Embed Size (px)
Citation preview

DNA PALINDROME METABOLISM IN MAMMALIAN CELLS
By
Tara Anne Belsito
A thesis submitted in conformity with the requirements for the degree of Master of Science
Graduate Department of Molecular Genetics University of Toronto
©Copyright by Tara Anne Belsito, 2009

ii
DNA Palindrome Metabolism in Mammalian Cells Master of Science 2009 Tara Anne Belsito Department of Molecular Genetics University of Toronto
Abstract
A DNA palindrome is a sequence of DNA followed by an exact inverted copy of itself.
Palindromes are associated with gross chromosomal instability in mammalian cells. This may be
related to their ability to extrude a double-stranded cruciform structure. In mammalian cells,
palindromes have been shown to undergo centre-directed rearrangements resulting in a central
region of asymmetry. This process occurs via a mechanism termed „centre break palindrome
revision‟. In this thesis, I have investigated palindrome revision in mammalian cells using two
existing assays. In the first, performed by transfection of an extrachromosomal palindromic
dimer, I have shown that joining of palindrome-mediated double-strand breaks does not
depend solely on NHEJ and instead relies heavily on an alternate end-joining pathway. Using
the second assay, the Line78 mouse model which contains a 15.4kb transgenic palindrome, I
have shown that small modifications near the centre of the palindrome prevent these centre-
directed rearrangements possibly by inhibiting cruciform formation.

iii
Acknowledgements
I would like to thank my supervisor, Dr. Susanna Lewis, for her advice, support, and
encouragement. I would also like to thank my committee members, Dr. Bri Lavoie and Dr.
Steve Meyn for their comments and suggestions. Finally, I would like to thank the members of
the Lewis Lab, past and present, especially Atina Côté for her technical assistance and good
company.

iv
Table of Contents
DNA PALINDROME METABOLISM IN MAMMALIAN CELLS I
ABSTRACT II
ACKNOWLEDGEMENTS III
TABLE OF CONTENTS IV
LIST OF FIGURES VII
LIST OF TABLES VIII
LIST OF ABBREVIATIONS IX
CHAPTER 1: INTRODUCTION TO DNA PALINDROMES 1
Disease-Causing Palindromes in Humans 2 Translocations 2 Deletions 5 Amplifications 5
Evidence for Cruciform Formation 7
Palindrome Metabolism in E. coli 11 SbcCD-dependent breakage 12 Replication slipped mispairing 14
Palindrome Metabolism in Yeast 14
Palindrome Metabolism in Mammalian Cells: Insight from Model Systems 18 Revision of extrachromosomal palindromes 18 Revision of integrated chromosomal palindromes – the Line78 transgenic mouse line 19
Centre Break Palindrome Revision: A Model For Preventative Palindrome Modification in Mammals 21
NHEJ and its Potential Involvement in Centre Break Palindrome Revision 21
Thesis Rationale 25

v
CHAPTER 2: MATERIALS AND METHODS 26
Note to Reader 27
Fetal Liver Harvest 30
Cloning and Subcloning by Limiting Dilution 31
Genomic DNA Preparation 31
Line 78 PCR conditions 31
Southern Analysis 35
Rate Analysis 36 Maximum likelihood estimate sample calculations 37 Likelihood ratio test 38
Palindromic Dimer DNA preparation 40
Transfection Procedure 42
Transformation Procedure 42
Revision Product Analysis 42
CHAPTER 3: PALINDROME REVISION IS A FORM OF NON-CANONICAL END-JOINING 44
Note to Reader 45
Introduction 46 Experimental approach 46
Results 51 Preliminary gene screen 51 Joint analysis reveals common features of palindrome revision between NHEJ-proficient and NHEJ-deficient cell lines 51 There is no difference in the quantity of palindrome revision joints between NHEJ-proficient and NHEJ-deficient cells 56
Discussion 61 Extrachromosomal mammalian palindrome revision occurs in the absence of NHEJ 61 A subset of palindrome revision joints are formed via microhomology 62 Palindrome revision may be a function of non-canonical end-joining 64 Inserted DNA is a common feature in both extrachromosomal and intrachromosomal palindrome revision 66 Future Directions 67

vi
CHAPTER 4: THE STABILIZING EFFECTS OF ∆4BP ON THE LINE78 PALINDROME 72
Note to Reader 73
Introduction 74 Experimental approach 77
Results 81
Discussion 85 Line78 Rearrangements do not occur via replication slipped mispairing 87 Future Directions 88
REFERENCES 90

vii
List of Figures
Chapter 1: Figure 1.1: DNA palindromes and inverted repeats can form secondary DNA structures 3 Figure 1.2: Model of palindrome-mediated amplification 6 Figure 1.3: Mechanism of cruciform formation 9 Figure 1.4: Model of SbcCD mediated palindrome deletions in E. coli 13 Figure 1.5: Palindrome deletion by replication slipped mispairing 15 Figure 1.6: Model of cruciform resolution 17 Figure 1.7: Centre break model of palindrome revision 22 Figure 1.8: Basic mechanism of the NHEJ pathway 24 Chapter 2: Figure 2.1: Maximum likelihood estimate 39 Figure 2.2: Schematic of plasmids used in this study 41 Chapter 3: Figure 3.1a: Schematic of the qualitative extrachromosomal palindromic dimer assay 48 Figure 3.1b: Schematic of the quantitative extrachromosomal palindromic dimer assay 49 Figure 3.1c: Schematic of the extrachromosomal V(D)J assay 50 Figure 3.2: Nucleotides retained in axis-inclusive joints at the SalI and BamHI axes 57 Figure 3.3: The number of nucleotides used at junctions formed via microhomology 58 Figure 3.4: Palindrome revision efficiency in NHEJ-deficient and NHEJ-proficient cells 60 Chapter 4: Figure 4.1: Schematic of Line78 variants relevant to this study 76 Figure 4.2: Schematic of the Line78 rate of rearrangement assay 78 Figure 4.3: Schematic of the Line78 transgene and outlying areas 79 Figure 4.4: Southern blot analysis of Line78 4bp rearrangements in the germline 82
Figure 4.5: Southern blot analysis of Line78 4bp rearrangements in somatic cells 83

viii
List of Tables
Chapter 2: Table 2.1: Source of cell lines used in this study 29 Table 2.2: Results of subcloning of Line 78 4bp by limiting dilution 32
Table 2.3: Primers used in this study 33 Chapter 3: Table 3.1: Selected monomer junction sequences 52 Table 3.2: Summary of qualitative revision product analysis 53 Table 3.3: Quantitative analysis of palindrome revision in NHEJ-proficient and
NHEJ-deficient cell lines 59
Table 3.4: Sequence of palindrome revision junctions 68 Chapter 4:
Table 4.1: Rate analysis of the 4bp Line 78 palindrome 84

ix
List of Abbreviations
DSB - double-strand break
PATRR - palindromic AT-rich repeat
HPFH - hereditary persistence of fetal hemoglobin
NHEJ - non-homologous end-joining
MMEJ - microhomology mediated end-joining
DNA-PKcs - catalytic subunit of DNA-PK
ATM - ataxia telangiectasia mutated
MRN - Mre11/Rad50/Nbs1 (human homolog)
MRX - Mre11/Rad50/Xrs2 (yeast homolog)
DC-PCR - digestion circularization PCR
MLE - maximum likelihood estimate
Ab-MLV - Abelson murine leukemia virus
dsDNA - double-stranded DNA
ssDNA - single-stranded DNA

1
Chapter 1: Introduction to DNA Palindromes

2
The term “palindrome” is often used to describe a variety of sequences arrangements
which include perfect palindromes, near-palindromes, inverted repeats and spaced inverted
repeats (reviewed in (Lewis and Cote 2006)). In this study, the term “palindrome” is used to
describe a perfect DNA palindrome, which is a sequence of DNA followed immediately by an
exact inverted copy of itself. The term “near-palindrome” is used to describe sequences which
are similar to perfect palindromes but contain small central spacers and/or small sequence
inconsistencies between the two arms. Perfect palindromes and near-palindromes behave
similarly to each other, particularly in their secondary structure forming abilities. They both
have the ability to self-anneal while single-stranded to form a hairpin structure as well as while
double-stranded to form a cruciform structure (Fig. 1.1). The term “inverted repeat” is used to
describe inverted DNA sequences separated by central spacers longer than approximately 10
bp. Inverted repeats only have the ability to self-anneal while single-stranded to form hairpins;
they cannot extrude a cruciform structure.
Disease-Causing Palindromes in Humans
DNA palindromes are actually quite common in the human genome ((Lu, Jia et al.
2007) and S. Lewis, in preparation); there are at least 3,000 near-palindromes in the human
reference sequence. Palindromes are increasingly becoming better understood as their
involvement in human disease comes to light. This involvement is a result of a palindrome‟s
ability to cause various types of chromosomal instability.
Translocations
Emanuel syndrome is an inherited genetic disorder characterized by severe mental
retardation, preauricular tag or sinus, ear anomalies, cleft or high-arched palate, micrognathia,
microcephaly, kidney abnormalities, heart defects, and genital abnormalities in males (Zackai
and Emanuel 1980). It is a result of a supernumerary der(22) t(11;22 ) translocation. The

3
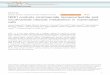
Figure 1.1: DNA palindromes and inverted repeats can form secondary DNA structures. A: A DNA Palindrome is a sequence of DNA followed immediately by an exact, inverted copy of
itself. B: DNA Palindromes have the ability to extrude a double-stranded cruciform structure. C: An inverted repeat is similar to a palindrome except that it contains a central region of
asymmetry. D: Inverted repeats are able to form single-stranded hairpin structures when single stranded,
such as on the lagging strand during replication. A palindrome is also able to form a single-stranded hairpin, however an inverted repeat cannot form a cruciform. Arrows indicate the direction of replication.
Okazaki Fragment
D
A C
B

4
breakpoints on chromosomes 11 and 22 map to the centres of large Palindromic AT-Rich
Repeats (PATRRs) located at 11q23 and 22q11 ((Kurahashi and Emanuel 2001) and see
(Kurahashi, Inagaki et al. 2006; Kurahashi, Inagaki et al. 2006) for reviews). A different PATRR
located at 17q11 is responsible for another recurrent constitutional translocation with 22q11;
again, the breakpoints map the centres of these near-palindromes (Kurahashi, Shaikh et al.
2003). Analysis of t(11;22) indicates that it occurs de novo at a high frequency during meiosis
in sperm samples, but not in lymphoblasts or fibroblasts (Kurahashi and Emanuel 2001).
Although the translocation occurs during meiosis, cytogenetics studies have determined that the
breakpoints on chromosomes 11 and 22 are not associated with MLH1 foci; MLH1 is involved
with reciprocal cross-overs, indicating that the breakpoints are not associated with
recombination hotspots (Ashley, Gaeth et al. 2006). Analysis of breakpoint sequences of
various translocations reveals that the breaks occur at the centres of the near-palindromes and
that joining is accompanied by small central deletions; thus joining is not dependent on
homology (Kurahashi, Inagaki et al. 2007).
It seems likely that the three PATRRs (22q11, 11q23, and 17q11) become involved in
translocations because of their ability to extrude cruciform structures. Using 2-D gel
electrophoresis and S1 nuclease digestion (techniques commonly used to detect cruciforms), it
was shown that plasmids containing the 22q11, 11q23, and 17q11 PATRRs can extrude
cruciforms in vitro (Kurahashi, Inagaki et al. 2004; Kogo, Inagaki et al. 2007). The 22q11
PATRR is the longest of the three studied at approximately 600 bp (the PATRR at 11q23 is
approximately 500bp and the 17q11 is the smallest at 200 bp) and is the PATRR which exhibits
the most homology between the two arms (Kurahashi, Inagaki et al. 2006). It is also the
PATRR most likely to undergo cruciform formation in vitro, followed by 11q23 (Kogo, Inagaki et
al. 2007). A small (9bp) spacer introduced into the centre of the 22q11 PATRR greatly reduces
its cruciform extrusion ability. The ability of the PATRRs to extrude in vitro compels one to

5
further examine the relationship between secondary structure formation and chromosomal
aberrations.
Deletions
A subset of thalassemia, hereditary persistence of fetal haemoglobin or HPFH, is caused
by deletions in the beta globin gene cluster; these deletions are all associated with a 160bp
palindrome. In a Belgian family, the 3‟ breakpoint is located at the centre of the palindrome
(Fodde, Losekoot et al. 1990). In an Indian family, the breakpoint is 4 bp downstream of the
centre (Henthorn, Mager et al. 1986) and in a Chilean family the breakpoint is 10 bp upstream
(Game, Bergounioux et al. 2003).
These palindrome-associated deletions are centre-directed (Henthorn, Mager et al.
1986; Fodde, Losekoot et al. 1990; Game, Bergounioux et al. 2003) as are the PATRR-mediated
translocations (Kurahashi, Inagaki et al. 2006; Kurahashi, Inagaki et al. 2006). It may be that
they are both a result of the same initiation event, presumably secondary structure formation.
Amplifications
Gene amplification is a common form of genome instability and is often associated with
tumour progression in mammals (see (Albertson 2006) for a review). In many characterized
cases in human cancers, the amplified regions contain large inverted duplications ((Tanaka,
Bergstrom et al. 2005; Myllykangas, Himberg et al. 2006; Tanaka, Bergstrom et al. 2006). One
proposed mechanism involves introduction of a double-strand break (DSB) adjacent to a short
inverted repeat. After formation of the DSB, 5‟-3‟ resection occurs followed by intrastrand
annealing at the short inverted repeat to create a hairpin structure. A large palindrome is
formed after one round of replication (Tanaka, Tapscott et al. 2002; Tanaka, Cao et al. 2007)
(Fig. 1.2). This can produce a dicentric chromosome which can then go on to cause further
rearrangements or amplifications through breakage-fusion-bridge cycles (McClintock 1941).

6
Figure 1.2: Model of palindrome-mediated amplification. Palindrome-mediated amplification occurs when a double-strand break occurs adjacent to a small inverted repeat (A). After the break occurs, 5‟-3‟ resection occurs, leaving a 3‟ overhang (B). The free end folds back on itself and intrastrand base-pairing occurs between the two arms of the inverted repeat (C). The gap is filled in, creating a hairpin end (D). After one round of replication, a large DNA palindrome is formed (E).
Inverted repeat
Double-strand break
5‟-3‟ resection
Foldback/ Intrastrand base-pairing
Gap fill-in
Replication
A
B
D B
C
E

7
In this case, the resultant palindromes serve as an intermediate for further large-scale
chromosomal amplification. There are also cases where only the palindromic sequence is
amplified. The nucleolus organizer regions (NORs) contain tandem arrays with hundreds of
copies of rRNA genes. Normally, these arrays contain a transcribed region followed by a non-
transcribed spacer. Molecular combing revealed that in normal individuals one third of the
repeats were arranged in an atypical fashion resulting in inverted repeats and/or near or perfect
palindromes. In Werner‟s syndrome patients approximately 50% of the repeats are inverted or
palindromic and they are found in clusters, implying that they are a result of repeat expansion
(Caburet, Conti et al. 2005) via a mechanism distinct from that causing gross chromosomal
amplification.
Based on the previous examples, it is fair to stay that palindromes are troublesome in
the human genome; often their effects are quite disastrous. Palindrome-induced chromosomal
rearrangements such as those discussed above also occur in other organisms. These organisms
also seem to have evolved mechanisms for preventing such damage.
Evidence for Cruciform Formation
Although palindromes and near-palindromes do have the ability to form a cruciform
structure, they do not do so readily. Approximately 17 kcal/mol of free energy are required to
drive cruciform extrusion in DNA (Courey and Wang 1983; Gellert, O'Dea et al. 1983; Lilley
1985; Benham, Savitt et al. 2002). This free energy is derived from torsional stress created by
negative supercoiling in the DNA (Lilley 1980; Panayotatos and Wells 1981; Mizuuchi, Mizuuchi
et al. 1982). Once the DNA has reached what‟s called its “superhelicity threshold” (Courey and
Wang 1988; Benham, Savitt et al. 2002), the excess energy created by the tightly under-wound
DNA results in melting of the interstrand hydrogen bonds at the centre of symmetry of the
palindrome (Murchie and Lilley 1987; Courey and Wang 1988; Zheng and Sinden 1988).
Approximately 10 base pairs are initially unwound, followed by intrastrand base-pairing and

8
branch migration, finally resulting in a fully extruded cruciform (Fig. 1.3) ((Murchie and Lilley
1987) and references therein). The formation of the cruciform results in the relaxation of
negative supercoils and a loss of the excess free energy that drove the initial change.
Therefore, when the DNA is past its superhelicity threshold, cruciforms are energetically
favoured and the process of cruciform formation is essentially irreversible.
In bacteria, the negative supercoiling required to drive cruciform extrusion may be
provided by DNA gyrase activity (Gellert, Mizuuchi et al. 1976) however recent work has shown
that supercoiling in the E. coli genome is tightly regulated by DNA gyrase, Topoisomerase I and
gene expression processes (i.e. transcription, see below) (Snoep, van der Weijden et al. 2002).
Given that negative supercoils in eukaryotic DNA are constrained by nucleosome formation, it
was originally thought that cruciform extrusion was not possible (Sinden, Carlson et al. 1980;
Mizuuchi, Mizuuchi et al. 1982). Subsequent studies have demonstrated that negative
supercoiling may occur during transcription (Liu and Wang 1987). Supercoils are induced in
plasmid DNA in prokaryotes in vivo (Wu, Shyy et al. 1988; Dayn, Malkhosyan et al. 1992) and in
vitro (Leng and McMacken 2002). This occurs as a result of the transcriptional machinery
progression; the DNA double-helix unwinds, the DNA ahead of the transcription machinery
becomes positively supercoiled and the DNA behind the transcription machinery becomes
negatively supercoiled. This is termed the “transcription-induced twin-supercoiled domain”
model (Liu and Wang 1987). Studies involving computer models indicate that the twin-domain
model generates sufficient stress to drive secondary structure transitions in DNA under
physiological conditions (Mielke, Fink et al. 2004).
Early evidence for cruciform formation in vitro came from electron micrographs of
plasmid DNA extruding cruciform structures in response to negative supercoiling (Mizuuchi,
Mizuuchi et al. 1982). The same study also demonstrated cruciform formation by showing the
loss of a restriction site located at the centre of the palindrome. Other in vitro studies make use
of the unique behaviour of the single-stranded nucleotides at the tips of the cruciform arms.

9
Figure 1.3: Mechanism of cruciform formation. In response to torsional strain, approximately 10 base pairs at the centre of the palindrome melt. Intrastrand base pairing occurs within the melted nucleotides. Branch migration occurs and the cruciform structure extrudes. Please see text for a more detailed description. Also see (Mizuuchi, Mizuuchi et al. 1982; Sullivan and Lilley 1986; Murchie and Lilley 1987) and references cited therein.
Lineform
Torsional strain and Local Melting
Intrastrand base-pairing and branch migration
Cruciform

10
Digestion of palindrome-containing DNA with S1 nuclease, which detects single-stranded
nucleotides at the cruciform tips, or Endonuclease VII, which cleaves across the base of the 4-
way junction, followed by electrophoresis and analysis of band size can reveal if the DNA
contains a nuclease-sensitive site (Panayotatos and Wells 1981). Single-strand bases at the tips
of cruciforms have been detected by chemical modification by chloroacetaldehyde (Dayn,
Malkhosyan et al. 1991), osmium tetroxide (McClellan, Boublikova et al. 1990) and
diethylpyrocarbonate (Scholten and Nordheim 1986)(reviewed in (Sinden 1994)). More
recently, cruciforms in DNA have been detected using atomic force microscopy (AFM)
(Chasovskikh, Dimtchev et al. 2005). Finally, trimethylpsoralen, which causes intrastrand
crosslinks to form, can be used to fix DNA in either the lineform or cruciform formation (Sinden,
Carlson et al. 1980). This assay has been used to demonstrate the ability of DNA to extrude a
cruciform structure in response to negative supercoiling (Sinden, Carlson et al. 1980) and the
rate of cruciform formation at different temperatures and at different ionic strengths (Sinden
and Pettijohn 1984).
Several of the above methods can be used to identify cruciform formation in vivo;
psoralen cross-linking (Zheng, Kochel et al. 1991), chemical modification of single stranded
bases (Dayn, Malkhosyan et al. 1992) and digestion by structure-specific nucleases in
protoplasts (del Olmo and Perez-Ortin 1993) have been used to demonstrate cruciform
formation in E. coli. Other in vivo studies that present more direct evidence for cruciform
formation involve use of an antibody that recognizes the cruciform structure (Frappier, Price et
al. 1987). These studies show that the cruciforms antibody binds DNA in living cells in G1/S
phase (Ward, Shihab-el-Deen et al. 1991). There is other indirect evidence for the existence of
cruciforms in vivo, mostly involving the location of site-specific DNA breaks; this will be
discussed later on.
The DNA sequence located at the centre of the palindrome has a drastic effect on its
ability to extrude a cruciform. Psoralen crosslinking assays have shown that palindromes with

11
AT stretches at their centres are more likely to form a cruciform in vitro (Zheng and Sinden
1988).
Studies involving plating efficiencies of phage containing palindromes show that those
palindromes with ATs at the centre have a decreased plating efficiency compared to those with
CGs at the centre (Davison and Leach 1994). Also, the plating efficiency increases if an
asymmetric spacer is inserted into the centre of the palindrome; the larger the spacer, the
greater the increase in plating efficiency (Chalker, Okely et al. 1993). Thus, one can say that
plating efficiency is correlated to cruciform formation; if the phage extrudes a cruciform, it is
not viable and the plating efficiency is decreased. Other studies examining imperfections in
palindrome-containing plasmids have shown that a 1-2 bp mutation in the centre 8-10 bp of
perfect palindromes can cause as much as a 2000-fold decrease in the kinetics of cruciform
formation (Murchie and Lilley 1987). Finally, studies on the energy needed to drive cruciform
formation show that for every unpaired base, the energy required increases substantially,
indicating that sequence imperfections in the near-palindrome have a major effect on cruciform
formation (Benham, Savitt et al. 2002). These effects decrease the further the imperfection is
from the centre of the near-palindrome.
Short DNA palindromes and inverted repeats are known to occur naturally, such as at
origins of DNA replication in plasmids and viruses in prokaryotes and eukaryotes (Jin and Novick
2001; Willwand, Moroianu et al. 2002). They have also been implicated in the regulation of
gene expression given the requirement for palindromes/inverted repeat symmetry for
transcription of some genes (Glucksmann, Markiewicz et al. 1992; Kim, Peng et al. 1998).
Palindrome Metabolism in E. coli
In E. coli, the first experiments regarding palindromic DNA involved palindrome-
containing plasmids in phage constructed in vitro and introduced in the cell (reviewed in
(Leach 1994)). Analysis of transformants revealed that palindromes any larger than 150-200 bp

12
were inviable (transformants did not contain any introduced DNA presumably due to a block in
replication) and palindromes as small as 20 bp were subject to partial or complete deletion,
referred to as instability. This instability increases with the length of the palindrome (up to 150-
200 bp). This instability or inviability in E. coli makes palindromic sequences very difficult to
analyze by standard molecular genetic techniques (incidentally, palindromes are also resistant
to amplification by standard PCR because of their ability to self-anneal). This „cloning barrier‟
explains why palindromes are under-represented and/or present in an incomplete form in the
human genome database (Lewis, Chen et al. 2005).
SbcCD-dependent breakage
There are two theories regarding palindrome metabolism in E. coli. The first theory is
that palindrome inviability in E. coli is dependent on the SbcCD complex, homologs of
eukaryotic Mre11/Rad50 (Naom, Morton et al. 1989; Gibson, Leach et al. 1992; Connelly and
Leach 1996). As SbcCD can cleave hairpin DNA molecules in vitro (Connelly, Kirkham et al.
1998), it likely exerts its effects by cleaving secondary structures formed in the cell (Leach,
Okely et al. 1997). A recent study (Eykelenboom, Blackwood et al. 2008) introduced a 246bp
closely spaced inverted repeat (24bp spacer) into the E. coli chromosome. Pulsed-field gel
electrophoresis revealed that DSBs are introduced in the vicinity of the inverted repeat centre.
The generation of these DSBs is both SbcCD- and replication-dependent. The physical and
genetic evidence presented by this study supports a model whereby a hairpin forms during
replication on the lagging strand after passage of the replication fork (Fig. 1.4). SbcCD cleaves
the hairpin and a DSB is produced. Further evidence indicates that the DSB is repaired via the
RecBCD recombination pathway.

13
Figure 1.4: Model of SbcCD mediated palindrome deletions in E. coli. During replication, palindromic DNA forms a hairpin structure on the lagging strand. Replication progresses past the hairpin, then the tips of the hairpin are cleaved by SbcCD. This results in one intact strand (formed from the leading strand) and a DSB in the other (formed from the lagging strand) (Eykelenboom, Blackwood et al. 2008).
SbcCD
Okazaki Fragment

14
Replication slipped mispairing
The second theory regarding palindrome metabolism in E. coli is also replication-
dependent and results in deletions (reviewed in (Leach 1994)). One group examined the
deleterious effects of a direct repeat flanked palindrome in the ampicillin resistance gene in
pBR322; deletion of the palindrome resulted in ampicillin resistant colonies. Sequence analysis
revealed that the deletion occurred via hybridization between the flanking repeats (DasGupta,
Weston-Hafer et al. 1987; Weston-Hafer and Berg 1989; Weston-Hafer and Berg 1991). They
propose a replication slipped mispairing model to explain their findings (Fig. 1.5). Hairpins are
formed in the lagging strand during DNA synthesis. When the replication machinery encounters
the hairpin, it stalls. The nascent strand dissociates and hybridizes with nearby complementary
sequence. This model is consistent with a similar, more recent study. A plasmid containing a
palindrome flanked by direct repeats was introduced into a sbcD mutant strain and sequence
analysis revealed deletion breakpoints at the direct repeats (Bzymek and Lovett 2001) implying
that these deletions are SbcCD-independent. Replication slipped mispairing can also occur
between a direct repeat located just inside the palindrome and a flanking direct repeat. This
implies that once the replication machinery encounters the hairpin, it progresses a short
distance into the hairpin before dissociating (Weston-Hafer and Berg 1989; Bzymek and Lovett
2001). A distinct feature of replication slipped mispairing is that the deletion always spans the
symmetry centre of the palindrome or inverted repeat.
Palindrome Metabolism in Yeast
Palindromes in yeast have a more varied behaviour than those in E. coli. There is
evidence that palindromes, inverted repeats and other hairpin-forming structures are deleted
during replication in a manner similar to replication slipped mispairing in E. coli (Gordenin,
Lobachev et al. 1993; Henderson and Petes 1993; Ruskin and Fink 1993; Tran, Degtyareva et
al. 1995; Lobachev, Shor et al. 1998). Short palindromes and inverted repeats can undergo

15
Figure 1.5: Palindrome deletion by replication slipped mispairing. A-B: A replication fork proceeds towards DNA containing a palindromic sequence. C: While single stranded, the palindromic DNA on the lagging strand forms a hairpin. The synthesizing lagging strand initially stalls at the hairpin, then dissociates from the template and re-anneals at a homologous sequence beyond the hairpin. D: Deletion of the palindrome can occur after a second round of replication. If only the central part of the palindrome takes place in hairpin formation, then the deletion will only include this central area. The deletion always includes the symmetry centre of the palindrome. Please see text for references.
microhomology
slippage
Okazaki
Fragment
A
B
C
D

16
amplification to create larger palindromes. For instance, budding yeast can maintain their
chromosome ends in the absence of telomerase or telomeric recombination by forming large
DNA palindromes; as such, this has been termed the PAL-mechanism of chromosome
maintenance ((Maringele and Lydall 2004) and reviewed in (Maringele and Lydall 2005)). The
PAL-mechanism occurs only in cells that have lost Exo1 activity, allowing the cells to overcome
checkpoint arrest and continue to divide. This type of palindrome amplification may be the
same as that which occurs in response to a DSB in yeast. The authors propose that the
inverted repeats can fold back on themselves to form a short hairpin and prime new DNA
synthesis. Break-induced replication of the hairpin-capped DNA creates a longer palindromic
sequence ((Butler, Yasuda et al. 1996)and reviewed in (Haber and Debatisse 2006)) (Fig. 1.2).
Amplification leading to giant palindromes occuring in response to chromosomal fragility
is observed at the location of an Alu inverted repeat in yeast chromosome V (320bp arms
separated by a 12bp spacer). The researchers observed that the inverted Alu repeats
stimulated a DSB at their centre and that the ends consisted of covalently closed hairpins
(Lobachev, Gordenin et al. 2002). The group proposes that the hairpin-capped DSB arises as a
result of cruciform extrusion in the Alu repeats followed by resolution of the four-way branch
structure. This occurs by diagonal cleavage across the base of the cruciform by a putative
nuclease in a manner analogous to Holliday junction resolution, hence a „cruciform resolution‟
model (Fig. 1.6). The MRX complex (Mre11-Rad50-Xrs2) and Sae2 are required to open the
hairpins and initiate single-strand resection of the DNA ends and that this can lead to the
formation of intrachromosomal amplicons (Narayanan, Mieczkowski et al. 2006; Narayanan and
Lobachev 2007). They propose that replication of the broken ends results in palindromic
dicentric chromosomes which break during anaphase. This stimulates break-induced
replication and recombination of the open-ended chromosome. Break-induced recombination

17
Figure 1.6: Model of cruciform resolution. When palindromic DNA extrudes a cruciform (A), a putative resolvase enzyme cleaves diagonally across the base of the cruciform (B). This leaves a hairpin-capped DSB (C) (Leach and Stahl 1983; Lobachev, Gordenin et al. 2002).
A
B
C

18
can occur intrachromosomally (Tran, Degtyareva et al. 1995; Narayanan, Mieczkowski et al.
2006) as well as interchromosomally, leading to translocations and gene conversion (Gordenin,
Lobachev et al. 1993; Tran, Degtyareva et al. 1995; Lobachev, Shor et al. 1998; Narayanan,
Mieczkowski et al. 2006).
Thus palindromes and inverted repeats in yeast can undergo simple deletions, such as
those seen in E. coli, or they can potentially lead to more extensive chromosomal aberrations
such as DNA amplification, although the specific mechanisms by which this amplification occurs
are speculative at this point.
Palindrome Metabolism in Mammalian Cells: Insight from Model Systems
Our lab has developed two systems for examining palindrome metabolism in mammalian
cells. The first is an extrachromosomal palindromic dimer assay. The second is an
intrachromosomal 15.4kb perfect palindromic transgene integrated into the mouse genome.
Examination of palindrome metabolism in these two systems has revealed that palindromes do
not always cause gross chromosomal instability leading to the idea that mammalian cells have
developed a system that modifies palindromes before they are able to cause damage.
Revision of extrachromosomal palindromes
The extrachromosomal palindromic dimer assay was developed in the course of
investigating hairpin DNA metabolism in eukaryotic cells (Lewis 1999). The specific details of
the assay and dimer construction will be discussed in Chapter 3. A palindromic dimer does not
transform E. coli, consistent with previous reports on large palindromes and their inviability
(reviewed in (Leach 1994)). When transfected into rodent cells, the dimer underwent
modifications to produce monomer sized DNA molecules capable of transforming E. coli. These
monomer products were DpnI resistant, indicating that they were unmethylated and thus had

19
been formed in the rodent cells. Sequence analysis revealed that breaks were induced quite
near the two symmetry axes of the dimer followed by rejoining; often the symmetry axes
remained intact. Monomer products also contained insertions which can be divided into two
categories: small insertions of fewer than 25bp of apparently random sequence, as well as
larger insertions, some of which contained vector-derived sequence. The joints also exhibited
variable sized deletions as well as joining apparently mediated by microhomology.
Revision of integrated chromosomal palindromes – the Line78 transgenic mouse line
The Line78 transgenic mouse line was originally created in a study that intended to
measure recombination in the male germline (Akgun, Zahn et al. 1997). The introduced
fragment contained two defective copies of the lacZ allele. Recombination between the two
alleles would result in a complete copy of the lacZ+ allele. One of the founder mice, mouse 78,
acquired two copies integrated in a tail-to-tail manner, thereby regenerating a PstI site at the
centre. The structure of the transgene was examined by Southern analysis (Akgun, Zahn et al.
1997) and DNA sequence analysis (Cunningham, Cote et al. 2003) and determined to be a
perfect palindrome.
A thorough analysis of the Line78 palindrome behaviour was conducted in the male
germline (Akgun, Zahn et al. 1997). Breeding analysis revealed that the Line78 transgene was
inherited in a Mendelian ratio. However, the transgene was not inherited stably. Novel
arrangements of the transgene were frequently detected in progeny of mice containing the
input palindrome – 15 to 56% of progeny were rearranged. Based on Southern analysis, it
appeared that the central region of the palindrome was particularly susceptible to
rearrangement; usually one, if not both, of the palindrome arms remained intact. The
variability of the bands seen by Southern analysis implied that this was due to some type of
nonhomolgous or illegitimate rearrangement. Rearrangement of the transgene was stabilizing,

20
so that the transgene was not observed to undergo any further rearrangements (Akgun, Zahn
et al. 1997).
Extensive Southern analysis of tail tip DNA from mice containing variants of the
transgene was performed; three classes of rearrangements were observed (Akgun, Zahn et al.
1997). The two relevant to this study were comprised of small (>200 bp) and large (1-3kb)
central deletions. These deletions often left the symmetry centre intact, indicating that they
occurred by a different mechanism than deletions in E.coli and yeast caused by replication
slipped mispairing. Both classes of deletions were stably inherited. The third class involved
homologous recombination between lacZ alleles and will not be discussed further.
In order to quantify the rate of rearrangement of the intact Line78 transgene in somatic
cells, permanent cell lines were established via Abelson Murine Leukemia Virus (Ab-MLV)
transformation of Line78 fetal liver cells (Cunningham, Cote et al. 2003). The specific details of
the rate assay will be discussed in Chapter 4. It was determined that rearrangement of the
intact Line78 transgene occurs at a rate of about 0.55% per population doubling. As in the
germline, the majority of the rearrangements involved non-homology based deletions extending
from the centre of the palindrome out to one or both arms.
The studies on the Line78 palindrome demonstrate that it can exist in the mouse
genome, although the intact palindrome undergoes frequent stabilizing rearrangements
affecting the symmetry centre. Although the intact Line78 palindrome is far from stable, it does
not appear to be associated with any gross chromosomal instability. For instance, there was no
unbridled amplification of the palindrome, only incremental expansion of the repeats (Zhou,
Akgun et al. 2001) and complete loss of the transgene was never seen (Akgun, Zahn et al.
1997; Cunningham, Cote et al. 2003).

21
Centre Break Palindrome Revision: A Model For Preventative Palindrome
Modification in Mammals
Our lab has proposed a model, termed Centre Break Palindrome Revision, to explain the
asymmetrical deletions and insertions observed in the Line78 palindrome as well as in the
extrachromosomal palindromic dimer assay (Fig. 1.7) (Lewis 1999; Cunningham, Cote et al.
2003). The model proposes that DNA containing a palindromic sequence extrudes a cruciform
structure. A nick is introduced at the tips of the hairpins. Branch migration and resorption of
the cruciform occur, resulting in a double strand break (DSB). The ends are processed by
nucleases and polymerases and eventually sealed. This results in an inverted repeat with a
central region of asymmetry, which is no longer able to extrude a cruciform structure. The
centre break palindrome revision model accounts for the structure of the joints seen in both the
intra- and extrachromosomal palindrome systems. The deletions encompassing only one arm of
the palindrome can be explained as a result of DSB formation at the symmetry centre of the
palindrome followed by processing. Similarly, the retention of the symmetry axes and
nucleotides past the symmetry centre provides evidence for centre-directed breaks, which can
arise from cruciform formation. The variable sized insertions observed can result from a free 3‟
end which can participate in strand invasion and extension. Finally, palindrome revision would
result in the palindrome containing sequence being modified, preventing further cruciform
extrusion and any possible destructive effects that may occur as a result.
NHEJ and its Potential Involvement in Centre Break Palindrome Revision
DSBs are extremely toxic to the cell. Some known causes are ionizing radiation, reactive
oxygen species, replication fork stalling, mechanical shearing at anaphase bridges, and during
V(D)J recombination and class-switch recombination (reviewed in (Sonoda, Hochegger et al.
2006; Soulas-Sprauel, Rivera-Munoz et al. 2007)). If not repaired, the broken chromosomes

22

23
can lead to deletions or translocations of chromosome fragments, potentially resulting in
inactivation of tumour suppressors and/or activation of oncogenes.
There are two well-characterized pathways that repair a DSB, homologous
recombination or NHEJ (reviewed in (Sonoda, Hochegger et al. 2006)). Homologous
recombination is a precise form of repair where a homologous sequence of DNA on a sister
chromatid serves as a template; for this reason homologous recombination occurs only in late S
and G2 phase. NHEJ on the other hand, is an imprecise mechanism active throughout the cell
cycle (reviewed in (Lieber 2008; Weterings and Chen 2008)).
The basic mechanism of NHEJ is outlined below (Fig. 1.8). The first step in NHEJ
involves recognition and binding of the broken DNA ends by Ku. Ku70 and Ku80 bind to either
hairpin-capped or open DNA ends with high affinity (Arosio, Cui et al. 2002) and form a
heterodimeric ring at the DNA ends (Walker, Corpina et al. 2001). Once Ku has bound to the
DNA, it attracts the catalytic subunit of DNA-PK (DNA-PKcs); the three proteins together from
the DNA-PK holoenzyme. DNA-PKcs, a kinase, may be involved in synapsis of the DNA ends
(DeFazio, Stansel et al. 2002). Next, incompatible ends are processed by nucleases and gap-
filling polymerases (TdT, polymerases and ) (reviewed in (Lieber, Lu et al. 2008). It is this
processing, resulting in the loss or gain of sequence, that makes NHEJ an imprecise form of
repair. Finally, DNA LigaseIV seals the break in conjunction with XRCC4 and XLF/Cernunnos
(Grawunder, Wilm et al. 1997; Grawunder, Zimmer et al. 1998; Ahnesorg, Smith et al. 2006).
NHEJ is especially important during development of the mammalian immune system; it
is responsible for joining the DSBs formed during V(D)J (reviewed in (Lieber, Ma et al. 2004)
(Soulas-Sprauel, Rivera-Munoz et al. 2007)). In V(D)J recombination during B-cell
development, DSBs are induced by the RAG proteins. At the coding ends, the RAG proteins
generate hairpin-capped ends that are opened by Artemis. Alone, Artemis acts as a 5‟-3‟
exonuclease, however upon phosphorylation by DNA-Pkcs, it acquires hairpin endonuclease

24
Figure 1.8: Basic Mechanism of the NHEJ pathway. When a DSB is formed, the DNA ends are recognized and bound by the Ku70/Ku80 heterodimer, which in turn attracts the catalytic subunit of DNA-PK (DNA-PKcs). The DNA-PK holoenzyme tethers the broken DNA ends together where they are processed by nucleases and polymerases. Once the ends are compatible they are sealed by LigIV in conjunction with XRCC4 and XLF. Redrawn from (Lees-Miller and Meek 2003).
DSB break
KU70/80
DNA-PKcs
LigIV/XRCC4/XLF
Restored DNA
DNA-Pk holoenzyme
Synapsis and processing

25
activity (Ma, Pannicke et al. 2002). The variability created by NHEJ during V(D)J recombination
leads to a greater diversity of antigen receptors.
On the face of it, NHEJ provides a mechanism for achieving centre break palindrome
revision for many reasons. NHEJ is highly variable because of the nuclease and polymerase
activity; there are a vast number of ways that NHEJ can repair the same DSB. This is
consistent with the variability observed in Palindrome Revision joints. Moreover, NHEJ accounts
for the opening of the hairpins (Artemis) and the joining of the processed ends (DNA LigaseIV
complex).
Thesis Rationale
Based on their involvement in translocations, deletions and amplifications, we know that
DNA palindromes and their propensity to form secondary structures have the ability to wreak
havoc in our tightly controlled genomes. A more comprehensive understanding of how cells
prevent palindrome-induced damage will help us understand how and why these palindromes
are so harmful when not properly maintained. Ultimately, we expect this knowledge will help
illuminate some of the mechanisms behind oncogenesis and other genetic diseases.
The focus of this thesis will be on elucidating the mechanism of palindrome revision in
mammalian cells. In the first half I have addressed the question „what DNA DSB repair factors
are implicated in centre break palindrome revision?‟. Using the extrachromosomal palindromic
dimer assay, I will present evidence that, contrary to junction appearance, palindrome revision
is not solely dependent on NHEJ and that other alternative end-joining factors are responsible
for a significant portion of the end-joining observed. In the second half, I will demonstrate how
small modifications near the symmetry centre of the Line78 palindrome stabilize it from further
rearrangements. The effect of these small modifications on Palindrome Revision provides
continuing evidence in support of the centre break palindrome revision model.

26
Chapter 2: Materials and Methods

27
Note to Reader
Lesley Cunningham performed the fetal liver harvests, Ab-MLV infection and cloning of the ATM
+/- and ATM -/- cell lines. Julia Appleby performed the fetal liver harvests and Ab-MLV
infection of the Ku70 +/- and Ku70 -/- cell lines. Ming Li performed fetal liver harvests and Ab-
MLV infection of the Line78 ∆4bp cell lines.

28
Mice and Cell Lines
The Line 78 mouse strain originated in Dr. Maria Jasin‟s lab (Sloane Kettering, NYC). It
contains a 15.4kb perfect DNA palindrome, as described in (Akgun, Zahn et al. 1997). We
obtained the line as a heterozygote for Line 78 in a (C57BL/6 X CBA/CA)F1 back-cross
background. Our Line 78 colony was maintained by crossing heterozygous males to Balb/C
(purchased from Charles River Laboratories) mice (Appleby 2005).
The Line 78 4bp originated in a female mouse heterozygous for Line78 (mouse
#B5383). #B5383 was then crossed with a male Balb/C; 3 of the female offspring (mouse #
78D1306, #78D1307 and #78D1310) were back-crossed to a male Balb/C (Table 2.1). Fetal
livers from these crosses were harvested and cell lines were generated by Ab-MLV infection, as
described below (Table 3.2). The 4bp genotype was confirmed by southern blotting, DC-PCR
and sequencing, as described below.
Atm +/- mice were purchased from Jackson Laboratories (129/SvEv-Atmtm1Awb) and bred
to C57B6/CBA, Balb/C and C.B.-17/lcrCR/-scidBR mice (Table 2.1). Ku70 null mice were
obtained from Dr. Yansong Gu (University of Washington, Seattle) (Gu, Jin et al. 1997).
The human pre-B cell line Nalm-6 (wild type) and NII4P2 (DNA LigIV null) cell lines were
obtained from Dr. Michael Lieber (University of Southern California, Los Angeles) (Table 2.1).
Briefly, to create the NII4P2 cell line, the open reading frame of the DNA Ligase IV gene in
Nalm-6 cells was disrupted by insertion of a neomycin or puromycin resistance gene, eliminating
DNA LigIV catalytic activity as well as suppressing binding to XRCC4 (Grawunder, Zimmer et al.
1998).
All cell lines used in this study were maintained in 5% CO2 at 37 in RPMI (Wisent or
HyClone) containing 10% heat-inactivated fetal bovine serum (Wisent or HyClone), and 50M
-mercaptoethanol (Sigma). Ab-MLV transformed cells were split 1:50 every 3 days, Nalm-6
and NII4P2 cells were split 1:20 every 3 days.

29
Table 2.1: Source of cell lines used in this study.
Genotype Cell Line Source (reference)
Line 78 4bp 78D1307FL-6-1 78D1307FL-1-2 78D1310FL-1-2
Generated by Ab-MLV infection of fetal
liver of Line 78 4bp mice generated in our lab.
ATM +/- M243FL-3(2)-1 Generated by Ab-MLV infection of mice fetal livers (Cunningham, 2002). ATM -/- M243FL-4-1
Ku70 +/- K2016FL-2 Generated by Ab-MLV infection of mice fetal livers (Gao et al., 1998). Ku70 -/- K2016FL-3
DNA LigIV +/+ Nalm-6
Grawunder et al., 1998. DNA LigIV -/- NII4P2

30
Fetal Liver Harvest
Breeding pairs were placed together on day 0 and separated by day 3. Pregnant females
were sacrificed on day 18, ensuring that the fetuses were between 16 and 18 days of gestation,
as this is when cells are sensitive to Ab-MLV infection and transformation (Rosenberg and
Baltimore 1976).
Under sterile conditions, uterine horns from pregnant females were removed. Each
fetus was then removed from the amniotic sac and placed in a separate 10cm dish containing
3ml of media (RPMI containing 20% heat-inactivated FBS, 50 uM B-mercaptoethanol and
100U/ml penicillin and 100 mg/ml streptomycin (Gibco)). Livers were dissected from the
fetuses and placed in 3cm dish containing 1ml of media.
In order to prepare a cell suspension, the livers and media were passed through a nylon
mesh (Cell MicroSieve, 70 micron pore size, Biodesign, or Nitex Nylon, 100 micron pore size,
Genesee Scientific). Cell suspensions were diluted to 2x106 cells/ml, and 1 ml was plated in a
6-well dish. Each well was then seeded with 1ml of Abelson murine leukemia virus (stocks were
prepared from a p-160 Ab-MLV producer cell line as previously described (Cho, Webber et al.
1999) and 4ug/ml polybrene (Sigma). Following infection, cells were rocked every 30 minutes
for 2 hours and incubated at 37C in 5% CO2. Five days post-infection, cells were fed with 2ml
of fresh media. Uninfected fetal liver cells were collected for DNA extraction and genotyping
(see below).
For the following 45-60 days, the cells undergo "crisis"; split ratios were initially only 1:2
every 3 days but gradually increased as the cultures because more confluent. Crisis is
considered complete when the cultures can be split 1:50 every three days and no apoptotic
cells were evident under the light microscope. At this time, cultures were switched to media
containing RPMI, 10% heat-inactivated FBS and 50uM B-mercaptoethanol. Cloned cell lines
were isolated by limiting dilution (see below).

31
Cloning and Subcloning by Limiting Dilution
Cells were plated on 96-well dishes at concentrations of 0.06, 0.19 or 0.31 cells per well
for ATM +/- and ATM -/- and for Ku70+/- and Ku70 -/- cell lines. Line 78 4bp cell lines were
plated at concentrations of 0.06 and 0.63 cells per well. After 12 days, wells with growth were
scored and select wells were expanded to cultures split 1:50 every 3 days (Table 2.2).
Genomic DNA Preparation
Approximately 2x107 cells from cell lines or liver tissue were collected at 100xg. Cells
were washed with RPMI and resuspended in 425 l of digestion buffer (100mM EDTA, 50mM
Tris pH 8.0, 1mg/ml Proteinase K (Sigma) and 0.5% SDS). When isolating DNA from mice tails,
approximately 0.5cm of tail was used.
Samples were rotated at 55C overnight, followed by one extraction with an equal
volume of phenol/chloroform/isoamyl alcohol (25:24:1) (500l) and one extraction with
chloroform in Phase Lock Gel 2.0ml heavy tubes (Eppendorf). The DNA was then precipitated
with an equal volume of isopropanol then pelleted. Precipitated DNA was resuspended in 200
to 500l TE and incubated overnight at 37C to reconsitute.
DNA concentration was determined using a Nanodrop (ND-1000) spectrophotometer
(Thermo Fisher Scientific).
Line 78 PCR conditions
PCR reactions were carried out using a PTC-200 thermal cycler (MJ research). Standard
reactions were comprised of 16mM (NH4)2SO4, 67mM Tris-HCl pH 8.8, 0.01% Tween-20, 25mM
MgCl2, 0.2mM of each primer, 0.2mM dNTPs and 0.5U of Taq (Bioron) in a total volume of 50l.
Primer sequences are listed in Table 2.3.

32
Table 2.2: Results of subcloning of Line 78 4bp by limiting dilution.
Cell Line Generation # of clones analyzed
(# of wells picked)
# of wells with growth
# of wells seeded
78D1307FL-1-2
66.7 7 7 96
96 4 15 96
148 24 24 384
78D1307FL-6-1 136 11 11 384
78D1310FL-1-2
52 9 23 96
81.3 3 5 96
133.3 29 29 384
Cultures were seeded at concentrations of 0.06 cells/ml and 0.63 cells/ml in 96-well plates.

33
Table 2.3: Primers used in this study.
Target Primer Name Sequence 5’-3’
Line 78 Left-flank (left-flank PCR)
AC174 CCG TGG CAG TAA CCA TTA AGA CC
DC5 GTC CAA CAA TCC AGC TTC AGG
Line 78 (DC-PCR)
Les1 CCT GTG TAG GTT CCA AAA TAT C
Erc1 AGC ATT ATC CTT ATC CAA AAC AGC
ACLF1 CTT TCC AGA TAC GAC GCA GG
ACR1 GGT GAC ATC ACA GTC TAA GGA GG
pGG52 monomer (sequencing)
TB-Fwd GAA AAG TGC CAC CTG ACG
TB-Rev GAG CAA CTG ACT GAA ATG CC
TB-004 AGA TCC AGT TCG ATG TAA CC
TB-005 ATG GCG GGA TCA ATT CGA GC
pJH298 monomer (sequencing)
lacprime CTC ATT AGG CAC CCC AGG CT
TB-001 CTA CAC AAC GAA GAG GTC C
TB-002 GAA CAC TAT CCC ATA TCA CC
TB-003 CGA TTC ATT AAT GCA GCT GG

34
The PCR conditions for Line 78 analysis are as follows: 94C for 3:00, 25 cycles at 94C
for 30sec, 58C for 45sec and 72C for 60sec, followed by a 10:00 extension at 72C. PCR
products were gel electrophoresed in a 2% agarose gel.
Mice were screened for the presence of Line 78 by using primers that spanned the 'left'
integration site (Left flank PCR), AC174 and DC5; a 200 bp band was indicative of transgene
inheritance.
DC-PCR
DC-PCR was used in order to determine the structure of the transgene in the area
adjacent to the centre of symmetry (Cunningham, Cote et al. 2003). 3g of DNA was digested
with 1U of Pst1 (Fermentas) overnight at 37C. The digestion reaction was then heat-
inactivated at 80C for 20min, then ligated with 80U T4 DNA ligase (NEB) in an 800 l reaction
for 2 hours at room temperature. The ligation reaction was then ethanol precipitated and
resuspended in 100l TE.
The left and right side of the Line 78 palindromic transgene were amplified in separate
nested reactions. The left side was amplified using the primers Les1/ACLF1 followed by
Erc1/ACLF1; the right side by using Les1/ACRF1 followed by Erc1/ACRF1. An intact transgene
resulted in a PCR product of approximately 200 bp from each side.
DC-PCR products were cloned into the pCR2.1-TOPO vector using the TOPO TA Cloning
kit (Invitrogen) using 1/3 of the manufacturers' recommended reaction volumes. The cloning
reaction was transformed into OneShot Top10 competent E. coli (Invitrogen) and plated onto
agar plates containing 100 g/ml of ampicillin. Plasmid DNA was prepared using the GeneJET
plasmid mini-prep kit (Fermentas) and sequenced by Macrogen Inc. (Seoul, Korea). Sequences
were aligned and analysed using BLAST 2 sequences (bl2seq program,
www.ncbi.nlm.nih.gov/blast/bl2seq/wblast2.cgi, (Tatusova and Madden 1999).

35
Southern Analysis
5g of genomic DNA was digested with 10U of the desired enzyme with the
recommended buffer in a total reaction volume of 30l, overnight at 37C. DNA samples were
electrophoresed in a 0.8% ultra-pure agarose gel (Invitrogen) with ethidium bromide in 1x TAE
at 57V for 4 hours.
After electrophoresis, the gel was rocked for 8min in 0.25M HCl to depurinate the DNA,
followed by 2 brief rinsings in H2O to remove residual HCl. The gel was then rocked in
denaturation solution 1 (0.5M NaOH, 1.5M NaCl) for 20 minutes, followed by 20 minutes in
denaturation solution 2 (0.25M NaOH, 1.5M NaCl).
Transfer was performed in denaturation solution 2 onto Genescreen transfer membrane
using upwards capillary transfer. After transfer, the membrane was rinsed with 2xSSC + 0.2M
Tris-HCL pH 8.0 and dried between 2 pieces of Whatmann paper.
Blots were probed with a labelled 3.5kb BamH1 fragment prepared from the Ercan 1.8
plasmid (Akgun, Zahn et al. 1997). Probe DNA was labelled using random primed synthesis;
50ng of probe DNA and 1 g of hexamers were boiled for 10min in 14l total volume. The
reaction was cooled on ice for 2min, then 5U of Klenow (NEB) with supplied buffer, 2.5mM
dNTP-dCTs and 50Ci -32P-dCTP were added to make a total volume of 25l. The reaction
was incubated at room temperature for 4 hours. The probe was purified using the ProbeQuant
G-50 Micro Columns (Amersham) according to the manufacturers' instructions.
Pre-hybridization was carried out for 4 hours at 42C in 25mL of Stark's buffer (5x SSC,
25mM NaPO4 pH 6.5, 5x Denhardt's, 50% formamide) supplemented with 6.25mg of boiled
torula RNA, 0.01% SDS and 125mg milk powder. The solution was first heated to 65C.
Hybridization was carried out in the same solution as above but with the addition of 1g
Dextran-Sufate Sodium Salt (Fisher Scientific). The probe was boiled for 10min and cooled on
ice for 2 min before addition to the hybridization solution. Hybridization was allowed to occur

36
overnight at 42C.
The hybridization solution was discarded and the membrane was rinsed briefly in 25mL
of Blot Wash Solution 1 (2x SSC, 0.1% SDS), then washed in same for 15min at 42C. Blot
Wash Solution 1 was discarded and the membrane was washed with Blot Wash Solution 2 (0.2x
SSC, 0.1% SDS) at 65C for 20 minutes, twice. Southern blots were then exposed on a
Phosphor screen (Molecular Dynamics) for 5-7 days and visualized using a PhosphorImager
Storm 860 (Molecular Dynamics) and ImageQuant computer program.
Rate Analysis
The rate of rearrangement of the Line 78 palindrome was calculated as in Cunningham
et al. (2003). Rates were based on the following equation:
x/y=(1-p)n
The fraction „x/y‟ is equal to the fraction of subclones with an input Line 78 transgene out of all
the subclones analyzed, and is determined by sampling subcultures via limiting dilution
subcloning. „n‟ is the number of population doublings since the subcloned line was seeded as a
single cell, and is calculated by the number of days that the cell line (since single cell) has been
in culture, given a population doubling time of 18 hours. Inputting these values into the above
equation allows us to solve for „p‟, which is the rate of rearrangement per population doubling.
Rate determinations from a culture at different generations, and/or from different
cultures of the same genotype (at different generations) can be pooled using the Maximum
Likelihood Estimate (MLE). MLE calculates the Likelihood (L) of a series of rates and determines
which rate best fits with the observed data. It also takes into consideration the limiting dilution
results (number of wells of growth/ total number of wells seeded, Table 4.3), as well as allows
calculation of 95% confidence intervals, even when the rate of rearrangement is 0 (Edwards
1972; Rosche and Foster 2000).

37
Maximum likelihood estimate sample calculations
Sample calculations to determine the Log Likelihood (LL) will be carried out using the
values for cell line 1307-1-2-1 at population doubling (n) 96, as in Table 4.4. The following
values were used: the estimate rate (R*) is equal to 0.003, x equals 3, y equals 4, the number
of wells seeded equals 96, and the number of wells with growth equals 15.
1. The fraction of intact Line78 transgenes (Fraction Intact, or FI) expected at a particular
doubling time (n) is calculated for an estimated rate (R*) according to the following
formula: FI = (1-R*)n, so FI=(1-0.003)96, and therefore FI=0.7494.
2. Next calculate the Revised Fraction (RF), which takes into account that a given well with
growth has a certain probability of being seeded by one cell, two cells, three cells etc.
The probability of each well being seeded by x number of cells follows a Poisson
distribution of rare events. It is calculated as follows: RF= (e-) [(e()(FI)-1)/( 1- e-
)], so RF=(0.844)[(e(0.1698)(0.7494)-1)/(1-0.844)], and therefore RF=0.7332.
The following was used to calculate in our sample: e-=1-(total # of wells with
growth/total # of wells seeded), so e-=1-(15/96)=0.844. Since =-ln(e-), then
=0.1698.
3. Log likelihood is then calculated using RF, x and y using the following equation:
LL=x(ln(RF/(1-RF)))+y(ln(1-RF)), so LL=3{ln[0.7332/(1-7332)]}+4[ln(1-0.7332)], and
therefore LL=(-2.25).
Steps 1 through 3 are then repeated for a series of estimated rates. The LLs are then
plotted against the estimated rates; the peak of the curve represents the rate most likely to fit
the data. The 95% confidence interval is bounded by the rates corresponding to the peak LL
+2 (LL=-4.25) (Rosche and Foster 2000).
Data from different experiments can be pooled by adding the LLs at each rate together
and then plotting. A sample curve is represented in Figure 2.1.

38
Likelihood ratio test
The likelihood ratio test can determine whether the data sets from different samplings
differ significantly from one another. The alternate hypothesis, that all experiments are
significantly different, is determined by summing the LL of the peak rate for each experiment
(LL1 + LL2 + LL3 +….). The null hypothesis, that the experiments are not significantly
different, is calculated as though all data were from one experiment (LL (1 + 2 + 3+…)) and
then determining the peak LL of this summation. The critical value is then determined as the
difference between the LL of the alternate and null hypothesis multiplied by two (2*([LL1 + LL2
+ LL3+…] – [LL(1 + 2 + 3 +…)])). Using the calculated critical value and setting the degrees
of freedom to the number of experiments being compared minus one, the p-value can be
determined by the 2 test. A p-value less than 0.05 allows for rejection of the null hypothesis.

39
Figure 2.1: Maximum likelihood estimate. The Log Likelihood (y) is calculated for a series of estimated rates (x). The peak rate represents the rate that best fits with the observed data. The 95% confidence interval is bounded at the rates corresponding to the peak LL+2 (see text).

40
Palindromic Dimer DNA preparation
The source plasmid for the palindromic dimer used to transfect human cell lines (Nalm-
6, NII4P2) was pGG52 (Gauss and Lieber 1996) (Fig. 2.2A). This plasmid was also used for
V(D)J assays in these cell lines. The source plasmid for the mouse (Ku70+/-, Ku70-/-)
palindromic dimer was pJH298 (Lieber, Hesse et al. 1988) (Fig. 2.2B). This plasmid does not
contain a functional RSS signal sequences, so p12x23 was used for V(D)J assays in these cell
lines. Plasmid DNA was purified from DH10B competent E. coli using a CsCl extraction.
The palindromic dimer was constructed in vitro in the following manner: 150g of DNA
was digested with 50U of BamH1 (Fermentas) overnight in 200l. The digestion was
precipitated and resuspended, then digested overnight with 50U of Sal1 (Fermentas), again, in
200l. A linear fragment was then purified from a 0.7% low-melting point agarose gel by
phenol/choloroform extraction. The recovered linearized DNA was ligated at 100g/ml with
approximately 20U of T4 DNA Ligase (NEB) per g of DNA in the suppliers‟s recommended
buffer at room temperature overnight. The ligation reaction was then electrophoresed in a
0.5% low-melting point agarose gel at 40V for approximately 24 hours at 4C and purified via
phenol/choloroform extraction. The purified dimer was transformed into E. coli to determine
the level of non-palindromic DNA (background levels). Only preps with less than 2000 ampicillin
resistant colonies per g of dimer prep were used.

41
Figure 2.2: Schematic of plasmids used in this study. Orange boxes represent the chloramphenicol resistance gene. Purple boxes represent the ampicillin resistance gene. Arrowheads indicate orientation. Black bars represents the bacterial origin of replication. Red bars represent the mammalian origin of replication. Elements are not to scale. Sizes of the linear fragments recovered and BamHI and SalI co-digestion are given. A: pGG52 is the source plasmid used for construction of the human palindromic dimer. It is derived from pBR328. It contains an SV40 ori for mammalian replication (Gauss and Lieber 1996). B: pJH298 is the source plasmid used for construction of the mouse palindromic dimer. It is derived from pUC13. It contains a polyoma ori for mammalian replication (Lieber, Hesse et al. 1988).
pJH298 8012kb
7722kb linear
SalI BamHI
pGG52
6277kb
5948kb linear
BamHI SalI
A
B

42
Transfection Procedure Transfection of pre-B cells was done by electroporation in the presence of DEAE-
Dextran. Transfection procedure was modified from the protocol described in Gauss and Lieber,
1992. Electroporations were performed using a BioRad Gene Pulser with 0.4cm electrode gap
cuvettes (BioRad or Cell Projects). 3.0x106 cells were pelleted, washed in RPMI then
resuspended in 800l RPMI, 10g/ml DEAE-Dextran (Pharmacia) and 100ng-1g DNA. The
suspension was then pulsed at 960FD and 0.3kV. Electroporated cells were then immediately
diluted with 4.2 ml of complete media (see above). Cells were incubated for 48 hours at 37C
in 5% CO2.
Transfected DNA was then harvested by a rapid alkaline lysis protocol as previously
described (Lewis 1994). In the case of control transfections (with source plasmids) or for V(D)J
assays, 3/5 of the harvested DNA was digested with 60U of Dpn1 (NEB) at 37C for 3 hours.
The digestion reactions were then precipitated and resuspended in TE.
Transformation Procedure
Transformation of recovered and/or Dpn1 digested DNA was performed by
electroporation of DH10B E. coli competent cells in a BioRad Gene Pulser at 200 , 25FD in
0.2cm electrode gap BioRad Gene Pulser cuvettes. Electroporated DH10B cells were incubated
with 500l of SOC in a 37C shaker for 1 hour at 220rpm then plated.
For V(D)J assays, cells were plated on LB-agar plates containing 100g/ml ampicillin, as
well as plates containing 100g/ml ampicillin and 10g/ml chloramphenicol. Chloramphenicol
resistant colonies were then streaked out on LB-agar plates containing 100g/ml ampicillin and
25g/ml chloramphenicol. For palindromic dimer assays, cells were plated on LB-agar plates
containing 100g/ml ampicillin.
Revision Product Analysis

43
DNA was isolated from ampicillin resistant colonies using a GeneJet Mini-Prep kit
(Fermentas). Monomer structure was then assessed by restriction enzyme digestion and a
suitable primer was chosen from those listed in Table 4.3. Sequencing was performed by
Macrogen Inc. (Seoul, Korea). Sequences were aligned and analyzed using BLAST 2 sequences
(bl2seq program, www.ncbi.nlm.nih.gov/blast/bl2seq/wblast2.cgi, (Tatusova and Madden
1999).

44
Chapter 3: Palindrome Revision is a Form of Non-Canonical End-Joining

45
Note to Reader
Screening for palindrome revision function in DNA-PKcs (+/+, +/-, -/-) and Artemis -/- was
performed by Julia Appleby, a previous MSc. student in the lab.

46
Introduction
The Line 78 mouse colony is a valuable tool for studying the effects of a chromosomally
located palindrome on genome stability. Attempts have been made to determine the enzymatic
activities necessary for palindrome processing by breeding the Line78 palindrome into mice
deficient for DNA repair, although these attempts have so far been unsuccessful. Fortunately, it
is possible to study palindrome processing in eukaryotic cells using the extrachromosomal
palindrome revision assay (Lewis 1999). I utilized this assay in cell lines deficient for NHEJ
components to determine if NHEJ is responsible for palindrome revision in eukaryotic cells.
Experimental approach
In this assay, the source plasmid from which the palindromic dimer is derived (Fig. 2.2)
is digested with BamHI and SalI to liberate the vector backbone fragment. The linearized
fragment is then ligated to obtain a palindromic dimer with a BamHI site at one axis of
symmetry and a SalI site at the other. The palindromic dimer itself is incapable of transforming
E. coli (Lewis 1999), but there is always a small residual amount of source plasmid in any dimer
preparation. When the prep is transformed into E. coli, only preps with less than 2000
ampicillin resistant colonies per µg of DNA (corresponding to residual source plasmid) were
used.
The extrachromosomal palindrome revision assay is illustrated in Figure 3.1A and is
briefly described below: 100 ng of dimer DNA was transfected into a given cell line.
Transfected cells were cultured for approximately 48 hours, at which time plasmid DNA was
harvested from the bulk pool of cells and transformed directly into E. coli. The palindromic
dimer cannot transform E. coli, but it is processed in eukaryotic cells to create a non-repeat
monomer circle. When harvested DNA is transformed into E. coli, transformants either contain
monomer DNA or source plasmid DNA; these can easily be distinguished from each other by

47
restriction enzyme digestion. Transformants containing monomers are scored, then DNA is
isolated and analyzed to assess the quality of palindrome revision junctions formed in all of the
cell lines studied.
It is also possible to measure differences in palindrome revision quantitatively (Fig.
3.1B). In these cases, 100 ng of unmanipulated source plasmid DNA is transfected in parallel
with dimer DNA to normalize for transfection efficiency. Harvested source plasmid DNA is then
Dpn1 digested to eliminate unreplicated plasmid and transformed into E. coli; transformants are
scored and used as a measure of transfection efficiency. There is no need to DpnI digest
recovered monomer products as we know they were processed in the mammalian cells by
virtue of their existence.

48
Figure 3.1A: Schematic of the qualitative extrachromosomal palindromic dimer assay. The palindromic dimer cannot transform E. coli. Upon transfection into mammalian cells, it undergoes site-specific cleavage at the symmetry axes and forms two monomer circles, which can transform E. coli. DNA is the isolated from the bacterial clones and the sequence of the revision joint is analyzed.
Transform
E. coli
(no colonies)
(colonies)
Palindromic dimer
Mammalian cells
Harvest monomers
Transform E. coli

49
Figure 3.1B: Schematic of the quantitative extrachromosomal palindromic dimer assay. The palindromic dimer and source plasmid are transfected in parallel. After harvesting, the source plasmid molecules are digested with DpnI to eliminate any unreplicated molecules. The remaining replicated molecules are transformed into E. coli and colonies are scored as a measure of transfection efficiency. Harvest monomers are transformed directly as in Fig. 3.1A. Individual revision events are scored and compared to the transfection efficiency to obtain a measurement of palindrome revision efficiency.
Revision events
Palindromic dimer
Mammalian cells
Harvest
Transform E. coli
DNA substrate
Transfection Efficiency
Dpn1 cut
B B S S
Source plasmid
B S

50
Figure 3.1C: Schematic of the extrachromosomal V(D)J assay. The V(D)J substrates contains a stop codon between the chloramphenicol resistance gene (camr) and the promoter, as such this substrate does not confer camr upon transformation into E. coli. Upon transfection into mammalian cells, the substrate can undergo V(D)J recombination, which deletes the sequence between the recombination signal sequences (blue and green arrow heads). The promoter is now located directly upstream of the camr gene. Harvested DNA contains 3 species of extrachromosomal DNA: rearranged (A), unreplicated unrearranged (B), and replicated unrearranged (C). DpnI digestion eliminates the unreplicated unrearranged species (B). The remaining DNA is transformed into E. coli. Plating on cam measures the amount of V(D)J rearrangement (A) and plating on amp measures the transfection efficiency (A + C). Colonies are scored and the values are compared to give a measure of V(D)J recombination efficiency.
V(D)J Substrate
Mammalian Cells
Harvest
Dpn1 Cut
cam to measure
V(D)J rearrangement amp to measure
transfection efficiency
(no camr colonies)
A B C
AMP

51
Results
Preliminary gene screen
To find genes involved in palindrome revision, a screen for candidate DNA repair
proteins involved in palindrome revision in mammalian cells was performed. DNA-PKcs (+/+,
+/- and -/-), Ku70 (+/-, -/-) and LigIV (+/+, -/-) were chosen based on their involvement in the
NHEJ pathway. Artemis (-/-) was chosen based on its hairpin-opening ability. ATM (ataxia
telangiectasia mutated)(+/-, -/-), a PIKK signalling kinase, was chosen because it is involved in
activating the signalling cascade resulting in cell-cycle arrest which provides the cell with
enough time to repair DSBs (reviewed in (Lobrich and Jeggo 2005; Lobrich and Jeggo 2005)).
All of the cell lines studied were able to support dimer-to-monomer resolution. A
qualitative analysis of monomers isolated from these cell lines was executed by restriction
digests and sequencing where possible. In total, 17 joints were sequenced from the Artemis -/-
cell line, 10 from DNA-PKcs +/+, 11 from DNA-PKcs +/-, 8 from DNA-PKcs -/-, 12 from ATM +/-
and 21 from ATM -/-. A selection of these sequences is displayed in Table 3.1, as well as a
selection from the Ku70 +/- and -/- and LigIV +/- and -/- cell lines (a complete list of these
junctions can be found in Table 3.S1). All of the monomers studied were variable in terms of
deletion size, insertion size and microhomology usage at the joint and there was no difference
in the degree of variability in any of the cell lines.
Joint analysis reveals common features of palindrome revision between NHEJ-proficient and
NHEJ-deficient cell lines
A more extensive study was conducted in the Ku70+/-, Ku70-/-, LigIV+/+ and LigIV -/-
cell lines (Table 3.2, Table 3.S1). In total, there were 77 monomers analyzed from LigIV+/+
cells, 67 from LigIV-/-, 73 from Ku70+/- and 51 from Ku70-/- cells. The junctions isolated from
NHEJ-deficient and NHEJ-proficient cells were compared based on 6 different criteria. The first
three categories were (1) the length of deletion from the SalI axis (0, <0.1, 0.1-1 or >1kb), (2)

52
Table 3.1: Selected monomer junction sequences.
Genotype
Input Palindrome Sequence
Symmetry Axis LEFT
Symmetry Axis RIGHT
ID#
AGCTTGGGCTGCAGGTC GACCTGCAGCCCAAGC AAGCTGATCCCCGGGGA TCCCCGGGGATCAGCTTG
Artemis -/-
AGCTTGGG GATCCCCGGGGA TCCCCGGGGATCAGCTTG 1
GGAT (-59) CAGCTTG 2
(-210) GCTTAGA (-21) 3
DNA-PK
-/-
AGCTTGGGCAGCAGG AGCTAAGTTTCTGAACTTAGGAGCTAAAGTTCCTGAA AACGTTTCAGTTTTAGTTCCTGAAAATCATGAATTGCAGGGTAT
T-CCCGGGATCAGCTTG 4
(-231) AGCTTG 5
DNA-PK +/-
AGCTTGGGCTGCAGGTC CCCGGGGATCAGCTTG 6
AGC (-39) G/T (-36) 7
DNA-PK +/+
(-208) CCCA(37bp)CTC GGGA TCCCCGGGGATCAGCTTG 8
AGCTTGG (-19) 9
ATM +/-
G (-217) (-1585) G 10
AGCTTGGGCTGCA TAGTTAAGCTGATCG CCCGGGGATCAGCTTG 11
ATM -/-
AGCTTGGGCTGCAGGTC GACCTGC CCAATCTAAATCTAAAGTATATATGAGTAAACTTGGTC (-90) C 12
AGCTTGGGCTGCAGGTC GACC ATG TCCCCGGGGATCAGCTTG 13
Ku70
+/-
AGCTTGGGCTGCAGGTC CCCGGGGATCAGCTTG 14
G (-14) (403bp)(277bp)(100bp)(~200bp)(349bp)(32bp)(165bp) (-635) T 15
AGCTTGGGCTGCAGGTC GACCT CTGG (-1115) T 16
AGCT (95bp) (22bp)A(193bp) (-1018) A 17
AGCTTGGGCTGCAGGTC GAC CTTCGGAAAAAGAGTTGGTAGCTCTTGATCC CCGGGGA TCCCCGGGGATCAGCTTG 18
Ku70 -/-
(-82) T
ATATAATATAATATATTATATATATATATATAATATATATATTATATATATAATAATATATA
TAATAAATATATTTATTATATATATAAGCA (-212) T
19
(-408) T TACG GGA TCCCCGGGGATCAGCTTG 20
AGCTTGGGCTGCAGGTC GAC TCTTCCATATT(420bp)TAG GGA TCCCCGGGGATCAGCTTG 21
AGCTTGGGCTGCAGGTC GACCTGCAGC (62bp)(226bp)ATT (-113) T 22
AGCTCA (-128) (-1504) G 23
(-263) G (263bp)TCCTCG GGGATCAGCTTG 24
Lig4 +/+
AGCTTGGCTGCAGGTC GAC T(675bp) CTAGCTGATCCCCGG GGA TCCCCGGGGATCAGCTTG 25
G (-98) 1.7kb (-201) A 26
GATCC (-417) CCGGGGATCAGCTTG 27
T (-234) (277bp)CCCCGGT (-69) T 28
AGC/TTTG/TG A (-521) 29
Lig4 -/-
AGCTTGGC GG (164bp)C (84bp) GGATCAGCTTG 30
TCCCCG/TG (-51) GGATCAGCTTG 31
AGCTTGGCTGC (-922) T 32
T (-235) (-863) A 33
AGCTTGGCTGCAGG (48bp)C (-876) C 34
AGCTTGGCTGCAGGTC GACCTGCAGCCAA (-61) T 35
Junction sequences from all genotypes studied are highly variable in terms of deletion size, insertion size and microhomology usage. Mutations from the reference sequence are indicated in purple, where the first base is the mutated base and the second is the original sequence. Insertion DNA of unknown origin is indicated in bold while insertion DNA of plasmid origin is indicated in red and insertion DNA of genomic origin is indicated in upper case blue. Underlined nucleotides indicate instances where the joint was formed using microhomology. In instances where the junction wasn‟t sequenced, the size of the deletions and insertions was estimated using restriction analysis and the approximate number of base pairs is indicated in green.

53
Table 3.2: Summary of qualitative revision product analysis. The number of junctions in each category is listed, followed by the percentage in brackets. Data from NHEJ-proficient and NHEJ-deficient cell lines was compared using a Fisher‟s Exact Test. a: all instances of joining via microhomology. For instance, if the left side of a junction was jointed to the insert via microhomology, and the insert was joined to the right side of the junction with microhomology, it was scored twice (see Table 3.1 #18 for an example). The following were considered significantly different: b:p=0.0170 c:p=0.0111
Cell
Line
Total
Left Side Deletion (kb) Right Side Deletion (kb) Insertion (kb) Intact
Sal1
site
Intact
BamH1
site
HOM
usagea 0 <0.1 0.1-1 >1 0 kb <0.1 0.1-1 >1 0 <0.1 0.1-1 >1
Lig4
+/+ 77
14
(18)
49
(64)
14
(18)
0
(0)
9
(11)
23
(30)
39
(51)
6
(8)
24
(31)
35
(45)
13
(17)
5
(7)
10
(13)
6
(8)
15
(19)
Lig4
-/- 67
22
(33)
35
(52)
10
(15)
0
(0)
16
(24)
23
(34)
27
(40)
1
(2)
29
(43)
29
(43)
6
(9)
3
(5)
18
(27)
11
(16)
20
(30)
Ku 70
+/- 73
29
(40)
35b
(48)
9
(12)
0
(0)
14
(19)
24
(33)
20
(27)
15
(21)
36
(49)
21
(29)
8
(11)
8
(11)
18c
(25)
4
(6)
34
(47)
Ku 70
-/- 51
34
(67)
9b
(17)
8
(16)
0
(0)
12
(24)
11
(22)
22
(43)
6
(11)
19
(37)
15
(29)
11
(22)
6
(12)
31c
(61)
8
(16)
25
(49)
53

54
length of deletion from the BamHI axis (0, <0.1, 0.1-1 or >1kb) and (3) length of insertion (0,
<0.1, 0.1-1 or >1kb). These criteria were chosen to reflect the variability in the joints while at
the same time allowing a simple statistical analysis. The other categories were whether the (4)
SalI or (5) BamHI site remained intact as a measure of the centre-specificity of palindrome
revision, and lastly (6) whether the joints were formed using microhomology to determine how
common its use was in palindrome revision.
There was no significant difference in any of these criteria between the LigIV+/+ and
LigIV-/- cell lines (Table 3.2). Two related categories were significantly different between the
Ku70+/- and Ku70-/- cell lines; in the Ku70-/- cell line, more monomers retained the restriction
site at the SalI axis and there were fewer monomers that had a <0.1kb deletion from the SalI
axis (Table 3.2, Fisher‟s Exact Test, p=0.0111 and 0.0170 respectively).
The sequencing results in Table 3.1 and Table 3.S1 revealed several important features
of palindrome revision in mammalian cells. Deletions in both NHEJ-proficient and NHEJ-
deficient cell lines range in size from no deletion to deletions of just over 400 base pairs from
the SalI axis. From the BamHI axis, deletions, when present, could be as large as 1.5kb.
Deletions larger than 400 base pairs from the SalI axis are not observed since either the
bacterial origin of replication (in the mouse monomer – Fig. 2.2B) or the ampicillin resistance
gene (in the human monomer – Fig. 2.2A) lies upstream; presumably, larger deletions may
occur but are excluded from analysis by the nature of the assay.
Palindrome revision joints can be formed simply from the two ends of the halved dimer
or instead may contain inserted sequences. In all 4 cases studied, the majority of the
recovered joints contained inserted DNA; 69% of the LigIV+/+, 57% of the LigIV-/-, 51% of
the Ku70+/- and 63% of the Ku70-/-. When inserted DNA is present in a joint, it can be as
little as a 1, 2 or 3 nucleotides (Table 3.1, #13), or as large as approximately 2kb (Table 3.1,
#26). The composition of the inserted DNA from one monomer to another varies. Inserted

55
DNA can be plasmid-derived (Table 3.1, #18) or genome-derived (Table 3.1, #8), but joints can
also contain insertions that appear to be neither genome- nor plasmid-derived (Table 3.1, #4).
Most monomers contain inserted DNA of mixed origin, for instance, stretches of plasmid-derived
DNA will be interspersed with stretches of untemplated DNA (Table 3.1, #15), or an insert may
contain both plasmid-derived and genome-derived DNA (Table 3.1, #30). Insertions larger than
approximately 800bp could not be sequenced in their entirety and their sizes are estimated by
restriction analysis; in these cases, the origin of the inserted DNA is not known (Table 3.1,
#15,26).
A major feature of palindrome revision is “axis-inclusive” breaks, breaks that retain the
axis of symmetry, as well as nucleotides beyond it; this is indicative of centre-directed
breakage. This occurred in all four genotypes studied, at both the SalI (Table 3.1, #12,13) and
BamHI (Table 3.1, #20,21) axes (Fig 3.2, Table 3.S1). The number of retained nucleotides at
the joints beyond the symmetry axes in all 4 cell lines was tallied and is presented as a
percentage of total joints in Figure 3.2. Up to 19 retained nucleotides were observed in any
given instance, but in the majority of the joints there were fewer than 10. There was no
significant difference between the LigIV+/+ and LigIV-/- genotypes in terms of the number of
axis-inclusive joints. There is also no significant difference in terms of the number of
nucleotides retained beyond the axis. Conversely, in both the Ku70+/- and Ku70-/- genotypes,
there are significantly more axis-inclusive joints at the Sal1 axis as opposed to the BamHI axis
(Fisher‟s exact test, p=0.0029 and 0.0015 respectively). In addition, there are also a greater
number of Ku70 -/- retained nucleotides beyond the SalI axis than Ku70+/- joints (Fisher‟s
exact test, p=0.045).
These retained nucleotides in axis-inclusive joints are reminiscent of the palindromic
nucleotides, or P-nucleotides, that arise when hairpin terminated ends are joined, as in the case
of V(D)J recombination (Lewis 1994). Thus, these retained nucleotides may be the result of

56
nicking hairpin arms arising from cruciform structures during centre break revision. The
difference in retained nucleotides between the BamHI and SalI axes may be sequence specific.
In a study which used related DNA substrates, Meier and Lewis (Meier and Lewis 1993) found
that during extrachromosomal V(D)J recombination in Ab-MLV transformed mouse cells, there
were more P-nucleotides retained at the coding end adjacent to a SalI site than that adjacent to
a BamHI site.
Joints exhibiting microhomology are common in palindrome revision (Table 3.1, #31).
All four genotypes had joints with some degree of microhomology; 17% of LigIV+/+, 31% of
LigIV-/-, 44% of Ku70+/- and 45% of Ku70-/-(Table 3.2). In each instance, the number of
homologous nucleotides was scored (Fig 3.3, Table 3.S1); the majority of microhomology-
derived joints hade between 1 and 3 homologous nucleotides. Use of 6 (Ku70 +/- and -/-) or
even 7 bases (LigIV-/-) did occur, but was rare.
There is no difference in the quantity of palindrome revision joints between NHEJ-proficient and
NHEJ-deficient cells
The efficiency of palindrome revision can be measured by performing side-by-side
transfections of the palindromic dimer and the source plasmid (Fig. 3.1B). Palindrome revision
efficiency is measured as the average number of palindrome revision events divided by the
average number of control colonies (Table 3.3). A large transfection-to-transfection variation in
palindrome revision efficiency was observed in all 4 genotypes. Revision efficiencies were
compared using a Student‟s T-test. There was no significant difference in palindrome revision
efficiency between LigIV+/+ and LigIV-/- genotypes (p=0.2) or between Ku70+/- and Ku70-/-
genotypes (p=0.7) (Fig. 3.4).

57
Figure 3.2: Nucleotides retained in axis-inclusive joints at the SalI and BamHI axes. A subset of revision joints formed were axis-inclusive, in that the symmetry axis remained intact. These joints retained a variable number of nucleotides past the symmetry axis. The number of joins that retained 1-19 nucleotides was tallied and is presented as a percentage of total junctions. Joints formed in Ku70-/- cells appear to retain more nucleotides at the SalI axis.
0
2
4
6
8
10
12
14
16
18
1 2 3 4 5 6 7 8 9 10 12 13 19
Percen
t o
f total
ju
nctio
ns
# of nucleotides retained from Sal1 axis
LigIV+/+
LigIV-/-
Ku70+/-
Ku70-/-
0
2
4
6
8
10
12
14
16
18
1 2 3 4 5 6 7 8 9 11 12 15 18
Percen
t o
f total
jun
ctio
ns
# of nucleotides retained from BamH1 axis
LigIV+/+
LigIV-/-
Ku70+/-
Ku70-/-
A
B

58
0
5
10
15
20
25
1 2 3 4 5 6 7
Pe
rce
nt
of
tota
l ju
ncti
on
s
# of nucleotides
LigIV+/+
LigIV-/-
Ku70+/-
Ku70-/-
Figure 3.3: The number of nucleotides used at junctions formed via microhomology. The number of junctions formed using a particular number of nucleotides of microhomology was scored and plotted as a percentage of total junctions analyzed. The majority of junctions formed using microhomology used only 1 or 2 nucleotides and no junctions used more than 7.

59
Table 3.3: Quantitative analysis of palindrome revision in NHEJ-proficient and NHEJ-deficient cell lines.
Genotype Average # control
coloniesa Average # palindrome
revision eventsb Palindrome revision
efficiencyc
LigIV +/+
103.1 3.825 0.03710
114.0 15.08 0.1323
11.83 6.825 0.5769
135.6 2.850 0.02102
18.48 2.725 0.1475
LigIV -/-
263.7 1.890 0.007167
12.63 1.590 0.1259
22.21 1.180 0.05313
74.53 1.540 0.02066
14.08 1.025 0.07280
Ku70+/-
2.525 1.070 0.4238
2.125 0.8200 0.3859
5.013 2.113 0.4215
Ku70-/-
0.1925 0.3175 1.649
9.750 0.1400 0.01436
0.4500 0.1500 0.3333
Palindrome revision efficiency between Lig4 +/+ and Lig4 -/- and Ku70+/- and Ku70-/- was compared by a Student‟s t-test. There was no significant difference between the NHEJ-proficient and NHEJ-deficient cell lines (Lig4, p=0.2; Ku70, p=0.7). a: Control transfections were done in duplicate. The DNA was harvested, Dpn1 digested to get rid of any unreplicated molecules, and then transformed in duplicate, for a total of 4 transformations. The number of colonies per transformation was counted and the average was taken. b: Dimer transfections were done in duplicate. The DNA was harvested from the mammalian cells and then transformed into E. coli in duplicate, for a total of 4 transformations. The number of independent revision junctions (as determined by sequencing of the junctions) was scored and the average was taken. c: Palindrome revision efficiency is measured as the average # of palindrome revision events divided by the average # of control colonies.

60
Figure 3.4: Palindrome revision efficiency in NHEJ-deficient and NHEJ-proficient cells. Palindrome Revision efficiency was compared using a Student‟s t-test. There was no significant difference between the NHEJ-proficient and NHEJ-deficient cell lines (LigIV, p=0.2; Ku70, p=0.7). Error bars represent the standard error of the mean.
Ku70-/- Ku70+/-
LigIV +/+ LigIV -/-
A
B

61
Discussion
Extrachromosomal mammalian palindrome revision occurs in the absence of NHEJ
The purpose of this study was to determine the role of NHEJ in palindrome revision in
mammalian cells. The NHEJ machinery is comprised of 8 key factors: Ku70, Ku80, DNA-PKcs,
Artemis, Pol X polymerases, XLF, XRCC4 and DNA ligase IV (reviewed in (Lieber, Ma et al.
2003)). I chose to examine two key NHEJ factors in this study. Ku70 is known to have many
diverse functions in the cell (i.e. telomere maintenance) (reviewed in (Tuteja and Tuteja
2000)); its function in NHEJ, in complex with Ku80, is to bind to the free DNA ends resulting
from a DSB, protecting them from degradation by nucleases and preventing binding of the
homologous recombination machinery (Pierce, Hu et al. 2001). DNA LigIV forms a complex
with XRCC4 and XLF and is responsible for re-joining the DNA ends after processing (Ahnesorg,
Smith et al. 2006); it has no other known function in the cell.
V(D)J recombination, a process known to occur via NHEJ, is severely crippled in the
absence of NHEJ-factors (Hesse, Lieber et al. 1989). The extrachromosomal V(D)J assays
revealed that while the LigIV+/+ and Ku70+/- were capable of producing rearranged V(D)J
products, there were no detected V(D)J rearrangement products in the LigIV -/- and Ku70 -/-
cell lines (Table 3.S1, Fig. 3.1C). Identical V(D)J assays using rearrangement substrates were
performed in the same LigIV+/+ and LigIV-/- cell lines (Grawunder, Zimmer et al. 1998). This
study reported a decrease in signal and coding joint formation of 100-1000 fold in the LigIV-/-
cell line. Moreover, in vivo plasmid end-joining assays in NII4P2 (LigIV-/-) cells and in vitro
using NII4P2 (LigIV-/-) cell extracts demonstrate a marked decrease in end-joining frequency
(~14-100-fold) as well as sequence fidelity; the joints had approximately twice as many deleted
nucleotides as the Nalm-6 (LigIV+/+) cell extracts (Smith, Riballo et al. 2003). Finally, as LigIV
has only one known function in the cell, and that is during NHEJ, it follows that if palindrome

62
revision were reliant upon LigIV and the remaining NHEJ factors, that it would be severely
crippled in its absence.
The same can be said of Ku70‟s involvement in palindrome revision. Although no
studies on V(D)J recombination in the absence of Ku70 have been performed in Ab-MLV
transformed cells, studies in ES cells and fibroblasts have shown a decrease in coding joint
formation (10-100 fold) (Zhu, Bogue et al. 1996; Bogue, Wang et al. 1997; Gu, Jin et al. 1997;
Ouyang, Nussenzweig et al. 1997). Also, plasmid end-joining studies in vitro and in vivo
demonstrate a requirement for Ku – in its absence, joints are accompanied by large deletions, a
reliance upon microhomology usage and a decreased frequency (Gu, Jin et al. 1997; Verkaik,
Esveldt-van Lange et al. 2002; Kuhfittig-Kulle, Feldmann et al. 2007).
Attempts to quantify palindrome revision were complicated (Table 3.3, Fig. 3.4), by the
variable transfection efficiencies in all four cell lines. Statistical analysis of this data did not
reveal any quantitative differences in palindrome revision efficiency but the results are
questionable due to this variation.
I observed no qualitative difference in the joints formed by palindrome revision between
NHEJ-proficient and NHEJ-deficient cell lines. In fact, in all genotypes studied (Artemis -/-,
DNA-PKcs +/+, +/- and -/-, ATM +/- and -/- as well as Ku70 +/- and -/- and LigIV +/+ and -/-)
several key features of palindrome revision became apparent: a broad spectrum of deleted DNA
from both sides of the symmetry axis, inserted DNA of multiple origins and varying lengths, use
of microhomologies and axis-inclusive joints. Although we can‟t rule out an involvement of
NHEJ entirely, it is undoubtedly not the sole and/or principal mechanism for palindrome revision
in mammalian cells.
A subset of palindrome revision joints are formed via microhomology
A microhomology-mediated end-joining (MMEJ) pathway was first identified in budding
yeast (Boulton and Jackson 1996); in yeast deficient for Ku70, end-joining occurred and was

63
accompanied by large deletions and short stretches of microhomology. Subsequent work in
NHEJ-deficient cells (Ku70, Ku80, LigIV) has further characterized this MMEJ pathway in
budding yeast (Ma, Kim et al. 2003) (Yu, Marshall et al. 2004) as well as in fission yeast
(Decottignies 2005; Decottignies 2007), Arabidopsis (Heacock, Spangler et al. 2004) and
mammals (Bentley, Diggle et al. 2004), (Liang, Deng et al. 2005; Kuhfittig-Kulle, Feldmann et
al. 2007; Liang, Deng et al. 2008). Typically, MMEJ joints contain terminal deletions up to
several hundred base pairs and somewhere between 3 and 16 bp of microhomology depending
on the study. In all of the above systems, few to no joints exhibiting defined MMEJ
characteristics are observed if NHEJ is fully functional.
Factors implicated in MMEJ in yeast are Mre11 and Rad50, Rad22, Exo1 and Pol4 (a
polymerase with gap-filling activity) (Ma, Kim et al. 2003), (Decottignies 2007). These factors
are also involved in single-strand annealing (SSA) in yeast. In humans, MMEJ is known to rely
upon the MRN complex (Mre11/Rad50/Nbs1), Fen-1 and DNA LigI and LigIII (Liang, Deng et al.
2005; Liang, Deng et al. 2008). Based on their in vitro studies with human Mre11, Paull and
Gellert (Paull and Gellert 2000) propose the following model for the function of Mre11 during
MMEJ. The model states that when a DSB occurs, the DNA ends are aligned by the MRN
complex. Mismatched ends stimulate Mre11‟s exonuclease activity, which degrades the DNA
until homology is reached. The single strand overhangs then pair with each other, preventing
further nucleation while a ligase seals one strand. The other strand undergoes further
processing until it too can be sealed and repair finished. In this model, the MRN complex
tethers the ends, preventing strand invasion of any 3‟ overhangs thereby preventing the
opportunity for filler DNA generation. Indeed, the majority of palindrome revision joints joined
by microhomology do not contain any inserted DNA (73% of LigIV+/+, 95% of LigIV-/-, 81% of
Ku70 +/- and 61% of Ku70 -/-) (Table 3.S1). In addition to its involvement in joining via
microhomologies, other activities of the MRN complex point to its potential involvement in

64
centre break palindrome revision; it is known to not only bind to hairpin ends, it also possesses
hairpin endonuclease activity (Paull and Gellert 1999) (Paull and Gellert 2000) in conjunction
with Sae2 (Lengsfeld, Rattray et al. 2007).
The majority of palindrome revision joints I observed did not use any microhomology;
those joints that did were often joined by only 1 or 2 nucleotides and none more than 7 (Fig.
3.3, Table 3.S1). This is fewer nucleotides than the typical MMEJ product although this may
reflect a difference in the experimental system – other end-joining studies in mammalian
systems were performed in vitro using cell free extracts and substrates with a defined regions
of microhomology (Bentley, Diggle et al. 2004); (Liang, Deng et al. 2005; Liang, Deng et al.
2008). If these palindrome revision joints are indeed formed by MMEJ, that only accounts for
approximately 36% of revision joints analyzed in this study. The remaining 64% must be
formed via another pathway (Table 3.S1).
Palindrome revision may be a function of non-canonical end-joining
A very small number of palindrome revision joints (approximately 10%, Table 3.4) were
joined with minimal deletions from both axes with no insertions, or small insertions that can be
explained by terminal deoxynucleotidyl transferase (TdT) activity (Mahajan, Gangi-Peterson et
al. 1999). While we can‟t rule out that in the NHEJ-proficient strains, these joints were formed
via canonical NHEJ, it is obviously not the primary end-joining pathway responsible for
extrachromosomal palindrome revision in mammalian cells.
A non-canonical end-joining pathway has been shown to be Ku-independent (Wang,
Perrault et al. 2003; Guirouilh-Barbat, Huck et al. 2004), DNA-PKcs-independent (Cheong,
Perrault et al. 1999; DiBiase, Zeng et al. 2000), DNA Ligase IV/XRCC4-independent (Wang,
Zeng et al. 2001; Smith, Riballo et al. 2003), and XL-F-independent (Zha, Alt et al. 2007). This
non-canonical end-joining pathway can join DSBs in vitro (Cheong, Perrault et al. 1999; Wang,

65
Perrault et al. 2003), in vivo (Wang, Zeng et al. 2001; Verkaik, Esveldt-van Lange et al. 2002)
as well as play a role in telomere fusion (Heacock, Idol et al. 2007), chromosomal translocations
(Weinstock, Brunet et al. 2007), IgH class switching and translocations (Yan, Boboila et al.
2007) and V(D)J recombination (Brown, Franco et al. 2002; Weinstock and Jasin 2006; Corneo,
Wendland et al. 2007). Non-canonical end-joining occurs with slower kinetics in vivo (DiBiase,
Zeng et al. 2000) than NHEJ and it is known to occur with decreased precision, resulting in
deletions. There is some evidence that non-canonical end-joining can rely on microhomology
(Corneo, Wendland et al. 2007; Audebert, Salles et al. 2008) implying that non-canonical end-
joining and MMEJ may be one and the same. Finally, when DNA-PK is fully functional, it
prevents non-canonical end-joining pathways by binding to DSB ends and shunting them
towards NHEJ (Perrault, Wang et al. 2004). It may be that in terms of DSB repair, NHEJ is the
preferred pathway; non-canonical end-joining is a back-up pathway in case NHEJ is non-
functional. Various studies have pointed to DNA Ligase III (Audebert, Salles et al. 2004; Wang,
Rosidi et al. 2005) and Poly(ADP-ribose) Polymerase-1 (Parp-1) (Audebert, Salles et al. 2004;
Audebert, Salles et al. 2008) as being involved in non-canonical end-joining.
In terms of palindrome revision, my results indicate that non-canonical end-joining is
likely to play a more important role than NHEJ. If this were the case, we would expect to see a
quantitative and/or qualitative difference between NHEJ-proficient and NHEJ-deficient cell lines.
Because this is not the case, we propose that this non-canonical end-joining pathway is the
intended mechanism for palindrome revision in mammalian cells. Furthermore, we propose that
palindrome revision may be the reason for the existence of this non-canonical end-joining
pathway. Under no other circumstances is joining a DSB with such decreased fidelity beneficial,
whereas with palindrome revision, a greater asymmetry at the joint is only further assurance
that the palindrome will not go on to cause chromosomal aberrations.

66
Inserted DNA is a common feature in both extrachromosomal and intrachromosomal palindrome
revision
Inserted, or filler, DNA is a feature of DSB repair by NHEJ in plants (Gorbunova and Levy
1997; Kirik, Salomon et al. 2000), yeast (Yu and Gabriel 1999; Haviv-Chesner, Kobayashi et al.
2007) and mammalian cells (Merrihew, Marburger et al. 1996; Lin and Waldman 2001; Lin and
Waldman 2001). In their studies on genomic DSBs, Lin and Waldman have observed inserted
DNA of both vector, genomic and unknown origin. Vector-derived inserts were the most
prevalent, occurring in approximately 50% of joints analyzed; they were approximately 150 to
490 bp in length. Inserts of unknown origin could be as large as 1490 bp. These results are
very similar to the inserted DNA found in palindrome revision joints in this study, both in terms
of origin and size (Table 3.2, Table 3.S1). Vector-derived inserted DNA was previously seen in
extrachromosomal palindrome revision joints in rodent cells (Lewis 1999).
The inserted DNA could be copied from either the Watson or the Crick strand, or even
both strands in the same insert; it could also be templated from distant or adjacent sequences
(data not shown). This hints at inserted DNA as originating from a strand invasion event as
those previously proposed (Merrihew, Marburger et al. 1996);(Gorbunova and Levy 1997);
(Zhou, Akgun et al. 2001). In the case of palindrome revision, if the hairpins are nicked
sequentially and 5‟ resection of the open end occurs, it would leave a 3‟ single stranded
overhang without another DNA end with which to interact. The 3‟ singled stranded DNA region
could then go on to invade a region of DNA via a short region of microhomology. DNA
synthesis occurs and the newly synthesized strand is immediately displaced from the template.
This newly synthesized DNA can then either join with another free end via end-joining or go on
to invade other regions of DNA, again via short microhomologous stretches. This last scenario
results in joints with short stretches of templated DNA pieced together, as well as joints with
templated DNA interspersed with non-templated DNA (Table 3.1, #15).

67
Finally, inserted DNA has been observed at rearrangement joints of two transgenic mice
lines containing inverted repeats (Collick, Drew et al. 1996). This same study observed
asymmetric deletions of variable sizes in both transgenes. This, in addition to the extensive
deletions and resulting asymmetry observed in the mouse Line78 transgenic palindrome
(Cunningham, Cote et al. 2003), indicates that the extrachromosomal palindrome revision assay
does indeed reflect processing of genomic palindromes.
Future Directions
In this study, I have demonstrated that NHEJ is not the main pathway responsible for
formation of palindrome revision joints in mammalian cells. Future research will focus on
determining which enzymes are specifically required, with particular interest being on the MRN
complex (Mre11, Rad50, Nbs1), Parp-1, LigIII and Fen-1 based on their involvement in non-
canonical end-joining pathways.

68
Table 3.S1: Sequence of palindrome revision junctions.
Genotype
Input Palindrome Sequence
Symmetry Centre LEFT
Symmetry Centre RIGHT
AGCTTGGGCTGCAGGTC GACCTGCAGCCCAAGC AAGCTGATCCCCGGGGA TCCCCGGGGATCAGCTTG
LigIV
+/+
C (-65) CT (-531) A
AGCTTGGCTGCAGGTC G G (-391) A
AGCTTGGCTGCAGG GG (-174) G
C (-46) (-28) C
AGCTTGGCTGCAGGTC GA G GCTGATCCCCGGGGA TCCCCGGGGATCAGCTTG
AGCTTGGCTGCAGGTC GACCT GA TCCCCGGGGATCAGCTTG
C (-35) CCCCC (-36) A
AG ATCAGCTTG
C (-22) (331bp) A (-191) A
C (-81) CAAGTCTCA (-264) A
AGCTTGGCTGCA CGG (-104) A
AGCTTGGCTGCAGGTC GACC CCT (-78) G
AGCTTGGCTGCAGGTC GACCTGCAG GGGA TCCCCGGGGATCAGCTTG
C (-81) CAAGTCTC (-263) A
AGCTTGGCTGCAG (-150) C
AGCTTGGC GGGGATCAGCTTG
AGCTTGGCTGCAG T A TCCCCGGGGATCAGCTTG
AGCTTGGCTGCAG TC (-446) C
AGCTTGGCTGCAGGT ATAAGCATATGTTCAGTAACAA GGGATCAGCTTG
AGCTTGGCTGCAGGTC GAC T(675bp) CTAGCTGATCCCCGGGGA TCCCCGGGGATCAGCTTG
C (-133) 400 bp (-1092bp)
T (-23) (-89) T
T (-14) AATTTTTTT (-719) A
G (-98) (154bp) 1.5kb (-201) A
AGCTTGGC GC (-427) A
AGCTTGGCTGCAGGTC GA (-177) G
(-150bp) 1kb (-131) T
T (-20) (-88) T
AGCTT (-206) A
AGCTTGGCTGCAG CC (-28) C
A (-24) GA (-185) G
T (-30) GGG (52bp) GGGAT (82bp) CCTGTTT (-149) C
T (-83) (-12) C
AGCTTGGCTGCAGGTC GACC GG (-283) G
G (-439) GA TCCCCGGGGATCAGCTTG
AGCTTGGCTGCAGGT GAC GGGGGAAG (-436) A
C (-15) CCAGCCCG GGGGATCAGCTTG
T (-22) GGCCCAAA (-183) A
AGCTTGGCTGCAGGTC G C (-15) C
AGCTTGGCTGCAGG GGGA TCCCCGGGGATCAGCTTG
AGCTTGGCTGC (-163) T
C (-418) CACAGTGGTAGTACTCCACTGTCTGGCTGTACAAAAACCCTCG GGA TCCCCGGGGATCAGCTTG
T (-202) CCCCGGGGA TCCCCGGGGATCAGCTTG
GATCC (-417) CCGGGGATCAGCTTG
A (-19) ACTAGTTAACTAGTTATAGTTAACTATAACTA (-126) T
T (-234) (277bp) CCCCGGT (-69) T
C (-93) 200bp (-265bp)
A (-74) CTCCT (-95) G
A (-97) (-200) T
T (-123) 700 bp (-265bp)
AGCTT (131bp) A GCTT
AGCTTGGCTGCA CA CCCCGGGGATCAGCTT
T (-76) (-152) G
AGCTTGGCTG GGATCAGCTT
AGCTTGGCTGC AGCG CCGGGGATCAGCTT
AGCTT (204bp) 1.3kb (-1092bp)
AGCTTGGCTGCAGG CCCTGG (-825) G
AGCT G (-526)
AGCTTGG A (-516)
AGCTTGGCTGCAGGTC GACCT AA CCCGGGGATCAGCTTG
(-90) A GA T (-213)
AGCTT T AGCTTG
AGCTTGG 600bp (-400bp)
(-200bp) 200 bp (-81) G
AGCTTGGCTGCAGGTC GACC 1kb (-1.2kb)
(-92) C T (-286) G
AGCTTGGCTGCAGGTC GACCTGCAGCCAAGCTGCG (-514) T
AGCTTGGCTGCAGGTC GACCTGCA TTGGTAACTGTCATACCACTTTTACACATATATACT(193bp)(312bp)1.5kb (-1.2kb)

69
G (-153) (-379) A
T (-267) (112bp)(104bp)(42bp)(233bp) TA (-1263) C
AGCTTGGCTGCAGGTC AT (-771) A
T (-83) CG (-515) T
T (-256) (361bp)(122bp) TACTT (-1263) C
C (-18) T (-347) A
G (-186) TTCAC (-315) C
AGCTTGGCTGCAGG (-371) A
G (-183) TTCAC (-315) C
LigIV
-/-
AGCTTGGC GG(164bp)C(84bp) GGATCAGCTTG
AGCTTGGCTGCA CGA (-108) A
AGCTTGGCTGCA CGG (-104) A
AGCTTGGCTGCAGGTC GACCTGCAGCCAA C (-14) C
A (-209) (-127) C
AGCTTGGCTGCAGG ACCGTTGATATATCCCAATGGCATCGTAAAGAACATTTTGAGGCATTTC (-876) C
AGCTT CGG (-687) G
AGCTTGGCTG (-779) A
AGCTTGGCTGCAGGT ATCAGCTTG
AGCTTGGCTGCAGGTC GAC GCTTG
AGCTTGGCTGCAGGTC GACCTG GGA (-97) A
AGCTTGGCTGC (-922) T
AGCTTGGCTGCAGGTC GACCTG GCATACCTCAAAATGTTCTTTA (-19) C
TCCCC(G/T)G (-51) GGATCAGCTTG
(-200bp) 200bp TCCCCGGGGATCAGCTTG
G (-17) GGGA TCCCCGGGGATCAGCTTG
AGCTTGGCTGC GTATAGTTCCGCACA (-671) A
A (-104) (-401) C
T (-235) (-863) A
AGCTTGGCTGCAGGTC GACCT TTT (-38) A
G (-332) TTTTT (-240) C
AGCTTGGCTGC (-860) C
AGCTTGGCTGCAGGTC 2.3 kb (-200bp)
C (-142) CTG CCGGGGATCAGCTTG
A (-25) TCCA CCGGGGATCAGCTTG
C (-23) GCCA (-540) C
G (-180) CGGAA (-280) A
TCCCCG(G/A) (-160) GGATCAGCTTG
AGCTTGG (-40) A
AGCTTG TGACCCT CGGGGA(T/C)CAGCTTG
C (-18) (-361) T
AGCTTGGCTGCAGGTC GAC (-591) A
AGCTTGGCTGCAGGTC GACCTGCAGCCAA (-61) T
AGCTTGGCTGCAGGTC GACCTGC TCCTGAAAATCTCGCCCCG GGGGA TCCCCGGGGATCAGCTTG
AGCTTGGCTGCAGGTC GAC (-583) A
AGCTTGGCTGCAGGTC GACCTG G(204bp)CCCCAGAGATCCGG(212bp) 2 kb (-875bp)
GT (-27) (-500) G
AGCTTGGCTGCAGGTC GACCTGCAG GTCGACCCCG GGGGA TCCCCGGGGATCAGCTTG
T (-50) A (-441) G
C (-18) CCTATACCATGGC GGGATCAGCTTG
C (-18) (157bp) 1.3kb (-1092bp)
AGCTTGGCTG GGA TCCCCGGGGATCAGCTTG
AGCTTGGCTGCAGGTC GACC ACAGTGCTACAGACTGGAACAAAAACCCTGCAGTCGACTCTCATCGATG GGGGATCAGCTTG
AGCTTGGCTGCAGGTC GACC CGGGGA TCCCCGGGGATCAGCTTG
AGCTTGGCTGCAGGTC GACCT AGGCCAAGGTTAAAAAAATGCTTTGCATTTTTTACATGCTTAAT G (-704)
AGCTTGGCTGCAG CA GA TCCCCGGGGATCAGCTTG
AGCTTGGCTGCAGGT A T (-35)
AGCTTGGCTGCAGGT T (-557)
AGCTTGGCTGCAGGTC GAC A (-148)
(-62) T GAGATAATTGACATACTGTTATGTCAATATA CCCGGGGA TCCCCGGGGATCAGCTTG
AGCTTGGCTGCAGG AAA ATCAGCTTG
AGCTTG A TCCCCGGGGATCAGCTTG
AGCTTGG A TCCCCGGGGATCAGCTTG
AGCTTGGCTGCAGGTC G T (-526)
AGCTTGGCTGCAGGTC G GGGA TCCCCGGGGATCAGCTTG
AGCTTGGCTGCAGGT GGGATCAGCTTG
AGCTTGGCTGCAGG GA TCCCCGGGGATCAGCTTG
AGCTTGGCTGCAGGTC GAC CCGGGGA TCCCCGGGGATCAGCTTG
(-100bp) 500 bp GGGA TCCCCGGGGATCAGCTTG
AGCTTGGCTGCAGGTC GAC CGGGGA TCCCCGGGGATCAGCTTG
AGCTTGGCT GGGG (-581) G

70
C (-94) CC (542bp)… …GGATCAGCTTG
A (-25) GGGC (-61) T
G (-183) TTCACCCTTGTT (-315) C
AGCTTGGCTGCAGGTC GACCT A(155bp)TTGGTCGAGG (-955) T
AGCTTGGC G GATCAGCTTG
Ku70+/-
A (-40) (778bp) (-939) T
AGCTTGGGCTGCAG 1.3kb (-489) A
AGCTTGGGCTGCA (-207) C
G (-14) (403bp)(277bp)(100bp)200bp(349bp)(32bp)(165bp) (-635) T
AGCTTGGGCTGC 4kb (-265bp)
(-50bp) 500bp (-265bp)
AGCTTGGGCTGCAGG GGATCAGCTTG
AGCTTGGGCTGCAGG GGGATCAGCTTG
AGCTTGGGCTGCAGGTC CCCGGGGATCAGCTTG
A(-133) ATTAATTTGAGTTAATGTGAGTTAGCATA CCCCGGGGATCAGCTTG
AGCTTGGGCTGCAGGTC GA TCCCCGGGGATCAGCTTG
A(-112) CTCAACCCCCGGGTTGTTTGTTATCCATACCA A TCCCCGGGGATCAGCTTG
(-435bp) TGTCAGACATCTCAAGATGAGTGTAG(363bp) 1.2bp CGGG(G/A)ATCAGC(G/T)T
G
(-435bp) 1275bp (-1169bp)
G (-111) 500bp (-265bp)
CA (-38) (-65) C
T(-33) (-29)C
AGCTTGGGCTGCAGGTC GAC (-20) C
AGCTTGGGCTGCAGGTC GACCTGCA (96bp)T GA TCCCCGGGGATCAGCTTG
AGCTTGGGCTGCAGGTC GACCTGCA TACAAGCTGTATGCTT CCGGGGATCAGCTTG
AGCTTGGGCTGCAGG CAAGATTTTCAAGATTTTGGG (-20) C
ATTA (-25) (-578) T
AGCTTGGGCTGCAGGTC GACCTG (670bp) 1.4kb (-265bp)
G (-11) A (-87) T
AGCTTGGGCTGCAG (672bp) (350bp) (-265bp)
AGCTTGGGC A (-180) T
AGCTTGGGCTGCAGGTC GACCT CTGG (-1115) T
AGCTTGGGCTGCAGGTC TACTGGGTTGCAATTTCGATCTGCTACCATG (-339) G
AGCTTGGGCTGCAGGTC GACCT CTGG T (-107)
TCC (-91) (-793) (G/C)
AGCTTGGGCTGCAGGTC GACC (27bp)(27bp)(36bp) GGA TCCCCGGGGATCAGCTTG
C (-18) TGCCTTACTAATGCCTTACTAATGACCGT A TCCCCGGGGATCAGCTTG
AGCTTGGGCTGCAGGTC GAC (728bp) 1.27kb (-265bp)
AGCTTGGGCTGCAGGTC T (-811)
A T (-27)
AGCTTGGGCTGCAGGTC GACCTGC TGGGCTGT (-1187) G
AGCTTGGGCTGCAGGTC GACC CGGGGATCAGCTTG
AGCTTGGGCTGCAGG CGCTTAAT G (-1445)
(-37) G A (-48)
AGCTTGGGC GTAATCAGATGGTAAGCACTGGGGCCAGATGGTAAGCCC GGGGA TCCCCGGGGATCAGCTTG
AGCTTGGGCTGCAG A (-1125)
(-83) A CTG A (-1600)
(-89) TTC(A/C)GG (-32) A
AGCTTGGGCTGCAGGTC GA TCCCCGGGGATCAGCTTG
AGCT (94bp)CTGGTGATATGGGATAGTGTTCA(193bp) (-1018) A
AGCTTGGGCTGCAGGTC GA TGTAATTTATGTAATTGA T (-1676)
AGCTTGGGCTGCAGGTC GACCTG A TCCCCGGGGATCAGCTTG
AGCTT T (-1807)
AGCTTGGGCTGCAGGTC GACCT GA TCCCCGGGGATCAGCTTG
(-97) T CCCGGGGATCAGCTTG
TGA (-31) (-759) A
G (-334) 500bp (-1.5kb)
AGCTTGG (-69) A
AGCTTGGGCTGCAGGTC G GGGGATCAGCTTG
AGCTTGGGCTGCAGGTC GACCT TTTTTTT (-716) A
AGCTTGGGCTGCAGGT G(109bp)TTACGCAAATCTCCCCGGCT GA TCCCCGGGGATCAGCTTG
AGCTTGGGCTGCAGGTC GACCTGC (40bp)TG C (-1510)
(-465bp) CCCCGGGGATCAGCTTG
AGCTTGGGCTGCAGGTC GACCT GA TCCCCGGGGATCAGCTTG
AGCTTGGGCTGCAGGTC GAC T (-1193)
G (-20) (778bp) 1kb (-265bp)
AGCTTGGGCTGCAGGTC GAC CTTCGGAAAAAGAGTTGGTAGCTCTTGATCC CCGGGGA TCCCCGGGGATCAGCTTG
AGCTTGGGCTGCAGGTC G (253bp) 1.5kb (-265bp)
AGCTTGGGCTGCAGGTC G GGGA TCCCCGGGGATCAGCTTG
(-25) A (-766) A

71
AGCTTGGGCTGCAGGTC GA T (-1480)
AGCTTGGGCTGCAGGTC CCCGGGGATCAGCTTG
(-54) CAA (56bp)AT CCCCGGGGATCAGCTTG
(-406) G (-500) T
(-349) A A (-1117)
AGCTTGG TTG
AGCTTGGGCTGCAGG CA(571bp)(51bp) C (-1675)
AGCTTGGGCTGCAGG A (-40)
AGCTTGGGCTGCAGGTC CGGGGATCAGCTTG
AGCTTGGGCTGCAGGTC GAC CCCCGGGGATCAGCTTG
Ku70-/-
AGCTTGGGCTGCAGGTC GACCTGCAGCCA TCGCAGCCTGAATGG(55bp)G GATCCCCGGGGA TCCCCGGGGATCAGCTTG
AGCTTGGCTGCAGGTC GACCTG AAAATCTCGCCAAGCTGATCCCC CCCGGGGATCAGCTTG
AGCTTGGCTGCAGGTC GACCTGC (-476) T
(-82) T ATATAATATAATATATTATATATATATATATAATATATATATTATATATATAATAATATATATAAT
AAATATATTTATTATATATATAAGCA
(-212) T
AGCTTGGCTGCAGGTC GAC AAGCT GA TCCCCGGGGATCAGCTTG
(-408) T TACG GGA TCCCCGGGGATCAGCTTG
AGCTTGGGCTGCAGGTC GACCTGC (-53) A
AGCTTGGGCTGCAGGTC GAC TCTTCCATAT(421bp)TAG GGA TCCCCGGGGATCAGCTTG
AGCTTGGCTGCAGGTC GACCTGCA (103bp)CTTAGCTCCTGAAAATCTCGCCAAGCTAATTTTTAACCAA (-37) A
AGCTTGGCTGCAGGTC GACCTGCA 100bp (-100bp)
AGCTTGGCTGCAGGTC GACCTGCAGC 800bp (-200bp)
AGCTTGGCTGCAGGTC GACCTG TA (-142) A
AGCTTGGCTGCAGGTC GAC (-510) C
(-405) GG ATCAGCTTG
AGCTTGGCTGCAGGTC GACCTGCAGCCA TCGCCAAGCTGATCCCCGGGGATC1.3kb (-265bp)
AGCTTGGGCTGCAGGTC GAC C (-564)
AGCTTGGCTGCAGGTC GACCTGCAGCCA TCGCCAAGCTGATCCCCGGGGATCC CCCGGGGA TCCCCGGGGATCAGCTTG
AGCTTGGGCTGCAGGTC GAC C (-110)
AGCTTGGCTGCAGGTC GAC C(856bp)650bp (-265bp)
AGCTTGGCTGCAGGTC GACCTGCA (686bp)900bp (-265bp)
AGCTTGGG (84bp)(219bp)GCTGAA (-475) T
AGCTTGGGCTGCAGGTC GACCTGCAGC (62bp)(226bp) ATT (-113) T
AGCTTGGGCTGCAGGTC GAC
(737bp)
750bp
(-265bp)
AGCTTGGG (159bp)G (-32) G
AGCTTGGGCTGCAGGTC GACCTG ACTATTC GGA TCCCCGGGGATCAGCTTG
AGCTTGGGCTGCAGGTC T (-45)
(-100bp) 1.5bp TTG
(-45) GGA T (-168)
AGCTTGGGCTGCAGGTC 600bp (-100bp)
A (-236) C (-1619) A
AGCTTGGGCTGCAGGTC GA (-573) A
AGCTCA (-128) (-1504) G
T (-93) (-85) A
AGCTTGGGCTGCAGGTC GACCTGCA TCGCCAAGCTGATCCCCGGGGATCC CCGGGGA TCCCCGGGGATCAGCTTG
AGCTTGGGCTGCAGGTC GACC 1.5kb (-265bp)
CC (-18) (-6) G
AGCTTGGGCTG 300bp GGGA TCCCCGGGGATCAGCTTG
AGCTTGGGCTGCAGGTC GACCTGC A (-141)
AGCTTGGGCTGCAGGTC GACCTGCAGCCCAAGCTG (123bp)AGAAAA (-983) G
(-278) A ATCCCCGGGGA TCCCCGGGGATCAGCTTG
AGC CCGGGGATCAGCTTG
AGCTTGGGCTGCAGGTC GACCTGCA 89bp A TCCCCGGGGATCAGCTTG
(-324) ACC C (-1441)
(-263) G (263bp)TCCTCG GGGATCAGCTTG
AGCTTGGGCTGCAGGTC GACCTGC CAGT G (-689)
AGCTTGGGCTG G (-67)
AGCTTGGGCTGCAGGTC GACCTGCAG TAATGTCATGATAATAATGGTTTCTTAGACGTCAGGTG C (-1510)
AGCTTGGGCTGCAGGTC GACCTGCA ACAACAACA A (-671)
AGCTTGGGCTGCAGGTC GACCTGCAG (38bp) CTTAGGAAATAGTAGA G (-1514)
AGCTTGGGCTGCAGGTC GAC C (-1759)
AGCTTGGGCTGCAGGTC GAC C (-438)
Mutations from the reference sequence are indicated in purple, where the first base is the mutated base and the second is the original sequence. Insertion DNA of unknown origin is indicated in bold while insertion DNA of plasmid origin is indicated in red and insertion DNA of genomic origin is indicated in upper case blue. Underlined nucleotides indicate instances where the joint was formed using microhomology. In instances where the junction wasn‟t sequenced, the size of the deletions and insertions was estimated using restriction analysis and the approximate number of base pairs is indicated in green.

72
Chapter 4: The Stabilizing Effects of ∆4bp on the Line78 Palindrome

73
Note to Reader
Line 78 ∆4bp germline analysis, fetal liver harvests and Ab-MLV infections were performed by
Ming Li, a technician in the lab.

74
Introduction
The Line78 transgenic mouse contains a 15.4kb perfect palindrome (Akgun, Zahn et al.
1997). The intact transgene undergoes centre-directed rearrangements, predominantly
deletional, in the germline (Akgun, Zahn et al. 1997) as well as in somatic cells, where it
rearranges at a rate of 0.55% per population doubling (Cunningham, Cote et al. 2003).
The centre break palindrome revision model, as described in Chapter 1, proposes that
DNA containing a palindromic sequence extrudes a cruciform structure and, in the case of the
Line78 palindrome, leads to transgene rearrangements. A nick is introduced at the tips of the
hairpins. Branch migration and resorption of the cruciform occur, resulting in a double strand
break (DSB). The ends are processed by nucleases and polymerases and eventually sealed
(Fig. 1.7). As we learned in Chapter 3, end-processing and joining during palindrome revision
occur via a non-homology based end-joining mechanism, either NHEJ or an alternate pathway.
However, the formation of a cruciform, a crucial step in centre break palindrome revision,
remained unconfirmed. A series of spontaneously arising mutations near the symmetry centre
provided an opportunity to further examine the centre break palindrome revision model in
particular, as well as palindrome metabolism in general.
The first spontaneously arising mutation occurred during attempts to determine the
enzymes responsible for rearrangements of the Line78 palindrome. Line78 mice were bred into
a WRN/hel background (Lebel and Leder 1998) with hopes of obtaining a mouse with an input
version of the Line78 transgene in a hel/hel background. A cell line thought to contain an
intact Line78 transgene in a WRN/hel background aroused suspicion because it was
anomalously stable. DNA sequence analysis was performed and a „tcc‟ mismatch near the
symmetry centre was identified (Fig. 4.1B) (Appleby 2005). This led towards a search for small
stabilizing modifications of the Line78 palindrome.

75
In the course of examining the origin of the „tcc‟ mismatch, it was determined that the
mutation arose from another spontaneous mutation of the Line78 perfect palindrome. The „g‟
insertion is between three and six base pairs away from the symmetry centre (Fig. 4.1C). It
originated in the germline of the wild type Line78 mouse colony and went undetected through
several generations. A cell line was created to analyze the rate of rearrangement of the „g‟
insertion palindrome in somatic cells; this cell line was also WRN/hel. The „g‟ insertion
palindrome was found to rearrange at a rate of 0.00189 events per population doubling
(95%CI:0.0012-0.0028). None of the „tcc‟ mismatch palindrome samples were rearranged, and
so are assigned a rate of 0.0 events per population doubling (95%CI: 0-0.00015) (Appleby
2005).
The purpose of this study is to determine how effective palindrome revision is at
preventing further Line78 rearrangements, or more specifically, the minimum size of asymmetry
that will stabilize Line78. In order to determine this, I examined the effects of a third small
mutation, a 4bp deletion in the Line78 palindrome 3 bp away from the symmetry centre on the
right side; this results in a 10 bp spacer (Fig. 4.1D). These three palindromes have grossly
different effects on palindrome revision, revealing new features of palindrome metabolism in
the mammalian system.

76
Figure 4.1: Schematic of Line78 variants relevant to this study. A. Intact Line78 transgene. B. The „g‟ insertion mutation. In this case, the inserted „g‟ is depicted as
the fourth in a short stretch of 4 „g‟s; in reality, it is not clear which „g‟ would bulge out of the hairpin stem. C. The „tcc‟ mismatch mutation. D. The ∆4bp mutation.
The left panel represents the central 18 nucleotides of the Line78 palindrome. Nucleotides surrounded by
the black box represent the central PstI site, which is conserved in all four mutations. The right panel the hypothetical structure that would form upon cruciform extrusion; for simplicity, only the top strand is
represented.
CAGGGTGTG CACACCCTG
G C
T A CG
CG CG
AT
CG AT
CG
G C
T A CG
CG CG
AT CG
AT
CG
>G
G C
T A
C G C G
C T A C
C C
AT CG
G C
T A C G
G
G T
G AT
CG
CAGGGTGTG CA - - - -CTG
CAGGTCCTG CACACCCTG
CAGGGGTGTG CACACCCTG
A
B
C
D

77
Experimental approach
The experimental system for analyzing rearrangements of the Line 78 palindrome is as
follows (Fig. 4.2):
Rearrangements of the Line78 palindrome that occur meiotically or post-meiotically in
germ cells or those that occur during very early embryonic development are revealed by
breeding analysis. Here, genomic DNA isolated from the tails of the original ∆4bp mouse and
its offspring and analyzed by Southern blotting and DC-PCR (Digestion-Circularization PCR) (Fig.
4.3 C and D). Rearrangements also occur in somatic cells; these are seen either by culturing
embryonic fibroblasts or Ab-MLV transformed pre-B cells(Cunningham, Cote et al. 2003). The
latter system allows for determination of the rate of rearrangement of the Line78 palindrome.
In order to determine the rate of rearrangement of the ∆4bp Line78 palindrome in
somatic cells, permanent cell lines were established by Abelson murine leukemia virus (Ab-MLV)
infection of ∆4bp Line78 fetal livers (Fig. 4.2, steps 1 & 2). Test cultures were generated by
seeding a cloning well with a single cell bearing the ∆4bp Line78 structure at day 0 (Fig. 4.2,
step 3). The test culture was carried continuously throughout the experiment. The test culture
was then sampled at various time points by generating single cell subclones via limiting dilution
(Fig. 4.2, step 4). Subclones were expanded, DNA was extracted and each was analyzed for
rearrangement via Southern blotting (for large rearrangements) and DC-PCR (to monitor
changes near the symmetry centre). A rate of rearrangement per population doubling is
determined according to the following equation, x/y=(1-p)n (Fig. 4.2, step 5) and refined using
the Maximum Likelihood Estimate (see Chapter 2). Analysis of conversion events in the ∆4bp
Line78 palindrome was performed by DC-PCR (Fig. 4.3 C and D) followed by sequencing.

78
1
2
3
1
4
x/y=(1-p)n
5
Figure 4.2: Schematic of the Line78 rate of rearrangement assay. Cells are harvested from Line78 fetal livers (1). Permanent cell lines are generated by Ab-MLV infection, resulting in transformed pre-B cells (2). A test culture is generated from a single cell (3). The test culture was sampled by limiting dilution (4). Genomic DNA was isolated from the subcloned samples. Samples were scored as either input or rearranged based on the banding pattern in a Southern blot. Rate of rearrangement was calculated according to the equation x/y=(1-p)n (5), where x/y is equal to the fraction of subclones with an input Line78 transgene out of the total subclones analyzed, n is the number of population doublings since the culture was seeded with a single cell, and p is the rate of rearrangement per population doubling.

79
A
D
SacI SacI
2.4 15.6 8.3
EcoRI EcoRI ecI
5.6 15 8
B
Les1 Erc1
(PstI)
(PstI) ecI
PstI PstI
PstI ecI
ACLF1
ACLR1
Les1 Erc1
C
PstI
Les1 Erc1 ACLF1
Left
Les1 Erc1
PstI
Right
ACLR1
(PstI) (PstI) PstI
PROBE
2.4 2.4 3.5 3.5
PstI BamHI BamHI BamHI BamHI
3.5 3.5 4.8
BamHI BamHI BamHI BamHI

80
Figure 4.3: Schematic of the Line78 transgene and outlying areas. A: Grey rectangles represent lacZ repeats; the two defective copies of lacZ are differentiated from each other by the black arrowhead. Wavy lines represent chromosomal sequences. Pst1 sites in brackets represent sites of transgene insertion. These sites were not conserved. The black box represents the probe fragment which contains the lacZ sequence. The hatched boxes represent probe binding sites. B: Map of the integrated Line 78 palindrome showing the diagnostic digests used in this study. Sizes of probe-positive fragments are given. Dashed lines indicate fragments where the cut site lies outside of the transgene. C: Location of primers used for DC-PCR. Transgene-specific primers are shown in gray. Chromosome-specific primers are shown in black; left and right side primers are not identical. D: DC-PCR strategy. Chromosome-specific primers ACLF1 or ACRF1 are used to amplify the left and the right side of the palindrome, respectively. A first PCR reaction is performed with Les1, followed by a second round with Erc1.

81
Results
The 4bp mutation originated in the germline. As with the „tcc‟ mismatch, it was
originally thought to be intact based on Southern blotting. Because it was highly unusual for a
Line78 mouse to produce even a single litter where all transgene-bearing offspring remained
unrearranged, the structure of the transgene in the parent was further investigated. Sequence
analysis was performed and the 4bp was detected. Three cell lines were generated from 4bp
containing mice and all three contain the mutation, as verified by sequence analysis.
The 4bp palindrome was observed through one generation in the germline. Of the 11
offspring who inherited the transgene, all 11 appeared unrearranged by Southern blot (Fig.
4.4). DC-PCR and sequence analysis revealed that they all contained the 4bp (data not
shown).
To determine the rate at which the 4bp palindrome rearranges due to palindrome
revision in somatic cells, 8 samplings were done with three test cultures. The first, 78D1307-1-
2-1, was sampled at 44, 66.7, 96 and 148 population doublings. The second, 78D1307-6-1-1,
was sampled at 136 population doublings. The final, 78D1310-1-2-1, was sampled at 29.3, 52
and 133.2 population doublings. There were a total of 96 samples analyzed and 95 appeared
unrearranged based on Southern blot. The calculated rate of rearrangement was 0.00065
rearrangement events per population doubling (95%CI: 0.00028-0.0013)(Table 4.1). A
Southern blot of genomic DNA isolated from a subset of the 96 samples is shown in Figure
4.5A. Each sample is represented twice, with a SacI digest in one lane and a BamHI and PstI
co-digest in the other. The sample represented in Lanes 7 and 8 is the single rearranged
sample; the others all exhibit an input Line78 band pattern. The same sample is represented
again in Lanes 3 and 4 of Figure 4.5B, digested with BamHI alone and EcoRI respectively. This
brief mapping exercise tells us that this transgene has an extensive deletion that spans into the
left and the right flanking regions, while leaving the centre of symmetry intact, as well as

82
Figure 4.4: Southern blot analysis of Line78 4bp rearrangements in the germline. A: A male mouse known to contain a 4 base pair deletion in the Line78 transgene adjacent to the symmetry centre was bred with a Balb/c female. Mouse tail DNA from the offspring was co-digested with BamHI and PstI and was probed as indicated above. An intact control in Lane 1 indicates the 3.5 kb and 2.4 kb bands. Offspring represented in lanes 4,7,8,11,12,14,15,17,19,20 and 21 exhibited input Line 78 transgene structure; those in lanes 2,3,5,6,9,10,13,16 and 18 did not contain the transgene. B: Digestion map showing BamHI (B) and Pst1 (P). Hatched boxes represent probe binding sites.
A
3.5kb
2.4kb
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21
B
2.4 2.4 3.5 3.5
B B B B P

83
Figure 4.5: Southern blot analysis of Line78 4bp rearrangements in somatic cells.
A: Cellular DNA was co-digested with BamHI and PstI (odd numbered lanes) or SacI (even numbered
lanes). An intact cell line in Lanes 1 and 2 indicates the 2.4kb band (BamHI/PstI and SacI) and 3.5kb band (BamHI/PstI only). Lanes 3-20 are samples. Lanes 7 and 8 represent a sample with a rearranged
transgene; the remaining lanes represent input Line78 structure. High molecular weight bands in the SacI digest lanes are fragments generated from a cut site that lies outside of the transgene.
B: Cellular DNA was digested with BamHI (Lanes 1 and 3) or EcoRI (Lanes 2 and 4). Lanes 1 and 2
represent an intact Line78 transgene. Lanes 3 and 4, the same sample as lanes 7 and 8 from 1B, representing a grossly rearranged transgene.
C: Digestion map showing BamHI (B) and Pst1 (P) co-digestion and SacI (S). Hatched boxes represent probe binding sites. Corresponds to 4.5A.
D: Digestion map showing EcoRI (E) and BamHI (B). Hatched boxes represent probe binding sites.
Corresponds to 4.5B.
1 2 3 4
15
8
5.6 4.8
3.5
1
11
3.5kb
2.4kb
2
11
3
11
4
11
5
11
6
11
7
11
8
11
9
11
10
011
11
0
11
12
0
11
13 14 15 16 17 18 19 20 A
B
3.5 3.5 2.4 2.4
2.4
3.5 3.5 4.8
B B B B P
C
S S
B B B B
E E
D
5.6 15 8

84
Table 4.1: Rate analysis of the 4bp Line 78 palindrome.
4bp
Mutation Cell
Line
Days in
Culture
Population
Doublingsa (n)
Input/Totalb
(x/y)
Fraction
Input
Ratec
95% CId
78D1307-1-2-1 33 44 6/6 1.0 0.0 [0 – 0.004]
78D1307-1-2-1 50 66.7 7/7 1.0 0.0 [0 – 0.002]
78D1307-1-2-1 72 96 3/4 0.75 0.003 [0.000175 – 0.0129]
78D1307-1-2-1 111 148 24/24 1.0 0.0 [0 – 0.0003]
78D1307-6-1-1 102 136 11/11 1.0 0.0 [0 – 0.0007]
78D1310-1-2-1 22 29.3 6/6 1.0 0.0 [0 – 0.006]
78D1310-1-2-1 39 52 9/9 1.0 0.0 [0 – 0.002]
78D1310-1-2-1 100 133.3 29/29 1.0 0.0 [0 – 0.00025]
Pooled Data 95/96 0.99 0.00065 [0.00028 –
0.0013]
The rate of rearrangement was determined using the Maximum Likelihood Estimate (MLE) as described in detail in the legend to Table 1. The samplings did not differ significantly from each other (Likelihood ratio test, p-value 0.2884). For further explanation, please see sample MLE calculations. aCalculated from a doubling time of 18h. bSubclones with an input Line 78 transgene/total number of subclones analyzed. cRate of rearrangement per population doubling, calculated by MLE. dThe 95% Confidence Interval, calculated by MLE.

85
potential insertions. Extensive mapping of rearranged transgenes has been done previously
(Cunningham, Cote et al. 2003) and several categories of rearrangement have been identified.
The rearranged transgene from the 4bp does not fit into any of those categories, providing
further evidence for its complexity.
DC-PCRs was performed on all the subclones generated from the 44, 66.7, and 96
generation sampling of test culture 78D1307-1-2-1 as well as the 29.3 and 52 generation
sampling of test culture 78D1310-1-2-1. All samples generated an input sized 200 bp band
from both the left and the right side (data not shown). Sequence analysis was performed on 19
of these samples. All 19 contained the ∆4bp on the correct side, including the one rearranged
transgene. These results indicate that the ∆4bp effectively prevents further rearrangement of
the palindrome.
Discussion
This study demonstrates the effects of a small mutation on the stability of a perfect
transgenic palindrome in the mouse genome. A single mutation, a ∆4bp near the symmetry
centre, was studied. It will be compared to results from the „g‟ insertion palindrome and the
„tcc‟ mismatch palindrome (Appleby 2005).
We know that the rate of rearrangement of a perfect Line78 palindrome in a wild type
background is 0.0055 (95%CI: 0.0045-0.0067) rearrangement events per population doubling
(Cunningham, Cote et al. 2003). The „g‟ insertion palindrome rearranges at a rate 3-fold less
than that of a perfect palindrome, suggesting that a single base disruption in the symmetry of a
palindrome can measurably affect, but not prevent, its rearrangement (Appleby 2005). The
„tcc‟ mismatch has a much more dramatic effect on the stability of Line78; it rearranges at a
rate at least 35-fold less than the perfect palindrome, comparable to the effect of the ∆270bp
palindrome.

86
The rate of rearrangement of the ∆4bp palindrome is more complicated to analyze. Out
of the 96 samples analyzed, all but one were intact. Rate of rearrangement calculations show
that the ∆4bp palindrome rearranges at a rate 8-fold less than that of the intact transgene. On
the surface, it seems that the ∆4bp has only a moderate affect on centre break palindrome
revision, however this may not be the case. The one rearranged sample was mapped more
extensively than samples which appeared intact (Fig. 4.5B). It exhibited extensive deletions
from both arms of the palindrome while leaving the centre of symmetry intact. This type of
rearrangement cannot be accounted for by the centre break palindrome revision model.
Therefore, including this rearrangement event in the rate calculations results in a measure of
palindrome rearrangement not related to centre break palindrome revision. If that entire
sampling (78D1307-1-2-1 at 72 days; Table 4.1) is eliminated from the rate calculation, the rate
of rearrangement of the ∆4bp palindrome is 0.0 (95%CI: 0.0-0.0006) events per population
doubling. This indicates that the ∆4bp does indeed have an effect on cruciform formation and
therefore on centre break palindrome revision.
While stabilizing rearrangements of the Line78 palindrome have been seen in the
germline (Akgun, Zahn et al. 1997) and in somatic cells (Cunningham, Cote et al. 2003)
previously, the resultant spacers are quite large when compared to the ∆4bp. In these cases,
the interruptions in the palindrome were approximately 200 to 300 base pairs, while the ∆4bp
has a mere 10bp spacer. This is the smallest stabilizing rearrangement of the Line78
palindrome identified, even smaller than the 14bp spacer created by the „tcc‟ mismatch. It also
provides a good basis for rate comparisons with an intact Line78 transgene (Cunningham, Cote
et al. 2003) because they are in the same genetic background; the „g‟ insertion palindrome and
„tcc‟ mismatch palindrome studied are both in a WRN/∆hel background (Lebel and Leder 1998).
The ∆4bp palindrome comes from a background that clearly is competent for rearrangement.
The original ∆4bp mouse had 9 siblings that had inherited the transgene. Three of the 9

87
appeared to have unrearranged transgenes according to DC-PCR and one of these was the
∆4bp; 7 out of the 9 siblings had a transgene that had been rearranged.
Line78 Rearrangements do not occur via replication slipped mispairing
It has been proposed that the centre break palindrome revision mechanism was
responsible for palindrome rearrangements in the Line78 transgene (Cunningham, Cote et al.
2003). However, this need not be an exclusive mechanism because in cases where the
symmetry centre was not intact, replication slipped mispairing provided a possible explanation.
There is good evidence for replication slipped mispairing as a mechanism for palindrome-
stimulated deletion in E. coli. Hairpin formation occurs preferentially from ssDNA on the
lagging strand (Trinh and Sinden 1991; Rosche, Trinh et al. 1995; Pinder, Blake et al. 1998;
Bzymek and Lovett 2001). As hairpins are known to stall replication forks (Lebofsky and
Bensimon 2005), this provides an opportunity for the nascent strand to misalign to short
homologous sequence on the opposite side of the hairpin, thereby deleting the majority of the
palindrome, including the axis of symmetry after the next round of replication (Weston-Hafer
and Berg 1989; Bzymek and Lovett 2001). The key difference between these two mechanisms
is that replication slipped mispairing only requires a hairpin to form from ssDNA, while centre
break palindrome revision requires a cruciform to form from dsDNA; thus only palindromes
capable of forming cruciforms undergo centre break palindrome revision. Neither the „g‟
insertion, „tcc‟ mismatch nor ∆4bp would be predicted to prevent rearrangement if the initiating
secondary structure occurs in ssDNA. Since only the ∆4bp and the „tcc‟ mismatch had a marked
stabilizing effect, we must conclude that rearrangement occurs by a centre break mechanism
and that a single „g‟ insertion is not enough of a sequence disruption to prevent cruciform
formation. Cruciform extrusion from dsDNA can be impeded by imperfections in a palindrome
or near-palindrome as the free energy cost of incorporating unpaired structures into a cruciform
must be met by a corresponding increase in negative superhelicity (Benham, Savitt et al. 2002).

88
The decrease in the rate of rearrangement for the „g‟ insertion mismatched palindrome suggests
that while it may have an increased free energy of cruciform formation cost, it is still below the
superhelicity threshold required to drive extrusion. In contrast, the increased free energy cost
for incorporating a 10bp or 14bp loop (∆4bp and „tcc‟ mismatch respectively) into a cruciform is
likely above threshold levels.
Another study examining inverted repeat transgenes in mice has yielded similar results.
The two transgenic lines, Line 109 and OX1-5, are prone to rearrangement at the site of the
inverted repeats, either insertions or deletions, and that these modifications stabilize the
transgene from further rearrangement in both the germline and in somatic cells (Collick, Drew
et al. 1996). In E. coli studies have shown that a plasmid containing a 101bp palindrome
undergoes deletions 10-fold more than a palindrome-free plasmid; when a 14bp spacer is
introduced into the centre of the palindrome, the stimulatory effect on deletion is eliminated
(Bzymek and Lovett 2001). Also, an 8 bp spacer alleviates the deleterious effects of a perfect
palindrome on propagation of palindrome-containing phage (Chalker, Okely et al. 1993). In a
similar study assaying E. coli propagation based on plaque size, it was found that the effects of
imperfections in the near-palindrome decreased as they moved away from the symmetry
centre. This implies that intrastrand base pairing after central melting is crucial to the formation
of cruciforms, which in turn is crucial to the centre break mechanism and Line78 palindrome
rearrangements.
Future Directions
The centre break mechanism is extremely efficient at producing stabilizing
rearrangements of the Line78 palindrome, as evidenced by the fact that a single four base pair
deletion, resulting in a 10 base pair loop, can confer such stability. In the future, breeding
experiments involving Line78 and various DNA repair strains, such as that which had been
attempted with the WRN/∆hel mice, will be undertaken. Alternatively, DNA repair factors can

89
be downregulated via siRNA in somatic cells. This will allow us to determine which proteins are
responsible for the centre break mechanism in mammalian cells.
An understanding of how cells prevent palindromic instability will help us understand
how such instability can lead to the various chromosomal aberrations such as translocations and
gene amplifications that can lead to genetic disease.

90
References
Ahnesorg, P., P. Smith, et al. (2006). "XLF interacts with the XRCC4-DNA ligase IV complex to promote DNA nonhomologous end-joining." Cell 124(2): 301-13.
Akgun, E., J. Zahn, et al. (1997). "Palindrome resolution and recombination in the mammalian germ line." Mol Cell Biol 17(9): 5559-70.
Albertson, D. G. (2006). "Gene amplification in cancer." Trends Genet 22(8): 447-55.
Appleby, J. H. (2005). DNA Palindrome Metabolism. Molecular and Medical Genetics. Toronto, University of Toronto. Master of Science: 127.
Arosio, D., S. Cui, et al. (2002). "Studies on the mode of Ku interaction with DNA." J Biol Chem 277(12): 9741-8.
Ashley, T., A. P. Gaeth, et al. (2006). "Meiotic recombination and spatial proximity in the etiology of the recurrent t(11;22)." Am J Hum Genet 79(3): 524-38.
Audebert, M., B. Salles, et al. (2004). "Involvement of poly(ADP-ribose) polymerase-1 and XRCC1/DNA
ligase III in an alternative route for DNA double-strand breaks rejoining." J Biol Chem 279(53): 55117-26.
Audebert, M., B. Salles, et al. (2008). "Effect of double-strand break DNA sequence on the PARP-1 NHEJ pathway." Biochem Biophys Res Commun 369(3): 982-8.
Benham, C. J., A. G. Savitt, et al. (2002). "Extrusion of an imperfect palindrome to a cruciform in
superhelical DNA: complete determination of energetics using a statistical mechanical model." J Mol Biol 316(3): 563-81.
Bentley, J., C. P. Diggle, et al. (2004). "DNA double strand break repair in human bladder cancer is error prone and involves microhomology-associated end-joining." Nucleic Acids Res 32(17): 5249-59.
Bogue, M. A., C. Wang, et al. (1997). "V(D)J recombination in Ku86-deficient mice: distinct effects on
coding, signal, and hybrid joint formation." Immunity 7(1): 37-47. Boulton, S. J. and S. P. Jackson (1996). "Saccharomyces cerevisiae Ku70 potentiates illegitimate DNA
double-strand break repair and serves as a barrier to error-prone DNA repair pathways." EMBO J 15(18): 5093-103.
Brown, M. L., D. Franco, et al. (2002). "Role of poly(ADP-ribosyl)ation in DNA-PKcs- independent V(D)J recombination." Proc Natl Acad Sci U S A 99(7): 4532-7.
Butler, D. K., L. E. Yasuda, et al. (1996). "Induction of large DNA palindrome formation in yeast:
implications for gene amplification and genome stability in eukaryotes." Cell 87(6): 1115-22. Bzymek, M. and S. T. Lovett (2001). "Evidence for two mechanisms of palindrome-stimulated deletion in
Escherichia coli: single-strand annealing and replication slipped mispairing." Genetics 158(2): 527-40.
Caburet, S., C. Conti, et al. (2005). "Human ribosomal RNA gene arrays display a broad range of
palindromic structures." Genome Res 15(8): 1079-85. Chalker, A. F., E. A. Okely, et al. (1993). "The effects of central asymmetry on the propagation of
palindromic DNA in bacteriophage lambda are consistent with cruciform extrusion in vivo." Genetics 133(2): 143-8.
Chasovskikh, S., A. Dimtchev, et al. (2005). "DNA transitions induced by binding of PARP-1 to cruciform structures in supercoiled plasmids." Cytometry A 68(1): 21-7.
Cheong, N., A. R. Perrault, et al. (1999). "DNA-PK-independent rejoining of DNA double-strand breaks in
human cell extracts in vitro." Int J Radiat Biol 75(1): 67-81. Cho, S. K., T. D. Webber, et al. (1999). "Functional characterization of B lymphocytes generated in vitro
from embryonic stem cells." Proc Natl Acad Sci U S A 96(17): 9797-802. Collick, A., J. Drew, et al. (1996). "Instability of long inverted repeats within mouse transgenes." EMBO J
15(5): 1163-71.
Connelly, J. C., L. A. Kirkham, et al. (1998). "The SbcCD nuclease of Escherichia coli is a structural maintenance of chromosomes (SMC) family protein that cleaves hairpin DNA." Proc Natl Acad Sci
U S A 95(14): 7969-74.

91
Connelly, J. C. and D. R. Leach (1996). "The sbcC and sbcD genes of Escherichia coli encode a nuclease
involved in palindrome inviability and genetic recombination." Genes Cells 1(3): 285-91. Corneo, B., R. L. Wendland, et al. (2007). "Rag mutations reveal robust alternative end joining." Nature
449(7161): 483-6. Courey, A. J. and J. C. Wang (1983). "Cruciform formation in a negatively supercoiled DNA may be
kinetically forbidden under physiological conditions." Cell 33(3): 817-29.
Courey, A. J. and J. C. Wang (1988). "Influence of DNA sequence and supercoiling on the process of cruciform formation." J Mol Biol 202(1): 35-43.
Cunningham, L. A., A. G. Cote, et al. (2003). "Rapid, stabilizing palindrome rearrangements in somatic cells by the center-break mechanism." Mol Cell Biol 23(23): 8740-50.
DasGupta, U., K. Weston-Hafer, et al. (1987). "Local DNA sequence control of deletion formation in Escherichia coli plasmid pBR322." Genetics 115(1): 41-9.
Davison, A. and D. R. Leach (1994). "The effects of nucleotide sequence changes on DNA secondary
structure formation in Escherichia coli are consistent with cruciform extrusion in vivo." Genetics 137(2): 361-8.
Dayn, A., S. Malkhosyan, et al. (1991). "Formation of (dA-dT)n cruciforms in Escherichia coli cells under different environmental conditions." J Bacteriol 173(8): 2658-64.
Dayn, A., S. Malkhosyan, et al. (1992). "Transcriptionally driven cruciform formation in vivo." Nucleic
Acids Res 20(22): 5991-7. Decottignies, A. (2005). "Capture of extranuclear DNA at fission yeast double-strand breaks." Genetics
171(4): 1535-48. Decottignies, A. (2007). "Microhomology-mediated end joining in fission yeast is repressed by pku70 and
relies on genes involved in homologous recombination." Genetics 176(3): 1403-15. DeFazio, L. G., R. M. Stansel, et al. (2002). "Synapsis of DNA ends by DNA-dependent protein kinase."
Embo J 21(12): 3192-200.
del Olmo, M. and J. E. Perez-Ortin (1993). "A natural A/T-rich sequence from the yeast FBP1 gene exists as a cruciform in Escherichia coli cells." Plasmid 29(3): 222-32.
DiBiase, S. J., Z. C. Zeng, et al. (2000). "DNA-dependent protein kinase stimulates an independently active, nonhomologous, end-joining apparatus." Cancer Res 60(5): 1245-53.
Edwards, E. (1972). Likelihood. Cambridge, Cambridge University Press.
Eykelenboom, J. K., J. K. Blackwood, et al. (2008). "SbcCD causes a double-strand break at a DNA palindrome in the Escherichia coli chromosome." Mol Cell 29(5): 644-51.
Fodde, R., M. Losekoot, et al. (1990). "Nucleotide sequence of the Belgian G gamma+(A gamma delta beta)0-thalassemia deletion breakpoint suggests a common mechanism for a number of such
recombination events." Genomics 8(4): 732-5.
Frappier, L., G. B. Price, et al. (1987). "Monoclonal antibodies to cruciform DNA structures." J Mol Biol 193(4): 751-8.
Game, L., J. Bergounioux, et al. (2003). "A novel deletion causing (epsilon gamma delta beta) degrees thalassaemia in a Chilean family." Br J Haematol 123(1): 154-9.
Gauss, G. H. and M. R. Lieber (1996). "Mechanistic constraints on diversity in human V(D)J recombination." Mol Cell Biol 16(1): 258-69.
Gellert, M., K. Mizuuchi, et al. (1976). "DNA gyrase: an enzyme that introduces superhelical turns into
DNA." Proc Natl Acad Sci U S A 73(11): 3872-6. Gellert, M., M. H. O'Dea, et al. (1983). "Slow cruciform transitions in palindromic DNA." Proc Natl Acad Sci
U S A 80(18): 5545-9. Gibson, F. P., D. R. Leach, et al. (1992). "Identification of sbcD mutations as cosuppressors of recBC that
allow propagation of DNA palindromes in Escherichia coli K-12." J Bacteriol 174(4): 1222-8.
Glucksmann, M. A., P. Markiewicz, et al. (1992). "Specific sequences and a hairpin structure in the template strand are required for N4 virion RNA polymerase promoter recognition." Cell 70(3):
491-500. Gorbunova, V. and A. A. Levy (1997). "Non-homologous DNA end joining in plant cells is associated with
deletions and filler DNA insertions." Nucleic Acids Res 25(22): 4650-7. Gordenin, D. A., K. S. Lobachev, et al. (1993). "Inverted DNA repeats: a source of eukaryotic genomic
instability." Mol Cell Biol 13(9): 5315-22.

92
Grawunder, U., M. Wilm, et al. (1997). "Activity of DNA ligase IV stimulated by complex formation with
XRCC4 protein in mammalian cells." Nature 388(6641): 492-5. Grawunder, U., D. Zimmer, et al. (1998). "DNA ligase IV is essential for V(D)J recombination and DNA
double-strand break repair in human precursor lymphocytes." Mol Cell 2(4): 477-84. Gu, Y., S. Jin, et al. (1997). "Ku70-deficient embryonic stem cells have increased ionizing radiosensitivity,
defective DNA end-binding activity, and inability to support V(D)J recombination." Proc Natl Acad
Sci U S A 94(15): 8076-81. Guirouilh-Barbat, J., S. Huck, et al. (2004). "Impact of the KU80 pathway on NHEJ-induced genome
rearrangements in mammalian cells." Mol Cell 14(5): 611-23. Haber, J. E. and M. Debatisse (2006). "Gene amplification: yeast takes a turn." Cell 125(7): 1237-40.
Haviv-Chesner, A., Y. Kobayashi, et al. (2007). "Capture of linear fragments at a double-strand break in yeast." Nucleic Acids Res 35(15): 5192-202.
Heacock, M., E. Spangler, et al. (2004). "Molecular analysis of telomere fusions in Arabidopsis: multiple
pathways for chromosome end-joining." EMBO J 23(11): 2304-13. Heacock, M. L., R. A. Idol, et al. (2007). "Telomere dynamics and fusion of critically shortened telomeres
in plants lacking DNA ligase IV." Nucleic Acids Res 35(19): 6490-500. Henderson, S. T. and T. D. Petes (1993). "Instability of a plasmid-borne inverted repeat in
Saccharomyces cerevisiae." Genetics 134(1): 57-62.
Henthorn, P. S., D. L. Mager, et al. (1986). "A gene deletion ending within a complex array of repeated sequences 3' to the human beta-globin gene cluster." Proc Natl Acad Sci U S A 83(14): 5194-8.
Hesse, J. E., M. R. Lieber, et al. (1989). "V(D)J recombination: a functional definition of the joining signals." Genes Dev 3(7): 1053-61.
Jin, R. and R. P. Novick (2001). "Role of the double-strand origin cruciform in pT181 replication." Plasmid 46(2): 95-105.
Kim, E. L., H. Peng, et al. (1998). "Cruciform-extruding regulatory element controls cell-specific activity of
the tyrosine hydroxylase gene promoter." Nucleic Acids Res 26(7): 1793-800. Kirik, A., S. Salomon, et al. (2000). "Species-specific double-strand break repair and genome evolution in
plants." EMBO J 19(20): 5562-6. Kogo, H., H. Inagaki, et al. (2007). "Cruciform extrusion propensity of human translocation-mediating
palindromic AT-rich repeats." Nucleic Acids Res 35(4): 1198-208.
Kuhfittig-Kulle, S., E. Feldmann, et al. (2007). "The mutagenic potential of non-homologous end joining in the absence of the NHEJ core factors Ku70/80, DNA-PKcs and XRCC4-LigIV." Mutagenesis 22(3):
217-33. Kurahashi, H. and B. S. Emanuel (2001). "Long AT-rich palindromes and the constitutional t(11;22)
breakpoint." Hum Mol Genet 10(23): 2605-17.
Kurahashi, H. and B. S. Emanuel (2001). "Unexpectedly high rate of de novo constitutional t(11;22) translocations in sperm from normal males." Nat Genet 29(2): 139-40.
Kurahashi, H., H. Inagaki, et al. (2007). "Molecular cloning of a translocation breakpoint hotspot in 22q11." Genome Res 17(4): 461-9.
Kurahashi, H., H. Inagaki, et al. (2006). "Chromosomal translocations mediated by palindromic DNA." Cell Cycle 5(12): 1297-303.
Kurahashi, H., H. Inagaki, et al. (2006). "Palindrome-mediated chromosomal translocations in humans."
DNA Repair (Amst) 5(9-10): 1136-45. Kurahashi, H., H. Inagaki, et al. (2004). "Cruciform DNA structure underlies the etiology for palindrome-
mediated human chromosomal translocations." J Biol Chem 279(34): 35377-83. Kurahashi, H., T. Shaikh, et al. (2003). "The constitutional t(17;22): another translocation mediated by
palindromic AT-rich repeats." Am J Hum Genet 72(3): 733-8.
Leach, D. R. (1994). "Long DNA palindromes, cruciform structures, genetic instability and secondary structure repair." Bioessays 16(12): 893-900.
Leach, D. R., E. A. Okely, et al. (1997). "Repair by recombination of DNA containing a palindromic sequence." Mol Microbiol 26(3): 597-606.
Lebel, M. and P. Leder (1998). "A deletion within the murine Werner syndrome helicase induces sensitivity to inhibitors of topoisomerase and loss of cellular proliferative capacity." Proc Natl Acad
Sci U S A 95(22): 13097-102.

93
Lebofsky, R. and A. Bensimon (2005). "DNA replication origin plasticity and perturbed fork progression in
human inverted repeats." Mol Cell Biol 25(15): 6789-97. Lees-Miller, S. P. and K. Meek (2003). "Repair of DNA double strand breaks by non-homologous end
joining." Biochimie 85(11): 1161-73. Leng, F. and R. McMacken (2002). "Potent stimulation of transcription-coupled DNA supercoiling by
sequence-specific DNA-binding proteins." Proc Natl Acad Sci U S A 99(14): 9139-44.
Lengsfeld, B. M., A. J. Rattray, et al. (2007). "Sae2 is an endonuclease that processes hairpin DNA cooperatively with the Mre11/Rad50/Xrs2 complex." Mol Cell 28(4): 638-51.
Lewis, S. M. (1994). "P nucleotide insertions and the resolution of hairpin DNA structures in mammalian cells." Proc Natl Acad Sci U S A 91(4): 1332-6.
Lewis, S. M. (1999). "Palindromy is eliminated through a structure-specific recombination process in rodent cells." Nucleic Acids Res 27(12): 2521-8.
Lewis, S. M., S. Chen, et al. (2005). "New approaches to the analysis of palindromic sequences from the
human genome: evolution and polymorphism of an intronic site at the NF1 locus." Nucleic Acids Res 33(22): e186.
Lewis, S. M. and A. G. Cote (2006). "Palindromes and genomic stress fractures: bracing and repairing the damage." DNA Repair (Amst) 5(9-10): 1146-60.
Liang, L., L. Deng, et al. (2005). "Modulation of DNA end joining by nuclear proteins." J Biol Chem
280(36): 31442-9. Liang, L., L. Deng, et al. (2008). "Human DNA ligases I and III, but not ligase IV, are required for
microhomology-mediated end joining of DNA double-strand breaks." Nucleic Acids Res. Lieber, M. R. (2008). "The mechanism of human nonhomologous DNA end joining." J Biol Chem 283(1):
1-5. Lieber, M. R., J. E. Hesse, et al. (1988). "The defect in murine severe combined immune deficiency:
joining of signal sequences but not coding segments in V(D)J recombination." Cell 55(1): 7-16.
Lieber, M. R., H. Lu, et al. (2008). "Flexibility in the order of action and in the enzymology of the nuclease, polymerases, and ligase of vertebrate non-homologous DNA end joining: relevance to
cancer, aging, and the immune system." Cell Res 18(1): 125-33. Lieber, M. R., Y. Ma, et al. (2003). "Mechanism and regulation of human non-homologous DNA end-
joining." Nat Rev Mol Cell Biol 4(9): 712-20.
Lieber, M. R., Y. Ma, et al. (2004). "The mechanism of vertebrate nonhomologous DNA end joining and its role in V(D)J recombination." DNA Repair (Amst) 3(8-9): 817-26.
Lilley, D. M. (1980). "The inverted repeat as a recognizable structural feature in supercoiled DNA molecules." Proc Natl Acad Sci U S A 77(11): 6468-72.
Lilley, D. M. (1985). "The kinetic properties of cruciform extrusion are determined by DNA base-
sequence." Nucleic Acids Res 13(5): 1443-65. Lin, Y. and A. S. Waldman (2001). "Capture of DNA sequences at double-strand breaks in mammalian
chromosomes." Genetics 158(4): 1665-74. Lin, Y. and A. S. Waldman (2001). "Promiscuous patching of broken chromosomes in mammalian cells
with extrachromosomal DNA." Nucleic Acids Res 29(19): 3975-81. Liu, L. F. and J. C. Wang (1987). "Supercoiling of the DNA template during transcription." Proc Natl Acad
Sci U S A 84(20): 7024-7.
Lobachev, K. S., D. A. Gordenin, et al. (2002). "The Mre11 complex is required for repair of hairpin-capped double-strand breaks and prevention of chromosome rearrangements." Cell 108(2): 183-
93. Lobachev, K. S., B. M. Shor, et al. (1998). "Factors affecting inverted repeat stimulation of recombination
and deletion in Saccharomyces cerevisiae." Genetics 148(4): 1507-24.
Lobrich, M. and P. A. Jeggo (2005). "Harmonising the response to DSBs: a new string in the ATM bow." DNA Repair (Amst) 4(7): 749-59.
Lobrich, M. and P. A. Jeggo (2005). "The two edges of the ATM sword: co-operation between repair and checkpoint functions." Radiother Oncol 76(2): 112-8.
Lu, L., H. Jia, et al. (2007). "The human genome-wide distribution of DNA palindromes." Funct Integr Genomics 7(3): 221-7.
Ma, J. L., E. M. Kim, et al. (2003). "Yeast Mre11 and Rad1 proteins define a Ku-independent mechanism
to repair double-strand breaks lacking overlapping end sequences." Mol Cell Biol 23(23): 8820-8.

94
Ma, Y., U. Pannicke, et al. (2002). "Hairpin opening and overhang processing by an Artemis/DNA-
dependent protein kinase complex in nonhomologous end joining and V(D)J recombination." Cell 108(6): 781-94.
Mahajan, K. N., L. Gangi-Peterson, et al. (1999). "Association of terminal deoxynucleotidyl transferase with Ku." Proc Natl Acad Sci U S A 96(24): 13926-31.
Maringele, L. and D. Lydall (2004). "Telomerase- and recombination-independent immortalization of
budding yeast." Genes Dev 18(21): 2663-75. Maringele, L. and D. Lydall (2005). "The PAL-mechanism of chromosome maintenance: causes and
consequences." Cell Cycle 4(6): 747-51. McClellan, J. A., P. Boublikova, et al. (1990). "Superhelical torsion in cellular DNA responds directly to
environmental and genetic factors." Proc Natl Acad Sci U S A 87(21): 8373-7. McClintock, B. (1941). "The Stability of Broken Ends of Chromosomes in Zea Mays." Genetics 26(2): 234-
82.
Meier, J. T. and S. M. Lewis (1993). "P nucleotides in V(D)J recombination: a fine-structure analysis." Mol Cell Biol 13(2): 1078-92.
Merrihew, R. V., K. Marburger, et al. (1996). "High-frequency illegitimate integration of transfected DNA at preintegrated target sites in a mammalian genome." Mol Cell Biol 16(1): 10-8.
Mielke, S. P., W. H. Fink, et al. (2004). "Transcription-driven twin supercoiling of a DNA loop: a Brownian
dynamics study." J Chem Phys 121(16): 8104-12. Mizuuchi, K., M. Mizuuchi, et al. (1982). "Cruciform structures in palindromic DNA are favored by DNA
supercoiling." J Mol Biol 156(2): 229-43. Murchie, A. I. and D. M. Lilley (1987). "The mechanism of cruciform formation in supercoiled DNA: initial
opening of central basepairs in salt-dependent extrusion." Nucleic Acids Res 15(23): 9641-54. Myllykangas, S., J. Himberg, et al. (2006). "DNA copy number amplification profiling of human
neoplasms." Oncogene 25(55): 7324-32.
Naom, I. S., S. J. Morton, et al. (1989). "Molecular organization of sbcC, a gene that affects genetic recombination and the viability of DNA palindromes in Escherichia coli K-12." Nucleic Acids Res
17(20): 8033-45. Narayanan, V. and K. S. Lobachev (2007). "Intrachromosomal gene amplification triggered by hairpin-
capped breaks requires homologous recombination and is independent of nonhomologous end-
joining." Cell Cycle 6(15): 1814-8. Narayanan, V., P. A. Mieczkowski, et al. (2006). "The pattern of gene amplification is determined by the
chromosomal location of hairpin-capped breaks." Cell 125(7): 1283-96. Ouyang, H., A. Nussenzweig, et al. (1997). "Ku70 is required for DNA repair but not for T cell antigen
receptor gene recombination In vivo." J Exp Med 186(6): 921-9.
Panayotatos, N. and R. D. Wells (1981). "Cruciform structures in supercoiled DNA." Nature 289(5797): 466-70.
Paull, T. T. and M. Gellert (1999). "Nbs1 potentiates ATP-driven DNA unwinding and endonuclease cleavage by the Mre11/Rad50 complex." Genes Dev 13(10): 1276-88.
Paull, T. T. and M. Gellert (2000). "A mechanistic basis for Mre11-directed DNA joining at microhomologies." Proc Natl Acad Sci U S A 97(12): 6409-14.
Perrault, R., H. Wang, et al. (2004). "Backup pathways of NHEJ are suppressed by DNA-PK." J Cell
Biochem 92(4): 781-94. Pierce, A. J., P. Hu, et al. (2001). "Ku DNA end-binding protein modulates homologous repair of double-
strand breaks in mammalian cells." Genes Dev 15(24): 3237-42. Pinder, D. J., C. E. Blake, et al. (1998). "Replication strand preference for deletions associated with DNA
palindromes." Mol Microbiol 28(4): 719-27.
Rosche, W. A. and P. L. Foster (2000). "Determining mutation rates in bacterial populations." Methods 20(1): 4-17.
Rosche, W. A., T. Q. Trinh, et al. (1995). "Differential DNA secondary structure-mediated deletion mutation in the leading and lagging strands." J Bacteriol 177(15): 4385-91.
Rosenberg, N. and D. Baltimore (1976). "A quantitative assay for transformation of bone marrow cells by Abelson murine leukemia virus." J Exp Med 143(6): 1453-63.
Ruskin, B. and G. R. Fink (1993). "Mutations in POL1 increase the mitotic instability of tandem inverted
repeats in Saccharomyces cerevisiae." Genetics 134(1): 43-56.

95
Scholten, P. M. and A. Nordheim (1986). "Diethyl pyrocarbonate: a chemical probe for DNA cruciforms."
Nucleic Acids Res 14(10): 3981-93. Sinden, R. R. (1994). DNA Structure and Function. San Diego, Academic Press, Inc.
Sinden, R. R., J. O. Carlson, et al. (1980). "Torsional tension in the DNA double helix measured with trimethylpsoralen in living E. coli cells: analogous measurements in insect and human cells." Cell
21(3): 773-83.
Sinden, R. R. and D. E. Pettijohn (1984). "Cruciform transitions in DNA." J Biol Chem 259(10): 6593-600. Smith, J., E. Riballo, et al. (2003). "Impact of DNA ligase IV on the fidelity of end joining in human cells."
Nucleic Acids Res 31(8): 2157-67. Snoep, J. L., C. C. van der Weijden, et al. (2002). "DNA supercoiling in Escherichia coli is under tight and
subtle homeostatic control, involving gene-expression and metabolic regulation of both topoisomerase I and DNA gyrase." Eur J Biochem 269(6): 1662-9.
Sonoda, E., H. Hochegger, et al. (2006). "Differential usage of non-homologous end-joining and
homologous recombination in double strand break repair." DNA Repair (Amst) 5(9-10): 1021-9. Soulas-Sprauel, P., P. Rivera-Munoz, et al. (2007). "V(D)J and immunoglobulin class switch
recombinations: a paradigm to study the regulation of DNA end-joining." Oncogene 26(56): 7780-91.
Tanaka, H., D. A. Bergstrom, et al. (2005). "Widespread and nonrandom distribution of DNA palindromes
in cancer cells provides a structural platform for subsequent gene amplification." Nat Genet 37(3): 320-7.
Tanaka, H., D. A. Bergstrom, et al. (2006). "Large DNA palindromes as a common form of structural chromosome aberrations in human cancers." Hum Cell 19(1): 17-23.
Tanaka, H., Y. Cao, et al. (2007). "Intrastrand annealing leads to the formation of a large DNA palindrome and determines the boundaries of genomic amplification in human cancer." Mol Cell
Biol 27(6): 1993-2002.
Tanaka, H., S. J. Tapscott, et al. (2002). "Short inverted repeats initiate gene amplification through the formation of a large DNA palindrome in mammalian cells." Proc Natl Acad Sci U S A 99(13):
8772-7. Tatusova, T. A. and T. L. Madden (1999). "BLAST 2 Sequences, a new tool for comparing protein and
nucleotide sequences." FEMS Microbiol Lett 174(2): 247-50.
Tran, H. T., N. P. Degtyareva, et al. (1995). "Replication slippage between distant short repeats in Saccharomyces cerevisiae depends on the direction of replication and the RAD50 and RAD52
genes." Mol Cell Biol 15(10): 5607-17. Trinh, T. Q. and R. R. Sinden (1991). "Preferential DNA secondary structure mutagenesis in the lagging
strand of replication in E. coli." Nature 352(6335): 544-7.
Tuteja, R. and N. Tuteja (2000). "Ku autoantigen: a multifunctional DNA-binding protein." Crit Rev Biochem Mol Biol 35(1): 1-33.
Verkaik, N. S., R. E. Esveldt-van Lange, et al. (2002). "Different types of V(D)J recombination and end-joining defects in DNA double-strand break repair mutant mammalian cells." Eur J Immunol
32(3): 701-9. Walker, J. R., R. A. Corpina, et al. (2001). "Structure of the Ku heterodimer bound to DNA and its
implications for double-strand break repair." Nature 412(6847): 607-14.
Wang, H., A. R. Perrault, et al. (2003). "Biochemical evidence for Ku-independent backup pathways of NHEJ." Nucleic Acids Res 31(18): 5377-88.
Wang, H., B. Rosidi, et al. (2005). "DNA ligase III as a candidate component of backup pathways of nonhomologous end joining." Cancer Res 65(10): 4020-30.
Wang, H., Z. C. Zeng, et al. (2001). "Genetic evidence for the involvement of DNA ligase IV in the DNA-
PK-dependent pathway of non-homologous end joining in mammalian cells." Nucleic Acids Res 29(8): 1653-60.
Ward, G. K., A. Shihab-el-Deen, et al. (1991). "DNA cruciforms and the nuclear supporting structure." Exp Cell Res 195(1): 92-8.
Weinstock, D. M., E. Brunet, et al. (2007). "Formation of NHEJ-derived reciprocal chromosomal translocations does not require Ku70." Nat Cell Biol 9(8): 978-81.
Weinstock, D. M. and M. Jasin (2006). "Alternative pathways for the repair of RAG-induced DNA breaks."
Mol Cell Biol 26(1): 131-9.

96
Weston-Hafer, K. and D. E. Berg (1989). "Palindromy and the location of deletion endpoints in
Escherichia coli." Genetics 121(4): 651-8. Weston-Hafer, K. and D. E. Berg (1991). "Limits to the role of palindromy in deletion formation." J
Bacteriol 173(1): 315-8. Weterings, E. and D. J. Chen (2008). "The endless tale of non-homologous end-joining." Cell Res 18(1):
114-24.
Willwand, K., A. Moroianu, et al. (2002). "Specific interaction of the nonstructural protein NS1 of minute virus of mice (MVM) with [ACCA](2) motifs in the centre of the right-end MVM DNA palindrome
induces hairpin-primed viral DNA replication." J Gen Virol 83(Pt 7): 1659-64. Wu, H. Y., S. H. Shyy, et al. (1988). "Transcription generates positively and negatively supercoiled
domains in the template." Cell 53(3): 433-40. Yan, C. T., C. Boboila, et al. (2007). "IgH class switching and translocations use a robust non-classical
end-joining pathway." Nature 449(7161): 478-82.
Yu, J., K. Marshall, et al. (2004). "Microhomology-dependent end joining and repair of transposon-induced DNA hairpins by host factors in Saccharomyces cerevisiae." Mol Cell Biol 24(3): 1351-64.
Yu, X. and A. Gabriel (1999). "Patching broken chromosomes with extranuclear cellular DNA." Mol Cell 4(5): 873-81.
Zackai, E. H. and B. S. Emanuel (1980). "Site-specific reciprocal translocation, t(11;22) (q23;q11), in
several unrelated families with 3:1 meiotic disjunction." Am J Med Genet 7(4): 507-21. Zha, S., F. W. Alt, et al. (2007). "Defective DNA repair and increased genomic instability in Cernunnos-
XLF-deficient murine ES cells." Proc Natl Acad Sci U S A 104(11): 4518-23. Zheng, G. X., T. Kochel, et al. (1991). "Torsionally tuned cruciform and Z-DNA probes for measuring
unrestrained supercoiling at specific sites in DNA of living cells." J Mol Biol 221(1): 107-22. Zheng, G. X. and R. R. Sinden (1988). "Effect of base composition at the center of inverted repeated DNA
sequences on cruciform transitions in DNA." J Biol Chem 263(11): 5356-61.
Zhou, Z. H., E. Akgun, et al. (2001). "Repeat expansion by homologous recombination in the mouse germ line at palindromic sequences." Proc Natl Acad Sci U S A 98(15): 8326-33.
Zhu, C., M. A. Bogue, et al. (1996). "Ku86-deficient mice exhibit severe combined immunodeficiency and defective processing of V(D)J recombination intermediates." Cell 86(3): 379-89.