Embed Size (px)
Citation preview

Cell Tiss. Res. 152, 513--523 (1974) �9 by Springer-Verlag 1974
Qualitative and Quantitative Studies on the Innervation of the Corpus luteum of Rat and Pig*
K. Unsicker * *
Department of Anatomy, University of Kiel (Head: Prof. Dr. Drs. h. c. W. Bargmann)
Received July 19, 1974
Summary. Qualitative and quantitative studies were made to determine the amount of nerve fiber supplying corpora lutea (CL) of rats during the oestrous cycle and preg- nancy and sow CL during days 4-6 after ovulation. Fluorescence microscopy of freeze-dried, paraformaldehyde treated (Falck-Hillarp method) rat ovaries reveals adrenergic nerve fibers which run along with vessels and form a network among interstitial gland cells. Nerve fibers do not enter the granulosa cell layer in follicles or CL. In the CL circumference both vascular and non-vascular nerves occur the latter being related to the fibromuscular layer and probably innervating smooth muscle cells. No striking differences exist between the innervation of the ovary in non-pregnant and pregnant rats. Bodian and methylene blue staining did not contribute to a more detailed knowledge of rat ovary nerve supply. Electron microscopic quantitative analysis of rat and pig CL (rat: day 18 of pregnancy;pig: day 4-6 after ovulation) revealed no axon profiles in 2.000 grid squares (one square measuring 2.25 • 10 -3 mm 2) of randomly taken CL sections. Thus it was possible to calculate an upper limit of 133 ~m of nerve fibers per 1 mm a CL tissue, in case there were any at all.
Key words: Ovary - - Corpus luteum - - Innervation - - Fluorescence histochemistry - - Electron microscopy - - Quantitative study.
Introduction
Corpora lutea (CL), of mammals at least, are generally thought to be non- inner- vated. This assumpt ion is not based on precise studies, ra ther it represents the common experience of endocrinologists as well as of electron microscopists who have never reported the occurrence of nerve fibers in the ma mma l i a n CL. On the other hand, morphologists using silver impregnat ion methods have described an abundance of what they believed to be nerve fibers in CL of various mammal i an
species (cf. StShr jr., 1957). This apparen t contradict ion has prompted the following a t t empt to ob ta in
objective results on CL innerva t ion by means of qual i ta t ive and quan t i t a t ive methods which have not yet been applied to this problem. This a t t e mp t seems to be even more justified since several endocrine glands formerly thought to have no nerve supply have recent ly been shown to contain varying quant i t ies of autonomic fibers (Bargmann, 1973). Even peripheral neuroendocrine contacts have been demonst ra ted on the u l t ras t ruc tura l level, e.g. in the adrenal cortex
Send o//print requests to: Dr. Klaus Unsicker, Anatomisches Institut der Universit~t, D-2300 Kiel, Neuc Universit~t, 01shausenstraBe 40/60, Federal Republic of Germany.
*Supported by a grant from the ,,Deutsche Forschungsgemeinschaft" (Un 34/1). **I am much obliged to Mm. K. Jacob, Mrs. A. L5we, and Mrs. R. Sprang for valuable technical assistance.

514 K. Unsicker
(Unsicker , 1969, 1971), t h e i n t e r r ena l g l and of birds (Unsicker , 1973b) and rept i les
(Unsicker , 1974), in t h e o v a r i a n in t e r s t i t i a l g l and of t he mouse (Unsicker , 1970)
and t h e d o m e s t i c fowl (Dahl , 1970), in t h e t c s t i cu la r i n t e r s t i t i a l t i ssue (cf. Un-
sicker, 1973a, for ref.), and in va r i ous non- s t e ro id secre t ing endoc r ine g lands .
The a c t u a l i t y of t he p r e sen t s t u d y is unde r l i ned b y a pe rsona l c o m m u n i c a t i o n
f r o m O w m a n a n d Svensson (1974a) who h a v e obse rved ad rene rg ic ne rves in a CL of a ca t in l a te p r egnancy .
Mater ia l s and Methods
A. Qualitative Studies
The ovaries of pregnant and non-pregnant rats (strain Wistar/ttannover), 3 4 months of age, and one sexually mature, non-pregnant sow (mini-pig, strain GSttingen), weighing approximately 50 kg, 71/2 months of age, were studied by light, fluorescence, and electron microscopic methods.
1. Fluorescence Histochemistry. 30 rats were bred and the day of copulation was designated day O. On days 1, 5, 10, 14, 16, 20 of pregnancy and day 1 after parturition ovaries were re- moved under ether anesthesia. 6 non-pregnant rats in various stages of the oestrous cycle served as controls. Most animals received 100 mg/kg nialamide (Niamid| Pfizer 1) i.p. 3-6 hours before killing. The left ovary of each rat was immediately frozen in liquid propane cooled by liquid nitrogen. The specimens were freeze-dried according to the method of Falck and Hillarp (for technical details cf. Falck, et al., 1962; Fuxe et al., 1970; Bj5rklund et al., 1972). The formaldehyde treatment was performed at --80~ for 1 hr with paraformaldehyde equili- brated in air at 50% or 70% relative humidity. The specimens were embedded in paraffin in vacuo and sectioned (8-15 ~m thickness). Sections were mounted in liquid paraffin or En- tellan | (Merck) and observed through a Zeiss fluorescence microscope with BG 12 (Schott) as lamp and Zeiss 47 + 50 as secondary barrier filters. Autofluorescent structures were identified by excluding the formaldehyde step from the procedure.
2. Bodian Silver Technique. Ovaries of two pregnant rats (day 16 and day 17) were fixed using a mixture of formalin, acetone, and acetic acid, and stained according to Bodian (double impregnation with 1% protargol).
3. Methylene Blue Staining. Three rats (16th, 17th, and 18th day of pregnancy) were per- fused via the heart with a solution containing I g methylene blue, 2 g glucose, 0.3 g Na- pyruvate, 1.5 g magnesiumbromide, 8.0 g NaC1, and 1.000 ml H20. After the perfusion the ovaries were removed and stored on cotton-wool impregnated with the same solution in a warm cupboard at 37~ for 3-6 hrs. Finally the organs were fixed in an 8% solution of ammonium- molybdate and sectioned (50 ~zm thickness).
4. Electron Microscopy. Two pregnant rats (day 18) and one non-pregnant sow were per- fused via the descending aorta or the heart, respectively, with isotonic Macrodex| (+37~ pH 7.7) in Nembutal| anesthesia for 10-20 sec. Then perfusion was continued with 3.5% glutaraldehydc in phosphate buffer (ice-cold, pH 7.7) and phosphate buffer. Slices of ovaries, 1 mm thick, were postfixed in unbuffered 2 % aqueous OsO 4 for 2 hrs, dehydrated with ethanol, and embedded in Araldite. Thin sections of CL were mounted on naked grids and stained with uranyl acetate and lead citrate for 5 min each.
Electron microscopes: Zeiss EM 9A and Siemens 101. The approximate age of the sow CL was determined on the basis of thick toluidine blue
stained sections and Araldite thin sections according to Corner (1921) and Bjersing (1967).
B. Quantitative Studies
CL of one pregnant rat (day 18) and one non-pregnant sow (4 6 days after ovulation) were chosen for quantitative analysis. Random sections were made from Araldite blocks the minimal distance of two consecutive sections being 0.3 ram. The total area of grid squares, one square measuring 2.25 • 10 2 mm 2, was 2.000 per gland and species.
1 Niamid| was kindly supplied by Pfizer Corp., U.S.A.

Innervation of Rat and Pig Corpus luteum 515
Considerations on Problems o/Quantification o/Nerve Fibers. The density of innervation is defined as the sum of the length L of nerves per total tissue volume examined:
L L v - V" (1)
~f nerve fibers may be regarded as equally distributed linear structures, the density of inner- ration can be calculated according to an equation given by Hennig (1963):
2n Lv = ~ - (2)
(n: number of axon profiles observed in a sample area of size F). Since the possibility that no axons will be found has to be included in our considerations,
we must know something about the probability of detecting no axon profile in q grid squares, though there is a definite number of axons. This probability is expressed by the equation
W = 2-q )' (3)
provided that the number of axon profiles per sample area follows the Poisson distribution (),: expected number of axon profiles per sample area).
Given the probability r162 for finding no axons although there is an innervation, it is possible to calculate an upper limit ~0 for 2. 20 is that expected number of axons crossing a grid square for which the probability of observing no axon profile in q sqares is exactly a. Hence
In �9 ~o = - - (4) q
By substituting (4) into (2) we obtain an upper limit for L:
2 ln~ L~ax -- F q
Results
Rat
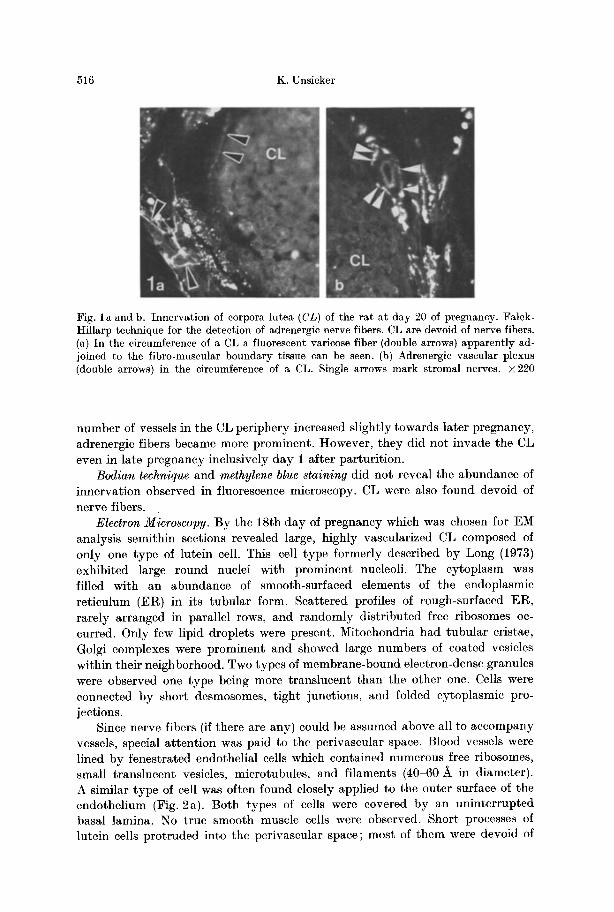
Fluorescence Histochemistry. Green fluorescent adrenergic nerve fibers were found enter ing the ovary by perivascular plexus in the hilar region. The major i ty of the fibers followed the course of the vessels; a considerable minori ty, however, was not adjoined to vessels, bu t formed a network among interst i t ial gland cells. Follicles, especially developing and atretic ones, received a dense innerva t ion in both the theca in t e rns and externa. Fluorescent nerve fibers did not penetrate the basement membrane of the s t r a tum granulosum. CL were never seen to be innerva ted (Fig. 1). Lute in cells contained autofluorescent t iny particles, p robably corresponding to lipofuscin-like granules. I nne rva t ion of the CL circumference was of dual na tu re (Fig. 1): first, adrenergic vascular plexus with their typical a r rangement of fibers (Fig. l b) surrounded arterial vessels obviously supplying CL. The inne rva t ion of those vessels stopped abrup t ly when they branched into smaller ones invading the parenchyma of CL. Second, varicose fibers ran in tangent ia l directions along the surface of CL (Fig. 1 a). They were contiguous with a f ibromuscular layer surrounding CL and independent from vessels. No nerve fibers existed in the circumference of CL extending towards the surface of the ovary. Only minor differences were observed when the innerva t ion of the ovary in pregnant rats was compared with tha t in non-p regnan t rats. Since the
516 K. Unsicker
ij e
cL "
Fig. la and b. Innervation of corpora lutea (CL) of the rat at day 20 of pregnancy. Falck- Hillarp technique for the detection of adrenergic nerve fibers. CL are devoid of nerve fibers. (a) In the circumference of a CL a fluorescent varicose fiber (double arrows) apparently ad- joined to the fibro-muscular boundary tissue can be seen. (b) Adrenergic vascular plexus (double arrows) in the circumference of a CL. Single arrows mark stromal nerves. • 220
number of vessels in the CL periphery increased slightly towards later pregnancy, adrenergic fibers became more prominent. However, they did not invade the CL even in late pregnancy inclusively day 1 after parturit ion.
Bodian technique and methylene blue staining did not reveal the abundance of innervation observed in fluorescence microscopy. CL were also found devoid o2 nerve fibers.
Electron Microscopy. By the 18th day of pregnancy which was chosen for EM analysis semithin sections revealed large, highly vascularized CL composed of only one type of lutein cell. This cell type formerly described by Long (1973) exhibited large round nuclei with prominent nucleoli. The cytoplasm was filled with an abundance of smooth-surfaced elements of the endoplasmic reticulum (ER) in its tubular form. Scattered profiles of rough-surfaced ER, rarely arranged in parallel rows, and randomly distributed free ribosomes oc- curred. Only few lipid droplets were present. Mitochondria had tubular cristae, Golgi complexes were prominent and showed large numbers of coated vesicles within their neighborhood. Two types of membrane-bound electron-dense granules were observed one type being more translucent than the other one. Cells were connected by short desmosomes, t ight junctions, and folded cytoplasmic pro- jections.
Since nerve fibers (if there are any) could be assumed above all to accompany vessels, special a t tent ion was paid to the perivascular space. Blood vessels were lined by fenestrated endothelial cells which contained numerous free ribosomes, small t ranslucent vesicles, microtubules, and filaments (40-60 A in diameter). A similar type of cell was often found closely applied to the outer surface of the endothelium (Fig. 2a). Both types of cells were covered by an uninterrupted basal lamina. No true smooth muscle cells were observed. Short processes of lutein cells protruded into the perivascular space; most of them were devoid of
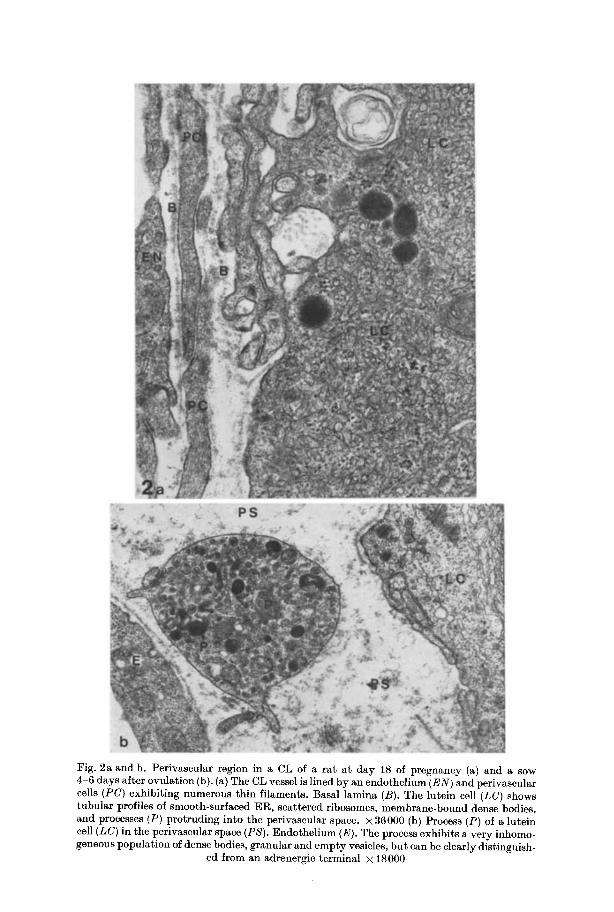
Fig. 2a and b. Perivascular region in a CL of a ra t a t day 18 of pregnancy (a) and a sow 4-6 days after ovulation (b). (a) The CL vessel is lined by an cndothclium (EN) and perivascular cells (PC) exhibit ing numerous th in filaments. Basal lamina (B). The lutein cell (LC) shows tubular profiles of smooth-surfaced ER, scat tered ribosomes, membrane-bound dense bodies, and processes (P) protruding into the perivascular space. • 36000 (b) Process (P) of a lutein cell (LC) in the perivaseular space (PS). Endothel ium (E). The process exhibits a very inhomo- geneous populat ion of dense bodies, granular and empty vesicles, bu t can be clearly distinguish-
ed from an adrenergic terminal • 18000
518 K. Unsickcr
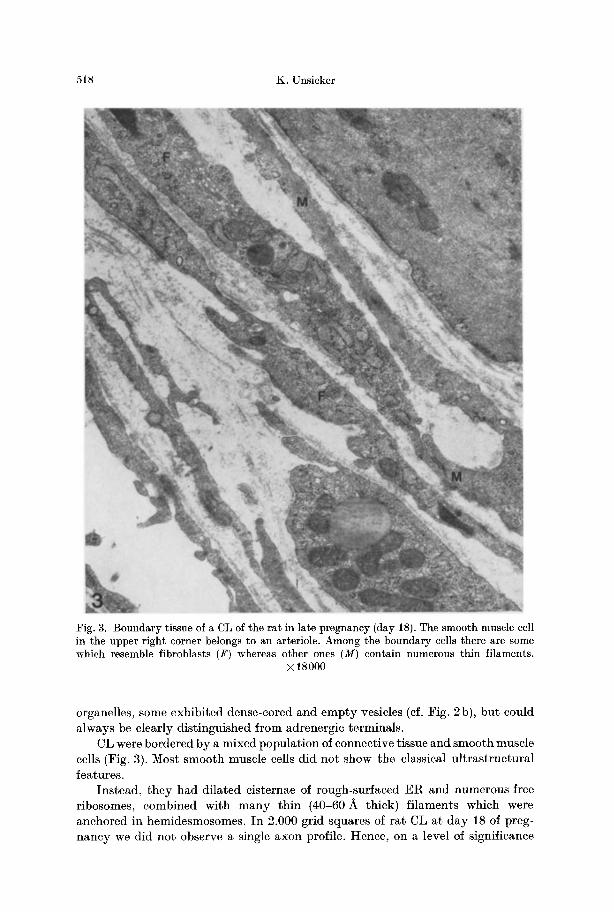
Fig. 3. Boundary tissue of a CL of the rat in late pregnancy (day 18). The smooth muscle cell in the upper right corner belongs to an arteriole. Among the boundary cells there are some which resemble fibroblasts (F) whereas other ones (M) contain numerous thin filaments.
• 18000
organelles, some exhibited dense-cored and empty vesicles (eft Fig. 2b), bu t could always be clearly dist inguished from adrenergic terminals.
CL were bordered by a mixed populat ion of connective tissue and smooth muscle cells (Fig. 3). Most smooth muscle cells did not show- the classical u l t ras t ructura l features.
Instead, they had dilated cisternae of rough-surfaced E R and numerous free ribosomes, combined with m a n y th in (40-60 A thick) f i laments which were anchored in hemidesmosomes. I n 2.000 grid squares of rat CL at day 18 of preg- nancy we did no t observe a single axon profile. Hence, on a level of significance

Innervation of Rat and Pig Corpus luteum 519
of 0.95 (a ~ 0.05) we get L~ ax = 0.133 ram, i.e. if there were any nerve fibers in CL, they could not have more than a maximum length of 133 fxm per 1 mm a tissue.
Pig
Electron Microscopy. Two types of endocrine cells could be distinguished well in sow CL. Type I exhibited the classical ultrastructural features of a steroid secreting cell and corresponded to the granulosa lutein cell 4-6 days after ovulation described by Bjersing (1967). The smooth-surfaced ER consisted of whorl-like figures with centrally located lipid droplets and diffusely distributed tubular profiles. Mito- chondria showed tubular cristae and were sometimes arranged along the long axis of granulosa lutein cells. Membrane-bound dense bodies which occasionally exhibited a finger-print-like inner structure were associated with Golgi complexes, but could also be found in close apposition to the cell membrane. Type I I was a small fusiform or stellate cell lying along blood vessels. This cell contained granular and agranular ER in approximately equal parts, mitoehondria with tubular and lamellar cristae, prominent Golgi complexes, and opaque lipid droplets in various numbers. Blood vessels inside the sow CL resembled those in rat CL; no smooth muscle cells were observed. No axon profiles were detected inside the sow CL. Sometimes processes of endo- crine cells exhibiting a very inhomogeneous population of dense bodies, round and elongated clear and dense-cored vesicles extended into the perivascular space (Fig. 2b), but their non-nervous character could always be established without a doubt. In 2.000 grid squares no nerve fibers were found. Thus we can say that there were--if any - -no t more than 133 ~m of nerves per 1 mm 3 CL tissue.
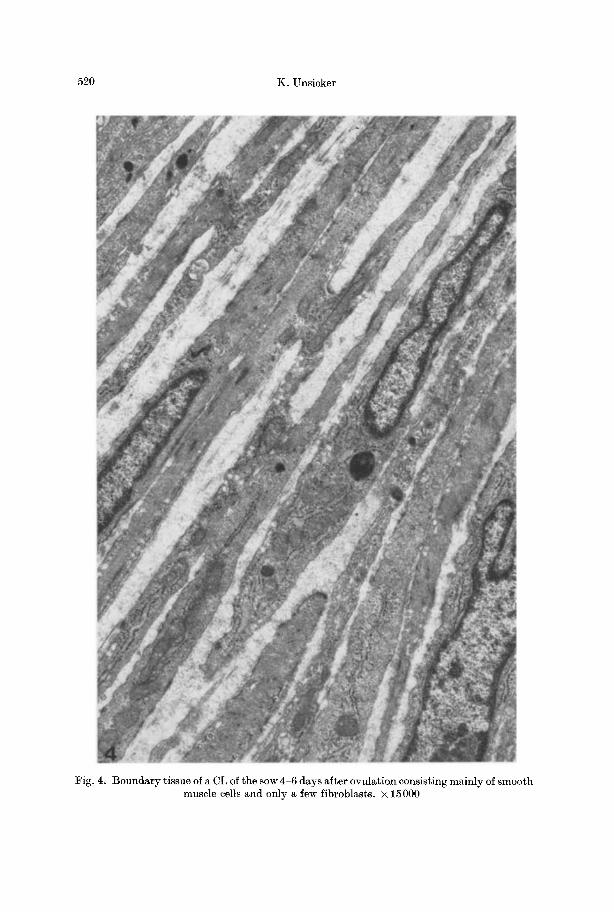
In the fibromuscular boundary tissue of the sow CL true smooth muscle cells preponderated (Fig. 4), but no nerve fibers were seen within this cell layer.
Discussion
The present study is the first to give highly probable evidence for the non-inner- ra t ion of CL in two mammalian species suggesting that hormonal control alone plays a role in the regulation of growth, secretory activity, and regression of CL. This evidence is based on combined qualitative and quantitative investigations applying several methods which are in use for the detection of nerve fibers.
Fluorescence histochemical results confirm the pattern of innervation in the non-pregnant rat ovary described by Burden (1972) who showed that adrenergic nerves do not penetrate the basement membrane surrounding granulosa cells in follicles and granulosa lutein cells in CL.
I t deserves mention, however, that the preparation procedure used by Burden (1972) and by us in the present study followed the classical methodological concept of Falck and Hillarp. Lindvall (cf. BjSrklund etal., 1972; Axelsson etal., 1973) has recently developed an improved method by introducing a glyoxylic acid t reatment instead of the formaldehyde step, and Owman, Svensson, and co-workers (1974b) have shown that the amount of fluorescent fiber in vibratome sections (el. Lindvall et al., 1973) of ovaries treated with glyoxylic acid is much higher
520 K. Unsicker
Fig. 4. Boundary tissue of a CL of the sow 4-6 days after ovulation consisting mainly of smooth muscle cells and only a few fibroblasts. • 15000

Innervation of Rat and Pig Corpus luteum 521
than in formaldehyde treated material. Our electron microscopic results, however, show that it is not necessary to use the glyoxylic acid method with regard to the problem of CL innervation in the rat. Supplementary to former investigations we can say that adrenergic nerve fibers in the circumference of rat CL innervate both blood vessels and the fibromuscular tissue. The latter probably forms some kind of theca externa which has been found to be innervated in several mam- malian species (cf. Bahr et al., 1974, for rev.). The significance of this ceil layer for the function of CL remains to be elucidated, but it cannot be excluded that adrenergic fibers in this layer are more or less vestigial structures.
No great differences could be observed in the adrenergic innervation when ov aries during the oestrous cycle were compared with those during pregnancy, though the number of adrenergically innervated blood vessels in the circumference of CL showed a moderate increase during pregnancy. This observation is in ac- cordance with that of Rosengren and SjSberg (1968) who found that the density of innervation by adrenergic fibers in the rabbit ovary was essentially unchanged during pregnancy, whereas chemical determinations showed an increase in content of transmitter in the oviduct at the same time. These differences in ovary and oviduct transmitter content can probably be explained by functional distinctions between short and long adrenergic neurons which make up the non-vascular adrenergic nerve supply in oviduct and ovary, respectively.
On the basis of our quantitative electron microscopic studies we can say with a very high degree of probability that CL in the pregnant rat at day 18 and CL in the sow 4-6 days after ovulation are not innervated. The possible existence of 133 ~m nerve fibers per 1 mm a tissue which cannot be totally excluded is of theoretical significance only. Since lutein cells have diameters up to 35 ,~m, such a scarce innervation would mean that the only axon of such a CL would not reach deeper into the gland than 4-5 lutein cell layers.
CL of pregnancy may be regarded as relatively stable structures compared to those during the oestrous cycle. If they are not innervated, there are only very few good reasons--if any- - to assume a nerve supply in menstrual CL. On the other hand, we cannot conclude from the apparent non-innervation of the sow menstrual CL 4-6 days after ovulation that CL of the same species receive no nerve supply during pregnancy.
The CL of the sow was chosen for the present study because it has two classes of luteal cells (Corner, 1915, 1919), the large polyhedral "granulosa" luteiu cell and the stellate cell which apparently differ in their ultrastructural aspects. The latter most likely arise from stromal cells which invade the early granulosa luteal mass along with the vessels (Mossman and Duke, 1973). The possibility that they could draw nerve fibers with them was estimated to be rather high, but apparently they do not.
From the present study we cannot draw the conclusion that the mammalian CL in general is not innervated. Perhaps this could be assumed with a certain degree of probability for those CL which are composed of "granulosa" lutein cells only. O'Donoghue (1963) has claimed that the luteal cells of the tree hyrax Dendro- hyrax arboreus are derived exclusively from theca interna cells. In case his view turns out to be right an innervated mammalian CL may be found, though Kayanja and Naeves (1973) in their report on the fine structure of the CL in two species

522 K. Unsicker
of rock hyrax (Procavia habessinica and Heterohyrax brucei) do not ment ion any nerve fibers. The chance of f inding nerve fibers in CL will probably be opt imal in very late stages of CL development. Accordingly, the observat ion of adrenergic nerves in a CL of a cat during late pregnancy (Owman and Svensson, 1974a) deserves an u l t ras t ruc tura l re-investigation.
References
Axelsson, S., Bj6rklund, A., Falck, B., Lindvall, O., Svensson, L.-A.: Glyoxylic acid con- densation: a new fluorescence method for the histochemical demonstration of biogenic monoamines. Aeta physiol, scand. 87, 57~62 (1973)
Bahr, J., Kao, L., Nalbandov, A. V.: The role of catecholamines and nerves in ovulation. Biol. Reprod. 10, 273-290 (1974)
Bargmann, W.: On the innervation of vertebrate endocrine organs. In: Endocrinology (ed. O. Scow), Proc. IV. Congress Endocrinol. Washington 1972, p. 220-223. Amsterdam: Excerpta Medica; New York: American Elsevier Publ. Co. 1973
Bjersing, L.: On the ultrastructure of granulosa lutein cells in porcine corpus luteum. With special reference to endoplasmic reticulum and steroid hormone synthesis. Z. Zellforsch. 82, 187-211 (1967)
BjSrklund, A., Falck, B., Owman, Ch.: Fluorescence microscopic and microspectrofluoro- metric techniques in the cellular localization and characterization of biogenic amines. In: S. A. Berson (ed.), Methods of investigative and diagnostic endocrinology, vol. 1: J. E. Rall and I. J. Kopin (eds.), The thyroid and hiogenic amines, p. 318-368. Amsterdam: North Holland Publ. Co. 1972
BjSrklund, A., Lindvall, 0., Svensson, L.-A.: Mechanisms of fluorophore formation in the histochemical glyoxylic acid method for monoamines. Histochemie 82, 113-131 (1972)
Burden, H. W. : Adrenergic innervation in ovaries of the rat and guinea pig. Amer. J. Anat. 133, 455-462 (1972)
Corner, G. W.: The corpus luteum of pregnancy, as it is in swine. Contr. Embryol. Carneg. Inst. 222, 69-94
Corner, G. W.: On the origin of the corpus luteum of the sow from both grauulosa and theca interna Amer. J. Anat. 26, 117 183 (1919)
Corner, G. W.: Cyclic changes in the ovaries and uterus of the sow, and their relation to the mechanism of implantation. Contr. Embryol. Carneg. Inst. 13, 117-146 (1921)
Dahl, E.: Studies on the fine structure of ovarian interstitial tissue. 3. The innervation of the thecal gland of the domestic fowl. Z. Zellforsch. 109, 212-226 (1970)
Falck, B., Hillarp, N.-/~., Thieme, G., Torp, A.: Fluorescence of eatechol amines and related compounds condensed with formaldehyde. J. Histochem. Cytochem. 10, 348-354 (1962)
Fuxe, K., HSkfelt, T., Jonsson, G., Ungerstedt, U.: Fluorescence microscopy in neuroanatomy. In: S. J. Nauta and S.O.E. Ebbesson (eds.), Contemporary research methods in neuroana- tomy, p. 275-314. Berlin-Heidelberg-New York: Springer 1970
Hennig, A.: Li~nge eines dreidimensionalen Linienzuges. Proc. I. Internat. Congr. Stereology 44, 1-8 (1963)
Kayanja, F.I.B., Naeves, W.B.: The fine structure of the corpus luteum in Hyrax. Z. Zell- forsch. 144, 475-487 (1973)
Lindvall, O., BjSrklund, A., H5kfelt, T., Ljungdahl, A.: Application of the glyoxylic acid method to vibratome sections for the improved visualization of central catecholamine neurons. Histochemie 35, 31 38 (1973)
Long, J. A.: Corpus luteum of pregnancy in the rat--ultrastructural and cytochemical obser- vations. Biol. Reprod. 8, 87-99 (1973)
Mossman, H.W., Duke, K.L.: Comparative morphology of the mammalian ovary. The University of Wisconsin Press, 1973
O'Donoghue, P. N.: Reproduction in the female hyrax (Dendrohyrax arborea ruwenzorii). Proc. zool. Soc. London 141, 207 237 (1963)
Owman, Ch., Svensson, K.-G.: Personal communication (1974a)

Innervation of Rat and Pig Corpus luteum 523
Owman, Ch., Svensson, K.-G.: Personal communication (1974b) Rosengren, E., SjSberg, N. O.: Changes in the amount of adrenergic transmitter in the fema|e
genital tract of the rabbit during pregnancy. Acta physiol, scand. 72, 412-424 (1968) StShr jr., Ph.: Mikroskopische Anatomie des vegetativen Nervensystems. In: Handbuch der
mikroskopischen Anatomie des Menschen, Bd. IV, Teil 5. Berlin-GSttingen-Heidelberg: Springer 1957
Unsicker, K.: Zur Innervation der Nebennierenrinde vom Goldhamster. Eine fluoreszenz- und elektronenmikroskopische Studie. Z. Zellforsch. 95, 608-619 (1969)
Unsicker, K.: Zur Innervation der interstitiellen Drfise im Ovar der Maus (Mus musculus L.). Z. Zellforsch. 109, 46-54 (1970)
Unsicker, K.: On the innervation of the rat and pig adrenal cortex. Z. Zellforsch. 115, 151-156 (1971)
Unsicker, K.: Innervation of the testicular interstitial tissue in reptiles. Z. Zellforsch. 146, 123-138 (19739)
Unsickcr, K.: Fine structure and innervation of the avian adrenal gland. V. Innervation of interrenal cells. Z. Zcllforsch. 146, 403-416 (1973b)
Unsicker, K.: Innervation of adrenal ceils in the lizards Lacerta dugesi und Lacerta pityusensis. Gen. comp. Endocr. (in press)
34 Cell Tiss. Res., Vol. 152





























