Embed Size (px)
Citation preview

Part III => METABOLISM and ENERGY
§3.2 Glucose Catabolism
§3.2a Glycolysis Pathway
§3.2b Glycolysis Regulation
§3.2c Fermentation

Section 3.2a:
Glycolysis

Synopsis 3.2a
- Dietary starch (eg bread, rice and potatoes) is hydrolyzed into glucose by the combined action of enzymes such as amylase (saliva) and maltase (small intestine)
- Glycolysis involves the breakdown/oxidation of glucose into pyruvate using a wide array of enzymes—and the free energy released in the process is either used to synthesize ATP or captured in the form of NADH
- In terms of chemical reactions, the glycolytic enzymes catalyze phosphorylation (transferase), isomerization (isomerase), bond cleavage (lyase), dehydrogenation (oxidoreductase), and hydrolysis (hydrolase)
- Of the six major classes/families of enzymes (see §2.5), all but ligase are involved in mediating glycolysis!
- The 10-reaction sequence of glycolysis is divided into two stages: Stage I Energy investment/expenditure Stage II Energy recovery/payoff
- Overall glycolytic reaction:Glucose + 2NAD+ + 2ADP + 2Pi <=> 2Pyruvate + 2NADH + 2ATP + 2H2O + 2H+
Glycolysis
Overview
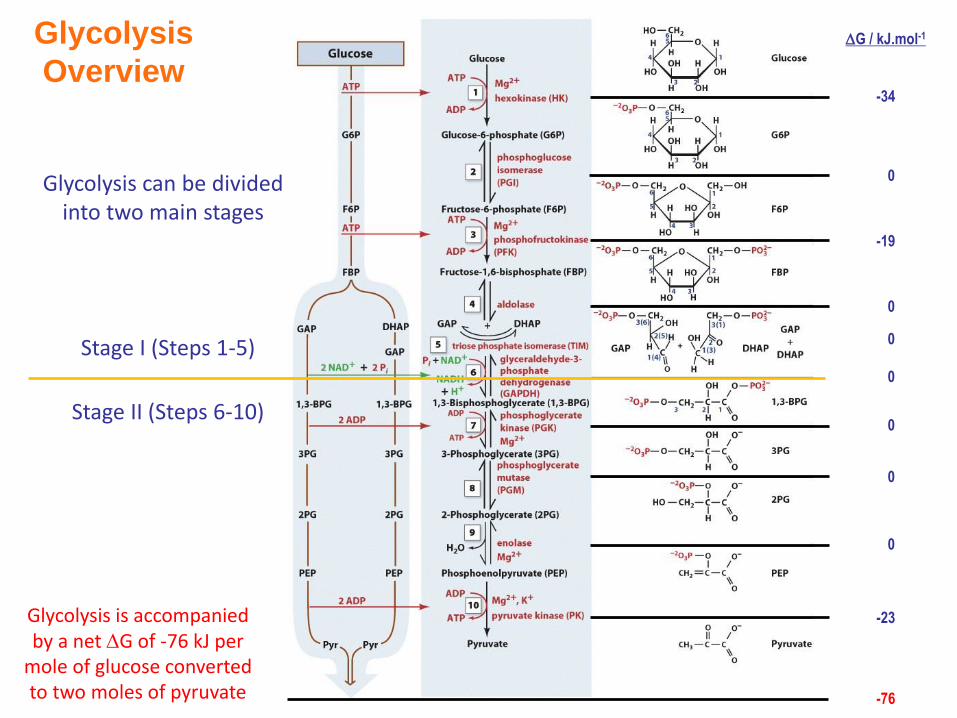
Glycolysis is accompanied by a net G of -76 kJ per
mole of glucose converted to two moles of pyruvate
Stage I (Steps 1-5)
Stage II (Steps 6-10)
G / kJ.mol-1
-34
0
-19
0
0
0
0
0
0
-23
-76
Glycolysis can be divided into two main stages
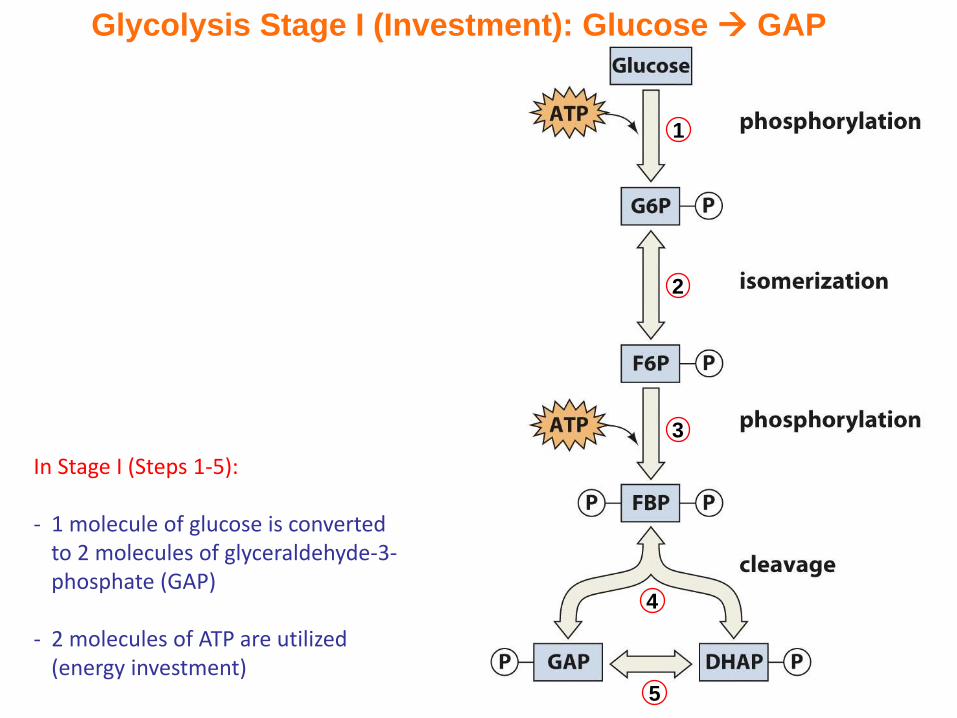
In Stage I (Steps 1-5):
- 1 molecule of glucose is converted to 2 molecules of glyceraldehyde-3-phosphate (GAP)
- 2 molecules of ATP are utilized (energy investment)
1
2
3
4
5
Glycolysis Stage I (Investment): Glucose GAP

(1) Glucose Glucose-6-Phosphate
- Transfer of the terminal phosphoryl group of ATP to glucose to generate G6P
- Thermodynamically favorable—powered by the free energy released due to ATP hydrolysis!
- Catalyzed by hexokinase (HK)—a non-specific enzyme that not only catalyzes the phosphorylation of glucose but also other hexoses such as mannose and fructose
- As is true for kinases in general, hexokinase requires Mg2+ divalent ions for catalytic activity—the Mg2+ ion is believed to shield the negative charges on - and -phosphate oxygen atoms within ATP, so as to render its -phosphate atom more accessible to nucleophilic attack by the –CH2OH group of glucose
G = -34 kJ/mol
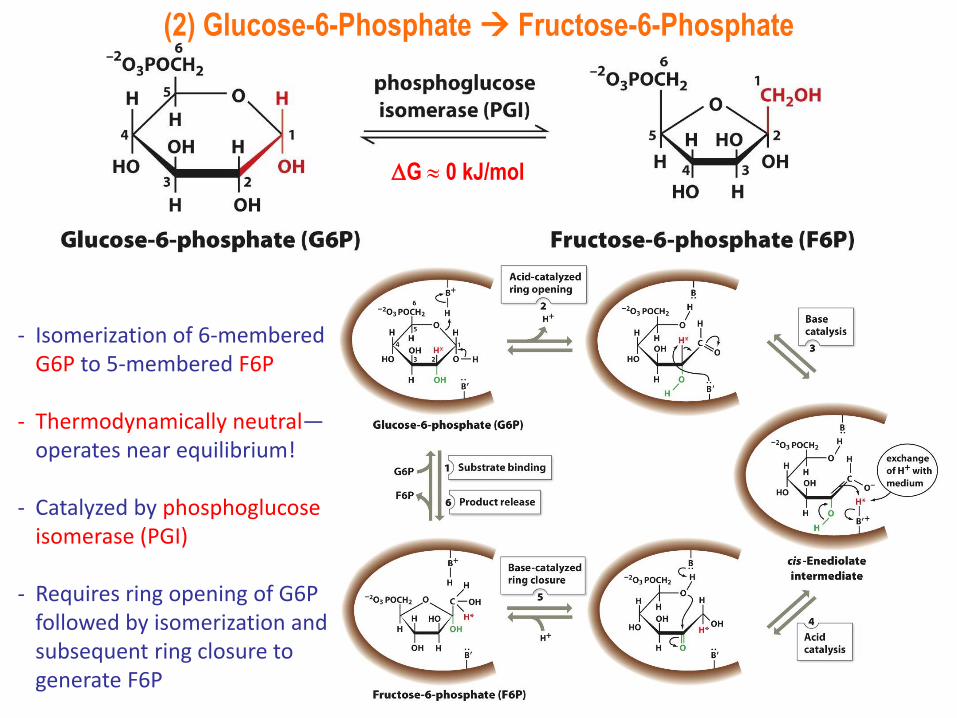
(2) Glucose-6-Phosphate Fructose-6-Phosphate
- Isomerization of 6-membered G6P to 5-membered F6P
- Thermodynamically neutral—operates near equilibrium!
- Catalyzed by phosphoglucoseisomerase (PGI)
- Requires ring opening of G6P followed by isomerization and subsequent ring closure to generate F6P
G 0 kJ/mol
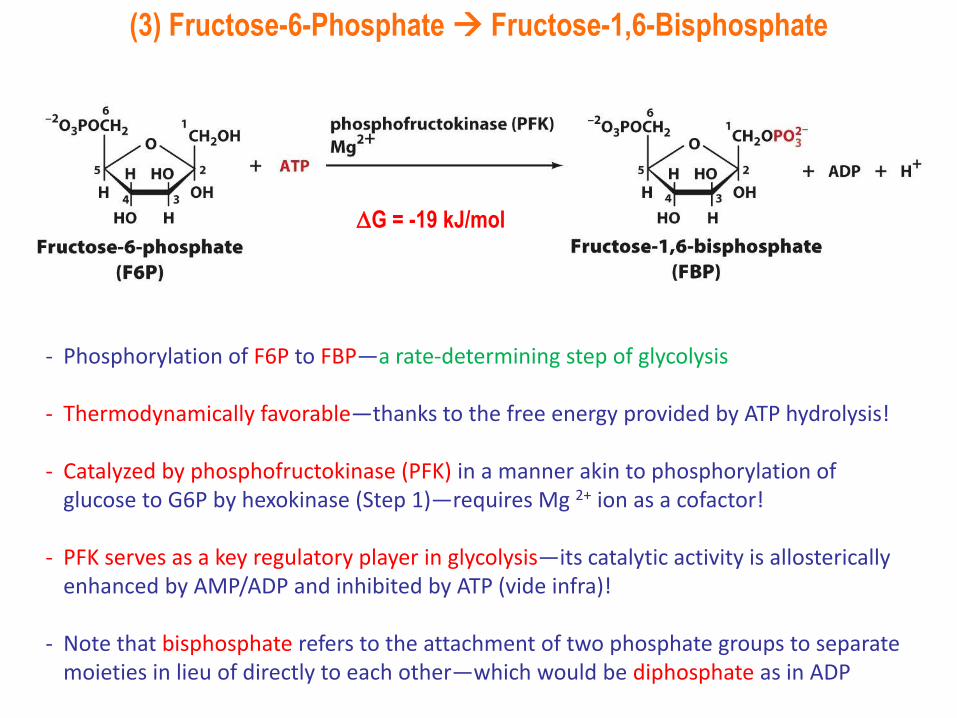
(3) Fructose-6-Phosphate Fructose-1,6-Bisphosphate
- Phosphorylation of F6P to FBP—a rate-determining step of glycolysis
- Thermodynamically favorable—thanks to the free energy provided by ATP hydrolysis!
- Catalyzed by phosphofructokinase (PFK) in a manner akin to phosphorylation of glucose to G6P by hexokinase (Step 1)—requires Mg 2+ ion as a cofactor!
- PFK serves as a key regulatory player in glycolysis—its catalytic activity is allostericallyenhanced by AMP/ADP and inhibited by ATP (vide infra)!
- Note that bisphosphate refers to the attachment of two phosphate groups to separate moieties in lieu of directly to each other—which would be diphosphate as in ADP
G = -19 kJ/mol

+
(4) Fructose-1,6-Bisphosphate GAP + DHAP
- Cleavage of a single 6-C compound (FBP) into two 3-C compounds (GAP and DHAP)
- Thermodynamically neutral—operates near equilibrium!
- Catalyzed by aldolase (ALD) into two interconvertible 3-C compounds—aldolase belongs to the lyase family that catalyze breaking/elimination of bonds by means other than hydrolysis!
- Aldolase is a portmanteau of aldol lyase—an aldol harbors both an hydroxyl and carbonyl group (ie aldol is an umbrella term for both aldoses and ketoses!)
- Note that the atom nomenclature changes upon cleavage of FBP—atoms 4/5/6 in FBP become atoms 1/2/3 in GAP, while atoms 1/2/3 in FBP become atoms 3/2/1 in DHAP
G 0 kJ/mol

(5) Dihydroxyacetone Phosphate Glyceraldehyde-3-Phosphate
- Interconversion of DHAP to GAP via an enediol intermediate
- Thermodynamically neutral—operates near equilibrium!
- Catalyzed by triose phosphate isomerase (TIM)—a “perfect” enzyme in that it operates near the diffusion-controlled limit with kcat/KM 109 M -1s-1 (see §2.7)
- DHAP and GAP are ketose-aldose isomers
- Only GAP continues along the glycolytic pathway—as GAP is siphoned off, more DHAP is converted to GAP due to “equilibrium shift”!
[GAP][DHAP]G 0 kJ/mol
triose
phosphate
isomerase

Glycolysis Stage II (Recovery): GAP Pyruvate
In Stage II (Steps 6-10):
- 2 molecules of GAP are converted to 2 molecules of pyruvate with concomitant generation of high-energy compounds (ATP and NADH)
- 4 molecules of ATP via substrate-level phosphorylation
- 2 molecules of NADH via reduction of NAD+
6
7
8
9
10

(6) Glyceraldehyde-3-phosphate 1,3-Bisphosphoglycerate
- Oxidation and phosphorylation of GAP to “high-energy” 1,3-BPG with concomitant release of NADH—cf the “high-energy” nature of mixed anhydride bond in 1,3-BPG vs the terminal phosphoanhydride bond in ATP!
- Hydrogen phosphate (HPO42-) used as a phosphate donor
- NAD+ used as an oxidizing agent to oxidize GAP
- Thermodynamically neutral—operates near equilibrium!
- Catalyzed by glyceraldehyde-3-phosphate dehydrogenase (GAPDH)—all dehydrogenases require a redox cofactor such as NAD+(H)/FAD(H2)
G 0 kJ/mol
HPO42-

(7) 1,3-Bisphosphoglycerate 3-Phosphoglycerate
- Dephosphorylation of 1,3-BPG to 3PG
- “High-energy” 1,3-BPG used as a phosphate donor to drive the synthesis of ATP from ADP via substrate-level phosphorylation (first ATP generation)
- Thermodynamically neutral—in spite of the free energy provided by the hydrolysis of “high-energy” 1,3-BPG, the reaction operates near equilibrium!
- Catalyzed by phosphoglycerate kinase (PGK)—requires Mg 2+ ion as a cofactor!
G 0 kJ/mol

(8) 3-Phosphoglycerate 2-Phosphoglycerate
- Isomerization of 3PG to 2PG in an intramolecular transfer of phosphate group
- Thermodynamically neutral—operates near equilibrium!
- Catalyzed by phosphoglycerate mutase (PGM)
- What do you call an isomerase that catalyzes the intramolecular transfer of a functional group from one position to another? Mutase!
G 0 kJ/mol
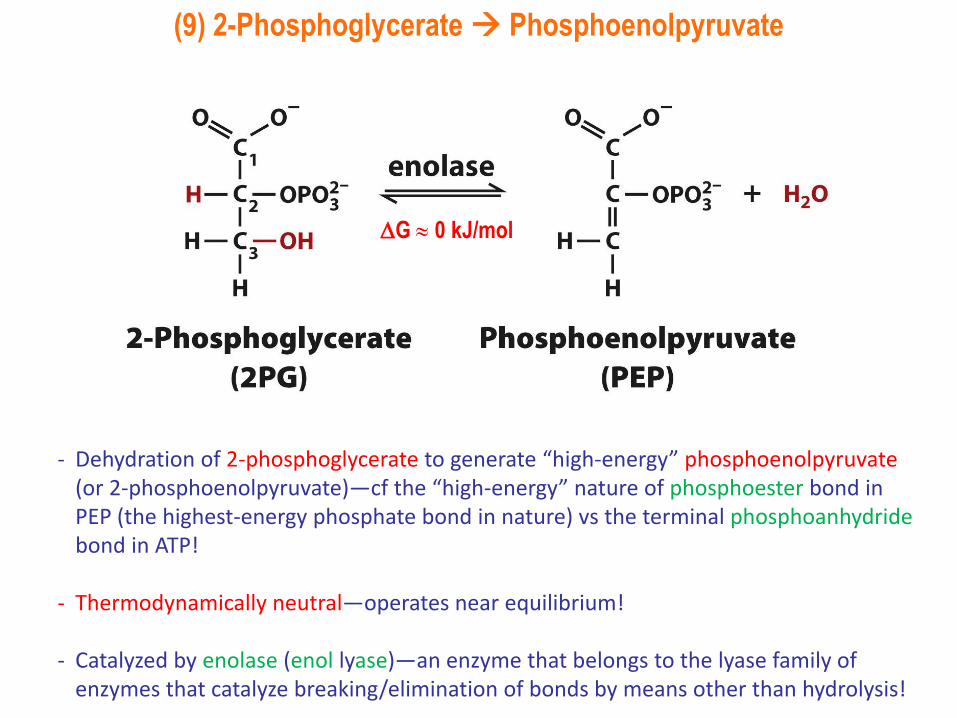
(9) 2-Phosphoglycerate Phosphoenolpyruvate
- Dehydration of 2-phosphoglycerate to generate “high-energy” phosphoenolpyruvate(or 2-phosphoenolpyruvate)—cf the “high-energy” nature of phosphoester bond in PEP (the highest-energy phosphate bond in nature) vs the terminal phosphoanhydridebond in ATP!
- Thermodynamically neutral—operates near equilibrium!
- Catalyzed by enolase (enol lyase)—an enzyme that belongs to the lyase family of enzymes that catalyze breaking/elimination of bonds by means other than hydrolysis!
G 0 kJ/mol
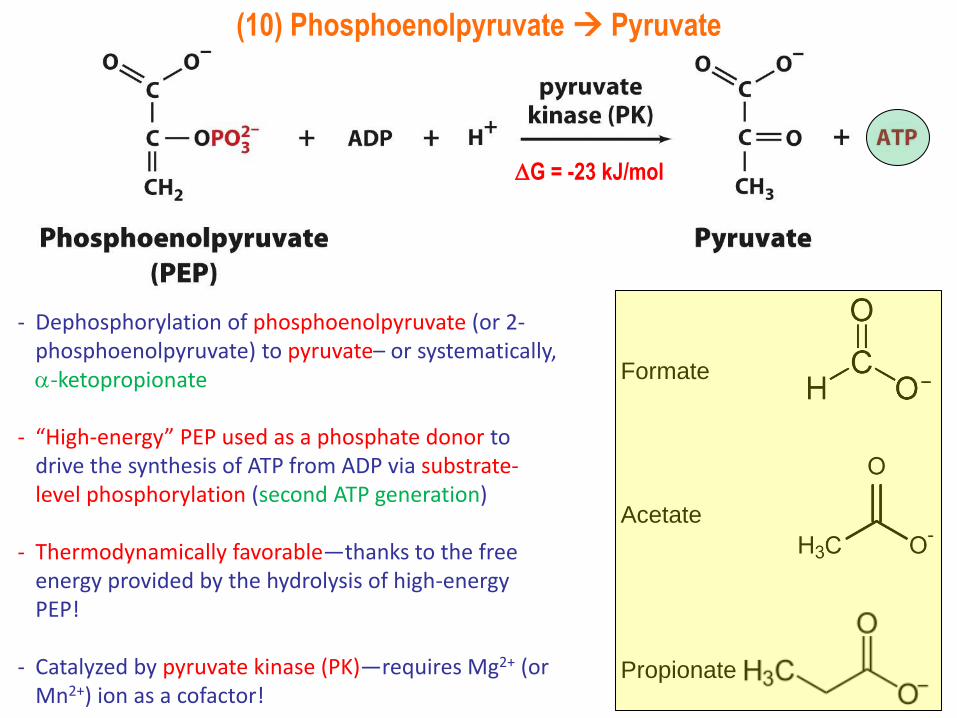
(10) Phosphoenolpyruvate Pyruvate
- Dephosphorylation of phosphoenolpyruvate (or 2-phosphoenolpyruvate) to pyruvate– or systematically,-ketopropionate
- “High-energy” PEP used as a phosphate donor to drive the synthesis of ATP from ADP via substrate-level phosphorylation (second ATP generation)
- Thermodynamically favorable—thanks to the free energy provided by the hydrolysis of high-energy PEP!
- Catalyzed by pyruvate kinase (PK)—requires Mg2+ (or Mn2+) ion as a cofactor!
G = -23 kJ/mol
Formate
Acetate
Propionate

Exercise 3.2a
- What happens during the two phases of glycolysis?
- How many ATP molecules are invested and how many are recovered from each molecule of glucose that follows the glycolytic pathway?
- Write the reactions of glycolysis, showing the structural formulas of the intermediates and the names of the enzymes that catalyze the reactions. Distinguish between thermodynamically neutral and favorable steps.
- Summarize the types of catalytic mechanisms involved. Do any glycolytic enzymes require cofactors?
- What “high-energy” compounds are synthesized during glycolysis?
Section 3.2b:
Glycolysis Regulation

Synopsis 3.2b
- Enzymes that function with a large negative G are candidates for flux-control points
- Phosphofructokinase (PFK), the major regulatory point for glycolysis in muscle, is allosterically inhibited by ATP and activated by AMP/ADP
- Substrate cycling allows the rate of glycolysis to respond rapidly to changing needs

Thermodynamics of Glycolysis (in erythrocytes)
- Recall that G = G + RT lnKeq (see §1.1)—where G is the actual free energy change under non-equilibrium (steady-state) conditions, and G is the standard free energy change @ equilibrium!
- Since cellular processes operate under steady-state rather than equilibrium setting, the free energy changes associated with various glycolytic steps are largely concerned with G!
- Of the 10 steps of glycolysis, only three (Steps 1/3/10) operate far from equilibrium (G << 0)—implying that they COULD be largely responsible for flux control!
Step Enzyme Name Enzyme Family G / kJ.mol-1 G / kJ.mol-1
1 Hexokinase (HK) Transferase -17 -34
2 Phosphoglucose isomerase (PGI) Isomerase +2 0
3 Phosphofrucktokinase (PFK) Transferase -14 -19
4 Aldolase (ALD) Lyase +24 0
5 Triose Phosphate Isomerase (TIM) Isomerase +8 0
6 Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) Oxidoreductase +6 0
7 Phosphoglycerate kinase (PGK) Transferase -19 0
8 Phosphoglycerate mutase (PGM) Isomerase +4 0
9 Enolase (ENO) Lyase +2 0
10 Pyruvate kinase (PK) Transferase -32 -23
Garrett R & Grisham CM (2005). Biochemistry (3rd ed). Belmont, CA: Thomson Brooks/Cole. pp 582–583.

G Profile for Glycolysis
Only Steps 1, 3 and 10 are associated
with large negative G!
PFK Is the Major Flux-Controlling Enzyme - Actual free energy changes (G) associated with various glycolytic
steps suggest that the major candidates for flux control are:- Hexokinase (HK) => Step 1- Phosphofructokinase (PFK) => Step 3- Pyruvate kinase (PK) => Step 10
- Of these three enzymes, only PFK plays a central role in controlling the rate of flow of metabolites (or flux) through glycolysis—why?!
- HK is not absolutely critical for glycolysis (much less serve as a regulatory point!)—since glycolysis in skeletal muscle often does not require HK (due to the breakdown of glycogen into glucose-6-phosphate via glycogenolysis)—see §3.3
- On the other hand, PK catalyzes the final step of glycolysis—thus its ability to control flux through glycolysis becomes somewhat moot
- Simply put, the conversion of F6P to FBP in Step 3 by PFK is the major rate-determining or rate-limiting (or the slowest) step of glycolysis
- How is PFK regulated? In PFK, ATP not only binds to the active site (to serve as a substrate) but also to an allosteric site (to serve as an allosteric inhibitor)
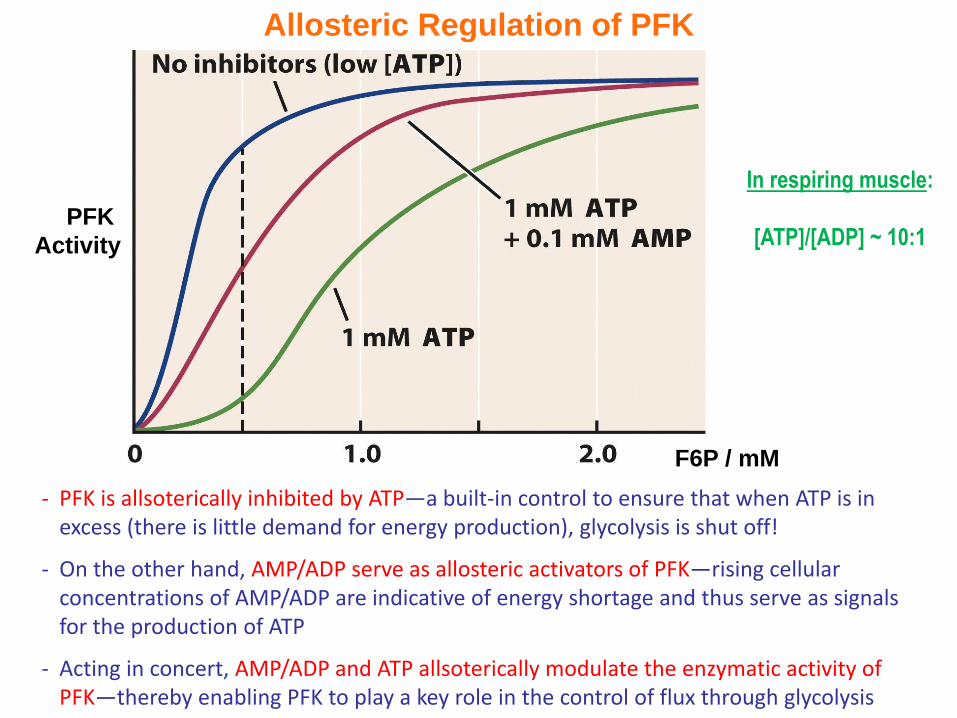
Allosteric Regulation of PFK
- PFK is allsoterically inhibited by ATP—a built-in control to ensure that when ATP is in excess (there is little demand for energy production), glycolysis is shut off!
- On the other hand, AMP/ADP serve as allosteric activators of PFK—rising cellular concentrations of AMP/ADP are indicative of energy shortage and thus serve as signals for the production of ATP
- Acting in concert, AMP/ADP and ATP allsoterically modulate the enzymatic activity of PFK—thereby enabling PFK to play a key role in the control of flux through glycolysis
F6P / mM
PFK
Activity
In respiring muscle:
[ATP]/[ADP] ~ 10:1
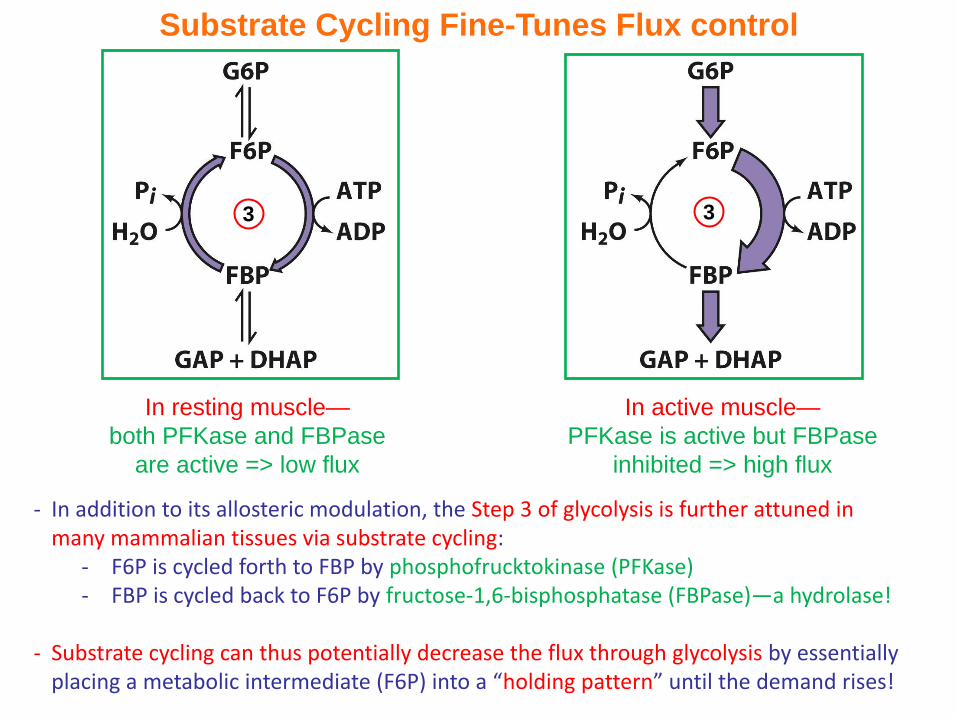
Substrate Cycling Fine-Tunes Flux control
- In addition to its allosteric modulation, the Step 3 of glycolysis is further attuned in many mammalian tissues via substrate cycling:
- F6P is cycled forth to FBP by phosphofrucktokinase (PFKase)- FBP is cycled back to F6P by fructose-1,6-bisphosphatase (FBPase)—a hydrolase!
- Substrate cycling can thus potentially decrease the flux through glycolysis by essentially placing a metabolic intermediate (F6P) into a “holding pattern” until the demand rises!
In resting muscle—
both PFKase and FBPase
are active => low flux
In active muscle—
PFKase is active but FBPase
inhibited => high flux
3 3

Exercise 3.2b
- Which glycolytic enzymes are potential control points?
- Describe the mechanisms that control phosphofructokinase activity
- What is the metabolic advantage of a substrate cycle?

Section 3.2c:
Fermentation

Synopsis 3.2c
- Under aerobic conditions, the glycolytic end-product pyruvate is completely oxidized to CO2 and H2O via the Krebs cycle (see §3.6)
- Under anaerobic conditions, the glycolytic end-product pyruvate is converted (or reduced) to either lactate or ethanol in a metabolic process referred to as “fermentation”
- Fermentation can be classified into two major groups:
(1) Homolactic (or lactic acid) fermentation:—largely occurs in the muscle cells of animals but also in some bacteria
(2) Alcoholic (or ethanol) fermentation:—largely occurs in yeast but also in some species of fish and plants
- In each case, the physiological significance of fermentation is to reoxidize/recycle NADH back to NAD+ so as to to ensure glycolytic continuity—thus fermentation plays a key role in the regeneration of energy under anaerobic conditions!

Metabolic Fate of Pyruvate
O2
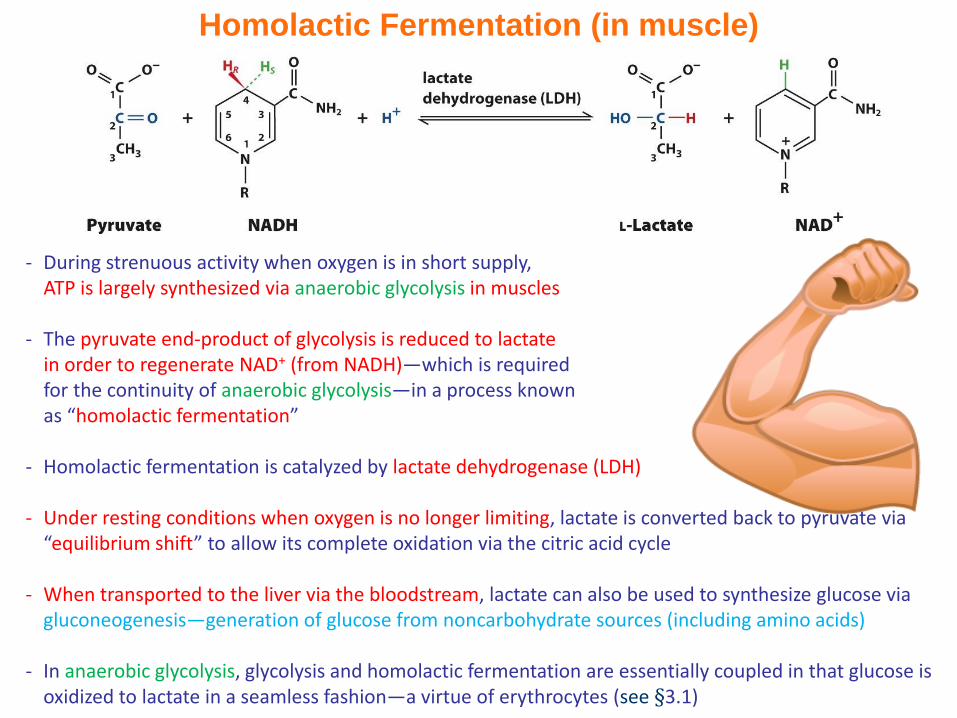
Homolactic Fermentation (in muscle)
- During strenuous activity when oxygen is in short supply, ATP is largely synthesized via anaerobic glycolysis in muscles
- The pyruvate end-product of glycolysis is reduced to lactate in order to regenerate NAD+ (from NADH)—which is required for the continuity of anaerobic glycolysis—in a process known as “homolactic fermentation”
- Homolactic fermentation is catalyzed by lactate dehydrogenase (LDH)
- Under resting conditions when oxygen is no longer limiting, lactate is converted back to pyruvate via “equilibrium shift” to allow its complete oxidation via the citric acid cycle
- When transported to the liver via the bloodstream, lactate can also be used to synthesize glucose via gluconeogenesis—generation of glucose from noncarbohydrate sources (including amino acids)
- In anaerobic glycolysis, glycolysis and homolactic fermentation are essentially coupled in that glucose is oxidized to lactate in a seamless fashion—a virtue of erythrocytes (see §3.1)

Alcoholic Fermentation (in yeast)
- Under anaerobic conditions, the major source of ATP in yeast is also obtained via anaerobic glycolysis
- The pyruvate end-product of glycolysis is ultimately converted via a two-step process called “alcoholic fermentation” to ethanol:
(1) Pyruvate decarboxylase (PDC) decarboxylates pyruvate to acetaldehyde using thiamine pyrophosphate (TPP) as a cofactor with concomitant release of CO2—which serves as a leavening agent in bread
(2) Alcohol dehydrogenase (ADH) subsequently reduces acetaldehyde to ethanol (the active ingredient in wine and beer) with concomitant regeneration of NAD+ (from NADH)—which is required for the continuity of anaerobic glycolysis
- Due to the release of CO2 as a by-product, alcoholic fermentation is commercially exploited in the production of alcoholic beverages as well as in the rising of bread (leavened bread)

Exercise 3.2c
- Describe the three possible fates of pyruvate
- Compare homolactic and alcoholic fermentation in terms of the products and the cofactors required