Embed Size (px)
Citation preview

Lectures in Plant Developmental Physiology, 3 cr.Lectures in Plant Developmental Physiology, 3 cr.Lectures in Plant Developmental Physiology, 3 cr.Lectures in Plant Developmental Physiology, 3 cr.
Kurt FagerstedtKurt FagerstedtDepartment of Biological and Environmental SciencesDepartment of Biological and Environmental Sciences
Plant BiologyPlant BiologyViikki Biocenter 3Viikki Biocenter 3

Leaf Development Leaf Development Lecture 5Lecture 5
Leaf Development Leaf Development Lecture 5Lecture 5

Emergence of the leaf Emergence of the leaf primordiaprimordia
Emergence of the leaf Emergence of the leaf primordiaprimordia

Hairy leaf of Coleus
1. adaxial epidermis2. abaxial epidermis3. epidermal cells4. trichomes 5. spongy parenchyma6. intercellular spaces7. palisade
parenchyma

Axis development in the leaf
• leaves are lateral organs.• leaves display consistent orientation and
polarity relative to the shoot i.e. axial information in the leaf does not arise de novo but depends on existing axial information.
• Angiosperm leaf is almost always a determinate organ.

Structural symmetry in the leaf
• simple leaves have three axes of symmetry.
• proximodistal axis from base of the leaf to the tip.
• adaxial-abaxial axis from the upper to the lower epidermis.
• centrolateral axis from the midrib to the margin.

Structural symmetry in the leaf

Adaxial-abaxial axis(dorsoventral axis)
• adaxial-abaxial asymmetry.• Dicot leaf primordium is initiated as a
radially symmetric outgrowth that rapidly acquires adaxial-abaxial asymmetry:– In tobacco P1 (the youngest visible leaf
primordium) is cylindrical whereas P2 has a flattened adaxial surface
• adaxial-abaxial polarity in the leaf depends on the radial axis of the shoot apical meristem.

Symmetry development in the leaf

Adaxial-abaxial polarity
• adaxial-abaxial polarity in the leaf depends on the radial axis of the shoot apical meristem.
• PHANTASTICA (from Antirrhinum)
• PINHEAD• ARGONAUTE1• PHABULOSA• YABBY

PHANTASTICA (from Antirrhinum), encodes a MYB-type transcription factor
• loss-of-function phan mutants develop leaves with variable loss of adaxial-abaxial asymmetry.
• it is expressed in apical meristems at the future sites of leaf initiation and in leaf primordia up until the P3 stage.
• PHAN expression is uniform along the adaxial-abaxial axis.
• PHAN does not itself provide adaxial-abaxial information but it might be that expression is needed by the primordia to be able to respond to the polarizing signal produced by the apical meristem.

Wild type Antirrhinum Radially symmetric phan leaf

PINHEAD& ARGONAUTE1
• PNH & AGO1 are needed for the development of adaxial leaf tissue.
• encode proteins with similarity to eukaryotic translation initiation factors but the biochemical functions are unknown.
• AGO1 is expressed ubiquitously in plants but PNH relates to the adaxial-abaxial axis.


PHABULOSA(transcription factor with homeodomain,
leucin zipper, sterol/lipid-binding domains)
• PHAB gene is also believed to act in promotion of adaxial leaf fates.
• In wild type plants PHAB is expressed uniformely across I1 (future leaf primordia, incipients) but becomes restricted to the adaxial region of the leaf by P2.

YABBY (transcription factors)
• YABBY gene family is required for the development of abaxial leaf tissue in Arabidopsis:– FILAMENTOUS FLOWER (FIL)– YABBY2 (YAB2)– YABBY3 (YAB3)
• Uniform expression begins at I2 in subepidermal cells but at P1 expression becomes restricted to the abaxial side. Expression disappears in the mature leaf.
• Signal from apical meristem promoting adaxial leaf fate inhibits directly or indirectly YABBY gene family expression in adaxial tissues.
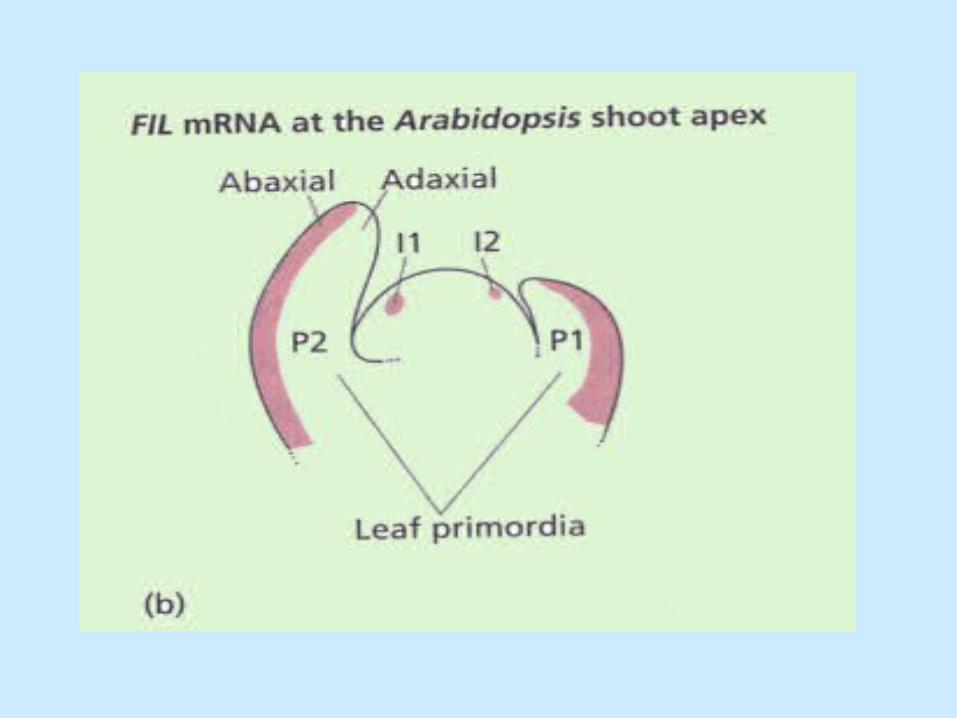

Maintenance of adaxial-abaxial axis
• The mechanims maintaining the axis are probably intrinsic to the leaf but little is known about this.
• LAM1 (DNA sequence?) is probably needed.
• In lam1 leaf primordia are indistinguishable from wild types. However, adaxial cell types are replaced by abaxial ones (and lamina fails to grow along the centrolateral axis).
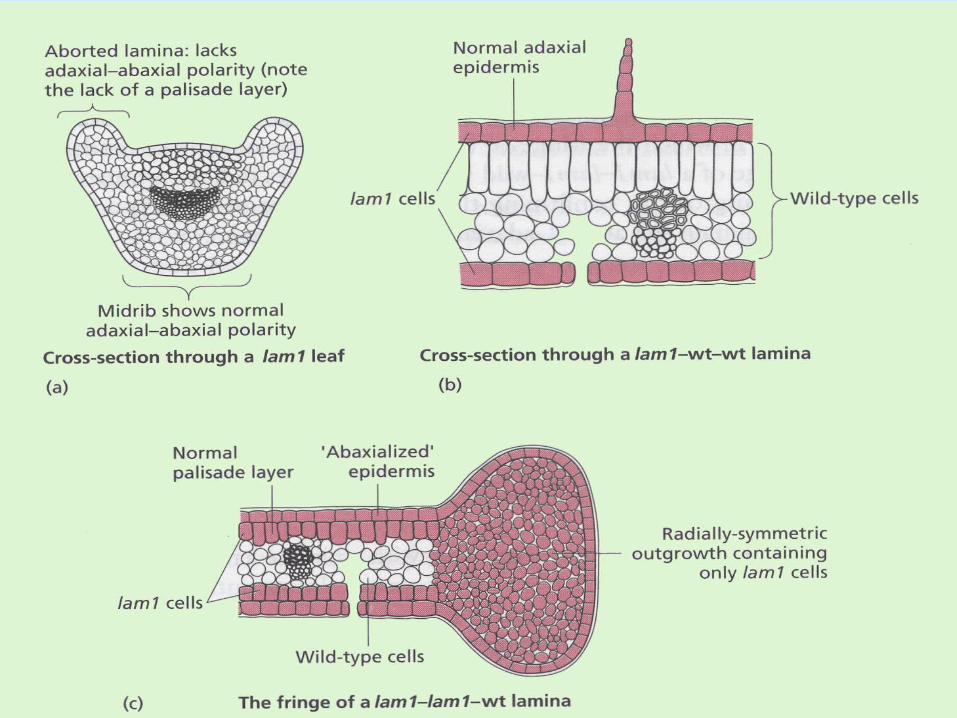

Centrolateral axis
• In dicots, the transition from a radially symmetric P1 leaf primordium to a flattened P2 primordium results in bilateral symmetry.
• At this stage centrolateral axis becomes apparent.
• The extension of lamina along the cenrolateral axis requires the juxtaposition of adaxial and abaxial cell types. – phan example

Adaxial leaf tissue and SAM
• Adaxial leaf tissue promotes the formation of axillary meristems and maintains the development of the primary shoot apical meristem.
• In wild type Arabidopsis leaf, an axillary meristem develops from adaxial cells at leaf base.


Proximodistal axis of the leaf
• Proximodistal differences between leaf cells are visible at the P3 stage.
• Leaf matures in a tip-to-base (basipetal) wave.
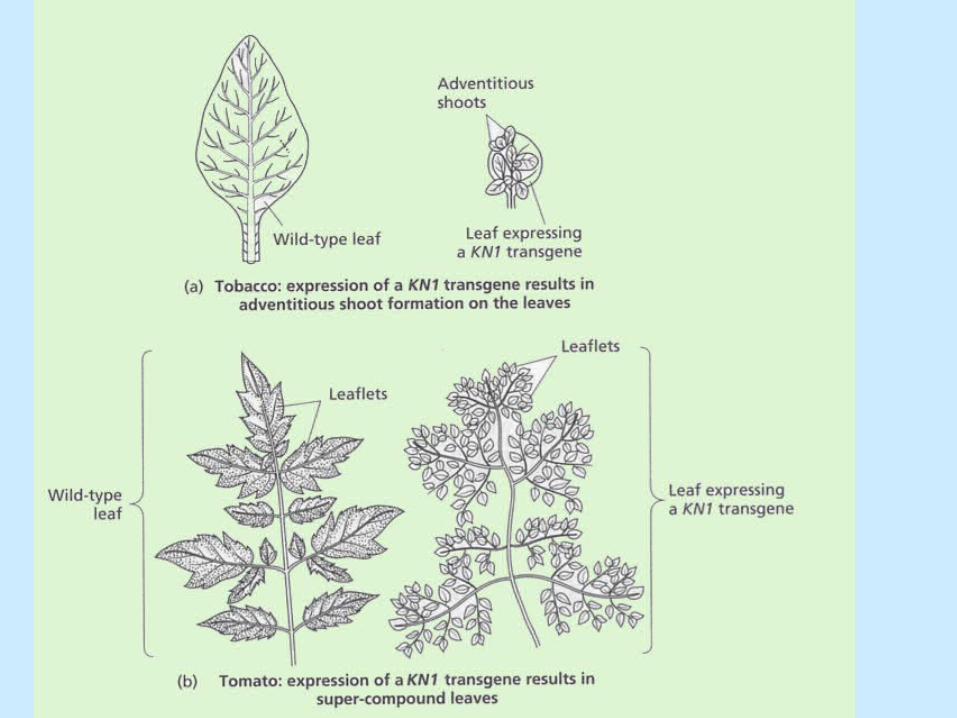
• knotted 1, consequence of gain of function.


Leaf development – determinate
• Loss-of function mutations in STM (SHOOTMERISTEMLESS, KNOX gene) lead to failure of meristem initiation during embryogenesis or premature meristem termination. KNOX genes are required to maintain indeterminate state of the apical meristems.
• High KNOX activity may induce SAMs on the leaf.
• Absence of KNOX activity contributes to the determinate nature of leaf development.
• compound leaves follow a less determinate pattern of development than simple leaves.

Stomatal development
• Epidermis – the interface between plant and the world.
• To maximize photosynthetic efficiency while minimizing water loss, stomatal pore size is modulated by the ion-driven swelling of the quard cells. Optimal gas exchange requires regulation of:– numbers and positions of stomata– the ability to open and close stomata

Cell signaling is critical to establisment of stomatal
pattern
• Stomata are formed through a stereotyped lineage of asymmetric cell divisions.
• Patterned locally so that two stomatal complexes never adjacent to each other = the one-cell-spacing rule.
• Overall numbers of stomatal complexes are controlled in response to environmental cues. (e.g. CO2)

Lineage pattern for quard cell formation in Arabidopsis
Current Opinion in Plant Biology 2004, 7:26-32.

Cell fate and cell signaling
• Protodermal cells distributed throughout the young leaf epidermis enter into the lineage pathway that leads to the formation of stomata.
• Lineage alone is not sufficient to ensure adherence to the one-cell-spacing rule.

Cell fate and cell signaling
• The major factors in determining the pattern of stomata are the signals from mature quard cells (or their precursors GMCs or meristemoids) to their neighboring cells.– cells that are in contact with a single stoma
are instructed to orient their future division planes such that divisions place the smaller cell distal to the pre-existing stoma.
– cells that are in contact with two or more stomata are instructed not to divide.
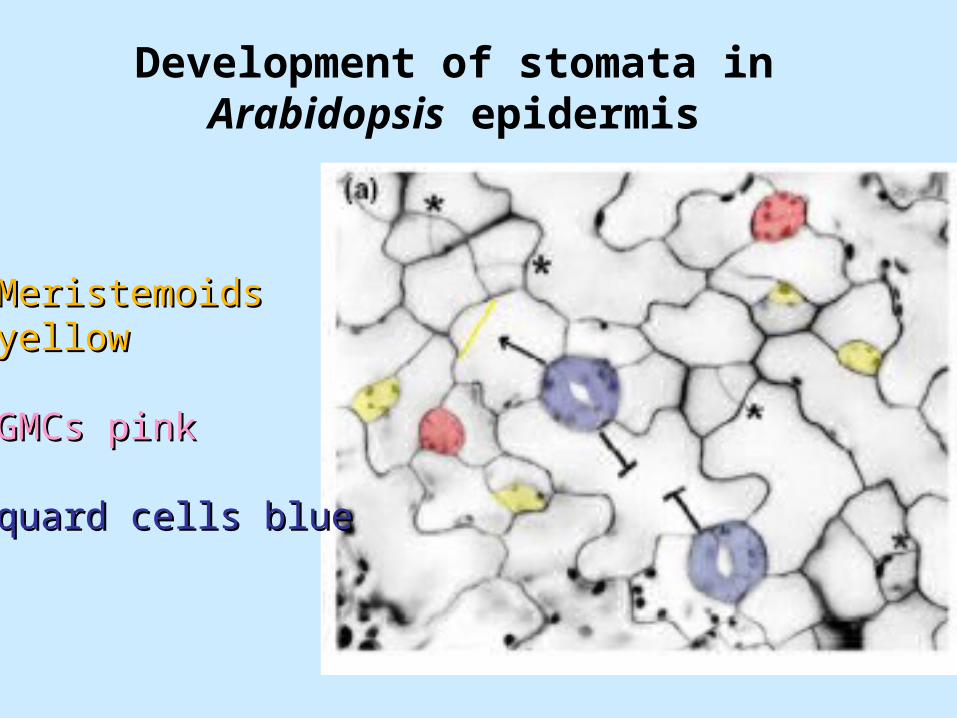
Development of stomata in Arabidopsis epidermis
MeristemoidsMeristemoidsyellowyellow
GMCs pinkGMCs pink
quard cells bluequard cells blue

Cell fate and cell signaling
• The gene products required for stomatal patterning are:– the leucine-rich-repeat –receptor-like protein
encoded by TMM (two many mouths).– serine protease encoded by SDD1 (stomatal
density and distribution).
• Mutations in TMM or SDD1 lead to an increase in stomatal index and a breakdown of the one-cell-spacing rule.
• TMM serves as a receptor for a signal generated by SDD1.

Hormonal control of stomata
• Application of GA in combination with auxin or ethylene > overproduction of stomata
• GA inhibitor > stomata were eliminated in hypocotyl but not in leaves – hypocotyl and leaves regulate
differently cell identity

Signals that direct stomatal pattern

Growth and cell size control in plants
• Cell-size increase in plants is driven by two very distinct processes:– cell growth involving an
increase in total cytoplasmic macromolecular mass.
– cell expansion involving increased cell volume through vacuolation.

Cell growth and cellexpansion

Cell growth and cellexpansion

Model ofsome of thekey processesthat regulatecell size.
cell size is independent of the cell number.

Leaf size
• The final size and shape of a leaf depends on the position of the leaf on the shoot and on environmental conditions.
• How a developing leaf can regulate its absolute size?– axis specific mechanisms?

Leaf size / competitionchimaeric Pelargonium
• Leaf cells compete to contribute to the leaf
Cell intrinsic information
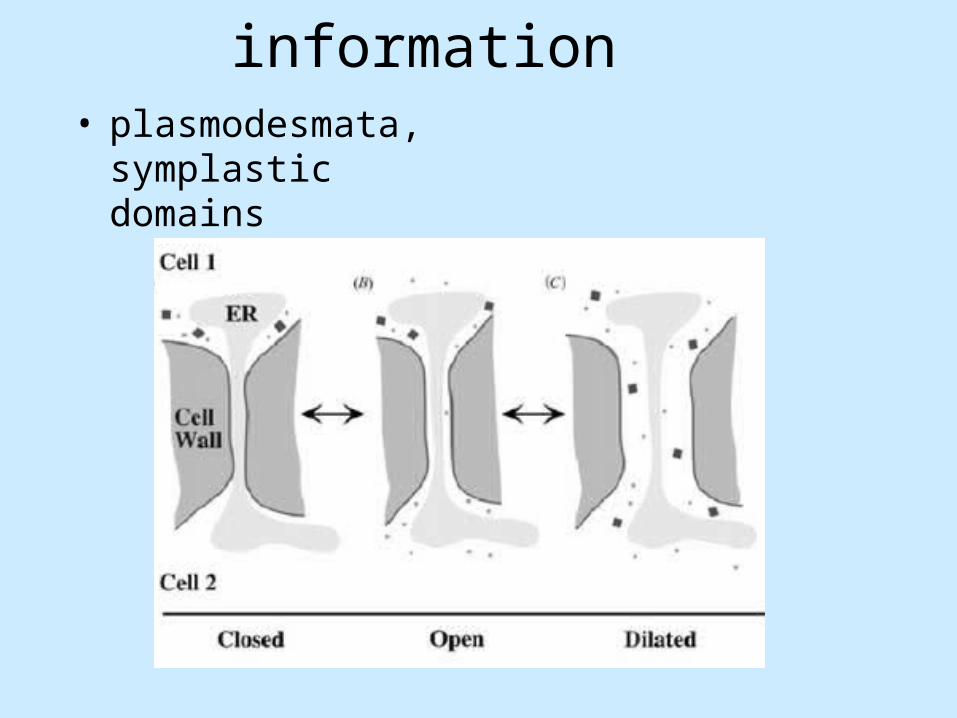
• plasmodesmata, symplastic domains






























