Embed Size (px)
Citation preview

UNIVERSITÀ DI PISA
Dipartimento di Scienze Agrarie, Alimentari e Agro-ambientali
Corso di Laurea Magistrale in Progettazione e Gestione del Verde
Urbano e del Paesaggio
RUOLO DELLE MOLECOLE SEGNALE
NELL’INDUZIONE DELLA MORTE CELLULARE IN
PIANTE DI QUERCUS ILEX ESPOSTE A OZONO
Candidato: Andrea Bianchi Relatore: Giacomo Lorenzini
Correlatore: Damiano Remorini
Anno Accademico 2014/2015

1
RIASSUNTO
Gli alberi presenti in ambito urbano svolgono differenti ruoli. Tra questi, uno dei più rilevanti
è legato al loro contributo nel miglioramento della qualità dell’aria e, di conseguenza, della
salute dell’uomo. In virtù degli effetti negativi indotti dalle odierne concentrazioni di ozono
troposferico (O3), in particolare in città, vi è la necessità di monitorare e valutare lo sviluppo di
specie arboree in previsione anche di scenari futuri critici. Scopo del presente lavoro è stato
quello di verificare il grado di sensibilità/resistenza di giovani piante (3 anni di età) di Quercus
ilex allo stress ossidativo, come quello imposto da O3 (200 ppb, 5 ore), simulando in ambiente
controllato le condizioni ambientali previste nel 2050. L’indagine ha preso in considerazione la
produzione di acqua ossigenata (H2O2) e anione superossido (O2•-), la risposta di etilene (ET),
acido abscissico (ABA), ac. salicilico (SA), ac. giasmonico (JA) e prolina (Pro) ai seguenti
tempi di analisi (1, 2, 5, 8, 24 ore dall’inizio del trattamento). L’esposizione all’O3 non ha
indotto la manifestazione di sintomi macroscopici su tessuti fogliari, sebbene dopo 5 e 24 ore
dall’inizio del trattamento sia possibile evidenziare, mediante colorazione con Evans blu e
DAB, rispettivamente gruppi di cellule morte e accumuli di H2O2. Nella prima ora di
trattamento si osserva un decremento (-27%) del contenuto di H2O2 rispetto ai controlli
mantenuti in aria filtrata. Successivamente tali valori rimangono inferiori o uguali a quelli
costitutivi fino al termine del periodo di recovery, dove si registra una evidente produzione
(+46%, rispetto ai controlli). Le concentrazioni di O2•- subiscono un significativo incremento
(+43%) già dopo un’ora dall’inizio della fumigazione, per poi mostrare un andamento
altalenante fino al termine del trattamento, in cui si nota un ulteriore aumento (+38%). I livelli
di ET e di ABA crescono durante l’intera durata dell’esperimento, raggiungendo un picco
rispettivamente al termine del periodo di recovery e nella prima ora di esposizione (+94 e
+188%). Le concentrazioni di SA (nella forma libera e coniugata) e JA subiscono un aumento
nelle prime ore di trattamento (circa tre e nove volte, rispettivamente) per poi diminuire al
termine del periodo di recovery (-58 e -73%, rispettivamente). Infine, i livelli di Pro registrano
un aumento nelle prime ore di esposizione (+47 e +37%, rispettivamente dopo 1 e 2 ore di
esposizione), per poi mostrare una progressiva stabilizzazione ai valori costitutivi. Solo al
termine dell’esperimento si nota un marcato decremento rispetto ai controlli mantenuti in aria
filtrata (-74%). Possiamo concludere che Q. ilex non mostra danni macroscopici indotti dal
trattamento con O3, sebbene a livello microscopico i risultati ottenuti dalla analisi istologiche
mostrino l’attivazione di morte cellulare programmata. L’H2O2 non svolge il suo noto ruolo di
molecola segnale durante il periodo di fumigazione, mentre O2•- mostra un andamento bifasico,
confermando la presenza di burst ossidativo a seguito del trattamento con l’inquinante. Gli
ormoni e le molecole segnale si attivano e interagiscono durante le varie fasi del trattamento,
documentando che l’O3 possa essere considerato alla stregua di un agente patogeno. In
particolare, si registra una produzione di ET per l’intera durata dell’esperimento coincidente
con quella di ABA che risponde in maniera più marcata e rapida, suggerendo che Q. ilex sia in
grado di attivare una strategia di esclusione (chiusura stomatica) in risposta all’inquinante. SA
e JA mostrano un andamento analogo facendo suppore che vi sia un’interazione sinergica che
regoli la diffusione della lesione.
Lavoro svolto nell’ambito del progetto PRIN 2010-2011 TreeCity.

2
INDICE
1. INTRODUZIONE ............................................................................................................................. 3
1.1. Il cambiamento climatico e lo scenario 2050 in ambiente urbano ......................................... 3
1.1.1. Ozono e cambiamento climatico ........................................................................................... 5
1.2. Inquinamento da ozono ............................................................................................................. 8
1.2.1. Formazione dell’ozono troposferico ..................................................................................... 8
1.2.2. Fitotossicità dell’ozono ....................................................................................................... 12
1.2.3. La risposta delle piante all’ozono ....................................................................................... 14
1.2.4. Ozono e morte cellulare programmata ............................................................................... 15
1.2.5. Interazioni tra le vie di segnalazione che influenzano la PCD ........................................... 16
1.3. Ormoni e molecole segnale in risposta allo ozono ................................................................. 19
1.3.1. Etilene ................................................................................................................................. 19
1.3.2. Acido abscissico .................................................................................................................. 20
1.3.3. Acido salicilico .................................................................................................................... 22
1.3.4. Acido giasmonico ................................................................................................................ 24
1.3.5. Prolina ................................................................................................................................. 27
1.4. Quercus ilex ............................................................................................................................... 28
2. SCOPO DELLA TESI .................................................................................................................... 28
3. MATERIALI E METODI .............................................................................................................. 31
3.1. Allevamento del materiale vegetale ed esposizione a stress idrico e/o ad ozono ................. 31
3.2. Liofilizzazione del materiale vegetale ..................................................................................... 32
3.3. Vitalità cellulare e localizzazione in situ di perossido di idrogeno ....................................... 32
3.4. Determinazione della produzione di radicali liberi ............................................................... 33
3.5. Determinazione del contenuto in fitormoni e molecole-segnale ........................................... 34
3.6. Trattamento statistico dei dati ................................................................................................ 37
4. RISULTATI E DISCUSSIONE ..................................................................................................... 37
4.1. Vitalità cellulare e localizzazione in situ di perossido di idrogeno ....................................... 37
4.2. ROS............................................................................................................................................ 39
4.3. Ormoni e molecole segnale ...................................................................................................... 40
5. CONSIDERAZIONI CONCLUSIVE ........................................................................................... 47
6. BIBLIOGRAFIA ............................................................................................................................. 48

3
1. INTRODUZIONE
1.1. Il cambiamento climatico e lo scenario 2050 in ambiente urbano
Il cambiamento climatico è un tema complesso, che è stato individuato come una delle
principali sfide che dovrà affrontare la società mondiale nel prossimo futuro. Sicuramente molti
dettagli non sono ancora noti, specialmente in relazione a (i) velocità con la quale avverranno i
mutamenti, (ii) precisa localizzazione dei fenomeni e (iii) distanza dai punti di non ritorno. In
ogni caso, si tratta di una certezza scientifica e vi sono evidenze che ne sia causa l’attività
antropica, in particolare attraverso l’emissione dei cosiddetti “gas serra” prevalentemente
prodotti da impianti di produzione di energia elettrica, cambiamenti nell’uso dei suoli
(deforestazione, in primo luogo), attività agricole intensive e soprattutto dai trasporti (Bussotti
et al., 2013). La preoccupazione per il XXI secolo è la rapidità con cui tale fenomeno si sta
verificando, portando alla crescente consapevolezza che l'attuale tasso di cambiamento
climatico può superare quello dei fenomeni adattativi e dei mutamenti evolutivi (Klein et al.,
2012).
Lo sviluppo delle città nel XX secolo è stato caratterizzato da eventi di urbanizzazione spinta
ed incontrollata, dovuti al rapido incremento della popolazione e allo sviluppo economico, con
conseguente degrado e, più spesso, scomparsa del verde. Già oggi, oltre la metà della
popolazione mondiale è classificata “urbana” e nel 2050 salirà al 70% (con punte dell’80% in
Europa). Le aree urbane, però, costituiscono soltanto il 3% della superficie totale del pianeta.
Nell’areale mediterraneo le diverse tipologie di microambienti influenzano considerevolmente
la vita delle piante in città (Gago et al., 2013). I principali aspetti di questi microclimi
(comunemente definiti “isole di calore”) sono di seguito riportati (Figura 1):
incremento delle temperature;
riduzione dell’escursione termica giornaliera;
distribuzione differenziale dei venti risultante dall’interferenza con le costruzioni e
conseguente canalizzazione di flussi d’aria lungo le vie di comunicazione;
quantità di acqua minore rispetto a quella presente nell’ambiente naturale.

4
Figura 1 - Fenomeno delle isole di calore urbane. I cambiamenti della copertura del suolo possono influire sulle superfici e
sulla temperature dell’aria (Giridharan et al., 2004).
A questo fenomeno, come detto, si devono aggiungere quei fattori su scala globale che
interferiscono sul regolare sviluppo delle specie vegetali. Tra questi, si annoverano: (i) crescenti
problemi legati all’inquinamento dell’aria; (ii) aumento delle temperature [con stime per il 2050
di un incremento di 1,3-1,8 °C della temperatura media mondiale e di 2-3 °C di quella del sud
dell’Europa (Christensen et al., 2007)]; (iii) riduzione delle risorse idriche disponibili [con
drastici cali delle precipitazioni medie annue in termini di frequenza ed intensità (Petit et al.,
2005; Rumukainen, 2012)]; (iv) deterioramento qualitativo delle acque (ad esempio la
progressiva salinizzazione delle falde, tossicità da metalli pesanti, etc.).
In questo contesto si riscontra la presenza di agenti di stress abiotici, che possono fortemente
influenzare la gestione delle colture (tanto da stimare che essi siano la causa di circa il 70%
delle perdite produttive, Boyer e Silk, 2004) e degli spazi verdi. In agricoltura, lo stress abiotico
è definito come “una qualunque pressione ambientale in grado di ridurre la produttività
potenziale di una coltura” (Vernieri et al., 2006). Nel caso delle specie ornamentali, per
riduzione della produttività normalmente si intende un calo in termini quantitativi (come accade
per fiori e/o fronde recise) e qualitativi (o meglio di valore estetico della pianta stessa). Solo
recentemente sono stati riaffermati i benefici sociali, ecologici, culturali ed economici del verde
ornamentale (Fini e Ferrini, 2007). In particolare, il ruolo degli alberi in città è legato ai loro
effetti sulla qualità dell’aria (Nowak et al., 2000). A tal proposito, essi garantiscono due ordini
di benefici (Cavanagh et al., 2009):
a) sequestrano e stoccano carbonio (C) atmosferico ed altri inquinanti gassosi;
b) garantiscono il miglioramento del microclima, riducendo i consumi energetici ed i
relativi carichi contaminanti (Nowak e Dwyer, 2007).

5
Tali funzioni sono spesso alterate da condizioni ambientali sfavorevoli, quali carenza idrica e
inquinamento dell’aria. Per questo motivo, l’approfondimento dei meccanismi morfologici e
fisiologici di risposta delle piante rappresenta uno strumento fondamentale per la comprensione
dei fenomeni d’acclimatazione e/o adattativi (in particolare, nelle specie arboree caratterizzate
da un lungo ciclo vitale, Ordòñez et al., 2010) già esistenti - quali (i) modifica della crescita e
delle fasi fenologiche per massimizzare il guadagno di C, evitando danni da gelo in inverno
(Cleland et al., 2007) ed (ii) alcune caratteristiche morfo-fisiologiche specifiche, che
permettono la sopravvivenza durante la siccità estiva (De Micco e Aronne, 2012) - e futuri,
come lo spostamento di areale di alcune specie e conseguente alterazione delle serie dinamiche
della vegetazione (Cheaib et al., 2012; Vitale et al., 2012).
Gli studi relativi alla risposta a stress abiotici, quali quelli sinora riportati, sono molti e indicano
come i meccanismi di risposta siano spesso simili e collegati tra loro, riconducendo agli stessi
processi di adattamento e acclimatazione. In particolare, carenza idrica, elevata salinità, freddo
o caldo eccessivo ed inquinamento dell’aria sono fattori primari di stress, ma che possono essere
tutti ricondotti ad un’unica tipologia: lo stress ossidativo (Vinocur e Altman, 2005). In generale,
l’adattamento delle piante a stress di natura ambientale è controllato da una cascata di segnali
biochimici che attivano meccanismi di risposta tali da ripristinare i processi omeostatici e di
riparazione (Wang et al., 2003).
Alla luce di tale situazione, è necessario conoscere in che modo il cambiamento climatico incida
sugli alberi, così da definire le linee guida sulla scelta e poter valutare le condizioni ambientali
di crescita.
1.1.1. Ozono e cambiamento climatico
Il cambiamento climatico è strettamente collegato all’incremento dell’inquinamento
atmosferico. Molti dei tradizionali inquinanti dell’aria e gas serra non solo hanno fonti comuni,
ma possono anche interagire sia fisicamente che chimicamente nell’atmosfera causando una
varietà di effetti ambientali su scala locale, regionale e globale (Bytnerowicz et al., 2007). Tra
questi, l’ozono (O3) troposferico rappresenta il più fitotossico, in virtù della sua distribuzione
capillare e della elevata attività biologica (Bytnerowicz et al., 2007). E’ stato stimato che la
concentrazione media diurna di questo contaminante durante la stagione fotochimica
nell’emisfero settentrionale (Fowler et al., 1999) risulta quattro volte maggiore rispetto a quella
rilevata nel periodo preindustriale (Volz e Kley, 1988). Modelli previsionali forniscono stime

6
riguardanti incrementi dei livelli di O3 del 20-25% tra il 2015 e il 2050 e di un ulteriore 40-60%
entro il 2100, a causa delle future condizioni climatiche (Meehl et al., 2007).
In particolare, le aree del bacino del Mediterraneo risultano sensibilmente esposte all’aumento
della concentrazione di O3, poiché caratterizzate da un’intensa radiazione luminosa ed elevate
temperature medie, oltre che dalla presenza di molte specie vegetali che emettono composti
organici volatili (COV). Tali condizioni risultano favorevoli per la formazione di questo
contaminante (Chameides et al., 1994).
Nella Figura 2 vengono riportati i valori massimi di O3 previsti per il 2050, secondo quanto
stimato da Community Atmosphere Model with Chemistry (CAM-Chem, Lei et al., 2012). Tale
studio mostra i cambiamenti climatici rappresentati in tre scenari e il rafforzamento dovuto alle
mutate condizioni meteorologiche, favorendo il rischio di episodi di inquinamento da O3 in
molte parti del mondo. La differenza della concentrazione di questo inquinante nelle varie
regioni per il 2050 dipende in larga misura dai mutamenti nelle emissioni antropogeniche dei
precursori.
Figura 2 - Simulazione della concentrazione di ozono durante i mesi estivi (giugno-agosto) tramite Community Atmosphere
Model with Chemistry (CAM-Chem), basato su concentrazioni massime giornaliere standard di ozono (espresse in ppb e
calcolare su 8 h). a) 2000; b) proiezione 2050 dello scenario B1 (limite superiore) come differenza dal 2000; c) proiezione
2050 dello scenario A1B (limite medio) come differenza dal 2000; d) proiezione 2050 dello scenario A1FI (limite inferiore)
come differenza dal 2000 (Lei et al., 2012). I differenti scenari climatici, distinti dall’IPCC per ottenere una comprensione più
completa dei possibili cambiamenti.

7
Il futuro incremento dei livelli di O3 non solo può indurre significative perdite in ambito
agricolo e forestale (UNECE, 2004; Hopkin, 2007; Chuwah et al., 2015), ma può aggravare
ulteriormente gli effetti derivanti dal cambiamento climatico, a causa del sinergismo con altri
inquinanti gassosi, in particolare l’anidride carbonica (CO2), come riportato da Stich et al.
(2007).
Nell’estate 2013, l’obiettivo a lungo termine per la protezione della salute umana (che fissa la
media massima giornaliera, calcolata su 8 ore nell’arco di un anno civile, alla massima
concentrazione di O₃ presente di 61 ppb) indicato nel D.Lgs. n. 155 del 2010 (il quale recepisce
la Direttiva 2008/50/CE) è stato superato per più di 25 giorni in buona parte dell’Europa. La
concentrazione di ozono ha superato il limite di guardia (122 ppb a 1 ora dall’esposizione) 27
volte in Austria, Belgio, Italia, Portogallo e Kosovo (EEA, 2013). Le analisi mostrano che:
a) i superamenti si verificano soprattutto nel nord Italia e in zone dove si registrano
abitualmente concentrazioni oltre i limiti di guardia;
b) più dell’80% delle stazioni prese in esame mostrano almeno un superamento della
media massima giornaliera durante il periodo estivo;
c) i superamenti dei limiti di guardia si sono concentrati tra i mesi di luglio e agosto,
mentre il periodo precedente alla metà di giugno e successivo all’otto agosto è stato
caratterizzato da bassissima incidenza di inquinamento;
d) l’estate 2013 continua a mostrare una tendenza positiva, registrando un abbassamento
del 2% nei fenomeni di superamento dei limiti di guardia e di media giornaliera fissata
per legge rispetto all’anno 2012 (rispettivamente da 28% a 26% e da 85% a 83% delle
stazioni totali);
e) le concentrazioni di O3 e dei suoi precursori non sono immobili, ma possono spostarsi
a lunga distanza rispetto al punto di origine. A causa del trasporto internazionale di
inquinanti da zone extra-europee verso il continente, la difficoltà di creare una
corrispondenza tra i dati ottenuti dalle analisi con le misure legislative attuate in
Europa per la riduzione delle emissioni di sostanze inquinanti è evidente.
L’inquinamento da O3 non può più essere considerato una questione di qualità dell’aria da
affrontare a livello locale, ma si tratta di un problema globale che richiede una soluzione globale
(Royal Society, 2008).

8
1.2. Inquinamento da ozono
1.2.1. Formazione dell’ozono troposferico
L’O3 è la forma triatomica dell’ossigeno, di colore azzurrino, di odore agliaceo pungente (il
limite di percezione olfattiva è circa 10 ppb; 1 ppb = 1,96 μg m-3 in condizioni standard di
temperatura e di pressione) e con un grande potenziale ossidoriduttivo (+2,07 V), in virtù del
quale attacca direttamente matrici animali e vegetali, nonché materiali non biologici. Dal 1851
è nota la sua azione irritante a carico dei polmoni; ad esso vengono attribuiti ruoli primari anche
nei numerosi casi di mortalità associati alle “ondate di calore” che spesso affliggono il nostro
Paese in estate (Lorenzini et al., 2014b). Perciò questo inquinante è da tempo oggetto di
interventi normativi in sede sia nazionale sia comunitaria. E’ presente in tutti gli strati
dell’atmosfera: sia superiori (stratosfera) che inferiori (troposfera). La maggior concentrazione
si trova tra 18-50 km dalla superficie terrestre, occupando circa il 97% del volume totale
dell’involucro gassoso. Nella stratosfera forma uno scudo protettivo contro la maggior parte
delle radiazioni ultraviolette (UV, bassa lunghezza d’onda) provenienti dal Sole,
potenzialmente dannose per la vita sulla biosfera a causa della loro azione mutagena. Qui,
secondo il meccanismo di Chapman (1930), il gas si forma per reazione dell’O2 con la sua
forma monoatomica (O) generata per fotolisi dai raggi UV, in presenza di una molecola (M),
in grado di assorbire l’energia in eccesso (Chameides et al., 1994):
O2 + hv (λ ≤ 242 nm) → O + O
O + O2 + M → O3 + M
Dove:
h: costante di Planck;
v: frequenza del fotone assorbito;
λ: lunghezza d'onda del fotone assorbito;
M: specie molecolare di fondo [di fatto, azoto (N2)].
A livello troposferico, l’O3 è prodotto da una reazione in equilibrio O2/O3, che coinvolge gli
ossidi di azoto (NOx) e la luce (Rao et al., 2000) e porta alla formazione e/o alla distruzione in
uguale quantità di O3 e monossido di azoto (NO), in quello che prende il nome di “ciclo
indisturbato del biossido di azoto (NO2)” (Figura 3a). Tale ciclo, che di per sé è naturale, si
verifica in zone non inquinate senza determinare episodi di inquinamento fotochimico. Possono
anche verificarsi fenomeni di intrusione del gas negli strati più bassi grazie a scambi tra
stratosfera e troposfera, che avvengono solo in particolari condizioni climatiche (Junge, 1962).

9
Questi e altri eventi, come le scariche elettriche durante i temporali, mantengono l’O3 “al suolo”
ad un livello di background naturale pari al massimo a 10-20 ppb.
Figura 3 - A) Ciclo fotolitico indisturbato del biossido di azoto; B) interazione con gli idrocarburi nella formazione dello smog
fotochimico (da Lorenzini e Nali, 2005).
Al contrario, in atmosfere inquinate la formazione di NO2 segue vie alternative che non
implicano la rimozione dell’O3, per cui si ha un accumulo notevole di questo inquinante.
Quando nella troposfera sono presenti anche specie chimicamente molto attive, quali gli
idrocarburi, il suddetto equilibrio viene spostato verso l’aumento dell’O3, attraverso la
sottrazione diretta di NO a formare altri composti (Figura 3b), anch’essi contaminanti. Inoltre,
la fotossidazione del monossido di carbonio (CO), la presenza di idrocarburi incombusti [come
il metano (CH4)], formaldeide e altre sostanze organiche non metaniche, possono contribuire
alla formazione di O3, nell’ambito del cosiddetto smog fotochimico (Fishman et al., 1985):
CO + 2O2 + hv → CO2 + O3
CH4 + 4O2 + 2hv → HCOH + H2O + 2O3
RH + 4O2 + 2hv → R’CHO + H2O + 2O3
Le dinamiche degli NOx rivestono dunque particolare importanza nei processi di formazione e
distruzione di questo inquinante. Da ciò, si evince che l’O3 non è soltanto il prodotto
quantitativamente più importante dei processi di smog fotochimico, ma è anche uno degli
A
B

10
elementi di innesco che attivano i processi stessi. Infatti, l’O3 subisce una dissociazione chimica
ad una lunghezza d’onda compresa tra 280 e 310 nm, formando atomi di O2 eccitati che
velocemente reagiscono con il vapore d’acqua formando radicale ossidrile:
O3 + hν → O2 + O•
O• + H2O → OH + OH•
Tutte queste reazioni conducono ad un’alterazione dell’equilibrio del ciclo foto-stazionario a
favore della produzione di O3 (Derwent et al., 1987) e permettono di comprendere per quale
motivo gli episodi acuti di smog fotochimico spesso persistono, con intensità crescente per più
giorni consecutivi.
Spesso i livelli più alti di polluzione si registrano in prossimità delle fonti di emissione, come
nelle aree urbane e/o industriali; tuttavia, considerata la facilità con la quale si spostano i
precursori, è frequente il caso in cui la concentrazione di O3 sia superiore in aree poste alcune
decine di chilometri sotto vento rispetto alle città, che non negli agglomerati stessi. Inoltre,
proprio nelle zone più inquinate si verificano con maggiore intensità, le reazioni che portano
alla diminuzione dell’O3 (“titolazione”) in particolare la combinazione con NO (Lorenzini e
Nali, 2005; Lorenzini et al., 2014b).
La possibilità di trasporto a lunghe distanze (anche a diverse centinaia di chilometri) dei
precursori e dello stesso O3 in aree remote, rurali e forestali (e in alto mare) è ben dimostrata.
Ad esempio, la maggior parte dell’inquinante che si riscontra sull’isola di Creta trova la sua
origine nella terraferma (Kouvarakis et al., 2000). Situazione analoga è stata riscontrata a
Tenerife (Isole Canarie, Spagna), dove i venti provenienti prevalentemente da nord-est
trasportano il contaminante da zone a latitudini nordiche, così da contribuire, insieme a quello
prodotto sull’isola, agli alti livelli di AOT40 (Accumulated exposure Over a Threshold of 40
ppb, Guerra et al., 2004).
Studi effettuati con tecniche di biomonitoraggio hanno evidenziato la presenza di significative
concentrazioni dell’ossidante nelle isole minori dell’Arcipelago Toscano (Gorgona e Capraia),
dove la produzione locale di precursori è irrilevante (Lorenzini et al., 1995). Quindi, nonostante
che la formazione di O3 sia tipica delle aree urbane con elevata densità di traffico, questa
polluzione è facilmente trasportato verso le zone suburbane e periferiche con risvolti negativi
sulla crescita e sullo sviluppo di colture agricole, essenze forestali e vegetazione spontanea
(Lefohn, 1992). I risultati di programmi di monitoraggio su scala internazionale hanno
11
confermato che il problema non è di soluzione locale, ma necessita di un intervento su scala
multi-regionale, attraverso piani di cooperazione.
Nel meccanismo di formazione dell’O3 troposferico svolgono un ruolo fondamentale la
componente antropogenica e quella meteoclimatica. Di conseguenza, questo inquinante è
caratterizzato da una distribuzione geografica peculiare, strettamente legata alla concentrazione
dei precursori e alle condizioni atmosferiche, e da un andamento sia stagionale che giornaliero.
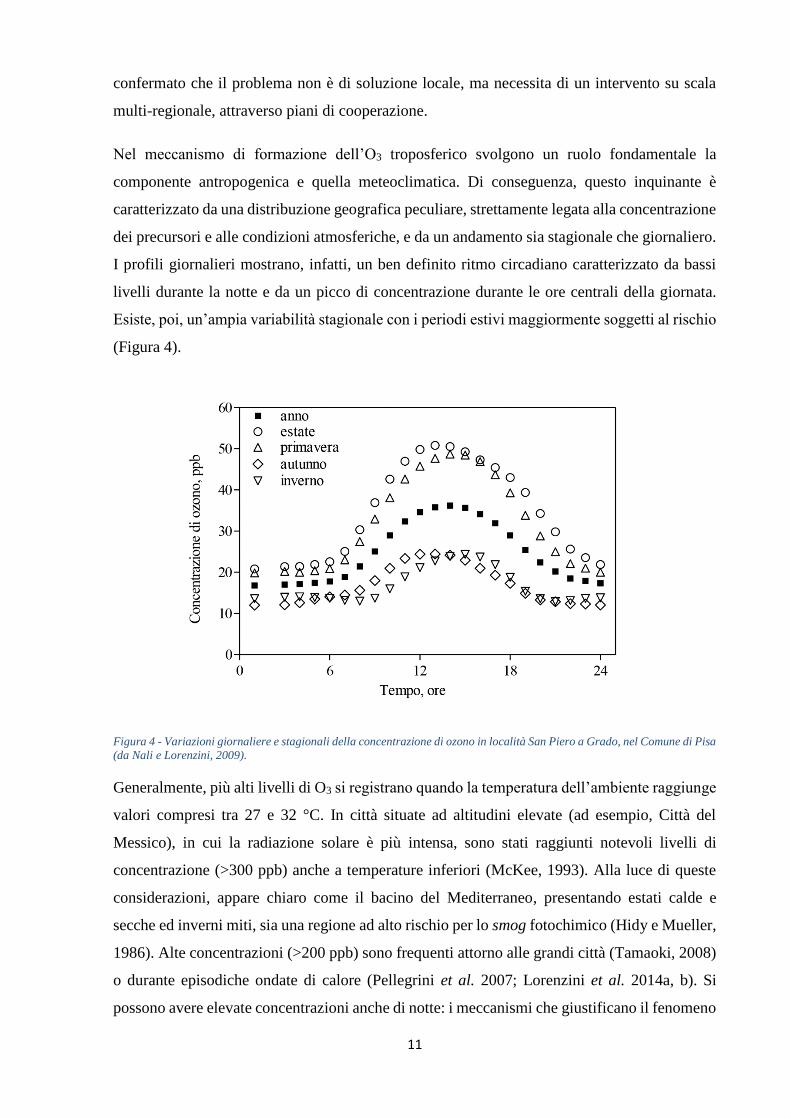
I profili giornalieri mostrano, infatti, un ben definito ritmo circadiano caratterizzato da bassi
livelli durante la notte e da un picco di concentrazione durante le ore centrali della giornata.
Esiste, poi, un’ampia variabilità stagionale con i periodi estivi maggiormente soggetti al rischio
(Figura 4).
Figura 4 - Variazioni giornaliere e stagionali della concentrazione di ozono in località San Piero a Grado, nel Comune di Pisa
(da Nali e Lorenzini, 2009).
Generalmente, più alti livelli di O3 si registrano quando la temperatura dell’ambiente raggiunge
valori compresi tra 27 e 32 °C. In città situate ad altitudini elevate (ad esempio, Città del
Messico), in cui la radiazione solare è più intensa, sono stati raggiunti notevoli livelli di
concentrazione (>300 ppb) anche a temperature inferiori (McKee, 1993). Alla luce di queste
considerazioni, appare chiaro come il bacino del Mediterraneo, presentando estati calde e
secche ed inverni miti, sia una regione ad alto rischio per lo smog fotochimico (Hidy e Mueller,
1986). Alte concentrazioni (>200 ppb) sono frequenti attorno alle grandi città (Tamaoki, 2008)
o durante episodiche ondate di calore (Pellegrini et al. 2007; Lorenzini et al. 2014a, b). Si
possono avere elevate concentrazioni anche di notte: i meccanismi che giustificano il fenomeno

12
sono tre: (i) trasporto orizzontale da altre località, (ii) discesa dalla stratosfera, (iii)
intrappolamento dell’O3 durante il giorno in uno strato di inversione termica (Lorenzini e Nali,
2005).
1.2.2. Fitotossicità dell’ozono
Il danno da O3 è il risultato di reazioni biochimiche e fisiologiche che avvengono nel mesofillo
fogliare, con ripercussioni in tutto l’organismo vegetale. Dato che la penetrazione attraverso
cuticola è insignificante (Kerstiens et al., 1989), gli stomi della pagina abassiale (Reich, 1987)
giocano un ruolo fondamentale per l’entrata dell’O3 nell’apoplasto. Il controllo delle aperture
stomatiche nella pianta e la restrizione del flusso nelle foglie si dimostrano “la prima linea di
difesa” (Vainonen e Kangasjärvi, 2015) per raggiungere l’acclimatazione (Hoshika et al.,
2013). L’O3 generalmente abbassa la conduttanza stomatica poche ore dopo l’inizio
dell’esposizione (inoltre restringe il flusso di CO2 nelle foglie), mantenendo questa condizione
per parecchie ore (Vahisalu et al., 2010). La sensibilità agli stimoli esterni è un importante
fattore nella regolazione dei movimenti stomatici e, conseguentemente, nel rapporto
sensibilità/tolleranza all’O3. Il controllo indotto dall’inquinante sulle aperture stomatiche via
etilene (ET) può promuovere la loro apertura e chiusura, in relazione a (i) stress aggiuntivi subiti
e (ii) intensità subita (Wilkinson et al., 2011). Superati gli stomi, l’O3 si trova all’interno della
foglia, si diffonde negli spazi intercellulari del mesofillo e nella parete cellulare e, raggiunto il
plasmalemma, attacca le molecole proteiche e quelle lipidiche (Rao et al., 2000).
L’O3 è particolarmente tossico per gli organismi vegetali per una serie di caratteristiche. Oltre
al suo alto potenziale redox, ha approssimativamente lo stesso coefficiente di diffusione della
CO2 e incontra, quindi, la stessa resistenza nella penetrazione fogliare e di membrana. Inoltre,
la sua solubilità in acqua (H2O) (e, quindi, nel liquido cellulare) è dieci volte maggiore dell’O2;
infine, reagisce con l’H2O maggiormente in ambiente alcalino (Colbeck e MacKenzie, 1994).
Secondo Gill e Tuteja (2010), l’O3 si dissocia rapidamente in soluzione acquosa, generando
specie attive dell’O2 (ROS), come l’anione superossido (O2•-) e il perossido di idrogeno (H2O2),
ma anche, secondariamente alla reazione con altre biomolecole, ossigeno singoletto (-1O2) e
radicale ossidrile (OH•). Quest’ultimo è il più attivo e potenzialmente dannoso per le strutture
biologiche, mentre H2O2 e O2•- sono meno reattivi in soluzione acquosa ed hanno la capacità di
spostarsi per poche distanze molecolari (Halliwell e Gutteridge, 1989). Inoltre, il decadimento
dell’O3 nel mezzo acquoso si realizza con un tasso significativo soltanto ad elevati livelli di pH
(circa 8) e, di conseguenza, la decomposizione diretta dell’inquinante è più lenta rispetto alle

13
reazioni con le molecole biologiche (Kanofsky e Sima, 1995), che assumono, quindi, un ruolo
principale nella produzione di ROS. Queste sono altamente reattive, tossiche e causano danno
a (i) proteine, (ii) lipidi, (iii) carboidrati e (iv) DNA, provocando la morte cellulare.
Sia l’O2•- che l’H2O2 reagiscono con i metalli di transizione nello stato ridotto (specialmente
Fe2+ e Cu+, rilasciati, ad esempio, dalle metallo-proteine dopo l’attacco ossidativo delle stesse
ROS), generando il radicale idrossile OH• attraverso le reazioni di Haber-Weiss e di Fenton:
H2O2 + Fe2+ → OH- + OH• + Fe3+ (reazione di Fenton)
O2•- + Fe3+ → O2 + Fe2+
H2O2 + O2•- → O2 + OH- + OH• (reazione di Haber-Weiss)
L’O3 interagisce anche con molecole organiche contenenti doppi legami C-C (Tamaoki, 2008),
andando così ad incrementare la produzione di radicali liberi sia attraverso l’addizione dell’O3
al doppio legame, con formazione dell’ozonide, sia tramite l’incorporazione di un atomo di O3
in un prodotto ossidato, attraverso la formazione di epossido e -1O2 (Kanofsky e Sima, 1995):
Sulla base della misurazione del flusso di O3 nella foglia (Laisk et al., 1989), il contaminante
non penetra profondamente negli spazi intercellulari, ma si decompone rapidamente a livello
della parete cellulare e della membrana plasmatica. Poiché entrambe le strutture contengono
numerose molecole organiche con doppi legami C-C (gruppi fenolici, composti olefinici,
proteine amidiche, ecc.), questo è il primo sito cellulare di reazione dell’O3. L’inquinante può
partecipare direttamente alla reazione di ozonolisi, attaccando le molecole lipidiche insature e
generando H2O2 e aldeidi, in particolare malondialdeide (MDA) e 4-idrossinonenale (HNE,
Figura 5), ma anche alcani, epossidi, alcoli e altri prodotti di degradazione che possono
facilmente interagire con DNA e proteine, con successiva formazione di pericolosi coniugati
(Heath, 1987; Davies, 2001; Fam e Morrow, 2003). In alternativa, può avvenire che l’OH•, o
un’altra ROS, rimuova un atomo di idrogeno da un gruppo metile della catena, producendo,

14
successivamente alla reazione con l’O2, un radicale perossile (Halliwell e Gutteridge, 1989) nel
processo che prende il nome di perossidazione lipidica:
L’O3 ha anche la capacità di inattivare le proteine di membrana attraverso reazioni di
ossidazione e tramite la modificazione dei gruppi funzionali degli aminoacidi (quali cisteina,
metionina e triptofano); nel primo, i gruppi –SH di due molecole vanno a formare ponti S-S
con conversione a cistina; nel secondo, l’atomo di zolfo è ossidato a sulfossido; nel terzo, si ha
la rottura dell’anello pirrolico. L’aggressione alle proteine coinvolge anche gli enzimi in seguito
ad alterazioni della struttura secondaria e terziaria (Kelly e Mudway, 2003), nonché le ATPasi
associate alla pompa ionica di membrana che possono essere inattivate, influenzando il
trasporto fino al dissesto dell’osmoregolazione (Singh et al., 2010). Il danno che ne deriva
favorisce l’ingresso degli ioni Ca2+ dall’apoplasto.
Figura 5 - L’attività delle ROS a carico dei lipidi di membrana porta a formazione di aldeidi (MDA, HNE) estremamente
reattive all’interno della cellula (da Gill e Tuteja, 2010).
1.2.3. La risposta delle piante all’ozono
La tossicità dipende essenzialmente da tre fattori: la penetrazione all’interno dei tessuti fogliari,
l’influenza dell’inquinante sui costituenti cellulari e, infine, le eventuali reazioni di difesa.
Riguardo al primo punto, è da tempo nota una notevole variabilità inter- e intraspecifica. Una
schematica rappresentazione dei meccanismi alla base di questi comportamenti differenziali,
riferita alla nota “Teoria generale della risposta agli stress” è riportata in Figura 6.

15
La prima possibile strategia (esclusione) prevede la chiusura degli stomi, con conseguente
diminuzione nell’assorbimento dei gas (compresa la CO2); questa via risulta percorribile nel
caso di esposizioni acute, quindi, limitate ad un periodo di tempo tale da non compromettere il
processo fotosintetico (Wieser e Matyssek, 2007). Il secondo meccanismo di difesa consiste
nell’attivazione di sistemi di detossificazione per le ROS (Castagna e Ranieri, 2009), così da
confinare il danno ossidativo.
Figura 6 - Rappresentazione schematica, riferita all’ozono, delle componenti della resistenza di un organismo a un fattore di
stress, in accordo con la teoria di Levitt (Lorenzini e Nali, 2005).
1.2.4. Ozono e morte cellulare programmata
L’esposizione acuta a concentrazioni relativamente alte di O3, che avviene durante la stagione
della crescita vegetativa della pianta in molte zone del mondo, comporta l’induzione di morte
cellulare e formazione di lesioni visibili su individui sensibili. La morte cellulare indotta da O3
è un processo attivo che condivide somiglianze con la morte cellulare programmata (PCD) in
piante con reazioni di ipersensibilità (HR) ai patogeni (Baier et al., 2005; Kangasjärvi et al.,
2005). L’HR, che spesso precede la resistenza sistemica acquisita (SAR), è caratterizzata
dall’induzione di (i) vari geni di protezione cellulare, (ii) morte cellulare nell’ospite e (iii)
restrizione dello sviluppo e diffusione del patogeno. La PCD è un processo fisiologico che
individua ed elimina selettivamente cellule non più necessarie o malfunzionanti perché vecchie,
infettate, danneggiate o dannose e influenza la risposta a vari stimoli biotici e abiotici (Rao et
al., 2000; Pellegrini et al., 2013; Vainonen e Kangasjärvi, 2015). Kangasjärvi et al. (2005) ha
analizzato il meccanismo di sensibilità all’O3, definibile nelle seguenti componenti:
1. regolazione del flusso di O3 nelle foglie;

16
2. degradazione di O3 a ROS (normalmente prodotto dal metabolismo cellulare della
pianta) e detossificazione [da ascorbato (AsA) nella maggior parte dei casi]
nell’apoplasto;
3. sensing/percezione dell’O3;
4. produzione attiva di ROS indotta da O3;
5. attivazione precoce di proteine chinasi attivate da mitogeni (MAPK) da O3;
6. regolazione ormonale indotta da formazione di lesioni da O3.
Un elemento in comune osservato tra O3 e altri stress biotici e abiotici è la produzione attiva di
ROS nell’apoplasto; quindi, l’inquinante può essere utilizzato come strumento non invasivo per
lo studio delle vie di segnalazione apoplastiche (Pellegrini et al., 2013; Vainonen e Kangasjärvi,
2015).
1.2.5. Interazioni tra le vie di segnalazione che influenzano la PCD
La capacità della pianta di adattarsi all’ambiente dipende dalla velocità e dall’efficienza con le
quali riesce a riconoscere uno stress, generare molecole-segnale e dare inizio ad una risposta
che induca il suo metabolismo a compensare i cambiamenti esterni (Rao et al., 2000). Dopo il
riconoscimento del fattore di stress biotico o abiotico, vengono attivate numerose segnalazioni
a catena, al fine di innescare le vie di difesa.
La sequenza di eventi che si verifica nelle piante esposte a O3 ha inizio con la penetrazione
dell’agente ossidante, tramite le aperture stomatiche, e sua degradazione a ROS nell’apoplasto.
Il loro accumulo è influenzato dall’azione di antiossidanti, quali AsA. Questa può essere
considerata come la prima linea di difesa (Vainonen e Kangasjärvi, 2015). Il ruolo di
antiossidante dell’AsA è strettamente dipendente dalla capacità delle cellule di mantenerlo allo
stato ridotto. Anche il glutatione (GSH) è un forte scavenger del ROS, ma solo nella sua forma
ridotta. Altri composti a basso peso molecolare (es. i fenilpropanoidi), presenti nell’apoplasto,
possono avere un ruolo nella difesa cellulare. Quando la capacità antiossidante dell’apoplasto
è superata, le ROS accumulate negli spazi extracellulari si propagano all’interno delle cellule
(Kangasjärvi et al., 2005).
La generazione di ROS è normalmente deleteria, ma molti studi la riconoscono come una
componente importante nelle vie di trasduzione del segnale che influenzano le risposte di difesa
della pianta e la PCD per molti stimoli, inclusi i patogeni (Lamb e Dixon 1997; Jabs 1999). La
produzione di ROS secondarie avviene in seguito ad un processo chiamato burst ossidativo,
che utilizza l’O3 accumulato nell’apoplasto per liberare radicali come H2O2 e O2•-, con un

17
procedimento simile a quello che si osserva durante la morte cellulare da HR (Baier et al. 2005).
Questo evento innesca una serie di segnali a cascata che lavorano simultaneamente,
alternativamente o in serie (Vaultier e Jolivet, 2015) grazie all’amplificazione dell’onda di ROS
in differenti comparti subcellulari (Figura 7). La generazione di ROS in risposta all’O3 è
bifasica: un picco anticipato distinguibile entro un’ora dall’esposizione di O3 e uno più ampio
sviluppato tra le nove e le dodici ore dopo l’inizio dell’esposizione. Una produzione simile e
associabile alla risposta a patogeni (Jones e Dangl, 2006).
Figura 7 - Via delle ROS e degli ormoni indotta da ozono (Wrzaczek et al., 2010). Abbreviazioni: ABA, acido abscissico; CRK,
cysteine-rich receptor-like kinases; DND1, defense no death 1; ET, etilene; H2O2, perossido di idrogeno; ICS1, isocorismato
sintasi 1; NPR1, non-expressor of pathogenesis-related genes 1; O2•-, anione superossido; O3, ozono; PAMP, pathogen-
associated molecular patterns; RBOH, plant respiratory burst oxidase homolog; ROS, specie reattive dell’ossigeno; SA, acido
salicilico; SOD, superossido dismutasi; JA, acido giasmonico.
Le cellule di guardia dei cloroplasti sono le prime ad avvertire l’accumulo di ROS,
probabilmente grazie ad un segnale a loro trasmesso. Una proteina G può essere il mediatore
della produzione bifasica di ROS in risposta a O3 (Joo et al., 2005; Booker et al., 2012). Le
ROS nell’apoplasto, formate dalla degradazione di O3 o da burst ossidativo, attivano molti
meccanismi simultanei, generando una complessa rete di segnali per la risposta all’O3. Ciò

18
include (i) cambiamenti nella omeostasi redox cellulare, (ii) percezione da proteine
apoplastiche, (iii) danni ossidativi di membrane e (iv) trasporto di H2O2 apoplastico per mezzo
di acquaporine attraverso la membrana plasmatica. Per questo, solo le ROS possono non essere
sufficienti a indurre PCD nella pianta, ma possono agire sinergicamente con altri componenti
per innescare la morte cellulare (Rao et al., 2000). Da qui, l’esposizione a O3 attiva varie vie di
segnalazione intercellulari che promuovono cambiamenti nelle risposte omeostatiche ormonali,
biochimiche e trascrizionali.
Le MPAK svolgono un ruolo fondamentale nella morte cellulare HR-like, suggerendo un loro
possibile coinvolgimento nella PCD indotta da ROS. I substrati MPK3/MPK6 sono collegati
alla biosintesi dell’ET (Meng et al., 2013). Inoltre, queste proteine possono essere attivate per
promuovere ET o regolare i suoi segnali e la senescenza fogliare (Liu et al., 2008; Zhou et al.,
2009; Yoo et al., 2008).
Un altro evento innescato dall’esposizione di O3 è l’incremento della concentrazione di ioni
calcio (Ca2+) nelle cellule di guardia (Evans et al., 2005) che agiscono come messaggeri
secondari, regolatori di una rete di segnali coinvolti nella risposta a differenti gamme di stimoli
allo sviluppo e ambientali (Tamaoki, 2008; Kudla et al., 2010; Short et al., 2012).
Gli ormoni regolano ogni aspetto dell’accrescimento e dello sviluppo della pianta, così come la
risposta a stress biotici e abiotici (incluso O3). Tra i regolatori di crescita vi sono i fitormoni
classici come ET, acido abscisico (ABA), auxine (IAA), citochinine (CK) e gibberelline (GA),
assieme ad altre molecole segnale, come acido salicilico (SA), acido giasmonico (JA), prolina
(Pro) e brassinosteroidi (BR). Inoltre, è confermata la partecipazione di molti fitormoni nella
risposta a stress acuto di O3. Recenti studi hanno dimostrato la presenza di cross-talk tra queste
vie (Tuominen et al., 2004; Gomi et al., 2005; Yaeno et al., 2006; Ludwikow et al., 2009;
Tamaoki, 2008). Una risposta anticipata all’O3 è la biosintesi dell’ET, seguita dalla produzione
di SA, JA e ABA. ET e SA promuovono una maggiore produzione di ROS e PCD, innescando
un ciclo (Figura 8). JA contrasta la produzione di ROS a seguito di (i) biosintesi di ET e (ii)
morte cellulare. L’ABA è importante specialmente come regolatore della chiusura stomatica e
dell’entrata di O3.

19
Figura 8 - Ciclo di morte cellulare ossidativa indotta da ozono (Kangasjärvi et al., 2005). Abbreviazioni: ABA, acido
abscissico; ET, etilene; SA, acido salicilico; JA, acido giasmonico.
1.3. Ormoni e molecole segnale in risposta allo ozono
1.3.1. Etilene
ET (C2H4) è essenziale nel (i) modulare la risposta delle piante a stress di varia natura (biotici
e abiotici) e (ii) regolare i processi di crescita e di senescenza (Lin et al., 2009). In presenza di
stress (carenza ed eccesso di acqua, danno meccanico, shock osmotico, O3 e attacchi da parte
di patogeni/insetti) il rapido aumento della produzione di ET (Wang et al., 2002; van Loon et
al., 2006) può favorire, di per sé o interagendo con altri ormoni, l’attivazione di vari meccanismi
cellulari di difesa (Overmyer et al., 2008; Wang et al., 2002; Trivellini et al., 2011).
ET è sintetizzato attraverso l’attività di due enzimi: 1-aminociclopropano-1-acido carbossilico
(ACC) sintasi (ACS) e ACC ossidasi (ACO), (Figura 9). ACS è coinvolta nella conversione
della S-adenosil-L-metionina (SAM) in ACC, mentre ACO catalizza la conversione di ACC in
ET con rilascio di CO2 e cianide (Wang et al., 2002).

20
Figura 9 - Via biosintetica dell’etilene in piante esposte ad ozono (Iqbal et al., 2014). Abbreviazioni: ACC, acido 1-
aminociclopropano-1-acido carbossilico; MAPK, proteina chinasi attivata da mitogeni; ROS, specie reattive dell’ossigeno;
SAM, S-adenosil-L-metionina.
L’induzione di ET è da considerarsi una delle più rapide risposte biochimiche e molecolari
osservate in piante sottoposte ad O3, poiché avviene nelle prime ore di esposizione (Nakajima
et al., 2001; Moeder et al., 2002; Pellegrini et al., 2013). La biosintesi è strettamente regolata
da processi di fosforilazione e degradazione proteica attraverso il controllo dell’attività
enzimatica delle ACS (necessario per l’aumento della produzione di ET).
E’ stato dimostrato che la sensibilità all’O3 in termini di sintomi fogliari risulta più marcata in
mutanti di Arabidopsis che producono elevate quantità di ET. Infatti, la precoce e rapida
induzione di ET durante un trattamento acuto con l’inquinante è in grado di stimolare la morte
cellulare interagendo in maniera sinergica ed antagonista con altre molecole segnale (SA e JA)
che regolano la risposta all’O3 (Rao et al., 2002).
1.3.2. Acido abscissico
Recentemente, particolare attenzione è stata rivolta al ruolo dell’ABA nella regolazione dei
meccanismi di risposta della pianta a stress di varia natura (biotici ed abiotici), durante i processi

21
di crescita e sviluppo, così come nella regolazione dell’apertura stomatica. È stato dimostrato
che in presenza di condizioni non ottimali per l’organismo, il contenuto di ABA aumenta nelle
cellule guardia, inducendo la chiusura degli stomi e la conseguente conservazione dello stato
idrico della foglia (Tardieu et al., 2010).
In specie sensibili all’O3, è stato dimostrato che la produzione di ET indotta dall’inquinante può
alterare la risposta degli stomi all’ABA, che tendono a restare molto più aperti (Wilkinson and
Davies, 2010).
In studi sperimentali recentemente condotti, è stato riscontrato che l’accumulo di ROS
nell’apoplasto indotto dalla presenza di stress, potrebbe essere coinvolto nei processi di
chiusura/apertura stomatica (An et al., 2008; Khokon et al., 2011), modulando la regolazione
degli stomi da parte delle cellule guardia e dell’ABA. In particolare, è stato osservato che le
ROS favoriscono l’aumento del contenuto di ABA nella forma libera, diminuendone la
degradazione o il suo rilascio a partire dalle forme coniugate (Song et al., 2014). Mittler e
Blumwald (2015) riportano che l’aumento delle ROS nell’apoplasto ed il conseguente
accumulo di ABA nelle cellule guardia potrebbero creare un loop positivo nel promuovere la
chiusura stomatica, considerando che un elevato contenuto di questo ormone potrebbe
anch’esso indurre una produzione di ROS.
La via di formazione dell’ABA è ben conosciuta, sebbene i meccanismi di signaling/percezione
risultino al momento alquanto ambigui, a causa del numero elevato di componenti
(metaboliti/enzimi) in essi coinvolti (Figura 10).

22
Figura 10 - Vie di formazione proposte per la biosintesi dell’acido abscissico nelle piante sottoposte ad ozono
(http://www.riken.jp). Abbreviazioni; ABA, acido abscissico; PP2C, proteina fosfatasi di tipo 2C; RCAR/PYR, componenti
regolatori dell’ABA/Recettori Pyrabactina resistenti; SnRK2, zuccheri non-fermentativi 1 relativi alla proteina chinasi
sottofamiglia 2.
Ad oggi, sono pochi gli studi relativi agli effetti dell’O3 sul contenuto di ABA, nonostante il
suo ruolo nella regolazione della produzione di ET in risposta all’inquinante. I livelli di ABA
subiscono un marcato aumento in piante di M. officinalis sottoposte a trattamento con O3 (200
ppb, 5 ore), che precede il secondo picco di produzione di ROS (Pellegrini et al., 2013):
Tuttavia, in alcune specie vegetali, l’inquinante non determina alcun effetto sulla
concentrazione endogena di questo ormone (Döring et al., 2014).
1.3.3. Acido salicilico
SA è un composto fenolico (acido o-idrossibenzoico), che può essere considerato come un
regolatore endogeno ormone-like (Iqbal et al., 2014). Tra le molecole segnale, SA ha ricevuto
particolare attenzione in relazione alla sua capacità di regolare differenti processi, coinvolti
nella risposta delle piante a stress di varia natura (biotici ed abiotici), attraverso l’interazione
con altre sostanze aventi funzione ormonale (Horváth et al., 2007). Nelle piante superiori, sono

23
state distinte due vie di formazione di SA: (i) fenilalanina ammonio-liasi (PAL) e (ii)
isocorismato. La prima avviene nel citoplasma e SA è sintetizzato a partire dalla fenilalanina,
attraverso l’attività della PAL, passando per l’acido t-cinnamico e l’acido benzoico (Figura 11).
La via dell’isocorismato (attualmente identificata e caratterizzata esclusivamente in
Arabidopsis; Ogawa et al., 2007) ha luogo nei cloroplasti e SA è prodotto dal corismato,
attraverso l’attività dell’isocorismato sintasi (ICS), passando per l’isocorismato (Wildermuth et
al., 2001). SA è sintetizzato nelle cellule vegetali a partire da differenti forme (biologicamente
inattive): glicosilate e/o metilate (tra cui SA o-β-glucoside e metil salicilato, che sono
rispettivamente le più importanti). Esperimenti su piante transgeniche esposte ad O3
confermano che in presenza dell’inquinante, la sintesi ed il metabolismo di SA risultano alterati
in quanto coinvolti in (i) sviluppo/propagazione dei sintomi e (ii) induzione della morte
cellulare HR-like (Örvar et al. 1997; Overmyer et al. 2005).
Figura 11 - Via proposta per la biosintesi e segnalazione dell’acido salicilico nelle piante esposte a ozono (Jayakannan et
al., 2015). Abbreviazioni; ICS, isocorismato sintasi; NPR1, non espressore di PR-1; PAL, fenilalanina ammonia-liasi; SAG,
acido 2-O-b-D-glucosilbenzoico.
Di conseguenza, è possibile affermare che la formazione di SA indotta dall’O3 agisce come
segnale per (i) il mantenimento dello stato redox delle cellule (Pellegrini et al., 2013), (ii)

24
l’induzione ed il potenziamento delle difese cellulari e (iii) la SAR (Di Baccio et al., 2012;
Pasqualini et al., 2002).
A livello funzionale, è dimostrato che la percezione di SA è in grado di amplificare il segnale
responsabile del processo di morte cellulare, analogamente a quello che si attiva durante
l’interazione pianta-patogeno (Tamaoki, 2008). SA è coinvolto in numerosi processi di sviluppo
in virtù del cross-talk con (i) altri ormoni (ad esempio ET e ABA), (ii) molecole con funzione
ormonale [basti pensare a JA e prolina (Pro)] e (iii) ROS (in particolare H2O2). A tal proposito,
è documentato che tali interazioni antagonistiche e/o sinergiche assumono un ruolo rilevante
nella (i) modulazione delle risposte delle piante durante l’esposizione all’O3 e (ii) regolazione
dei danni fogliari a seguito del trattamento con l’inquinante. Per quanto l’argomento richieda
ulteriori approfondimenti, è noto che ET e SA sono necessari per lo sviluppo/propagazione dei
danni visibili indotti da O3 attraverso l’amplificazione del segnale ossidativo (Kangasjärvi et
al., 2005). Appare, quindi, evidente come SA sia fondamentale per l’attivazione della PCD
attraverso il controllo/potenziamento del burst ossidativo assieme ad ET. L’azione di SA risulta
strettamente dipendente dalla sintesi/percezione di ET, oltre che dalla via di segnalazione
ROS/ET (Rao et al., 2002; Vahala et al., 2003; Pellegrini et al., 2013).
Ad oggi, scarsa attenzione è stata rivolta all’interazione ABA-O3 e per questo motivo i possibili
ruoli/interazioni di SA e ABA nella regolazione dello stress indotto dall’inquinante sono quasi
completamente sconosciuti. In generale, è noto che SA esplichi un’azione antagonista nei
confronti di ABA durante la SAR (Jayakannan et al., 2015), sebbene insieme possono ricoprire
un ruolo sinergico nel processo di chiusura stomatica. Recentemente, Xia et al., (2015) hanno
introdotto il concetto di “self-amplifying feedback loop” allo scopo di chiarire e spiegare la
possibile interazione di SA e H2O2 in presenza di stress. H2O2 può indurre l’accumulo di SA
(catalizzando la conversione di acido benzoico in SA), mentre SA può aumentare i livelli di
H2O2 inibendo l’attività della catalasi o inducendone la formazione attraverso una perossidasi
fenolo-dipendente (Diara et al., 2005).
1.3.4. Acido giasmonico
JA è un ormone considerato come un regolatore endogeno che svolge un ruolo molto importante
in (i) processi di sviluppo e (ii) risposta a vari fattori di stress (di natura biotica ed abiotica;
Okada et al., 2015). Dal punto di vista chimico/biologico, JA è un acido grasso (derivato del
ciclopentanone) sintetizzato mediante l’attività di numerosi enzimi (presenti nei plastidi,

25
perossisomi e citoplasma) attraverso la via di formazione degli octadecanoidi (Figura 12; Dar
et al., 2015).
Figura 12 - Via di formazione dell’acido giasmonico in piante sottoposte ad ozono (Dar et al., 2015). Abbreviazioni: 12-OPDA,
acido 12-ossifitodienoico; AOS/DDE2, allene ossido sintasi/deiscenza ritardata 2; LOX6, lipossigenasi 6; MYC2, OPR3, 12-
ossifitodienoato reduttasi 1; PLD, fosfolipasi D; TaAOC1, Triticum aestivum allene ossido ciclasi 2; TaOPR1, Triticum
aestivum 12-ossifitodienoato reduttasi 1; JA, acido giasmonico; JAR1, giasmonato resistente.
JA ed il suo estere metilico (metil giasmonato, MeJA) sono le molecole segnale derivate
dall’acido linoleico più studiate nel mondo vegetale e che sono nel complesso definite come
ossilipine o giasmonati (Dar et al., 2015). La loro biosintesi risulta essere stimolata
dall’esposizione all’O3 (Rao et al., 2000): è stato, infatti, riportato che reagendo con i lipidi
insaturi della membrana plasmatica, l’inquinante induce l’attivazione di processi di

26
perossidazione e, conseguentemente, la formazione di substrati (come ad esempio l’acido α-
linoleico) per le vie degli octadecanoidi (Mudd, 1997).
Durante il processo di morte cellulare indotto da stress ossidativo, i giasmonati partecipano
attivamente a (i) contenimento della propagazione delle lesioni causate dalla presenza delle
ROS (Devoto e Turner, 2005; Kangasjärvi et al., 2005), (ii) protezione dei tessuti dalla perdita
di vitalità cellulare (Overmyer et al., 2000) e (iii) opposizione agli effetti negativi legati ad ET
e SA (Rao et al., 2002). Alcuni studi confermano che la formazione di JA avviene
principalmente in corrispondenza di ferite/danni meccanici (ad esempio ai margini delle lesioni
fogliari) ed il suo accumulo si osserva esclusivamente nelle piante danneggiate durante
l’esposizione a O3 (Tuominen et al., 2004; Di Baccio et al., 2012; Pellegrini et al., 2013). Anche
a livello genico è stato dimostrato che JA può modulare il processo di PCD (in particolare negli
ultimi stadi), limitando e contenendo lo sviluppo delle lesioni (Castagna et al., 2007; Di Baccio
et al., 2012).
Generalmente, JA svolge un ruolo fondamentale nell’attivazione di meccanismi di difesa contro
gli stress (in particolare di natura biotica, quali patogeni necrotrofi). Inoltre, Rohwer e Erwin
(2008) hanno dimostrato che il trattamento con JA è da considerarsi una parte integrale nei
processi di difesa delle piante contro erbivori e predatori. Nel caso di stress abiotici (come ad
esempio l’O3), Örvar et al. (1997) riportano che il pre-trattamento con JA in tabacco inibisce il
processo di morte cellulare indotto dalla seguente esposizione all’inquinante. Risultati simili
sono stati documentati in Arabidopsis (Rao et al., 2000) e pioppo (Kock et al., 2000). Tuttavia,
è stato osservato che mutanti di Arabidopsis JA-insensivite (jin1) risultano tolleranti a
trattamenti acuti con O3 (250 ppb, 6 h), suggerendo così un duplice ruolo da parte di JA
(Nickstadt et al. 2004). JA può (i) avere la meglio sull’ET in termini di generazione/produzione
delle ROS, contenendo così la diffusione delle lesioni (Santino et al., 2013), o (ii) agire come
regolatore negativo della morte cellulare, promuovendo la morte cellulare HR-like e la
senescenza.
Generalmente JA agisce come antagonista nei confronti di ET e SA (Kangasjärvi et al., 2005).
E’ noto che ET stimola la diffusione della morte cellulare mentre, al contrario, JA la inibisce
(Kazan, 2015). A tal proposito, Tuominen et al. (2004) documentano un precoce e rapido
accumulo di ET durante la formazione delle lesioni fogliari in piante di A. thaliana sottoposte
a trattamento con O3, che tende così a contrastare le funzioni protettive del JA durante la fase
di propagazione. Pellegrini et al. (2013) riportano che in piante di M. officinalis esposte
all’inquinante, la produzione di ET è indotta nella prima ora di trattamento, mentre l’accumulo

27
di JA nelle 3 ore successive al termine della fumigazione. Tale risultato suggerisce che JA
svolge un’azione antagonista nei confronti di ET in termini di contenimento delle lesioni indotte
dall’O3. Analoghe conclusioni sono state riportate in pomodoro (Di Baccio et al., 2012),
Arabidopsis (Overmyer et al., 2000) e betulla (Vahala et al., 2003).
Le interazioni antagoniste tra JA e SA sono state oggetto di numerosi studi condotti su piante
esposte ad O3. In particolare, la regolazione delle ROS prodotte potrebbe giocare un ruolo
fondamentale nel cross-talk tra questi due ormoni (Xia et al., 2015). Infatti, JA potrebbe
proteggere i tessuti fogliari dal processo di morte cellulare indotto dalle ROS opponendosi agli
effetti di SA ed ET (come riportato in precedenza). Inoltre, le relazioni tra JA e SA possono
essere regolate dalle concentrazioni relative di ciascuna molecola: basse concentrazioni di SA
sono in grado di agire sinergicamente con JA nell’induzione di meccanismi di difesa (in accordo
con quanto osservato da Koch et al. 2000 e Pellegrini et al. 2013). Di conseguenza, è possibile
affermare che JA ricopre un ruolo nella modulazione dei processi di diffusione delle lesioni e
che esistono importanti similitudini tra le risposte pianta-patogeno e pianta-O3.
1.3.5. Prolina
Pro è un aminoacido che svolge differenti ruoli nelle piante superiori (Szabados e Savouré,
2010); è sintetizzato nei mitocondri, mediante l’azione sequenziale di differenti enzimi (Pro
deidrogenasi e ossidasi (Iqbal et al., 2014). Per lungo tempo, Pro è stata considerata un’osmolita
inerte in grado di proteggere le strutture subcellulari e le macromolecole in presenza di stress
osmotico. Tuttavia, recentemente alcuni studi hanno confermato il suo possibile
coinvolgimento nei meccanismi di adattamento, recovery e signaling. Infatti, Pro può (i) agire
come enzima in grado di proteggere l’integrità delle proteine e di intensificare l’attività di altri
composti simili (Filippou et al., 2014), (ii) agire come scavenger dei radicali ossidrili e
quencher dell’ossigeno singoletto prevenendo il danno ossidativo causato dalle ROS (Rejeb et
al., 2014) e (iii) bilanciare lo stato redox cellulare, stabilizzando l’omeostasi delle cellule e delle
strutture subcellulari che forniscono carbonio e azoto utili per la crescita della pianta (Szabados
e Savouré, 2010). Il metabolismo di Pro può inoltre influenzare la PCD, agendo come molecola
segnale per (i) modulare le funzioni micondriali, (ii) influenzare la proliferazione cellulare e
(iii) innescare l’espressione di specifici geni (Iqbal et al., 2014). Questi ruoli sono stati
ampiamenti studiati e dimostrati in piante sottoposte a stress idrico, salino e da metalli pesanti
a livello funzionale e genico. Tuttavia, il ruolo di Pro in risposta all’O3 necessita di ulteriori
studi.

28
1.4. Quercus ilex
L’areale del leccio comprende buona parte del Mediterraneo. La specie è maggiormente diffusa
in Algeria, Marocco, in tutta la penisola iberica, nella Francia mediterranea e in Italia. Nel
nostro Paese, lo troviamo in boschi misti e, più raramente, in popolamenti puri. E’ uno dei
rappresentanti più tipici e importanti dei querceti perenni mediterranei. Si tratta di un albero
sempreverde, a crescita lenta, longevo, che può arrivare a 25 m di altezza. Se cresce in ambienti
rupestri può assumere un aspetto cespuglioso. La corteccia è liscia e grigia nella fase giovanile,
scurisce e indurisce con il passare del tempo, fino a diventare nerastra e screpolata. I giovani
rami sono pubescenti e grigi, ma dopo poco tempo diventano glabri e grigio-verdastri. Le foglie
sono semplici, ovali, lanceolate, coriacee, con la pagina superiore verde scuro, lucida e quella
inferiore grigiastra e tomentosa. E’ eterofilla, ovvero sono presenti due tipi di germogliazione
nelle plantule. Questa specie preferisce il pieno sole, ma sopporta anche la mezz’ombra. Nel
nostro territorio si trova anche fino a 1000 m. Ha un apparato radicale inizialmente fittonante;
in seguito, sviluppa radici laterali, che gli permettono una buona resistenza alla siccità,
riuscendo a intercettare l’H2O in profondità. Non tollera i terreni molto argillosi e teme il
ristagno idrico, predilige terreni alcalini, ma cresce bene anche su substrati acidi. Viene
ampiamente utilizzata per arricchire giardini boscosi o per viali alberati, poiché tollera molto
bene le potature, tanto da essere una pianta adatta alle forme obbligate e nelle regioni costiere
viene impiegata come barriera frangivento.
2. SCOPO DELLA TESI
L’andamento generale della concentrazione di O3 troposferico è da tempo in continua e
progressiva crescita. Le piante sono i recettori più sensibili alla sua azione tossica e possono
manifestare sintomi (clorosi e necrosi fogliari) e/o fenomeni subliminali responsabili di
riduzione della crescita e della produzione quali-quantitativa. Le somiglianze riscontrate tra i
meccanismi attivati dall’inquinante e la HR osservata nell’interazione incompatibile pianta-
patogeno hanno suggerito che l’O3 possa comportarsi alla stregua di uno stress biotico. Infatti,
gli effetti di questo contaminante si esplicano attraverso una cascata di eventi regolati
dall’induzione di ormoni e molecole segnale, che vanno a mediare la stimolazione di risposte a
livello genico e funzionale.
Le specie quercine sono state oggetto di numerosi studi per quanto riguarda la loro interazione
col cambiamento climatico, in quanto sono molto diffuse nell’areale mediterraneo e sono molto
utilizzate in ambito urbano. Inoltre, le piante appartenenti a questo genere sono note per le alte

29
emissioni di COV e se da un lato sequestrano gli inquinanti, da un altro, sono esse stesse
emettitrici di precursori volatili per la formazione di smog fotochimico.
Attualmente molte ricerche hanno affrontano la tematica dello stress da O3 su Q. ilex con
trattamento cronico, mentre poche analisi sono state compiute per quello puntiforme. Per questo
motivo, lo scopo del presente lavoro è stato quello di verificare il grado di sensibilità/resistenza
di giovani piante (3 anni di età) di Q. ilex allo stress ossidativo, come quello imposto da O3 (200
ppb, 5 ore), sulla base dell’ipotesi formulata da Kangasjärvi et al. (2005), riproducendo in
ambiente controllato scenari previsti dai modellisti del global change per il 2050. L’indagine
ha preso in considerazione (i) la produzione di H2O2 e O2•- e (ii) la risposta di ET, ABA, SA,
JA e Pro nei seguenti tempi di analisi (1, 2, 5, 8, 24 ore dall’inizio del trattamento).
Il lavoro è svolto nell’ambito del progetto PRIN 2010-2011 TreeCity, che vede la
partecipazione di otto unità nazionali tra cui l’Università di Pisa in veste di coordinatore dei
lavori. E’ supportato da dodici istituzioni di ricerca a titolo gratuito (9 delle quali straniere) ed
è caratterizzato dalla forte connotazione interdisciplinare. Si articola in quatto sottoprogetti
operativi (WP), ciascuno delle quali, a sua volta, organizzato in azioni. Questo lavoro si pone
all’interno del primo gruppo (WP1: SISTEMI VEGETALI URBANI, UN LABORATORIO A
CIELO APERTO PER VALUTARE LA QUALITÀ URBANA E LE CONDIZIONI DI
STRESS) che si occupa del monitoraggio delle funzioni ecosistemiche in aree urbane ed extra-
urbane campione, misure di parametri ecofisiologici, indagini istopatologiche, studi molecolari
e tecniche di remote sensing, per tracciare un quadro relativo alle capacità dei sistemi vegetali
urbani di adattarsi alle attuali condizioni ambientali. Particolare attenzione è dedicata agli studi
sui componenti del sistema di difesa antiossidante (nei confronti delle ROS prodotte a seguito
dell’induzione dello stato di stress), specialmente quelli relativi al metabolismo secondario,
l’induzione di molecole segnale (ET, ABA, SA, JA) e signaling ossidativo (profilo di ROS e
NO). Il progetto TreeCity si pone obiettivi di natura conoscitiva, ben coniugabili con aspetti di
ordine pratico-applicativo nel tema degli alberi e la loro capacità di migliorare le condizioni
ambientali delle città, con particolare riferimento ai fenomeni di cambiamento globale previsti.
Tra i traguardi fissati, opera per giungere alla definizione di criteri e strategie per la scelta di
specie ornamentali legnose da impiegare nelle aree urbane del futuro, con l’acquisizione di
importanti conoscenze sui meccanismi intimi di risposta degli alberi allo stress, con particolare
riferimento ai processi ossidativi, specialmente nel campo del metabolismo secondario e sui
fenomeni di trasduzione del segnale stress-dipendente. A fini dello studio, è importante che i
risultati ottenuti siano utili per stabilire le migliori strategie di gestione della vegetazione per

30
migliorarne le potenzialità di rimozione degli inquinanti e di mitigazione del cambiamento
climatico.

31
3. MATERIALI E METODI
3.1. Allevamento del materiale vegetale ed esposizione a stress idrico e/o ad
ozono
Giovani individui di leccio (3 anni di età) sono stati sottoposti per brevi periodi a condizioni di
crescita sfavorevoli, riprodotte in ambiente controllato, consistenti in una temporanea
insufficiente disponibilità idrica del terreno, in concomitanza, o meno, con esposizioni
puntiformi ad O3. L’indagine è stata condotta su semenzali di tre anni di età, forniti dal vivaio
“La Torraccia” di UmbriaFlor s.r.l. (Gubbio, PG), presso le strutture del Dipartimento di
Scienze Agrarie, Alimentari e Agro-ambientali (DiSAAA-a) dell’Università di Pisa.
Sino all’avvio della sperimentazione, il materiale è stato mantenuto all’aperto, in condizioni
idriche e nutritive ottimali, in vasi in PVC da 6,5 l, contenenti (in un rapporto volumetrico 3,5:1)
terriccio ED 63 della Einheitserden e sabbia di fiume vagliata (da 0,6 mm e 2,67 g cm-3 di
peso specifico). Dopo 14 giorni, piante omogenee per taglia, stadio di sviluppo e in buono stato
vegetativo sono state selezionate, numerate e trasferite in idonee camere di crescita in perspex
(1,50 × 1,50 × 1,00 m), ventilate in continuo con aria filtrata attraverso carbone attivo, in
condizioni climatiche naturali. La minima e la massima temperatura media del periodo era di
15,8 e 28,0 °C, l’umidità relativa (U.R.) minima e massima media di 32,6 e 83,6% e la
radiazione solare diurna minima e massima media di 9,7 e 30,1 MJ m-2.
Dopo un periodo di acclimatamento di due settimane, il lotto è stato suddiviso in due gruppi
omogenei, di identica numerosità, di cui uno ha continuato a beneficiare di adeguato
approvvigionamento idrico; l’altro è stato sottoposto ad un regime irriguo scarso, ricevendo un
volume di adacquamento giornaliero ridotto al 30% del corrispettivo evapotraspirato.
Quest’ultimo era stato preventivamente calcolato, nelle medesime condizioni di crescita, sulla
base dei volumi d’acqua raggiunti alla capacità di campo e al punto di appassimento.
Dopo due settimane alle condizioni sopra esposte, le piante (26 esemplari) sono state trasferite
in una camera di crescita, impostata a 23±1 °C, 85±5% U.R., 530 µmol di fotoni m-2 s-1 di
radiazione fotosinteticamente attiva (PAR: l = 400-700 nm) e fotoperiodo di 14 ore, e
acclimatate per tre giorni alle medesime disponibilità idriche. All’interno di questa si trova un
box in perspex (1,30 × 0,75 × 1,00 m), continuamente ventilato con aria filtrata, in cui è
possibile l’arricchimento controllato con O3. L’inquinante è generato a partire da ossigeno puro
(O2), attraverso la scarica elettrica prodotta da un generatore (Fisher 500), la cui energia scinde

32
la molecola biatomica in atomi i quali, collidendo con un’altra molecola di O2, formano O3. E’
possibile impostare la concentrazione di O3, regolando l’intensità della scarica elettrica e/o la
portata d’ingresso dell’O2. La miscela di aria-O3, in corrispondenza della parte epigea delle
piante, è campionata in continuo attraverso un tubo in teflon e “letta” da un rilevatore
fotometrico a raggi ultravioletti (UV) (Monitor Labs, mod. 8810), controllato da uno specifico
software.
Alla fine del periodo di acclimatamento, i due lotti di piante (13 esemplari per parte, diversi per
i regimi idrici, di cui sopra) sono stati ulteriormente suddivisi in due gruppi da esporre o no a
O3. Quindi, le tesi sono quattro, come di seguito specificato: 1) regime idrico giornaliero pari
al 30% dell’evapotraspirato; 2) esposizione all’inquinante (200 ppb per 5 ore consecutive); 3)
stress combinato (O3 + carenza idrica); 4) controllo mantenuto in aria filtrata e a regime idrico
giornaliero pari al 100% dell’evapotraspirato.
3.2. Liofilizzazione del materiale vegetale
Il materiale composto da foglie raccolte, divise in gruppi, pesate (5 g per replica) e conservate
a -20 °C, è stato sottoposto a liofilizzazione per una migliore conservazione. Questo processo
permette l’eliminazione dell’acqua all’interno di una sostanza organica, con trascurabile
deterioramento della struttura e dei componenti della sostanza stessa. I campioni sono stati posti
in provette aperte e inseriti nella camera (Standard Chamber, 215 × 300 mm) del liofilizzatore
(Telstar LyoQuest -55), precedentemente raffreddata a -15 °C. Il processo è stato protratto per
tre giorni, al termine dei quali il materiale è stato macinato.
3.3. Vitalità cellulare e localizzazione in situ di perossido di idrogeno
Dopo 1, 2, 5, 8 e 24 ore dall’inizio della fumigazione, foglie asintomatiche sono state
campionate allo scopo di saggiare in vivo la vitalità cellulare e di localizzare eventuali siti di
accumulo di H2O2, seguendo il protocollo di Iriti et al. (2003).
Con l’ausilio di un bisturi, sono state ricavate porzioni di circa 1 cm2 di area fogliare, avendo
avuto preventivamente cura di eliminare la nervatura principale e i margini. Per la vitalità
cellulare, le sezioni sono state immerse per un minuto in una miscela colorante bollente
composta da una soluzione di Evans blu in H2O distillata (20 mg ml-1), fenolo, acido lattico e
glicerolo, nel rapporto volumetrico 1:1:1:1 (preparata immediatamente prima dell’uso). Dopo
un preliminare risciacquo (per tre volte) con H2O distillata e con una soluzione diluita di cloralio

33
idrato (2,5 mg ml-1), i frammenti sono stati lasciati a decolorare per tutta la notte in una
soluzione di cloralio idrato più concentrata (2,5 g ml-1 in H2O distillata).
Per visualizzare in vivo i siti di accumulo di H2O2, altre porzioni di lamina fogliare sono state
sottoposte a colorazione con 3,3-diaminobenzidina (DAB). Immerse per 8 ore in una soluzione
acquosa di DAB-HCl (1 mg ml-1), portata a pH 5,6 con idrossido di sodio (NaOH), esse sono
state risciacquate in alcool etilico a 70 °C e chiarificate con una soluzione decolorante di
cloralio idrato (2,5 g ml-1). Le sezioni sottoposte alle colorazioni sopra descritte sono state
montate su vetrino, osservate e fotografate al microscopio ottico (Ingrandimento 10x,
DM4000B, Leica, dotato di fotocamera Olympus SH-60).
3.4. Determinazione della produzione di radicali liberi
Il contenuto in H2O2 è stato determinato mediante l’uso di una sonda fluorimetrica 10-acetil-
3,7-diidrossifenoxazina, particolarmente sensibile a basse concentrazioni della molecola
(Mohanty et al., 1997), con cui essa reagisce (in presenza di perossidasi), con un rapporto
stechiometrico 1:1, formando resofurina, un prodotto di ossidazione colorato e fluorescente
(Zhao et al., 2012). Venticinque mg di materiale liofilizzato e macinato sono stati estratti in
1,250 ml di tampone potassio fosfato (K/P) 20 mM (pH 6,5). Dopo centrifugazione a 13.200 g
a 4 °C per 20 minuti, il surnatante è stato utilizzato per misurare il contenuto in H2O2, secondo
il protocollo Amplex Red Hydrogen peroxide/peroxidase assay kit (Molecular Probes,
Invitrogen). I campioni sono stati incubati e mantenuti in continua agitazione per 30 minuti a
25 °C, in assenza di luce. L’emissione di resofurina è stata determinata a 590 nm dopo
eccitazione a 530 nm, mediante uno spettrofluorimetro a piastre (Victor3 1420 Multilabel
Counter, Perkin Elmer). La concentrazione di H2O2, espressa in µmol g-1 di peso secco (PS), è
stata determinata in riferimento ad una curva standard, ottenuta con il medesimo protocollo,
sostituendo l’estratto con soluzioni a concentrazione nota di H2O2 da 0 a 20 µmol.
Il protocollo seguito per la determinazione della concentrazione di O2•- è in accordo con Lei et
al. (2011). Il saggio prevede l’uso del sale sodico 2,3-bis(2-metossi-4-nitro-5-sulfofenil)-2H-
tetrazolo-5-carboxanilide (XTT) che, in presenza di ossidanti quali O2•-, si riduce a formazano
solubile, un composto che può essere quantificato in soluzione con tecniche spettrofotometriche
grazie alla variazione di colore causata dalla reazione di riduzione (Able et al., 1998). Ogni
campione, accuratamente pesato (50 mg), è stato estratto con un tampone di K/P 50 mM (pH
7,5), in rapporto di estrazione 50:500 (peso/volume). Dopo centrifugazione a 12.000 g a 4 °C
per 15 minuti, il surnatante è stato recuperato e posto in ghiaccio per la misurazione del

34
contenuto in O2•-. La soluzione finale (1,5 ml) letta allo spettrofotometro (Jenway 6505)
conteneva: 1435 µl di 50 mM Tris-HCl (pH 7,5), 50 µl di estratto e 0,5 mM XTT. Per il bianco,
è stato sostituito all’estratto il tampone di estrazione. La riduzione di XTT è stata seguita ogni
60 minuti per 3 ore consecutive misurando l’assorbanza a 470 nm. La concentrazione in O2•- è
stata determinata in base al coefficiente di estinzione molare (e) 2.16 × 104 M−1 cm−1 di XTT a
470 nm ed è stata espressa in nmol O2•- min-1 g-1 PS.
3.5. Determinazione del contenuto in fitormoni e molecole-segnale
La quantità di ET è stata valutata secondo il protocollo messo a punto da Mensuali-Sodi et al.
(1992). Il materiale vegetale (3 g) è stato posto in contenitori di vetro da 80 ml aventi un tappo
a vite forato e dotato di setto di caucciù. Questi sono stati chiusi ermeticamente e la produzione
dell’ormone è stata valutata a distanza di 1 ora (tempo necessario ad accumulare all’interno
delle vial una quantità determinabile di etilene) a temperatura ambiente, prelevando un
campione di aria di circa 2,5 ml mediante l’ausilio di una siringa ipodermica. La determinazione
di ET è stata effettuata mediante gas-cromatografo (Hewlett e Packard, mod. 5890A),
utilizzando un detector FID (Flame Ionization Detector) e una colonna metallica (Ø 150 × 0,4
cm impaccata con Hysep T). La sua quantificazione è stata ottenuta con concentrazione nota
(1000 ppm) di uno standard di etilene iniettate all’inizio di ogni analisi. La temperatura della
colonna e del detector erano rispettivamente di 70 e 350 °C. Come gas di trasporto è stato
utilizzato N2 a 30 ml min-1.
Il contenuto in ABA è stato valutato mediante immunoassay (ELISA) indiretto, basato
sull’utilizzo dell’anticorpo monoclonale DBPA1 per S(+)-ABA (Vernieri et al., 1989;
Trivellini et al., 2011). L’ELISA test è stato effettuato secondo il metodo descritto da Walker-
Simmons (1987), con piccole modifiche. L’ABA è stato misurato su materiale vegetale
liofilizzato (100 mg) dopo estrazione in H2O distillata (rapporto H2O:tessuto 10:1,
peso/volume) una notte a 4 °C. Le piastre sono state sensibilizzate con 200 µl per pozzetto di
ABA-4’-BSA, incubate per tutta la notte a 4 °C e, quindi, lavate tre volte con tampone PBS 75
mM (pH 7,0), contenente 1 g l-1 di BSA e 1 ml l-1 di Tween 20, mantenendo la terza soluzione
di lavaggio per 30 minuti a 37 °C. Successivamente, sono stati aggiunti a ciascun pozzetto 100
µl di soluzione standard di ABA o del campione, quindi 100 µl di soluzione DBPA1 (terreno
di coltura cellulare liofilizzato diluito in tampone PBS contenente 10 g l-1 di BSA e 0,5 ml l-1
di Tween 20, ad una concentrazione finale di anticorpo pari a 50 mg ml-1), lasciando avvenire
la competizione per 30 minuti ad una temperatura di 37 °C. Le piastre sono state lavate come
descritto precedentemente; sono stati, poi, aggiunti 200 µl per pozzetto del secondo anticorpo

35
(anti-topo coniugato con fosfatasi alcalina, Sigma, in PBS + 10 g l-1 di BSA, alla diluizione
finale di 1:2.000) e incubato per 30 minuti a 37 °C. Le piastre sono state nuovamente lavate e
vi sono stati aggiunti 200 µl, per pozzetto, di p-Nitrofenile e incubate per 30 minuti a 37 °C. Le
letture di assorbanza sono state effettuate a 415 nm, utilizzando un lettore di micropiastre
(Perkin-Elmer, mod. MDL 680).
I livelli di SA e JA sono stati determinati, previa opportuna estrazione, mediante tecniche di
cromatografia liquida ad alta prestazione (HPLC, Dionex). Venti microlitri (volume di
iniezione) di ciascun estratto sono stati analizzati in fase inversa (RP), tramite un sistema di
pompa quaternaria a bassa pressione P680 A LPG-4, colonna C18 (Acclaim 120), rilevatori a
luce ultravioletta/visibile (UVD 170U UV/VIS) e a fluorescenza (Fluorescence Detector RF-
2000). I dati cromatografici sono stati processati e registrati mediante software (Chromeleon
della Chromatography Management System, versione 6.60-2004).
L’analisi di SA è stata effettuata in accordo con Wang et al. (2013). Il saggio permette di
determinare la presenza del composto fenolico in forma libera e coniugata. A 300 mg di
materiale vegetale liofilizzato è stato aggiunto 1 ml di metanolo per HPLC al 90% in H2O ultra-
pura. Il tutto è stato sonicato per tre volte (ciascun ciclo aveva una durata di 10 minuti) e
successivamente centrifugato a 16.000 g a temperatura ambiente per 20 minuti. Una volta
recuperato il surnatante, sono stati aggiunti 0,5 ml di metanolo puro per HPLC al pellet. Dopo
aver ripetuto i processi di sonicatura e centrifugazione, il surnatante (riunito al precedente) è
stato portato a secco a 40 °C, con l’ausilio di un evaporatore rotante (Laborota 4000). L’estratto
secco è stato disciolto e recuperato in una miscela eterogenea composta da 0,25 ml di una
soluzione acquosa di acido tricloroacetico (TCA) al 5% (peso/volume) e 0,8 ml di solventi
apolari (etilacetato e cicloesano, nel rapporto volumetrico di 1:1). Dopo energica agitazione, la
miscela è stata centrifugata al fine di favorire una netta separazione di fase nelle due
componenti, quella idrofoba, più leggera, e l’altra idrofila, più pesante. La fase sovrastante,
contenente SA libero, è stata recuperata e portata a secco, ripresa con 0,4 ml del tampone sodio
acetato 0,2 M (pH 5,5) e analizzata direttamente all’HPLC, previa filtrazione (Iso-disc Filters
PTFE-13-2, Ø 0,2 µm). La fase sottostante (contenente SA coniugato) è stata acidificata con
0,3 ml di acido cloridrico (HCl) 8M e riscaldata a bagnomaria a 80 °C per 60 minuti, al fine di
favorire l’idrolisi dei legami glicosidici e ottenere l’ormone in forma libera. La soluzione
(contenente, dunque, SA coniugato) è stata sottoposta alla stessa procedura estrattiva di cui
sopra, con la medesima miscela eterogenea di TCA al 5% ed etilacetato e cicloesano (1:1). La
fase apolare sovrastante è stata recuperata, portata a secco, ripresa anch’essa con 0,4 ml del

36
tampone sodio acetato, filtrata e iniettata all’HPLC. Il composto è stato eluito isocraticamente,
usando come fase mobile il tampone sodio acetato 0,2 M (pH 5,5) ad un flusso di 0,8 ml min-1,
per 20 minuti e rilevato in fluorescenza operando a 305 e 407nm, rispettivamente lunghezze
d’onda di eccitazione ed emissione. L’analita è stato identificato mediante l’uso di uno standard
interno e quantificato in riferimento alla retta di calibrazione ottenuta iniettando soluzioni del
medesimo standard a concentrazioni crescenti (0,375-3 µg ml-1).
L’analisi di JA è stata effettuata in accordo con Kramell et al. (1999). Aliquote di campione
liofilizzato e macinato (150 mg) sono state sottoposte a estrazione per 12 ore al buio in 0,75 ml
di etilacetato a 4 °C, previa sonicazione in ghiaccio (3 cicli di 10 minuti ciascuno). Dopo
l’aggiunta di 0,75 ml di una soluzione allo 0,2% (v/v) di acido acetico in H2O ultra pura e
vigorosa agitazione, i campioni sono stati centrifugati a 16.000 g a temperatura ambiente per
20 minuti, al fine di favorire la separazione di fase. Il surnatante filtrato è stato direttamente
iniettato all’HPLC. Il composto è stato eluito isocraticamente, ad un flusso di 1 ml min-1, per
20 minuti, usando come fase mobile due eluenti al 50% ciascuno: la soluzione allo 0,2% (v/v)
di acido acetico in H2O ultra pura e metanolo puro (MeOH). I picchi sono stati acquisiti a 210
nm. L’analita è stato identificato mediante l’uso di uno standard interno e quantificato in
riferimento alla retta di calibrazione ottenuta iniettando soluzioni del medesimo standard a
concentrazioni crescenti (0,062-1 mg ml-1).
Il contenuto di Pro è stato determinato secondo il metodo di Bates et al. (1973). Al materiale
liofilizzato (100 mg) sono stati addizionati 1 ml di acido sulfosalicilico 3%. Il tutto è stato
successivamente sottoposto a sonicatura a 70 °C (10 minuti per tre volte). Questo è stato
centrifugato a 16.000 g per 20 minuti a 20 °C. A 0,4 ml di surnatante sono stati aggiunti 0,4 ml
di acido acetico glaciale e 0,4 ml di reagente, ottenuto sciogliendo 1,25 g di ninidrina in 30 ml
di acido acetico glaciale e 20 ml di acido fosforico 6 M. Successivamente, è stato posto a
bagnomaria per 1 ora a 100 °C e subito dopo raffreddato in ghiaccio. Alla miscela sono stati
aggiunti 0,8 ml di toluene (ottenendo una separazione di fase) ed è stata agitata vigorosamente
per 20 secondi. Sono state eseguite le letture allo spettrofotometro a 520 nm su fase organica
contenente toluene. Dai valori di assorbanza ottenuti è stata determinata la concentrazione di
Pro, utilizzando una curva di taratura calcolata usando concentrazioni note di Pro in acido
sulfosalicilico 3% (0-0,015 mg ml-1).

37
3.6. Trattamento statistico dei dati
L’esperimento è stato ripetuto tre volte, utilizzando ogni volta un minimo di tre replicazioni per
tesi. I dati sono stati archiviati mediante il programma Microsoft® Office Excel 2013. Per
l’elaborazione statistica è stato utilizzato il metodo di analisi della varianza (ANOVA) a due
vie, considerando i fattori “trattamento” e “tempo”. Dove l’interazione “trattamento x tempo”
è risultata significativa, le differenze statistiche tra le medie sono state determinate tramite il
test della differenza minima statistica (LSD) al livello di significatività (P ≤ 0,05). Sono stati
utilizzati i software NCSS 2007 per la statistica e SigmaPlot 12 per la rappresentazione grafica.
4. RISULTATI E DISCUSSIONE
4.1. Vitalità cellulare e localizzazione in situ di perossido di idrogeno
In accordo con quanto riportato da Calatayud et al. (2011), in uno studio condotto su giovani
piante di Q. ilex sottoposte a trattamento cronico con O3 (aria non filtrata + 30 ppb, 8 ore d-1
per 7 giorni consecutivi), anche in conseguenza di un episodio puntiforme (come nel nostro
studio) non sono stati riscontrati sintomi fogliari durante e al termine del periodo di esposizione.
Figura 13 - Tessuto fogliare di Quercus ilex esposto a ozono (200 ppb, 5 ore): controllo (A); cellule morte del mesofillo,
colorate con Evans blu dopo 1, 5 e 24 ore dall’inizio del trattamento (B-D). Barra = 50 µm.
A
D C
B
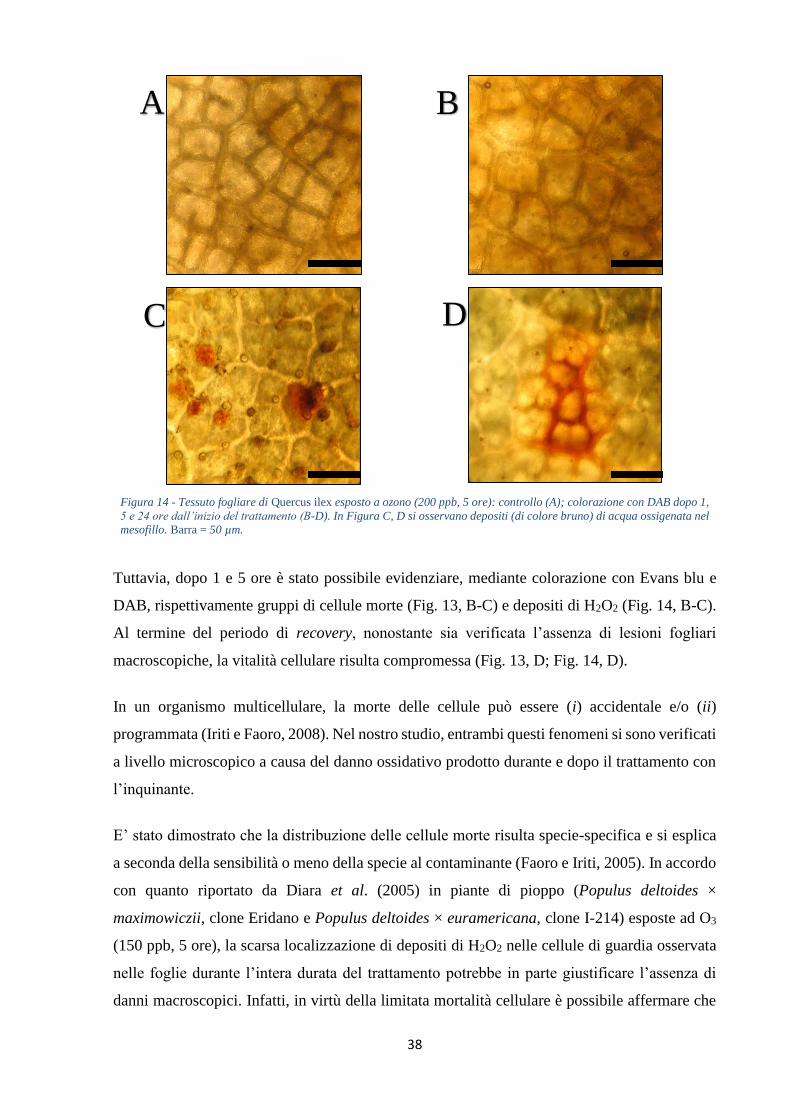
38
Figura 14 - Tessuto fogliare di Quercus ilex esposto a ozono (200 ppb, 5 ore): controllo (A); colorazione con DAB dopo 1,
5 e 24 ore dall’inizio del trattamento (B-D). In Figura C, D si osservano depositi (di colore bruno) di acqua ossigenata nel
mesofillo. Barra = 50 µm.
Tuttavia, dopo 1 e 5 ore è stato possibile evidenziare, mediante colorazione con Evans blu e
DAB, rispettivamente gruppi di cellule morte (Fig. 13, B-C) e depositi di H2O2 (Fig. 14, B-C).
Al termine del periodo di recovery, nonostante sia verificata l’assenza di lesioni fogliari
macroscopiche, la vitalità cellulare risulta compromessa (Fig. 13, D; Fig. 14, D).
In un organismo multicellulare, la morte delle cellule può essere (i) accidentale e/o (ii)
programmata (Iriti e Faoro, 2008). Nel nostro studio, entrambi questi fenomeni si sono verificati
a livello microscopico a causa del danno ossidativo prodotto durante e dopo il trattamento con
l’inquinante.
E’ stato dimostrato che la distribuzione delle cellule morte risulta specie-specifica e si esplica
a seconda della sensibilità o meno della specie al contaminante (Faoro e Iriti, 2005). In accordo
con quanto riportato da Diara et al. (2005) in piante di pioppo (Populus deltoides ×
maximowiczii, clone Eridano e Populus deltoides × euramericana, clone I-214) esposte ad O3
(150 ppb, 5 ore), la scarsa localizzazione di depositi di H2O2 nelle cellule di guardia osservata
nelle foglie durante l’intera durata del trattamento potrebbe in parte giustificare l’assenza di
danni macroscopici. Infatti, in virtù della limitata mortalità cellulare è possibile affermare che
C D
B A
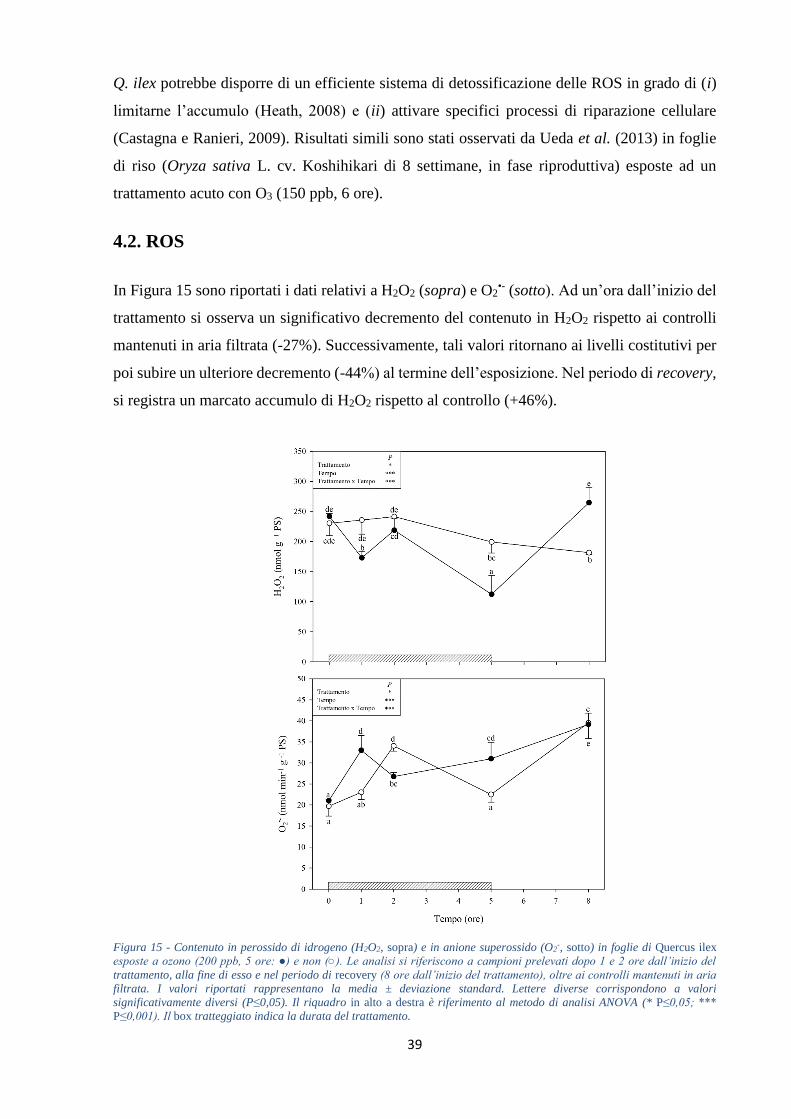
39
Q. ilex potrebbe disporre di un efficiente sistema di detossificazione delle ROS in grado di (i)
limitarne l’accumulo (Heath, 2008) e (ii) attivare specifici processi di riparazione cellulare
(Castagna e Ranieri, 2009). Risultati simili sono stati osservati da Ueda et al. (2013) in foglie
di riso (Oryza sativa L. cv. Koshihikari di 8 settimane, in fase riproduttiva) esposte ad un
trattamento acuto con O3 (150 ppb, 6 ore).
4.2. ROS
In Figura 15 sono riportati i dati relativi a H2O2 (sopra) e O2•- (sotto). Ad un’ora dall’inizio del
trattamento si osserva un significativo decremento del contenuto in H2O2 rispetto ai controlli
mantenuti in aria filtrata (-27%). Successivamente, tali valori ritornano ai livelli costitutivi per
poi subire un ulteriore decremento (-44%) al termine dell’esposizione. Nel periodo di recovery,
si registra un marcato accumulo di H2O2 rispetto al controllo (+46%).
Figura 15 - Contenuto in perossido di idrogeno (H2O2, sopra) e in anione superossido (O2-, sotto) in foglie di Quercus ilex
esposte a ozono (200 ppb, 5 ore: ●) e non (○). Le analisi si riferiscono a campioni prelevati dopo 1 e 2 ore dall’inizio del
trattamento, alla fine di esso e nel periodo di recovery (8 ore dall’inizio del trattamento), oltre ai controlli mantenuti in aria
filtrata. I valori riportati rappresentano la media ± deviazione standard. Lettere diverse corrispondono a valori
significativamente diversi (P≤0,05). Il riquadro in alto a destra è riferimento al metodo di analisi ANOVA (* P≤0,05; ***
P≤0,001). Il box tratteggiato indica la durata del trattamento.

40
Sulla base dei risultati ottenuti possiamo osservare che l’H2O2 non rappresenta una prima linea
di difesa attivata in risposta all’inquinante [come riportato in precedenti studi condotti su piante
di tabacco (Pasqualini et al., 2002), pomodoro (Di Baccio et al., 2012) e melissa (Pellegrini et
al., 2013)], ma piuttosto un marker di danno ossidativo.
Modifiche significative interessano anche l’O2•-: nella prima ora di esposizione, si nota un
marcato incremento del suo contenuto rispetto ai controlli (+43%), seguito da una lieve
riduzione dopo 2 ore (-21%) e da un ulteriore aumento (+38%) al termine della fumigazione.
Nel periodo di recovery, non si osservano differenze statisticamente significative nel materiale
vegetale sottoposto o meno a trattamento. Tale risultato suggerisce che la pianta potrebbe aver
attivato durante la fumigazione meccanismi in grado di regolare la formazione di H2O2
(nonostante l’accumulo di O2•-) tali da contenere il burst ossidativo (Vahala et al., 2003).
Tuttavia, il seguente accumulo in vitro e in vivo di H2O2 denota che tali processi regolatori non
sono stati più in grado di evitare la formazione di lesioni e la conseguente morte cellulare, in
accordo con quanto osservato mediante i saggi istologici (Overmyer et al., 2000).
4.3. Ormoni e molecole segnale
In Figura 16 sono riportati i livelli di ET (sopra) e ABA (sotto). La produzione di ET aumenta
significativamente durante l’intera durata dell’esperimento, per raggiungere un picco al termine
del periodo di recovery (+94%, rispetto ai controlli). Secondo lo studio condotto da Moeder et
al. (2002) su cultivar di pomodoro (Lycopersicon esculentum) esposte ad un trattamento acuto
con O3 (250 ppb, 5 ore), la sintesi di questo ormone risulta necessaria per la
diffusione/progressione delle lesioni. E’ noto, infatti, che la produzione di ET permette
l’accumulo di ROS e la successiva induzione di morte cellulare (Moeder et al., 2002, Vahala et
al., 2003).
L’aumento dell’ormone registrato alla fine del periodo di recovery risulta coincidente con il
picco di H2O2, suggerendo una possibile interazione sinergica tra queste due componenti. E’
importante sottolineare come il possibile ruolo di ET nei principali meccanismi di risposta agli
stress è stato oggetto di pochi studi, in virtù della sua complessità. Infatti, questo ormone svolge
non solo un importante ruolo nell’induzione della PCD, ma instaura anche una complessa rete
di relazioni con varie vie di segnalazione (Vainonen e Kangasjärvi, 2015).

41
Figura 16 - Contenuto in etilene (ET, sopra) e in acido abscissico (ABA, sotto) in foglie di Quercus ilex esposte a ozono (200
ppb, 5 h: ●) e non (○). Le analisi si riferiscono a campioni prelevati dopo 1 e 2 ore dall’inizio del trattamento, alla fine di esso
e nel periodo di recovery (8 ore dall’inizio del trattamento), oltre ai controlli mantenuti in aria filtrata. I valori riportati
rappresentano la media ± deviazione standard. Lettere diverse corrispondono a valori significativamente diversi (P≤0,05). Il
riquadro in basso e in alto a destra è riferimento al metodo di analisi ANOVA (** P≤0,01; *** P≤0,001). Il box tratteggiato
posto in ascissa indica la durata del trattamento.
La risposta di ABA è stata rilevata sin dall’inizio del trattamento: si osserva, infatti, un marcato
aumento della sua concentrazione (+188 e +97%, rispettivamente dopo 1 e 2 ore di
esposizione). Nelle ore successive, i livelli di questo ormone si mantengono significativamente
maggiori rispetto a quelli del materiale di controllo (+123 e +44%, rispettivamente al termine
del periodo di trattamento e di recovery). Appare evidente come l’ABA rappresenti una prima
linea di difesa in virtù della sua rapida produzione (il suo picco si registra precedentemente a
quello di ET, Roychoudhury et al., 2013). In virtù del ben noto ruolo dell’ABA nella
regolazione stomatica (Kangasjärvi et al., 2005), possiamo ipotizzare che il marcato e celere
accumulo di ABA possa influenzare l’apertura/chiusura degli stomi in risposta al trattamento
con O3 (Ederli et al., 2006), consentendo a Q. ilex di adottare una strategia di esclusione nei
confronti dell’inquinante. Simili risultati sono stati riportati da Ahlfors et al. (2004) in piante
di Arabidopsis thaliana (Col-0, naturale e mutante) sottoposte ad un trattamento acuto con O3
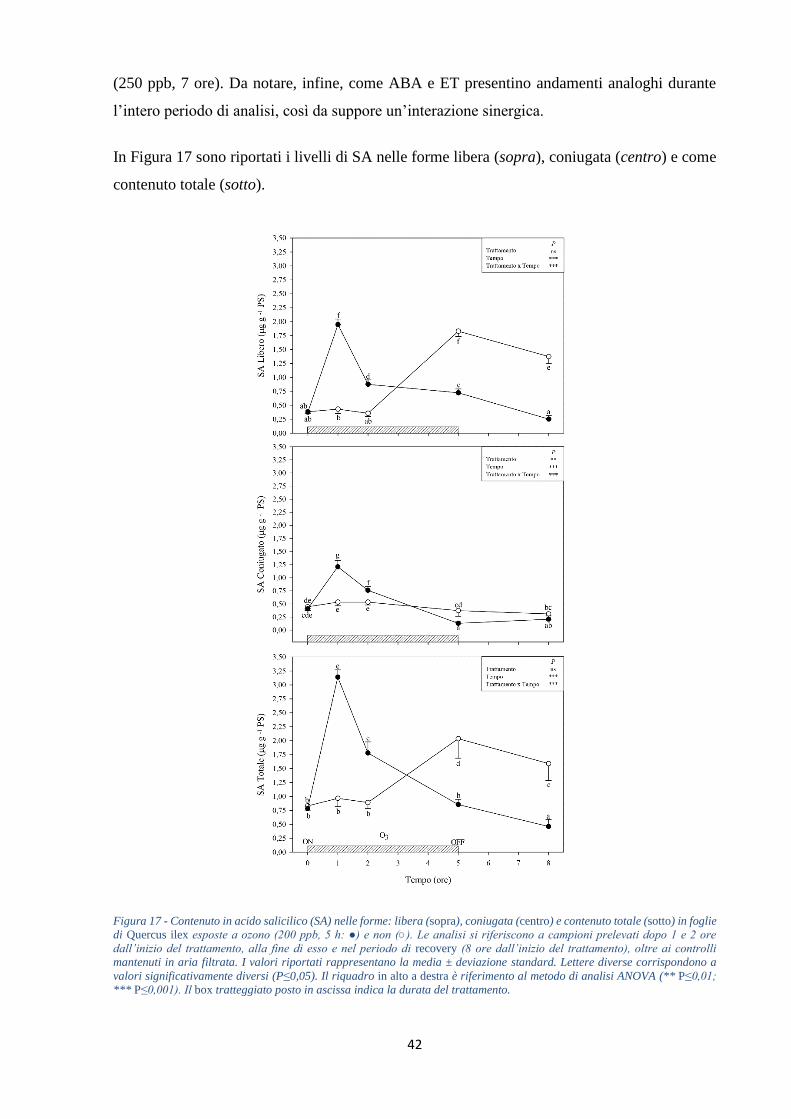
42
(250 ppb, 7 ore). Da notare, infine, come ABA e ET presentino andamenti analoghi durante
l’intero periodo di analisi, così da suppore un’interazione sinergica.
In Figura 17 sono riportati i livelli di SA nelle forme libera (sopra), coniugata (centro) e come
contenuto totale (sotto).
Figura 17 - Contenuto in acido salicilico (SA) nelle forme: libera (sopra), coniugata (centro) e contenuto totale (sotto) in foglie
di Quercus ilex esposte a ozono (200 ppb, 5 h: ●) e non (○). Le analisi si riferiscono a campioni prelevati dopo 1 e 2 ore
dall’inizio del trattamento, alla fine di esso e nel periodo di recovery (8 ore dall’inizio del trattamento), oltre ai controlli
mantenuti in aria filtrata. I valori riportati rappresentano la media ± deviazione standard. Lettere diverse corrispondono a
valori significativamente diversi (P≤0,05). Il riquadro in alto a destra è riferimento al metodo di analisi ANOVA (** P≤0,01;
*** P≤0,001). Il box tratteggiato posto in ascissa indica la durata del trattamento.

43
Nelle prime ore di esposizione, le piante rispondono allo stress ossidativo aumentando il
contenuto di SA (+352 e +146%, rispettivamente dopo 1 e 2 ore di trattamento per la forma
libera, +126 e +42% per quella coniugata). Nelle ore successive, le concentrazioni di SA
subiscono un marcato decremento (-60 e -65% dopo 5 ore, rispettivamente per la forma libera
e coniugata). Al termine del periodo di recovery, tali valori (e di conseguenza il contenuto
totale) si mantengono significativamente inferiori rispetto ai controlli (-82 e -33%,
rispettivamente dopo 8 ore). E’ noto in letteratura come il trattamento con O3 sia in grado di
stimolare la produzione di SA (Kangasjärvi et al., 2005), il quale svolge un importante ruolo
nella regolazione della risposta della pianta mediante un intenso scambio di informazioni con
altri ormoni (Durner et al., 1997; Tamaoki, 2008; Bartoli et al., 2013). In particolare, con la sua
influenza (i) regola la sintesi di ET, (ii) previene la diffusione della morte cellulare (Vahala et
al., 2003), (iii) attiva meccanismi di risposta antiossidanti, (iv) mantiene un stato redox cellulare
ottimale (Tamaoki et al., 2008; Pellegrini et al., 2013). Secondo Overmyer et al. (2005), studi
compiuti su piante di A. thaliana (Cvi-0 e rcd1, mutanti) sottoposte ad un trattamento acuto con
O3 (250 ppb, 6 ore) hanno documentato una maggiore tolleranza in termini di sviluppo dei
sintomi in presenza di ridotte concentrazioni di SA. Altri lavori hanno evidenziato l’importante
funzione di questa molecola nel prevenire la formazione di lesioni fogliari, come riportato da
Pasqualini et al. (2002) su piante di tabacco sottoposte ad un trattamento acuto con O3 (200
ppb, 3 ore) e Di Baccio et al. (2012) su pomodoro (200 ppb, 5 ore). A tal proposito, Pellegrini
et al. (2013) riportano un picco dei livelli di SA dopo 8 ore dall’inizio del trattamento in
individui di Melissa officinalis (200 ppb, 5 ore).
In Figura 18 sono riportati i livelli di JA. Nelle prime ore di esposizione, si osserva un
significativo incremento del contenuto di questo ormone rispetto ai controlli mantenuti in aria
filtrata (un aumento di nove e due volte, rispettivamente dopo 1 e 2 ore di trattamento). Al
termine della fumigazione, si nota un marcato decremento (-58%, rispetto ai controlli), con una
successiva stabilizzazione dei livelli di JA ai livelli dei controlli. E’ noto come l’esposizione a
O3 sia in grado di stimolare la produzione di JA (Rao et al., 2000), attraverso la formazione di
substrati derivati dalla perossidazione lipidica (in particolare acido α-linoleico). Nel ciclo di
PCD, JA (i) partecipa al contenimento delle lesioni prodotte da ROS (Kangasjärvi et al., 2005),
(ii) protegge i tessuti dalla morte cellulare (Overmyer et al., 2000) e (iii) interferisce
negativamente con la biosintesi di ET e l’accumulo di SA (Tamaoki, 2008).
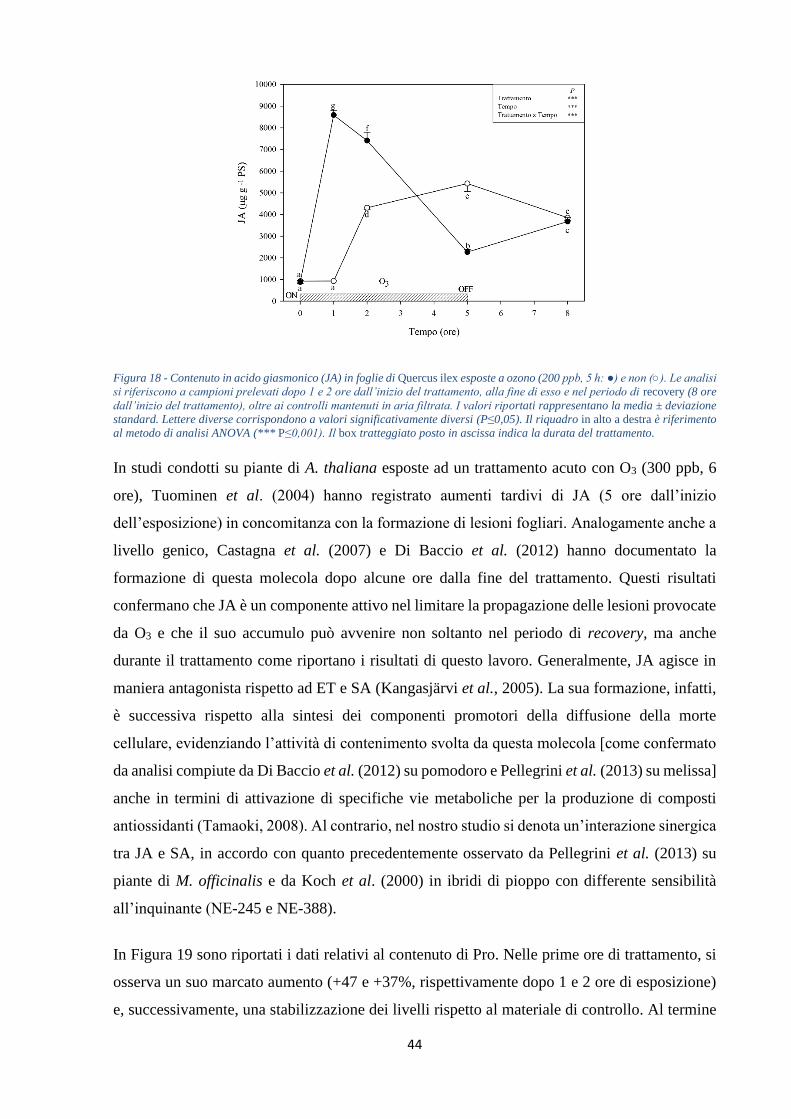
44
Figura 18 - Contenuto in acido giasmonico (JA) in foglie di Quercus ilex esposte a ozono (200 ppb, 5 h: ●) e non (○). Le analisi
si riferiscono a campioni prelevati dopo 1 e 2 ore dall’inizio del trattamento, alla fine di esso e nel periodo di recovery (8 ore
dall’inizio del trattamento), oltre ai controlli mantenuti in aria filtrata. I valori riportati rappresentano la media ± deviazione
standard. Lettere diverse corrispondono a valori significativamente diversi (P≤0,05). Il riquadro in alto a destra è riferimento
al metodo di analisi ANOVA (*** P≤0,001). Il box tratteggiato posto in ascissa indica la durata del trattamento.
In studi condotti su piante di A. thaliana esposte ad un trattamento acuto con O3 (300 ppb, 6
ore), Tuominen et al. (2004) hanno registrato aumenti tardivi di JA (5 ore dall’inizio
dell’esposizione) in concomitanza con la formazione di lesioni fogliari. Analogamente anche a
livello genico, Castagna et al. (2007) e Di Baccio et al. (2012) hanno documentato la
formazione di questa molecola dopo alcune ore dalla fine del trattamento. Questi risultati
confermano che JA è un componente attivo nel limitare la propagazione delle lesioni provocate
da O3 e che il suo accumulo può avvenire non soltanto nel periodo di recovery, ma anche
durante il trattamento come riportano i risultati di questo lavoro. Generalmente, JA agisce in
maniera antagonista rispetto ad ET e SA (Kangasjärvi et al., 2005). La sua formazione, infatti,
è successiva rispetto alla sintesi dei componenti promotori della diffusione della morte
cellulare, evidenziando l’attività di contenimento svolta da questa molecola [come confermato
da analisi compiute da Di Baccio et al. (2012) su pomodoro e Pellegrini et al. (2013) su melissa]
anche in termini di attivazione di specifiche vie metaboliche per la produzione di composti
antiossidanti (Tamaoki, 2008). Al contrario, nel nostro studio si denota un’interazione sinergica
tra JA e SA, in accordo con quanto precedentemente osservato da Pellegrini et al. (2013) su
piante di M. officinalis e da Koch et al. (2000) in ibridi di pioppo con differente sensibilità
all’inquinante (NE-245 e NE-388).
In Figura 19 sono riportati i dati relativi al contenuto di Pro. Nelle prime ore di trattamento, si
osserva un suo marcato aumento (+47 e +37%, rispettivamente dopo 1 e 2 ore di esposizione)
e, successivamente, una stabilizzazione dei livelli rispetto al materiale di controllo. Al termine
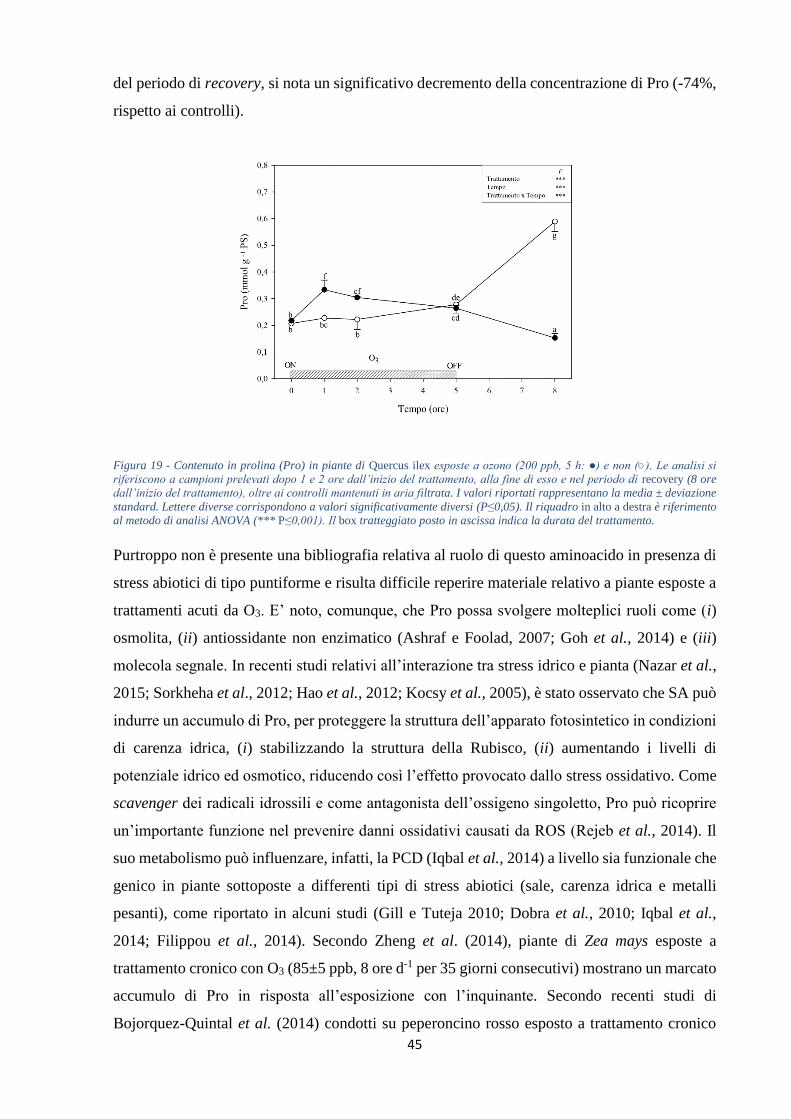
45
del periodo di recovery, si nota un significativo decremento della concentrazione di Pro (-74%,
rispetto ai controlli).
Figura 19 - Contenuto in prolina (Pro) in piante di Quercus ilex esposte a ozono (200 ppb, 5 h: ●) e non (○). Le analisi si
riferiscono a campioni prelevati dopo 1 e 2 ore dall’inizio del trattamento, alla fine di esso e nel periodo di recovery (8 ore
dall’inizio del trattamento), oltre ai controlli mantenuti in aria filtrata. I valori riportati rappresentano la media ± deviazione
standard. Lettere diverse corrispondono a valori significativamente diversi (P≤0,05). Il riquadro in alto a destra è riferimento
al metodo di analisi ANOVA (*** P≤0,001). Il box tratteggiato posto in ascissa indica la durata del trattamento.
Purtroppo non è presente una bibliografia relativa al ruolo di questo aminoacido in presenza di
stress abiotici di tipo puntiforme e risulta difficile reperire materiale relativo a piante esposte a
trattamenti acuti da O3. E’ noto, comunque, che Pro possa svolgere molteplici ruoli come (i)
osmolita, (ii) antiossidante non enzimatico (Ashraf e Foolad, 2007; Goh et al., 2014) e (iii)
molecola segnale. In recenti studi relativi all’interazione tra stress idrico e pianta (Nazar et al.,
2015; Sorkheha et al., 2012; Hao et al., 2012; Kocsy et al., 2005), è stato osservato che SA può
indurre un accumulo di Pro, per proteggere la struttura dell’apparato fotosintetico in condizioni
di carenza idrica, (i) stabilizzando la struttura della Rubisco, (ii) aumentando i livelli di
potenziale idrico ed osmotico, riducendo così l’effetto provocato dallo stress ossidativo. Come
scavenger dei radicali idrossili e come antagonista dell’ossigeno singoletto, Pro può ricoprire
un’importante funzione nel prevenire danni ossidativi causati da ROS (Rejeb et al., 2014). Il
suo metabolismo può influenzare, infatti, la PCD (Iqbal et al., 2014) a livello sia funzionale che
genico in piante sottoposte a differenti tipi di stress abiotici (sale, carenza idrica e metalli
pesanti), come riportato in alcuni studi (Gill e Tuteja 2010; Dobra et al., 2010; Iqbal et al.,
2014; Filippou et al., 2014). Secondo Zheng et al. (2014), piante di Zea mays esposte a
trattamento cronico con O3 (85±5 ppb, 8 ore d-1 per 35 giorni consecutivi) mostrano un marcato
accumulo di Pro in risposta all’esposizione con l’inquinante. Secondo recenti studi di
Bojorquez-Quintal et al. (2014) condotti su peperoncino rosso esposto a trattamento cronico
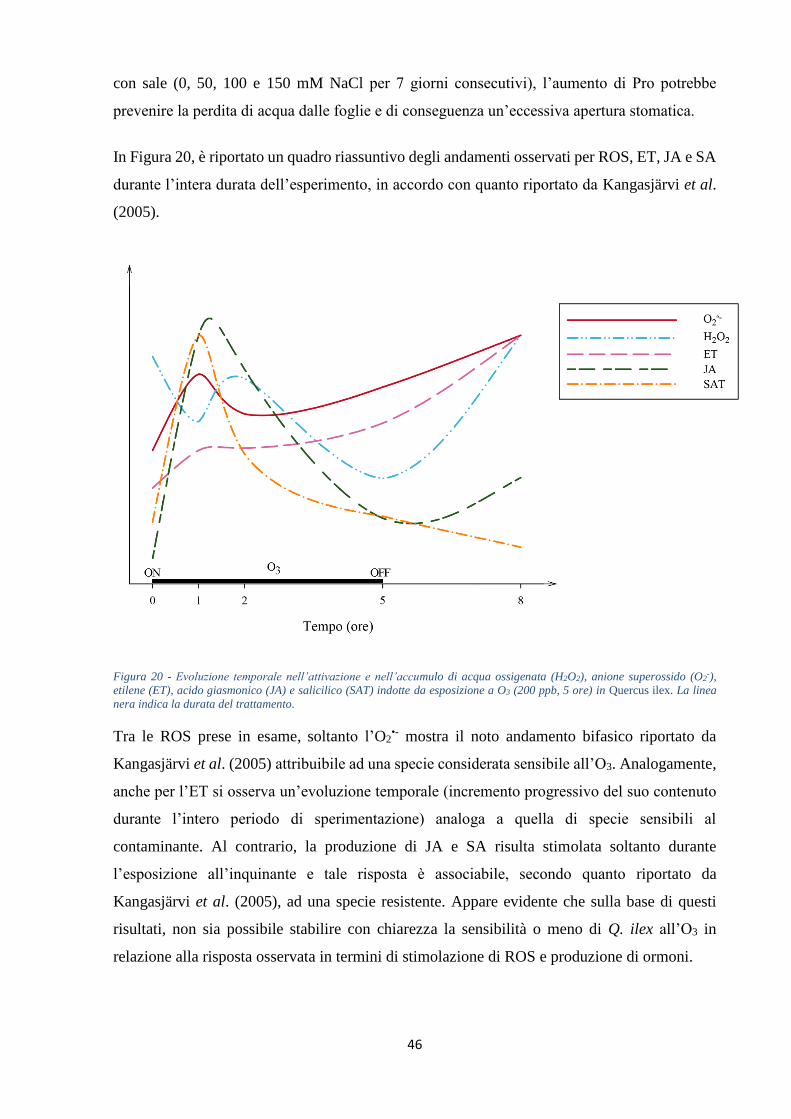
46
con sale (0, 50, 100 e 150 mM NaCl per 7 giorni consecutivi), l’aumento di Pro potrebbe
prevenire la perdita di acqua dalle foglie e di conseguenza un’eccessiva apertura stomatica.
In Figura 20, è riportato un quadro riassuntivo degli andamenti osservati per ROS, ET, JA e SA
durante l’intera durata dell’esperimento, in accordo con quanto riportato da Kangasjärvi et al.
(2005).
Figura 20 - Evoluzione temporale nell’attivazione e nell’accumulo di acqua ossigenata (H2O2), anione superossido (O2-),
etilene (ET), acido giasmonico (JA) e salicilico (SAT) indotte da esposizione a O3 (200 ppb, 5 ore) in Quercus ilex. La linea
nera indica la durata del trattamento.
Tra le ROS prese in esame, soltanto l’O2•- mostra il noto andamento bifasico riportato da
Kangasjärvi et al. (2005) attribuibile ad una specie considerata sensibile all’O3. Analogamente,
anche per l’ET si osserva un’evoluzione temporale (incremento progressivo del suo contenuto
durante l’intero periodo di sperimentazione) analoga a quella di specie sensibili al
contaminante. Al contrario, la produzione di JA e SA risulta stimolata soltanto durante
l’esposizione all’inquinante e tale risposta è associabile, secondo quanto riportato da
Kangasjärvi et al. (2005), ad una specie resistente. Appare evidente che sulla base di questi
risultati, non sia possibile stabilire con chiarezza la sensibilità o meno di Q. ilex all’O3 in
relazione alla risposta osservata in termini di stimolazione di ROS e produzione di ormoni.

47
5. CONSIDERAZIONI CONCLUSIVE
Ad oggi, scarsa attenzione è stata rivolta allo studio dei possibili effetti di vari agenti di stress
(biotici ed abiotici) su specie quercine. Il numero di lavori sperimentali volti ad analizzare gli
eventuali meccanismi attivati in presenza di O3 è assai ridotto (e.g. Calatayud et al., 2011,
Guerrero et al., 2013).
In virtù delle analogie osservate tra le risposte indotte dall’O3 e da un patogeno, è possibile
affermare che questo inquinante agisca similmente ad un agente biotico (Rao et al., 2000).
Recentemente, è stato dimostrato che tali somiglianze si riscontrano non soltanto a livello
macroscopico con la comparsa di sintomi visibili (quali necrosi e clorosi), ma anche in termini
fisiologici, biochimici e molecolari. Di conseguenza, l’analisi di tutti gli eventi che si innescano
e si susseguono prima della manifestazione del danno può risultare utile nell’individuazione e
nello studio di quelle molecole responsabili dell’attivazione dei meccanismi di difesa
(Pellegrini et al., 2013).
Possiamo concludere che Q. ilex non mostra danni macroscopici indotti dal nostro trattamento
con O3, sebbene a livello microscopico i risultati ottenuti dalla analisi istologiche evidenziano
l’attivazione di PCD nei tessuti. L’H2O2 non svolge il suo noto ruolo di molecola segnale
durante il periodo di fumigazione, mentre O2•- mostra un andamento bifasico confermando la
presenza di un burst ossidativo a seguito del trattamento con l’inquinante. Gli ormoni e le
molecole segnale si attivano e interagiscono durante le varie fasi del trattamento, documentando
come l’O3 possa agire come un agente patogeno. In particolare, si registra una produzione di
ET per l’intera durata dell’esperimento coincidente con quella di ABA che risponde in maniera
più marcata e rapida suggerendo che Q. ilex sia in grado di attivare una strategia di esclusione
(chiusura stomatica) in risposta all’inquinante. SA e JA mostrano un andamento analogo,
lasciando supporre che vi sia un’interazione sinergica che regoli la diffusione della lesione.
Questi risultati suggeriscono che Q. ilex in conseguenza di un episodio puntiforme di O3 non
mostra sintomi macroscopici, pur subendo morte cellulare a causa del burst ossidativo prodotto
dal trattamento a cui è in grado di rispondere attivando PCD. Per valutare l’effettivo utilizzo
come pianta ornamentale in città (in particolare quelle caratterizzate da livelli medio-alti di
inquinamento da O3), è necessario condurre ulteriore studi, approfondendo anche l’analisi delle
possibili risposte ad altre tipologie di stress, prese in esame singolarmente o in combinazione
con O3.

48
6. BIBLIOGRAFIA
Able A.J., Guest D.I., Sutherland M.W. (1998) - Use of a new tetrazolium-based assay to
study the production of superoxide radicals by tobacco cell cultures challenged with avirulent
zoospores of phytophthora parasitica var nicotianae - Plant Physiol. 117: 491-499.
Ahlfors R., Macioszek V., Rudd J., Brosché M., Schlichting R., Scheel D., Kangasjärvi J.
(2004) - Stress hormone-independent activation and nuclear translocation of mitogen-activated
protein kinases in Arabidopsis thaliana during ozone exposure - The Plant Journal 40:4: 512-
522.
An Z., Jing W., Liu Y., Zhang W. (2008) - Hydrogen peroxide generated by copper amine
oxidase is involved in abscisic acid-induced stomatal closure in Vicia faba - J Exp Bot 59: 815-
825.
Ashraf M., Fooladb M.R. (2007) - Roles of glycine betaine and proline in improving plant
abiotic stress resistance - Environmental and Experimental Botany 59:2: 206-216.
Baier M., Kandlbinder A., Golldack D., Dietz K.J. (2005) - Oxidative stress and ozone:
perception, signalling and response - Plant Cell Environ 28: 1012-1020
Bartoli C.G., Casalongué C.A., Simontacchi M., Marquez-Garcia B., Foyer C.H. (2013) -
Interactions between hormone and redox signalling pathways in the control of growth and cross
tolerance to stress - Environmental and Experimental Botany 94: 73–88.
Bates L.S., Waldren R.P., Teare J.D. (1973) – Rapid determination of free proline for water
stress studies – Plant and Soil 39: 205-207.
Beauchamp J., Wisthaler A., Hansel A., Kleist E., Miebach M., Niinemets Ǜ., Schurr U.,
Wildt J., (2005) - Ozone induced emissions of biogenic VOC from tobacco: relationships
between ozone uptake and emission of LOX products - Plant, Cell and Environment 28: 1334-
1343
Bojórquez-Quintal E., Velarde-Buendía A., Ku-González Á., Carillo-Pech M., Ortega-
Camacho D., Echevarría-Machado I.,1 Pottosin I., Martínez-Estévez M. (2014) -
Mechanisms of salt tolerance in habanero pepper plants (Capsicum chinense Jacq.): Proline
accumulation, ions dynamics and sodium root-shoot partition and compartmentation – Plant
Sci. 5: 605.
Booker F.L., Burkey K.O., Jones A.M. (2012) – Re-evaluating the role of ascorbic acid and
phenolic glycosides in ozone scavenging in the leaf apoplast of Arabidopsis thaliana L. – Plant,
Cell and Environment 35: 1456-1466.
Boyer J.S., Silk W.K. (2004) - Hydraulics of plant growth - Func. Plant Biol. 31: 761-773.
Bussotti F., Ferrini F., Pollastrini M., Fini A. (2013) - The challenge of Mediterranean
sclerophtllous vegetation under climate change: From acclimatation to adaptation -
Enviromental and Experimental Botany xxx (2013) xxx-xxx (article in press).
Bytnerowicz A., Omasa K, Paoletti E. (2007) - Integrated effects of air pollution and climate
change on forests: a northern emisphere perspective - Enviromental Pollution 147: 348-445.
Calatayud V., Cerveró J., Calvo E., García-Breijo F-J., Reig-Armiñana J., Sanz M.J.
(2011) - Responses of evergreen and deciduous Quercus species to enhanced ozone levels -
Environmental Pollution 159: 55-63.
Castagna A., Ederli L., Pasqualini S., Mensuali-Sodi A., Baldan B., Donnini S., Ranieri A.
(2007) - The tomato ethylene receptor LE-ETR3 (NR) is not involved in mediating ozone
sensitivity: casual relationships among ethylene emission, oxidative burst and tissue damage -
New Phytol 174: 342-356.

49
Castagna, A., Ranieri, A. (2009) - Detoxification and repair process of ozone injury: From O₃ uptake to gene expression adjustmen - Environmental Pollution 157: 1461-1469.
Cavanagh J.A., Zwar- Reza E., Wilson J.G. (2009) - spatial attenuation of ambient particulate
matter air pollution within an urbanized native forest patch - Urban Forestry and Urban
Greening 8(1): 21-30.
Chameides W.L., Kasibhatla P.S., Yienger J., Levy H. (1994) - Growth of continental scale
metro-agro-plexes, regional ozone pollution, and world food production - Science 264: 74-77.
Cheaib A., Badeau V., Boe V., Chuine I., François C., Gritti E.S., Legay M., Pagè C.,
Thuiller W., Viovy N., Leadley P. (2012) - Climate change impacts on tree ranges: model
intercomparison faciclitates understanding and quantification of uncertainty - Ecology Letters
15: 533- 544.
Christensen J.H., Hewitson B., Busuioc A., Chen a., Gao X., Held I., Jones R., Kolli R.K.,
Kwon W.T., Laprise R., Magaña Rueda V., Mearns L., Menendez C.G., Räisänen J.,
Rinke A., Sarr A., Whetton P. (2007) - Regional climate projections - Climate change 2007:
The Physical Science Basis. Contribution of Working Group 1 to the Fourth Assesment Report
on Intergovernmental Panel on Climate Change - Cambridge University Press, UK and NY,
USA: Chapter 11.
Chuwah C., van Noijea T., van Vuurenb D.P., Stehfestb E., Hazelegera W. (2015) - Global
impacts of surface ozone changes on crop yields and land use - Atmospheric Env. 106: 11–23.
Cleland E.E., Chuine I., Menzel A., Mooney H.A., Schwartz M.D. (2007) - Shifting plant
phenology in response to global change - Trends Ecology and Evolution 22: 357 – 365.
Colbeck I., MacKenzie A.R. (1994) - Air pollution by photochemical oxidants. In: Air Quality
Monogrphs (Elsevier Science Ed.) - London, pp. 376.
Dar T.A., Uddin T., Khan M.M.A., Hakeem K.R., Jaleel H. (2015) - Jasmonates counter
plant stress: a review - Environ Exp Bot 115: 49-57.
Davies K.J.A. (2001) - Oxidative stress, antioxidant defenses, and damage removal, repair, and
replacement systems - International Union of Biochemistry and Molecular Biology Life 50:
279-289.
De Micco V., Aronne G. (2012) - Morpho- anatomical traits for plant adaptation to drought.
In: Aroca R. (Ed.), Plant responses to Drought stress - Springer Verlag, Berlin: 37-61.
Derwent, R.G., Apling, A.J., Ashmore, M.R, Ball, D.J., Clark, P., Cocks, A.T., Cox, R.A.,
Fowler, D., Gay, M., Harrison, M.R., Jenkins, G.J., Kay, P.J.A., Laxen, D.P.H., Martin,
A., McKenna, D., Penkett, S.A., Williams, M.L., Woods S, P.T. (1987) - Ozone in the United
Kingdom - Department of Environment e Department of Transport, London.
Devoto A., Turner J.G. (2005) - Jasmonate-regulated Arabidopsis stress signaling network -
Physiol Plant 123: 161-172.
Di Baccio D., Ederli L., Marabottini R., Badiani M., Francini A., Nali C., Antonelli M.,
Santangelo E., Sebastiani L., Pasqualini S. (2012) - Similar foliar lesions but opposite
hormonal patterns in a tomato mutant impaired in ethylene perception and its near isogenic wild
type challenged with ozone - Environ Exp Bot 75: 286-297.
Diara, C., Castagna, A., Baldan, B., Mensuali Sodi, A., Sahr, T., Langebartels, C.,
Sebastiani, L., Ranieri, A. (2005) - Differences in the kinetics and scale of signaling molecule
production modulate the ozone sensitivity of hybrid poplar clones: the roles of H2O2, ethylene
and salicylic acid - New Phytologist 168, 351–364.
Dobra J., Motyka V., Dobrev P., Malbeck J., Prasil I., Haisel D. (2010) - Comparison of
hormonal responses of heat, drought and combined stress in tobacco plants with elevated
proline content - J Plant Physiol 167: 1360-1370.

50
Döring A.S., Pellegrini E., Campanella A., Trivellini A., Gennai C., Petersen M., Nali C.,
Lorenzini G. (2014) - How sensitive is Melissa officinalis to realistic ozone concentrations? -
Plant Physiol Biochem 74: 156-164.
Durner J., Shah J., Klessig D.F. (1997) - Salicylic acid and disease resistance in plants -
Trends in Plant Science 2:7: 266–274.
Ederli L., Morettini R., Borgogni A., Wasternack C., Miersch O., Reale L., Ferranti F.,
Tosti N., Pasqualini S. (2006) - Interaction between nitric oxide and ethylene in the induction
of alternative oxidase in ozone-treated tobacco plants - Plant Physiol. 142:2: 595-608.
EEA (2013) - Air quality in Europe, 2013 report - September 2013.
Evans N.H., McAinsh M.R., Hetherington A.M., Knight M.R. (2005) - ROS perception in
Arabidopsis thaliana: the ozone-induced calcium response - Plant J 41: 615-626.
Fam S.S., Morrow J.D. (2003) - The isoprostanes: unique products of arachidonic acid
oxidation-a review - Current Medicinal Chemistry 10: 1723-1740.
Faoro F., Iriti M. (2005) - Cell death behind invisible symptoms: early diagnosis of ozone
injury - Biologia Plantarum 49: 585–592.
Filippou P., Bouchagier P., Skotti E., Fotopoulos V. (2014) - Proline and reactive
oxygen/nitrogen species metabolism is involved in the tolerant response of the invasive plant
species Ailanthus altissima to drought and salinity - Environ Exp Bot 97: 1-10.
Fini A., Ferrini F. (2007) - Influenza dell’ambiente urbano sulla fisiologia e la crescita degli
alberi - Italus Hortus 14 (1): 9-24.
Fishman J., Vukovich F.M., Browell E.V. (1985) - The photochemistry of synoptic-scale
ozone synthesis: implications for the global ozone budget - Journal of Atmospheric Chemistry
3: 299-320.
Fowler D., Cape J.N., Coyle M., Flechard C., Kuylenstierna J., Hicks K., Derwent D.,
Johnson C., Stevenson D. (1999) - The global exposure of forets to air pollutants - Water, Air,
and Soil Pollution, 116: 5-32.
Gago E.J., Roldan J., Pacheco- Torres R., Ordonez J. (2013) - The city and urban heat
islands: A review of strategies to mitigate adverse effects - Renewable and Sustainable Energy
Reviews 25: 749- 758.
Gill S.S., Tuteja N. (2010) - Reactive oxygen species and antioxidant machinery in abiotic
stress tolerance in crop plants - Plant Physiology and Biochemistry 48: 909-930.
Giridharan R., Ganesan S., Lau S.S.Y. (2004) - Daytime urban heat Island effect in high-rise
and high- density residential developments in Hong Kong - Energy and buildings 36: 525- 34.
Goh E.J., Kim J., Kim W., Ha B., Kim S.H., Kang S., Seo Y.W., Kim D.S. (2014) -
Physiological changes and anti-oxidative responses of Arabidopsis plants after acute and
chronic c-irradiation - Radiat Environ Biophys 53: 677–693.
Gomi K., Ogawa D., Katou S., Kamada H., Nakajima N., Saji H., Soyano T., Sasabe M.,
Machida Y., Mitsuhara I., Ohashi Y., Seo S. (2005) - A mitogen-activated protein kinase
NtMPK4 activated by SIPKK is required for jasmonic acid signaling and involved in ozone
tolerance via stomatal movement in tobacco - Plant Cell Physiol 46: 1902-1914.
Guerra J.C., Rodrìguez S., Arencibia M.T., Garcìa M.D. (2004) - Study on the formation of
ozone in relation to the air quality management and vegetation protection in Tenerife (Canary
Islands) - Chemosphere, 56: 1157-1167.
Guerrero C.C., Günthardt-Goerg M.S., Vollenweider P. (2013) - Foliar symptoms triggered
by ozone stress in irrigated holm oaks from the city of Madrid, Spain - PLoS ONE 8:7:e69171.
doi: 10.1371/journal.pone.0069171.

51
Halliwell, B., Gutteridge, J.M.C. (1989) - Free radicals in biology and medicine - Clarendon
Press, Oxford. Second Edition, pp. 851.
Hao J.H., Yi Y., Shang Q.M., Dong C.J., Zhang Z.G. (2012) - Effects of exogenous salicylic
acid on membrane lipid peroxidation and photosynthetic characteristics of Cucumis sativus
seedlings under drought stress - Ying Yong Sheng Tai Xue Bao, 23: 717–723.
Heath R.L. (1987) - The biochemistry of ozone attack on the plasma membrane of plant cells
- Recent Advances in Phytochemistry 21: 29-54.
Heath R.L. (2008) - Modification of the biochemical pathways of plants induced by ozone:
What are the varied routes to change? - Environmental Pollution 155: 453–463,
doi:10.1016/j.envpol.2008.03.010.
Hidy G.M., Muller P.K. (1986) - The sulfur ossido-particulate matter complex - Air pollutants
and their effects on the terrestrial ecosystems (A.H. Legge, S.V. Krupa Eds.) - Wiley, New
York, pp. 51-104.
Hopkin M. (2007) - Carbon sinks threatened by increasing ozone - Nature 448: 396-397.
Horváth E, Szalai G, Janda T (2007) - Induction of abiotic stress tolerance by salicylic acid
signaling. J Plant Growth Regul 26: 290-300.
Hoshika Y., Watanabe M., Inada N., Koike T. (2013) - Model-based analysis of avoidance
of ozone stress by stomatal closure in Siebold’s beech (Fagus crenata) - Ann. Bot. 112: 1149-
1158.
IPCC (2013) Fifth Assessment Report. http://www.ipcc.ch/report/ar5/index/.shtml
Iqbal N, Umar S, Khan NA, Khan MIR (2014) - A new perspective of phytohormones in
salinity tolerance: regulation of proline metabolism. Environ Exp Bot 100: 34-42.
Iriti M., Faoro F. (2008) – Oxidative stress, the paradigm of ozone toxicity in plants and
animals - Water, Air, and Soil Pollution 187:1: 285-301.
Iriti M., Rabotti G., De Ascensao A., Faoro F. (2003) – Benthiadiazole-induced resistance
modulates ozone tolerance - Journal of Agricultural and Food Chemistry 51: 4308-4314.
Jabs T. (1999) - Reactive oxygen intermediates as mediators of programmed cell death in
plants and animals - Biochem Pharmacol 57: 231-245.
Jayakannan M., Bose J., Babourina O., Shabala S., Massart A., Poschenrieder C., Rengel
Z. (2015) - The NPR1-dependent salicylic acid signalling pathway is pivotal for enhanced salt
and oxidative stress tolerance in Arabidopsis - J. Exp. Bot. 66:7: 1865-1875.
Jones J.D.G., Dangl J.L. (2006) - The plant immune system - Nature 444: 323-329.
Joo J.H., Wang S., Chen J.G., Jones A.M., Fedoroff N.V. (2005) - Different signaling and
cell death roles of heterotrimeric G protein alpha and beta subunits in the Arabidopsis oxidative
stress response to ozone - Plant Cell 17: 957-970.
Junge C.E. (1962) - Global ozone budget and exchange between stratosphere and troposphere
- Tellus, 14: 363-377.
Kangasjärvi J., Jaspers P., Kollist H. (2005) - Signalling and cell death in ozone-exposed
plants – Plant, Cell and Environment 28: 1021-1036.
Kanofsky G., Sima H. (1995) - Singlet oxygen generation from the reaction of O3 with plant
leaves - Journal of Biological Chemistry 270: 7850-7852.
Kazan K. (2015) - Diverse roles of jasmonates and ethylene in abiotic stress tolerance - Trends
Plant Sci 20: 219-229.
Kelly F.J., Mudway I.S. (2003) - Protein oxidation at the air lung interface - Amino Acids 25:
375-396.
Kerstiens G., Lendzian K.J. (1989) – Interactions between ozone and plant cuticles. 1. Ozone
deposition and permeability - New Phytologist 112: 13-19.

52
Khokon AR, Okuma E, Hossain MA, Munemasa S, Uraji M, Nakamura Y, Mori IC,
Murata Y (2011) - Involvement of extracellular oxidative burst in salicylic acid-induced
stomatal closure in Arabidopsis - Plant Cell Environ. 34: 434-443.
Klein T., di Matteo G., Roenberg E., Cohen S., Yakir D. (2012) - Differential
ecophysiological response of a major Mediterranean pine species across a climatic gradient -
Tree physiology 33: 26- 36.
Koch J.R., Creelman R.A., Eshita S.M., Seskar M., Mullet J.E., Davis K.R. (2000) - Ozone
sensitivity in hybrid poplar correlates with insensitivity to both salicylic acid and jasmonic acid.
The role of programmed cell death in lesion formation - Plant Physiol. 123:2: 487-496.
Kocsy G., Laurie R., Szalai G., Szilagyi V., Simon-Sarkadi L., Galiba G., De Ronde, J.A.
(2005) - Genetic manipulation of proline levels affects antioxidants in soybean subjected to
simultaneous drought and heat stresses - Physiol. Plant., 124: 227–235.
Kouvarakis G.K., Tsigaridis M., Kanakidou, Mihalopoulos N. (2000) - Temporal variations
of surface regional background ozone over Crete Island in southeast Mediterranean - Journal
of Geophysical Research, 105: 4399-4407.
Kramell R., Miersch O., Schneider G., Wasternack C. (1999) - Liquid Chromatography of
Jasmonic Acid Coniugates - Chromatographia: 49, 42-46.
Kudla J, Batistic O, Hashimoto K (2010) - Calcium signals: the lead currency of plant
information processing - Plant Cell 22: 541-563.
Laisk A., Kull O., Moldau H. (1989) - Ozone concentration in leaf intercellular air spaces is
close to zero - Plant Physiology 90: 1163-1167.
Lamb C., Dixon R.A. (1997) - The oxidative burst in plant disease resistance - Annu Rev Plant
Physiol Plant Mol Biol 48: 251-275.
Lefohn A.S. (1992) - Exposure-yield response relationships. In: Surface level ozone exposures
and their effects on vegetation (AS. Lefohn Ed.) - Lewis Publishers, London, pp. 236-239.
Lei H., Wuebbles D.J., Liang X.Z. (2012) - Projected risk of high ozone episodes in 2050 -
Atmospheric environment 59: 567- 577.
Lin Z., Zhong S., Grierson D. (2009) - Recent advances in ethylene research - J Exp Bot 60:
3311-3336.
Liu H., Wang Y., Xu J., Su T., Liu G., Ren D. (2008) - Ethylene signalling is required for the
acceleration of cell death induced by activation of AtMEK5 in Arabidopsis - Cell Research 18:
422-432.
Lorenzini G, Nali C., Pellegrini E. (2014a) - Summer heat waves, agriculture, forestry and
related issues: an introduction (Editorial) - Agrochimica, LVIII, Special Issue: 3-19.
Lorenzini G., Nali C. (2005) - Le Piante e l’inquinamento dell’aria - Sprinter-Verlag: 256.
Lorenzini G., Nali C., Biagioni M. (1995) - Long–range transport of photochemical ozone
over the Tyrrhenian Sea demonstrated by a new miniaturized bioassay with ozone–sensitive
tobacco seedlings - The Science of the Total Environment 166: 193-199.
Lorenzini G., Pellegrini E., Campanella A., Nali C. (2014b) - It’s not just the heat and the
drought: the role of ozone air pollution in the 2012 heat wave - Agrochimica, LVIII, Special
Issue: 40-52
Ludwików A., Kierzek D., Gallois P., Zeef L., Sadowski J. (2009) - Gene expression
profiling of ozone-treated Arabidopsis abi1td insertional mutant: protein phosphatase 2C ABI1
modulates biosynthesis ratio of ABA and ethylene - Planta 230: 1003-1017
McKee D.J. (1993) - Tropospheric ozone - Lewis Publishers: 223 pp.
Meehl G.A., Stocke T.F., Collins, W.D., Friedlingstein P., Gaye A.T., Gregory J.M., Kitoh
A.,Knutti R., Murphy J.M., Noda A., Raper S.C.B., Watterson I.G., Weaver A.J., Zhao

53
Z.C. (2007) - Global Climate Projections - Climate Change (2007): The Physical Science Basis,
Contribution of Working Group 1 to the Fourth Assessment Report of the Intergovernmental -
- Cambridge University Press.
Meng X., Xu .J, He Y., Yang K.Y., Mordorski B., Liu Y., Zhang S. (2013) - Phosphorylation
of an ERF transcription factor by Arabidopsis MPK3/ MPK6 regulates plant defense gene
induction and fungal resistance - Plant Cell 25: 1126-1142.
Mensuali-Sodi A., Panizza M., Tognoni F. (1992) - Quantification of ethylene losses in
different container-seal systems and comparison of biotic and abiotic contributions to ethylene
accumulation in tissue cultures - Physiologia Plantarum 84: 472-476.
Mittler R., Blumwald E. (2015) - The roles of ROS and ABA in Systemic Acquired
Acclimation - Plant Cell 27: 64-70.
Moeder W, Barry CS, Tauriainen AA, Betz C, Tuomainen J, Utriainen M, Grierson D,
Sandermann H, Langebartels C, Kangasjärvi J (2002) - Ethylene synthesis regulated by
biphasic induction of 1-aminocyclopropane-1-carboxylic acid synthase and 1-
aminocyclopropane-1-carboxylic acid oxidase genes is required for hydrogen peroxide
accumulation and cell death in ozone-exposed tomato - Plant Physiol 130:1918-1926
Mohanty J.G., Jaffe J.S., Schulman E.S., Raible D.G. (1997) - A highly sensitive fluorescent
micro-assay of H2O2 release from activated human leukocytes using a dihydroxyphenoxazine
derivative - Journal of Immunological Methods 202: 133–141
Mudd J.B. (1997) - Biochemical basis for the toxicity of ozone - Plant response to air pollution
- Wiley: 267-284
Nakajima N, Matsuyama T, Tamaoki M, Saji H, Aono M, Kubo A, Kondo N (2001) -
Effects of ozone exposure on the gene expression of ethylene biosynthetic enzymes in tomato
leaves - Plant Physiol Biochem 39: 993-998.
Nali C., Lorenzini G. (2009) - L’ozono, causa di gravi malattie alle piante - Protezione delle
colture 2: 4-11.
Nazar R., Umar S., Khan N.A., Sareer O. (2015) - Salicylic acid supplementation improves
photosynthesis and growth in mustard through changes in proline accumulation and ethylene
formation under drought stress - South African Journal of Botany 98: 84–94.
Nickstadt A., Thomma B.P.H.J., Feussner I., Kangasjärvi J., Zeier J., Loeffler C., Scheel
D., Berger S. (2004) - The jasmonate-insensitive mutant jin1 shows increased resistance to
biotrophic as well as necrotrophic pathogens - Mol Plant Pathol 5: 425-434.
Nowak D. J., Civerolo K.L., Rao S.T. Sistla G., Luley C.J., Crane D.E. (2000) - A modeling
study of the impact of urban trees on ozone - Atmosferic environment 34: 1601- 1613.
Nowak D.J., Dwyer J.F. (2007) - Understanding the benefits and costs of urban forest
ecosystems - Urban and Community Forestry in the Northeast: 25-46.
Ogawa D., Nakajima N., Tamaoki M., Aono M., Kubo A., Kamada H., Saji H. (2007) -
The isochorismate pathway is negatively regulated by salicylic acid signaling in O3-exposed
Arabidopsis - Planta 226: 1277-1285.
Okada K., Abe H., Arimura G. (2015) - Jasmonates induce both defense responses and
communication in monocotyledonous and dicotyledonous plants - Plant Cell Physiol 56: 16-
27.
Ordòñez C., Duinker P.N., Steenberg J. (2010) - Climate change Mitigation and Adaptation
in Urban Forest: a Framework for Suistainable Urban Forest Management - Paper prepared for
presentation at 18th commonwealth Forestry Ceonference, Edinburgh.
Örvar B-L., McPherson J., Ellis E.B. (1997) - Pre-activating wounding response in tobacco
prior to high-level ozone exposure prevents necrotic injury - Plant J 11: 203-212.

54
Overmyer K., Brosché M., Pellinen R., Kuittinen T., Tuominen H., Ahlfors R., Keinänen
M., Saarma M., Scheel D., Kangasjärvi J. (2005) - Ozone-Induced Programmed Cell Death
in the Arabidopsis radical-induced cell death Mutant - Plant Physiol. 137:3: 1092-1104.
Overmyer K., Kollist H., Tuominen H. (2008) - Complex phenotypic profiles leading to
ozone sensitivity in Arabidopsis thaliana mutants - Plant Cell Environ 31: 1237-1249.
Overmyer K., Tuominen H., Kettunen R., Betz C., Langebartels C., Sandermann Jr. H.,
Kangasjärvi J. (2000) - Ozone-sensitive Arabidopsis rcd1 mutant reveals opposite roles for
ethylene and jasmonate signaling pathways in regulating superoxide-dependent cell death – The
Plant Cell 12:10: 1849-1862.
Pasqualini S, Della Torre G, Ferranti F, Ederli L, Piccioni C, Reale L, Antonielli M (2002)
- Salicylic acid modulates ozone-induced hypersensitive cell death in tobacco plants - Physiol
Plant 115: 204-212.
Pellegrini E., Lorenzini G., Nali C. (2007) – The 2003 European heat wave: which role for
ozone? Some data from Tuscany, Central Italy - Water, Air and Soil Pollution 181: 401-408.
Pellegrini E., Trivellini A., Campanella A., Francini A., Lorenzini G., Nali C., Vernieri P.
(2013) - Signaling molecules and cell death in Melissa officinalis plants exposed to ozone -
Plant Cell Rep 32: 1965-1980.
Petit R.J., Hampe A., Cheddadi R. (2005) - Climate Change and tree phylogeography in the
Mediterranean - Taxon 54: 877-885.
Rao M.V., Koch J., Davis K.R. (2000) - Ozone: a tool for probing programmed cell death in
plants - Plant Molecular Biology 44: 345-358.
Rao M.V., Lee H-I., Davis K.R. (2002) - Ozone-induced ethylene production is dependent on
salicylic acid, and both salicylic acid and ethylene act in concert to regulate ozone-induced cell
death - Plant J 32: 447-456.
Reich P.B. (1987) - Quantifying plant response to ozone: a unifying theory - Tree Physiology
3: 63-91.
Rejeb K.B., Abdelly C., Savouré A. (2014) How reactive oxygen species and proline face
stress together - Plant Physiol Biochem 8: 278-284.
Rohwer C.L., Erwin J.E. (2008) - Horticultural applications of jasmonates: a review - J Hortic
Sci Biotech 83: 283-304.
Royal Society (2008) - Ground-level ozone in the 21st century: future trends, impacts and
policy implication - Science Policy, Report 15/8.
Roychoudhury A.,Saikat P., Basu S. (2013) - Cross-talk between abscisic acid-dependent and
abscisic acid-independent pathways during abiotic stress – Plant Cell Reports 32:7: 985-1006.
Rumukainen M. (2012) - Changes in climate and weather extremes in the 21st Century - Wiley
Interdisciplinary reviews: Climate Change 3: 115- 119.
Santino A., Taurino M., De Domenico S., Bonsegna S., Poltronieri P., Pastor V., Flors V.
(2013) - Jasmonate signaling in plant development and defense response to multiple (a)biotic
stress - Plant Cell Rep 32: 1085-1098.
Short E.F., North K.A., Roberts M.R., Hetherington A.M., Shirras A.D., McAinsh M.R.
(2012) - A stress-specific calcium signature regulating an ozone-responsive gene expression
network in Arabidopsis - Plant J 71: 948-961.
Singh S., Kaur D., Agrawal S.B., Agrawal M. (2010) - Responses of two cultivars of
Trifolium repens L. to ethylene diurea in relation to ambient ozone - Journal of Environmental
Sciences 22: 1096-1103.
Song Y., Miao Y., Song C.P. (2014) - Behind the scenes: the roles of reactive oxygen species
in guard cells - New Phytol 201: 1121-1140.

55
Sorkheha K., Shiran B., Khodambashi M., Rouhi V., Mosavei S., Sofo A. (2012) -
Exogenous proline alleviates the effects of H2O2-induced oxidative stress in wild almond
species - Russ. J. Plant Physiol., 59: 788–798.
Stich S., Cox P.M., Collins W.J., Huntingford C. (2007) - Indirect radiative forcing of climate
change through ozone effects on land-carbon sink - Nature 448: 791-794.
Szabados L., Savouré A. (2010) - Proline: a multifunctional amino acid - Trends Plant Sci 15:
89-97.
Tamaoki M. (2008) - The role of phytohormone signaling in ozone-induced cell death in plants
- Plant Signaling & Behavior 3:3: 166-174.
Tardieu F., Parent B., Simonneau T. (2010) - Control of leaf growth by abscisic acid:
hydraulic or non-hydraulic processes? - Plant Cell Environ 33: 636-647
Trivellini A., Ferrante A., Vernieri P., Serra G. (2011) - Effects of abscisic acid on ethylene
biosynthesis and perception in Hibiscus rosa-sinensis L. flower development - J Exp Bot 62:
5437-5452.
Tuominen H., Overmyer K., Keinänen M., Kollist H., Kangasjärvi J. (2004) - Mutual
antagonism of ethylene and jasmonic aid regulates ozone-induced spreading cell death in
Arabidopsis - Plant J 39: 59-69.
Ueda Y., Uehara N., Sasaki H., Kobayashi K., Yamakawa T. (2013) - Impacts of acute
ozone stress on superoxide dismutase (SOD) expression and reactive oxygen species (ROS)
formation in rice leaves - Plant Physiology and Biochemistry 70: 396-402.
UNECE (2004) - Revised manual on methodologies and criteria for mapping critical
levels/loads and geographical areas where they are exceeded - www.icpmapping.org (February
12, 2006).
Vahala J., Ruonala R., Keinänen M., Tuominen H., Kangasjärvi J. (2003) - Ethylene
insensitivity modulates ozone-induced cell death in birch – Plant Physiol. 132: 95-185.
Vahisalu T., Puzorjova I., Brosché M., Valg E., Lepiku M., Moldau H., Kollist H. (2010)
– Ozone-triggered rapid stomatal response involves the production of reactive oxygen species,
and is controlled by SLAC1 and OST1 – The Plant Journal 62: 442-453.
Vainonen J.P., Kangasjärvi J. (2015) – Plant signaling in acute ozone exposure – Plant, Cell
and Environment 38: 240-252.
van Loon L.C., Geraats B.P.J., Linthorst H.J.M. (2006) - Ethylene as a modulator of disease
resistance in plants - Trends Plant Sci 11: 184-191.
Vaultier N.V., Jolivet Y. (2015) - Ozone sensing and early signaling in plants: An outline from
the cloud - Environ Exp Bot 114: 144-152.
Vernieri P., Ferrante A., Mugnai S., Paradossi A. (2006) - Stress abiotici e bilancio ormonale
nelle piante coltivate - Italus Hortus 13 (1): 19-31.
Vernieri P., Perata P., Armellini D., Bugnoli M., Presentini R., Lorenzi R., Ceccarelli N.,
Alpi A., Tognoni F. (1989) - Solid phase radioimmunoassay for the quantitation of abscisic
acid in plant crude extracts using a new monoclonal antibody - Journal Plant Physiology 134:
441-446.
Vinocur B., Altman A. (2005) - Recent advances in engineering plant tolerance to abiotic
stress: achievements and limitations - Current Opinion in Biotechnology 16:123– 132.
Vitale M., Mancini M., Matteucci G., Francesconi F., Valenti R., Attorre F. (2012) -
Model-based assessment of ecological adaptations of three forest tree species growing in Italy
and impact on carbon and water balance at National scale under current and future climate
scenarios – iForest - Bioscience and Forestry 5: 235- 246.

56
Volz A., Kley D. (1988) - Evaluation of the montsouris series of ozone measurements made in
the 19th-century - Nature 332: 240-242.
Walker-Simmons M. (1987) - ABA levels and sensitivity in developing wheat embryos of
sprouting resistant and susceptible cultivars - Plant Physiology, 84: 61–66.
Wang B., Vincour A., Altman A. (2003) - Plant responses to drought, salinity and extreme
temperatures: towards genetic engineering for stress tolerance - Planta 218: 1-14.
Wang K.L., Li H., Ecker J.R. (2002) - Ethylene biosynthesis and signaling networks - Plant
Cell 14: 131-151.
Wang Y., Wen T., Hu J., Han R., Zhu Y., Guan Y., Zhu S. (2013) – Relationship between
endogenous salicylic acid and antioxidant enzyme activities in maize seedlings under chilling
stress – Expl Agric. 49:2: 295-308.
Wieser G., Matyssek R. (2007) - Linking Ozone Uptake and Defense towards a Mechanistic
Risk Assessment for Forest Trees - New Phytologist 174: 7-9.
Wildermuth M.C., Dewdney .J, Wu G., Ausubel F.M. (2001) - Isochorismate synthase is
required to synthesize salicylic acid for plant defense - Nature 414: 562-565.
Wilkinson S., Davies W.J. (2010) - Drought, ozone, ABA and ethylene: new insights from cell
to plant to community - Plant Cell Environ 33: 510-525.
Wilkinson S., Mills G., Illidge R., Davies W.J. (2011) - How is ozone pollution reducing our
food supply? - J Exp Bot, 1-10. doi:10.1093/jxb/err317.
Wrzaczek M., Brosché M., Salojärvi J., Kangasjärvi S., Idänheimo N., Mersmann S.,
Robatzek S., Karpiński S., Karpińska B., Kangasjärvi J. (2010) - Transcriptional regulation
of the CRK/DUF26 group of Receptor-like protein kinases by ozone and plant hormones in
Arabidopsis - Plant Biol 10: 95-114.
Xia X-J., Zhou Y-H., Shi K., Zhou J. Foyer C.H., Yu J.Q. (2015) - Interplay between oxygen
species and hormones in the control plant development and stress tolerance - J Exp Bot 66:
2839-2856.
Yaeno T., Saito B., Katsuki T., Iba K. (2006) - Ozone-induced expression of the Arabidopsis
FAD7 gene requires salicylic acid, but not NPR1 and SID2 - Plant Cell Physiol 47: 355-362.
Yoo S-D., Cho Y-H., Tena G., Xiong Y., Sheen J. (2008) Dual control of nuclear EIN3 by
bifurcate MAPK cascades in C2H4 signalling - Nature 451: 789-795.
Zhao B., Summers F.A, Mason R.P. (2012) – Photooxidation of Amplex red to resorufin:
Implications of exposing the Amplex red assay to light - Free Radical Biology and Medicine
53: 1080-1087.
Zheng Y., Cheng D., Simmons M. (2014) - Ozone pollution effects on gas exchange, growth
and biomass yield of salinity-treated winter wheat cultivars - Sci Total Environ 499: 18-26.
Zhou C., Cai Z., Guo Y., Gan S. (2009) - An Arabidopsis mitogen-activated protein kinase
cascade, MKK9–MPK6, plays a role in leaf senescence - Plant Physiol 150: 167-177.





























