Embed Size (px)
Citation preview

Charles B. Halpern,Shelley A. Evans, Cara R. Nelson, Don McKenzie, and DeniseA. Liguori, D vls on of EcosysternScience Co ege oi Forest Resoufces Box352100 Un versity ol Wash ngton, Seatl le Washngton9B1952100
David E. Hibbs and Melora G. Halal, Departrnenl of Forest Scence, Oregon State Univefsty Corva s, Oregon97331 5705
Response of Forest Vegetation toVarying Levels and Patterns ofGreen-tree Retention: An Overview of a Long-term Experiment
AbstractTimbcr harvest wiih fetentid oflive ( grccn') trees. snags, and krg\ is now r shndard praclice on fedeml lllan ix" hnds $ ithirthe rangc of lhe northern spotted owl. Although specill c guide I ines have been adoplcd for thc leve I s and spat ial conllgu fations olretained structures. neithef the ecologic.rl :rssumptions ihat undeflie these recommendations no. lhe outcomes of these pfactices
hrlc bccn rigorously tested. The Demonnralion of Ecosy\ren Management Oplion\ (DEM() stud,v exrmines the responses oflbrcsr ccos_r'srems in the Paciiic Norrh$cs! to varying levels (pefcentage oi basal arca) alrd patterns (dispefsed vefsus rsgresttlcd)ol green-tree ferenrion. ln rhis papcr $edescribe vegetalion studies that lorm the ibundatioo ofthis l(xlg-term. iDlerdi\ciplinar)cxperiment. \\re fe!ielr the rcsulls ofretrospectile anallses and simulation models lr'hich suggesl that even mirinal lc\cl! ofrclc tion m ay have inpo|lant ellccB on ibrest de!e]opment. \!'e descibc thc characleri siics of ihe DEIUO sites. lhe erpcrimcnl aldcsign. ard the pfincifal lrlrirbles of inlcrcs! (stand structufe. tree regeneralion and growth. and undeNtor) conposilion rnddi!ersir,"-). \!c speculate rbout the silviculrural and ecological fesponses of lores6 to \aryjng levels and paftems of retention andtbcus ir panicula. on the d! namics of the ibrcsl undcrnory. \\'e anticiprte strolg c|llrtra s|s anr ong lreannents in $e e \tabl i\hnentof e|rly sclal. opcn site sfecies. aDd in the pcrsislcnce of shxde-toleranl plants associalcd $it| older forests or fore\t-interiofeD!ironmL'nts. Short-rerm fesponses arc likcl) to be driven by \'.rfiation iD the distribulion and intensit! of harvest distufbance.Longcr lcfln rends afe erpecled lo rcllcct the elltcrs of contfasting paltem\ of canop,v retention. Wt conclude b) discussins\omc ofdre scientific challenges iaccd in designing rnd inplenertins largc scalc. interdisciplinar), experiments.
lntroduction
Traditionally. management of fcderal forests inwestern Oregon and Washington has fbcused onthc production. utilization. and regenemtion oftimber resourccs. Until fairlv recently (late 1980s),clearcut logging and broadcast buming werc stan-dard practices. Clearcut units were dispersed inspace and timc, logging slash was burned to re-duce fire hazard and/or competilion from non-crop species, and trccs rvere planted with unifirrmsprc ing a t h igh den. i t ie . . l t h r r . bccorne inerc r . -ingly apparent, ho$'ever, that these harvest prac-tices do not produce the same complexity of pat-tern as arises throu-qh natural disturbance-acomplexity that is thought to underlie thc recoYery of forest ecosystems tir l lowing large scaledisturbanca (e.9.. Franklin et al. 1995. Perry andAmaranthus 1997). For examplc, historic wild-lires in the wcstem Cascade Range \\'ere cpisodic,varying in size, tiequency. and iDtensity $'ith el-evation, topography, and local climate (Tecnsma1987. Morrison and Srvanson l990,Agee 1993.).Even where natural tires wcre catastrophic. indi-vidual trees or -troups of trees often survived and
large amounts of $,oody debris remained as logsand snags. It has also become apparcnt that thestructural and conpositional attribules thirt der elop lhrnuph ., 'nr cnri, 'nal nrJnapcrncnt pri lctice\difl'er markcdly liom those shaped b)' naturalsuccessional processes (Spies et al. 1988. Hansenet al. 1991. Spies and Franklin 1991. Halpem andSpies 1995, Tappeiner et al. 1997).
Federal forest management policics and prlc-tices have changed signil icantly in the last decade in response to glowing concems that tradi-tional approaches have led to widespread loss andfragnentation of old-growth ecosystcnls rnd 1oattendant declines in biological diversit) (FEMAI1993. Thomas et al. l993,Tuchmanncral. 1996).A new framewortri is emerging, characterized bya shift in ernphasis f'r1)m nranagement of a singlecommodity resourcc to a broader considerationof ecosystem conpoDents and tunctions. L(x)sclydetlned as 'ecosystem nranagemcnl.'- this lpproachseeks to incorporate ecological knowledge intoforest nanagement to sustain biological diversity. as well as tinrber production (Kessler et al.l992. Grurnbine 199:1, Christensen et al. 1996).
Northwest Science, Vol. 73, Special Issue. 1999
i l 9 9 L r b l n r c \ o n h \ e n S c r e , \ l i . i ! h n , . \ . r 1 . d
21

Consequently. f()rest managers have bcgun toimplement silvicultural prescriptions that are in-tended to more closely mimic the plocesses andoutcoltlcs of natur_al disturbance and successt0nby retaining stluctural characteristics ofthe fomcrtbrest-in particul
harvest, the species composition of a dispersedretention unit (27 trees/ha) rcscmbled that of anadjacelt c]earcut to a greater degree than that ofan adjacent, intact forest (reflecting abundant es-tablishment of early successionai species). However. shade-tolerant forest species wcre more di-verse and abundant in the dispersed unit than inthe clcarcut, suggesting that fbrest understory taxamay benefit from the increased shading and/orreduced disturbance aftbrded by retained trees.Untbrtunately, these short term, unrcplicatedobservations affbrd only a limited view of thepossible ellects of green tree retention.
Studies ofunderstory responses to thinning invounger tbrests may otler additional insights. Ithas been found that thinning generally leads togreater plant species diversitl' (in large pan dueto the establishment ofinvasive or exotic species)and to increased plant cover (Witler l975.Alabackand Herman 1988. Bailey 1996). although thenature of these responses may vary with the veg-etation, environment. or intensity ofthinning. Forexample, rvhere thinning leads to increased dominance by clona) shrubs (e.g.. Tappeiner et al. 1991 .Huffman et al. 1994). thcrc nray be a resultantdccline in species diversity (Alaback and Hennan1988). There has been litt lc consideration of theeff'ects ofthinning on plant species that arc ass(Fciated with late-seral environments o[ that aresensitive to logging disLurbance (but see BaiJeytree6l).
lnsights into the potential long-tefm consequences of green-tree retentioD for understoryconnunitics may be aided by retrospective analyses ofnarural. two tiered (two-agcd) stands. Theseyoung-to-mature torests support an emergcntcanopy ofolder, scattered residual trees and havebeen described as the natural analogues of fbr-esls thal may dcvelop fi) l lowing retention harvest(e.g., Rose and Muir l996, Peck and McCune 1997,Zenner et al. 1998). Traut (199.1) described difterences in understory species composition in plots\\ ith and without natwal, residual trees arrd at-tribuled thesc differences. in part, to the effectso l re r idua l t ree \ on lhe den. i t ) and eonrpos i t i , ' nofthe rcgcnerating tree layer Unfbfunately. causalmechanisms cannot be deduced through retrospcc-tive analysis, nor is it possible to distinguish di-rect effects (e.g., shading by residual trecs) fromindirect ones (e.g., species interactions or localvariation in disturbance history. init ial conposition. or other site features). Replicated, prc{reat-
ment measurements in the DEMO study u'ill provide a basis for separating the potential effects ofretained trees tiom initial species composition andother sitelevel vadation.
Considerably more atlention has been devotedto thc effects of rcsidual trees on forsst stand de-velopment and tree growth. Forexample, throughanalysis of annual growth rings North et al. (1996)observcd that 1br 6 yr alter dispersed retentionharvest, the basal area increment ofretained Douglas-fn (.Pseudot.sugq melr..iesii') was 157r lowerthan that in an adjacent. uncut forest. This is notthe rcsponse typically obsen'ed in younger thinnedstands (reviews in Banett It980], Bailey [l9961.North et al. u9961); it may ret'lect a 'thinning
shock" in which resources are differentially allo-cated to roots (Oliver and Larson 1990). Longterm [leasurements of tree growth among repli-cate DEMO blocks may suggest whether theseare local orgeneral phenomena, and whethcr theyrepresent short-term or more persistent efTects.
Retrospective studies of natural, two tieredstands also provide insights into the long-terrnefftcts of dispersed retention on forest develop-ment and tree growth. Regional comparisons of70- to 1lo-yr-old stands with and without older(>200 yr) residual trees suggestthat above athrcsh-old density (ca. l5 trees/ha), residuals ctrn aft'ectthe composition of younger cohorts and reducetheir basal area growth (Rose and Muir 1996).Negative eflects on basal area growth have alsobeen repor ted l rom pr i r (d -p lo r . lud ie . in ue \ r -ern Oregon. Here, however, the strongest effectper unit basal area of residual trces occurred atthe lowest levels ofresidual tree density (5 trees/ha: Zenner et al. 1998)orbasal area(5-10m']/ha;Acker et al. l99li).
S tund te rpon.e . to Brccr - l ree re renL ion mavalso include increased mortality due to windthrowof isolated residual trees. Removal of 30 607c ofthe initial volume of rnature and old growth for-ests in nofihwestem British Colunrbia resultedin rates of windthrow of0.3 to 8.27c of trces within2 yr after harvest (Coates l997). However, nor-tality can vary narkedly from site te site. rcflecting variation in the lcvel ofretention, initial standdensity and composition. stand age, local weatherconditions, and landfbrrn. For example, among4.1 green-tree retention sites in western Oregon,rates of windthrow-related mortalityranged ftom0 to nearly 607o (mean of 16%). with the greatestIosses observed soon atier harvest (Adler u99z1l
Vegetation Response to Green-trcc Retention 29

and Hunter I 1995I cited in Franklin et al. | 1997l).By comparison, annual ratgs of mortalit) ' inunmanaged mature and old growth folests ofrvestem Oregon and Washington average <17c(Franklin et al. 1987). The randomized blockJc . rpn , ' i th . D t \4O . rud) u i l l pernr i r sepur . r -tioD of treatnenl effects (level or pattcrn of rc-tentioD) on mofiality from effects associated witl'lsitc-spccifi c t 'rctors (e.9.. bpography. soils).
Forest gap and stand-levcl growth and yicldmodels have also been used to explore the pos-sible effects of varying levels ofdispersed reten-tion on fbrest stand development and wood production. Some ofthe predictions ofthese modelsare intuitive (indeed some are predetemlined bygrowth equations, McKenzie 199,1). For example.the degree to \\,hich models simulatc thc dcvcl-opment of old-grorvth stand characteristics (e.g.,tree size class structure) improves with the nurr-ber of trees retained through harvest (Hansen etl l . l Q Q 5 r . r n J I c l r t l \ . 1 , \ j l e r r . ' u l l r r g g i n g . r . u n r u -lative basal alea and volune growth are reducedwhen live trees are retuined (Birch and Johlson1992. Hansen et al. 1995). Other simulation re-sults, although less intuit ive. are supported byr-effospective analyses of natural stands. In parlicular. rcductions in stand basal area grou,th perresidual tree are predicted to be strongest at thelowest levels of retention (5 trees/ha: Hansen etal. 1995). Sti l l olhsr simulation results are quitesurprisilg and seen to coutradict empirical ob-servations. Rrr cxamplc. under low levels ofdis-persed retention (as feu'as 5 trees/ha) and givensufficient t ime (240 yr). naturally establishing.shade-tolerant species such as westem hemlockl.Tsuga heterophtlLa) and western redcedar (Tftfnplicatal can oulcompetc plantcd Douglas-fir(Hansen et al. 1995). To what extent the spatialconfigLuation ofresidual |rccs may influcncc standdevelopment has not been examined empiricallyor through simulation. despite the fact that ag-gregated retention is now a staldard practice onfcdcral matrix lands in the region (USDA and USDT199.1). Conparisons of aggregated and dispcrscdretention in the DEMO design explicit ly test forsuch effects.
Methods
Study Areas
To establish a broad geographic and ccolo-qicalbase ofinterence. eight study blocks rvere selected
to represent a diversity of forest types in westemOregon (Umpqua N. F) and Washington (GiffordPinchot N. F. and Washington DNR) (see Aubryet al. 1999). Blocks encompass markedly differ-ent physical environnents, stand ages, and foreststructures (Table l). Sites were chosen 1() mini-mize the variation in environment and vegetationamong treatment units within each block, withinthe constraints imposed by current landscape con-figurations and past management activities (Aubryct al. 1999). To varying degrees this goal wasachieved (but see a discussion of this issue in theChallenges section. belor').
Across blocks, elevations range ti-om ca. 2001700 m. slopes vrry trom gentle to steep, and nearlyrl l aspects are represented (Table l). Most siteslie $'ithin the westeln hemlock (fsir.qrihetetopltllla) forest zone (Franklin and Dyrness1973). Two blocks, LittleWhite Salnon and DogPrairie are located in the more eastrly grand t'ir(Abies grandis) and whitc t\r (Alties concoLor)zones. respcctively. and a third, Paradise Hil ls,occupies the colder, $etter, silver f ir (ADiesornab i lis) ane. P s e uclot s uga menie.!il doninatesall sites but associated tree species vary fiom blockto block (Table 1).
Forest stfuctwe varies anong blocks. reflect-ing diffelences in stand age (c.g.. Litt le River vs.Layng Creek). regeneration and disturbance his-tory, and physical eDvironment (Table 1). Forestunderstory composition also valies: sone blockshave depauperatc ground layers (e.g., Dog Plairie) while others suppo dense herb and shrubcommunities (e.g.. Layng Creek. Litt le WhiteSalmon). Despite this heterogeneity, many blocksshare the same dominant taxa (e.g., vinc maplelAcer circinatuml, Oregongrape lBerberi.taervosaf. salal lGaultheria .r/rdfurnl. and huckleberty fVat: cittiu r n spp. I ; Table 1.1. Ditlerencesin stand sffucture and composition among treat-ment units within each block are generally smallcompared to difterences among blocks (Table 1;but see Challenges section and Table ,l).
511.5 h,s had varying histoies of rccent dis-turbance (Aubry et al. 1999). Forexample, LayngCreek and Capitol Forest are second-rotation stands(i.e., clearcut but regenenled naturally) rnd alltbur blocks onthe UmpquaN. F. have bcen thinnedor salvage logged. In contrast, thc three blockson the Gifibrd Pinchot N. F. supporl mature, un-disturbed forest.
-10 Hrlpern et rl.

: : = : !5 - : i . - gi = l i :: . i ; a
: = > : :: = * i ! -i , : i = j= : " : < |: :S s = s :=
--L
i > _ : ' j a
i : + . . i ' i: a - , e . :
= d " i :t ; s - i - '; * . : 3 . .; . i ! :
t . ? : =E ! : s :: i z 2 ?: : ; ! : :; : : . i y .; : i . 6 ) -
< ; : : ; s :i . : = a : < 1 , ,:'^ = : : : : ,; i ; - i i :r : - ! f r =' . 0 : i - , : y '? . : i a + 9 !: l i < i i i ,- I { , : t ; :
: ( , : : : : :: . s : =
< : = : . i PL L I i - . i > e
< - € " J : + a. a ! : ! - . > n
; 1 . o \ : - i
: : a : E . . ;i ! i i " : : ;
5 i i : ; i ' 4l , : : i - : . i l: 5 s i : i ' :; > - . i f - | =
' ' i i : i i i j€ : - - : , ; : ; =5 7 - t : i - = = : =- - ) . . ! r : Z : a; / i " S i : =i i i , : : : : !f t s ! i d i = = - : A. - F : < . - -
_ : .- - i - 1 = < t t i= _ , q z a a = i t^ _ ^ e - ^ L ^
r + - i ; r i b : x ;J + 3 J E : : ^ i I
t : :! ^ : = z - ^ ia l i ! = ; i K n 1 [= : - \ - + - l == ' r - : ; . E f , ; j = l =
i ! _ F
T = E +
: = . 7 : i i = a i' ' = i Q - ' . ! = 2 n t r- + Z : 4 1 = 2 2 = E z! : = 7 2 4 = . - i 4 +c ' - " a - ' " t J
i ; :
3 lVegetation Response to Green-trce Retention
a.
' a
: 5
: ;8 , ,
a
.2
a
2
i
. 9

Experimental and Samp ng Designs
Aubry et al. (1999) describe the DEMO experimental design and harvcst prcscriptions rn an rn-troductory paper in this volume. To rcview. ateach of the eight study blocks (replicates). sixharvest treatments have been randomly assignedto l3-ha experimental (treatment) units. Trcat-ments dift-er in the level (percentage ofbasal area)and/or spatial pattern (dispersed vs. aggregated)ofretained trees as follows: (l) 100% rctention(control): (2) 75%, retention (haryested as threecircular. 1-ha gaps-hencetbfth the "gap" trcat-ment); (3) 407. dispersed retention: (4) 40% ag-gregated retention (as five. cilcular l-ha reten-tion patches); (5) 15'la dispersed retention; and(6) 157. aggregated retention (as two, circular Iha retention patches) (see Figure 1). By design,these treatments produce distinctly different pat-terns of vadation in stand structure and environ-ment: relativcly homogeneous conditions in thecontrol and dispersed retention treatments, andt\\,o contrasting conditions (intact tbrest and har-vested area) in thc aggregated retention and "gap"
treatments. We empl0y tvo sanrpling designs to. : rp tu rc lh ( con l rJ . l \ and gr ld ien t . in rcce t r t ion
rcsponse that ffe expected to arise fiom these treatnlenls.
Sampling Design for Treatment Effects
The first sampling design, which perrnits analy-sis of treatment eft 'ects (e.g., Hypotheses l-3,below). uses a set of permanent vegetation plotsanayed across a grid system at each site (63 or64 gdd points $,ith 40-m spacing, Figure I ). Thenumber and spatial distributions of sample plotsvary by treatnent, however. ln the dispersed re-tention and control treatments, where stand conditions wil l be relatively homogenous,32 plotsare placed systematically at altemate grid points(Figure la). In the aggregated retention and gaptreahnents, where we expect two distinct post-har-vest environments. plots are placed at all gridpoints within the aggregates (or gaps), and at asubset ofpoints in the surrounding matrix (yielding 32 or 37 plots per treatment, Figure 1b).
Strong contrasts in harvest treatments will prc-duce marked d i l fe rence: in lhe cornpo: i r ion . \ i /estructure, and spatial distribution ofoverstory trees.We arc pafiicularly interested in the elfects ofthesctrcatments on volume and basal area growth: stem
a. Disoersed retention treatment b. Aggregated retention treaknent
Figure L Plot locations (open circle\) lbf srmplimg (a) relati\el) homogeneous conditions iD the dispersed feteDtion tfeatment\and (b) contr.rsting conditi(ms in the aggregated retention treatnents. Control units are sanpled as in (a)i "gap' (757.
retentior) units as in (b). Spatial grrdients \i illin aggregated retentiL'n units are sanpled using ca. 100 n long traDsects(d.rk iires ir b. see also figurc l).
32 Halpem et al.

TABI-E 2. Variablcsofi ei: n and derived statistics fbr the two pfnnaD sampling affroaches (see Ni ethods and Figurc 2 ). X =
variable sampled;blank = variablc no! samplcd.
Treatnent-level Within-treatmenreiiecb gradienrs Deri\'ed statistics
Physical Sitc Charact€risticsElclation. aspecl. slopeTopogr.rphic position, slope coDfiguratxnrGround lurfacc condidon\ (c.g.. mineral
soi l . s lone. l r t ter , log)Coare woody debris
Post-harvest disturbance(e.g.. vegetarion damage, soil disturbance)
Ovefitory VcgetationCanop) co!erTree dia clcrs (>5 cm dbh)
Trcc characteristics (e.9., carop) class, vigor.cro\'" n ralio. damagc)
'liee mortality: phvsical conditions and
Tree heighrs: rotal. canop! depth
Snags: diameGr. height. decay class
Understory Veg€tationLichcnsi prcscnce/ilbsence
Bryophytcs: prcscnce/absence
Herbsi co\ er. hcighl
Tall shrubs: cover. height
Undcrslor) fccsSeedl ings (<10 cm ta l l ) : no. ncmsSapl ings (>10 cm t . l . <5 cn dbh):
coler, no. slens, height class, ofigilr(nalural !s. plunred)
XXX
XXX
XX
rolunre and dcnsity (b,,- species and decay
XX
XXX
XX
densit). basal area. \,olurre. and diame|erdistributions (by species)
fates and causes of nrortalit! (b)'. spccics)
heighi and canopy depth distributions (by
density (b,v specics. djamercr. deca). aDdheight c1a\s)
specles liequencv and divcrsil)species frequenc! and di!cr\illspecies frequenc!. diversiry. and hcighl
d is l f ibut idr \species liequenc). dilersity. and height
distribulions
fiequency xnd density (b,v species)ircqucncy. densill. size structure, and ratesofnoriaLrt) (b) species and size cl.rs\)
"Conduclcd in the.10% uggregated retentiur treatments at Bufte and Pafadise Hills blocks only.
density: diameter, height. and canopy-depth distributions of canopy and sub-canopy trees: ratesand causes of morlality; and recruitment and fatesof snags (Table 2). A series of nested circularplots (Figure 2a) will be used to sample theseoverstory rcsponses at 3 5 yr inteNals. Pefma-nently tagged trees will be measuled for diam-eter, canopy position, structuml attributes, andmoftality. Tree height and crown depth will bemeasurcd tirr a subset of sterns. Snags wil l bemeasured for diametet height, decay class, andangle of lean.
As with the overstory, the forest undersk)ry isl ikely to respond to strong contrasts in harvest-
rclated disturbance (and resulting canopy condi-tions) among treatments. We are paticularly in-terestcd in treatment e1l'ects on plant species dir'er-ritl r richne:s anJ er enne\. r: c!,mmunit) compo:itionand structure (the proponional representation ofdifferent plant groups or 'functional' types); andthe establishment, growth, and nrertality of regenerating trees (Table 2). We will also consider un-derstory attributes that are known to influence thcabundance and distribution of u'ildlile (e.g., plantcover and vefiical stratification. recruitrnent anddyramics of corusc woody debris) (Table 2). Aseries of belttrmsects, intcrcept iines, and nicroplotsnested within each tree plot will be used to sanple
Vegetation Response to Creen-tree Retention 33

Fi:ure:. Plol r.d rrrnscct designs fof sampling rfe.Ltment efiects (a, b) and spatial gradicnl\ (c, d) $llhin aggregated retentionunils. For lreannent efi_ects. olefstory chltracterisrics (a) arc mcasurcd witli. nested circular plots: 0.01 ha for trees5.0 11.9 cm dbh (T,) .0.0.1 ha lbr t rees >15.0 cm dbh ( l ) . and 0.08 ha for snags (S). UndeNbry \ ,egel . r r iorr (b) issampled r \ i rh: bel t r f rnsects (B) lbr dcnsi ly o l \apl ings(t rees>10cJn ta l l .<5 cmdbh)r intercept l ines (L) fbr covethr'ighl oftall shrubs and r.rplings. dislurbance asscssmcnts. and volume ofcoarse woody debri\l and lllicr'oflds ($. 0.1x 0.5 nr) ibr g.ound su| face ch.mctefistics, presence ofbryophycs and macrolichens. coler and heighl olherbs xnd lollshrubs.anddcnsir )ofreeseedl ing, t (< l0cmtal l ) . OverstLi r ) canop," 'co!er is cninarcd $i th a sphcr icaldensiomclcr a!rhe end poinls (c) ol cich inrercept Iine. For spatial gfadients in uggregaled rercndon |rcarlncnls (c). undcrnory \ cgctat ion is srmplcd wrrh l3 b inds per t ransect . With in each brnd (d) t t re: subplots (sp. 1 r I n) ior colcr ,41cjghl o l a l lspecies and dcn\i!icr of tree seedlings/saplingsr Dricfopbts (lrp, 0.2 r 0.5 nl) lor prcsencc/abscncc of brtophlt!'st andcnd poi.rs (c) lor o\'erslor] crnopy cover. See Table 2 ibr a compl.lc lin ol \ariablcs.
lhe responses of undcrstory species. changes inground surt'acc characteristics, and the types andlcvcls of hawest related disturbance (see Figurc2 caption tbr details). The ncstcd plot design willalso pcnnil direct conelation of overstory andunderstory chilracteristics.
Sampling Design for Spatial GradientsW i th i n Ag g reg ate d B ete n ti o n Tre atme nts
A sccond design wil l be used to sample fine-scalegradients in undersk)rv rcsponse within the ag
grcgated retention treatments (e.9., Hypotheses4-7. below). This approach is l inited to the,10%retention treatments at Buttc and Paradisc Hil ls.Here our tbcus is on the extent to which the I -haaggregates rgtain conrponents of the original un-derstory and/or amelioratc microclimatic condi-tions in thc surrounding matrix. Bands of salrpleplots arc arrayed along four o hogonal transectsofca. 100 rn in lcngth that span harvested. ecotonal(edge). and intact tbrest envir()nments (Figurc 2c).Th< peryend icu lu r p laLetnent r ' l - l r rn .e , ts le rn r i t 'examination of the effects of aspect on spatial
3,1 Halpern et irl.

gradients in vegetation. Bands are spaced moreclosely (every 5 m) near the edges (ca. fiIst 15-25 m) ofthe 1-ha aggregates where we anticipatesharyer gadients in vegetation response, and morcwidely (every l0 m) in the harvested matrix andforest interior (Figure 2c). A series of sub-plotsand microplots within each band (Figure 2d) willbe used to sample changes in understory compo-sition and structure (Table 2; see Figure 2 cap-tion for details).
Hypotheses, Mechanisms, andAnalytical Approaches
It is not possible in this paper to describe the fullrange of expected ecological or silvicultural re-sponses to varying levels or patterns of green-tree retention. However. to stimulate future dis-cussion and to guide ongoing inquiry and analysis,we have summarized our expectations in a tableof predicted, qualitative responscs to the six experimental treatments (Table 3). These predic-tions draw upon the results of ernpirical studiesof natural disturbance and succcssion. plant environment relationships, plant I ite history strate-gies, and vegetation rcsponses to more conven-tional methods ofsilviculture (i.e.. clea-rcut loggillg.commercial thinning). Others are more specula-tive and rcflect general theory that is untested inthese forest ecosystems.
In the lbllowing sections, \\,e explore a set ofpredictions in depth as examples of the types ofquestions that we have posed in this study. Wefocus on the response ofthe forestunderstory whichcontains the virst majority olplant spccies in thesetbrests. and in which we anticipate short terrnchanges to be nost dynamic. Our hypothesesconsider three groups of taxa for which we an-ticipate distinct pattems of response. The lirstare "fbrest-interior" specics: subordinate foresthcrbs that are not restdcted to. but are fbund nrostliequently and with greatest abundance in, forest-interior and late-successional environments(e.g., coral rcotlCordllorhizq spp.l, prince s pine
lChim a p hikt utn b e l I q tal. rattlcsnake-plantainlGootlyera obkrtgifo lial, twayblade [lstera spp.].pyro\a fPtrola pictal; Spics 1991. Halpern andSpies lo05r . l \4 rn ) o l thc .e herh \ i r re \en . i l i reto disturbances associated with logging and broad-cast burning (R5 species of Halpern u9891; seealso Halpern et al. [1992]) and are thought to rc-qu i re l , ' ne pcr rL 'd . r r f t ime fo r popu la t ion . to re
cover fbllowing large-scale disturbance (Halpernand Spies 1995). The second group consists of"early-sera1" or "open-site" species. These in-clude annual, biennial, and perennial herbs (na-tive and exotic) that typically dominate recentlydisturbed and etuly successional communities (Il14 species of Halpem [1989]; see also Dyrness[9731, Schoonmaker and McKee [l988], Halpemand Franklin [1990]). These taxa are characterizedby long-distance seed dispersal, rapid growthrates, and high fecundities. The third group ofinterest, "forest dominants," are woody and her-baceous species that dominate mature and old-growth forest understodes. These taxa are toler-ant of disturbance (R3 species of Halpern | 19891;e.g., Acer circinatum, Berberis nervosa, Gaultheria shallon, and Polystichum munitum) awl canoccupy a broad range of canopy conditions andseral stages (Spies 1991).
Hypothes zed Responses AmongTreatments
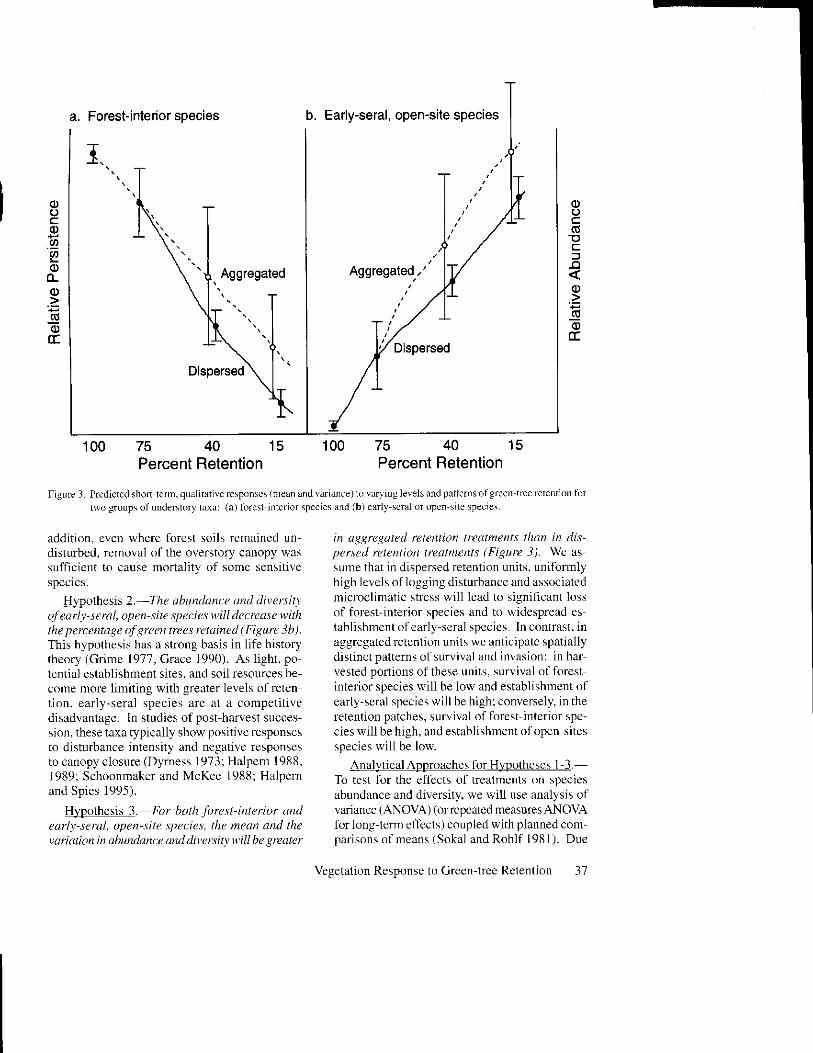
An underlying premise of green-tree retention isthat the suNival of organisms associated withtbrest-interier environments will increase with thepropofiion of live trees retained through harvest(Franklin et al. 1997). Conversely, invtrsion olearly-successional or open-site species shoulddecline rvith increased levels ofretention. To date.these assumptions have not been tested empiri-cally. nor have $e fomrs of these relationshipsbeen afi iculated (e.g., as l inear or non-linear func-l ions . o r as th re .ho ld re .ponse. ) . Ther '< i s e rengreater unceft ainty regarding possiblc diffcrcnccsin responsc within aggregated and dispersed retention (Franklin et al. 1997). We prcdict thatlhe-e ! ruup. u i11 re .p , rn , l I ' l i ' l l , ' u r :
Hyoothesis 1.-Tlre qbtotdence and dirersit)of forcst interior species will hcrease v,ith thepercentage oJ green trees retained (Figure 3a).This prediction rests on two likely outcomes ofincreasing levels of retention: (l) reduced levelsofdisturbance to soils and unde$tory vegetation.and (2) greater amounts ofshading liom retainedtrees. The results of long{cm, succcssional sludiesfollowing clearcut logging and burning suggestthat both conditions should pronote survival offbrest-interior species (Halpern 1989. Halpern eta1. 1992. Halpem and Spics 1995). In thcsc studics.species extinctions induced by harvest were correlated with the intensitv of soil disturbance. In
Vesetation Response to Green-ffee Retention 35

_ 3 :
c =
3 ;
. : }
! i a
E r a ;
7 9 - E
i 2 + a E == . - 4
9 ; :
t = ' - ' 2 - :- | - . v u a d1 . - 1 1 " ' r = Ei 1 ! 4 F E r. ? - a : 9 . a .= / ' . t f l . t
: 2 z i c ! , :
t - = i i = . a- t i a e E ; ' ,. 2 2 = aa i , .= - d 7 r = , t , , ,i = : j ' ! t :
E ' Z ? = ^ =
. i E = € + . :i 1 7 4 = ! id a i z Z = I' - r 4
2 Y . f " : P - Hi :
1 7 E z ? t z
_ 2
z 9 : E z t a ! t
I " 4- :
" .?t e 9,1+ . E 2 ' - ' u / . t ^ / : - i i t Z
< = E 2. i l E
, >= - - ; 1 ; = - - 2 - . ; . ; . i t = " "' - = = - E - r : . ; ; : a ;
ca - e
t i l -
a i - 2 r i E 4 ,-
, a L ;= = ' ' - - . a. ; - - . : a i : ^= r : i - v : - i 7 ; -- = E y - !- ! ' - . = - 1 t ! = _ _
L " - i = = e " ; , 1 2 i E ; = 2. i L . - E i z t , ! y n , 2 . = - P e =' i > : = : . i > ! 1 . a : . i ; ' ; ij t . ; i . * 2 " . ! = = iF 2 ; i e E 2 t ; i =- 4 i ; i t,4 : a E ; i : = r 2 = = = = z : zF : 2 ) : - : = ! : ; ; t . - LY 2 Y ' , ! - - 2 - = : i - : : l s : !? = = : = = = = = : = - y . ! . = y
? = Z 2 r Y - = = Z = ? Z i : = ii t s t t i - t E 5 P - r & - i I
a
s
a
i
l
IEg-t
:i s
iIIlI
f
Halpem et al.36
=e 1v . a
E i i
- .
P E
i . .
i a =
a t ;
; = :
a z a
ooco.9.
o
c)
so(r
a. Forest-intenor species
I
100 75 40 15Percent Retention
addition, even where forest soils remained un-disturbed, removal of the overstory canopy wassulficient to cause mortality of somc sensitivespecies.
Hypothesis 2.-Ilre abundance dnd diversittof earl r- se ru| open-site qteci e s u,ill dec rease w iththe percentdge of green tees ret.lined (Figure 3b).This hypothesis has a strong basis in life historytheory (Grime 1977. Grace 1990). As light, po-tential establishment sites, and soil resources be-come more limiting with greater levels of reten-tion. early-seral species are at a competit lvedisadvantage. In studies of post haryest succes-sion, these taxa typically show positivc responsesto disturbanca intensity and negative responsesto canopy closure (Dyrness 1973; Halpem 1988.1989; Schoonmaker and McKee 1988; Halpernand Spies 1995).
Hypothesis 3. For both forest-interior dndearl-t-seral, open-site species, the mean and theyaiation in abtndance ttrul diversitt will be greater
c)
(d
c
c)
(5
o)E
100
Figure 3. Prcdicted short-tenn. qualitative responscs (mean and lafiance) to rarying levels and pattcrns ofgreen-tree relenlion fi)l
tlvo groups ofunderslory iaxa: (a) lbren inlcrior species and (b) early-seral or op!n 'ile .pe.:re(.
75 40 15Percent Retention
in dggregdted rete tion treatments tlnn in dis-persed retentio tre1tmefits (Figure 3). \Ne assume that in dispersed retention units. uniformlyhigh levels oflogging disturbance and associatedmicroclimatic stress will lead to signiticant lossof forest-interior species and to widespread establishment of early-seral species. In conlrast. inaggregated retention units we anticipate spatiallydistinct pattens of survival and invasion: in harvested portions of these units, survival of forestinterior species will be low and establishment ofearly-seral species will be high; conversely, in theIetention patches, survival of forestinterior spe-cies will be high, and establishmcnt of open sitesspecies will be low.
Analytical Approaches for Hvootheses I -3.-
lo te.t lor the el'fect. ol trealments , 'n speciesabundance and diversity, we will use analysis ofvariance (ANOVA) (or repeated measures ANOVAtbr long-term effects) coupled with planned com-parisons of means (Sokal and Rohlf I981). Due
Vesetation ResDonse to Green-tree Retention
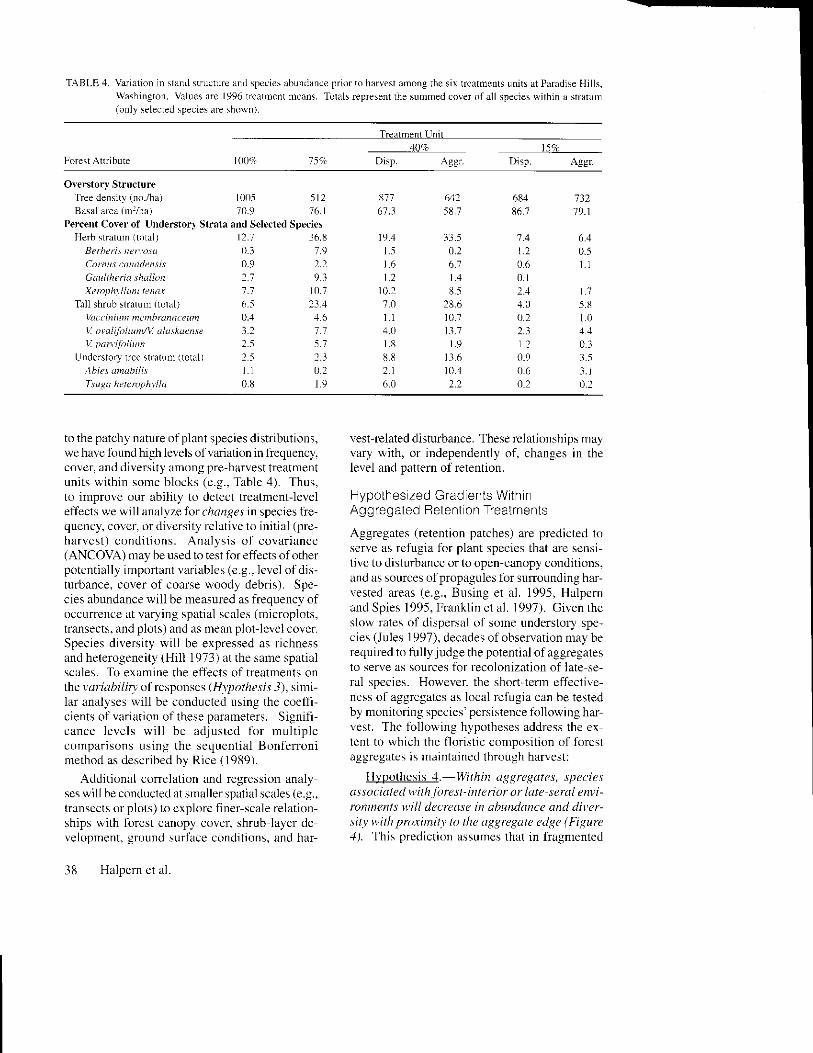
TABLE :1. Varirtion in sland struclure and species abundance pfror to harvest among lhe six lrcatmen|s units at Pafrdise Hills.Washi ngton. Values are | 996 r.car mcnl ncans. Tokls repre\ent the su mmed cover of al I species wilhin a stratu Drr(nrh selected sDecies afe \ho$,n).
Treatment l-rnit
Disp. AgC. Disp.
Percent Coyer of Understor) Strata and Sclccted SpeiesHerb srraturn ltoral) 12.7 16.8
0.3 t.9Cotn l ts tu t l tdLn \ t \ 0 .9
Olerstory Structure'Iiee denslt,v (no.,4ra)Basal area (Drr/hal
Cauhhetia shdllonXenph\llLtL tenar
5 1 216 .1
2 .29 .3
10 .723..1
1 .72 .5 5 .1t.5 l .- lt . ] 4 . 2o.r i 1.9
r00510.9
2.',l1.',l
81',767.1
19..1t . 5
1 . 210 .21.01 . 1
1 . 88 .82 . 16.0
61258.7
33 .50.26.1
u .528.610 .711 .7
t . 9l t . 610..12 .2
6lJ,l86.7
1.41 . 20.6t J . l2.1
o.22 .3t . 20.9
0.2
13279.1
0.5
\ . 15 .81 .0
0.33 .53 . 10.1
Tall shrub slratum (olal) 6.5Uutl innor n(nbruntk?un 0.1\/. onliPliunt/r: uhskutnse :1.2
Undcrstory trcc \lralum (told)
7.\uga h(k'rophrlla
to the patchy nature of plant species distributions,we have found high levels of variation in ftequency.cover, and diversity among pre-harvest treatmentunits within sone blocks (e.g.. Table 4). Thus,to irnprove our ability to detect ffeatmentleveleff'ects we will analvze 1br c/ratges in species tie-quency, cover, or diversity relative to initial (pre-harvest) conditions. Analysis of covarrance(ANCOVA) may be used to test for effects ofotherpotcntially impoftant variables (e.g.. level of dis-turbance, coyer ol coarse woody debris). Spc-cies abundance will be measured as fiequency ofoccurrence rt varying spatial scales (micropiots,transects. and plols) and as mean plotJcvcl cover.Species diversity wil l be expressed as richnessand heterogeneity (Hill 1973) at the salre spatialscales. To examine the effects of treatments onthe r'.r,'i.rlrilil ofresponses (Hrpolhesls 3). simi-Iar analyses *ill be conducted using the coeffi-cients of variation of these parameters. Signifi-cance levels wil l be adjusted for multiplecomparisons using the sequential Bonferronimethod as described by Rice ( 1989)
Additional correlation and regression analyses will be conducted at smaller spatial scales (e.9.,transects or plots) to explorc finer-scale relation-ships with tbrest canopy cover. shrub layer development. ground surtace conditions, and har-
vest-related disturbance. These rclationships mayvary with, or independently of. changes in thelevel and pattem of retention.
Hypothes zed Gradients WithinAgg'egaled Relel l o.
- eaLme' l rs
Aggregates (retention patches) are predicted toserve as refugia fbr plant species that are sensi-tive k) disturbance or to open-canopy conditions,and as sources ofpropagules for sunounding har-vested areas (e.g., Busing et al. 1995, Halpernand Spies 1995, Franklin et al. 1997). Given theslow rates of dispersal of sotne understorJ spe-cies (JuJes 1997), decades ofobservation nray berequired to fullyjudge the potential of aggregalesto serve as sources tbr recolonization of late se-ral species. However, the short-tenn effective-ness of aggregates as local refugia can be testedby monitoring species' persistence following har-vest. Thc tbllowing hypotheses address the extent to which the tloristic composition of forestaggregates is maintained through harvest:
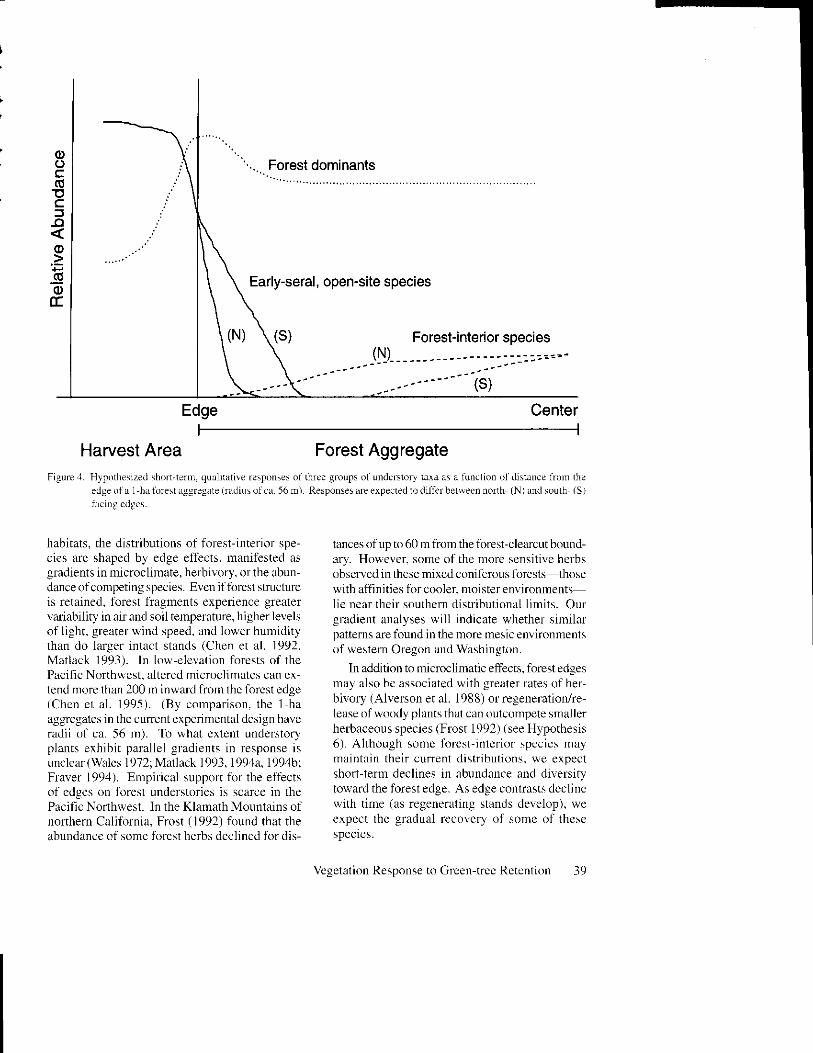
Hvp, t the . i . r t . - lV l th i t uqqegut t , , :1 ' r ' , i , tassociated w,ith lbrest-i terior or lute seral enyi-romnants vill decreuse in abntdance and diver-sil with proxinih to the aggregctte edge (FigLtre4r. This prediction assumes that in ftagmented
38 Halpcrn ct al.
Q'o(U
o
_goE
Forest dominants
Early-seral, open-site species
Forest-interior species
Center
Harvest Area Forest AggregateFigufe'1. H)pothesized sho 'terln, qualitati!e responses of three groups of undcrstor) uxa rs a iunction |)1 disunce lioln the
edge of r I +r ibfe\t rLggregate (radi u s of ca. 56 m). Responses are expecGd to diller berwecn nonh (N ) and sou rh (S )tircing edges.
habitats, the distibutions of fbrest-interior spe-cies arc shapcd by edge efl'ects. manifested asgladients in microclimate, herbivory. or the abun-dance ofcompeting species. Evcn iftbrest sfuctureis retained. forest fragments experience greatervariability in air and soil temperuture, higher levelsof l ight. greater wind speed. and lowcr huniditythan do larger intact stands (Chen et al. 1992.Matlack 1993). [n low-elevation forests of thePacific Northwest. altered nicroclimates can ex-tend more than 200 rn inward flom thc forcst edge(Chcn et al. 1995). (By comparison, the I haaggregates in the current experimental design haveradii of ca. 56 m). To what cxtent understoryp lan ts e rh ib i t p r rc l ie l ! r ' rd ienr . in re .pon.e isunclear(Walcs 1972; Matlack 1993, 1994a, 1994btFraver 1994). Empirical suppert lbr the etlectsof edges on lbrest understories is scarce in thePacific Northrvest. In the Klamath Mountains ofnorthern Calitbrnia, Frost ( 1992) fbund that theabundance t-'f somc fbrcst hcrbs dcclincd fbr dis-
tances ofupto 60 m fiom the tbrest-clearcut beund-ary. However, some of the more sensitive herbsobsen'ed in these nixed coniferous forests thosewith affinities f or cooler. moister environmentslie near their southem distributional l imits. Ourgradient analyses wil l indicate whether similarpfl lcm: are iound in thc morc rnc:i( en\ ironmcnl:of western Oregon and Washington.
ln addition to microclimatic effects, forest edgesmay also bc associatcd with greater rates ef her-bivory (Alvenon et al. 1988) or regeneration/re-lease ofwoody plants that can outcompete smallerherbaceous species (Flost 1992) (see Hypothesis6) . A I lhouBh \omc I i \ rc : l - rn lc r i , ' r .pee ie . rn r lmaintain their current distributions, $e expectshot-tenn declines in abundalce and divelsitytoward the forest edge. As edge contrasK dcclincwith time (as regenerating stands develop). weexpcct the gradual rccovcrl of some of thesespecies.
Vegetation Response to Grccn-trcc Retcntion 39

HJpothesis 5. Eurlr--seral, open-site.\pecieswill declhe in abundance (ud dfiersih with dislatrce into the forest aggregqte; gradients m abLln-dance and diversin )rill be steeper and shorterthan those of lbrestinterior species (Figure 1).It has been observed in other forest ecosystemsthat changes in micrcclimate and local disturbanceregime along the lbrest edge may promote theinvasion of exotic or ruderal taxa (Ranney et al.1981. Laurance 1991, Brothers and Spingam 1992,Frost 1992). The distances to which early-seralspecies invade may be more closely linked to thedepth of edge-related disturbance than to gradi-ents in microclimate, thus we expect relativelyshort, steep gradients in response. As the effectsofdisturbance diminish with time, we expect thesegradients in seral specjes distributions to becomeless apparent.
Hyoothesis 6. The dominantJbre.tt understoryspecies will shovt ltositive responses lo the creation of edges, pettking in abtntlance within ashort distance of the forest qg,grcgate-haryestednntrix bounclar,v ( Figirre 4). Many of our domi-nant understory taxa (e.g., Vaccb'rium spp., Gault-heria shalLon, andAce,..ift ir..rtrrr) possess mor-pho log i . a l o r phys io log ica l t r r r i t . r e .9 . . . ' r t cns i r erhizome systems. variable leaf morphology, and/or potential for rapid vegetative spread) that con-fer resistance to disturbance and enable vigorouserpans ion on(e re \our (c cond i t ion . improre(Alaback and Hennan 1988; Halpern 1988, 1989iHuffman et al. 1994; O'Dea et al. 1995). It islikely that increases in Iight along the boundaiesofnewly created forest aggregates will induce rapidlateral and vefiical growth of these taxa.
Hvpothesis 7. tdSe effects, as tlelinecl by thedistances to v,hit:h earl.\,-seral species penetrateor forest-interior species are lost (and bt thenognitude oftlrcse changes), will be greateru,hereenvironmental conditions (te more stressJul (i.e.,akng south awl west Jacfug edges as opposedto north- qn(l eastfacing edges ) ( Figure 1). Thrsprediction is based on the assumption that envi-ronmental conditions that inhibit fbrest-interiorspecies and enhance cstablishment of open-sitespecies are accentuated along south- and wcst-t-acin g aspects (Wales I 972, Ranney I 977, Matlack1993. Chen et al. 1995, Murcia 1995). In thePacific Northwest. these aspect-related ell-ects maybe magnified by greater rates ofteetall or canopydamage along south- and west-lacing edges dueto the strongly directional nature ofwinter storms.
Anal),tical Approaches for Hl'potheses,+ 7.Analyses of spatial gradients within aggregatedretention treatments will be used to elucidate howspecies abundance (frequency and cover) and diversity vary with distance from the forest-aggre-gate edge (Hypotheses 4-6, Figure,l). Abundanceor diversity measures will be ht to linear or non-linear regression models (Sokal and Rohlf 1981,Zar 1981). For species that exhibit a signiticantcorrelation between abundance and distance fromedge, the form of the most appropriate model (e.g.,linear, logarithmic, or exponential) may provideinsight into the mechanism(s) of decline or ad-vance (e.g., invasion or die-back as isolated indi-viduals orin a wave-tiont; sers, Matlack 1994a).To examine possible aspect-related differences inthese effects (Hypothesis 7, Figure 4), we willtest for the significance of an aspect term in mul-tiple regressions of abundance on distance andaspect. Multiple regression wil l also be used inexploratory fashion to test for relationships withother biotjc anryor abiotic factors that vtlly at finerspatial scales and that may influence, or be influ-enced by, edge-related phenomena (e.9., tree cover.ground surface conditions, local disturbance. orassociatedvegetation). The sequential Bonfenonirnethod will be used to adjust significance levelsfor multiple comparisons (Rice 19891.
The l-ha sized aggregates in the current de-sign are relatively large with respect to the rangeofprescribed patch sizes (0.2 to >l.0 ha.)forgrcen-tree rctention units on federal matrix lands (USDAandUSDI 1994). Howevet they are much smallerthan arc necessary to escape microclimatic effectsassociated with edges (Chen et al. 1995). Althoughour studies do not explicitly test for the effects ofaggregate size, the results of these gradient analysesmay suggest whether smaller aggregates mightseNe as plant rcfugia, or whether edge effects aresuch that larger retention units are necessary tomaintain pxrticular species or groups of species.
The Scientitic Challenges of Large-scale, Interdisciplinary Experiments
Numerous challenges arise in designing and implemenring lffge \cale. inlerdi.ciplinury e\ferimenls.particularly when these studies are motivated byconcerns over natural resource management orpolicy. The DEMO experimental design and itscomponent studies represent the products ofa longand iterative process in which we have incorporated
40 Halpem et al.

diverse ecologicd interests, a rigorous experimentaldesign, and the needs and concems of land man-agers and the public (Abbott et al. 1999, Aubryet al. 1999, Franklin et aJ. 1999). Many of ourdecisions represent tradeoffs: they resolve particular problems but in turn may limit statisticalpower, ecological or silvicultural inf'erence, orharvest elliciency. In this section. we provideseveral examples of the conceptual and method-ological chailenges that have shaped our studies.
Sample area antl sanple size: inherent trudeoffsin intenlisciplinary studie.\. If the spatial scalesof sampl ing in th is experiment were not constrainedby interdisciplinary linkages, vegetation studiescould employ greater replication of smaller-sizedtreatment units, thus cnhancing statistical powerHowever, vegetation measurements are integralto interpreting launal responses (Lehmkuhl et al.1999) which, by necessity, must be exrmined atlarger spatial scales ( l3 ha). As a consequence.our ability to detect teatment ellects tbr somecomponents ofthe fbrest community may be com-promisedby the relativcly small numbers of sampleunits.
Experimental rigor versus site-speciJic man-agement qpproache,s. To minimize the potentialconfounding of post-harvest activities with retention treatments, our silvicultural prescriptionsrequire that site-preparation and tree-plantingspecifications are consistent among all harrest unitswithin a block (Aubry et al. 1999). However, whereerperimenlal unit ' r.r i. upy JiFlerent enr ir, 'nment:.suppoft different plant communities, or generatedifferent densities of slash. this consistency maylead to a prescdption that is less than optimal forrrlr iculturrl objectire'. Ecological interpretation..as well as the applicability of pafiicular resultsfor managers, must be tempered by these consid-erahons.
Naturul variation in vegetation compositionond s t ruc lu re . Desp i te . r t templs l i , In in m, /ewithin-block variability, at sqne sites topographicconfigurations, land designations, and past man-agement activit ies (e.g., adjacent harvest units orroads) precluded the establishnent of six, 13-haexperimental units within a forest ofhomogeneoustopography, vegetation, and stand history. Con-sequently, stand age, tbrest structure, and speciescomposition vary among experimental units priorto treatment (Table 4). This heterogeneity hasimplications for interprcting the experimental feat-
ments and resulting ecological responses. Forexample, because the total basal area retained inforest aggregates dictates the total basal area re-tained in dispersed units. pre-treatment di11'erenceswill skew the true propofiion of basal area retaired.
Variation in vegetation structure and composition prior to treatment (Table 4) must also beconsidered in interpreting post har.,'est responses.To minimize these effccts, we will fbcus on cfirrge.tin abundance/diversity between pre- and postharvest communities. Nonetheless, where com-pe l i l i \e in le r rc l ion . a re i rnponant de ter rn inant .of species rcsponses, and where there are largedifferences in the init ial abundance of competi-tors. effects of treatments may be difficult to de-tect.
Separttthg effects ofbgging disturbance fromellects of retention: short-tenn versus long-termconsiderations. A primary goal of our researchis to elucidate the effects of varying levels andpattems ofgreen-tree retention on lbrest community development. Although there has been con-certed effbrt to mjnimize eftects of confoundingvariation (Aubry et al. 1999). operationally, lev-els ofdisturbance will change markedly with theproportion of basal area removed and with thepattem in which it is removed. Thus, it is l ikelythat short-term responses of vegetation will reflect differential patterns of logging disturbancaas much as, or more than. pattems of retention.Sampling of post-harvest disturbance character-istics will permit an analysis of these effects. Asdisturbance efttcts diminish with time, canopyeffects may dominate understory responses, al-though legacies of difterential disturbance maycontinue to shape successional trajectories (Halpern1988). Thus, green-tree reiention strategies in-corporate more than variation in the leveland spatialarangement ofretained structwes they encom-pass an array ofco-varying paftems and processes.To the extent that we can distinguish disturbanceellects from those associated with canopy struc-tures, we may better understand the mechanismsthat shape vegetation response and more succcss-fully guide new strategies tbr fbrest management.
Acknowledgements
D. Berg and J. Franklin conceived ofthe originaldesign tbr a large-scale expedment on green-treeretention in the Pacific Nonhwest. R. Cunis and
Vecetation ResDonse to Green tree Retention 4l

D. Minore contributed to the initial vegetationsampling design. G. Spycherhas been insh'unentalin developing the database nanagement systen.The goals and approaches presented in this paperreflect the ideas and suggestion ofmany individuals: R. Abbott, M. Amaranthus. K. Aubry, L.Bednar, D. Berg, R. Bigley. N. Christensen, D.DeBell, N. Diaz. D. Ford, J. Franklin. M.Hemstrom, A. Hofion, D. Neeley. L. Noris, M.Raphael. J. Reynolds, G. Smith, F. Swanson, J.White. and B. Woodard. We thank S. Acker, K.Aubry P Burton, D. Luoma,A. Olson, M. Raphael,J. White, and an anonymous reviewer for commcnts on carlicr vcrsions of this manuscript. Fortheir t ireless effbrts we gratetully acknowledge
Literature CitedAbbott. R. S.. J. D. White. rnd B. L. $ixxl.rd. 1999. The
benefits and chrlleDges of large'\cale sil!iculturalexperinents: perspecti\es fioln tbrest managers onthe Demonstr.rlion oi lcLrsysleln MaMgement Optrors(DEMO) nudy. Nonh$. Sci .7 l (Spcc. ls \ . ) : l l8 125.
Acker. S. A. . E. K. Zenner. and w. H. E'n 'n ingham. 1998.Structufe . rnd y ie ld of r$o-rged dands or theWilldmett NrdN.rl lorc!r Oregor: implicrdurs lorgrccn trcc rctcntion. Can. J. For. Rcs. l8i7,l9 758.
Adlcr. P B. 199,1. Nell forestr! in pracrice: A suney ofr ' ^ r ' J l i l \ r r p r c c l r c ( r c r c l r o r h r n c r r , n i t . . u c . tem Cascades. Oregon. Hanard Universiq'. Cambridge.N{llsiachusetts. B.A. Thesis.
Agee.J.K. 1991. Fi feecolog! ofPaci f ic Norhnest forests.Islrnd Press. \\hshhgton. D.C.
Alaback. P 8.. and F. R. Hcnnan. 1988. I -ong rcrm rcspon\cof unde.\iory \'egeration Io sland densitt in Pnr.dTrrgd fbrests. Crn. J. For. Res. 18:1522-1530.
Al \ 'erson. $ ' . S. . D. M. Wal lef . and S. L. Solheim. 1988.Forests t(xr deef: edge efiects in nonhem Wi\consin.Consen. Brc l . 2: -1.18'358.
Aubr) . K. B. , V P Annranrhus. C. B. Hulpern, J. D. White.B. L. \\'oodrrd. C. E. Pcr.Ison. C. A. Lagoudakis. andA. J. Horton. 1999. Evaluadng the efitcrs of\aryingle\els and p.rtterns of green-tfee ferenlion: e\pefi-ment.rl design of the DEN{O stud). Noth$. Sci. 73(Spec. lss.) :12 26.
B.r i le) . J . D. 1996. l l lects of s l rnd densi ty reduct ion onslructural dcvclopman( in lvc\tcm C)lcgoi DouglasI l l lbrcsls A rccon\ l ruct ion stud! . Orcgon StatcL , \ e , . , ) . ( ^ r ' . l l . P \ l ) . D : . . e r J , , , , , .
tsarrett. J. $'. 1980. RegionalSihicultLrrc ofrheLrnited States.Second edirion. John $'iley and Sons. Ne$ York.
Bcrg. A. . B. Ehnstrdm. I - . Cusla l ! ion. T. I {a lhngbict . M.Jonsel l , and. l . \vesl ie. . I 99 '1. I hreaiened plant . animrl . and fungur species in Sxedish forests: d ist r ibut ion and habi tat associat ions. Consen. ts io l .8:7187 3 1 .
oul f-leld assistants, too numerous to namc indi-vidually. Forlogistical support we thank R.Abbott.D. Allau'ay, D. Anderson. R. Bigley, D. Couchc.B. Fontaine. L. Fitzer. F. Henchel, W Kleckner,S. Macmeeken. J. Nakae, E. Tompkins. J.Walmslcy, and J. White.
This is a product of the Demonstration ofEcosystem Management Options (DEMO) study. ajoint eftbrt ofthe USDA Forest Service Region 6and Pacific Northwest Research Station. Researchpaftners include the University of Washington.Oregon State University, University of Orcgon,Gittord Pinchot and Umpqua National Forests,and the Washington State Department of NatumlResources.
Bn!h, K. R., and K. N. Johnson. 1992. Standle|el woodprodnclion cosrs L'f le.rviDg live. mature tree\ rt fe-generalion harlcsl incoaslal Douglas lir stands. \\'est.J. Appl . For.7165 68.
Brothem. T. S.. and A. Spingarn. 1992. Forcst liagmcn|rt ion and al ien plant in lasion of centra l lndiana oldgrolvrh lbresls. Conserv. Bio l . 6:91-100.
Busing, R. L. C. B. Halpern. and T. A. Spic\ . 1995. Ecology ofPacificYew (7iiurs r,?rlirlid) in lvestern Oregon and$'ashingion. Consef! . Bio l . 9: l 199-1207.
Carc) , .A.8. . and \ { . L. JohnsLrD. 1995. Smal lmanmals inmanrgcd. nrturall) young, rnd old-gro$th fi)rests.Ecol . Appl . 5: l l6 352.
Chen.J. . J . F. Frankl in. and T. A. Spjcs. 1992. Vcgctat ionresponses to edge enllronments in old gro$tl Douglrs tir lbrests. Ecol. Appl. 2:387-396.
. 1993. Contrasting nicroclimarcs ilnong clearcut.edge, and inlefi(n of old-sroqth Douglas iir forcsl.,A.gf. Fof. Nleleof. 63:2 1 9-237.
-. 1995. Gro$nrg-serson microclim.rtic grudients r-romcleur cut edges irtLi old-gro\\'th Dougl.r,i-lir forests.Ecol. Appl. 5:74 86.
Chr is iensen. N. l - . . A. N{. Bartuska. J. H. Bro$n. S. CarpeDter, C. D Antonio, R. Francis. .1. H Frankl in. J . A.MacN1ahot. R. n Noss. D. J. Parsons. C. H. Petefson.M. G. lurner. and R. C. Woodmanscc. 1996. Thereto . ot rh( | .o lngr.J l \^c 'cr) J t \m. r ( - (urnm
tee on the scieniific basis forecosystem manag.m.nr.Ecol . Appl . 6:665-691.
Coates. K. D. 1997. Windthro\| damage 2 )ea|s afier pardal
culling tl| thc Dxte Creek silvicultural syslems stud,\'in lhe interior cedar hemlock forcst\ ol northwcnernBr i t i \h Columbia. C.rn. J. For Res.27:1695 l l0 l .
Dlrness. Cl. T. 1971. Early s[ges of p]ant succession fol-lo\l'ing logging and slashbumilg ir rhe westem Crs-cadcs ofOrcgon. Ecologl 5.1:57 68.
12 Halpern et al.

FEMAT. 1993. Foresr ecos,"-s(Jn anagement: .lll ecological. economic. ,lnd social assessment. Report of thcForest Ecosyslcnr N4anagemenl Assessmenl T,i.rm.USDA For Ser\'. r USDC Nat. Ocem. Atnos. Admin.aDd Nar. Mar. Fish. SeN.r LISDI Bur. L.]l1d Nianage..fish wildl. Sen.. and Nat. Park Sen.: and US FIPA.Washinglon. D.C.
Franklin. J. F. 1989. To!'ards a new forestq AmeL for. 951
Franklin. J. F.. and C. T. Dyrncs\. 1973. Naiurrl legelationof Oregon and [ashington. USDA For Sen.. Ger.Tech. Rep. PNw li.
Ffanklin. J. F.. P NI. Frcnzen. and F. J. S\\'aDson. 1995. Rccreation ofccosystems ar Nt(nnn St. Helens: conirrstsir rlilicial and natutal rpproaches. 1, J. Cdms. Jr(ed.). R.habilinling Damaged Ecoslstems. Sccondedi t i (D. Le\ i is Publ ishcrs. Boca Ratm. Pp. 281 333.
Frankl in, J . F. , H. H. Shugarr and \ ' I . F l . Hammn. 1987. Treedeath .Ls an ecological proccss. Bioscience 37:55().5 5 6 .
Frankl in. J . F. L. A. Norr i \ . D. R. Berg. and G R. Smith.l o o . ' I n ( \ r . t r ' \ . n l \ l n . r i \ p e ( r l I n r .generation har\cst of nofih$estem li)rest ecosvslcms.\or th$. Sci . l3 (Spcc. lss.) : l - l L
Frankl in. J . F. . D. R. Bcrg. l ) . A. Thombufgh, and J. C.Tappeinef. 1997. Alcmalivc silvicultural rpprcachesto timber harvesting: \ariable retention han'est svs-tems. ./l1 K. A. Kohln. rnd J. F. Fianklin (eds.). Cre-ating a Fofestry ibr thc 2l'' Ccntury: Ihe Science ofEcosystem N{dnagcmcnl. lsland Pfess, washinglon.D . C . P p . 1 1 1 1 3 9 .
Fr.rnklm. J. n. K. Cromack. Jr., W Derison. A. McKcc. C.Nhser. J. Scdcl l . F. Sranson. . rnd G. Juday. 1981.Ecologicrl chrractcrjnics of old-growth Douglas ilrfofests. USDA For. Scr!.. Cen. l'ech. Rep. PN\\'-118.
Fraver. S. 199.1. Vegetation r.sponses along edge-ro-inte'rior gfadient\ in the l1ixcd hardqood lbresls ol theRomoke Rivef B$in. Nol1l Carolina. Consen'. Blol.8:822- i132.
Frost .E.J. 1992. Thc c l lcc ls of ibre, i l -c leafcur edges on lhcstnrclurc and conrporition of 01d-grorvth mi),cd coni ler stxnd\ in l le \ \estern Kl .Lmrth Nlountai rs.Hunboklt S(alc Univcriit,v, Afcata. Calilbrnia. Nr.S.Thesi \ .
Grace. J. ts . 1990. On the re la l ionship bet \ |een planl t fa i tsand comfet i t i le abi l i t ) . 1 l1 J. B. Crace. and D. Ti lman(eds.) . Pefspertnes on PhnlCo pctr t ion. AcrdelnicPfes\ , Srn Dieso. Pp. 5 l 65.
Grime. J. P 1917. [vidcncc lirr the eristence of three primary strllegics in plants and its relevaDce to ecologrcal and eloludonary iheor_1. r\'n. Nfi. I I l:1169 ll9.+.
Gruinbine. R. E. 199,1. Whal i \ cco\ ls ten manrgement lC o n s e n . B i d . 8 : 2 7 3 8 .
H.llpcnr. C. B. 1988. ElLlly ruccessional palhwa,vs and theresisrance rnd rc\ilience of l orcsl com munities. Ecol-og,v 69:170-:- 1715.
Halpcrn. C. B.. and T. A. Spies. 1995. Plant spccics divcrsiq in naturaland ln naged lorests ofrhc Paciiic Norlhwest . Ecol . Appl . 5:91.1 93,+.
Halpern.C.B..J. F. Frankl in.andA. N4cKee. 1992. Changesin plaDt species diversi!y after hanest of Douglas-firfbrests. Nonhw. Environ. J. 8:205-207.
Hansen. A. J. , T ' A. Spics, F J. S$anson. and J. L. Ohmrnn.1991. Conser\ing biodivcnily i, managed forests.BioScience. l l :381-392.
Hanscn. A. J.. S. L. Gafman, J. F. \\'eigand. D. L. Uftan. WC. Mccomb,.rnd \1. G. Raphael . 1995. Al lcmat i \esil\'icultufal regimes in rhe Pacilic Northwc!u rimLrlations ofecological and cconorrjc eftlcts. Bcol.Afpl.5:535'55'1.
Harmon. \ f . E. , J . F. Frunkhn. F. J. S$anson. P Sol l ins. S. VGregory. J. D. Lal l in . N. H. Anderson. S. P Cl ine. \ .G. Aumen. J. R. Sedel l , G. W. L icnkacmper. K.Cromack. Jr , and K. W Cumlnins. 1986. Ecologyofcoarse nood! debrs in tempefute ecos)s|en\. Ad!.Ecol . Res. l5: l l l -302.
Hi l l . M. O. 1973. DiveI \ i t ) and evenness: a uni l l ing notation and its consequerces. Ecdogl 5.1:,121 ,112.
Hop\\'ood, D. 1991. Principlcs and pracdces of nelv foFe\iry. L.rnd N{lln. Rep.7l. B.Cl. Nijn. For., Victofia.ts.c.
Hutfman. Il. W, J. C. nrppeirer, ll. and J. C. Zasada. 199.1.Regeneration of srl.rl (Cdlrlu,rr7 rr.rll.)r) in thc ccntralCloast Range fofests of Oregon. Can. J. Br't. 121395 l
Hunrer, l\'L 1995. Residual trees as biological legalrles.Management Colnlnu qu6 No. 2. Clascade Center forEcoslsten Nllrnugcncnl. Corvallls.
Jules,E.K. 1997. Consequenccs of I orcst t iagmentat io n forthe u ndef\tof) planL ?ri lliun o\rnun (l .iliaceael. ln' l ' .
N. Ka]e. A. L iston, R. M. Lo\e. D. L. l -uoma. R.J. N{elnke. and NL V \ \ i lson (eds.) . Conscr!atronandN{ anage ment of Nalive Plants rndFungi. \ati\c PlanlSociety of Oregon. Corvallis. Pp. 101 206.
Kcssler. w B.. H. Salr.rsser. C. W Cunwright. Jr.. and J.Caflan. 1992. New perspecri\ c\ lbr susrai.able naturalresource lnunagel1lcl1t. Ecol. Appl.l:221 225.
LaLrrance. W F. 1991. Edge cllccs in rropical forest frag-ments: .Lppliearion of .r modcl 1br dcsign of na$rereserves. Bio l . Con\en. 57:2{15 219.
Lchmkull. J. I-1. S. ll. West. C. L. Chanbers. \{ C. McCoiIb.D. A. Niaru\ \a l , K. B. Aubr! , J . L. Ef ick$n. R. A.Ci l^n, and M. Leu. 1999. A\sessing $i ld l i ie response Io larying le\els and prtterns of $.cn lrccretention in $estern Oregon trnd wlshinglon. \ori|w.Sci . 73 (Spec. Iss.) : ,15 63.
Nlaser. C., R. F. Trrmnt. J. Nt. Trappe. and J. F Fr,l klin (tech.
eds.) . 1988. Frm lhe lbrest to lhe sea: l sror ] oflallcn lrees. t-rSDA For Serv.. Gen. Tech. Rep. PNwGTR 229.
\{all.rck. G. R. 1993. Microenlifonmenr v.ui.rtioD $ ithirand a olrg foresi edge sites in the Eastern LlnitedStalcs. Bio l . Con\eI \ . 66: 185- 19.1.l o c o . L r f l ) - . . ( . ' r " r J l p . . r r r ' l . o f f r i . . p e -
c ies: inter.rcrions o I li I c I i\!ort traits and distufbance.Ecolost 70:70.1 720.
Halpefn, C. 8. , . rnd J. F Frankl in. 1990. Phlsrognonrc dc!elotment of PJ.rdi).llrg.r lbresls in relati(m to ini-t ia l s t ruclure.Lnd disrurbrncc jn icrs i r ) . .1. \ tg. Sci .l i , {75 ,182.
l o o l r . P t r n r . p e ' ' ' e r i i r . , r ' . r ' n - n i ' . r l ' i n ' :forest landscape ln eastern Noih Arncrica. Ecolog,v7 5 : 1 1 9 1 - 1 5 0 2 .
199.1b. \regeration dr-ranics oflhc lbrc\t edgelrends in sprce and successional l imc. J.Ecol . Slr l l lI2-1.
Vesetation ResDonsc to Green tree Retention .13

NlcKcnzie.D. 199,1. Diameter growth eqnations and growth.1Dd yield prciecti(nrs lbrtlvo-iiered stands in rveslemWirshington. University of \Vashington. Sca(lc. Ivl.S.Thesis.
Morrisor. P H.. and F. J. Swan\on. 1990. fire history andpaltcm in a Cascade Range landscape. USDA For.Sen.. Gen. Tech. Rep. PNW GI R 25,1.
Nlurcia. C. 1995. Edgc cllccls in liagmenred lbrests: inpli-cat iors lbr conser\r t ion. Trends Ecol . Evol . 10r5862.
North, N{. 199-1. Sland slrucrurc and lrullle abundance asso-cialcd witb northcrn spotted owl habitat. Uni\ersn.voi $' slingtor, Seritie. Ph.D. Dissetatron.
North, M., J. Chen, G. Smith, L. Krakowiak. and J. Fiankl in.1996. Initial response of undcrstory pianl diversiryand olcrslory uee diumelef glowlh n) a green tree re-r e f l i , ' n h - r \ e . r . \ o r L u S c i .
- 0 : ' l l :
O'Dea. \'I.. i. C. Zasada. and J. C. Tappeiner. 1995. Viremuple clonal development in coastalDouglas-fir rbresrs. Ecol . Appl . 5:63-13.
Olnef. C. D.. and ts. C. Larson. 1990. Fbrest Stand Dlnanics. Nlc(;ra$ Hill, Ncw York.
Peck, J. E.. and B. l\'lcCunc. I991. Renn.rnl lrees and canopylichen conmuDitie\ ln $e\tern Oregon: r retfospect i \ e a p p f o a c h . E c o l . A p p l . 7 : l l 8 l 1 1 8 7 .
Perry. D. A. 199,1. Forcsl Ecoslstems. John\ Hopkin\ Uni-!ersill Press. B.rltilnofe.
Pern'. I). A.. and M. P Anaranthus. 1997. DistLrbance.reco\ef i . and sr . rh i l i ry . / , K. A. Kohm. and J. F.Franklin (eds.). Crcaring a Forcnr] lbr |he 21'' Cen1ur, , - : The Science of Ecos]srem \4anagemenL lsland Press, Wal l r ingur. D.C. Pp.3l 56.
Perry. D.,4.. N,l. P Amaranthu\, J. G. BorcheN. and R. F-.Brainc . lgE9. Boolstrapping in eco\yslems.ts ioScience 39:130 237.
Ranne), J. \{ 1977. Fofest i\land edges-their structure.dr\elopmcnl. and inportance t|) rcgional lbrest eco-s lsten d!nl l l l r ics. En! . Sci . Di ! . Publ . No. l06c), OalRrdge National l-aborarory. Oak Ridgc. TenDessee.
RItnnc). L W. \ ,L C. Bruner, rnd J. B. Leven$n. 1981. Ihermportance of edge in thc slructurc and dynamics oftirest islancls. 1" R. L. Burgess, and D. \4. Sharpe(cds.). Forcst lslard Dynalnics in Vrn-dominatedLandscape!. Springcr Vcrlag. Ncs York. Pp. 67 95.
Rice. $'. R. 1989. Analyzing tables of strtistical teirs. Evolul ion,13r l l3 225.
R,^c. e. R. . . .nJ P S. \ lur . . loo. i . C-rrr r ree -r . . f l r^nconsequences lirr trmbcr producrion in lbrests oI the$esrem Cascades. Oreg(m. Ecol . Appl . 7:2 i )9-217.
Ruggiero. l-. F.. K. B. Aubr!. A. B. Carc). and Nl. H. HulT(tech coords.) . 1991. Wi ld l }e and vegetat ion ofunnanugedDouglas-firtbrests USDAFoT Sen..Gen.'1ech. Rep. PNW GTR 185.
SchooDnuker, P, and A. NIcKee. 1988. Species compo\i!ion and di\crsil,"" durng second.rl successrcn olco-nilerous fofests in rhe western Cascade Mou.lains C)1'Oregon. For. Sci .3,1:960 979.
Scho$altcr. T. D. 19U9. C nop)- althropod communitv structure and herbivory in old gro$th and regeneratingfbrests in lvestern Oregon. Can. J. For Rcs. l9 i3 l8l2t .
Sokal . I t . R. . and F. J. Rohl i . 1981. Bioner,"- . W H. Free-n n and Comprn). San Francisco.
Spies. T. A. l99l. Plant species diversiiy and occuffence inyoung. mature. and old grolrth Douglas-fif stands in$estern Oregon and \\'ashington. 1, L. F Ruggiero.K. B. Aubr,v. A. B. Care) ' . . ard M. H. HuU (ech.coords.). Wildlife and vegetalion of onmanaged Dou,glas-fir forestli. USDA l-or Ser\'.. Ccn Tcch. Rep.PN\\ ' -GTR-185. Pp. l l l l2 l .
Spies, T. A.. and J. Fl l-rankljn. 1991. The structure of natu-ral young. maiurc. and old grcwth Douglas-fir for-esls in Orcgon and \\hshington. /, L. F. Ruggiero,K. B. Aubry. A. B. Carey. ,tnd M. H. Huff (coords.).Wildlife and vegelalion of unnanaged Douglas-firforcsis- USDAFoT Serv..Cen. Tech. Rep. PNW-GTR-2 8 5 . P p . 9 l - 1 0 9 .
Spies, T. A., J. F Franklin. and T. B. Thomas. 1988. Coarsewood]' deb.is in Douglas fir forcsts ol western Or-egon and Washington. Ecoiogy 69:1689-1702.
Tappeiner.J. C. . J . C. Zasada. P Ryan, andM. Newton. 1991.Salmonb€rr]' clonal and population structurei the basisibr a persistenl co!cr. Ecolog] 72:609-618.
Tappeiner. J. C-. D. Lavendcr, J. \lilslad, R- O. Cufiis, andD. S. DeBel l . 1997. Si lv icul lural systems and regen-eration methods: cunent practices and new iltema,lives. /, K. A. Kohm. and J. F. Franklin (eds.). Cre-ating a Forestry for the 2l" Cenlury: The Science ofEcosvstem Managemcnt. lsland Press, Washington,D.C. Pp. l5 l 16.1.
Teensma.PD.A. 1987. Fi rchis lory and l i re regines of thecentral western Cascadcs c'l Oregon. Universit), ofOfegoo. Eugene. Ph.D. Disscrtalion.
Thonas, J. $1, N,l. G. Raphael. R. G. Anthon),. E. D. forsnan,A. G. Gunderson. R. S. Hol thausen. B. C. Marcor c.H. Ree\es. J. R. Sedel l , and D. \4. Sol is . 1993. Viability assessment,i and manageincnt consideratiLrnsfor species associaled with lalc successional and old-growth foresls of lhc Paciic North$esi. A repof ofthe scientific anallsis lcam. USDA For. Serv., PoIt-land. Oregon. 530 p.
Traut. B. H. 199.1. Efttcts ofrariation in ccos)sten cuTvoleron biodrvenity and connnuni$ slructuii ol lbrestlloorbrlophyres rncl understory vascular planlsi a rerospecrile rppro.rch. Ofegon Stae Univcrsity. Clor!.tllis.IU.S. Thesis.
Tuchnrann. E. T.. K. P Connaughton. L. E. FreedmaD, and C.ts. N{oriwaki. 1996. Thc Northwest Forest Plan. Areporrto the Presjdcnt and Congres s. USDA Otr. For.Bcon. Assl.. Porlland. Oregon. 153 p.
US DA and U S D L l 99:1. Record o l dccision lbr anendnentsto Foresr Senice and Bureau of Land N{magenentpluning documents within the range of$e norfiernspotted o$1. USDA F'or. Serv.. Po and, Oregon.
Wales, B. A. 1912. Vegetation anallsis ol nLrrth aDd southedges ln a aturc oak hickor] lbrest. Ecol. Monogr..12:151 ,17l .
Willer. J. W. 1975. Thc elect ol thinning intensit] uponundersnry g(Nrh and species conposiiim in an Oregon Coas! Ra gc Douglas lir sraDd. Ofegon StateUniler\iry, Corlallis. [,i. S. Thcsis.
Zar. J . H. 198,1. Bio\rar is t ical Analysis. Sccond edi t ion.Prent ice-Hr l l . Inc. . Engle\ \ood Cl i l \ .
Zennef. E. K.. S. A. Acker. and W. H. Emmingham. 1998.(;rowth reduclion in hancsr age. conilefous fore\islr'ith rcsidual rccs in |he western central CascadeRange ofOrcgon. F 'or 0col . Munage. 102:75-88.
11 Halpern et al



![A Proposed Probabilistic Extension of the Halpern and ... · been proposed by Halpern ([2008], pp. 200–5), Halpern and Hitchcock ([2010], pp. 389–94, 400–3), and Halpern and](https://img.pdfslide.net/doc/110x75/6057db13fe4a5562be12ee7a/a-proposed-probabilistic-extension-of-the-halpern-and-been-proposed-by-halpern.jpg)



![DANIEL HALPERN-LEISTNER - Columbia Universitydanhl/derived_equivalences... · DANIEL HALPERN-LEISTNER In [BO], Bondal and Orlov made the following Conjecture 0.1 (D-equivalence)](https://img.pdfslide.net/doc/110x75/6057dbfe94cc0e1ab62d2580/daniel-halpern-leistner-columbia-danhlderivedequivalences-daniel-halpern-leistner.jpg)





















