Embed Size (px)
DESCRIPTION
Cognitie Si Emotie
Citation preview

This article was downloaded by: [79.116.219.66]On: 05 May 2012, At: 05:41Publisher: Psychology PressInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: MortimerHouse, 37-41 Mortimer Street, London W1T 3JH, UK
Journal of Cognitive PsychologyPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/pecp21
Neural correlates of emotion–cognitioninteractions: A review of evidence from brainimaging investigationsFlorin Dolcos a b c , Alexandru D. Iordan b & Sanda Dolcos aa Psychology Department, University of Illinois Urbana-Champaign, Urbana, IL, USAb Neuroscience Program, University of Illinois Urbana-Champaign, Urbana, IL, USAc Beckman Institute for Advanced Science & Technology, University of IllinoisUrbana-Champaign, Urbana, IL, USA
Available online: 22 Sep 2011
To cite this article: Florin Dolcos, Alexandru D. Iordan & Sanda Dolcos (2011): Neural correlates of emotion–cognitioninteractions: A review of evidence from brain imaging investigations, Journal of Cognitive Psychology, 23:6, 669-694
To link to this article: http://dx.doi.org/10.1080/20445911.2011.594433
PLEASE SCROLL DOWN FOR ARTICLE
Full terms and conditions of use: http://www.tandfonline.com/page/terms-and-conditions
This article may be used for research, teaching, and private study purposes. Any substantial orsystematic reproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distributionin any form to anyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representation that thecontents will be complete or accurate or up to date. The accuracy of any instructions, formulae, anddrug doses should be independently verified with primary sources. The publisher shall not be liable forany loss, actions, claims, proceedings, demand, or costs or damages whatsoever or howsoever causedarising directly or indirectly in connection with or arising out of the use of this material.

Neural correlates of emotion�cognition interactions: Areview of evidence from brain imaging investigations
Florin Dolcos1,2,3, Alexandru D. Iordan2, and Sanda Dolcos1
1Psychology Department, University of Illinois Urbana-Champaign, Urbana, IL, USA2Neuroscience Program, University of Illinois Urbana-Champaign, Urbana, IL, USA3Beckman Institute for Advanced Science & Technology, University of Illinois Urbana-
Champaign, Urbana, IL, USA
Complex dynamic behaviour involves reciprocal influences between emotion and cognition. On the onehand, emotion is a ‘‘double-edged sword’’ that may affect various aspects of our cognition and behaviour,by enhancing or hindering them and exerting both transient and long-term influences. On the other hand,emotion processing is also susceptible to cognitive influences, typically exerted in the form of emotionregulation. Noteworthy, both of these reciprocal influences are subjective to individual differences thatmay affect the way we perceive, experience, and eventually remember emotional experiences, or respondto emotionally challenging situations. Understanding these relationships is critical, as unbalancedemotion�cognition interactions may lead to devastating effects, such as those observed in mood andanxiety disorders. The present review analyses the reciprocal relationships between emotion andcognition, based on evidence derived from brain imaging investigations focusing on three main topics: (1)the impact of emotion on cognition, (2) the impact of cognition on emotion, and (3) the role of individualdifferences in emotion�cognition interactions. This evidence will be discussed in the context ofidentifying aspects that are fundamental to understanding the mechanisms underlying emotion�cognitioninteractions in healthy functioning, and to understanding changes associated with affective disorders.
Keywords: Affective�cognitive interactions; Amygdala; Event-related potentials; Functional magneticresonance imaging; Medial-temporal lobe memory system; Neural circuitry; Prefrontal cortex.
The ability to process emotional stimuli withincreased efficacy depends on neural mechanismsthat allow detection, identification, and proces-sing of stimuli and situations that are importantfor survival (e.g., finding food, avoiding preda-tors) (Anderson & Phelps, 2001; LeDoux, 1996;Ohman, Flykt, & Ludqvist, 2000). These mechan-isms have evolved during evolution and arecontinuously modelled during ontogeny. At abasic level, these mechanisms overlap with thoseinvolved in the generalised response to stressfulsituations, which consist of coordinated neurohor-monal changes that are set in motion when a
stressor of any kind exceeds specific thresholds(Arnsten, 1998; Selye, 1985). The engagement ofthe stress system as a result of exposure touncontrollable stressful situations sets off a chainof events that not only ensure immediate adaptiveresponses but also trigger mechanisms that willdetermine our responses in future similar situa-tions.
Emotions may affect various aspects of ourcognition and behaviour, by enhancing or hinder-ing them and by exerting both transient and long-term influences. Possibly as a result of theirrelevance for survival, emotional stimuli tend to
Correspondence should be addressed to Florin Dolcos, Assistant Professor of Psychology & Neuroscience, 2057 Beckman
Institute for Advanced Science & Technology, University of Illinois at Urbana-Champaign, 405 North Mathews Avenue, Urbana, IL
61801, USA. E-mail: [email protected]
JOURNAL OF COGNITIVE PSYCHOLOGY, 2011, 23 (6), 669�694
# 2011 Psychology Press, an imprint of the Taylor & Francis Group, an Informa businesshttp://www.psypress.com/ecp http://dx.doi.org/10.1080/20445911.2011.594433
Dow
nloa
ded
by [
79.1
16.2
19.6
6] a
t 05:
41 0
5 M
ay 2
012

capture our attention more easily than none-motional stimuli, and thus may affect differentlevels of our cognition, from lower level (e.g.,perceptual) to higher level (e.g., mnemonic andexecutive) cognitive processes. The enhancedsignificance of emotional stimuli can benefitcognitive processes (e.g., better memory foremotional events) (Dolcos, 2010; Dolcos &Denkova, 2008; Dolcos, LaBar, & Cabeza, 2006;McGaugh, 2004; Phelps, 2004), but can also havedetrimental effects on behaviour (e.g., increaseddistractibility to task-irrelevant emotional stimuli)(Dolcos, Diaz-Granados, Wang, & McCarthy,2008; Dolcos, Kragel, Wang, & McCarthy, 2006;Dolcos & McCarthy, 2006). Also, although someof these effects are transient, influencing onlineperceptual and executive processes, others pro-duce long-term effects (Dolcos, LaBar, & Cabeza,2005) that can last for a lifetime (Markowitsch,2008; Moscovitch, Nadel, Winocur, Gilboa, &Rosenbaum, 2006; Rubin, 2005). Evidence con-cerning these aspects will be discussed in the firstsection of this review.
Emotion processing is also susceptible tocognitive influences, as complex behaviour in-volves reciprocal interactions between affectiveand cognitive processes. Indeed, the neurobiolo-gical theories of cognitive�affective interactionswould not be complete without an account of themechanisms underlying the effect of cognitiveprocesses on emotional processing. Particularlyimportant in this context is the ability to promptlydeploy cognitive control of emotion in order toresist momentary emotional distraction and, in alonger run, the ability to cope with emotionalresponses as a result of recollecting memories ofunpleasant events. The engagement of thesemechanisms can occur automatically, probablyas a result of gradual development during onto-geny, and depend on the attributed personalsignificance to the potentially emotional situa-tions. For instance, when witnessing a tragicaccident some individuals may be more distressedand hence experience and express intense emo-tions, whereas others are better able to controltheir emotions (e.g., by reappraising the situation,or by concealing their emotions), and henceoverall reduce their emotional experience. Be-cause emotion regulation strategies have a sub-stantial impact on one’s emotional experience,leading to different behavioural, cognitive, andneural results (Gross, 2008; Ochsner & Gross,2005; Richards, 2004), they are an important partof emotion�cognition interactions. Evidence con-
cerning these aspects will be discussed in thesecond section.
Understanding the mechanisms underlying thereciprocal interactions between emotion and cog-nition is critical for understanding fundamentals ofhealthy functioning, as well as changes associatedwith psychological and emotional disorders. Animportant aspect in this context concerns the roleof individual differences in emotion�cognitioninteraction, and, related to this, individual varia-tions in the vulnerability to affective disorders, aseven within the realm of healthy emotional re-sponses some individuals may tend to be moreemotionally responsive and ‘‘anxious’’, whereasothers can control better their emotions. Under-standing the role of individual differences canprovide insights into the factors that may influencethe susceptibility to affective disorders, in whichunbalanced interactions between emotion andcognition may lead to devastating effects, such asthose observed in depression and anxiety. Forexample, the tendency to ruminate on negativeemotions and memories observed in depressedpatients and the intrusive recollection of traumaticmemories observed in patients with posttraumaticstress disorder affect tremendously the way thesepatients think and behave. Therefore, it hasbecome apparent that finding cures for thesedisorders depends on understanding the brainmechanisms that are responsible for such dramaticchanges in the ways emotion interfaces withcognition, as well as on identifying the role ofindividual differences in mediating these interac-tions. Evidence concerning these aspects will bediscussed in the third section.
To summarise, the present review analyses thereciprocal interactions between emotion andcognition, as derived from brain imaging investi-gations focusing on the three main topics high-lighted earlier: (1) the impact of emotion oncognition, (2) the impact of cognition on emotion,and (3) the role of individual differences inemotion�cognition interactions. First, we willdiscuss evidence concerning the neural circuitryunderlying the enhancing/beneficial and impair-ing/detrimental impact of emotion on cognitiveprocesses, by considering findings from studiesfocusing on both lower level perceptual proces-sing and higher level executive functions, and onboth transient and longer lasting effects of emo-tion. Conversely, we will consider the reverse sideof these effects, namely the impact of cognitionon affective processing, by addressing evidenceconcerning the neural correlates of emotion
670 DOLCOS, IORDAN, DOLCOS
Dow
nloa
ded
by [
79.1
16.2
19.6
6] a
t 05:
41 0
5 M
ay 2
012

regulation and coping with emotional distraction.Finally, we will also discuss the reciprocal inter-actions between emotion and cognition in thecontext of highlighting the role of individualdifferences (i.e., related to personality, sex, andage).
THE IMPACT OF EMOTION ONCOGNITION
The impact of emotion on perceptionand attention
In this section, we will review two main polarisedpositions concerning emotion processing: onesupporting a traditional view according to whichemotion processing is automatic and does notdepend on the availability of processing resources(Vuilleumier, Armony, Driver, & Dolan, 2001),and the other supporting a competing view, whichposits that emotion processing depends on atten-tional resources (Pessoa, McKenna, Gutierrez, &Ungerleider, 2002). In the following paragraphs,we will discuss the main evidence supportingthese two opposing views, along with their re-ciprocal critiques, as well as emerging evidencesupporting the idea that the two views are notmutually exclusive.
The traditional view proposes that, probablybecause of their relevance for survival, processingof emotional stimuli is generally automatic. Anautomatic process is fast, unintentional and un-conscious, therefore it is not subject to control,and cannot be avoided or interrupted in its course(Schneider & Shiffrin, 1977). Evidence comesfrom studies showing that emotional stimuli canbe detected and processed with increased efficacy,and that this privileged processing depends on theamygdala (AMY), a main brain structure asso-ciated with emotion processing. For instance,visual search paradigms reveal more rapid andaccurate detection of schematically depictedthreatening faces presented among similar dis-tractors, an effect called the ‘‘face in the crowd’’,‘‘snake in the grass’’, or ‘‘pop-out’’ effect (e.g.,Ohman, Lundqvist, & Esteves, 2001). Also, evi-dence from AMY patients shows that intactAMY is needed to observe enhancement ofattention by emotional stimuli, which eliminate‘‘attentional blinks’’ during processing of rapidlysucceeding stimuli (Anderson & Phelps, 2001).Further supporting the traditional view and the
role of the AMY, brain imaging evidence showsthat facial stimuli with emotional expressions(e.g., expressing fear) can be processed even inthe absence of awareness (Whalen et al., 1998)
This and other evidence led to the generallyaccepted notion that processing of emotionalinformation occurs automatically, and does notdepend on available attentional resources(LeDoux, 1996; Morris, Ohman, & Dolan, 1999;Ohman, Flykt, & Esteves, 2001; Vuilleumier et al.,2001). Although the exact mechanisms that allowfor such automatic processing of emotional in-formation are not fully understood, direct sub-cortical pathways that reach the AMYindependently of the typical cortical connectionssubserving various sensorial modalities are im-plicated (Vuilleumier, 2005). This is consistentwith the evidence that emotional stimuli benefitfrom enhanced processing due to their ability to‘‘capture attention’’ and reallocate processingresources (Anderson & Phelps, 2001; LeDoux &Phelps, 2000; Vuilleumier, 2005). As a result ofthis ‘‘parallel processing’’ that allows for bothautomatic, nonconscious, preattentional proces-sing and enhanced conscious processing boostedby the engagement of attentional mechanisms,emotional stimuli are processed with increasedefficacy (see also Balconi, this issue 2011; Shaw,Lien, Ruthruff, & Allen, this issue 2011; Sutton &Altarriba, this issue 2011).
Strong brain imaging support for the idea thatemotion processing is independent of attentioncomes from an influential fMRI study byVuilleumier and colleagues (2001). In this study,attention was manipulated by asking subjects toattend either to pairs of houses or faces, whichwere presented in a four-picture display eccen-trically to a fixation point. The pictures in thehouse-pairs were either identical or different,whereas the faces were either fearful or neutral,and the subjects were asked to attend either tohouses or faces and to make same/differentjudgements. Supporting the view that emotionprocessing occurs automatically and indepen-dently of the attentional focus, fMRI resultsrevealed increased AMY activity to the fearfulfaces regardless of whether they were attended ornot. Also consistent with the traditional view,response times to houses were slower whenfearful faces were displayed as distractors.
Although much of the available evidencesupports the traditional view concerning theautomaticity of emotion processing, there is alsoevidence supporting an emerging competing view,
EMOTION, COGNITION, AND THE BRAIN 671
Dow
nloa
ded
by [
79.1
16.2
19.6
6] a
t 05:
41 0
5 M
ay 2
012

according to which emotion processing is notautomatic, but rather dependent on attentionalresources (Pessoa, 2005). Representative evi-dence for this view comes from work by Pessoaet al. (2002), which challenged Vuilleumier et al.’s(2001) findings and hence triggered an interestingdebate concerning the role of attention in earlyemotion processing (compare Pessoa et al. andVuilleumier et al.). The main methodologicalcriticism raised by Pessoa et al. was that previousstudies failed to reveal evidence for modulationof emotion processing by attention because thetasks used were not demanding enough to reducethe availability of processing resources to beengaged by emotional information*hence, thefindings supporting the automaticity of emotionprocessing.
To address this limitation, Pessoa et al. (2002)devised a more difficult task, in which subjectshad to fixate on centrally displayed faces (malesor females, with fearful or neutral expressions)and either make a gender judgement or specify iftwo peripherally displayed bars were oriented inthe same direction or not. Thus, similar toVuilleumier et al.’s (2001) task, the attentionalfocus was alternating between stimuli with orwithout emotional content. As expected, in thegender judgement condition (attending faces),fearful faces evoked greater response in a net-work of regions associated with emotion proces-sing, including the AMY. However, thisdifferential activation was not present whensubjects performed the more difficult peripheral(bar-orientation) task, and thus did not attend tofaces. Also, there were no differences in responsetimes related to the fearful expression of the facedistractors. Based on these findings, Pessoa et al.concluded that emotional stimuli can ‘‘captureattention’’ if there are enough attentional re-sources ‘‘to be captured’’ and not engaged byother tasks at hand.
Since their initial publication, when the evi-dence supporting the two views was seen inradical opposition (Pessoa et al., 2002; Vuilleu-mier et al., 2001), the positions of their propo-nents have became milder relative to thecompatibility of the two views (see Pessoa, 2005;Vuilleumier, 2005). Nevertheless, discussionsabout the automatic versus attention-dependentnature of emotion processing continue to be amatter of current debate. One possible cause forthis continuing debate is the fact that emotionalcontent and task demands were not systematicallyand impartially manipulated within the same
study. Studies providing support for automaticemotion processing could be criticised for notusing challenging enough tasks to deplete atten-tional resources, but those studies supporting theattention-dependent view could be criticised fornot using powerful enough emotional stimuli to‘‘capture’’ attention. The apparent inconsistencybetween the findings resulted from these investi-gations could be reconciled by studies concomi-tantly manipulating both the task demands andthe emotional content (Shafer & Dolcos, 2010).Such investigations may not only solve this debatebut could also provide evidence that the twoviews concerning basic emotion processing arenot mutually exclusive, and that depending onthe circumstances emotional information can beprocessed automatically and can also benefitfrom engaging attentional resources, if available(Shafer & Dolcos, 2010).
The impact of emotion on long-termmemory
Emotion has not only transient effects on cogni-tive processing, by influencing initial perceptualprocesses and attention paid to emotional stimulior to details surrounding emotional events, butalso long-lasting effects, which will eventuallylead to better memory for those events. Vividmemories for emotionally charged personalevents support this notion, but there is alsoempirical evidence that emotional events arebetter remembered than neutral events (e.g.,Bradley, Greenwald, Petry, & Lang, 1992; Chris-tianson, 1992). Emotional stimuli (including pic-tures, words, and faces) are better rememberedthan neutral stimuli, an effect that tends to besimilar for positive and negative stimuli. Thisfinding suggests that the memory-enhancementeffect of emotion is driven by emotional arousal(i.e., emotional intensity) rather than by emo-tional valence (Dolcos, LaBar, & Cabeza, 2006).Also, emotional memories tend to be accompa-nied by a sense of re-experiencing (Dolcos et al.,2005; Ochsner, 2000), and these recollectionbenefits are also augmented relative to neutralmemories over time (Anderson, Yamaguchi,Grabski, & Lacka, 2006; Ritchey, Dolcos, &Cabeza, 2008; Sharot & Yonelinas, 2008).
Of particular interest are the neural mechan-isms believed to make emotional events morememorable, and the extent to which the brain
672 DOLCOS, IORDAN, DOLCOS
Dow
nloa
ded
by [
79.1
16.2
19.6
6] a
t 05:
41 0
5 M
ay 2
012

regions involved in initial stages of processingalso contribute to the long-term effects of emo-tion. The modulation hypothesis, inspired fromanimal research (McGaugh, 2000, 2002, 2004),suggests that emotional events are rememberedbetter than neutral events because the AMYenhances the function of memory-related brainregions.1 Recent studies involving event-relatedfMRI in humans provide further support for thisidea (Dolcos, LaBar, & Cabeza, 2004b; Kensinger& Corkin, 2004; Kilpatrick & Cahill, 2003;Richardson, Strange, & Dolan, 2004; Ritcheyet al., 2008; Sergerie, Lepage, & Armony, 2006).These studies are particularly relevant becausethey involved methods that allowed identificationof (1) memory-specific brain activity from activityassociated with more general perceptual proces-sing, as a result of using the so-called subsequentmemory paradigm (SMP) (Dolcos et al., 2004b;Kensinger & Corkin, 2004; Paller & Wagner,2002; Ritchey et al., 2008; Sergerie et al., 2006;Shafer, Iordan, Cabeza, & Dolcos, 2011) (alsoreviewed in Dolcos, 2010; Dolcos & Denkova,2008; Dolcos, LaBar, & Cabeza, 2006; LaBar &Cabeza, 2006), (2) nearby medial temporal lobe(MTL) subregions involved in the memory-en-hancing effect of emotion, as a result of using theso-called anatomical region of interest (ROI)approach (Dolcos et al., 2004b), and (3) supportfor the idea that the memory-enhancing effect ofemotion involves interactions between emotion-based (AMY) and memory-based brain regions(Dolcos et al., 2004b; Kilpatrick & Cahill, 2003;Richardson et al., 2004; Ritchey et al., 2008).
By using the SMP in conjunction with fMRI,we showed that emotion enhances long-termepisodic memory (i.e., memory for personalevents) by modulating activity in two mainmemory-related brain regions, i.e., the MTLmemory system (Dolcos et al., 2004b) and theprefrontal cortex (PFC) (Dolcos, LaBar, & Ca-beza, 2004a). Further, we showed that theseeffects can occur during one or more memorystages, including the initial encoding of an emo-tional event (Dolcos & Cabeza, 2002; Dolcos,Graham, LaBar, & Cabeza, 2003; Dolcos et al.,
2004a, b) and the retrieval and reexperiencing ofemotional memories (Dolcos et al., 2005). Forinstance, one of our studies focusing on the role ofthe AMY and MTL memory system (Dolcos etal., 2004b) during encoding yielded two mainfindings supporting the modulation hypothesisin neurologically intact humans (also reviewedin Dolcos, 2010; Dolcos & Denkova, 2008; Dolcos,LaBar, & Cabeza, 2006). Using anatomicallydefined ROI to precisely quantify the MTLactivity, and the SMP to identify brain activityspecifically predicting memory performance, wefound enhanced memory for emotional informa-tion to be associated with enhanced memory-related activity in (and interactions between) theAMY and the MTL memory regions. Consistentwith animal findings, the greatest differencesbetween the emotional and the neutral encodingsuccess activity were found in the basolateralnucleus of the AMY (BLA), the anterior part ofthe hippocampus (HC) and the entorhinal cortex(EC). Moreover, encoding success activity inAMY and MTL memory regions was morestrongly correlated with each other for emotionalthan neutral stimuli. This finding suggests thatthese regions interact more intimately during theencoding of emotional stimuli than duringthe encoding of neutral stimuli, thus providingfurther support for the modulation hypothesis(McGaugh, 2002).
Evidence from a recent study involving reten-tion interval manipulations also provided insightinto the effect of emotion consolidation, byshowing that AMY�MTL interactions initiatedduring encoding contribute to the persistence ofemotional memories over time (Ritchey et al.,2008). In this study, participants were scannedwhile encoding negative and neutral pictures,then postscanning recognition memory was testedafter a short delay (20 min after encoding) and along delay (1 week after encoding). This manip-ulation revealed that AMY�MTL connectivityduring encoding was greater for items retrievedafter a longer delay, thus suggesting that AMY�MTL interactions underlie consolidation andpersistence of the emotional memories overtime (Ritchey et al., 2008).
Studies of memory retrieval also showed thatemotional memory is particularly resilient totime, with laboratory enhancements being re-ported up to 1 year after encoding (Dolcoset al., 2005). Our retrieval study (Dolcos et al.,2005) provided behavioural and functional neu-roimaging evidence concerning the effect of
1Another influential hypothesis concerning the specific
role of the AMY is the so-called plasticity hypothesis (Dolcos
& Denkova, 2008), which has also been formulated based on
animal studies (LeDoux, 2000). This hypothesis proposes that
the AMY itself is a site of plasticity underlying learning and
memory of fear conditioning. For a comparison of both
hypotheses see Dolcos and Denkova (2008).
EMOTION, COGNITION, AND THE BRAIN 673
Dow
nloa
ded
by [
79.1
16.2
19.6
6] a
t 05:
41 0
5 M
ay 2
012

emotional arousal on memory retrieval processesafter a longer retention interval than typicallyinvolved in laboratory-based studies (minutes,days, weeks) (e.g., Cahill, Babinsky, Markowitsch,& McGaugh, 1995; LaBar & Phelps, 1998). In thisstudy, participants encoded high-arousing emo-tional (pleasant and unpleasant) and low-arousingneutral pictures, and 1 year later memory wastested while undergoing fMRI. Notably, subjectsperformed a recognition task that distinguishesbetween recollection-based and familiarity-basedresponses (Tulving, 1985). One year after initialencoding, emotionally arousing pictures wereremembered better and elicited greater recollec-tion than neutral pictures, and fMRI resultsindicated that emotional content enhances activ-ity in the AMY and MTL memory systems (HCand EC) related to successful retrieval of indivi-dual items from long-term storage (hits vs.misses). These findings support the idea thatsuccessful retrieval of emotional memories in-volves MTL mechanisms similar to those identi-fied during successful emotional encoding (Cahillet al., 1996; Dolcos et al., 2004b). Also, in theAMY and HC (but not in the EC), the emotioneffect was greater for recollection than forfamiliarity, which provided initial evidence con-cerning the neural correlates of the differentialeffect of emotion on these two types of memoryretrieval (Ochsner, 2000).
Although the majority of studies have focusedon the interaction between the emotion-basedAMY system and memory-related MTL system,the role of the PFC in emotional memory forma-tion has also been investigated. The extant evi-dence concerning the role of the PFC in emotionalmemory encoding suggests both arousal-driveneffects (e.g., Dolcos et al., 2004a; Sergerie, Lepage,& Armony, 2005) and valence-driven effects (e.g.,Kensinger & Corkin, 2004). For instance, we foundthat emotional arousal enhanced successful en-coding activity (Dm effect) in two subregions ofthe left lateral PFC (i.e., one dorsolateral*Brod-man Area [BA] 9/6 and the other ventrolateral*BA 47), in which the Dm effect for highly arousingpictorial items (both pleasant and unpleasant) wasgreater than for low arousing and neutral items(Dolcos et al., 2004a). Similarly, using a wordencoding task with high- and low-arousing nega-tive and neutral stimuli, Kensinger and Corkin(2004) also obtained a regionally specific modula-tion of PFC activity by valence. These findings arefurther supported by studies providing evidencefor AMY�PFC interactions during emotional
memory formation (e.g., Canli, Desmond, Zhao,& Gabrieli, 2002; Kilpatrick & Cahill, 2003).Given the role of left ventrolateral PFC (vlPFC)regions in semantic processing (Poldrack et al.,1999) and the role of the dorsolateral PFC (dlPFC)regions in working memory operations (D’Espo-sito, Postle, & Rypma, 2000), these findingssuggest that arousing events enhance memory inpart by promoting semantic (vlPFC) and workingmemory (dlPFC) processing.
Finally, complementary evidence concerningthe mechanisms underlying the formation ofemotional memories comes from studies investi-gating the timing of processing linked to thememory-enhancing effect of emotion (e.g., Dol-cos & Cabeza, 2002; Palomba, Angrilli, & Mini,1997). Using event-related potential (ERP) re-cordings during encoding, in conjunction with theSMP, we identified that the emotional Dm effectoccurs faster than the neutral Dm (Dolcos &Cabeza, 2002): Emotional Dm was greater duringan early epoch (400�600 ms poststimulus onset),whereas the Dm effects were similar for emo-tional and neutral stimuli during a late epoch(600�800 ms). Although given the spatial resolu-tion of ERPs it was difficult to identify the neuralgenerators of this effect, faster emotional Dmsuggests that emotional stimuli have privilegedaccess to processing resources leading to subse-quent memory, which could be another mechan-ism conferring a memory advantage to emotionalinformation (Dolcos & Cabeza, 2002).
In sum, the previously mentioned studiesprovide strong evidence for the notion thatemotion enhances long-term episodic memoryby modulating activity in two main memory-related brain regions, the MTL memory systemand the PFC, which also benefits from fasterencoding. The effects of emotion on MTL andPFC regions, however, may be related to differentmechanisms. The evidence concerning the func-tions typically attributed to these brain regions(Moscovitch, 1992; Simons & Spiers, 2003) sug-gests that in the MTL emotion enhances basicencoding, consolidation, storage, and retrieval ofmemory representations, whereas in the PFCemotion enhances semantic, strategic, and work-ing memory processes. These led to the proposalthat AMY and MTL are part of basic/directneurohormonal mechanisms underlying the mem-ory-enhancement effect of emotion, whereasPFC has an indirect/mediated involvement inthe formation of emotional memories, by enhan-cing strategic, semantic, and working memory
674 DOLCOS, IORDAN, DOLCOS
Dow
nloa
ded
by [
79.1
16.2
19.6
6] a
t 05:
41 0
5 M
ay 2
012

processes (LaBar & Cabeza, 2006) (Figure 1).
Finally, evidence also suggests that overall the
memory-enhancing effect of emotion is driven by
the intensity of emotional events, rather than by
their valence, although there is evidence that
valence is also a contributing factor (Kensinger
& Corkin, 2004; Mickley Steinmetz & Kensinger,
2009; Shaw, Brierley, & David, 2005).
The impact of emotion on workingmemory and decision making
In addition to influencing initial stages of proces-
sing and their long-term consequences (reviewed
previously), emotional information also impacts
other higher level processes, such as working
memory and decision making, which are essential
to goal-directed behaviour. Evidence concerning
the impact of emotion on these executive pro-
cesses will be discussed in turn.
Working memory. Working memory (WM) isinvolved in the active maintenance and manip-ulation of task-relevant information (Baddeley,1996). As suggested earlier, maintaining emo-tional information in WM may be one of themechanisms contributing to the enhancing effectsof emotion on long-term memory (Dolcos et al.,2004a; Kensinger & Corkin, 2004). Although thismay imply that emotional stimuli also produce anenhancing effect on WM, the evidence fromstudies investigating this matter by comparingWM performance for emotional versus neutralstimuli did not yield conclusive findings (Ken-singer & Corkin, 2003). Instead, emerging evi-dence concerning the impact of emotion on WMand the associated neural correlates comes fromstudies in which emotional stimuli are presentedas task-irrelevant distractors (Chuah et al., 2010;Denkova et al., 2010; Dolcos et al., 2008; Dolcos& McCarthy, 2006; Wong et al., in press).
Distraction challenges our ability to maintainfocus on goal-relevant information and thus mayimpair cognitive performance. The existence ofneural mechanisms that allow for privilegedaccess of emotional information to processingresources raises the possibility that emotionalinformation may also be a very potent distraction,particularly when task irrelevant. However, upuntil recently, the neural correlates of the detri-mental effects of emotion on cognitive functionshad received relatively less attention (Johnson etal., 2005; Most, Chun, Widders, & Zald, 2005). Adefault assumption, based on the findings con-cerning the enhancing effect of emotion onmemory (where emotion enhanced activity inmemory-related brain regions), is that the impair-ing effect of emotion may be linked to reducedactivity in brain regions subserving the functionsimpaired by emotion. This assumption is sup-ported by evidence from both clinical and non-clinical groups.
Models of affective�cognitive interactions in-spired by clinical studies point to dysfunctionalinteractions between a dorsal executive neuralsystem and a ventral affective system, and pro-pose that impaired executive control and en-hanced emotional distractibility observed indepression are linked to hypofunction of thedorsal executive and hyperfunction of the ventralaffective systems (Drevets & Raichle, 1998; May-berg, 1997) (Figure 2). The dorsal system includesbrain regions typically associated with ‘‘cold’’executive (ColdEx) functions, such as the dlPFCand the lateral parietal cortex (LPC), which are
Figure 1. Main mechanisms involved in the memory-enhan-
cing effect of emotion. Emotion enhances long-term episodic
memory by modulating activity in two main memory-related
brain regions, the medial-temporal lobe (MTL) memory
system and the prefrontal cortex (PFC). However, the effects
of emotion on MTL and PFC regions may be related to
different mechanisms: AMY and MTL are part of basic/direct
neurohormonal mechanisms underlying the memory-enhance-
ment effect of emotion (bottom-up mechanism), whereas PFC
is part of a mechanism (also including the parietal cortex*PC) that has an indirect/mediated involvement in the forma-
tion of emotional memories, by enhancing strategic, semantic,
working memory, and attentional processes (top-down me-
chanism). Adapted from LaBar and Cabeza (2006); courtesy
of Dr. Roberto Cabeza. [To view this figure in colour, please
visit the online version of this Journal.]
EMOTION, COGNITION, AND THE BRAIN 675
Dow
nloa
ded
by [
79.1
16.2
19.6
6] a
t 05:
41 0
5 M
ay 2
012
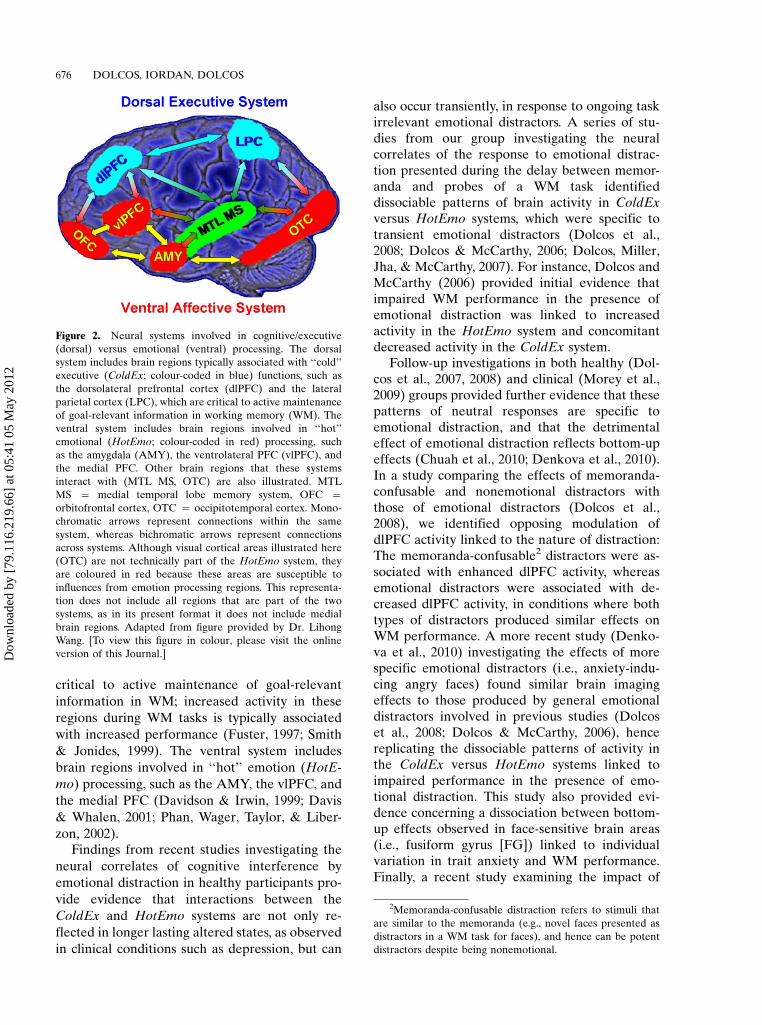
critical to active maintenance of goal-relevant
information in WM; increased activity in these
regions during WM tasks is typically associated
with increased performance (Fuster, 1997; Smith
& Jonides, 1999). The ventral system includes
brain regions involved in ‘‘hot’’ emotion (HotE-
mo) processing, such as the AMY, the vlPFC, and
the medial PFC (Davidson & Irwin, 1999; Davis
& Whalen, 2001; Phan, Wager, Taylor, & Liber-
zon, 2002).Findings from recent studies investigating the
neural correlates of cognitive interference by
emotional distraction in healthy participants pro-
vide evidence that interactions between the
ColdEx and HotEmo systems are not only re-
flected in longer lasting altered states, as observed
in clinical conditions such as depression, but can
also occur transiently, in response to ongoing taskirrelevant emotional distractors. A series of stu-dies from our group investigating the neuralcorrelates of the response to emotional distrac-tion presented during the delay between memor-anda and probes of a WM task identifieddissociable patterns of brain activity in ColdExversus HotEmo systems, which were specific totransient emotional distractors (Dolcos et al.,2008; Dolcos & McCarthy, 2006; Dolcos, Miller,Jha, & McCarthy, 2007). For instance, Dolcos andMcCarthy (2006) provided initial evidence thatimpaired WM performance in the presence ofemotional distraction was linked to increasedactivity in the HotEmo system and concomitantdecreased activity in the ColdEx system.
Follow-up investigations in both healthy (Dol-cos et al., 2007, 2008) and clinical (Morey et al.,2009) groups provided further evidence that thesepatterns of neutral responses are specific toemotional distraction, and that the detrimentaleffect of emotional distraction reflects bottom-upeffects (Chuah et al., 2010; Denkova et al., 2010).In a study comparing the effects of memoranda-confusable and nonemotional distractors withthose of emotional distractors (Dolcos et al.,2008), we identified opposing modulation ofdlPFC activity linked to the nature of distraction:The memoranda-confusable2 distractors were as-sociated with enhanced dlPFC activity, whereasemotional distractors were associated with de-creased dlPFC activity, in conditions where bothtypes of distractors produced similar effects onWM performance. A more recent study (Denko-va et al., 2010) investigating the effects of morespecific emotional distractors (i.e., anxiety-indu-cing angry faces) found similar brain imagingeffects to those produced by general emotionaldistractors involved in previous studies (Dolcoset al., 2008; Dolcos & McCarthy, 2006), hencereplicating the dissociable patterns of activity inthe ColdEx versus HotEmo systems linked toimpaired performance in the presence of emo-tional distraction. This study also provided evi-dence concerning a dissociation between bottom-up effects observed in face-sensitive brain areas(i.e., fusiform gyrus [FG]) linked to individualvariation in trait anxiety and WM performance.Finally, a recent study examining the impact of
Figure 2. Neural systems involved in cognitive/executive
(dorsal) versus emotional (ventral) processing. The dorsal
system includes brain regions typically associated with ‘‘cold’’
executive (ColdEx; colour-coded in blue) functions, such as
the dorsolateral prefrontal cortex (dlPFC) and the lateral
parietal cortex (LPC), which are critical to active maintenance
of goal-relevant information in working memory (WM). The
ventral system includes brain regions involved in ‘‘hot’’
emotional (HotEmo; colour-coded in red) processing, such
as the amygdala (AMY), the ventrolateral PFC (vlPFC), and
the medial PFC. Other brain regions that these systems
interact with (MTL MS, OTC) are also illustrated. MTL
MS � medial temporal lobe memory system, OFC �orbitofrontal cortex, OTC � occipitotemporal cortex. Mono-
chromatic arrows represent connections within the same
system, whereas bichromatic arrows represent connections
across systems. Although visual cortical areas illustrated here
(OTC) are not technically part of the HotEmo system, they
are coloured in red because these areas are susceptible to
influences from emotion processing regions. This representa-
tion does not include all regions that are part of the two
systems, as in its present format it does not include medial
brain regions. Adapted from figure provided by Dr. Lihong
Wang. [To view this figure in colour, please visit the online
version of this Journal.]
2Memoranda-confusable distraction refers to stimuli that
are similar to the memoranda (e.g., novel faces presented as
distractors in a WM task for faces), and hence can be potent
distractors despite being nonemotional.
676 DOLCOS, IORDAN, DOLCOS
Dow
nloa
ded
by [
79.1
16.2
19.6
6] a
t 05:
41 0
5 M
ay 2
012

sleep deprivation on the response to emotionaldistraction (Chuah et al., 2010) also pointed tobottom-up effects of emotional distraction, byshowing that increased distraction by negativeemotional pictures following sleep deprivationwas associated with increased AMY activationand reduced functional connectivity between thisregion and cognitive control brain regions.
Collectively, these findings provide strongevidence that the outcome of task-irrelevantemotional distraction depends on dynamic inter-actions between neural systems that allow theability to stay focused on task-relevant informa-tion and systems involved in the processing ofemotional information that may compete with theavailable processing resources. Possibly as a resultof their salience, emotional distractors may pro-duce a bottom-up impact on processing of goal-relevant information by reallocating processingresources (Vuilleumier et al., 2001) and impairingperformance. However, as we will see later, thedisadvantageous outcomes of this bottom-up im-pact of emotional distraction can be mitigated by‘‘top-down’’ interventions from cognitive controlregions, which are engaged to regulate emotionalresponses and cope with emotional distraction(Dolcos, Kragel, et al., 2006; Gray, Braver, &Raichle, 2002; Pessoa, 2008).
Decision making. Recently, the fact that emo-tion provides essential input for high-level com-plex processes, like decision making, has becomemore largely accepted. Initially developed toaccount for problems of decision making dis-played by patients with ventromedial PFC(vmPFC) damage, the influential so-called ‘‘so-matic marker hypothesis’’ (SMH) (Damasio,1994, 1996) argues that decision making is aprocess shaped by multiple sources of input, notonly cognitive, but also somatic-related feedbackwith emotional/motivational relevance.
Generally speaking, a somatic marker is a typeof nonspecific ‘‘feeling’’ that has been linkedthrough learning to predicted outcomes in thefuture (Damasio, 1994, 1996). Somatic markerscan be related to either primary inducers (e.g.,seeing a snake or losing a large sum of money) orsecondary inducers (e.g., the perspective of losinga large sum of money), which send a biasing signalthat crudely affects choice (Damasio, 1995).Using the Iowa Gambling Task (IGT; Bechara,Damasio, Damasio, & Anderson, 1994; Bechara,Tranel, Damasio, & Damasio, 1996), Damasio andcolleagues showed that successful performance in
the IGT is related to the ability to displayanticipatory skin conductance responses (i.e.,somatic markers), which are typically present innormal subjects but absent in patients withvmPFC/orbitofrontal cortex (OFC) damage. Theimplicit ability to use these responses in makingadvantageous decisions (Bechara, Damasio, Tra-nel, & Damasio, 1997) suggests that emotion-related signals are important for decision makingeven when they reside at a nonconscious level.The SMH underlines the contributions of theAMY and vmPFC as components of a neuralnetwork with an essential role in judgement anddecision making (Damasio, 1994). However, thecontributions of the two structures are at differentlevels*not only do both of them contribute to theintegration of exteroceptive and interoceptivesomatic/emotional information, but also AMY iscritical for processing primary inducers and OFCis critical for processing secondary inducers (Be-chara, Damasio, & Damasio, 2003).
Emotional influences on decision-making pro-cesses is further supported by fMRI studies usingthe ultimatum game (e.g., Sanfey, Rilling, Aron-son, Nystrom, & Cohen, 2003), whereby twoplayers respond to fair and unfair proposals tosplit money. Unfair offers elicited activity in brainareas related to both emotion (anterior insula)and cognition (dlPFC). If the decisions are madepurely on cognitive grounds, acceptance of eventhe most unfair offers would lead to gains,whereas in case of rejected offers neither of theplayers receives anything. Typically, this does nothappen, as the tendency for players is to rejectunfair offers, despite a detriment to their owngains. Interestingly, activation in the anteriorinsula (which is commonly associated with pro-cessing of negative emotions, particularly disgust),increased for rejected offers, suggesting that inthose cases emotion ‘‘overruled’’ cognitive pro-cesses, thus highlighting an important role ofemotional responses in decision making (seealso Nguyen et al., this issue 2011).
THE EFFECT OF COGNITION ONEMOTION PROCESSING
Up to this point, we have reviewed the impact ofemotion on cognitive processing. In this section,we focus on the reciprocal influence, namely theeffect of cognition on emotion processing. Princi-pally, we review evidence from studies of emotion
EMOTION, COGNITION, AND THE BRAIN 677
Dow
nloa
ded
by [
79.1
16.2
19.6
6] a
t 05:
41 0
5 M
ay 2
012

regulation and investigations of the neural corre-lates of coping with emotional distraction.
Emotion regulation*background
Understanding the mechanisms by which cogni-tion affects emotion is the focus of a recentlyemerging field investigating the neural under-pinnings of cognitive control of emotion (oremotion regulation), which is an important copingmechanism in the face of emotional stressors(Ochsner & Gross, 2005). Despite recent progressin identifying the neural correlates of emotionregulation, a number of questions remain. Chiefamong these is the role of different types ofemotional control and their associated neuralcorrelates. Most of the previous emotion regula-tion research has focused on conscious/deliberateforms of regulation, such as instructed reappraisal(Kim & Hamann, 2007; Ochsner, Bunge, Gross, &Gabrieli, 2002), suppression (Jackson et al., 2003),and cognitive distraction (Denkova, Chakrabarty,Dolcos, & Dolcos, in press; Levesque et al., 2003).A large body of evidence shows that the delib-erative forms of emotion regulation broadlyinvolve the PFC, especially the ventrolateralaspects of the PFC, as well as the posteriorportion of the dorsomedial PFC (dmPFC)/super-ior frontal gyrus (BA 6/8) (Goldin, McRae,Ramel, & Gross, 2008; Ochsner et al., 2002). Inaddition, a few studies also found activation in thedlPFC (Goldin et al., 2008; Levesque et al., 2003),and there is inconsistent support for more poster-ior regions such as the posterior temporal gyri andinferior parietal lobules (Goldin et al., 2008;Ochsner et al., 2004).
Different than deliberate emotion regulation,the nonconscious/automatic forms of emotionregulation and the associated neural correlateshave received relatively little empirical attention(cf. Mauss, Cook, & Gross, 2007). Nonconsciousemotion regulation is an important part of ourbehaviour that, similar to conscious forms ofemotional control, undergoes ontogenetic trans-formations. Although emotions are displayed veryearly in ontogeny, the ability to control them isnot fully developed in early childhood andadolescence. As we age, we learn that displayingcertain emotions may not be appropriate inspecific contexts, and often we find ourselvesautomatically adjusting, or regulating, our emo-tional responses to fit the current circumstances.Recent evidence suggests that nonconscious emo-
tion regulation could be as effective as deliberateemotion regulation, while being less costly be-cause of the automatic effortless engagement(Bargh & Williams, 2007). However, becauseprevious research tended to separately investigatethe two forms of emotion regulation (Goldin etal., 2008; Mauss et al., 2007) it is not clear whetherthey are equally effective in controlling theresponse to emotionally challenging situations,or whether they are differentially effective de-pending on the intensity of emotional challenge.Moreover, brain imaging studies investigating theneural correlates of nonconscious emotion regu-lation are generally lacking (but see Dolcos, Sung,Denkova, Dixon, & Dolcos, in press).
Although the brain imaging studies of emotionregulation mentioned above provided evidence ofincreased activity in cognitive control regions as aresult of engaging emotion regulation strategies,these studies could not establish the precise roleof these regions in controlling the impact ofemotion on ongoing cognitive processes. On theother hand, brain imaging studies in which emo-tional information was presented as task-irrele-vant distraction provided such evidence byidentifying neural correlates of coping with dis-tracting emotions. In the next section, we willfocus on findings from such studies highlightingthe role of lateral and medial PFC regions.
Cognitive control of emotionaldistraction
Understanding the impact of cognitive control onemotion processing requires identification of theneural mechanisms associated with the ability tocope with distracting emotions. A series ofinvestigations from our group (Chuah et al.,2010; Denkova et al., 2010; Dolcos et al., 2008;Dolcos, Kragel, et al., 2006; Dolcos & McCarthy,2006) provided evidence that coping with task-irrelevant emotional distraction entails increasedactivity in and interactions between brain regionsinvolved in basic emotion processing (the AMY)and brain regions associated with cognitive con-trol (particularly lateral and medial PFC). Theengagement of the AMY can be seen as havingthe role of an ‘‘emotional detector’’ that signalsthe cognitive control regions the presence ofpotentially distracting, emotional stimuli andthus the need to control their possible detri-mental effects on cognitive performance (Dolcos,
678 DOLCOS, IORDAN, DOLCOS
Dow
nloa
ded
by [
79.1
16.2
19.6
6] a
t 05:
41 0
5 M
ay 2
012
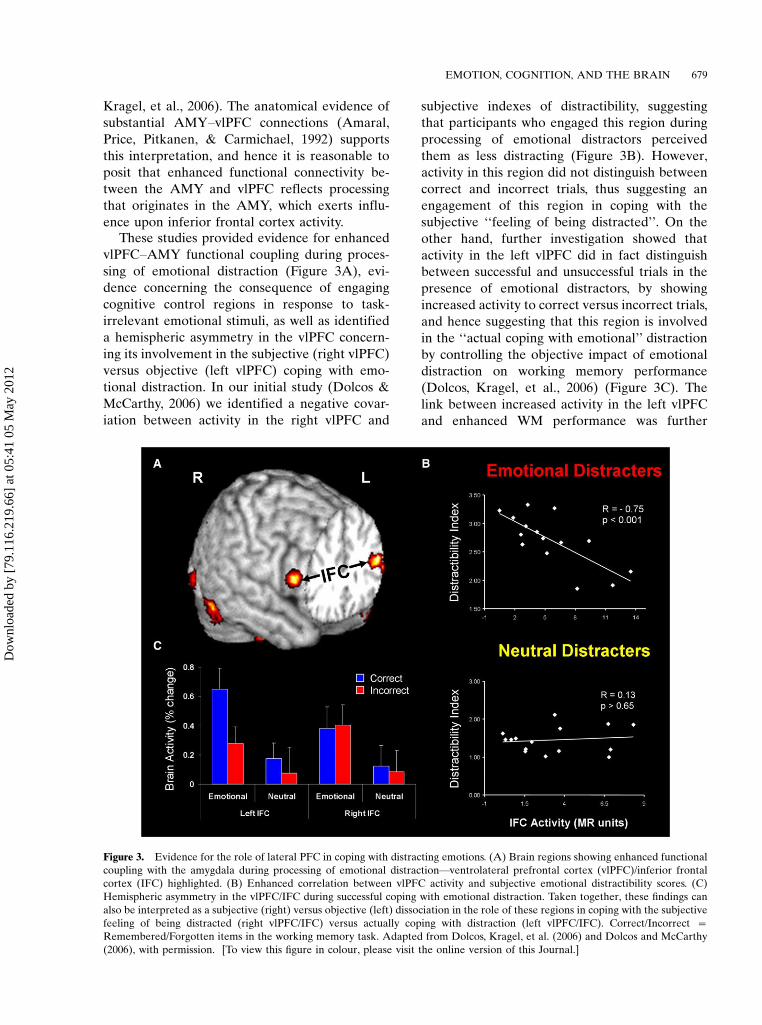
Kragel, et al., 2006). The anatomical evidence of
substantial AMY�vlPFC connections (Amaral,
Price, Pitkanen, & Carmichael, 1992) supports
this interpretation, and hence it is reasonable to
posit that enhanced functional connectivity be-
tween the AMY and vlPFC reflects processing
that originates in the AMY, which exerts influ-
ence upon inferior frontal cortex activity.These studies provided evidence for enhanced
vlPFC�AMY functional coupling during proces-
sing of emotional distraction (Figure 3A), evi-
dence concerning the consequence of engaging
cognitive control regions in response to task-
irrelevant emotional stimuli, as well as identified
a hemispheric asymmetry in the vlPFC concern-
ing its involvement in the subjective (right vlPFC)
versus objective (left vlPFC) coping with emo-
tional distraction. In our initial study (Dolcos &
McCarthy, 2006) we identified a negative covar-
iation between activity in the right vlPFC and
subjective indexes of distractibility, suggesting
that participants who engaged this region during
processing of emotional distractors perceived
them as less distracting (Figure 3B). However,
activity in this region did not distinguish between
correct and incorrect trials, thus suggesting an
engagement of this region in coping with the
subjective ‘‘feeling of being distracted’’. On the
other hand, further investigation showed that
activity in the left vlPFC did in fact distinguish
between successful and unsuccessful trials in the
presence of emotional distractors, by showing
increased activity to correct versus incorrect trials,
and hence suggesting that this region is involved
in the ‘‘actual coping with emotional’’ distraction
by controlling the objective impact of emotional
distraction on working memory performance
(Dolcos, Kragel, et al., 2006) (Figure 3C). The
link between increased activity in the left vlPFC
and enhanced WM performance was further
Figure 3. Evidence for the role of lateral PFC in coping with distracting emotions. (A) Brain regions showing enhanced functional
coupling with the amygdala during processing of emotional distraction*ventrolateral prefrontal cortex (vlPFC)/inferior frontal
cortex (IFC) highlighted. (B) Enhanced correlation between vlPFC activity and subjective emotional distractibility scores. (C)
Hemispheric asymmetry in the vlPFC/IFC during successful coping with emotional distraction. Taken together, these findings can
also be interpreted as a subjective (right) versus objective (left) dissociation in the role of these regions in coping with the subjective
feeling of being distracted (right vlPFC/IFC) versus actually coping with distraction (left vlPFC/IFC). Correct/Incorrect �Remembered/Forgotten items in the working memory task. Adapted from Dolcos, Kragel, et al. (2006) and Dolcos and McCarthy
(2006), with permission. [To view this figure in colour, please visit the online version of this Journal.]
EMOTION, COGNITION, AND THE BRAIN 679
Dow
nloa
ded
by [
79.1
16.2
19.6
6] a
t 05:
41 0
5 M
ay 2
012

confirmed by other studies (Dolcos et al., 2008;Denkova et al., 2010). For instance, in anotherrecent study we showed that increased activity inthe left vlPFC was linked to increased WMperformance in the presence of emotional, butnot neutral, distractors (Denkova et al., 2010).
Taken together, these investigations providedevidence linking the engagement of activity inthese lateral and medial PFC regions not only incontrolling the subjective emotional responseinduced by potentially distracting emotional sti-muli (Ochsner & Gross, 2005) but also in dimin-ishing the actual negative impact of distractingemotions on ongoing cognitive processes (Chuahet al., 2010; Denkova et al., 2010; Dolcos et al.,2008; Dolcos, Kragel, et al., 2006). Moreover, theextant evidence also suggests that this engage-ment involves enhanced functional coupling withactivity in the AMY (Chuah et al., 2010; Dolcos,Kragel, et al., 2006).
THE ROLE OF INDIVIDUALDIFFERENCES IN EMOTION�COGNITION INTERACTIONS
An important aspect in the investigation ofemotion�cognition interactions is identificationof individual differences that may affect theresponse to emotional challenge and thus thedifferential vulnerability to affective disorders. Aspresented in the previous parts, the interactionsbetween emotion and cognition are apparent atdifferent levels of analysis of human behaviour.The final output of these interactions defineshuman behaviour as a whole, and is reflected inindividual differences, which in the domain ofemotion processing seems to be the rule ratherthan the exception (Eugene et al., 2003). As such,all the effects that have been discussed so far maybe influenced by individual differences in theresponse to emotional and cognitive challenge. Agiven emotional situation can evoke a wide rangeof emotional responses across individuals. Thissuggests that, not only are our responses toemotional contexts influenced by present emo-tional states, but also our behaviour in the face ofemotional challenge is influenced by our person-ality traits.
In this context, it is important to emphasisethat the response to emotional challenge, and thusvulnerability to affective disorders (i.e., depres-sion- and anxiety-related), depend on individual
variation in both emotional and cognitive do-mains, and that the final behavioural outcomedepends on the overall ‘‘affective�cognitive pro-file’’. It is known, for instance, that the negativeaffective bias in processing emotional informa-tion, the increased susceptibility to emotionaldistraction, and the affective dysregulation ob-served in affective disorders are also accompaniedby overall impaired cognitive/executive control(e.g., Bishop, 2009; Eysenck, Derakshan, Santos,& Calvo, 2007). Also, these changes could affectnot only activity in higher level cognitive controlbrain regions but also in lower level perceptualprocessing regions (e.g., Pujol et al., 2009).Investigation of individual differences is alsocritical for elucidating the neural bases of thesephenomena, as it has become apparent thatfinding cures for these clinical conditions dependsnot only on integrative understanding of thefactors that influence the susceptibility to emo-tional and cognitive challenge, but also alterationsof the brain mechanisms that accompany patho-logical changes in the emotion�cognition interac-tions.
In this section, we review behavioural andfMRI findings from studies investigating the roleof individual characteristics, such as personality-,sex-, and age-related, in mediating the interac-tions between emotion and cognition. Investiga-tion of the role of such factors that mediate theresponse to emotional challenge may provecritical for understanding affective disorders,which are characterised by increased emotionalsensitivity and distractibility, have increased in-cidence in women, and are associated withdysfunctional emotion regulation or emotiondysregulation. Earlier brain imaging investiga-tions have linked individual variation in person-ality measures reflecting general emotionprocessing (e.g., extraversion and neuroticism)to brain activity associated with general proces-sing of emotional information (e.g., Canli &Amin, 2002). However, more recent studies(e.g., Denkova et al., 2010) also provided evi-dence linking individual variation in personalitytraits reflecting specific emotions (social anxiety)to neural changes in responses to transientlyinduced specific emotions (e.g., anxiety-inducingdistraction). Similarly, studies investigating sex-related differences in emotion processing re-ported differences in brain activity associatedwith enhanced emotional reactivity and emo-tional memory in females compared to males(Cahill, Uncapher, Kilpatrick, Alkire, & Turner,
680 DOLCOS, IORDAN, DOLCOS
Dow
nloa
ded
by [
79.1
16.2
19.6
6] a
t 05:
41 0
5 M
ay 2
012

2004; Canli, Desmond, et al., 2002; Lang, Green-wald, Bradley, & Hamm, 1993). Finally, recentstudies investigating changes in emotion proces-sing associated with ageing point to a functionalshift consistent with a positivity bias in healthyageing (Carstensen, Mikels, & Mather, 2006;Mather & Carstensen, 2005), possibly linked toenhanced ability to control emotions (St. Jacques,Dolcos, & Cabeza, 2009; Dolcos, & Cabeza,2010), as opposed to emotion dysregulation ob-served in affective disorders. Evidence concerningthe role of individual differences in these threemain aspects in the context of investigating theneural correlates of emotion processing, in general,and of emotion�cognition interactions, in particular,will now be discussed in turn.
The role of personality-relateddifferences
Recent studies have shown that personality traits,such as extraversion and neuroticism, influencethe neural correlates of emotion processing acrossa range of emotional processes, including experi-ence, perception, attention, and memory (Canli,Sivers, Whitfield, Gotlib, & Gabrieli, 2002; Ha-mann & Canli, 2004; Touryan et al., 2007). It hasalso been suggested that individual differences inemotional biases linked to personality might berooted in an attentional network driven primarilyby AMY reactivity during the encoding of emo-tional stimuli (Haas & Canli, 2008). Extraversionand neuroticism have both been associated withattentional biases (Derryberry & Reed, 1994) butin opposite directions (Rusting, 1999). For exam-ple, individuals higher on extraversion exhibitgreater attention towards positive- rather thannegative-valenced emotional stimuli (Derryberry& Reed, 1994), coupled with longer reactiontimes when responding to positive relative toneutral words (Haas, Omura, Amin, Constable,& Canli, 2006). Functional MRI studies havedemonstrated that regions of the PFC and theAMY exhibit differential responses to emotionalstimuli in individuals who describe themselves ashighly extraverted versus neurotic (Canli &Amin, 2002; Canli et al., 2001). In those studies,extraversion was positively correlated with theactivity of the lateral and medial PFC (includingthe cingulate gyrus) and the AMY in response toviewing positive (vs. negative) stimuli. Neuroti-cism, in contrast, was positively correlated with
the activity in the PFC when viewing negative (vs.positive) stimuli. Extraversion and neuroticismalso affect recollection of personal autobiogra-phical memories. In a recent behavioural study wefound that extraversion contributed to remember-ing more positive personal experiences and tomaintaining a positive state, while neuroticismpredicted the phenomenological characteristics ofnegative autobiographical memories (Denkova,Dolcos, & Dolcos, 2011).
Personality-related individual differences werealso identified in conditions where emotionalinformation was presented as task-irrelevant dis-traction. Based on recent findings suggestingindividual variation in the response to emotionaldistraction (Dolcos et al., 2008), in a series offollow-up studies we investigated the factors thatmay underlie these differences. For instance, aninvestigation of task performance in conjunctionwith personality data (Dolcos, 2009) revealed thatparticipants that were more susceptible to emo-tional distraction in the WM task not only tendedto perceive emotional distractors as more emo-tional (as measured by subjective ratings of thedistractors) but also to have higher scores forattentional impulsivity (as measured with theBarratt Impulsiveness Scale [BIS]; Spinella,2007). Moreover, analyses linking these beha-vioural differences to brain activity showed thatindividual variation in the susceptibility to emo-tional distraction was linked to enhanced activityin brain regions associated with emotional pro-cessing (AMY) and to disrupted activity inregions associated with cognitive control (dlPFC)(Dolcos, 2009).
Relationships between brain activity and per-sonality-related differences were identified notonly for traits reflecting general aspects of cogni-tive processing, such as attentional impulsivitydiscussed earlier, but also for traits reflectingdifferences in processing and experiencing speci-fic emotions, such as anxiety. Investigations of therole of anxiety-related individual differencesshow that increased reactivity to potential threatconveyed by socially relevant stimuli (e.g., angryfaces) is associated with exacerbated activity inthe AMY in both clinical patients with socialanxiety disorder, linked to individual variations inthe severity of the symptoms (Evans et al., 2008;Goldin et al., 2008; Phan, Fitzgerald, Nathan, &Tancer, 2006), and in nonclinical individuals,linked to variations in the level of anxiety(Bishop, Duncan, & Lawrence, 2004; Ewbanket al., 2009; Stein, Simmons, Feinstein, & Paulus,
EMOTION, COGNITION, AND THE BRAIN 681
Dow
nloa
ded
by [
79.1
16.2
19.6
6] a
t 05:
41 0
5 M
ay 2
012
2007). These findings suggest that altered func-tioning of the AMY is not only disorder specific,but can also be observed in individuals prone todevelop anxiety disorders (Hariri et al., 2002;
Morey et al., 2011). In one of our recent studies(Denkova et al., 2010), we identified brain regionswhere activity in response to transient anxiety-inducing distraction covaried with individual
differences in trait anxiety and cognitive perfor-mance. Specifically, activity in the left visualcortex (including the left FG, BA 37) and vmPFCwas positively correlated, and activity in the right
dlPFC and dmPFC was negatively correlated withanxiety scores. These findings suggest that en-hanced trait anxiety is associated with increasedsensitivity to transient anxiety-inducing stimula-tion, which results in enhanced activity in brain
regions associated with the perception and ex-periencing of emotions (FG and vmPFC, respec-tively), and impaired activity in brain regionsassociated with the ability to maintain focus on
goal-relevant information (dlPFC).Another important source of interindividual
variation in emotion processing is individuals’tendency to spontaneously engage in emotionregulation, particularly in a context when thereis no explicit instruction or awareness of emotion
regulation. Spontaneous emotion regulation istypically assessed by evaluating the use of regula-tion strategies such as reappraisal and suppres-sion, as indexed by the Emotion RegulationQuestionnaire (ERQ; Gross & John, 2003).
Spontaneous reappraisal tendencies have beenassociated with lower levels of negative affect,greater interpersonal functioning, and greater
psychological and physical well-being (Gross &John, 2003). At the brain level, similar withinstructed reappraisal, individual differences inhabitual engagement of reappraisal are associatedwith decreased activation in ventral emotiongenerative regions, such as AMY, and increasedactivation in prefrontal control regions, such asdlPFC and dmPFC, in response to negativestimuli (Drabant, McRae, Manuck, Hariri, &Gross, 2009).
The role of sex-related differences
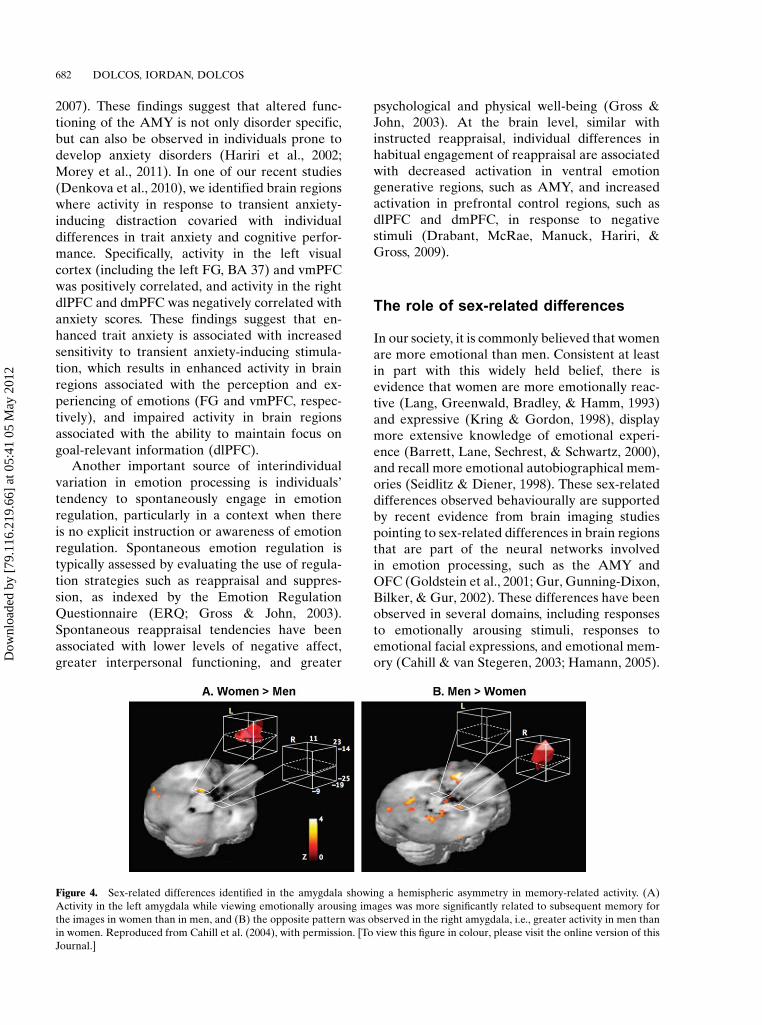
In our society, it is commonly believed that womenare more emotional than men. Consistent at leastin part with this widely held belief, there isevidence that women are more emotionally reac-tive (Lang, Greenwald, Bradley, & Hamm, 1993)and expressive (Kring & Gordon, 1998), displaymore extensive knowledge of emotional experi-ence (Barrett, Lane, Sechrest, & Schwartz, 2000),and recall more emotional autobiographical mem-ories (Seidlitz & Diener, 1998). These sex-relateddifferences observed behaviourally are supportedby recent evidence from brain imaging studiespointing to sex-related differences in brain regionsthat are part of the neural networks involvedin emotion processing, such as the AMY andOFC (Goldstein et al., 2001; Gur, Gunning-Dixon,Bilker, & Gur, 2002). These differences have beenobserved in several domains, including responsesto emotionally arousing stimuli, responses toemotional facial expressions, and emotional mem-ory (Cahill & van Stegeren, 2003; Hamann, 2005).
Figure 4. Sex-related differences identified in the amygdala showing a hemispheric asymmetry in memory-related activity. (A)
Activity in the left amygdala while viewing emotionally arousing images was more significantly related to subsequent memory for
the images in women than in men, and (B) the opposite pattern was observed in the right amygdala, i.e., greater activity in men than
in women. Reproduced from Cahill et al. (2004), with permission. [To view this figure in colour, please visit the online version of this
Journal.]
682 DOLCOS, IORDAN, DOLCOS
Dow
nloa
ded
by [
79.1
16.2
19.6
6] a
t 05:
41 0
5 M
ay 2
012

Particular attention have received sex-relateddifferences showing a hemispheric asymmetry inmemory-related activity in the AMY (Figure 4),i.e., left-lateralised activity in women and right-lateralised activity in men (Cahill, 2006; Cahillet al., 2004). Further evidence suggests that thislateralisation is confined to the BLA, and isobserved only when memory is tested after adelay (e.g., 2 weeks later), but not when testedimmediately (Mackiewicz, Sarinopoulos, Cleven,& Nitschke, 2006). Interestingly, these hemi-spheric differences were not observed for dorsalAMY (Mackiewicz et al., 2006) where activitywas associated with anticipatory processes andpredicted immediate, but not delayed recognitionmemory (see also Talmi, Anderson, Riggs,Caplan, & Moscovitch, 2008). These findingssuggest that the sex-related lateralisation is spe-cifically related to the BLA and to memoryconsolidation.
Given the function attributed to the left andright hemisphere in verbal/local versus nonverbal/global processing, it is possible that the patternrevealed by these studies could be due to the use ofverbal/local strategies during emotional encodingby women and the use of visual spatial/globalstrategies by men (Cahill, 2006). However, recentevidence suggests a social account for sex differ-ences in emotional memory. Consistent with thisinterpretation, recent evidence suggests that thesex-related lateralisation of AMYactivity dependson the sex of the faces, in that left AMY ispreferentially involved in successful encoding offemale faces in women, whereas right AMY islinked to successful encoding of male faces in men(Armony & Sergerie, 2007). This finding highlightsthe possible social/biological relevance of therelationship between the perceiver’s sex and thestimulus properties. Related to this, there isevidence that women seem, indeed, to have agreater preference for socially relevant stimuli(faces and persons vs. scenes; Proverbio, Zani, &Adorni, 2008), and that it is the feminine andmasculine roles as established by the society,rather than the biological sex per se, that seem toinfluence emotional memory (Cahill et al., 2004).
Although investigation of sex-related differ-ences in the cognitive control of emotion havereceived less attention, the available evidencepoints to possible differences in both emotionregulation strategies and the associated neuralcorrelates. Indeed, emotional responses and thesubsequent memory can be influenced not only bydifferences in emotional reactivity per se, but also
by differences in how emotions are regulated, orsome interaction between emotional reactivityand emotion regulation (McRae, Ochsner, Mauss,Gabrieli, & Gross, 2008). Consistent with the ideaof possible dissociable engagement of emotionregulation strategies in women and men, in astudy by Koch et al. (2007), during a taskinvolving emotion regulation females mainlyrecruited brain regions associated with emotionprocessing (i.e., AMY and the OFC), whereasmales mainly recruited brain regions typicallyinvolved in cognitive processing (i.e., prefrontaland superior parietal regions). Also, a studyinvestigating sex-related differences during cog-nitive reappraisal found that men showed greaterdecreases in AMY activity, along with lessercontrol-related prefrontal activity, whereas wo-men showed greater ventral striatal activity(McRae et al., 2008). These results may suggestthat men may be able to use cognitive regulationwith less effort than women and that women mayuse positive affect in the service of down-regulat-ing negative affect to a greater extent than men.
Sex-related differences were also identified inthe relationship between the habitual engagementof emotion regulation strategies and the retrievalof emotional autobiographical memories, withreappraisal being related to positive personalmemories in men and suppression with negativepersonal memories in women (Denkova et al.,2011). These findings extend the evidencethat men and women differ not only in termsof general emotional processing and memorybut also in the engagement of emotion regulationstrategies (Matud, 2004; Thayer, Rossy, Ruiz-Padial, & Johnsen, 2003).
In sum, the extant evidence suggests thatdifferential processing of and memory for specificemotional events, along with the associated neuralcorrelates, differ in men and women, and that thesedifferences may be related to variations in the useof encoding and emotion regulation strategies.
The role of age-related differences
Although older age is associated with increasingphysical ailments, psychological stress, sociallosses, and increased dependency, most of thepeople are in general satisfied in old age andexperience relatively high levels of emotionalwell-being. Supporting this idea, a large body ofresearch indicates that negative affect decreases(Gruhn, Jacqui, & Baltes, 2005) and positive
EMOTION, COGNITION, AND THE BRAIN 683
Dow
nloa
ded
by [
79.1
16.2
19.6
6] a
t 05:
41 0
5 M
ay 2
012

affect increases or remains stable throughout thelife span (Carstensen, Isaacowitz, & Charles,1999). This unexpected phenomenon, charac-terised by improved subjective emotional experi-ence in older age, is called the ‘‘paradox’’ ofageing (Carstensen et al., 1999; Mather et al.,2004) and understanding it in the context ofelucidating the role of individual differences inemotion�cognition interactions warrants furtherinvestigation (Allen et al., 2005).
Abundant evidence suggests an overall cogni-tive decline with ageing (Daselaar, Dennis, &Cabeza, 2007; Park et al., 2002; see also Timpe,Rowe, Matsui, Magnotta, & Denburg, this issue2011), but there is also evidence suggesting theexistence of compensatory mechanisms, the en-gagement of which allow healthy elderly to per-form in cognitive tasks as well as their youngercounterparts (Cabeza, Dolcos, Graham, & Ny-berg, 2002; Dolcos, Rice, & Cabeza, 2002). How-ever, evidence from the affective domain suggestsnot only overall preservation of ability to processemotional information with ageing (Keightley,Winocur, Burianova, Hongwanishkul, & Grady,2006; Mather & Knight, 2006), but also anenhanced ability to control emotion (Gross etal., 1997), which is reflected in a positive affectivebias, i.e., the tendency to attenuate negativeemotions and enhance positive emotions (Mather& Knight, 2006).
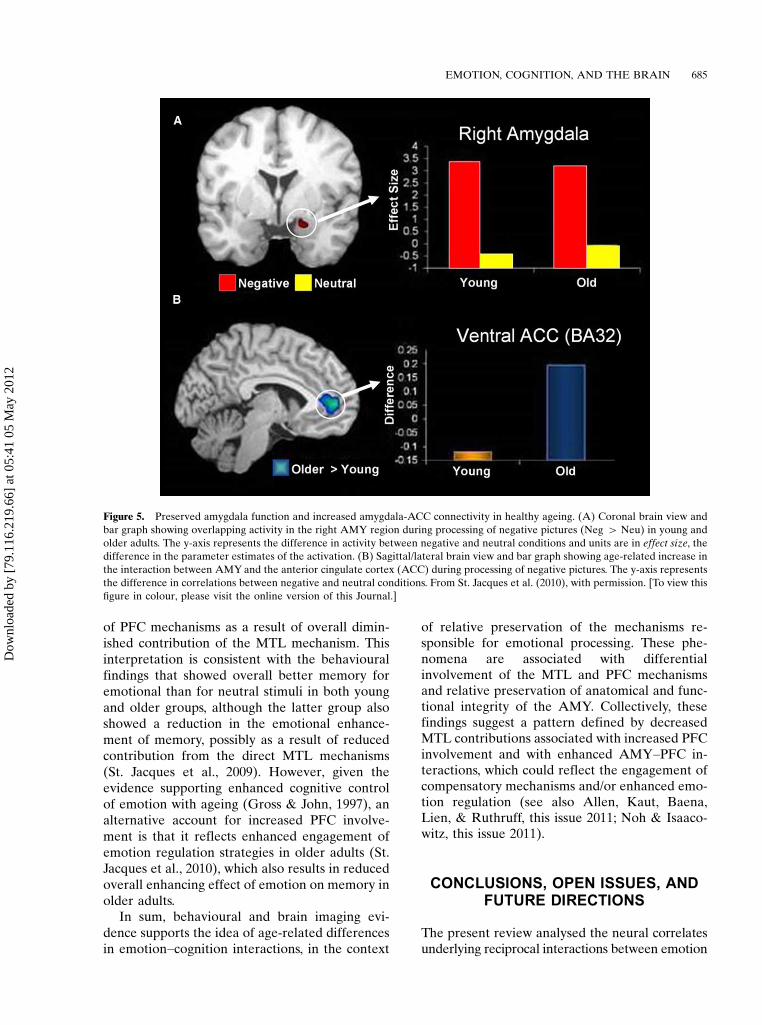
Age-related positivity effects have been docu-mented in how people perceive and appraiseemotional material, as well as how they remem-ber emotional memories (Mather et al., 2004).Functional neuroimaging evidence suggests thatage-related changes in general emotion proces-sing are linked to changes in AMY and frontalcortex activity, and in their interaction. Althoughsome reports suggest age-related decrease inAMY activation (Fischer et al., 2005; Tessitoreet al., 2005), others show that diminished AMYactivity is limited to negative stimuli, as opposedto positive stimuli (Mather et al., 2004). In arecent investigation of functional connectivitywithin the emotional network (St. Jacques et al.,2010), we identified evidence for age-relatedincrease in AMY�frontal interactions duringprocessing of negative stimuli, in the context ofpreserved AMY function (Figure 5A). Specifi-cally, older adults showed enhanced connectivitybetween activity in the AMY and the anteriorcingulate cortex (ACC) while rating the emo-tional content of negative pictures (Figure 5B).Interestingly, these age-related differences in
AMY�ACC interactions were associated withchanges in the perceived emotional content ofnegative pictures, as shown by more ‘‘neutral’’responses given by the older participants to thenegative pictures (St. Jacques et al., 2010). Otherstudies that explicitly manipulated emotion reg-ulation in older adults also found a couplingbetween the AMY and both the medial andlateral PFC (Urry et al., 2006; Winecoff, Labar,Madden, Cabeza, & Huettel, 2011)
Other reports focusing on age-related changesin emotional memory also provided evidence thatpreserved emotional enhancement of memory inageing is associated with relative preservation ofAMY structure and function, as well as withchanges in the neural circuitry underlying emo-tion processing in healthy older adults (Kensinger& Schacter, 2008; St. Jacques et al., 2009). In ourstudy investigating age-related differences inemotional memory, for instance (St. Jacques etal., 2009), we identified similar involvement ofAMY in emotional memory encoding in bothyounger and older participants, i.e., greater en-coding success activity for emotional than forneutral stimuli. However, the study also identifiedage-related differences in the formation of emo-tional memories in both MTL and PFC regions,i.e., a decrease in the MTL contribution (alongwith other posterior brain regions, includingvisual cortex) coupled with an increase in thedlPFC involvement. Moreover, these opposingeffects of ageing on MTL and dlPFC activity werealso observed in the functional connectivity ofthese brain regions with AMY: decreased for HC,and enhanced for dlPFC during encoding ofmemories for negative pictures.
These findings suggest an age-related reduc-tion in the contribution of the direct AMY�MTLmechanisms, which is compensated by the en-hanced contribution of AMY�PFC mechanismsto the formation of emotional memories. Theseage-related differences in the emotional networkare consistent with a more general pattern ofPosterior-Anterior Shift in Ageing (the PASAmodel), i.e., diminished engagement of the poster-ior brain regions compensated by enhanced con-tribution of the PFC regions (Davis, Dennis,Daselaar, Fleck, & Cabeza, 2008). Given thatthe PASA pattern observed in the cognitivedomain is interpreted as reflecting the engage-ment of compensatory mechanisms (Davis et al.,2008; Grady et al., 1994), the findings in theaffective domain mentioned above could also beinterpreted as reflecting compensatory engagement
684 DOLCOS, IORDAN, DOLCOS
Dow
nloa
ded
by [
79.1
16.2
19.6
6] a
t 05:
41 0
5 M
ay 2
012
of PFC mechanisms as a result of overall dimin-ished contribution of the MTL mechanism. Thisinterpretation is consistent with the behaviouralfindings that showed overall better memory foremotional than for neutral stimuli in both youngand older groups, although the latter group alsoshowed a reduction in the emotional enhance-ment of memory, possibly as a result of reducedcontribution from the direct MTL mechanisms(St. Jacques et al., 2009). However, given theevidence supporting enhanced cognitive controlof emotion with ageing (Gross & John, 1997), analternative account for increased PFC involve-ment is that it reflects enhanced engagement ofemotion regulation strategies in older adults (St.Jacques et al., 2010), which also results in reducedoverall enhancing effect of emotion on memory inolder adults.
In sum, behavioural and brain imaging evi-dence supports the idea of age-related differencesin emotion�cognition interactions, in the context
of relative preservation of the mechanisms re-sponsible for emotional processing. These phe-nomena are associated with differentialinvolvement of the MTL and PFC mechanismsand relative preservation of anatomical and func-tional integrity of the AMY. Collectively, thesefindings suggest a pattern defined by decreasedMTL contributions associated with increased PFCinvolvement and with enhanced AMY�PFC in-teractions, which could reflect the engagement ofcompensatory mechanisms and/or enhanced emo-tion regulation (see also Allen, Kaut, Baena,Lien, & Ruthruff, this issue 2011; Noh & Isaaco-witz, this issue 2011).
CONCLUSIONS, OPEN ISSUES, ANDFUTURE DIRECTIONS
The present review analysed the neural correlatesunderlying reciprocal interactions between emotion
Figure 5. Preserved amygdala function and increased amygdala-ACC connectivity in healthy ageing. (A) Coronal brain view and
bar graph showing overlapping activity in the right AMY region during processing of negative pictures (Neg � Neu) in young and
older adults. The y-axis represents the difference in activity between negative and neutral conditions and units are in effect size, the
difference in the parameter estimates of the activation. (B) Sagittal/lateral brain view and bar graph showing age-related increase in
the interaction between AMY and the anterior cingulate cortex (ACC) during processing of negative pictures. The y-axis represents
the difference in correlations between negative and neutral conditions. From St. Jacques et al. (2010), with permission. [To view this
figure in colour, please visit the online version of this Journal.]
EMOTION, COGNITION, AND THE BRAIN 685
Dow
nloa
ded
by [
79.1
16.2
19.6
6] a
t 05:
41 0
5 M
ay 2
012

and cognition, focusing on three main topics: (1)the impact of emotion on cognition, (2) theimpact of cognition on emotion, and (3) the roleof individual differences in emotion�cognitioninteractions. As discussed, emotion can produceboth enhancing and impairing effects on cogni-tion, can have both immediate and long-termeffects that may impact not only lower level (e.g.,perceptual) but also higher level (e.g., mnemonic)cognitive processes. An important aspect relatedto the impact of cognition on emotion processingis the capacity to deploy cognitive control in orderto resist momentary emotional distraction and, ona long run, the ability to cope with longer lastingemotions and feelings. The evidence concerningthe role of individual differences in these inter-actions points to factors that may influencesusceptibility to mood and anxiety disorders,which are characterised by unbalanced interac-tions between emotion and cognition that lead tothe devastating effects associated with thesepsychiatric conditions.
Overall, the extant evidence concerning theneural substrate of various aspects of emotion�cognition interactions points to dynamic inter-plays between separate but interconnected neuralsystems involved in emotional and cognitive/executive functions. This evidence also highlightsthe role of the AMY as central to the impact ofemotion on cognition, although its engagementimpacts differently activity in brain regions in-volved in the enhancing (increased) versus im-pairing (decreased) effects of emotion. Amygdalaactivity is also the target of top-down control ofemotion, exerted by lateral and medial PFCregions. Finally, activity in these neural systemsmay be linked to individual variation in aspectsthat potentially influence the sensitivity to emo-tional information, such as neuroticism, andfunctional alterations in the way they interfacewith each other may lead to emotional andcognitive disturbances, such as those observed inaffective disorders. Despite a rapidly growingbody of literature providing clarification of theneural correlates of emotion�cognition interac-tions, a number of issues are still unclear. We willnow briefly introduce them in relationship withaspects reviewed in this paper.
1. Regarding the impact of emotion on cogni-tion, although the findings reviewed hereprovide compelling evidence that emotioncan produce both enhancing and impairingeffects on cognition, the link between these
two effects is not clear. For instance, it isunclear to what extent certain brain regionscommonly identified in studies investigatingthe enhancing and impairing effects of emo-tion on cognitive processing (e.g., AMY)contribute to both effects, or may specificallycontribute to one or the other of the effects,depending on the context in which emotionalstimuli are presented (i.e., as task relevant ortask irrelevant). Simultaneous investigationof the neural correlates of the two opposingeffects of emotion on cognition (i.e., theenhancing vs. impairing) would allow identi-fication of the neural mechanisms underlyingprocessing that links the immediate disrupt-ing effects of emotional distraction on short-term/working memory to the enhancingeffect of emotion on long-term episodicmemory. It is plausible to expect that reallo-cation of processing resources in the presenceof task-irrelevant emotional distraction leadsto both impaired WM and enhanced episodicmemory for the distractors themselves, butprobably through modulation of brain activ-ity in different neural systems. Investigationof these effects within the same subjects iscritical, as they tend to cooccur in bothnormal and clinical conditions.
2. Another open question refers to the role ofemotional valence and arousal in both im-mediate and long-term effects of emotion oncognition. For instance, the majority ofstudies investigating the impact of task-irrelevant emotional distraction on perfor-mance in short-term/working memory tasks(Chuah et al., 2010; Denkova et al., 2010;Dolcos et al., 2008; Dolcos & McCarthy,2006) used only high-arousing unpleasantstimuli as emotional distractors, and henceit is not known whether the observed effectscould be generalised to all emotional stimuli,regardless of valence and level of arousal.Similarly, although most of the evidencefrom studies of episodic memory suggeststhat arousal is the main factor influencingthe memory-enhancing effect of emotion,the neural correlates underlying the contri-bution of emotional valence are less speci-fied (but see Mickley Steinmetz &Kensinger, 2009; Talmi, Schimmack, Pater-son, & Moscovitch, 2007). Given that posi-tive and negative emotions have evolved tosubserve different functions, it is reasonableto expect that their contribution to the
686 DOLCOS, IORDAN, DOLCOS
Dow
nloa
ded
by [
79.1
16.2
19.6
6] a
t 05:
41 0
5 M
ay 2
012

memory-enhancing effect of emotion may beassociated with different neural mechanisms,which may work in synchrony with themechanisms underlying the more generaleffect of emotional arousal. In this context,it would also be interesting to investigatewhether similar mechanisms contribute tothe memory-enhancing effect of emotion inthe case of specific emotions that may shareone or both of the basic emotion propertiesof arousal and valence (e.g., anger vs. sad-ness, anger vs. fear).
3. Regarding the neural correlates of emotioncontrol, chief among the open questions isunderstanding the role of different types ofemotional control and their associated neur-al correlates. Most of the previous emotionregulation research has focused on con-scious/deliberate forms of regulation, suchas conscious suppression of emotional re-sponses (Jackson et al., 2003), which mayrequire effort in controlling emotional re-sponses, but little is known about noncon-scious/automatic forms of emotionregulation and the associated neural corre-lates (see however Dolcos et al., 2009;Mauss, Evers, Wilhelm, & Gross, 2006).Manipulating the inducement of the goal tocontrol emotional response would be apractical way to directly compare the twoforms of emotion regulation. It is reasonableto expect that although automatic regulationmight be less effortful and less costly, itmight also be less efficient in the face ofhighly challenging transient emotional situa-tions. It is important to note, however, thattypically most of the emotions that we haveto deal with in daily life situations are rather‘‘mild’’, and thus it is very likely that auto-matic emotion regulation is more frequentlyengaged as a regulation strategy in normalcircumstances, as opposed to the engage-ment of deliberate strategies that may be-come critical in ‘‘crisis’’ circumstances.Therefore, it is important to investigatewhether nonconscious and deliberate emo-tion regulation strategies have similar im-mediate versus long-term effects onbehaviour, whether they share the sameneural correlates, and whether they aresimilarly affected in the case of chronicexposure to emotional challenge, leading toaffective dysregulation observed in affectivedisorders.
4. Concerning the role of individual differencesin understanding emotion�cognition interac-tions and the associated neural correlates,important open questions refer to their linkto individual variation in the susceptibility toaffective disorders. One way to address thisissue is by linking investigations of taskperformance in conjunction with personalitydata to investigations of brain activity. Also,in the context of a rapidly accumulatingcorpus of evidence pointing to sex-relateddifferences in emotion�cognition interac-tions, an important open question concernsthe link between emotional reactivity anddistractibility. Specifically, it is not knownwhether the enhanced emotional reactivityobserved in female participants also leads toenhanced emotional distractibility, andwhether individual variations in emotionalreactivity are linked to specific patterns ofactivity in the neural network involved in thecognitive control of emotion. Thus, it isimportant to investigate whether sex-relateddifferences in behaviour are reflected indifferent patterns of brain activity in brainregions associated with emotion processingand cognitive control.
5. Another important area of investigationconcerns identification of alterations in theneural correlates of cognition�emotion inter-action in clinical conditions, such as majordepression and posttraumatic stress disorder(Hayes et al., 2011; Ritchey, Dolcos, Edding-ton, Strauman, & Cabeza, 2011; Wang,LaBar, et al., 2008), and direct comparisonsof findings from populations showing con-trasting affective biases. An example of suchan ‘‘extreme’’ comparison in the domain ofemotion regulation is between healthy age-ing, associated with enhanced ability tocontrol emotion, and depression, associatedwith impaired emotion regulation, or emo-tion dysregulation. Existing evidence sup-ports the idea of age-related differences inemotion�cognition interactions, in the con-text of relative preservation of the mechan-isms responsible for emotion, and point toAMY�PFC interactions that may reflect theengagement of enhanced emotion regulation(St. Jacques et al., 2009, 2010). However, lessis known about the nature of specific auto-matic engagement of emotion regulationstrategies used by the elderly that generatespositivity biases in emotion processing and
EMOTION, COGNITION, AND THE BRAIN 687
Dow
nloa
ded
by [
79.1
16.2
19.6
6] a
t 05:
41 0
5 M
ay 2
012

how they may be affected in depression.Recent findings from our studies suggest thatsimilar neural circuitry, which appears to beengaged under conditions of enhanced cog-nitive control of emotion (in healthy ageing)(St. Jacques et al., 2010), malfunctions inconditions where regulation of emotion isdysfunctional (e.g., in depression) (Wang,Krishnan, et al., 2008; Wang, LaBar, et al.,2008), and that these effects are reversed bytreatment (Ritchey et al., 2011). Thus,further investigations and direct comparisonsof the neural mechanisms underlying theseopposing affective biases could prove fruitfulin determining whether the opposing affec-tive biases observed behaviourally are alsoreflected in the neural responses associatedwith differences in the ability to controlemotions observed in these two groups.These investigations have the potential toalso provide an explanation for the evidencesuggesting reduced incidence of depressionin the elderly (Christensen et al., 1999; Jorm,2000).
6. Finally, another open issue concerns theinvestigation of higher level affective phe-nomena, as well as emotion�cognitioninteractions in social contexts. Recent meth-odological refinements have encouraged theempirical approach of emotion�cognitioninteractions involving superior emotions,such as love (Zeki, 2007), and complexphenomena, such as social interaction(Adolphs, 2009). In both cases, the ability todecode emotional expressions and body lan-guage is crucial for the assessment of others’intentions. Studies investigating the neuralcorrelates of social cognition usually tendedto use facial stimuli (e.g., Todorov, 2008).However, there are clear advantages to usingalso whole-body stimuli, as they can berecognised as reliably as facial expressions,can direct attention to a person’s actions, andoverall are more ecologically valid (deGelder, 2009 Sung et al., 2011). Using emo-tion manipulations with dynamic whole-bodystimuli that mimic more closely real-lifesituations may contribute not only to betterunderstanding of the neural mechanisms ofsocial interactions in healthy behaviour, butalso to gaining insight into possible causes ofdeficits in social behaviour in such clinicalconditions as social anxiety and autism (Pel-phrey & Morris, 2006).
REFERENCES
Adolphs, R. (2009). The social brain: Neural basis ofsocial knowledge. Annual Review of Psychology, 60,693�716.
Allen, P. A., Kaut, K. P., Baena, E., Lien, M.-C., &Ruthruff, E. (2011). Individual differences in posi-tive affect moderate age-related declines in episodiclong-term memory. Journal of Cognitive Psychology,23(6), 768�779.
Allen, P. A., Kaut, K. P., Lord, R. G., Hall, R. J.,Grabbe, J. W., & Bowie, T. (2005). An emotionalmediation theory of differential age effects inepisodic and semantic memories. ExperimentalAging Research, 31(4), 355�391.
Amaral, D. G., Price, J. L., Pitkanen, A., & Carmichael,S. T. (1992). The amygdala: Neurobiological aspectsof emotion, memory, and mental dysfunction. NewYork, NY: Wiley-Liss.
Anderson, A. K., & Phelps, E. A. (2001). Lesions of thehuman amygdala impair enhanced perception ofemotionally salient events. Nature, 411, 305�309.
Anderson, A. K., Yamaguchi, Y., Grabski, W., & Lacka,D. (2006). Emotional memories are not all createdequal: Evidence for selective memory enhancement.Learning and Memory, 13(6), 711�718.
Armony, J. L., & Sergerie, K. (2007). Own-sex effects inemotional memory for faces. Neuroscience Letters,426(1), 1�5.
Arnsten, A. F. (1998). The biology of being frazzled.Science, 280(5370), 1711�1712.
Baddeley, A. (1996). The fractionation of working -memory. Proceedings of the National Academy ofSciences of the USA, 93(24), 13468�13472.
Balconi, M. (2011). Frontal brain oscillation modula-tion in facial emotion comprehension: The role ofreward and inhibitory systems in subliminal andsupraliminal processing. Journal of Cognitive Psy-chology, 23(6), 723�735.
Bargh, J. A., & Williams, L. E. (2007). On thenonconscious regulation of emotion. In J. Gross(Ed.), Handbook of emotion regulation (Vol. 1,pp. 429�445). New York, NY: Guilford Press.
Barrett, L. F., Lane, R., Sechrest, L., & Schwartz, G.(2000). Sex differences in emotional awareness.Personality and Social Psychology Bulletin, 26,1027�1035.
Bechara, A., Damasio, A. R., Damasio, H., & Ander-son, S. W. (1994). Insensitivity to future conse-quences following damage to human prefrontalcortex. Cognition, 50(1�3), 7�15.
Bechara, A., Damasio, H., & Damasio, A. R. (2003).Role of the amygdala in decision-making. Annals ofthe New York Academy of Sciences, 985, 356�369.
Bechara, A., Damasio, H., Tranel, D., & Damasio, A.R. (1997). Deciding advantageously before knowingthe advantageous strategy. Science, 275(5304),1293�1295.
Bechara, A., Tranel, D., Damasio, H., & Damasio, A.R. (1996). Failure to respond autonomically toanticipated future outcomes following dama-ge to prefrontal cortex. Cerebal Cortex, 6(2),215�225.
688 DOLCOS, IORDAN, DOLCOS
Dow
nloa
ded
by [
79.1
16.2
19.6
6] a
t 05:
41 0
5 M
ay 2
012

Bishop, S. J. (2009). Trait anxiety and impoverishedprefrontal control of attention. Nature Neuroscience,12(1), 92�98.
Bishop, S. J., Duncan, J., & Lawrence, A. D. (2004).State anxiety modulation of the amygdala responseto unattended threat-related stimuli. Journal ofNeuroscience, 24(46), 10364�10368.
Bradley, M. M., Greenwald, M. K., Petry, M. C., &Lang, P. J. (1992). Remembering pictures: Pleasureand arousal in memory. Journal of ExperimentalPsychology: Learning, Memory, and Cognition,18(2), 379�390.
Cabeza, R., Dolcos, F., Graham, R., & Nyberg, L.(2002). Similarities and differences in the neuralcorrelates of episodic memory retrieval and workingmemory. Neuroimage, 16(2), 317�330.
Cahill, L. (2006). Why sex matters for neuroscience.Nature Reviews Neuroscience, 7(6), 477�484.
Cahill, L., Babinsky, R., Markowitsch, H. J., &McGaugh, J. L. (1995). The amygdala and emotionalmemory. Nature, 377(6547), 295�296.
Cahill, L., Haier, R. J., Fallon, J., Alkire, M. T., Tang, C.,Keator, D., et al. (1996). Amygdala activity atencoding correlated with long-term, free recall ofemotional information. Proceedings of the NationalAcademy of Sciences of the USA, 93(15), 8016�8021.
Cahill, L., Uncapher, M., Kilpatrick, L., Alkire, M. T.,& Turner, J. (2004). Sex-related hemispheric later-alization of amygdala function in emotionally influ-enced memory: An FMRI investigation. Learningand Memory, 11(3), 261�266.
Cahill, L., & van Stegeren, A. (2003). Sex-relatedimpairment of memory for emotional events withbeta-adrenergic blockade. Neurobiology of Learn-ing and Memory, 79(1), 81�88.
Canli, T., & Amin, Z. (2002). Neuroimaging of emotionand personality: Scientific evidence and ethicalconsiderations. Brain and Cognition, 50(3), 414�431.
Canli, T., Desmond, J. E., Zhao, Z., & Gabrieli, J. D.(2002). Sex differences in the neural basis ofemotional memories. Proceedings of the NationalAcademy of Sciences of the USA, 99(16),10789�10794.
Canli, T., Sivers, H., Whitfield, S. L., Gotlib, I. H., &Gabrieli, J. D. (2002). Amygdala response to happyfaces as a function of extraversion. Science,296(5576), 2191.
Canli, T., Zhao, Z., Desmond, J. E., Kang, E., Gross, J.,& Gabrieli, J. D. (2001). An fMRI study ofpersonality influences on brain reactivity to emo-tional stimuli. Behavioral Neuroscience, 115(1),33�42.
Carstensen, L. L., Isaacowitz, D. M., & Charles, S. T.(1999). Taking time seriously: A theory of socio-emotional selectivity. The American Psychologist,54(3), 165�181.
Carstensen, L. L., Mikels, J. A., & Mather, M. (2006).Aging and the intersection of cognition, motivationand emotion. In J. E. Birren & K. W. Schaie (Eds.),Handbook of the psychology of aging (pp. 343�362).San Diego, CA: Academic Press.
Christensen, H., Jorm, A. F., Mackinnon, A. J., Korten,A. E., Jacomb, P. A., Henderson, A. S., et al. (1999).Age differences in depression and anxiety symp-
toms: A structural equation modelling analysis ofdata from a general population sample. Psychologi-cal Medicine, 29(2), 325�339.
Christianson, S. A. (1992). Emotional stress and eye-witness memory: A critical review. PsychologicalBulletin, 112(2), 284�309.
Chuah, L. Y. M., Dolcos, F., Chen, A. K., Zheng, H.,Parimal, S., & Chee, M. W. L. (2010). Sleepdeprivation and interference by emotional distrac-ters. SLEEP, 33(10), 1305�1313.
Damasio, A. R. (1994). Descartes error: Emotion,reason and the human brain. New York, NY: Avon.
Damasio, A. R. (1995). Toward a neurobiology ofemotion and feeling: Operational concepts andhypotheses. The Neuroscientist, 1, 19�25.
Damasio, A. R. (1996). The somatic marker hypothesisand the possible functions of the prefrontal cortex.Philosophical Transactions of the Royal Societyof London: Biological Sciences, 351B(1346),1413�1420.
Daselaar, S. M., Dennis, N. A., & Cabeza, R. (2007).Ageing: Age-related changes in episodic and work-ing memory. In S. A. R. B. Rombouts, F. Barkhof, &P. Scheltens (Eds.), Clinical applications of func-tional brain MRI (pp. 115�148). New York, NY:Oxford University Press.
Davidson, R. J., & Irwin, W. (1999). The functionalneuroanatomy of emotion and affective style.Trends in Cognitive Sciences, 3(1), 11�21.
Davis, M., & Whalen, P. J. (2001). The amygdala:Vigilance and emotion. Molecular Psychiatry, 6(1),13�34.
Davis, S. W., Dennis, N. A., Daselaar, S. M., Fleck, M.S., & Cabeza, R. (2008). Que PASA? The posterior-anterior shift in aging. Cerebral Cortex, 18(5),1201�1209.
De Gelder, B. (2009). Why bodies? Twelve reasons forincluding bodily expressions in affective neu-roscience. Philosophical Transactions of the RoyalSociety of London: Biological Sciences, 364B(1535),3475�3484.
Denkova, E., Chakrabarty, T., Dolcos, S., & Dolcos, F.(in press). Brain imaging investigation of the neuralcorrelates of emotional autobiographical recollec-tion. Journal of Visualized Experiments. doi: 10.3791/2396.
Denkova, E., Dolcos, S., & Dolcos, F. (2011). Relivingpersonal memories: Affective biases linked to per-sonality and sex-related differences. Manuscriptunder review.
Denkova, E., Wong, G., Dolcos, S., Sung, K., Wang, L.,Coupland, N., et al. (2010). The impact of anxiety-inducing distraction on cognitive performance: Acombined brain imaging and personality investiga-tion. PLoS ONE, 5(11), e14150.
Derryberry, D., & Reed, M. A. (1994). Temperamentand attention: Orienting toward and away frompositive and negative signals. Journal of Personalityand Social Psychology, 66(6), 1128�1139.
D’Esposito, M., Postle, B. R., & Rypma, B. (2000).Prefrontal cortical contributions to working mem-ory: Evidence from event-related fMRI studies.Experimental Brain Research, 133(1), 3�11.
EMOTION, COGNITION, AND THE BRAIN 689
Dow
nloa
ded
by [
79.1
16.2
19.6
6] a
t 05:
41 0
5 M
ay 2
012

Dolcos, F. (2009). Neural correlates of opposing mod-ulation of emotion on episodic and working memoryprocesses. Paper presented at the 2009 annualmeeting of the Memory Disorders Research Society.
Dolcos, F. (2010). The impact of emotion on memory:Evidence from brain imaging studies. Saarbrucken,Germany: VDM Verlag.
Dolcos, F., & Cabeza, R. (2002). Event-related poten-tials of emotional memory: Encoding pleasant,unpleasant, and neutral pictures. Cognitive, Affec-tive and Behavioral Neuroscience, 2(3), 252�263.
Dolcos, F., & Denkova, E. (2008). Neural correlates ofencoding emotional memories: A review of func-tional neuroimaging evidence. Cell Science Reviews,5(2), 78�122.
Dolcos, F., Diaz-Granados, P., Wang, L., & McCarthy,G. (2008). Opposing influences of emotional andnon-emotional distracters upon sustained prefrontalcortex activity during a delayed-response workingmemory task. Neuropsychologia, 46(1), 326�335.
Dolcos, F., Graham, R., LaBar, K., & Cabeza, R.(2003). Coactivation of the amygdala and hippo-campus predicts better recall for emotional than forneutral pictures. Brain and Cognition, 51, 221�223.
Dolcos, F., Kragel, P., Wang, L., & McCarthy, G. (2006).Role of the inferior frontal cortex in copingwith distracting emotions. Neuroreport, 17(15),1591�1594.
Dolcos, F., LaBar, K. S., & Cabeza, R. (2004a).Dissociable effects of arousal and valence on pre-frontal activity indexing emotional evaluation andsubsequent memory: An event-related fMRI study.Neuroimage, 23(1), 64�74.
Dolcos, F., LaBar, K. S., & Cabeza, R. (2004b).Interaction between the amygdala and the medialtemporal lobe memory system predicts better mem-ory for emotional events. Neuron, 42(5), 855�863.
Dolcos, F., LaBar, K. S., & Cabeza, R. (2005).Remembering one year later: Role of the amygdalaand the medial temporal lobe memory system inretrieving emotional memories. Proceedings of theNational Academy of Sciences of the USA, 102(7),2626�2631.
Dolcos, F., LaBar, K. S., & Cabeza, R. (2006). Thememory-enhancing effect of emotion: Functionalneuroimaging evidence. In B. Uttl, N. Ohta, & A. L.Siegenthaler (Eds.), Memory and emotion: Interdis-ciplinary perspectives (pp. 107�133). Malden, MA:Blackwell Publishing.
Dolcos, F., & McCarthy, G. (2006). Brain systemsmediating cognitive interference by emotional dis-traction. Journal of Neuroscience, 26(7), 2072�2079.
Dolcos, F., Miller, B., Jha, A., & McCarthy, G. (2007).Regional brain differences in the effect of distrac-tion during a delay interval of a working memorytask. Brain Research, 1152, 171�181.
Dolcos, F., Rice, H. J., & Cabeza, R. (2002). Hemi-spheric asymmetry and aging: Right hemispheredecline or asymmetry reduction. Cognitive andBiobehavioral Reviews, 26, 819�825.
Dolcos, S., Sung, K., Denkova, E., Dixon, R. A., &Dolcos, F. (in press). Brain imaging investigation ofthe neural correlates of emotion regulation. Journalof Visualized Experiments. doi: 10.3791/2430.
Drabant, E. M., McRae, K., Manuck, S. B., Hariri, A.R., & Gross, J. J. (2009). Individual differences intypical reappraisal use predict amygdala and pre-frontal responses. Biological Psychiatry, 65(5),367�373.
Drevets, W. C., & Raichle, M. E. (1998). Reciprocalsuppression of regional cerebral blood flow duringemotional versus higher cognitive processes: Impli-cations for interactions between emotion and cogni-tion. Cognition and Emotion, 12(3), 353�385.
Eugene, F., Levesque, J., Mensour, B., Leroux, J. M.,Beaudoin, G., Bourgouin, P., et al. (2003). Theimpact of individual differences on the neuralcircuitry underlying sadness. Neuroimage, 19(2. Pt.,1), 354�364.
Evans, K. C., Wright, C. I., Wedig, M. M., Gold, A. L.,Pollack, M. H., & Rauch, S. L. (2008). A functionalMRI study of amygdala responses to angry sche-matic faces in social anxiety disorder. Depressionand Anxiety, 25(6), 496�505.
Ewbank, M. P., Lawrence, A. D., Passamonti, L.,Keane, J., Peers, P. V., & Calder, A. J. (2009).Anxiety predicts a differential neural response toattended and unattended facial signals of anger andfear. Neuroimage, 44(3), 1144�1151.
Eysenck, M. W., Derakshan, N., Santos, R., & Calvo, M.G. (2007). Anxiety and cognitive performance:Attentional control theory. Emotion, 7(2), 336�353.
Fischer, H., Sandblom, J., Gavazzeni, J., Fransson, P.,Wright, C. I., & Backman, L. (2005). Age-differen-tial patterns of brain activation during perception ofangry faces. Neuroscience Letters, 386(2), 99�104.
Fuster, J. M. (1997). Network memory. Trends inNeurosciences, 20(10), 451�459.
Goldin, P. R., McRae, K., Ramel, W., & Gross, J. J.(2008). The neural bases of emotion regulation:Reappraisal and suppression of negative emotion.Biological Psychiatry, 63(6), 577�586.
Goldstein, J. M., Seidman, L. J., Horton, N. J., Makris,N., Kennedy, D. N., Caviness, V. S., Jr., et al. (2001).Normal sexual dimorphism of the adult human brainassessed by in vivo magnetic resonance imaging.Cerebral Cortex, 11(6), 490�497.
Grady, C. L., Maisog, J. M., Horwitz, B., Ungerleider, L.G., Mentis, M. J., Salerno, J. A., et al. (1994). Age-related changes in cortical blood flow activationduring visual processing of faces and location.Journal of Neuroscience, 14(3, Pt. 2), 1450�1462.
Gray, J. R., Braver, T. S., & Raichle, M. E. (2002).Integration of emotion and cognition in the lateralprefrontal cortex. Proceedings of the National Acad-emy of Sciences of the USA, 99(6), 4115�4120.
Gross, J. J. (2008). Emotion regulation. In M. Lewis, J.M. Haviland-Jones, & L. F. Barrett (Eds.), Hand-book of emotions (pp. 497�512). New York, NY:Guilford Press.
Gross, J. J., Carstensen, L. L., Pasupathi, M., Tsai, J.,Skorpen, C. G., & Hsu, A. Y. (1997). Emotion andaging: Experience, expression, and control. Psychol-ogy and Aging, 12(4), 590�599.
Gross, J. J., & John, O. P. (1997). Revealing feelings:Facets of emotional expressivity in self-reports, peerratings, and behavior. Journal of Personality andSocial Psychology, 72(2), 435�448.
690 DOLCOS, IORDAN, DOLCOS
Dow
nloa
ded
by [
79.1
16.2
19.6
6] a
t 05:
41 0
5 M
ay 2
012

Gross, J. J., & John, O. P. (2003). Individual differencesin two emotion regulation processes: Implicationsfor affect, relationships, and well-being. Journal ofPersonality and Social Psychology, 85(2), 348�362.
Gruhn, D., Jacqui, S., & Baltes, P. B. (2005). No agingbias favoring memory for positive material: Evi-dence from a heterogeneity�homogeneity list para-digm using emotionally toned words. Psychologyand Aging, 20(4), 579�588.
Gur, R. C., Gunning-Dixon, F., Bilker, W. B., & Gur, R.E. (2002). Sex differences in temporo-limbic andfrontal brain volumes of healthy adults. CerebralCortex, 12(9), 998�1003.
Haas, B. W., & Canli, T. (2008). Emotional memoryfunction, personality structure and psychopathology:A neural system approach to the identification ofvulnerability markers. Brain Research Reviews,58(1), 71�84.
Haas, B., Omura, K., Amin, Z., Constable, R. T., &Canli, T. (2006). Functional connectivity with theanterior cingulate is associated with extraversionduring the emotional Stroop task. Social Neu-roscience, 1(1), 16�24.
Hamann, S. (2005). Sex differences in the responses ofthe human amygdala. The Neuroscientist, 11(4),288�293.
Hamann, S., & Canli, T. (2004). Individual differencesin emotion processing. Current Opinion in Neuro-biology, 14(2), 233�238.
Hariri, A. R., Mattay, V. S., Tessitore, A., Kolachana,B., Fera, F., Goldman, D., et al. (2002). Serotonintransporter genetic variation and the response of thehuman amygdala. Science, 297(5580), 400�403.
Hayes, J. P., LaBar, K. S., McCarthy, G., Selgrade, E.,Nasser, J., Dolcos, F., et al. (2011). Reducedhippocampal and amygdala activity predictsmemory distortions for trauma reminders in com-bat-related PTSD. Journal of Psychiatric Research,45(5), 660�669.
Jackson, D. C., Mueller, C. J., Dolski, I., Dalton, K. M.,Nitschke, J. B., Urry, H. L., et al. (2003). Now youfeel it, now you don’t: Frontal brain electricalasymmetry and individual differences in emotionregulation. Psychological Science, 14(6), 612�617.
Johnson, M. K., Raye, C. L., Mitchell, K. J., Greene, E.J., Cunningham, W. A., & Sanislow, C. A. (2005).Using fMRI to investigate a component process ofreflection: Prefrontal correlates of refreshing a just-activated representation. Cognitive, Affective andBehavioral Neuroscience, 5(3), 339�361.
Jorm, A. F. (2000). Does old age reduce the risk ofanxiety and depression? A review of epidemiologi-cal studies across the adult life span. PsychologicalMedicine, 30(1), 11�22.
Keightley, M. L., Winocur, G., Burianova, H., Hon-gwanishkul, D., & Grady, C. L. (2006). Age effectson social cognition: Faces tell a different story.Psychology and Aging, 21(3), 558�572.
Kensinger, E. A., & Corkin, S. (2003). Effect ofnegative emotional content on working memoryand long-term memory. Emotion, 3(4), 378�393.
Kensinger, E. A., & Corkin, S. (2004). Two routes toemotional memory: Distinct neural processes for
valence and arousal. Proceedings of the NationalAcademy of Sciences of the USA, 101(9), 3310�3315.
Kensinger, E. A., & Schacter, D. L. (2008). Neuralprocesses supporting young and older adults’ emo-tional memories. Journal of Cognitive Neuroscience,20(7), 1161�1173.
Kilpatrick, L., & Cahill, L. (2003). Amygdala modula-tion of parahippocampal and frontal regions duringemotionally influenced memory storage. Neuro-image, 20(4), 2091�2099.
Kim, S. H., & Hamann, S. (2007). Neural correlates ofpositive and negative emotion regulation. Journal ofCognitive Neuroscience, 19(5), 776�798.
Koch, K., Pauly, K., Kellermann, T., Seiferth, N. Y.,Reske, M., Backes, V., et al. (2007). Gender differ-ences in the cognitive control of emotion: An fMRIstudy. Neuropsychologia, 45(12), 2744�2754.
Kring, A. M., & Gordon, A. H. (1998). Sex differencesin emotion: Expression, experience, and physiology.Journal of Personality and Social Psychology, 74(3),686�703.
LaBar, K. S., & Cabeza, R. (2006). Cognitive neu-roscience of emotional memory. Nature ReviewsNeuroscience, 7(1), 54�64.
LaBar, K. S., & Phelps, E. A. (1998). Arousal-mediatedmemory consolidation: Role of the medial temporallobe in humans. Psychological Science, 9(6),490�493.
Lang, P. J., Greenwald, M. K., Bradley, M. M., &Hamm, A. O. (1993). Looking at pictures: Affective,facial, visceral, and behavioral reactions. Psycho-physiology, 30(3), 261�273.
LeDoux, J. (1996). The emotional brain: The mysteriousunderpinnings of emotional life. New York, NY:Simon & Schuster.
LeDoux, J. (2000). Emotion circuits in the brain.Annual Review of Neuroscience, 23, 155�184.
LeDoux, J., & Phelps, E. A. (2000). Emotional net-works in the brain. New York, NY: Guilford Press.
Levesque, J., Eugene, F., Joanette, Y., Paquette, V.,Mensour, B., Beaudoin, G., et al. (2003). Neuralcircuitry underlying voluntary suppression of sad-ness. Biological Psychiatry, 53(6), 502�510.
Mackiewicz, K. L., Sarinopoulos, I., Cleven, K. L., &Nitschke, J. B. (2006). The effect of anticipation andthe specificity of sex differences for amygdala andhippocampus function in emotional memory. Pro-ceedings of the National Academy of Sciences of theUSA, 103(38), 14200�14205.
Markowitsch, H. J. (2008). Autobiographical memory:A biocultural relais between subject and environ-ment. European Archives of Psychiatry and ClinicalNeuroscience, 258(Suppl. 5), 98�103.
Mather, M., Canli, T., English, T., Whitfield, S., Wais, P.,Ochsner, K., et al. (2004). Amygdala responses toemotionally valenced stimuli in older and youngeradults. Psychological Science, 15(4), 259�263.
Mather, M., & Carstensen, L. L. (2005). Aging andmotivated cognition: The positivity effect in atten-tion and memory. Trends in Cognitive Sciences,9(10), 496�502.
Mather, M., & Knight, M. R. (2006). Angry faces getnoticed quickly: Threat detection is not impaired
EMOTION, COGNITION, AND THE BRAIN 691
Dow
nloa
ded
by [
79.1
16.2
19.6
6] a
t 05:
41 0
5 M
ay 2
012

among older adults. Journal of Gerontology: Psy-chological Sciences and Social Sciences, 61B(1),54�57.
Matud, M. P. (2004). Gender differences in stress andcoping styles. Personality and Individual Differences,37, 1401�1415.
Mauss, I. B., Cook, C. L., & Gross, J. J. (2007).Automatic emotion regulation during anger provo-cation. Journal of Experimental Social Psychology,43, 698�711.
Mauss, I. B., Evers, C., Wilhelm, F. H., & Gross, J. J.(2006). How to bite your tongue without blowingyour top: Implicit evaluation of emotion regulationpredicts affective responding to anger provocation.Personality and Social Psychology Bulletin, 32(5),589�602.
Mayberg, H. S. (1997). Limbic-cortical dysregulation: Aproposed model of depression. Journal of Neurop-sychiatry and Clinical Neurosciences, 9(3), 471�481.
McGaugh, J. L. (2000). Memory: A century of con-solidation. Science, 287(5451), 248�251.
McGaugh, J. L. (2002). Memory consolidation and theamygdala: A systems perspective. Trends in Neuro-sciences, 25(9), 456�461.
McGaugh, J. L. (2004). The amygdala modulates theconsolidation of memories of emotionally arousingexperiences. Annual Review of Neuroscience, 27,1�28.
McRae, K., Ochsner, K. N., Mauss, I. B., Gabrieli, J. J.D., & Gross, J. J. (2008). Gender differences inemotion regulation: An fMRI study of cognitivereappraisal. Group Processes and Intergroup Rela-tions, 11, 143�163.
Mickley Steinmetz, K. R., & Kensinger, E. A. (2009).The effects of valence and arousal on the neuralactivity leading to subsequent memory. Psychophy-siology, 46(6), 1190�1199.
Morey, R. A., Dolcos, F., Petty, C. M., Cooper, D. A.,Hayes, J. P., LaBar, K. S., et al. (2009). The role oftrauma-related distractors on neural systems forworking memory and emotion processing in post-traumatic stress disorder. Journal of PsychiatricResearch, 43(8), 809�817.
Morey, R. A., Hariri, A. R., Gold, A., Hauser, M. A.,Munger H. J., Dolcos, F., et al. (2011). Serotonintransporter gene polymorphisms and brain functionduring emotional distraction from cognitive proces-sing in posttraumatic stress disorder. BMC Psychia-try, 11(1), 76, http://www.biomedcentral.com/1471-244X/11/76, doi:10.1186/1471-244X-1-76.
Morris, J. S., Ohman, A., & Dolan, R. J. (1999). Asubcortical pathway to the right amygdala mediating‘‘unseen’’ fear. Proceedings of the National Acad-emy of Sciences of the USA, 96(4), 1680�1685.
Moscovitch, M. (1992). Memory and working-with-memory: A component process model based onmodules and central systems. Journal of CognitiveNeuroscience, 4(3), 257�267.
Moscovitch, M., Nadel, L., Winocur, G., Gilboa, A., &Rosenbaum, R. S. (2006). The cognitive neu-roscience of remote episodic, semantic and spatialmemory. Current Opinion in Neurobiology, 16(2),179�190.
Most, S. B., Chun, M. M., Widders, D. M., & Zald, D. H.(2005). Attentional rubbernecking: Cognitive con-trol and personality in emotion-induced blindness.Psychonomic Bulletin and Review, 12(4), 654�661.
Noh, S. R., & Isaacowitz, D. M. (2011). Age differencesin the emotional modulation of attention: Effects ofemotional cues on attentional engagement anddisengagement. Journal of Cognitive Psychology,23(6), 709�722.
Nguyen, C., Koeniqs, M., Yamada, T., Teo, S.-H.,Cavanaugh, J., Tranel, D., et al. (2011). Affectivefactors predict economic decision-making. Journalof Cognitive Psychology, 23(6), 748�759.
Ochsner, K. N. (2000). Are affective events richlyrecollected or simply familiar? The experience andprocess of recognizing feelings past. Journal ofExperimental Psychology: General, 129(2), 242�261.
Ochsner, K. N., Bunge, S. A., Gross, J. J., & Gabrieli, J.D. (2002). Rethinking feelings: An FMRI study ofthe cognitive regulation of emotion. Journal ofCognitive Neuroscience, 14(8), 1215�1229.
Ochsner, K. N., & Gross, J. J. (2005). The cognitivecontrol of emotion. Trends in Cognitive Sciences,9(5), 242�249.
Ochsner, K. N., Ray, R. D., Cooper, J. C., Robertson, E.R., Chopra, S., Gabrieli, J. D., et al. (2004). Forbetter or for worse: Neural systems supporting thecognitive down- and up-regulation of negative emo-tion. Neuroimage, 23(2), 483�499.
Ohman, A., Flykt, A., & Esteves, F. (2001). Emotiondrives attention: Detecting the snake in the grass.Journal of Experimental Psychology: General,130(3), 466�478.
Ohman, A., Flykt, A., & Ludqvist, D. (2000). Uncon-scious emotion: Evolutionary perspectives, psycho-physiological data and neuropsychologicalmechanisms. In R. D. Lane & L. Nadel (Eds.),Cognitive neuroscience of emotion (pp. 296�327).New York, NY: Oxford University Press.
Ohman, A., Lundqvist, D., & Esteves, F. (2001). Theface in the crowd revisited: A threat advantage withschematic stimuli. Journal of Personality and SocialPsychology, 80(3), 381�396.
Paller, K. A., & Wagner, A. D. (2002). Observing thetransformation of experience into memory. Trendsin Cognitive Sciences, 6(2), 93�102.
Palomba, D., Angrilli, A., & Mini, A. (1997). Visualevoked potentials, heart rate responses and memoryto emotional pictorial stimuli. International Journalof Psychophysiology, 27(1), 55�67.
Park, D. C., Lautenschlager, G., Trey, H., Davidson, N.S., Smith, A. D., & Smith, P. K. (2002). Models ofvisuospatial and verbal memory across the adult lifespan. Psychology and Aging, 17(2), 299�320.
Pelphrey, K. A., & Morris, J. P. (2006). Brain mechan-isms for interpreting the actions of others frombiological-motion cues. Current Directions in Psy-chological Science, 15(3), 136�140.
Pessoa, L. (2005). To what extent are emotional visualstimuli processed without attention and awareness?Current Opinion in Neurobiology, 15(2), 188�196.
Pessoa, L. (2008). On the relationship between emotionand cognition. Nature Reviews Neuroscience, 9(2),148�158.
692 DOLCOS, IORDAN, DOLCOS
Dow
nloa
ded
by [
79.1
16.2
19.6
6] a
t 05:
41 0
5 M
ay 2
012

Pessoa, L., McKenna, M., Gutierrez, E., & Ungerleider,L. G. (2002). Neural processing of emotional facesrequires attention. Proceedings of the NationalAcademy of Sciences of the USA, 99(17),11458�11463.
Phan, K. L., Fitzgerald, D. A., Nathan, P. J., & Tancer,M. E. (2006). Association between amygdala hyper-activity to harsh faces and severity of social anxietyin generalized social phobia. Biological Psychiatry,59(5), 424�429.
Phan, K. L., Wager, T., Taylor, S. F., & Liberzon, I.(2002). Functional neuroanatomy of emotion: Ameta-analysis of emotion activation studies in PETand fMRI. Neuroimage, 16(2), 331�348.
Phelps, E. A. (2004). Human emotion and memory:Interactions of the amygdala and hippocampalcomplex. Current Opinion in Neurobiology, 14(2),198�202.
Poldrack, R. A., Wagner, A. D., Prull, M. W., Desmond,J. E., Glover, G. H., & Gabrieli, J. D. (1999).Functional specialization for semantic and phonolo-gical processing in the left inferior prefrontal cortex.Neuroimage, 10(1), 15�35.
Proverbio, A. M., Zani, A., & Adorni, R. (2008).Neural markers of a greater female responsivenessto social stimuli. BMC Neuroscience, 9(1), 56.doi:10.1186/1471-2202-1189-1156
Pujol, J., Harrison, B. J., Ortiz, H., Deus, J., Soriano-Mas, C., Lopez-Sola, M., et al. (2009). Influence ofthe fusiform gyrus on amygdala response to emo-tional faces in the non-clinical range of socialanxiety. Psychological Medicine, 39(7), 1177�1187.
Richards, J. M. (2004). The cognitive consequences ofconcealing feelings. Current Directions in Psycholo-gical Science, 13(4), 131�134.
Richardson, M. P., Strange, B. A., & Dolan, R. J. (2004).Encoding of emotional memories depends on amyg-dala and hippocampus and their interactions. NatureNeuroscience, 7(3), 278�285.
Ritchey, M., Dolcos, F., & Cabeza, R. (2008). Role ofamygdala connectivity in the persistence of emo-tional memories over time: An event-related fMRIinvestigation. Cerebral Cortex, 18(11), 2494�2504.
Ritchey, M., Dolcos, F., Eddington, K. M., Strauman, T.J., & Cabeza, R. (in press). Neural correlates ofemotional processing in depression: Changes withcognitive behavioral therapy and predictors oftreatment response. Journal of Psychiatric Research,5(5), 577�587. doi:10.1016/j.jpsychires.2010.09.007
Rubin, D. C. (2005). A basic-systems approach toautobiographical memory. Current Directions inPsychological Science, 14, 79�83.
Rusting, C. L. (1999). Interactive effects of personalityand mood on emotion-congruent memory andjudgment. Journal of Personality and Social Psychol-ogy, 77(5), 1073�1086.
Sanfey, A. G., Rilling, J. K., Aronson, J. A., Nystrom, L.E., & Cohen, J. D. (2003). The neural basis ofeconomic decision-making in the Ultimatum Game.Science, 300(5626), 1755�1758.
Schneider, W., & Shiffrin, R. M. (1977). Controlled andautomatic human information processing: I. Detec-tion, search, and attention. Psychological Review,84(1), 1�66.
Seidlitz, L., & Diener, E. (1998). Sex differences in therecall of affective experiences. Journal of Personalityand Social Psychology, 74(1), 262�271.
Selye, H. (1985). The nature of stress. Basal Facts, 7(1),3�11.
Sergerie, K., Lepage, M., & Armony, J. L. (2005). Aface to remember: Emotional expression modulatesprefrontal activity during memory formation. Neu-roimage, 24(2), 580�585.
Sergerie, K., Lepage, M., & Armony, J. L. (2006). Aprocess-specific functional dissociation of the amyg-dala in emotional memory. Journal of CognitiveNeuroscience, 18(8), 1359�1367.
Shafer, A., & Dolcos, F. (2010, October). FunctionalMRI investigation of the role of processing load,emotional content, distraction time, and individualdifferences in the impact of distraction on a visualperceptual task. Paper presented at the fourthannual meeting of the Social and Affective Neu-roscience Society, Chicago, IL.
Shafer, A., Iordan, A., Cabeza, R., & Dolcos, F. (2011).Brain imaging investigation of the memory-enhan-cing effect of emotion. Journal of Visualized Experi-ments, 51, doi: 10.3791/2433
Sharot, T., & Yonelinas, A. P. (2008). Differential time-dependent effects of emotion on recollective experi-ence and memory for contextual information. Cog-nition, 106(1), 538�547.
Shaw, K., Lien, M.-C., Ruthruff, E., & Allen, P. A.(2011). Electrophysiological evidence of emotionperception without central attention. Journal ofCognitive Psychology, 23(6), 695�708.
Shaw, P., Brierley, B., & David, A. S. (2005). A criticalperiod for the impact of amygdala damage on theemotional enhancement of memory? Neurology,65(2), 326�328.
Simons, J. S., & Spiers, H. J. (2003). Prefrontal andmedial temporal lobe interactions in long-termmemory. Nature Reviews Neuroscience, 4(8),637�648.
Smith, E. E., & Jonides, J. (1999). Storage and execu-tive processes in the frontal lobes. Science,283(5408), 1657�1661.
Spinella, M. (2007). Normative data and a short form ofthe Barratt Impulsiveness Scale. International Jour-nal of Neuroscience, 117(3), 359�368.
St. Jacques, P. L., Dolcos, F., & Cabeza, R. (2009).Effects of aging on functional connectivity of theamygdala for subsequent memory of negative pic-tures: A network analysis of fMRI data. Psycholo-gical Science, 20(1), 74�84.
St. Jacques, P. L., Dolcos, F., & Cabeza, R. (2010).Effects of aging on functional connectivity of theamygdala during negative evaluation: A networkanalysis of fMRI data. Neurobiology of Aging, 31,315�327.
Stein, M. B., Simmons, A. N., Feinstein, J. S., & Paulus,M. P. (2007). Increased amygdala and insula activa-tion during emotion processing in anxiety-pronesubjects. American Journal of Psychiatry, 164(2),318�327.
Sung, K., Dolcos, S., Flor-Henry, S., Zhou, C., Gasior,C., Argo, J., et al. (2011). Brain imaging investiga-tion of the neural correlates of observing virtual
EMOTION, COGNITION, AND THE BRAIN 693
Dow
nloa
ded
by [
79.1
16.2
19.6
6] a
t 05:
41 0
5 M
ay 2
012

social interactions. Journal of Visualized Experi-ments, 53. doi: 10.3791/2379.
Sutton, T. M., & Altarriba, J. (2011). The automaticactivation and perception of emotion in wordprocessing: Evidence from a modified dot probeparadigm. Journal of Cognitive Psychology, 23(6),736�747.
Talmi, D., Anderson, A. K., Riggs, L., Caplan, J. B., &Moscovitch, M. (2008). Immediate memory conse-quences of the effect of emotion on attention topictures. Learning and Memory, 15(3), 172�182.
Talmi, D., Schimmack, U., Paterson, T., & Moscovitch,M. (2007). The role of attention and relatedness inemotionally enhanced memory. Emotion, 7(1),89�102.
Tessitore, A., Hariri, A. R., Fera, F., Smith, W. G., Das,S., Weinberger, D. R., et al. (2005). Functionalchanges in the activity of brain regions underlyingemotion processing in the elderly. Psychiatry Re-search, 139(1), 9�18.
Thayer, J. F., Rossy, L. A., Ruiz-Padial, E., & Johnsen,B. H. (2003). Gender differences in the relationshipbetween emotional regulation and depressive symp-toms. Cognitive Therapy and Research, 27, 349�364.
Timpe, J. C., Rowe, K. C., Matsui, J., Magnotta, V. A.,& Denburg, N. L. (2011). White matter intensity, asmeasured by diffusion tensor imaging, distinguishesbetween impaired and unimpaired older adultdecision makers. Journal of Cognitive Psychology,23(6), 760�767.
Todorov, A. (2008). Evaluating faces on trustworthi-ness: An extension of systems for recognition ofemotions signaling approach/avoidance behaviors.Annals of the New York Academy of Sciences, 1124,208�224.
Touryan, S. R., Johnson, M. K., Mitchell, K. J., Farb, N.,Cunningham, W. A., & Raye, C. L. (2007). Theinfluence of self-regulatory focus on encoding of,and memory for, emotional words. Social Neu-roscience, 2(1), 14�27.
Tulving, E. (1985). Memory and consciousness. Cana-dian Psychology, 26, 1�12.
Urry, H. L., van Reekum, C. M., Johnstone, T., Kalin,N. H., Thurow, M. E., Schaefer, H. S., et al. (2006).Amygdala and ventromedial prefrontal cortex areinversely coupled during regulation of negativeaffect and predict the diurnal pattern of cortisolsecretion among older adults. Journal of Neu-roscience, 26(16), 4415�4425.
Vuilleumier, P. (2005). How brains beware: Neuralmechanisms of emotional attention. Trends in Cog-nitive Sciences, 9(12), 585�594.
Vuilleumier, P., Armony, J. L., Driver, J., & Dolan, R. J.(2001). Effects of attention and emotion on faceprocessing in the human brain: An event-relatedfMRI study. Neuron, 30(3), 829�841.
Wang, L., Krishnan, K. R., Steffens, D. C., Potter, G. G.,Dolcos, F., & McCarthy, G. (2008). Depressive state-and disease-related alterations in neural responsesto affective and executive challenges in geriatricdepression. American Journal of Psychiatry, 165(7),863�871.
Wang, L., LaBar, K. S., Smoski, M., Rosenthal, M. Z.,Dolcos, F., Lynch, T. R., et al. (2008). Prefrontalmechanisms for executive control over emotionaldistraction are altered in major depression. Psychia-try Research, 163(2), 143�155.
Whalen, P. J., Rauch, S. L., Etcoff, N. L., McInerney, S.C., Lee, M. B., & Jenike, M. A. (1998). Maskedpresentations of emotional facial expressions mod-ulate amygdala activity without explicit knowledge.Journal of Neuroscience, 18(1), 411�418.
Winecoff, A., Labar, K. S., Madden, D. J., Cabeza, R.,& Huettel, S. A. (2011). Cognitive and neuralcontributors to emotion regulation in aging. SocialCognitive and Affective Neuroscience, 6(2), 165�176.
Wong, G., Dolcos, S., Denkova, E., Morey, R. A., Wang,L., McCarthy, G., Dolcos, F. (in press). Brainimaging investigation of the impairing effect ofemotion on cognition. Journal of Visualized Experi-ments. doi: 10.3791/2434
Zeki, S. (2007). The neurobiology of love. FEBS[Federation of European Biochemical Societies]Letters, 581(14), 2575�2579.
694 DOLCOS, IORDAN, DOLCOS
Dow
nloa
ded
by [
79.1
16.2
19.6
6] a
t 05:
41 0
5 M
ay 2
012





























