Embed Size (px)
Citation preview

THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 243, No. 10, Issue of May 25, pp. 2587-2596, 1968
Printed in U.S.A.
Kinetics of Escherichia coli B D-Lactate Dehydrogenase and Evidence for Pyruvate-controlled Change in Conformation*
(Received for publication, September 28, 1967)
E. M. TARMY~ AND NATHAN 0. KAPLAN
From the Graduate Department of Biochemistry, Brandeis Un,iversity, Waltham, Massachusetts 0215.J
SUMMARY
Kinetic properties of D( -)-specific lactate dehydrogenase (D-lactate : diphosphopyridine nucleotide oxidoreductase, EC 1.1.1.28) from Escherichia coli B indicate the following.
1. The reaction catalyzed is essentially unidirectional, the oxidation of D( -)-lactate with diphosphopyridine nucleotide as coenzyme proceeding at only about 0.01% of the rate of pyruvate reduction.
2. The enzyme shows Michaelis-Menten kinetics for re- duced diphosphopyridine nucleotide, but pyruvate activates it.
3. A lag in the rate of oxidation of the reduced diphospho- pyridine nucleotide is eliminated by prior incubation with pyruvate. The duration of this lag is independent of enzyme concentration, suggesting that pyruvate causes a change in the conformation of the enzyme rather than in its state of aggregation. The identity of the molecular weight of the enzyme at a very low concentration in the absence and in the presence of the substrate confirms this fmding.
4. Two lines of evidence indicate that there are two types of binding sites for pyruvate. The variation of Hill plots with pH is interpretable only in terms of two sites with differ- ent binding properties. Furthermore, the two sites bind different substrate analogues. Oxamate is an inhibitor of this enzyme, but does not substitute for pyruvate as an ac- tivator. On the other hand, a-ketobutyrate does not in- hibit the catalysis but activates the enzyme, converting the substrate saturation curve from sigmoidal to hyperbolic.
5. The kinetic properties of the catalytic site appear to be similar to those of vertebrate L-lactate dehydrogenases, although the dissociation constants for both pyruvate and reduced diphosphopyridine nucleotide are considerably larger for the E. coli D-lactate dehydrogenase.
6. The kinetic properties of this enzyme are consistent with the course and regulation in vivo of pyruvate metab- olism.
* Publication 555 from the Graduate Department of Biochem- istry, Brandeis University. This study was supported by Re- search Grant P77-J from the American Cancer Societv. bv Grant CA-03611 from the National Institutes of Health, ancI b$ Grant GB-5708 from the National Science Foundation.
$ Present address, Department of Biochemistry, University of Leicester, Leicester, England.
In a preliminary report we have described a DPNH-dependent lactate dehydrogenase from Escherichia coli B that differs markedly from vertebrate lactate dehydrogenases in its catalytic properties (1). This enzyme has now been purified to homo- geneity. The method of purification and some of the chemical properties of the enzyme have been described in the preceding paw (2).
This enzyme had been described erroneously as catalyzing the formation of L-lactate from pyruvate (1); the product of the reaction is now known to be u-lactate. The remainder of the results described in the preliminary report have been confirmed: this n-lactate dehydrogenase (n-lactate:DPN+ oxidoreductase, EC 1.1.1.28) has unusually high Michaelis constants for pyruvate and DPNH, it exhibits sigmoidal kinetics with respect to pyruvate, and it is inhibited by oxamate, which, however, is not an allosteric effector. These kinetic studies have been expanded and are described in this report.
There are, in E. coli, three distinct enzymes concerned with the pyruvate-lactate interconversions. One of these is the D-
lactate dehydrogenase, specifically dependent upon DPNH as cofactor and, as noted above, forming the D( -) isomer of lactate. Most likely, this enzyme is functionally a unidirectional enzyme in vivo as well as in vitro. The other two enzymes concerned with this interconversion are enzymes, not dependent on pyridine nucleotide, catalyzing the conversion of lactate to pyruvate, each enzyme specific for one isomer of lactate (3, 4). These enzymes (called “lactate dehydrogenase” or “lactate oxidase” in the literature) are probably flavoproteins (3), and are perhaps related to yeast cytochrome bz.
Whereas L-lactate oxidase and, to a lesser extent, n-lactate oxidase are inducible enzymes in E. coli (3), the specific activity of the u-lactate dehydrogenase of E. coli 58-161 does not change markedly with conditions of growth. That the E. coli u-lactate dehydrogenase is an allosteric enzyme with pyruvate acting as the activator at a site distinct from the catalytic site is suggested by the present studies. As a consequence of the properties of these sites, the activity of this enzyme appears to be controlled by the cellular concentrations of pyruvate and H+. No other metabolites were observed to affect the activity of the enzyme in vitro, except adenine nucleotides, which are effecters at non- physiological concentrations, and a-ketobutyrate, which prob- ably acts as a substrate analogue.
by guest on July 16, 2020http://w
ww
.jbc.org/D
ownloaded from

2588 E. coli D-Lactate Dehydrogenase Kinetics Vol. 243, No. 10
TABLE I Stereospecificity of E. coli lactate dehydrogenase
Oxidation of DPNH and formation of D- and n-lactates with limiting concentrations of pyruvate, after incubation with the enzyme for 1 hour. The concentration of DPNH was 4.5 mM.
Pyruvate concentration DPNH oxidizeda D-Lactateb formed L-Lactateb formed
?nM ?nM ?&If 7n.M
0 0.05 0.4 0.38 0.25 0.6 0.55 0.55 1.0 0.94 0.95 0.0
a Determined from absorption at 340 rnp. b Assayed by the method of Dennis and Kaplan (6).
TABLE II
Reduction of DPN+ by lactate
The reaction mixture (1 ml) contained lactate (0.24 M), DPN+ (0.7 mm), sodium pyrophosphate buffer (0.03 M), pH 8.9, and lactate dehydrogenase (95 units).
Isomer of lactate Rate of reaction
AAAlpa/minlml
DL 0.037 D 0.13 L 0.001
EXPERIMENTAL PROCEDURE
Materials
The purification of the n-lactate dehydrogenase from E. coli B is described in the preceding paper (2). For some of the kinetic studies, impure enzyme solutions were used. In no case were interfering activities present. Crystalline chicken Hq lactate dehydrogenase was prepared by the method of Pesce et al. (5). Leuconostoc mesenteroides n-lactate dehydrogenase was partly purified by Mrs. R. Garland.’ Egg white lysozyme was purchased from Worthington.
Coenzymes and adenine nucleotides were purchased from P-L Biochemicals, Milwaukee, Wisconsin, sodium pyruvate from Sigma, lithium D( -)-lactate and L(+)-lactate from Miles Laboratory, Elkhart, Indiana, and cr-ketobutyrate from K $ K Laboratories, Inc., Brooklyn. Oxamic acid, purchased from Matheson Coleman and Bell (Division of the Matheson Com- pany, Inc., East Rutherford, New Jersey), was neutralized with NaOH prior to use. Reagent solutions were always pre- pared each day and kept at 4”.
Sephadex G-200 was purchased from Pharmacia, Uppsala. An addition product between DPN+ and pyruvate, the re-
duced form of the binary complex, was prepared by Dr. R. H. Sarma of our laboratory.2
Methods
Determination of Stereospec$city-The stereospecihcity of this enzyme was determined with crude extracts and with 1% pure, 20% pure, and homogeneous preparations. Limiting amounts of pyruvate and excess DPNH were used, and DPNH oxidation was measured spectrophotometrically until the amount of DPNH
1 R. Garland and N. 0. Kaplan, unpublished results. 2 R. H. Sarma and N. 0. Kaplan, unpublished results,
that remained became constant (after about 1 hour), at which time the pyruvate was completely reduced. Samples were then assayed enzymatically with n-lactate dehydrogenase from L. mesenteroides and Hd n-lactate dehydrogenase from chicken heart for the formation of D-lactate and n-lactate, respectively (6).
Kinetic Studies-Most assays were performed on a Cary model 14 recording spectrophotometer. Rapid reactions were measured with a stop-flow apparatus (7), which consisted of two syringes containing the reactants connected to the cell of a Beckman DU spectrophotometer. The reactants were mixed rapidly in the spectrophotometric cell with an automatic plunger. The rate of reaction was recorded both on a Gilford recorder and on a Tektronix type 564 storage oscilloscope with a differential amplifier.
All assays were performed at room temperature, or about 23”. The ionic strength for all experiments was 0.24.
Molecular Weights-The molecular weight of the catalytic unit of the enzyme was determined by gel filtration (8) through Sephadex G-200, both in the absence and in the presence of pyruvate.
Optical Rotatory Dispersion-The optical rotatory characteris- tics of this lactate dehydrogenase were determined, both in the absence and in the presence of pyruvate, with a Cary 60 spec- tropolarimeter.
RESULTS
Stereospecificity-When this enzyme was used to catalyze the complete reduction of small amounts of pyruvate in the presence of excess DPNH, the pyruvate was stoichiometrically converted to the D( -) isomer of lactate. Results of a typical experiment are given in Table I.
The reduction of DPN+ by the enzyme was studied with a racemic mixture of lactate and pure D( -)- and L( +)-lactates. The assay mixture (1.0 ml) contained lactate (0.24 M), DPN+ (0.7 mu), sodium pyrophosphate buffer, pH 8.9 (0.03 M), and E. co& lactate dehydrogenase (45 units). The rate of reduction for each form of lactate is given in Table II. The reaction in this direction was very slow, and was significant with DL- or n-lactate but not with n-lactate. There was no reduction of DPN+ in the absence of n-lactate or of the enzyme. The rate of reduction observed with the complete reaction mixture was about 0.01% of the rate of oxidation of DPNH by this enzyme.
Dependence of Rate upon Pyruvate Concentration-In a prelimi- nary report we noted that a plot of rate against pyruvate concen- tration is a curve with sigmoidal character (1). This observation with enzyme preparations of 1 to 100% purity has been made numerous times.
The curve shown in Fig. 1 is representative of the results of every experiment in which the pyruvate concentration was varied. The rate was shown to be directly proportional to enzyme concentration at several concentrations of pyruvate.
When a Lineweaver-Burk plot is made of these data, the points fall on a curved line. However, when only those points obtained with relatively large concentrations of pyruvate were plotted, a straight line was generated (Fig. 2), with intercepts corresponding to K, = 7.2 no and V,, = 286 pmoles of DPNH oxidized per min per mg of lactate dehydrogenase. The apparent K, calculated in this way was very close to the concen- tration of pyruvate at which the velocity is half of the observed maximal velocity. The Vmax calculated from this Lineweaver-
by guest on July 16, 2020http://w
ww
.jbc.org/D
ownloaded from

Issue of May 25, 1968 E. M. Tarmy and N. 0. Kaplan 2589
Burk plot was about 20% higher than the rate observed under optimal conditions. This discrepancy is probably due to sub- strate inhibition.
The turnover number, at pH 7.5 and 0.3 mM DPNH, was 3.4 x lo4 moles of DPNH oxidized per min per mole of lactate dehydrogenase.
Hill Plots-It is expected that a Hill plot for an enzyme with multiple, interacting binding sites will be sigmoidal, with the slope approaching 1.0 at the extremes of low and high substrate concentrations and being greater than 1 at intermediate concen- trations. The Hill parameter, nn, is defined as the slope of a tangent to the curve at half-maximal velocity, i.e. at v/(V,, - v) = 1.0 (9).
For E. coli lactate dehydrogenase, Hill plots for pyruvate are indeed sigmoidal, the slope approaching 1 at very low and at relatively high pyruvate concentrations. However, from the plot for pyruvate at pH 7.5, shown in Fig. 3, it was calculated that nn is 1.20. In other words, the slope approached 1 at a lower value of v/(Vm, - V) than expected from the theory of Hill plots, described by Wyman (9). Indeed, at the pyruvate concentration for which v/(V,, - V) is about 0.7, the curve is already approaching a slope of unity. At lower values of vI(vlnax - v), the slope equals 1.9. A new parameter, n’n, has been defined as the slope of the curve at v/(Vm, - v) =
100
j
-=
2 a 90
n/ /
z
80 -
70 -
60 -
50 -
40 -
30 -
20 -
25
20
15
IO
5
0 5 IO I5 20 25 30 Pyruvate Concentration ( m MI
FIG. 1. Pyruvate saturation curve for E. coli lactate dehydro- genase at pH 7.5. The DPNH concentration was 0.33 mM and the ionic strength, 0.24. The inset is the section from 0 to 3 rn~ pyruvate on an expanded scale. Each symbol represents the mean of five assays, with vertical bars to indicate the standard error of the mean. Different symbols are used to indicate different concentrations of enzyme.
-15 -10 -5 0 5 IO I5 20 I/[Pyruvate] (mM-‘1 (~10~)
FIG. 2. Lineweaver-Burk plot for pyruvate at pH 7.5; 0.33 mM DPNH.
1.0
_ 0.1 5 1 :: E
> Y > m 0
.Ol
.OOl
0.1 1.0 IO log [Pyruvatc] (mM)
100
FIG. 3. Hill plot for pyruvate at pH 7.5, 0.33 mM DPNH, and ionic strength 0.24. Different symbols are used to indicate differ- ent concentrations of enzyme.
0.5, in order that the results obtained with this enzyme may be described numerically.
Dependence of Rate upon pH-The relation between rate and pyruvate concentration has been examined at several pH values. These results are summarized in Fig. 4 and Table III. Lineweaver-Burk plots indicate that the K, was relatively constant between pH 6.4 and pH 7.5, whereas the V,, was a
by guest on July 16, 2020http://w
ww
.jbc.org/D
ownloaded from

E. coli D-Lactate Dehydrogenase Kinetics Vol. 243, No. 10 2590
100
so
N I 80 0 I 70 .- c
z 80 Ii .- E
G 50
lo
z 40
5! 30
z .-
g 20 z
IO 5
0
, pH 8.5
0 2 4 8 12 I8 20 24 28 32 36
Concrntrotion of Pyruvota (mM)
FIG. 4. Dependence of the rate of reaction upon pyruvate concentration at several pH values. In each case the concentra- tion of DPNH was 0.33 mM and the ionic strength was 0.24. For each curve, different symbols are used to indicate different con- centrations of enzyme.
Lineweaver-Burk plots for pyruvate at different constant concentrations of DPNH (Fig. 6Q show that K, = 7 mM at pH 7.5. The characteristics of Hill plots for pyruvate did not vary with changing DPNH concentrations in the range, 0.086 to 0.57 mu DPNH.
TABLE III Kinetic constants for pyruvate reduction at different pH values
PH K0.P Gb V,,Xb Vm.x/W+lb a'&
?nM gyrwate X10-
6.4 2 5 457 1.1 1.8 6.7 2.5 4.4 343 1.7 2.0
7.5 5 7.2 286 9.0 1.9
0 Ko.5 is defined as the pyruvate concentration that gives 50% of the observed maximal velocity.
b K,,, and V,,, were determined from Lineweaver-Burk plots. e n’n was determined from Hill plots.
Ozamate 1&ibition--Oxamate, an inhibitor of animal lactate dehydrogenases (12), also was found to be an inhibitor of the E. coli enzyme. The oxamate concentration was varied between 0 mu and 30 111~. The DPNH concentration was 0.33 mM; the buffer was 0.1 M potassium phosphate, pH 7.5. Experiments in which oxamate concentration was varied were done at a number of constant pyruvate concentrations between 4 and 30
function of PH. Hill plots all show the same characteristics, with n’n essentially constant in this pH range. However, the range of pyruvate concentration through which cooperative binding was operative appeared to become narrower as the pH was lowered, as shown in Fig. 5.
I f the observed decrease in Vmax with pH was due to an increase in the K, for DPNH in this pH range, it would indicate that the coensyme is rate limiting in alkaline buffers. However, it is known from experiments described below that the coenayme is present at saturating levels at pH 7.5, the most alkaline pH for which V,,,ax and K, could be calculated.
If this observed increase of Vmax with H+ concentration was due only to the inclusion of a proton from the medium in the V max term, as indicated by the equation for the over-all reaction
Pyruvate + DPNH + H+ --+ n-lactate + DPN+
then Vmax/(Hf concentration) would be constant as the pH was varied. As is shown in Table III. this was not the case.
-I 0 I
Logarithm of Pyruvate Concentration
FIG. 5. Hill plots for pyruvate at several pH values. 0, pH 6.4; A, pH 6.7; El, pH 7.5.
It may be noted (Fig. 4) that it was not possible to determine meaningful values for V,, and Km of this enzyme at pH 8.5, because the highest pyruvate concentration that could be used was 33 mM. Consequently, it was also impossible to make a Hill plot requiring the value of the V,, in its construction. However, the sigmoidal character of this curve is readily apparent in Fig. 4. Also, the apparent K, for pyruvate is considerably greater than 7 mu since, at that concentration, the rate was only about 10% of that observed at 33 mu.
Dependence of Rate upon DPNH Concentration-The E. coli
n-lactate dehydrogenase appears to follow Michaelis-Menten kinetics for DPNH at all pyruvate concentrations between 3 mu and 30 mu. Lmeweaver-Burk plots for DPNH at several constant concentrations of pyruvate are shown in Fig. 6A.
The apparent K, for DPNH increases when the pyruvate concentration is reduced to levels below the apparent K, for pyruvate. Just as has been found for animal lactate dehydro- genase, the DPNH concentration at the common point for intersection is higher than any of the individual apparent K, values (10, 11). The V,, at saturating levels of pyruvate was 322 pmoles of DPNH oxidized per min per mg of lactate dehy- drogenase, in good agreement with the Vmax value obtained when pyruvate was varied at saturating levels of DPNH.
by guest on July 16, 2020http://w
ww
.jbc.org/D
ownloaded from

Issue of May 25, 1968 E. M. Tarmy and N. 0. Kaplan 2591
mM. The results are shown in Fig. 7, A and I?. Fig. 7A is a plot of l/v against oxamate concentration at different, constant pyruvate concentrations at different, constant levels of oxamate. The inhibition by oxamate was of the “mixed” type. The appar- ent Ki for oxamate was about 18 mM. Hill plots for pyruvate at the different oxamate concentrations gave n’n values of 1.5 to 1.9, with no indication that there was any regular change in n’n as the oxamate concentration was increased.
With aspartate transcarbamylase, low concentrations of maleate activate the enzyme at low aspartate concentration, presumably by causing the same conformational change usually
A
-4 -2 0 2 4 6 8
I/[DPNH] (mM-‘)
0 5 IO 15 20
I/[Pyruvate] (mhl-‘1 (x Id)
FIG. 6. A, Lineweaver-Burk plots for DPNH at different, constant concentrations of pyruvate with an ionic strength of 0.24. The pyruvate concentrations were A, 3 maa; 0, 6 mu; V, 9 mM; X, 12 mu; and 8, 15 mu. B, Lineweaver-Burk plots for pyruvate at different constant concentrations of DPNH. The DPNH concentrations were Q, 0.86 mM; A, 0.14 mu; 8, 0.17 mM; v, 0.34 mM; and X, 0.57 mu. The point on the abscissa corresponding to the intersection of the curves at different con- .-- . centrations of substrate is -l/K, (10). nucleotides do not appear to be inhibitors.
36 r-
Symbol [PY~] I 2 ::
4mM 5mM
3 0 7mM 4 A IO m M 5 x 15 m M 6 0
A -
0 5 IO 15 20 25 30
Concentration Oxamata (mM)
36
Symbol [~xamatc] f7 I I x 2 0 3 0 4 A 5 0 6 0 7 .
B -
OmM 32 5mM
lO.mM 15 m m
28 ,
20mM 25mM 30 mM
0 4 6 12 16 20 24 26
I / [Pyruvate] mM (x 100)
FIG. 7. A, reciprocal velocity plotted against the concentration of oxamate at several constant concentrations of pyruvate (pyr). The DPNH concentration was 0.33 mM. B, Lineweaver-Burk plots for pyruvate at several constant concentrations of oxamate at the same DPNH concentration.
induced by the substrate, aspartate (13). No such activation by oxamate was found for E. coli lactate dehydrogenase at a low pyruvate concentration (Fig. 8).
a-Ketobutyrate Inhibition--+Ketobutyrate is a very poor substrate for this enzyme; the rate of DPNH oxidation with 20 mM ar-ketobutyrate was about 1 To of that with pyruvate as substrate. However, when 20 mu a-ketobutyrate was added to the assay mixture and pyruvate was varied, the substrate satura- tion curve was hyperbolic, the Lineweaver-Burk plot was linear, and a Hill plot indicated that the reaction was first order in pyruvate (Fig. 9).
ATP Inhibitzon-E. wli n-lactate dehydrogenase is inhibited by ATP, as are the n-lactate dehydrogenases of Butyribacterium ret&pi (14) and Lactobacillus pluntarum (15) and the n-lactate dehydrogenase of the latter organism (15). Other adenine
by guest on July 16, 2020http://w
ww
.jbc.org/D
ownloaded from

E. coli D-Lactate Dehydrogenase Kinetics Vol. 243, No. 10
0 2 4 6 8 IO
Concentration of Oxamate 1 m M 1
FIG. 8. Inhibition of pyruvate reduction by oxamate at 1 rnnl pyruvate and 0.33 mM DPNH, pH 7.5.
ATP was a poor inhibitor at a concentration of 1 mu. At 4 mM, the triphosphate appeared to be competitive with DPNH (Fig. 10). Hill plots for pyruvate and for pyruvate with 4 mM ATP were virtually identical in shape although displaced from one another along the abscissa, indicating that ATP does not affect the interaction (or interactions) between pyruvate and the enzyme.
conditions the apparent Ki was 0.013 mM, which is lower than the apparent Km for DPNH.
Activation by Pyruvate-The suggestion that the conformation of the active site of the E. coli lactate dehydrogenase changes upon binding of pyruvate is derived from an examination of the effect of pyruvate upon the initial rate of its reaction with DPNH. We observed that there was a pronounced lag in the rate of reaction at low initial concentrations of pyruvate (Fig. 12). As the pyruvate concentration was increased, the duration of the lag decreased, until the lag could not be observed with a Cary model 14 recording spectrophotometer.
300
200
7 \ m
.fI 5
100
0 3 6 9 12 15
[Pyruvate] (m Ml
B
. ..’ . . . .
I%oduct Inhibition-When the activity of a sample of lactate dehydrogenase is measured by the standard assay (2), the rate begins to decrease after about 0.15 Mmole of DPNH has been oxidized. This decrease was thought to be caused either by product inhibition by DPN+ or n-lactate or, by analogy with the animal lactate dehydrogenases (16), by the formation of an abortive ternary complex among enzyme, pyruvate, and DPN+.
The enzyme was incubated for 2 min with DPN+ (0 to 0.67 mu), pyruvate (15 or 30 mM), and potassium phosphate buffer (0.1 M), pH 7.5. The reaction was initiated with 0.33 mu DPNH. No inhibition was observed when initial rates were measured.
D-Lactate is virtually ineffective as an inhibitor of pyruvate
1.0
0.5
reduction. The enzyme was incubated with n-lactate (0 to 30 mM), DPNH, and potassium phosphate buffer at the same concentrations as mentioned above. The assays were initiated after 2 min by the addition of 15 mM pyruvate. The apparent Ki was about 0.25 M.
Inhibition by Reduced Binary Complex-As already mentioned, we believe that the substrate inhibition that we observed with vertebrate lactate dehydrogenases is caused by the formation of an abortive ternary complex (16-B). A binary complex of pyruvate and DPN+ has been synthesized that acts as an inhibitor of vertebrate lactate dehydrogenases and is presumed to mimic the observed substrate inhibition.
> L 0.1 \ 5
0.05
The purified reduced binary complex was shown to be an inhibitor of the E. coli n-lactate dehydrogenase. A plot of recip-
0.01 L
rocal rate against concentration of inhibitor is shown in Fig. 11. I 5 IO
The enzyme was incubated for 5 min with complex (0 to 26 PM), Pyruvate ( m Ml
pyruvate (30 mM), and potassium phosphate buffer (0.1 M), pH FIG. 9. A, substrate saturation curve for pyruvate, pH 7.5, in
7.5. The reaction was initiated with 0.33 mu DPNH. Initial the absence (X) and in the presence (0) of 20 mM cr-ketobutyrate.
rates of reaction were measured. Under these experimental B, Hill pIots of these data. The slope, n’n, at v/(V - v) = 0.5 was decreased from 1.7 to 1.0 by or-ketobutyrate.
by guest on July 16, 2020http://w
ww
.jbc.org/D
ownloaded from

Issue of May 25,1968 E. M. Turmy and N. 0. Kaplan 2593
If this lag results from some conformational change in the lactate dehydrogenase upon binding of the substrate, giving a more active form of the enzyme, a lag should also be observed at a saturating concentration of pyruvate and should be eliminated by prior incubation with the substrate. This was observed with a stop-flow apparatus.
The results of three related experiments done with the stop- flow apparatus are summarized in Fig. 13 and Table IV.
In the first experiment, the lactate dehydrogenase was placed in one syringe and pyruvate and DPNH in the other prior to mixing. In this way, the enzyme was not exposed to either reactant until the reaction was initiated. As shown in Fig. 13, Curve A, a distinct lag was observed: 5 see elapsed before a constant rate of oxidation of DPNH was observed. This con- stant rate amounted to 1.19 pmoles of DPNH oxidized per min, and was reproducible within about 1 TO in five trials.
In the next experiment, lactate dehydrogenase and DPNH were placed in one syringe and pyruvate in the other.prior to mixing. In this case, the enzyme was incubated with coenzyme
0 5 IO 15
I/[DPNH] (IJIM-‘)
20
FIG. 10. Lineweaver-Burk plots for DPNH with (X) and with- out (0) 4 mM ATP.
20 X/
‘6 - n
0 5 IO 15 20 25
Concentration of Binary Complex (PM)
FIG. 11. Reciprocal rate plotted against concentration of the pyruvate-DPN+ complex at 30 mM pyruvate and 0.33 mM DPNH.
c \ 0 zl
a
t
\ \ \
\
\ \
t 0.1
L
\
1 I I I
I5 45 75 105
Seconds FIQ. 12. Rate of oxidation of DPNH at 1 mM pyruvate, 0.33
mM DPNH, and pH 7.5. The solid Zinc was traced from the Cary 14 recording spectrophotometer. The dashed line has been added to show, under these conditions, the extent of the lag over 1 min.
FIQ. 13. Oscilloscope patterns showing initial rate of oxidation of pyruvate by E. coli lactate dehydrogenase. The ordinate is Aaro; one division = 0.05 optical density units. The abscissa is time; one division = 1 sec. CzLrve A represents the initial rate of reaction when the enzyme was incubated separately from the substrate and coenzyme prior to mixing, and Curve B the rate when the enzyme was incubated with pyruvate. The assay mix- ture contained pyruvate (25 mM), DPNH (0.33 my), potassium phosphate buffer (0.1 M), pH 7.5, and purified lactate dehydrogen- ase (4.5 pg). The curve that was obtained when the enzyme was incubated with DPNH was qualitatively similar to that obtained when it was incubated alone.
and the reaction was started by the addition of pyruvate upon mixing. A lag was observed, again lasting for about 5 sec. However, the rate calculated from the apparently linear portion of the curve was not reproducible but, rather, slowly fell as the time of incubation increased. The rate decreased 5% in 8 min; this decrease would not have been observed with a less repro- ducible method of assay. When the rate was extrapolated to zero time of incubation, it was 1.14 pmoles of DPNH oxidized
by guest on July 16, 2020http://w
ww
.jbc.org/D
ownloaded from

2594 E. wli D-Lactate Dehydrogenase Kinetics Vol. 243, No. 10
TABLE IV
Effects of substrate and coenzyme upon initial lag
Assays were performed with stop-flow apparatus as described in the text. The concentrations of the various reactants were lactate dehydrogenase, 4.5 rg per ml; pyruvate, 25 mM; and DPNH, 0.22 mM. Potassium phosphate buffer, 0.1 M, pH 7.5, was present in both syringes at a concentration of 0.1 M.
A
B
C
Syringe 1
Lactate dehydrogen- ase
Lactate dehydrogen- ase and DPNH
Lactate dehydrogen- ase and pyruvate
Syringe 2
Pyruvate and DPNH
Pyruvate
DPNH
DllrZLti0n of lag
set
5
5
>>0.2
Rate of reaction
m~moles DPNH
oxidized/ min
1.19
1.14a
1.43
a Extrapolated to 0 min of prior incubation; see text for details.
0.12
0 ~
IO 20 30 40
0.24
0.06 5 2
0
Elution Volume (ml)
FIG. 14. Elution of 3.2 rg of E. co&i lactate dehydrogenase per ml from Sephadex G-200. X, blue dextran marker; 0, lactate dehydrogenase alone; A, lactate dehydrogenase plus 33 mM pyru- vate. The arrows represent the calibration curve, in which A is the elution peak of blue dextran, B that of chicken Hq lactate dehydrogenase, and C that of egg white lysozyme. The Sephadex G-209 column was 49.5 X 1.1 cm, and the flow rate was about 6 ml per hour. After equilibration with potassium phosphate buffer (0.i M), pH 7.5, containing mercaptoethanol (6.1 M), a sample (0.4 ml) containing 3.2 ~g of E. coli lactate dehvdrogenase ner ml and blue dextran 2&O ii applied and eluted with the same buffer. The total volume of each fraction was measured. Because of the extremely low concentration of enzyme in the fractions, l-ml portions were used in the assay mixture (2) and the rates were followed for 10 min on a Zeiss model PM& II spectrophotometer. The rates appeared to be linear. The column was then calibrated with a sample (0.4 ml) containing blue dextran 2000, chicken HI lactate dehydrogenase, and egg white lysozyme. Afterward, the column was equilibrated with the phosphate buffer, containing pyruvate (30 mM) as well as mercaptoethanol (0.1 M). Then a sample (0.4 ml) containing E. coli lactate dehydrogenase (3.2 pg per ml), pyruvate (30 mM), and blue dextran 2000 was applied and eluted with the buffer containing pyruvate. The fractions were analyzed as described above, except that less pyruvate was put into the assay mixture before addition of the enzyme sample, to compensate for the pyruvate that was being added together with the enzyme.
per min. This is in good agreement with the rate calculated for the first experiment.
In the final experiment, lactate dehydrogenase and pyruvate
were in one syringe and DPNH in the other prior to mixing. In this case, the enzyme was incubated with pyruvate and the reaction was initiated with coenzyme. No lag was observed in five trials; results of one trial are shown in Fig. 13, Curve B. Again, the rate observed was reproducible. However, it was about 25% higher than that observed in the previous experi- ments, being 1.43 pmoles of DPNH oxidized per min.
The higher rate found in this experiment, in which there was no lag, probably is the “true” maximal rate of reaction. The lower rate found in the other two experiments may represent a portion of the curve, which appears to be linear, between the initial lag and the later slowing down of the reaction; this de- crease in rate is presumably due to formation of the abortive ternary complex.
Further studies with the stop-flow apparatus showed that the duration of the lag did not vary as the enzyme concentration was changed between 1.8 and 9 pg per ml. The pseudo first order rate constants calculated for the activation depended on the concentration of pyruvate, below 20 mu. The observed dependence suggests that the change is second order in pyruvate. This is in agreement with the indication from Hill plots that the over-all reaction is at least second order with respect to pyruvate.
Molecular Weight of Catalytic Unit-The observation that the half-time of the lag was independent of the enzyme concentration suggested that there is no change in the state of aggregation of the subunits of the enzyme upon binding of pyruvate. This was confirmed by the determination of the molecular weight of a catalytic concentration of enzyme in the absence and in the presence of pyruvate.
The elution peak of the E. coli lactate dehydrogenase in the absence of pyruvate corresponds to a molecular weight of 130,000 f 15,000 (Fig. 14). This value is in excellent agreement with that obtained by gel filtration with a higher concentration of enzyme (2). Furthermore, the elution pattern was the same in the presence of pyruvate, indicating that there is no change in the molecular weight of this enzyme upon binding of the substrate.
Optical Rotatory Dispersion--An attempt was made by optical rotatory dispersion to detect a change in the peptide Cotton effect upon binding of pyruvate. However, there were no differ- ences apparent in the spectra between 225 and 350 rnp with or without pyruvate present.
DISCUSSION
The simplest interpretation of the anomalous kmetics observed when pyruvate was varied is that there are two types of binding sites for the substrate on the enzyme. One type, with a rela- tively low dissociation constant, is associated with the activation of the enzyme by pyruvate. The second, with Km = 7 mM at pH 7.5, is at the catalytic site of the enzyme.
If this hypothesis is correct, it would be expected that, at low pyruvate concentrations, the reaction would appear to be second order in pyruvate. However, at higher concentrations, the conformational, or allosteric, site would be wholly occupied, the enzyme would be fully activated, and the reaction would appear to be first order in the substrate. This hypothesis fits the observation that at low pyruvate concentrations the slope of the Hill plot for pyruvate is about 2, whereas at higher concentrations it is only 1.
If there are two distinct pyruvate-binding sites with diierent binding constants, the observation that the concentration range for which the slope of a Hill plot is 2 becomes smaller as the pH
by guest on July 16, 2020http://w
ww
.jbc.org/D
ownloaded from

Issue of May 25, 1968 E. M. Tarmy and N. 0. Kaplan 2595
is lowered suggests that the K, for the allosteric site decreases more, or more rapidly, than does the K, for the catalytic site as the pH is lowered. Thus, H+ would appear to be acting as an allosteric effector, by lowering the concentration of pyruvate at which the interactions are apparently abolished. It is note- worthy that Wittenberger and Fulco have recently published evidence suggesting that the n-lactate dehydrogenase from Butyribacterium rettgeri not only is activated by pyruvate but may also be activated by H+ (14).
Since the enzyme appears to be fully activated at concentra- tions of pyruvate about 5 mM, it is possible to compare the kinetics observed with relatively high concentrations of pyruvate with what is known about vertebrate lactate dehydrogenases. The turnover numbers are similar (19). The K, for pyruvate (catalytic site) is affected similarly by a change in pH (20). Substrate inhibition decreases with pH for both enzymes (21). Oxamate inhibits both enzymes in a similar manner (12). The K, for DPNH is greater than any of the apparent K, values, and K, = Km for pyruvate for both enzymes (11). Both enzymes are affected in the same way by the reduced binary complex.3 From these observations it would appear that the mechanism whereby pyruvate is reduced by the bacterial enzyme may be chemically similar to that which is operative in vertebrate lactate dehydrogenases. This similarity would not be surprising.
With regard to the activation by pyruvate, the lag in the initial rate of oxidation of DPNH is significant at saturating as well as at low concentrations of substrate. This lag is eliminated by prior incubation of the enzyme with the substrate, suggesting that the binding of pyruvate effects a change, either in the state of aggregation of the subunits or in the conformation of the enzyme. The former possibility has been ruled out by studies showing that the molecular weight of a very low concentration of enzyme is the same both in the absence and in the presence of pyruvate.
The conformational change probably is subtle, inasmuch as it is not detected by optical rotatory dispersion studies. This fact is not surprising, for x-ray crystallographic studies on hemoglobin have suggested that the Bohr effect is caused by a very small change in conformation, on the order of 7 A (22).
The studies with substrate analogues are interesting with regard to the hypothesis that there are two distinct pyruvate sites. Oxamate, an inhibitor, competes with pyruvate for the catalytic site on the enzyme; however, it does not activate the enzyme at low concentrations of pyruvate. Another analogue, ol-ketobutyrate, does not appear to bind to the catalytic site very well if at all; it is a very poor substrate and does not inhibit significantly the reaction of pyruvate at 20 mM. However, this analogue does act as an allosteric activator, changing the sub- strate saturation curve from sigmoidal to hyperbolic. This fact suggests that this compound may be replacing pyruvate at a second, allosteric site. This behavior of theE. coli lactatedehydro- genase with respect to ol-ketobutyrate is quitesimilarto that found by Wittenberger and Fulco for the n-lactate dehydrogenase from B. rettgeri with this analogue, except that it is an inhibitor of the B. rettgeri enzyme (14).
It is not surprising that the activity of the E. coli lactate dehydrogenase appears to be determined by the metabolic en- vironment of the enzyme, since this organism is a facultative anaerobe and the fate of pyruvate differs markedly depending upon conditions of growth.
3 J. Everse and N. 0. Kaplan, unpublished results.
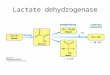
FIG. 15. Diagram of pyruvate metabolism in E. coli. The major fermentation products of glucose metabolism are n-lactate, acetatft, formate, ethanol, COZ, and succinate; they are under- lined m the figure. PEP, phosphoenolpyruvate; D-LDH, D-
lactate dehydrogenase.
A diagram of the metabolism of pyruvate in E. coli is shown in Fig. 15. Amarasingham and Davis have shown that this orga- nism obtains its energy from glycolysis when grown aerobically as well as when grown anaerobically (23). It may be that when the concentration of pyruvate in the cell is low the lactate dehy- drogenase is inactive (wholly or partly) and pyruvate is available for other reactions; when the pool of pyruvate is higher, only a portion of the substrate is used for oxidation of DPNH because the K, for the catalytic site of the enzyme is so high. The pyruvate pool is thereby not depleted completely by what is only one of the metabolic reactions of pyruvate.
Because of the observed dependence of this enzyme on pH, it would be expected that if the pool of pyruvate is below about 10 mM, which seems reasonable, more lactate would be produced in tivo at acid pH. This is true, as was shown by Tikka (24). He studied the fermentation products of E. coli suspensions in defined media at diierent pH values. At pH 6.3, 40 to 50% of the glucose that fermented was converted to lactate; at pH 7, only 20 y0 was fermented to lactate; and at pH 7.5, only 3 y0 of the glucose was converted to lactate. As the amount of lactate fermentation decreased, there was a parallel increase in fermenta- tion to acetate plus formate, probably arising by way of the phosphoclastic reaction. The ethanolic fermentation was con- stant, at about 20% of the total.
The present studies on the effect of pH are thus consistent with the course of fermentation in vivo.
In vertebrates, the following roles have been proposed for the two types of subunits of lactate dehydrogenase found in animals (25). The form of lactate dehydrogenase predominating in muscle, Mq lactate dehydrogenase, is active under physiological conditions when the concentration of pyruvate rises, thus per- mitting the oxidation of DPNH under the anaerobic conditions that obtain during muscle contraction. On the other hand, in highly aerobic tissues, such as the heart, Hd lactate dehy- drogenase predominat,es. This form of lactate dehydrogenase is inhibited in the presence of relatively high concentrations of pyruvate because of the formation of the inactive, ternary com- plex. Therefore, in such tissues as the heart, the pyruvate is converted not to lactate but to acetyl-CoA and is channeled into the Krebs cycle, generating necessary metabolic energy for these
by guest on July 16, 2020http://w
ww
.jbc.org/D
ownloaded from

E. coli D-Lactate Dehydrogenase Kinetics Vol. 243, No. 10
tissues. The function of the H-type subunit may be to convert lactate to pyruvate.
In E. coli it appears that the regulation of the pyruvate-lactate interconversion is effected by the presence of a totally distinct enzyme, o-lactate dehydrogenase, which acts essentially as a pyruvate reductase, and by flavin-linked lactate oxidases, which appear to be inducible enzymes (3). Hence, in E. co& the regulation is controlled by both flavin and pyridine nucleotide- linked enzymes.
Acknowledgments-We gratefully acknowledge the able assist- ance of Reba Chakrabarti during a portion of these studies, and thank Ronald Barnett and David Barker, who performed the experiments with the stop-flow apparatus. We are indebted to Professor William P. Jencks for helpful discussions during the course of these studies.
1.
2.
TARMY, E. M., AND KAPLAN, N. O., Biochem. Biophys. Res. Commun., 21, 379 (1965).
TARMY, E. M., AND KAPLAN, N. O., J. Biol. Chem., 243, 2579 (1968).
3. KLINE, E. S., AND MAHLER, H. R., Ann. N. Y. Acad. Sci., 119, 905 (1965).
4. HAUG~ARD, N., Biochim. Biophys. Acta, 31, 66 (1959). 5. PESCE. A.. MCKAY. R. H.. STOLZENBACH. F. E.. CAHN. R. D..
6. 7.
AND’KAPLAN, N.‘O., J.‘BioZ. Chem., 2$9, 1753 (1964). ’ DENNIS, D., AND KAPLAN, N. O., Biochem. Z., 338,485 (1963). STURTEVANT, J., in B. CHANCE, R. H. EISENHARDT, Q. H.
REFERENCES
8. 9.
10. 11.
12.
13.
14.
15.
16. 17.
18. 19.
20. 21.
22.
23.
24. 25.
GIBSON, AND K. K. LONBERG-HOLM (Editors), Rapid mixing and sampling techniques in biochemistry, Academic Press, New York, 1964, p. 89.
ANDREWS, P., Biochem. J., 91, 222 (1964). WYMAN, J., Cold Spring Harbor Symp. Quant. Biol., 23, 483
(1963). FRIEDEN, C., J. Amer. Chem. Sot., 79, 1894 (1957). HAKALA, M. T., GLAID, A. J., AND SCHWERT, G. W., J. Biol.
Chem., 221, 191 (1956). NOVOA, W. B., WINER, A. D., GLAID, A. J.. AND SCHWERT. G.
W., J. BioZ..Chem., 434. 1143 (1959). ’ GERHART. J. C.. AND PARDEE. A. B.. Cold Swrina Harbor
Symp. &ant. &ok, 28, 491 (i963). ’ * ” WITTENBERGER, C. L:, AND F&co, J. G., J. Biol. Chem., 242,
2917 (1967). MIZUSH~MA,‘S., HIYAMA, T., AND KITAHARA, K., J. Gen. Appl.
Microbial., 10, 33 (1964). WINER, A. D., Acta Chem. &and., 17, S293 (1963). KAPLAN, N. O., EVERSE, J., AND ADMIRAL, J., Ann. N. Y.
Acad. Sci., in press. LEE, H. A., AND WINER, A. D., Fed. Proc., 26, 557 (1967). PESCE, A., FONDY, T. P., STOLZENBACH, F., CASTILLO, F.,
AND KAPLAN, N. O., J. Biol. Chem., 242, 2151 (1967). NYGAARD, A. P., Acta Chem. Stand., 10, 408 (1956). WINER, A. D., AND SCHWERT, G. W.. J. Biol. Chem.. 231, 1065
(1958). PERUTZ, M. F., BOLTON, W., DIAMOND, R., MUIRHEAD, H., AND
WATSON, H. C., Nature. 203, 687 (1964). AMARASIN~HAM, 6. R., AND DAVIS, B. D.., J. Biol. Chem., 240,
3664 (1956). TIKKA, J., Biochem. Z., 279, 264 (1935). DAWSON, D. M., GOODFRIEND, T. L., AND KAPLAN, N. O.,
Science, 143, 929 (1964).
by guest on July 16, 2020http://w
ww
.jbc.org/D
ownloaded from

E. M. Tarmy and Nathan O. KaplanPyruvate-controlled Change in Conformation
B d-Lactate Dehydrogenase and Evidence forEscherichia coliKinetics of
1968, 243:2587-2596.J. Biol. Chem.
http://www.jbc.org/content/243/10/2587Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/243/10/2587.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on July 16, 2020http://w
ww
.jbc.org/D
ownloaded from