Embed Size (px)
Citation preview

THE JOURNAL OF COMPARATIVE NEUROLOGY 375303-320 (1996)
Neuropeptides and Neurotrophin Receptor mRNAs in Primary Sensory Neurons of
Aged Rats
ESBJORN BERGMAN, HANS JOHNSON, XU ZHANG, TOMAS HOKFELT, AND BRUN ULFHAKE
Department of Neuroscience, Karolinska Institutet, S-171 77 Stockholm, Sweden
ABSTRACT Neuropeptides and neurotrophin receptors are regulated in primary sensory neurons in
response to axonal injury, and axonal lesions are characteristic stigmata of aging primary sensory neurons. We have therefore examined the expression of neuropeptides and neurotrophin receptor mRNAs in 30-month-old (median survival age) Sprague-Dawley rats to see if similar adaptive mechanisms operate in senescence. The content of neuropeptides was examined with immuno- histochemistry (IHC) and in situ hybridization (ISH), and the cellular mRNA expression of neurotrophin receptors was studied with ISH. All of the aged rats had symptoms of hind limb incapacity (posterior paralysis), but fore limbs did not seem affected. The size-distribution of neuronal profiles in cervical and lumbar dorsal root ganglia (DRGs) was similar in aged and young adult (2-3 months old) rats. In aged rats, the DRG neurons showed an increase in both immunolabelling and mRNA content of neuropeptide tyrosine (NPY), as well as an increased cellular expression of galanin mRNA. In the same animals, there were decreased cellular levels of calcitonin gene-related peptide (CGRP; IHC and ISH) and substance P (SP; IHC and ISH), while the difference in neuronal somatostatin (IHC and ISH) was small. The distribution of neuropeptide immunoreactivities in the dorsal horn of the corresponding spinal cord segments revealed a decreased labelling for CGRP-, SP-, and somatostatin-like immunoreactivities (LI) in the aged rats at both cervical and lumbar levels. NPY- and galanin-LI had a similar distribution in aged and young adult rats. NPY-immunoreactive fibers were also encountered in the dorsal column of aged but not young adult rats. ISH revealed that most of the primary sensory neurons express mRNA for the p75 low-affinity neurotrophin receptor (p75-LANR) and that there was no discernible difference between young adult and aged rats. The labelling intensity for mRNA encoding high-affinity tyrosine kinase receptors (TrkA, TrkB, and TrkC) was decreased in aged rat DRG neurons, wh’le the percentage of neuronal profiles expressing mRNA for TrkA/B/C was similar in young adult and aged rats. The changed pattern of neuropeptide expression in primary sensory neurons of aged rats resembled that seen in young adult rats subjected to axonal injury of peripheral sensory nerves and may, thus, indicate aging-related lesions of sensory fibers. Since NPY is primarily present in large and galanin in small DRG neurons, the stronger effect on NPY as compared to galanin expression may indicate that aging preferentially affects neurons associated with mechanoreception (Aa and A@ fibers) as compared to nociceptive units (A6 and C fibers). Furthermore, the observed changes in neuropeptide expression were most pronounced in lumbar DRGs, that harbors the sensory neurons supplying the affected hindlimbs of the rats. o 1996 Wiley-Liss, Inc.
Indexing terms: aging, axonopathy, neurotrophins, Trk, dorsal root ganglia
Rats show a progressive deterioration of motor behaviour during aging (Berg et al., 1962; van Steenis and Kroes, 1971; Burek et al., 1976; Mitsumori et al., 1981; Krinke,
sensory systems have been considered to contribute to the decline in motor functions seen in aged subjects (see Koller
Accepted June 21, 1996. 1983; Johnson et al., 1993, 19951, with symptoms usually
during the third year Of life and essential1y being Address reprint requests to Dr, Brun Ulfh&e, Department of Neurosci- confined to the hind limbs (“posterior paralysis”). Along ence. Dlv. Neuroanatomv and Neuronal Plasticitv. Karolinska Institutet, with motoneuron-muscle dysfunction, changes in somato- S-17177 Stockholm, Sweden. E-mail: brun.ulfhak& neuro.ki.se
o 1996 WILEY-LISS, INC.

304
et al., 1985; Woollacott, 1986; and references therein). Axon lesions such as dystrophy and degeneration have been reported to occur in both peripheral and central sensory pathways (Berg et al., 1962; van Steenis and Kroes, 1971; Gilmore, 1972; Samorajski, 1974; Sharma et al., 1980; Mitsumori et al., 1981; Krinke, 1983; Mittal and Logmani, 1987; Fujisawa, 1988; Knox et al., 1989).
Primary sensory neurons synthesize neuromodulatory peptides (reviewed by e.g., Dalsgaard, 1988; Willis and Coggeshall, 1991; Lawson, 1992) such as calcitonin gene- related peptide ( - 40-50% of all dorsal root ganglion [DRGJ neurons; CGRP), substance P ( - 20%; SP), somatostatin ( -&lo%), and galanin ( < 10%). The expression of neuro- peptides in sensory neurons can be influenced by several factors. Thus, nerve injury, such as crush or transection, induces a down-regulation of CGRP and substance P, while vasoactive intestinal polypeptide (VIP), galanin, and neuro- peptide tyrosine (NPY) are upregulated (see Hokfelt et al., 1994). This pattern of neuropeptide regulation has been considered important for adaptation to the deficits induced by lesioning sensory neurons in adult rats (see Hokfelt et al., 1994). Although a number of investigations have dealt with axonal lesions of primary sensory neurons in aged subjects (see above), little is known about neuropeptide expression in primary sensory neurons during aging.
In the adult, experimental data indicate that neuro- trophic factors (reviewed by Barde, 1989; Thoenen, 1991, 1995; Korsching, 1993; Lindsay et al., 1994) and their receptors (e.g., reviews by Chao et al., 1992; Meakin and Shooter, 1992; Ip et al., 1993; Lindsay et al., 1994) influence neuronal phenotype, neuronal regeneration, and degenera- tive processes (Brewster et al., 1994; Lindsay et al., 1994; Lindvall et al., 1994; Rylett and Williams, 1994). With regard to sensory neurons, it has been demonstrated that axonal injury also influences expression of neurotrophin receptors (Verge et al., 1989, 1992). Moreover, several studies have shown that the expression of a number of neuropeptides such as CGRP, SP, and NPY in sensory neurons can be influenced by neurotrophic factors (e.g., Goedert et al., 1981; Schwartz et al., 1982; Fitzgerald et al., 1985; Lindsay and Harmar, 1989; Wong and Oblinger, 1991; Jones et al., 1994; Mulderry, 1994; Verge et al., 1995; Watson, 1995). Based on these evidences, it has been suggested that the changes in neuropeptide expression seen in sensory neurons following axonal lesions may at least in part be the result of decreased neurotrophic signalling.
Taken together, it is well established that the expression of both neuropeptides and neurotrophin receptors are regulated in response to axonal injuries of sensory neurons in young adult rats. Against the background that axonal lesions are characteristic stigmata of aging in primary sensory pathways, we have investigated the expression of neuropeptides and neurotrophin receptor mRNAs in sen- sory neurons of aged rats, to see if similar adaptive mecha- nisms operate in senescence. Thus, we have examined the labelling for CGRP, SP, somatostatin, galanin, and NPY in cervical and lumbar DRGs as well as in the dorsal horn of the corresponding spinal cord segments by using immuno- histochemistry (IHC). The cellular expression of mRNA for these peptides in DRGs was investigated by use of in situ hybridization (ISH). In addition, the mRNA expression in DRG neurons of the p75 low-affinity neurotrophin receptor (p75-LANR) and the high-affinity tyrosine kinase receptors (TrkA, TrkB, and TrkC) was analysed.
E. BERGMAN ET AL.
MATERIALS AND METHODS Experimental animals
Ten aged (30-months-old) male Sprague-Dawley rats (SD; strain: Bkl: SD; bwt 550-650 g) and ten male young adult rats (same strain; 2-3-months-old; bwt 200-250 g) were used. The rats were delivered by a local breeder (B&K, Stockholm, Sweden) at an age of 2 months and thereafter kept under standardized barrier conditions at our depart- ment, on a 12 hour day/ 12 hour night cycle and ad libitum access to water and food (Lactaminm, with reduced protein
h I Cervical DRGs
z 0 500 1000 1500 2000 Cell profile cross-sectional area (pm2) A
h .cI I Lumbar DRGs b' w 20
% 10 Young adult
a 2 0
h 10
rA W CI
L CI CI
U
iu 0
W .a
20 7 7
I I I I u 0 500 Cell profile cross-sectional area (pm2)
c
1000 1500 2000 B Fig. 1. Size frequency histograms showing the distribution of cell
profile cross-sectional areas in cervical (A) and lumbar (B) dorsal root ganglia (DRGs) of both young adult and aged rats. Contingency table analysis yielded x2 = 32.9 (cervical DRGs; P < 0.05) and x2 = 25.2 (lumbar DRGs; P = n.s.1, respectively, indicating that the cell profile distribution was similar in the two age groups. A total of 2,300 to 3,400 cell profiles were measured in each age category and on each level.

PRIMARY SENSORY NEURONS IN AGED RATS 305
perfused transcardially with warm (37”C), Ca2+-free Ty- rode’s solution followed by cold fixative (4°C) containing 4% wlv paraformaldehyde and 0.2% wlv picric acid in 0.1 M phosphate-buffered saline (PBS; pH 7.2) (Pease, 1962; Zamboni and DeMartino, 1967) for 6 minutes. The spinal cord segments and DRGs were quickly dissected out, im- mersed in fresh fixative for 90 minutes, and then stored in 10% sucrose buffer solution containing 0.01% sodium azide (Sigma, St. Louis, MO) and 0.02% Bacitracin (Bayer, Leverkusen, Germany) overnight at 4°C. The spinal cord (cervical and lumbar segments) and DRGs (cervical and lumbar) were cut at 12-14 km thickness in a cryostat (Dittes, Heidelberg, Germany) and thawed onto gelatin/ chrome-alum coated slides. The sections were then pro- cessed according to the indirect immunofluorescence tech- nique of Coons and collaborators (see Coons, 1958). Briefly, the sections were rehydrated in PBS and then incubated for 18 or 72 hours at 4°C in a humid chamber with the following primary antiseralantibodies, diluted in PBS con-
content). Under these conditions, the median life span is 30 months ( 2 2 months across cohorts) and the maximum life-span is >36 months (see also, Gutman and Hanzli- kowa, 1972; Burek and Hollander, 1980; Masoro, 1980; Algeri et al., 1983) for both males and females. Based on this, the 30-month-old rats are defined as “aged.” All aged rats used in this study disclosed clinical signs of hind limb incapacities, described in detail elsewhere (see Johnson et al., 1993,1995). Briefly, the symptoms were confined to the hind limbs with increased stride-width, adduction insufi- cience, weakness, muscle atrophy, decreased or obsolete placing reaction, and in the most severe cases a complete hind limb paralysis. All experiments were approved by the Local Ethical Committee (Stockholms Norra Djurforsok- setiska Namnd; proj. no. 75/93).
Immunofluorescence histochemistry Seven aged and seven young adult rats were deeply
anaesthetized with chloral hydrate (300 mg kg-I i.p.1 and
Fig. 2. Confocal images of aged (Ag) ( A X ) and young adult (Ad) (D) rat lumbar DRGs after incubation with neuropeptide tyrosine (NPY) antiserum. In young adult rats no NPY-immunoreactive (NPY- IRj cell profiles were observed (D), hut NPY positive fibres were seen (arrows in D). In aged rats a moderate number of NPY-IR DRG neurons, mainly of medium or large size, were encountered (C). To remove autofluorescence contribution to the images, a confocal micro- scope with dual detectors was used. The fluorescein isothiocyanate
(FITCj-labelled specimen was excited with the 488-nm line from an argon ion laser, and the fluorescence emission light was collected in the dual detector system. Each recording thus resulted in two images, one containing the FITC-fluorescence and autofluorescence (A), and the other containing autofluorescence (B). The autofluorescence (arrow- heads in A and Bj was then removed by subtraction of image B from image A, to obtain an image devoid of autofluorescence (arrowheads in C). (For details see Johnson et al., 1995.) Scale bar = 50 pm.

306 E. BERGMAN ET AL.
Figure 3

PRIMARY SENSORY NEURONS IN AGED RATS 307
or sheep anti-mouse (1:lO; Amersham Ltd., Amersham, UK) antibodies. The sections were rinsed in PBS, mounted in glycerol/PBS (3: 1) containing 0.1% P-phenylenediamine in order to retard fading (Johnson and de C Nougueira Araujo, 1981; Platt and Michael, 1983) and examined in either a Nikon Microphot-FX epifluorescence microscope equipped with the proper filters for FITC-fluorescence (filter cube B-2A; 510 nm dichroic mirror; 450-490 nm excitation filter; 520-560 nm barrier filter, and 550 nm extra barrier shortpass filter), or a Sarastro 1000 dual- detector confocal microscope (CLSM). As described else- where (Johnson et al., 19951, the dual-detector system allows separation of specific fluorescence from that deriving from lipofuscin granulae. Photomicrographs were taken with black-and-white Kodak Tri-X film (400 ASA) in the epifluorescence microscope, and digital images recorded in the CLSM were printed on a Kodak printer.
The specificity of the respective antiserum/antibody has been discussed elsewhere, and control experiments con- ducted by preabsorption of the antisera with the correspond- ing peptide at M resulted in no detectable labelling of the sections (Zhang et al., 1993a).
Fig. 4. Immunofluorescence photomicrographs of young adult (Ad) (A) and aged (Ag) (B) rat cervical spinal cord showing the dorsal columns after incubation with NPY antisera. The young adult rats disclosed no NPY-LI in the dorsal column (A), whereas in the aged rats numerous NPY-IR profiles were found in the lateral portion of the gracile fasciculus (arrow in B). Arrowhead points dorsally. Scale bar = 100 bm.
taining 0.3% Triton X-100 (Hartman et al., 19721, sodium azide (0.01%), and Bacitracin (0.02%): (1) rabbit anti-CGRP antiserum (1:400; Orazzo et al., 1992); (2) rabbit anti-SP antiserum (1:400; Christensson-Nylander et al., 1986); (3) rabbit anti-galanin antiserum (1:400; Peninsula Laborato- ries, Belmont, CA); (4) rabbit anti-NPY antiserum (1:400; Lundberg et al., 1984); (5) mouse anti-somatostatin mono- clonal antibodies (1:250; Buchan et al., 1985; kindly pro- vided by Dr. J. Brown and Dr. A. Buchan). After incubation with the primary antiseralantibodies, the sections were rinsed several times in PBS, transferred to a humid cham- ber, and incubated at 37°C for 30 minutes with fluorescein isothiocyanate (F1TC)-conjugated goat anti-rabbit (1230; Boehringer Mannheim Scandinavia, Stockholm, Sweden)
Fig. 3. Immunofluorescence photomicrographs of young adult (Ad) (A, C, E, G, I) and aged (Ag) (B, D, F, H, J) rat lumbar dorsal horn after incubation with NPY (A, B), galanin (GAL) (C, D), somatostatin (SOM) (E, F), substance P (SP) (G, HI, and calcitonin gene-related peptide (CGRP I, J) antisera. NPY-LI was found in a fairly dense fibre plexus in laminae I and I1 of both age groups; (A, B). Note the slightly increased fibre density in the deeper laminae of the aged rat (arrow in B). Abundant galanin-IR fibres were found in the superficial dorsal horn with no major difference between the young adult and aged rats (C, D). A very dense fibre plexus with somatostatin-11 was observed in laminae 1 and I1 of the young adult rats (El, a marked reduction in the number of somatostatin-IR fibres was found in the superficial dorsal horn of the aged rats (F). In young adult rats both substance P and CGRP immunoreactive fibres were found in large numbers in laminae I and I1 of the dorsal horn, with gradually lower numbers in deeper laminae (G, I). The aged rats disclosed, with regard to both substance P and CGRP, a lower fibre density in the superficial dorsal horn (H, J). Scale bar = 150 pm.
In situ hybridization histochemistry Three deeply anaesthetized animals in each age group
were sacrificed by decapitation. The cervical and lumbar DRGs were rapidly removed and frozen on dry ice. Serial transverse sections (12 Fm) were cut in a cryostat and thaw-mounted onto “Probe on” slides (Fisher Scientific, Pittsburgh, PA) and stored in sealed boxes at -20°C until hybridization. To reduce variability, cervical and lumbar DRGs from both age groups were mounted on the same slide.
Oligonucleotide probes were synthesized by Scandina- vian Gene Synthesis AB (Koping, Sweden). The sequences were complementary to a-CGRP (bp 664-698; Amara et al., 1985), substance P (bp 145-192; Krause et al., 19871, somatostatin (bp 351-399; Goodman et al., 19831, NPY (bp 1671-1714; Larhammar et al., 1987), galanin (bp 152-199; Vrontakis et al., 19871, p75-LANR (bp 765-812; Radeke et al., 1987), TrkA (bp 1025-1075; Martin-Zanca et al., 19861, TrkB (bp 2576-2617; Middlemas et al., 19911, and TrkC (bp 1189-1239; Lamballe et al., 1991). The TrkB probe detects mRNA encoding the full length TrkB receptor, and not the truncated forms. The oligonucleotides were labelled at the 3’-end with C X - ~ ~ S - ~ A T P (New England Nuclear, Boston, MA) by using terminal deoxynucleotidyltransferase (IBI, New Haven, CT) in a buffer containing 10 mM CoC12, 1 mM dithiothreitol (DTT; LKB; Bromma, Sweden), 300 mM Tris base, and 1.4 M potassium cacodylate (pH 7.2). The label- ling mixture was incubated for 3 hours and then purified through NENSORB-20 columns (New England Nuclear). DTT was added to a final concentration of 10 mM. The specific activity of the radiolabelled probes was 6-9 x los cpm Fg-l.
Our hybridization procedure followed previously de- scribed protocols (see Young, 1990; Dagerlind et al., 1992). Without any pretreatment the sections were thawed, put in humidified boxes, and incubated for 16-18 hours at 42°C with lo6 cpm of the radiolabelled probe per 100 ~1 of a hybridization solution containing: 50% formamide (J.T. Baker Chemicals BW, Deventer, The Netherlands); 4 x SSC (1 x SSC = 0.15 M sodium chloride and 0.015 M sodium citrate); 1 x Denhardt’s solution (0.02% each of polyvinyl pyrrolidone, Ficoll and bovine serum albumin);

Fig. 5. Darkfield (A-F) and brightfield (G, H) photomicrographs of young adult (Ad) (A, C, EJ and aged (AgJ (B, D, F, G, H) rat cervical (A, B, G), and lumbar (C, D, E, F, H) DRGs hybridized with oligonucleotide probes complementary to mRNA encoding NPY (A-D) and galanin (GAL,) (E-H). In young adult rats, only occasional weakly NPY mRNA-positive neurons were observed (A, C) whereas no galanin expressing cells could be detected (EJ. In the aged rats, significantly
more DRG cells expressed NPY mRNA, at higher levels, mainly in medium-sized and large cells (B, D). A rostrocaudal gradient was found with more positive neurons at the lumbar level. A weak galanin signal could be detected in a low number of cells in both cervical (G) and lumbar (F, H) DRGs in the aged rat. Arrowheads (G, H) point to small, arrows (G, H) point to medium-sized or large neurons. Scale bars = 150 pm (A-F), 50 pm (G, HI.

PRIMARY SENSORY NEURONS IN AGED RATS 309
90% sequence homology. The probes were aligned to a sequence databank (Gen Bank), and no mRNA was found with more than 80% sequence homology. Hybridization with addition of an excess of unlabelled probe ( 1 0 0 ~ ) resulted in abolished signal. The oligonucleotides used have been employed successfully in other studies (see e.g., Zhang et al., 1993a; Wetmore and Olson, 1995).
1% Sarcosyl (N-lauryl sarcosine, Sigma); 0.02% sodium phosphate (pH 7.0); 10% dextran sulphate (Pharmacia, Uppsala, Sweden); 500 p.g ml-l sheared and heat denatur- ated salmon testis DNA (Sigma); 250 p.g ml-1 yeast tRNA (Sigma), and 200 mM DTT. Following hybridization, sec- tions were rinsed 4 x 15 minutes in 1 x SSC at 56"C, brought to room temperature in the final rinse, dipped twice in distilled water, transferred through ascending concentrations of ethanol, and air dried. Thereafter NTB2 nuclear track emulsion (Kodak), diluted 1: 1 in distilled water, was applied by dipping followed by 3-5 weeks of exposure in the dark at 4°C. The slides were developed in D-19 (Kodak) for 3 minutes, fixed in Kodak 3000 A&B for 7 minutes, rinsed in distilled water, and mounted with glycerol. Some sections were after the initial analysis including densitometry of silver grains, counterstained with Toluidine Blue and coverslipped with Entellan (Merck, Darmstadt, Germany). The sections were examined in a Nikon Microphot-FX microscope equipped for bright- and darkfield illumination. All photomicrographs were taken with Kodak Technical Pan black-and-white film.
The hybridization stringency conditions used here do not allow cross-reaction to any related mRNA with less than
Quantitative analyses To determine changes in the labelling intensity for
mRNA encoding the different neuropeptides and Trk (A, B, C) receptors, 8 to 13 DRG sections from each spinal cord level and age group were analyzed. The data were collected using a Nikon microphot-FX microscope with a x 10 objec- tive lens, equipped with a DAGE-MTI CCD-72 series cam- era, and connected to a Macintosh IIx computer. To process images, Image version 1.52 (courtesy of Dr. Wayne Ras- band, National Technical Information Services, NIH, Bethesda, MD) was used. The sensitivity of the intensity scale (256 grey levels) was adjusted in order to prevent pixels from becoming saturated. The silver grain density over individual labelled cell profiles were recorded by using
Fig. 6. Confocal images of young adult (Ad) (A, C ) and aged (Ag) (B, D) rat lumbar DRGs after incubation with CGRP (A, B) and SP (C, D) antisera. In the young adult rats CGRP was expressed in a large number of cells (A), whereas SP-LI was found in a moderate number of cells (C). A significant reduction in the number of positive cell profiles
was observed in the aged rats for both CGRP (B) and SP (D). With regard to the distribution of positive cell profiles on different cell size categories or the subcellular localization of the immunoreactivities, no difference was observed between the age groups. Scale bar = 50 pm.

310 E. BERGMAN ET AL.
a window measuring 10 pm x 10 pm. The optical density obtained for each labelled cell profile was corrected by subtracting the value for the surrounding background labelling and image noise measured over an area devoid of labelled cells in the corresponding region. A cell profile was considered positive, if the corrected optical density exceeded the background with at least 100%. The results are pre- sented as mean optical density ? SEM. In addition, follow- ing counterstaining (see above) all positive cell profiles were counted in each section and expressed as a percentage of the total number of cell profiles in the section.
To determine the size distribution of DRG cell profiles and the percentage of profiles in each size bin that express mRNA encoding p75-LANR, TrkA, TrkB, and TrkC, respec- tively, two slides for each of the different neurotrophin receptors were analyzed, in both cervical and lumbar DRGs. The sections were examined under brightfield illumination with a x40 objective lens and positive cell profiles (see above) were recorded. The cross-sectional area of all cell profiles in each section were measured on photomontages by using a digitizing tablet (Summagraphie) interfaced with a computer equipped with BioQuant system IV@ (R&M Biometrics Inc., Nashville, TN). For each neuro- trophin receptor, a size-frequency histogram was con- structed showing the relationship between labelled and unlabelled cell profiles in each size bin. Small, medium, and large DRG neurons correspond here to neuronal profiles with an area of < 750 pm2, 750-1750 pm2, and > 1750 pm2, respectively.
Statistical tests Statistical analysis was performed by using the nonpara-
metric Mann-Whitney U-test (two-sample test) or conti- gency tabel analysis (x2; bin data). The level of significance was set a t P < 0.05. In the histograms, the level of significance has been indicated as follows: n s . = non significant; * = P < 0.05; ** = P < 0.01; *** = P < 0.001.
RESULTS Size-distribution of neuronal profiles in
aged DRGs Cell profile cross-sectional area histograms of cervical
and lumbar DRG neurons revealed a similar size distribu- tion in aged and young adult rats (Fig. 1).
Neuropeptide Y NPY-LI could be detected in both cervical and lumbar
DRG neurons of aged rats (Fig. 2). The number of NPY- immunoreactive (IR) cell profiles was larger in lumbar than in cervical DRGs. NPY-LI was only very infrequently seen in DRG neurons of young adult rats, while NPY-IR axon fibers were encountered more often (Fig. 2).
The labelling for NPY-LI in the dorsal horn, of the corresponding spinal cord segments, was similar in aged and young adult rats, at both cervical (not shown) and lumbar levels (Fig. 3). There were possibly more NPY fibers in lamina I11 of aged rats as compared to young adult rats. In aged rats, NPY-IR fibers could also be seen in the dorsal column of the spinal cord (Fig. 4).
In situ hybridization showed neurons expressing NPY mRNA in every section from aged rat DRGs (Fig. 5 ) . In young adult rats, very few DRG neurons were NPY mRNA positive, and in such cases the labelling was weaker than in
aged rats (Fig. 5). Consistent with the NPY-LI labelling in aged rat DRG neurons, the number of neuronal profiles positive for NPY mRNA was larger in lumbar ( - 13%) than in cervical DRGs ( - 3%) (Fig. 5 ) .
Galanin Galanin-LI was detected in a small number of DRG
neurons in both aged and young adult rats, and no clear difference could be detected between the age groups. The labelling pattern for galanin-LI in the dorsal horn was similar in aged and young adult rats (Fig. 3).
The expression of galanin mRNA was increased in aged rats in both cervical and lumbar DRG neurons. As for NPY a rostrocaudal gradient was observed with more cells labelled in lumbar ( - 7%) than in cervical ( - 4%) DRGs (Fig. 5 ) .
CGRP, substance P, and somatostatin The labelling for CGRP- and SP-LI showed a reduced
number of positive neuronal profiles in both cervical and lumbar DRGs of aged rats (Fig. 61, while no difference could be detected between the age groups for somatostatin-LI (not shown). Also the subcellular distribution of the neuro- peptide immunoreactivities was similar in aged and young adult rats (Fig. 7).
Consistent with the labelling pattern in the DRGs, a decrease in both CGRP-LI and SP-LI was seen in the dorsal horn of the aged rat (Fig. 3). However, also the labelling for somatostatin-LI was decreased as compared to young adult rats (Fig. 3).
Fig. 7. A confocal, -0.8 km thick, optical section recorded with a x loo/ 1.3 oil immersion objective showing the subcellular distribution of CGRP in aged rat DRG neurons. The immunostaining is concen- trated in the perinuclear region, presumably corresponding to the Golgi apparatus, and throughout the cytoplasm in small dot-like structures, likely to represent large dense-core vesicles (arrows). Framed area is a confocal image recorded with a ~4011.0 oil immersion objective show- ing the same cells. Scale bars = 5 pm.

PRIMARY SENSORY NEURONS IN AGED RATS 311
Fig. 8. Brightfield photomicrographs of young adult (Ad) (A, C ) and aged (Ag) (B, D) rat cervical (A, B) and lumbar (C, D) DRGs hybridized with a probe complementary to mRNA encoding CGRP. The young adult rats expressed CGRP in a large number of mainly small DRG cells, but occasional positive medium-sized and large cells were also
found (A, C). In aged rats a significant decrease in the number of CGRP positive neurons was observed (B, D), the downregulation being most pronounced in lumbar DRGs (D). Arrowheads (A-D) point to small, arrows (A-C) point to medium-sized or large neurons. Scale bar = 50 pm.
In situ hybridization (Figs. 8 and 9) disclosed a difference between aged and young adult rats (Fig. lo), with a smaller percentage of profiles expressing mRNA for CGRP, SP, and somatostatin in lumbar DRGs ( P < 0.01). The labelling intensity appeared to be increased in somatostatin mRNA positive neurons of the aged rat (Figs. 8 and 10).
decreased expression of Trk mRNAs (Fig. 14). The possible significance and mechanism(s) behind these changes are discussed below.
DRG neurons of aged rats show lower levels of CGRP and SP, and an upregulation of NPY and galanin (Fig. 14). These changes were most pronounced in lumbar DRGs.
p75-LANR and Trk mRNAs in primary sensory neurons
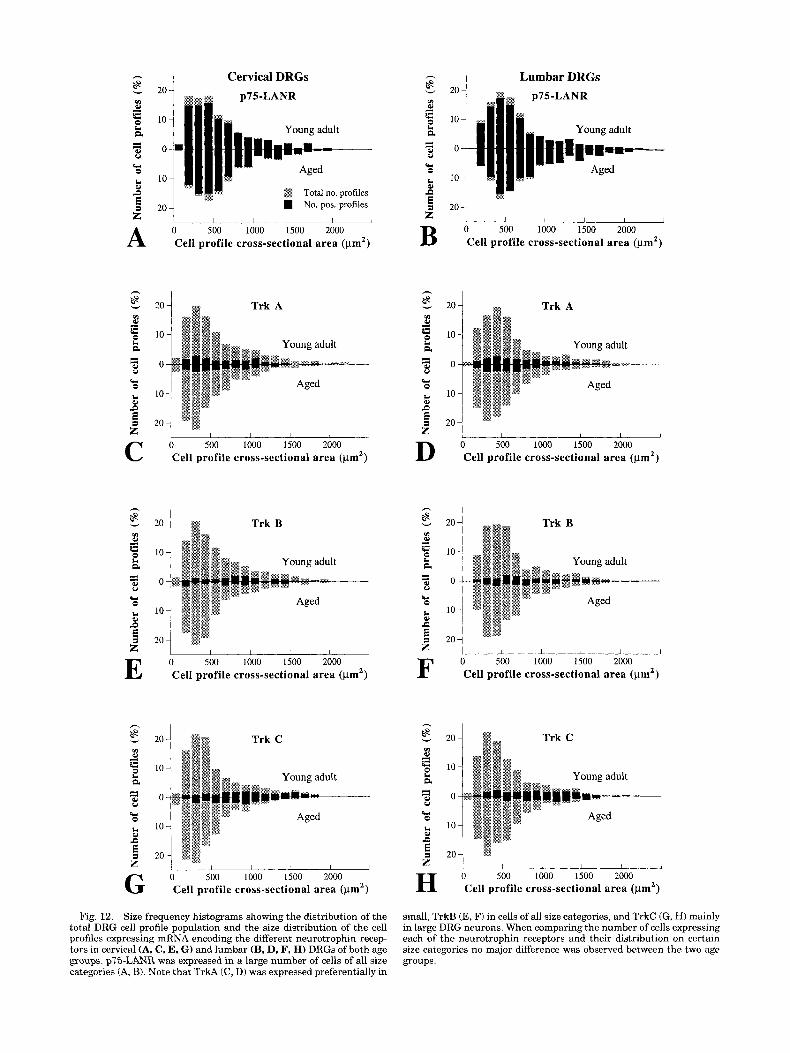
The labelling pattern for p75-LANR, TrkA, TrkB, and TrkC mRNA, respectively, is shown in Figure 11. Most (87 to 97%) of the neurons, in both cervical and lumbar DRGs, expressed p75-LANR mRNA in aged and young adult rats (Figs. 11 and 12). No difference could be detected between the age groups. The labelling for TrkA, TrkB, and TrkC mRNAs, respectively, was decreased in both cervical and lumbar DRGs of aged rats (Fig. 11). The difference con- cerned only the labelling intensity (Fig. 131, whereas the percentage of labelled neuronal profiles (Fig. 12) was similar in aged and young adult rats (10-20% of the profiles).
DISCUSSION In this study we show that primary sensory neurons in
30-month-old SD rats with hind limb motor incapacities have a change in their content of neuropeptides and a
With the exception for somatostatin, there was a good concordance between the findings in the DRG somata, and the labelling for the different peptides in the dorsal horn of the corresponding spinal cord segment. A major difference between SP and CGRP on one hand, and somatostatin on the other hand, is that in the dorsal horn 80% of the somatostatin is localized to intrinsic, dorsal horn neurons (Hunt, 1983), whereas most SP and CGRP originate from DRG neurons. The marked decrease in somatostatin-LI in aged rats dorsal horn may therefore, at least in part, be due to a downregulation of this peptide in neurons intrinsic to the dorsal horn (Todd and Spike, 1993).
Interestingly, no difference was seen in the dorsal horn (lamina I and 11) of aged and young adult rats with regard to NPY-LI, which also is known to be present almost exclu- sively in dorsal horn neurons under normal conditions (Gibson et al., 1984). Different populations of dorsal horn interneurons may therefore react in a nonuniform way to aging processes. Although somatostatin was changed in the dorsal horn, no certain effect was observed in the DRGs. Earlier studies suggested that somatostatin is present in a

312 E. BERGMAN ET AL.
Fig. 9. Darkfield photomicrographs of young adult (Ad) (A, C) and aged (Ag) (B, D) rat lumbar DRGs hybridized with probes complemen- tary to mRNA encoding substance P (SP) (A, B) and somatostatin (SOM) (C, D). In the young adult rats substance P was found in a large number of mainly small cells (A), whereas somatostatin mRNA was
expressed in a low number of exclusively small neurons (C). The aged rats disclosed a significant reduction in the number of cells positive for both substance P (B) and somatostatin (D). Note however the increased labelling intensity in the cells expressing somatostatin (D). Scale bar = 150 pm.
population different from SP and CGRP (Hokfelt et al., 1976; Tuchscherer and Seybold, 1985; Zhang et al., 1995) which could be related to the differential effect of aging. More recent studies show that somatostatin mRNA can be detected in SP and CGRP mRNA positive neurons (Kashiba et al., 1996). On the other hand, Verge et al. (1995) have shown that SP and CGRP, but not somatostatin DRG neurons are NGF responsive.
In many respects, the changes in peptide expression in aged rats resemble those seen in young adult rats subjected to sensory nerve injury (Fig. 14). As mentioned in the Introduction, transection and crush of peripheral sensory nerves induce marked changes in the expression of pep- tides, with a decrease of CGRP, SP, and somatostatin (see Hokfelt et al., 1994 for references) and an upregulation of galanin (Hokfelt et al., 1987; Villar et al., 19891, VIP (Shehab and Atkinson, 1986; Villar et al., 19891, and NPY (Wakisaka et al., 1991, 1992; Zhang et al., 1993b). I t has been suggested that the changed pattern of neuropeptide expression in response to axonal injury serves to attenuate dorsal horn transmission and to promote regenerative processes of the lesioned sensory pathway. Evidence indi- cate that the upregulation of VIP and galanin occurs mainly in small DRG neurons that normally express CGRP and SP (Xu et al., 1990; Doughty et al., 1991; Kashiba et al., 19921, and that the inhibitory function of galanin in the dorsal horn is enhanced after axotomy (Wiesenfeld-Hallin et al.,
1992). These similarities may suggest that the changes seen here in aged rats are caused by aging-related axonal lesions.
In peripheral nerves of aged animals both losses and degenerative changes have been reported for aminergic, cholinergic, and peptidergic nerve fibers (Cowen et al., 1982; Dhall et al., 1986; Mione et al., 1988; Cowen and Thrasivoulou, 1990; Navarro and Kennedy, 1990; Abdel- Rahman and Cowen, 1993). These data are consistent with morphological studies of peripheral sensory nerves and dorsal roots demonstrating axonal dystrophy, demyelin- ation, axon degeneration, as well as loss of fibers during aging (Berg et al., 1962; van Steenis and Kroes, 1971; Samorajski, 1974; Burek et al., 1976; Sharma et al., 1980; Thomas et al., 1980; Cotard-Bartley et al., 1981; Mitsumori et al., 1981; Krinke, 1983). The aging-related lesions seem to be most prevalent in nerves supplying the hind limbs and tail (Berg et al., 1962; van Steenis and Kroes, 1971; Burek et al., 1976), and resemble those seen in different experimen- tal and disease induced peripheral neuropathies (Cavanagh, 1964; Jellinger, 1973). As mentioned in the Introduction, a vast majority of the aging rats develop hind limb incapaci- ties (“posterior paralysis”) during their third year of life, and there seems to be a fairly good agreement between the distribution of aging-related axonal lesions and the behav- ioral symptoms of the animals (e.g., Berg et al., 1962; Johnson et al., 1993, 1995). The etiology of these distur- bances remains, however, unclear. According to Berg (1959)

PRIMARY SENSORY NEURONS IN AGED RATS 313
Fig. 10. Histograms showing the percentage f SEM (n = 2,393- 4,801) (A, C, E) and the mean optical densities f SEM (n = 304-737) (B, D, F) of cell profiles expressing CGRP (A, B), substance P (C, D), and somatostatin (E, F) in cervical and lumbar DRGs of both age groups. The number of cell profiles expressing mRNA for each of the three peptides was reduced in lumbar DRGs of the aged rats (A, C, E),
and for CGRP we also found a lower number of positive neurons at the cervical level (A). With regard to labelling intensity an increased signal was found in somatostatin positive neurons of the aged rats (F), whereas no difference was observed for CGRP (B) or substance P (D). Levels of significance are indicated as described in Materials and Methods.
vitamin E supplementation had no effect on the progressive nerve lesions, and the extensive engagement of the tail nerve (King and Thomas, 1982) argues against “pressure” as a main factor to the lesions. Studies using mutant mice indicate that both aging-related axonal dystrophy and degeneration may be linked to certain gene mutations
(Kikuchi et al., 1990; Bronson, 1992). Axonal lesions during aging are not confined to the peripheral nervous system, but they occur with high frequency also in the dorsal column system that contains central projections of proprio- ceptive sensory neurons (e.g., Brannon et al., 1967; Sroka et al., 1969; Fuijisawa and Shiraki, 1978; Fujisawa, 1988). It is
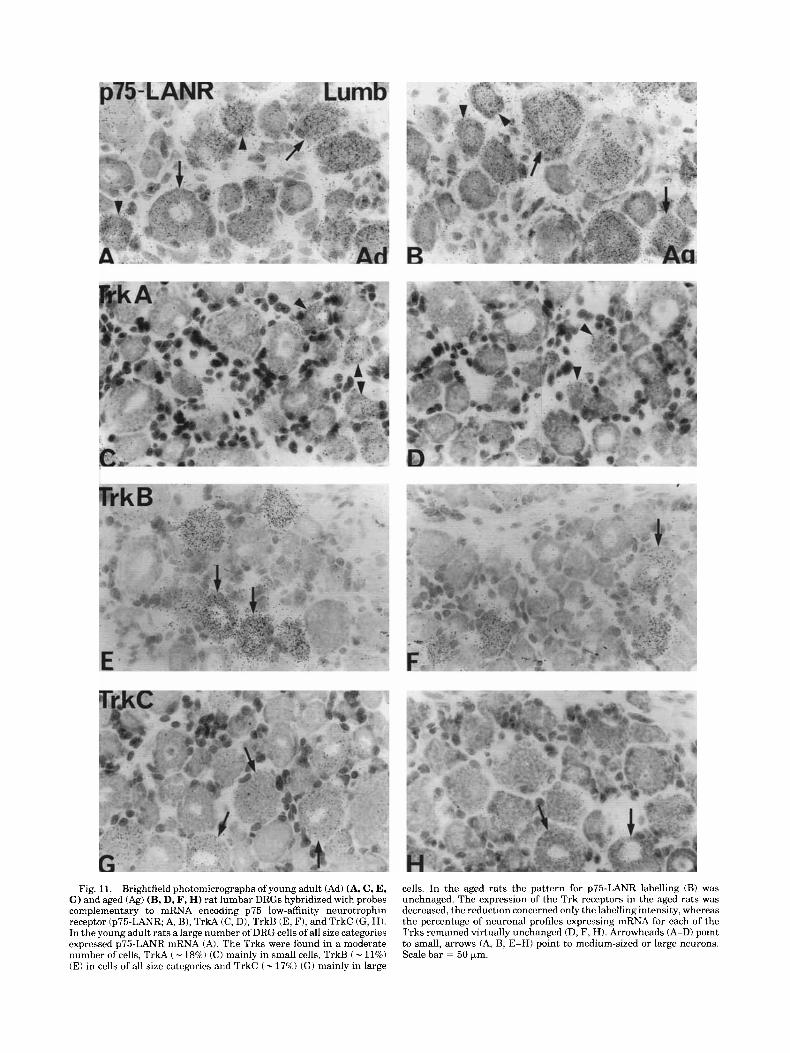
Fig. 11. Brightfield photomicrographs ofyoung adult (Ad) (A, C, E, G) and aged (Ag) (B, D, F, H) rat lumbar DRGs hybridized with probes complementary to mRNA encoding p75 low-affinity neurotrophin receptor (p75-LANR; A, B), TrkA (C, D), TrkB (E, F), and TrkC (G, H). In the young adult rats a large number of DRG cells of all size categories expressed p75-LANR mRNA (A). The Trks were found in a moderate number of cells, TrkA ( - 18%) (C) mainly in small cells, TrkB ( - 11%) (E) in cells of all size categories and TrkC ( - 175%) (GI mainly in large
cells. In the aged rats the pattern for p75-LANR labelling (B) was unchnaged. The expression of the Trk receptors in the aged rats was decreased, the reduction concerned only the labelling intensity, whereas the percentage of neuronal profiles expressing mRNA for each of the Trks remained virtually unchanged (D, F, H). Arrowheads (A-D) point to small, arrows (A, B, E-HI point to medium-sized or large neurons. Scale bar = 50 pm.
h .Q I Cervical DRGs Lumbar DRGs n .a I
v)
6 20 6 20 v)
10 z 10 e a i - I
0 0 0 0
L 10 CI 10
(r
I I
(r
B @ Total no profiles B No pas profiles B 20 B 20 c z
I
0 500 1000 1500 2000 Cell profile cross-sectional area (prnz) A
6 I I I I I I 0 500 1000 1500 2000 Cell profile cross-sectional area (pmz) B
Trk A !5 204 ~ Trk A
I I
0 500 1000 1500 2000 Cell profile cross-sectional area (prnz) C
I 1 1 I I I
0 500 1000 1500 2000 Cell profile cross-sectional area (prnz) D
Trk B - 1 5 z o i
h
I I I I I I 0 500 1000 1500 2000 E Cell profile cross-sectional area (pmz)
h
5 20
g 10
k
v)
- I 0 0
h 10 n
z
(r 0
$ 20
0 500 1000 1500 2000 G Cell profile cross-sectional area (prnz)
Fig. 12. Size frequency histograms showing the distribution of the total DRG cell profile population and the size distribution of the cell profiles expressing mRNA encoding the different neurotrophin recep- tors in cervical (A, C, E, G ) and lumbar (B, D, F, H) DRGs of both age groups. p75-LANR was expressed in a large number of cells of all size categories (A, B). Note that TrkA (C, D) was expressed preferentially in
I I I I I J 0 500 1000 1500 2000 F Cell profile cross-sectional area (prnz)
h I
! I I !
0 500 1000 1500 2000 H Cell profile cross-sectional area (prnz)
small, TrkB (E, F) in cells of all size categories, and TrkC (G, HI mainly in large DRG neurons. When comparing the number of cells expressing each of the neurotrophin receptors and their distribution on certain size categories no major difference was observed between the two age groups.

316 E. BERGMAN ET AL.
Fig. 13. Histograms showing the mean optical densities f SEM of cell profiles expressing mRNA for each of the Trk neurotrophin receptors in cervical and lumbar DRGs of both age groups. The aged rats disclosed a significant decrease in labelling intensity for all of the Trk-receptors in both cervical and lumbar DRGs. Levels of significance are indicated as described in Materials and Methods.
now well established that whereas VIP and galanin are upregulated mainly in small DRG neurons of C and AS type (see Hokfelt et al., 19941, NPY is mainly increased in large DRG neurons, i.e., proprioceptive neurons from muscle and joints (Wakisaka et al., 1991, 19921, projecting in the dorsal column to the gracile nuclei (Zhang et al., 1993b). In
agreement NPY could be seen in the dorsal columns of aged rats in the present study. In fact, NPY seemed more affected by aging than galanin, supporting the view of aging affecting proprioceptive systems more severely.
Although the mechanisms causing degeneration of sen- sory fibers in aged rats remain unclear, their presence may thus explain the changed peptide expression in DRG neu- rons observed here. Consistently the changes found in this study were most pronounced in lumbar DRG neurons that supplies the hind limbs, indicating a correlation to the behavioral symptoms of the rats.
Target derived neurotrophic factors (nerve growth fac- tor, NGF; brain-derived neurotrophic factor, BDNF; neuro- trophin-3, NT-3; neurotrophin-4/5, NT-4/5) have well es- tablished roles during prenatal development. Mice knockouts have elegantly illustrated the specificity of the different neurotrophins and their respective Trk-signalling pathway (Klein et al., 1993, 1994; Ernfors et al., 1994a,b; Farifias et al., 1994; Jones et al., 1994; Smeyne et al., 1994; Conover et al., 1995; Liu et al., 1995). The expression of both neuro- trophins and their receptors persists beyond the develop- mental stage (see e.g., Ernfors et al., 1990; Merlio et al., 1992; Verge et al., 1992; Altar et al., 1994; Funakoshi et al., 19951, and in the adult rat the expression of p75-LANR and Trk mRNA remains sensitive to perturbations of axonal integrity as well as levels of accessible ligands. Thus, peripheral axotomy and crush injury induce a downregula- tion of p75-LANR and Trk mRNA, and a decreased binding of NGF (Verge et al., 1992). This pattern could be reversed by exogenous NGF, and, moreover, p75-LANR mRNA was upregulated in response to exogenous NGF also in intact DRG neurons. These data may indicate a sustained respon- siveness to low-levels of target-derived trophic factors(s) in the adult, and that would also be consistent with the significant loss of DRG neurons following axonal severance ( - 30%) (Ygge and Aldskogius, 1984).
At large our data on neurotrophin receptor expression in young adult DRGs are in agreement with earlier work (Verge et al., 1992; McMahon et al., 1994; Kashiba et al., 1995); however, we found a higher frequency of p75-LANR positive neurons (see also Wetmore and Olson, 1995). At this point it is unclear if hybridization conditions and/or probe sensitivity may have contributed to these differences. The data from aged rats show that both the percentage of p75 and Trk mRNA expressing DRG neurons, and the size-distribution of positive neurons remain unchanged during aging, while the level of Trk A/B/C mRNA expres- sion is lower in the aged rat. I t cannot be ruled out that the lowered levels of Trk mRNA expression seen in aged rats may have been due to a decreased access to Trk-ligands (Figs. 14). However, it needs to be shown that the Trk mRNA expression in aged DRG neurons corresponds to a lower cellular content of Trk receptors, and that the gene expressions remains responsive to axonal lesions and neuro- trophin supplementation. Thrasivoulou and Cowen (1995) showed that the 50% decrease in sympathetic nerve fiber innervation of cerebral arteries seen in aged rats could be restored to normal by exogenous NGF, indicating a main- tained responsiveness to target derived neurotrophins throughout life (see also reviews by Brewster et al., 1994; Lindsay et al., 1994; Rylett and Williams, 1994).
Neurotrophins can also influence the cellular expression of neuropeptides such as CGRP, SP, and NPY (e.g., Goedert et al., 1981; Schwartz et al., 1982; Fitzgerald et al., 1985; Lindsay and Harmar, 1989; Wong and Oblinger, 1991;

PRIMARY SENSORY NEURONS IN AGED RATS
Glutamate Neuropeptides (CGRP, SP, SOM,VIP, NPY, Galanin) p75-LANR, Trk A/BK BDNF, NT3, other?
317
+ NTS
Aging /
+ Glutamate (+) CGRP/SP (+) Galanin (+/-) NPY (-?) VIP (?>
& (?) +
+ NPY pept. t NPY -A +- Galanin pept. & mRNA J. CGRP pept. 62 mRNA J. SP pept. & mRNA J. Trk mRNA J. p75mRNA ? BD"T3 ?
t N T S -+
4 - ->
7
Wderian degeneration Tmgaglionic degeneration of fibers distal to hjq. Decreased access to target-derived NTS. haeased expression of NTs in supportive ceh. galanin.
of fibers in the dorsal horn. Decreased IR to CGRP, SP, SOM in the dorsal horn. Increased inhibitory effect of
d NPY pept. & NPY mRNA 4FGalaninmRNA .L CGRP pept. & CGRP mRNA J. SP pept. & SP mRNA
Axonal dystrophy/ degeneration. Decreased access to target-derived NTs ? J. TrkmRNA
f, p75mRNA ? BDNFMT3
Axonal dystrophy/
SP, SOM in the dorsal horn.
? degeneration? + Decreased IR to CGRP, - ->
N T S
/ \
1' \.
Fig. 14. Schematic drawing showing the effects on primary sensory neurons of aging and axotomy, respectively.
Jones et al., 1994; Mulderry, 1994; Verge et al., 1995; Watson, 1995). It can therefore not be excluded that the changed expression of neuropeptides during aging, or as a result of axotomy, is mediated by changes in the availability of neurotrophins and/or the expression of the neurotrophin receptors.
Finally, our data show that the expression of p75-LANR mRNA is maintained in aged animals. Consequently, the cellular ratio between p75 and Trk mRNA is increased in aged DRG neurons. The precise interaction of p75-LANR and Trk receptors is still not fully clarified (Chao and Hempstead, 1995); however, evidence indicates that coex- pression of these receptors may enhance responsiveness to neurotrophins. Of importance here is that increased levels/ overexpression of p75 may induce cellular susceptibility to cell degenerative stimuli/apoptosis (Rabizadeh et al., 1993). Hence, increased levels of unbound p75-LANR, which may be the case in aged DRG neurons, may prove to be harmful, assuming that the changed mRNA expression ratio corre- spond to a changed receptor ratio (Fig. 14).
ACKNOWLEDGMENTS
Institutet, Stockholm, Sweden (CGRP, NPY, SP) and J. Brown and A. Buchan, University of British Columbia, Vancouver, Canada (somatostatin) for generous supply of antiseralantibodies.
LITERATURE CITED Abdel-Rahman, T.A., and T. Cowen (1993) Neurodegeneration in sweat
glands and skin of aged rats. J. Auton. Nerv. Syst. 46:55-63. Algeri, S., G. Calderini, G. Toffano, and F. Ponzio (1983) Neurotransmittor
alterations in aging rats. In D. Samuel (ed): Aging of the Brain. New York: Raven Press. pp. 227-243.
Altar, C.A., J.A. Siuciak, P. Wright, N.Y. Ip, R.M. Lindsay, andS.J. Wiegand (1994) In situ hybridization of trkB and trkC receptor mRNA in rat forebrain and association with high-affinity binding of 1125IlBDNF, 1125I]INT-4/5 and 1125IlNT-3. Eur. J. Neurosci. 6:1389-1405.
Amara, S.G., J.L. Arriza, S.E. Leff, L.W. Swanson, R.M. Evans, and M.G. Rosenfeld (1985) Expression in brain of a messenger RNA encoding a novel neuropeptide homologous to calcitonin gene-related peptide. Sci- ence 229: 1094-109 7.
Barde, Y.A. (1989) Trophic factors and neuronal survival (Review). Neuron 21525-1534.
Berg, B.N. (1959) Study of vitamin E supplements in relation to muscular dystrophy and other diseases in aging rats. J. Gerontol. 14:174-180.
This study was supported by grants from the swedish Medical Research 10820 and 2887)' Karo- linska Institutet, f i ren S t i f t e k L and H O&Wmans fond for medicin& forskning, Stohnes S t i f t e k A &.
Berg, B.N., A. Wolf, and H.S. Simms (1962) Degenerative lesions of spinal roots and peripheral nerves in aging rats. Gerontologia (Basel) 6:72-80.
Brannon, W., W. McCormick, and P. Lampert (1967) Axonal dystrophy in the gracile nucleus of man. Acta. Neuropathol. (Berlin) 9tl-6.
Brewster, W.J., P. Fernyhough, L.T. Diemel, L. Mohiuddin, and D.R. bergs Stiftelse. We thank Profs. L. Terenius, Karolinska Tomlinson (1994) Diabetic neuropathy, nerve growth factor and other

318
neurotrophic factors [see comments] (Review). Trends Neurosci. 17r321- 325.
Bronson, R.T., H.O. Sweet, C.A. Spencer, and M.T. Davisson (1992) Genetic and age related models of neurodegeneration in mice: Dystrophic axons. J. Neurogenet. 8:7143.
Buchan,A.M.J., L.K.J. Sikora, J.G. Levy, C.H.S. McIntosh, I. Dyck, and J.C. Brown (1985) An immuno-cytochemical investigation with monoclonal antibodies to somatostatin. Histochemistry 83:175-180.
Burek, J.D., and C.F. Hollander (1980) Experimental gerontology. The Laboratory Rat, Vol. 11. New York: Academic Press, pp. 149-159.
Burek, J.D., A.J. van der Kogel, and C.F. Hollander (1976) Degenerative myelopathy in three strains of aging rats. Vet. Pathol. 13,321-331.
Cavanagh, J.B. (1964) The significance of the “dying back” process in experimental and human neurological disease. Int. Rev. Exp. Pathol. 3:2 19-267.
Chao, M.V. (1992) Neurotrophin receptors: a window into neuronal differen- tiation (Review). Neuron 9583-593.
Chao, M.V., and B.L. Hempstead (1995) p75 and Trk: A two-receptor system. Trends Neurosci. 18:321-326.
Christensson-Nylander, I., M. Herrera-Marschitz, W. Staines, T. Hokfelt, L. Terenius, U. Ungerstedt, A.C. Cuello, W.L. Oertel, and M. Goldstein (1986) Striato-nigral dynorphin and substance P pathways in the rat: I. Biochemical and immunohistochemical studies. Exp. Brain Res. 64:164- 192.
Conover, J.C., J.T. Erickson, D.M. Katz, L.M. Bianchi, W.T. Poueymirou, J. McClain, L. Pan, M. Helgren, N.Y. Ip, P. Boland, B. Friedman, S. Wiegand, R. Vejsada, A.C. Kato, T.M. DeChiara, and G.G. Yancopoulos (1995) Neuronal deficits, not involving motor neurons, in mice lacking BDNF and/or NT4. Nature 375:235-238.
Coons, A.H. (1958) Fluorescent antibody methods. In J.F. Danielli (ed): General Cytochemical Methods. New York: Academic Press, pp. 399- 422.
Cotard-Bartley, M.P., J. Secchi, R. Glomot, and J.B. Cavanagh (1981) Spontaneous degenerative lesions of peripheral nerves in aging rats. Vet. Pathol. 18: 110-1 13.
Cowen, T., A.J. Haven, C. Wen &in, D.D. Gallen, F. Franc, and G. Burnstock (1982) Development and ageing of perivascular adrenergic nerves in the rabbit. A quantitative fluorescence histochemical study using image analysis. J. Auton. Nerv. Syst. 5:317-336.
Cowen, T., and C. Thrasivoulou (1990) Cerebrovascular nerves in old rats show reduced accumulation of 5-hydroxytryptamine and loss of nerve fibres. Brain Res. 513:237-243.
Dagerlind, A, K. Friberg, A.J. Bean, and T. Hokfelt (1992) Sensitive mRNA detection using unfixed tissue: Combined radioactive and non-radioac- tive in situ hybridization histochemistry. Histochemistry 98:39-49.
Dalsgaard, C.J. (1988) The sensory system. In A. Bjorklund, T. Hokfelt, and C. Owman (eds): Handbook of Chemical Neuroanatomy. Amsterdam: Elsevier Science Publishers B.V., 6. pp. 599-636.
Dhall, U., T. Cowen, A.J. Haven, and G. Burnstock (1986) Perivascular noradrenergic and peptide-containing nerves show different patterns of change during development and ageing in the guinea-pig. J. Auton. Nerv. Syst. 16:109-126.
Doughty, S.E., M.E. Atkinson, and S.A. Shehab (1991) A quantitative study of neuropeptide immunoreactive cell bodies of primary afferent sensory neurons following rat sciatic nerve peripheral axotomy. Regul. Pept. 35:59-72.
Ernfors, P., K.F. Lee, and R. Jaenisch (1994a) Mice lacking brain-derived neurotrophic factor develop with sensory deficits. Nature 368:147-150.
Ernfors, P., K.F. Lee, J. Kucera, and R. Jaenisch (199413) Lack of neuro- trophin-3 leads to deficiencies in the peripheral nervous system and loss of limb proprioceptive afferents. Cell 77:503-512.
Ernfors, P., C. Wetmore, L. Olson, and H. Persson (1990) Identification of cells in rat brain and peripheral tissues expressing mRNA for members of the nerve growth factor family. Neuron 5:511-526.
Farinas, I., K.R. Jones, C. Backus, X.Y. Wang, and L.F. Reichardt (1994) Severe sensory and sympathetic deficits in mice lacking neurotrophin-3. Nature 369:658-661.
Fitzgerald, M., P.D. Wall, M. Goedert, and P.C. Emson (1985) Nerve growth factor counteracts the neuro-physiological and neurochemical effects of chronic sciatic nerve section. Brain Res. 332:131-141.
Fuiisawa. K. (1988) Studv of axonal dvstrouhv. 111. Posterior funiculus and
E. BERGMAN ET AL.
~ posterior column of‘aging and old rats. Acta Neuropathol. (Berlin) 76:115-127.
Fujisawa, K., and H. Shiraki (1978) Study of axonal dystrophy. 1. Pathology of the neuropil of the gracile and the cuneate nuclei in aging and old rats: A stereological study. Neuropathol. Appl. Neurobiol. 4: 1-20.
Funakoshi, H., N. Belluardo, E. Arenas, Y. Yamamoto, A. Casabona, H. Persson, and C.F. Ibanez (1995) Muscle-derived neurotrophin-4 as an activity-dependent trophic signal for adult motor neurons. Science 268:1495-1499.
Gibson, S.J., J.M. Polak, J.M. Allen, T.E. Adrian, J.S. Kelly, and S.R. Bloom (1984) The distribution and origin of a novel brain peptide, neuropeptide Y, in the spinal cord of several mammals. J. Comp. Neurol. 227:78-91.
Gilmore, S.A. (1972) Spinal nerve root degeneration in aging laboratory rats: A light microscopic study. Anat. Rec. 174r251-257.
Goedert, M., K. Stoeckel, and U. Otten (1981) Biological importance of the retrograde axonal transport of nerve growth factor in sensory neurons. Proc. Natl. Acad. Sci. USA 78:5895-5898.
Goodman, R.H., D.C. Aron, and B.A. Roos (1983) Rat pre-prosomatostatin. J. Biol. Chem. 258:5570-5573.
Gutman, B., and V. Hanzlikova (1972) Age Changes in the Neuromuscular System. Bristol, Scientechnica Ltd.
Hartman, B.K., D. Zide, and S. Udenfriend (1972) The use of dopamine p-hydroqlase as a marker for the central noradrenergic nervous system in rat brain. Proc. Natl. Acad. Sci. USA 692722-2726.
Hunt, S.P. (1983) Cytochemistry of the spinal cord. In P.C. Emson (ed): Chemical Neuroanatomy. New York: Raven Press, pp. 53-84.
Hokfelt, T., R. Elde, 0. Johansson, R. Luft, G. Nilsson, and A. Arimura (1976) Immunohistochemical evidence for separate populations of so- matostatin-containing and substance P-containing primary afferent neurons in the rat. Neuroscience I:131-136.
Hokfelt, T., Z. Wiesenfeld-Hallin, M.J. Villar, and T. Melander (19871 Increase of galanin-like immunoreactivity in rat dorsal root ganglion cells after peripheral axotomy. Neurosci. Lett. 83:217-220.
Hokfelt, T., X. Zhang, and Z. Wiesenfeld-Hallin (1994) Messenger plasticity in primary sensory neurons followingaxotomy and its functional implica- tions. Trends Neurosci. 17:22-30.
Ip, N.Y., T.N. Stitt, P. Tapley, R. Klein, D.J. Glass, J. Fandl, L.A. Greene, M. Barbacid, and G.D. Yancopoulos (1993) Similarities and differences in the way neurotrophins interact with the Trk receptors in neuronal and nonneuronal cells. Neuron 10:137-149.
Jellinger, K. (1973) Neuroaxonal dystrophy: Its natural history and related disorders. In H.M. Jimmernan (ed): Progress in Neuropathology. New York, London: Grune & Stratton, pp. 129-180.
Johnson, G.D., and G.M.d.C. Nogueira Araujo (1981) A simple method of reducing the fading of immunofluorescence during microscopy. J. Immu- nol. Methods 43:349-350.
Johnson, H., B. Ulfhake, A. Dagerlind, G.W. Bennett, K.C. Fone, and T. Hokfelt (1993) The serotoninergic bulbospinal system and brainstem- spinal cord content of serotonin-, TRH-, and substance P-like immunore- activity in the aged rat with special reference to the spinal cord motor nucleus. Synapse 15:63-89.
Johnson, H., K. Mossberg, U. Arvidsson, F. Piehl, T. Hokfelt, and B. Ulfhake (1995) Increase in alpha-CGRP and GAP-43 in aged motoneurons: A4 study of peptides, growth factors, and ChaT mRNA in the lumbar spinal cord of senescent rats with symptoms of hindlimb incapacities. J. Comp. Neurol. 359:69-89.
Jones, K.R., I. Farinas, C. Backus, and L.F. Reichardt (1994) Targeted disruption of the BDNF gene perturbs brain and sensory neuron development but not motor neuron development. Cell 76:989-999.
Kashiba, H., E. Senba, Y. Ueda, and M. Tohyama (1992) Co-localized but target-unrelated expression of vasoactive intestinal polypeptide and galanin in rat dorsal root ganglion neurons after peripheral nerve crush injury. Brain Res. 58247-57,
Kashiba, H., K. Noguchi, Y. Ueda, and E. Senba (1995) Coexpression of trk family members and low-affinity neurotrophin receptors in rat dorsal root ganglion neurons. Mol. Brain Res. 30:158-164.
Kashiba, H., Y. Ueda, and E. Senba (1996) Coexpression of preprotachyki- nin-A, a-calcitonin gene-related peptide, somatostatin, and neurotrophln receptor family messenger RNAs in rat dorsal root ganglion neurons. Neuroscience 70:179-189.
Kikuchi, T., M. Mukoyama, K. Yamakazi, and H. Moriya (1990) Axonal degeneration of ascending sensory neurons in gracile axonal dystrophy mutant mouse. Acta Neuropathol. (Berlin) 80:145-151.
King, R.H.M., and P.K. Thomas (1982) Ultrastructural changes in periph- eral nerve with aging. Ninth meeting of Swiss neuropathologists (with international participation), St Moritz, Switzerland.
Klein, R., I. Silos-Santiago, R.J. Smeyne, S.A. Lira, R. Brambilla, S. Bryant, L. Zhang, W.D. Snider, and M. Barbacid (1994) Disruption of the

PRIMARY SENSORY NEURONS IN AGED RATS
neurotrophin-3 receptor gene trkC eliminates la muscle afferents and results in abnormal movements (see comments). Nature 368249-251,
Klein, R., R.J. Smeyne, W. Wurst, L.K. Long, B.A. Auerbach, A.L. Joyner, and M. Barbacid (1993) Targeted disruption of the trkB neurotrophin receptor gene results in nervous system lesions and neonatal death. Cell 75:113-122.
Knox, C.A., E. Kokmen, and P.J. Dyck (1989) Morphometric alteration ofrat myelinated fibers with aging. J. Neuropathol. Exp. Neurol. 48:119-139.
Koller, W.C., S.L. Glatt, and J.H. Fox (1985) Senile gait: A distinct neurologic entity. Clin. Geriatr. Med. 1:661-669.
Korsching, S. (1993) The neurotrophic factor concept: a reexamination (Review). J. Neurosci. 13:2739-2748.
Krause, J.E., J.M. Chirgwin, M.S. Carter, Z.S. Xu, and A.D. Hershey (1987) Three rat preprotachykinin mRNAs encode the neuropeptides substance P and neurokinin A. Proc. Natl. Acad. Sci. USA84:881-885.
Krinke, G. (1983) Spinal radiculoneuropathy in aging rats: demyelination secondary to neuronal dwindling? Acta Neuropathol. (Berlin) 59.63-69.
Lamballe, F., R. Klein, and M. Barbacid (1991) trkC, a new member of the trk family of tyrosine protein kinases, is a receptor for neurotrophin-3. Cell 66:967-979.
Larhammar, D., A. Ericsson, and H. Persson (1987) Strncture and expres- sion of the rat neuropeptide Y gene. Proc. Natl. Acad. Sci. USA 84:2068-2072.
Lawson, S.L. (1992) Morphological and biochemical cell types of sensory neurons. In S.E. Scott (ed): Sensory Neurons: Diversity, Development, Plasticity. New York: Oxford Univ. Press, pp, 27-59.
Lindsay, R.M., and A.J. Harmar (1989) Nerve growth factor regulates expression of neuropeptide genes in adult sensory neurons. Nature 337:362-364.
Lindsay, R.M., S.J. Wiegand, A. Altar, and P.S. DiStefano (1994) Neuro- trophic factors: from molecule to man. Trends Neurosci. 17:182-190.
Lindvall, O., Z. Kokaia, J. Bengzon, E. Elmer, and M. Kokaia (1994) Neurotrophins and brain insults (Review). Trends Neurosci. 17:490- 496.
Liu, X., P. Ernfors, H. Wu, and R. Jaenisch (1995) Sensory but not motor neuron deficits in mice lacking NT4 and BDNF. Nature 375:238-241.
Lundberg, J.M., L. Terenius, T. Hokfelt, and K. Tatemoto (1984) Compara- tive Immunohistochemical and biochemical analysis of pancreatic poly- peptide-like peptides with special reference to presence of neuropeptide Y in central and peripheral neurons. J. Neurosci. 4.2376-2386.
Martin-Zanca, D., S.H. Hughes, and M. Barbacid (1986) A human oncogene formed by the fusion of truncated tropomyosin and protein tyrosine kinase sequences. Nature 319:743-748.
Masoro, E.J. (1980) Mortality and characteristics of rat strains commonly used in agingresearch. Exp. Aging Res. 6:219-233.
McMahon, S.B., M.P. Armanini, L.H. Ling, and H.S. Phillips ( 1994) Expression and coexpression of Trk receptors in subpopulations of adult primary sensory neurons projecting to identified peripheral targets. Neuron 12: 1161-1 17 1.
Meakin, S.O., and E.M. Shooter (1992) The nerve growth family of receptors. Trends Neurosci. 15:323-331.
Merlio, J.P., P. Ernfors, M. Jaber, and H. Persson (1992) Molecular cloning of rat trkC and distribution of cells expressing messenger RNAs for members of the trk family in the rat central nervous system. Neurosci- ence 51:513-532.
Middlemas, D.S., R.A. Lindberg, and T. Hunter (1991) trkB, a neural receptor protein-tyrosine kinase: Evidence for a full-length and two truncated receptors. Mol. Cell. Biol. 11.143-153.
Mione, M.C., K.K. Dhital, F. Amenta, and G. Burnstock 11988) An increase in the expression of neuropeptidergic vasodilator, but not vasoconstric- tor, cerebrovascular nerves in aging rats. Brain Res. 460:103-113.
Mitsumori, K., K. Maita, and Y. Shirasu (1981) An ultrastructural study on spinal nerve roots and dorsal root ganglia in aging rats with spontaneous radiculoneuropathy. Vet. Pathol. 18:714-726.
Mittal, K.R., and F.H. Logmani (1987) Age-related reduction in 8th cervical ventral nerve root myelinated fiber diameters and number in man. J. Gerontol. 42:8-10.
Mulderry, P.K. (1994) Neuropeptide expression by newborn and adult rat sensory neurons in culture: Effects of nerve growth factor and other neurotrophic factors. Neuroscience 59:673-688.
Navarro, X., and W.R. Kennedy 11990) Changes in sudomotor nerve territories with aging in the mouse. J. Auton. New. Syst. 31:101-107.
Orazzo, C., V.A. Pieribone, S. Ceccatelli, L. Terenius, and T. Hokfelt (1992) CGRP-like immunoreactivity in A1 1 dopamine neurons projecting to the
319
spinal cord and a note on CGRP-CCK cross reactivity. Brain Res. 600:39-48.
Pease, P.C. (1962) Buffered formaldehyde as a killing agent and primary fixative for electron microscopy. Anat. Rec. 142342.
Platt, J.L., and A.F. Michael (1983) Retardation of fading and enhancement of intensity of immunofluorescence hy p-phenylenediamine. J. Histo- chem. Cytochem. 31:840-842.
Rabizadeh, S., J. Oh, L.T. Zhong, J. Yang, C.M. Bitler, L.L. Butcher, and D.E. Bredesen (1993) Induction of apoptosis by the low-affinity NGF receptor. Science 261 :345-348.
Radeke, M.J., T.P. Misko, C. Hsu, L.A. Herzenberg, and E.M. Shooter (1987) Gene transfer and molecular cloning of the rat nerve growth factor receptor. Nature 325593-596.
Rylett, R.J., and L.R. Williams (1994) Role of neurotrophins in cholinergic- neurone function in the adult and aged CNS (Review). Trends Neurosci. 17:486-490.
Samorajski, T. (1974) Age differences in the morphology of posterior tibial nerves of mice. J. Comp. Neurol. 1571439-445.
Schwartz, J.P., J. Pearson, and E.M. Johnson (1982) Effect of exuosure to anti-NGF on sensory neurons of adult rats and guinea pigs. Brain Res. 244:3 78-38 1.
Sharma, A.K., S. Bajada, and P.K. Thomas 11980) Age changes in the tibial and plantar nerves of the rat. J. Anat. 130:417-428.
Shehab, S.A., and M.E. Atkinson (1986) Vasoactive intestinal polypeptide (VIP) increases in the spinal cord after peripheral axotomy of the sciatic nerve originate from primary afferent neurons. Brain Res. 3723744.
Smeyne, R.J., R. Klein, A. Schnapp, L.K. Long, S. Bryant, A. Lewin, S.A. Lira, and M. Barbacid (1994) Severe sensory and sympathetic neuropa- thies in mice carrying a disrupted TrkiNGF receptor gene (see com- ments). Nature 368:246-249.
Sroka, C., B. Bornstein, N. Strulovici, and U. Sandbank (1969) Neuroaxonal dystrophy: its relation to age and central nervous system lesions. Isr. J. Med. Sci. 5:373-377.
Thoenen, H. (1991) The changing scene of neurotrophic factors (Review). Trends Neurosci. 14:165-170.
Thoenen, H. (1995) Neurotrophins and neuronal plasticity. Science270:593- 598.
Thomas, P.K., R.H. King, and A.K. Sharma (1980) Changes with age in the peripheral nerves of the rat. An ultrastructural study. Acta Neuropathol. (Berlin) 52:1-6.
Thrasivoulou, C., and T. Cowen (1995) Regulation of rat sympathetic nerve density by target tissues and NGF in maturity and old age. Eur. J. Neurosci. 7.381-387.
Todd, A.J., and R.C. Spike (1993) The localization of classical transmitters and neuropeptides within neurons in laminae 1-111 of the mammalian spinal dorsal horn (Review). Prog. Neurobiol. 41.609-645.
Tuchscherer, M.M., and V.S. Seybold 11985) Immunohistochemical studies of substance P, cholecystokinin-octapeptide and somatostatin in dorsal root ganglia of the rat. Neuroscience 14:593-605.
Van Steenis, G., and R. Kroes (1971) Changes in the nervous system and musculature of old rats. Vet. Pathol. 8t320-332.
Verge, V.M., R.J. Riopelle, and P.M. Richardson (1989) Nerve growth factor receptors on normal and injured sensory neurons. J. Neurosci. 9:914- 922.
Verge, V.M., J.P. Merlio, J. Grondin, P. Ernfors, H. Persson, R.J. Riopelle, T. Hokfelt, and P.M. Richardson 11992) Colocalization of NGF binding sites, trk mRNA, and low-affinity NGF receptor mRNA in primary sensory neurons: responses to injury and infusion of NGF. J. Neurosci. 12:40114022.
Verge, V.M., P.M. Richardson, Z. Wiesenfeld-Hallin, and T. Hokfelt (1995) Differential influence of nerve growth factor on neuropeptide expression in vivo: a novel role in peptide suppression in adult sensory neurons. J. Neurosci. 15:2081-2096.
Villar, M.J., R. Cortes, E. Theodorsson, Z. Wiesenfeld-Hallin, M. Schalling, J. Fahrenkrug, P.C. Emson, and T . Hokfelt (1989) Neuropeptide expres- sion in rat dorsal root ganglion cells and spinal cord after peripheral nerve injury with special reference to galanin. Neuroscience 33:587-604.
Vrontakis, M.E., L.M. Peden, M.L. Duckworth, and H.G. Friesen (1987) Isolation and characterization of a complementary DNA (galanin) clone from estrogen-induced pituitary tumor messenger RNA. J. Biol. Chem. 262:16755-16758.
Wakisaka, S., K.C. Kajander, and G.J. Bennett (1991) Increased neuropep- tide Y (NPY)-like immunoreactivity in rat sensory neurons following peripheral axotomy. Neurosci. Lett. 124.200-203.

320
Wakisaka, S., K.C. Kajander, and G.J. Bennett (1992) Effects of peripheral nerve injuries and tissue inflammation on the levels of neuropeptide Y-like immunoreactivity in rat primary afferent neurons. Brain Res. 598:349-352.
Watson, A., E. Ensor, A. Symes, J. Winter, G. Kendall, and D. Latchman (1995) A minimal CGRP gene promoter is inducible by nerve growth factor in adult rat dorsal root ganglion neurons but not in PC12 phaeochromacytoma cells. Eur. J. Neurosci. 7:394400.
Wetmore, C., and L. Olson (1995) Neuronal and nonneuronal expression of neurotrophins and their receptors in sensory and sympathetic ganglia suggest new intercellular trophic interactions. J. Comp. Neural. 353:143- 159.
Wiesenfeld-Hallin, Z., T. Bartfai, and T. Hokfelt (1992) Galanin in sensory neurons in the spinal cord [Review). Front. Neuroendocrinol. 13319- 343.
Willis J r , W.D., and R.E. Coggeshall (1991) Sensory Mechanisms of the Spinal Cord. New York: Plenum Press.
Wong, J., and M.M. Oblinger (1991) NGF rescues substance P expression but not neurofilament or tubulin gene expression in axotomized sensory neurons. J. Neurosci. 11:543-552.
Woollacott, M.H. (1986) Gait and postural control in the aging adult. In W. Bles and T.H. Brandt (eds): Disorders of Posture and Gait. Amsterdam: Elsevier, pp. 325-336.
Xu, X.J., Z. Wiesenfeld-Hallin, M.J. Villar, and T. Hokfelt (1990) Intrathecal
E. BERGMAN ET AL.
galanin antagonizes the facilitatory effect of substance P on the nocicep- tive flexor reflex in the rat. Acta Physiol. Scand. 137:463-464.
Ygge, J., and H. Aldskogius (1984) Intercostal nerve transection and its effect on the dorsal root ganglion. A quantitative study on thoracic ganglion cell numbers and sizes in the rat. Exp. Brain Res. 55:402408.
Young HI, W.S. (1990) In situ hybridization histoehemistry. In A. Bjorklund, T. Hokfelt, F.G. Wouterlood and A.N. van den Pol (eds): Handbook of Chemical Neuroanatomy: Analysis of Neuronal Microcircuits and Synap- tic Interactions. Amsterdam: Elsevier Science Publishers B.V., 8 pp. 481-512.
Zamboni, I., and C. De Martino (1967) Buffered picric acid-formaldehyde: A new, rapid fixative for electron microscopy. J. Cell Biol. 35:148A.
Zhang, X., G. Ju, R. Elde, and T. Hokfelt (1993a) Effect of peripheral nerve cut on neuropeptides in dorsal root ganglia and the spinal cord of monkey with special reference to galanin. J. Neurocytol. 22.342-381.
Zhang, X., B. Meister, R. Elde, V.M. Verge, and T. Hokfelt (1993b) Large calibre primary afferent neurons projecting to the gracile nucleus express neuropeptide Y after sciatic nerve lesions: An immunohistochemi- cal and in situ hybridization study in rats. Eur. J. Neurosci. 5:1510- 1519.
Zhang, X., Z.Q. Xu, L. Bao, b. Dagerlind, and T. Hokfelt (1995) Complemen- tary distribution of receptors for neurotensin and NPY in small neurons in rat lumbar DRGs and regulation of the receptors and peptides after axotomy. J. Neurosci. 15:2733-2747.





























