Embed Size (px)
Citation preview

Lectures in Plant Developmental Lectures in Plant Developmental Physiology, 2 cr.Physiology, 2 cr.
Lectures in Plant Developmental Lectures in Plant Developmental Physiology, 2 cr.Physiology, 2 cr.
Kurt FagerstedtKurt FagerstedtDepartment of Biological and Environmental SciencesDepartment of Biological and Environmental Sciences
Plant BiologyPlant BiologyViikki BiocenterViikki Biocenter
Spring 2006Spring 2006

Embryo development Embryo development Lecture 3Lecture 3
Embryo development Embryo development Lecture 3Lecture 3

Time-table and organisationMon 13.3. Orienteering and Introduction to plant developmental
biology. Cell-intrinsic information. Prof. mvs. Kurt Fagerstedt
Wed 15.3. Embryo development (primary axis development). Prof. mvs. Kurt Fagerstedt
Mon 20.3. Shoot apical meristems. Prof. mvs. Kurt Fagerstedt
Wed 22.3. Leaf development, stomata. Prof. Jaakko Kangasjärvi
Mon 27.3. Root apical meristems, root development. Prof. Ykä Helariutta
Wed 29.3. Flower development. Prof. Teemu Teeri
Mon 3.4. Hormonal control of development, Prof. Ykä Helariutta
Wed 5.4. Developmental responses to light. Prof. Jaakko Kangasjärvi
Mon 10.4. Environmental information other than light. Prof. mvs. Kurt Fagerstedt
Wed 12.4. Coordination of development, Prof. mvs. Kurt Fagerstedt
Mon 17.4. No lecture (Easter)
Wed 19.4. Open examination on the lectures and additional reading.

Primary axis developmentradial axis & longitudinal axis
• axes are polar (mature as well as developing axes).
• the acquisition of polarity has been studied extensively in the seaweed Fucus.
• Fucus produces free-floating eggs which are fertilized by motile sperm. After fertilization the zygote attaches itself to a rock and commences embryogenesis.
• The Fucus egg is spherical and apolar. • The zygote acquires longitudinal polarity
largely in response to environmental cues.

Thallus
Rhizoid
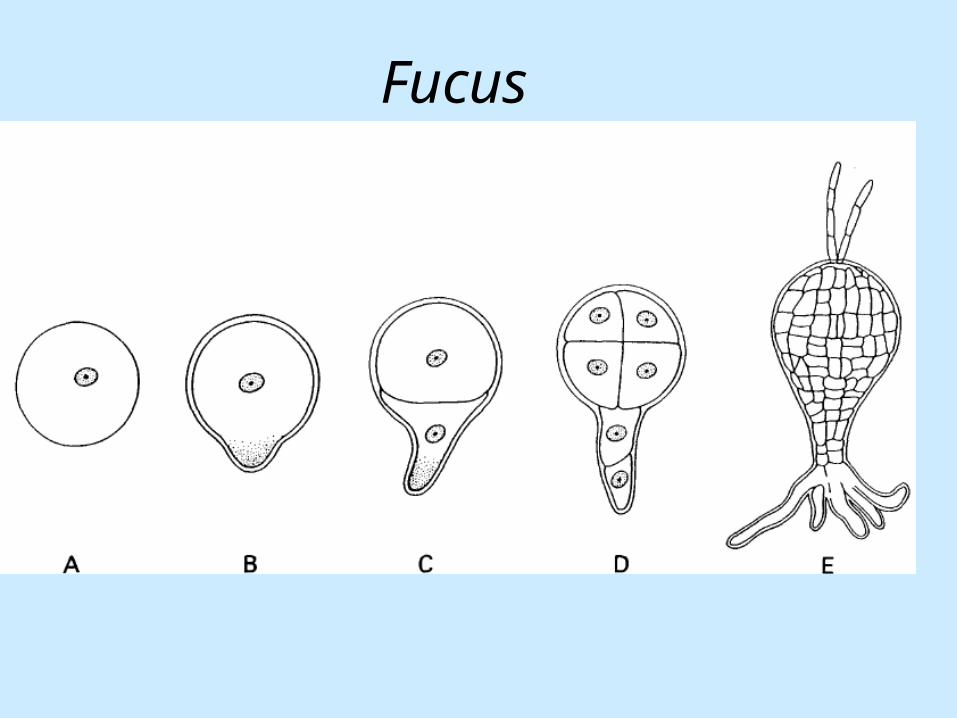
Fucus

Establishing polarity in the zygote
• The Fucus egg has no cell wall and is apolar.
• The first sign of polarity in the zygote occurs within minutes of fertilization as a patch of F-actin accumulates at the site of sperm entry. In the absence of polarized environmental cues, this site will become the rhizoid pole of the zygote. Usually the longitudinal axis is oriented relative to external information.

Establishing polarity in the zygote
• Environmental cues affecting polarity are directional light, gravity, water currents, and temperature gradients.
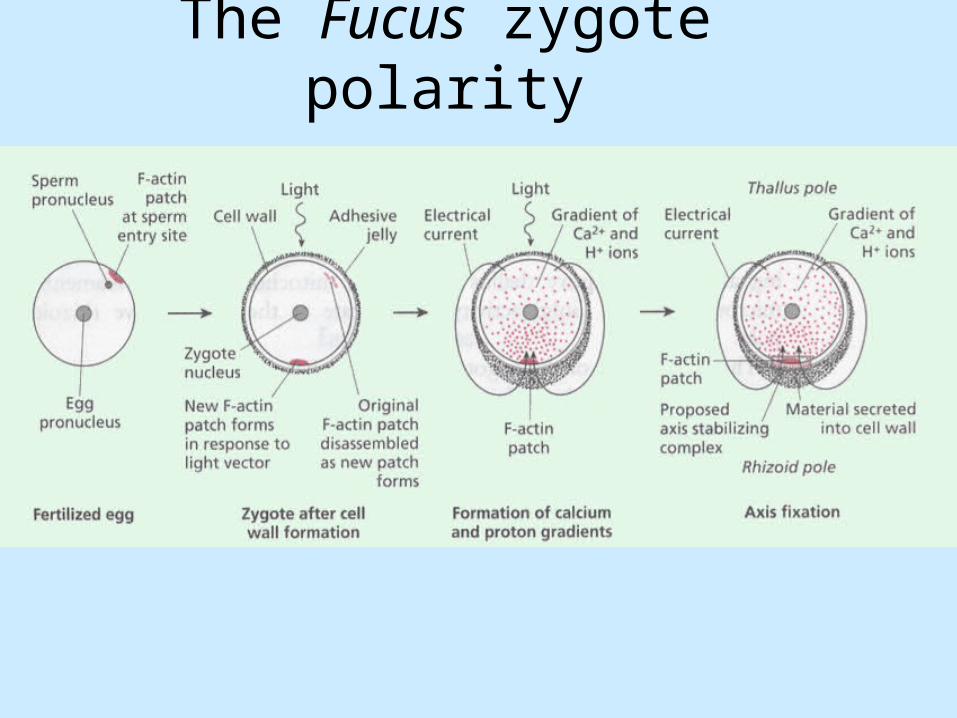
The Fucus zygote polarity

Establishing polarity in the zygote - the mechanism ?
• If environmentally determined axis is oriented differently from the axis defined by site of sperm entry, the F-acting patch marking the sperm entry is disassembeld and a new F-actin patch accumulates at the new rhizoid pole.
• Electric current that flows out of the zygote at the thallus pole and into the zygote at the rhizoid pole can be detected. Current involves movement of calcium and / or hydrogen ions through asymmetrically distributed pumps and channels in the zygote plasma membrane.
• Asymmetric distribution is directed by the polarized distribution of F-actin i.e. cytoskeleton is in a central role.

The role of cell wall in maintaining longitudinal axis
• Axis fixation: Several hours after fertilization, the longitudinal axis becomes fixed and the positions of the future thallus and rhizoid cannot be altered by external cues.
• Axis fixation involves interactions between the cytoplasm and cell wall.
• Axis stabilizing complex, actin filaments and substances in the cell wall.

The role of cell wall in maintaining longitudinal axis
• A simple axis-stabilizing complex might explain how the polarity of the zygote is fixed but more complex positional information is also secreted into the cell wall.
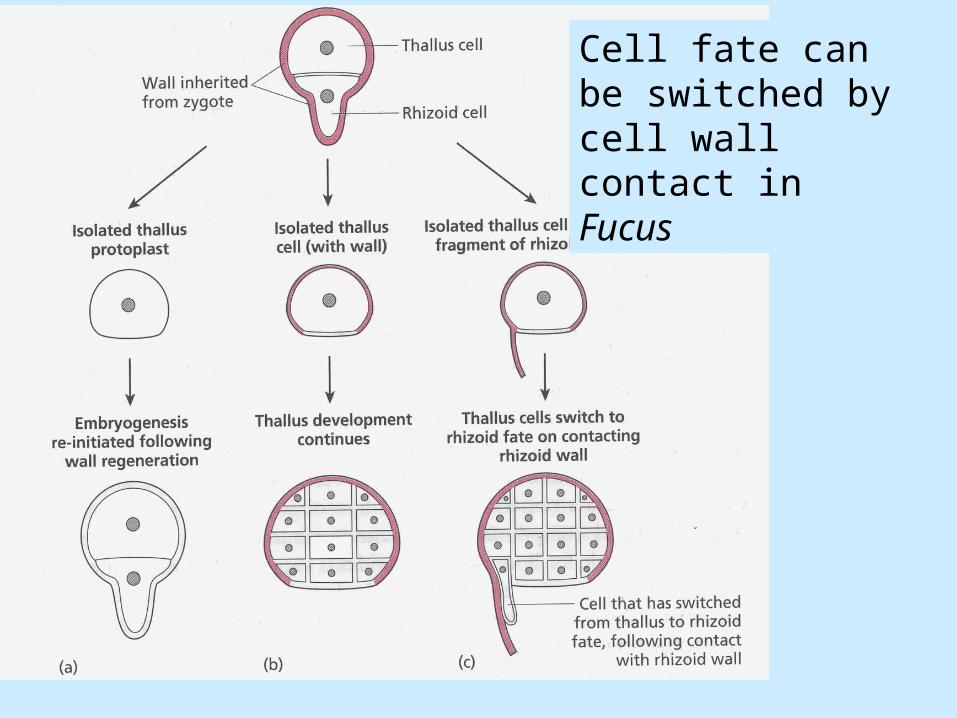
Cell fate can be switched by cell wall contact in Fucus
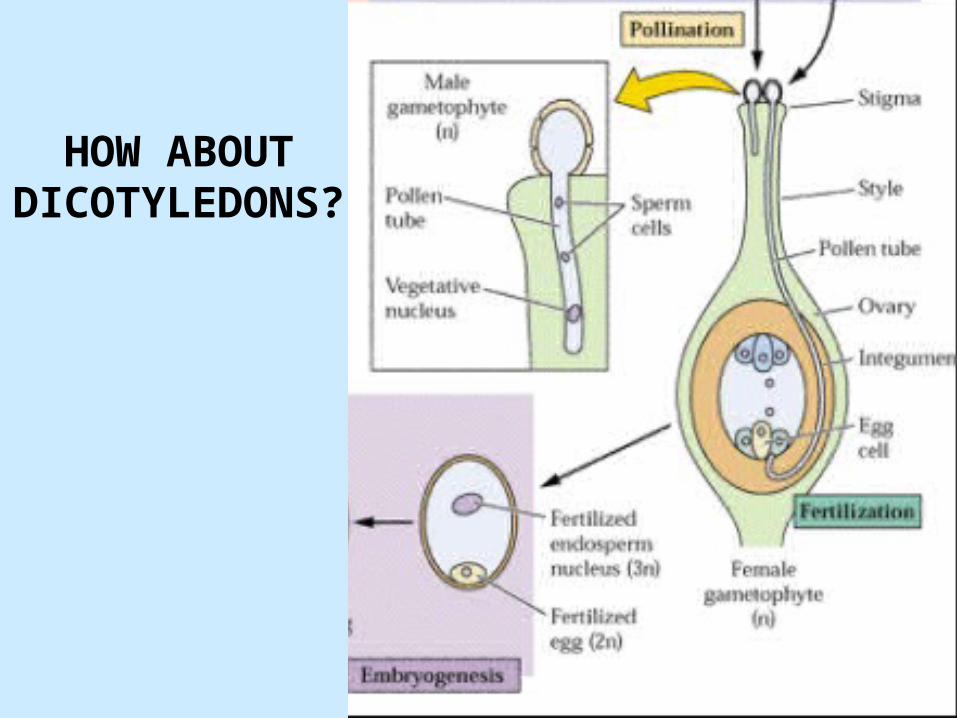
HOW ABOUT DICOTYLEDONS
?
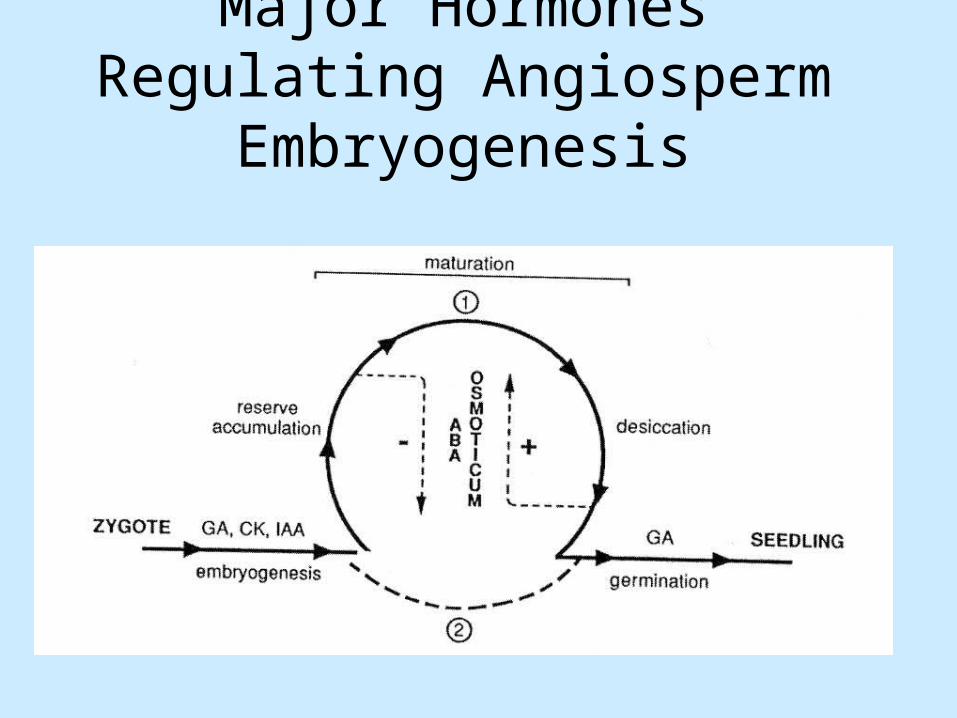
Major Hormones Regulating Angiosperm Embryogenesis

Development of the sporophyte- embryogenesis
Embryogenesis in Arabidopsis thaliana

Pattern formation in Arabidopsis embryo
Thick lines present division lines separating apical (A), central (C) and basal (B) embryo regions.

Arabidopsis embryoasymmetry of the zygotic division is not required to establish the longitudinal axis of the embryo
proper
• AtLTP1 in protoderm, encodes a lipid transfer protein involved in formation of cuticle. Marker of embryo polarity.
• GNOM gene encodes a protein with similarity of yeast proteins involved in secretion.
• gnom-phenotype, GNOM protein is required to direct wall materials to the sites of cell wall deposition. If not directed accurately > abnormal division orientations in gnom embryos.

Arabidopsis embryoThe polarity of the zygote >
longitdinal axis of the embryo

Arabidopsis gnom embryo

Asymmetry of the zygotic division is not required to establish the longitudinal axis of the embryo proper
• Embryo polarity can be expressed despite abnormal division plane > there is a signal or a gradient defining the apical-basal axis that is independent of the cellular architecture of the-embryo.
• initial orientation of the signal or gradient depends on polarity inherited from the egg cells.
• reverse longitudinal axes suggest that after original embryo polarity has been lost, a new longitudinal axis can arise de novo.

Polar auxin transport is a prominent feature of the shoot-to-root axis

Role of polar auxin transport in the embryo
• Auxin transport along longitudinal axis is a universal feature of higher plant embryos.
• From the globular stage onwards, auxin transport can be detected in the shoot-to-root direction and this is in correlation in the distribution of PIN1 (PIN-FORMED1) protein, which is a component of an auxin efflux carrier.
• In early embryos PIN1 has an apolar distribution.
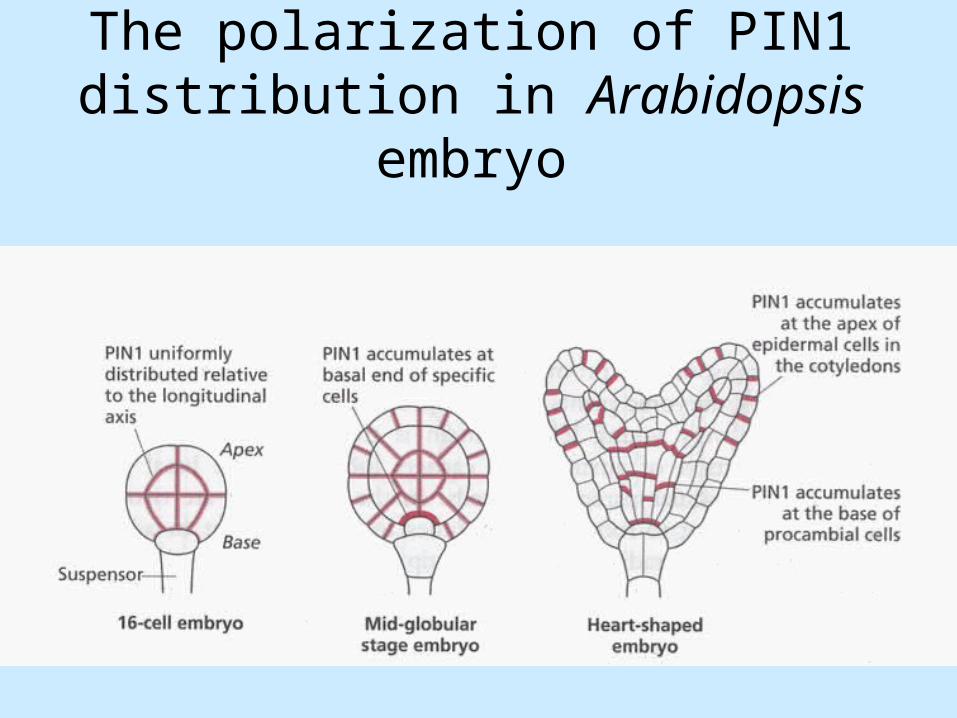
The polarization of PIN1 distribution in Arabidopsis embryo

PIN localisation and auxin transport in A. thaliana embryo

Role of polar auxin transport in the development of embryogenic axes
• PIN1 / longitudinal axis in gnom embryos remains random
• > GNOM is required for the polarization of PIN1
• > gnom embryos have reduced auxin transport ?

PIN1 / longitudinal axis in mp embryos
• The development of mp embryos is abnormal from the two-cell stage onwards. MONOPTEROS (MP) gene encodes an Auxin Response Factor (ARF). ARFs bind to promoters of auxin-inducible genes and regulate their transcription.
• Rate of auxin transport is significantly less than in wild type i.e. it might be that a reduction in auxin transport causes the partial failure of the longitudinal axis in mp embryos.

MP expression and the effects of the mp mutation in Arabidopsis embryos.

RAM & SAM apical meristem formation• Auxin concentration gradient either induces or modulates
the development of RAM= root apical meristem.• ”Auxin maximum” is required for RAM development.• Several genes necessary for shoot apical meristem
(SAM) function have been identified. SHOOT- MERISTEMLESS (STM), WUSCHEL (WUS), CLAVATA1 (CLV1) & CLAVATA3 (CLV3).
• The first indication of of SAM development is the expression of WUS in cells at the apex at the early globular stage. STM is expressed in the apex of late globular embryo. CLV1 and CLV3 are expressed at the site of presumptive SAM in the early heart-shaped embryos.
• SAM becomes histologically distinquishable at the torpedo stage.

Radial axis of A. embryos
• Radial axis becomes apparent later than longitudinal axis
• Radial axis in all parts of the plant is under similar control during both embryonic and post-embryonic development.

Roles of localised auxin transport
BDL=Auxin repressorARF=Auxin Response ElementMP=Auxin response factor

Major regulators of maturation





























