Embed Size (px)
Citation preview

Int. J. Cancer: 55,692-699 (1993) 0 1993 Wiley-Liss, Inc.
PJbhcatlon of tne nterna:lona, Urion Agains: Carccr PLol cat on oe ;union mernat onale Contre 1e Cancer
NERVE GROWTH FACTOR EFFECTS ON HUMAN AND MOUSE MELANOMA CELL INVASION AND HEPARANASE PRODUCTION Dario MARCHETT11s3, Dave MENTER', Li JIN*, Motowo NAKAJIMA~ and Garth L. NICOLSON' 'Department of Tumor Biology, University of Texas M.D. Anderson Cancer Center, Houston, TX 77030, USA; and 2Division of Bioactive Substances, Institute of Applied Microbiology, University of Tokyo, Tokyo 113, Japan.
The role of growth factor networks in regulating the progres- sion of human melanocytes towards tumorigenicity and ulti- mately the malignant phenotype is poorly understood. In particu- lar, the autocrine and paracrine influences that modulate cellular invasion and extracellular matrix degradative enzymes of melanoma cells remain undefined at the molecular level. We report here that nerve growth factor (NGF) can modify some metastasis-associated cellular properties of human and mouse melanoma cells. Treatment of early-passage human metastatic melanoma cells (MeWo) or their variants (3S5, 70W) with biologically active 2.5s NGF resulted in (a) delayed density- dependent inhibition of melanoma cell growth: (b) increased in vitro invasion through a reconstituted basement membrane: and (c) time- and dose-dependent induction of heparanase, a heparan-sulfate-specific endo-B-D-glucuronidase associated with human melanoma metastasis. These effects of NGF were most marked in the 70W brain-colonizing cells (70W > MeWo > 3S5). The NGF enhancement of heparanase secretion was not species- specific, since it was also observed in murine B I6 melanoma cells; the highest NGF stimulation of heparanase was found in brain-colonizing murine B 16-8 15b variant (B 16-8 15b > B 16- BL6, B 16-F 10, B 16-F I). NGF also increased the invasive capac- ity of the human 70W and murine BI6-BI5b sublines in a chemoinvasion assay performed with filters coated with purified heparan sulfate proteoglycan (HSPG). The enhancement of chemotactic response and heparanase production was detected at NGF concentrations sufficient to fully saturate both low- and high-affinity NGF receptors (NGFR), the neurotrophin receptor (p75) and the trkA gene product, respectively. The results suggest that, in addition to the effects of NGF on cellular development and differentiation within the peripheral and central nervous systems, NGF can exert changes in the invasive properties of neuroectoderm-derived melanoma cells. o 1993 Wiley-Liss, Inc.
Tumor-cell metastasis is a complex process involving detach- ment from the primary tumor, invasion and destruction of adjacent host tissue barriers, intravasation and extravasation of the vascular system, and colonization and growth at distant sites while evading host immune defenses. Critical to several of these steps is the ability of malignant cells to invade basement membranes (BM). Liotta et al. (1986) proposed a 3-step process for tumor-cell invasion of BM, consisting of cell attachment to BM, local dissolution of BM components, and tumor-cell migration through the degraded zone of BM. These steps appear to require multiple receptors and enzymes (Nicolson, 1989).
The responses of malignant cells to paracrine growth and differentiation factors that are differently expressed in differ- ent organs and tissues are important in metastatic colonization and probably determine to a great degree the organ preference of metastatic cell growth (Nicolson, 1993). The first paracrine peptide differentiation factor to be purified and characterized was nerve growth factor (NGF). NGF is important for the survival and differentiation of sympathetic and some sensory neurons in the peripheral nervous system and of certain subpopulations of cholinergic neurons in the central nervous system (Levi-Montalcini, 1987). NGF also stimulates the differentiation in vitro of the pheochromocytoma PC-12 cell line into cells that resemble sympathetic neurons (Greene and Tischler, 1976).
Malignant melanoma is of particular interest because of its neuroendocrine origin and high frequencies of metastasis to the brain. Several clinical and histopathological studies have affirmed that malignant melanomas develop through a se- quence of steps, progressing from benign proliferative lesions to laterally-invasive primary melanomas, to vertically-invasive primary lesions and, finally, to metastases (Herlyn, 1990). The expression of NGF receptors during melanoma development and progression to the metastatic phenotype has been corre- lated with an increase in NGF receptor levels. NGF receptors are expressed in metastatic melanoma cells at levels 10-fold higher than the number of receptors in sensory and sympa- thetic neurons and PC-12 cells (Herlyn, 1990; Fabricant et al., 1977). To study the effects of NGF on melanoma cells we have used a model system consisting of a selected human melanoma cell line (MeWo) and its wheat-germ-agglutinin-selected vari- ants (3S5 and 70W) (Ishikawa et al., 1988a,b). The MeWo brain-metastasizing 70W cell line, selected from the MeWo cell line by exposure to stepwise increases in concentration of wheat-germ agglutinin (WGA) was the first reported brain metastasis of a human tumor in nude mice. Since the number of NGF-binding sites increases during progression of mela- noma to more invasive, metastatic phenotypes, we studied the effects of NGF on metastasis-associated properties of mela- noma cells. The results indicate that exposure to NGF results in enhanced ability of MeWo cells and, most notably, the highly brain-metastatic 70W cells, to penetrate a basement membrane-like matrix in vitro by stimulating BM degradation.
MATERIAL AND METHODS Cell lines, tissue culture and NGF treatment
The human MeWo melanoma line and its WGA-resistant variants 3S5 and 70W (Ishikawa et al., 1988a,b) were main- tained as monolayer cultures in a 1:l (v/v) mixture of DMEMI F12 medium (GIBCO, Grand Island, NY) suppiemented with 5% FBS (GIBCO) at 37°C in a humidified 5% coz-%% air atmosphere. Cells were removed from substratum by trypsin- EDTA treatment and subcultured every 3-4 days, except for the 70W cell line whose medium was changed daily. B16 melanoma sublines (B16-F1, -F10, -BL6 and -B15b) were grown to subconfluence in a 1:l (v/v) mixture of DMEM/F12 medium supplemented with 5% FBS. Cells were rinsed twice in medium without serum, detached in Ca2+- and Mg2+-free PBS containing 2 mM EDTA, and rinsed again with the same solution prior to use. Pheochromocytoma (PC-12) cells were grown in 85% RPMI-1640 medium (GIBCO), 10% FBS on type-IV collagen-coated culture dishes (Collaborative Re- search, Bedford, MA). The cells were diluted at a ratio of 1:3 to 1:4 in culture medium and subcultured weekly. Only subconfluent cells of less than 5 passages from original frozen stocks were used. Cell cultures (2-8 x lo5 cells in 35-mm
3To whom correspondence and reprint requests should be sent, at the Department of Tumor Biology, Box 108, University of Texas M.D. Anderson Cancer Center, 1515 Holcombe Blvd., Box 108, Houston, TX 77030, USA. F a : (713) 794-0209.
Received: July 2, 1993.

NGF AND MELANOMA CELL INVASION 693
culture dishes) were fed and maintained in medium without serum and were left untreated or treated with 50 ng/ml ( 5 2 nM) purified mouse salivary gland 2.5s NGF (Collaborative Research) for 72 hr. NGF bioassays using PC-12 cells were performed to confirm biological activity of purified NGF (Greene and Tischler, 1976). Twenty-four hours before NGF treatment the medium was replaced by serum-free medium. Following incubation with NGF for various times, the cells were scored for differentiation if cellular projections were longer than 2 cell diameters and/or if a branch point was observed. Number, length and density of such processes continued to increase during 2 weeks of NGF treatment, until at least 80% of the cells responded. The processes extended by PC-12 cells in response to NGF treatment resembled those produced by cultured primary sympathetic neurons. The mela- nin content of the cells was quantitatively assayed as described by Miner et al. (1982) with slight modifications. Briefly, cells (1-3 x lo6) were suspended in dd H 2 0 (100 pl), then frozen and thawed 3 times to disrupt the cells. The supernatants were acidified to 0.5 N perchloric acid and extracted twice with cold (4°C) ethyl alcoho1:ethyl ether (3:1 v/v) and once with ethyl ether. Precipitates were dissolved in 1 ml of 0.85 M KOH by boiling the mixture for 10 rnin at 100°C. After insoluble materials were cooled down and removed, absorbance of supernatants was determined at 400 nm. All cell lines were periodically checked for Mycoplasma contamination, and only Mycoplasma-free cell lines were used in this study. Cell growth analysis
A fluorescence cell viability assay was used for the cell growth studies. Briefly, cells in 96-well plates (Corning, Corn- ing, NY) were exposed to 1.17 pM calcein-AM (Molecular Probes, Eugene, OR) for 45 rnin at 37°C. Calcein-AM hydrody- sis by intracellular esterases yielded the intensely fluorescent product calcein. Following incubation (100 pl volume), the cells were rinsed gently with 1,000 vol of tissue-culture grade Dulbecco's PBS without Ca2+, Mg2+ or pH indicator (DPBS). After sequential washes, 100 ml of DPBS were added and the plates were scanned using a Cytofluor 2300 (Millipore, Bed- ford, MA) with emission filters set to detect calcein fluores- cence (530 nm region). Cells numbers were also determined by counting (Coulter Model ZM-6602318, Hialeah, FL) to obtain an optimal ratio between fluorescence and cell number for each of the cell lines. Fluorescence microscopic studies were performed to show that all calcein-AM-treated cells possessed an intense, uniform, green fluorescence. Stock solutions of calcein-AM were kept at -20°C inside a sealed bag containing a dessicant protected from light, and single aliquots (50 pg) were used after thorough solubilization in 50 ml of highly- purified DMSO (Fisher, Pittsburgh, PA). Before initial plating (6,600 cells/well in 96-well plate) the instrument was cali- brated for each cell line. A linear relationship between cell number and fluorescence was observed with sensitivity down to 50 cells/well. R-squared values ranged from 0.85 to 0.91. Cell growth conditions were similar for all 3 cell lines with the exception of the 70W line where the medium was removed every 24 hr to prevent formation of clumps and detachment from the well surface, probably due to secretion of cytotoxic factor(s) in the extracellular environment. In 5% FBS, 2-3 cell-population doublings occurred over a period of 8 days with a doubling time of approximately 24 hr. In the absence of serum, cell growth diminished over the same time interval by 38-50%, according to the cell line used. Heparanase assay
Heparanase activity was determined by degradation of radiolabelled HS using high-speed gel permeation chromatog- raphy (Nakajima et al., 1986) with some modifications. Briefly, subconfluent cells (2 x lo8) were harvested and solubilized in 50 ml of 50 mM Tris-HC1, pH 7.5, containing 1 mM phenylmeth- ylsulphonylfluoride (PMSF; Sigma, St Louis, MO), 5 mM
N-ethylmaleimide (NEM; Sigma), 0.05% sodium azide, 0.5% Triton X-100 at 4°C for 30 min. The cell extract was centri- fuged at 12,000 g for 30 min at 4"C, and protein concentration was determined using the Pierce (Rockford, IL) Coomassie blue protein assay protocol. The cell extract (20-70 p,g protein) was incubated with 5 pg N-t3H]acetylated HS in 100 ml of 0.2 M sodium acetate, pH 5.0, at 37°C for up to 24 hr. The reaction was terminated by heating the samples at 95°C for 5 min. Following a centrifugation step, high-speed gel permeation chromatographic analysis was performed by reverse-phase HPLC. Supernatants (50-100 pl) were injected into a Waters (Milford, MA) 600E HPLC System equipped with a Bio-Gel SEC 30-XL gel filtration column (BioRad, Richmond, CA). A flow rate of elution of 0.50 ml/min was used and fractions were collected every 30 sec. Radioactivity was determined using a liquid scintillation counter (LKB-Pharmacia, Uppsala, Swe- den, 75% efficiency).
Chemoinvasion assay The invasive properties of tumor cells following NGF
treatment were assayed by means of Transwell cell culture chambers (Albini et al., 1987) and monitored by fluorescence plate scanner analysis using the Cytofluor 2300 (Millipore). Reconstituted basement membrane (Matrigel; Collaborative Research) was diluted 1:30 in cold DMEMIF12 without phenol red (Sigma) and applied (approx. 20 mg/100 ml/filter) onto the upper surface of Transwell (6.5 mm diameter; 8 pm pore size) polycarbonate filter inserts (Costar, Cambridge, MA). Alternatively, purified heparan sulfate proteoglycan (HSPG) was applied to the filter at various concentrations (0.125-0.5 mg/ml in 28 M a deionized H20). HSPG was obtained from Becton Dickinson Labware (Bedford, MA). It was EHS-tumor-derived and pure as judged by 4%-20% gradient SDS-PAGE and Western blotting using specific HSPG MAbs. The lower chamber contained a mixture of 0.5% low-gelling agarose (FMC Bio Products, Rockland, ME), 0.5% gelatin (Sigma) with addition of 2.5-10 nM of the chemotactic peptide N-formyl-met-leu-phe (Sigma) and 25% phenol-red- free fresh human brain endothelial cell-conditioned medium (MBE-CM, 48 hr). Total volume in the lower chamber was 380-390 pl, depending on the degree of gel drying. The bottom of each well (24-well plates, Costar) was previously coated with 5 pg/cm2 of fibronectin (Collaborative Research). Cells sus- pended in serum-free, phenol-red-free DME/F12 were seeded into the upper compartment at 2.0 x lo4 cells/filter. After 72 hr incubation at 37°C with varying amounts of purified 2.5s NGF (0-100 ng/ml), the upper chambers were carefully removed and the bottom chambers treated with 1 pM cal- cein-AM for l hr at 37°C. After several washes the plates were analyzed for fluorescence using the Cytofluor 2300 fluores- cence plate scanner (BB4 settings), and the formation of calcein was monitored by measuring the increase in fluores- cence at 530 nm. Controls were performed to optimize conditions with regard to calcein diffusion and hydrolysis. Invasive cells were also visualized by fluorescence microscopy at 530 nm. The accuracy of the invasion assay was assessed by replicate determination of cell number by measuring specific fluorescence of the lower side of the Transwell filters. Trypsin/ EDTA solution (0.6 ml) was added to the bottom chamber and incubation proceeded for 10 rnin at 37°C. To ensure that all cells were removed, 100 pl of a trypsin/EDTA solution were placed on top of each filter for 10 min. At the end of this incubation the cover on the plate of the Transwell was removed, and the plate was sharply hit on its side to ensure that all cells were detached from the membrane. Transwells were also observed under a microscope to see whether all the cells had been removed. A second trypsinization was usually not required. Once the cells became rounded and detached, the trypsin/EDTA was removed from the top, bottom and sides of the Transwell after a quick rinse of the membranes.
694 MARCHETTI ET AL.
The cells in the lower chamber were counted by calcein-AM methodology. We generally observed that approximately 10% of the invasive cells attached to the lower side of the Transwell filters. This percentage did not vary significantly among the 3 MeWo cell lines. It is possible that some of the fluorescence detectable in the assay is associated with diffusion and slow hydrolysis of solubilized calcein-AM. We therefore performed experiments in which calcein-AM was added alone to the wells. Cell suspensions were added either on top of a layer of low-gel agarose mixture or into the bottom well. Calcein-AM was added for the various periods of time and fluorescence monitored by the Cytofluor system. Background fluorescence was noted as well as a decrease in fluorescence when cells were plated on the bottom versus the top of the filter. These control experiments helped us to quantify background fluorescence.
Solubilization of NGF receptors and NGFR binding assays NGF receptors (NGFR) were extracted by incubating the
cells, with continuous mixing, in phosphate-buffered saline (PBS), 2% Nonidet-P40 (NP-40) for 18 hr at 4°C as described (Marchetti and McManaman, 1990). The extracts were then centrifuged to remove insoluble material. The pellet was resuspended in PBS-NP-40 and centrifuged a second time, then supernatants were combined and diluted 1:l with 20 mg/ml cytochrome c in PBS containing 0.5% NP-40. Receptor binding assays were subsequently performed. Briefly, 100 yI of solubilized NGFR were incubated with 100 yl of [1251]-NGF ranging in concentration from 9.0 x lo-'* M to 3.0 x M, for 120 min (at which time the steady state had been reached at all concentrations) at 25°C. The reaction was filtered through GF/C filters using a cell harvester (M-12, Brandel, Gaithers- burg, MD) and washed under vacuum with 40 ml of ice-cold PBS. Non-specific binding was made by including a 2,000-fold excess of corresponding unlabelled NGF in parallel tubes. The filters were dried under heat lamps and counted in a gamma spectrometer. To reduce the background and improve the flow rates of the filtering system, the GF/C filters were soaked in a solution of 20 mg/ml cytochrome c and 0.01% polyethylen- imine for 2 hr at 25"C, drained, and dried in an oven at 65°C prior to use. Background binding to GF/C filters, prepared in this way, was less than 1%. Scatchard analysis was then performed by analyzing binding data with the help of the LIGAND program. To examine the sensitivity of our binding assay and to verify that the extraction and assay procedures effectively solubilized NGFR we examined the binding of [1251]-NGF to solubilized control preparations of PC-12 cells, known to possess both high- and low-affinity NGF binding (positive control), A-875 human melanoma, known to possess only low-affinity binding (positive control for low-affinity NGFR) and A-375 human melanoma, known not to have any appreciable NGF binding activity (negative control; Fabricant et al., 1977). After extraction in 2% NP-40, about 80% of the specific NGF binding sites were found in the soluble superna- tant and re-extraction of the membrane pellet did not result in the release of additional binding sites. In agreement with previous results, we found solubilized preparations of PC-12 cells to have both high-affinity (K, = 10-1lM) and low-affinity (K, = M) NGF binding sites in a ratio of 1:18, respec- tively, A-875 cells to have only low-affinity NGF binding, and A-375 cells to be devoid of NGF binding sites as determined by Scatchard analysis and LIGAND program. The limit of detec- tion of NGF-binding sites in this assay is 25 fmoles.
RESULTS Effect of NGF on the prolife.ation of Me Wo, 3S5, and 70W melanoma cells
The 3 MeWo cell lines were exposed to physiological concentrations (2 nM) of purified, biologically active 2.5s NGF. This specific concentration was used because of its
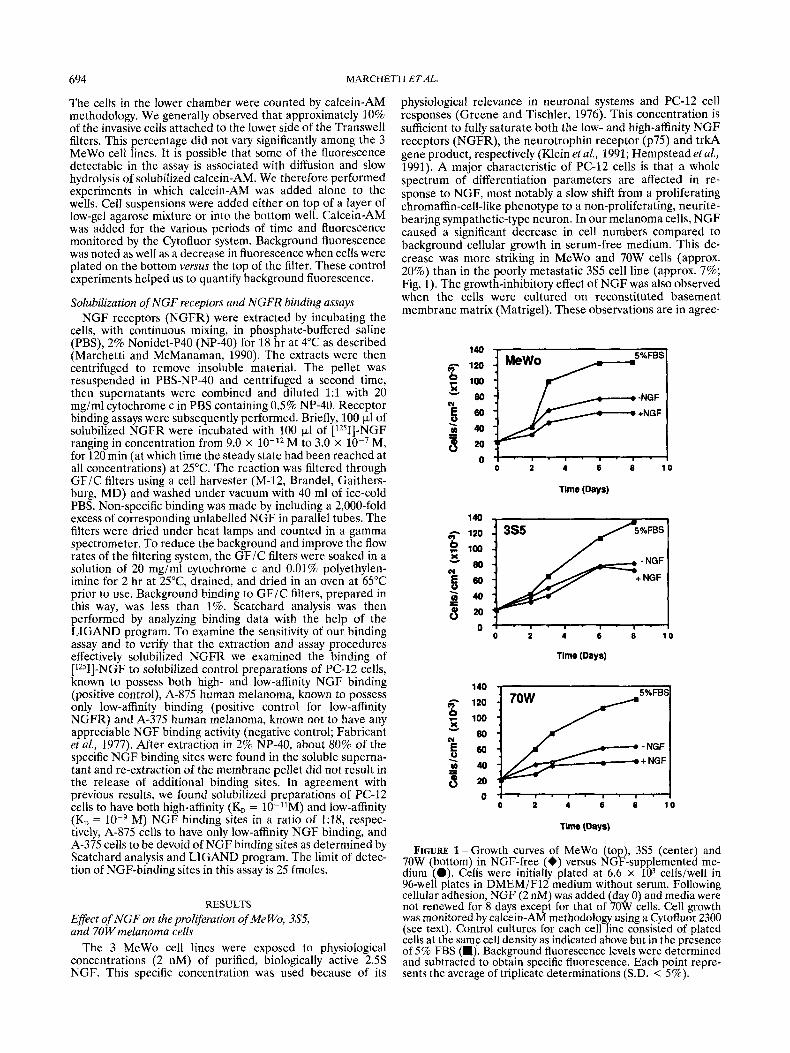
physiological relevance in neuronal systems and PC-12 cell responses (Greene and Tischler, 1976). This concentration is sufficient to fully saturate both the low- and high-affinity NGF receptors (NGFR), the neurotrophin receptor (p75) and trkA gene product, respectively (Klein et al., 1991; Hempstead et al., 1991). A major characteristic of PC-12 cells is that a whole spectrum of differentiation parameters are affected in re- sponse to NGF, most notably a slow shift from a proliferating chromaffin-cell-like phenotype to a non-proliferating, neurite- bearing sympathetic-type neuron. In our melanoma cells, NGF caused a significant decrease in cell numbers compared to background cellular growth in serum-free medium. This de- crease was more striking in MeWo and 70W cells (approx. 20%) than in the poorly metastatic 3S5 cell line (approx. 7%; Fig. 1). The growth-inhibitory effect of NGF was also observed when the cells were cultured on reconstituted basement membrane matrix (Matrigel). These observations are in agree-
S%FBS
-NGF
+NGF
I I . I . I ' I . 1 .
0 2 4 6 8 1 0
Time (Days)
5XFBS
I 1 . I . t . I . I .
0 2 4 6 8 10
Time (Days)
FIGURE 1-Growth curves of MeWo (top), 3% (center) and 70W (bottom) in NGF-free (+) versus NGF-supplemented me- dium (.). Cells were initially plated at 6.6 x lo3 cells/well in 96-well plates in DMEM/F12 medium without serum. Following cellular adhesion, NGF (2 nM) was added (day 0) and media were not renewed for 8 days except for that of 70W cells. Cell growth was monitored by calcein-AM methodology using a Cytofluor 2300 (see text). Control cultures for each cell line consisted of plated cells at the same cell density as indicated above but in the presence of 5% FBS (m). Background fluorescence levels were determined and subtracted to obtain specific fluorescence. Each point repre- sents the average of triplicate determinations (S.D. < 5%).
NGF AND MELANOMA CELL INVASION 695
ment with the notion that NGF is not acting as a mitogen capable of stimulating the growth of MeWo melanoma cells.
In addition to studying motility and invasive properties of melanoma cells in response to NGF treatment, we determined whether NGF affected some of the differentiation parameters of the melanocytic cell lineage seen in melanoma, such as altered adhesive properties of these cells in response to their exposure to NGF and possible effects of N G F on melanin production (Miner et al., 1982). Despite several attempts, we did not observe significant NGF-mediated changes in cell morphology, nor in the adhesive properties of these cells to various matrix components (data not shown). We also found no changes in melanin content when cells where exposed to physiological levels of NGF (data not shown).
Enhancement of in vitro invasive capacities of melanoma cells by NGF
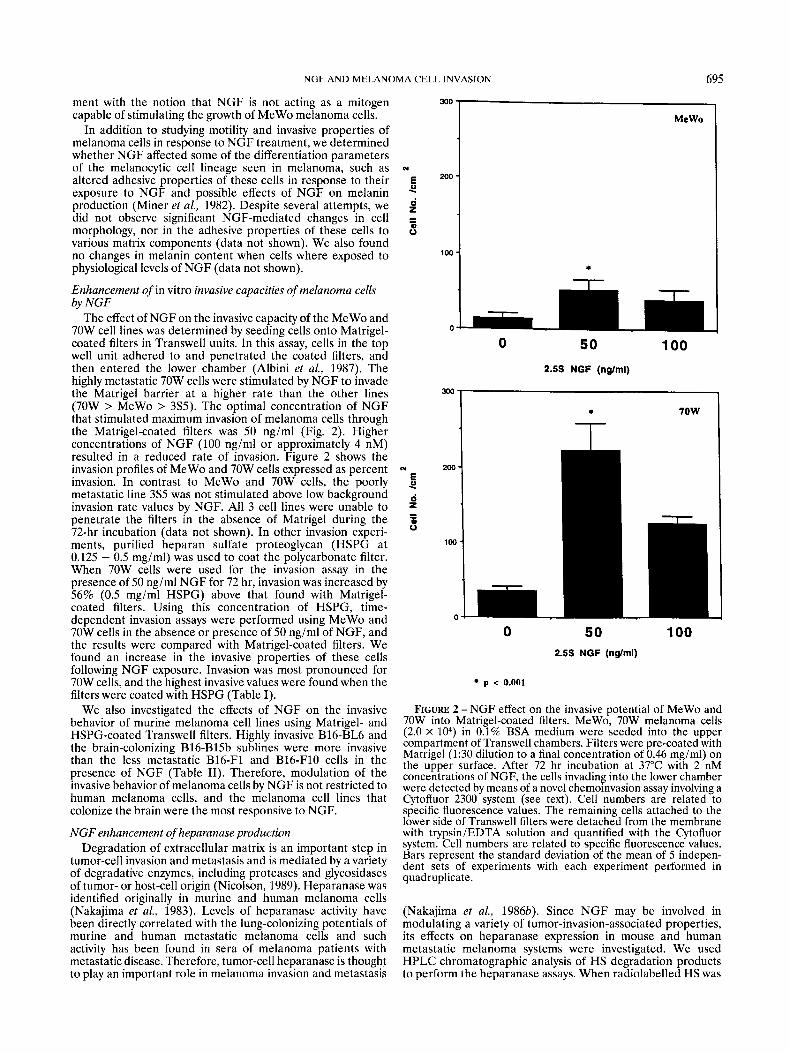
The effect of NGF on the invasive capacity of the MeWo and 70W cell lines was determined by seeding cells onto Matrigel- coated filters in Transwell units. In this assay, cells in the top well unit adhered to and penetrated the coated filters, and then entered the lower chamber (Albini et al., 1987). The highly metastatic 70W cells were stimulated by NGF to invade the Matrigel barrier a t a higher rate than the other lines (70W > MeWo > 3S5). The optimal concentration of NGF that stimulated maximum invasion of melanoma cells through the Matrigel-coated filters was 50 ng/ml (Fig. 2). Higher concentrations of NGF (100 ng/ml or approximately 4 nM) resulted in a reduced rate of invasion. Figure 2 shows the invasion profiles of MeWo and 70W cells expressed as percent invasion. In contrast to MeWo and 70W cells, the poorly metastatic line 3S5 was not stimulated above low background invasion rate values by NGF. All 3 cell lines were unable to penetrate the filters in the absence of Matrigel during the 72-hr incubation (data not shown). In other invasion experi- ments, purified heparan sulfate proteoglycan (HSPG at 0.125 - 0.5 mg/ml) was used to coat the polycarbonate filter. When 70W cells were used for the invasion assay in the presence of 50 ng/ml N G F for 72 hr, invasion was increased by 56% (0.5 mg/ml HSPG) above that found with Matrigel- coated filters. Using this concentration of HSPG, time- dependent invasion assays were performed using MeWo and 70W cells in the absence or presence of 50 ng/ml of NGF, and the results were compared with Matrigel-coated filters. We found an increase in the invasive properties of these cells following NGF exposure. Invasion was most pronounced for 70W cells, and the highest invasive values were found when the filters were coated with HSPG (Table I).
We also investigated the effects of NGF on the invasive behavior of murine melanoma cell lines using Matrigel- and HSPG-coated Transwell filters. Highly invasive B16-BL6 and the brain-colonizing B16-Bl5b sublines were more invasive than the less metastatic B16-F1 and B16-Fl0 cells in the presence of NGF (Table 11). Therefore, modulation of the invasive behavior of melanoma cells by NGF is not restricted to human melanoma cells, and the melanoma cell lines that colonize the brain were the most responsive to NGF.
NGF enhancement of heparanase production Degradation of extracellular matrix is an important step in
tumor-cell invasion and metastasis and is mediated by a variety of degradative enzymes, including proteases and glycosidases of tumor- or host-cell origin (Nicolson, 1989). Heparanase was identified originally in murine and human melanoma cells (Nakajima et al., 1983). Levels of heparanase activity have been directly correlated with the lung-colonizing potentials of murine and human metastatic melanoma cells and such activity has been found in sera of melanoma patients with metastatic disease. Therefore, tumor-cell heparanase is thought to play an important role in melanoma invasion and metastasis
300
MeWo
200
100
* I
0 * 0 50
2.5s NGF (nglml)
100
t
-r 70W
0 50 100 2-55 NGF (ng/ml)
p c o m
FIGURE 2 - NGF effect on the invasive potential of MeWo and 70W into Matrigel-coated filters. MeWo, 70W melanoma cells (2.0 X lo4) in 0.1% BSA medium were seeded into the upper compartment of Transwell chambers. Filters were pre-coated with Matrigel (1:30 dilution to a final concentration of 0.46 mg/ml) on the upper surface. After 72 hr incubation at 37°C with 2 nM concentrations of NGF, the cells invading into the lower chamber were detected by means of a novel chemoinvasion assay involving a Cytofluor 2300 system (see text). Cell numbers are related to specific fluorescence values. The remaining cells attached to the lower side of Transwell filters were detached from the membrane with trypsin/EDTA solution and quantified with the Cytofluor system. Cell numbers are related to specific fluorescence values. Bars represent the standard deviation of the mean of 5 indepen- dent sets of experiments with each experiment performed in quadruplicate.
(Nakajima et al., 19866). Since N G F may be involved in modulating a variety of tumor-invasion-associated properties, its effects on heparanase expression in mouse and human metastatic melanoma systems were investigated. We used HPLC chromatographic analysis of HS degradation products to perform the heparanase assays. When radiolabelled HS was
696 MARCHETTI ET AL.
TABLE I - TIME-DEPENDENT INVASION OF MATRIGEL- OR HSPG-COATED FILTERS IN THE ABSENCE OR PRESENCE OF NGF
Matrigel HSPG p values p values (0.5 mgiml) Cell Time (0.46 mgiml)
+NGF - NGF +NGF *we (hr) -NGF
~ ~~
MeWo 0 1.00 f 0.01’ 1.60 f 0.01 - 1.00 2 0.01 1.50 t 0.01 - 24 1.20 k 0.10 1.86 -t- 1.14 < O . l 1.20 t 0.07 1.64 f 0.19 <0.25 48 1.10 k 0.22 2.30 k 0.06 <0.001 1.022 0.19 2.24 f 0.21 <1.07 72 1.30 _t 0.06 2.75 f 0.16 <0.01 1.30 2 0.23 2.80 2 0.12 <0.001
n = 42 n = 4 -NGF +NGF -NGF +NGF
70W 0 1.00 f 0.01 1.00 f 0.01 - 1.00 2 0.01 1.00 f 0.01 - 24 1.20 t 0.05 5.52 t 0.07 <0.1 1.08 2 0.06 3.51 t 0.01 <0.001 48 1.24 t 0.01 7.16 t 0.01 < O . O O l 1.18 2 0.01 7.18 * 0.02 <O.OOl 72 1.12 t 0.03 8.84 f 0.09 <0.01 1.16 2 0.05 13.79 t 0.04 <0.001
n = 4 n = 4
’MeWo AFU (Arbitrary Fluorescence Units) at time 0 considered equal to 1.00.-2n, Data displayed are means of 4 independent experiments with quadruplicate assays performed in each experiment.
TABLE 11 - INVASION O F MATRIGEL- OR HSPG-COATED FILTERS IN MURINE MELANOMA CELLS: EFFECTOF NGF
cell Matrigel (0.46 mgiml) HSPG (0.50 mgiml) type -NGF +NGF -NGF +NGF
B16-FI 1.00 0.03’ 1.00 t 0.01 1.00 _t 0.01 1.00 t 0.02 B16-FlO 1.25 2 0.09 1.10 k 0.12 1.05 t 0.19 1.15 f 0.15 B I ~ - B L ~ 0.68 k 0.08 3.41 f o x * * 0.67 2 0.13 3.59 2 o.71* B16-BI5b 2.27 t 0.56 8.98 f 0.33** 2.15 f 0.46 7.02 t 0.66**
n = 22 n = 22 ~~
‘B16-Fl AFU at time 0 considered equal to 1.00.-2n, data displayed are means of 2 independent experiments with quadrupli- cate assays performed in each experiment.-*p < 0.06; **p < 0.001.
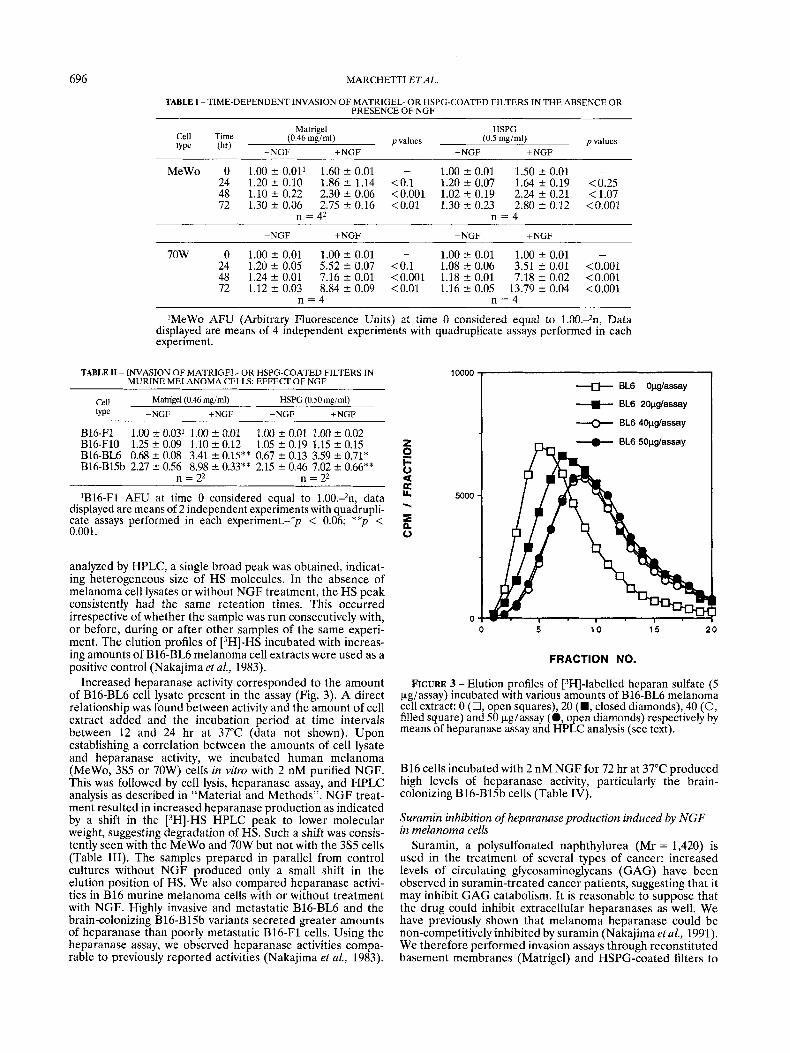
analyzed by HPLC, a single broad peak was obtained, indicat- ing heterogeneous size of HS molecules. In the absence of melanoma cell lysates or without NGF treatment, the HS peak consistently had the same retention times. This occurred irrespective of whether the sample was run consecutively with, or before, during or after other samples of the same experi- ment. The elution profiles of [3H]-HS incubated with increas- ing amounts of B16-BL6 melanoma cell extracts were used as a positive control (Nakajima et al., 1983).
Increased heparanase activity corresponded to the amount of B16-BL6 cell lysate present in the assay (Fig. 3). A direct relationship was found between activity and the amount of cell extract added and the incubation period at time intervals between 12 and 24 hr at 37°C (data not shown). Upon establishing a correlation between the amounts of cell lysate and heparanase activity, we incubated human melanoma (MeWo, 3% or 70W) cells in vitro with 2 nM purified NGF. This was followed by cell lysis, heparanase assay, and HPLC analysis as described in “Material and Methods”. NGF treat- ment resulted in increased heparanase production as indicated by a shift in the [3H]-HS HPLC peak to lower molecular weight, suggesting degradation of HS. Such a shift was consis- tently seen with the MeWo and 70W but not with the 3S5 cells (Table 111). The samples prepared in parallel from control cultures without NGF produced only a small shift in the elution position of HS. We also compared heparanase activi- ties in B16 murine melanoma cells with or without treatment with NGF. Highly invasive and metastatic B16-BL6 and the brain-colonizing B16-Bl5b variants secreted greater amounts of heparanase than poorly metastatic B16-F1 cells. Using the heparanase assay, we observed heparanase activities compa- rable to previously reported activities (Nakajima et al., 1983).
10000
z c 0 U R U 5000
P
0
* EL6 Opglassay
--)- BL6 2Opglassay
+ EL6 4Opglassay
FRACTION NO.
FIGURE 3 - Elution profiles of [3H]-labelled heparan sulfate (5 &assay) incubated with various amounts of B16-BL6 melanoma cell extract: 0 (0, open squares), 20 (W, closed diamonds), 40 (0, filled square) and 50 kg/assay (0, open diamonds) respectively by means of heparanase assay and HPLC analysis (see text).
B16 cells incubated with 2 nM NGF for 72 hr at 37°C produced high levels of heparanase activity, particularly the brain- colonizing B16-Bl5b cells (Table IV).
Suramin inhibition of heparanase production induced by NGF in melanoma cells
Suramin, a polysulfonated naphthylurea (Mr = 1,420) is used in the treatment of several types of cancer: increased levels of circulating glycosaminoglycans (GAG) have been observed in suramin-treated cancer patients, suggesting that it may inhibit GAG catabolism. It is reasonable to suppose that the drug could inhibit extracellular heparanases as well. We have previously shown that melanoma heparanase could be non-competitively inhibited by suramin (Nakajima et aZ., 1991). We therefore performed invasion assays through reconstituted basement membranes (Matrigel) and HSPG-coated filters to

NGF AND MELANOMA CELL INVASION 697
TABLE 111 -HEPARIN SULFATE DEGRADATION ACTIVITY IN HUMAN MELANOMA CELLS
Heparan sulfate degradation activity
-NGF +NGF Cells’ (Mean cpm t SD)
3% 1,409 k 140 1,828 f 235 MeWo 1,978 f 114 6,008 f 187* 70W 1.397 f 127 4.696 f 146*
Tells were exposed to NGF (2 nM) for 72 hr at 37 “C. Data shown represent subtraction of control values (no cellular extract in HS assay) from data of test samples.-*p < 0.01 for n = 4 separate experiments.
TABLE N - HEPARANASE ASSAY REVERSE-PHASE HPLC RETENTION TIMES
Mean difference in % increase in Initial mean retention time retention time
Cell type retention time following NGF following NGF (sec) L SD treatment treatment
fsec) k SD i SD
B16-F1 157.6 f 5.6 0 0 B16-Fl0 168.5 f 24.3 0 0 B16-BL6 189.1 f 6.7 10.9 f 18.7* 5.6 f 9.6* B16-Bl5b 205.4 f 23.0 36.6 k 8.6** 18.0 k 4.4**
*p < 0.5 for n = 3 experiments when comparing k NGF treatment.-**p < 0.02 for n = 3 experiments when comparing k NGF treatment.
determine whether the observed NGF-dependent increase in invasion of B16 cells, especially the brain-metastatic line B16-B15b, was due to NGF-stimulation of heparanase produc- tion that could be blocked with suramin. Control B16-F1 and B16-Bl5b cells were treated with different concentrations of suramin in the absence or presence of 2 nM NGF, seeded onto Matrigel- or HSPG-coated filters, and allowed to invade the coated filters. The results from 2 independent experiments indicated that the invasion process was effectively inhibited by suramin at concentration above 1.5 pM (Table V). Interest- ingly, the presence of both suramin and NGF in the upper Transwell chambers did not result in an increase in cell penetration through the matrix layer with either Matrigel- or HSPG-coated filters. These results further demonstrate that the stimulation of heparanase activity is one of the causes of extracellular matrix degradation by melanoma cells. Addi- tional causes could be the regulation by NGF of expression of melanoma matrix metalloproteinases such as type-IV collage- nases (Hermann et al., 1993).
NGF binding activities in solubilized preparations of human and murine melanoma cells
The binding characteristics of NGFR in human melanoma cells were studied in solubilized preparations using a filtration assay procedure (see “Material and Methods”). There are 2 known receptors for NGF, a low-affinity (KO approx. M or Type-I1 NGFR) receptor class, thought to be a no-signal- transducing receptor, and a high-affinity (KO approx. M or Type-I NGFR) signal-transducing receptor class (Hemp- stead et al., 1991; Klein et al., 1991; Marchetti and McMana- man, 1990). We considered the possibility that the greater degree of matrix invasion responsiveness exhibited by the 70W variant subline after 2.5s NGF treatment resulted from increased expression of the low-affinity class (p75 NGFR) and/or the high-affinity class corresponding to the tyrosine kinase proto-oncogene product pl4OtrkA (p140trkA NGFR; Hempstead et al., 1991; Klein et al., 1991). To examine this possibility, NGF receptors were extracted by incubating the cells of the MeWo and B16 series with continuous mixing in PBS containing 2% NP-40 for 18 hr at PC, and the NGF
receptor binding assays were performed. Several solubilized preparations from other cells were used as positive and negative controls: (a) A875 human melanoma cells that are known to express only the low-affinity NGFR (Fabricant et al., 1977), (b) A375 human melanoma cells known to be negative for NGFR content (Fabricant et al., 1977) and (c) PC-12 rat pheochromocytoma cells well known to possess both low- and high-affinity NGF binding sites (Halegoua et al., 1991). Scatchard analyses using the LIGAND program revealed the presence of defined NGF binding sites as expected in the control cells and, most important, the presence of both low- (Type-11) and high-affinity binding sites (Type-I NGFR) in the MeWo and the brain-colonizing 70W and B16-Bl5b cells. Detectable NGF binding in the Scatchard analysis was not observed for 3S5 and B16-F1 cell lines (Table VI). The Scatchard plots for MeWo and the highly invasive, brain metastatic 70W and B16-Bl5b cells were biphasic, approach- ing linearity at both extremes of [1251]-NGF concentrations. The K,, values and the number of NGF binding sites per cell, as estimated for all the human and murine melanoma cells considered in this study, are shown in Table VI.
DISCUSSION
It is thought that metastasis is dependent on certain host properties, particularly host organ microenvironments and immunologic and non-immunologic responses against tumor cells. One property that should be important to metastatic cell colonization is the ability to respond to local concentrations of hormones and growth factors at various organ microenviron- mental sites (Nicolson, 1993).
Tumor metastasis to the brain occurs fairly frequently in some cancers, such as melanoma, lung, mammary and gastroin- testinal carcinomas, and some lymphomas. Malignant mela- noma patients have the highest percentage of brain metastases (Posner, 1980). The NGF receptor (at least the p75 or low-affinity NGFR) is abundantly expressed in many mela- noma cell lines (Fabricant et al., 1977), and an augmented NGFR expression occurs at the later stages of melanoma progression (Herlyn, 1990).
We found that exposure of the MeWo human malignant melanoma line and its spontaneous WGA-resistant variants 3S5 and 70W with physiological concentrations of purified, biologically active NGF resulted in: (a) delayed density- dependent inhibition of cellular growth; (b) significant enhance- ment of tumor-cell invasion through reconstituted basement membranes, especially by the brain-metastatic human 70W and mouse B16-Bl5b cells, and (c) enhancement of heparan- ase production. Our data indicate that NGF could be involved in some aspect of melanoma progression to the metastatic phenotype. In agreement with differentiation induced by NGF in neurons (Halegoua et al., 1991), cell-growth studies indicate that NGF acts as a cytostatic or differentiation factor rather than a mitogenic factor in human melanoma cells. Previous results had shown that NGF acts as a survivalldifferentiation factor for melanoma cells (Fabricant et al., 1977) and human melanocytes (Peacocke et al., 1988). It is of interest that cultured neural-crest-derived Schwann cells do not require NGF for growth or survival and do not show morphological changes in response to NGF, yet perineural Schwann cells express NGFR in adult skin, as do rat sciatic nerves after axotomy (Taniuchi et al., 1986). Disruption of contacts be- tween axons and Schwann cells appears to stimulate expres- sion of p75 NGFR (Lemke and Chao, 1988). When Schwann cells regain contact with neurites, NGFR expression is again suppressed. It has been suggested that NGFR may immobilize neurotrophic factors secreted in a paracrine fashion and thus provide a substratum laden with NGF which, in turn, attracts
698 MARCHETTI ETAL.
TABLE V - INHIBITION OF B16 MELANOMA CELL INVASION BY SURAMIN IN THE ABSENCE O R PRESENCE OF NERVE GROWTH FACTOR
Suramin Matrigel (0.46 rnglml) HSPG (0.50 mg/rnl) (CLM) -NGF +NGF % -NGF +NGF %
B16-F1 cells 0 1.5 15 150
B16-Bl5b cells n i.5 15 150
1.00 k 0.03' 0.40 2 0.14
1.06 k 0.05 0.33 ? 0.01*
0 0 0 0
n = 2l
2.87 2 0.01 2.25 2 0.03
11.35 k 0.02 2.05 f 0.05**
0 0 0 0
n = 2
0 31.1
100 100
0 18.0
100 100
0.92 k 0.15 1.00 k 0.08 0 N.D. N.D. N.D.
0 0 100 0 0 100
n = 2
3.15 k 0.15 11.02 k 0.02 0 0.37 k 0.04 1.14 k 0.08*** 10.3
0 0 100 0 0 100
n = 2
'B16-Fl AFU at time 72 hr considered equal to l.OO.-*n, data displayed are means of 2 independent experiments with quadruplicate assays performed in each experiment.-*p < 0.005, **p < 0.07, * * p < 0.001.
TABLE VI - NGF-BINDING PROPERTIES OF HUMAN AND MURINE MELANOMA CELLS
KD value (M) NGFR sitesicell ( x lo3) Cell line
TYPC I Type I1 Type I Type I1 Ratio
MeWo 4.8 fk2.8) x 10-l' 1.1 fk0.2) x lo-' 1.05 k 0.5 5.67 f 1.2 54.0 3s5 ' N.D.1 ' N.D. N.D. N.D. - 70W 4.5 (20.6) x lo-" 5.1 (f0.4) x 29.7 k 3.2 192 f 8.3 6.46 B16-F1 ~ N.D. N.D. N.D. N.D. - B16-Bl5b 2.0 (20.6) x 1.5 (20.3) x lo-' 13.5 2 2.2 150 2 4.7 11.1
n = 22 n = 2 'N.D. = Not detectable.4, Data displayed are the means of 2 independent experiments with
triplicate assays performed in each experiment.
and guides the neuronal growth cone to the proper site of innervation after damage (Taniuchi et al., 1988). These changes may be paralleled by melanoma cells as they progress to more invasive phenotypes.
The NGF effects described here were most pronounced in the brain-colonizing 70W and B16-Bl5b subclones. The in- creased ability of the 70W line to degrade extracellular barriers during the metastatic process may be due, in part, to its sensitivity to physiological levels of NGF (and high NGFR levels; see Hermann et al., 1993) within the CNS. We found contrasting results when poorly metastatic 3S5 cells were incubated with NGF, supporting the notion of the importance of microenvironmentally-induced changes affecting pheno- typic behavior. The 3S5 cells are relatively refractory to such changes, and they are not metastatic to the brain. Thus, the 70W melanoma line may offer an excellent model for studying neurotrophic factors involved in brain metastasis. Similarly, we found that the invasion of murine-brain-colonizing B16 mela- noma cells were most affected by NGF.
We have examined the relationship between metastatic properties and the ability of human and mouse melanoma lines with various invasive characteristics to enzymatically degrade HS. The mechanisms by which tumor cells invade neuronal tissues are not known, but hydrolytic and collagenolytic en- zymes could facilitate tumor invasion. Heparanase plays an important role in melanoma invasion and metastasis (Na- kajima et al., 1983, 1986b). Among the several BM compo- nents, HSPG is of unique importance, because it is a major constituent of endothelial BM (Ruoslahti, 1988). Therefore, destruction of the HSPG barrier by heparanase can be of particular importance in BM invasion by tumor cells.
We found that NGF treatment of the highly invasive 70W cells (but not poorly metastatic 3S5 cells) increased heparan- ase production. Such an effect was not confined to human
melanoma cells and was also observed with highly metastatic, brain-colonizing murine melanoma (B16-Bl5b) cells. NGF has been reported to induce the metalloproteinase stromelysin (MMP-3) in PC-12 cells (Machida et al., 1991). NGF effects on melanoma tumor invasion were not confined to an increased heparanase production: results from our laboratory indicate that NGF treatment results in increased secretion of the 72-kDa type-IV collagenase (in its fully active 68-kDa form) paralleled by an inhibition of the 92-kDa type-IV collagenase. Such an effect was again most pronounced in the 70W cells (Hermann et al., 1993).
Neoplastic cells are capable of responding to their microen- vironments and, although these responses may not be normal, they may mimic normal cell responses that occur during cellular differentiation and development. It is highly unlikely that a single molecule can regulate these responses, particu- larly those involved in tumor invasion and metastasis. Nonethe- less, because of NGF production and its plethora of effects in the brain, it may influence the malignant properties of cancer cells during brain metastasis. NGF and other uncharacterized factors may serve to regulate the invasive capacity of tumor cells in the CNS.
ACKNOWLEDGEMENTS
We thank Mr. D. Kraus, Quality Control Division, Collabo- rative Research, Irvine, CA, for providing data regarding 2.5s NGF purification procedures. We thank Dr. R. Kapoor, Becton Dickinson, for helpful discussions concerning HSPG purification procedures and biology. The secretarial help of Ms. J. North and Ms. L. Small is acknowledged. These studies were supported by grants from the National Institutes of Health (R35-CA 44352) and the National Foundation for Cancer Research (to G.L.N.).

NGF AND MELANOMA CELL INVASION
REFERENCES
699
ALBINI, A., IWAMOTO, Y., KLEINMANN, H.K., MARTIN, G.R., AARON- SON, S.A., KOZLOWSKI, J.M. and MCEWAN, R.N., A rapid in vitro assay for quantitating the invasive potential of tumor cells. Cancer Res., 47, 3239-3245 (1987). FABRICANT, R.N., DE LARCO, J.E. and TODARO, G.L., Nerve growth factor receptors on human melanoma cells in culture. Proc. naf. Acad. Sci. (Wash.), 74,565-569 (1977). GREENE, L.A. and TISCHLER, AS., Establishment of a noradrenergic clonal line of rat adrenal pheochromocytoma cells which respond to nerve growth factor. Proc. naf. Acad. Sci. (Wash.), 73, 2424-2428 (1976). HALEGOUA, S., ARMSTRONG, R.C. and KREMER, N.E., Dissecting the mode of action of a neuronal growth factor. Cur. Top. Microbiol. ImmunoL, 165,119-170 (1991). HEMPSTEAD, B.L., MARTIN, Z.D., KAPLAN, D.R., PARADA, C.F. and CHAO, M., High-affinity NGF binding requires coexpression of the trk proto-oncogene and the low-affinity NGF receptor. Nature (Lond.), 350,678-683 (1991). HERLYN, M., Human melanoma: development and progression. Can- cerMetast. Rev., 9,101-112 (1990). HERMANN, J.H., MENTER, D.G., MARCHETTI, D., HAMADA, J. and NICOLSON, G.L., A functional role for the low-affinity 75 nerve growth factor receptor in human melanoma progression to t l!e invasive phenotype. Mol. Biol. Cell, submitted (1993). ISHIKAWA, M., DENNIS, J.W., MAN, M.S. and KERBEL, R.S., Isolation and characterization of spontaneous wheat germ agglutinin-resistant human melanoma mutants displaying remarkably different metastatic profiles in nude mice. CancerRes., 48,665-670 (1988~). ISHIKAWA, M., FERNANDEZ, B. and KERBEL, R.S., Highly pigmented human melanoma variant which metastasizes widely in nude mice, including to skin and brain. CancerRes., 48,4897-4903 (19886). KLEIN, R., JING, S., NANDURI, V., O’ROURKE, E. and BARBACID, M., The trk proto-oncogene encodes a receptor for nerve growth factor. Cell, 85,189-197 (1991). LEMKE, G. and CHAO, M., Axons regulate Schwann cell expression of the major myelin and NGF receptor genes. Development, 102,499-504 (1988). LEVI-MONTALCINI, R., The nerve growth factor 35 years later. Science, 237,1154-1164 (1987). LIOTTA, L.A., RAO, C.N. and WEWER, U.M., Biochemical interactions of tumor cells with the basement membrane. Ann. Rev. Biochem., 55, 1037-1057 (1986). MACHIDA, C.M., SCOTT, J.D. and CIMENT, G., NGF-induction of the
metalloproteinase transinistromelysin in PC-12 cells: involvement of multiple protein kinases. J. Cell Biol., 114,1037-1048 (1991). MARCHETTI, D. and MCMANAMAN, J.L., Characterization of nerve growth factor binding to embryonic rat spinal cord and neurons. J. Neurosci. Res., 27,211-218 (1990). MINER, K.M., LOTAN, R. and NICOLSON, G.L., Metastatic and melano- genic properties of in vivo selected B16 melanoma sublines and their clonal derivatives. Cancer Res., 42,4631-4638 (1982). NAKAJIMA, M., DECHAVIGNY, A., JOHNSON, C.E., HAMADA, J., STEIN, C.A. and NICOLSON, G.L., Suramin. J. biol. Chem., 266, 9661-9666 (1991). NAKAJIMA, M., IRIMURA, T., DI FERRANTE, D., DI FERRANTE, N. and NICOLSON, G.L., Heparan sulfate degradation: relation to tumor invasive and metastatic properties of mouse B16 melanoma sublines. Science, 220,611-613 (1983). NAKAJIMA, M., IRIMURA, T. and NICOLSON, G.L., A solid phase substrate of heparanase: its application to assay of human melanoma for heparan sulfate degradative activity. Anal. Biochem., 157, 162-171 (1986~). NAKAJIMA, M., IRIMURA, T. and NICOLSON, G.L., Heparanase and tumor metastasis. J. cell. Biochem., 36,157-167 (19866). NICOLSON, G.L., Metastatic tumor cell interactions with endothelium basement membrane and tissue. Curr. Opinion Cell Biol., 1, 1009-1019 (1989). NICOLSON, G.L., Cancer progression and growth: relationship of paracrine and autocrine growth mechanisms to organ preference of metastases. Exp. Cell Res., 204,171-180 (1993). PEACOCKE, M., YAAR, M., MANSUR, C.P., CHAO, M. and GILCHREST, B.A., Induction of nerve growth factor receptors on cultured human melanocytes. Proc. naf. Acad. Sci. (Wash.), 85,5282-5286 (1988). POSNER, J.B., Brain metastasis: a clinician’s view. In: L. Weiss, H.A. Gilber and J.B. Posner (eds.), Brain metastasis, pp. 2-29, Hall, Boston (1980). RUOSLAHTI, E., Structure and biology of proteoglycans. Ann. Rev. Cell Biol., 4,229-255 (1988). TANIUCHI, M., CLARK, H.B. and JOHNSON, E.M., JR., Induction of nerve growth factor receptor in Schwann cells after axotomy. Proc. naf. Acad. Sci. (Wash.), 83,4094-4098 (1986). TANIUCHI, M., CLARK, H.B., SCHWEITZER, J.B. and JOHNSON, E.M., JR., Expression of nerve growth factor receptors by Schwann cells of axotomized peripheral nerves: ultrastructural location, suppression by axonal contact and binding properties. J. Neurosci., 8,664-681 (1988).





























