Embed Size (px)
Citation preview

YeastYeast 2004; 21: 703–746.Published online in Wiley InterScience (www.interscience.wiley.com). DOI: 10.1002/yea.1113
Review
A history of research on yeasts 7: enzymic adaptationand regulationJames A. Barnett*School of Biological Sciences, University of East Anglia, Norwich NR4 7TJ, UK
*Correspondence to:James A. Barnett, School ofBiological Sciences, University ofEast Anglia, Norwich NR47TJ, UK.E-mail: [email protected]
Received: 18 December 2003Accepted: 20 January 2004
Keywords: history; yeast research; enzymic adaptation; enzyme induction andrepression; galactose pathway; GAL genes; Frederic Dienert; Solomon Spiegelman;Luis Leloir
The phenomenon of enzymatic adaptation may be simply stated in the following terms: apopulation of cells placed in contact with some substrate acquires, after a lapse of some time,the enzymes necessary to metabolize the added substrate (Spiegelman, 1946 [248] p. 256).
Contents
Introduction: the scope of this article 704Dienert’s work on adaptation of yeast to galactose 707Enzymic adaptation or selection of mutants? 709Galactose fermentation by yeasts 713Monod’s work on lactose utilization by Escherichia coli 714
Gratuitous induction 716‘Permeases’: transport of metabolites into the cells 718Carbon catabolite repression 720The operon 722
The galactose pathway in yeasts: work of Leloir and others 723Uridine diphosphate sugars 725
Genetic regulation of the galactose pathway 729Regulation of the GAL genes 730
Genetic regulation of the utilization of other substrates 732Maltose utilization: the MAL genes 732Sucrose utilization: invertase and the SUC genes 733
Conclusion 733References 733Appendix: sugar structures 743
Copyright 2004 John Wiley & Sons, Ltd.

704 J. A. Barnett
Introduction: the scope of this article
The present article concerns mechanisms by which yeasts regulate the amount of certain enzymes in theircells. Such regulations enable microbes to adapt to the different nutrients available to them in changingenvironments. The history of research on these regulatory processes in yeasts is the subject discussedbelow.
Most of the work which has explained the general phenomenon of enzymic adaptation1 has been doneby studying the microbial utilization of two sugars, lactose and D-galactose, both of which have long beenrelatively easy to obtain and purify.2 Accordingly, this article concentrates on the research done on theutilization of these two sugars. The structures of these and other sugars and sugar derivatives discussedherein are shown in an appendix at the end of this article.
Frederic Dienert3 (Figure 1), who worked on galactose utilization by yeasts in 1900, gave the first clearaccount of enzymic adaptation. His research eventually led to two major advances — in biochemistryand in molecular biology. (a) In the late 1940s and early 1950s, the great Argentinian biochemist,Luis Leloir4 (Figure 2) worked out the pathway of galactose catabolism in yeasts. He found that thispathway involved nucleoside diphosphate sugars and demonstrated the importance of these compoundsin biosynthetic pathways. (b) Although Leloir studied mainly Kluyveromyces marxianus, his work on thegalactose pathway made it possible to establish the details of how that pathway is controlled genetically inSaccharomyces cerevisiae. In the latter yeast, the GAL gene system, which controls galactose utilization,became the most extensively studied mechanism of biochemical–molecular genetic regulation in anyeukaryote. In turn, this work on galactose metabolism and its control was of medical significance,explaining the biochemistry of the severe human autosomal recessive disorder, galactosaemia.5
A great deal of the research on enzymic adaptations by yeasts was made possible by the researchescarried out at the Institut Pasteur in Paris in the 1940s and 1950s by Jacques Monod6 and his colleagues.By studying the β-galactosidase7 activity of Escherichia coli, they elucidated the general characteristics ofenzyme induction [54,183,189]. Accordingly, some of their work is described below. In his biographicalmemoir of Monod, Andre Lwoff8 explains:
The enzyme β-galactosidase . . . was only a tool for the understanding of the relation betweengenes and enzymes . . . When the work on the induced synthesis of enzymes was started in1941, nothing was known except the phenomenon; the concepts developed essentially from1948 on. In the first phase, biochemical, Melvin Cohn played a determining role. In the second,genetical and regulatory, Francois Jacob’s intervention had been essential . . . Between 1948
1 Even today, ‘adaptation’, the action of adapting, is still sometimes written ‘adaption’ as it was by the Anglo-Irish satirist JonathanSwift (1667–1745) in A Tale of a Tub (1704) [212] and in 1790 by the English chemist Charles Blagden (1748–1820) ([28] p. 344).2 When the water is evaporated from whey, which is a by-product of cheese-making, crystalline lactose (milk sugar) is deposited and thisis easily purified by recrystallization ([32] pp. 708–709; [113] p. 496). Pasteur discovered D-galactose (which he called ‘lactose’) in 1856by hydrolysing lactose [214]. Indeed, it is usually prepared by the acid hydrolysis of lactose forming glucose and galactose, the latterthen being separated from the glucose by direct fractional crystallization. The glucose may also be removed by a non-galactose-utilizingyeast and the galactose then crystallized ([32] pp. 602–603; [245] p. 89).3 Frederic Vincent Dienert (1874–1948), French biologist, is known for his public health work on purifying water. He became InspectorGeneral of the Paris Water Surveillance Service, honorary professor at the Institut Agronomique and, in 1946, President of the FrenchAcademy of Agriculture [4,33,156,157].4 Luis Federico Leloir (1906–1987), Argentinian biochemist, received the Nobel Prize for chemistry in 1970 ‘for his discovery of sugarnucleotides and their role in the biosynthesis of carbohydrates’. He became professor at Buenos Aires University. [209,305].5 Patients with galactosaemia are unable to metabolize galactose, because of a deficiency of one of the enzymes of the galactose pathway.D-Galactose 1-phosphate accumulates, damaging liver, central nervous system and other parts of the body [239,240].6 Jacques Monod (1910–1976) was French and one of the greatest of microbiologists.Working at the Institut Pasteur in Paris, he was afounder of molecular microbiology and largely responsible for the concept of allostery [163,164].7 β-Galactosidase is the enzyme which hydrolyses lactose to D-glucose and D-galactose.8 Andre Michel Lwoff (1902–1994), French microbiologist, worked at Institut Pasteur, Paris, from 1921, was professor at the Sorbonnefrom 1959 to 1968 and shared the Nobel Prize in Physiology or Medicine with Monod and Jacob. He worked on the genetics ofbacteriophages and, like Monod, was one of the originators of molecular biology [123].
Copyright 2004 John Wiley & Sons, Ltd. Yeast 2004; 21: 703–746.

History of research on yeasts 7 705
Figure 1. Frederic Dienert. Photograph from La Technique Sanitaire et Municipale, vol 3 (1948). Reproduced by kindpermission of the Secretaire perpetuel, Academie d’Agriculture de France
Figure 2. Luis Leloir (1970). The Nobel Foundation
Copyright 2004 John Wiley & Sons, Ltd. Yeast 2004; 21: 703–746.

706 J. A. Barnett
and 1963, the main problems posed by the induced synthesis of enzymes (that is regulation)were solved, and molecular biology was created ex nihilo ([163] p. 399).
Melvin Cohn9 writes that it was he who decided to work with β-galactosidase because ‘it was the onlyenzyme that could withstand fractionation at the summer temperature of Paris. Monod was on vacationand the Pasteur Institute had no cold rooms, a primitive centrifuge, but it did have amonium sulfate’ (M.Cohn, personal communication).
The workers at the Institut Pasteur made very considerable advances in understanding the physiologicaland molecular control of (a) enzyme induction and repression, and (b) the mechanisms by whichsubstrates enter microbial cells. In their studies of the regulation of metabolism, Monod and FrancoisJacob10 (Figure 3) established the role of allosteric enzymes [190]. The activity of such enzymes dependson their conformation (or shape) which is changed by the binding (attachment or detachment) of anothermolecule, an effector.
Monod and Jacob coined the expression ‘allosteric inhibition’; that is, to use Monod’s words, whenan enzyme:
. . . inhibitor is not a steric analogue of the substrate [of that enzyme]. We propose thereforeto designate this mechanism as ‘allosteric inhibition’ ([190] p. 391).
and he emphasized:
Figure 3. Jacques Monod and Francois Jacob (1966). Photograph from Paris-Match
9 Melvin Cohn (b. 1922), American microbial biochemist, worked at the Institut Pasteur, Paris from 1949 to 1955 and is now at the SalkInstitute for Biological Studies, La Jolla, California. He was professor at Washington University (1955–1958) and at Stanford University(1959–1961). His more recent work has been on immunology (M. Cohn, personal communication).10 Francois Jacob (b. 1920), French microbial geneticist and molecular biologist, shared the Nobel prize for physiology or medicine in1965 with Lwoff and Monod. When the Germans occupied France in 1940, Jacob came to London and joined the Free French forces,serving in North Africa. In 1944, he was badly wounded in Normandy and received the Croix de la Liberation. In 1964, Jacob becameprofessor at the College de France ([Nobel Lectures Physiology or Medicine 1963–1970] p. 172).
Copyright 2004 John Wiley & Sons, Ltd. Yeast 2004; 21: 703–746.

History of research on yeasts 7 707
. . . the regulatory role of allosteric proteins is absolutely fundamental; it explains everything:hormonal action, repressor function, non-Michaelian enzyme kinetics . . . (Monod, 1961;quoted in [276] p. 167).
The binding of an effector molecule at a second (non-catalytic) site alters the enzyme’s activity andso provides a means for its regulation [186].
Furthermore, in 1960, the Institut Pasteur researchers also published [127] the concept of the operon,a unit of coordinated gene activity, which made it possible to understand the underlying regulation ofinducible and repressible enzymes. They commented that up to that time:
. . . the word ‘gene’ . . . [was used to designate] a DNA molecule whose specific self-replicatingstructure can, through mechanisms unknown, become translated into the specific structure ofa polypeptide chain ([125] p. 318).
Monod and his remarkable colleagues were responsible for some outstanding achievements. Theirexcitement and enthusiasm is abundantly clear in their publications, occasionally obscuring someof the experimental detail! The work was, however, described in many extensive and clearlywritten reviews, many by Monod himself, making the subject both well-known and well understood[35,50,51,54,56,125,126,180,182,189,220,249,252,260,277].
Dienert’s work on adaptation of yeast to galactose
In the middle of the nineteenth century, Louis Pasteur11 was the first to report alterations in the behaviourof yeast in response to an environmental change, when he found that switching between aerobic andanaerobic conditions affected the rate of sugar fermentation [215,216]. Although the great chemist, EmilFischer, had studied the utilization of a number of sugars by various yeasts in the 1890s (see [23]), it wasnot until 1900 that Dienert gave the first lucid account of the special adaptations which yeasts undergo,enabling them to use certain sugars. In a 50-page paper on galactose fermentation, which constituted histhesis for a doctorate, Dienert used the word ‘l’accoutumance’ (habituation) for these adaptations. Withastonishing foresight, Dienert compared the adaptation he described to mammalian antibody production,12
as did Macfarlane Burnet13 over 50 years later [34] and Monod considered Dienert’s paper to be ‘one ofthe classics’ on enzymic adaptation ([180] p. 231).
Many microbial biochemists adopted Henning Karstrom’s14 term ‘adaptive’, which he proposed in 1938for microbial enzymes produced in specific response to the presence of an appropriate substrate [132].The term referred to the general and widespread phenomenon, which Dienert had studied: that an adaptiveenzyme is one synthesized by cells only when they are in contact with a substrate for the metabolizingof which the enzyme is necessary. By contrast, those enzymes which are present, irrespective of theavailability of any substrate, were called ‘constitutive’. Later, several kinds of physiological regulatorymechanism (Table 1) have been found to be responsible for enzymic ‘adaptivity’. At the molecular level,
11 Some accounts of the following are given in earlier articles: E. Fischer [23], H. M. Kalckar [21], C. C. Lindegren [23], P. M. Nurse[25], L. Pasteur [20], M. Stephenson [23], H. K. A. S. von Euler-Chelpin [21], Ø. Winge [23].12 C’est ce qui arrive egalement chez les animaux qui perdent de leur immunite contre une toxine, des que la toxine cesse d’agir ([69]p. 152).13 Frank Macfarlane Burnet (1899–1985), Australian virologist, immunologist and Nobel Prize winner, worked on the characteristicsand replication of bacteriophages and animal viruses and generated the clonal selection theory of antibody production. The third editionof his Natural History of Infectious Disease (1962; the last edition of which he was sole author) is still marvellous to read [87,208].14 Henning Karstrom (1899–1989) was, with the Nobel prize winner Artturi Virtanen, a pioneer of Finnish biochemistry. He worked onfood chemistry and nutrition but, in the 1940s, gave up science for religion (information kindly given by Anna-Maija Pietila, HelsinkiUniversity Library).
Copyright 2004 John Wiley & Sons, Ltd. Yeast 2004; 21: 703–746.

708 J. A. Barnett
Table 1. Mechanisms underlying enzymic adaptations involving changes in the amount of enzyme in thecell (after Entian and Barnett [81])
Kind ofregulation Physiological observation Enzymic or molecular mechanism
Induction Increase in enzymic activity in response topresence of inducer (i.e. substrate orstructurally similar compound)
Inducer evokes activation of transcription
Derepression Increased specific activity after removingrepressing substrate
De-inhibition of transcription
Inactivation Irreversible loss of enzymic activity Specific proteolysis of the enzyme
control can be exerted at the level of (a) transcription, affecting mRNA synthesis, or (b) translation,affecting protein synthesis from the mRNA transcript.
Reports of apparent enzymic adaptations had appeared even before Dienert’s work was published. In1882, Julius Wortmann15 found that ‘Bacterium termio’ did not produce amylases unless starch was inthe growth medium [300] and in 1898, Julius Katz of the Botanical Institute at Leipzig noted that theamylase (diastase) of a strain of a Penicillium species increased when growth was in the presence ofstarch [133]. There was also a report, based on inadequate experimental evidence, of yeasts adapting tosucrose [75]. It was Dienert, however, who in 1899 and 1900, when working under Emile Duclaux,16
published the first clear evidence of enzyme induction [67,68,69]. Using washed cells of brewer’s yeastsand also of yeasts which fermented lactose, Dienert reported adaptation to galactose in the absence ofcell division. Hence he was not selecting mutants: the yeast cells which were present were adapting tothe sugar. They fermented D-galactose after a few hours of adaptation, whereas glucose was fermentedwithout delay. Furthermore, he found that some yeasts which he had adapted to galactose, had alsoadapted to lactose. The kind of flask Dienert used for his adaptation experiments is shown in Figure 4.
The following were Dienert’s salient findings. (a) The rate of galactose fermentation depends on whichsugars are present in the growth medium; but the rate of glucose fermentation varies very little, irrespectiveof what the yeast is grown on. (b) When galactose-grown cells are re-grown in medium containing glucosethey lose their ‘galactozymase’ (that is, the enzymic mechanism by which galactose is catabolized).(c) When yeasts are grown in medium containing galactose, lactose or melibiose, galactozymase activityis maximal; for those grown with sucrose or maltose, the galactozymase activity is slight; yeasts grown
a
Figure 4. Flask used for adaptation experiments by Dienert [69]. Yeast accumulates in the little bulb (a) in the base ofthe vessel
15 Julius Wortmann (1856–1924), German botanist, worked inter alia on Saccharomyces cerevisiae in relation to vinification at theGeisenheim research station for viticultural problems [194].16 Emile Duclaux (1840–1904), French chemist and bacteriologist, helped Pasteur in his work on diseases of silk-worms and succeededhim as director of the Institut Pasteur, Paris [169].
Copyright 2004 John Wiley & Sons, Ltd. Yeast 2004; 21: 703–746.

History of research on yeasts 7 709
on glucose or fructose have no galactozymase. (d) Yeasts grown on either melibiose or galactose formα-galactosidase (melibiase). He wrote:
Galactose fermentation is possible only when the yeast is adapted to this sugar. The time ofadaptation varies with different yeasts . . . Adapted yeasts ferment glucose about 1.6 timesfaster than galactose . . . An adapted yeast loses its adaptation little by little if it is given asugar other than galactose, lactose or melibiose. If growth occurs, the adaptation is destroyedafter a few hours . . . Adaptation does not affect the morphological characteristics of the yeasts. . . Certain substances prevent adaptation without preventing the fermentation of glucose (e.g.borate and toluene).17
In 1936 Marjory Stephenson and John Yudkin18 were not as generous as Monod, when they commented:
These experiments of Dienert, though highly suggestive, are difficult to interpret owingprincipally to their non-quantitative nature, rates of fermentation, quantity of yeast and changein cell numbers not being recorded ([266] p. 506).
Table 2 gives the results of some of Dienert’s experiments and in 1908 Arthur Slator19 confirmedthose findings (Table 3). At about the same time, Edward Armstrong20 named several yeasts which didnot adapt to galactose [14] (Table 4), although all could ferment glucose, fructose and mannose withoutadaptation. In 1910 Arthur Harden confirmed that ‘some yeasts can be trained to ferment galactose bycultivation in a medium containing that sugar’ and that ‘such a trained yeast yields a juice capable offermenting galactose’ ([111] p. 649).
Dienert’s publications also stimulated work on enzymic adaptations in multicellular organisms. Forexample, in 1906 Ernst Weinland21 reported that repeated intravenous injections of sucrose inducedinvertase activity in a dog’s plasma [288].
Enzymic adaptation or selection of mutants?
Yudkin pointed out:
As would be expected, when organisms possessing a newly acquired enzyme are grown inthe absence of the substrate, enzymes arising by adaptation are readily lost whilst enzymesarising by mutation and selection tend to be permanent (Yudkin, 1938 [301] p. 104).
Indeed, as there were often difficulties in distinguishing between enzymic adaptation and the selectionof genetic variants, many experiments were done to decide whether microbial adaptive adjustments were
17 La fermentation du galactose n’est possible que lorsque la levure s’est acclimatee a ce sucre. La duree de l’acclimatation varie avec leslevures . . . Chez les levures acclimatees, le glucose fermente environ 1,6 fois plus vite que le galactose . . . Une levure acclimatee perdpeu a peu son acclimatation si on lui offre un autre sucre que du galactose, du lactose ou du melibiose. Si on favorise la multiplication,la perte de l’acclimatation se produit au bout de quelques heures . . . L’effet est nul sur les proprietes morphologiques des levures . . .
Certaines substances empechent l’acclimatation sans empecher la fermentation du glucose (BoO3−, toluene) ([69] p. 187).18 John Yudkin (1910–1995) English biochemist and nutritional physiologist, worked with Marjory Stephenson on enzymic adaptation inyeast and bacteria for about 5 years and simultaneously studied medicine. From 1946, he was professor of physiology at King’s Collegeof Household and Social Science of London University (later Queen Elizabeth College) (M. D. Yudkin, personal communication).19 Arthur Slator (1879–1953), English brewing specialist, published work on rates of activities of various microbes and became chiefchemist to Bass brewers at Burton-on-Trent [5,128].20 Edward Frankland Armstrong (1878–1945), English carbohydrate chemist with wide scientific interests, obtained his doctorate at theUniversity of Berlin and published six papers on sugar chemistry with Emil Fischer. Armstrong had a career in industries concernedwith biscuits, soup, dyestuffs, coal and gas [94,232].21 Ernst Weinland (1869–1932), German medical physiologist, worked on anatomy, physiology and biochemistry at Munich, Tubingenand Leipzig. He held a chair in physiology at Erlangen from 1913 to 1932 [103].
Copyright 2004 John Wiley & Sons, Ltd. Yeast 2004; 21: 703–746.

710 J. A. Barnett
Table 2. Results of some of Dienert’s experiments [69]. In each flask ofthe type shown in Figure 4, the yeast was grown in yeast extract to whicha sugar was added as indicated in the table. After growth, the medium wasdrawn off and the yeast, which accumulated in the bulb at the base of theflask, was washed in sterile water. The washing water was then replaced bya galactose solution. Incubation was at 25 ◦C; the time was noted when 1 gasbubble per second was produced and this time was treated as the beginningof fermentation
YeastSugar added togrowth medium
Delay before beginningof fermentation
A yeast growing on lactose Sucrose 2 hGalactose 1 hLactose 0.5 hMaltose 2 h
‘Frohberg’ yeast (a Glucose 2 daysbottom-fermenting beer Sucrose 2 daysyeast ([105] pp. 233–234) Fructose 2 days
Melibiose 1 hGalactose 1 h
Table 3. Rates of fermentation of galactose or glucose by two yeastspecies when grown in either (a) brewer’s wort (an aqueous extract ofmalt) or (b) a solution of hydrolysed lactose + a trace of wort. Results ofSlator in 1908 ([242] p. 224)
Relative rates
Yeast Growth medium Glucose Galactose
Saccharomyces cerevisiae Wort 100 <1Hydrolysed lactose 100 21–77
Saccharomyces pastorianus Wort 100 <1Hydrolysed lactose 100 25–155
caused by selecting a minority of cells which already had the ability to use the substrate in question, orby the action of that substrate on most of the cells present.
Such questions were asked when ‘training’ Salmonella typhi to use lactose (1907) [275] or galactitol22
(1911) [218]. Working with yeasts, several authors in the 1920s concluded that new cells had to beformed for these adaptations to occur. Various methods were used to stop cell division, such as adding0.5% phenol [279,284] or measuring enzymic activity at 38 ◦C [244]. However, such experiments wereusually inconclusive, as the techniques may well have also stopped synthesis of the required enzyme.
In 1933, Stephenson and her colleagues showed that adaptive enzyme synthesis occurs without cellreproduction in both yeasts and bacteria, for example in the presence of formate, E. coli synthesizes‘formic hydrogen lyase’23 without cell multiplication [265]. Three years later, they also detected adaptiveformation of galactozymase in non-proliferating cells of S. cerevisiae [266]. When washed cells, deficientin galactozymase, were suspended in galactose solution, galactozymase activity developed within anhour, that is before appreciable cell division could occur. Cell counts confirmed that enzyme productionoccurred without the numbers of cells increasing. The criterion used for assessing enzymic activity was
22 Galactitol (also called dulcitol or dulcite) is the reduction product of galactose (Appendix).23 ‘Formic hydrogen lyase’, catalysing formic acid → CO2 + H2, is made up of two enzymes [217,223,286]: (a) formate dehydrogenase[EC 1.2.1.2], which catalyses formate + NAD+ → CO2 + NADH; (b) hydrogen dehydrogenase [EC 1.12.1.2], which catalyses H2 +NAD+ → H+ + NADH [264].
Copyright 2004 John Wiley & Sons, Ltd. Yeast 2004; 21: 703–746.
History of research on yeasts 7 711
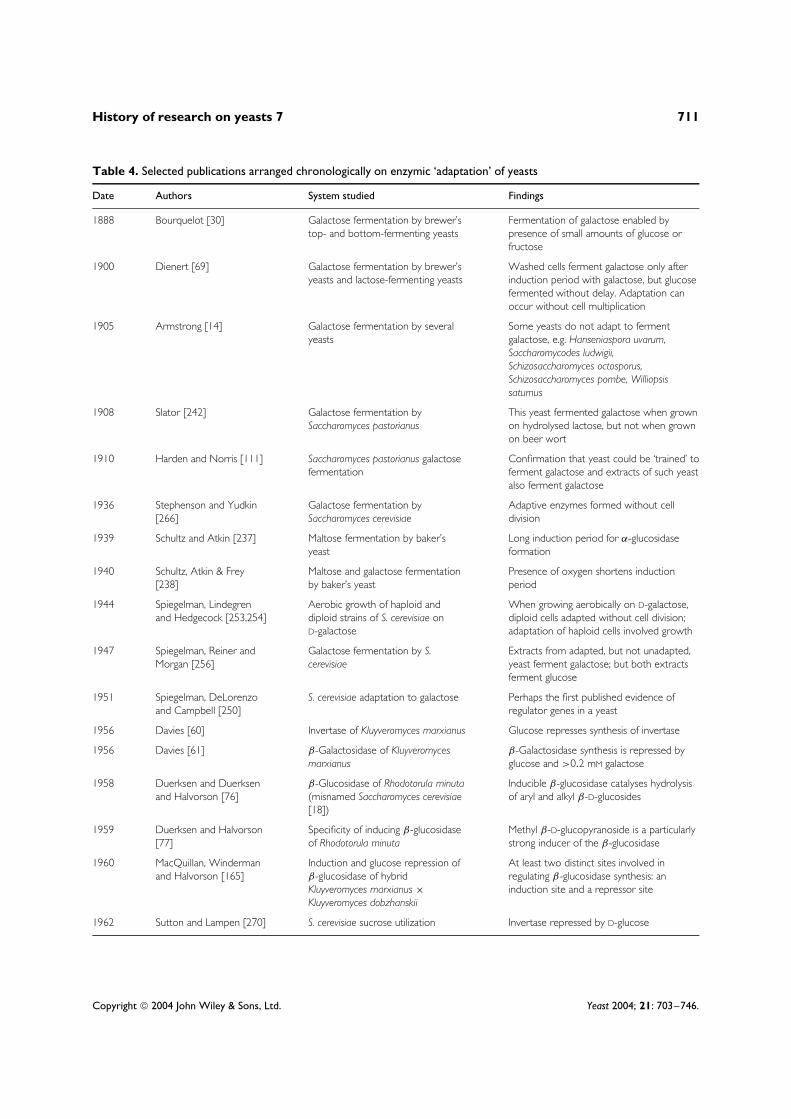
Table 4. Selected publications arranged chronologically on enzymic ‘adaptation’ of yeasts
Date Authors System studied Findings
1888 Bourquelot [30] Galactose fermentation by brewer’stop- and bottom-fermenting yeasts
Fermentation of galactose enabled bypresence of small amounts of glucose orfructose
1900 Dienert [69] Galactose fermentation by brewer’syeasts and lactose-fermenting yeasts
Washed cells ferment galactose only afterinduction period with galactose, but glucosefermented without delay. Adaptation canoccur without cell multiplication
1905 Armstrong [14] Galactose fermentation by severalyeasts
Some yeasts do not adapt to fermentgalactose, e.g. Hanseniaspora uvarum,Saccharomycodes ludwigii,Schizosaccharomyces octosporus,Schizosaccharomyces pombe, Williopsissaturnus
1908 Slator [242] Galactose fermentation bySaccharomyces pastorianus
This yeast fermented galactose when grownon hydrolysed lactose, but not when grownon beer wort
1910 Harden and Norris [111] Saccharomyces pastorianus galactosefermentation
Confirmation that yeast could be ‘trained’ toferment galactose and extracts of such yeastalso ferment galactose
1936 Stephenson and Yudkin[266]
Galactose fermentation bySaccharomyces cerevisiae
Adaptive enzymes formed without celldivision
1939 Schultz and Atkin [237] Maltose fermentation by baker’syeast
Long induction period for α-glucosidaseformation
1940 Schultz, Atkin & Frey[238]
Maltose and galactose fermentationby baker’s yeast
Presence of oxygen shortens inductionperiod
1944 Spiegelman, Lindegrenand Hedgecock [253,254]
Aerobic growth of haploid anddiploid strains of S. cerevisiae onD-galactose
When growing aerobically on D-galactose,diploid cells adapted without cell division;adaptation of haploid cells involved growth
1947 Spiegelman, Reiner andMorgan [256]
Galactose fermentation by S.cerevisiae
Extracts from adapted, but not unadapted,yeast ferment galactose; but both extractsferment glucose
1951 Spiegelman, DeLorenzoand Campbell [250]
S. cerevisiae adaptation to galactose Perhaps the first published evidence ofregulator genes in a yeast
1956 Davies [60] Invertase of Kluyveromyces marxianus Glucose represses synthesis of invertase
1956 Davies [61] β-Galactosidase of Kluyveromycesmarxianus
β-Galactosidase synthesis is repressed byglucose and >0.2 mM galactose
1958 Duerksen and Duerksenand Halvorson [76]
β-Glucosidase of Rhodotorula minuta(misnamed Saccharomyces cerevisiae[18])
Inducible β-glucosidase catalyses hydrolysisof aryl and alkyl β-D-glucosides
1959 Duerksen and Halvorson[77]
Specificity of inducing β-glucosidaseof Rhodotorula minuta
Methyl β-D-glucopyranoside is a particularlystrong inducer of the β-glucosidase
1960 MacQuillan, Windermanand Halvorson [165]
Induction and glucose repression ofβ-glucosidase of hybridKluyveromyces marxianus ×Kluyveromyces dobzhanskii
At least two distinct sites involved inregulating β-glucosidase synthesis: aninduction site and a repressor site
1962 Sutton and Lampen [270] S. cerevisiae sucrose utilization Invertase repressed by D-glucose
Copyright 2004 John Wiley & Sons, Ltd. Yeast 2004; 21: 703–746.
712 J. A. Barnett
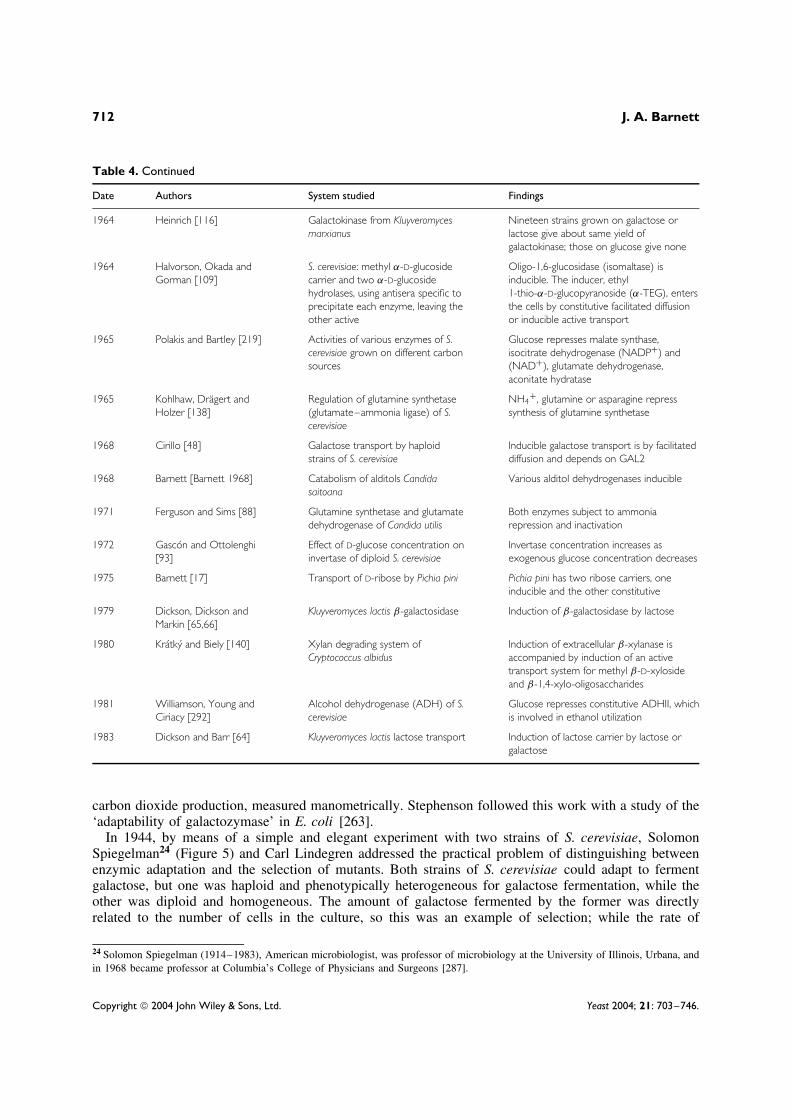
Table 4. Continued
Date Authors System studied Findings
1964 Heinrich [116] Galactokinase from Kluyveromycesmarxianus
Nineteen strains grown on galactose orlactose give about same yield ofgalactokinase; those on glucose give none
1964 Halvorson, Okada andGorman [109]
S. cerevisiae: methyl α-D-glucosidecarrier and two α-D-glucosidehydrolases, using antisera specific toprecipitate each enzyme, leaving theother active
Oligo-1,6-glucosidase (isomaltase) isinducible. The inducer, ethyl1-thio-α-D-glucopyranoside (α-TEG), entersthe cells by constitutive facilitated diffusionor inducible active transport
1965 Polakis and Bartley [219] Activities of various enzymes of S.cerevisiae grown on different carbonsources
Glucose represses malate synthase,isocitrate dehydrogenase (NADP+) and(NAD+), glutamate dehydrogenase,aconitate hydratase
1965 Kohlhaw, Dragert andHolzer [138]
Regulation of glutamine synthetase(glutamate–ammonia ligase) of S.cerevisiae
NH4+ , glutamine or asparagine repress
synthesis of glutamine synthetase
1968 Cirillo [48] Galactose transport by haploidstrains of S. cerevisiae
Inducible galactose transport is by facilitateddiffusion and depends on GAL2
1968 Barnett [Barnett 1968] Catabolism of alditols Candidasaitoana
Various alditol dehydrogenases inducible
1971 Ferguson and Sims [88] Glutamine synthetase and glutamatedehydrogenase of Candida utilis
Both enzymes subject to ammoniarepression and inactivation
1972 Gascon and Ottolenghi[93]
Effect of D-glucose concentration oninvertase of diploid S. cerevisiae
Invertase concentration increases asexogenous glucose concentration decreases
1975 Barnett [17] Transport of D-ribose by Pichia pini Pichia pini has two ribose carriers, oneinducible and the other constitutive
1979 Dickson, Dickson andMarkin [65,66]
Kluyveromyces lactis β-galactosidase Induction of β-galactosidase by lactose
1980 Kratky and Biely [140] Xylan degrading system ofCryptococcus albidus
Induction of extracellular β-xylanase isaccompanied by induction of an activetransport system for methyl β-D-xylosideand β-1,4-xylo-oligosaccharides
1981 Williamson, Young andCiriacy [292]
Alcohol dehydrogenase (ADH) of S.cerevisiae
Glucose represses constitutive ADHII, whichis involved in ethanol utilization
1983 Dickson and Barr [64] Kluyveromyces lactis lactose transport Induction of lactose carrier by lactose orgalactose
carbon dioxide production, measured manometrically. Stephenson followed this work with a study of the‘adaptability of galactozymase’ in E. coli [263].
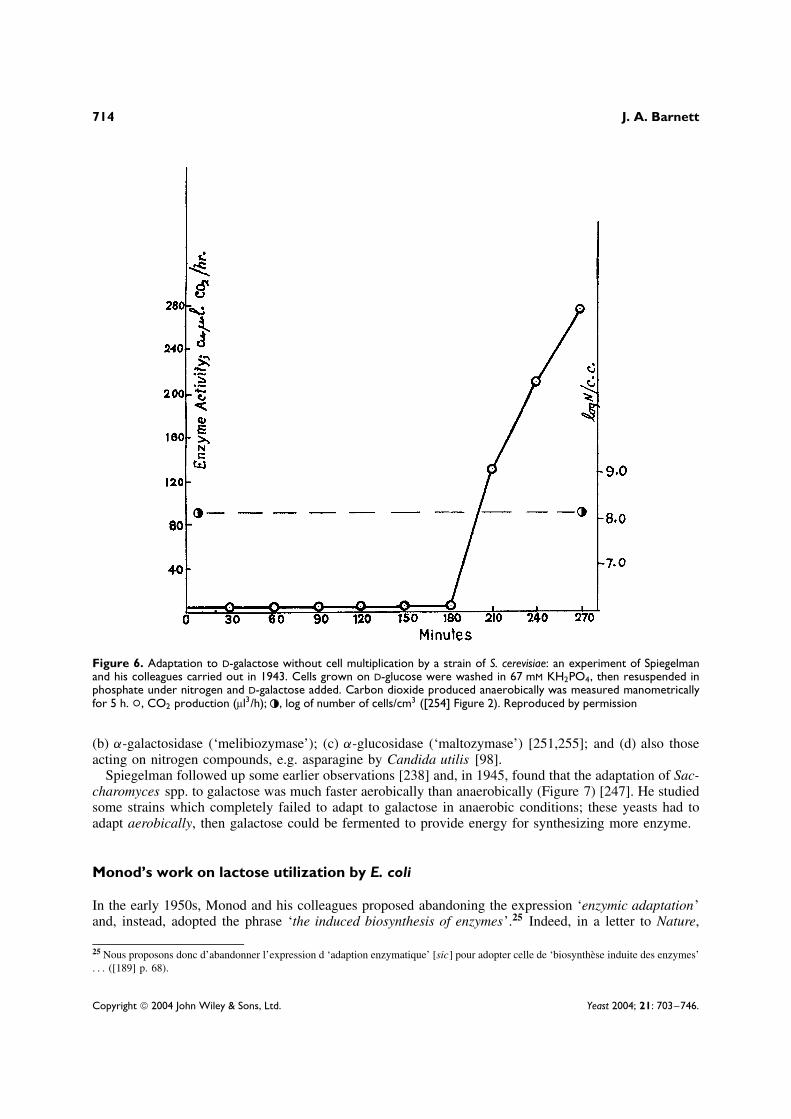
In 1944, by means of a simple and elegant experiment with two strains of S. cerevisiae, SolomonSpiegelman24 (Figure 5) and Carl Lindegren addressed the practical problem of distinguishing betweenenzymic adaptation and the selection of mutants. Both strains of S. cerevisiae could adapt to fermentgalactose, but one was haploid and phenotypically heterogeneous for galactose fermentation, while theother was diploid and homogeneous. The amount of galactose fermented by the former was directlyrelated to the number of cells in the culture, so this was an example of selection; while the rate of
24 Solomon Spiegelman (1914–1983), American microbiologist, was professor of microbiology at the University of Illinois, Urbana, andin 1968 became professor at Columbia’s College of Physicians and Surgeons [287].
Copyright 2004 John Wiley & Sons, Ltd. Yeast 2004; 21: 703–746.

History of research on yeasts 7 713
Figure 5. Soloman Spiegelman. Photo courtesy of the University of Illinois at Urbana-Champaign Archives; record seriesnumber 26/4/1, Alumni and Faculty File
galactose fermentation by the latter strain was almost independent of the number of cells and hence, asthere was no growth, this was a case of physiological adaptation (Figure 6) [254].
Galactose fermentation by yeasts
By the late 1940s, the fermentation of galactose by yeasts had become the most thoroughly studiedsystem of enzymic adaptation and the then current findings may be summarized as follows:
1. Yeasts which could ferment galactose did so only after being incubated with galactose [69,111,242,246,266].
2. This adaptation did not depend on cell multiplication and, hence, selection was not involved[69,246,266].
3. Cell-free extracts from adapted yeast could ferment galactose; extracts from unadapted cells did not doso, although they fermented glucose [111,256]. These observations showed that the adaptation couldnot be explained simply in terms of changes in the permeability of the cells.
In the course of his work on adaptation to galactose, Spiegelman reported a singular observation onSchizosaccharomyces pombe, which is usually held to be galactose-negative [14,141,143,161,262]. Hefound that when cultures composed of >20% asci (as meiotic products, the ascospores would be haploid)were inoculated into a medium of 8% galactose and 2% glucose, some strains could be isolated whichutilized galactose. Consistent with Spiegelman’s finding, a more recent work on yeast systematics recordsthat some strains can use galactose after a delay of more than 7 days ([24] p. 678). However, thenature of this ‘adaptation’, if indeed it occurs, does not seem to have been investigated and Paul Nurse(personal communication, 2003) comments that if indeed Spiegelman selected mutants, such mutantswould probably be rare.
By 1950, microbiologists had examined adaptations of many other enzymes ([249] pp. 268–270), suchas those by yeasts for: (a) invertase, studied by Hans von Euler-Chelpin and his colleagues [278,280–284];
Copyright 2004 John Wiley & Sons, Ltd. Yeast 2004; 21: 703–746.
714 J. A. Barnett
Figure 6. Adaptation to D-galactose without cell multiplication by a strain of S. cerevisiae: an experiment of Spiegelmanand his colleagues carried out in 1943. Cells grown on D-glucose were washed in 67 mM KH2PO4, then resuspended inphosphate under nitrogen and D-galactose added. Carbon dioxide produced anaerobically was measured manometricallyfor 5 h. °, CO2 production (µl3/h); , log of number of cells/cm3 ([254] Figure 2). Reproduced by permission
(b) α-galactosidase (‘melibiozymase’); (c) α-glucosidase (‘maltozymase’) [251,255]; and (d) also thoseacting on nitrogen compounds, e.g. asparagine by Candida utilis [98].
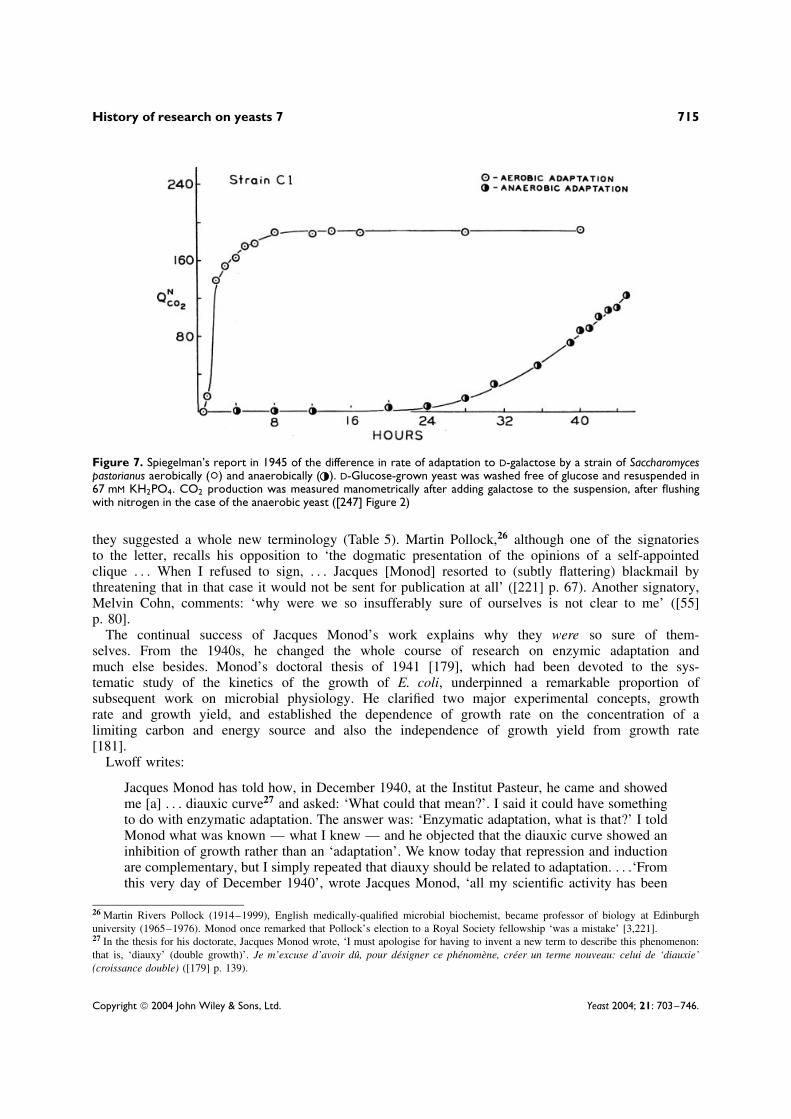
Spiegelman followed up some earlier observations [238] and, in 1945, found that the adaptation of Sac-charomyces spp. to galactose was much faster aerobically than anaerobically (Figure 7) [247]. He studiedsome strains which completely failed to adapt to galactose in anaerobic conditions; these yeasts had toadapt aerobically, then galactose could be fermented to provide energy for synthesizing more enzyme.
Monod’s work on lactose utilization by E. coli
In the early 1950s, Monod and his colleagues proposed abandoning the expression ‘enzymic adaptation’and, instead, adopted the phrase ‘the induced biosynthesis of enzymes’.25 Indeed, in a letter to Nature,
25 Nous proposons donc d’abandonner l’expression d ‘adaption enzymatique’ [sic] pour adopter celle de ‘biosynthese induite des enzymes’. . . ([189] p. 68).
Copyright 2004 John Wiley & Sons, Ltd. Yeast 2004; 21: 703–746.
History of research on yeasts 7 715
Figure 7. Spiegelman’s report in 1945 of the difference in rate of adaptation to D-galactose by a strain of Saccharomycespastorianus aerobically (°) and anaerobically ( ). D-Glucose-grown yeast was washed free of glucose and resuspended in67 mM KH2PO4. CO2 production was measured manometrically after adding galactose to the suspension, after flushingwith nitrogen in the case of the anaerobic yeast ([247] Figure 2)
they suggested a whole new terminology (Table 5). Martin Pollock,26 although one of the signatoriesto the letter, recalls his opposition to ‘the dogmatic presentation of the opinions of a self-appointedclique . . . When I refused to sign, . . . Jacques [Monod] resorted to (subtly flattering) blackmail bythreatening that in that case it would not be sent for publication at all’ ([221] p. 67). Another signatory,Melvin Cohn, comments: ‘why were we so insufferably sure of ourselves is not clear to me’ ([55]p. 80].
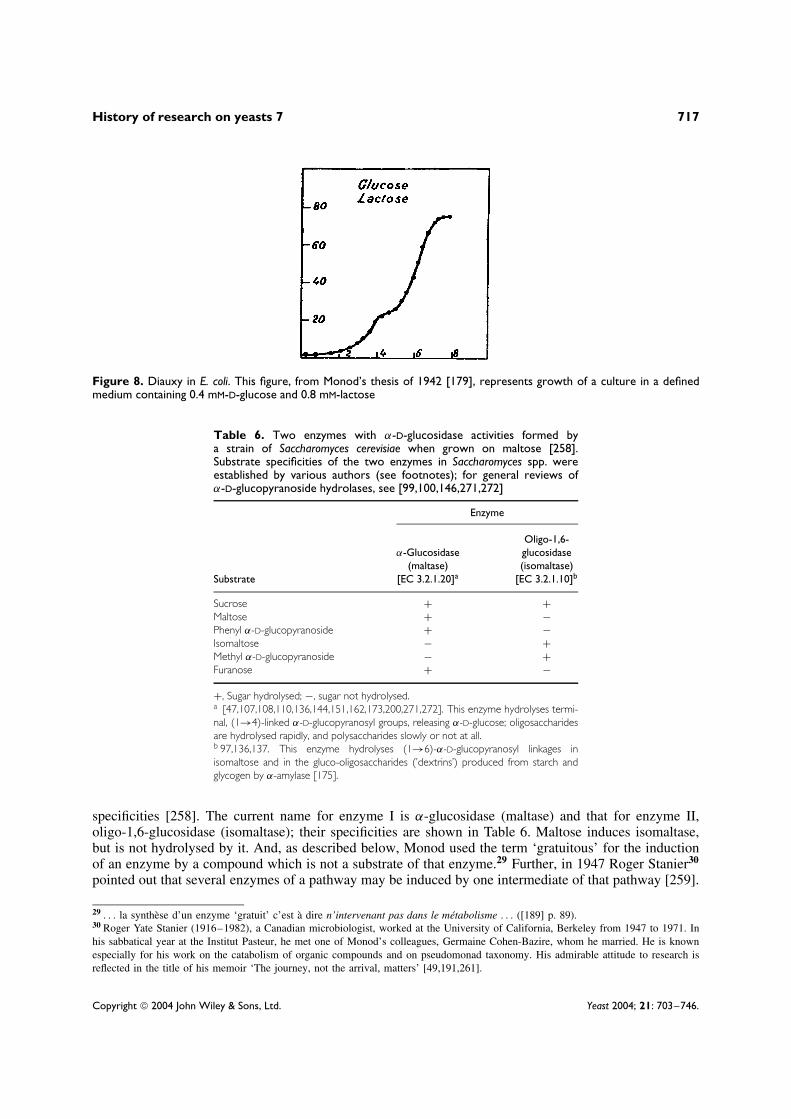
The continual success of Jacques Monod’s work explains why they were so sure of them-selves. From the 1940s, he changed the whole course of research on enzymic adaptation andmuch else besides. Monod’s doctoral thesis of 1941 [179], which had been devoted to the sys-tematic study of the kinetics of the growth of E. coli, underpinned a remarkable proportion ofsubsequent work on microbial physiology. He clarified two major experimental concepts, growthrate and growth yield, and established the dependence of growth rate on the concentration of alimiting carbon and energy source and also the independence of growth yield from growth rate[181].
Lwoff writes:
Jacques Monod has told how, in December 1940, at the Institut Pasteur, he came and showedme [a] . . . diauxic curve27 and asked: ‘What could that mean?’. I said it could have somethingto do with enzymatic adaptation. The answer was: ‘Enzymatic adaptation, what is that?’ I toldMonod what was known — what I knew — and he objected that the diauxic curve showed aninhibition of growth rather than an ‘adaptation’. We know today that repression and inductionare complementary, but I simply repeated that diauxy should be related to adaptation. . . .‘Fromthis very day of December 1940’, wrote Jacques Monod, ‘all my scientific activity has been
26 Martin Rivers Pollock (1914–1999), English medically-qualified microbial biochemist, became professor of biology at Edinburghuniversity (1965–1976). Monod once remarked that Pollock’s election to a Royal Society fellowship ‘was a mistake’ [3,221].27 In the thesis for his doctorate, Jacques Monod wrote, ‘I must apologise for having to invent a new term to describe this phenomenon:that is, ‘diauxy’ (double growth)’. Je m’excuse d’avoir du, pour designer ce phenomene, creer un terme nouveau: celui de ‘diauxie’(croissance double) ([179] p. 139).
Copyright 2004 John Wiley & Sons, Ltd. Yeast 2004; 21: 703–746.

716 J. A. Barnett
Table 5. Terminology of enzymic adaptation, proposed in 1953 by Cohn, Monod, Pollock,Spiegelman and Stanier [57]
Term Meaning
’Enzyme induction’ Increase in rate of synthesis of a specific apo-enzyme resultingfrom exposure to a chemical substance
’Enzyme inducer’ Any substance inducing enzyme synthesis
’Inducible enzyme’ An enzyme-forming system which can be activated by anexogenous inducer
’Induced enzyme’ Enzyme formed by induction
’Constitutive enzyme’ One formed in ‘considerable amounts’ in the absence of anexogenous inducer. The amount is often increased by specificinduction
’Constitutivity’ and ‘inducibility’ Properties of enzyme-forming system, not of enzymes per se
’Sequential induction’ A single substance may induce a sequence of enzymes
Note: Certain substrates of induced enzymes do not induce them; and some inducers are not substratesof the enzymes they induce
devoted to the study of enzymatic adaptation.’ Yet during the dark years, he had joined theunderground. He had even been arrested by the Gestapo, but cleverly managed to escape([163] p. 388).
And Monod tells us:
Lwoff’s intuition was correct. The phenomenon of ‘diauxy’ that I had discovered was indeedclosely related to enzyme adaptation, as my experiments, included in the second part of mydoctoral dissertation, soon convinced me. It was actually a case of the ‘glucose effect’ . . .today better known as ‘catabolite repression’ . . . ([184] p. 189).
Monod’s assessment is well-illustrated by the results of an experiment he published in 1946, when heand a colleague worked with a lactose-utilizing strain of E. coli, which showed a typical diauxic curvewhen grown in a medium containing a mixture of glucose and lactose (Figure 8). Here was an early, butclear, representation of the ‘glucose effect’ or ‘carbon catabolite repression’, which is discussed below.Suspensions of this strain grown (a) on lactose (‘adapted’) or (b) on glucose (‘non-adapted’) were testedfor the ability to respire lactose. Adapted cells respired fast; non-adapted cells respired no faster thanthe negative control cells without exogenous substrate, and the adaptive enzyme systems were shownto be determined genetically [185]. Glucose repression was clearly an economy measure: unnecessaryenzymes, such as β-galactosidase, are not synthesized when so readily catabolized a compound as glucoseis present.
Gratuitous induction
In 1947, concluding that microbial enzymes which attack exogenous substrates are usually ‘adaptive’,Monod defined ‘enzymatic adaptation’ as ‘apo-enzyme28 formation induced by a specific substrate’ ([180]p. 226). However, not long afterwards, certain enzymes were shown to be induced by compounds whichare not substrates of those enzymes. Spiegelman and his colleagues found that a strain of S. cerevisiae,grown on or otherwise adapted to maltose, formed two enzymes (I and II) which had different substrate
28 Apoenzyme is the protein component of an enzyme; an enzyme with a tightly bound cofactor (prosthetic group) is a holoenzyme[1,205].
Copyright 2004 John Wiley & Sons, Ltd. Yeast 2004; 21: 703–746.
History of research on yeasts 7 717
Figure 8. Diauxy in E. coli. This figure, from Monod’s thesis of 1942 [179], represents growth of a culture in a definedmedium containing 0.4 mM-D-glucose and 0.8 mM-lactose
Table 6. Two enzymes with α-D-glucosidase activities formed bya strain of Saccharomyces cerevisiae when grown on maltose [258].Substrate specificities of the two enzymes in Saccharomyces spp. wereestablished by various authors (see footnotes); for general reviews ofα-D-glucopyranoside hydrolases, see [99,100,146,271,272]
Enzyme
Substrate
α-Glucosidase(maltase)
[EC 3.2.1.20]a
Oligo-1,6-glucosidase(isomaltase)
[EC 3.2.1.10]b
Sucrose + +Maltose + −Phenyl α-D-glucopyranoside + −Isomaltose − +Methyl α-D-glucopyranoside − +Furanose + −
+, Sugar hydrolysed; −, sugar not hydrolysed.a [47,107,108,110,136,144,151,162,173,200,271,272]. This enzyme hydrolyses termi-nal, (1→4)-linked α-D-glucopyranosyl groups, releasing α-D-glucose; oligosaccharidesare hydrolysed rapidly, and polysaccharides slowly or not at all.b 97,136,137. This enzyme hydrolyses (1→6)-α-D-glucopyranosyl linkages inisomaltose and in the gluco-oligosaccharides (’dextrins’) produced from starch andglycogen by α-amylase [175].
specificities [258]. The current name for enzyme I is α-glucosidase (maltase) and that for enzyme II,oligo-1,6-glucosidase (isomaltase); their specificities are shown in Table 6. Maltose induces isomaltase,but is not hydrolysed by it. And, as described below, Monod used the term ‘gratuitous’ for the inductionof an enzyme by a compound which is not a substrate of that enzyme.29 Further, in 1947 Roger Stanier30
pointed out that several enzymes of a pathway may be induced by one intermediate of that pathway [259].
29 . . . la synthese d’un enzyme ‘gratuit’ c’est a dire n’intervenant pas dans le metabolisme . . . ([189] p. 89).30 Roger Yate Stanier (1916–1982), a Canadian microbiologist, worked at the University of California, Berkeley from 1947 to 1971. Inhis sabbatical year at the Institut Pasteur, he met one of Monod’s colleagues, Germaine Cohen-Bazire, whom he married. He is knownespecially for his work on the catabolism of organic compounds and on pseudomonad taxonomy. His admirable attitude to research isreflected in the title of his memoir ‘The journey, not the arrival, matters’ [49,191,261].
Copyright 2004 John Wiley & Sons, Ltd. Yeast 2004; 21: 703–746.

718 J. A. Barnett
The workers at the Institut Pasteur used a number of non-metabolizable synthetic glycosides in order tostudy induction. Substituting sulphur for oxygen in the galactoside linkage converts an O-glycoside intoan S -glycoside [Figure 9; and cf. methyl β-D-galactopyranoside with methyl 1-thio-β-D-galactopyranoside(TMG) in Appendix]. Although β-galactosidase does not hydrolyse thio-galactosides, it has about thesame affinity for them as for the corresponding O-galactosides.
These ‘gratuitous’ inducers, such as TMG, produced for Monod’s laboratory in the chemical institute ofBonn University [117], made it practicable to study enzyme induction separately from enzymic activity.Although it is not hydrolysed, TMG strongly induces the synthesis of β-galactosidase and at constant rate,as the synthesis is not limited by metabolic dependence on the inducer itself. Thus, many experimentson the induced synthesis of β-galactosidase were done with E. coli under conditions of gratuity: neitherinducer, nor the enzyme it induced, gave the cells any advantage. Before 1950, ‘adaptive’ enzymes hadbeen thought to be produced only in response to the presence of their substrates; but a systematic studyof the utilization of β-galactosides by E. coli enabled Monod and his colleagues to make the followinggeneralizations [188,189]:
1. Some enzymic substrates are inducers; others do not induce.2. Some β-galactosides for which β-galactosidase has no affinity are powerful inducers of that enzyme.3. Some inducers are not substrates of the induced enzyme, but inhibit its activity competitively.
By means of a series of elegant experiments on the kinetics of incorporation of sulphur into the β-galactosidase molecule during its induced synthesis, Monod’s group showed that induction of the enzymeinvolves its synthesis de novo, rather than the activation of a single enzyme precursor [119].
‘Permeases’: transport of metabolites into the cells
In the 1950s, Monod and his colleagues isolated many mutants from E. coli. Some of these mutantswere ‘cryptics’,31 that is they could not catabolize exogenously-supplied β-galactosides, such as lactose,despite possessing β-galactosidase. Such crypticity had already been reported for a number of yeasts.For example: (a) intact brewer’s yeast fermented maltose at 4 ◦C, but not methyl α-D-glucopyranoside,although extracts fermented the latter at that temperature [152]; (ii) β-glucosidase, which hydrolysescellobiose to two glucose molecules, was present in a strain of Kluyveromyces marxianus,32 but intactyeast did not ferment cellobiose [204]; (iii) pressed yeast (Preßhefe) contained α,α-trehalase, but didnot ferment α,α-trehalose [196]. Such observations indicated that the substrates were inaccessible to theenzymes across cell membranes.
By the mid-1950s, some studies had been published on the passage of various substrates acrossthe plasma membrane of yeasts. An example is the work on the uptake of sugars by yeasts [233].In addition, there was evidence for the active transport of monosaccharides across the membranes of
Figure 9. Exchanging a sulphur atom for the oxygen of the bridge converts an O-glycoside to a thioglycoside. From Barnett,1981 [19]
31 The state of ‘crypticity’: certain cells are unable to metabolize a given substrate which is supplied exogenously, although they possessthe relevant enzyme system [51].32 This work and much of the later work on galactose metabolism was done with ‘Saccharomyces fragilis’, here called Kluyveromycesmarxianus (see [24]).
Copyright 2004 John Wiley & Sons, Ltd. Yeast 2004; 21: 703–746.

History of research on yeasts 7 719
primate erythrocytes by means of ‘a temporary complex formed between the sugars and a cell-surfacecomponent during the transfer’ ([150] p. 135).
Georges Cohen33 and Monod argued that this entry of organic substrates into microbial cellsis ‘mediated by more or less selective permeation systems’ ([51] p. 169), which they proceededto characterize. In 1956, they published their seminal findings, which led them to the concept of‘permeases’.34 Monod and his colleagues wrote:
In this report we describe a system characterized by the property of accumulating exogenousgalactosides in the cells of Escherichia coli. The discovery of this inducible system, distinctfrom β-galactosidase, but which controls in vivo the activity of this enzyme as well as itsinduction, gives a solution to numerous problems posed by the metabolism of galactosides andby the induction of β-galactosidase in E. coli, and provides experimental confirmation of thehypothesis, often discussed, that stereospecific and functionally specialized catalytic systems,distinct from metabolic enzymes themselves, govern the penetration of certain substances intomicrobial cells.35
Two observations provided their primary evidence: (a) cells may be cryptic towards one sugar, yet quitenormal towards others; and (b) some cells have the capacity to accumulate certain substrates internally.Accordingly, they thought it necessary to explain such observations in terms of a number ‘of specificpermeation systems for which no positive evidence existed, and towards which no direct experimentalapproach seemed open’ ([51] p. 171). The alternative interpretation, that specific crypticity was due toinactivity of the intracellular enzyme, soon had to be abandoned.
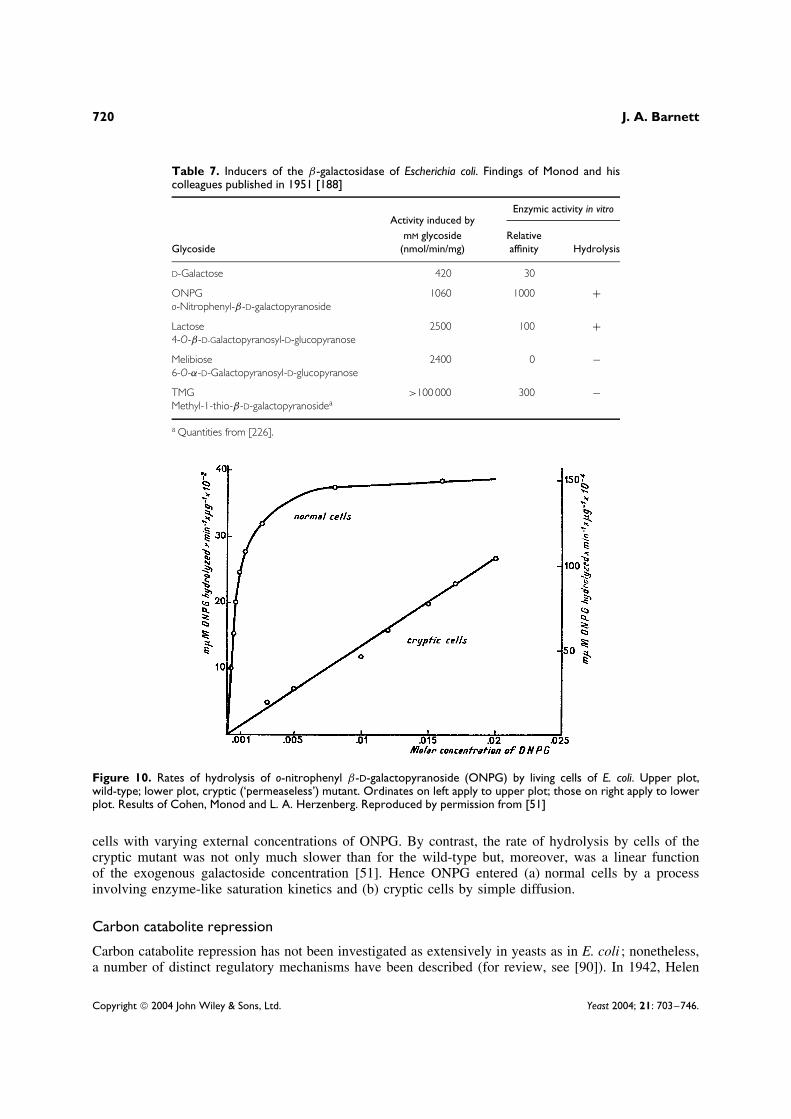
The workers in Monod’s laboratory made use of various synthetic glycosides (Table 7) for theirinduction experiments, notably o-nitrophenyl β-D-galactopyranoside36 (ONPG) as well as the thio-galactoside, TMG (Appendix). Joshua Lederberg37 had already introduced ONPG in 1950 for studyingβ-galactosidase activity [148]; this enzyme acts on ONPG, for which the enzyme has a high affinity,38
and liberates o-nitrophenol which is easy to measure spectrophotometrically:39
β-galactosidase
o-nitrophenyl β-D-galactopyranoside (ONPG) −−→ D-galactose + o-nitrophenol
In a momentous and much quoted experiment, using ONPG, Monod and his disciples compared theβ-galactosidase activity of intact (a) wild-type cells and (b) cells of a cryptic mutant of E. coli [51,118](Figure 10). They showed that uptake of a non-hydrolysed thio-β-galactoside had enzyme-like saturationkinetics [134]. This observation was consistent with the observed rates of hydrolysis by intact wild-type
33 Georges N. Cohen (b. 1920), French biochemist and molecular geneticist, Directeur de Recherche Emerite, Centre National de laRecherche Scientifique and Professeur Honoraire a l’Institut Pasteur [7].34 ‘Permeases’, also called ‘carriers’ or ‘transportases’ (the present writer favours ‘carriers’), are associated physically with the plasmamembrane. They bind the specific solute to be transported and undergo a series of conformational changes, thereby transferring thebound substrate across the membrane.35 Nous decrivons, dans ce memoire, un systeme caracterise par la propriete d’accumuler, dans les cellules d’Escherichia coli, lesgalactosides exogenes. La decouverte de ce systeme inductible, distinct de la β-galactosidase, mais qui commande in vivo l’activitede cet enzyme ainsi que son induction, donne une solution a de nombreux problemes que posaient le metabolisme des galactosideset l’induction de la β-galactosidase chez E. coli, et apporte une confirmation experimentale a l’hypothese, souvent envisagee, quedes systemes catalytiques steriquement specifiques et fonctionellement specialises, distincts des enzymes metaboliques proprement dits,gouvernent la penetration de certains substrats dans les cellules microbiennes ([226] p. 829).36 ONPG was prepared for Lederberg by chemists at Wisconsin [241].37 Joshua Lederberg (b. 1925), American microbial geneticist was, with Edward Tatum (1909–1975), the first to demonstrate sexualreproduction in bacteria [149]. Lederberg, who shared the Nobel Prize for physiology or medicine in 1958 with Tatum and GeorgeBeadle (1903–1989), was professor of genetics in the School of Medicine at Stanford University, California, 1959–1978 [Who’s Whoin America, 2003].38 Dissociation constants of E. coli β-galactosidase: 1.4 × 10−3 M-lactose and 1.4 × 10−4 M-ONPG ([148] p. 388).39 ONPG has a λmax of 420 nm at pH 10.2.
Copyright 2004 John Wiley & Sons, Ltd. Yeast 2004; 21: 703–746.
720 J. A. Barnett
Table 7. Inducers of the β-galactosidase of Escherichia coli. Findings of Monod and hiscolleagues published in 1951 [188]
Activity induced byEnzymic activity in vitro
GlycosidemM glycoside
(nmol/min/mg)Relativeaffinity Hydrolysis
D-Galactose 420 30
ONPG 1060 1000 +o-Nitrophenyl-β-D-galactopyranoside
Lactose 2500 100 +4-O-β-D-Galactopyranosyl-D-glucopyranose
Melibiose 2400 0 −6-O-α-D-Galactopyranosyl-D-glucopyranose
TMG >100 000 300 −Methyl-1-thio-β-D-galactopyranosidea
a Quantities from [226].
Figure 10. Rates of hydrolysis of o-nitrophenyl β-D-galactopyranoside (ONPG) by living cells of E. coli. Upper plot,wild-type; lower plot, cryptic (‘permeaseless’) mutant. Ordinates on left apply to upper plot; those on right apply to lowerplot. Results of Cohen, Monod and L. A. Herzenberg. Reproduced by permission from [51]
cells with varying external concentrations of ONPG. By contrast, the rate of hydrolysis by cells of thecryptic mutant was not only much slower than for the wild-type but, moreover, was a linear functionof the exogenous galactoside concentration [51]. Hence ONPG entered (a) normal cells by a processinvolving enzyme-like saturation kinetics and (b) cryptic cells by simple diffusion.
Carbon catabolite repression
Carbon catabolite repression has not been investigated as extensively in yeasts as in E. coli ; nonetheless,a number of distinct regulatory mechanisms have been described (for review, see [90]). In 1942, Helen
Copyright 2004 John Wiley & Sons, Ltd. Yeast 2004; 21: 703–746.

History of research on yeasts 7 721
Epps and Ernest Gale40 described the ‘glucose effect’: ‘the presence of glucose in the medium during thegrowth of E. coli suppresses the formation of certain enzymes’ [85]. Monod confirmed this observationin 1947 for several enzymes responsible for the breakdown of other sugars and their derivatives. In 1960,Harlyn Halvorson41 explained that ‘The mechanism of these ‘glucose effects’ is little understood’ ([106]p. 121) and a year later Boris Magasanik42 renamed Gale’s ‘glucose effect’ as ‘catabolite repression’[166].
In 1956, A. Davies43 showed that glucose represses the production of constitutive invertase44 byKluyveromyces marxianus. He detected the greatest activity of this enzyme in yeast which had its growthrate much restricted, that is when it was grown in <60 µM-glucose [60]. In the same yeast, invertaseformation was shown to be inhibited too by its own substrate, sucrose, unless the sucrose concentrationwas low: in this case, repression probably occurs because hydrolysis of the sucrose produces excesshexose. In addition, Davies observed the glucose repression of β-galactosidase in K. marxianus [61].
The first unequivocal evidence that the ‘glucose effect’ involved the specific inhibition of enzymesynthesis was discovered only in 1953 [56,187]. Furthermore, Monod and his colleagues soon realizedthat, although repression and induction produced opposite effects, they were strikingly similar and withsimilar kinetics. Both are highly specific and control the rate of enzyme synthesis, although unrelatedto substrate-specificity. Functionally associated enzymes were often found to be co-induced or co-repressed and, hence, induction and repression were thought to have similar underlying mechanisms[125]. Moreover, since specificity of induction or repression of an enzyme is not associated with itsstructural specificity, the structural genes could not also be responsible for regulation. In the words ofMonod and Jacob:
Since the specificity of induction or repression is not related to the structural specificity of thecontrolled enzymes, and since the rate of synthesis of different enzymes appears to be governedby a common element, this element is presumably not controlled . . . by the structural genesthemselves. This inference . . . is confirmed by the study of certain mutations which convertinducible or repressible systems into constitutive systems ([125] p. 328).
Juana Maria Gancedo’s excellent review of the extensive research on carbon catabolite repressionin S. cerevisiae describes its complex regulatory system and also considers some of the work on afew other species [90]. Another valuable review of the subject is that of Karl-Dieter Entian and Hans-Joachim Schuller [83]. Unlike in E. coli [29], catabolite repression in yeasts is not associated with lowconcentrations of cAMP [86]. Gancedo points out that ‘Catabolite repression can be exerted not only bythe three related sugars glucose, mannose and fructose, but also by other types of sugars like galactose[219] or maltose’ [86].
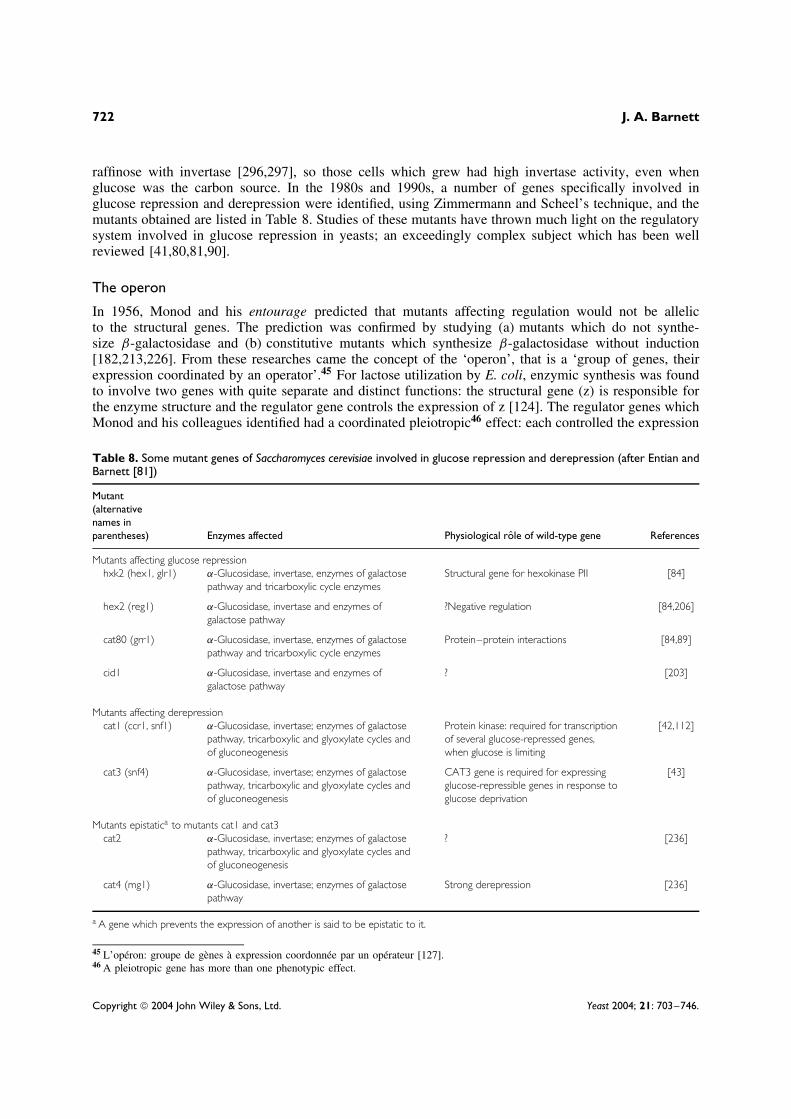
Many of the components of the repression mechanisms were identified by obtaining mutants inwhich catabolite repression had been abolished. Zimmermann and Scheel, of the Technische HochschuleDarmstadt, made an ingenious technical advance with the selection system they introduced in 1977[303]. They plated S. cerevisiae, which was growing exponentially on glucose as carbon source, onmedia containing 2-deoxy-D-glucose (2-deoxy-D-arabino-hexose) plus raffinose. Selection depends ontwo particular characteristics of this sugar which, unlike D-glucose, is not used for growth but, likeD-glucose, represses the synthesis of a number of catabolic enzymes [299]. S. cerevisiae hydrolyses
40 Ernest Frederick Gale (b. 1914), English microbial biochemist, was professor at Cambridge University 1960–1981 [12].41 Harlyn Odell Halvorson (b. 1925), American microbial biochemist, was professor at the University of Wisconsin (1956–1971) and atBrandeis University since 1971 [8].42 Boris Magasanik (b. 1919), American microbiologist, emigrated from the USSR to Austria in 1921 and thence to America in 1938,becoming professor at Massachusetts Institute of Technology from 1960 [9,11,168].43 A. Davies (1931–?1994) was a student in the Biochemistry Department of Cambridge University. His thesis (1955) was entitled ‘TheEffect of Environment on the Enzyme Constitution of Yeast’ and subsequently he worked for Imperial Chemical Industries (informationkindly supplied by Elizabeth Stratton, Archivist, Selwyn College, Cambridge).44 Davies’s ‘invertase’ may well have been inulinase (EC 3.2.1.7), not β-fructofuranosidase (EC 3.2.1.26) [243] (discussed in [18]pp. 187–188).
Copyright 2004 John Wiley & Sons, Ltd. Yeast 2004; 21: 703–746.
722 J. A. Barnett
raffinose with invertase [296,297], so those cells which grew had high invertase activity, even whenglucose was the carbon source. In the 1980s and 1990s, a number of genes specifically involved inglucose repression and derepression were identified, using Zimmermann and Scheel’s technique, and themutants obtained are listed in Table 8. Studies of these mutants have thrown much light on the regulatorysystem involved in glucose repression in yeasts; an exceedingly complex subject which has been wellreviewed [41,80,81,90].
The operon
In 1956, Monod and his entourage predicted that mutants affecting regulation would not be allelicto the structural genes. The prediction was confirmed by studying (a) mutants which do not synthe-size β-galactosidase and (b) constitutive mutants which synthesize β-galactosidase without induction[182,213,226]. From these researches came the concept of the ‘operon’, that is a ‘group of genes, theirexpression coordinated by an operator’.45 For lactose utilization by E. coli, enzymic synthesis was foundto involve two genes with quite separate and distinct functions: the structural gene (z) is responsible forthe enzyme structure and the regulator gene controls the expression of z [124]. The regulator genes whichMonod and his colleagues identified had a coordinated pleiotropic46 effect: each controlled the expression
Table 8. Some mutant genes of Saccharomyces cerevisiae involved in glucose repression and derepression (after Entian andBarnett [81])
Mutant(alternativenames inparentheses) Enzymes affected Physiological role of wild-type gene References
Mutants affecting glucose repressionhxk2 (hex1, glr1) α-Glucosidase, invertase, enzymes of galactose
pathway and tricarboxylic cycle enzymesStructural gene for hexokinase PII [84]
hex2 (reg1) α-Glucosidase, invertase and enzymes ofgalactose pathway
?Negative regulation [84,206]
cat80 (grr1) α-Glucosidase, invertase, enzymes of galactosepathway and tricarboxylic cycle enzymes
Protein–protein interactions [84,89]
cid1 α-Glucosidase, invertase and enzymes ofgalactose pathway
? [203]
Mutants affecting derepressioncat1 (ccr1, snf1) α-Glucosidase, invertase; enzymes of galactose
pathway, tricarboxylic and glyoxylate cycles andof gluconeogenesis
Protein kinase: required for transcriptionof several glucose-repressed genes,when glucose is limiting
[42,112]
cat3 (snf4) α-Glucosidase, invertase; enzymes of galactosepathway, tricarboxylic and glyoxylate cycles andof gluconeogenesis
CAT3 gene is required for expressingglucose-repressible genes in response toglucose deprivation
[43]
Mutants epistatica to mutants cat1 and cat3cat2 α-Glucosidase, invertase; enzymes of galactose
pathway, tricarboxylic and glyoxylate cycles andof gluconeogenesis
? [236]
cat4 (mg1) α-Glucosidase, invertase; enzymes of galactosepathway
Strong derepression [236]
a A gene which prevents the expression of another is said to be epistatic to it.
45 L’operon: groupe de genes a expression coordonnee par un operateur [127].46 A pleiotropic gene has more than one phenotypic effect.
Copyright 2004 John Wiley & Sons, Ltd. Yeast 2004; 21: 703–746.

History of research on yeasts 7 723
of several structural genes, which were closely associated with each other and encoded enzymes of thesame biochemical sequence.47 Jacob and his fellow-authors explained the mechanism of this effect interms of a new genetic entity, the operator [127]. This operator, adjacent to a group of genes, (a) wouldcontrol their activity and (b) would be responsive to the repressor produced by a particular regulatorgene (Figure 11). This hypothesis of how enzymic activity is regulated formed the basis of valid laterexplanations, including that of the molecular regulation of the galactose pathway in yeasts.
The galactose pathway in yeasts: work of Leloir and others
Working in Buenos Aires between 1948 and 1952, Leloir and his colleagues elucidated the pathway ofD-galactose catabolism in Kluyveromyces marxianus. In this pathway (Figure 12), the isomerization ofD-galactose 1-phosphate to D-glucose 1-phosphate involves uridine diphosphate (UDP) intermediates, andnot the direct isomerization of the galactose phosphate molecule (Figure 13).
As long ago as 1935, glucose and fructose phosphates had been found to accumulate during galactosefermentation by top and bottom brewing yeasts [101]. Furthermore, in 1943 Hans Kosterlitz48 hadsuggested that D-galactose 1-phosphate49 had a role in galactose fermentation as it is fermented byextracts from a galactose-adapted brewing yeast [139]. He made the suggestion, which later proved to betrue, that adaptation to galactose involved the formation of two new enzymes catalysing the followingreactions:
(i) D-galactose + ATP −−→ D-galactose 1-phosphate + ADP(ii) D-galactose 1-phosphate −−→←−− D-glucose 1-phosphate
Although in his 1946 review, Spiegelman was still writing of the adaptation of ‘galactozymase’[248], the next year he and his colleagues found that the mechanism by which galactose iscatabolized by S. cerevisiae involves more than one (unspecified) enzyme [256]. Then in 1949 JohnWilkinson,50 working in Cambridge, showed that Dutch top yeast (S. cerevisiae) produces galactokinase
Figure 11. Diagram of the operon and the mechanisms for synthesis and regulation of enzymes, suggested in 1961 byJacob and Monod ([125] p. 344). Copyright 1961, reproduced with permission from Elsevier
47 Les genes regulateurs identifies jusqu’a ce jour presentent la propriete remarquable d’exercer un effet pleiotrope coordonne, chacungouvernant l’expression de plusieurs genes de structure, etroitement lies entre eux, et correspondant a des proteines-enzymes appartenanta une meme sequence biochimique ([127] p. 1727).48 Hans Walter Kosterlitz (1903–1996), German-born British pharmacologist, professor at Aberdeen University (1968–1973), Directorof Unit for Research on Addictive Drugs from 1973; discovered (1975), with John Hughes, enkephalins, two potent naturally-occurringopiates in the brain [6,10].49 Not galactose 6-phosphate, as might have been expected by analogy with glucose fermentation.50 John Frome Wilkinson (b. 1925) received his doctorate at Cambridge in 1950, one of his examiners having been Jaques Monod.Wilkinson worked at Edinburgh University from 1949 to 1991, where he became professor of microbiology (personal communicationsfrom B. E. B. Moseley and J. F. Wilkinson).
Copyright 2004 John Wiley & Sons, Ltd. Yeast 2004; 21: 703–746.

724 J. A. Barnett
Figure 12. The galactose pathway (Barnett, 1976 [18]). NB phosphoglucomutase is now EC 5.4.2.7 [286]
Figure 13. The change in configuration of the C-4 of D-galactose 1-phosphate to D-glucose 1-phosphate achieved in theLeloir pathway
adaptively. Extracts of galactose-adapted yeast formed ‘an easily hydrolysable phosphoric ester fromgalactose and adenosinetriphosphate’. Some evidence was also presented ‘that galactose-1-phosphate isfurther transformed into glucose-6-phosphate’ [291]. Wilkinson named the adaptively-produced enzyme‘galactokinase’, as had Leloir and his colleagues a year earlier for an enzyme which they detected inlactose-grown Kluyveromyces marxianus (‘Saccharomyces fragilis’) [36]. Clearly, these were two quiteindependent observations, since the Biochemical Journal received Wilkinson’s script in July 1948, so hewould not have seen the earlier publication in Enzymologia before submitting his paper.
What led Leloir, working in Buenos Aires in the 1940s, to study galactose utilization by K. marxianus?He relates:
. . . it was known . . . that glycogen could be formed from glucose-1-phosphate . . . and it hadbeen shown . . . that sucrose could be formed from glucose-1-phosphate and fructose with anenzyme of bacterial origin . . . Consequently the idea that lactose originated from glucose-1-phosphate and galactose was floating in the air . . . we started studies with a lactose-utilizingyeast (Saccharomyces fragilis) which grew on whey in large milk cans . . . This led us to thestudy of galactose utilization ([155] p. 28).
Copyright 2004 John Wiley & Sons, Ltd. Yeast 2004; 21: 703–746.

History of research on yeasts 7 725
His laboratory was not well equipped:
The most sophisticated piece [of equipment] was a Beckman DU spectrophotometer, run bydry cells which were very difficult to get. We also had a Klett–Summerson photocolorimeter, amicroscope and a home-made Warburg respirometer. No refrigerated centrifuge was availablefor a long time and of course all the initial basic discoveries on sugar nucleotides were madewithout the help of radioactive materials ([155] p. 27).
And further:
. . . we used to have around the lab all kinds of fancy bottles, that were originally containersof perfumes, shampoos or prescriptions. Many came from Dr Leloir’s home and he insisted instoring reagents in them with the idea that the non-uniformity of the shapes and colors helpedto avoid mistakes ([155] p. 34).
In 1949 Leloir and his colleagues found that K. marxianus converts D-glucose 1-phosphate to D-glucose6-phosphate by the action of phosphoglucomutase51 [39], so they now had evidence that ‘galactozymase’involves the following series of reactions:
D-galactose + ATP
↓ galactokinase
D-galactose 1-phosphate + ADP
↓ thermostable factor
D-glucose 1-phosphate
↓ phosphoglucomutase
D-glucose 6-phosphate
Uridine diphosphate sugars
A year later, Leloir and his colleagues announced a finding of major significance when they described‘uridine diphosphate glucose: the coenzyme of the galactose–glucose phosphate isomerization’ [37,40].Still working with Kluyveromyces marxianus, they detected a thermostable factor necessary for convertingD-galactose 1-phosphate to D-glucose 1-phosphate. They determined the activity of this ‘coenzyme’ byusing galactose 1-phosphate as substrate with excess phosphoglucomutase and determining the glucose6-phosphate formed:
‘coenzyme’ phosphoglucomutase
galactose 1-phosphate −−→ glucose 1-phosphate −−→ glucose 6-phosphate
Uridine diphosphate (UDP) was extracted with ethanol from baker’s yeast, fractionally precipitated withmercuric acetate, absorbed on charcoal, eluted with ethanol and treated with a cation-exchange resin [37].The authors comment that ‘The mechanism by which uridine diphosphate accelerates the conversion ofgalactose into glucose will require further investigation’ ([40] p. 192). This remarkable reaction involves
51 The Coris, in whose laboratory Leloir worked in the 1940s, discovered phosphoglucomutase in 1938 [58,59], which was purified frommuscle in 1948 by Victor Assad Najjar [197]. This enzyme converts glucose 1-phosphate to glucose 6-phosphate. The catalytic site of themutase includes a phosphorylated serine residue. This phosphoryl group transfers to the C-6 hydroxyl group of glucose 1-phosphate toform glucose 1,6-bisphosphate, the C-1 phosphoryl group of which attaches to the same serine residue, so forming glucose 6-phosphateand regenerating the mutase [198,224,225].
Copyright 2004 John Wiley & Sons, Ltd. Yeast 2004; 21: 703–746.

726 J. A. Barnett
a change in configuration at C-4 of the galactose molecule to form glucose 1-phosphate (Figure 13).The enzyme responsible for the inversion was called ‘galactowaldenase’52 in order to avoid the word‘isomerase’, which had been used for many other enzymes [131,154]. The enzyme producing the inversionis now called uridyl transferase (Table 9).
In 1950, Leloir and his colleagues isolated from baker’s yeast ‘the coenzyme of the galactose-1-phosphate → glucose-1-phosphate transformation’. They wrote: ‘The substance contains uridine, twophosphate groups and glucose, and has therefore been named uridine-diphosphate-glucose’ (UDPGlc)([37] p. 349). Leloir tells us that ‘this was rather exciting, because at that time uridine was known onlyas a nucleic acid constituent’ ([155] p. 30). In the following year, he described this compound (seeFigure 14) as a ‘glucose-1-phosphate molecule attached to uridine 5′-phosphate forming a pyrophosphatelink’ [153].
Leloir and his colleagues measured the molecular mass of uridine diphosphate glucose using aKuhlmann microbalance (Figure 15), which weighs to about 1 µg [79,222]. This remarkably sensitiveinstrument has a double case to minimize the effects of room temperature changes and air movements;but it is not easy to use. Like all research balances made before the mid-1930s, it is not air-damped, so the
Table 9. Enzymes of the galactose pathway
Enzyme and Enzyme Commission number Reaction
Galactokinase 2.7.1.6 D-galactose + ATP → D-galactose-1-phosphate + ADP
UTP-hexose-1-phosphate uridylyltransferase 2.7.7.10(galactose-1-phosphate uridylyltransferase)
α-D-galactose-1-phosphate � UDP-galactose + pyrophosphate
UDP-glucose-hexose-1-phosphate uridylyltransferase 2.7.7.12(uridyl transferase)
UDP-glucose + α-D-galactose-1-phosphate �α-D-glucose-1-phosphate + UDP-galactose
UDP-glucose-4-epimerase 5.1.3.2 UDP-glucose � UDP-galactose
UTP-glucose-1-phosphate uridylyltransferase 2.7.7.9 α-D-glucose-1-phosphate � UDP-glucose + pyrophosphate
Phosphoglucomutase 5.4.2.2 α-D-glucose-1-phosphate � α-D-glucose-6-phosphate
Figure 14. Uridine 5′-diphospho-α-D-glucopyranose (UDP-glucose) and uridine 5′-diphospho-α-D-galactopyranose(UDP-galactose)
52 In 1950, Leloir and his colleagues had written that galactose 1-phosphate → glucose 1-phosphate consists in a Walden inversion of C-4and is catalysed ‘by an enzyme which is currently called “galactowaldenase” in this laboratory’ ([37] p. 333). The ‘Walden inversion’,i.e. the conversion of one optical isomer into a derivative of the other, was discovered in 1895 by the Latvian chemist, Paul Walden(1863–1957) [285].
Copyright 2004 John Wiley & Sons, Ltd. Yeast 2004; 21: 703–746.

History of research on yeasts 7 727
Figure 15. A Kuhlmann microbalance. From Pregl [222]
balance does not remain at rest, but continues oscillating like a pendulum for a long time. Consequently,it is necessary to observe and take a mean of the extreme positions of the pointer53 and estimating theamplitude of the oscillations is a time-consuming occupation ([155] p. 31).
Treatment of UDPGlc with an extract of K. marxianus produced a galactose-containing compound,uridine diphosphate galactose (UDPGal). Hence, Leloir was able to suggest two sequential steps to explainthe conversion of D-galactose 1-phosphate into D-glucose 1-phosphate:
(i) UDPGlc + D-galactose 1-phosphate −−→←−− UDPGal + D-glucose 1-phosphate(ii) UDPGal −−→←−− UDPGlc
So by the early 1950s, the old ‘galactozymase’, by which galactose is converted to glucose, had beenpartly unravelled [131,153,192] as follows:
(1)
galactokinaseD-galactose + ATP −−→ D-galactose 1-phosphate + ATP
‘galactowaldenase’(2) D-galactose 1-phosphate + UDPGlc −−→ UDPGal + D-glucose 1-phosphate
In addition, Herman Kalckar and his colleagues reported [131] two other reactions effected by extractsof K. marxianus, viz. a reaction between uridine triphosphate (UTP) and D-galactose 1-phosphate:
(3) UTP + D-galactose 1-phosphate −−→←−− UDPGal + D-glucose 1-phosphate
and another by which UDPGlc is converted to D-glucose 1-phosphate and UTP:
(4) UDPGlc + pyrophosphate −−→ D-glucose 1-phosphate + UTP [192]
The enzyme responsible for reaction (3) was later named UTP-hexose-1-phosphate uridylyltransferase[EC 2.7.7.10] and that for reaction (4) UTP-glucose-1-phosphate uridylyltransferase [EC 2.7.7.9]. Theauthors’ illustration of these reactions with UDPGlc, published in 1953, is reproduced in Figure 16.
53 The method of weighing using the swinging movement of non air-damped balances is described, for example, on page 8 of reference[273].
Copyright 2004 John Wiley & Sons, Ltd. Yeast 2004; 21: 703–746.

728 J. A. Barnett
Figure 16. The points of enzymic attack on uridine diphosphoglucose, as illustrated by Kalckar and his colleagues in 1953.Reprinted with permission from Nature [192]. Copyright (1953) Macmillan Magazines Limited
Figure 17. The reactions of the galactose pathway as represented in 1958 by Huguette de Robichon-Szulmajster [62]
Writing about the important contributions of Kalckar and his colleagues to understanding the galactosepathway, Leloir says:
Although our laboratories worked on similar lines there was never any rivalry or resentmentamong us as often happens. We exchanged information quite freely ([155] p. 33).
In 1958, Huguette de Robichon-Szulmajster54 summarized the galactose pathway (formerly knownsimply as ‘galactozymase’) [62] as comprising the reactions shown in Figure 17. Reaction 4 of thispathway shows the initial formation of a catalytic amount of UDPGlc which is necessary to start reaction(2). Working with galactokinase-negative haploid mutants, she found that, as well as galactokinase, twoother enzymes of the galactose pathway in S. cerevisiae are ‘adaptive’. These are galactose-1-phosphateuridyltransferase and UDP-glucose 4-epimerase, which are formed when growth is on galactose but notwhen on glucose; the same is true for the epimerase in K. marxianus [63]. Her findings (Table 10) werenot consistent with Stanier’s concept of sequential adaptation, mentioned above, since the free galactoseappeared to be a ‘multi-inducer’. She and a colleague purified UDP-glucose 4-epimerase in 1960 [174].Table 9 lists the enzymes of the galactose pathway.
54 Huguette de Robichon-Szulmajster worked at the Laboratoire d’Enzymologie, Centre national de la Recherche scientifique, Gif-sur-Yvette, France.
Copyright 2004 John Wiley & Sons, Ltd. Yeast 2004; 21: 703–746.

History of research on yeasts 7 729
Table 10. Activities of enzymes of the galactose pathway in yeasts grown on D-glucose orD-galactose. Results of de Robichon-Szulmajster [62,63] (after Barnett [18])
Galactokinaseless strain ofSaccharomyces cerevisiae
Kluyveromycesmarxianus
Carbon source in growth medium
Enzyme D-Glucose D-Galactose D-Glucose D-Galactose
Galactokinase 0 0 0 26 000
UDP-glucose-hexose-1-phosphateuridylyltransferase
0.01 8.96 10 3680
UDP-glucose-4-epimerase 0 5.70 0 333 000
UTP-glucose-1-phosphateuridylyltransferase
9.17 12.50 1770 2220
Specific activities, expressed in (nmoles of substrate reacted) min−1 (mg of protein)−1.
Genetic regulation of the galactose pathway
The GAL genes55
In the yeast Saccharomyces cerevisiae, the interplay between Gal3p, Gal80p and Gal4pdetermines the transcriptional status of the genes needed for galactose utilization [27].
In the 1940s and 1950s, several authors reported on their studies of the genetics of the ability of S.cerevisiae to ferment galactose [95,114,115,158,160,193,230,231,250,257,293]. The Lindegrens were thefirst to describe the genetic control of galactose fermentation in 1947 [160]. When they crossed haploidfermenters with non-fermenters, each ascus of the diploid hybrids produced two fermenters and twonon-fermenters. Their conclusion that galactose fermentation was determined by a single pair of alleleswas confirmed 4 years later by Herschel Roman56 and his colleagues [230]. The non-fermenting haploidswere found to differ from each other genetically, being recessive for the galactose fermenting genes atdifferent loci. One of them, G2 (later called GAL2 ), was described as probably ‘involved in the transportof galactose into the cell’ [70], which was confirmed by Vincent Cirillo57 in 1968 [48].
Donald Hawthorne58 found three dominant genes to be necessary for fermenting galactose rapidlyand designated the recessive alleles59 as g-1, g-2 and g-3, a slow fermenter of galactose having genesG-1G-1 G-2G-2 g-3 g-3. However, Lindegren drew attention to the difficulties in being certain of thedistinction between slow-fermenters and non-fermenters [159].
Øjvind Winge had already reported in 1948 that the ‘long-term adaptation’ of S. cerevisiae to galactoseis associated with presence of gene gs [293], and this was re-named60 ga-3 in 1963 [74] by Howard
55 The ways in which the genes were written changed over the years, at any one time varying from one author to another.56 Herschel Lewis Roman (1914–1989), Polish-born American yeast geneticist, was professor in the Department of Genetics at theUniversity of Washington, Seattle [91]. He wrote, in 1947, that Carl Lindegren’s ‘unorthodox interpretations presented a challenge thatwas largely responsible for my choosing yeast as an experimental organism’ ([229] p. 3).57 Vincent P. Cirillo worked at the State University of New York.58 Donald C. Hawthorne (1926–2003), American yeast geneticist, worked at the Department of Genetics of Washington University from1950 onwards, first as a student, later becoming a professor. Apart from his work on the genetics of galactose utilization, he is knownespecially for his work on chromosome mapping and on various suppressors. Bonny Brewer writes that he never had a telephone at homenor used a computer, that he walked to work and ‘he foraged for food . . . harvesting watercress from Ravenna Creek and collectingfennel and mushrooms from where he planted them in his walks around the city’ [304].59 Alleles or allelomorphs are different forms of a gene which are alternative to one another at the same locus. The term ‘allelomorph’was coined by Bateson and Saunders in 1902 ([26] p. 126) from the Greek: αλλος�′ (other); µορφη′ (form).60 ‘Hawthorne writes that g-3 is “allelic with” gs, but from his text it appears that he means ‘identical with’ ([298] p. 136).
Copyright 2004 John Wiley & Sons, Ltd. Yeast 2004; 21: 703–746.

730 J. A. Barnett
Douglas, who made major contributions to the understanding of the genetic regulation of galactosemetabolism. In 1976, he showed this gal3 mutation to be pleiotropic, also impairing the utilization ofmelibiose and maltose [135].
In 1968, Vincent Cirillo found that galactose enters galactokinaseless haploid cells of S. cerevisiae byfacilitated diffusion61 [48]. He showed the GAL2 gene (Roman’s G2) to be responsible for this (inducible)transport of which, for some strains, D-fucose and L-arabinose are gratuitous inducers. Mutants alteringthe inducibility of galactose pathway enzymes also affected the inducibility of galactose transport.
Regulation of the GAL genes
When D-galactose is the sole source of carbon, S. cerevisiae activates the GAL genes which encode theenzymes of the Leloir pathway. The main features of the regulation of the GAL genes were worked out byDouglas, Hawthorne and their colleagues in the 1960s [70–74] (Figure 18 and Table 11). They showedthat the closely linked genes GAL1, GAL7 and GAL10 encode galactokinase, galactose-1-phosphateuridylyltransferase and UDP-glucose 4-epimerase, respectively. They also found that the mutation gal4blocks the synthesis of these enzymes, since GAL4 is necessary for expressing GAL1, GAL7 and GAL10.[71,72]. In 1966, these two authors suggested that the regulatory mechanism of the galactose pathwayconforms with concept of the operon of Jacob and Monod [72,126]. Thus, for example, the GAL80 geneforms a repressor which represses the expression of the GAL4 gene when galactose is absent by affectingthe GAL81 site which, much like Monod’s lac operator, is the site of repressor recognition, controllingtranscription of the contiguous structural gene [170].
Subsequent work refined the Douglas–Hawthorne scheme and, by the 1990s, the mechanism of theregulation of the galactose pathway in S. cerevisiae had been shown to be as represented in Figure 19.The Gal4 protein activates GAL genes when galactose is in the medium. When it is absent, induction isprevented by the regulatory protein Gal80 [120] (Figure 18). Regions of yeast DNA have been isolatedcontaining genes, the expression of which depends on the nature of the exogenous carbon source[267], and one of these regions contains DNA sequences encoding three galactose-inducible RNAs,transcription of which is galactose-specific and depends on GAL4. The region encoding these RNAsis the GAL7–GAL10–GAL1 gene cluster [268]. D-Galactose activates the Gal3 protein, galactokinase,which interacts with the Gal80p–Gal4p complex, alleviating repression by Gal80p, hence allowingGal4p to activate transcription [15]. Glucose repression was found to be mediated by GAL4, which
Figure 18. Regulation of the galactose pathway in S. cerevisiae. Symbols: , gene encodes; , stimulates activity;, inhibits activity; ∗, repressed by D-glucose
61 Facilitated diffusion is carrier-mediated movement across a membrane which depends on a concentration gradient and not expenditureof metabolic energy (for review, see [78]).
Copyright 2004 John Wiley & Sons, Ltd. Yeast 2004; 21: 703–746.

History of research on yeasts 7 731
Table 11. Galactose pathway genes of Saccharomyces cerevisiae (afterJohnston and Carlson [130])
Gene Encodes/function References
GAL1 Galactokinase [268]GAL2 Galactose carrier [48,70,202,274]GAL3 Inducer [15,31]GAL4 Transcriptional activator,
mediating glucose repression[102,147,170]
GAL5 Phosphoglucomutase [210]GAL7 Uridyl transferase [268]GAL10 UDP-glucose-4-epimerase [268]GAL80 Inhibition of Gal4 protein [122]GAL81 Part of Gal4 protein [170]GAL82 Glucose repression [171,172]GAL83 Glucose repression [171,172]
Figure 19. The enzymes of the galactose pathway in S. cerevisiae and the genes encoding them (after Johnston [129]). Theenzymes are galactokinase (encoded by GAL1), UDP-glucose–hexose-1-phosphate uridylyltransferase (encoded by GAL7),UDP-glucose-4-epimerase (encoded by GAL10), phosphoglucomutase (encoded by GAL5)
Copyright 2004 John Wiley & Sons, Ltd. Yeast 2004; 21: 703–746.

732 J. A. Barnett
encodes the transcriptional activator of GAL genes [102], although the mechanisms appear to be complex[145].
In summary: (a) the protein encoded by the GAL4 gene activates the transcription of each regulatedGAL gene; (b) the Gal80 protein binds to the Gal4 protein, preventing its activating activity; and(c) galactokinase, the Gal3 protein, prevents the inhibitory action of the Gal80 protein (Figure 18).
In the 1980s, some of the studies on the GAL system were extended to another yeast, Kluyveromyceslactis, which also utilizes galactose by inducing enzymes of the galactose pathway. The GAL4 of S.cerevisiae was shown to activate the lactose–galactose operon (or regulon62) of K. lactis [228], whichhas similar GAL1, GAL4, GAL7, GAL10 coding to that of S. cerevisiae [177,178,227,234]. Inductionand repression of the galactose pathway enzymes in K. lactis were studied further in the 1990s, somemechanisms involving Gal4p [142,302]. The regulation of lactose and galactose metabolism by K. lactiswas reviewed in 1996 [289].
Genetic regulation of the utilization of other substrates
Adaptations to other kinds of substrate have been studied, such as that to nitrogen compounds (for review,see [167]). Here, it seems apposite to write briefly about some of the work which has been published onthe genetic regulation of two additional substrates, maltose and sucrose.
Genetic regulation of maltose utilization: the MAL genes
Brewers require S. cerevisiae to ferment maltose and maltotriose. Hence, along with galactose and sucrose,maltose was among the first markers to be used for genetic studies of yeasts. Indeed, in the 1940s and1950s, the pioneer yeast geneticist at the Carlsberg Laboratory in Copenhagen, Øjvind Winge, publishedwork on the genetics of α-glucosidase activity in brewing yeasts [293–295]. He identified four genes(MAL1–MAL4 ) for maltose fermentation in S. cerevisiae and a single gene (MAL6 ) in Saccharomycespastorianus (formerly S. carlsbergensis), any of which would encode α-glucosidase (Winge crossed thetwo species). The structural genes for maltose utilization are only expressed after induction by maltoseand are also subject to glucose repression (for reviews, see [82,83]).
Maltose utilization requires the functional presence of both a maltose carrier and a cytosolic α-glucosidase. In 1976, Gennadi Naumov reported that MAL loci contained both regulatory and structuralgenes, as well as three genes encoding the maltose carrier [199]. In addition, a transcriptional activatorprotein was described in the 1980s [44,52,53,201]. Maltose is generally taken into the cells of S. cerevisiaeby proton symport (for review, see [78]) and each molecule is hydrolysed in the cytosol by α-glucosidaseto give two molecules of D-glucose. The maltose carrier is encoded by the MAL61 gene [45], with thefirst indication of any MAL gene encoding this carrier coming in 1983 from the identification of aMAL1-linked temperature-sensitive maltose transport mutation [96].
Qi Cheng and Corinne Michels explain the nomenclature of the MAL genes:
Each [MAL] locus is a complex locus containing three genes required for maltose fermentation. . . We have established a two digit numbering system in order to distinguish the GENE 1, 2,and 3 functions mapping to the different MAL loci. The first digit indicates the locus positionand the second the GENE function . . . Thus the MAL61 gene is the GENE 1 function mappingto the MAL6 locus ([45] p. 477).
Two years later, those authors reported that the inducible, high-affinity maltose carrier of S. cerevisiaeis encoded by genes MAL11 and/or MAL61 [46]. Entian and Schuller review some of the intricacies ofthe genetics of the regulation of maltose utilization by S. cerevisiae [82].
62 A regulon is a system in which the coordinated regulation of two or more structural genes or operons, each with its own promoter, isachieved by a common regulator molecule.
Copyright 2004 John Wiley & Sons, Ltd. Yeast 2004; 21: 703–746.

History of research on yeasts 7 733
Genetic regulation of sucrose utilization: invertase and the SUC genesInvertase (β-fructofuranosidase) catalyses the hydrolysis of sugars, such as sucrose and raffinose, whichhave a terminal, unsubstituted β-D-fructofuranosyl group [195] (Appendix). The enzyme is located outsidethe plasma membrane and is associated physically with the cell wall (for review, see [19]). In 1967,Santiago Gascon and Paul Ottolenghi found that the invertase concentration varies by a factor of about1000 with the concentration of exogenous D-glucose [92] and, consistent with this finding, derepressionof this enzyme at <6 mM D-glucose was reported some 7 years later [176].
From the 1950s to the 1970s genetic analysis demonstrated that at least five genes, designatedSUC1–SUC5, encode invertase, any one of which is sufficient for invertase synthesis and, hence, theutilization of sucrose or raffinose [95,104,211,295]. Invertase molecular genetics was studied extensivelyin the 1980s and 1990s and regulation was found to be at transcription [235]. Mark Johnston and Mar-ian Carlson have reviewed this subject, as well as the regulation of the utilization of other organiccompounds [130].
Conclusion
The small beginning in 1900 of studying the adaptation of yeasts to D-galactose was followed up throughthe twentieth century, eventually to generate considerable understanding of the complexities of themolecular regulation of enzyme synthesis. The enormous volume of work published on the regulation ofenzymes and transport carriers, and on the molecular control of both, for S. cerevisiae and also for E.coli, using increasingly refined techniques, has made it impracticable to give here more than an outlineof how this study developed. Only mechanisms underlying the regulation of the amount of enzymes inthe cells have been discussed; other important systems of enzymic regulation, such as interconversionby covalent modification and systems affecting enzymic activity, allosteric activation and deactivation,have been ignored. The genetic regulatory mechanism of S. cerevisiae, acting on the GAL genes whichencode the enzymes of galactose utilization, has been the most intensively studied and has become thebest understood genetic regulatory mechanism in any eukaryote.
Kevin Struhl tells us:
. . . classical and molecular yeast genetics has permitted the discovery and functionalcharacterization of transcriptional regulatory proteins that were not identified in biochemicalstudies. Thus, genetic analysis in yeast has often generated information complementary to thatobtained from biochemical studies of transcription in vitro, and it has provided unique insightsinto mechanisms of eukaryotic transcriptional regulation ([269] p. 651).
And work on this mechanism continues today [2].An account of the history of research on aspects of more general metabolic regulation in yeasts, such
as the Pasteur and Kluyver effects, the regulation of glycolysis and aerobic metabolism, will be given ina later article.
AcknowledgementsThe errors are entirely mine; but I thank the following most warmly for all the help they have given me: B. J.Brewer, Melvin Cohn, H. B. F. Dixon, K.-D. Entian, Peter Gray, Robert Hauer, Alexandre Herlea, M. C. Kielland-Brandt, B. E. B. Moseley, Andrea Munsterberg, P. M. Nurse, Georges Pedro, N. M. Temperley, J. F. Wilkinson, M. D.Yudkin, F. K. Zimmermann. I am also very much indebted to L. K. Barnett for extensive criticisms of the text and muchhelp with the figures, as well as to the Royal Society for a research grant.
References
1. Albers H. 1936. Wesen und Wirkung der Fermente. Angewandte Chemie 49: 448–455.2. Alberti A, Lodi T, Ferrero I, Donnini C. 2003. MIG1-dependent and MIG1-independent regulation of GAL gene expression in
Saccharomyces cerevisiae: role of Imp2p. Yeast 20: 1085–1096.
Copyright 2004 John Wiley & Sons, Ltd. Yeast 2004; 21: 703–746.

734 J. A. Barnett
3. Ambler RP, Murray K. 2002. Martin Rivers Pollock. Biographical Memoirs of Fellows of the Royal Society 48: 359–373.4. Anon. 1948. Prof. Frederic Dienert. Chemistry and Industry: 398.5. Anon. 1953. Arthur Slator. Journal of the Royal Institute of Chemistry 77: 460–461.6. Anon. 1987. Kosterlitz Hans W. Who’s Who in Science in Europe, 5th edn, vol 2. Longman: Harlow; 1329.7. Anon. 1987. Cohen Georges N. Who’s Who in Science in Europe, 5th edn, vol 1. Longman: Harlow; 429.8. Anon. 1989. Halvorson, Harlyn Odell. American Men and Women of Science 1989–90, 17th edn, vol 3. Bowker: New York; 452.9. Anon. 1989. Magasanik, Boris. American Men and Women of Science 1989–90, 17th edn, vol 5. Bowker: New York; 135.
10. Anon. 2001. Kosterlitz, Hans Walter. Who Was Who 1996–2000. Palgrave: New York; 331.11. Anon. 2003. Magasanik, Boris. Who’s Who in America 2003, 57th edn, vol 2. Marquis Who’s Who; New Providence; 3299.12. Anon. 2003. Gale, Prof. Ernest Frederick. Who’s Who 2003, 155th edn. Black: London; 775–776.13. Anon. 2003. Jacob (Francois). Who’s Who in France (Qui est Qui en France) 2002–2003. Lafitte: Levallois-Perret; 998.14. Armstrong EF. 1905. Studies in enzyme action. VIII. The mechanism of fermentation. Proceedings of the Royal Society of London
B 76: 600–605.15. Bajwa W, Torchia TE, Hopper JE. 1988. Yeast regulatory gene GAL3 : carbon regulation; UASGal elements in common with
GAL1, GAL2, GAL7, GAL10, GAL80, and MEL1 ; encoded protein strikingly similar to yeast and Escherichia coli galactokinases.Molecular and Cellular Biology 8: 3439–3447.
16. Barnett JA. 1968. The catabolism of acyclic polyols by yeasts. Journal of General Microbiology 52: 131–159.17. Barnett JA. 1975. The entry of D-ribose into some yeasts of the genus Pichia . Journal of General Microbiology 90: 1–12.18. Barnett JA. 1976. The utilization of sugars by yeasts. Advances in Carbohydrate Chemistry and Biochemistry 32: 125–234.19. Barnett JA. 1981. The utilization of disaccharides and some other sugars by yeasts. Advances in Carbohydrate Chemistry and
Biochemistry 39: 347–404.20. Barnett JA. 2000. A history of research on yeasts 2: Louis Pasteur and his contemporaries, 1850–1880. Yeast 16: 755–771.21. Barnett JA. 2003. A history of research on yeasts 5: the fermentation pathway. Yeast 20: 509–543.22. Barnett JA. 2003. A history of research on yeasts 6: the main respiratory pathway. Yeast 20: 1015–1044.23. Barnett JA, Lichtenthaler FW. 2001. A history of research on yeasts 3: Emil Fischer, Eduard Buchner and their contemporaries,
1880–1900. Yeast 18: 363–388.24. Barnett JA, Payne RW, Yarrow D. 2000. Yeasts: Characteristics and Identification, 3rd edn. Cambridge University Press:
Cambridge.25. Barnett JA, Robinow CF. 2002. A history of research on yeasts 4: cytology part II, 1950–1990. Yeast 19: 745–772.26. Bateson W, Saunders ER. 1902. Experimental studies in the physiology of heredity. Reports of the Evolution Committee of the
Royal Society 1: 1–160.27. Bhat PJ, Murthy TVS. 2001. Transcriptional control of the GAL/MEL regulon of yeast Saccharomyces cerevisiae. Molecular
Microbiology 40: 1059–1066.28. Blagden C. 1790. Report on the best method of proportioning the excise upon spirituous liquors. Philosophical Transactions of the
Royal Society of London 80: 321–345.29. Botsford JL. 1981. Cyclic nucleotides in procaryotes. Microbiological Reviews 45: 620–642.30. Bourquelot E. 1888. Sur la fermentation alcoolique du galactose. Comptes Rendus Hebdomadaires des Seances de l’Academie des
Sciences, Paris 106: 283–286.31. Broach J. 1979. Galactose regulation in Saccharomyces cerevisiae. The enzymes encoded by the GAL7, 10, 1 cluster are coordinately
controlled and separately translated. Journal of Molecular Biology 131: 41–53.32. Browne CA. 1912. A Handbook of Sugar Analysis. Wiley: New York.33. Brunotte R. 1948. Frederic Dienert. La Technique Sanitaire et Municipale 3 (Nos. 3–4, March–April).34. Burnet FM. 1956. Enzyme, Antigen and Virus. Cambridge University Press: Cambridge.35. Buttin G. 1961. Some aspects of regulation in the synthesis of the enzymes governing galactose metabolism. Cold Spring Harbor
Symposia on Quantitative Biology 26: 213–216.36. Caputto R, Leloir LF, Trucco RE. 1948. Lactase and lactose fermentation in Saccharomyces fragilis . Enzymologia 12: 350–355.37. Caputto R, Leloir LF, Cardini CE, Paladini AC. 1950. Isolation of the coenzyme of the galactose phosphate–glucose phosphate
transformation. Journal of Biological Chemistry 184: 333–350.38. Caputto R, Leloir LF, Trucco RE, Cardini CE, Paladini A[C]. 1948. A coenzyme for phosphoglucomutase. Archives of Biochemistry
18: 201–203.39. Caputto R, Leloir LF, Trucco RE, Cardini CE, Paladini AC. 1949. The enzymatic transformation of galactose into glucose
derivatives. Journal of Biological Chemistry 179: 497–498.40. Cardini CE, Paladini AC, Caputto R, Leloir LF. 1950. Uridine diphosphate glucose: the coenzyme of the galactose–glucose
phosphate isomerization. Nature 165: 191–192.41. Carlson M. 1999. Glucose repression in yeast. Current Opinion in Microbiology 2: 202–207.42. Celenza JL, Carlson M. 1986. A yeast gene that is essential for release from glucose repression encodes a protein kinase. Science
233: 1175–1180.43. Celenza JL, Eng FJ, Carlson M. 1989. Molecular analysis of the SNF4 gene of Saccharomyces cerevisiae: evidence for physical
association of the SNF4 protein with protein kinase. Molecular and Cellular Biology 9: 5045–5054.44. Charron MJ, Dubin RA, Michels CA. 1986. Structural and functional analysis of the MAL1 locus of Saccharomyces cerevisiae.
Molecular and Cellular Biology 6: 3891–3899.
Copyright 2004 John Wiley & Sons, Ltd. Yeast 2004; 21: 703–746.

History of research on yeasts 7 735
45. Cheng Q, Michels CA. 1989. The maltose permease encoded by the MAL61 gene of Saccharomyces cerevisiae exhibits bothsequence and structural homology to other sugar transporters. Genetics 123: 477–484.
46. Cheng Q, Michels CA. 1991. MAL1 and MAL61 encode the inducible high-affinity maltose transporter of Saccharomyces cerevisiae.Journal of Bacteriology 173: 1817–1820.
47. Chiba S, Sugawara S, Shimomura T, Nakamura Y. 1962. Comparative biochemical studies of α-glucosidases Part I. Substratespecificity and transglucosidation action of an α-glucosidase of brewer’s yeast. Agricultural and Biological Chemistry 26: 787–793.
48. Cirillo VP. 1968. Galactose transport in Saccharomyces cerevisiae I. Nonmetabolized sugars as substrates and inducers of thegalactose transport system. Journal of Bacteriology 95: 1727–1731.
49. Clarke PH. 1986. Roger Yate Stanier. Biographical Memoirs of Fellows of the Royal Society 32: 543–568.50. Cohen GN. 1965. Regulation of enzyme activity in microorganisms. Annual Review of Microbiology 19: 105–126.51. Cohen GN, Monod J. 1957. Bacterial permeases. Bacteriological Reviews 21: 169–194.52. Cohen JD, Goldenthal MJ, Buchferer B, Marmur J. 1984. Mutational analysis of the MAL1 locus of Saccharomyces: identification
and functional characterization of three genes. Molecular and General Genetics 196: 208–216.53. Cohen JD, Goldenthal MJ, Chow T, Buchferer B, Marmur J. 1985. Organization of the MAL loci of Saccharomyces. Physical
identification and functional characterization of three genes at the MAL6 locus. Molecular and General Genetics 200: 1–8.54. Cohn M. 1957. Contributions of studies on the β-galactosidase of Escherichia coli to our understanding of enzyme synthesis.
Bacteriological Reviews 21: 140–168.55. Cohn M. 1979. In memoriam. In Origins of Molecular Biology. A Tribute to Jacques Monod, Lwoff A, Ullmann A (eds). Academic
Press: New York; 75–87.56. Cohn M, Monod J. 1953. Specific inhibition and induction of enzyme biosynthesis. Adaptation in Micro-organisms. Third
Symposium of the Society for General Microbiology ; 132–149.57. Cohn M, Monod J, Pollock MR, Spiegelman S, Stanier RY. 1953. Terminology of enzyme formation. Nature 172: 1096.58. Cori GT, Colowick SP, Cori CF. 1938. The formation of glucose-1-phosphoric acid in extracts of mammalian tissues and of yeast.
Journal of Biological Chemistry 123: 375–380.59. Cori GT, Colowick SP, Cori CF. 1938. The enzymatic conversion of glucose-1-phosphoric ester to 6-ester in tissue extracts. Journal
of Biological Chemistry 124: 543–555.60. Davies A. 1956. Invertase formation in Saccharomyces fragilis . Journal of General Microbiology 14: 109–121.61. Davies A. 1956. Some factors affecting lactase formation and activity in Saccharomyces fragilis . Journal of General Microbiology
14: 425–439.62. de Robichon-Szulmajster H. 1958. Induction of enzymes of the galactose pathway in mutants of Saccharomyces cerevisiae. Science
127: 28–29.63. de Robichon-Szulmajster H. 1958. Uridine-diphosphogalactose-4-epimerase, an adaptive enzyme in Saccharomyces cerevisiae.
Biochimica et Biophysica Acta 29: 270–272.64. Dickson RC, Barr K. 1983. Characterization of lactose transport in Kluyveromyces lactis . Journal of Bacteriology 154: 1245–1251.65. Dickson RC, Dickson LR, Markin JS. 1979. Purification and properties of an inducible β-galactosidase isolated from the yeast
Kluyveromyces lactis . Journal of Bacteriology 137: 51–61.66. Dickson RC, Markin JS. 1980. Physiological studies of β-galactosidase induction in Kluyveromyces lactis . Journal of Bacteriology
142: 777–785.67. Dienert [F]. 1899. Sur la fermentation du galactose. Comptes Rendus Hebdomadaires des Seances de l’Academie des Sciences, Paris
128: 569–571.68. Dienert [F]. 1899. Sur la fermentation du galactose. Comptes Rendus Hebdomadaires des Seances de l’Academie des Sciences, Paris
128: 617–618.69. Dienert F. 1900. Sur la fermentation du galactose et sur l’accoutumance des levures a ce sucre. Annales de l’Institut Pasteur 14:
139–189.70. Douglas HC, Condie F. 1954. The genetic control of galactose utilization in Saccharomyces . Journal of Bacteriology 68: 662–670.71. Douglas HC, Hawthorne DC. 1964. Enzymatic expression and genetic linkage of genes controlling galactose utilization in
Saccharomyces . Genetics 49: 837–844.72. Douglas HC, Hawthorne DC. 1966. Regulation of genes controlling synthesis of the galactose pathway enzymes in yeast. Genetics
54: 911–916.73. Douglas HC, Hawthorne DC. 1972. Uninducible mutants in the gal i locus of Saccharomyces cerevisiae. Journal of Bacteriology
109: 1139–1143.74. Douglas HC, Pelroy G. 1963. A gene controlling inducibility of the galactose pathway in Saccharomyces . Biochimica et Biophysica
Acta 68: 155–156.75. Dubourg E. 1899. De la fermentation des saccharides. Comptes Rendus Hebdomadaires des Seances de l’Academie des Sciences,
Paris 128: 440–442.76. Duerksen JD, Halvorson H. 1958. Purification and properties of an inducible β-glucosidase of yeast. Journal of Biological Chemistry
233: 1113–1120.77. Duerksen JD, Halvorson H. 1959. The specificity of induction of β-glucosidase in Saccharomyces cerevisiae. Biochimica et
Biophysica Acta 36: 47–55.78. Eddy AA. 1982. Mechanisms of solute transport in selected eukaryotic microorganisms. Advances in Microbial Physiology 23:
1–78.
Copyright 2004 John Wiley & Sons, Ltd. Yeast 2004; 21: 703–746.

736 J. A. Barnett
79. Emich F. 1910. Uber Mikrochemie, mit besonderer Berucksichtigung der Arbeiten von H. Behrens. Berichte der DeutschenChemischen Gesellschaft 43: 10–45.
80. Entian K-D. 1986. Glucose repression: a complex regulatory system in yeast. Microbiological Sciences 3: 366–371.81. Entian K-D, Barnett JA. 1992. Regulation of sugar utilization by Saccharomyces cerevisiae. Trends in Biochemical Sciences 17:
506–510.82. Entian K-D, Schuller H-J. 1997. Genetics of di- and trisaccharide utilization. In Yeast Sugar Metabolism, Zimmermann FK,
Entian K-D (eds). Technomic: Lancaster, PA; 225–233.83. Entian K-D, Schuller H-J. 1997. Glucose repression (carbon catabolite repression) in yeast. In Yeast Sugar Metabolism,
Zimmermann FK, Entian K-D (eds). Technomic: Lancaster, PA; 409–434.84. Entian K-D, Zimmermann FK. 1980. Glycolytic enzymes and intermediates in carbon catabolite repression mutants of
Saccharomyces cerevisiae. Molecular and General Genetics 177: 345–350.85. Epps HMR, Gale EF. 1942. The influence of the presence of glucose during growth on the enzymic activities of Escherichia coli :
comparison of the effect with that produced by fermentation acids. Biochemical Journal 36: 619–623.86. Eraso P, Gancedo JM. 1984. Catabolite repression in yeasts is not associated with low levels of cAMP. European Journal of
Biochemistry 141: 195–198.87. Fenner FJ. 1987. Frank Macfarlane Burnet. Biographical Memoirs of Fellows of the Royal Society 33: 101–162.88. Ferguson AR, Sims AP. 1971. Inactivation in vivo of glutamine synthetase and NAD-specific glutamate dehydrogenase: its role in
the regulation of glutamine synthesis in yeasts. Journal of General Microbiology 69: 423–427.89. Flick JS, Johnston M. 1991. GRR1 of Saccharomyces cerevisiae is required for glucose repression and encodes a protein with
leucine-rich repeats. Molecular and Cellular Biology 11: 5101–5112.90. Gancedo JM. 1992. Carbon catabolite repression in yeast. European Journal of Biochemistry 206: 297–313.91. Gartler S, Stadler D. 1990. Herschel L. Roman (1914–1989). Genetics 126: 1–3.92. Gascon S, Ottolenghi P. 1967. Invertase isozymes and their localization in yeast. Comptes Rendus des Travaux du Laboratoire
Carlsberg 36: 85–93.93. Gascon S, Ottolenghi P. 1972. Influence of glucose concentration of the medium on the invertase content of a strain of
Saccharomyces bearing the SUC2 gene. Comptes Rendus des Travaux du Laboratoire Carlsberg 39: 15–24.94. Gibson CS, Hilditch TP. 1948. Edward Frankland Armstrong. Obituary Notices of Fellows of the Royal Society 5: 619–633.95. Gilliland RB. 1949. A yeast hybrid heterozygotic in four fermentation characters. Comptes Rendus des Travaux du Laboratoire
Carlsberg, Serie Physiologique 24: 347–356.96. Goldenthal MJ, Cohen JD, Marmur J. 1983. Isolation and characterization of a maltose transport mutant in the yeast Saccharomyces
cerevisiae. Current Genetics 7: 195–199.97. Gorman J, Halvorson H[O]. 1966. Isomaltase from yeast. Methods in Enzymology 8: 562–565.98. Gorr G, Wagner J. 1933. Uber das Amidspaltungsvermogen der Torula utilis, eine Untersuchung uber die Abhangigkeit pflanzlicher
Enzymausbildung von der Stickstoffernahrung. Biochemische Zeitschrift 266: 96–101.99. Gottschalk A. 1950. α-D-Glucosidases. In The Enzymes. Chemistry and Mechanism of Action, vol 1, Sumner JB, Myrback K (eds).
Academic Press: New York; 551–582.100. Gottschalk A. 1950. Principles underlying enzyme specificity in the domain of carbohydrates. Advances in Carbohydrate Chemistry
5: 49–78.101. Grant GA. 1935. The metabolism of galactose. I. Phosphorylation during galactose fermentation and its relation to the
interconversion of hexoses. Biochemical Journal 29: 1661–1676.102. Griggs DW, Johnston M. 1991. Regulated expression of the GAL4 activator gene in yeast provides a sensitive genetic switch for
glucose repression. Proceedings of the National Academy of Sciences of the USA 88: 8597–8601.103. Groß K. 1933. Zum Gedachtis an Ernst Weinland. Sitzungsberichte der Physikalisch-Medizinischen Sozietat zu Erlangen 63/64:
357–370.104. Grossmann MK, Zimmermann FK. 1979. The structural genes of internal invertases in Saccharomyces cerevisiae. Molecular and
General Genetics 175: 223–229.105. Guilliermond A. 1920. The Yeasts. Wiley: New York.106. Halvorson HO. 1960. The induced synthesis of proteins. Advances in Enzymology and Related Subjects of Biochemistry 22: 99–156.107. Halvorson H[O]. 1966. α-Glucosidase from yeast. Methods in Enzymology 8: 559–562.108. Halvorson H[O], Ellias L. 1958. The purification and properties of an α-glucosidase of Saccharomyces italicus Y1225. Biochimica
et Biophysica Acta 30: 28–40.109. Halvorson HO, Okada H, Gorman J. 1964. The role of an alpha-methylglucoside permease in the inducted synthesis of isomaltase
in yeast. In The Cellular Functions of Membrane Transport, Hoffman J (ed.). Prentice Hall: Englewood Cliffs, NJ; 171–191.110. Halvorson HO, Winderman S, Gorman J. 1963. Comparison of the α-glucosidases of Saccharomyces produced in response to five
non-allelic maltose genes. Biochimica et Biophysica Acta 67: 42–53.111. Harden A, Norris RV. 1910. The fermentation of galactose by yeast and yeast-juice. Proceedings of the Royal Society of London B
82: 645–649.112. Hardie DG, Carling D, Carlson M. 1998. The AMP-activated/SNF1 protein kinase subfamily: metabolic sensors of the eukaryotic
cell? Annual Review of Biochemistry 67: 821–855.113. Hassid WZ, Ballou CE. 1957. Oligosaccharides. In The Carbohydrates. Chemistry, Biochemistry, Physiology, Pigman W (ed.).
Academic Press: New York; 478–535.
Copyright 2004 John Wiley & Sons, Ltd. Yeast 2004; 21: 703–746.

History of research on yeasts 7 737
114. Hawthorne DC. 1955. The use of linear asci for chromosome mapping in Saccharomyces . Genetics 40: 511–518.115. Hawthorne DC. 1956. The genetics of galactose fermentation in Saccharomyces hybrids. Comptes Rendus des Travaux du
Laboratoire Carlsberg Serie Physiologique 26: 149–16.116. Heinrich MR. 1964. The purification and properties of yeast galactokinase. Journal of Biological Chemistry 239: 50–53.117. Helferich B, Turk D. 1956. Synthese einiger β-D-Thiogalactoside. Chemische Berichte 89: 2215–2219.118. Herzenberg LA. 1959. Studies on the induction of β-galactosidase in a cryptic strain of Escherichia coli . Biochimica et Biophysica
Acta 31: 525–538.119. Hogness DS, Cohn M, Monod J. 1955. Studies on the induced synthesis of β-galactosidase in Escherichia coli : the kinetics and
mechanism of sulfur incorporation. Biochimica et Biophysica Acta 16: 99–116.120. Hopper JE, Broach JR, Rowe LB. 1978. Regulation of the galactose pathway in Saccharomyces cerevisiae: induction of uridyl
transferase mRNA and dependency on GAL4 gene function. Proceedings of the National Academy of Sciences of the USA 75:2878–2882.
121. Hovland P, Flick J, Johnston M, Sclafani RA. 1989. Galactose as a gratuitous inducer of GAL gene expression in yeasts growingon glucose. Gene 83: 57–64.
122. Igarashi M, Segawa T, Nogi Y, Suzuki Y, Fukasawa T. 1987. Autogenous regulation of the Saccharomyces cerevisiae regulatorygene GAL80 . Molecular and General Genetics 207: 273–279.
123. Jacob F, Girard M. 1998. Andre Michel Lwoff. Biographical Memoirs of Fellows of the Royal Society 44: 255–263.124. Jacob F, Monod J. 1959. Genes de structure et genes de regulation dans la biosynthese des proteines. Comptes Rendus
Hebdomadaires des Seances de l’Academie des Sciences, Paris 249: 1282–1284.125. Jacob F, Monod J. 1961. Genetic regulatory mechanisms in the synthesis of proteins. Journal of Molecular Biology 3: 318–356.126. Jacob F, Monod J. 1961. On the regulation of gene activity. Cold Spring Harbor Symposia on Quantitative Biology 26: 193–210.127. Jacob F, Perrin D, Sanchez C, Monod J. 1960. L’operon: groupes de genes a expression coordonnee par un operateur. Comptes
Rendus Hebdomadaires des Seances de l’Academie des Sciences, Paris 250: 1727–1729.128. Johnston JHS. 1953. Dr. Arthur Slator. Chemistry and Industry: 943–944.129. Johnston M. 1987. A model fungal gene regulatory mechanism: the GAL genes of Saccharomyces cerevisiae. Microbiological
Reviews 51: 458–476.130. Johnston M, Carlson M. 1992. Regulation of carbon and phosphate utilization. In The Molecular and Cellular Biology of the Yeast
Saccharomyces, Jones EW, Pringle JR, Broach JR (eds). Cold Spring Harbor Laboratory Press: New York; 193–281.131. Kalckar HM, Braganca B, Munch-Petersen A. 1953. Uridyl transferase and the formation of uridine diphosphogalactose. Nature
172: 1038.132. Karstrom H. 1938. Enzymatische Adaptation bei Mikroorganismen. Ergebnisse der Enzymforschung 7: 350–376.133. Katz J. 1898. Die regulatorische Bildung von Diastase durch Pilze. Jahrbucher fur Wissenschaftliche Botanik 31: 599–618.134. Kepes A, Monod J. 1957. Etude du fonctionnement de la galactoside-permease d’Escherichia coli . Comptes Rendus Hebdomadaires
des Seances de l’Academie des Sciences, Paris 244: 809–811.135. Kew OM, Douglas HC. 1976. Genetic co-regulation of galactose and melibiose utilization in Saccharomyces . Journal of
Bacteriology 125: 33–41.136. Khan NA, Eaton NR. 1967. Purification and characterization of maltase and α-methyl glucosidase from yeast. Biochimica et
Biophysica Acta 146: 173–180.137. Khan NA, Haynes RH. 1972. Genetic redundancy in yeast: non-identical products in a polymeric gene system. Molecular and
General Genetics 118: 279–285.138. Kohlhaw G, Dragert W, Holzer H. 1965. Parallel-Repression der Synthese von Glutamin-Synthetase und DPN-abhangiger Glutamat-
Dehydrogenase in Hefe. Biochemische Zeitschrift 341: 224–238.139. Kosterlitz HW. 1943. The fermentation of galactose and galactose-1-phosphate. Biochemical Journal 37: 322–326.140. Kratky, Z, Biely P. 1980. Inducible β-xyloside permease as a constituent of the xylan-degrading enzyme system of the yeast
Cryptococcus albidus . European Journal of Biochemistry 112: 367–373.141. Kudryavtsev VI. 1954. Yeast Systematics. Akademii Nauk SSSR: Moscow [kUDRQWCEW wi. 1954. sucmeMamuKa dROVVEu
aKADEMII nAUK].142. Kuger P, Godecke A, Breunig KD. 1990. A mutation in the Zn-finger of the GAL4 homolog LAC9 results in glucose repression
of its target genes. Nucleic Acids Research 18: 745–751.143. Kurtzman CP, Fell JW. 1998. The Yeasts, a Taxonomic Study, 4th edn. Elsevier: Amsterdam.144. Lai H-Y, Axelrod B. 1975. The specificity of the synthetic reaction of two yeast α-glucosidases. Biochimica et Biophysica Acta
391: 121–128.145. Lamphier MS, Ptashne M. 1992. Multiple mechanisms mediate glucose repression of the yeast GAL1 gene. Proceedings of the
National Academy of Sciences of the USA 89: 5922–5926.146. Larner J. 1960. Other glucosidases. In The Enzymes, 2nd edn, vol 4, Boyer PD, Lardy H, Myrback K (eds). Academic Press: New
York; 369–378.147. Laughon A, Gesteland RF. 1984. Primary structure of the Saccharomyces cerevisiae GAL4 gene. Molecular and Cellular Biology
4: 260–267.148. Lederberg J. 1950. The beta-D-galactosidase of Escherichia coli, strain K-12. Journal of Bacteriology 60: 381–392.149. Lederberg J, Tatum EL. 1946. Gene recombination in Escherichia coli . Nature 158: 558.
Copyright 2004 John Wiley & Sons, Ltd. Yeast 2004; 21: 703–746.

738 J. A. Barnett
150. LeFevre PG. 1954. The evidence for active transport of monosaccharides across the red cell membrane. Symposium of the Societyfor Experimental Biology 8: 118–135.
151. Legler G, Lotz W. 1973. Funktionelle Gruppen am aktiven Zentrum einer α-Glucosidase aus Saccharomyces cerevisiae. Hoppe-Seyler’s Zeitschrift fur Physiologische Chemie 354: 243–254.
152. Leibowitz J, Hestrin S. 1939. The direct fermentation of maltose by yeast. Enzymologia 6: 15–26.153. Leloir LF. 1951. The enzymatic transformation of uridine diphosphate glucose into a galactose derivative. Archives of Biochemistry
and Biophysics 33: 186–190.154. Leloir LF. 1953. Enzymic isomerization and related processes. Advances in Enzymology and Related Subjects of Biochemistry 14:
193–218.155. Leloir LF, Paladini AC. 1983. The discovery of sugar nucleotides. In Comprehensive Biochemistry, vol 35, Neuberger A, van
Deenen LLM, Semenza G (eds). Elsevier: Amsterdam; 25–42.156. Le Strat A. 1948. Eloge funebre prononce le 9 avril 1948, aux obseques de M. F. Dienert. La Technique Sanitaire et Municipale 3:
(Nos. 3–4, March–April).157. Le Tourneur S. 1967. Dienert (Frederic-Vincent). Dictionnaire de Biographie Francaise 11: 306.158. Leupold U, Hottinguer H. 1954. Some data on segregation in Saccharomyces . Heredity 8: 243–258.159. Lindegren CC. 1955. The identification of fermentative phenotypes in Saccharomyces . Journal of Bacteriology 70: 127.160. Lindegren CC, Lindegren G. 1947. Mendelian inheritance of genes affecting vitamin-synthesizing ability in Saccharomyces . Annals
of the Missouri Botanical Garden 34: 95–99.161. Lodder J, Kreger-van Rij NJW. 1952. The Yeasts, a Taxonomic Study. North-Holland: Amsterdam.162. Lovgren T, Siro M-R. 1972. Studies on baker’s yeast. VII. Isolation of α-glucosidases from baker’s yeast. Acta Academiae Aboensis
Series B 32: 1–10.163. Lwoff A. 1977. Jacques Lucien Monod. Biographical Memoirs of Fellows of the Royal Society 23: 385–412.164. Lwoff A, Ullmann A (eds). 1979. Origins of Molecular Biology. A Tribute to Jacques Monod. Academic Press: New York.165. MacQuillan AM, Winderman S, Halvorson HO. 1960. The control of enzyme synthesis by glucose and the repressor hypothesis.
Biochemical and Biophysical Research Communications 3: 77–80.166. Magasanik B. 1961. Catabolite repression. Cold Spring Harbor Symposia on Quantitative Biology 26: 249–256.167. Magasanik B. 1992. Regulation of nitrogen utilization. In The Molecular and Cellular Biology of the Yeast Saccharomyces,
Jones EW, Pringle JR, Broach JR (eds). Cold Spring Harbor Laboratory Press: New York; 283–317.168. Magasanik B. 1994. A charmed life. Annual Review of Microbiology 48: 1–24.169. Martin CJ. 1904. Emile Duclaux. Nature 70: 34–35.170. Matsumoto K, Adachi Y, Toh-e A, Oshima Y. 1980. Function of positive regulatory gene gal4 in the synthesis of galactose pathway
enzymes in Saccharomyces cerevisiae: evidence that the GAL81 region codes for part of the gal4 protein. Journal of Bacteriology141: 508–527.
171. Matsumoto K, Toh-e A, Oshima Y. 1981. Isolation and characterization of dominant mutations resistant to carbon cataboliterepression of galactokinase synthesis in Saccharomyces cerevisiae. Molecular and Cellular Biology 1: 83–93.
172. Matsumoto K, Yoshimatsu T, Oshima Y. 1983. Recessive mutations conferring resistance to carbon catabolite repression ofgalactokinase synthesis in Saccharomyces cerevisiae. Journal of Bacteriology 153: 1405–1414.
173. Matsusaka K, Chiba S, Shimomura T. 1977. Purification and substrate specificity of brewer’s yeast α-glucosidase. Agricultural andBiological Chemistry 41: 1917–1923.
174. Maxwell ES, de Robichon-Szulmajster H. 1960. Purification of uridine diphosphate galactose-4-epimerase from yeast, and theidentification of protein-bound diphosphopyridine nucleotide. Journal of Biological Chemistry 235: 308–312.
175. McCann AK, Barnett JA. 1986. The utilization of starch by yeasts. Yeast 2: 109–115.176. Meyer J, Matile P. 1974. Regulation of isoenzymes and secretion of invertase in bakers’ yeast. Biochemie und Physiologie der
Pflanzen 166: 377–385.177. Meyer J, Walker-Jonah A, Hollenberg CP. 1990. GAL1 of Kluyveromyces lactis is required for the induction of the Leloir enzymes
and can complement the gal3 long term adaptation-phenotype in Saccharomyces cerevisiae. Yeast 6: S591.178. Meyer J, Walker-Jonah A, Hollenberg CP. 1991. Galactokinase encoded by GAL1 is a bifunctional protein required for the induction
of the GAL genes in Kluyveromyces lactis and able to suppress the gal3 phenotype in Saccharomyces cerevisiae. Molecular andCellular Biology 11: 5454–5461.
179. Monod J. 1942. Recherches sur la Croissance des Cultures Bacteriennes. These Doctorat es Sciences, Paris. Hermann: Paris.180. Monod J. 1947. The phenomenon of enzymatic adaptation and its bearing on problems of genetics and cellular differentiation.
Growth 11: 223–289.181. Monod J. 1949. The growth of bacterial cultures. Annual Review of Microbiology 3: 371–394.182. Monod J. 1956. Remarks on the mechanism of enzyme induction. In Enzymes: Units of Biological Structure and Function,
Gaebler OH (ed.). Academic Press: New York; 7–28.183. Monod J. 1959. Biosynthese eines Enzyms. Information, Induktion, Repression. Angewandte Chemie 71: 685–691.184. Monod J. 1965. From enzymatic adaptation to allosteric transitions. In Nobel Lectures Physiology or Medicine 1963–1970. Elsevier:
Amsterdam [published 1972].185. Monod J, Audureau A. 1946. Mutation et adaptation enzymatique chez Escherichia coli-mutabile. Annales de l’Institut Pasteur 72:
868–878.186. Monod J, Changeux J-P, Jacob F. 1963. Allosteric proteins and cellular control systems. Journal of Molecular Biology 6: 306–329.
Copyright 2004 John Wiley & Sons, Ltd. Yeast 2004; 21: 703–746.

History of research on yeasts 7 739
187. Monod J, Cohen-Bazire G. 1953. L’effet inhibiteur specifique des β-galactosides dans la biosynthese ‘constitutive’ de la β-galactosidase chez E. coli. Comptes Rendus Hebdomadaires des Seances de l’Academie des Sciences, Paris 236: 417–419.
188. Monod J, Cohen-Bazire G, Cohn M. 1951. Sur la biosynthese de la β-galactosidase (lactase) chez Escherichia coli. La specificitede l’induction. Biochimica et Biophysica Acta 7: 585–599.
189. Monod J, Cohn M. 1952. La biosynthese induite des enzymes (adaption enzymatique). Advances in Enzymology and RelatedSubjects of Biochemistry 13: 67–119.
190. Monod J, Jacob F. 1961. General conclusions: teleonomic mechanisms in cellular metabolism, growth, and differentiation. ColdSpring Harbor Symposia on Quantitative Biology 26: 389–401.
191. Morris JG. 1983. Roger Yate Stanier, 1916–1982. Journal of General Microbiology 129: 255–261.192. Munch-Petersen A, Kalckar HM, Cutolo E, Smith EEB. 1953. Enzymic production of uridine triphosphate: uridine
diphosphoglucose pyrophosphorolysis. Nature 172: 1036–1037.193. Mundkur BD, Lindegren CC. 1949. An analysis of the phenomenon of long term adaptation to galactose by Saccharomyces .
American Journal of Botany 36: 722–727.194. Muth F. 1925. Julius Wortmann. Berichte der Deutschen Botanischen Gesellschaft 43: 112–142.195. Myrback K. 1960. Invertases. In The Enzymes, 2nd edn, vol 4, Boyer PD, Lardy H, Myrback K (eds). Academic Press: New York;
379–396.196. Myrback K, Ortenblad B. 1936. Trehalose und Hefe. I. Biochemische Zeitschrift 288: 329–337.197. Najjar VA. 1948. The isolation and properties of phosphoglucomutase. Journal of Biological Chemistry 175: 281–290.198. Najjar VA. 1962. Phosphoglucomutase. In The Enzymes, 2nd edn, vol 6, Boyer PD, Lardy H, Myrback K (eds). Academic Press:
New York; 161–178.199. Naumov GI. 1976. Comparative genetics of yeast XVI. Genes for maltose fermentation in Saccharomyces carlsbergensis N.C.Y.C.
74. Genetika 12: 87–100 [nAUMOW gi. 1976. sRAWNITEL_NAQ GENETIKA DROVVEJ XVI. gOOB]ENI GENY SBRAVIWANIQ MAL_TOZY UDROVVEJ. Saccharomyces carlsbergensis N.C.Y.C. 74. gENEmuka 12: 87–100].
200. Needleman RB, Federoff HJ, Eccleshall TR, Buchferer B, Marmur J. 1978. Purification and characterization of an α-glucosidasefrom Saccharomyces carlsbergensis . Biochemistry 17: 4657–4661.
201. Needleman RB, Kaback DB, Dubin RA, et al. 1984. MAL6 of Saccharomyces: a complex genetic locus containing three genesrequired for maltose fermentation. Proceedings of the National Academy of Sciences of the USA 81: 2811–2815.
202. Nehlin JO, Carlberg M, Ronne H. 1989. Yeast galactose permease is related to yeast and mammalian glucose transporters. Gene85: 313–319.
203. Neigeborn L, Carlson M. 1987. Mutations causing constitutive invertase synthesis in yeast: genetic interactions with snf mutations.Genetics 115: 247–253.
204. Neuberg C, Hofmann E. 1932. Neue Beobachtungen uber β-glucosidase. Biochemische Zeitschrift 256: 450–461.205. Neuberg C, v[on] Euler H. 1931. Zur Nomenklatur der an der alkoholischen Garung beteiligten Katalysatoren. Biochemische
Zeitschrift 240: 245.206. Niederacher D, Entian K-D. 1991. Characterization of Hex2 protein, a negative regulatory element necessary for glucose repression
in yeast. European Journal of Biochemistry 200: 311–319.207. Nobel Lectures, Physiology or Medicine 1963–1970 [published 1972]. Elsevier: Amsterdam; 172.208. Nossal GJV. 1985. Sir Frank MacFarlane Burnet (1899–1985). Nature 317: 108.209. Ochoa S. 1990. Luis Federico Leloir. Biographical Memoirs of Fellows of the Royal Society 35: 203–208.210. Oh D, Hopper JE. 1990. Transcription of a yeast phosphoglucomutase isozyme gene is galactose inducible and glucose repressible.
Molecular and Cellular Biology 10: 1415–1422.211. Ottolenghi P. 1971. A comparison of five genetically distinct invertases from Saccharomyces. Some enzymatic characteristics.
European Journal of Biochemistry 18: 544–552.212. Oxford English Dictionary, 2nd edn, vol 1. 1989. 139.213. Pardee AB, Jacob F, Monod J. 1959. The genetic control and cytoplasmic expression of ‘inducibility’ in the synthesis of β-
galactosidase by E. coli. Journal of Molecular Biology 1: 165–178.214. Pasteur [L]. 1856. Note sur le sucre de lait. Comptes Rendus Hebdomadaires des Seances de l’Academie des Sciences, Paris 42:
347–351.215. Pasteur [L]. 1861. Influence de l’oxygene sur le developpement de la levure et la fermentation alcoolique. Bulletin de la Societe
de Paris (Resume de Seance du 28 juin 1861); 79–80.216. Pasteur L. 1861. Experiences et vues nouvelles sur la nature des fermentations. Comptes Rendus Hebdomadaires des Seances de
l’Academie des Sciences, Paris 53: 1260–1264.217. Peck HD, Gest H. 1957. Formic dehydrogenase and the hydrogenlyase enzyme complex in coli–aerogenes bacteria. Journal of
Bacteriology 73: 706–721.218. Penfold WJ. 1911. Studies in bacterial variation. With special reference to the chemical functions of the members of the typhoid–coli
group. Journal of Hygiene 11: 30–67.219. Polakis ES, Bartley W. 1965. Changes in the enzyme activities of Saccharomyces cerevisiae during aerobic growth on different
carbon sources. Biochemical Journal 97: 284–297.220. Pollock MR. 1959. Induced formation of enzymes. In The Enzymes, 2nd edn, vol 1, Boyer PD, Lardy H, Myrback K (eds). Academic
Press: New York; 619–680.
Copyright 2004 John Wiley & Sons, Ltd. Yeast 2004; 21: 703–746.

740 J. A. Barnett
221. Pollock M[R]. 1979. An exciting but exasperating personality. In Origins of Molecular Biology. A Tribute to Jacques Monod,Lwoff A, Ullmann A (eds). Academic Press: New York; 61–73.
222. Pregl F. 1917. Die Quantitative Organische Mikroanalyse. Springer: Berlin.223. Quayle JR. 1966. Formate dehydrogenase. Methods in Enzymology 9: 360–364.224. Ray WJ, Peck EJ. 1972. Phosphomutases. In The Enzymes, 3rd edn, vol 6, Boyer PD (ed.). Academic Press: New York; 407–477.225. Ray WJ, Roscelli GA. 1964. A kinetic study of the phosphoglucomutase pathway. Journal of Biological Chemistry 239: 1228–1236.226. Rickenberg HV, Cohen GN, Buttin G, Monod J. 1956. La galactoside-permease d’Escherichia coli . Annales de l’Institut Pasteur
91: 829–857.227. Riley MI, Dickson RC. 1984. Genetic and biochemical characterization of the galactose gene cluster in Kluyveromyces lactis .
Journal of Bacteriology 158: 705–712.228. Riley MI, Hopper JE, Johnston SA, Dickson RC. 1987. GAL4 of Saccharomyces cerevisiae activates the lactose–galactose regulon
of Kluyveromyces lactis and creates a new phenotype: glucose repression of the regulon. Molecular and Cellular Biology 7: 780–786.229. Roman H. 1986. The early days of yeast genetics: a personal narrative. Annual Review of Genetics 20: 1–12.230. Roman H, Hawthorne DC, Douglas HC. 1951. Polyploidy in yeast and its bearing on the occurrence of irregular genetic ratios.
Proceedings of the National Academy of Sciences of the USA 37: 79–84.231. Roman H, Phillips MM, Sands SM. 1955. Studies of polyploid Saccharomyces. I. Tetraploid segregation. Genetics 40: 546–561.232. Ronneberg CE. 1970. Armstrong, Edward Frankland. In Dictionary of Scientific Biography, vol 1, Gillispie CC (ed.). Scribner:
New York; 286–287.233. Rothstein A. 1954. Enzyme systems of the cell surface involved in the uptake of sugars by yeast. Symposium of the Society for
Experimental Biology 8: 165–201.234. Salmeron JM, Johnston SA. 1986. Analysis of the Kluyveromyces lactis positive regulatory gene LAC9 reveals functional homology
to, but sequence divergence from, the Saccharomyces cerevisiae GAL4 gene. Nucleic Acids Research 14: 7767–7781.235. Sarokin L, Carlson M. 1985. Upstream region of the SUC2 gene confers regulated expression to a heterologous gene in
Saccharomyces cerevisiae. Molecular and Cellular Biology 5: 2521–2526.236. Schuller H-J, Entian K-D. 1991. Extragenic suppressors of yeast glucose derepression mutants leading to constitutive synthesis of
several glucose-repressible enzymes. Journal of Bacteriology 173: 2045–2052.237. Schultz AS, Atkin L. 1939. Fermentation of maltose. Journal of the American Chemical Society 61: 291–294.238. Schultz AS, Atkin L, Frey CN. 1940. Influence of oxygen on the fermentation of maltose and galactose. Journal of the American
Chemical Society 61: 2271–2272.239. Segal S. 1998. Galactosaemia today: the enigma and the challenge. Journal of Inherited Metabolic Disease 21: 455–471.240. Segal S, Berry GT. 1995. Disorders of galactose metabolism. In The Metabolic and Molecular Bases of Inherited Disease,
Scriver CR, Beaudet AL, Sly WS, Valle D (eds). McGraw-Hill: New York; 967–1000.241. Seidman M, Link KP. 1950. o-Nitrophenyl-β-D-galactopyranoside and its tetraacetate. Journal of the American Chemical Society
72: 4324.242. Slator A. 1908. Studies in fermentation. Part II. The mechanism of alcoholic fermentation. Journal of the Chemical Society 93:
217–242.243. Snyder HE, Phaff HJ. 1960. Studies on a beta-fructosidase (inulinase) produced by Saccharomyces fragilis . Antonie van
Leeuwenhoek 26: 433–452.244. Sohngen NL, Coolhaas C. 1925. Die Galaktosegarung durch Saccharomyces cerevisiae. Centralblatt fur Bakteriologie,
Parasitenkunde und Infektionskrankheiten, Zweite Abteilung, 66: 5–11.245. Sowden JC. 1957. Occurrence, properties, and synthesis of the monosaccharides. In The Carbohydrates. Chemistry, Biochemistry,
Physiology, Pigman W (ed.). Academic Press: New York; 76–137.246. Spiegelman S. 1945. The physiology and genetic significance of enzymatic adaptation. Annals of the Missouri Botanical Garden
32: 139–163.247. Spiegelman S. 1945. The effect of anaerobiosis on adaptation to galactose fermentation by yeast cells. Journal of Cellular and
Comparative Physiology 25: 121–131.248. Spiegelman S. 1946. Nuclear and cytoplasmic factors controlling enzymatic constitution. Cold Spring Harbor Symposia on
Quantitative Biology 11: 256–277.249. Spiegelman S. 1950. Modern aspects of enzymatic adaptation. In The Enzymes. Chemistry and Mechanism of Action, vol 1,
Sumner JB, Myrback K (eds). Academic Press: New York; 267–306.250. Spiegelman S, DeLorenzo WF, Campbell AM. 1951. A single-cell analysis of the transmission of enzyme-forming capacity in
yeast. Proceedings of the National Academy of Sciences of the USA 37: 513–524.251. Spiegelman S, Dunn R. 1947. Interactions between enzyme-forming systems during adaptation. Journal of General Physiology 31:
153–173.252. Spiegelman S, Halvorson HO. 1953. The nature of the precursor in the induced synthesis of enzymes. Symposium of the Society
for General Microbiology 3: 98–131.253. Spiegelman S, Lindegren CC. 1944. A comparison of the kinetics of enzymatic adaptation in genetically homogeneous and
heterogeneous populations of yeast. Annals of the Missouri Botanical Garden 31: 219–233.254. Spiegelman S, Lindegren CC, Hedgecock L. 1944. Mechanisms of enzymatic adaptation in genetically controlled yeast populations.
Proceedings of the National Academy of Sciences of the USA 30: 13–23.
Copyright 2004 John Wiley & Sons, Ltd. Yeast 2004; 21: 703–746.

History of research on yeasts 7 741
255. Spiegelman S, Lindegren CC, Lindegren G. 1945. Maintenance and increase of a genetic character by a substrate-cytoplasmicinteraction in the absence of the specific gene. Proceedings of the National Academy of Sciences of the USA 31: 95–102.
256. Spiegelman S, Reiner JM, Cohnberg R. 1947. The relation of enzymatic adaptation to the metabolism of endogenous and exogenoussubstrates. Journal of General Physiology 31: 27–49.
257. Spiegelman S, Sussman RR, Pinska E. 1950. On the cytoplasmic nature of ‘long-term adaptation’ in yeast. Proceedings of theNational Academy of Sciences of the USA 36: 591–606.
258. Spiegelman S, Sussman M, Taylor B. 1950. Isolation and characterization of two adaptive enzymes formed by yeast in responseto maltose. Federation Proceedings 9: 120.
259. Stanier RY. 1947. Simultaneous adaptation: a new technique for the study of metabolic pathways. Journal of Bacteriology 54:339–348.
260. Stanier RY. 1951. Enzymatic adaptation in bacteria. Annual Review of Microbiology 5: 35–56.261. Stanier RY. 1980. The journey, not the arrival, matters. Annual Review of Microbiology 34: 1–48.262. Stelling-Dekker NM. 1931. Die sporogenen Hefen. Verhandelingen der Koninklijke Akademie van Wetenschappen te Amsterdam,
Tweede Sectie, 28: 1–547.263. Stephenson M, Gale EF. 1937. The adaptability of glucozymase and galactozymase in Bacterium coli . Biochemical Journal 31:
1311–1315.264. Stephenson M, Stickland LH. 1931. Hydrogenase: a bacterial enzyme activating molecular hydrogen. Biochemical Journal 25:
205–214.265. Stephenson M, Stickland LH. 1933. Hydrogenlyases. III. Further experiments on the formation of formic hydrogenlyase by Bact.
coli. Biochemical Journal 27: 1528–1532.266. Stephenson M, Yudkin J. 1936. Galactozymase considered as an adaptive enzyme. Biochemical Journal 30: 506–514.267. St John TP, Davis RW. 1979. Isolation of galactose-inducible DNA sequences from Saccharomyces cerevisiae by differential plaque
filter hybridization. Cell 16: 443–452.268. St John TP, Davis RW. 1981. The organization and transcription of the galactose gene cluster of Saccharomyces . Journal of
Molecular Biology 152: 285–315.269. Struhl K. 1995. Yeast transcriptional regulatory mechanisms. Annual Review of Genetics 29: 651–674.270. Sutton DD, Lampen JO. 1962. Localization of sucrose and maltose fermenting systems in Saccharomyces cerevisiae. Biochimica
et Biophysica Acta 56: 303–312.271. Terui G, Okada H, Oshima Y. 1959. Studies on the correlation of alpha-glucosidase formation with genotypic composition in
Saccharomyces (I). Technology Reports of the Osaka University 9: 237–259.272. Terui G, Okada H, Oshima Y. 1959. Correlation of α-glucosidase formation and genotypic composition in Saccharomyces I [in
Japanese]. Hakko Kogaku Zasshi 37: 177–189 [Chemical Abstracts 54: 15520d (1959)].273. Treadwell FP, Hall WT. 1942. Analytical Chemistry, 9th edn, vol 2. Wiley: New York.274. Tschopp JF, Emr SD, Field C, Schekman R. 1986. GAL2 codes a membrane-bound subunit of the galactose permease in
Saccharomyces cerevisiae. Journal of Bacteriology 166: 313–318.275. Twort FW. 1907. The fermentation of glucosides by bacteria of the typhoid-coli group and the acquisition of new fermenting
powers by Bacillus dysenteriae and other micro-organisms. Proceedings of the Royal Society of London B 79: 329–336.276. Ullmann A. 1979. Being around. In Origins of Molecular Biology. A Tribute to Jacques Monod, Lwoff A, Ullmann A (eds).
Academic Press: New York; 165–170.277. Virtanen AI. 1948. On the adaptive formation of enzymes by micro-organisms. Svensk Kemisk Tidskrift 60: 23–38.278. [von] Euler H, Cramer H. 1914. Zur Kenntnis der Invertasebildung in Hefe. Biochemische Zeitschrift 58: 467–469.279. v[on] Euler H, Jansson B. 1927. Uber die Anpassung von frischen Kulturhefen an Galaktose. Hoppe-Seyler’s Zeitschrift fur
Physiologische Chemie 169: 226–234.280. [von] Euler H, Johansson D. 1912. Untersuchungen uber die chemische Zusammensetzung und Bildung der Enzyme. IV. Uber die
Anpassung einer Hefe an Galaktose. Hoppe-Seyler’s Zeitschrift fur Physiologische Chemie 78: 246–265.281. [von] Euler H, Johansson D. 1913. Untersuchungen uber die chemische Zusammensetzung und Bildung der Enzyme. VIII.
Mitteilung. Uber die gleichzeitige Veranderung des Gehaltes an Invertase und Garungsenzymen in der lebenden Hefe. Hoppe-Seyler’s Zeitschrift fur Physiologische Chemie 84: 97–108.
282. v[on] Euler H, Laurin I, Pettersson A. 1921. Anpassung einer Oberhefe an das Garsubstrat Galaktose. Biochemische Zeitschrift114: 277–291.
283. [von] Euler H, Meyer H. 1912. Untersuchungen uber die chemische Zusammensetzung und Bildung der Enzyme. V. Mitteilung.Zur Kenntnis der Invertasebildung. Hoppe-Seyler’s Zeitschrift fur Physiologische Chemie 79: 274–300.
284. v[on] Euler H, Nilsson R. 1925. Uber die Galaktosevergarung durch Hefe nach Vorbehandlung mit dieser Zuckerart. Hoppe-Seyler’sZeitschrift fur Physiologische Chemie 143: 89–107.
285. Walden P. 1895. Ueber optisch active Halogenverbindungen. Berichte der Deutschen Chemischen Gesellschaft 28: 1287–1297.286. Webb EC. 1992. Enzyme Nomenclature 1992. Recommendations of the Nomenclature Committee of the International Union of
Biochemistry and Molecular Biology on the Nomenclature and Classification of Enzymes. Academic Press: San Diego, CA.287. Webster B. 1983. Dr. Sol Spiegelman, Microbiologist at Columbia and geneticist, dies. New York Times January 22: 16.288. Weinland E. 1906. Uber das Auftreten von Invertin im Blut. Zeitschrift fur Biologie 47: 279–288.289. Wesolowski-Louvel M, Breunig KD, Fukuhara H. 1996. Kluyveromyces lactis . In Nonconventional Yeasts in Biotechnology, Wolf K
(ed.). Springer: Berlin; 139–201.
Copyright 2004 John Wiley & Sons, Ltd. Yeast 2004; 21: 703–746.

742 J. A. Barnett
290. Who’s Who in America, 57th edn. 2003. Marquis Who’s Who: New Providence, USA.291. Wilkinson JF. 1949. The pathway of the adaptive fermentation of galactose by yeast. Biochemical Journal 44: 460–467.292. Williamson VM, Young ET, Ciriacy M. 1981. Transposable elements associated with constitutive expression of yeast alcohol
dehydrogenase II. Cell 23: 605–614.293. Winge O, Roberts C. 1948. Inheritance of enzymatic characters in yeasts and the phenomenon of long-term adaptation. Comptes
Rendus des Travaux du Laboratoire Carlsberg Serie Physiologique 24: 263–313.294. Winge O, Roberts C. 1950. The polymeric genes for maltose fermentation in yeasts, and their mutability. Comptes Rendus des
Travaux du Laboratoire Carlsberg Serie Physiologique 25: 35–83.295. Winge O, Roberts C. 1952. The relation between the polymeric genes for maltose, raffinose, and sucrose fermentation in yeasts.
Comptes Rendus des Travaux du Laboratoire Carlsberg Serie Physiologique 25: 141–171.296. Winge O, Roberts C. 1956. Complementary action of melibiase and galactozymase on raffinose fermentation. Nature 177: 383–384.297. Winge O, Roberts C. 1957. A genetic analysis of melibiose and raffinose fermentation. Comptes Rendus des Travaux du Laboratoire
Carlsberg Serie Physiologique 25: 419–459.298. Winge O, Roberts C. 1958. Yeast genetics. In The Chemistry and Biology of Yeasts, Cook AH (ed.). Academic Press: New York;
123–156.299. Witt I, Kronau R, Holzer H. 1966. Repression von Alkoholdehydrogenase, Malatdehydrogenase, Isocitratlyase und Malatsynthase
in Hefe durch Glucose. Biochimica et Biophysica Acta 118: 522–537.300. Wortmann J. 1882. Untersuchungen uber das diastatische Ferment der Bakterien. Hoppe-Seyler’s Zeitschrift fur Physiologische
Chemie 6: 287–329.301. Yudkin J. 1938. Enzyme variation in micro-organisms. Biological Reviews 13: 93–106.302. Zachariae W, Kuger P, Breunig KD. 1993. Glucose repression of lactose/galactose metabolism in Kluyveromyces lactis is
determined by the concentration of the transcriptional activator LAC9 (K1GAL4). Nucleic Acids Research 21: 69–77.303. Zimmermann FK, Scheel I. 1977. Mutants of Saccharomyces cerevisiae resistant to carbon catabolite repression. Molecular and
General Genetics 154: 75–82.304. http://www.gs.washington.edu/news/hawthorne/index.htm (2003).305. http://www.nobel.se/chemistry/laureates/1970/leloir-bio.html
Copyright 2004 John Wiley & Sons, Ltd. Yeast 2004; 21: 703–746.
History of research on yeasts 7 743
Appendix. The structures of some sugars and sugar-derivatives
Copyright 2004 John Wiley & Sons, Ltd. Yeast 2004; 21: 703–746.
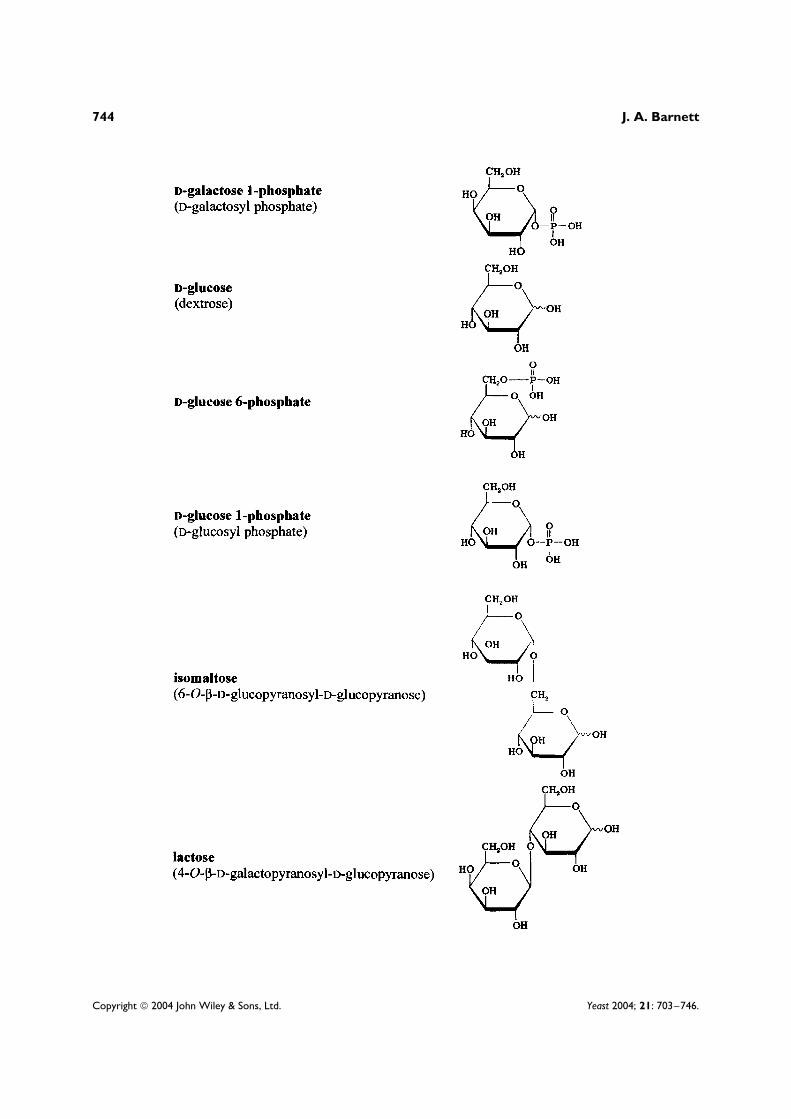
744 J. A. Barnett
Copyright 2004 John Wiley & Sons, Ltd. Yeast 2004; 21: 703–746.
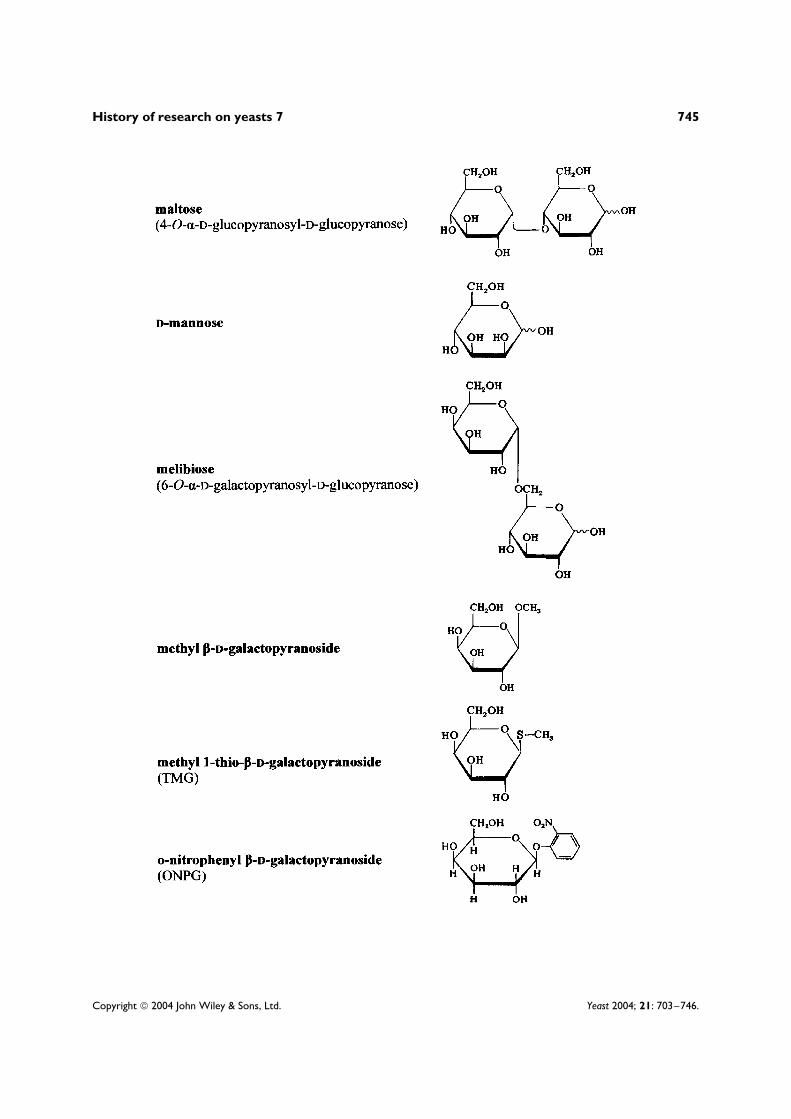
History of research on yeasts 7 745
Copyright 2004 John Wiley & Sons, Ltd. Yeast 2004; 21: 703–746.
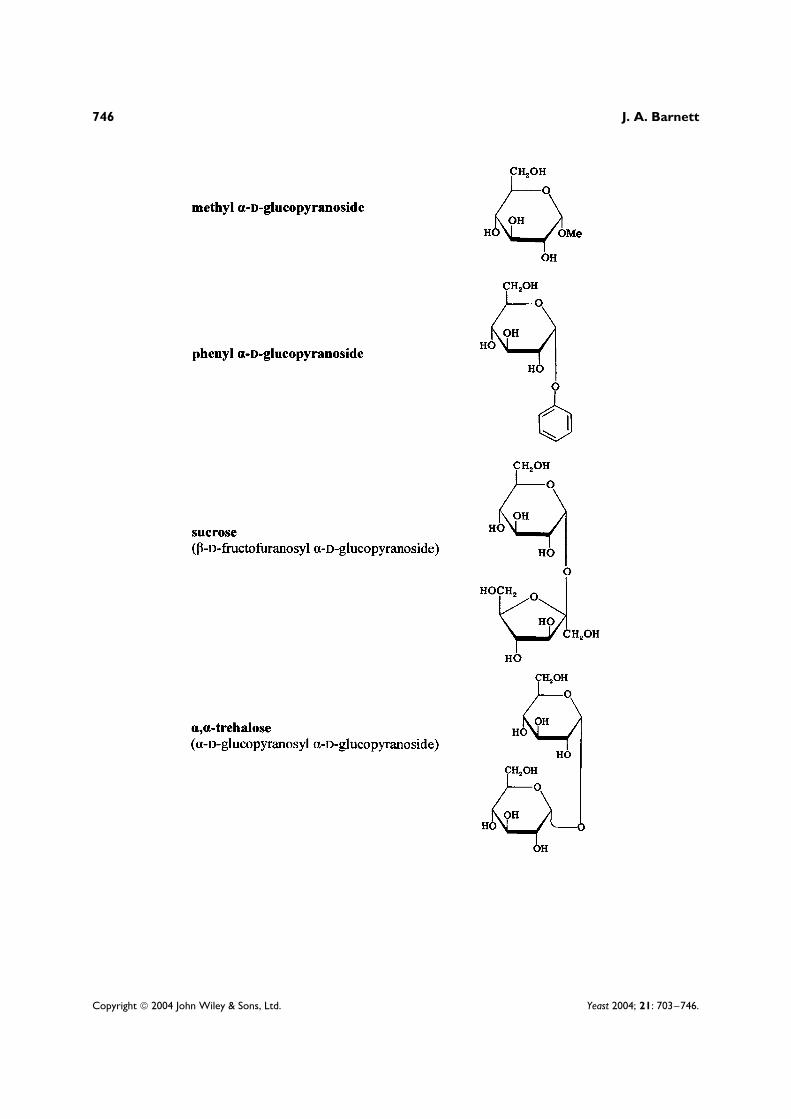
746 J. A. Barnett
Copyright 2004 John Wiley & Sons, Ltd. Yeast 2004; 21: 703–746.





























