Embed Size (px)
Citation preview

BRAIN RESEARCH 305
STEADY POTENTIAL CORRELATES OF POSITIVE REIN F O RCEMEN T AND SLEEP ONSET IN THE CAT; 'REWARD CO N TIN G EN T POSITIVE VARIATION' (RCPV)
T. J. MARCZYNSKI, J. L. YORK, S. L. ALLEN, J. H. RICK AND C. J. SHERRY
Department of Pharmacology, University of Illinois at the Medical Center, Chicago, Ill. 60612 (u .s .A.)
(Accepted September 15th, 1970)
INTRODUCTION
Epicortical steady potential (SP) shifts occur during electrical stimulation of the brain stem reticular formation or non-specific thalamic nuclei% 16, sensory stimula- tion 19,2°,29, and they also closely reflect shifts in the sleep-wakefulness spec- trum 5,6,27,42,67. They have been shown to gradually develop in response to conditioned stimuli reinforced aversivelyV,3a,56, as or positively 34 as well as after the establishment of a conditional reflex to electrical stimulation of the midline thalamic nuclei4L Con- ditional auditory or flash stimuli reinforced with food produce large SP shifts if pre- sented to a food-deprived subject34, 55. The magnitude of these SP shifts gradually diminishes as a function of volume of food eaten. It was therefore suggested that their magnitude reflects the level of drive and motivation25,26, 34,55.
A negative SP shift, maximal at the vertex, occurs in human subjects during performance of a variety of stimulus-response tasks26,33,34,6a, 64. In a typical experi- mental paradigm, pairs of stimuli were presented at random time intervals. The first stimulus (S-I) preceded the second one (S-2) by a fixed time interval (usually 1-2 sec), and a manual response was required to S-2. If the subject was not instructed to respond to S-2, the amplitude and duration of the negative SP shift in response to S-1 was negligible or absent, Since the occurrence of a fully developed negative shift was contingent upon the significance of an association between S-1 and S-2, it was termed 'contingent negative variation' by Walter et al. 64. Later, Walter ~8 termed this negative SP 'expectancy wave', since its magnitude was found to be a function of two factors: probability of occurrence of S-2 and 'preparation to respond'. Kornhuber and Deecke 3° also described negative SP shifts and termed it 'Bereitschaftspotential' (i.e.,
'readiness potential') occurring over the contralateral Rolandic cortex before a voluntary'movement, and they noted that its amplitude was related to intentional engagement on the part of the subject. Subsequent studies26,3~,34, 43,44,49,5° have shown that during the time interval between the 'warning' stimulus (S-I) and the 'imperative' one (S-2) the level of motivation to perform a required volitional act plays
Brain Research, 26 (1971) 305-332

306 T.J. MARCZYNSKI et al.
a crucial role in triggering the negative SP shift. Low et al. 33,34 have therefore suggest- ed that this SP should be renamed 'conative negative variation', since it reflects a specific mental state of conation defined as a conscious drive to perform a volitional act. Walter 63,64, Low et al. 33, and more recently Hillyard and Galambos 23 were able
either to rule out or to differentiate the 'artifactual ' influences generated by involun- tary eye movements, galvanic skin resistance and respiratory or cardiovascular corre- lates, thus demonstrating that the conditioned SP shifts are of cerebral origin.
In a food-deprived cat trained to press a lever for milk reward, the electro- corticogram (ECoG) over the visual cortex shows dramatic fluctuations from de- synchronized to highly synchronized patterns of high-voltage slow-wave activity (5-8 c/sec) after the presentation and during the consumption of reward. It was suggested that these bursts of alpha activity, termed post-reinforcement synchronization (PRS) by Sterman et al. 59 and Clemente et aL s, reflect an active process of Pavlovian internal inhibition and result from a transient but powerful depression of the reticular activating systemS,3s,41,51, 60 (see also Discussion). I t may be suggested that the de-
synchronized ECoG patterns observed during non-rewarded performance and prior to the rewarded lever press reflect high level motivation, a state of conation and expectancy; a delivery of reward, and particularly its consumption associated with a burst of PRS, may be related to a sudden and transient termination of this state.
The present study on SP was designed to shed more light on ECoG phenomena associated with positive reinforcement by investigating the time course and possible relationship between the PRS phenomena and SP shifts. The SP correlates of PRS were compared to those occurring during: (a) non-rewarded lever press performance, and (b) onset of sleep in satiated subjects. Since PRS activity is contingent on the visual input as well as on the appropriateness and desirability of food re- ward 4,s,ss,aa,42,6°, the effects of changes in the intensity of ambient light and in the
quality of food reward were also tested.
METHODS
Thirteen friendly adult cats (2.5-4 kg) of either sex, kept on a 23 h food and water deprivation schedule, were trained in a soundproof cage (provided with one-way window and light) to press a lever for 0.5-1.0 ml of milk reward. Delivery was programmed on a variable interval schedule on which the lever press produced the reward aperiodicaUy, an average once per 12-16 sec. The intensity of illumination of the test chamber was set at approximately 25 candles/sq.m. Nine animals were finally selected which served in 166 experimental sessions (each lasting 30-60 min) during which each subject was allowed to press the lever until satiation.
Under pentobarbital anesthesia, epidural Ag-AgC1 electrodes were implanted over the frontal and parieto-occipital cortex of both hemispheres. These electrodes were marked as filled circles on cortical diagrams. In most subjects two reference electrodes were implanted in the bone marrow of the skull: one over the frontal sinus and another in the occipital crest. In 4 subjects additional reference electrodes were implanted 3-4 m m beneath the cortical surface in the white matter subjacent to a
Brain Research, 26 (1971) 305-332

STEADY POTENTIALS-REINFORCEMENT 307
corresponding epicortical electrode in the posterior marginal, medial suprasylvian and anterior ectosylvian gyri for transcortical recording of SP shifts. These electrodes were marked as stars on cortical diagrams. In order to monitor directional changes in the corneo-retinal potential field, i.e., the electrooculogram (EOG), and to observe the relationship between the eye movements and the occurrence of lambda waves over the parieto-occipital cortex that could contaminate the SP responses recorded over the same cortical region during operant behavior, an Ag-AgC1 electrode was implanted in the frontal sinus in the bone behind the eyeball, and another over the posterior aspect of the bone overlaying the frontal sinus. Potentials caused by lapping or licking were recorded with a similar pair of electrodes placed at the bottom of the frontal sinus and in the temporal bone. In several experiments the latter electrodes were connected with a Grass AC amplifier using an electrostatically 'noisy' lead whose shield was disconnected from a 'common ground'. Such a lead usually portrayed both the tap- ping activity and head movements. Potentials caused by eye movements could be easily differentiated from those caused by head movements or lapping, since the former were characterized by a much higher amplitude, irregular occurrence and sharp peaks (Fig. 1, middle and bottom).
The Ag-AgC1 electrodes were formed by compressing AgC1 with pure Ag under vacuum. A 1 mm diameter pair, 1 cm apart in normal saline, showed the following characteristics: ZC 10 kc/sec, 1000 f2; phase angle,--10°; potential difference 1 mV; cumulative drift < 1 #V/min*. Another series of electrodes were made according to the method described by Bond and Ho3; these electrodes showed characteristics comparable to those just described. The electrodes were soaked for 48 h in Ringer solution prior to the implantation. In order to prevent their drying out after implanta- tion, a miniature strip of gauze was used to secure a connection between their tips and the adjacent bone marrow. They were connected to a miniature socket and fixed to the skull using dental cement. Low noise cable 61 was used to feed the signal into a 4- channel Grass polygraph equipped with model 5P1 low-level DC amplifiers. All recordings were made using 200,000 f2 input impedance. During 11 experimental sessions the SP shifts were integrated and quantified by feeding the signal from stage J5 and J6 of the driver amplifier into a 5U-2 integrator preamplifier coupled with a model UI-1 unit integrator. Its output was in turn fed into a Grass DC driver amplifier ~ and written out on a separate channel. The integration system was adjusted to produce 20 mrn pen deflection in response to 100/~V positive SP shift lasting I sec. This value was accepted as one unit of RCPV (see ref. 42). During the initial period of several~non-rewarded lever presses, i.e., during the maximum negative SP shifts, the baseline of the driver amplifier, whose output was fed into the DC integrator, was adjusted to provide an approximately zero output from the DC integrator.
Recording convention
In all drawings of the cat's cortex the surface epidural electrodes have been
* Several electrodes were generously supplied by Dr. H. W. Bond of Research Laboratories, Parke Davis and Company, Ann Arbor, Mich.
Brain Research, 26 (1971) 305-332
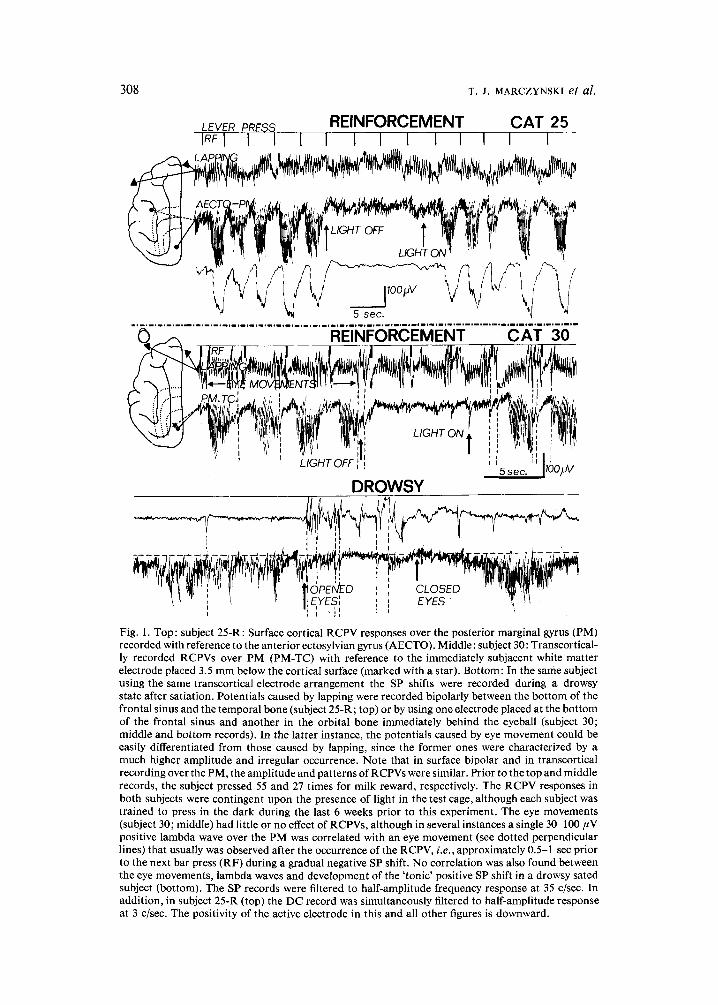
308 T. J. MARCZYNSKI et al.
LEVER PRESS REINFORCEMENT CAT 25 IRE I I I I I I I I I I I I I
a s e e . I ' v W -~v
DROWSY
Fig. 1. Top: subject 25-R: Surface cortical RCPV responses over the posterior marginal gyrus (PM) recorded with reference to the anterior ectosylvian gyrus (AECTO). Middle: subject 30: Transcortical- ly recorded RCPVs over PM (PM-TC) with reference to the immediately subjacent white matter electrode placed 3.5 mm below the cortical surface (marked with a star). Bottom: In the same subject using the same transcortical electrode arrangement the SP shifts were recorded during a drowsy state after satiation. Potentials caused by lapping were recorded bipolarly between the bot tom of the frontal sinus and the temporal bone (subject 25-R; top) or by using one electrode placed at the bot tom of the frontal sinus and another in the orbital bone immediately behind the eyeball (subject 30; middle and bottom records). In the latter instance, the potentials caused by eye movement could be easily differentiated from those caused by lapping, since the former ones were characterized by a much higher amplitude and irregular occurrence. Note that in surface bipolar and in transcortical recording over the PM, the amplitude and patterns of RCPVs were similar. Prior to the top and middle records, the subject pressed 55 and 27 times for milk reward, respectively. The RCPV responses in both subjects were contingent upon the presence of light in the test cage, although each subject was trained to press in the dark during the last 6 weeks prior to this experiment. The eye movements (subject 30; middle) had little or no effect of RCPVs, although in several instances a single 30-100 #V positive lambda wave over the PM was correlated with an eye movement (see dotted perpendicular lines) that usually was observed after the occurrence of the RCPV, i.e., approximately 0.5-1 sec prior to the next bar press (RF) during a gradual negative SP shift. No correlation was also found between the eye movements, lambda waves and development of the ' tonic ' positive SP shift in a drowsy sated subject (bottom). The SP records were filtered to half-amplitude frequency response at 35 c/sec. In addition, in subject 25-R (top) the DC record was simultaneously filtered to half-amplitude response at 3 c/sec. The positivity of the active electrode in this and all other figures is downward.

STEADY POTENTIALS--REINFORCEMENT 309
marked with filled dots; the white matter electrodes subjacent to the epidural ones (used in transcortical recording) have been marked with stars; in all recordings, dotted lines connect to the reference G-l, and the continuous lines to the 'active' G-2 elec- trode. The positivity of the latter with respect to G-1 is always down.
Control 'acute' experiments
(1) Contamination from the electroretinogram (ERG). The main purpose of this series of experiments was to test the reliability and the 'indifference' of the reference electrode located over the frontal sinus which was used in 'chronic' preparations. Six adult cats of either sex weighing 2.5-3.0 kg were used. Tracheotomy, cannulation of the cephalic vein and craniotomy were carried out under ether anesthesia. Ether administration was discontinued after infiltrating the scalp and pressure points with 2 ~o solution of Xylocaine. The animals were then immobilized with gallamine triethi- odide (Flaxedil) and artificially respired. It was necessary to avoid anesthetics since they affect the SP shifts1, 46. Ag-AgC1 electrodes (similar to those used in 'chronic' preparation) were placed epidurally over the posterior marginal gyrus and in the posterior aspect of the bone overlaying the frontal sinus, i.e., in the same location in which the reference electrodes were placed in the chronically implanted subjects. A third electrode was applied to the cornea to monitor the electroretinogram (ERG) with reference to frontal sinus, posterior marginal gyrus or occipital crest. After complet~ mydriasis was achieved by topical application of homatropine, the effect of switching on and off the light was tested on the ERG and SP shifts over the posterior marginal gyrus. A surgical lamp directed towards both eyes served as light source whose intensity was comparable to that of the illumination of the test chamber in which the behavioral experiments were performed.
(2) Contamination from the electrooculogram (EOG). In 3 cats anesthetized with Pentobarbital (35 mg/kg, i.p.) the influences of directional changes in the standing corneo-retinal potential associated with passive eye movements were explored for all epicortical and white matter electrode arrangements that closely corresponded to those used in freely moving subjects. A thread was attached to each cornea, and both eyes were simultaneously rotated by approximately 45 ° or 90 ° either in vertical or horizontal direction. The average peak-to-peak amplitude (with standard errors) of the EOG contamination for a particular electrode arrangement was based on at least 10 measurements.
RESULTS
Electroretinogram (ERG) and epicortical SP shifts
In the 'acute' anesthetized preparation the ERG in response to 'light on' or 'light off', recorded between the cornea and the reference placed over the posterior marginal gyrus, ranged from 350 to 400 #V (measured peak to peak). The electrode located over the bone of the frontal sinus picked up approximately 25-28 ~ of this voltage with
Brain Research, 26 (1971) 305-332

310 T.J. MARCZYNSKI et al.
reference to the posterior marginal gyrus. It was found that the following two pairs of epidural or white matter electrodes, oriented in rostro-caudal direction and covering a relatively long distance, may pick up the ERG of 70-100 ~V: (1) between the anterior ectosylvian and the posterior marginal gyri that corresponded to those used in the behavioral study shown in Fig. 1, top (channel AECTO-PM), Fig. 4 (channels 2 and 3), Fig. 5 (channel 2); and (2) another pair between the ectosylvian gyrus and the frontal sinus, as shown in Fig. 5 (channel WE-FS). No ERG contamination was found in transcortical recordings (Fig. 1, middle and bottom, channel PM-TC; Fig. 4, channel 1 ; Fig. 5, channels 1 and 3; Fig. 6, top and left bottom, channel marked *-4). These ERG influences have been substantially reduced to 20--40 #V or even abolished by a procedure subsequently adopted in all 'chronic' preparations. This procedure con- sisted of : (a) placing of the frontal sinus electrode over the most posterior aspect of the sinus; (b) making a wide transversal incision in the bone overlying the frontal sinus; (c) removing most of the mucose membrane; and (d) filling the sinus cavity with dental cement. In such 'acute' preparations relatively free from ERG influences, 'light on' usually caused a negative or positive-negative SP shift of 60-100/~V over the striate and parastriate cortex in recordings with reference to the occipital crest or subjacent white matter. Similar SP shifts were also sporadically observed in the chronic prepara- tion during lever pressing performance in which 'light on' caused a negative SP swing lasting 2-3 sec prior to the development of PRS and associated positive SP shifts. In the 'acute' preparation over the striate and parastriate cortex, the SP shifts to 'light on' were sometimes associated with bursts of relatively low amplitude (70-90 /~V) 14-18 c/sec regular waves reminiscent of those induced by a continuous tone over the auditory projections described by K6hler et al. 29, and Gumnit and Grossman 2°. Other parameters of light stimuli, e.g., frequency of light flashes and their intensity were not explored in this preparation.
Cortical SP shifts and eye motion potentials, i.e., EOG
In agreement with observations made by others26, as,s4,5°,53, it was found in anesthetized subjects (whose frontal sinus was prepared as described above) that the EOG influences caused by passive vertical eye rotation (but not by a horizontal one) may 'contaminate' cortical records of SP shifts obtained from electrode pairs oriented in the rostro-caudal direction, provided that the distance between such electrodes was relatively long, e.g., between those located over the anterior ectosylvian and posterior marginal gyri (used in experiments illustrated in Fig. 1, top) or between the frontal sinus and the ectosylvian gyrus (Fig. 5, channel WE-FS). A vertical 45 ° rotation of both eyes caused an average shift of 97 :t: 2/~V and 85 :L 3 #V in each of the two electrode pairs respectively, the downward rotation increasing the negativity of the rostrally located electrode. A vertical rotation of 90 ° (unlikely to occur during operant behav- ior or sleep onset) approximately doubled these EOG influences. In all instances, however, the possible contribution of the EOG to cortical SP shifts observed in freely moving subjects could be differentiated or ruled out by comparing the SP records liable to pick up EOG influences with: (1) transcortical records that have been found free
Brain Research, 26 (1971) 305-332
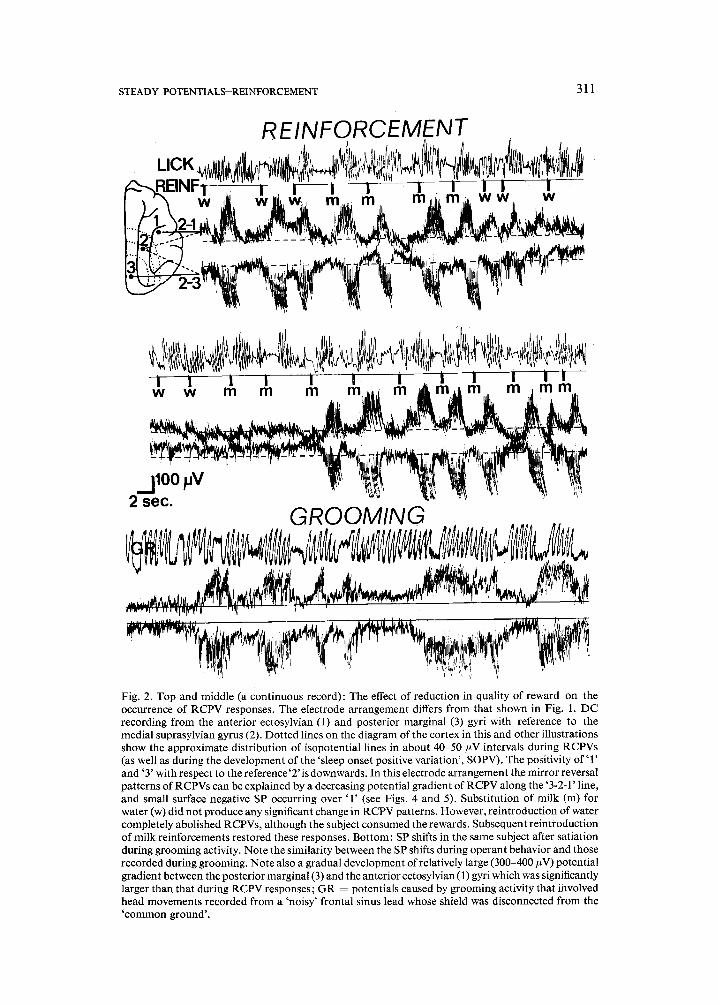
STEADY POTENTIALS-REINFORCEMENT
REINFORCEMENT
311
m
GROOMING II . ' 1
r , ~ , , ~ , , , l ~ i ~ , ~ ,,,~ . . . . . i '
, , L
Fig. 2. Top and middle (a continuous record): The effect of reduction in quality of reward on the occurrence of RCPV responses. The electrode arrangement differs from that shown in Fig. 1. DC recording from the anterior ectosylvian (1) and posterior marginal (3) gyri with reference to the medial suprasylvian gyrus (2). Dotted lines on the diagram of the cortex in this and other illustrations show the approximate distribution of isopotential lines in about 40-50 #V intervals during RCPVs (as well as during the development of the 'sleep onset positive variation', SOPV). The positivity of '1' and '3' with respect to the reference'2' is downwards. In this electrode arrangement the mirror reversal patterns of RCPVs can be explained by a decreasing potential gradient of RCPV along the '3-2-1' line, and small surface negative SP occurring over '1' (see Figs. 4 and 5). Substitution of milk (m) for water (w) did not produce any significant change in RCPV patterns. However, reintroduction of water completely abolished RCPVs, although the subject consumed the rewards. Subsequent reintroduction of milk reinforcements restored these responses. Bottom: SP shifts in the same subject after satiation during grooming activity. Note the similarity between the SP shifts during operant behavior and those recorded during grooming. Note also a gradual development of relatively large (300-400 #V) potential gradient between the posterior marginal (3) and the anterior ectosylvian (1) gyri which was significantly larger than that during RCPV responses; GR = potentials caused by grooming activity that involved head movements recorded from a 'noisy' frontal sinus lead whose shield was disconnected from the 'common ground'.

312 T.J. MARCZYNSKI et al.
from any discernible EOG contamination even to a 90°C vertical rotation of both eyes (such electrodes were used in experiments illustrated in Fig. 1, middle and bottom, and in Fig. 5, channel 3); (2) records obtained in the absence of light, when cortical SP shifts were suppressed, but the eye movements and the lever pressing performance were not (Fig. 1, top and middle); and (3) with SP shifts that occurred during absence of eye movements in a relaxed subject (Fig. 1, bottom right).
In another rostro-caudal electrode arrangement (used in experiments illustrated in Figs. 2 and 3), the SPs were recorded with reference (G-l) to the medial suprasylvian gyrus (2) simultaneously over the anterior ectosylvian (1) and posterior marginal (3) gyri (G-2). In the anesthetized preparation a passive vertical rotation of 45 ° caused in either channel a potential shift of 40-50/zV, the polarity of the EOG being reversed (180 ° out of phase) as that of the true and larger (150-200 #V) epicortical SP shift observed during consumption of reward or sleep onset (Fig. 3, bottom). It may be assumed, however, that the EOG contamination in this electrode arrangement did not significantly contribute to the cortical SP shifts, since an almost isoelectric record was usually observed following substitution of water for mild rewards (Fig. 2, top and middle). In most instances, such change in the quality of reward did not discourage the subject from lever pressing and consuming the apparently less gratifying reward, as judged from signals caused by lever pressing and lapping. During a prolonged non- rewarded lever pressing performance (Fig. 3, top), the restless subject usually walked from one side of the delivery cup to another, and sporadically licked it; this activity caused SP shifts that could be attributed to cable artefacts rather than to EOG con- tamination, since most shifts of the baseline in both channels were in phase, i.e., in contrast to the EOG influences which were 180 ° out of phase in a comparable electrode arrangement explored in anesthetized subjects.
ECoG and SP shifts during simple alimentary behavior
In confirmation of our previous observations 41 and those by other investiga- tors17,24,51, consumption of milk from a bowl in the presence of light was associated with high voltage, 120-200 #V and 6-9 c/sec alpha bursts over the posterior marginal and suprasylvian gyri over which PRS and sleep onset alpha activity were recorded. This synchronized ECoG was associated with large surface positive SP shifts compa- rable to those observed during PRS (see below). Interestingly enough, free consump- tion of semisolid canned cat food was associated with only sporadic bursts of alpha activity and positive SP shifts; in most trials high-frequency (15-20 c/sec) low-voltage ECoG patterns prevailed during which no positive SP shifts were observed.
Post-reinforcement synchronization (PRS) and 'reward contingent positive variation' (RCPV)
All 9 subjects kept on a 23 h food and water deprivation schedule, when placed in the illuminated test chamber, displayed high-frequency (16-24 c/sec) low-voltage (20-30 #V) ECoG activity in frontal (anterior and posterior sigmoid, anterior marginal
Brain Research, 26 (1971) 305-332
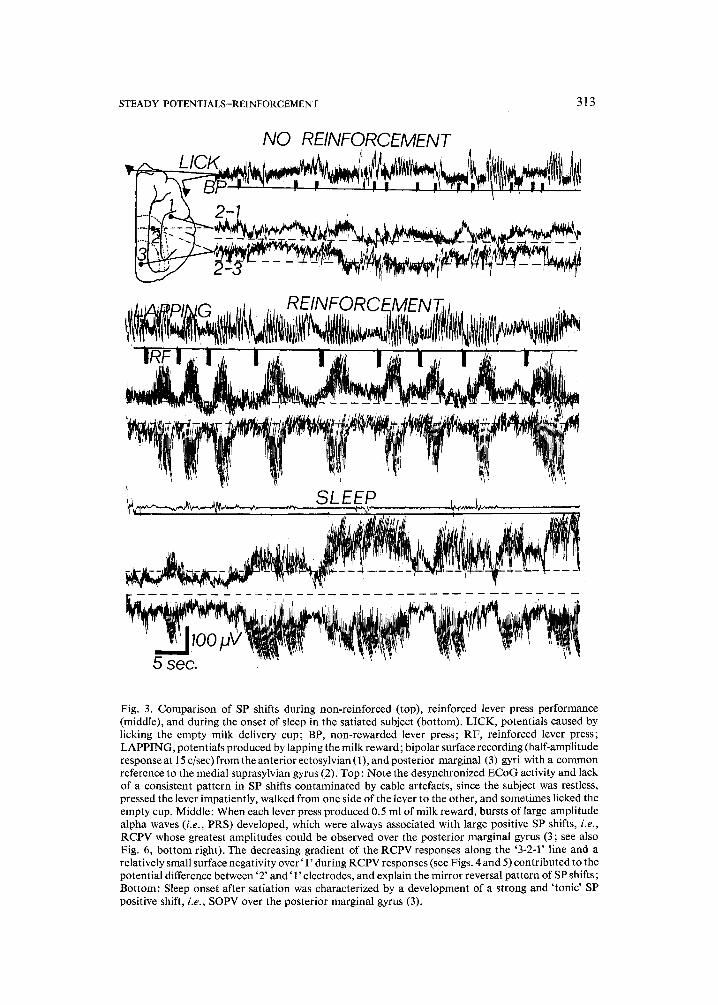
STEADY POTENTIALS--REINFORCEMENT 313
. . . . ° . ,
NO REINFORCEMENT L I C K I ~ ' ' " ' '
" ' P I G " R E I N F O R C E M E N T , j . ~.,~ .
~, , ,. S L E E P I - -
, ' ' i " ' ~ ' ~ ' ~ ~ "- - - - l l ' I
Fig. 3. Comparison of SP shifts during non-reinforced (top), reinforced lever press performance (middle), and during the onset of sleep in the satiated subject (bottom). LICK, potentials caused by licking the empty milk delivery cup; BP, non-rewarded lever press; RF, reinforced lever press; LAPPING, potentials produced by lapping the milk reward; bipolar surface recording (half-amplitude response at 15 c/sec) from the anterior ectosylvian (1), and posterior marginal (3) gyri with a common reference to the medial suprasylvian gyrus (2). Top: Note the desynchronized ECoG activity and lack of a consistent pattern in SP shifts contaminated by cable artefacts, since the subject was restless, pressed the lever impatiently, walked from one side of the lever to the other, and sometimes licked the empty cup. Middle: When each lever press produced 0.5 ml of milk reward, bursts of large amplitude alpha waves (i.e., PRS) developed, which were always associated with large positive SP shifts, i.e., RCPV whose greatest amplitudes could be observed over the posterior marginal gyrus (3; see also Fig. 6, bottom right). The decreasing gradient of the RCPV responses along the '3-2-1' line and a relatively small surface negativity over ' 1' during RCPV responses (see Figs. 4 and 5) contributed to the potential difference between '2' and ' 1' electrodes, and explain the mirror reversal pattern of SP shifts; Bottom: Sleep onset after satiation was characterized by a development of a strong and 'tonic' SP positive shift, i.e., SOPV over the posterior marginal gyrus (3).

314 T.J. MARCZYNSKI et al.
gyri) and parieto-occipital leads (medial and posterior suprasylvian, anterior, medial and posterior ectosylvian, medial and posterior marginal gyri). During the non-re- inforced and prior to a reinforced lever press the ECoG remained desynchronized, and the SP baseline showed little or no significant fluctuations. However, after 5-10 re- inforcements, characteristic bursts of large amplitude (180-200 #V) alpha activity of 5-9 c/sec, i.e., a typical PRS, gradually developed over the parieto-occipital region during consumption of reward. During PRS the frontal cortex remained desyn- chronized. Simultaneously with PRS bursts, large positive SP shifts, i.e., RCPV of 150-400/~V occurred over the same cortical region recorded with reference to either frontal sinus, frontal cortex, occipital crest or white matter subjacent to an 'active' surface electrode. A simultaneous monitoring of PRS and RCPV over the posterior marginal gyrus by using two separate DC channels, one set for half amplitude response at 15 c/sec and the other at 3 c/sec, showed that the RCPV resulted from a temporal summation of single and relatively small 10-30 #V positive potentials carried by each alpha wave (Fig. 1, top). Thus, each RCPV response represented a faithful 'envelop' of a PRS burst. After consumption of reward, both PRS and RCPV disappeared and the tracing returned to the negative baseline. The negativity usually reached the peak just prior to or during the lever press.
The RCPV responses recorded transcortically over the posterior marginal gyrus with reference to the subjacent white matter (Fig. 1, middle) displayed similar patterns and comparable amplitude to those observed in surface bipolar recordings between the 'active' posterior marginal gyrus and the relatively 'inactive' site over the anterior ecto- sylvian gyrus, as shown in Fig. 1 (top). During operant performance as well as during non-operant behavior in a relaxed or drowsy subject, in many instances eye move- ments could be correlated with 50-100 #V positive Iambda waves over the visual cortex (as indicated by vertical dashed lines in Fig. 1, middle and bottom). The lambda waves did not significantly contribute to the development of RCPV responses, since in most instances the former were recorded after a fully developed RCPV response, i.e., during a gradual negative SP shift occurring prior to a subsequent bar press. Neither was a correlation found between the gradually developing 'tonic' positive SP shift over the visual cortex in drowsy subjects and the occurrence of lambda waves (Fig. 1, bottom).
During operant performance in an illuminated cage, the introduction of novel en- vironmental stimuli (such as knocking the cage, fish odor, or cigarette smoke blown into the cage through the ventilation system) abolished both PRS and RCPVresponses, and the SP baseline showed a sustained negative shift, although the operant perform- ance was undisturbed.
The RCPV and visual input; lack of correlation between eye closure and R C P V
Both PRS and RCPV occurred only in an illuminated environment, although all subjects were trained to perform in complete darkness. Therefore, "light off' could not be considered as a novel environmental change. During several experimental sessions the subjects were allowed to perform for 10-30 min in the dark. None of
Brain Research, 26 (1971) 305-332

STEADY POTENTIALS-REINFORCEMENT 315
them developed RCPV or PRS responses, provided that all possible seemingly negligible light sources had been eliminated. During this performance, a variant of PRS gradually developed; it consisted of lower amplitude (70-90 #V) and higher frequency (15-19 c/sec) bursts of synchronization 21, which were not associated with positive SP shifts. Thus, its synaptic mechanisms presumably differed from those of PRS (see Discussion).
Gumnit and Grossman 20 have shown in freely moving cats that auditory stimuli, devoid of conditional properties, cause a sustained negative SP shift over specific auditory projections. Similarly, we have observed that 'light on' caused, in a relaxed subject and sometimes during operant performance, a negative SP shift over the parieto-occipital cortex prior to the development of RCPV responses. Since Row- land 5~ suggested that phasic eye closure and resulting reduction of visual input may be responsible for the occurrence of PRS, an attempt was made to investigate this possibility. A gradual reduction of light intensity in the test chamber from approxi- mately 26-2.6 candles/sq.m did not abolish PRS and RCPV responses, nor did it cause a significant 'tonic' SP shift over the striate and parastriate cortex. Therefore, it is unlikely that a partial eye closure, even if it reduced light input to 1/10, should significantly affect the PRS and RCPV phenomena. Most subjects, when offered a bowl of milk, tended to close their eyes during lapping activity and showed bursts of large amplitude alpha waves associated with SP shifts comparable to RCPVs, but there was no correlation between the onset of the ECoG phenomena and eye closure. During operant behavior, well-trained subjects, if allowed to perform on a schedule in which almost every lever press produced a reward, developed a certain behavioral automatism and kept one paw on the lever and the mouth less than 2 cm above the delivery cup; their eyes usually remained constantly half-closed during both consump- tion associated with PRS-RCPV response and prior to the lever press associated with desynchronization and a negative SP swing. In many instances, during a well-devel- oped PRS-RCPV response the subject's eyes were even opened wider than during the preceding lever press associated with a negative SP swing and desynchronized ECoG. Furthermore, topical (5 ~o solution) or systemic (0.08 mg/kg, i.m.) administration of scopolamine methylbromide, resulting in mydriasis, markedly enhanced PRS-RCPV responses. In addition, a marked enhancement (up to 600 ~) of evoked potentials to flash stimuli presented during PRS-RCPV responses was observed in the optic tract, dorsal lateral geniculate and striate cortex as compared to those recorded during non- rewarded lever pressing or emotional excitement during which the eyes were usually widely opened21, 22. All the above-mentioned observations indicate that: (1) partial eye closure, and the presumable change in the photic influx, were not the determining factors in the PRS-RCPV phenomena; and (2) photic input through half-closed eyes was sufficient for full development of these ECoG phenomena in a test chamber in which the intensity of light could be manipulated within the wide range of 26-2.6 candles/sq.m.
Brain Research, 26 (1971) 305-332
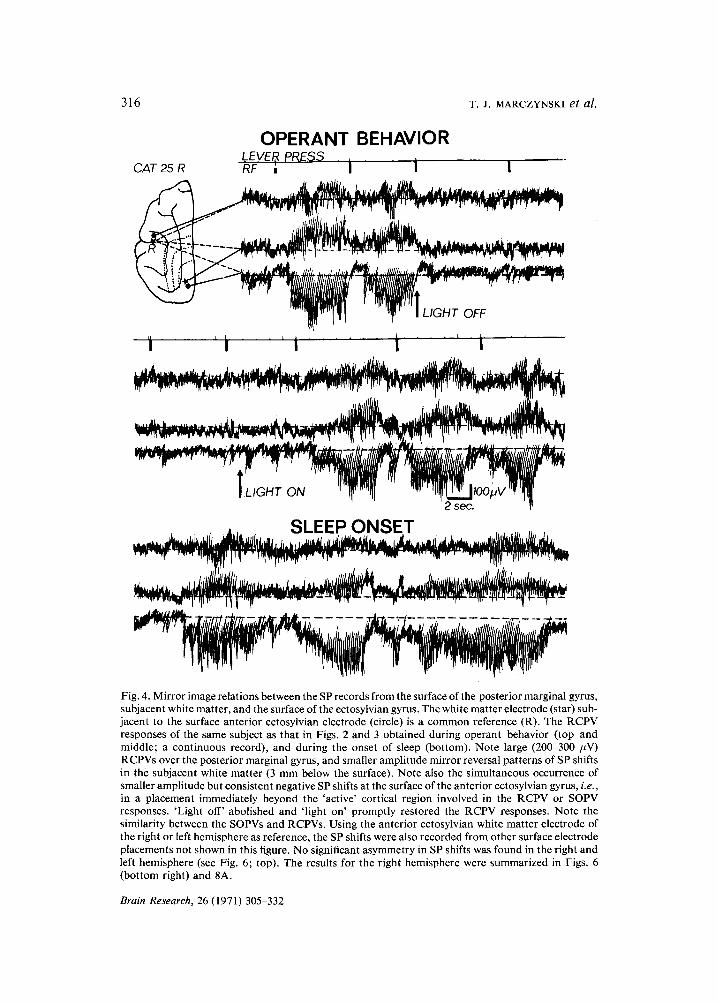
316 T .J . MARCZYNSKI et al.
OPERANT BEHAVIOR I,.EVEI~ PRESS
CAT 25 R RF , i i |
Fig. 4. Mirror image relations between the SP records from the surface of the posterior marginal gyrus, subjacent white matter, and the surface of the ectosylvian gyrus. The white matter electrode (star) sub- jacent to the surface anterior ectosylvian electrode (circle) is a common reference (R). The RCPV responses of the same subject as that in Figs. 2 and 3 obtained during operant behavior (top and middle; a continuous record), and during the onset of sleep (bottom). Note large (200-300 FV) RCPVs over the posterior marginal gyrus, and smaller amplitude mirror reversal patterns of SP shifts in the subjacent white matter (3 mm below the surface). Note also the simultaneous occurrence of smaller amplitude but consistent negative SP shifts at the surface of the anterior ectosylvian gyrus, i.e., in a placement immediately beyond the 'active' cortical region involved in the RCPV or SOPV responses. 'Light off' abolished and 'light on' promptly restored the RCPV responses. Note the similarity between the SOPVs and RCPVs. Using the anterior ectosylvian white matter electrode of the right or left hemisphere as reference, the SP shifts were also recorded from other surface electrode placements not shown in this figure. No significant asymmetry in SP shifts was found in the right and left hemisphere (see Fig. 6; top). The results for the right hemisphere were summarized in Figs. 6 (bottom right) and 8A.
Brain Research, 26 (1971) 305-332

STEADY POTENTIALS-REINFORCEMENT 317
The RCPV and quality of reinforcement
In all our subjects reduction in quality of reinforcement affected both PRS and RCPV responses. When water was used as reinforcer from the beginning of an exper- imental session, the food and water deprived subject usually developed large PRS and RCPV responses after 5-15 reinforcements. Using Student's t-test no significant differ- ences were found between the mean values of the integrated RCPV responses (ex- pressed in RCPV units; for definition, see Methods) induced by milk or water (P < 0.01)*. A substitution of milk rewards for water during an experimental session had no significant effect (P < 0.01). However, when water was substituted for milk during the same experimental session, in most experiments there was abolishment or a significant reduction of RCPV responses when compared to experiments in which initially water or milk was used (P < 0.001), although the subject usually consumed the water rewards. Reintroduction of milk rewards promptly restored both responses. In Fig. 2 (top and middle) the effect of reduction in quality on RCPV responses was recorded simultaneously from the surface of the anterior ectosylvian (1) and from the posterior marginal gyrus (3) with reference to the medial suprasylvian gyrus (2). In this electrode arrangement a mirror reversal pattern of RCPVs was always obtained. During grooming activity in the same satiated subject similar SP shifts were observed in the same electrode arrangement (Fig. 2, bottom). Measuring the SP difference between the anterior ectosylvian and posterior marginal gyri (by summing the ampli- tude of the negative SP in the '2-1' tracing and the positive SP in the '2-3' tracing) values of 250-700 #V were observed during operant performance as well as during grooming activity. The latter seemed to be most effective in the genital region (Fig. 2, bottom).
The degree of fluid deprivation obviously played a significant role in this para- digm, since in subjects deprived only of fluid (milk and water) for a longer time period than 23 h a substitution of water for milk rewards, and vice versa, did not abolish the occurrence of large PRS and RCPV responses; the amplitude and duration of these SP shifts observed after reintroduction of water rewards were comparable to those recorded after milk (P > 0.1).
Lack of correlation between the rhythm of lapping activity and PRS waves
Since Sadowski and Longo .57 have reported 'masticatory waves' in the ECoG of rabbit, an attempt was made to test a possible correlation between the lapping move- ments and the frequency of alpha waves in PRS bursts. No such correlation was found: the Pearson's correlation coefficient for these two rhythms (calculated from series of 20 pairs of PRS and lapping responses) ranged from 0.03 to 0.06 (t < 0.2). The fre- quency of PRS waves remained usually very stable during a time period of several months and was characteristic of an electrode pair in a particular subject. Average frequency calculated from 30 PRS bursts usually showed less than l ~ standard deviation, provided that the intensity of light in the test chamber was kept constant,
* An integrated RCPV record was published in Science, 163 (1969) 302; see also ref. 39.
Brain Research, 26 (1971) 305-332

318 T.J. MARCZYNSKI et al.
and the subject was allowed to perform almost every day. The frequency of PRS waves recorded over the posterior marginal gyrus (with reference to the subjacent white matter or the distant anterior ectosylvian gyrus) ranged from 5 to 9.5 c/sec across all subjects (average 7.8; S.E. j- 0.6 c/sec). In contrast, the frequency of lapping movements in the same subject varied from one trial to another or even significantly changed during the same experimental session; it was usually faster at the beginning, and then gradually slowed down toward the end of the trial, when the subject became satiated. The average frequency of lapping movements ranged from 1.5 to 2.5 c/sec in the same subject and from 1.5 to 3.3 c/sec across all subjects used in this study.
In approximately 97Yo of instances, the PRS and RCPV occurred 0.5-2.5 sec after the onset of lapping. In approximately 2 ~ of responses, the PRS and RCPV occurred after a latency of more than 3 sec, i.e., at the end of consumption, when the subject licked the empty cup at a much slower pace (1-2 c/sec). Also during grooming, associated with much slower rhythmic (0.8-1.8 c/sec) tongue and head movements as compared to lapping, the patterns and frequency of alpha activity and the concom- itant positive SP shifts were essentially undistinguishable from those observed during lapping (Fig. 2) or sleep onset (Figs. 3, 4 and 5).
RCPV and 'sleep onset positive variation' (SOPV)
During the onset of sleep in satiated subjects, bursts of high voltage 180-200/zV alpha activity 5-9 c/sec were consistently observed prior to the development of delta wave sleep patterns. This activity always started over the posterior marginal gyrus in both hemispheres, and gradually spread to the parieto-occipital and to the frontal cortices. The alpha burst over the posterior marginal gyrus was always associated with large positive SP shifts, i.e., SOPVs of about 200-700 #V over the same area in both hemispheres in referential recording to occipital crest, frontal sinus, in bipolar surface recording with reference to the frontal or parietal (e.g., anterior ectosylvian gyrus) cortices, or in transcortical recording with reference to the subjacent white matter. These SOPVs displayed patterns indistinguishable from those of RCPVs, as well as SP shifts recorded during grooming. Fig. 3 shows representative samples of SP shifts during non-reinforced (top), reinforced lever press performance in the illuminated cage (middle) and during sleep onset in the same satiated subject (bottom). The mirror reversal patterns of SP shifts (obtained with the electrode arrangement shown in the diagram of the cortex) suggest (in conjunction with results presented in the next paragraph) that a strong positive potential field developed over the region of the posterior marginal gyrus after each reinforcement as well as during each 6-8 c/sec alpha burst associated with the onset of sleep; it also indicates that a relatively strong potential gradient developed in the anterior-lateral direction along the '3-2-1' line (Fig. 3).
Topographical distribution of RCPVs and SOP Vs
Large amplitude (200--700 #V) RCPV and SOPV responses over the posterior
~3rain Research, 26 (1971) 305-332
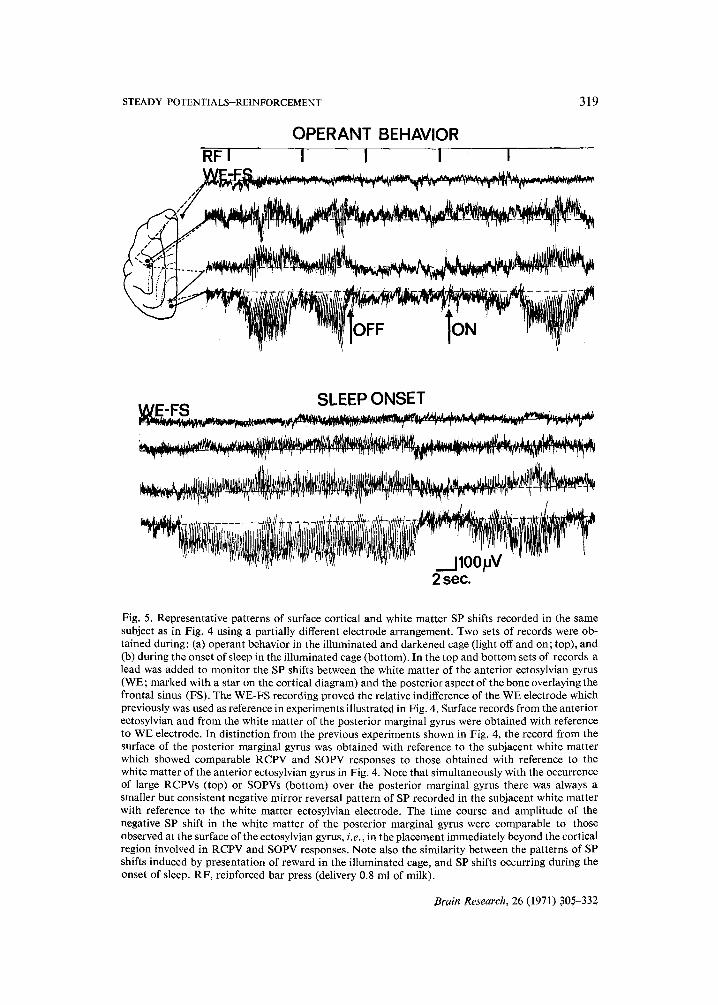
STEADY POTENTIALS-REINFORCEMENT
OPERANT BEHAVIOR RFI I I I
319
SLEEP ONSET -FS ' ~ . ~ , , ~ % ~
' 'm,{ ~,',m,,~,',,t~",'fr"~ -,i~7,
2 see.
Fig. 5. Representative patterns of surface cortical and white matter SP shifts recorded in the same subject as in Fig. 4 using a partially different electrode arrangement. Two sets of records were ob- tained during: (a) operant behavior in the illuminated and darkened cage (light off and on; top), and (b) during the onset of sleep in the illuminated cage (bottom). In the top and bot tom sets of records a lead was added to monitor the SP shifts between the white matter of the anterior ectosylvian gyrus (WE; marked with a star on the cortical diagram) and the posterior aspect of the bone overlaying the frontal sinus (FS). The WE-FS recording proved the relative indifference of the WE electrode which previously was used as reference in experiments illustrated in Fig. 4. Surface records from the anterior ectosylvian and from the white matter of the posterior marginal gyrus were obtained with reference to WE electrode. In distinction from the previous experiments shown in Fig. 4, the record from the surface of the posterior marginal gyrus was obtained with reference to the subjacent white matter which showed comparable RCPV and SOPV responses to those obtained with reference to the white matter of the anterior ectosylvian gyrus in Fig. 4. Note that simultaneously with the occurrence of large RCPVs (top) or SOPVs (bottom) over the posterior marginal gyrus there was always a smaller but consistent negative mirror reversal pattern of SP recorded in the subjacent white matter with reference to the white matter ectosylvian electrode. The time course and amplitude of the negative SP shift in the white matter of the posterior marginal gyrus were comparable to those observed at the surface of the ectosylvian gyrus, i.e., in the placement immediately beyond the cortical region involved in RCPV and SOPV responses. Note also the similarity between the patterns of SP shifts induced by presentation of reward in the illuminated cage, and SP shifts occurring during the onset of sleep. RF, reinforced bar press (delivery 0.8 ml of milk).
Brain Research, 26 (1971) .305-332

320 T.J. MARCZYNSKI et al.
marginal gyri of bo th hemispheres were recorded not only with reference to the imme-
diately subjacent white mat ter or relatively distant surface of the medial suprasylvian
and anter ior ectosylvian gyri (as shown in Figs. 1, 2 and 3), bu t also with reference (R)
to the white mat ter of the lat ter gyrus (Fig. 4). Using the white mat ter of the anter ior
ectosylvian gyrus as a reference (R), large (200-600 #V) RCPVs were recorded over
the posterior marginal gyrus, and a relatively small (50-70 #V) bu t consistent inver-
sion of this SP in the subjacent white mat ter (star). A similar inversion was observed
on the surface at the periphery of the striate and parastr iate cortex involved in RCPV
responses, e.g., over the anter ior ectosylvian gyrus (solid circle). The ampli tude and
pat terns of SP shifts associated with the onset of sleep, both at the surface and in the
white matter, were indist inguishable from those occurring after the presentat ion of
food rewards (Fig. 4, bot tom). The only difference between these responses in this and
other electrode ar rangements was that the RCPVs were cont ingent upon an adequate
i l luminat ion of the test chamber, whereas the SOPVs were not , a l though the develop-
ment of the latter was somehow facilitated by ambient light.
To prove that the white mat ter of the anter ior ectosylvian gyrus was a relatively
reliable reference, this electrode (WE) in the same subject was paired with a frontal
Fig. 6. Topographical distribution of RCPV and SOPV. Top left: During sleep onset the absence of significant SP shifts in bipolar 'transcallosal' recording between the left and right homotopical elec- trode placements over the anterior ectosylvian (1), medial suprasylvian (2) and posterior marginal (3,4) gyri. Note that a simultaneous transcortical recording from the left posterior marginal gyrus (4) with respect to the white matter electrode (located 3 mm below the cortical surface and marked with a star) shows large amplitude sleep onset positive variation (SOPV) associated with bursts of high- voltage slow-wave activity of approximately 8 c/sec. Top right: The same subject in the same 'trans- callosal' electrode arrangement shows no substantial SP shifts during operant behavior. This contrasts again with the simultaneous occurrence of large amplitude RCPV responses in the transcortical record- ing over the posterior marginal gyrus (4). Note that the gain in channels 1, 2 and 3 in both top figures is approximately twice as high as that in channels 4 and 5. RF, reinforcement. Bottom left: A comparison between the surface bipolar and the transcortical recording from the right hemisphere in the same subject as above during operant behavior. Note the absence of substantial SP shifts in 2-1 recording at the periphery of the cortical region involved in RCPV responses (a tangential orientation of the 2-1 electrodes with respect to the tentative isoelectric line of RCPV responses). A simultaneous transcorti- cal recording from the point '2' with respect to the subjacent white matter (,-2 lead) shows relatively small but consistent negative SP shifts that coincide with large RCPVs in transcortical recording over the posterior marginal gyrus (lead *-4). The small negativity over '1' (not shown in the left bottom but in right bottom figure) was comparable to that recorded over '2', as it can also be inferred from the lack of SP shift between '1' and '2'. These surface negative shifts indicate that the region of the anterior ectosylvian, anterior suprasylvian and anterior marginal gyri becomes a 'sink' for a current flow from the strong positive 'source' over the parieto-occipital region during RCPV and SOPV responses. The topographical distribution of RCPVs and SOPVs is outlined by tentative isoelectric lines based on other experiments. Note also that in bipolar surface recording the anterior ectosylvian with respect to the medial suprasylvian gyrus (3-2 lead), and the latter with respect to posterior marginal gyrus (4-3 lead) became negative (the larger distance 4-3 as compared to 3-2 being approximately proportional to the amplitude of the negativity). The reversal of polarity in 4-3 recording as compared to the trans- cortical one over '4' (,-4 lead) can be explained by the maximum RCPV amplitude occurring over '4', and a decreasing amplitude of RCPV responses along the 4-3-2-1 line as shown in the bottom right figure in which in the same subject the RCPVs were referentially recorded with respect to the common reference (R) located in the white matter of the anterior ectosylvian gyrus. This reference was pre- viously shown to be relatively 'indifferent' with respect to the bone overlaying the posterior aspect of the frontal sinus (Fig. 5; top and bottom).
Brain Research, 26 (1971) 305-332
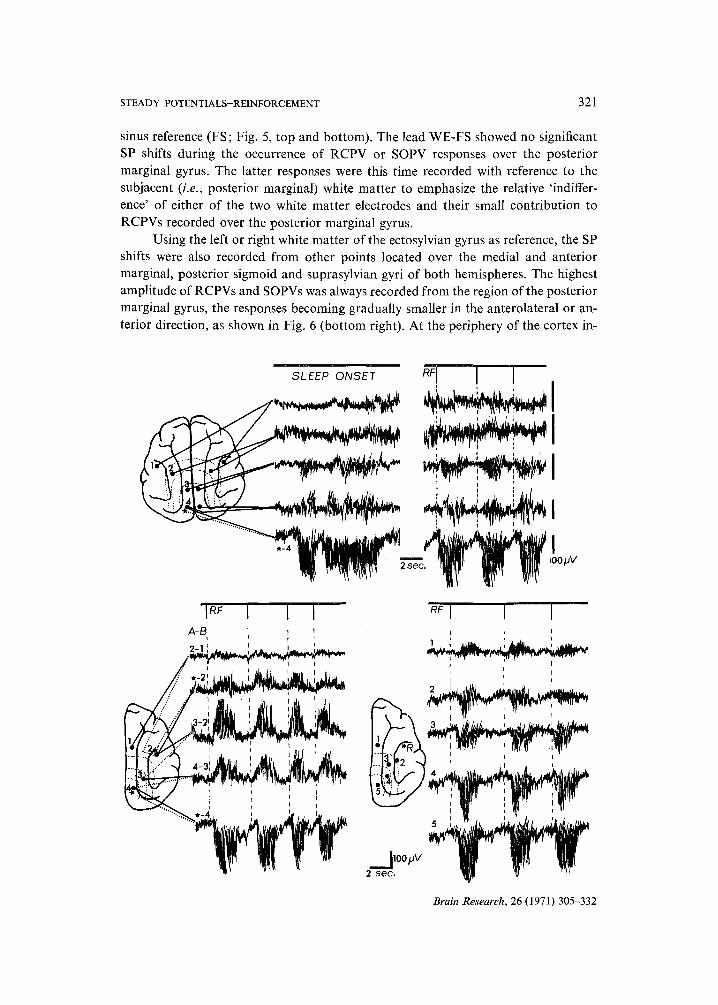
STEADY POTENTIALS-REINFORCEMENT 321
sinus reference (FS; Fig. 5, top and bottom). The lead WE-FS showed no significant SP shifts during the occurrence of RCPV or SOPV responses over the posterior marginal gyrus. The latter responses were this time recorded with reference to the subjacent (i.e., posterior marginal) white matter to emphasize the relative 'indiffer- ence' of either of the two white matter electrodes and their small contribution to RCPVs recorded over the posterior marginal gyrus.
Using the left or right white matter of the ectosylvian gyrus as reference, the SP shifts were also recorded from other points located over the medial and anterior marginal, posterior sigmoid and suprasylvian gyri of both hemispheres. The highest amplitude of RCPVs and SOPVs was always recorded from the region of the posterior marginal gyrus, the responses becoming gradually smaller in the anterolateral or an- terior direction, as shown in Fig. 6 (bottom right). At the periphery of the cortex in-
I I
I RF I I I A - B ', , ,
t T I ~ _ i I I I I
I I I I
/ ~ i *-211 I I l
..: i I I I I * .: ~ I I I
.:" - 2 1 1 i • ¢ i r
I k - 4 I I , [
RFI I I I i I
I I I I I I
~, ' \ I 'l[ ~ 1 .
..... } ,, , ,
2 s e c . I W 1,~ "
Brain Research, 26 (1971) 305-332

322 T.J. MARCZYNSKI et al.
volved in RCPV and SOPV responses, over the anterior marginal gyrus (electrode 1), an inversion of RCPV was observed comparable to that previously shown on the surface of the anterior ectosylvian gyrus (in Figs. 4 and 5). The measurements of maximum amplitude of RCPV and SOPV responses over the frontal and parieto- occipital region were carried out in both hemispheres with reference to white matter of the anterior ectosylvian gyrus. The mean values based on twenty such measurements were plotted (with two standard deviations represented by vertical lines) in Fig. 8, Diagram A. The method of measuring the maximum amplitude of RCPVs and SOPVs from records filtered to half-amplitude response at 3 c/sec is shown in the inserts at the left and right bottom of Fig. 8. There were no significant quantitative differences be- tween the amplitudes and distribution of RCPV and SOPV responses; both displayed maximum amplitudes over the region of the posterior marginal gyrus, a small but con- sistent mirrer reversal image over the anterior marginal and anterior ectosylvian gyri, and in the white matter of the posterior marginal gyrus. It is noteworthy that the occipital crest bone marrow electrode that was meant as a reference (OC 1) was markedly influenced in all subjects by positive SP shifts presumably occurring over the posterior marginal gyri.
The symmetry in distribution of RCPVs and SOPVs over both hemispheres was demonstrated in simultaneous 'transcallosal' recording from homotopical surface electrodes over the ectosylvian, medial suprasylvian, and posterior marginal gyri, (Fig. 6, top left and right). In spite of the relatively large cortical area subtended between the ectosylvian (1) and medial suprasylvian (2) electrodes, and significantly increased gain in the first top 3 channels, relatively small SP shifts were recorded in these leads as compared to large SOPVs and RCPVs obtained simultaneously in transcortical recording over the posterior marginal gyrus (last channel marked ,-4). In other electrode arrangements, e.g., using as reference (R) the surface anterior ectosylvian electrode (Fig. 8B), or white matter of the posterior marginal gyrus (Fig. 8C), an approximately symmetric distribution of RCPV and SOPV responses on both hemispheres was quantitatively demonstrated, the maximum amplitude being observed over the posterior marginal gyri of both hemispheres, with a sharply falling potential gradient in the anterolateral and anterior direction.
A joint consideration of SP patterns shown in the left and right bottom illustra- tions of Fig. 6 is necessary for a proper interpretation of the previously shown surface bipolar records of large RCPVs over the posterior marginal gyrus obtained with reference to the distant anterior ectosylvian gyrus (Fig. 1, top), and of the surface mirror image inversion of RCPVs and SOPVs recorded with reference to the medial suprasylvian gyrus over the posterior marginal and anterior ectosylvian gyri (Figs. 2 and 3). In surface recording with reference (R) to the white matter of the anterior ectosylvian gyrus (Fig. 6, bottom right), a gradually decreasing amplitude of RCPVs was observed in the antero-lateral direction of electrode placements ('5-4-3-2-1' line). Over the anterior marginal gyrus (1) a small but consistent inversion of RCPV was observed (average 20-30 #V negative as shown in Fig. 8A, lead 6); a comparable or somewhat larger inversion was recorded over the anterior ectosylvian gyrus as shown in the bottom left illustration of Fig. 6, lead ,-2 (the average negativity ranged from 40
Brain Research, 26 (1971) 305-332
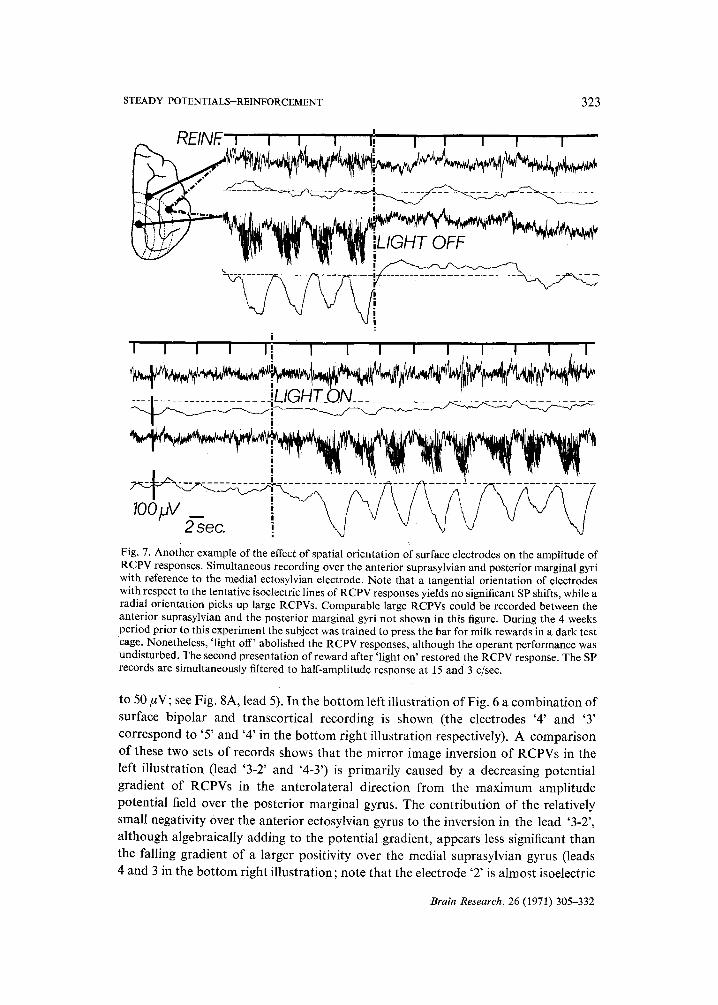
STEADY POTENTIALS-REINFORCEMENT 323
_ REINE,.,, . . . . . . . J . . / .. I . . , . .
~- . . . . . . . . . . . . . . . . . . . . . . . . i . . . . . . . . . .
i
. . . . . . - J L / - G H - T - . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . - - - _ i _ . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
2"~ec. ! Fig. 7. Another example of the effect of spatial orientation of surface electrodes on the amplitude of RCPV responses. Simultaneous recording over the anterior suprasylvian and posterior marginal gyri with reference to the medial ectosylvian electrode. Note that a tangential orientation of electrodes with respect to the tentative isoelectric lines of RCPV responses yields no significant SP shifts, while a radial orientation picks up large RCPVs. Comparable large RCPVs could be recorded between the anterior suprasylvian and the posterior marginal gyri not shown in this figure. During the 4 weeks period prior to this experiment the subject was trained to press the bar for milk rewards in a dark test cage. Nonetheless, 'light off' abolished the RCPV responses, although the operant performance was undisturbed. The second presentation of reward after 'light on' restored the RCPV response. The SP records are simultaneously filtered to half-amplitude response at 15 and 3 c/see.
to 50 #V; see Fig. 8A, lead 5). In the b o t t o m left i l lus t ra t ion o f Fig. 6 a combina t ion o f
surface b ipo la r and t ranscor t ica l recording is shown (the electrodes '4 ' and '3 '
co r respond to '5 ' and '4 ' in the b o t t o m r ight i l lus t ra t ion respectively). A compar i son
o f these two sets o f records shows tha t the mi r ro r image invers ion o f RCPVs in the
left i l lus t ra t ion ( lead '3-2' and '4-3') is p r imar i ly caused by a decreasing po ten t ia l
g rad ien t o f RCPVs in the an te ro la te ra l d i rec t ion f rom the m a x i m u m ampl i tude
poten t ia l field over the pos te r io r marg ina l gyrus. The con t r ibu t ion o f the relat ively
small negat ivi ty over the an te r ior ectosylvian gyrus to the inversion in the lead '3-2' ,
a l though a lgebraical ly adding to the po ten t ia l g rad ien t , appea r s less significant than
the fall ing g rad ien t o f a larger posi t iv i ty over the media l suprasylvian gyrus (leads
4 and 3 in the b o t t o m r ight i l lus t ra t ion; note tha t the e lectrode '2 ' is a lmos t isoelectric
Brain Research, 26 (1971) 305-332

324 T.J . MARCZYNSKI et al.
A 25R B 25 C 30;16 D . . . . . . . . . . . . . . . . . . . . . - i . . . . . . . . . . . . . . . . .
: RCPV ~ ~ ~
' o~_.-.SOPV 100 - - ~ . . . . . . . . . .
_"_~ _
R P SOPV ~ /
! ~ r,?, ,, .'¢
1 ,~=WHITE MATTER
Fig. 8. Topographical distribution of RCPV and SOPV responses. In the upper part of the figure the mean maximum RCPV and SOPV (with two standard deviations) are represented on the voltage scale by solid circles-continuous line and by open circles-dotted line respectively. The mean maxi- mum values are based on twenty measurements for each cortical (solid circles) and subjacent white matter (star) locations shown on the cortical diagrams below in three different electrode arrangements: A, B, and C. Method of taking the maximum amplitude measure is shown in the left and right bottom inserts (the upper record was filtered to half-amplitude response at 35 c/sec, the lower one, from which amplitude was measured, at 3 c/sec; triangles above the RCPVrecord denote delivery of milk reward). Electrode arrangement 'A' shows the distribution of SP shifts on the right hemisphere with reference (R) to the white matter electrode located in the anterior ectosylvian gyrus; diagram 'B' shows re- sponses from both hemispheres with reference to the surface electrode (R) located over the left anterior ectosylvian gyrus; diagram 'C' summarizes responses from both hemispheres recorded with reference (R) to the white matter or the left posterior marginal gyrus. Cortical diagram 'D ' summarizes the distribution of surface positive and negative points during RCPV and SOPV responses obtained from 39 electrodes in 5 subjects. Note that during SOPV and RCPV responses positive SP shifts could also be recorded from the bone, e.g., occipital crest (OC) overlying the cerebellum (diagrams A and D). Note that in all three (A, B, and C) electrode arrangements the region of the posterior marginal gyrus of both hemispheres shows the highest amplitude of positive shifts; the white matter electrodes (stars) subjacent to the most 'active' cortex shows relatively small but consistent polarity reversal of RCPV and SOPV responses (diagram A, electrode 2, and diagram B, electrode 5); similar polarity inversion was recorded from surface electrodes at the immediate periphery of the 'active' cortex involved in RCPV and SOPV responses (diagrams A and C); between homotopical surface electrode locations on the left and right hemisphere with reference (R) to anterior ectosylvian gyrus (diagram B) or with reference to the white matter of the posterior marginal gyrus comparable SP shifts were recorded, thus implying an approximately symmetrical distribution of RCPV and SOPV responses on both hemi- spheres.
and is loca ted on the reversa l l ine). I n a g r e e m e n t w i th t he genera l d i s t r i bu t i on pa t t e rn s
o f R C P V s and S O P V s (Fig. 8A, B and C), a rad ia l o r i e n t a t i o n o f e l ec t rodes o v e r the
pa r i e to -occ ip i t a l c o r t e x wi th respec t to t he m a x i m u m p o t e n t i a l field o v e r the p o s t e r i o r
m a r g i n a l gyrus y ie lded large responses , w h e r e a s s i m u l t a n e o u s r e c o r d i n g f r o m t angen-
t ial ly o r i en ted e l ec t rodes s h o w e d ne i the r c o n s p i c u o u s P R S n o r R C P V responses as
Brain Research, 26 (1971) 305-332

STEADY POTENTIALS-REINFORCEMENT 325
illustrated in Fig. 7; the same holds for SOPVs and sleep onset alpha activity (not shown in this figure).
The tentative isoelectric lines, drawn in approximately 60-80 #V steps on all cortical diagrams (in a concentric manner with respect to the maximum potential field over the posterior marginal gyri) are based on quantitative data obtained from 4 subjects (25, 25-R, 30 and 16) in which a total of 29 varying electrode locations were considered (Fig. 8A, B, C and D).
All patterns of both surface and white matter SP shifts were stable and repro- ducible over a time period of more than 3 months in 5 out of 9 subjects. The remaining 4 subjects also showed consistent patterns of surface and white matter distribution of SP shifts, except for a rather inconsistent polarity inversion over the anterior ecto- sylvian gyrus when recorded with reference to either frontal sinus, or subjacent white matter. The reason for this discrepancy is not known. The most likely cause is a misplacement of the epidural electrode (these subjects have not yet been sacrificed) or an individual small difference in the spread of RCPV and SOPV responses over the cortical surface.
Other foci of ECoG synchronization and SP shifts
In relaxed but waking subjects, prior to development of large amplitude SOPVs, smaller amplitude (70-90 #V) SP shifts associated with spindle-like (10-18 c/sec) ECoG synchronization were observed over the parastriate cortex (medial supra- sylvian and medial marginal gyri) and over the auditory projections (medial and anterior ectosylvian gyrus). These foci occurred either spontaneously or upon an introduction of mild auditory stimuli (gentle knocking on the test chamber or in response to laboratory noise). They disappeared with the development of large amplitude alpha SOPV response over the striate cortex and subsequent spread of the latter over the entire parieto-occipital cortex. The exact topographical distribution of the small SP loci was not systematically studied.
DISCUSSION
There are at least two categories of changes in SP between the brain and the extracerebral reference. One category is characterized by slow onset (approximately 1-10 min), diffuse distribution in the cortex and subcortical structures, and lack of polarity inversion as an electrode is inserted through the cortex. It is believed that these SPs are determined by changes in blood-brain barrier potential 6z, blood osmolali- ty H,zs, regional metabolism and blood flow 6s, or CO2 concentration 7°. Another category of SP is believed to depend solely on changes in patterns of regional synaptic activity2,5,6,15,46, 5~, and has accordingly different characteristics. The following ob- servations indicate that the RCPV and SOPV as well as SP associated with grooming were induced by phasic changes in the synaptic activity of cortical neurons: (1) a relatively fast time course of both phenomena; they may develop within a few seconds and as quickly disappear either spontaneously or in response to a novel environmental
Brain Research, 26 (1971) 305-332

326 T. J, MARCZYNSKI et al.
stimulus; (2) characteristic topographical distribution on the surface of the parieto- occipital cortex approximately identical with the specific and non-specific visual pro- jections; (3) two polarity reversals and dipoles were generated by the parieto-occipital cortex involved in RCPV and SOPV responses: the stronger was oriented vertically to the surface; it had a 'source' at the cortical surface of the posterior and medial marginal and posterior and medial suprasylvian gyri, and a 'sink' in the subjacent white matter; the second dipole was oriented parallel to the surface; its 'source' was identical with the previous one, but its 'sink' was located at the immediate periphery of the cortex involved in RCPV and SOPV responses, i.e., over the anterior ectosylvian, anterior suprasylvian, and anterior marginal gyri; (4) contingency of RCPV upon relative quality and taste of food reward; (5) high susceptibility of RCPV and PRS responses to peripheral administration of anticholinergic drugs: atropine sulfate, scopolamine (but not methylscopolamine) abolished, and physostigmine promptly restored these responses35,~7,39,4°; (6) lack of correlation between the PRS-RCPV responses (including similar ECoG phenomena during grooming or sleep onset) on the one hand, and the rhythm of lapping activity, and the eye and head movements on the other; and (7) contingency of RCPV upon visual input, even in subjects previously trained in the dark (a diffuse, unpatterned visual input turned out to be sufficient in experiments in which the subjects, wearing translucent contact lenses, performed in an illuminated environment39,40).
Although no laminar recording was attempted, and the records were obtained only from the surface and subjacent white matter, it may be assumed (on the basis of what is known of the functional relationship between single unit activity and surface potentials 9a,9b,l°,66) that the synaptic events which caused current flow at the surface and in the white matter occurred in the gray matter. Our observations seem to be most consistent with the view that the RCPV responses result from temporal summation of widespread hyperpolarizing inhibitory postsynaptic potentials that invade apical dendrites from cell bodies located in the deeper cortical layers. One can assume that the vertical dipole of the RCPV and SOPV responses resulted from thousands of small parallel dipoles generated by single neurons and their apical dendrites. The indirect evidence and arguments that lend support to this postulate and also tentatively explain why the SOPV, in contrast to the RCPV, does not depend on visual input, have been discussed elsewhere36, 39.
The second dipole observed during RCPV and SOPV, oriented parallel to the surface, with a 'source' over the striate and parastriate cortex, and a 'sink' over the immediate periphery, can be visualized as resulting from a current flow through the glia substrate. There are indications that glia are joined in a syncytium by tight electri- cal junctions 1s,48, and that current may flow through glia tissue, as shown in lower animals al,zz. Studies on the relationship between the slow intracortica112 and/or surface positivity on the one hand, and the rate and probability of neuronal firing and time course of the inhibitory postsynaptic potentialsga,gb, 1°,66 on the other (with the exception of results by one group of investigators lz,14 who probably have mis- interpreted their observations9b), lend some support to the view that RCPV may be caused by hyperpolarizing inhibition of neurons in the striate cortex 39. Hence, it can
Brain Research, 26 (1971) 305-332

STEADY POTENTIALS-REINFORCEMENT 327
be assumed that during the peak of the RCPV or SOPV responses the overall number of neuronal discharges is relatively greater at the periphery of the RCPV or SOPV focus. Neuronal depolarization and associated release of K + cause also a prolonged local depolarization of glia al. Thus, one can postulate a current flow from the 'source' glia of the RCPV focus to the 'sink' located at the periphery, e.g., to the anterior ectosylvian gyrus over which a consistent negative SP reversal was observed during the peak of RCPV or SOPV responses.
Contrary to our results, Grandstaff 17 reported that PRS of 7 c/sec occurred over the striate cortex in split-brain cat preparation in which visual input to the ipsilateral eye was 'totally' blocked. Since an opaque corneal occluder in a presumably illumi- nated test chamber was used, it can be assumed that a reduced light input, sufficient to maintain PRS activity, entered the 'occluded' eye. This possibility should be consid- ered, since we have found that even one-tenth of the usual light intensity in the test chamber, i.e., approximately 2.6 candles/sq.m, was sufficient to maintain the PRS bursts in a well-trained subject2Z, 2a. Grandstaff 17 also suggested that PRS may be simply determined by a specific state generated by lapping activity, and that a proprio- ceptive input associated with tongue movements may play the role of a conditional stimulus. These suggestions are not plausible, since: (1) the PRS-RCPV phenomena depend on the desirability, and thus presumably on the hedonic character of reward as well as on the absence of disturbing environmental stimuli8,41,60; (2) there was no correlation between the changing rhythm of lapping activity and the relatively stable rhythm of PRS; and (3) the PRS and RCPV patterns were indistinguishable from those of alpha bursts and SP shifts that occurred during grooming or sleep onset (see also Roth et a l ) l ) . Hence, it seems that bursts of alpha activity and the concomitant positive SP shifts reflect mechanisms which are common to all these behavioral states, i.e., phasic internal inhibition, and a 'quantum-like' reduction of the tonus of the reticular activating system, as suggested by Sterman and Wyrwicka 6°, and others41,sL This interpretation of RCPV and SOPV phenomena is in agreement with Caspers '5,6 view that surface depolarization of apical dendrites and the negative SP shift are related to arousal, desynchronization and an increased tonus of the brain stem reticular activating system, and that positive surface SP shifts reflect hyperpolarizing mechanisms related to synchronization and reduced wakefulness.
Our observations on RCPV are compatible with more recent results obtained by Irwin and Rebert 25, Low et alfl TM, Kornhuber and Deecke 3°, McAdam and Seals 44 as well as with the original observations made in man by Walter et al. 64,65. The 'contingent negative variation' of Walter and the 'conative negative variation' of Low et al. that occurred in man after the 'warning' signal, seem to correspond to the negative baseline observed in the cat during non-rewarded lever pressing performance or prior to the rewarded lever press. On the other hand, the sudden termination of the state of expectancy or conation that is reflected as' an abrupt termination of the CNV in man upon a successful completion of the task, seem to correspond to the RCPV triggered by a successul lever press and consumption of reward. The obvious differ- ence between the termination of the CNV and the patterns of RCPV is that the latter was always associated with a burst of alpha activity, i.e., PRS. On the other hand, both
Brain Research, 26 (1971) 305-332

328 T.J. MARCZYNSKI et al.
the PRS-RCPV phenomena and the termination of the CNV were associated with a conspicuous enhancement of cortical evoked potentials to sensory stimuli21,z2,38,41, 65.
The question of why the bursts of alpha activity and positive SP shifts were so consistently observed during lapping of milk from a bowl, but occurred only sporadi- cally during eating semisolid canned cat food, is difficult to answer. It can be suggested, however, that meat reward has a different hedonic character and that chewing large chunks of meat, in contrast to lapping, is usually related to a successful hunt and/or fight, and therefore may 'set' a different eleztrophysiological and emotional state of violence and increased tonus of the arousal system.
In the cat, mirror image relations in SP shifts were recorded from the cortical surface and subjacent white matter during consumption of high incentive food; they disappeared or even reversed in polarity after satiation, and depended on the sense of taste, since anesthetization of the mouth blocked the SP shifts but not the con- summatory behavior 54. Topographically restricted surface SP shifts were also observed in female cats during estrus in response to genital stimulation, and in rats during intracranial self-stimulation 54. In rats during self-stimulation in the caudal hypothalamus or rostral midbrain, negative epicortical SP shifts were observed by Wurtz 69, which are characteristic of orienting reaction and arousal 5,6,27,46,54. If both Wurtz's and our data, although seemingly contradictory, reflect effects generated by reward substrates, it may be assumed that rewarding electrical stimulation in the hypothalamus, and sensory input provided by conventional food reward act on different substrates or utilize different pathways in triggering the reward-related electrophysiological phenomena. On the basis of results obtained by Routtenberg and Olds 52 (c f Olds47), a functional distinction can be made between the paleo- cortico-septal and the medial forebrain bundle reward substrates, since stimulation of the former has a pronounced quieting effect, and strongly suppresses both the aversive and brain stem arousal systems; whereas stimulation of the latter is strongly excitatory, increases the arousal level and behavioral output elicited by aversive stim- uli. Therefore, it may be suggested that the RCPV and PRS phenomena (as well as positive SP shifts observed during grooming activity) are triggered by influences that originate in the paleocortico-septal reward system rather than the medial forebrain bundle substrates.
SUMMARY
Nine cats were implanted with epidural non-polarizable Ag-AgC1 electrodes over the parieto-occipital and frontal cortex. In addition, similar electrodes were implanted in subjacent white matter of the posterior marginal, medial suprasylvian, and anterior ectosylvian gyri for transcortical recording. Two bone marrow reference electrodes were also used: one in the posterior aspect of the bone overlaying the frontal sinus, and another in the occipital crest. In 'acute' anesthetized preparations it was found that the transcortical recording was free from contamination caused by eye-motion potentials. The chronically implanted subjects, kept on 23 h food and water deprivation schedule, were trained to press a lever for 0.5-1 ml of milk reward.
Brain Research, 26 (1971) 305-332

STEADY POTENTIALS-REINFORCEMENT 329
Potentials caused by lapping or licking were recorded from an electrode implanted at the bottom of the frontal sinus or in the temporal bone. Eye movements were moni- tored from an electrode placed in the orbital bone immediately behind the eyeball. After 2-3 weeks of training, all subjects displayed bursts of high-voltage slow-wave activity of 5-9.5 c/sec over the primary and secondary visual projections during consumption of milk reward i.e., characteristic post-reinforcement synchronization (PRS). The PRS activity was always associated with 200-700 #V positive steady potential (SP) shift over the visual cortex in either transcortical or surface bipolar recording with reference to the anterior ectosylvian gyrus or frontal sinus. This SP shift was con- tingent upon the presence of light in the test chamber, although the subjects were trained and habituated to perform in the dark; the SP shift was also contingent upon the relative quality and taste of food reward. Therefore, it was termed 'reward contingent positive variation' (RCPV). Eye movements, which in several instances could be correlated with relatively small (30-60 #V) positive lambda waves over the visual cortex, did not significantly contribute to the RCPV. In satiated subjects during sleep onset or grooming activity a similar phasic positive SP shift occurred over the same cortical area; it was always associated with or shortly followed by a burst of high- voltage slow-wave activity of 6-8 c/sec; the latter SP shift was termed 'sleep onset positive variation' (SOPV). The occurrence of the SOPV was facilitated by the presence of ambient light, but, in distinction to the RCPV responses, it was not absolutely contingent on it. Two relatively small amplitude but rather consistent mirror image reversals of RCPV and SOPV responses were observed: one in the subjacent white matter 3 mm beneath the striate and parastriate cortex, and an- other at the surface in the immediate periphery of the visual projections, i.e., over the anterior portions of the ectosylvian, suprasylvian and marginal gyri. The phasic occurrence of SOPV immediately prior to the development of delta slow-wave sleep patterns resulted usually in a cumulative and more stable positive SP shift over the striate cortex. During the peak of a cumulative SOPV a potential difference of approximately 600-700 #V was observed, when measured epidurally across the parieto-occipital cortex between the anterior ectosylvian and posterior marginal gyri. This potential became more diffuse and partially dissipated with the development of slow-wave delta sleep patterns.
The PRS, RCPV and SOPV phenomena have been interpreted as processes sub- serving internal inhibition.
ACKNOWLEDGEMENTS
This work was supported by the National Institutes of Health Grant NB 06385; a preliminary report was published in Science, 163 (1969) 301-304; the results were also presented at the Annual Meeting of the American College of Neuropsycho- pharmacology held in San Juan, Puerto Rico, in December, 1968.
J. L. Y. and J. H. R. are trainees supported by the National Institutes of Health Grant GM 81-09.
Brain Research, 26 (1971) 305-332

330 T .J . MARCZYNSKI et at,.
REFERENCES
1 ALADJALOVA, N. A., Slow Electrical Processes in the Brain, Progress in Brain Research, Vol. 7, Elsevier, Amsterdam, 1964, 243 pp.
2 ARDUINI, A., MANCIA, M., AND MECHELSE, K., Slow potential changes elicited in the cerebral cortex by sensory and reticular stimulation, Arch. ital. Biol., 95 (1957) 127-138.
3 BOND, W., AND HO, P., Solid miniature silver-silver chloride electrodes for chronic implantation, Electroenceph. clin. Neurophysiol., 28 (1970) 206-208.
4 BUCHWALD, N. A., HORWATH, F. E., WYERS, E. J., AND WAKEFIELD, C., Electroencephalogram rhythms correlated with milk reinforcement in cats, Nature (Lond.), 201 (1964) 830-831.
5 CASPERS, H., Uber die Beziehungen zwischen Dendritenpotential und Gleichspannung an der Hirnrinde, Pfliigers Arch. ges. Physiol., 269 (1959) 157-181.
6 CASPERS, H., Relations of steady potential shifts in the cortex to the wakefulness-sleep spectrum, In M. A. B. BRAZIER (Ed.), Brain Function, UCLA Forum in Medical Sciences, No. 1, Univ. of California Press, Los Angeles, Calif., 1963, pp. 177-213.
7 CHIORINI, J. R., Slow potential changes from cat cortex and classical aversive conditioning, Elec- troenceph, clin. NeurophysioL, 26 (1969) 399-406.
8 CLEMENTE, D. C., STERMAN, M. B., AND WYRWICKA, W., Post-reinforcement LEG synchronization during alimentary behavior, Electroenceph. clin. Neurophysiol., 16 (1964) 355-365.
9a CREUTZFELDT, O., WATANABE, S., AND LUX, H. D., Relations between LEG phenomena and potentials of single cortical cells. I. Evoked responses after thalamic and epicortical stimulation, Electroenceph. clin. Neurophysiol., 20 (1966) 1 - 18.
9b CREUTZEELDT, O., ROSINA, A., ITO, M., AND PROBST, W., Visual evoked response of single cells and of the LEG in primary visual area of the cat, J. NeurophysioL, 32 0968) 127-139.
10 CREUTZFELDT, O., WATANABE, S., AND LUX, H. D., Relations between LEG phenomena and potentials of single cortical cells. II. Spontaneous and convulsoid activity, Electroenceph. clin. Neurophysiok, 20 (1966) 19-37.
11 FELDMAN, M. H., AND GOLDRING, S., Osmotically induced changes in brain steady potential and auditory-evoked response, Electroenceph. clin. NeurophysioL, 26 (1969) 588-596.
12 Fox, S. S., AND O'BR1EN, J. H., Duplication of evoked potential waveform by curve of probability of firing of a single cell, Science, 147 (1965) 888-890.
13 FROMM, G. H., AND BOND, H. W., Slow changes in the electrocorticogram and the activity of cortical neurons, Electroenceph. clin. Neurophysiol., 17 (1964) 520-523.
14 FROMM, G. H., AND BOND. H. W., The relation between neuron activity and cortical steady poten- tials, Electroenceph. clin. Neurophysiol., 22 (1967) 159-166.
15 GOLDRING, S., Negative steady potential shifts which lead to seizure discharge. In M. A. B. BRAZIER (Ed.), Brain Function, Vol. 1, Univ. California Press, Los Angeles, Calif., 1963, pp. 215 236.
16 GOLDRING, S., AND O'LEARY, J. L., Cortical DC changes incident to midline thalamic stimulation Electroenceph. clin. NeurophysioL, 9 (1957) 577-584.
17 GRANDSTAFF, N. W., Frequency analysis of LEG during milk drinking, Electroenceph. clin. Neuro- physiol., 27 (1969) 57-65.
18 GRAY, E. G., Tissue of central nervous system. In S. M. KURTZ (Ed.), Electron Microscopic Anatomy, Academic Press, New York, 1964, pp. 369-471.
19 GUMNIX, R. J., DC potential changes from auditory cortex of cat, J. Neurophysiol., 9 (1960) 667-675.
20 GUMNIT, R. J., AND GROSSMAN, R. G., Potentials evoked by sound in the auditory cortex of the cat, Amer. J. Physiol., 200 (1961) 1219-1225.
21 HACKETT, J. T., AND MARCZYNSKI, T. J., Post-reinformcement electrocortical synchronization and enhancement of cortical photic evoked potentials during instrumentally conditioned appetitive behavior in the cat, Brain Research, 15 (1969) 447-464.
22 HACKETT, J. T., AND MARCZYNSKI, T. J., Positive reinforcement and visual evoked potentials in the cat, Brain Research, 26 (1971) 57-70.
23 HILLYARD, S. A., AND GALAMBOS, R., Eye movement artifact in the CNV, Electroenceph. clin. Neurophysiol., 28 (1970) 173-182.
24 HESS, R., JR., KOELLA, W. P., AND AKERT, K., Cortical and subcortical recordings in natural and artificially induced sleep in cats, Electroenceph. clin. Neurophysiol., 5 (1953) 75-90.
25 IRWIN, D. A., AND REBERT, C. S., Slow potential changes in cat during classical appetitive condi-
Brain Research, 26 (1971) 305-332

STEADY POTENTIALS-REINFORCEMENT 331
tioning of jaw movements using two levels of reward, Electroenceph. clin. Neurophysiol., 28 (1970) 119-126.
26 IRWIN, D. A., KNOTT, J. R., MCADAM, D. W., AND REBERT, C. S., Motivational determinants of the 'Contingent Negative Variation', Electroenceph. clin. Neurophysiol., 21 (1966) 538-543.
27 KAWAMtrRA, H., ANO SAWYER, C. H., D-C potential changes in rabbit brain during slow wave and paradoxical sleep, Amer. J. Physiol., 207 (1964) 1379 1386.
28 KAWAMURA, H., WHITMOYER, D. I., AND SAWYER, C. H., D-C potential changes recorded between brain and skull in the rabbit after eating and drinking, Electroenceph. din. NeurophysioL, 22 (1967) 337-347.
29 KOHLER, W., NEEF~ W. D. AND WEOENER, J., Currents of auditory cortex in the cat, J. cell. comp. PhysioL, 45 (1955) Suppl. 1-24.
30 KORNHtmER, H. H., UNO DEECKE, L., Hirnpotential/inderungen bei Willktirbewegungen und passiven Bewegungen der Menschen: Bereitschaftspotential und reafferente Potentiale, Pfliigers Arch. ges. Physiol., 248 (1965) 1-17.
31 KOFFLER, S. W., AND NICHOLS, J. G., The physiology of neuroglia cells, Ergebn. Physiol., 57 (1966) 1-90.
32 KUFFLER, S. W., AND POTTER, D. D., Glia in the leech central nervous system: physiological prop- erties and neuron-glia relationship, J. Neurophysiol., 27 (1964) 290-320.
33 Low, M. D., BORDA, R. P., FROST, J. D., AND KELLAWAY, P., Surface negative slow potential shift associated with conditioning in man, Neurology (Minneap.), 16 (1966) 771-782.
34 Low, M. D., BORDA, R. P., AND KELLAWAY, P., Contingent negative variation in the rhesus mon- keys: an EEG sign of a specific mental process, Percept. Motor Skills, 22 (1966) 443 446.
35 MARCZYNSKI, T. J., Postreinforcement synchronization and cholinergic system, Fed. Proc., 28 (1969) 132-134.
36 MARCZYNSKI, T. J., The effect of photic stimuli on sleep onset and epicortical steady potentials during instrumentally conditioned behavior in cat, Brain Research, in preparation.
37 MARCZYNSKI, T. J., AND HACKETT, J. T., Cholinergic drugs and epicortical D.C. potentials during instrumentally conditioned behavior in the eat, Pharmacologist, 10 (1968) 204.
38 MARCZYNSKI, T. J., AND HACKETT, J. T., Post-reinforcement electrocortical synchronization and facilitation of cortical somatosensory evoked potentials during instrumentally conditioned appetitive behavior in the cat, Electroenceph. clin. Neurophysiol., 26 (1969) 41-49.
39 MARCZYNSKI, T. J., HACKETT, J. T., SHERRY, C. J., YORK, J. L., RICK, J. H., AND ALLEN, S. L., Diffuse visual input and cholinergic transmission determine the occurrence of 'Reward Contingent Positive Variation' (RCPV) in cat, Brain Research, 27 (1971) in press.
40 MARCZYNSKr, T. J., HACKETT, J. T., SHERRY, C. J., YORK, J. L., AND RICK, J. H., Unpatterned visual input and cholinergic transmission determine the occurrence of reward contingent positive variation (RCPV), Fed. Proe., 29 (1970) 252.
41 MARCZYNSKL T. J., ROSEN, A. J.~ AND HACKETT, J. T., Post-reinforcement electrocortical syn- chronization and facilitation of cortical auditory evoked potentials in appetitive instrumental conditioning, Electroenceph. clin. Neurophysiol., 24 (1968) 227-241.
42 MARCZYNSKL T. J., YORK, J. L., AND HAC~ETT, J. T., Steady potential correlates of positive rein- forcement: Reward Contingent Positive Variation, Science, 163 (1969) 301-304.
43 McADAM, D. W., IRWIN, D. A., REnERT, C. S,, AND KNOTT, J. R., Conative control of the contin- gent negative variation, Electroeneeph. olin. Neurophysiol., 21 (1966) 194-195.
44 McADAM, D. W., AND SEALS, D. M., Bereitschaftspotential enhancement with increased level of motivation, Electroenceph. clin. Neurophysiol,, 27 (1969) 73-75.
45 MORRELL, F., Microelectrode and steady potential studies suggesting a dendritic locus of closure, Eleetroenceph. clin. Neurophysiol., Suppl. 13 (1960) 65-79.
46 O'LEARY, J. L., AND GOLDRIN~, S., D-C potentials of the brain, Physiol. Rev., 44 (1964) 91-125. 47 OLDS, J., The mechanisms of voluntary behavior. In R. G. HEATH (Ed.), The Role of Pleasure in
Behavior, Harper and Row, Hoeber Med. Division, New York, 1964, pp. 23-53. 48 PETERS, A., Plasma membrane contracts in the central nervous system, J. Anat. (Lond.), 96 (1962)
237-248. 49 REBERT, C. S., MCADAM, D. W., KNOTT, J. R., AND IRWIN, D. A., A slow potential change in
human brain related to level of motivation, J. comp. physiol. Psychol., 63 (1967) 20-23. 50 REBERT, C. S., AND IRWIN, D. A., Slow potential changes in cat brain during classical appetitive
and aversive conditioning of jaw movement, Electroenceph. clin. Neurophysiol., 27 (1969) 152-161. 51 ROTH, S. R., STERMAN, M. B., AND CLEMENTE, D. C., Comparison of LEG correlates of reinforce-
ment, internal inhibition and sleep, Electroenceph. din. Neurophysiol., 23 (1967) 509-520.
Brain Research, 26 (1971) 305-332

332 T . J . MARCZYNSKI et ak
52 ROUTTENBERG, A., AND OLDS, J., The attenuation of responses to an aversive brain stimulation by concurrent rewarding septal stimulation, Fed. Proe., 22 (1963) 515.
53 ROWLAND, V., Steady potential phenomena of cortex. In G. C. QUARTON, T. MELNECHUK AND F. O. SCHMITT (Eds.), The Neurosciences, Rockefeller Univ. Press, New York, 1967, pp. 482-495.
54 ROWLAND, V., Cortical steady potentials direct current potentials in reinforcement and learning. In Progress in Physiological Psychology, Vol. 2, Academic Press, New York, 1968, pp. 1-77.
55 ROWLAND, V., AND GOLDSTONE, M., Appetitively conditioned and drive-related bioelectric base- line shift in cat cortex, Electroenceph. din. Neurophysiol., 15 (1963) 474-485.
56 RUSINOV, V. S., General and localized alterations in the electroencephalogram during the forma- tion of conditioned reflexes in man. In H. H. JASPER AND G. D. SMIRNOW (Eds.), The Moscow Colloquium on Electroencephalography of Higher Nervous Activity, Electroenceph. clin. Neuro- physiol., Suppl. 13 (1960) 311-320.
57 SADOWSr~L B., AND LONGO, V. G., Electroencephalographic and behavioral correlates of an instrumental reward conditioned response in rabbit : a physiological and pharmacological study, Electroenceph. clin. Neurophysiol., 14 (1962) 465-476.
58 SHVETS, T. B., (cited by Rusinov), Conference on Electrophysiology at Higher Nervous Activity, Abstracts, Moscow, 1958, pp, 138-140.
59 STERMAN, M. B., ROTH, S. R., AND CLEMENTE, C. D., LEG correlates of instrumental performance, Fed. Proc., 22 (1963) 399.
60 STERMAN, M. B., AND WVRWXCKA, W., LEG correlates of sleep : evidence for separate forebrain substrates, Brain Research, 6 (1967) 143-163.
61 STRAW, R. N., MCADAM, D., BERRY, C. A., AND MITCHELL, C. L., A simple cable for reduction of movement artifacts in LEG recordings, Electroenceph. din. Neurophysiol., 22 (1967) 90-92.
62 TSCHIRGI, R. D., AND TAYLOR, J. L., Slowly changing bioelectric potentials associated with blood- brain barrier, Amer. J. Physiol., 195 (1958) 7-22.
63 WALTER, W. G., Brain responses to semantic stimuli, J. psychonom. Res., 9 (1965) 51-61. 64 WALTER, W. G., COOPER, R., ALDR1DGE, V. J., MCCALLUM, W. C., AND WINTER, A. L., Contingent
negative variation: an electric sign of sensorimotor association and expectancy in the human brain, Nature (Lond.), 203 (1964) 380-384.
65 WALTER, W. G., COOPER, R., CROW, H. J., MCCALLUM, W. C., WARREN, W. J., ALDRIDGE, V. J., STORM VAN LEEUWEN, W., AND KAMP, A., Contingent negative variation and evoked responses recorded by radio-telemetry in free-ranging subjects, Electroenceph. din. Neurophysiol., 23 (1967) 197-206.
66 WATANABE, S., KONISHI, M., AND CREUTZFELDT, O. D., Postsynaptic potentials in the cat's visual cortex following electrical stimulation of afferent pathways, Exp. Brain Res., 1 (1966) 272-283.
67 WURTZ, R. H., Steady potential shifts during arousal and deep sleep in the cat, Electroenceph. clin. Neurophysiol., 18 (1965) 649-662.
68 WURTZ, R. H., Steady potential fields during sleep and wakefulness in the cat, Exp. Neurol., 15 (1966) 274-292.
69 WURTZ, R. H., Steady potential correlates of intraeranial reinforcement, Electroenceph. clin. Neurophysiol., 20 (1966) 59-67.
70 WURTZ, R. H., AND O'FLAHERTY, J. J., Physiological correlates of steady potential shifts during sleep and wakefulness. I. Sensitivity of steady potential to carbon dioxide, Electroenceph. clin. Neurophysiol., 22 (1967) 30-42.
Brain Research, 26 (1971) 305-332





























