabbit, an animal model of familial hypercho-lesterolemia. Proc NatJ Acad Sci USA 1981;78:2268-2272
30. Bachorlk PS, Franklin FA, Virgil DG, Kwtterovlch PO Jr.High-affinity uptake and degradation of apofipoprotein E freehigh-density lipoprotein and low-density lipoprotein in cul-tured porcine hepatocytes. Biochemistry 1982;21:5675-5684
31. Hul DY, Innerartty TL Mahley RW. Upoproteln binding tocanine hepatic membranes. Metabollcally distinct apo-E andapo B,E receptors. J Biol Chem 1981:256:5646-5655
32. Van Lenten BJ, Fogelman AM, Jackson RL Shapiro S,Haberiand ME, Edwards PA. Receptor-mediated uptake of
remnant lipoprotelns by cholesterol-loaded human monocyte-macrophages. J Biol Chem 1985;260:8783-8788
33. Basu SK, Brown MS, Ho YK, Goldstein JL Degradation oflow density lipoprotein dextran sulfate complexes associatedwith deposition of cholesteryl esters In mouse macrophages.J Biol Chem 1979:254:7141-7146
34. Randolph RK, St Clalr RW. Pigeon aortic smooth musclecells lack a functional low density lipoprotein receptor path-way. J LJpid Res 1984:25:888-902
35. Soutar AK, Knight BL Degradation of lipoprotelns byhuman monocyte-derived macrophages. Evidence for twodistinct processes for the degradation of abnormal very-low-density lipoprotein from subjects with type III hyperilp-Idaemla. Blochem J 1984;218:101-111
36. Glass CK, Plttman RC, Keller GA, Steinberg D. Tissuesites of degradation of apoprotein A-l in the rat J Biol Chem1983:258:7161-7167
37. Rlnnlnger F, Plttman RC. Regulation of the selective uptakeof high density lipoproteln-assodated cholesteryl esters. JUpid Res 1987:28:1313-1325
38. Azhar S, Cooper A, Tsal L Matfe W, Reaven E. Charac-terization of apo B,E receptor function in the luteinlzed ovary.J Upld Res 1988:29:869-882
39. Ptttman RC, Knecht TP, Rosenbaum MS, Taylor CA Jr. Anonendocytotic mechanism for the selective uptake of highdensity llpoproteln-assoclated cholesterol esters. J Biol Chem1987:262:2443-2450
40. Lottersdorf E, Israeli A, Stein O, Elsenberg S, Stein Y.The role of apolipoprotelns of HDL In the selective uptake ofcholesteryl linoleyl ether by cultured rat and bovine adrenalcells. Blochlm Biophys Acta 1986:878:320-329
41. Slotte JP, Chart A, Blerman EL Cholesterol accumulationin aortic smooth muscle cells exposed to low density lipopro-telns. Contribution of free cholesterol transfer. Arteriosclero-sis 1988:8:750-758
42. Bachorlk PS, Franklin FA, Virgil DG, Kwtterovlch PO Jr.Reversible high-afflnlty uptake of apo E-free high densitylipoprotelns In cultured pig hepatocytes. Arteriosclerosis1985:5:142-152
43. Tabas I, Tall AR. Mechanism of the association of HDL3with endothelial cells, smooth muscle cells, and fibroblasts.Evidence against the role of specific ligand and receptorproteins. J Biol Chem 1984:259:13897-13905
44. Rothblat GH, Arbogast LY, Ray EK. Stimulation of esteri-fied cholesterol accumulation in tissue culture cells exposedto high density lipoproteins enriched in free cholesterol. JUpid Res 1978:19:350-358
45. Arbogast LY, Rothblat GH, Leslie MH, Cooper RA. Cel-lular cholesterol ester accumulation Induced by freecholesterol-rich lipid dispersions. Proc Nati Acad Sci USA1976:73:3680-3684
46. Granot E, Tall A, Tabas I. Enhanced HDL cholesteryl esteruptake by model liver cells mediated by cholesteryl estertransfer protein [abstr]. Arteriosclerosis 1986;6:538a
47. Tollefson JH, Faust R, Albers JJ, Chart A. Secretion of aUpid transfer protein by human monocyte-derived macro-phages. J Biol Chem 1985;260:5887-5890
48. Schmltz G, Robenek H, Lohmann U, Assmann G. Inter-action of high density lipoproteins with choiesteryl ester-laden macrophages: biochemical and morphological charac-terization of cell surface receptor binding, endocytosis andresecretion of high density lipoprotelns by macrophages.EMBO J 1985:4:613-622
Index Terms: atherosclerosis • 0-VLDL receptors • rabbit 0-VLDL • lipoproteinsWhite Carneau pigeon • Show Racer pigeon
by guest on July 21, 2018http://atvb.ahajournals.org/
Dow
nloaded from

S J Adelman and R W St Clairdifferent potentials promoting cholesterol accumulation.
Beta-VLDL metabolism by pigeon macrophages. Evidence for two binding sites with
Print ISSN: 1079-5642. Online ISSN: 1524-4636 Copyright © 1989 American Heart Association, Inc. All rights reserved.
Avenue, Dallas, TX 75231is published by the American Heart Association, 7272 GreenvilleArteriosclerosis, Thrombosis, and Vascular Biology
doi: 10.1161/01.ATV.9.5.6731989;9:673-683Arterioscler Thromb Vasc Biol.
http://atvb.ahajournals.org/content/9/5/673World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://atvb.ahajournals.org//subscriptions/
at: is onlineArteriosclerosis, Thrombosis, and Vascular Biology Information about subscribing to Subscriptions:
http://www.lww.com/reprints
Information about reprints can be found online at: Reprints:
document.Permissions and Rights Question and AnswerFurther information about this process is available in theis being requested is located, click Request Permissions in the middle column of the Web page under Services.Clearance Center, not the Editorial Office. Once the online version of the published article for which permission
can be obtained via RightsLink, a service of the CopyrightArteriosclerosis, Thrombosis, and Vascular Biology Requests for permissions to reproduce figures, tables, or portions of articles originally published inPermissions:
by guest on July 21, 2018http://atvb.ahajournals.org/
Dow
nloaded from





























