Embed Size (px)
Citation preview

Vol. 165, No. 2, 1989
December 15, 1989
BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
Pages 575-579
NADH- AND NADPH-LINKED AQUACOBALAMIN REDUCTASES OCCUR IN BOTH MITOCHONDRIAL AND
MICROSOMAL MEMBRANES OF RAT LIVER
Fumio Watanabe*, Yoshihisa Nakano#, Shinichi Maruno#, Noriyuki Tachikake#, Yoshiyuki Tamura, and Shozaburo Kitaoka#
Laboratory of Nutrition and Food Science, College of Hagoromo-gakuen, Sakai, Osaka 592, Japan
#Department of Agricultural Chemistry, University of Osaka Prefecture, Sakai, Osaka 591, Japan
Received October 30, 1989
SUMMARY: To elucidate the mechanism for the synthesis of the coenzyme forms of cobalamin in mammals, rat liver aquacobalamin reductase was partially characterized. Rat liver contained both NADH- and NADPH-linked aquacobalamin reductases. The NADH-linked enzyme was distributed in the mitochondria (approx. 40%) and microsomes (60%), identical to the distribution of the NADPH-linked enzyme. The two mitochondrial NADH- and NADPH-linked enzymes were located inside of the outer membrane. ® 1989 Academic Press, Inc.
Aquacobalamin reductase catalyzes the reduction of OH-Col to cob(ll)alamin in the
synthesis of the coenzyme forms Ado- and Me-Cbls (i). The enzyme has been reported to
occur in Clostridium tetanomorphum (i) and Eu$1ena sracilis(2). The Clostridium and
Euglena enzymes are linked to NADH and NADPH, respectively. Although the overall con-
version activity of 0H-Cbl to Ado-Cbl has been found in the mitochondrial fraction of
mammmlian cells (3, 4), the enzymological information on the m~mmalian aquacobalamin
reductase is not available yet. We describe the subeellular distribution of the enzyme
and discuss the synthesis of the coenzyme forms in rat liver.
EXPERIMENTAL PROCEDURES
Animals: Male albino rats of the Wistar strain, housed in individual cages, were fed normal laboratory stock diet, EC-2 (Clea Japan Inc.), ad libitum and used when they attained a body weight of about 250 g.
Subcellular fractionation: The livers were resected from rat starved for 24 h, washed with chilled 0.9% NaCI solution, cut into small pieces with a razor, and homogenized in about 4 volumes of chilled 0.25 M sucrose solution (adjusted to pH 7.5 with 0.i M KOH) by using a Potter-Elvenhjem-type glass homogenizer with a Teflon pestle (two strokes at a low speed of 200-300 rpm). The homogenate was filtered through a double layer of gauze to remove unbroken tissues and then centrifuged at 500 x g for i0 min to remove unbroken cells. A portion (i.0 ml) of the supernatant was layered on the top of a 9.0 ml linear sucrose gradient (20-60% w/w) and centrifuged at i00,000 x g for 2.5 h in a Hitachi 55P centrifuge with a swing-out bucket rotor (RPS 40T). Fractions (0,3 ml) were collected from the bottom of the centrifuge tube. All procedures were done at 4°C.
*Correspondence to: Fumio Watanabe, Ph D., Laboratory of Nutrition and Food Science, College of Hagoromo-gakuen, Sakai, Osaka 592, Japan.
The abbreviations used are: Cbl, cobalamin; OH-Cbl, hydroxocobalamin; Me-Cbl, Methyl- cobalamin; Ado-Cbl, 5'-deoxyadenosyleobalamin; Hepes, N-2-hydroxyethylpiperazine-N- 3-propanesulfonic acid.
675
0006-291 X/89 $1.50 Copyright © 1989 by Academic Press, Inc.
All rights of reproduction in any form reserved.

Vol. 165, No. 2, 1989 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
Submitochondrial fractionation: The supernatant fraction obtained by centrifugation (500 x g for i0 min) of the liver homogenate was centrifuged at I0,000 x g for i0 min and the precipitate, which contained the mitochondria, was suspended in 25 mM glycyl- glycine (Glygly)-KOH buffer, pH 7.5, containing 0.25 M sucrose, and then subjected to Percoll density gradient (i0, 30, and 50% v/v Percoll in 25 ~i Glygly-KOH buffer, pH 7.5, containing 0.25 M sucrose and 3% Ficoll) centrifugation. After centrifugation at 7,500 x g for 30 rain, the fraction banding between 30 and 50% Percoll was collected and washed with 25 mM Glygly-KOH buffer, pH 7.5, containing 0.25 M sucrose to remove the Percoll. The Percoll-purified mitochondria were partially disrupted by sonic treatment according to Sottocasa et al., (5). A pomtion (i.0 ml) of the treated mitochondria was layered on the top of a 9.0 ml linear sucrose gradient (20-60% w/w) and centrifuged at i00,000 x g for 2.5 h. Fractions (0.3 ml) were collected from the bottom of the centrifuge tube. All procedures were done at 4@C.
Enzyme assay: The activities of NADH- and NADPH-linked aquacobalamin reductases were assayed by spectrophotometric estimation of the amount of OH-Cbl converted to cob(ll)- alamin at 50°C. The reaction mixture (i.0 ml) contained 50 mM Tris-acetate buffer, pH 7.0, 0.i mM OH-Cbl, 0.2 mM NAD(P)H and enzyme. The cob(ll)alamin formed was assayed by measuring the decrease in absorbance of OH-Cbl at 525 nm and calculated on the basis of the differential molecular extinction coefficient of OH-Cbl to cob(ll)alamin (5.57
x 103 M -I cm-l).
The following enzyme activities were assayed by the methods described in the cited references: Lactate dehydrogenase (6), a cytosolic maker enzyme; rhodanese (7), a mito- condrial marker enzyme; glueose-6-phosphatase (8), a microsomal marker enzyme; catalase (9), a peroxisomal marker enzyme; rotenone-insensitive NADH-cytochrome e reduetase (i0) and monoamine oxidase (ii), marker enzymes of the outer membrane of mitochondria; adenylate kinase (5), a marker enzyme of the intermembrane space of mitochondria; suc- cinate dehydrogenase (12), a marker enzyme of the inner membrane of mitochondria; and
isocitrate dehydrogenase (NAD +) (13), a marker enzyme of mitochondrial matrix.
Protein assay: Protein was measured by the method of Bradford (14) with ovalbumin as a standard.
Materials: OH-Cbl and CN-Cbl were obtained from Sigma. FAD and FMN were purchased from Pharmacia P-L Biochemicals.
RESULTS AND DISCUSSION
The rat liver homogenate contained both NADH- and NADPH-linked aquacobalamin re-
ductases with specific activities of 11.67 and 3.69 nmol/min/mg protein, respectively.
Non-enzymatic reduction of OH-Cbl by reducing substances of either high or low molecular
weight in the homogenate was not detected.
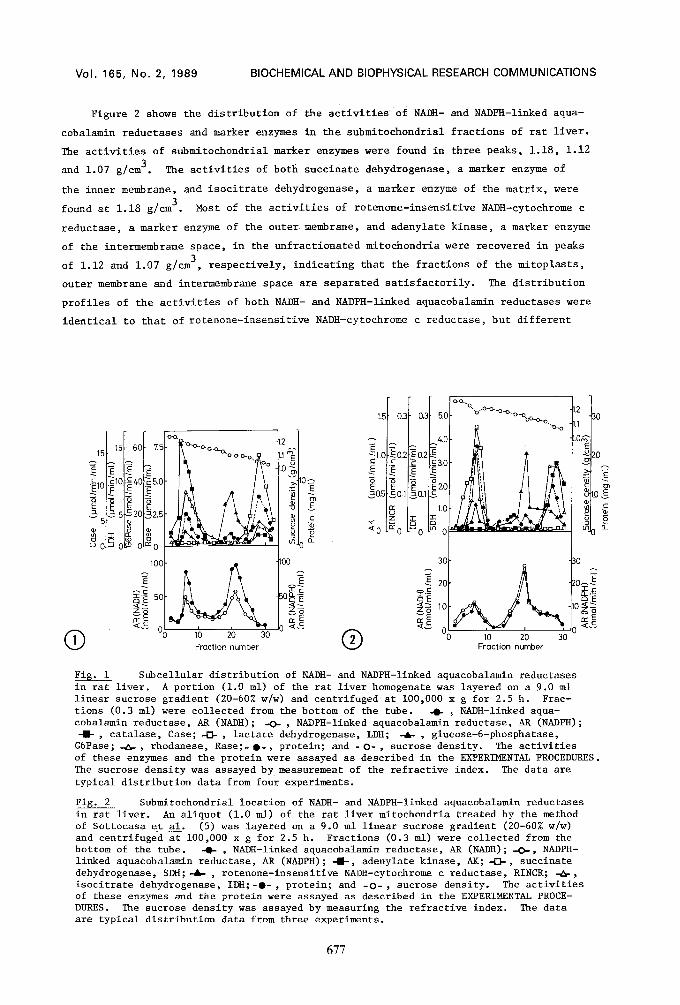
A cell homogenate of rat liver was separated by linear sucrose gradient (20-60% w/w)
centrifugation (Fig. i). Peroxisomes, mitochondria, microsomes and cytosol were sepa-
rated satisfactorily, since the activities of catalase, a peroxisomal marker enzyme;
rhodanese, a mitochondrial marker enzyme; and glucose-6-phosphataae, a mierosomal marker
enzyme, were found in peaks corresponding to densities of 1.20, 1.18 and 1.08 g/cm 3, res-
pectively; and that of lactate dehydrogenase, a cytosolie marker enzyme, was found in
the top fraction. Both activities of NADH- and NADPH-linked aquacobalamin reductases
were found in two peaks (1.18 and 1.08 g/cm3), which were identical to those of the
activities of rhodanese and glucose-6-phosphatase, respectively, but not to that of the
activity of catalase or lactate dehydrogenase. Approximately 40% of the activities of
NADH- and NADPH-linked aquacobalamin reductases in the homogenate was recovered in the
mitochondrial fraction, and the remaining activity found in the microsomal fraction.
These results show that both mitochondria and microsomes contain the two types of re-
ductases.
676
Vol. 165, No. 2, 1989 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
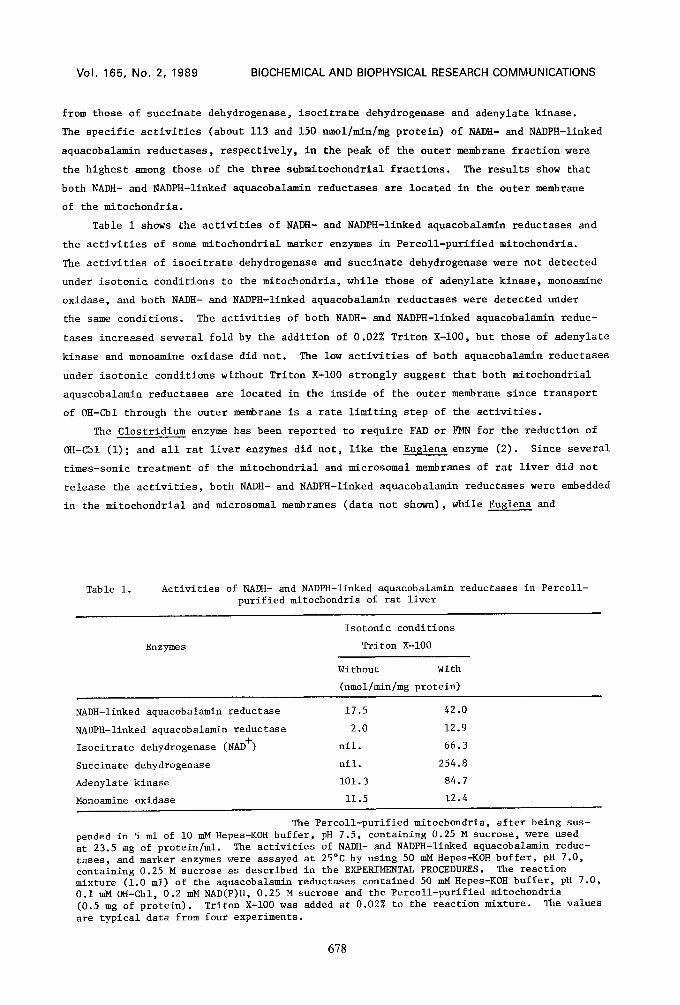
Figure 2 shows the distribution of the activities of NADH- and NADPH-linked aqua-
cobalamin reductases and marker enzymes in the submitochondrial fractions of rat liver.
The activities of submitochondrial marker enzymes were found in three peaks, 1.18, 1.12
and 1.07 g/cm 3. The activities of both suecinate dehydrogenase, a marker enzyme of
the inner membrane, and isocitrate dehydrogenase, a marker enzyme of the matrix, were
found at 1.18 g/cm 3. Most of the activities of rotenone-insensitive NADH-cytochrome c
reductase, a marker enzyme of the outer membrane, and adenylate kinase, a marker enzyme
of the intermembrane space, in the unfractionated mitochondria were recovered in peaks
of 1.12 and 1.07 g/cm 3, respectively, indicating that the fractions of the mitoplasts,
outer membrane and intermembrane space are separated satisfactorily. The distribution
profiles of the activities of both NADH- and NADPH-linked aquacobalamin reductases were
identical to that of rotenone-insensitive NADH-cytochrome c reductase, but different
15F 60 75 15.
• ~1o "~1o~.~4o~s.o
:~ ~ 5J-S20L~2.5 5 8
~.0 IO ~ o-~ o~ o~=o /
i00[
< c o (D 10 20 30 Fraction number
I0~
~10 o_
~2
®
1.5" 02
~o.5 ~o.1
8
~0 ~0
0,"
0.2
0.1
5C
4.C
gC
2£
1.0
0
o-~. D~,o/O-O'--O-.~.~... 0
DO-C~,o, "0--0
.2 30 .1
.Oc
20
m
30~ ~30
A . ~ 20
O0 10 20 30 Fraction number
Fig. i Subcellular distribution of NADH- and NADPH-linked aquacobalamin reductases in rat liver. A portion (i.0 ml) of the rat liver homogenate was layered on a 9.0 ml linear sucrose gradient (20-60% w/w) and centrifuged at i00,000 x g for 2.5 h. Frac- tions (0.3 ml) were collected from the bottom of the tube. -O- ~ NADH-linked aqua- cobalamin reductase, AR (NADH); -O- , NADPH-linked aquacobalamin reductase, All (NADPH); -I- , catalase, Case; -0- , lactate dehydrogenase, LDH; -4&- , glucose-6-phosphatase, G6Pase; -~-, rhodanese, Rase;- O-, protein; and - O- , sucrose density. The activities of these enzymes and the protein were assayed as described in the EXPERIMENTAL PROCEDURES. The sucrose density was assayed by measurement of the refractive index. The data are typical distribution data from four experiments.
Fig. 2 Submitochondrial location of NADH- and NADPH-linked aquacobalamin reductases in rat liver. An aliquot (I.0 ml) of the rat liver mitochondria treated by the method of Sottocasa et al. (5) was layered on a 9.0 ml linear sucrose gradient (20-60% w/w) and centrifuged at i00,000 x g for 2.5 h. Fractions (0.3 ml) were collected from the bottom of the tube. -0- , NADH-linked aquacobalamin reductase, All (NADH); -O-, NADPH- linked aquacobalamin reductase, AR (NADPH); -m-, adenylate kinase, AK; -{3-, succinate dehydrogenase, SDH;-4&- , rotenone-insensitive NADH-cytochrome c reductase, RINCR; -~-, isocitrate dehydrogenase, IDH;-o- , protein; and -O- , sucrose density. The activities of these enzymes and the protein were assayed as described in the EXPERIMENTAL PROCE- DURES. The sucrose density was assayed by measuring the refractive index. The data are typical distribution data from three experiments.
677
Vol. 165, No. 2, 1989 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
from those of succinate dehydrogenase, isocitrate dehydrogenase and adenylate kinase.
The specific activities (about 113 and 150 nmol/min/mg protein) of NADH- and NADPH-linked
aquacobalamin reduetases, respectively, in the peak of the outer membrane fraction were
the highest among those of the three submitochondrial fractions. The results show that
both NADH- and NADPH-linked aquacobalamin reductases are located in the outer membrane
of the mitochondria.
Table i shows the activities of NADH- and NADPH-linked aquacobalamin reductases and
the activities of some mitochondrial marker enzymes in Percoll-purified mitochondria.
The activities of isocitrate dehydrogenase and succinate dehydrogenase were not detected
under isotonic conditions to the mitochondria, while those of adenylate kinase, monoamine
oxidase, and both NADH- and NADPH-linked aquacobalamin reductases were detected under
the same conditions. The activities of both NADH- and NADPH-linked aquacobalamin reduc-
tases increased several fold by the addition of 0.02% Triton X-IO0, but those of adenylate
kinase and monoamine oxidase did net. The low activities of both aquacobalamin reductases
under isotonic conditions without Triton X-100 strongly suggest that both mitochondrial
aquacobalamin reductases are located in the inside of the outer membrane since transport
of OH-Col through the outer membrane is a rate limiting step of the activities.
The Clostridium enzyme has been reported to require FAD or FMN for the reduction of
OH-Cbl (i); and all rat liver enzymes did not, like the Eu$1ena enzyme (2). Since several
times-sonic treatment of the mitochondrial and microsomal membranes of rat liver did not
release the activities, both NADH- and NADPH-linked aquacobalamin reductases were embedded
in the mitochondrial and microsomal membranes (data not shown), while Eu$1ena and
Table i. Activities of NADH- and NADPH-linked aquacobalamin reductases in Percoll- purified mitochondria of rat liver
Enzymes
Isotonic conditions
Triton X-100
Without with
(nmol/min/mg protein)
NADH-linked aquacobalamin reductase 17.5
NADPH-linked aquacobalamin reductase 2.0
Isocitrate dehydrogenase (NAD +) nil.
Succinate dehydrogenase nil.
Adenylate kinase 101.3
Monoamine oxidase 11.5
42.0
12.9
66.3
254.8
84.7
12.4
The Percoll-purified mitochondria, after being sus- pended in 5 ml of i0 mM Hepes-KOH buffer, pH 7.5, containing 0.25 M sucrose, were used at 23.5 mg of protein/ml. The activities of NADH- and NADPH-linked aquacobalamin reduc- tases, and marker enzymes were assayed at 25°C by using 50 mMHepes-KOH buffer, pH 7.0, containing 0.25 M sucrose as described in the EXPERIMENTAL PROCEDURES. The reaction mixture (i.0 ml) of the aquacobalamin reductases contained 50 mM Hepes-KOH buffer, pH 7.0, 0.I mM OH-Cbl, 0.2 mM NAD(P)H, 0.25 M sucrose and the Percoll-purified mitochondria (0.5 mg of protein). Triton X-100 was added at 0.02% to the reaction mixture. The values are typical data from four experiments.
678

Vol. 165, No. 2, 1989 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
Clostridium enzymes have been found in the soluble fractions of mitochondria and cell
homogenate, respectively (i, 2).
By the labeling rat hepatocytes with radioactive OH-Cbl, most of the Ado-Cbl syn-
thesized and Me-Cbl are accumulated in the mitochondrial and cytosolic fractions, res-
pectively (15). The mitochondrial Ado-Cbl synthetic system has been reported to be
present in rat liver (16), human skin fibroblasts (17), guinea pig mucosa (4) and E.
$racilis (2), suggesting that the mitochondrial NADH- and/or NADPH-linked aquacobalamin
reductases function in Ado-Cbl synthesis. Although there are some observations (15, 18)
suggesting that the microsomal NADH- and/or NADPH-linked enzymes are involved in the
synthesis of Me-Cbl, which would be supplied to the Me-Cbl dependent methionine synthase
(19) located in rat liver cytosol, the detailed enzymatic systems of Me-Cbl synthesis
are still unknown.
REFERENCES
I. Walker, G. A., Murphy, S., and Huennekens, F. M. (1969) Arch. Biochem. Biophys., 134, 95-102.
2. Watanabe, F., Oki, Y., Nakano, Y., and Kitaoka, S. (1987) J. Biol. Chem., 262, 11514-11518.
3. Pletsch, Q. A., and Coffey, J. W. (1971) J. Biol. Chem., 246, 4619-4629. 4. Peters, T. J., and Hoffbrand, A. V. (1970) Br. J. Haematol., 19, 369-382. 5. Sottocasa, G. L., Kuylenstierna, B., Ernster, L., and Bergstrand, A. (1967)
Methods Enzymol., i0, 448-463. 6. Stolzenbach, F. (1966) Methods Enzymol., 9, 278-288. 7. Weng, L., Heirikson, R. L., and Westley, J. (1979) J. Biol. Chem., 253, 8109-8119 8. de Duve, C., Pressman, B. C., Gianetto, R., Wattiaux, T. G., and Appelmans, F.
(1955) Biochem. J., 60, 604-617. 9. Chance, B., and Maehly, C. (1955) Methods Enzymol., 2, 764-775.
i0. Sottocasa, G. L., Kuylenstierna, B., Ernster, L., and Bergstrand, A. (1967) J. Cell Biol., 32, 415-438.
ii. Weissbach, H., Smith, T. E., Daly, J. W., Witkop, B., and Udenfriend, S. (1960) J. Biol. Chem., 235, 1160-1163.
12. Ackrell, B. C., Kearney, E. B., and Singer, T. P. (1978) Methods Enzymol., 53, 466-483.
13. Plaut, G. W. E. (1969) Methods Enzymol., 13, 34-42. 14. Bradford, M. M. (1976) Anal. Biochem., 72, 248-253. 15. Mellman, I. S., Youngdahl-Turner, P., Willard, H. F., and Rosenberg, L. E. (1977)
Proc. Natl. Acad. Sci. USA, 74, 916-920. 16. Fenton, W. A., and Rosenberg, L. E. (1978) Arch. Biochem. Biophys., 189, 441-447. 17. Mahoney, M. J., Hart, A. C., Steen, V. D., and Rosenberg, L. E. (1975) Proc. Natl.
Acad. Sci. USA, 72, 2799-2803. 18. Mellman, I. S., Willard, H. F., Youngdahl-Turner, P., and Rosenberg, L. E. (1979)
J. Biol. Chem., 254, 11847-11853. 19. Kolhouse, J. F., and Allen, R. H. (1977) Proc. Natl. Acad. Sci. USA, 74, 921-925.
679





























