Embed Size (px)
DESCRIPTION
Metabolizam i procesi metabolizma
Citation preview

Poljoprivredno- prehrambeni fakultet
Univerzitet u Sarajevu
METABOLIZAM MIKROORGANIZAMA I
Sarajevo, februar 2013. god

METABOLIZAM MIKROORGANIZAMA
Metabolizam je skup hemijskih reakcija u kojim ćelija proizvodi i izgradjuje veliki broj različitih jedinjenja neophodnih za njezino održavanje.
U osnovi razlikujemo dva procesa metabolizma:
a) Katabolizam- To su reakcije u kojima se proizvodi energija, tj. reakcije u metabolizmu koje uključuju enzimatsku razgradnju organske materije u jednostavnija organska i anorganska jedinjenja pri čemu dolazi do oslobađanja energije.
b) Anabolizam- To su reakcije koje zahtjevaju energiju za sintezu ili izgradnju organskih materija, odnosno to je proces u kojem se energija iskorištava.
U reakcijama katabolizma razgrađuju se šećeri, aminokiseline, masne kiseline i ostala srodna jedinjenja. Na ovaj način dobivene jednostavne molekule mogu učestvovati u reakcijama anabolizma u kojima se sintetiziraju enzimi, strukturni proteini, nukleinske kiseline, ugljikohidrati i ostala biohemijska jedinjenja potrebna organizmu.
Kao ugljikohidratni izvori energije značajni su: skrob i glikogen, različite pentoze i heksoze, te disaharidi (laktoza, saharoza, maltoza). Da bi iskorištavala polisaharide ( skrob i glikogen) ćelije moraju sintetizirati ekstracelularne ( vanćelijske ) enzime radi cijepanja složenih jedinjenja na manje molekule koje mogu ući u ćeliju. Npr. enzim amilaza cijepa skrob na jedinice maltoze, a zatim enzim maltaza cijepa maltozu na dvije jedinice glukoze. Tek nakon toga glukoza će biti upotrijebljena u reakcijama metabolizma.
Biološki katalizatori- ENZIMI
U biološkim organizmima neprekidno se odvija veliki broj hemijskih reakcija koje djeluju na brz porast potomstva i veliko povećanje broja ćelija koje rastu na odgovarajućim hranjivim podlogama u veoma kratkom vremenskom razdoblju. Navedene promjene zahtjevaju brze hemijske reakcije za dobivanje energije i oblikovanja ćelija. Stoga, biološki organizmi zahtjevaju katalizatore koji mogu funkcionirati u uslovima koji su bitni za život.
Biološki katalizatori- enzimi ubrzavaju hemijske reakcije smanjujući energiju aktivacije i pravilno usmjeravajući molekule koje se sudaraju. Tako se, bez povećanja temperature ili pritiska ( tj. bez razaranja ili ubijanja žive ćelije ) u ćeliji odvija veliki broj hemijskih reakcija.
2

Materije koje mogu ubrzati hemijsku reakciju, a da se pri tome same ne mijenjaju,nazivaju se katalizatori. U živim ćelijama enzimi služe kao biološki katalizatori. U optimalnim uslovima enzimi kataliziraju reakcije koje su od 108 do 1010 puta ( više od 10 milijardi puta) brže od reakcija koje se odvijaju bez enzima.
Enzimi su po hemijskoj strukturi protein. Oni sudjeluju u metabolizamskim reakcijama, disanju, u pretvaranju i prenošenju energije između živih organizama i u sintezi različitih molekula i ćelijskih sastojaka. Do sada je opisano oko 2000 različitih enzima, pri čemu svaki od njih može katalizirati specifičnu reakciju.
Struktura ( građa) enzima
Mnogi enzimi su građeni samo od proteina i nazivaju se jednostavni enzimi. Za razliku od njih, enzimi koji sadrže i neproteinske komponente nazivaju se konjugirani ( spojeni ) enzimi. Proteinski dio se naziva apoenzim, a ne proteinski dio koenzim (kofaktor). Zajedno, apoenzim i koenzim stvaraju enzim koji se naziva holoenzim.
Kada je koenzim ( kofaktor) metalni jon ( npr. cink, željezo, bakar ili mangan) onda je riječ o aktivatoru. Mnogi koenzimi potiču od vitamina iz hrane. Bolesti koje su posljedica nedostatka vitamina često su povezane sa nedostatkom sposobnosti određenog enzima da funkcioniše pri normalnim metabolizamskim aktivnostima.
Slika 1. Sastavni dijelovi holoenzima. Mnogi enzimi zahtijevaju i apoenzim ( proteinski dio ) i koenzim ( kofaktor), neproteinski dio radi započivanja svoga djelovanja. Kofaktor može biti jon metala, ili neka organska molekula što se naziva koenzim( kao što je na slici prikazano ). Apoenzim i kofaktor zajedno tvore holoenzim ili potpuni enzim. Supstrat je reactant što djeluje na enzim.
3

Brzina enzimatske reakcije zavisi od pH vrijednosti i temperature. Navedeni parametric utiču ne samo na privlačne sile između pojedinih aminokiselinskih ostataka u molekulu enzima, nego in a reakciju između enzima i materija ( supstrata) na koje enzim djeluje i koje se djelovanjem enzima hemijski mijenja.
Budući da različita denaturirajuća hemijska jedinjenja ( kiseline, radijacija i ektremne temperature) inhibiraju ili razaraju enzyme, navedeni uslovi se mogu iskoristiti za suzbijanje rasta mikroorganizama.
Nakon katalizacije određenih procesa enzimi ostaju nepromjenjeni. Neki enzimi su specifični pa kataliziraju samo određenu reakciju.
Nomenklatura enzima
Enzim dobija ime prema meteriji ( supstratu ) na koju djeluje. Enzimi se također mogu imenovati na osnovu njihovog zajedničkog djelovanja. Tako npr. enzimi klasificirani kao dehidrogenaze prenose vodonik sa supstrata na koenzim, a one koji prenose ugljen dioksid nazivamo dekarboksilaze. Nastavak –aza na kraju označava ime svih enzima.
Hemijske reakcije koje se odvijaju u živoj ćeliji, a u kojima se povezuju jednostavna jedinjenja u složenije molekule, zajednički se nazivaju anabolizam. Anabolitički procesi često obuhvataju dehidratacijske reakcije sinteze ( u kojima se oslobađa H2O) i zahtjevaju energiju za stvaranje novih hemijskih veza. Npr. nastajanje protein iz aminokiselina, nukleinskih kiselina iz nukleotida i polisaharida iz prostih šećera. U ovim reakcijama biosinteze proizvode se materije neophodne za rast ćelije.
Hemijske reakcije u kojim se cijepaju složene organske molekule u jednostavnije nazivaju se katabolizam. Katabolitičke reakcije su najčešće hidrolitičke ( reakcije u kojima se koristi molekul H2O za cijepanje hemijskih veza). U navedenim reakcijama se oslobađa energija koja se pohranjuje u hemijske veze. Primjer katabolizma je kada ćelija cijepa nepotrebne protein u pojedinačne aminokiseline za dalju upotrebu.
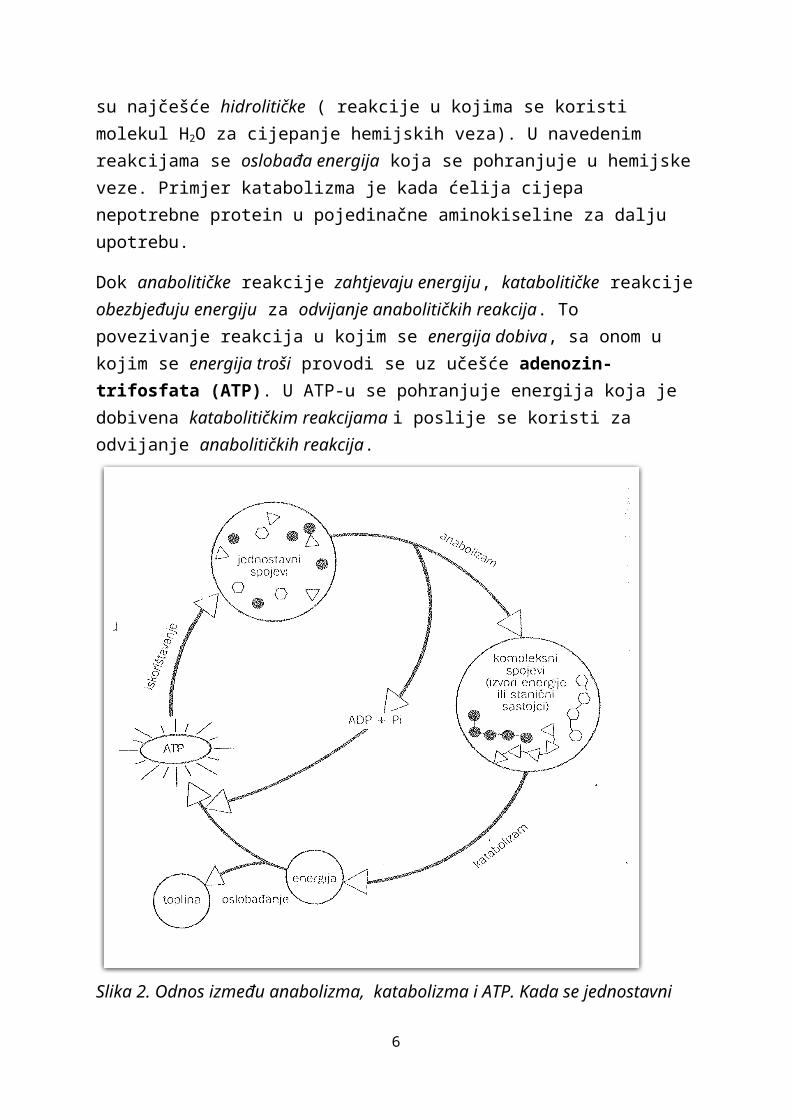
Dok anabolitičke reakcije zahtjevaju energiju, katabolitičke reakcije obezbjeđuju energiju za odvijanje anabolitičkih reakcija. To povezivanje reakcija u kojim se energija dobiva, sa onom u kojim se energija troši provodi se uz učešće adenozin-trifosfata (ATP). U ATP-u se pohranjuje energija koja je dobivena katabolitičkim reakcijama i poslije se koristi za odvijanje anabolitičkih reakcija.
4

Slika 2. Odnos između anabolizma, katabolizma i ATP. Kada se jednostavni spojevi povezuju u složene spojeve ( anabolizam), potrebnu energiju z ate sinteze osigurava ATP. Kada se složeni slojevi neovisno cijepaju( katabolizam), oslobađa se kao toplina velika količina energije. Značajna se količina energije upotrijebi za tvorbu ATP, a potom za odvijanje reakcija anabolizma.
5
ENERGETSKI METABOLIZAM
( obezbjeđivanje energije za živu ćeliju )
Živa ćelija energiju dobiva iz hranjivih materija u nizu hemijskih reakcija, u kojima se energija pohranjena u hemijskim vezama prenosi na ostale nove oblike hemijskih veza. Veliki broj tih reakcija čine oksidacijske reakcije. Tokom oksidacijskih reakcija oslobađa se energija i tada mogu nastati visokoenergetske veze, poput onih u ATP-u, radi uskladištenja oslobođene energije.
Rekacije oksidacije i redukcije- Kada god atom, jon ili molekul izgube jedan ili više elektrona u nekoj reakciji, proces se naziva oksidacija, a za materije koje učestvuju u tom procesu kaže se das u oksidirane. Elektroni koji se odvijaju od oksidirane materije ne putuju bez cilja. Oni su reaktivni i odmah se vežu na drugu molekulu. Prihvatanje jednog ili više elektrona naziva se redukcija, a za materiju koja je prihvatila elektron kaže se da je reducirana. Prema tome u oksidacijsko- redukcijskim reakcijama elektroni se prenose sa jedne molekule na drugu. U mnogim procesima oksidacije istovremeno se uklanjaju dva elektrona i dva protona, tj. dva atoma ( H+). Stoga se biološke oksidacije opisuju i kao dehidrogenacije. Kao tipičan primjer može se navesti oksidacija glicer-aldehid-3-fosfata u 1,3- difosfoglicerinsku kiselinu.
U biološkim organizmima reakcije oksidacije i redukcije uvijek su udružene ( istovremene ) i opisuju se kao oksidacijsko-redukcijske reakcije. Ta povezanost reakcija može se uočiti na primjeru EMP puta- pri oksidaciji glicer-aldehid-3-fosfata uz istovremenu redukciju koenzima nikotinamid-adenin-dinukleotida (NAD+).Koenzimi NAD, zajedno sa NADP služe u biološkim sistemima za prenošenje elektrona i vodikovih atoma. Npr. pri oksidaciji gliceraldehid-3-fosfata koenzim NAD prima dva eletrona i vodikov jon za stvaranje redukovanog oblika NADH. Preostali vodikov jon se otpušta kao Slobodan H+ jon u supstrat.
Proizvodnja energije kod mikroorganizama
Akumuliranje i oslobađanje energije u upotrebljivim oblicima u procesu katabolizma često se naziva respiracija i fermentacija. To su dva velika mehanizma za nastajanje energije oksidacijom hemijskih jedinjenja. Uopšteno, respiracija se opisuje kao oksidativni put koji proizvodi energija ili kao putevi u kojim anorganska jedinjenja služe kao primaoci elektrona ( elektron akceptor) u seriji reakcija.
Aerobna respiracija odnosi se na takve puteve u kojim je kiseonik glavni akceptor elektrona pa najčešće nastaje voda.
U oksidativnim putevima anaerobne respiracije koriste se drugi anorganski elektron akceptori kao npr. nitrati, karbonati i sulfati.
6

Fermentacija ili vrenje se odvija pri takvim oksidativnim putevima u kojim organska jedinjenja služe i kao elektron donor i kao elektron akceptor. Ova oksidacija se odvija u odsustvu ma kojih vanjskih elektrona akceptora. Mnogobrojni proizvodi koji nastaju tokom fermentacije imaju praktičnu primjenu, a to su : vinsko sirće, pivo, vino, jogurt.
Struktura i funkcija adenozin-trofosfata (ATP)
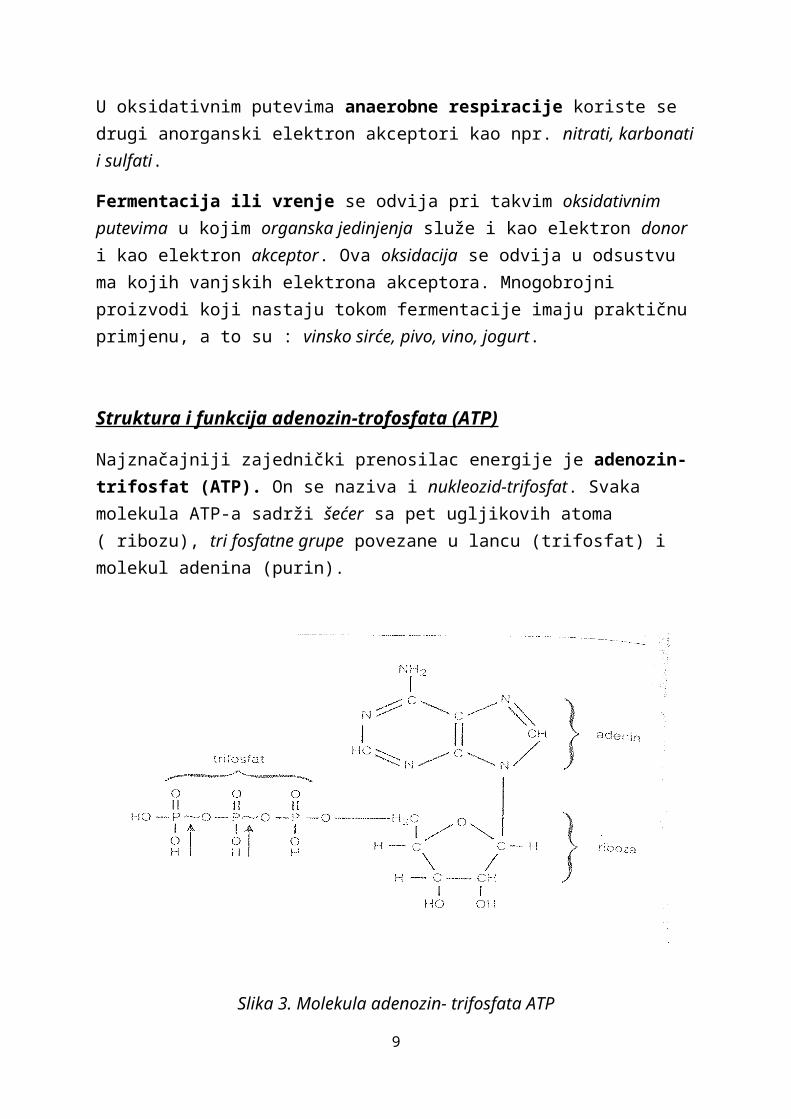
Najznačajniji zajednički prenosilac energije je adenozin-trifosfat (ATP). On se naziva i nukleozid-trifosfat. Svaka molekula ATP-a sadrži šećer sa pet ugljikovih atoma ( ribozu), tri fosfatne grupe povezane u lancu (trifosfat) i molekul adenina (purin).
Slika 3. Molekula adenozin- trifosfata ATP
7

Ciklus ATP ↔ ADP glavni je put u biološkim sustavima u kojim se energija transformiše iz jednog oblika u drugi.
Slika 4. Glavni put izmjene u biološkim sistemima
ATP je poznat kao energetski bogat molekul, jer ima energetski bogate veze između prve i druge, te između druge i treće fosfatne grupe. Ove veze omogućavaju ATP da ima puno veću količinu energije, nego druga jedinjenja. Energetski bogate fosfatne veze obično se označavaju simbolom ( ~), a obične hemijske veze samo crticom ( - ).
Najčešće se vrši prenošenje krajnje ili terminalne fosfatne grupe sa ATP-a ka drugoj molekuli. Sam proces prenošenja fosfatne grupe naziva se fosforilacija, a enzim koji učestvuje tom procesu naziva se kinaza. U reakcijama fosforilacije određena energija fosfatne grupe sa molekulu ATP-a biva prenesena na fosforilirano jedinjenje koje u tome novom energetskom stanju može učestvovati u kasnijim reakcijama.
Fosforilacija- nastajanje i stvaranje ATP
U procesu nastajanja ATP učestvuje ADP i anorganski fosfor. Za odvijanje ove reakcije potrebno je 30-34kJ ( 7-8 kcal) da bi nastala jedna molekula ATP. U toku metabolizma ATP nastaje putem tri fundamentalna biohemijska mehanizma:
a) fosforilacijom na nivou supstratab) oksidativnom fosforilacijomc) fotofosforilacijom
8
a) Fosforilacija na nivou supstrata- Pri ovoj reakciji energija koja se oslobađa oksidacijom supstrata iskorištava se za fosforilaciju ADP i nastajanje ATP. Ovo nastajanje molekule ATP odvija se u citoplazmi prokariota i eukariota.
b) Oksidativna fosforilacija- U oksidativnoj fosforilaciji materija koja se oksidira naziva se elektron- donor, dok se supstrat koji je elektron primio naziva elektron akceptor, a niz elektrona se naziva elektron- transportni lanac. Ovaj lanac je uključen u oksidaciju organskih jedinjenja, a sastoji se od niza prenosnika elektrona i koenzima koji prenose elektrone od jednog do drugog jedinjenja.
Elektron- transportni lanac predstavlja zajednički put za iskorištavanje elektrona tokom različitih metabolizamskih reakcija. U prvom redu učestvuju u aerobnom metabolizmu, međutim neki dijelovi mogu isto reagovati i u anaerobnim organizmima. Ti organizmi ne koriste kiseonik kao akceptor, nego neke druge anorganske akceptore elektrona. Takvi procesi slični su respiraciji, a pošto se odvijaju bez kiseonika nazivaju se anaerobna respiracija.
c) Fotofosfotilacija- Treći mehanizam fosforilacije odvija se u fotosintetskim ćelijama. Svjetlosnu energiju apsorbuju pigmenti ( hlorofil kod fotoautotrofa) pri čemu se oslobađa elektron koji se potom prenosi preko serije elektrona- akceptora. Pri svakom prenošenju se oslobađa energija za nastajanje ATP-a. U cikličnoj fosforilaciji elektroni se oslobađaju iz molekula hlorofila pomoću svjetlosne energije i ponovo se vraćaju u molekul hlorofila nakon prolaska kroz elektrontransporni lanac. Energija dobivena prelaskom elektrona pretvara se u ATP.
Slika 5. U cikličnoj fosforilaciji elektroni se oslobađaju iz molekule hlorofila uz pomoć svjetlosne energije i vraćaju u molekulu hlorofila nakon prolaska kroz elektrontransportni lanac. Energija dobivena prelaskom elektrona pretvara se u ATP.
9

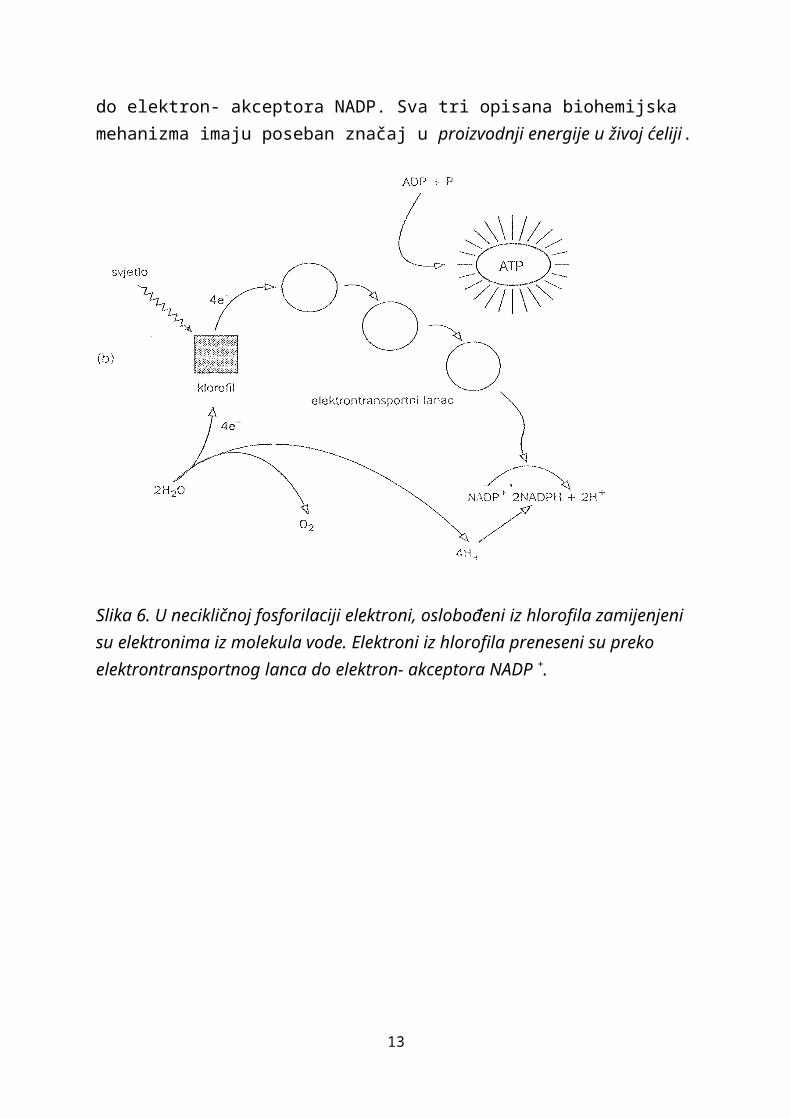
U necikličnoj fosforilaciji elektroni oslobođeni iz hlorofila zamjenjuju se elektronima iz molekula ovde. Dok se elektroni iz hlorofila prenose preko transportnog lanca do elektron- akceptora NADP. Sva tri opisana biohemijska mehanizma imaju poseban značaj u proizvodnji energije u živoj ćeliji.
Slika 6. U necikličnoj fosforilaciji elektroni, oslobođeni iz hlorofila zamijenjeni su elektronima iz molekula vode. Elektroni iz hlorofila preneseni su preko elektrontransportnog lanca do elektron- akceptora NADP +.
10

FOTOSINTEZA- Fiksiranje ili fotosintetsko vezivanje ugljik-dioksida
Fotosinteza je jedan od najstarijih procesa života i ona je odgovorna za nastajanje svih organskih molekula što ih iskorištavaju heterotrofi za obavljanje svojih različitih aktivnosti. Stalnost života na zemlji zavisi o ponovnom vraćanju ugljika u tom procesu, a i o proizvodnji O2 za aerobnu respiraciju. U ovom kružnom toku učestvuju cijanobakterije, alge i zelene biljke. Svi mikroorganizmi koji vrše proces fotosinteze posjeduju zeleni pigment hlorofil, a drugi imaju i dopunske različito obojene pigmente. Kod eukariotskih mikroorganizama hlorofil i dopunski pigmenti su smješteni u posebnim organelama – hloroplastima, a kod prokariotskih mikroorganizama pigmenti se nalaze u tvorevinama koje se nazivaju hromatofore. Alge i cijanobakterije sadrže hlorofil a i hlorofil b. Oni se razlikuju po sposobnosti apsorpcije svjetlosti različite talasne dužine. Hlorofil a najbolje apsorbuje svjetlost talasne dužine 665 nm, a hlorofil b 645 nm. Oni pored hlorofila posjeduju i dopunske pigmente koji se razlikuju od hlorofila po tome što apsorbuju svjetlost dugih talasnih dužina. Najčešći dopunski pigmenti su : karotinoidi( pigmenti žute boje prisutni u ćelijama mnogih algi ) , fikoeritrin ( pigment crvene boje i nalazi se u ćelijama crvenih algi) i fikocijanin ( pigment plave boje i prisutan je u ćelijama cijanobakterija).
Ukupna reakcija fotosinteze, može se prikazati izrazom :
6CO2 + 12H2O → C6H12O6 + 6O2 + 6H2O
ugljendioksid voda enzimi glukoza kiseonik voda
Fotosinteza je složen proces i obuhvata tri različita tipa hemijskih reakcija:
1) Reakcije na svijetlu ; 2) Reakcije u mraku ;3) Regeneracija fotosintetskog pigmenta
U prvom fotosintetskom procesu sunčevo je svjetlo upotrijebljeno za izbacivanje elektrona iz molekula donora ( hlorofil ), što dovodi do nastajanja ATP. Reakcije na svjetlu obuhvataju fotosintetske membrane i odvijaju se samo u prisutnosti svjetla. To su serije reakcija kataliziranih enzimima koji koriste novostvorene ATP i NADPH za nastajanje organskih molekula tokom vezivanja atmosferskog CO2.
Vezivanje ugljika u reakcijama fotosinteze u mraku zavisi od prisutnosti molekula na koje se CO2 može vezati. Glavno jedinjenje u tom ciklusu je ribuloza-1,5-difosfat.
11

Putevi u kojima se proizvodi energija
Prvi korak u katabolizmu organskih molekula je razgradnja složenih molekula u jednostavnija jedinjenja ( depolimerizacija). Tako npr. skrob se razgrađuje na jednostavnije molekule šećera kao što su glukoza, proteini na aminokiseline, masti na masne kiseline i glicerol, a nukleinske kiseline u nukleotide. Iako se pri ovim procesima ne proizvodi značajnija upotrebljiva energija, ovim putem se osigurava veliki broj malih molekula koji čine početni materijal za metabolizam. Tokom oksidacije ovih organskih molekula, organizam proizvodi energiju aerobnom respiracijom, anaerobnom respiracijom i vrenjem. Iako se veliki dio ove energije gubi u obliku toplotne energije, ćelije ostatak energije koriste u obliku ATP za obavljanje brojnih aktivnosti.
Katabolizam ugljikohidrata
Većina mikroorganizama da bi obezbjedila neophodnu ćelijsku energiju, oksidira ugljikohidrate. U ćelijskom metabolizmu izuzetno važno cijepanje molekula ugljenih hidrata ( katabolizam ugljikohidrata) radi stvaranja neophodne energije.U ćeliji se najčešće katabolizira glukoza radi dobivanja neophodne energije. Da bi obezbjedili neophodnu energiju mnogi mikroorganizmi također kataboliziraju i različite lipide i proteine.
Neophodnu energiju iz glukoze mikroorganizmi dobijaju na dva načina:
1) respiracijom (disanjem)2) vrenjem (fermentacijom )
U oba navedena procesa elektroni, proizvedeni oksidacijom glukoze, bivaju preneseni na NAD+. Krajnja sudbina elektrona zavisna je o procesima razgradnje glukoze.
GLIKOLIZA- Mikrobiološka razgradnja glukoze
Razgradnja glukoze se odvija u dvije faze. Prva faza u razgradnji glukoze i kod respiracije i kod vrenja je oksidacija glukoze do piruvata u procesu koji se naziva glikoliza.
Postoji više biohemijskih puteva za razgradnju glukoze do piruvata, a Embden-Majerhof-Parnas-ov (EMP) put je put najpoznatijih serija katabolitičkih reakcija u kojem se glukoza pretvara u 2 molekula piruvata ( pirogrožđane kiseline) kroz 10 reakcija ili 4 stadija i to :
Stadij a- stadij aktivacije glukoze, a vrši se uz pomoć ATP i obuhvata prve tri reakcije (1,2,3)
12
Stadij b- stadij cijepanja glukoze na dvije molekule triozo fosfata i obuhvata 2 reakcije ( 4,5).
Stadij c- je stadij oksidacije i on obuhvata uklanjanje elektrona i prenos na molekule nikotinamid-adenin-dinukleotid NAD, obuhvata jednu reakciju ( 6 ).
Stadij d- stadij regeneracije ATP, to je stadij u kojem se stvara ATP i piruvat i obuhvata 4 reakcije ( 7,8,9,10).
Proces glikolize se odvija u 10 reakcija :
1) U prvoj reakciji glikoloznog puta prisutnog kod svih tipova glikolize se vrši fosforilacija glukoze pomoću enzima heksokinaze i ATP pri čemu nastaje glukoza-6-fosfat.
2) U drugoj reakciji glukoza-6-fosfat izomerizacijom uz pomoć enzima
fosfoglukozoizomeraze prelazi u fruktoza-6-fosfat.3) U tećoj reakciji iz fruktoza-6-fosfata uz učešće ATP, tj. fosforilacije nastaje
fruktoza-1,6-difosfat.4) i 5) U četvroj i petoj reakciji fruktoza -1,6-difosfat se pod uticajem enzima
aldolaze razlaže na gliceraldehid-3-fosfat i dihidroksiaceton-fosfat.
6) Reakcija šest je prva reakcija oksidacije koja nastaje konverzijom gliceraldehida-3-fosfata u 1,3 difosfoglicerat.
13

7) U sedmoj reakciji 1,3- difosfoglicerat se defosforiliše i prelazi u 3-fosfoglicerat.
8) U osmoj reakciji 3-fosfoglicerat pod uticajem enzima fosfogliceromutoze prelazi u 2-fosfoglicerat.
9) U devetoj reakciji 2-fosfoglicerat u procesu dehidracije prelazi u fosfoenolpiruvat, uz učešće enzima enolaze pri čemu se izvaja molekul H2O.
10) U desetoj reakciji defosforilacijom fosfoenolpiruvata pod uticajem enzima piruvatkinoze u prisustvu ADP koji prelazi u ATP nastaje enolpiruvat koji se spontano transformiše u stabilan oblik piruvat.
Iako se u procesu glikolize proizvodi mali broj molekula ATP za ćelijsku aktivnost, ove reakcije omogućavaju stvaranje malih molekula koje čine osnovu u strukturi ćelija prokariota i eukariota. Najveći ostatak energije povezan je sa piruvatom, koji je dominantno jedinjenje ovog procesa. Na osnovu konačne oksidacije jedinjenja izvedenih iz piruvata energija se može ponovo dobiti natrag.
Drugi katabolički putevi ugljikohidrata
Postoji više biohemijskih puteva za razgradnju glukoze do piruvata. Pored EMP puta pojavljuju se i druge važne reakcije- putevi katabolizma. Dva su tipična primjera tih puteva :
a) Entner-Duodoroff-ov (ED) put- glukoza se prevodi u pirogrožđanu kiselinu( piruvat) u manjem broju reakcija nego što je slučaj sa glikozom.
b) pentozo-fosfatnim putem (PFP)- u kojem se glukoza prevodi u pentozne jedinice koje se koriste za sintezu nukleinskih kiselina.
14

Entner-Duodoroffov (ED) put
Slično glikolizi i ovaj put kao krajnji produkt pri razlaganju glukoze proizvodi piruvat. ED obuhvata početnu fosforilaciju isto kao kod glikolize, a zatim procesom oksidacije glukozo-6-fosfat prelazi u 6-fosfoglukonsku kiselinu.
Dehidratacijom 6-fosfoglukonske kiseline nastaje 2-keto-3-dezoksi-6-fosfoglukonska kiselina. Navedena se kiselina pod uticajem enzima aldolaze cijepa na 2 trioze: pirogrožđanu kiselinu ( piruvat) i gliceraldehid-3-fosfat koji se oksidacijom uz istovremenu redukciju NAD može prevesti u piruvat. Ovaj put je osnovni energetski put kod nekih Gram negativnih bakterija i to prvenstveno kod vrsta iz rodova: Pseudomonas, Rhizobium, Agrobacterium. Ovaj put nije tipičan za Gram pozitivne bakterije.
Pentozo fosfatni put ( PFP)
On se, sa glikolizom, susreće kod velikog broja bakterija. U ovom katabolitičkom putu se proizvodi riboza koja se koristi za sintezu nukleinskih kiselina i NADPH koji se koristi za ostale reakcije sinteze, uključujući i vezivanje CO2 u proces fotosinteze.
Prva reakcija u PFP put je fosforilacije glukoze do glukoza-6-fosfata i ista je kao kod glikolize i ED-puta. U sljedećoj reakciji dolazi do provođenja glukoza-6-fosfata u 6-fosfoglukonsku kiselinu, isto kao kod ED-puta. Nakon toga slijedi njena transformacija u ribuloza-5-fosfat u procesu oksidacije i dekarboksilacije, a zatim ribuloza 5-fosfat prevodi u riboza-5-fosfat. Ribuloza-5-fosfat se također spaja sa CO2 u reakciji što se odvija u mraku tokom fotosinteze. Pri tome se NADP+ reducira, a jedinice pentoze mogu biti pretvorene u dva različita međuprodukta.
Tako PFP put služi kao veza sa glikolizom za nastajanje pentoznih jedinica i reduktiranog koenzima NADPH+ .
15

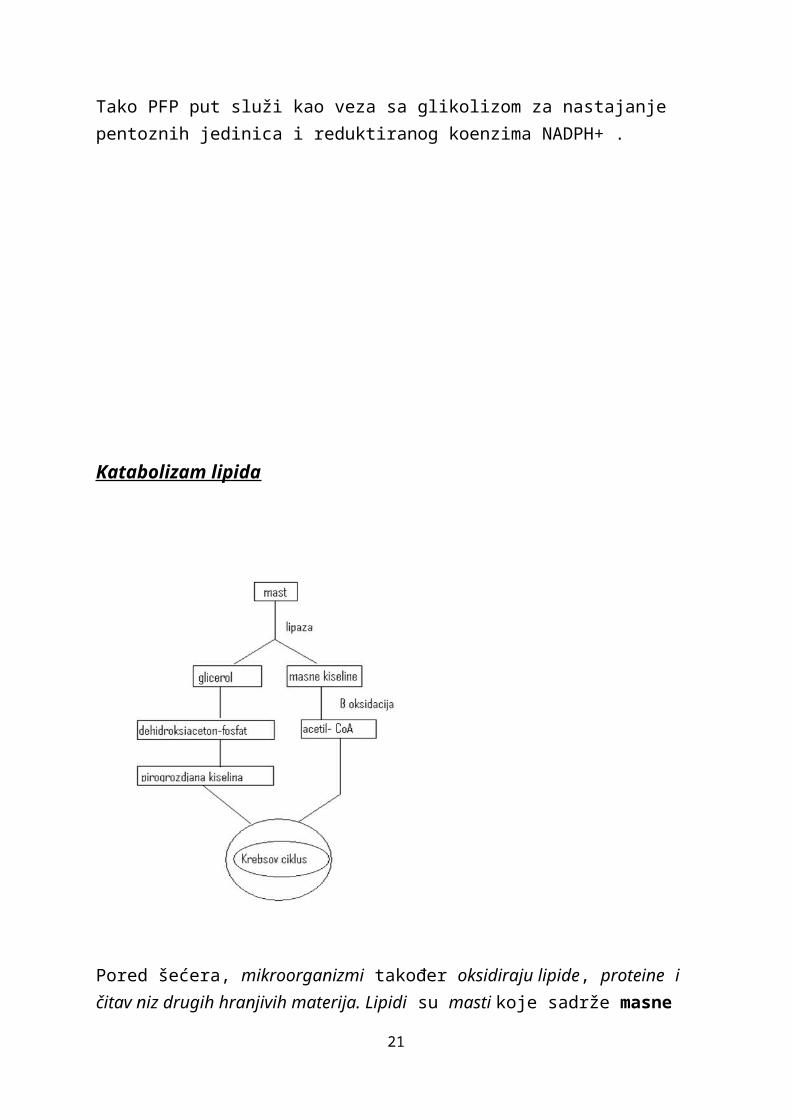
Katabolizam lipida
Pored šećera, mikroorganizmi također oksidiraju lipide, proteine i čitav niz drugih hranjivih materija. Lipidi su masti koje sadrže masne kiseline i glicerol. Mikroorganizmi sintetiziraju ekstracelularne enzime, nazvane lipaza, koji cijepaju lipide na masne kiseline i glicerol. Svako od nastalih jedinjenja se potom posebno metabolizira.
Glicerol veliki broj različitih mikroorganizama može prevesti u dihidroksi-aceton-fosfat, jedan od međuprodukata sa tri ugljikova atoma koji nastaje tokom glikolize.Dihidroksi-aceton-fosfat se potom metabolizira tako što se prevodi u gliceraldehid-3-fosfat koji se dalje katabolizira glikoliznim putem do pirogrožđane kiseline.
Masne kiseline se kataboliziraju nešto drugačijim putem. Mehanizam oksidacije masnih kiselina naziva se β-oksidacija. Pri ovom procesu se cijepaju dijelovi dugačkog lanca masne kiseline, tako da se odvajaju po dva ugljikova atoma, u obliku acetil-CoA. Tako nastale molekule acetil-CoA dalje ulaze u Krebsov ciklus, a također i molekule acetil -CoA koje su nastale u procesu oksidacije pirogrožđane kiseline. Na osnovu navedenog proizilazi da je oksidacija glicerola i masnih kiselina direktno povezana sa oksidacijom glukoze pomoću Krebsovog ciklusa.
16

Katabolizam proteina
Proteini su makromolekule koje bez pomoći( tj. razgradnje) ne mogu da prođu kroz citoplazmatsku membranu. Mikroorganizmi sintetiziraju ekstracelularne proteaze i peptidaze, a navedeni enzimi cijepaju proteine na manje podjedinice- aminokiseline, koje mogu biti prenesene kroz citoplazmatsku membranu. Međutim, prije nego što aminokiseline budu katabolizirane, one moraju biti prevedene u jedinjenja koja mogu ući u Krebsov ciklus.
U transformaciji, nazvanoj dezaminacija, uklanja se sa aminokiseline- aminogrupa u obliku amonijum jona, koji se izlučuje iz ćelije. Preostali dio aminokiseline može se uključiti u Krebsov ciklus.
Druga transformacija obuvata dekarboksilaciju( uklanjanje COOH grupe) i dehidrogenaciju ( uklanjanje H+ atoma). Neophodno je istaći da se aminokiseline različitim postupcima mogu prilagoditi da bi mogle ući u Krebsov ciklus.
AEROBNA RESPIRACIJA
( Krebsov ciklus- Ciklus limunske kiseline )
Ovaj ciklus je dobio ime po Hans Krebsu, koji je otkrio neka jedinjenja koja se pojavljuju u ovom ciklusu, a sastoji se od niza hemijskih reakcija katalizitanih enzima.
Taj niz reakcija se opisuje kao ciklus zato što jedinjenje koje nastaje na kraju ovih reakcija, oksaloacetat, je identično sa jedinjenjem sa početka ove reakcije- oksalacetatom.
17

Molekule piruvata, dobivene glikolizom ili nekim drugim putem, neprekidno se uključuju u ovaj ciklus. Međutim, prije početka Krebsovog ciklusa iz pirogrožđane kiseline se enzimatskim pute mora ukloniti CO2. Taj prelazni korak između dva ciklusa omogućuje jedinjenju sa dva ugljika- acetilu da se poveže sa koenzimom A u oblik acetil ≈CoA, radi početka sljedeće reakcije u Krebsovu ciklusu.
Devet reakcija u ciklusu limunske kiseline mogu se podijeliti u tri stadija:
1) stadij A- pripremni2) stadij B- izdvajanje energije3) stadij C- regeneracija početne materije.
Ovaj ciklus započinje i završava se oksalacetatom koji se povezuje sa acetil- CoA. Potom se acetil-CoA oksidira do CO2 i H2O, a zatim se CO2 prenosi u okolinu kao gasoviti produkt aerobne respiracije. Ovaj se ciklus može opisati kao “energetski krug ćelijskog metabolizma “ jer neprekidno nadoknađuje energiju potrebnu ćeliji.
Stadij A- pripremni stadij- obuhvata tri reakcije:
18

Acetil CoA se spaja sa oksaloacetatom ( 4 C atoma) u limunsku kiselinu-citrat ( 6 C atoma). U ostalim dvjema reakcijama vrši se premještanje hidroksilne grupe, pa nastaje akonitat, a zatim izocitrat ili izocitronska kiselina.
Stadij B- stadij izdvajanja ( oslobađanja) energije- obuhvata 4 reakcije:
U četvrtoj reakciji se vrši redukcija NAD+ u NADH, što uzrokuje oksidativno uklanjanje CO2( dekarboksilaciju) s molekule izocitronske kiseline i nastanak α-keto-glutarne kiseline ( 5 C atoma). U petoj reakciji se α-ketoglutarna kiselina oksidira i dekarboksilira i u reakciji s CoA stvara sukcinil-CoA (4 C atoma). U ovoj reakciji se također izdvaja i elektronski par koji reducira NAD+ u NADH. Veze što spaja sukcinil-grupu i CoA bogata je energijom i sliči onoj u ATP. U šestoj reakciji se cijepa energijom bogata veza i otpušta se energija, što uzrokuje fosforilaciju guanozin-difosfata( GDP) u guanozin- trifosfat( GTP). Sukcinat ili jantarna kiselina predstavlja kraj reakcije. U sedmoj reakciji se oksidira jantarna kiselina ( sakcinat) u fumarnu kiselinu- fumarat. U ovoj reakciji sudjeluje drugi elektron-akceptor flavinadenindinukleotid( FAD+) koji se redukuje u FADH2.
Stadij C- stadij regeneracija početne materije- Ovaj stadij Krebsovog ciklusa obuhvata dvije reakcije. U osmoj reakciji se molekula vode dodaje fumarnoj kiselini pri čemu nastaje malat ( jabučna kiselina). U devetoj reakciji malat potom biva oksidirana i ponovo se stvara oksaloacetat. U ovoj reakciji oksidacije također se oslobađaju dva elektrona, koji reduciraju NAD+ u NADH. Regeneracijom oksaloacetata Krebsov ciklus može ponovo započeti povezivanjem sa novom molekulom acetil- CoA.
Svaka molekula piruvata, koja se oksidira putem ovog ciklusa, uzrokuje obrazovanje 12 molekula ATP. Pošto se od svake molekule glukoze može proizvesti 2 molekula piruvata, iz jedne molekule glikoze dobija se 24 molekula ATP.
Prokariotski aerobni mikroorganizmi stvaraju ukupno 38 molekula ATP iz molekula glukoze tokom aerobne respiracije glukoze. U procesu glikoze nastaje 2 ATP supstratnom fosforilacijom, a u sličnom se procesu dobijaju još 2 ATP u Krebsovom ciklusu. Ukupno 10 NADH proizvodi 30 ATP, a 2 FADH mogu proizvesti 4 ATP sve u toku oksidativne fosforilacije, što ukupno iznosi 38 molekula ATP.
19

Aerobne fermentacije
Neki mikroorganizmi vrše nepotpunu oksidaciju glukoze tako da se kao krajnji proizvodi dobijaju neke organske kiseline i različit broj molekula CO2, NADH i FADH. Ove respiracije su nazvane aerobne fermentacije. Najčešće su prisutne sirćetna, fumarno-ćilibarna i limunska fermentacija.
Sirćetna fermentacija
20
Pri ovoj fermentaciji se kao krajnji proizvod dobija sirćetna kiselina. Razlaganje glukoze do pirogrožđane kiseline odvija se po EMP putu. Pirogrožđana kiselina se zatim dekarboksilira do acetaldehida, a njegovom oksidacijom nastaje sirćetna kiselina. Pored glukoze supstrat za stvaranje sirćetne kiseline može biti i etil alkohol koji se oksidiše do acetaldehida, a daljom oksidacijom acetaldehida nastaje sirćetna kiselina.
Mikroorganizmi koji imaju ovaj tip respiracije pripadaju bakterijama i nazivaju se sirćetne bakterije, a najpoznatije vrste su: Acetobacter aceti, Acetobacter xylinoides, Acetobacter suboxydans, Acetobacter acidophylum koje su sposobne i da svoj proizvod oksidišu u potpunosti do CO2 i H2O i Gluconobacter oxydans koji nema navedenu sposobnost. Ove bakterije se koriste za proizvodnju sirćetne kiseline ( vinsko, voćno, alkoholno) s tim da koncentracija alkohola nije veća od 10-13%.
Fumarno-ćilibarna fermentacija
Kao krajnji produkt respiracije gljiva iz roda Mucor, Rhizopus, Cunningamella i Circinella iz glukoze nastaju fumarna, ćilibarna i jabučna kiselina. Transformacija glukoze se vrši po EMP putu do pirogrožđane kiseline koja se uključuje u Krebsov ciklus.
Limunska fermentacija
U procesu respiracije nekih gljiva, među kojima je najpoznatija vrsta Aspergillus niger, iz glukoze nastaje limunska kiselina. U kiseloj sredini pirogrožđana kiselina se oksidiše u koenzim uz pomoć NAD, a zatim se dekarboksilizira ( izdvaja se CO2) i sjedinjuje se sa CoA pri čemu nastaje acetil-CoA. Međusobnom reakcijom acetil CoA i oksalsirćetne kiseline nastaje limunska kiselina. Zahvaljujući ovom tipu respiracije vrsta Aspergillus niger se koristi u industrijskoj proizvodnji limunske kiseline.
Ostale aerobne fermentacije
Gljive, u zavisnosti od vrste, mogu u toku respiracije proizvesti i druge različite kiseline. Tako neke vrste iz roda Aspergillus kao što su Aspergillus itaconicus i Aspergillus terreus, iz glukoze stvaraju itakonsku kiselinu, a neke vrste iz roda Aspergillus i Penicillium stvaraju glukonsku kiselinu.
21

Svi ovi mikroorganizmi koji imaju nepotpunu aerobnu respiraciju značajni su za transformaciju monosaharida u tlu. Proizvodi njihove respiracije koriste drugi mikroorganizmi pri čemu se vrši sinteza nove organske materije čijom mineralizacijom nastaju biljni asimilativi.
Lanac disanja i oksidativna fosforilacija
Vodik koji se u toku razgradnje glukoze, u Krebsovom ciklusu i aerobnim fermentacijama vezao za NAD i FAD prenosi se na lanac disanja koji predstavlja niz enzima koji prenose vodik od supstrata do akceptora. Kod aerobnog disanja krajnji akceptor vodika i elektrona jeste kisik, a kao rezultat oksidacije nastaje voda i energija koja se akumulira u ATP. Enzimi koji čine lanac disanja kod mikroorganizama su flavoprotein, hinoni i citohromi. Lanac disanja nije isti kod svih mikroorganizama, a naročito su uočene razlike u redoslijedu i broju citohroma. Kod eukariotskih mikroorganizama lanac disanja smješten je u mitohondrijama, a kod prokariota u citoplazmatskoj membrani.
Vodik se sa NADH-a i FADH-a prvo prenosi na flavoprotein, a zatim na hinon. U ovoj fazi se vodik razlaže na proton i elektron. Proton se izvaja iz lanca, a elektron se dalje prenosi preko sistema citohroma do kisika. Kisik nakon što primi elektron prelazi u anionski oblik i potom reaguje sa protonom vodika pri čemu nastaje voda. Prilikom prenošenja vodika i elektrona oslobađa se energija. Oslobađanje energije je postepeno i slično kaskadama vodopada u kome se ukupna energija slapa oslobađa u dijelovima.
Oslobođena energija se akumulira u ATP, a proces se naziva oksidativna fosforilacija.
Dokazano je da se oksidacijom jednog molekula NADH oslobodi maksimalno 3 molekula ATP, oksidacijom jednog FADH maksimalno 2 ATP, dok je energija koji
22

ima GTP identična energiji ATP. Mikroorganizmi sa kraćim lancem disanja dobivaju manje energije, a što je obično slučaj sa bakterijama i gljivama koje respiraciju vrše po tipu aerobne fermentacije.
ANAEROBNA RESPIRACIJA
Uporediv anaerobni proces sa biološkim oksidacijama, u kojim kisik nije upotrijebljen kao konačan akceptor elektrona, naziva se anaerobna respiracija. U anaerobnoj respiraciji se u prvom redu deduciraju jedinjenja kao npr. karbonati, nitrati, sulfati. Veliki broj fakultativno anaerobnih bakterija može reducirati nitrate u nitrite u anaerobnim uslovima. Ovaj tip reakcije omogućava stalan rast organizama kada nema slobodnog kisika, ali se tada nagomilavaju nitriti, proizvedeni redukcijom nitrata, koji mogu biti toksični za organizme.
Neke vrste iz rodova Bacillus i Pseudomonas mogu kontinuirano reducirati nitrite do molekularnog azota. Taj proces se naziva dentrifikacija. Taj proces je prisutan samo kada te aerobne bakterije rastu u anaerobnim uslovima . Organizmi koji reduciraju sulfate i karbonate su obligatni anaerobi. Vrsta Desulfovibrio desulfuricans reducira sulfate do vodiksulfida, a također oksidira i ugljikohidrate do sirćetne kiseline, dok vrsta Methanobacterium bryantii vrši redukciju CO2 do metana. Ove reakcije se pojavljuju u prirodi u procesima ponovnog vraćanja navedenih elemenata u kružni tok materija.
23





























