Embed Size (px)
Citation preview

Journal of Biotechnology, 10 (1989) 161-170 Elsevier
JBT 00380
161
Biological delignification of pulp by Phanerochaete chrysosporium
J. Pellinen, J. Abuhasan, T.W. Joyce and H.-M. Chang
North Carolina State University, Department of Wood and Paper Science, Box 8005, Raleigh, NC 27695-8005, U.S.A.
(Received 9 November 1988; accepted 22 February 1989)
Summary
Biological delignification of wood pulp was studied using the white-rot fungus Phanerochaete chrysosporium in liquid cultures. Kraft pulp and chemithermo-mecha- nical pulp (CTMP) were used as substrates. Both types of pulp could readily be delignified by the shaking cultures of the fungus: the kappa number of kraft pulp decreased from 33 to less than 10 in two weeks and the Klason lignin content of CTMP decreased from 26.5 to 21.3% in the same time. Stationary cultures did not delignify pulps effectively. During the fungal treatment, the strength of CTMP increased substantially. At a freeness level of about 350 ml, the tensile and tear indices increased by 20% and the burst index by 40% as compared to the original pulp. Unfortunately, CTMP turned dark during the treatment. The ISO brightness was originally 52.5, but after the incubation with the fungus it was only 17.9. After peroxide bleaching the brightness was still low, only 41.6.
Biodelignification; Kraft pulp; Chemithermo-mechanical pulp; White-rot fungus; Phanerochaete chrysosporium
Introduction
White-rot fungi are known for their ability to degrade all the main components of wood, including lignin. In addition, some of them can degrade a variety of synthetic, aromatic and chlorinated compounds (Bumpus et al., 1985). The degrada-
Correspondence to: J. PeUinen STFI, Box 5604, S-114 86 Stockholm, Sweden.
0168-1656/89/$03.50 © 1989 Elsevier Science Publishers B.V. (Biomedical Division)

162
tion of lignin occurs during secondary metabolism which is triggered by a limitation of an essential nutrient such as carbon or nitrogen. In secondary metabolism, the fungus produces many extracellular, nonspecific lignin peroxidases which initiate the degradation of lignin (Tien and Kirk, 1983; Glenn et al., 1983; for a review see Kirk and Farrell, 1987).
The ability of the white-rot fungi to degrade lignin makes them a logical choice for study of the biological delignification of pulp. Delignification of unbleached kraft pulp and thermo-mechanical pulp (TMP) as well as treatment of wood chips before refining has been studied using Phanerochaete chrysosporium (also known as Sporotrichum pulverulentum) and Dichomitus squalens (Kirk and Yang, 1979; Yang et al., 1980; Eriksson and Vallander, 1982, Bar-Lev et al., 1982; Johnsrud et al., 1987; Tran and Chambers, 1987; Myers et al., 1988). The fungus degrades lignin only to gain access to carbohydrates since they cannot utilize lignin as their carbon and energy source. The carbohydrates can be protected by adding glucose to the cultures to repress the production of cellulose-degrading enzymes. Eriksson and Vallander (1982) and Johnsrud et al. (1987) used a cellulase-less mutant of the fungus as another way to prevent degradation of carbohydrates.
We studied delignification of chemithermo-mechanical pulp (CTMP) and un- bleached kraft pulp in a similar way as Bar-Lev et al. (1982), Yang et al. (1980) and Kirk and Yang (1979). Our results differed in that we achieved delignification only in shaking cultures and that also softwood pulp was delignified by P. chrysosporium in our study. Unbleached kraft pulp was used mainly to screen culture conditions.
Materials and Methods
Microorganism Phanerochaete chrysosporium Burds. (BKM F-1767) was obtained from the
U.S.D.A. Forest Products Laboratory, Madison, WI, U.S.A. It was cultured on 2% malt agar and later also on sterile rice. A spore suspension of 3-4-week-old cultures was used as the inoculum. The density of the suspension was adjusted to give an absorbance of 0.5 at 650 nm, corresponding to about 2.5 × 10 6 spores per ml.
Culture conditions The culture medium contained the following: 94 ml nutrient solution, 63 ml
mineral solution, 0.8 g benzyl alcohol, 1.0 g Tween 80, 10 ml penicillin-streptomy- cin-neomycin mixture or 0.20 g chloramphenicol (Sigma, St. Louis, MO, U.S.A.), 10.00 g glucose, 0.032 g NH4C1 (0.6 mM, unless otherwise indicated) and distilled water to a total of 1000 ml. The pH was adjusted to 4.5 and buffered in some cases with 30 mM sodium tartrate.
The nutrient solution contained per liter of distilled water: 2.0 g KH2PO 4, 0.5 g MgSO4.TH20, 0.1 g CaC12 and 1.0 mg thiamine-HC1. The mineral solution contained per liter of distilled water: 1.5 g nitrilotriacetic acid, 0.5 g MnSO 4 • H20, 0.1 g FeSO 4. 7H20, COSO4 and ZnSO 4, and 0.01 g CuSO 4. 5H20, AIK(SO4) 2, H3BO 3 and NaMoO 4.

163
The incubation was done in 500 ml flasks with 90 ml of the culture medium, 10 ml of spore suspension, and 1.0 g of oven dry pulp. The solutions, flasks, and pulp samples were sterilized in an autoclave (30 min, 121°C). The control samples contained distilled water instead of the spore suspension and 1 ml of cycloheximide solution (10 mg per ml in ethanol; Sigma) to prevent the growth of microorganisms.
The flasks were incubated at 39°C either in a rotary shaker or stationary. The shaker speed was 150 rpm for the first two days and then 100 rpm for the rest of the time; the amplitude was 2.5 cm. The shaker speed was changed to adjust the size of the mycelium-pulp pellets (at a low speed, the pellets were very large or did not form at all). The flasks were flushed with oxygen every day.
Substrates The unbleached kraft pulp sample (kappa number 33) was obtained from a
commercial mill. The chemithermo-mechanical pulp (CTMP) A2 was an experimen- tal pulp from a pilot plant of C-E Sprout-Bauer, Inc.: Loblolly pine pin chips were impregnated with 5.1% Na 2SO3 under pressure followed by a short (20 rain) cook in a rapid cycle digester at 160 °C with 5% Na2SO3. Refining was done with a Model 418 pressurized double disc refiner (12 s, 2.1 kg cm -2, 47 kWh) with a yield of 83.5%. The pulp was stored at 4 ° C after washing.
Analytical methods After the incubation with the fungus, the pulp samples were filtered with a 400
mesh nylon screen and washed. Residual lignin was determined as kappa number for kraft pulp or as Klason lignin for mechanical pulps (TAPPI standards T 222 and T 230) after alkali extraction (3% NaOH on pulp, 70 o C, 1 h).
Strength properties (tensile, tear and burst strength) were determined according to TAPPI standards T 205, 220, 236, 248, 403, 411, and 414 after beating with a PFI mill.
Brightness was determined with an Elrepho 2000 instrument (Datacolor, Switzer- land) according to ISO 3688 standard. The reflectance spectra were recorded with a Pye Unicam SP8-100 spectrophotometer (Pye Unicam, Cambridge, U.K.).
Bleaching conditions were the following: pH 11.0, 4% (w/w) H202, consistency 12%, temperature 60 o C, time 2 h. Before bleaching the pulp was treated with a DTPA solution (0.5%) at 3% consistency to remove metal ions (Singh, 1979; Daneault et al., 1987). Sodium silicate (0.05%) and MgSO 4 (5%) were used to stabilize the peroxide and to prevent degradation of carbohydrates.
Results
Effect of pH and nutrient nitrogen on delignification The preliminary experiments were conducted with unbleached kraft pulp to
screen for the conditions of cultivation. The results indicated that consistency, flask size, liquid volume and shaking speed were key variables for delignification. It was, however, difficult to determine the optimum conditions regarding these factors
164
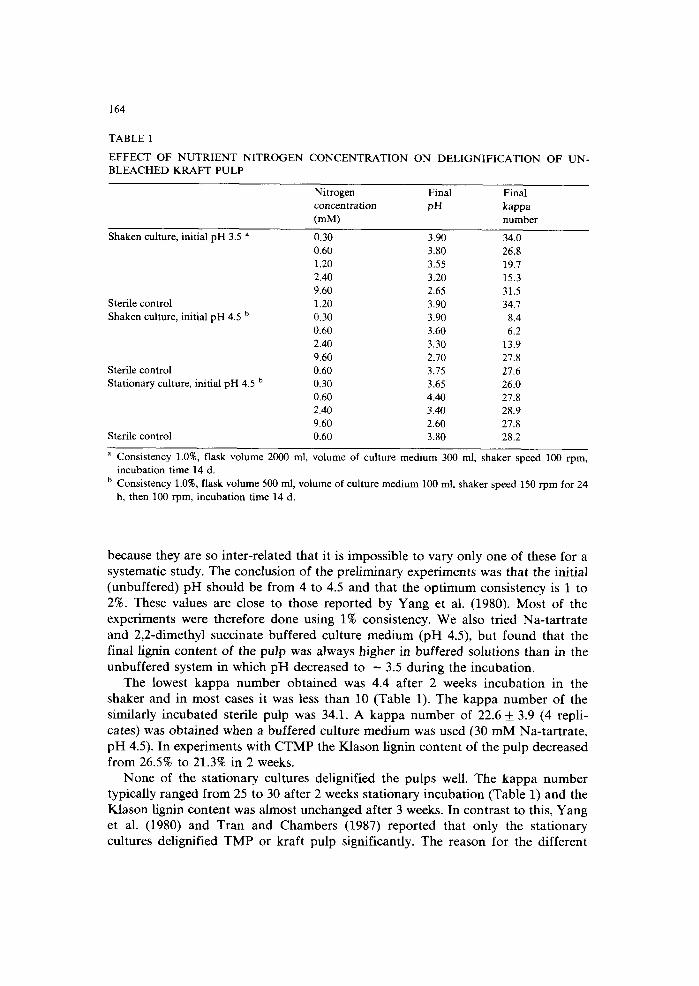
TABLE 1
EFFECT OF NUTRIENT NITROGEN BLEACHED KRAFT PULP
CONCENTRATION ON DELIGNIFICATION OF UN-
Nitrogen Final Final concentration pH kappa (mM) number
Shaken culture, initial pH 3.5 a 0.30 3.90 34.0 0.60 3.80 26.8 1.20 3.55 19.7 2.40 3.20 15.3 9.60 2.65 31.5
Sterile control 1.20 3.90 34.7 Shaken culture, initial pH 4.5 b 0.30 3.90 8.4
0.60 3.60 6.2 2.40 3.30 13.9 9.60 2.70 27.8
Sterile control 0.60 3.75 27.6 Stationary culture, initial pH 4.5 b 0.30 3.65 26.0
0.60 4.40 27.8 2.40 3.40 28.9 9.60 2.60 27.8
Sterile control 0.60 3.80 28.2
a Consistency 1.0%, flask volume 2000 ml, volume of culture medium 300 ml, shaker speed 100 rpm, incubation time 14 d.
b Consistency 1.0%, flask volume 500 ml, volume of culture medium 100 ml, shaker speed 150 rpm for 24 h, then 100 rpm, incubation time 14 d.
because they are so in te r - re la ted that it is imposs ib le to vary on ly one of these for a sys temat ic s tudy. The conclus ion of the p re l imina ry exper imen t s was that the ini t ia l (unbuffered) p H should be f rom 4 to 4.5 and that the o p t i m u m cons is tency is 1 to 2%. These values are close to those r epor t ed by Yang et al. (1980). M o s t of the exper iments were therefore done using 1% consis tency. W e also t r ied N a - t a r t r a t e and 2,2-dimethyl succinate buf fe red cul ture m e d i u m (pH 4.5), bu t found that the f inal l ignin conten t of the pu lp was a lways h igher in buf fe red solut ions than in the unbuf fe red sys tem in which p H decreased to - 3.5 dur ing the incuba t ion .
The lowest k a p p a number ob t a ined was 4.4 af ter 2 weeks incuba t ion in the shaker and in mos t cases it was less than 10 (Table 1). The k a p p a n u m b e r of the s imilar ly i ncuba ted steri le pu lp was 34.1. A k a p p a n u m b e r of 22.6 + 3.9 (4 repl i - cates) was ob t a ined when a buffered cul ture m e d i u m was used (30 m M Na- t a r t r a t e , p H 4.5). In exper iments wi th C T M P the K l a s o n l ignin con ten t of the pu lp decreased f rom 26.5% to 21.3% in 2 weeks.
N o n e of the s ta t ionary cul tures del ignif ied the pu lps well. The k a p p a n u m b e r typica l ly ranged f rom 25 to 30 af ter 2 weeks s t a t ionary incuba t ion (Table 1) and the K la son l ignin con ten t was a lmos t unchanged af ter 3 weeks. In con t ras t to this, Yang et al. (1980) and Tra i l and Chamber s (1987) r epor t ed tha t on ly the s t a t iona ry cul tures del ignif ied T M P or kraf t pu lp s ignif icant ly. The reason for the d i f ferent

165
results may in part be connected to the use of Tween 80 in the present study (J~iger et al., 1985; Asther et al., 1987; see Discussion).
It is well known that nitrogen-limited conditions cause the fungus to shift to secondary metabolism (Kirk, 1978). We, therefore, determined the optimum nitro- gen concentration for delignification in shaking cultures using kraft pulp as the substrate. This optimum was found to be 0.6 mM NH~ in the solution at an initial pH of 4.5 and 2.4 mM at an initial pH of 3.5 (Table 1). No conclusions on the optimum nitrogen level could be made in the case of stationary cultures as there was practically no delignification observed.
Rate of delignification The rate of delignification was followed in separate experiments. The kappa
number of kraft pulp decreased from 33 to 21 in 6 d and to 8.9 in 14 d (Fig. 1). From a practical viewpoint, it might be enough to delignify pulp only slightly using fungal treatment and then use some chemical bleaching process to reach the brightness desired.
In contrast to the relatively fast delignification of kraft pulp during the first week, no decrease of lignin in CTMP had taken place by day 6 (Fig. 1). After 14 d the lignin content of CTMP had decreased by 20%, In 3 weeks, 28% of the lignin was degraded and the pulp had a Klason lignin content of 19.1% (Fig. 1). Practically no lignin degradation took place in the stationary cultures in 21 d.
J2 E
40
30
20
10
t\ , \
\ .,, \ •
D~ D • ~D
' I I I ' I ' I
5 i0 15 20
_ 3 0
26
22
18
14
Tlrne (d)
Fig. 1. Rate of delignification of kraft pulp (D, kappa number) and CTMP (11, Klason lignin) by Phanerochaete ehrysosporium.
166
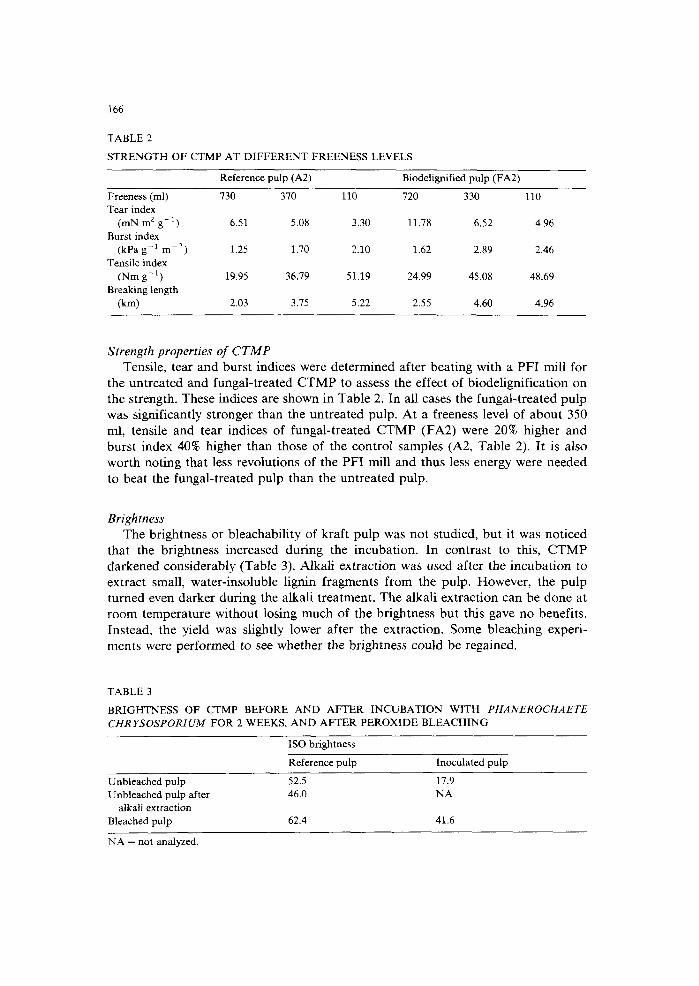
TABLE 2
S T R E N G T H OF CTMP AT D I F F E R E N T FREENESS LEVELS
Reference pulp (A2) Biodelignified pulp (FA2)
Freeness (ml) 730 370 110 720 330 110 Tear index
(mN m 2 g - 1 ) 6.51 5.08 3.30 11.78 6.52 4.96 Burst index
(kPa g-1 m 2) 1.25 1.70 2.10 1.62 2.89 2.46 Tensile index
(Nm g - l ) 19.95 36.79 51.19 24.99 45.08 48.69 Breaking length
(km) 2.03 3.75 5.22 2.55 4.60 4.96
Strength properties of CTMP Tensile, tear and burst indices were determined after beating with a PFI mill for
the untreated and fungal-treated CTMP to assess the effect of biodelignification on the strength. These indices are shown in Table 2. In all cases the fungal-treated pulp was significantly stronger than the untreated pulp. At a freeness level of about 350 ml, tensile and tear indices of fungal-treated CTMP (FA2) were 20% higher and burst index 40% higher than those of the control samples (A2, Table 2). It is also worth noting that less revolutions of the PFI mill and thus less energy were needed to beat the fungal-treated pulp than the untreated pulp.
Brightness The brightness or bleachability of kraft pulp was not studied, but it was noticed
that the brightness increased during the incubation. In contrast to this, CTMP darkened considerably (Table 3). Alkali extraction was used after the incubation to extract small, water-insoluble lignin fragments from the pulp. However, the pulp turned even darker during the alkali treatment. The alkali extraction can be done at room temperature without losing much of the brightness but this gave no benefits. Instead, the yield was slightly lower after the extraction. Some bleaching experi- ments were performed to see whether the brightness could be regained.
TABLE 3
BRIGHTNESS OF CTMP BEFORE A N D AFTER I N C U B A T I O N WITH PHANEROCHAETE CHR YSOSPORIUM F OR 2 WEEKS, A N D AF TER PEROXIDE B L E A C H I N G
ISO brightness
Reference pulp Inoculated pulp
Unbleached pulp 52.5 17.9 Unbleached pulp after 46.0 NA
alkali extraction Bleached pulp 62.4 41.6
NA = not analyzed.

167
80
ORIGiAL PULP
g 6o
~ 4o
~ 2o re-
AFTER FUNGAL TREATMENT
0 250 350 450 550 650
Wavelength (rim) Fig. 2. Reflectance UV-spectra of CTMP original pulp (A2) and fungal-treated pulp (FA2).
The reflectance spectra (Fig. 2) show that the biodelignified CTMP (FA2) reflected less light over the entire range of the spectrum than the original pulp (A2). Thus, the FA2 pulp contains many different chromophores, ranging from quinones to other highly conjugated systems and possibly chelates. Using energy dispersive X-ray analysis, we found that the pulps A2 and FA2 contained similar amounts of iron, nickel and copper, and that A2 contained more aluminum, whereas FA2 contained more calcium. Manganese was not present in the samples although it was added to the cultures. As the metal contents of fungal-treated pulp and control pulp were similar, it seems unlikely that the presence of chelates or deposited minerals is responsible for the darkening of the pulp.
Discussion
The results reported here differ in some cases considerably from those obtained earlier by Kirk and Yang (1979), Yang et al. (1980), and Tran and Chambers (1987). We achieved marked delignification of both unbleached kraft pulp and CTMP, but not in stationary cultures as they did. Our finding was rather surprising since until 1985 all reports showed that lignin is degraded in stationary but not in agitated cultures.
At present, there are several reports showing that ligninolytic activity can be obtained in shaken cultures (Reid et al., 1985; Leisola and Fiechter, 1985). It seems that the use of a detergent (like Tween 80) is important in agitated cultures (J~iger et al., 1985; Asther et al., 1987). In our study the fungus grew well in stationary cultures, but lignin was hardly degraded. Stationary cultures without the detergent showed no difference to this nor did the use of varying amounts of free liquid in the flasks (Tran and Chambers, 1987).
The pH of the unbuffered culture medium decreased from 4.5 to about 3.5 during the incubation, probably because of acidic lignin degradation products which were released. This was apparently advantageous since the fungus grows optimally at pH

168
of 4.5, but the optimal delignification occurs at lower pH values (Tran and Chambers, 1987). The delignification was less complete in buffered cultures.
The optimum concentration of nutrient nitrogen was found to be 0.6 mM NH~ (0.084% on dry pulp basis) which is less than that obtained by Yang et al. (1980; 0.12-1.2%). In their study, only up to 3.8% of the lignin in softwood TMP (western hemlock) was degraded in 2 weeks, which was low compared to the 30% degradation of lignin in hardwood TMP (red alder) found in the same study. We did not use hardwood pulp, but lignin degradation of softwood pulp (Loblolly pine) was 20% in 14 d.
The darkening of mechanical pulp during the incubation is the most serious drawback of the biodelignification method. Our results regarding the brightness of pulp are similar to those reported earlier (Yang et al., 1980; Eriksson and Vallander, 1982; Johnsrud et al., 1987; Myers et al., 1988). Bar-Lev et al. (1982) reported that biodelignified TMP retained its brightness. In this case a short incubation time was used (5 d) after which only 3% of the lignin had been degraded. It seems possible that the pulp could have turned darker later. Eriksson (1988) reported increase in both brightness and viscosity of kraft pulp made of rotted wood.
It was shown in a related study (Abuhasan et al., 1988) that a 80% increase in tensile index was achieved in 5 d with the final ISO brightness still at 33. It seems, therefore, that a practical optimum may be found for the strength increase, loss of brightness, and treatment time.
Deposition of manganese in the form of MnO 2 was considered as one explanation for the darkening of pulp (Blanchette, 1984), but the results of the X-ray analysis disproved this hypothesis as the content of metals was approximately the same for both the untreated and the fungal-treated pulp. However, the metals might form different colored complexes with the original lignin of pulp and that modified by the fungus. Another possible explanation is the formation of melanin - a dark pigment composed of amino acids and phenolic residues that is often associated with fungal mycelium (Linhares and Martin, 1978). In part, the discoloration of mechanical pulp seems to be connected to the presence of sulfonate groups as kraft pulp did not turn dark. In a separate study for effluent purification we found that also the wastewater from CTMP production became darker when it was treated with P. chrysosporium (Pellinen, unpublished), whereas the spent bleach liquor from kraft pulping could effectively be decolorized in the system (Pellinen et al., 1988).
The increase of strength of CTMP observed in the present study was quite significant. It may in part be due to the increase in the number of carboxyl and hydroxyl groups of lignin as a result of the fungal attack (Chen and Chang, 1985). Eriksson and Vallander (1982) reported only small changes in the strength of TMP after fungal treatment of the chips with wild-type P. chrysosporium. More signifi- cant strength increase for CTMP or TMP after fungal treatment has also been reported (Johnsrud et al., 1987; Myers et al., 1988), but considerable carbohydrate degradation has been found in other studies (Kirk and Yang, 1979).
The results of this study as well as those obtained earlier indicate that white-rot fungi can be used to delignify wood or pulp so that the pulp strength is improved and energy consumption reduced. The biodelignification of kraft pulp may be

169
overest imated because of the l ignin de te rmina t ion method used (kappa number) . According to W u and Glasser (1979), the kappa n u m b e r of biodel ignif ied pulp is from 6 t o 20% too low. The error in Klason l ignin is p robab ly smaller. For C T M P the impor tan t point is the observed increase of strength, not the extent of delignifi- cation.
The disadvantages of the method include long incuba t ion t ime and loss of pulp brightness. By further developing the fungal strains, condit ions, and appara tus as well as by combin ing biological and chemical processes this approach may become economical ly competitive.
Acknowledgements
The authors wish to thank Mrs. Elizabeth Wilson for her help in the analyt ical work and C.-E. Sprout-Bauer, Inc. for the f inancial suppor t and for the C T M P
samples.
References
Abuhasan, M.J., Pellinen, J., Joyce, T.W. and Chang, H.-M. (1988) Biodelignification of chemi-thermo- mechanical pulp by a fungal treatment. Proceedings of the AIChE Annual Meeting: New Develop- ments in Pulping and Bleaching, Washington D.C., Nov. 27-Dec. 2.
Asther, M., Corrieu, G., Drapron, R. and Odier, E. (1987) Effect of Tween 80 and oleic acid on ligninase production by Phanerochaete chrysosporium INA-12. Enzyme Microb. Technol. 9, 245-249.
Bar-Lev, S.S., Kirk, T.K. and Chang, H.-M. (1982) Fungal treatment can reduce energy requirements for secondary refining of TMP. TAPPI J. 65, 111-113.
Blanchette, R.A. (1984) Manganese accumulation in wood decayed by white rot fungi. Phytopathology 74, 725-730.
Bumpus, J.A., Tien, M., Wright, D. and Aust, S.D. (1985) Oxidation of persistent environmental pollutants by a white rot fungus. Science 228, 1434-1436.
Chen, C.L. and Chang, H.-M. (1985) Chemistry of lignin biodegradation. In: Higuchi, T. (Ed.), Biosynthesis and Biodegradation of Wood, San Diego, CA, Academic Press, pp. 535-556.
Daneault, C., Law, K.N. and Valade, J.L. (1987) Brightening of ultra-high yield larch pulp. Proceedings of the 4th International Symposium on Wood and Pulping Chemistry, Paris, Apr. 27-30.
Eriksson, K.-E. (1988) Microbial delignification - basis, potentials and applications. In: Aubert, J.P., Beguin, P. and Millet, J. (Eds.), Biochemistry and genetics of lignocellulose degradation, Academic Press, London, p. 285-302.
Eriksson, K.-E. and Vallander, L. (1982) Properties of pulps from thermomechanical pulping of chips pretreated with fungi. Sven. Papperstidn. 85, 34-38.
Glenn, J.K., Morgan, M.A., Mayfield, M.B., Kuwahara, M. and Gold, M.H. (1983) An extracellular H202-requiring enzyme preparation involved in lignin biodegradation by the white rot Basidiomycete Phanerochaete chrysosporium. Biochem. Biophys. Res. Commun. 114, 1077-1083.
Johnsrud, S.C., Fernandez, N., Lopez, P., Gutierre, I., Saez, A. and Eriksson, K.-E. (1987) Properties of fungal pretreated high yield bagasse pulps. Nordic Pulp Paper Res. J. 2, 47-52.
J~iger, A., Croan, S. and Kirk, T.K. (1985) Production of ligninases and degradation of lignin in agitated submerged cultures of Phanerochaete chrysosporium. Appl. Environ. Microbiol. 50, 1274-1278.
Kirk, T.K. (1978) Influence of culture parameters on lignin metabolism by Phanerochaete chrysosporium. Arch. Microbiol. 117, 277-285.

170
Kirk, T.K. and Farrell, R.L. (1987) Enzymatic "combustion": The microbial degradation of lignin. Annu. Rev. Microbiol. 41,465-505.
Kirk, T.K. and Yang, H.H. (1979) Partial delignification of unbleached kraft pulp with ligninolytic fungi. Biotechnol. Lett. 1, 347-352.
Leisola, M.S.A. and Fiechter, A. (1985) Lignin production in agitated conditions by Phanerochaete chrysosporium. FEMS Microbiol. Lett. 29, 33-36.
Linhares, L.F. and Martin, J.P. (1978) Decomposition in soil of the humic acid-type polymers (melanins) of Eurotium echinulatum, Aspergillus glaucus sp. and other fungi. Soil Sci. Soc. Am. J. 42, 738-743.
Myers, G.C., Leatham, G.F., Wegner, T.H. and Blanchette, R.A. (1988) Fungal treatment of aspen chips improves strength of refiner mechanical pulp. TAPPI J. 71, 105-108.
Pellinen, J., Yin, C.-F., Joyce, T.W. and Chang, H.-M. (1988) Treatment of chlorine bleaching effluent using a white-rot fungus. J. Biotechnol. 8, 67-76.
Reid, I.D., Chao, E.E. and Dawson, P.S.S. (1985) Lignin degradation by Phanerochaete chrysosporium in agitated cultures. Can J. Microbiol. 11, 88-90.
Singh, R.P. (1979) The bleaching of pulp, 3rd edition, TAPPI Press, Atlanta, GA, pp. 211-252. Tien, M. and Kirk, T.K. (1983) Lignin-degrading enzyme from the Hymenomycete Phanerochaete
chrysosporium Burds. Science 221,661-663. Tran, A.V. and Chambers, R.P. (1987) Delignification of an unbleached hardwood kraft pulp by
Phanerochaete chrysosporium. Appl. Microbiol. Biotechnol. 25, 484-490. Wu, L.C.-F. and Glasser, W.G. (1979) Utility of oxidative lignin determination methods for biodegraded
lignocellulosic substrates. Biotechnol. Bioeng. 21, 1679-1683. Yang, H.H., Effland, M..I. and Kirk, T.K. (1980) Factors influencing fungal degradation of lignin in a
representative lignocellulosic, thermomechanical pulp. Biotechnol. Bioeng. 22, 65-77.





























