Embed Size (px)
Citation preview

Z. Zellforsch. 145, 417--442 (1973) © by Springer-Verlag 1973
Fine Structure and Innervation of the Avian Adrenal Gland* I I . Cholinergic I n n e r v a t i o n of Ad r e na l Chromaff in Cells
K. Unsieker
Department of Anatomy, University of Kiel (Head: Prof. Dr. Drs. h.e.W. Bargmann)
Received June 6, 1973
Summary. According to their ultrastrueture and histochemistry three types of efferent nerve fibers can be distinguished in the bird's adrenal gland. The main part is made up of eholinergic fibers recognizable by a positive reaction for acetyleholinesterase and two specific populations of granules within the synaptic ending. Synaptie vesicles measuring 300 to 500 A in diameter and dense-cored vesicles with a diameter of about 1000 ~k are discernible.
In the periphery of the gland cholinergie axons for the innervation of adrenal cells form large bundles surrounded by a perineural sheath. The bundles cross the capsule and are situated within the adrenal ehromaffin cords or at their periphery. Finally small groups of fibers enter a group of chromaffin cells which are surrounded by a basal lamina and which consist of about a dozen or more cells producing adrenaline and noradrenaline. Synaptie en- dings occur, above all in passeriform species, in the center of a ehromaffin cell complex. They are either attached to the innervated cells or their dendrite-like processes, or embedded into the cells, or connected to short spines of the innervated cells. Synaptie and dense-cored vesicles leave the bouton by exoeytosis. One synaptie terminal may innervate up to three A- or NA-cells. The existence of different types of synapses for A- and NA-cells cannot be excluded.
Key words : Adrenal ehromaffin cells - - Birds - - Cholinergie innervation - - Histoehemistry - - Electron microscopy.
Introduction
I t is well established now tha t adrenal chromaffin cells of higher ver tebrates are innerva ted by preganglionie sympathet ic nerve fibers (Feldberg and Minz, 1933 ; Feldberg et al., 1934; Goodman and Gilman, 1970). I n mammals the greatest par t of these fibers reach the adrenal gland via the splanchnie nerves ; this is indi- cated by the results of numerous degenerat ion experiments (Elliott, 1913; Botar and O'Shaughnessy, 1936; t Iol l inshead, 1936 ; Holl inshead and Finkelstein, 1937; Swinyard, 1937; Young, 1939; MacFa r l and and Davenpor t , 1941; IIi l larp, 1947; Kiss, 1951; a.o.) as well as by the invest igat ions of secretion after s t imula t ion of the splanehnie nerves (Maycoek and tIeslop, 1939 ; Marley and Prout , 1965, 1968 ; a.o.). The cell bodies of the preganglionic neurons are s i tuated p redominan t ly in the ipsilateral nucleus intermediolateral is between Th 4 and L 2 (Cummings, 1969) and leave the spinal cord by the vent ra l root (Lobko, 1969). We cannot say yet for certain whether all these results hold t rue for the innerva t ion of the av ian adrenal gland as well (el. Pet t i t , 1896a, b; Brauer, 1932). Coupland (1965) described the pa t t e rn of d is t r ibut ion of nerve fibers in the adrenal medul la of the ra t ; bu t after this excellent work nobody has yet tr ied to elucidate whether this pa t t e rn is
* Supported by a grant from the "Deutsche Forsehungsgemeinschaft" (Un 34/1).

418 K. Unsicker
representat ive for other mammal ian species and other kinds of vertebrates. One could suppose tha t different kinds of ar rangement of adrenal chromaffin cells as observed in the adrenal gland of several bird species (Unsieker, 1973a) would provoke different pat terns of distr ibution of nerve fibers. Other questions con- cerning the innervat ion of adrenal chromaffin cells which are current ly unsolved are the different innervat ion of adrenaline (A)- and noradrenaline (NA)-storing cells (Grynszpan-Winograd, 1969; Lewis and Shute, 1969) and the formation and fate of the large dense-cored vesicles in eholinergie terminals. The problem of non-eholinergie nerves in the avian adrenal gland is to be excluded from this work and dealt with in the next paper.
Materials and Methods
1. Animals Studied (/or Number, Sex, Maturity, and Date o/Fixation See Unsicker, 1973a)
Domestic Fowl, Domestic Goose, Peking Duck, Larus ridibundus (Black-headed Gull), Larus argentatus (Herring Gull), Larus marinus (Black-backed Gull), Rissa tridactyla (Kitti- wake), Uria aatge (Guillemot), Domestic Pigeon, Corvus ]rugilegus (Rook), Corvus monedula (Jackdaw), Turdus merula (Blackbird), Sturnus vulgaris (Starling), Domestic Sparrow, Fringilla coelebs (Chaffinch).
2. Methods Paraffin sections were made from adrenal glands of the Domestic Fowl, Duck, Black-
headed Gull, Black-backed Gull, Kittiwake, Guillemot, Domestic Pigeon, Rook, Starling, Domestic Sparrow, and Chaffinch, and stained with Heidenhain's azan and chromalum hema- toxylin-phloxine; semithin sections were taken from the whole araldite-embedded material.
For histochemical and electron microscopical methods see Unsicker (1973 a). In the course of an attempt to solve the problem of a different type of innerwtion of A- and NA-cells, the number and size of dense-cored vesicles, the number of synaptic vesicles and the length of postsynaptic membrane thickenings were determined in 53 synapses on A-cells and 58 synapses on NA-cells.
Observations
Domestic Fowl
Light Microscopy
Nerves to the adrenal gland enter the capsule in tangential directions. Before they reach the gland they often cross one of the periadrenal ganglia. They spread out within the superficial layer of chromaffin cells (cf. Unsieker, 1973a) or reach the interior of the organ as large bundles. Semithin sections show nerve fibers between chromaffin and interrenal cell cords, near sinusoid blood vessels or inside chromaffin cell complexes (Fig. 10). Sections incubated with aeety]thiocholine show positive reactions of the large nerve t runks inside the capsule and along chromaffin cords as well as positive reactions of a fine network of fibers which surround ehromaffin cells (Fig. 1).
Electron Microscopy
Large Nerve Trunks without Intimate Relationship to Chroma/]in and Inter- renal Cells. Large nerves in the pericapsular tissue and in the capsule are com-

Fine Structure and Innervation of the Avian Adrenal Gland 419
Fig. 1. Pekin Duck. Demonstration of acetylcholinesterase. A positive reaction can only be seen in adrenal chromaffin cell groups (A) and in a small ganglion (G). Cords of interrenM
cells (I)
posed of u n m y e l i n a t e d and mye l i na t ed fibers in a ra t io of abou t 10:1. Some of the large nerve t runks only exhib i t u n m y e l i n a t e d axons. Nerves are su r rounded b y a comple te or incomple te per ineura l sheath, the th ickness of which depends on the d i ame te r of the nerve sur rounded. I n most eases, however, there are only one or two layers of f l a t t ened per ineura l cells. They eontMn numerous vesicles and are su r rounded b y a basa l l amina which shows a super imposed layer of connect ive t issue cells. Collagenous fibers which are p r e d o m i n a n t l y a r ranged in a longi tud ina l d i rect ion are found be tween the different cell layers. Axons are a lways su r rounded b y sate l l i te cells. The number of axons which make up a bundle is g rea te r the smal ler the axons are. The d i ame te r of u n m y e l i n a t e d axons varies f rom 0.2 to 1.8 ~m, m y e l i n a t e d axons measure 1.5-2.8 ~m. Myel ina ted axons nea r ly a lways possess a satel l i te cell of thei r own. The n u m b e r of mesaxon loops is be tween 2 and a p p r o x i m a t e l y 20, bu t axons rich in myel in prevai l . Somet imes a g ranu la r ma te r i a l of high e lect ron dens i ty can be observed in the in terce l lu lar space be tween inner and ou te r mesaxon. Axons and satel l i te cells are su r rounded b y an u n i n t e r r u p t e d basa l lamina.
Nerves and Axons within Chroma/lin Cell Cords be/ore Synaptic Contacts with Chroma/[in Cells Are Made. The s t ruc ture of nerve t runks which enter subcapsu la r regions of the adrena l g land mos t l y correspond comple te ly to the nerves descr ibed above. The deeper t h e y come wi th in the g land the smal ler t h e y are. The peri- neur ium d isappears so t h a t only basa l lamina, collagenous fibers and some fibro- b las ts separa te nerves f rom chromaff in cells or the per ivascu la r space. Mye l ina ted axons decrease in number and the d i ame te r of single axons diminishes.

420 K. Unsieker
Terminal or collateral branches of such bundles invade groups of adrenaline (A)- and noradrenaline (NA)-storing cells, often in such a manner tha t the nerve can be compared to a stalk, bearing synapses and ehromaffin cells like buds and umbels (Fig. 2, 10). To reach the effeetor cells satellite cells and associated axons have to penetrate the basal lamina which surrounds single groups of ehromaffin cells. As groups of ehromaffin cells are addit ionally surrounded, as a rule, by their own satellite cells inside and a layer of collagenous fibers and fibroblasts outside the basal lamina, three layers mus t be penetra ted in all. Mostly, this penetrat ion does not occur abrupt ly near the basis of the nerve stalk entering the cell group, but satellite sheath and basal lamina which belong to the group slide in with the stalk as well. Somewhere along the stalk the basal lamina of the nerve fuses with that of the chromaffin cell group. Then, the axons of the nerve stalk enter the "somatic" satellite sheath of the ehromaffin cells.
Accordingly, the distance covered by an axon between entrance into the adrenal gland and formation of synapses on a chromaffin cell can be divided up schematiely in the following manner (Fig. i0):
i. pericapsular and capsular section; 2. perivascular or interehromaffin section; 3. nerve stalk of a small ehromaffin group;
4. preterminal section I 5. synapse t see below.
Axons belonging to sections 1, 2, or 3 essentially show the already known ul t ras t ruetural features (Fig. 4). They are covered by a tr i laminar unit membrane, 60-80 A thick, where mieropinocytosis vesicles bud off. Similar vesicles can be found inside the axon.
Neurotubules and -filaments, mixed in a varying degree, are the integrat ing parts of these axons. Tubules m ay have filaments as their continuation, in which ease the fi lament forms an extension of a par t of the wall of the tubule (Fig. ¢d). Fi laments are about 60 A, tubules 240-270 ti, thick. Tubules have a tr i laminar wall, which measures about 60 ~. They are filled with a material of medium electron densi ty and exhibit a centrally located filament. The ends of tubes some- times show buds and constrictions (Fig. 4a), f rom which vesicles with a mean diameter of 300 A can obviously develop. I t is difficult to say what rules deter- mine the distr ibution of tubules and filaments in the axon. I t is striking, however, tha t regions with m a n y filaments do not show any tubules, and vice versa (Fig. 4 a). Sometimes tubules are arranged in a ring around mitoehondria (Fig. 4b). Struc- tures which could be a hint to the existence of an axoplasmie retieulum, are rarely seen. Particles which look like ribosomes can be found, but eisternae to which they are sometimes bound, seem to be artifacts in the sections 1-3. Further- more, axons contain long slender mitoehondria, about 0.1 ~m wide: the eristae are embedded into a homogenous electron-dense matr ix and usually run in a longitudinal direction. Often, mitoehondria are t ransversed by a single longi- tudinal erista. Granular vesicles (diameter about 900 It) can be seen in m a n y axons. Within the nerve stalk of a ehromaffin group, however, they occur more regularly. I n nearly all eases the outline of the dense core is sharply demarcated, its position is str ict ly in the center of the vesicle. Clear vesicles with a diameter of 300-500 A are found less often; usually they lie in aggregates. The axoplasm
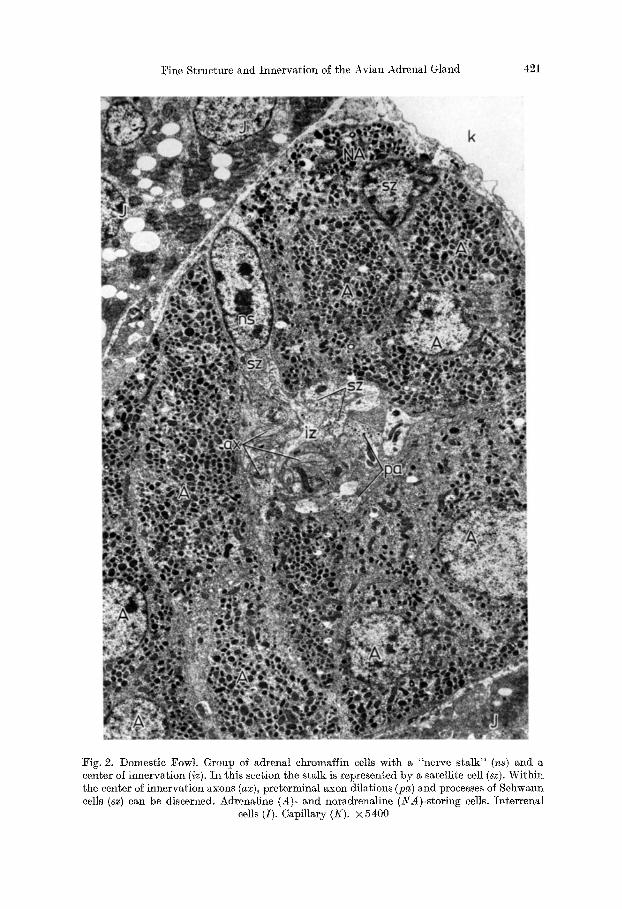
Fine Structure and Innervation of the Avian Adrenal Gland 421
Fig. 2. Domestic Fowl. Group of adrenal chromaffin cells with a "nerve stalk" (ns) and a center of innervation (iz). In this section the stalk is represented by a satellite cell (sz). Within tile center of innervation axons (ax), preterminal axon dilations (pa) and processes of Schwann cells (sz) can be discerned. Adrenaline (A)- and noradrenaline (_YA)-storing cells. Interrenal
cells (I). Capillary (K). × 5400

Fig. 3. Corvus ]rugilegus. Peripheral satellite (Schwann) cells with whorls of double mem- branes (din) and membrane-bounded cytoplasmic lamellae (cs). These lamell~e as well as the central cytoplasmic area sometimes contain dense-cored vesicles. Interrenal cell (I). Axon (a) with clear synaptic and large dense-cored vesicles contacts the surface of the interrenal cell.
× 18000
Fig. 4a - -d . Domestic Fowl. a) Axon with neurotubules (t) and -filaments ([). The end of one tubule (te) shows buds and constrictions. Note the different degree of electron density of the aXOlOlasm in the two halfs where tubules or filaments prevail. × 90000. b) Ring-like arrangement of neurotubules around a mitochondrion. × 18 000. c) Axon with myelin bodies.
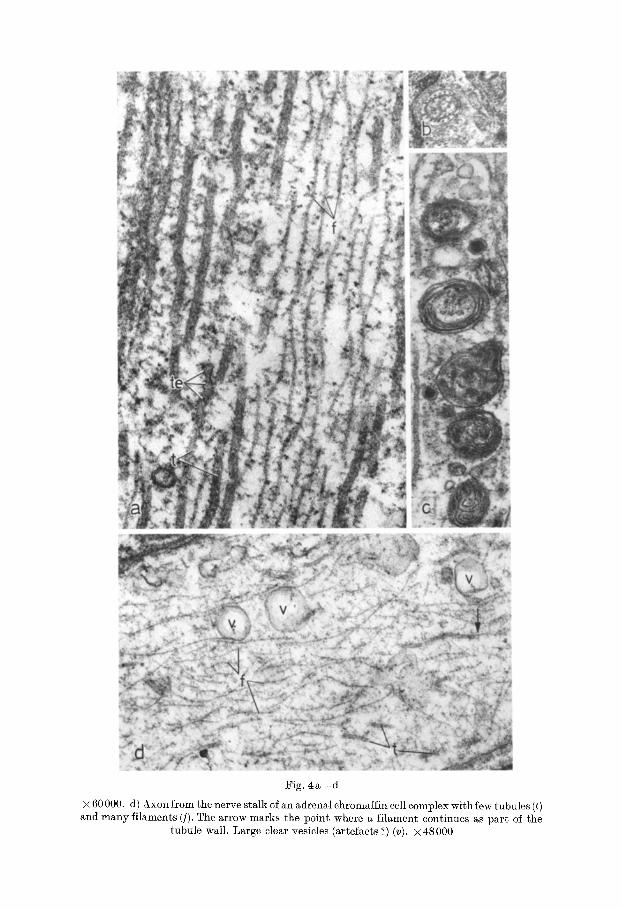
Fig. 4 a - - d
× 60000. d) Axon from the nerve stalk of an adrenal ehromaffin cell complex with few tubules (t) and many filaments (J). The arrow marks the point where a filament continues as part ol the
tubule wall. Large clear vesicles (artefacts ?) (v). ?<48000

424 K. Unsicker
which shows a low electron density on the average seems to be of higher density in such regions where tubules occur, and of lower density where filaments are situated (Fig. 4a). The intercellular cleft between axons or axons and satellite/ chromaffin cells is about 200 A wide. Apart from these components axons contain different forms of dense bodies: they are always surrounded by a unit membrane and exhibit a granular dense matrix into which myelin-like figures can be embedded (Fig. 4c). Most myelinated and non-myelinated axons show a positive reaction to acetylcholinesterase on the outer surface. I t cannot be excluded that some of the negative axons represent non-cholinergic fibers (see Unsicker, 1973 c). Schwann (satellite) cells (Fig. 2) appear "dark" or "light" according to the density of their hyaloplasm. They contain lots of rough-surfaced ER, which often forms dilated cisternae, and free ribosomes. Mitoehondria sometimes resemble those seen in axons; often, however, they are less even and show an irregular arrangement of their cristae. The Golgi apparatus is well developed. Near it one or two eentrioles can be discovered. Filaments can be observed ending within half-desmosomes. The nucleus is placed according to the possibilities given by the embedded axons. I ts profile is round or elongated and often shows indentations.
Preterminal Sections and Synapses on Chroma//in Cells. In the course of the preterminal section (Fig. 10) of an axon neurotubules and -filaments diminish in comparison with clear and dense-cored vesicles and groups of mitochondria. Synapses on chromaffin cells (Figs. 5, 6, 9, 10) are characterized by pre- and post- synaptic membranes. There is no definite border between the two regions. Some- times preterminal sections and synapses can already be seen near the base of the nerve stalk. The more the axon bundle invades the cell group, the more synapses can be observed. Axons which give rise to preterminal sections and synapses, have a diameter of 0.2-1 ~m. Presynaptie sections either have the same diameter as the axon from which they arise or show slight dilations. As a rule synapses are formed as boutons, oval in shape, and up to 2.5 ~zm in diameter. There can be a prevalence of clear synaptie vesicles either in the preterminal or in the synaptic ending (Fig. 5-7). The synaptic bouton is not always a true ending (Fig. 5). Often, it gives rise to another preterminal section which ends as a synapse. I t could not be clearly determined how many synaptie boutons can follow successively.
Concerning the connexions between synaptic boutons and chromaffin cells several variations can be observed (Fig. 6). Synaptie contacts can either be established at the cell body of the effeetor cell or at its dendritic processes (el. pigeon, Fig. 8). The latter possibility is not seen so often in the chicken. If the contact takes place on a process, the attachment zone is smooth and rectilinear. If the synapse contacts the perikaryon the following morphological variations can be distinguished: 1. the contact zone is rectilinear (Fig. 9a), 2. the synapse lies within a fold of the cell (el. rook, Fig. 6b). 3. short spines of the ehromaffin cell penetrate into the bouton (Fig. 6a). In such eases, pre- and postsynaptic mem- branes are either located beside the spine or on the spine itself. A single bouton may contact up to three cells. However, we have never seen a simultaneous innervation of A- and NA-cells.
The characteristic features of synaptic membranes can be clearly studied in Fig. 9 a. Dense particles which pile up on the inner lamella of the unit membrane build up the thickenings of the pre- and postsynaptic membrane. On the postsynaptie

Fine Structure and Innervation of the Avian Adrenal Gland 425
Fig. 5. Corvus/rugilegus. Synapse on a chromaffin cell (A). Formation of the synapse occurs at the margin of a passing axon (ax). A preterminM section is absent. Neurotubules (t) and -filaments ([). Clear synaptic vesicles (sv). Pre- and postsynaptic membrane thickenings (s).
Spine of a chromaffin cell (sp). x 60000
side par t ic les form small spikes and a hi l ly relief. I n numbers : the th ickness of the pos t synap t i c m e m b r a n e varies f rom 150 to 200 A, t h a t of the p resynap t i c side f rom 80 to 450 A. The synap t i c cleft often exhibi ts a med ian lamella , 30-50 A thick. I n some places f i laments or canalieul i seem to br idge the cleft. The length of the pos t synap t i c m e m b r a n e th ickening of ~NA-cells is 0.54 ~m on the average (58 synapses measured) , t h a t of A-cells 0.47 btm (53 synapses measured) . However , the v a r i a b i l i t y is so great , t h a t this resul t cannot be e s t ima ted s ta t i s t ica l ly . Clear synap t i c vesicles are nea r ly a lways found in close p r o x i m i t y to the p resynap t i c

Fig. 6a--d . Domestic Fowl (a) and Corvus /rugilegu8 (b--d). Several types of cholinergie synapses on chromaffin cells, a) Bouton (b) on an adrenaline (A)-storing cell. Pre- and post- syaaptic membrane thickenings (8). Spine of the chromaffin cell (sp). Invaginated part of the bouton (ba) inside the chromaffin cell. × 24000. b) Deep invagination of two synaptie endings (b) showing synaptic and large dense-cored vesicles, mitochondria and glycogen- particles within a noradrenaline (NA)-storing cell. Pre- and postsynaptic membrane thicken- ings (8). × 18000. c) Presynaptic terminal (ps) with a bouton-like synaptic protrusion (s) extending into a :NA-cell. × 18000. d) Innervation of two NA-cells by a single synaptic
terminal (8). Presynaptie terminal (ps). × 18000

Fine Structure and Innervation of the Avian Adrenal Gland 427
Fig. 7. Fringilla coelebs. Clusters of voluminous loresynaptic and synaptic terminals in the center of a chromaffin cell group. Noradrenaline (NA)-storing cells. × 18000
membrane . Most ly t hey are e l l ip toid in shape, the smal les t d i ame te r is 300-500 A. Some vesicles touch the m e m b r a n e or fuse wi th it, so t h a t the in ter ior communi- cates wi th the synap t i c cleft (cf. rook, Fig. 9d). The number of synap t i c vesicles in A-synapses is 80.5 on the average, t h a t in NA-synapses 85.1. A stat is t icM evMution was not possible.
Dense-cored vesicles are ano ther popu la t ion of vesicles ob l iga to ry for synap t i c te rmina ls (Fig. 6). I n the dense core g lobular par t ic les can often be discerned. Be tween the dense core and the uni t m e m b r a n e a 60-120 • wide cleft is inter- posed. Somet imes the uni t m e m b r a n e bears a fuzzy coat. Mostly, g ranu la r vesicles are round and cannot be d is t inguished f rom those wi th in o ther segments of the
29a Z. Zel]forsch., ]3d. 145

Fig. 8. Domestic Pigeon. Synaptie endings (s) on a dendrite-like process (dA) and on a "peri- karyon" (pA) of two chromaffin cells. × 30000
Fig. 9. a) Domestic Fowl. Synaptic terminal on a noradrenaline (NA)-storing cell. Synaptic vesicles (sv). Pre- (pr) and postsynaptlc (pc) membrane thickenings. The synaptic cleft contains a th in lamella and is bridged by fine filaments and canaliculi (/). × 120 000. b) Domestic Fowl. Synaptic terminal with a formation which resembles a Golgi apparatus (g). A coated vesicle sits on a cisterna like a cap (8k), another cisterna contains electron-dense material (Ze). Synaptic (sv) and dense-cored vesicles (dv). Pre- and postsynaptic membrane thickenings (s). × 60000. c) Synaptic ending with a "Golgi equivalent" . Remnants of a coated vesicle (sk) and the beginning deposition of electron-dense material (ze) can be seen. × 120000. d) Corvus /rugilegus. The contents of a synaptic vesicle (Sv) communicates with the synalotic cleft (sp). × 180000. e) Domestic Fowl. Exocytosis of a dense-cored vesicle ( ~ ) i n the preterminal
portion of an axon. × 60000

Fine S t ruc t u r e a n d I n n e r v a t i o n of t he A v i a n Adrena l Gland 429
29b Z. Zellforsch., Bd. 145
Fig. 9 a - - e

430 K. Unsicker
axon. Dense-cored vesicles of A-synapses have a mean diameter of 980 • (variabili- ty 541-1303 ~), those of NA-synapses a mean diameter of 907 • (variability 733-1209 A). The differences in the variability are striking. Granular vesicles can probably be extruded from the axon by exocytosis (Fig. 9 e). Clear vesicles measuring 500-1500 A are only seldom found. Furthermore, mieropinoeytosis and coated vesicles occur in synapses. Their small invaginations can be observed all along the plasmalemma. Inside the synaptic terminal mieropinocytosis vesicles often cannot be discerned from synaptie vesicles. Coated vesicles are apparently trans- ported to a synaptie equivalent of the Golgi apparatus, where they can fuse with dilated eisternae containing an electron-dense material (Figs. 9b, e). The mito- ehondria of the terminals can often be distinguished from those in other axonal sections by their compact shape and disintegrated matrix. Only sometimes does a subsynaptic web occur (Fig. 9d).
Exocytosis of synaptie vesicles increases after application of 6-hydroxydop- amine. Synaptie terminals show on their surface a positive reaction for aectyl- thioeholinesterase. The interior is free from reaction products.
Other Bird Species. The non-passeriform birds show no variations concerning the eholinergic innervation of ehromaffin cells and the distribution pat tern of nerve fibers.
In Corvus ]rugilegus (Rook) the principle of chromaffin cell innervation is particularly clear. Chromaffin cell cords show bundles of longitudinally oriented nerve fibers from which smaller groups of axons branch off to innervate single A- and NA-cells. The light- and electron microscopical demonstration of aeetyl- eholinesterase facilitates the recognition of this mode of innervation. Subsynaptic webs can often be observed (Fig. 9d). Some Schwann cells contain eontinous whorls of double membranes which surround a central cytoplasmic area. The membrane-bounded cytoplasmic lamellae sometimes show dense-cored vesicles (Fig. 3). These formations resemble similar systems of membranes in the super- ficiM glia covering of the cat 's visual cortex (I-Iaug, 1972).
Corvus moneclula (Jackdaw), Turdus merula (Blackbird), and Sturnus vulgaris (Starling) show no peculiarities in the cholinergie innervation of chromaffin tissue.
In the Domestic Sparrow non-myelinated axons and Sehwann cells enter the follicles (Unsieker, 1973a) and form synaptie endings on the surface of ehromaffin cells.
Among all bird species investigated Fringilla coelebs exhibits most distinctly the mode of innervation typical for avian ehromaffin cells. Electron microscopical surveys demonstrate dusters of voluminous presynaptic sections and synaptic endings in the colloid-filled center of ehromaffin cell groups (Fig. 7), so tha t poles of innervation and secretion are dear ly discernible.
Discussion
Our light microscopical results on the entrance of nerves to the avian adrenal gland and their distribution within the organ generally confirm the former results of Giaeomini (1898), Kura (1927), Knouff and Ha r tman (1951) and Shioda and Nishida (1967). Bundles of nerves enter the capsule tangentially and join the chromaffin cell cords. Dense meshworks of nerve fibers in the capsule of the

Fine Structure and Innervation of the Avian Adrenal Gland 431
adrenal gland are a common characteristic feature of birds and mammals (for the latter see Ph. StShr jr., 1957). Furthermore, our ultrastruetural and histoehemical results confirm tha t most nerve fibers are non-adrcnergic, non-peptidergic (purinergic) (cf. Unsicker, 1973 c), which means, with a high degree of probability, cholinergic. The morphological identification of eholinergic axons is mainly a proof per exclusionem (no small dense-cored vesicles, no varieosities, no degenerative alterations following application of 6-hydroxydopamine, no granular vesicles as large as p-granules (ef. Unsicker, 1973e), negative results with the Falek-Hillarp fluorescence method).
However, the diagnosis "cholinergie" can only be made, when presynaptic sections and synapses are observed where the typical features of a cholinergie fiber can more easily be recognized.
A positive reaction to acetyleholinesterase (ache) may be regarded as a hint for the cholinergic nature of an axon. Nerve trunks lying outside, inside or beneath the capsule as well as nerve fibers which are in close relation to adrenal ehromaffin cell cords show in all bird species studied an ache-positive reaction, which is characteristic, too, for the same structures in mammals (Koelle, 1950, 1951; Er/tnk6, 1959). When ache is demonstrated with the electron microscope ache- negative axons can be observed only exceptionally (el. Lewis and Shute, 1969; adrenal medulla of the rat). This result may give a hint to the possible existence of non-eholinergie fibers in the avian adrenal gland (see Unsieker, 1973 c). However, it must be admit ted tha t an ache-positive axon need not be a cholinergie one. Often non-eholinergic nerves are ache negative, for example adrenergie nerves to vessels of pancreas and spleen (Esterhuizen et al., 1968; Graham et al., 1968; Lever et al., 1970), but adrenergic and other non-cholinergic fibers can also show positive reactions to ache (Graf, 1967; Hajos et al., 1970; Gwyn and Flumerfeld, 1971). These results demonstrate tha t ache is certainly an essential, but not highly specific component of cholinergie nerves.
The pa th of an axon beginning with the entrance to the adrenal gland and ending at the synapse formed with an A- or NA-cell, can be divided up into several sections, with certMn topographical relations to the capsule, to vessels, and interrenal- and adrenal ehromaffin cell cords. In section 1 (peri- and intra- capsular nerves) large bundles with myelinated axons dominate. As in the rat (Coupland, 1965) we must suppose tha t in the avian adrenal gland reat numbers of preganglionic axons are non-myelinated. The perineurium separating endo- neural and interstitial space has been described by Shantaveerappa and Bourne (1962), Kerjasehki and Stockinger (1970) and Klemm (1970) as a diffusion barrier. Therefore, an influence of nervous elements on surrounding structures cannot be expected until the perinenral sheath has disappeared.
In the course of section 2 nerve fibers come into relation to chromaffin cell cords. They are either situated inside the cord or at its periphery. Then, small groups of axons leave the large bundle and form section 3, comparable to a stalk, before they establish presynaptic sections and synapses.
This mode of innervation cannot be imagined as a fixed scheme; rather there are variations between different species and even within the adrenal gland of a single animal which may contribute to a better or worse understanding of the

432 K. Unsieker
principle. This depends on the fact, for instance, whether the stalk is long or not and whether synapses are distributed Mong the whole length of the stalk or whether they are situated at the end of the stalk.
The mode of innervation described seems to be realized most distinctly in passeriform birds, but even there more distinctly in the Domestic Sparrow and Fringilla than in Corvidae. These birds show clearly a central arrangement of synaptie endings within the ehromaffin cell bale, and a polarization of the cells (of. Unsicker, 1973a). The connexion between central innervation and the occur- rence of folhcles filled with a colloid like substance is worth mentioning. Both phenomena can be studied particularly well in Domestic sparrow and Fringilla. The ehromaffin part of the adrenal gland of these spezies reminds one of the hamster adrenal medulla, where follicles (Graumann, 1956 ; Franzen, 1964 ; Unsicker, 1969 ; A1-Lami, 1970 ; Benedeczky and Smith, 1972) and polarized chromaffin cells occur as well. Whether this mode of innervation and the formation of colloid are causally connected to each other remains uncertain. The discovery of Bennet (1941) that ehromaffin cells in the adrenM medulla of the cat, where follicles do not occur, are columnar and polarized, and my own investigations concerning the formation of colloid in the adrenal medulla of the pig (Unsicker, in preparation) do not speak in favour of such a concept.
Sometimes, even the light microscopical demonstration of ache gives a good impression of the finer distribution of nerve fibers. So, for instance, in Corvidae, which often show a central nerve trunk inside a cord of cells from which fine meshworks of fibers resembling bells arise.
One of the variations in the innervation of ehromaffin cell groups has to do with the behaviour of the basal laminae. In the course of section 2 axons and Sehwann cells are surrounded by a basal lamina of their own. Chromaffin cells and related Schwann cells possess a basal lamina of their own as well. When axons approach ehromaffin cells it becomes necessary that the two basal laminae fuse. This occurs somewhere in the course of section 3 (nerve stalk). Thus, the chromaffin cell complex and the nerve fibers, by which it is supplied, can be regarded as a unit with a single basal lamina stocking. Investigations by Marley and Prout (1965, 1968) show that such a morphological unit is a functional unit as well: different nerve bundles which enter the adrenal gland supply different complexes of chromaffin cells. There is a quantitative relation between the amount of catecholamines secreted and the number of nerve fibers excited. An axon need not be covered by a "somatic" Sehwann cell until it forms a synaptic ending; often there is an extensive contact between axon and plasmMemma of a ehromaffin cell already before the preterminM sections can be recognized.
The pattern of innervation of avian adrenal chromaffin cells resembles the mammalian situation in that in both cases a group of ehromaffin cells is innervated by one nerve bundle. This pattern finds its embryological explanation in the mode of immigration and in the first arrangement of sympathopheoehromoblasts in the future adrenal gland. During this process bales of 10-12 chromaffin cells are formed which fuse later to become the definite adrenal medulla (Wiesel, 1901, 1902; t tett , 1925).
Axons which innervate ehromaffin cells exhibit the well-known ultrastructurM features. Neurofilaments and -tubules make up the main part of the axonM inner

Fine Structure and Innervation of the Avian Adrenal Gland 433
Fig. 10. Cholinergie innervation of adrenal chromaffin cells in birds. The path of an axon beginning with the entrance to the adrenal gland (E) and ending at the synapse (5) formed with an adrenaline (A)- or noradrenaline (NA)-storing cell, can be divided up into several sections, with certain topographical relations to the capsule, to vessels (K) and interrenal (I) and adrenal chromaffin (d) cell cords. In order to give a clear demonstration of the pattern of innervation Schwann cells surrounding the outer surface of nerve bundles and chromaffin cell groups were marked as a continuous black layer. Other satellite cells, basal laminal and perineural sheath were left out. I peri- and intracapsular nerves. 2 nerve fibers between groups of chromaffin cells and a capillary. 3 "nerve stalk" of a chromalfin cell group. ~ preterminal section of nerve fibers. 5 synaptic endings on chromaffin cells. X fusion of peripheral and somatic satellite cells. (Ka) small axons without an individual satellite sheath in the periphery of a chromaffin cell group which possibly represent afferent endings (of. Unsicker, 1973c).
(axi) axons for the inncrvation of interrenal cells (cf. Unsicker, 1973e)
s~ructures. Their ultrastrueture and biochemistry has been studied repeatedly (Silver and McKinstry, 1967; Schmitt, 1968; Frisch, 1969; Olmsted et al., 1970; a.o.) and shall not be discussed here. I t should be mentioned, however, that filaments can be a continuation of part of the tubule wall; this may be taken as another proof of the fragility of tubules (ef. Schlaepfer, 1971). A ring-like arrange-

434 K. Unsicker
ment of neurotubules around mitochondria has already been described by Raine et al. (1971) in the central and peripheral nervous system. The authors suppose that neurotubules take par t in the transport of mitochondria.
The fine structure of preterminal sections and synaptie endings essentially resembles tha t of other eholinergic endings (Barton and Causey, 1958; Taxi, 1962, 1964; Elfvin, 1963; Bargmann and Lindner, 1964; Coupland, 1965; Gray and Guillery, 1966; Grillo, 1966; Unsieker, 1967; Kudo, 1971; a.o.), which are characterized by two different populations of vesicles. The smaller ones (diameter 300-500 A) exhibit a homogeneous lucent contents and are called synaptie vesicles because they occur almost regularly near the presynaptie membrane (Robertson, 1956; Birks et al., 1960; Barrnett , 1062; v. Diiring, 1968; Hubbard and Kwan- bunbumpen, 1968). Probably these vesicles do not represent a homogeneous population (Fig. 11). For the first we can assume tha t at least some of them contain acetylcholine (aeh). This assumption can be derived from results obtained from synaptosomes of the CNS (Gray and Whittaker, 1962; De Robertis et al., 1963; Whit taker et al., 1963; Whittaker, 1965. 1966) and from the investigations of De Robertis and Vaz Ferreira (1957) on the adrenal medulla and those of Jones and Kwanbunbumpen (1968) and Hubbard and Kwanbumbumpen (1968) on the rat diaphragm, which show that there is a relation between the number of synaptic vesicles and the frequency of excitation of a eholinergie nerve. Additional to the experiments mentioned above we can show tha t the number of synaptic vesicles leaving the bouton by exoeytosis increases significantly after the application of 6-OHDA. This phenomenon may be taken to be a vain effort to compensate the depleted amounts of NA in adrenergie varieosities by NA of ehromaffin cells. Another portion of the so-called synaptie vesicles may be involved in miero- pinocytosis processes, which can be observed all along the plasmalemma of the bouton. We have demonstrated that the dense core of large granulated vesicles may leave the bouton or the preterminal section of the axon by exocytosis (see below). There may be a mechanism which brings the surplus membrane material back into the bouton since the surface can not enlarge infinitely. In neuroseeretory axons small vesicles resembling synaptie vesicles can be observed, which, according to Holmes and Knowles (1960), Bunt (1969), Norman (1969), and Smith (1971), are involved in the mechanism of membrane retrieval. Probably a similar mecha- nism accounts for the membrane retrieval in eholinergic terminals (A.D. Smith, 1971) and gives rises to a certain number of "synaptie vesicles", which are free from ach.
Synaptic vesicles are flattened or elliptoid in shape as a rule. Such vesicle profiles have been described for inhibitory synapses (Uehizono, 1965; Bodian, 1966; Carramendi et al., 1967; Lemkey-Johnston and Carramendi, 1968). However, their occurrence in synapses of adrenal chromaffin cells indicates that this shape of synaptic vesicles depends more on the fixation than on the type of synapse.
The opinions concerning the origin of aeh-eontaining synaptie vesicles are controversial as yet. Authors who assume a formation within the cell body claim that after ligation of any type of axon vesicles accumulate above the ligature (Breemen et al., 1958; Dahlstr6m and H~ggendal, 1966; Kapeller and Mayor, 1967; Geffen and Rush, 1968; Zelena and Gutman, 1968; Pellegrino de Iraldi and De Robertis, 1968; Banks et al., 1969).

Fine Structure and InnervatioI1 of the Avian Adrenal Gland 435
If synaptic vesicles are formed within the axon, they could have budded off from neurotubules (De Robertis and Bennett, 1955; De Robertis, 1964; Pellegrino de Iraldi and De Robertis, 1968), from the "axoplasmic retieulum" (Palay, 1956, 1968; Andres, 1964; v. D/iring, 1968), or from mitochondria (Dyatchkowa et al., 1962). We often observed vesicle aggregations within axons which could come from the perikarya, but we also found vesicles which bud off from ncurotubules in the presynaptie region. Biochemical studies on the aeh-synthesis (Marehbanks, 1968, 1969; Whittaker, 1968; Guth, 1969; Igitchie and Goldberg, 1970) and careful analysis of the vesicle membrane (Whittaker and Sheridan, 1965; De Carlo, 1967; Eiehberg et al., 1964; Hosei, 1965; Whittaker, 1968) do not permit a definite conclusion on the vesicle origin either.
Large dense-cored vesicles which are regularly found in eholinergic neurons (Katz, 1961; Taxi, 1961, 1962; Grillo and Palay, 1962; Elfvin, 1963; Pick, 1963; Yamamoto, 1963; Bargmann and Lindner, 1964; Richardson, 1964; Uchizono, 1964; Coupland, 1965; I-I6kfelt and Nilsson, 1965; Grillo, 1966) are another population of vesicles typical of synapses on adrenal chromaffin cells. They can be distinguished from large granular vesicles in adrenergie neurons by the round even profile of the dense core and the constant width of the light halo lying between dense core and unit membrane.
Dense-cored vesicles may leave the terminal sections of the axon by exocytosis, but their origin is as unclear as that of synaptie vesicles. Perhaps the existence of a membrane system in the bouton, which, because of its dilated cisternae and the occurrence of coated vesicles, resembles a Golgi apparatus, may give a hint, where at least part of the dense-cored vesicles come from (Fig. 11). Contradictory to this mode of origin is perhaps the fact that after a cervical sympathectomy boutons containing dense-cored vesicles cannot be observed any longer in the superior cervical ganglion (Grillo and Palay, 1962 ; Grillo, 1966). As to the contents of these vesicles in eholinergic axons, no answer can be given as yet. The extrusion by cxocytosis probably speaks in favour of a protein. Liillmann-Raueh (1971) has argued that dense-cored vesicles in motor endplates contain trophic factors, because they can be seen to increase in number during ontogenesis and regenera- tion (Ter~vginen, 1968 ; Kelly and Zacks, 1970 ; Lfillmann-Rauch, 1971 ; Lentz, 1967).
Invaginations of coated vesicles on the surface of the bouton can regularly be observed in peripheral autonomic and motor endings (el. Grillo, 1966 ; v. D/iring, 1967; Nickel et al., 1967; A.D. Smith, 1971). In general, it can be assumed that they incorporate protein (Roth and Porter, 1962; Kanaseki and Kadota, 1969), in some eases perhaps lipid as well (Palay and Karlin, 1959). I t is not probable that they are involved in the ach-synthesis (Andres, 1964; Andres and v. D/iring, 1966). Certainly, a carrier system is necessary for the uptake of choline because of the hydrophilie character of the molecule. However, molecules of this diameter probably need not be transported by coated vesicles.
Organelles which were regularly found in synaptic endings on ehromaffin cells are mitochondria and lysosome-like bodies. In mitochondria disintegration of the matrix need not necessarily be caused by fixation; it could rather be an expression of the large amount of energy required in synapses (cf. Gray and Guillery, 1966). Lysosomes are probably involved in the elimination of surplus vesicle material (Holtzman, 1971; A.D. Smith, 1971).

436 K. Unsicker
...-.---.. m 0 •
n 13,-i
Fig. 11. Synaptic ending of a cholinergie axcn (ax) on an adrenal chromaffin cell (A). Synaptic (sv) and dense-cored (dv) vesicles leave the axon by exocytosis. (Hypothetical) origin of synap- tic vesicles: 1. cluster of vesicles in the axon (ha). 2. vesicles which bud cff from neurotubules (bt). 3. vesicles, which resemble micropinocytosis vesicles (bp). (Hypothetical) origin of dense- cored vesicles: 1. dense-cored vesicles from the perikaryon (da). 2. formation of dense-cored vesicles in a Golgi apparatus-like structure (g[) in the bouton. Coated vesicles are apparently transported to this synaptic equivalent of a Golgi apparatus, where they can fuse with dilated cisterrtae containing an electron-dense material Pre- and pestsynaptic membranes (pro).
Intersynaptic lamella and filaments (i])
Pre- and pos t synap t i c membrane th iekenings follow the classical model as descr ibed b y Gray and Gui l lery (1966), for instance. F i l amen t s and eanalieuli , which br idge the cleft, and the in t e r synap t i c lamel la have often been ~ subjec t of controversy . Pfenninger (1971) showed t h a t the two opposi te opinions - f i laments on the one hand (De Rober t i s et al., 1961; Van der Lees, 1963; De Rober t i s , 1967), i n t e r synap t i e lamel la on the o ther hand (Pease, 1966; W e s t r u m and Lund , 1966; Bloom and Agha jan ian , 1968; Ake r t st al., 1969; Meyer, 1969; Pfenninger et al., 1969) - are no t con t r ad i c to ry b u t supp lemen ta ry .

Fine Structure and Irmervation of the Avian Adrenal Gland 437
The a t t a chmen t of synapses to chromaffin cells seems to be less uni form in
birds t h a n in mammals . P robab ly this can be explained by the more dis t inct format ion of dendri t ic processes and the occurrence of short spines on chromaffin
cells. As to differences in the inne rva t ion of A- and NA-cells (Grynszpan-Winograd,
1969) final conclusions cannot be made as yet because the n u m b e r of synapses on which the different parameters have been measured is too small. Nevertheless, the differences in the var iab i l i ty of the diameter of dense-cored vesicles in A- and NA-synapses are worth ment ioning.
References
Akert, K., Moor, H., Pfenninger, K., Sandri, C.: Contributions of new impregnation methods and freeze etching to the problems of synaptic fine structure. Progr. Brain ges. 31. 223- 240 (1969)
A1-Lami, F.: Follicular arrangements in hamster adrenomedullary cells: light and electron microscopic study. Anat. Rec. 168, 161-177 (1970)
Andres, K.H.: Mikropinozytose im Zentralnervensystem. Z. Zellforsch. 64, 63-73 (1964) Andres, K.H., Diiring, M. v.: Mikropinozytose in motorischen Endplatten. Naturwissen-
schaften 53, 6115-616 (1966) Banks, P., Mangnall, D., Mayor, D. : The redistribution of eytochrome oxidase, noradrenaline
and adenosine triphosphate in adrenergic nerves constricted at two points. J. Physioh (Lond.) 200, 745-762 (1969)
Bargmann, W., Lindner, E.: Uber den Feinbau des Nebennierenmarkes des Igels (Erinaceus europaeus L.). Z. Zellforseh. 64, 868-912 (1964)
Barmett, R.J.: The fine structural localization of aeetyleholinesterase at the myoneural junction. J. Cell Biol. 12, 247-262 (1962)
Barton, A.A., Causey, G.: Electron microscopic study of the superior cervical ganglion. J. Anat. (Lond.) 92, 399-407 (1958)
Benedeezky, I., Smith, A.D.: Ultrastructural studies on the adrenal medulla of golden hams- ter: origin and fate of secretory granules. Z. Zellforsch. 124, 367 386 (1972)
Bennett, H.W.: Cytological manifestations of the secretion in the adrenal medulla of the cat. Amer. J. Anat. 69, 333-382 (1941)
Birks, R.I., Huxley, A.E., Katz, B.: The fine structure of the neuromuscular junction of the frog. J. Physiol. (Loud.) 150, 134-144 (1960)
Bloom, F.E., Aghajanian, G.K.: Fine structural and eytochemieal analysis of the staining of synaptie junctions with phosphotungstie acid. J. Ultrastruet. l~es. 22, 361-376 (1968)
Bodian, D.: Electron microscopy: Two major synaptie types on spinal motoneurones. Science 151, 1093-11094 (1966)
Borer, I., O'Shaughnessy, L.: L'innervation de la glande surr6nMe. Bull. Ass. Anat. (Nancy) 31, 77-78 (1936)
Brauer, A.: A topographical and cytological study of the sympathetic nervous components of the suprarenal of the chick embryo. J. Morph. 53, 277-325 (1932)
Breemen, L. van, Anderson, E., Reger, J.R.: An attempt to determine the origin of synaptie vesicles. Exp. Cell. l%es., Suppl. 5, 1153-167 (1958)
Bunt, A.H.: Formation of coated and "synaptic" vesicles within neuroseeretory axon ter- minals of the crustacean sinus gland. J. Ultrastruct. Res. 28, 411-421 (1969)
Coupland, R.E.: Electron microscopic observations on the structure of the rat adrenal medulla. II. Normal innervation. J. Anat. (Lond.) 99, 255-272 (1965)
Cummings, J.F.: Thoraeolumbar preganglionic neurons and adrenal irmervation in the dog. Acta anat. (Basel) 73, 27-37 (1969)
Dahlstr5m, A., tt~ggendal, J.: Studies on the transport and life span of amine storage granules in a peripheral adrenergic system. Aeta physiol, scand. 67, 278-288 (1966)
De Robertis, E.: Histophysiology of synapses and neurosecretion. Oxford: Pergamon Press 1964

438 K. Unsicker
De l~obertis, E.: Ultrastructure and cytochemistry of the synaptic region. Science 156, 907-914 (1967)
De l~obertis, E., Bennett, H.S.: Some features of the submicroscopic morphology of synapses in frog and earthworm. J. biophys, biochem. Cytol. 1, 47-58 (1955)
De Robertis, E., De Iraldi, A.P., De Lores Arnaiz, G.R., Salganicoff, L.: Electron microscope observations on nerve endings isolated from rat brain. Anat. Rec. 189, 220-221 (1961)
De l~obertis, E., De Lores Arnaiz, G.R., Salganicoff, L., De Iraldi, A.P., Zieher, L.~.: Iso- lation of synaptic vesicles and structural organization of the acetylcholine system with brain nerve endings. J. Neurochem. 10, 225-235 (1963)
De Robertis, E., Ferreira, V.A.: Submicroscopic changes of the nerve endings in the adrenal medulla after stimulation of the splanchnic nerve. J. biophys, biochem. Cytol. 3, 611-614 (1957)
Diamond, I., Kennedy, E.P.: Carrier-mediated transport of choline into synaptie nerve endings. J. biol. Chem. 224, 3258-3263 (1969)
DiCarlo, V.: Ultrastructure of the membrane of synaptic vesicles. Iqature (Lend.) 213, 833-835 (1967)
Diiring, M. v.: Uber die Feinstruktur der motorischen Endplatte yon h6heren Wirbeltieren. Z. Zellforsch. 81, 74-90 (1967)
Dyatehkowa, L.N., Dawidowa, T.W., Yakobson, N. R.: The role of mitochondria in forming synaptic vesicles [Russisch]. Dokl. Akad./qauk. SSSI% 147, 1467-1469 (1962)
Eichberg, J., Whittaker, V.P., Dawson, R.M.C.: Distribution of lipids in subcellular particles of guinea pig brain. Biochem. J. 92, 91-100 (1964)
Elfvin, L.-G.: The ultrastructure of the superior cervical ganglion of the cat. IL The structure of the preganglionic end fibers and the synapses as studied by serial sections. J. Ultra- struet. Res. 8, 441476 (1963)
Elliott, T.R.: The innervation of the adrenal glands. J. Physiol. (Lend.) 46, 285-290 (1913) ErEnkS, 0.: Specific demonstration of acetylcholinesterase and nonspecific cholinesterase in
the adrenal gland of the rat. Histochemie 1, 257-267 (1959) Esterhuizen, A.C., Spriggs, T.L.B., Lever, I.D.: Nature of islet cell innervation in the eat
pancreas. Diabetes 17, 33-36 (1968) Feldberg, W., !V[inz, B.: Des Auftreten eines azetylcho]inartigen Stoffes im Nebennieren-
venenblut bei Reizung der Nervi splanchnici. Pflfigers Arch. ges. Physiol. 283, 657-682 (1933)
Feldberg, W., ~V[inz, B., Tsudzimura, H.: The mechanism of the nervous discharge of adrena- line. J. Physiol. (Lend.) 81, 286-304 (1934)
Fischer, H.: Vergleichende Pharmakologie yon Ubertr~gersubstanzen in tiersystematischer Darstellung. Handbnch der exp. Pharmakologic, Bd. XXVI. Berlin-Heidelberg-New York: Springer 1971
Franzen, D.: Beitr~ge znr Merphelogie und Chemohistologie des Nebennierenmarks des Goldhamsters (Mesocricetus auratus). Anat. Anz. 115, 35 58 (1964)
Frisch, D.: A photegraphic reinforcement analysis of neurotubules and cytoplasmic mem- branes. J. Ultrastruct. l~es. 29, 357-372 (1969)
Geffen, L.B., Rush, I~.A.: Transport of noradrenaline in sympathetic nerves and the effect of nerve impulses on its contribution to transmitter stores. J. Neurochem. 15, 925-931 (1968)
Gershon, ]V[.D., Drakontides, A.B., Ross, L.L.: Serotonin: Synthesis and release from my- enteric plexus of the mouse intestine. Science 149, 197-199 (1965)
Gershon, M.D., Ross, L.L.: Badioisotopic studies of the binding, exchange, and distribution of 5-hydroxytryptamine synthesized from its radioactive precursor. J. Physiol. (Load.) 186, 451-476 (1966)
Giacomini, E.: Sur les terminaisons nerveuses duns le capsules surr6nales des oiseanx. Arch. ital. Biol. 29, 482-483 (1898)
Goodman, L.S., Gilman, A.: The pharmacological basis of therapeutics; fourth ed. New York: Macmillan Co. 1970
Gordon, M.K., Benisch, K.G., Deanin, G.G., Gordon, M.W.: Histochemical and biochemical study of synaptic lysosomes. :Nature (Lend.) 217, 523-527 (1968)

Fine Structure and Innervation of the Avian Adrenal Gland 439
Graf, J.: Elektronenmikroskopiseher Nachweis der Acetylcholinesterase in den postganglio- ngren sympathisehen Nerven des Iterzens. J. Neuroehem. 14, 893-897 (1967)
Graham, J.D.P., Lever, J.D., Spriggs, T.L.B.: An examination of adrenergic axons around pancreatic arterioles of the cat for the presence of aeetylcholinesterase by high resolution autoradiographic and histochemical methods. Brit. J. Pharmaeol. 33, 15-20 (1968)
Graumann, W.: Bildung und Sekretion perjodatreaktiver Stoffe im Nebennierenmark des Goldhamsters. Z. Anat. Entwickl.-Gesch. 119, 415430 (1956)
Gray, E. G.: Electron microscopy of presynaptic organelles of the spinal cord. J. Anat. (Lend.) 97, 101-106 (1963)
Gray, E.G., Guillery, R.W.: Synaptic morphology in the normal and degenerating nervous system. Int. gev. Cytol. 19, 111-182 (1966)
Gray, E.G., Whittaker, V.P.: The isolation of nerve-endings from brain; an electron micro- seopy study of cell fragments derived by homogenization and eentrifugation. J. Anat. (Lend.) 96, 79-88 (1962)
Grille, M.A.: Electron microscopy of sympathetic tissues. Pharmaeol. Rev. 18, 387-399 (1966) Grille, M.A., Palay, S.L.: Granule-containing vesicles in the autonomic nervous system.
V. Intern. Congr. Electron-Microscopy, Philadelphia, vol. 2, U. p. New York: Academic Press 1962
Grynszpan-Winograd, O.: Diffgrences dans l'innervation des (~cellules £ adr6naline~) et des <~cellules £ noradr4naline~) de la m6dullo-surr6nale du Hamster. C. R. Acad. Sei. (Paris) D 268, 1420-1422 (1969)
Guth, P.S.: Acetyleholine binding by isolated synaptic vesicles in vitro. Nature (Lend.) 224, 384-385 (1969)
Gwyn, D.G., Flumerfelt, B.A.: Aeetyleholinesterase in noncholinergic neurons: a histo- chemical study of dorsal root ganglion cells in the rat. Brain l~es. 34, 193-198 (1971)
Hajos, F., Priymak, E.K., Kerpel-Fronius, S.: The electron microscopic demonstration of acetylcholinesterase activity in some cholinergie and non-cholinergic synapses of the rat brain. Acts histochem. (Jena) 35, 114-122 (1970)
Heft, g.: Ein Beitrag zur Histogenese der mensehlichen Nebenniere. Z. mikr.-anat. Forsch. 3, 179-282 (1925)
Hillarp, N.-A.: Innervation of the adrenal medulla in the rat. Aeta anat. (Basel) 3, 153-161 (1947)
H6kfelt, T., Nilsson, O.: Electron microscopy of the adrenergic and eholinergic innervation of the iris muscle. J. Ultrastruet. l~es. 12, 237 (1965)
Hollinshead, W.H.: The innervation of the adrenal glands. J. comp. Neurol. 64, 449-467 (1936)
I-Iollinshead, W.H., Finkelstein, H.: I~egeneration of nerves to the adrenal gland. J. comp. Neuro]. 67, 215-220 (1937)
Holnms, R.L., Knowles, F.G.W.: "Synaptic vesicles" in the neurohypophysis. Nature (Lend.) 185, 710 711 (1960)
Holtzman, E.: Cytochemical studies of protein transport in the nervous system. Phil. Trans. B 261, 407 (1971)
Hosei, R.J.A.: The localization of adenosine triphosphatases in morphologically characterized subcellular fractions of guinea-pig brain. Biochem. J. 96, 404-412 (1965)
Hubbard, J.I.: Mechanism of transmitter release. Progr. Biophys. 21, 33-124 (1970) Hubbard, J.I., Kwanbunbumpen, S.: Evidence for the vesicle hypothesis. J. Physiol. (Lend.)
194, 407-421 (1968) Jones, S.F., Kwanbunbumpen, S.: On the role of synaptic vesicles in transmitter release.
Life Sci. 7, 1251-1255 (1968) Kanaseki, T., Kadota, K.: The "vesicle in a basket". A morphological study of the coated
vesicle isolated from the nerve endings of the guinea pig brain, with special reference to the mechanism of membrane movements. J. Cell Biol. 42, 202-220 (1969)
Kapeller, K., Mayor, D.: The aceumulation of noradrenaline in constricted sympathetic nerves as studied by fluorescence and electron microscopy. Prec. roy. See. B 167, 282-292 (1967)
Katz, B.: The terminations of the afferent nerve fibre on the muscle spindle of the frog. Phil. Trans. B 2411, 221-240 (1961)

440 K. Unsicker
Kelly, A.M., Zacks, S.I.: The fine structure of motor endplate morphogenesis. J. Cell Biol. 42, 154-169 (1970)
Kerjaschki, D., Stockinger, L.: Zur Struktur und Funktion des Perineuriums. Die Endigungs- weise des Perineuriums vegetativer Nerven. Z. Ze]lforsch. 110, 386400 (1970)
Kiss, T.: Experimentell-morphologische Analyse der Nebenniereninnervation. Acta anat. (Basel) 13, 81-89 (1951)
Klemm, K.: Das Perineurium als Diffusionsbarriere gegeniiber Peroxidase bei epi- und endo- neuraler Applikation. Z. Zellforsch. 108, 431445 (1970)
Knouff, I~.A., Hartman, F.A.: A microscopic study of the adrenal of the brown pelican. Anat. l~ee. 109, 161-187 (1951)
Koelle, G.B.: The histoehemical differentiation of types of cholinesterases and their locali- zations in tissues at the eat. J. Pharmacol. exp. Thor. 100, 158-179 (1950)
Koelle, G. B.: The elimination of diffusion artifacts in the histoehemical localization of cholin- esterases and a survey of their cellular distributions. J. Pharmacol. exp. Thor. 103, 153- 171 (1951)
Kudo, S.: Fine structure of autonomic ganglion in the chicken pancreas. Arch. histol, jap. 32, 455497 (1971)
Kura, N.: (Jber die Innervation der Nebenniere [Japanese]. Mitt. ivied. Akad. Kyotv 1, 107-124 (1927)
Larramendi, L.M.H., Fickenscher, L., Lemkey-Johnston, N.: Synaptic vesicles of inhibitory and exhibitory terminals in the cerebellum. Science 156, 967-969 (1967)
Lemkey-Johnston, N., Larramendi, L.M.H.: Type and distribution of synapses upon basket and stellate cells of the mouse cerebellum. An electron microscope study. J. comp. Neurol. 134, 73-112 (1968)
Lentz, T.L.: Fine structure of nerves in the regeneration limb of the newt Triturus. Amer. J. Anat. 121, 647-670 (1967)
Lever, J.D., Spriggs, T.L.B., Graham, J.D.P., Irons, C.: The distribution of 3H-noradrenaline and aeetylcholinesterase (ACHE) proximal to constrictions of hypogastric and splenic nerves in the eat. J. Anat. (Lend.) 107, 407-418 (1970)
Lewis, P.R., Shute, C.C.D.: An electron-microscopic study of eholinesterase distribution in the rat adrenal medulla. J. Microscopy 89, 181-193 (1969)
Lobko, P.I.: Sources of sympathetic preganglionic fibres of splanchnic nerves and ganglia of splanehnic plexus. Arkh. Anat. 56, 52-54 (1969)
Lees, H. van der: Fine structure of synapses in the cerebral cortex. Z. Zellforsch. 60, 815-825 (1963)
Liillmann-Rauch, 1~.: The regeneration of neuromuscular junctions during spontaneous re- innervation of the cat diaphragm. Z. Zellforsch. 121, 593 603 (1971)
MacFarland, W.E., Davenport, It.E.: Adrenal innervation. J. comp. Neurol. 75, 219-233 (1941)
Marchbanks, I~.M.: Exchangeability of radioactive acetylcholine with the bound acetyl- choline of synaptosomes and synaptic vesicles. Bioehem. J. 106, 87-95 (1968)
Marchbanks, 1%. M.: Biochemical organization of cholinergic nerve terminals in the cerebral cortex. In: Cellular dynamics of the neuron (ed. S.tt. Barondes). New York: Academic Press 1969
Marley, E., Prout, G.J.: Physiology and pharmacology of the splanchnic-adrenal medullary junction. J. Physiol. (Lend.) 180, 483 513 (1965)
Marley, E., Prout, G.J.: Innervation ef the cats adrenal medulla. J. Anat. (Lend.) 102, 257-273 (1968)
Maycock, W.A., Heslop, T.S.: An experimental investigation of the nerve supply of the adrenal medulla of the cat. J. Anat. (Lend.) 73, 551-558 (1939)
Meyer, W. J.: Phosphotungstic acid section staining of synaptic junctions of rat brain. J. Cell Biol. 43, 929 (1969)
Nickel, E., Vogel, A., Wasser, P. G.: Coated vesicles in der Umgebung yon neuro-muskul/iren Synapsen. Z. Zellforsch. 78, 261-266 (1967)
Norman, T.C.: Experimentally induced exocytosis of neuroseeretory granules. Exp. Cell Res. 55, 285-287 (1969)

Fine Structure and Innervation of the Avian Adrenal Gland 441
Olmsted, J.B., Carlson, K., Klebe, g., t~uddle, F., Rosenbaum, J.: Isolation of microtubule protein from cultured mouse neuroblastoma cells. Proc. nat. Acad. Sci. (Wash.) 65, 129- 136 (1970)
Palay, S.L.: Synapses in the central nervous system. J. biophys, biochem. Cytol. 2, Suppl. 2, 193-201 (1956)
Palay, S.L.: The morphology of synapses in the central nervous system. Exp. Cell Res., Suppl. 5, 275-293 (1958)
Palay, S.L., Karlin, L.J.: An electron microscopic study of the intestinal villus: II. The path- way of fact absorption. J. biophys, biochem. Cytol. 5, 373-383 (1959)
Pearse, A.G.E.: Histochemistry. Theoretical and applied. London: I. + A. Churchill. Ltd. 1961
Pellegrino de Iraldi, A., Robertis, E. de: The neurotubular system of the axon and the origin of granulated and nongranulated vesicles in regenerating nerves. Z. Zellforsch. 87, 330 344 (1968)
Pettit, A.: Sur les capsules surr~nales et la circulation porte surr4nale des oiseaux. Bull. Mus. d'hist, nat. Paris 2, 87-88 (1896a)
Pettit, A.: Recherches sur les capsules surr~nales. Th~se de Paris (S4r. A, Nr. 254, Nr. d'ordre 886) (1896b)
Pfenninger, K.I-L: The cytochemistry of synaptic densities. II. Proteinaceous components and mechanism of synaptic connectivity. J. Ultrastruct. Res. 35, 451475 (1971)
Pferminger, K., Sandri, C., Akert, K.: :Neue Darstellung yon Membranen im :Nervensystem. Acta anat. (Basel) 711, 305 (1969a)
Pfenninger, K., Sandri, C., Akert, K., Eugster, C.H.: Contribution to the problem of struc- tural organization of the presynaptie area. Brain l~es. 12, 10-18 (1969b)
Pick, J.: The submicroscopic organization of the sympathetic ganglion in the frog (Rana pipiens). J. eomp. Neurol. 120, 409~1~62 (1963)
Raine, C.S., Ghetti, B., Shelanski, M.L.: On the association between microtubules and mito- chondria within axons. Brain Res. 34, 389-393 (1971)
Richardson, K.C.: Fine structure of the albino rabbit iris with special reference to the identi- fication of adrenergic and cholinergic nerves and nerve endings in its intrinsic muscles. Amer. Anat. 114, 173-205 (1964)
Ritchie, A.K., Goldberg. A.M.: Vesicular and synaptoplasmic synthesis of acetylcholine. Science 169, 489-490 (1970)
Robertson, J.D.: The ultrastructure of a reptilian myoneural junction. J. biophys, biochem. Cytol. 2, 381-394 (1956)
Roth, T.F., Porter, K.R.: Specialized sites on the cell surface for protein uptake. In: V. Int. Congr. for Electron Microscopy (ed. SS. Breese, jr.) 2: LL 4. New York: Academic Press 1962
Schlaepfer, W.W.: Experimental alterations of neurofilaments and neurotubules by calcium and other ions. Exp. Cell Res. 67, 75-80 (1971)
Schmitt, F.O.: Fibrous proteins--neuronal organelles. Proc. nat. Acad. Sci. (Wash.) 60, 1092-1101 (1968)
Shanthaveerappa, T.R., Bourne, G.tt.: The "perineural epithelium", a metabolic active, continuous, protoplasmic celI barrier surrounding peripheral nerve fasciculi. J. Anat. (Loud.) 96, 527-537 (1962)
Shioda, T., Nishida, S.: The innervation of the adrenal cortex. Arch. histol, jap. 28, 23-43 (1967)
Silver, M.D., McKinstry, 1. E.: Morphology of microtubules in rabbit platelets. Z. Zellforsch. 81, 12 17 (1967)
Smith, A.D.: Summing up: some implications of the neuron as a secreting cell. Phil. Trans. B 261, 423-437 (1971)
Smith, U.: Uptake of ferritin into neurosecretory terminals. Phil. Trans. B 261, 391 (1971) StShr, Ph., jr.: Mikroskopische Anatomic des vegetativen Nervensystems IV/5. In: ttandbuch
der mikroskopischen Anatomie des Menschen. W. Bargmann Hrsg. Berlin-G6ttingen- Heidelberg: Springer 1957
Swinyard, C.A.: The innervation of the suprarenal glands. Anat. Rec. 68, 417-429 (1937)

442 K. Unsicker
Taxi, J.: Etude de l'ultrastructure des zones synaptiques duns les ganglions sympathiques de la Grenouille. C. R. Avcad. Sci. (Paris) D 252, 174-176 (1961)
Taxi, J.: Etude au microscope 61ectronique de synapses ganglionaires chez quelques Vert6br6s. Prec. IV. Int. Congr. Neuropathol., Mfinchen 1961, ed. H. Jacobs, vol. 2, p. 197-203. Stuttgart: Thieme 1962
Taxi, J.: Etude de certaines synapses interneuronales du systbme nerveux autonome. Acta neuroveg. (Wien) 26, 360-372 (1964)
Ter~Lv/~inen, H.: Development of the myoneural junction in the rat. Z. Zellforseh. 87, 249-265 (1968)
Tucek, S.: On the question of the localization of choline aeetyltransferase in synaptie vesicles. J. Neuroehem. 13, 1329-1332 (1966)
Uchizono, K.: On different types of synaptic vesicles in sympathetic ganglia of amphibia. Jap. J. Physiol. 14, 210-219 (1964)
Uehizono, K.: Characteristics of excitatory and inhibitory synapses in the central nervous system of the cat. Nature (Lend.) 207, 642-643 (1965)
Unsicker, K.: Uber die Ganglienzellen im Nebennierenmark des Goldhamsters (Mesocricetus auratus). Ein Beitrag zur Frage der peripheren Neurosekretion. Z. Zellforsch. 76, 187-219 (1967)
Unsicker, K.: Follikel aus Ganglienzellen im Nebennierenmark des Goldhamsters (Meso- cricetus auratus). Z. Zellforsch. 95, 86-101 (1969)
Unsicker, K.: Fine structure and innervation of the avian adrenal gland. I. Fine structure of adrenal chromaffin cells and ganglion cells. Z. Zellforsch. 145, 389-416 (1973a)
Unsicker, K.: Fine structure and innervation of the avian adrenal gland. III. Non-cholinergie nerve fibers. Z. Zellforsch. (in press) (1973c)
Westrum, L.E., Lund, R.D.: Formalin perfusion for correlative light- and electron-micro- scopical studies of the nervous system. J. Cell Sci. 1, 229-238 (1966)
Whittaker, V.P.: The application of subeellular ffactionation techniques to the study of brain function. Progr. Biophys. melee. Biol. 15, 39-96 (1965)
Whittaker, V.P.: Some properties of synaptic membranes isolated from the central nervous system. Ann. N.Y. Acad. Sci. 137, 982-998 (1966)
Whittaker, V.P.: Synaptic transmission. Prec. nat. Acad. Sci. (Wash.) 60, 1081-1091 (1968) Whittaker, V.P., Michaelson, I.A., Kirkland, R.J.A.: The separation of synaptic vesicles
from nerve ending particles ("synaptosomes'). Biochem. J. 90, 293-303 (1963) Whittaker, V.P., Sheridan, M.N.: The morphology and acetylcholine content of isolated
cerebral cortical synaptic vesicles. J. Neurochem. 12, 363-372 (1965) Wiesel, I.: Uber die Entwicklung der Nebenniere des Schweines, besonders der Marksubstanz.
Anat. H. 16, H. 50, 115-150 (1901) Wiesel, L: Beitr~ge zur Anatomie und Entwieklung der menschlichen Nebenniere. Anat. It.
19, It. 63, 481-522 (1902) Yamamoto, T.: Some observations on the fine structure of the sympathetic ganglion of
bullfrog. J. Cell Biol. 16, 159-170 (1963) Young, J.Z.: Partial degeneration of the nerve supply of the adrenal. A study in autonomic
innervation. J. Anat. (Lend.) 7~, 540-550 (1939) Zelena, J., Gutman, E.: Accumulation of organelles in central and peripheral stumps of
interrupted neurons. In: lYlacromolecules and the function of the neuron, p. 156-166. New York: John Wiley & Sons 1968
Priv.-Doz. Dr. K. Unsieker Anatomisehes Institut Universit~t Kiel D-2300 Kiel Federal Republic of Germany

![Adrenal Imaging - University of Floridaxray.ufl.edu/files/2010/02/Adrenal-Imaging.pdfadrenal glands [3], and a metastasis might ... CT, adrenal imaging, adrenal lymphoma imaging, adrenal](https://img.pdfslide.net/doc/110x75/5b26814c7f8b9a8c0f8b4820/adrenal-imaging-university-of-glands-3-and-a-metastasis-might-ct-adrenal.jpg)












![Muscle Innervation Chart II[1]](https://img.pdfslide.net/doc/110x75/55241db64a7959da488b45f0/muscle-innervation-chart-ii1.jpg)