Embed Size (px)
Citation preview

Journal of the American Mosquito Control Association, 12(2):194-198, 1996Copyright O 1996 by the American Mosquito Control Association, Inc.
LACTATE DEHYDROGENASE AS A MARKER OF PLASMODIUMINFECTION IN MALARIA VECTOR ANOPHELES
M.-E RIANDEY' C. SANNIER,I G. PELTRE,'N. MONTENY3 eNn M. CAVALEYRA3
ABSTRACT Lactate dehydrogenase (llh) electrophoresis showed the presence of Plasmodium yoeliiyoelii in Anopheles stephensi and An. gambiae. The Mh appeared as an additional band (pLdft) whoseactivity was more intense with 3-acetyl pyridine adenine dinucleotide as coenzyme than with p nicotin-amide adenine dinucleotide. Several allelic forms occurred both in the vector and the host. The isoelectricpoint of Uh, similar in the vector and host, differed from those of ldh from mosquito and mouse. Thepresence of pldh was detected from the 2nd to the 28th day of infection. T'he pl^dh appeared to beproportional to the number of sporozoites present in infected salivary glands. However, pldh was notfound in salivary glands or midguts, but it was detected in the rest of the corresponding mosquito. Theorigin and use of pldh as a marker of Plasmodium in its vector is discussed.
INTRODUCTION
Throughout the world, about 2 million peopledie of malaria annually (Gentilini 1991), with120 million clinical cases/year and 300 millionpeople carrying the parasite (World Health Or-ganization 1992). Different strategies are used tofight this disease and one involves control of theAnopheles vectors. A rapid method to diagnosemalaria-infected and malaria-infective mosqui-toes is needed.
Plasmodium can be detected in human bloodusing Giemsa-stained smears, whereas in mos-quitoes, microscopic examination of midgutsand/or salivary glands is necessary. This lattermethod is labor intensive and time consumingand many mosquitoes have to be dissected todetermine sporozoite rates. Other techniqueshave thus been developed, including the en-zyme-linked immunosorbent assay (ELISA)(Wirtz et al. 1985) and DNA probes (Delves etal. 1989, Ponglikitmongkol et al. 1994).
Analysis of Plasmodium enzymes (Broun1961; Sherman 196l; Carter 1973:' Walliker etal. 1971, 1973) showed that host- and parasite-specific enzymes could be distinguished. InPlasmodium falciparum, the coenzyme for lac-tate dehydrogenase (llh) is preferably 3-acetylpyridine AD (APAD), whereas the activity ofhuman llh requftes B nicotinamide adenine di-nucleotide (NAD). This property permits mea-surement of parasite llh and malarial diagnosisin man (Makler and Hinrichs 1993). Our goalwas to adapt this method to reveal pMh of Plas-
rLaboratoire de Lutte contre les Insectes Nuisibles(LIN), Centre de recherches ORSTOM Ile-de-France,32, Av. Varagnat,93143, Bondy, Cedex, France.
2 Laboratoire d'Immunoallergie, Institut Pasteur75724.Pais Cedex 15. France.
3 Laboratoire de Lutte contre les Insectes Nuisibles(LIN), Parc Scientifique Agropolis, 34397, MontpellierCedex 5, France,
modium yoelii yoelii in our model vector Anoph-eles stephensi Liston in order to develop a rapidand reliable technique to detect the parasite inmosquitoes.
MATERIAL AND METHODS
Parasite: Plasmodium y. yoelii (Killick-Ken-drick 1974), strain 17X, a rodent malaria para-site, was obtained from the lab of Biologie Par-asitaire, Helminthologie et Protistologie du Mu-seum National d'Histoire Naturelle, Paris (I.Landau). It was stored at -80"C in mouse bloodwit}r 5Vo glycerol.
Vectors: The An. stephensi strain used in thisstudy was collected in Iraq in 1973 and rearedat 2O-24'C, 67 + 2Vo RH, and a 12/12 photo-period. Three strains of Anopheles gambiae Gi-les were included in the study: "Bobo," origi-nating from Burkina Faso, was obtained fromInstitut Pierre Richet (IPR) at Bouak6 (Cdted'Ivoire), and 2 strains obtained from the lab ofEcologie des Systbmes Vectoriels (Institut Pas-teur, Paris). The latter two strains included astrain from The Gambia that was sensitive toPlasmodium (G3) and a second strain (Blue) thatwas selected from G3 for its refractoriness toPlasmodium cynomolgi (Collins et al. 1986,l99l; Vernick and Collins 1989).
Mosquito samples: Infected mice were cagedovernight with An. stephensi, resulting in an87Vo infection rate. Bloodfed mosquitoes werecollected the following day. Every 2 days fromday 2 to day 28 following the blood meal, 10mosquitoes were frozen at -80'C and stored un-til electrophoresis analysis. The presence of oo-cysts in midguts (from day 6) and sporozoites insalivary glands (from day 14) was checked bydissection and microscopic examination.
Sporozoite purffication: After checking posi-tive for the presence of sporozoites (day 17),20O mosquitoes were ground in RPMI medium
194



JuNe 1996 DETEcrroN oF Mar-ep.h-IxrscrED MoSQUIToES
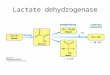
PARASITE LDH
I
i
A/ \
) | 6 \
/ \
/ \
EcIl|)
oo o.t
mosouTo LoH
CONTBOL
LOW llilFECTlON (1+)
l{GH INFECnOil (3+)
DISTANCE IN CII
Fig. 4. Lactate dehydrogenase zymogr:rm of Anopheles stephensi debris. Densitometric recording of the Ldhintensity.
oA
le
between infected and infectious mosquitoes be-cause the infection stage could not be deter-mined; however, there was good correlation be-tween the cause (parasite) and the effect @InD.Therefore, pldh is a good marker for the Plas-modium parasite. The technique is rapid; a singleperson can test 100 mosquitoes daily. It is notexpensive as only small quantities of APAD, themost expensive chemical, are needed (50 mg/100 mosquitoes). The ELISA and DNA proberequire specialized equipment. more expensivechemicals. and several long successive process-es. Furthermore, the ELISA has to be calibratedto avoid false-positive or -negative responses.This electrophoretic method is suitable for fielduse. Nevertheless, further work is required to ex-tend this method to other species of mosquitoesinfected by other Plasmodium species.
A number of questions were raised by the factthat pldh was only detected in the vector, not inthe parasite. Is this enzyme really synthetizedbythe parasite or the mosquito as a response to theparasite infection? If so, what is the origin of the"pldh" present in the blood of infected mice?If these "p" bands are synthetized either by thehost or the vectot does this only occur as a re-sponse to the presence of Plasmodium or also asa response to other aggressions? Sherman (1962)
showed that another parasite (Babesia rodhaini)did not cause this phenomenon in the mouse, butgiven the size of the parasite, he suggested thatthe detection technique is perhaps limited. Insome diseases, additional Mh is considered asevidence of disease (Yamaoka and Kameya1972, Beyer et al. 1990).
Howeve! pldh activity was more intense withthe APAD coenzyme than with NAD and the pIlevel was the same in mouse and mosquito.These results are in agreement with the sup-posed synthesis of this enzyme by the parasite.The localization of pllh outside Plasmodium iscomparable to another type of enzymatic induc-tion: lipolysis in host tissues, which provides li-pidic acids for intraerythrocytic growth of theparasite under control of a lipase originatingfrom the parasite (Vial and Ancelin 1992).ln tl:resame way, pllh seems to be active outside ofthe parasite, allowing ATP production by thehost for the glycolytic pathway of Plasmodium(Dnbeli et al. 1990).
REFERENCES CITED
Babiker, H. A., A. M. Creasey, B. Fenton, R. A. L.Bayoumi, D. E. Arnot and D. Walliker. 1991. Ge-netic diversity of Plasmodium falciparum in a vil-

198 JoupNer oF THE AMERICAN Moseurro Cotvrnol Assocranor.l Vor-. 12. No. 2
lage in eastern Sudan 1. Diversity of enzymes, 2D-PAGE proteins and antigens. Tlans. R. Soc. Trop.Med. Hyg. 85:572-577.
Beyer, C., M. Hofland and D. G. Que. 1990. Lactaredehydrogenase isoenzyme 6 in serum of two pa-tients with severe pre-eclampsia. Clin. Chem. 36:4 t t - 412 .
Broun, G. 1961. Enzymes 6rythrocytaires et infesta-tion d Plasmodium berghei chez la souris. Rev. Fr.Erud. clin. Biol. 6:695-699.
Carter, R. 1973. Enzyme variation in Plasmodiumberghei and Plasmodium vinckei. Parasitology 66:297-3U.
Collins, E H., S. M. Paskewitz and A. E. Crews-Oyen.1991. A genetic study of Plasmodium susceptibilityin the african malaria vector Anopheles gambiae.Ann. Soc. Belge Med. Trop. 71(Suppl. l):225-231.
Collins, F H., R. K. Sakai, K. D. Vernick, S. Paske-witz, D. C. Seeley, L. H. Miller, W. E. Coltins, C.C. Campbell and R. W. Gwadz. 1986. Genetic se-lection of a Plasmodium-refractory strain of the ma-laria vector Anopheles gambiae. Science 234:6O7-610.
Delves, C. J, M. Goman, R. G. Ridley, H. Marile, T,H. W. Lensen, T Ponnudurai and J. G. Scaife. 1989.Identification of P. falciparum infected mosquitoesusing probe containing repetitive DNA. Mol. Bio-chem. Parasitol. 32:lO5-1 12.
Dcibeli, H., A. Tlzeciak, D. Gillessen. H. Matile. I. K.Srivastava, L. H. Perrin, P E. Jakob and U. Certa.1990. Expression, purification, biochemical char-acterization and inhibition of recombinant Plasmo-dium falciparun aldolase. Mol. Biochem. Parasitol.41:259-268.
Gentilini, M. 1991. G6n6ralit6s et Histoire du palu-disme, p. 16. In: M. Danis and J. Mouchet (eds.).Paludisme. Universit6s Francophones, Ellipses/Au-pelf, Paris.
Killick-Kendrick, R. 1974. Parasitic protozoa of theblood of rodents. II. Haemogregarines, malaria par-asites and piroplasms of rodents: an annotatedchecklist and host index. Acta Trop. (Separatum) 3 1 :28.
Mak]er, M. T, and D. J. Hinrichs. 1993. Measurementofthe lactate dehydrogenase activity of Plasmodiumfalciparum as an assessment of parasitemia. Am. J.Trop. Med. Hyg. 48:2O5-21O.
Ponglikitmongkol, M., D. Nicomrat, V. Boonsaeng, PWilairat and S. Panyim. 1994. Field evaluation ofsimplified radioactive and nonradioactive DNAprobe methods for the diagnosis of falciparum ma-laria. Southeast Asian J. Tiop. Med. Public Health25:43O-435.
Riandey, M.-E 1993. Le Systdme des est6rases car-boxyliques chez Anopheles stephensi. C. R. Acad.Sci . Par is Sci . Vie 316:1183-1187.
Rosenberg, R., R. A. Wirtz, I. Schneider and R. Burge.1990. An estimation of the number of malaria spo-rozoites ejected by feeding mosquito. Trans. R. Soc.Tiop. Med. Hyg. 84:2O9-212.
Second, G. and P Trouslot. 1980. Electrophorbsed'enzymes de riz. No. 120, Tfavaux et Documentsde I'ORSTOM, Bondy, France.
Selande6 R. K., M. H. Smith, S. Y. Yang, W. E. John-son and J. B. Gentry. 1971. Biochemical polymor-phism and systematic in the genus Peromyscus. l-variation in the old field mouse (Peromyscus po-lionotus). Stud. Gene. 6:49-9O.
Sherman, I. W. 1961. Molecular heterogeneity of lac-tic dehydrogenase in avian malaria (Plasmodium lo-phurae). J. Exp. Med. 114:1O49-1O62.
Sherman, l. W. 1962. Heterogeneiry of lactic dehy-drogenase in intra-erythrocytic parasites. Trans. N.Y. Acad. Sci. 24(Ser. 2\:944-953.
Vernick, K. D. and E H. Collins. 1989. Associationof a Plasmodium-refractory phenotype with an es-terase focus in Anopheles gambiae. Am. J. Tfop.Med. Hyg. 4O:593-597.
Vial, H. J. and M. L. Ancelin. 1992. Malarial lipids.An overview. Subcell. Biochem. 18:259-3O6.
Walliker, D., R. Carter and S. Morgan. 1971. Geneticrecombination in malaria parasites. Nature (Lond.)232:561-562.
Walliker, D., R. Carter and S. Morgan. 1973. Geneticrecombination in P lasmodium be rghei. Parasitology66:3O9-32O.
Walliker, D., R. Carter and A. Sanderson. 1975. Ge-netic studies on Plasmodium chabaudi: recombina-tion between enzyme markers. Parasitology 70:19-n /
Williams, C. B. 1973. The use of logarithms in theinterpretation of certain entomological problems.Ann. Appl. Biol. 24:4M-414.
Wirtz, R. A., T R. Burkot, R. G. Andre, R. Rosenberg,W. E. Collins and D. R. Roberts. 1985. Identifica-tion of Plasmodium vivax sporozoites in mosquitoesusing an enzyme-linked immunosorbent assay. Am.J. Trop. Med. Hyg. 34:1048-1054.
World Health Organization. 1992. Strat6gie mondialede lutte anti-paludique. Conf6rence ministdrielle surle paludisme, Amsterdam, Netherlands.
Yamaoka, S. and T, Kameya. 1972. Studies on senrmlactate dehydrogenase (S-Ldh) isoenzyme in race-horses especially on relationship between appear-ance of S-Ldh, isoenzyme and hepatic dysfunction.Exp. Rep. Equine Health Lab. 9:55-56.