Embed Size (px)
Citation preview

(
STIMULATION-PRODUCED ANALGESIA IN THE FORMALIN AND
--TAIL-FLICK TESTS:- A COMPARISON OF BRAINSTEM AND
FORE-BRAIN SITES IN THE RAT.
Michael John Morgan
Department of Psychology
McGill Uni versi ty, Montreal
A thesis submitted to the Faculty of Graduate Studies and
Research in partial fulfillment of the requirements' for the
degree. of Master of Science.
, .
(§) Michael Morgan, 1986
.,

--
t ~.,.""
.URIVERS,ITE MçGILL
. ~ . 'ACULTE DES.&unES AVANCf!S ET '1)! LA IlECItF.RèaE
..
~te _____ '~2~S~I/~C~~~!~~_b __ _
**HL'~Tam: ____ ~H~(è~~_A __ È_~ ____ ~~,V~M~N __ ~M~~~~~A~N~ ______________ ~~~f ____ _ DiP~: __ ~l~f:~~~~_c~~C~~~~_4~1~ ________ ~c~G~~~~!~: __ ~~~ ________ ~ ____________ __
TÎTI! ~E LA misE: _______________________________ _
1.
2.
j
.... Par la pd,-nte, l'auteur accorde 1 l'universitE McGill l' a~torisat1on de mettre cette thi.e l la/ disposition des lecteurs dans une b1blioth~que de McGill ou une autre blbllotbaque. soit sous sa forme actuelle, soit sous fonae d'une rEproduction. L'auteur d'tient cependant 'les autres droits de publications. Il est entendu, par ailleurs, que ni la thlse, ni le8 longs straits cie cett, thèse ne pourront être impr1mEs:' ou reproduit. par d'autres moyens sana l'autorisation Ecrite.d~ l'auteur.
La prE. ente autorisation entre en vigueur 1 la date indiquEe ci-dessus l moins que le CQIIitE exEoutif du conseil,n'ait votE de difUrer cette.date. Dans ce cas, la date diffErEe aera le .
;
, (
~ignature de l'auteur
Adresse permanente: • J. ~
--o
8ipature du doyen si un. date fiaure l 1'.linEa 2.:
(JDa1:lah on ravene) -'

, . .
J
~i:!, ,
, .
"
,
. -\ .
\
CI
1
. '
t
STlHUL'ATION-PRODUCEl> ANALGESIA IN THE FORHALlN AND J
TAIL-FLICK TESTS'" h '.!lrAltiSO" 8' 8_tl"ll3" Ulli
\'
1 .. '
J'

--
o
.. !"""._~.'1~. ---- _. -- -~._ ....... ~ ........ --... _--- "-~7- "T_~·~~"·""': -~.~ ----- ~ .. ><"·,\i~.·
_- .. ) , t
• ABSTRACT
..
The effect of electrical stimulation of brainstem and forebrsin
sites on t11e rats res'ponse to paTn was examined with the- taÏl-flick ,.
and formalin tests. The principal regions that supported strong
sti~ulation-produced analgesia (SPA), in both pain tests, were the
peri-aqueductal gray (PAG) and the, internal capsule (IC). The
_occurrence of SPA following stimulation of the lC was found to be )
strongly a8sociated with the occurrence of convulsive reactions to
stimulation. In a sample of lC sites at which stimulation elicited
'both SPA and convulsions, SPA was found to be accompanied by
catalepay. lt was suggested that ·post-i.ctal SPA from IC sites may be •
partially confounded with inhibition of mot,or responsivity to pain.
In both pain tests, across all sites, there was an association
between SPA thresholds and thresholds for aversive reactions to \ the stimul"tion. This association was interpreted as evidence that
SPA observed may represent a form of stress-induced analgesia!
r
)
-.
"
1
--~

o
- ..
,
o " , .. \
L;.~ .. ~_~._~.~~~ .... .1
JI L IX
ii
RESUME
, , , L'Effet de la stimulation. electdque locale du tronc cerebral e.t du
c
cerveau ant~rieur sur la r:ponse ~ la douleur chez le ra'f fut :tuc!i: lU
moyen des _ test s du l'et rai t de, la queue et de la Forma line.
prinC,ipales r~gions ou l'on rencontra une forte analg:sie
Les
post-stimulatoire (APS) ~taient la substance grise periaqueducale (SGP)
et la capsule int~rne (C!). L'apparition d'une APS suite ~ la
" "", 'stimulation de la CI fut demontree comme etant etroitement associee ~ .
avec l'apparition de convulsions eri ~ponse ~ la stimulation. Dana un
:chantillon de sites de la Clou la stimulation li:sultait en de l'APS
1 • 1 et des convulsions, on trouva que l'APS etait accompagnee par de la
1 , '\ catalepsie. Il eet suggere que l'APS post-ictale due> a des 'sites dé' la
CI peut :tre confondue, en partiê" avec une inhibition des r:ponse,s
• ""v 1 :' motr1ces a la douleur. Dans l,es deux tests, independamment du site de
. . \ stimulation, on avait une association entl'e les seui ls d' APS et de
'. IJ'" react10ft aver81ve a la stimulation.
)
J
: 214

~j ••
J ..
r ~.
t.~ .... :- .. _,:..
_... fIT- -l"-'--
iii
ACKNOWLEDGEMENTS • (
1 wish to tbank Dr. Keith Franklin and Dr. Ronald Melzack for
tbeir suggestions and support over tbe course of this research. 1 also .
wish to thank Dr. Francis Abbott and Dr. Andy Tasker for reading tbe
manuscript and Barry Connell for his expert assistance and d>ral
support. This research was supported by NSERC grant 8 A6303 to K. B.J.F.
and A7891 to R.M.
-. .
• ..

-'0 TABLE OF CONTENTS
Abstract . · . . · · · · Resume . · . . · · · · · . . Acknowledgement s . · . .
" List of figures · · · · . · . . List of tables . · · · ·
Introduction
Review of SPA in animals and man .,.,. . Pain tests • ~ • · . . . . . . Ph8rmacologi~l basis of SPA •
t>
Neural syste,s underlying SPA
Aim of the present study · . . · . . 1
Method
Subjects and materialB · · · Procedure · . . · · ·
" Design . .. . . . · · ·
· · · · . Data analys i s
Results
RistQlogy · . . · • · Group resul~ · . . . · · · Individual r~"lt •• · • · ,
Discussion • . . . . . . • · · • · · '. References • . . . . . . • ·
~O
~ ..
· l
· ii · Hi
· iv
· v
• 1
• 10
.13
• 16
18
20
· 21
· 26
t8
32
· 39
· 57
· 62
· 82
4"";'
/
A "'JUt@',1 ','
---:;-
" :~ . .,..

-t;> i ,ca • .. \
)
•
iv
.- "
LI ST OF FIGURES
• Il, l
Figure lA • 34
Figure lB ~ 35
Figure le 36
Figure ID .' . 37
Fbgure lE 38 / Figure 2 • 40 a_ ..- ...
Figure. 3 • 43
Figure 4 • 46
Figure 5 • 48 F' , 19ure 6 . ",. a • 49
Figure 7 52
Figure 8 54
F'lgure 9 61
,
. /-
o ...

.. f~------'-----------""!""'-lIIIt"'"""~-~---"'."""'-IIIII!I-II!I.IIJII; Il. 1119"'I!I. ------IIIII#IIIJ i o
/'
v
o ., ..
L"I ST OF TABLES 1
,. ..
'-Table 1 • 33
Tab le Z- '" . • 58
Table 3/"" • 60
/
o 'V

(
(
o
, L ":- ,'),
1
INTRODUCTION
St imulat ion-produced ana 1gesia (SPA)
The discovery, in the 1960's, that pain can be relieved by focal
electrical stimulation of the brain, is attributable to research within
tvo distinct disciplines - clinical neurosurgery and physi~'logical
psychology.
Stimul~tion-produced ~nalgesia (SPA) in humans was first noted in two
early applications of clinical brain stimulation. In the fi rst of these ~
studies, Heath and Hickle (1960) stimulated the brain for brief periode
vith electrodes that had been implanted to record electrical activity in
patients, most of whom vere 'schizophrenics. AlDOng their patients were a
fev non-psychotics with intractable pain who reported pain relief when
the septa 1 are a of the d iencepha Ion wa s st imu la ted.
Perha~s more credit must go ta Maurs, Roge and Mazars (1960), who ..... ,,'
not only independently discovered the phenomenon of SPA, but then
proceeded to develop it as a form of therapy. In the course of making
• 1e8ions in the midbrain to control chronic pain, Hazars and his
c01leagues noted that, vhen the rostral end of the spinothalamic tract
va. IItimulated ta provid,e physiological confirmation of the target site,
several patients vith post herpetic neuralgia reported that their pain
di sappeared.
The independent discovery of SPA by animal researchers had its
oriains in a study by Melzack, Stot 1er and Livingstone (1958). Melzack
and his colleagues accidentally discovered an area in the mid-brain of
the cat that appeared to be a pathway for pain inhibit ion. This finding
--, -- -

o
o
o
il" ,
2
led Reynolds (1969) to test the hypothesis that electrieal stimulet ion
of this region, the central tegmental-Iateral 'periaqueductal gray, might
au~!"ent the tonie inhibition of pain signaIs and thus produce analgesia.
His results indieated that stimulation did indeed produee analgesia that
was profound enough to allow surgery on awake rats with~~t any ehemical
anaesthetic. These rats remained eonscious, able to walk, and could be
disturbed by loud noises or sudden movements during stimulation.
Unfortunately however, despite a repLication of his results (Reynolds,
1970}, . .,these ob servat ions we re met wi th seept ici sm and were genera lly
ignored.
A recognition of the significance of SPA, by both the clinica!" and
basic researeh communities, did not occur until the independent
rediscoveryof the phenomenon by Mayer, Wolfle, Akil, Cardner and
Liebeskind, in 1971. This event provided the impetus for the subsequent
extensive research into SPA, in both disciplines. One of the fiut
issues examined by these ruearchers was the question of the specificity
of the effect t'o particular brain regions.
SPA from brainstem ,sites ln animaIs
The peri-aqueductal gray (PAG) region (including the dorsal raphe
nucleus) of the medial brainatem, has been confirmed as a principal
anatomical substrate for SPA in rat (e.g. Reynolds, 1969; Hayer, Akil
and Liebeskind et al, 1971; Ralagura and Ralph. 1973; Mayer and
Liebeskind, 1974; Soper, 1976), in cat (e.g. Liebeakind. Guilbaud,
Besson and Oliveras. 1973; Melzack and Melinkoff. 1974; Oliver .. ,
,

(
..
..
3
Besson. Guilbal1d and Liebeskind. 1974), and in rhe8u8 monkey (e.g. 1
Goodman and Holcombe. 1975; Ruda. Haye8, Priee, Hu and Dubner. 1976). <
Additionsl active 8ites have been reported in more rostral regions
ineluding. the periventricular gray (PVG). exçending to the pre-tecta1
region of the me.8o-dlencephalic junetion and the posterior hypothalamus
(e.g Hayer. Akil and Liebeskind et al, 1971; Hayer and Liebeskind, 1974;
Soper. 1976; ,Rhodes and Liebesklnd, 1978).
More recently, Fardin, Oliveras and Besson (1984, 1 & II) have
claimed that in the rat "pure" SPA, without any concomitant behavioral
signa of aver8ion to 8timulation, is restricted to a specifie portion of
• the PAG (ventral PAG). On the other hand, Prado and Roberts (1985) found
that ventral, a8 weIl 88 dorsal, PAG stimulation indueed aversion that '.
~ was correlated with SPA. -,Ooly /two sites ventrolateral to the PAG
. provided good SPA without concomitant behavioral signs of averS10n.
3PA from bra instem si tes in h~mans
" In man, ~timulation of the PAG, and its more rostral counter-part the
peri-ventricular gray (PVG). was undertaken to exa.mine -the, clinieal
utility of the diseoveries in basic researeh. Richardson and Akil
(1977s) vere the first to asseS8 stimulation st the8e sites in five
patients undergoing surgery to produce thalamic lesions •• Stimulation was
found to reduee chronic pai n.
ln a follov up 8t'Jdy, electrodes deligned for long term use were
_ implanted ioto the PVG of patients with chronic pain (Richardson and
Akil, 1977b). The same good pain relief that was previously observed in
brief intraoperative episodes of stimulation ensued in 6 out of 8 .'
•

, (
o
o
o
4
patients. In the same year, Hosobuchi, Adams and Linchitz, (1977) .. reported pain relief in 6 patiénts stimulated at PVG and Pite situ.
More recent cl inical studies have corr<1borated the efficacy of
/
brainstem SPA. Over a"9 year period, Hoaobuchi (l9~O) obtained relief
from pain of· peri pheral origin in 16 of 22 pat ients wi th PAG .)
stimulation. Simi18rily, Plotkin (1982), reported that 38 of 48 p.tie-nts
with peripheral pain syndromes responded weil to PVG stimulation.
Two important, attributes of SPA from human PAG/PVG sites should be
noted. Firstly, the above studies indicate that stimulat.ion of these
sites is most effective for peripheral p~ins of noèiceptive origine
Secondly, the degree of analgesia produced lS comp~rable vith that
obtained from a moderate dose of morphine. The surgical ~nae~5Jt~tic , level-"chieved in animals (Reynolds, 1969) has not been reproduced in
hu·mana.
SPA trom diencephalic aite. in humans'
Following their discovery of an SPA effect in man, ..taurs, Merienne
..,
and Cioloca (1974) conducted a fo 110v up atudy in t,lhich elect rode. vere.
implanted in the (sensory) nucleus ventralia posterolate'ralis (VPL) o(
thalamus. The electrodes ~ere left in place for 88 10ng.88 si" week ••
Between 1962 and 1972, 17 patients with chronic pain responded vell to
this treatment.
A~ electrophysiological. fi.nding by Richardson (1970) prompted triat.
by Hoeobuchi, Adams and Rutkin, (1973) to st imu late the VPL for the
control of anaeathesia dolorOI8 of the face. Initial .ucc~ .. e. led the_
.'

"'
c.
(
)
o
..
" br,,,,· ..... '~d-L ;. a,
5
ta implant electrodes for long term use in 5 further patients, 4
report~d pain relief whenever they used ~he st imulator.
More recentIy, Mazars, Merienne and Cioloca (1979), stimulated the
VPL ta provide good pain relief in 83 of 93 patients with
deafferentation pains. Turnball, Shu Iman and Woodhurst (1980), treated
18 patients vith pain due to sensory pathway damage by stimulating the
sensory nudei of thalamus. Complete or partial relief,occurred in 12
patients. Seigfried (1982) found stimulation 'Of the medial counter-part
of the VPH, nucleus ventroposteromedialis (VPH) produced pain-relfef in
8 of 10 patients vith 'post-herpetic neuralgia of ~he face.
These studies demonstrate a diencephalic substrate for SPA 'in humans,
concentrated at the sensory nue lei 'of thalamus (VPH and VPL), also kno,wn . ~ . as the vent robasal "thalamuB (VBT). They <Il 18 0 indicatè that clinical SPA
from this site is especially effective for facial and deafferèntation ',,
pains.
..
SPA from ventr'obasal thàlamue in .nimals 1
In contrast. ta' the clinical finding8, thet'e is no ct)nvincing evidence
Chat the VBT ia an effective 8ubstrate for SPA in animah. Richardson
(1970), demonlt rated that stimulati'on of this region in the cat could"
black evoked reeponus ta nociceptive acimulatoion re~orded dt, the ~dial '.
t'halamic nucleue.. Tsubokawa, Yamamoto, Katayallla and' MOriy~8U, (1982), •
.h~ved that Iti:mul,tion of the VPL in cats accivates raphe spinal
neurone as powerfully as PAG st imulat ion ~es. Similarily, Willis,
Gerhart, Willcocksol'l, Yflrzierski ,nd Cargill, -'fl984) demon.trate~ ~
.-xcitation of raphe spiql neurons folloving ,sti .. ulation of the VPL in ... .
. .
"
'.
"

6
o monkeys.'Fi~lly, Dickenson J1983), demon.trated that
in the rat could powerfully inhibit the activitie. of neurone responding ,
pnly,to noxious .stimulation.
However, aIl of these studies relied on an electrophYliological
.meàsure, rather than a beha~ioral test for araalgesia. Furthermore, they
aIl examined neuronal xeaponsivity in the anaethestized animal
(anaesthesia is known to alter neuronal responsivity: e.g Bowman and • • •
,Ran~, 1980). In one of the few behavioral studie~ ,that incidentally ,.
included some electrode placements in rat VST, Hayer and Liebeskind ,/
,
(1974), found ttlat none of - the 5 rat S ln quest ion~hibited SPA.
Similarily, Goodmaft and HQlcombe (1976), reported the results fot' the
sti~u1ation of J elE!ctrod~ in rhesus monkey VBT. Again, this site did
o . not produce an appreciable SPÂ effect. In.a recent study, Prado and
Robert,s (1985) inr:luded 3 electrodes ~n' rat VBT. Stimulation d-id not
produce analgesia at any of these sites (although significa~t .version
to stim\Jlation was noted). Only Balagura and Ralph 09!3), reported 'any
direc ln animal VBT. In this cale 'Only moderc.r-. SPA , .
was f t the single electrode placement tested.
, . ,.'
SPA fr internaI ca sùle in humans ,',
Enco~raged by. their initial clinic~,l SUce88 vith diencepalic (YBT)
stimulation, Adams,' H080buchi and Fields (1974), explored the
..1
po88ibility that simi1ar stimulation of the ~omato.en.ory neurone in the
retion of the po.terior limb of Che internaI capsule (IC), mi,bt provide ,
pain telief. Thil, region compd'el a co.pact bând of fibru chat radiae.
o from the, thalamu .. to cortex.and provid*, reciprocal cdnn~ctionl b.t ••• n
. ,

\
, 1
o
7
the tvo are .. (Carpenter, 1972). They found that patients with lesions
of the centrai nervoua aystem could be succeufully relieved of
.. soc iated severe spontaneous pa i n by IC st imu la t ion.
Maurs, Herienne and Cio loca, (1976) refuted the Adams et al (1974)
finding. In reference to a recent study in which the y had shown
aigni ficant SPA in the VPL, they stated that "1 t ia our opinio!, that the
electrodes placed by Adams et al~ in cases of chr,?nic pain were in the . ,
VPL.and fiot iri the intert1al capsule". Their skepticism of le a8 an
effective substr~te for SPA, continued,. so that in a 1979 paper, Haurs
et al, still 88serted that "it is ou'!- bpinion that the beneficia1 cases
of internai capsule stimulation shou1d be set to çhe credit of
stimul .. tion of the VPL" - (Hazers et al, 1979). . ,. Despite this skeptifiam concerning the find,ings of Adams et al,
further evidenèe for posterior IC as a locus for SPA was. forthcoming.
Cooper, Upton and AUJ'in, (1980) reported that sfimul,tion of the
polterior limb of the IC produced complete re'lÏef hom deafferentation~
pain in one patient. Turn6ul! (1982) found the lC to be an effective
1 -
.timulation site for two patients suffering fr'om thalamic pain. A more Il;>
extensive clinica1 study by Namba, Nakao, Sakutai, HatsulD()to, Ohmôto and
NÏlhimoto, (1984) demonstrated that stimula~ion of IC produ.,ced relief , '
frOID thalami~ pain, in 5 of 6 patients. That the le is no longer
considered to be a disputed site for SPIln humans is' cIear trom a
.tateaent by Richard Ion (1985):-- "l n generaI, the interna 1 capsu le ia .,
bitter targe.t site than the laterai thalamu8-, because it doea not '
produce •• n,ory los. vith electrode placeaaent, and it leems to produce
bitter pain relief".- It is important to note, hovever, that the ab ove
, .

o
,
"
o· t"
o
t..: "
studies indicate that clinica1 IC stimulation'has been u,ed to
particulsr advantage in the- é1-eatme~t of pain «;>f central origine
. , SPA !tom the internai capsulé in animaIs
. In 1983 in Ylliologic8l study, with cats, Nakao, teported
that neuronal activity in H of the thalamus evoked by contralateral
tOQth pulp stimulation was inhibited"by fipetitive stimulation of the
8
le. In a follow-up st'udy, Ni§hintoto, N.!lJDba, Nakao, HatStllll\to ,nd Ohmoto, ,J
O~84) 'repli~a'ted this finding.
As in the caslE! of the YBT electro~hy8iological IItudie.a, nowevér, i
thesé studi~s do not provide direct evidence that the IC is an effective ,
substrate for SPA in animaIs, as well aS humens. Neither IIta,ady employed
a behavioral test of analg~sia and both were eonducte-d ~ith ,>
anaesthestized cata. 'l'hua they both suffer from the lame Ihortcomings ..
their VBT counterparta. Currently, the question as to the effieacy of, , ,
the le .as a substrate for SPA in animal,." remains unanswered.
the rat as a model of. clinicai SPA ( ,
. .:From the above review, it ia cleu that a functional homology, ih
terms of SPA, exista between PAG, sites in man and r~t. It woù1d eppe.r,
therefore, that at this brainstem site, the rat is a use fuI model of
~linieal SPA. However, th,is, functional ho'mology does not lIeem to extend
to late!;.l diencephalic st ructures. Both the VBT and the le have proven
to be clinieaUy. effective substrate. for SPA. In eont rait, there 'is
very little behavioral evidence th~. the- YBT i, an' e,ffective 'site,'for .
:a
r •

\. \ \
(
o
)
,-9
SPA in the rat, and the issue of the efHcacy of rat le as a subst rate
for SPA has not been addr~ssed. ~
The main aim of the present study was to explicit 1y- adClress the' issue
. of the suitability of thé rat as a ~del of cli.nical SPA at' both /'
brainatem.and lateral diencephalic sit~s. Of the latter, the le was the
main focus of, interest, both bec8Ui8e of the controversy about the
clinical significance of this site and because it remains unexplored as ~
a aubltrate for SPA in the rat. Should SPA in the rat prove to be a ... suitahle model of clinical SPA at eirher, or both. of the two lateral .
, "'
dienc~phalic sites to be explored (VBt and le) an analysis of the
mechanisma of SPA would be facilitated.
The problem of pain asseaament in animals
In ex~loring animai modela of SPA a major problem is the aasessment
of pain. In the clinical ~tuation pain arises from injured or diseased
tissue and parn 'can be' a1l~essed from verbàl report 8S weil as behavioral l'
observation. Both ethifal a,pd practical c~nsiderations preclude a , .
complete duplication of theae clinical atates in animal subjects.
Inste4d, animal SPA must be examined with pain that is neither too
severe 001' too prolqnged to be ethically unacceptable. However, the
po •• ibility arises that the painful state induced in an animal subject
" ... y be of '. type onrdated to painful states present in clinical studies
/
of SPA. Therefore, telt induced pain in animals may not be affected by
~. brain Iti.ulation in the ume way as cliJlical pàin. A brief review of
SPA in teras of animal pain tests and the pain ~pes they measure ilia,' ,
clar if y thÏl illue. \. ( ~ 1
l~ , .,'
..... ~IiIilIII~ ...... ' .. ' , _____ .:..," ..... ' -"'"' ......... '"'-............. ;, .... ',~-"'~ -' -~ •• ~. ..

' .. !j,
't
o
o \
o
• • 10
Pain tests
The response to à vide range of noxious stimuli appears to be
-inhibited by SPA in animale. t·lectrical st imulat ion of medial brain
... "l.
structures has been dhown to inhibit responding to:- painfu1 electric . shock (e.g Mayer, AldI, Liebeskind et el, 1971)', electrieal stimulation
of the tooth pulp (e.g Seule, Dubner, Greenwood and Lucier, 1976),
pinching of the extremeties (Goodman and Hole6mbe, 1975; Liebeskind ~t >
al,o 1973; 'Hayer, AkH: Liebes.kind et al, 1971; $oper 1976) and pinprick
(Balagura and Ralph, 1973; Soper, 1976).
ito~ever, two other pain tests have proved to be the most popu laI' 1.n
SPA basic research, these are the tail-fliek tes~ (D'Armour and Smith,
1941) and the hot-plate test (Woolfe and Macdonald, 1944). Both testa-
measure the withdrawal response to thermal pain. In the former, a rat's
tail ia stimulatéd by hot water (a~proximately 55 degrees) or radiant
heat, and pain is inferred when the rat, reflexively, flicks its tait
8side. In the latter, the 'rat is placed on a hot p'late (approximately 52 • - J
degreee) until it lieks its paws or jumps up, signalling that it ia
experiencing pain.
These,threshold level pains, are very different from the intense,
p;'olo'nged type of pai~ typically auociated vith tiseue damage or
pathology. Dennis a~d Melzack (~979) argued that thi, difference May be 4
qualitatl.ve'as wri;. aIS quantitative. In 1968, Beecher had found that
laboratory paitT: in human., produced by pricking, pinching or radiant .
beat, appeared to be immune to the specifie an.lge.ic effect. of
,morphine. Beecher overcalle tbJa dift'iculty by uling a .tbod that - .
, ' .. , '
•

11
involved exer~âing an ischeamic limb. The steady, burning, pain this / '
produced val readily lusceptible to morphine analgesia. Thus, 'the
analgesic action of morphine appeared ta reveal a distinction betveen
s~ort, sharp pain and more prolonged pain associated vith in jury. If
this vas the case, then the animal researc~ers were limiting themselves ,
to pain telts that only measured other types of pain - the less
clinically significant forms that, are 1e88 susce'ptible to morphine -f
. " measured by the tail-flick and hot-plate tests.
Melzack and his collaborators set out to redress this situation by
creating ail' animal model of more prolonged, "burning" pain, resulting of'
from localized tissue damage. The resultant test was published by ,
Dubuisson and Dennis in 1977 under the title of the "formalin Test". The
test consisted of in je ct ing a small amount of di lute formalin under. the
skin of the rat's paw, and then recording a distinct set of behavioral
res~onles that were assigned numerical values.
ln 1980, Dennis, Choiniere and Melzack, 'conducted a series of studies
which demonltr.ted Chat the formalin test did, indeed, reveal different
components of pain. TWey reported that when tbe PAG'was Itimulated, much
le •• ëlectrical current wal required ta produce analgeaia in the
formalin telt than in tail-flick and hot-plate tests. A pharmacologieal
.... analyais indic.ted that the specific cpiate ancagon~st, naloxone, did
not reverse formalin test SPA a. it typically does in. the t.il-flick
teat. Further evidence fat a phar~cological dissociation betveen the
. forulin teat pain and tail-flick test pain wa' provided by Abbott, ...
o Franklin, Ludvic~ and Meback (1981) who found rapid and strong . .
tolerance to .,rphïne analguia in the tail-flick', test, but littl. or no
..
Q
iL...,~" .... '

----..
12
(
o tolerance to morphine in the formalin telt. Dennis and Helzack (1979) -coined the terme "phasift" and "tonie" pain, respeetively to eharaeterize
the qualitative distinction betveen transient tai1f1iek test pain and
the continuous pain of the formalin test.
Pha,sic versus T9nic SPA in the rat; ,
A subsidiary aim of the present study was to examine the l'ole of pain
type in determining the strength of the SPA effeet at each of the rat
brain target si tes (PAG, le and VBT). From the reviev above it ia cleu , .
that the majority of' animal SPA studie8 have been limited to an
evaluation of "pha8ic" pain (i.e tail-flick te8t pa-in). In eontrUt molt
clinical SPA studies are concerned with "tonie" SPA in the form of
o relief from the pain of tissue damage or pathology. Since the preaelU
s~u.dy sought to compare clinical SPA vith SPA from correaponding lite.
in the rat brain, a behaviora1 mea.sure of toqie SPA vas required. The
formalÏn test vas seleeted 'for this purpose.
• • • The tatl-filek test, vas seleeted as a second measure of SPA to allov
a direct compal'ison of the results of this study vith other animal
studies. More important 1y, uti l ization of both the ta il-flick and
formalin tests prov~des the basis for an examination of the .uggeltion
. (e.g Abbott and Melzack, 1983) that the8e tvo tests 'revea1 a
.\ dissociation in the pharmacologtcal and neural syltems underlying SPA.
if this dissociation is common to both rat and man, then a comparative
analYli8 of the tvo pain testl .provides a quali tative,. as vell . SI a
quantitative, measure of the luitability of the rat a. a IIIOdel of
0 cl iaical SPA at lateral dieneephalie .nd br.inecc!. Itiaulation ait ••• t'
An ~
,
--- , .;

o
o
13
evaluation of ti}e hypothesis t.bat pain tests can differentiate pain ,
typee that are mediated by distinct neurochemical/neural systems,
~equires a brief review of the current understanding pf ~he
pharmacologics 1 and neural basis of SPA in' rat and man.
Pharmacologicsl buis of SPA
(
imal etudies •
SPA exhibits several striking features that parallel morphine .. analgesia. One set of studies (e.g. Berz, Albus, Met ys, Schubert and
Teschemacher, 1970) showed that micro-injection of morphine i~ the PAG ,
produced significant analgesia sU8gesting that morphine and SPA may have
a, ehared site of action. Samenin and Valzelli (1971) showed that
sub-analgeeic doses of morphine combined wLth sub-analgesic PAG
stimulation, synergise to produce significant SPA. Other parallels with
morphine analgesia are that SPA, may depend upon the integrity of. (/
serotonergttc transmission (Aki 1 and Mayer, 1972), ,tolerance develops p
with SPA ~nd, more significantly, cross-tolerance between SPA and
morphine analgesia occurs (Mayer and Baye~. 1975). Final1y, Akil, Mayer
and Lieoeekind (1976) showed that (tail-flick»)SPA is partially reversed
by na loxone.
A neuroc~emical basis for a link between SPA and morphine analgesia
va •• uggested when Hughes""'(1975) discovered an endogenous, morphine-like
factor in the brain (tel'1Ded "enkephalin':), described its peptidic nature
aad .ynthe.ized it (Hughee, Smith, Kosterlitz, Fothergill, Morgan and
Morri., 1975). Intta-ventricular micro-injection of this substance was -
thea .hova to produce poverful analge.ia ia the rat {e.g. Belluzi,
. , .
\

o
o
,
14
Grant, Garsky. Sarantakis. Wise and St'ein. 1976; Malick and Go1datein,
1977; Pert. Simantov. and Snyder. 1977). A second type of opioid peptide
• - endorphin - was found in the pituitary gland of the pig (Teschemacher.
Blasig and Kromer, 1916). This was a1so demonstrated to have analge.ie
act ion when micro-injected int ra-ventricu 1ari 1y. Finally. elevated ,
levels of' endorphin-like substances were found to be assoeiated vith SPA
in the rat (Akil, Watson and Barchas, 1916).
These findings suggested to Liebeskind and Paul (1917) that an . ,
( endogenous substrate of antinociception existed vithin the medial p
brainstem. They assumed that stimulation both activates this substrate .
directIy and also indirectIy via the release of endogenous opioids
(opiate-like peptides). Hovever, ~his elegant concep\ion was primarily
based on experiments that used the tail-flick test~
As indicated above, Dennis, Choiniere and Melzack (1980) reported
that th~ neurochemical systems underlying formaiin test analgesia are
not the same as those involved in the tail~lick test. ,
In contrast to .
typical tailflick test SPA. formaJin test SPA from this site was not
reveraed by ~aloxone. Thus. in this case, brainstem SPA .doe. not appear
,to be mediated by opioid neurotransmitters. More generaIly, this
suggests that the involvement of nellrochemical .ystems common 1:0 both
IPA and morphine afia1.gesla, may be determined by pain type.
o • The relation.hip between' systems activated by stilllulation, and the
type of SPA they mediate, may differ vith other rat brain .timulation
site.. Nevertheless, the .evide"'ee for IIOre than one. phanueololically .'
distinct, type of SPA, at brain.te. _he. ,uI8e~t. 'Chat mu than ODe
o

15
type of pal.n test LS necessary for a thorough examination of the rat as
a model of clinica1 SPA.
Clinical studies
It h;s been shown that stimulation of the PAG and PVG in humans, at
intensities sufficient to provide analgesia for chronic pain of
periphera1 origin, is an effect that is reversed by naloxone (Hosobuchi
et al, 1977>. This parallels the na1oxone reversability of (tail-flick)
SPA in the animal PAG and PVG, and with the evidence for elevated Beta
endorphin leve1s accompanying human SPA in this area (e.g. Hosobuchi,
R08sier, Bloom and Guillemin, 1979), it wou1d appear that endogenous
opioids play an intermediary role in clinical SPA. However, there is
some evidence that conf1icts with this hypothesls.
Whi1e PAG stimulation in humans e1evates the concentration of Beta
endorphin in ventricular fluid, le stimulation does not (Hosobuchi et
'al, 1979). Similarly, cross-tolerance between SPA at the PVG and
'narcotics develops readily (Hosobuchi et al, 1977) but has not been
observed wi th stimu lat ion of the somatosensory pathway. Tsuboka";a et al,
(1982) noted a lesser ine rease in ventricu Ur Beta eadorphin in 6 '
'patients who received VBT stimulation, than in 0Fhers who were
stimulated at the PAG. This corresponds well with this teams
experimental finding that PAG st imulation-induced excitat ion of r'aphe ",
~ c. neurone 11 reveraed by naloxone, while VBT stimulation-induced
• excitation of raphe neurone is not. Thus, it appears that stimulation of
YBT and 'IC in humane scti vates neural mechanisGls that do not involve
\

o
1\ o
• • ./
endogenouli opioids, in contr8st to the system activated'by stimulation
of PAG and PVG sites •.
Neural systems underlying SPA
In a recent paper, Basbaum and Fields (1984) outli-ned eurrel)t
undetstanding of the intricate structure and funetion of brainstem
descending systems. Descending fibres from the PAG projeet to the more
caudal nucleus raphe magnus (NRM) syste; and surrounding nuclei in the
rostral medulls. These nucl~hen projeet, via the dorsolateral
funiculus, to the spinal dorsal horn, where they inhibit nociceptive
neurons. The inhibitory action at the cord may be via 'direct
16
post-synaptlc inhibition, or via an opioid peptide containing, endorphin
interneuron. There are thought to be other endorphi nergic inputs st the
PAG and rostral medulla levels. An important nor,adrenergic system ia . inwlicated in the excitation of t~~e NRM system at the rostral medulla
level., Serotonin is be 1 ieved ta be the main neuro-t ransmitter for the
NRM - Spinal, descending inhibitory pathway.
The basic structure of "the PAG descending inhibitory system has been r-
;' known for some ti~e (e.g. Basbaum and l'ields, 1978). ,More recently. it
has b~en demonstrated that the PAG la.. pivotally located to transmit
cortical and dien~ephalic inputs to the lower brainstem. For ex.mpl~,
Beckstead . {19791'reported 2 part iculari ly interesting cases (FC8 .nd
FClO), in a retrograde tran'port study in rats. Labelled amino acid VII ,
deposited in the lateral portion of the rat pre-frontal cortex (PFC).
Labelled fibres we re found to project caadally via the media 1 exten'l: of
t'he IC. and separately, via the medill nucleu. of thalamul, ta the
\

c
•• c
o
17
dorej.t. hypothalamus and on through the ventral PAG to lover brainstem • .. ~
This FYC - PAG descendlng psthway led Hardy and Hsigler (1985) to ~
suspect that PFC might provide an effective, rostral, substr.ate for SPA
in the rat. ln an initial electrophysiological study. these werken
found that midbrain neurons altered their fi ring pat terns in response to
PFC J. imulat ion. Horeover, the majority of nocicept ive neurons vere ~ \ . ~I es sed and micro-iontophoretic administ rat ion of met-enkephal i n and
, noradrenalin st the PAG leve 1, mimicked the effect of PFC stimulat ion to,
varying degrees. In a follow-up study Hardy (1985) used tail-flick and
hot-plate test sand demonstrated that PFC st imulat ion did indeed produce
'" analgesia in rats.
Although 'there is no direct evidence that lC is an effective neural
/
substrate for SPA in rat s, these Btud ies suggest that the lC may b,Ç part
of a descending (excitatory?) system that projects to the lover ,
brainstem. From the work of Tsubokswa et al (1982) it is possible to
apeculate that this input't1l.en activate8 the raphe nuclei, either
directly or indirectly via enkëphalinergic and noradrenergic neurons,
tbereby activating the descending inhibitory system. However, it is
important to note that PFC st imulation has only. proven to produce '"
an.lgesia in phasic pain tests. 1
There ie increasing evidence "'that when the tonie pain of the formalip
teat is studied. ascend ing projections to the fore-brain are more'
important than/descend ing projections tc? the spinal cord. Destruct ion of
the NaM or the caudal PAG attenuates analgesi. prodOced by morphine. or
by Itiaaulation of the midbrain. in the tai l-fl ick test, but bas no
.ffact on ati.ulation-produeed. or .,rphine analaesia in the farmalin
\. ,
•

o
o
0
, 'oL L:: .•
c ..
test (Abbott, MehacK, Sa'mue1 et al, 1982; Abbott and Mel,zack, 1983>, Furthermorè" destruction of the median raphe nucleus (KR) hu no effect
in the taÏl-flick test (Buxbaum, Yarbrough and Carter, 1?73; Lorenl and
Yunger, 1974; York and Haynert, 1978; Abbott ànd Helzack, 1982), but
18
caU8f!s either a potentiation of morphine analgesia in the f,Mma 1 in t~lt,
or possibly a reduct ion in the' amount of pain, prod~ce~ br ye fO[1II.Ilin .
(Abbott, Melzack et al, 198.2; Abbott and Helzack, 1983). The MR, unlike
the NaM, is known to project rostra11y to fore-brain structures (Conr~~,
Leonard and Pfaff, 1974; Bobillier, Seiguin, Petitjean, Sa1vert, Touret
and Jouvet, 1976), ..
In conclusion, current knowledge o( the functional anatorny underly.ing
• SPA suggests that tai I-flick SPA is mediated by the activation of
descending systems. ln contrast, formalin test SPA appears to be
mediated by an effect of st imu1at ion upon the ·conduction of nociceptive
input via med ia 11y ascending systems.
Aim. of the present atudy
T~e present study explores the efficacy of a' series of rat brain l
The fi rst area to be examined, the 1 regions a8 possible si tes for SPA.
peri-aqueducta·1 grey region of the mid-bra i n (PAG),. VII se lected bec.ule
it ia an establÏshed site for SPA in both the animal and cl ini:cal
-literature (Bee above for a reviev). A replication. of this property,
) . would serve to confirm the appropriateneas of the et i.u1_t ion par ... t.r. . . and procedul;'e emp10yed in thie Itudy. Secondly, it va. ',xpected that
'\ .f If .. ..
.:
- ~

c
. - .
1 \
(
o
. "
r' 19
.ti~ulAti_.()n of this, site would produce andgesia wllich could be compared ,
.'witft analgesic effects, if any, obtained from ot-her stimulation sites. , . • J
The posterior- limb of the internal capsule OC) was the second site
tb be explored. There ie s~me, somewhat controversial, evidence that
1:his region is a site for clinical SPA (see above for a review), but_
. there bas bElen no research into'l:his question in the rat.
The ventrobasal thalamus (VBT) comprÏslng tbe two v'éntro-pqsterior
nùclei (the VPH and VPL) of thalamus, was the third target site for SPA
expl'orat ion'. The VBT is alao relatively well established as a substrate ,
for SPA iri.c.linical etudies, but there is mea.gre evidence for a similar
potent iaI .in anima·~ etud ies (see above for a review). -..
The princiJ1al objec~ive of this study was, therefox:-e, to determine i!
t~ rait can be regarded as. a~ appropriate 'animal model of clinical SPA
at thue ,two diencephalic' loci (VBT and lC). Thesè regions occupY 'he
same plane perpendicular toJ
the anterior/JlOsterior axis, in the rat-
brain. The VBT 18 situâted media 11y to the le, and is separated from it
by • relat i vely narrow border region comprising 1) the medullary lami na
"" of t;he VPL, (ML) and 2) the zona incerta (Zl). The relative proximity of
the VBT and the- le, in the saDie pla~e, provided the pos sibil i ty of
c,l.rify~ng the. role of spread of , current between adjacent st ructures.
Cinaulate cortex (CING.) was selected a~ t'he fourth and' fi~al target
lite because of its l .. ck of association vith neural system! thought to
underlie the SPA effect. Furthermore, A,bbott and Helzack (1978) did ~ot
find SPA in a .ample of 5 rats stimulated at thi. site.t. Thu. t thia area , -It.-.... --judaed to provide a suitabie_ èontrol for .ny generalÏsed ~ffects of
Itiaul.tion, in order to elucidate the .ite specifie effects.
1
' .

o
o
....
o . , ,. j
•
.
- o
-20
METlIOo. ,
SUBJEC;rS
Subjects were 64, naive, male, Long Evans hooded rats which weiahed ...
275 ta- 300 grams on arrivai. They vere housed 4 ta • cage in, the .,n,imal
c<?lony room, for 3 days prior ~o surgery, with free accels to food and' . .. , ,
vater. , .
W:U:R1ALS
Stimulation apparat'us. j _ ''>;.,-
Stimulation "aB provided by a Grass SS8 stlUlulator '(model ,S88B) via ( ,
a Gru:;8 constant current unit (model CCUIA). Current "as me.aurecl by an'
oscilloscope displa,ying the voltage 'dr~p acro~s a 30 K ohm reaÏltor in
series with the ra~.
For ~dQliniJtration of stimulation rets were placed in the 32 x 32 x
32 J)lf'xig1asa box used for fonaalin. cesting. This app.ratua vas fitt,d
with a plexiglass lid (vith a slit for eléctrode leads), t.o prevent •
esc'ape. A stopvatch vas used to determine total Itiaulation'duratien and
,post st imulat ion intérvala.
.
Tait-Pl ic~ tes't apparat.à.
A thet-mostatically controlled "ater bath VII uaed to keep "eter (the
thermal stiQiulus), at a conatant tellper.ture (55, + or - 0.2, deare ••
C.).' A 500 ml pla8ti~ be.ker '!Jas used to s •• ple ".ter for e.ch t •• ~. An - , ·electron.ic IIÜlisecond tiller, vith foot avitch, VII u.ed t?b .... ure CaU
flick latel'cies.
\ , ...
,-
\

Ci
,-
c
-")
L
)
o.
!rh 5
'7 -~
0
c'
..
Poraalin test .pparatu. -', t
Rats in the for_lin test con~ition , vere observe'd in -the large
plexiglall box in vhich they had received stimul.tion. Bel\eath the
tran.par~nt floo'r, a large mirror vas 1IIOunted at a 45 degue angle,
, permiiting unhi~dered observation of the racls )lMis. A TRS80 (Radio
Shack, Pe2), micro computer vas employed to record and calculate
formalin test scores. 'VIII
CatalepaI test apparatua
21
. . Apparatus for the cata lepsy:-test comprised .a steei bar (l0 cm long
and 5 I11III in diameter>, which was sl,lpported 'firm~y, 10 cm above, and
hori~opta1 to, the surface of a table. (>
< •
PROCEDURE
• Surgery
Bipolar electrodes (Plastic Producta, Roanoke; Va.) vere cu't to
lenlths appropriate for their target sité •. The exposed tips ~t th.? cut
. end of the electrode vere aeparated by .2 an. After the rats vere • \1'
Inaeathetizéd with sodium pento-barbital (60 mg/kg IP), the electrodes , ,
vere i.planted using standard stereotaxie techniq~es. The ~oordi~ate8
for eleetrodé placelllenta vere talten from an atlas of t~e rat, brain
(Paxino. and Watlon, 1982). CoorcHnates relative. to the midline of the
~ ,) 'd • .. ... ,+11 an the lnteraural line were aa followl: ~ • -6.5 1l1li APt -0.1 1l1li
Lit •. -5.~ DV; !Q lot -3.5 1l1li AP, 4~O am Lat, -6.5 _ DV;.'y!! • -3.5 am
AP; 2.75 _ Lat, ~6.5 ... DV; CING. • 1.2 11118 AP" 1.0 1IID Lat, -1.0 _ DV.b
. " The firat 10 rats received double. implanta (at PAG and 'IC target
. , .it •• )', to .ini.he the nu.be~ of ,rats required, but this pro~edure was
, . ", •• r ••
~
o

o
o
o
l' •
f9.
22
judged too costly in terms of accuracy and. tilDe. The .remaininl S4 ratl
received single implants. 2 out' of t-he total of 64 ratl died during
surgery. . 1.
HabituatioQ
" Rats vere allowed at Leut one week to recover from surgery and "ere·
t~en habituated to the sti.mulat ion p~o~edure 'without actually
experiencing stimulation. For this purpose, 'f!!'Ach rat was l:aken from its
individual cag~ and was plac'ed. one at 8 time, into the stimulation
container. The electrode lead was then ~bnnected to the fat' B head-cap
so that it could experience the restriction of a lead attached to its
• head. During habituation the electrode lead W8'S not èonnected to the
stimulator. Each rat expe~ienced at least 5 daily 20 minute seuions of
habituation to the stimulation containèr.
, St imulat ion
At the bèginn ing of a test session, each rat was taken from ita home
.~ cage and waS placed in the IItimulation container. the electrode lead w ..
.connected to the rat' 8 ~e,d-cap ,and the iid was, pl,aced over the
container. Current was switched on and W811 immediate 1; adj,ulted to the
desired întensity level as indicated on the oscilloscope screen. This
adjustment required a maximum of 3 ae<1Onds. When the delired current
level was attained a stopwatch was àtarted to tiale the total I,timulation
duration (30 seconds). AU behavipural relpon.es during Itimulation vere ~ . ~ ,
noted. If beha-v.(our became violent, indiclting severe aversion or
aeizure act ivlty. the sti~ulation period wu eut Ihort. At It i.ul~t ion , ,
'offset (30 seconda after atimulation onset), the Itop"atch va. stopped •
and ÎlDllled ia tely re-Itarted to record the polt":,,st i.u1at ion inte rvall.

---
c
o
\
23
St illulat ion Parametere ' -\ In co~trut to ,pre~ious anÏlul SPA studiea,. this s.tudy i! primarily \
concerned with a comparison of the SPA efficacy of anatomicaUy .
, homoLogous brain sites in rat and man. Collsequently, the rationale for , ' r
the selection of stimulation parameters was that the y should he
re'presentative of tholle used in clinical,. rather than animal, SPA
studies (e.g Adams et al, 1974; Riehar?8on and Akil, 1977a; Namba et al,
1984). The parameters seleeted were: - 30 sec trains of 50 Hz, 0.5 ln see
pulse duration, square wave stimulation. Based on preli~tnary trials, . current ateps for the four target laite groups, were:- 0, 40, 80 and 160
uAmpa for the le and' CING. groups; 0, 20, 40, 80 and 160 uAmps for the
PAG group; and finally, 0, 80, 160, 320.and 640' uAmps for the \TST group •.
Lateralitf of stimulatins ·electrodes
On1y the formalÏn test is latera'lity -specifie and therefore the
Ï8sué of optimum laterality of el~ctrode placements on1y arose with this '.
teat. Althou&h there is current 1y. no direct evidence for the
1âteralisation of formalin test SPA from PAG sites, the proximity of
these sites to the mid-line ~uggests that their lateraFtr, relative tG ..
,the foraalin-injected hindpaw, ia unlikely to be a criticd faccor in
,> deterliining optimal SPA. Nevertheless, simplicity of de8ig~ demanded ".,a
choice bel!ween .ides. therefore, it wa8 decided arbit radly . that dl PAG
,tectrode placements should be located ipsilateral to the
fonaali'n.-injected paw. -'
Âbi .. ~ etudies provide no indication of the l.ùerality of possible
fonaalin ,test SPA ai either the VST or le sites. But 80me clinical
1
.t·udi •• Su.a ••• t that if. tonie, pain SPA. is lateralized. it uy be
, \
,'>
. '
" il •
~,

-0
o
o , ,
24
restricted to' the contralateral aide of the bo~y (Turnbal1, 1984). Thui • .. in the absence of Any further evidence, it vu decided tbat the
formalin-injected pav should be sited 'cont ra1ateral to the YBT and IC
electrode placementa.
PAIN TESTS ,
1) The taÏl-flick test procedure
1 Th;' s .test vas ~ modificat ion by Janssen. Niemegeers aad Dony·.
(963) of tbBt originally proposed by D'Armour a"d Smith (1941). The rat
vas removed from ·the st imulat ion box, wrapped in a" ~mal1 tovel, leavillJ
the tail exposed, ..... and carried (a distance of approximately 4 metres) to
th~' tail":;f~ick test-ing apparatus in an adjoining room. This part of the <IJ.:
prpcedure was complete&' within 30 secs of ,stimulation offaet. At each
test time po:"lt (l, 3, and 5 min post-stimulation), the distal 5 cm of
the rats tail vas dipped in 55 degree C. water. The tai l-flick latency _ -t,
vas recorded. If the rat had not withdrawn ies tail by.10 sec&'j' the tail . /
vas removed to prevent tiuue damage. The taÏl was then dried and the
rat wat'l held gently untÏl the next tail-flick test time point, or vu , .
returned -to eith,er the stimulation box or its home-cage. The 2 min
inter"':test interval wa.s adopted to mi ni1Vtze carry over effects of the . ,
thel'1llal stimulation.
2) The formalin' test procedure
Each rat vas placed in, the stimu lat ion box at leaet 20 ain pri9r co
formalin ~nject ion Co allov habituat ion to the apparatu.. After thie
geriod, the plantar surface of one of the hindpa"l va. injected vitb "
0.0~5 III of 2.5% forma lin.
q

. ' •
25 r
The behavioral pain rating scale of Dûbuis80n and Dennis (1977) va8
modified for use vith' the hindpav8. Rating score8 could range from 0.0
3.0. A rating of 0, corre8ponding to complete analge8ia, was recorded
vhen the rat rested its weight equally on both hindpaws. A rating of 1
was recorded when the rat showed a preference for resting its veight on
the uninjected hindpaw. A rating of 2 vas recprded when the rat lifted
the injected hindpav off the floor. Finally, a rating of 3,
corre8ponding to maximal formalin test pain, was recorded when the rat
resorted to licking or chewing the injected hindpaw. Each momentary
rating was entered into a TR~~O micro computer, vhich was .programmed to
print a mean rating score for every minute post stimulation. Data were .
normally collected only for the first five minutes post stimulation.
This vas followed by 4-5 minute urest intervals before the next
stimulation periode Occasionally, pain scores did not rettKn to le!,s
than 1 rating from baseline, by the five minutes post stimulation "'-
interval. In these ca.sei the period of data collection vas extended (to
ensure an approximate return to ba8eline before testing at the next
intensity level commenctd).
The fonulin te8t produces a moderate pa}n which diminishes in the ,-firat 5-10 min and then rises steadily until it reaches a relatively
conllt.nt plateau at 20-30 min post ,injection. This constant level of
alge.ic behavior la8ts fro. about 3,0 min. to 120 ~IL..JJst-injeC'tion •
Telt. for the effect of stimulation occurred within this period (i.e
fro. 30-75 min after fonaalin injection). For IC, CING. and VBT groups
th. hindpa. contraI.terd to the electrode placement was used, for the
•

o
o
. ""'*-
o
\.
26
PAG group, the ipsi,lateral hindpaw was used (aee Laterality aection
above for rationale).
, The Catalepsy test ... During testing it was observed that 160uA stimulation of tc sites
elicited seizure behavior. These rats exhibited post-ictal analgeaia in
conjunction with a depression of motor activity. To explore the nature J
of IC post-ictal behavioral depression, the catalepsy test (e.g CoataU •
and Naylor, 1974) was incorporated into the testing procedure of a
sample of 7 rat8. from the le group. Immediately~fter offset of
8timulation (l60uA) in the tail-flick test condit~n, the rat was
removed from the stimùlation box and te8ted for catalepsy. The rats
hindpaw8 vere placed on the table and the forepaws on a bar 10 cm above
the tablEJ. The time taken, for the rat to remove both forepaws, was
measured. If the rat kept both of its forepavs on the bar for more than
45 sec it wa8 j1lliged to be cata leptic. This criterion ie 15 sec more
conservati ve than that employed by KofW __ Berney and Hornt!iewicz
(1978) in their study of morphine-induced catalepsy.
Design .. o ~ "
Rats vere provisionally allocated to groups on the buis of the
target site of theilt electrode implants. Ali rats were then telted for \
.the effects of brain stimulation on pain uling both the tail-flick ,test
and the· formalin test •
{'
Controlling for' order eff/cts acrou pain test •
.. Each target wite group val rando.ly divided into tvo, equal Rt
lub-groupa. One eub group perfonaed the tail-flick test tiret and tbe
... " , .'

(;
o . .....
27
foralin test second, the 8e[ond ~ub-group wa8 tested in reverse order.
At lesst 1 week elapsed between testing "ith the -two pain tests.
Controlling for order effects across levels of stimulation intensity
In the course of each pain testing procedure, each rat was tested at
four or five stimulation int~nsities depending on test and target site
group. To control for carry-over effects from stimulation, rats received
the different stimulation intensities ln a pre-determined random
sequence, L~lanced within groups.
Controlling for order effects across post stimulation intervals
Since order effect sand t ime are inherent ly eonfounded, an ana lysis
of the ti~e iacfor assumes constant levels of baseline responding over
time, in both pain tests. A ailot study sUPPQrted previous findings (e.g
Dubui880n and Dennis, 1977) that basel ine levels of responding in the
formalin test. do indeed remain relatively constant throughout the test" ..
period (35 - 45 mins). However, another pi lot study demonst rated that
this is not true of 'the tail-flick test. The latter test comprised a
bloctk of three measures aeross time (l, 3 and 5 mins post st imulat ion)
and four or five intensity lev.els. In. the pilot baselÏne analysis, it
was diacovered that tai l-flick latencies were signi ticant ly shorter
. during the first time interval. Becsuse order and the time factor are
confounded, a randomiaat ion solution to this problem 1US inappropriate:
Accordingly, the taU flick test proc'edu're was divided into two more
condition.:- 1) s no stimulation control condition, and 2) the
aci.ulation condition. The order of performance of
counhr-balanced within groups. The difference the tail-fl ick
latanci •• in the Iti.ulation and the no was used
.J,

o
o
o
t
.. 18
to provide a Ille88Ure (wttich minimÏ!:ed spurious order effects) of the
effect of stimulation on tail-flic" latenciea.
DATA ANALYSI S
Group Analyses
Rats were assigned to electrode pla~ment groups on the basi. of
histological results. Ail rats experienc-ed 4 or 5 Levels of stimulation CI
(dependlng upop group and pain test - see Table 1. in the results) in
bath the tail-flick and the rforlll8lin test. ln the taÏl-flick test, data
were recorded at 1,3 and 5 minutes post-stimulation. In the formalin
test data were recorded at five time points post stimulation (l, 2, 3,.4
and 5 minutes). In some cases rats had to be excluded from testing at
the higher current intensities due to aversion to stimlation. In such
cases the' means of the remaining animals tested at thue intenait ies
were used ta replace missing data points.
The results from the tail-flick test and tbe formalin test were
"' ànalyzed sepa rately. An ana lysis of the 0 Amp t baseline data t comparing
the effects of time and stimulation site group, was conducted with a ..... 2-way, 1 repeated~f8ctor,'ANOVA, for each pain test. Subsequent".n.ly.es
of the effect of stimula.tiofl intensity and post-stimulation time point
on pain test scores, vere conducted for each histologically defined
group (and operationally defined sub-group) vith 2-way, 2 repeated
factor, ANOVA's, again for both.pain tests. r
In the case of the taÏl-flick tesuata, a lack C?f hOalOgeneity of (
. ~'. 1
variance, and correlatÏ'ôn of-vai;.an<es vith the .an difference .core.,
vas observed. For chis reason, AROVA·. vere perfo~d on .quare-root
\

/"
y'
~''''' ... '
29
tranlformed data. In some cases, the 2-vay ANOVA stimulation intensity x
time interaction term proved significant. In most cases this was a
straight-forward consequence of an increase in the duration of the SPA
effect vith increments in stimulation intenslty. If this was the case, a •
further l-wsy analysis of the components of the interaction term was
de~d unnecessary. 1 f, however, the source of the interact ion was
unclear, I)urther l-way analysis of the stimulation compone nt of the
interaction term was carried out. To examine the significance ol the
stimulation factor aione, the analysis vas reduced to a l-way ANOVA, by
averaging scores ac ross t ime.
Individual Analyses
Si tes su SPA and imulation intensities at which SPA
occurred
To determine the st imulation intensity that supported significant
SPA for each sub ject, the pain scores at each 'st imulat ion intensity were
- compared to the mean baseline scores for a11 rats tested at the same
site. Pain scores were averaged across time and the mean and standard
âeviatio=, of the scores for the baselÏne test were ca lculated.
Signifieant SPA was defined as the turrent at which:- A) the tail-flick
test score was more than 4 standard deviations above baseline; B) the
formalin.. telt pain score was lIIOre than 4 standard deviat ions below

o
o
o
o \,
30
Sites and stimulation intensities st vhich' aversion to stimulation
occurred
Aversion to st imulat ion occurred at many stimulat ion sites in both
pain test conditions. The behavioral definition of aversion employed in
this study required that the rat should display one of the following
behaviors during stimulat ion :- 1) irttense escape involving jumping up
and hitting the !id of the stimulation box; 2) escape accompanied by
vocalizing; 3) escape followed by major tonic/clonic convulsions.
The st,imulation intensity at vhich these aversive reactions occurred
vere noted for each stimulation site, in both pain test conditions.
Also,r parallel ing the SPA ind ividual si te ana lyses, the number of si tes
'" per ~roup at which aversive reac"tions to stimulation occurred V88
calculated for't~:7. stimulation intensity tested.
These tvo setJJ~tndividual analyses provided the buis for a site
by site comparison of the s~imulation intensit ies at vhich SPA and
aversion to stimu lat ion occurred.
Comparison of the relative strength of SPA in the two pain tests ,
In order to make a direct comparison of ta i l-flick and fonulin telt
SPA at each stimulation site, individual ,taÏl-flÏck latency scores and
Il formalin test pain scores, àt the 160uA stimulation level, vere
converted into percentage anaigesia (% SPA) scores uBing the follovina ,
transform: - (test score-baseline score/maximum teU score-ba.e 1 ine
score) x 100.
Histology ,
Rats vere deeply anaestvtiz~d vith chloral hydrate. The, 1I.r. tben
perfused intra-cardially fint vith 0.09% .aline. and the~ vitb 10%
,-

,
- ..
_ .. =
31
• foraalin., The electrode head cap was removed from the sltull, the' brain
wu eJttracted and stored in 10% formalin for at least 24 hourà. Bl"aitl8
.Oi
\ \,
". rt. were then 'sectioned on a free~ing mict"'otome. The s1ides of IIections were
1
stained according to the Kluver-Barrera procedure. Stained sections were
examined, under a microscope, ta identlfy the positioo& of electrode ~
tracks •
•
1
(
,
-

o
/~.-
"
,
•
•
RESULTS
o
e • Hi sto1ogy
A total of 74 electrodes were' implanted in 64 rata (this number-
included 10 rats vith ~uble implant s). 10 of the placements vere
10lt due to the death of the rat, or to dislocation of the electrode r
during testing. Of the remaining 64 placements, 51 vere "on tU'get"
within their original target sites ,and 13 vere "off target" in 4,
adjacent regions. The· posit ion of e1ectrode' placements within the
PAG, IC, VBT, CINe. target sites, or one of the off-target regions
32
(OT, ML, RT, Zl), are illustrated in Fig.s lA-lE. The compositiçm of
stimulation site groups, and the current intensities used in the
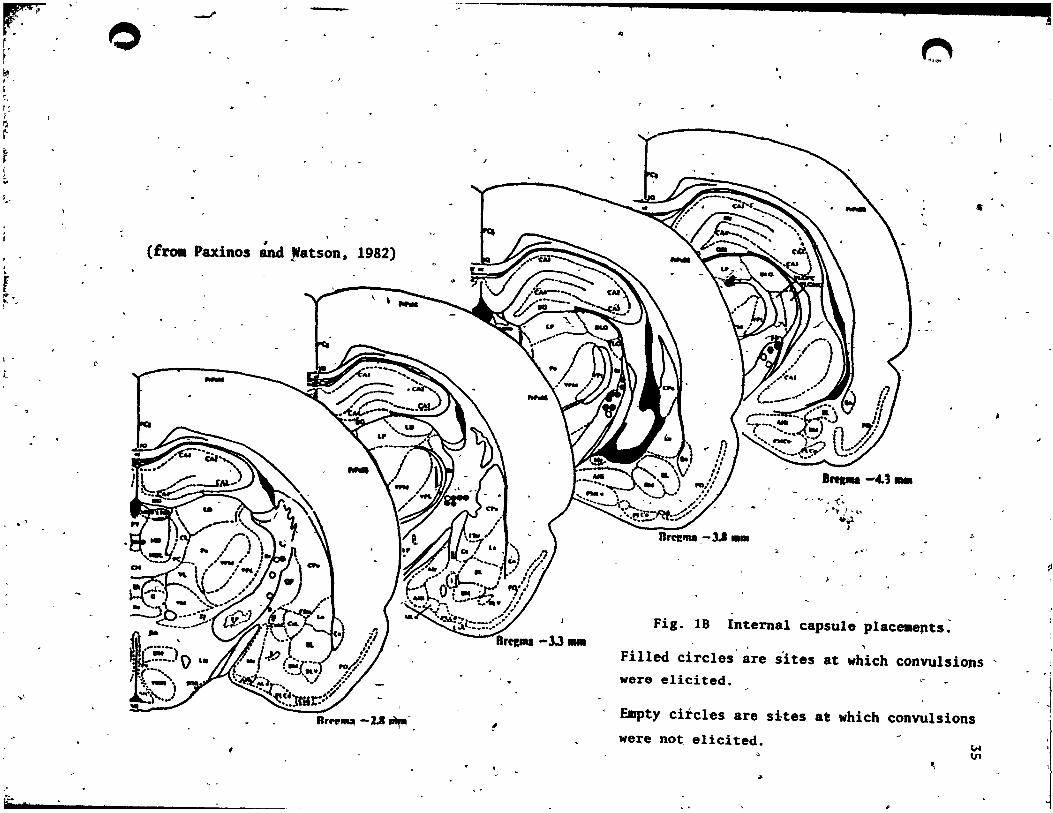
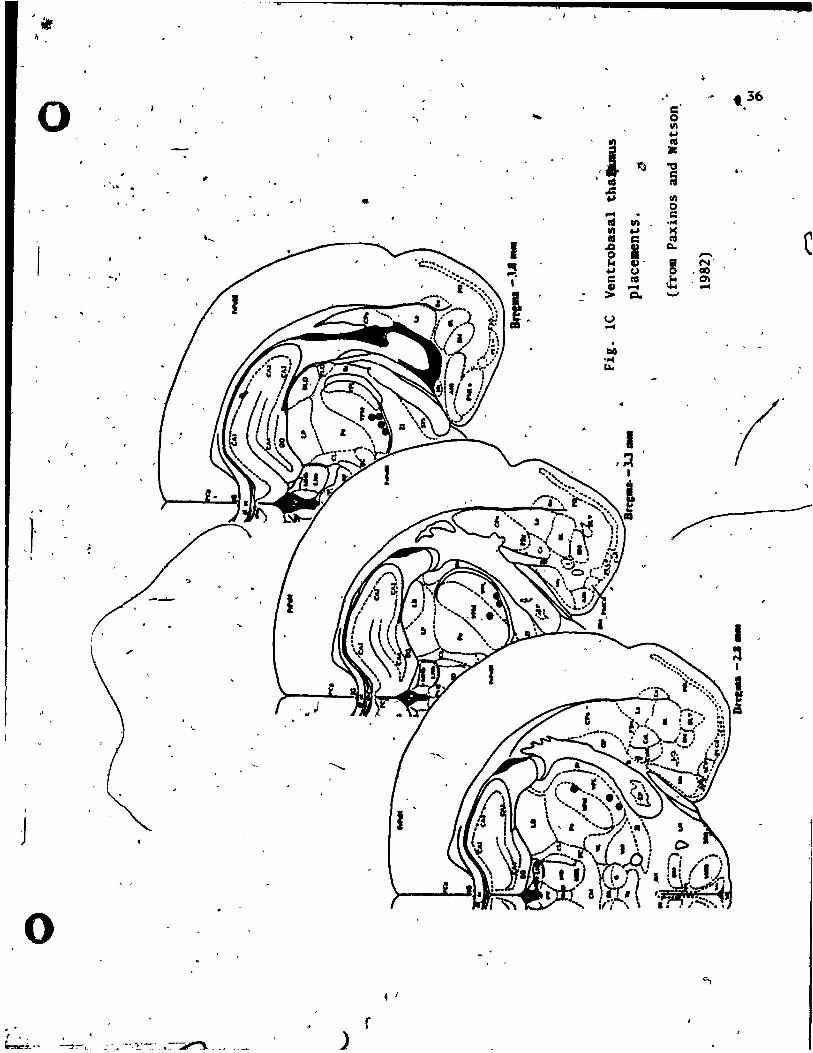
testing of each group, are t.ab~latea, in Tab le 1. • ·"<t
The lC and VBT groups vere divided into_ sub-groups on the basis of
test reBults. The number of ratl chat were Cested (in each pain tut ,
condition) in each group, and subgroup" are aho tabulated in Tab le
1.
Rueline analys.s
A 2 vay ANOVA vu performed on baseline (0 uA) taU flick
difference' Icores (test condit'ion mnus b.aeline condition).
lndependent variables were:- 1) a Site factor, co~priaing the 4 "on
taraet" group.:- 1) PACt 2) IC, 3) VBT, 4) CING., and 2) a Time
factor, coubtina of the J,time intervals poat- atimulation (1, J
and 5 .inut .. poet .timulation). Thi. revealed tbat there ven DO
difference. betveen baselines acro •• aroupa !(J, 47) • 1. 79, liS; DOr
( 1

•... _-r
• .; II<.~ ,
, '
33
• .. ~LE 1
Composition of groups tested for-SPA in the tail-flick (TIF) ,
and, fonulin (F)· tests and ,~imulation intensities used for each \ ,
group
Number of Stimulation . On target groups sites test-ed intensitieà and sub-sroues T!F F used (QA)
PAG \. 10 07 o , 20 ,40 , 80 , 1,60 '*
" Ie 23 23 -0,40,80,160
lC rats exhibiting seizut:es 13 15 o ,40 , 80 , 160
èxhibiting " . , 0,40,80 lC rats selzutes at 160uA only 09 11 ,
l
lC rat Il not ,exhibiting sei~ures ,10 08 0,40,80,160
VBT (standard intensity range) 10 09 0,80,160 ---:-"'...a.
(extended intensi ty range) 10 09 O,80.~60,320;640
CING. 08 08 0 14° 180 1 160, : .~
r .. Off tarlet sroue'
OT 03 03 0,40,80,160
HL " '
,03 03 o ,40 ,80 , 160
RT 03 02 O,~~O, 160 -~
Zl 04 04 o ,80, 160
.' * Tbe 20uA Iti.ulatioD iotènsi~y va. ooly tested in the tail-flick .
tut condition.
1 •
o - 1
.... ' 1Ir..,... __

o ~
,
~. >--~
) . ( .;-.., ,
-Brc:gma- -6.3 mm
fie. lA Periaqueduc~al gray place~nts.
(f~ Paxinos #nd Wats~._198~
,< o '"
\#0
, .
"
r
..
Bregma - 6.8 mm
Triangles represent sites w~th hÎgh thresholds of
aversion. J
Squares represent sL~es with low thresholds of ~ers1on.
(,0-A
r· --" -0
r , ~ , ' i :, r: ; >:1
~
~ ~
[ , .. ; .'
~ (f l' ra. Paxino5 and patson, 1982)
~
,
t ,... ~. ~
, -t
-3.3_
... '
!
,
1ft: __
.. ~
.~r. CAN~ j" ........ --
- /,~"~ \ , •
-(. ' .... J .. t,.,
*o.l , "'
" ."
Fig. 18 InternaI capsule place.epts~
Filled circles are s'ites at which convulsions ' were elicited .
Empty circles are sites at which convulsions we~e not, elicited.
#
..
t.
~ VI
~
,
.. il , 1
0 . , .,..
~ , ~ ". .
• 1 •
' .....
~. ,
. -
, . J
o 4 1
.. r ~-~j.
,1 ~ ' III
.c:: +.1
~ . 1'0 ln III .., " c: ,Q
" 0 ,.. ~.
+.1 (J
i CO .... > Q.
U .... . QG ....
LW
1 ,::t
1
. c:' 0 11\ +.1 1'0 • -g 1'0
11\ 0 c: ."" = CI.
~ ~ '-'
-N' • CIO .0\
..-4
t 36
. ,
1 :s 1
!
$

,_ . . • • •
37 ,.... "1 ..
("II . . . 00' G ~ . 01
." .-4 ... . = .. ~
i s:: 0 III U ~
., as
= ... 0-4
~ /
~ "C:I
0 ra ... 9 k III
, 0 0 U .. s:: G
.P'4 ... =
« as Co 0-4 :::s
~ 00
= ct 0f'4 U -• Q .-4
00 ..... "" \.. & •
; . "': 0
1 ! lE
CD .B "'!
" .B ~ ! '\ .
1 CD
"': -.. Il .' . I! ! =
"
~.
o ..
o ..
Cl"
:. ,
\ . ,
4 •
-.1.3 ...
(fra. Paxinos and Watson. 1982)
,
.>-~. ---"._--"->.. :...-..... ~ __ J'
.•• OT
• ML
.,
" • _ ..... , '0
..
.~
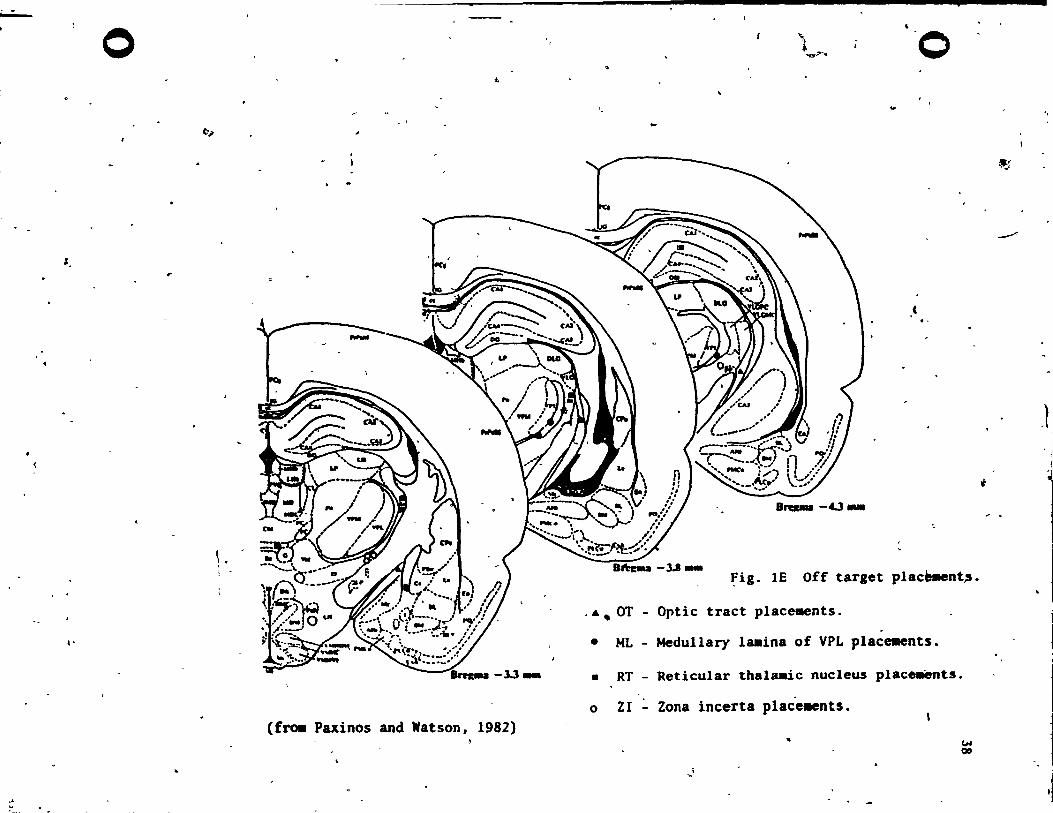
-3.1_ ~ig. lE Off target plac~nt~.
Optic tract place.ents . .
Medul1ary laaina of VPL plac~nts.
• RT - Reticular thala.ic nucleus place.ents.
o ZI - Zona incerta place.ents.
,
~I
'" Olt
,..;
.-/
J
~

\ -
." •
()
1
1
•
..:f ,.,
'"
- ~ ~
-ovér ti_ !(2, 6) • 1.67., NS; and the interaction tertl va. not
d,nificant !(6, 94) • 1.04, RS. ,/5'
ln a .imilar analy.is of formalin te.~ data for the OuA intenaity 1 Ir
• level, a,ain, there vere no differences betveen baae1inea acrOI.
group. !O, 43) - 1.66, NS; nor acrou time !(4, 12)·· 0.90., NS. and
the interaction term was insignificant !(12, 172) - 0.82, NS.
The.e ana1y.e. confi~ed that, in the absence of stimulation, both
pain meaaure. wer~ stable across time and comparable ~cross groups.
Group Re.ulta
Tbe PAG Group (N-I0)
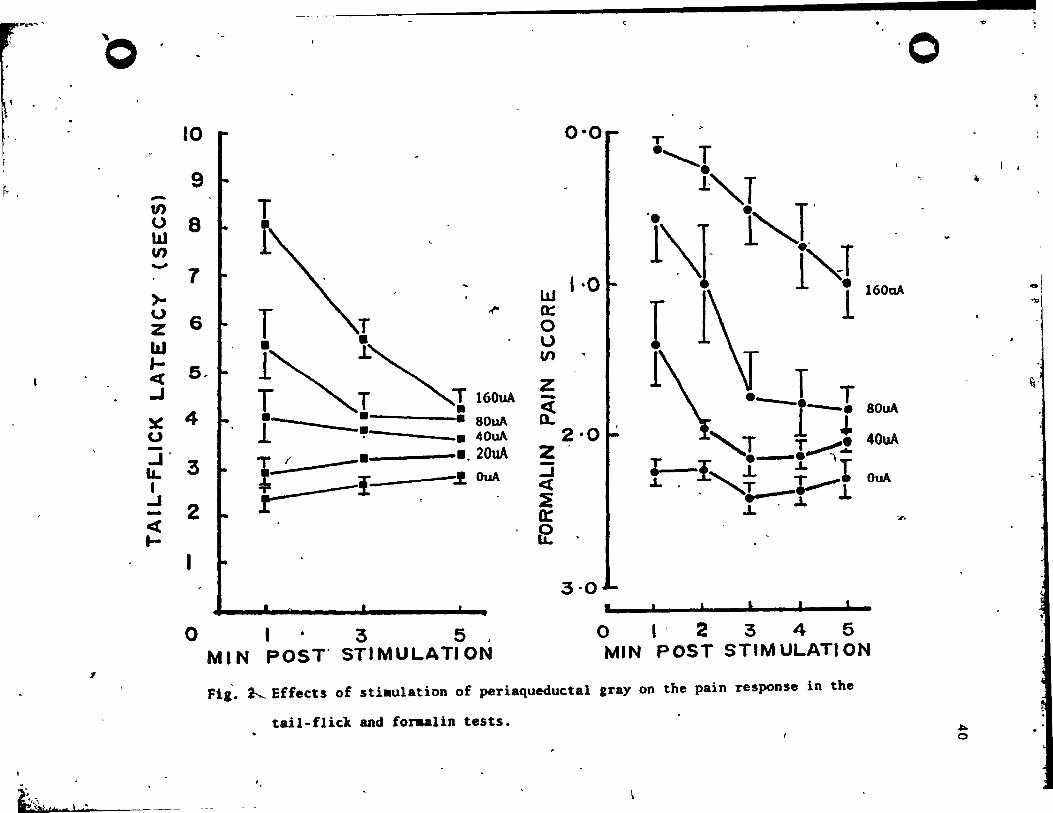
The re,u1ts for the PAG aroup are plotted in Fig. 2. Fro" the
fiaure, it is c1ear that 'PAG .timulati~n produced analges'ia which
increa.ed vith the intensity of stimulation in both the tail-flick
te.t !(4, 36) • ~2.55, ~<.001. and the formalin test !(3, 18) -
31.58, ~<.001. It i. also apparent that thi. SPA effect decayed
rapid1y over the 5 minute poet-stimulation,period. The decay of
.na1aeaia over time va. highly .ignificant in the tai1-flick test
!(2, 18) - ~3.90, ,2<.001. and the formalin telt !(4, 24) - 13.61,
R<.OOI. rinally, there vas •• ignificant interaction betveen
.ti.ulation and time f.ctors in ,the tail-flick test !(8, 72) - 9.02,
R<.OOI. and the forulin tut !(12, 72) - 4.45, ,2<.001. , In both
" c •••• , this va. evidently a con.equence of an increase in. the
duration of the SPA effect vith increment. in .timulation inten.ity.
lt .hould he noted that in the tail-fl~c1t te.t J' 2 out of 10 rat.
vere dropped out at 80uA and. further 3 vere~dropped out .t 160uA
. .
59 t
~~'111'"
o
~ .
~
t ~{\ -. /~a},*. ) .ete
10
9 -~ a w VJ
- 7 )-u Z W Je(
.J
6
5.
x 4 o -..J' lL 3 1 .J - 2 ct ....
1
T ,r . ~ ,
~~;~T160uA .- . . l • 80uA ---_.40uA
".
1--" _. .,20uA
}.:.-.___----!: -.! OuA
0135, MIN POST' STIMULATION
0·0
w 1·0 a: o U Cf)
z -~
2'0 Z :::i <t ~ 0:: f2
3'0
'0
T ·"r -l"T 1\}. 1"-1 . l~T T 1\ 1 l
1600A
T T ------. 1 * ., T T~ ....
"'--- T .L • T ,r,J.. T ............. .L • .x= T __ .L . .,.L
.J.
80uA
40uA
OuA
~
, , o l' 2345 MIN POST STIM ULATI ON
Fi,_ ~ Effects of stiaulation of periaqueductal gray on the pain response in the
tail-flick and for.alin tests. ~ o
l
'"
-""
ft,

• , " ~
,'" .
o
,
'J
..
becau.e of aversion to stimulation (see criteria for aversion in
# •
_thoa .ection: page, 30). In. the formalin test 3 out of 10 rats
dropped out at 80uA and a further 2 vere dropped from the 160uA level
cella. To maintain equal n's, muing data points vere replaced by
the _an of the available scores. Accordingly, the results for the
higher current levels should be interpreted vith caution.
In a subsidiary analysis, the possibility of an association
between PAG stimulation induced aversion and dorsal/ventral elèctrode
po.ition (Fardin et a~, 1984) vas examined. Rats vere divided into
tvo groups on the buis of electrode position (ventral or dorsal to ., the midline of aqueduct) and the stimulation intensity level required
to induce aversion (Lov aversion threshold • 40uA, High aversion
threshold - 80 and l60uA, see Fig. 2). No association vas found
between aver.ion -threshold and dorsal versus ventral stimulating
__ --...:e:..;l;.;;ectrode polition in either pain test Fisher exact .f - .300 for both
.. ----
pain te.ts. ..
SPA froID. tbe' i nterul cap.ule
The ruulu for ratl vith IC implants, vere cOllplicated by tbe
discovery of an a •• ociatipn betveen SPA and escape behavior
accompanied by .ei&urea. Accordingly, the reluIts of the IC group
vere fir.t analy.ed a. a whole. and then separately, for tvo , .
sub-Iroup. compri.ina Oret. vbicb eitber did, or did not, display an
.. cape+ •• i.ure re.pon •• to (160uAJ nilDulation. \.
o
41

o
o
-'~-_.~--------------------~,-,----~;~----------"~~,~.~ ... :::s~~.q •
42
The lC Group.(N-23)
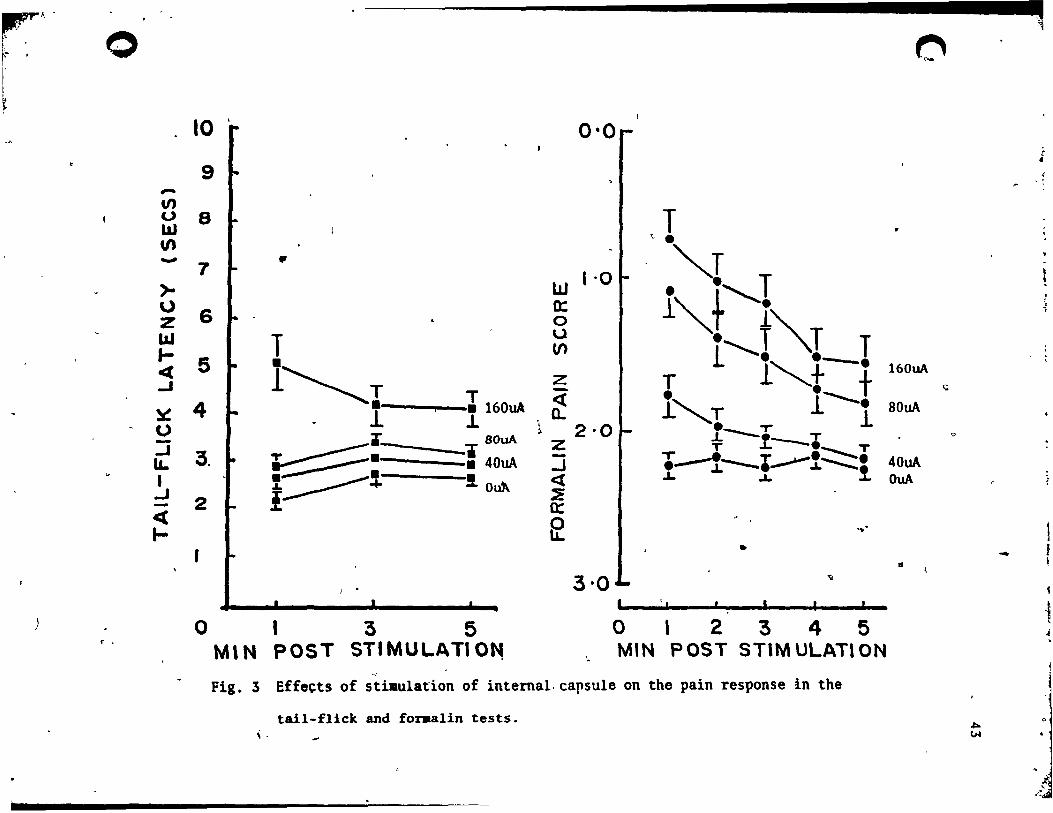
The effecta of IC stimulation are illultrated in Fig. 3. From the •
figure, it is clear that IC 'stimulation elevated tail-flick latenciel
IO, 66) - 20.40, .2<'001. Thia effect did not alter over time !(2,
-44) • 3.20, NS. but the interaction between stimulation and time wa.
significant !(6, 132) - 6.87, ~<.OOI. Because the inter~ction wa. not
readily interpretable. the effect of stimulation was reanalYled by
averaging across;the time intervals. Under this condition, the
stimulation ef/ect remained significant !O, 66) - 20.98, ~<'OOl. .('
From Fig. 3 it i8 also clear tbft lC stimulation reduced the pain
scores in the fOl"malin test !(3, 66) - 23.19, ~<.OOI. this effect
declined vith time !.(4, 88) - 24.16, ,E<.OO1. and there wa. a
, significant interaction between stimulation intensity and time !( 12, ,
264) - 4.66, .2<.001. In thh case the interaction term was a atraight
forward consequence of an increase in the ,duration of the SPA effect
vith increments in stimulation intenaity.
lC Stimulation, Seizures and CataleplY
In the co~rse of teating, it was noted that the majority of the
rata in the lC group (13 in the tail-flick and 15 in the formalin
test) exhibited escape and leizure behavior at the 160uA level of
stimulation intensity. Within 10 seconds of the onset of 160 uA
stimulation, these ra,ts pawed the air vith the contralateral foupaw,
tben ahowed rapid contraveuive cirelingpand vocaUsing. Between 5-15
aeconds after stimulation onaet, the tat exhibited an explosive
eacape re.pon •• - it vould juap up and bit the lid of tb., .tiaulatioll
J
"
,.
~~- ~- -~
"'1 ,
0 ~ b ' r . l' 1
l , ~ ,
10 ' 0'0 ,. r :t
-9 ~ , 1 ; . -V) (.) 8 T bJ \1) " . - 7 • "'T .
1 ·0 t >- w l"-J'I \,) 6 0:
Z 0 LLI T u ... ,J""T T l- CI)
5 • l 1~+-. 160uA ct l~T ..J z T • t Q
l 160uA -
4 • <t 1, T 1------1 BOuA ~ 1 .L 0-0
\ 2·0 ~ • ___ T
~.- -1' 80uA .L .t T - Z T ----. T ..J 3. T :::::::----:. - .--1----.~!:::::::i 40uA l&. • 40uA .J 1 .~. !.Otd\ <t ~ ~ O~
eL + ..J ! ~ - 2 a: ct ft .... 3.J
.... f • -Il
~
'. 1 -1
) 0 1 3 5 0 , 2 3 4 5 J r ,
POST STIMUL.ATI O~ MIN POST STIMULATION ,
MIN ~
~
Fig. 3 Effects of stiaulation of internal.capsule on the pain response in the
tail-flick and foraalin tests. "" , - ~
f. ~.

o
o
box, often succeeding in forcing its vay out through the lIlit for the
electrode lead~. lanediately after this apparently directed e.cape
behavior, the rat invariablY exhibited a "grand mal" tonic-clonic
seizure. Stimulation was always terminated at this po'trrt '. Convulsions
usually abated as soon as the current was switched off (except in one
case in which they outlasted stimulation offset by approximately 30
secs). AH rats that exhibited seizures app~ared to ~f;eI from \1
post-ictal depression in the form of immobility, often in unnatural
postures. After convulsive reactions to le stimulation, rats in the
formalin test condition, would often remain ln unnatural postures for
a period ranging from 45 seconds to 5 mi nutes. The amount of wei~ht
placed on the forœalin injected paw appeared to be determined more by
the chance posture assumed than by any at tempt to avoid psinful
contact with the floor. A sample of 7 rats which showed convulsive
reactions to le stimulation were tested for post-ietal eatalepsy with
the standard bar test (e.g Costall and Naylor, 1974). AU 7 rats left
thei r forepaws on the bar for st least 45 secs post-st imulat ion
offset, and therefore aU were judged eataleptiç (see Method, page 26
for criteria for bar tet catalepsy).
IC Seizure/SPA association
The occurrence of escape+seizure be'havior appeared to be a
crit ieal determinant .of "analgesia ft. To examine thia associat ion, IC
implanted rats vere clauified according to the presence or absence
of convulsive bebavior and signifieant SPA (IIOre than 4 S.D.'. froa
ba.eline). at the 160uA .t1.ulation intenaity. Cbi2 analy.ia ,
"
44
d
-

o
indicated thatlh~re vas a strong a88ociation betwéen the occurrence
XL • of aehures and 8ignificant SPA O,.!! • 23) • 13.08, l<.OOI in the &
tai l-fliclt test and in the formalin te8t x: (1, N· 23) • 18.86
Accordingly, animais in the le group vere partitioned into tvo
eub-groups on the basÎ8 of eeizure occurrence at the 160uA i'ntensity
leve!. These were:- 1) rats which exhibited seizures - 13 rat .. in the , tai l-flick test condition'-and 15 in the formalin test condition, and
45
2) rats which :lid not show seizures - 10 and 8 rats in the tail-flick 1
and formalin ten conditions, respectively.
SPA in rats exhibit;ng seizures
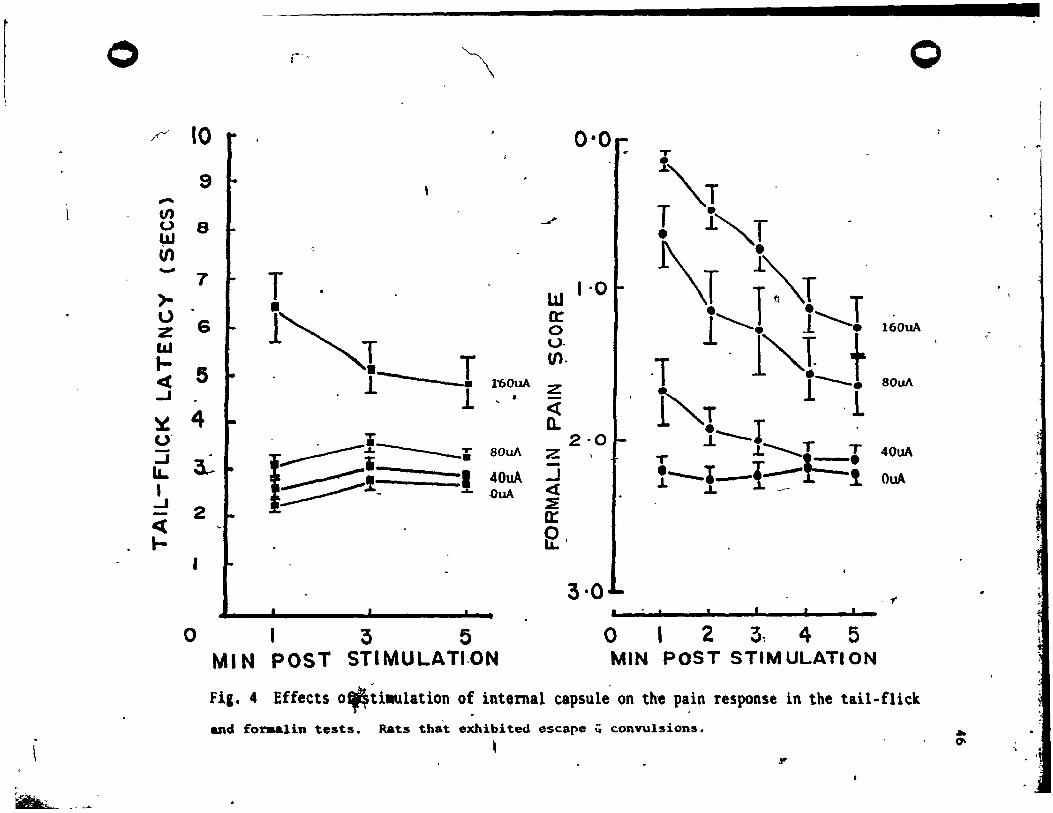
The resu lts for the IC sub-group that exhibited seizures are
plotted in Fig. 4. Tail-flick lateneies inereased Vith stimulation
!( 3. 36) • 35.95, ,E<.OOl., this effeet did not vary over time !( 2,
24) • 2.51", MS,. but interaction term was signifieant .!( 6, 72) • 6.08,
l<.OOI. When the resu~t. were collapsed aeross time, the increase in
an.lgesia vith st imulation inten8ity remained signific.nt .!( 3, 36) -
36 .99, l' 00 1.
From Fia. 4, i t i. elear tbat IC .t imulation reduced pain 8cores
in the formalin "test !(3. 42) • 37.14: ,E<.OOI. J:hia effeet declined
over tilDe P(4, 56) • 39.97, l<'OOI and there vas a sianifieant
interaction betveen stimUla/ion i~tenaity and time !(12, 168) • 4.71, .
l<.OOI. 'As before, the interaction evidently reau/ta from an increa8e \ .
in the dura-tion of the SPA effect vith incr •• enta in stimulation,
o ~
" V) u ~ V) -)-\) z laJ I-ct ..J
~ u' -..J li.. 1 ..J -« l-
~-
r-' \ o
10 r . 0·0. or
9 ~"T .../ r l""T 8
T 7 1 ·0 • l1J
6 l~T 0: 0
. 1"'-1 t (J-
5 T (Il.
8-. l -. 1'60uA Z -T 1-- .0uA l · <i . l''r ,1 4 Q. !---I ... 2-0 - .
~ ~î -----. 80uA Z ..L-...........T T 40uA
3..- T l _e 1:::::::::::::1 1 40uA - l---I--!-:- 1 OuA ...J <2:
2 * ~ ~uA ~ ... n:
~, I
3·0 - 1" •
o 1 3 5 o 1 2 3~ 4 5 MIN POST STIMULATI.ON MIN POST STIM ULATI ON
fil. 4 Effects o~~iaulation o~ internal capsul~ on the p~in response in the tail-flick
and foraalin tests. Rats that exhibited escape q convulsions. , It'
~ 0-
, -
• ~I
1 1

c •
, 1
41
thus an independant analysis of, ~he stimulation main effect va.
deemed unnecel8ary. , '
Of tb~ 13 rats in th~ tail-flick IC (seburé) sub-group, 4 exhibited
eJcape+aeizurea at both the 80uA and the 160uA" intenaity lev~la •. The
remaining 9 rata only exhibited theae reaponaes 'to stimulation at the,
160uA level. The data for these 9 rats vere exalDined at the aub-seizure
inten.ity levela (0, 40 & 80uA) to ascertain if there vas any SPA effect . ' at current levels that did not e-l ieit overt seizure act i vity. The reaults
are plotted in Fig. 5. There waa a small, but atati'stically significant,
- " SPA effect F(2, 16) - 4.18, R<.05. this effect did not vary over time !(2,
16) - 1.67, NS. and the intera~tion term was not significant !(4, 32) -
,2.35, NS.
Silllilà,rly, the 15 rats in the formalÏn test le (seizure) sub.-group,
included 4 which exhibited ~eizures at both the 80uA 8'nd the 160uA
, inten.ity levela. Theae vere exc luded and the data fÇYf the remaining 11
rats vere analyzed for the sub-seizure intenaity levels (0, 40 & 80uA).
In contrut to the marginal effect iri the tail-fl iek teat, in the
formalin test' there vas a clear increase in analgesia a!J stimulation
current increased F(2, 20) - 11.74, R<'OO,l. This effect decay.ed with ~
time !(4, 40) • 7.19, R<·OOI and there was a signi Hcane interaction
t.ra !(S, 80) • 4.43, R<·OOI again due to an increase in the iuration "
the SPA effect vith • 1 ....
lncrement. ln st imu lat ion intensity.
Rat. not exhibittng se!zures " ,
The rIBult. for this lub-group are plot~ed in Pig. 6. From t.he
fiau-re, it is c;lear that' rats vhicb did not .xhibit seizures at the
-' -
of
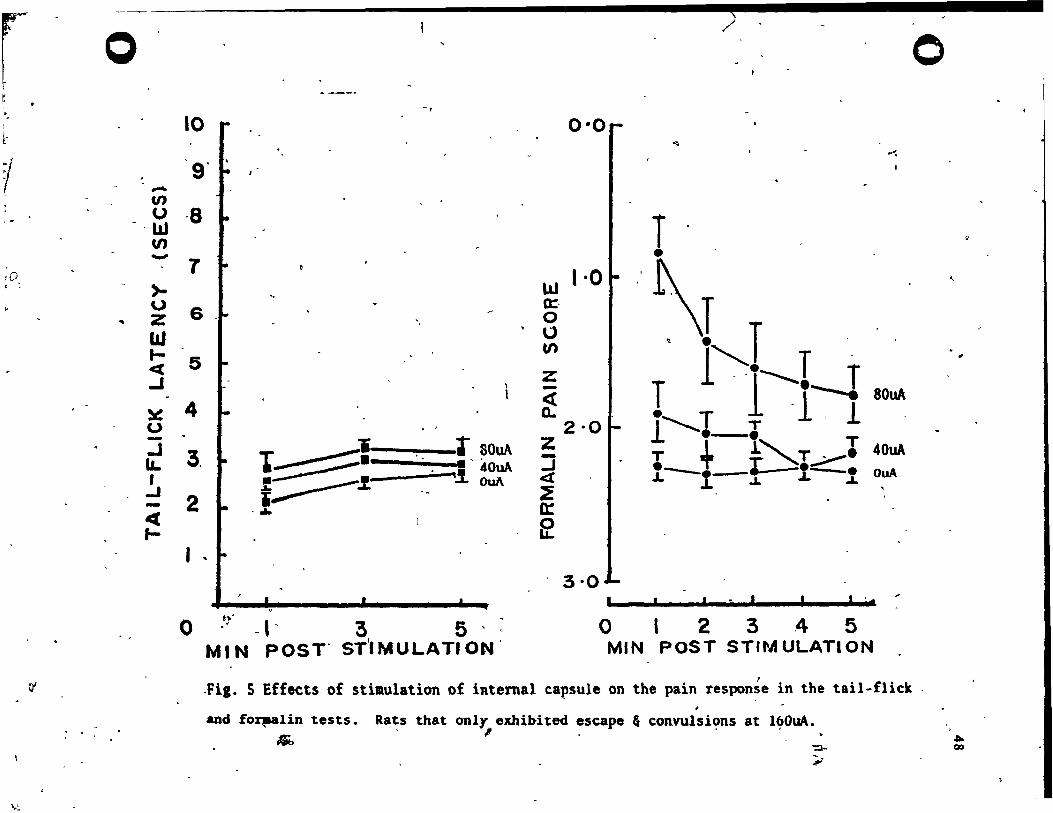
r r
~.
1
rD , ~
0'
\.~
"\
o / o -,
10 0·0 ~
~-:
g' -VI _ (J ,8 ·w T '
U) -)-\,)
.. z l&J 1-~ .J
~ u -.J IL. r
..J -~ f-
T
6 '.
5
4
3,
2
T ~'i r .. ~. .,~OuA f- ____ .1.- :!. 40uA
1"""'-- OuA ....
lLJ 0:. o
'0 tf)
Z -~
1 ·0
2·0 Z --J « ~ 0: El
\T · ." T T 1 . ·l·~.--""'" Tl. 1
r----I-~"-.. T...--1 T + L """1"--1 1--1-.1.
BOllA
40uA OuA
l '
o h" ,. -t' 3 5 ' MIN POST" S~MULATION'
3·0 , .
o , 23,4 5 MIN POST STIMULATION
.Fil. S Effects of stimulation of internal capsule on the pain respon~e in the tail-flick .
and fO~lin tests.
~
Ra~s that onl" exhibited ~scape fi convulsi~ns at l~OuA.
':'l-
"
"
,
". 00
;;
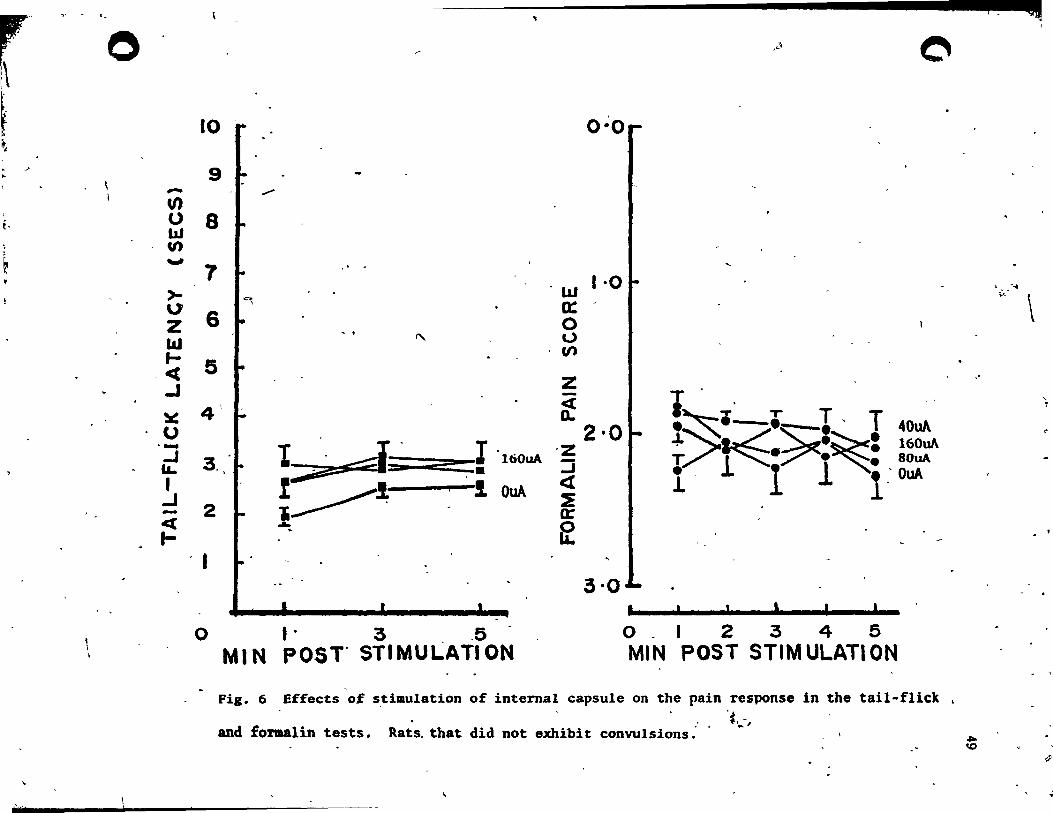
'-
o
, \
" L .. ,.. ......
• ,~\ o
0·0
w 1·0 0:/ o U CIl
z f
2·0 ·z l '160uA :::i
;;: ,- <[ --"1-"""""1 OuA ~
0:
~
3·0
o 1- 3 _5 MIN POST' STIMULATION
I~.~._ T, T 40uA
!?~ ~:~: 160w\ . .~.. 80uA l l 1 l rOuA
, , 0_1 2345 MIN POST STIM ULATI ON
Fig. 6 Effects of ~timulation of internaI capsule on the pain response in the taiI-fIlet . • . ·1, ... ,
and foraalin tests. Rats. that did not exhibit convulsions:' -
"
".. ID
"'J'IJj
\:L:'" \
"
..
.po
cO

o
..
"
o
{ ',. J .... SA I . ..J. ~ ... j t $Ct. en 2 '" ,c a , .. :
\
l6..,OuA lev.e1, did Dot display .tilllulati~produced analaeaia in the
taU-flick test !e3, 27). 2.28, NS. or t'he forma1..in tut F(3, 21) • • .
1.46, MS. The lack of a SPA effect, in either pain test, in thia
sub-group" supports the previous evidence for an associat ion between '/' 1
seizure occurrence and SPA at le ~i tes ~
The VBT group (N-lO) . .
A pi lot study of vBT stimùlation sites, demonlt rated that ,
stilllulat ion did not, produce appreciable analania at the 160uA , .. intensity level. Consequently, a vider range of intensity levela vu
chosen f-er testing at this site (0, 80, 160, 320, & 640uA) to lee if
an effect could be obtained at the liighe~ levela of stimulation
intensity.
1\ YBT stimulation over the standard range of intensities (0-160u'\)
did l'lot induce the aversion characteristic of PAG stimulat ion. ,
Inltead, 160uA stimulation of the VBT resulted in a pattern of
relponse8 wbich appeared ta be of ~ pretmina~tlY motor, rather than
avenive, nature. Shortly after atimula\ion onset rata vould exhib.it
contravershe head-turnina which in SOlle caae. developed into
contraversive circling. Otber relponae. to 160uA atimulation included
rhytbmic mouth movementl Oa" openinl) ~nd head-noddin,.· " .
. At the higher stimulation intensitie. (320 & 640uA) tvo relat,ively
di.tinct .etl of behavior. vere ob.erved:- 1) mouth IIOve';nta, b.ad ï
noddinl and forelillb cionui leadial to rearin, and taUinl b.ckvard., - "' , a1id 2) contr.ver.be circU"1 of .uch iatenaity tbat it often 1ed to
"rallinl over" and culminated in full tonic exten.ion and 10 .. of
.j
so

.\
.'
,
~~
"
po.tural control. ne.pite the .everity of thele behavior8, hovever, ,
even the hiahest inten8ity of YBT stimulation did DOt produce the
degree of aversion seen vith much lower stimulation inten8ities at
the PAG.
From Fig. 7, it is elear that even at the tvo higher in~f.ity leveh there wa8 only a relat i vely weak SPA effect ÎCl both pain tests
(eompared to tha t found in the PAG and le groups). The tail-fliek o
test re8ults indicated' a stimulation effect !(4, 36) • 3.:59, .E<.05 .. ,
this effect did Dot vary over time F(2, 18) • 1:.75, NS. and the
interaction term vas not significant F(S, 72) • 0.89, NS. Formalin
test re8ults demonstrated an effect of stimulation !(4, 32) • 12.75,
,2<.001., this decayed over time !(4, 32) • 7.51, .e< .001. and the
interaction term 'vas 8ignifieant !(l6, 128'-'· 2.41, .E<.01. When
resulu vere eollapsed acrOS8 time, the inerease in SPA vi th
.timulation remained 8ignifieant !(4, 32) - 12.75, ,2<.00l. J' ,
For comparison purpo8el, the data from thè YBT gr~up va8
re-analyzed for the 0, 80 and 160uA leveh, only (these intenaity
leveh correlpond to the maximum range eumined in an other groups
and lub-group.). Under thie constr.int, there vas no SPA in the
t~il-flick test !(2, 18) • 1.30,. MS.
With the fonulin te8t data for the 0, 80 and 160uA intensity
leveh, tbe Itimulation effèet reuined .ilnificant-'U, 16) • 8.60,.'
,2<.01., tbia effect .howed a marginal variatio'n over time .!(4, 32) •
2.74, .2<.-05; the~.etion tera va. DOt .iloifieant,!(,8, 64) •
1.68, MS.
51

0 0 '"
10 t- 0·0 f - 0
1 \.
9 C· -.n
0 8 • • ~
ut U) - 7 , ·0 r >- " w '0
6 0: Z 0 l&J u 1- (fJ T 5 ct • ...J z
~'\ -.... ,w\ 4 «
~ a.. u 640uA 2·0 ~i-i~f-I 640uA - T l z .-.:=- --.J 3. • tC:::::::!-t/Ï boA la. ~;r 1 -..J , « .L, ............... T T
.J 2 ~ . 1----..~1 . - !, .
0:: Cl OuA ~ ~ 1-'
1 3.01 • •
0 1 3 5 0 1 2 3 4 5 # MI N POST' ·STIMULATI ON MIN POST STIM ULATI ON
Fi,. 7 Effects of sti.ulation of ventrobasal' thalaaus on the pain response in the tail-fliek
. and fO~lln tests. l ' ~ •• ~\ J
v ,
~ ta. ~ "_
en f"J
...
t i f
. !
1 •
~

(}
o
. .. r~' +" .... _. .. "_~_-''''''~''''''''''''_.~~II'''J'!''''''_ ...• -_ ..• _ ...... .,.
-The-CING.(Controt):Croup (&-8)
•
The 8 rats vith electrodel in the -cinlulate IYrus, were tested r ~ U acrOIl the standard range of Itimul\j:ion intensitiel (0 - 160uA). Tbe
reluits are plot ted in Fig. 8. FroID tbe figure, it is elear that in
the tail-fliek test there was no effect of stimulation !(3, 21) -
1.43, HS., there was, howl:ver, a marginal, but statistieally
lignificant, variation 01 tail-fliek scores over time !(2, 14) •
4.14, .E<'OS. but the interaction was insignifieant !(6, 42) • 0.98, ,
MS. / From Fig. 8, it is alao apparent that there was no effect of
Itimula~ ion in the formalin test !(3, 21) • 1.30, MS. J no variation
over tilDe !(4, ~ - 0.72, NS. and the interaction term was not
significant !( 12, 84) - 0.89, MS.
Relation.hip between SPA and medio-Iateral location of electrode
sites in the VBT and IC, 1-26.) 1
Four of ~he histologieally determined groups (VIT, ZI, ML, le)
Iha~ed t)le .ame anterior and dorsal targlt coordinates (frolD:-
Paxinos and Watlon, 1982). The.e vere 3.5 1IIID posterior to Bregma and
6.S .. ventral to skull surface, respectively. The target
coordinatel, for the electrodes im;tanted in rats in theae 4 group'
d~ffered onl1 ).n. the medial-lateral extent - froa the medial VBT
(2.75 _ lateral to the aid-line), via the border region of the Zl
and the ML, to the lateral IC (4.0 ... lateral to tbe mid-line). The
continuity of thi, _dial-lateral di.enaion, lent itlelf to a
croll-aite, analyai. of tbe relative effieacy of VIT and IC for SPA.
53
..
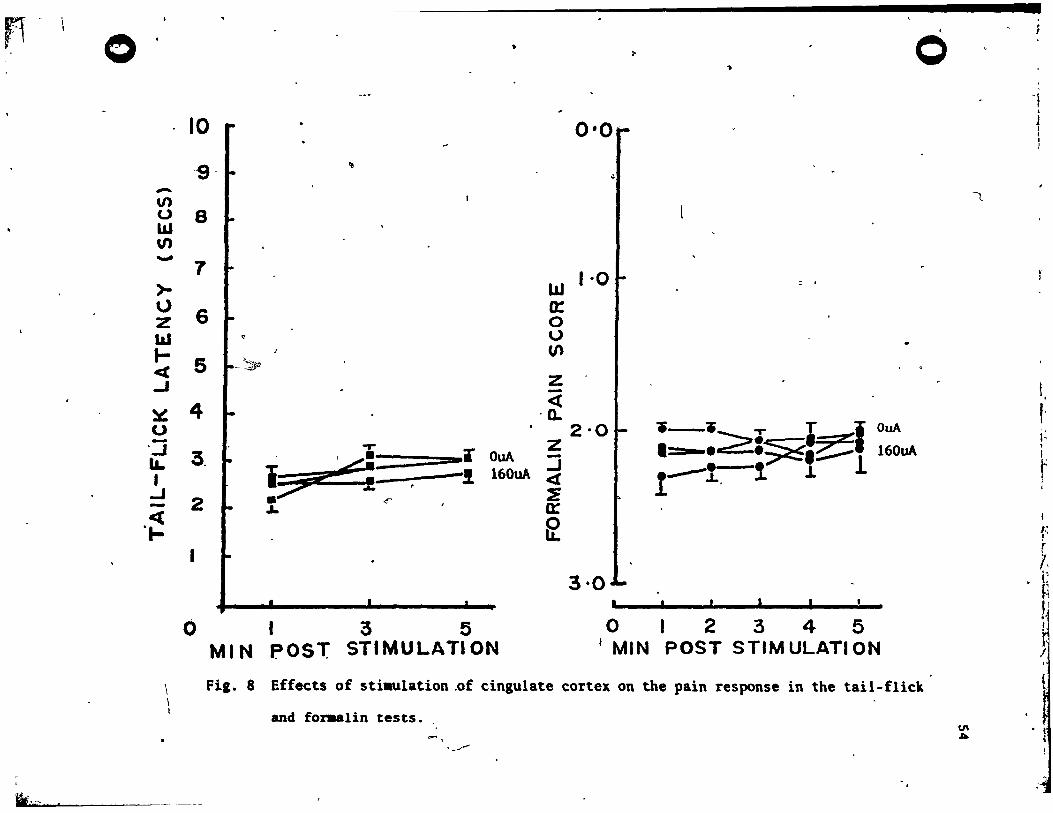
f1 o
ItN"
10
-9. -~ 8 lLI U')
-- 1 >-
6 1
"
Û Z lLI 1-e( 5 .. -~~ -1
~ U ':J 1&. , -1 -ct
1-
4
3.
2
1
()
q,
-r 1 • i~..- L..: ;- OuA .;;:> .!_ -1 160uA
... ~
1 3 5 MIN POST STIMULATION . .
0'0'
w 1'0 0= o U (Il
Z -« 'Q.
2'0 Z :J c:(
~ 0:
~
3·0
4
,
;;>
..
e-e .=--.~:.c:::::;'~1 r--t-rt:;"--l
o 123 4 5
< o
OuA
160uA
1 MIN POST STIM ULATI ON
\ \
Fig. 8 Effects of sti.ulation .of cingulate cortex on the pain response in the tail-flick
and foraalin tests. -,
.----U1 ~
l
'\ • ~ • 1 1
t. f· 1
i; 1
1
t
J.; !
7, i~ . ~ t !

o
o
Actual pl.cement. (.ee Fig.'. lB, le & lE) v.ried around the . tlraet coordinate. in ail 'three dimensions (ineluding 5 placements in
. . the ZI/HL border area). Since tbe dimension of interest vas tbe
.dial - lateral one, limit s vere placed around the other tvo, to
exclude:- A) the placements that shoved extreme variation in the -
otber dimensions, and' B) those that vere in other "off target" sites
(RT & OT). The inclusion parameters vere:- 3.1-4.0 DIO posterior to
Brea", 2.3-4.3 DIO lateral to the mid-line and 6.0-7.2 1IIIIl ventral to
the .tull IUrface. 26 placements were included within these
bound.riu (7 VBT, 5 ZI & ML ,14 IC).
The magnitude of the mean SPA effect at 160uA, vas ranked for e~ch
p~lcement in both the tajl-flick and formalin ~t8. Placement
po.ition vas ranked in terms of medial-lateral position. The rank cl
correlation (Spearman), between SPA 'lnd medial-Iateral position, vas
highly .ignificant for the tail-flicrnst data r; · 0.477, ,2<.001.
The r.nk correlation for the foruaalÏn-test data' waa alao highly
" aignificant r-; • 0.686, ~<'001. In both cases, SPA was grelter at
• more l.terll .ites.
A second an.lysis of tbese 26 placement. was conducted to
dltendne the deal'ee of .. sodetion between .eizure occurrence and
the occurrence of SPA (defined by the criteria .. pecified in the
Mthod .Iction, PI 30), in both pain tute. In tbe teil-fIiek. test,
•• hur. occurrencl va. aianificantly aasociated vith SPA x'< 1 , !! • 10
26) • 16.25, ,E<.OOI. Tbe .... we. trul for the formalin test XO, N
• 26) • 22.29 J ,E<.OOI.
ss
,

-0
. ---- ~
0,
i
"
. " l » e Q lU $1
A final analysis vas con~ucted to determine the degree of
association betveen seizure occurrence and later.lity of the (13 moat
medially ranked vs 13 \DOst laterally ranked) placements. Thia
- X2-association aleo proved significant in the tai l-flick test (l, N -
26) - 10.40,2.<.005 and in the formalin testX~I,.!! - 26) • 12.46,
,2<. ~Ol.
The OT Group (N-3)
3 rats were found to have placement a, latera 1 to the lC, (aee Fig.
lE) in the optic tract region (OT). Tbere vas no effect of
stimulation in either the tail-flick test !O, 6) • 0.68, NS. or in
the formalin test F(3, 6) • 2.02, NS.
The HL Group (N-3)
3 rats had placements in the meduUary lamina of the
ventro-posterior nucleua of thalamua (ML). vhich lies between the lC
and VBT target sites (see Fig. lE). There vas no eff~ct ?f
stimulation in either the tail-flick test !(3. 6) • 1.08, NS •• or the
the formal,in test F,C 3, 6) - 3.94. NS.
The RT Group (N-3)
3' rats had placementa in the reticular nucleus of thalamus region
(RT). vhich' lies betveen the doraal lilDit of the YBT and the IC
regions (see Fig. lE). There v.s no effect of .timulation in either ,
the t.il-flick test F(3. 6) • ,1. 73. NS. or the fonulÏn tut FO, 6) .... • 0.45 J NS.
The Zl Group (N-4)
4 r.t. had pl.c .... nt. in the Zob Incerta re.ion (ZI) betve.n th.
IC and vaT rellons (aea Pi •• lE).
J _
Por 2 of th ••• rat. th. oriaiaal
• f. 0
k ...I.;";_.:c. ........ ~ ...... ,J:# ..... t~ ..... -._ .... ~.
i ,.:;
S6
--

-,
o
- J
target va8 the YBT. Con8equent ly, they vere teated over the range of
etimul .. t ion intenaity Ievela u8ed for that target ai te group (0, 80,
160, 320, 640uA). Of these intenaity levela, only the first 3 vere
comparable to those us.ed "WÏ.th the other tvo rats. Therefore, the
analysi. of this group was rest ricted to these 3 intensity levels (0,
80, 160uA). Again, there wu no effect of st imulation ~ the
tai l-flick test F(2 j 6) • 4.06, NS. or the fornlllin test !( 2, 6) -
2.40, NS.
Incidence of SPA and/or aversion at 160uA .
The number of stimulation si tes in each group and 8ub-group, that
showed:- A) significant SPA. and B) aver~ion; (according to the
cri teria outlined in the Method section: page 29) in each pain test
are tabulated in Table 2. For aU groups (and le sub-groups) the8e
incidences vere calculated for the 160uA stimulation intensity level.
The reBulte of two sub~idiary analyaes at other stimulation
intenaitie. are aho tabulated in the table in rovs identified as:-
80uA:- the incidence of analge.ia and aversion at the 80u!
.timulation intenaity, in the le rats that exhibited .eizure. at
l60uA. and 640uA: - the incidences of analgesi. and aversion in VBT
rat. at the 64"OuA atmulation intenaity. The re8ults for the four "off
tal::let" aroupa are diltinguisbed from the "on target" result. by a
broken line. Tbe total incidence of site. from which signifieant SPA
va. obt'dned .. and the total incidence of .ites at vbicb .timulation ./
va. Averai ve. are tabulated for ea~b pain test Bt tbe foot of tbe
tabl~.
--.. '
57
, .
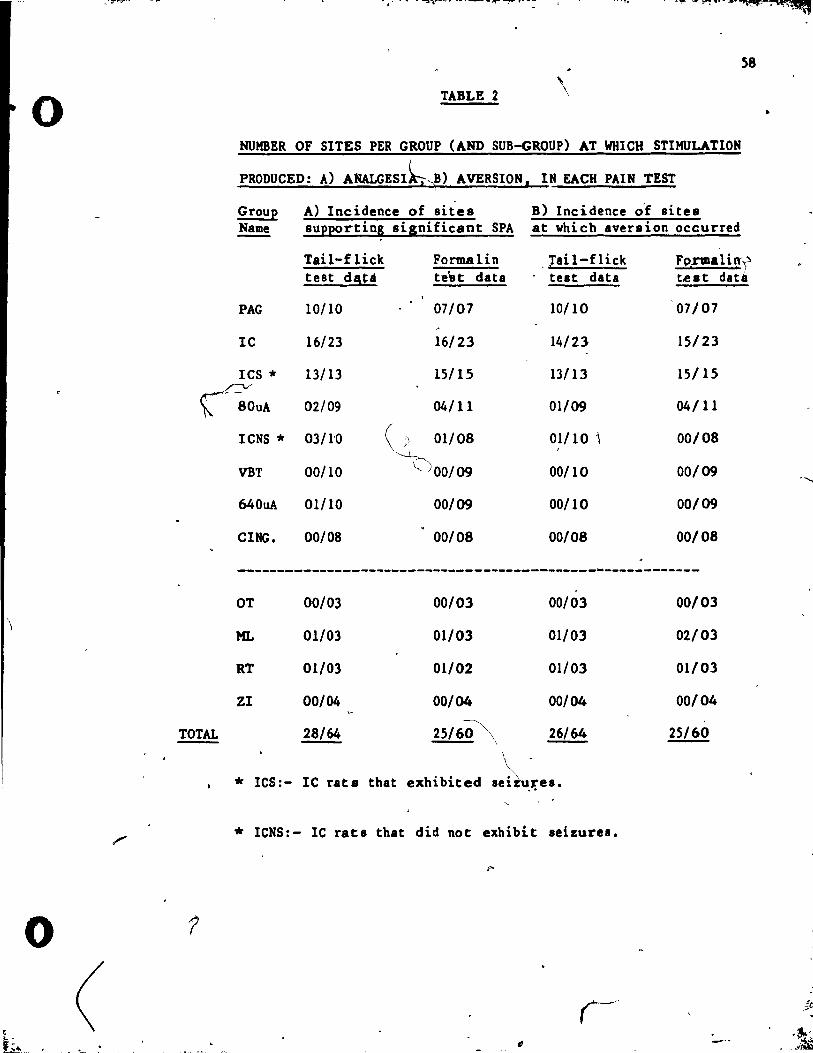
.. ··~·T"'rr·
0
\
0
ç
( ... -f~ ..
'fi,,!. ' •
58
TABLE 2 \
HUMBER OF SITES PER GROUP (AND SUB-GROUP) AT WHICH STIMULATION
PRODUCED: A) AMALGESlk"B) AVERSION, IN EACH PAIN TEST i
Group A) Incidence of sites B) Incidence o-f sites Name supporting significant SPA at which aversion occurred
Tail-flick Forma lin Tail-flick .. Fp"rmali rr,;"1 test d!\U te"st data test data t.e s t dadl
PAG 10/10 07/07 10/10 07/07
lC 16/23 16/23 14/23 15/23
lCS * 13/13 15/15 13/13 15/15
00UA 02/09 04/ Il 01/09 04/ Il
TOTAL
?
. ~ .....•.. .'
ICNS * 03/l'0 (,;l 01/08 01/10 1 00/08 ~
,
VBT 00/10 (JOO/09 00/10 00/09
64 OuA 01/10 00/09 00/10 00/09
CING. 00/08 00/08 00/08 00/08
____________________________________________ w ___________ _
OT 00/03 00/03 00/03
ML 01/03 01/03 01/03
RT 01/03 01/02 01/03
Zl 00/04 00/04 00/04
28/64 25/6.Q\ 26/64
* ICS:- IC rats that exhibited \
Se1ZUJ'e8. , .
* ICNS:- lC rat8 that did not exhibit seizure8.
,--"
00/03
02/03
01/03
00/04
25/60
-'
•
-'-
~ ",
1 -~ 1
.j -~

-
o ..
fL ," . +' f~# i ;'.
Compari.on of intensities that elicited SPA or aversion or both
Thé possibility of an a8lociation between SPA and aversion was
examined by determining vhether st imulation intensities at which
aversion occurred were below, equal to, or above those at which SPA
59
occurred, for each site at which either or both behaviors were elicited.
The incidence of sites, in each pain test, corresponding to each of
these catergories is, tabulated i,. Table 3. The IC group is represented
in 'the table by lts two sub-groups! le rats that exhibited seizures (lc
SElZ) and lC rats that did not exhibit seizures (iC NO SEIZ). The total •
incidence of sites, in each pain test, at which stimuIa:-ion produced
aversion at intensities below, equal to, or above that which eHcited
SPA, is tabulated at the foot of the table. An 'analysis of the
occu.rrence of SPA and aversion at the same stimulation intensities,
across a11 sites, showed that these behaviors vere associated in the \ 2.
tail-flick test condition X(2, ! · 29) • 10.27, .f<.01. and in the ( ~
formalin test conditionX(2, !'- 27) - 16.67, ~<.001.
SPA in the tail-fliek and formalin test compared
the means of the individual tail-flick and formalin test scores (at
the 160uA stimulat ion intensity unless otherwise specified) were
converted int9 mean % SPA scores (see Method section, page 30, for
transfona) to faciHtate a comparison of tail-flick and formalin test
result. within groupa. Tail-flick. and formalin test % SPA means, Ând
Itandard errors of the mean, are represented graphieally for each group,
lub-I1:0UP and .ubsidiary analYlia (le BOuA and YBT 640uA) in .Fig. 9. 1 j \
---- - ~!
"
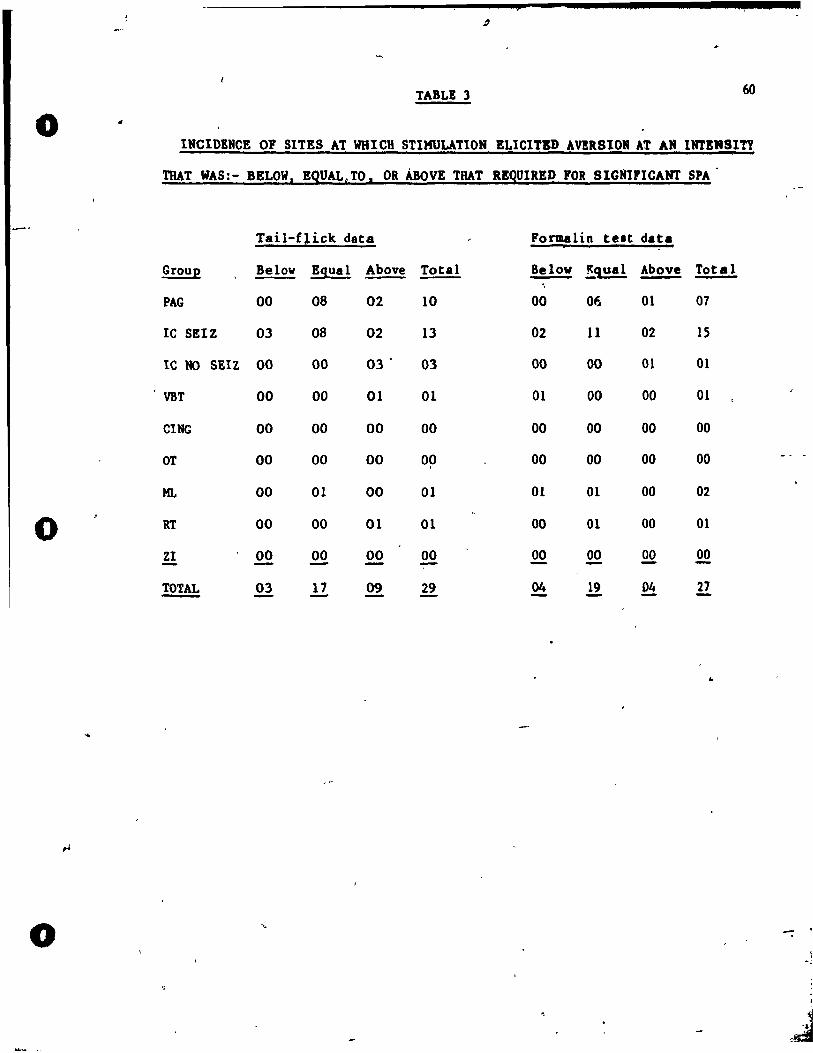
o
0
o
TABLE 3 60
INCIDENCE OF SITES AT WHICH STIMULATION ELIetTE» AVERSION AT AN INTSltSITY
THAT WAS:- BELOW, EQUALe TO. OR ABOVE THAT RBQUIRE» FOR SIGNIPICAtn' SPA·
Tail-flick data Formalin telt data
Group Below Bqual Above Total Below !qual Above Total -PAG 00 08 02 10 00 Of.. 01 07
IC SEIZ 03 08 02 13 02 11 02 15
IC NO SEIZ 00 00 03· 03 00 00 01 01
VBT 00 00 01 01 01 00 00 01
CING 00 00 00 00 00 00 00 00
DT 00 00 00 Op 00 00 00 00
ML 00 01 00 01 01 01 00 02
RT 00 00 01 01 00 01 00 01
ZI 00 00 00 00 00 00 QQ 00 - - - -~ 03 17 09 29 04 .!! ~ 27 - -
..
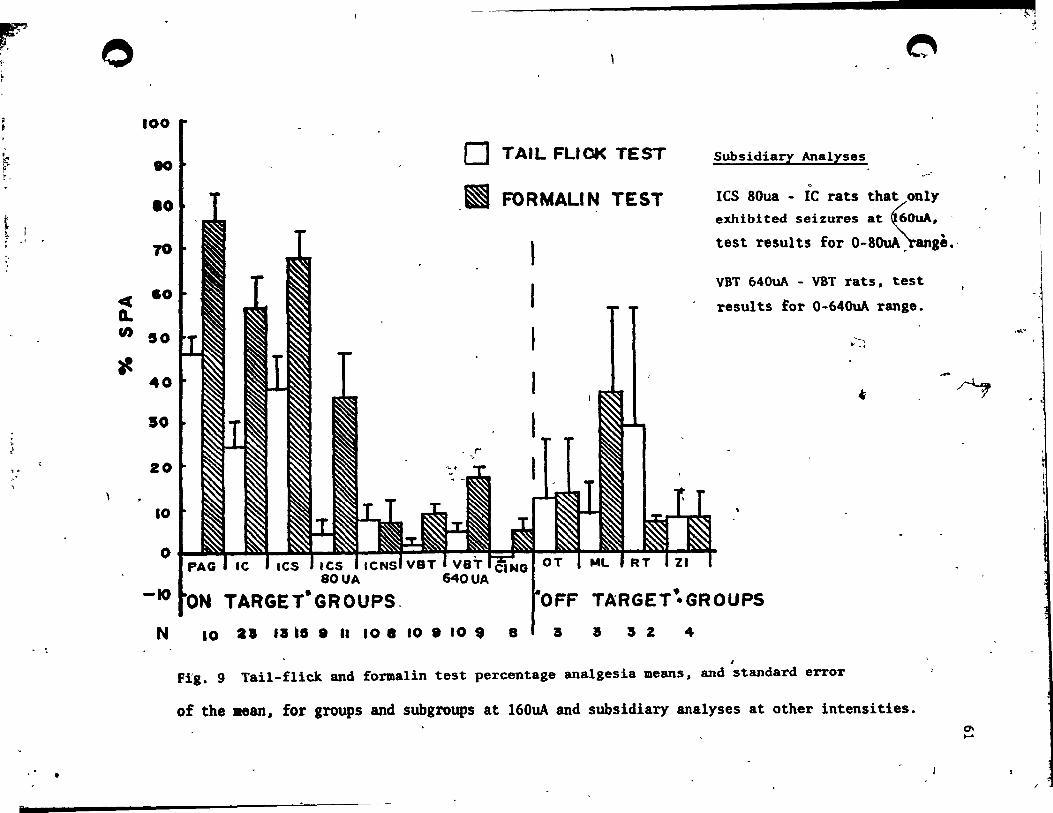
~ !!:".
r
k ( r ..
t ~ . '.
" "
1 1
,1
•
o 100
" .0 70
« .0
G-U) 50
t-40
50
20
10
o TAIL FLIOK TEST
~ FORMALIN TEST
~
Subsidiary Analyses ~.
~
les SOua - le rats that only exhibited seizures at ~OuA •• '
test results for 0-80uA';range.,
VBT 640uA - VBT rats, test
results ~or 0-640uA range.
........ .. . ,
~ -~
o 1 1t."'J '\~ F'N\ .'»1 es
-10
N
ICS 80UA
TARGET-GROUPS.
10 al UI 10 • .. 10 8 10 • 10 $ 8
OFF TARGET~GROUPS a a a 2 4
• Fig. 9 Tail-flick and formaI in test percent age analgesia means, and standard errOT
of the .ean, for groups and subgroups at 160uA and subsidiary analyses at other intensities. 0-....
;~~~
.' .'
,",'
'"

o
o
~ .. -
62
Discus8ion
rrom the results of both group and individual analyses. it is
clear that this 'study provides further evidence for the efficacy of
PAG as a site for SPA in the rat. 'l'he strength of SPA at the PAG
confirms the appropriatenestl of the stimulation parametera employed
and provides the basia for a comparison with SPA obtained (rom the
other stimulation sites. .
The most striking aspect of the PAG stimulation reaulta was that
in order to obtain strong SPA it W8S necessary to apply stimulation
intensities that concomitafltly proauced aversive resctions to
stimulation. The first signs of aversion included urination and
"freezing" behavior which appeared in some rats vith stimulation
intensities as low as 20uA. At the 40uA stimulation intensley level.
signs of aversion included escape and vocalization that, in 2 cases
in the taÏl-flick test and 3 casee ir. the formalin test" were severe
enough to prohibit further testing. Ae the 80uA intensity level mo.t
rats showed more intense escape (jumping up) and vocalization
responses which necessitated the exclu.ion of 3 more rat 8 from
further testing in both pain tests. All remaining rat. a~owed aevere
aversive reactions to 160uA 8timulation.
The 8timulation intenaity threshold for excluaion from te.tiaa at
higher intensities provided an index of aver.ion to .ti.ulation vhich
allowed the association of aver.ion vith PAG .t~.ulating electrode ~
position to be examined. Tbe .tudie. by Vardin et al (1984. l & Il)
had auggested that 1titbin the ventral PAG of the rat there are tvo
-
il
.1
;l,

i'
o
63
"
"pure analaelic" regions, dorso.edia! dorsal r.p,he nucleus (DRN)' anà
ventrolateral PAG, from which SPA can be obtained without concomitant ---aversion to stimulation.' ln the present study, aversiveness of
stimulation wu found to not be associated with the dorso-ventral
location of PAG stimulation sites. However, this apparent discrepaney
may be partially explained by Fardin et al 's obser:vation that the
"pure" ana 1gesia elicited from ventral PAG ~as mainly limited to the
stimulation period and was almost absent after stimulation offset.
Since both pain tests employed in the present study were appliea
after the offset of stimulation, it is unlikely that either would be
.ensitive to "pure" SPA of such limited duration. Furthermore, a more
detaHed analysis of the ventral PAG stimulation sites revealed that
none of the 6 placements were within the two "pure" analgesic regions
defined by Fardin et al. Thus, the association of SPA with aversion,
corresponds well with other recent findings (e.g Fardin et al, 1984;
Ptado and Roberts, 1985).
Althouah it may be difficult to demonstrate a cause-effect
relatidnship, the existance of an association bet'ween the stimulation
inten.itie. at which aver.ion and SPA occurred, for' aIl stimulation
aite., suaaeets that in many ca.es, stimulation may not activate
aotioodceptive .ysteas directly. Instead, the .nalge..,.ia obeerved. may
be medilted by the ,tres.fulne .. of brain stigpllation, sa that .uch {
SPA .. , actuany repusent an instance of the more generai phenomenon
of .tre .. -induced anelgesia (SIA). This effect is often induced
experi .. otaUy by subjectiOS an animal to a period of intense,
iD •• capable, foot-abock. Afçer stimulation, animals exhibit analgelia
'.

o
.... 1 .. _
.... - ! ' ." • "L
o
_5
that, depending upon the temporal parailetere of iu app!ication, '"
or 1118.1 not be med iated by endogenou8 opioUa (e.1 Levis, Cannon and
Liebeskind, 1980; Levis, Sherman and Liebeskind, 1981) •
. The establishment of a distinct "pure" SPA requires the
demonstration of the existence of stimulation sites from which
analg~sia without concomitant aversion can be obtained. If, instead
64
of meeting this criterion, an association between SPÂ and aversion is \,.
observed, this would appear to support the hypothesis that 8uch SPA
comprises a form of SO.. Simi lar cane lusions were reachEd by Fardin
et al (1984, 1) who suggested that, in su ch cases, SPA ~y be a ,
.• consequence of aversio!" and by Prado and Roberts (1985), who sta'ted
"
,Chat "there can be litt le justification in assigning a cprimary
antinociceptive role to a brain area which evoku atrons eacape
reactions when st imulated."
Both the group and individual rea,ulta indicated that dngul.te
-C\lrtex did not support SPA at intenaitiea ranging up to 160uA. This
demonltrates that at this site there is a negligible generalized
effeet of stimulation and prov:ides support for the site-apecificity
o~ SPA. Interestingly, stimulation of dngulate cortex did pot induce
Appreciable aversive reactions.
Group and individusl results of VBT Itimulation over the extended
range of stimulation intenlities (O-640uA), indicated veak SPA in
,both pain tests. However, even et the 640uA st imulat ion intendtr.
the overall magnitude 'of the group 'SPA effect. for bath pain tuu ,
vas lus than that seen at the 40uA .t imul.tion intenaity at the ,AG. Indivi~ual relulte Ihoved tbae, 1 of 10 rat. in t~ tail-fliek te.e
r
• -'

Ct
l >, ,.. \ ~
<]ti,.:#. '.
.-
65
1
condition and 0 of 9 in the formalin te':i: condition, supported
lignifieant SPA It 640uA. ,
A correlation WIS found to exist between tbe magnitude of SPA and
the' latentity of diencephalic stimulation sites. SPA inerelSed from
inlignificant levela at the IDOst medilll VBT sites to intermediate (
levels at Zl and ML sites (tloff target ll regions which lie between VBT -"'>
and le) to a maximum at sites in the lateral le. This suggests that
the 'Îlngle case of tail-flick test SPA obtained from the VBT site at
, -the 640uA stimulation intensity may be due, to spread of current ,to
the adjacent le. Thus, the present result s do not cont rad iet the few
put studies which examined VBT as a site for SPA in animais and
!ound it ineffective (e.g Mayer and Liebeskind, 1974; Goodman and
Bolcombe, 1916; Prado and Roberts, 1985).
VIT stimulat ion over the standard range of intensities (O-160uA) ,
did not induce the extreme aversion characteristic of PAG
stimulation. lnstead, as indicated in 'the results, 160uA stimulation"
of the YBT resulted in a pat tern of responses which appeared to be of
• predominant ly mOtor, rather than aversi ve, nature. Thi s pattern of
relpon8es, which included rhythmic mouth movement (jaw opening) and
head-nodding, corresponds weIl with descriptions of the intermediate , ' .
shgll of limbic motor seÏEure behavior (e.g Racine, 1972; Racine,
Livina8tone and Joaq~,in, 1975; Collins, Tearse and Lothman, 1983).
None ,of the YBT rata progreased to the final stages of limbic , ".
IIOtor seizure behavior at the 160uA intensity level, but at 640uA the
ujority of rats exhibited full motor seÏEures. The behavioral
unifestations of these eehures included rearing and falling
• 1

o
o
o
.... -
66
backvards and "roll ing over" le.ding to full tonie extension and 10 ..
of postu~l control. ~,
Despite the severity of these behaviors.
however. even the higheet in'tenaity of VBT stimulation did not
produee the degree of aversion seen with much lower stimulation
intensities at the PAG. 1
The results for the IC group as a whole. indicate that, the IC is
indeed an effective site for SPA in the rat, 88 it is claimed to be
in man (e.g Adams et al, 1974; Turnball, 1982; Namba et al 1984;
RiChardson. 1985). However, this interpretation is tempered by other
reeults which indicate that SPA at the IC is contingent upon the
occurrence of escape followed by convuls~ve responses to 160uA
stimulation. The st imulation induced seizure behavior wu remsrkable
in its explosive _qp.set and the violence of subsequent convulsion ••
Unlike the relatively mild and aversion-free progres~ion of lim'ie
motor seizure behavior "resulting from VBT stimulation, IC seizure
activity appeared to model the "grand mal" respanse to
electro-convulsive shock (ECS). Such a grand mal tonie-clonie
response to 160uA stimulation oceurred with stimulation of 15 of the
23 IC stimulation sites Cin one or both pain tests) and 1 other .ite
in t'he adjacent reticular nucleus of thalamus (RT) region. The
occurrènce of major convulsive re.ponses folloving le stimulation i.
cODsi,tent with the previous observation that IC hu one ~f the
lovest convulsion thresholds in the cat br.in (Gibbs and Gibb ••
19~6)' Simi !arly, tl\e low convulsive thre.hold of the neighborina IT
resion corroborates a study by 8er,_nl), Co.tin and Gut .. n (963) who
found a "low convulsive thre.hold area" in the .. aeneephalic
\

o
o
67
reticular foraation of the rabbit. Reverthelell, it remains
reurkable tbat this relatively low level of IC 8timulation wu
capable of triggering an ECS-like level of convulsive reaponse.
The reluits for the two behaviorally' defined sub-groups of lC
group rat s (sei zure vs no sei zure sub-groups) emphas ize the
..... ociation between overt seizure activity and reduced responsivic·y
•
to painful stim'l1,i. The results of the no seizure sub-group indicated
• lack of SPA in the group analyses of both pain tests and individual
reluits showed that SPA occurred in only 3 of 10 rats in the
tail ... flick test condition and 1 of 8 rats in the formalin test
condition. On the other hand, for thf! IC seizur:e sub-group, both
group and individual results showed a strong SPA effect tha t almost (
co.parel in magnitude to that found vith the same intensity of PAG
stimulat ion (see Fig. 2, Fig. ,4 and Table 2). This 8uggest s that SPA
from the IC may be contingent upon the occurrence of stimulation
induced convulsions. However, another result suggests that this may
not be entirely true. The analysis of SPA in seizure sub-gro~p rats
at a pre-convulsive stimulation intensity (BOuA) indicated marginal
SPA in the tail-fli.ck test and a relatively strong SPA in the
formalin test. There are at least two alternative interpretations of
thil finding. The pre-convulsive SPA may be associated with covert
.eizure activity or instead it may represent a veak analgesia that is 1
di •• ociable from the Itrpnler seizure-allociated effect.
An a~.oci.tion between stiaulation-produced seizure activity and , ~
aaalael\a bal been docu"nted in leveral relatively recent paperl. In ~
1978, Abbott and Nel •• ct reported that Iti.ulation of tvo li~e

o
o
o
-_.....- " '. j t 4.4 '. $ '.-1
.. 68
structures, the la-teral septal region .nd the dor.al hippoc:.mpu., .
produced .naigeaia in the formalin test. This .naigeai. oc:c:urred only
when stimulation produced eleptiform .fter-discharges in thele are'l.
However, the behavioral expression of })re-kindled after-disch.rgel
was li~ited to immobility and oeeasional "vet dog shakes" (a
eharacterist ic of the esrly stages of limbic motôr seizurel>.
Repeated stimulatiorl of the lateral septum kindled convulsion. and
abolished analgesia. Frenk and Yitzhaky (1981) a1so found that
analgesia accompanied pre-convulsive', but not post-convulsive
after-discharges in amygda1oid-kind led rat s. CIe. rly then, limbic
kindling activates a different set of responsea to those found vith
IC stimulation. Stimulation of limbic structures produces only
pre-convulsive analgesia without aversion, vhereas IC stimulation
produces strong aversion and major convulsions (without kindling)
folloved by analgesia.
Another seizure/analgesia auoc:iation was reported by Holaday,
Belensky, Loh and Meyerhoff (977) who found th.t, in rate, BCS
elicited analgesia that was attenuated by the specific opiate
antagonist, na loxone. As ind icated previousl~ 1 leS indueee the .. Ille
type of major convulsions observed vith IC stimulation and both
produce post-ictal analgesia. Therefore, BCS would leem to provide a
better model of IC stimulation than the kindling of li';bie
structures. Urca, Vit zhaky and Frenk (1981) reproduce4 the Holaday et
al finding and confirJDed that the poet-ictal analaeeia va. partiall,
reveraed by naioxone. -Both Bolada, et al and Urc:a et al interpretéd \
their finding. al evidenc:e tbat !CS-induc:ed analaeaia il .. diated bJ
--

o
..
o
,#'-, -
69
endolenou. opioide. Purthec .upport for this hypothesis was provided
by Levis, Cannon, Chudler and Liebe.ltind (1981) who found that .. naloxone revereed ECS analgesia in hypophyseetomized rats, implying
that the effect is mediated by opioids of central origine
ln the present study it was observed that a second behavioral
effect accompanied post-ictal analgesia in rats stimulated at the IC.
AU rat. that exhibited convulsions showed a marked post-ietal
behavioral depression that appeared to resemble pharmacologieally
induced eatalepsy. The results of the bar test (e.g Costall and
Naylor, 1974) appeared to confirm the cataleptic nature of this
phenomenon.
It was suggested 'above chat 10 stimulation < resembles ECS both in
\
---terme of the type of convulsive reeponse induced and the occurrence
of post-ietal analgesia. Further evidence for a similarity between
thue two effeets is provided by the occurrence of post-ictal
catalep.y in rats that had received ECS (e.g Holaday, Belensky, Loh
and Meyerhoff, 1978; Belensky and Holaday, 1981; Urca, Yitzhaky and
Frenlt, 1981). Interestingly, a catalepay-like post-ieta 1 depression .P
<a. oppo.ed to pre-ictal analgesia) was 8lso observed in
a.ygdaloid-kind1ed rats (e.1 Prenk, Engel, Ackermann, Shavit ~llld.
Liebeakind, 1979; Yitzhaky, Frenlt and Urca, 1982).
Tbere vere, however, important inconsi stencies within, and
bet ••• n, the ECS and .. yzdaloid-kindling paradigme. In their' < 1_981)
BCS stud" for ex •• pIe, Belensky and Holaday measured catalepay vith
a bar tait li.ilar to that used in the preeent study, while in tbe
- !CS Itudiel of Urca et al (1981) and Frenlt and Stein (1984), the

ii --
70
o lat~ney ot post-ietal "spontaneous dghting" was used a. a mea.ure of
eatalepsy. In these latter studies rata diaplayed latencie. of
between 1 - 2 min post-BCS before righting th~mselves when plaeed on
their backs. In an earlier definition of eatalepsYl, Munkvad.
Pakkenberg and Randrup (1968) specified that cataleptie animaIs right
themselves when plaeed on their baeks. Costall and Naylor (1974)
operationalised this definition by adding that such righting should
oeeur within 10 secs. thus raisifig doubts about the validity of ,
delayed righting as an index of eatalepsy. Finally, in their
amygdaloid kindling studies,'F~enk et al (1979) and Yitzhaky et al
(1982) adopted another potentially ambiguoua me.sure which indexed .. the offset of post-ietal depression by the rats Hut step forward.
o These inconsisteneies would have little signifieanee if it wa.
clear that 'catalepsy comprises a unitary ph nomenon that lS not /\-/'
behaviorally dissociable. Unfortunately h
'" pharmacologieal studies suggest that catal compriaes two
behaviorally and pharmacologieally distinct syndromes: a
\neuroleptic-induced "flaccid catalepsy" and an opiate-induced "riaid"
eatale~sy that is more properly termed "catatonia" (e.g Cott.ll and
Raylor, 1974; De Ryek, Schallert and Teitelbaum, 1980; Rondeau,
Turcotte, Young and Herbert, 1982).
This distinction allow8 a reinterpretation of the leS work of
rese_rebets who used the 8pontaneou~ht ing meaaure, and .u"eata
that aeeording to the. definition of -catal~p.y of Munkvad et al (1968)
they vere, actually mea.uring eatatonia rather than catalep." aa
o tbey auppo.ed. Furthermore. the distinction led Mya!obodsky and Minlk
~ ~-ir:. • th r...,:M., ~ .. •

/Pu
r'
(1981) to tut the ECS po.Jt-ictsl effect to 8ee if it was more
accurate1y eharaeterized 8S "true" cata1epsy or eatatonia. Their
re.ults indieated lhst post-ic~a1 rats showed symptoms of both
behaviors. although catatonia appeared to be predominant after ,
repeated ECS. It appesrs then. that an opiat~-like catatonia may
comprIse a signi ficant portion [r0f 'the ECS motor effect \ This
interpretation LS criticsl because Hys1obodsky and Minsk (1981). and
both righting measure studies (Urca et al. 198'1; Frenk and Stein.,
71
1984). shoved that ECS induced "catatonia" vas mediated I>-y endogenous
opioids. Therefore. ECS may concurrent ly indu ce anMlgésic and
catatonie behaviors whi,~h are both mediated by endogenous opioids.
Frenk et al (1979) and Yttzhaky et ~1 (1982) c1aimed that bath ~
pre-convulsive analgesia and post-ict~l depression. in
"' amygdaloid-kind led rats. were opioid mediated effect 8. Hovever, the
c!1ear differences ln sequence and intensity of these behaviors,
compared to those seen vith lC st imulat ion, suggest that the former "
provide fev clues about the neurochemical basis of t~e latter.
~ ________________________ ~_T_h_e __ s_i_m_i_1_a_r_i_t~y_betveen the ~ehavior81 effects of ECS __ a_nd~I~C~ ________ __
\
t d • t "1
stimulation raises two issues. The first concerns the p6ssibiiity
o that both effects share 8imi~8r neurochemical mechanisms. The second, \"
perhaps more fundamental issue, concerns the interpretation of
behavioral melllures that are a,.sumed to diseriminate between the
sensory (attalgesia) and mator (catalepsy) effeç,t8 of IC stimulation.
The urked behavioral reseablence betveen the effècts of ECS and
le sei.ulation i ... rred on1y. by the attribution of a ca~atonic l
COIlpODlnt to the' ses et fect vbUe the bar tes t of t'he le st imu lat i,On

o
o
o • 1
z q : J il e
72
effect appeared to indicate the occurence of catalepay. Recent ly
however, De Ryck. and Teite1baum (1984) have noted that the bar telt
cannot differentiate neuroleptic catalepay from opiate/opioid .. catatonia. Thus, it - remains possible that the behavioral profile. of
the ECS and le post-ictal effects are congruent in terme of both
~nalgesia and catatonie components. If this is the case it would
suggest that both of the IC effects may also be mediated by
endogenous opioids.
An argument against direct opioid mediation of the le post-ietal
effects is provided by pharmacologic"al evidence which shows that .,
there i8 little opiate/opioid receptor binding in large fibre bundles h
such 8S the le (e.g Kuhar, Pert and Snyder, 1973). Nevertheleu. the
~gnitude of the le st imulat ion-induced convulsive effect suggut.
that there is considerable spread of seizure act,ivity to otifr brain
regions. Thus, site-specific receptor characteristics are probably
irrelevant to the issue of the neurochemica 1 basis of the le
post-ictal effects. Therefore, it remains possible that both of the f
lC post-ictal effects are mediated· by en~~u.nou~-opi&i-4&r -C-au-ly,
f~rthërruearcb îS- required to elucidate this issue. Future studiee
should include: pharmacological manipulations which can diacriminate
the degree and specificity of opioid involvement in lC poet-ietal .
effeeta ~ a me8sure of electrophyaiological activity to provide a IDOre
accurate measure of seizure activity, and behavioral _ .. urel vhich
can disc,rillli'nate be~ween the catalept ic and catatonic componenu of
the general effect of "catalepsy".

rt' ". ~"
A8 indicated previously, the coexistance of post-ietal analgesia
and catalepsy/catatonia, in both the ECS and lC stimulation \
paradigms, raises the more complex issue of the ~havioral
diaaociability of these two effects. The presence of both of these
73
states il inferred from reduced motor resporlsivity. Since pain tests
such as the tail-flick and formaI in test are limited to a measure of
reduced motor responsivity to a painful stimulus it would appear that
reliance 00 su ch measures might lead to a confound ing of SPA vi th
mator dYlfunction.
SOlDe evidence that components of analgesia and "catalePjY" are
dissociable ia- pro,:,ided by the ECS study of Hyslobodsky and Mintz
(1981). They reported that during post-ictal immobility rats had
rapid righting responses and responded to tail-pinch. The presence of
righting defines the immobilityas neuroleptic-type catalepsy, rather
thaD opiate-like catatonia (e.g Hunkvad et al, 1968) and the lack of
tail-pinch analgesia is evidence that catalepsy and analgesia are
behaviorally dissociable. They also reported that a naloxone
____ reversible, "rigid" catatonia built up over days of repeated ECS. An
analgesia to tail-pinch, that was not reversed by naloxone, also
built up after repeated ECS, roughly paralleling the development of
catatonia. Thul, opiate-like catatonia is evident ly behaviorally
dillociable from non-opioid mediated analgesia. The critical iaaue
re_ioing ia vhether the opioid mediated, and therefore presumably,
opiate-induceable, forms of analgesia and "catalepsy" are
behaviorally diaaociable. Studies by Thorn-gray, Levitt, Hill and
Ward (1981) and Thor"a-gray and Levitt (I983) addre8sed this iuue. In

o
o
. (
o
.. te,
74
their 1983 study, Thorn-gray and Levitt con~ed that
opiate-el icited al'lalgesia and catatonia may be neuro-anatolllic:ally
distinct phenomena. At 28 rat brain sites, analgesia (mea.ured by the
fl inch jump test of Eva118, 1961) and catatonia (bar test) vere
produced concurrently by micro-,injections of etorphine. At a further
29 sites, etorphi ne produced catatonia only and at 9 si tes it
produced analgesia only. Interestingly, e,torphine did not elicit
either behavior when micro-injected into the IC and dual behavioral
effeets' occurred most frequently with micro-injections into the PAG
and its vicinit,.
If the le post-ictal effects are opioid mediated the resulte of
the Thorn-gray abd Levitt study suggests two points. Firstly, it
provides further evidence that IC post-ietal ef fects do not involve
local opioid systems and mU$t instead involve other brain regions,
perhaps including PAa- Secondly, if this is the case, then it might
not be possible to behaviorally differentiate opio id medi.ted
analgesia and catatonia. If, hoveyer, eithe~ or both of the IC
post-ietal effeets are not opioid mediated,J previou8 etudies 8uggUt ~
that they should be behaviorally dissociable. Clearly, thi. complu
issue remains unresolved. Pharmaèological and eleetrophysiologieal
analyses combined vith improved measures of catalepsy and catatonia
are neee8sary to ~etermine whether le post-ietal ~\nalgesia and
"cat.-t'epsYU are confounded.
ln the present etudy, the results of the group analyse. of the \
four "off t.rget" group. (optie tract .i te.:- OT, medull.ary la .. in. of
,VPL sitea:- HL, reticular nucleus of thala.ul .ites:- KT and zona
j

G
. .
c
o
incerta eites: - zI) ind icated that GOne of these si tes support SPA.
However. the lack of signifieance of these results may be partially
attribut:able to the sma11 number of an~ma1s per group (see Table' 1).
The individual reluIts show that 2 of 13 rats showed sigificant SPA
(more than 4 S.D. 's above baseline) in the tail-flick test and 2 of
12 showed signifieant SPA (more than 4 S.D.· s below baseline) in the
formalÏn test. For these groups, the strongest SPA was found in the
ML and KT groups and the weakest was found in the ZI group •
Similarly, ML and RT sites showed the greatest aversion to
et imulat ion whi le those in the Zl grou,p showed the least aversion to .'
stilllulat ion.
7S
As outlined in the introduction, the primary aim of this study was
to evaluate the rat as a behavioral model of clinical SPA at
brainstem and lateral diencephalic sites. To sU1llllarise the relevL-~-~~
points of the discussion:- group and individual results showed that ~
st:rong SPA, over the '()-160uA range of stimulat ion intensites J was / •
. limited to PAG sites, le sites from which convulsive seizure activity'
.. wàs elicited and a minority of "off target" sites adjacent to le. VBT
at: imulat ion over this range did not prt1duce appreciab le SPA and, even
over the 0-640uA intensity range stimulàti~n of VBT produced only
weak SPA that was associated with limbic seizures. Np SPA was
obeerved at cingulate cortex sites or at le sites that did not
aupport seizurea.
AU regions that supported st rong SPA aho ahowed sÏ'gns of a
concollitant strong aversion to atimulation. Therefore, parsimony
luagelU that in thue cases, SPA may represent a, case of the IDOre

o
" "
o
o
;
76
general phenOlllenon of stresa-induced analge.ia (SIA: e.g Levie,
Cannon and Liebeskind, 1980) rather than the dist inet, aversion-free,
form observed by Fardin et al (1984): Furthermore, a critical
analy8i8 of the lC po8t-ictal SPA effect 8uggeats that it repreaentl
an SrA effect that msy be partially confounded b~ concurrent
seizure-induced inhibition of motor responsivity.
Thus, the findings of the present 8tudy appear to be at vadance \,
with the majority of previous animal, and clinical, SPA studies,
which" haye generally, either diacounted, or faited to report an
a8sociation between stimulation-produced analgesia and ~ver.ion. The
• paucity of prevlous reports of the averslveness of SPA may be
explained by the fact that, 1.n cont ra8,t to mo8t past studies of SPA
\ in the rat (e.g Akil and Hayer, 1972; MaY6J:" and Hayes, 1975; Hayer
and Liebeskind, 1974; Hayer, Akil, Liebe8kind\t al, 1971; Rhodes and
'--Liebeskind, 1978), the present study examined SPA in freely moving
, ,
rats. In previou8 studie8, rats vere often restrained rats during
- pain testing, which mey have pre-~ented the observat ion of the
behavioral manife8tat ion8 of aversion to st imulat ion. Nevertheless,
it remain8 supri8ing that the occurrence of aversive reactions to the v
antinociceptive'effects of mesencephalic stimulation in the rat, have
only been reported by four authors (Pert and Walter, 1976; Yeung,
Yaksh and Rudy, 1917; Fardin et al, 1 & II, 1984; Prado and Robert.,
1985). Of these, only the AKrn~&.. of the two most recent etudiee bave
conducted 8ystematic investigations which considered the occurrence
of aversive reactio~ limitin condition in the production of
"pure" analgeaja.) Thu., s~pport Jor the finding. of tyt pre.ent
. L--
li.!

c
o
1tM" ... _
"
1,< i
-
etudy, and for the hypothe.i. that SPA 'ob.erved in rats may be
••• ociated with the .tress ~f stimulation, ha. only appeared very
recently (Fardin et al, 1984; Prado and Roberts, 1985).
-77
The appaunt diacrepancy between the reluits of the present study
and those of clinical SPA studies, resu lts from the fact that the
majority of the latter do Qot report an 'association between analgesia
and aversion. However, a few clinical SPA studies have noted
unple8lant side effects to stimulation. For example, SPA from the.
PAG. bas been reported to be accompanied by 8~nsation8 of nausea, ,
nystagmus and vertigo (e.g Richardson and Akil, 1977a; eosyns and
Gybels, 1979). In a more extensivé analysis, Richardson (1982)
stimulated medial sites in man, ranging from PAG to basal
hypothalamus. Basal hypotha14mic stimulation produced partial pain :?
o '1 relief togetber with side effects auch a8 fluahing and diplopia,
whi le inferior PAG sites produced dist ressing side effects below,the
Itimulation level required for pain relief. Thus, for mesencephdic en
sites at least, there is some evidence tha't the analgesia/aversion
auociation observed in the present study is supported by clinical
findinss. The rat may, therefore, be a relatively good model of
clinical SPA, and it8 side effects, 'at mesencephalic sites. Further
investigation of this region may provide information that will allow
clinicians to develop techniqu'ès to minimise the aversive side
effecte of clinica 1 SPA.
\ With laterai diencephalic sites J however, a search of the
literature reveals an absence of any reports of aversive side effects
to SPA. This .. y be a consequence of the definition of clinical ,
1 j
, .

o
o
o
78
succesa, which precludes the occurrence of severely averai ve side
effects to stimulation. The aversive aide effectlJ reported vith SPA
from PAG aites in man, are aU relatively mild, and therefore
clinically acceptable -if compenaated for by a sufficiently effective
SPA. The same effects may be more stressful to the rat because the
rat h~s no comprehension of the situat ion and cannot relliEe the ,
beneficial effects of the stimulation. It is also pouible that
aversive side ef fects of le and VBT st imulat ion in man, may include
severe motor and convulsive behaviors that would pre vent a report of
clinical success, despite the occurrence of analgeaia.
Nevertheless, it is possible that the absence of reports of
aversive side effects to clinical SPA. from lateral diencephalic
sites. Dl8Y simply be because these effects do not occur in the
clinical situation. If this ia the case. then the reaults of the <il
present study remain discrepant with cl ioicai research and would
" suggest that rat and man are not functionally homologoulJ in terme of
SPA from IC and VBT stimulation sites. However, even this
interpretation must be tempered by the recognition that the pain
assessed in this study is ,not necessarily comparable to that
described hl man. In man stimulation is applied to retieve chroni~
pain of pathological origin and ia generally ineffect i ve againat
acute pain (e.g Cosyns a~d Gyb.,ls, 1979; Richardson and Aki l, 1977a).
This principle' was cited in the introduction as the rationale for the
'l"" inc lus ion of the forma li n test' in the present study. lt vu allo
ootect. in the introduction that clioical SPA frOIl lateral dienceph.lic
sites is particularily effective for .enaory de_fferent.tion and \
a

,
( " o
79
-central pains whilè PAG stimulation i_ more appropriate for
peripheral pains. The formaI in test -may be a useful model of chronic
pain of peripheral pathologieal origin, but it ia, less libly to be a
good model of central and deafferentation types of pain. Thus,
although it can be concluded that the pain tests employed in this
.tudy show that th~ rat is not a suitable model of clinical SPA at
lateral diencephalic sites, it is not possible to conclude that thi s
will remain true for aU future animal models of clinical pain.
A subsidiary aim of the present study was to examine the raIe of
pain type by contrasting the relative strength of tail-flick and ~
formalin test SPA at each target site. To facilitate this comparison,)
individual resu lts were transformed ioto percentage analgesia scores.
These vere then averagéd to provide group means, and standard errors
-of the mesn (see Fig. 9). From the figure, it is elear that the
etrongest SPA was found at PAG and IC sites. In all analyses of these
tvo groups and their sub-groups, in which st rong SPA occurred, the
results show st rength of the formalin test SPA effect to be
approximately double that of the tait-flielc test effect. In the
pre-ictal analysis of the lC (seizure) group the differential between
taÏl-flick- and formalin test re8ults is even greater, but in the same
direction. Only in the case of the IC (no se izure) group, for which
no lignifieant SPA oceurred in either test, was there a minimal
di-fferent ial betveen tail-flick. and formalin tests. The figure dso
i'liultrates the relative inefficacy of rat VBT as a site for SPA,
even when the et illulation inteneity ~as increBsed to 640uA. Despite
the ",.kne .. of YBT effectl, hovever, the saae pattern of a stonger

o
o
,/
foraHn SPA /tfect is evident in 'the results of the 160uA and the
640uA analyses. Finally, the control group (Cing.) did not exhibit
SPA in either test and therefore did not show a large diffential
betwe~n tests.
, The "offftarget" groups do not show a consistent differential
between tai I-flick and formalin test SPA, but the standard errara of
these sma11 N groups cautions against drawing any conclusions from
this set of data. The conc lusion that can be drawn, il that in "on
target" groups, if there is any SPA at al l, SPA in the formalin test
appears to be ~tronger than that in the tail-flick test.
'This relatively constant differential has significance for the
hypotl)esis that the,se two tests dif ferent iate
mediated by distinct neurochemica l and neural
betweer Î
systeJbs
pa in types
(Abbott and
Helzack, 1983). A clear dissociation of pain test perce,ntage
analgesia results between sites, would be evidenced by a reversal,
across groups, of the relat i ve st rengths of tail-fl ick and for .. lÏn
SPA. lnstead, it is clear from Fig. 9 that no such reversaIs occur
for any of the analyses of "on target" groupa which support
~ignificant SPA. Therefore, the data do not support the hypothesia
,that tail-flick and formalin test pain types, are relieved by
different st imulation-acÙvated mechani8ms'~ at PAG and le situ.
Strong SPA was limited to the PAG and le regions, and of these.
only the PAG supported strong analgesia wi thout el icitit;'1 .e~zure ••
ThuI, a1though for both the le and PAG groups the foraal in tut va.
80
IIIOre sensitive to SPA than the taï I-flick test, the aide effects vere,
different. If the si.ilar pain test differentials reflect the
\

()
o
81
activity of COlBlOn, analgesic systems, this difference appears to be o
contradictory. Bovever, the hypothesis that both IC-post-ictal, and
PAG effects actually constitute streas-induced anaigesia, suggests a"
cOlIIIIOnaHty betveen the mechanisms underlying these effects.
The fact that in the formalin test, SPA is about double that seen
in the tail-flick test, relllains to be explained. If PAG and IC SPA
did indeed invol ve common systems vith a predisportition tovards
mediation of formalin test SPA, this would explain the pattern of
results obeerved. However, the fd'rmalin test may simply be more
sensitive to SPA. In this case, the apparent differential betveen the
results of the tvo tests can be regarded as the consequence of a
scaling artifact. For example, a simple manipulation of the
tail-flick test data would raise tait-fl ick percentage SPA scores to
parity with formaI in test scores. This could be achieved by assessing
tail-flick latencies with a cut-ôff close to 7, rather than 10,
seconds. Alternaci vely, the differential might be removed by reducing \
the dose of formalin injected into the rats hindpaw. Thus, despite
the use of percentage SPA scores, the reBults of the two pain tests
are not readily 'cogaparable in absolute terms • . -- ~ "
In conclusion, the lack of any reversaI of pain test.sensitivity
betveen IC and PAG group results, and th~ likelihood of a scaling
proble., preclude any definite conclusions as to the signifieance of
"pha.ic" vereua "tonie" pain types in SPA effects at IC and PAG. The
lack of bebaviora! dis.odation of the Analgesie effects of
Ici.ulation of tbeee two regions lUIy reflect t~e activation of •
co-.on .treee-induced 'llalgeai •• ,ste •• c

-
o
Co )
he
R.eferencea
Abbott, F. V., FraoklinK. B. J., Ludwick, R. J., & Mehack, R. (1981). Apparent lack of tolera-nce in the formalin te.t IUlaelt differeot mechani 8ma for 1IIOrphine analae.ia in different type. of pain. Pharmacology. BiochemÎltry and 1Sehavior, 11,' 637-640.
Abbott, F. V., & Helzack, R. (1978),. Analaesia produced by electrieal stimulation of limbic. structures and itl re lation to epileptiform after-dia~harges. Experimental Neurology, ~, 720-734.
82
1 Abbott, F. V., & Htlzack, R. (1982). Brainstem lesions dis80ciate
neural mechanisms of ~rphine analgesia in different kinds of pain. Brain Research, lli, 1~9-1SS.
Abbott, F. V., & H*.zaek. R. (983). Dill8ociation of the mechanisms of stimulat' n-produced analgesia in tests of tonic and phasic pain. In J. J. Boniea & A. 199o (Eds.), Advanceà in pain research and therapy (Vol. 5, pp. 401-409). New York: Raven Press.
(
Abbott, F. V., & Helzack, R., & Samuel, C. (1982). Horphine analgesia in the tail-flick and formalÏn teats ~s mediated by different neural systems. Experimenta 1 Neurology, 21., 644-651.
Adama, J. E., H080~ucbi, Y., & Fields, H. L. (1974). Thalamic syndrome and e1ectrical stimulation of the internai capsuie. Journal of Neuro8urgery, ~, 740-744.
AkÏl, B., & Mayer, D. J. (1972). Antagonism of stimulation-produced analaesia by p-CPA, a aerotonin synthesis inhibitor. Brain Rese.rch, 44, 692-697.
Akil, B., Hayer, D. J., & Liebeskind, J. c. (1976). Antagonism of stimulation-produced analaesia by naloxone, a narcot i c antagonist. Science, l2.!, 961-962.
Akil, B •• Watson, S., & Barchal, J. (1976). lncrease in opiate-like factors upon a~algetic electrical stimulation in rat-brain. Neuroscience Abstracts, 1, 563.
Balagura, S., & Ralph, T. (1973). The analgesie effect of electrieal stimulation of 'the diencephalon and mesencephalon. ~ Research, 60, 369-381.
Basbaum, A.!., & Fields, H. L. (1978). Endogenoua pain contrai mechanisms: review and hypothesia. Annala of Neurolol!, !, 451-462.
Baab.um, A. 1., & Fieldl, H. L. (1984). Endolenoua pain control .y.teml: brainlte .. apin.l pathvay. and endorphin circuitary. Annual lleviev of Neuro.cience. 2., 309-338.
Becltatead, R. H. (1979). An autoradioaraphic: eu.in.tion of cortico-cortical and ,ub-cortical projection. of the
$

o
o , r.
,. . _dio-dor.at-projection (prefrontal) cortex in rat. Journal of C~parative NeurololY, ~, 43-62.
Beecher, B. It. (1968). The meuurement of pain in Man: A rein.pection of the work of the Harvard group." ln SOUlairai, A., Cahn, J. and Charpentier, J., (Eds), Pain, Academie Press, London, 201-213..
83
Belensky, G. L. & Boladay, J. W. (1981). Repeated electroconwhive shock' (BCS) and IIIOrhpine tolerance: demonstration of crou-.ensitivity in the rat. Life Sciences, 29, 553-563.
Belluzi, J. D., Grant, N., Ganky, V., Sarantakis, D., Wise, C. D. &" Stein, L. (1976). Analgesia induced in vivo by central adminÏltration of enkephalin in the rat. Nature, 260, p.625oo
Bergmann, F., Costin, A. & Gutman, J. (963). A low convulsive thre.hold aru in the rabbits mellencephalon. Electroencephalography and Clinical Neurophysiology, 11, 683-690.
Bobillier, P., Seguin, S., Petitjean, F., Salvert, D., Touret, M. & Jouvet, M. (1976). The raphe nuclei of the cat brain stem: a topographical at lall of their efferent project ions as revealed by autoradiography. Brain Research, 113. 449-486.
Bowman, W. G. & Rand, M. J. (1980). Textbook of pharmacology, ~ Edi tion, Blackwe Il Scient ific Publications, Oxford, London, Edinburgh, Melbourne. - -
< Buxbaum, D. M., Yarbrough, G. G. & Carter, M. E. (1973). Biogenic amine. and narcotic effects. 1 Modication of morphine induced analgesia and motor activity after alteration of cerebral amine levela. Jour.nal of Pharmacological and Ex'perimental Therapeutics, li, 317-327.
eupenter, M. B. (1972). Core text of .uroanatomy, The Williams and wH ki n. Campa ny. Bal t imore • r-
Collin •• R. C. Teuse, R. G. & Lotbman, E. W. (1983). Functional anatolll)' of limbic sehures: Focal discharges from medial entorhinal cortex in rat. Brain Research, 280, 25-40.
Conrad, L. C. A., Leonard, C. K. & Pfaff, D. W. (1974). Connections of the median and doreal raphe nue lei in the rat: an autoradiolraphic and degeneration st,udy. Journal of Comparat ive leurololJ, 156, 179-206.
Cooper, 1. S •• Upton. A. R. M. & Amin, 1. (1980). lleversibility of chronic neur010aic defect.: • 101De" effectl of electrical stimulation of th. thal •• uI a~d internai caplule in man. Applied NeurophYlÎolol1. !!1. 244-258.
. '

o /
o
---- ._-~-------------""'III!I"II!!!II$!
*
84
CostaU, B. & Nay10r, R. .J. (1974). On catalep.y and catatonia and the predictability of the catalepsy test for neuroleptic activity. Paychopharmacologia (JSerlin), 34, 233-241.
Cosynl, P. & Gybels, J. (1979). stimulation for pain in man. ln J • Liebesk.ind (Eda.), Advanèee in pain 511-514). New Yo-rk: Raven Press.
E1ectrica1 central gray .J. Bonica, D. Albe-Fea .. rd & .J. c. rFsurcb and therapy, (Yol. 3, pp. ,
D'Armour, F. E. and Smith, D. L. (1941). A method of determining the loss of pain sensation. .Journal of Pharmacology Experimental Therapeutics, 72, 74,-79.
Dennis, S. G. and Helzack, R. (1919). Comparison of phasic and tonic pain in animaIs. In J. J. Bonica, D. Albe-Fessard & J. C. Liebeskind (Eds.), Advances in pain research and therapy, (Yol. 3. pp. 747-760). New York: Raven Press.
Dennis, S. G., Choiniere, M. and Melzack, R. (1980). Stimulation-produced analge.sia in rats: assessment by two pain tests. and correlation with self-stimulation. Experimental Neurology, 68, 295-309 •
De Ryck, M., Schallert, T. & Teitelbaum, P. (1980). Morphine venus haloperidol cataleps1 in the rat: A behavioral an4,lysi 1 of postural support mechanisms. Brain Research, lQ.!., 143-172.
De Ryck, M. & Teitelbaum, P. (1984). Morphine catalepey as an adaptive reflex Iltate in rats. 8ehavioral Neuroscience, 98, No. 2, 243-261.
Dickenson, A. H. (1983). _, The inhibitory effedts of thalamie etimulat ion on the spinal transtV.uion of nocicepti ve informatiOon in the rat. Pain, Q, 213-224.
Dubuislon, D. and Dennis, S. G. (1977).. The formalin test: A quantitative study of the ana·lgesic effects of morphine, meperidine and brain-stem stimulation in rats and cat.. Pain, 4, 161-174.
Evans, W. A. (1961). A nev technique for the investigation of some ana.l,esic drug. -.n a reflexive behavior in the rat. Psychopharmacologia,.!, ~18-325.
'ardin, Y. Oliveraa, J. L. & Bea.on, J. M. (1984). A reinve.tigation of the analaeaic effecta induced by .-tilllulation of tht periaqueductal gray matter in the rat. l The production of b.ha.vioral aide tlffecta toaether vith ,anal,e.ia. Brain ats.arch. 1Q!. 105-123.
'Ardin, Y., 'Olivera., J. L. & Be •• oD, J. K. (1984). A reinve.tiaation of the anal, •• ic .ff.cU induced by Itiaulation of th, periaqueductal aray .. tter in th, rat. 11 Different i.1 charact.ri.t les of tbe ana11uia illduc.d by Yentral and' dor.al .tillUhtion. araiD .... arch, 1Q!, 125-139. i
..

o
o
85
Frenk, H., Engel, J. (Jr.), Ackermann, R. F., Shavit, Y. & Lie~skind, J. C. (1979). Endogenous opiods may mediate post-ietal behavioral depression in amygdaloid-kindled rat~. Brain Research, 167, 435 ... 440. -
Frenk, H., & Stein, B. E. (984). Endogenous opioids mediate ECS-indueed catalepsy at supraspinal levels. Brain Rèsearch, 303~' 109-112. ~ -
Frenk, H. & Yitzhaky, J. (1981). Effects of amygdaloid kindling on the pain threshold of the rat. kxperimental Neurology,.Il, 487-496.
Gibbs, F. A. &Vibbs, E. L. (1936). The convulsion threshold of various parts of ~thè cats brain. Archives 'of Neurology and Psyehiatry, 35, 109-116.
--Goodman, S. J. & Holeombe, V. (1975). Selective and analgesia in monkey resulting from brain stimulation. In and D. Albe-Fessard CEds.) Advances in Pain Research and (Vol. 1, pp. 495-502), Raven Press, New York.
prolonged J. J. Boniea
Therapy,
Hardy, S. G. P. (1985). Analgesia elicited by prefrontal '-''-st imulat ion. Brain Researeh, 339, 281-284.
Hardy, S. G. P';'--b Haigler, upon the midbrain: a possible Research, 339, 285-293.
H. J. (985). Prefrontal influences route for pain modulation. Brain ---..-
Heath, R. G. & Hickle, W. A. Evaluation of seven years experienee vith depth electrode studies in human patients. In Ramey, E. R., O'Doherty, D. S. (Eds.). Electrical studies on the unanesthetized brain. Paul B. Hoebner, New York, p. 214.
Herz, A., Alhus, K., Hetys, J. Schubert, P. & Teschemacher, H. J. (1970). On the central sites for the antiinociceptive action of morphine and fentanyl. Neuropharmacology, 9, 539-551.
Holaday, J. W., Belensky, G. L., Loh, H. H. & Heyerhof-f, J. L. (1977). Evidence for endorphi-ne release during electroeonvulsive shoek. ,·Neuroscience Abstract s,l, 293. /)'
H08obuchi, Y. (1980). The eurrent status of analgesfe brain stimulation. Acta Neurochirurgiea (supplement) 30, 219-227.
Hoaobuchi, Y., Adams, J. E. & Linchitz, R. (1977). Pain relief by electrical stimulation of the central gray mat ter in humans and i t s reversaI by naloxone. SeÏènce, 197, 183-186.
H08obuchi, Y., Adams, J. E. & Rutkin, B. (1973). Chronic thalamic stimulation for the central of facial anaesthesia dolorosa. Archi ves of Neurology, 29, 158-161.
H080buchi, Y., Rossier, J., Bloom, F. E., & Gui llemin, R. (1979). Stimulation of human periaqueductal gray for pain relief increues
~
-( ,

c
i '" • c '-.1"
iuanunoreactive beta-endorphin in ventricular fluid. Science, 203, 279-181.
Hughes, J. (1975). Isolation of an endogenous compound from the brain wi th pharmaco logical propert ies simi lar to morphine. Brain Research, 2., 29?-308.
Hughes, Jo;. Smith, T. W., Kosterlitz, H. W., Fothergill, L. A., Morgan, B. A. & Horris, H. R. (1975). Identification of two related pentapeptides from the brai n with potent opiate agonist activity. Nature, 258, 577-579.
86
Janssen, P. A., Niemegeers, C. J. E. & Dony. J. G. H. (1963). The inhibitory effect of fentanyl and other morphine-like analgesics on the wa rm water induced ~ail withdrawal ref lex in rats. Arzneimittel-Forschung, Q, 502-507.
Roffer, K. B., Berney, S. & Hornykiewicz, O. (1978). The role of the corpus striatum in neuroleptic-and narcotic-induced catalepsy. European Journal of Pharmae~, 47, 81-86.
Ruhar, M., Pert, C. &. Snyder, S. (1973). Regional distrïbution of opiate receptor binding in human and monkey brain. Nature, 245, 447-450.
Lewis, J. W., Cannan, J. T., Chudler, E. H. & Liebeskind, J. C. (1981). Effects of naloxone and hypophysectomy on elect ro-convulsive shock induced analgesia. Brain Research, 208, 230-233.
Lewis, J. W., Cannon,' J. T., & Liebeskind, J. C. (1980). Opioid and non-opioid mechanisms of stress analgesia. Science, 208, 623-625.
Lewis, J. W., Sherman, J. E., & Liebeskind, J •. C. (1981). Opioid and non-opioid stress analgesia: as sessment of to lerance and cross-toleranee with morphine. Journal of Neuroscienee, l, 358-36j.
Liebeskind, J. C., Guilbaud, C., Besson, J. H. & Olive.ras, J. L. (1973). Analgesia from electrieal stimulat ion of the peri-aquedu1al gray matter of the eat: Behavioural observations and inhibitory effects on the spinal eord interneurons. Brain Research, 50,441-446 •
. ' Liebeskind. J. C. & Paul, L. A. (1977). Psychologieal and phyaio1ogical meehanisms of f,ain. Annual Review of Ps:yehology, '28, 41-60 .~
Lorens, S. A. & Yunger. L'. M. {1974). Morphine analgesia, two-way avoidance, and eonaummatory behav.1or following lelJ'tbliS" in the midbrai'n raphe nuclei of the rat. Pharmacology, Biochemistry and Behavior, !, 215-221.
Kalick, J. B. & Goldstein, J. H. (1977). Analgesie activity of enkepbalinl following intracer~1)ral administration in the rat. Life Science. t,!!!. 827-832. -
o
•

o
r
o
ik.
, 1
87
Mayer, D. J. and Hayes. R. (1975). St imulat ion-produced analgeaia: development of tolerance and' cross-tolèrance to morphi ne. In S. Snyder and S. Hathysee (Eds.), Opiate receptor mechanisms: Neuroacienee Researeh Program Bulletin. Q, 94-100. •
Mayer, eleetrical analysia.
D. J. and Liebeskind, J. C. (1974). Pain reduction by focal s-..illlulation of the brain: an anatomiesl and behavioral Brain Researeh. 68, 73-93.
Mayer, D. J., Wolfle, T. L., Akil, M., Cardner, B. & Liebeskind, J. c. (1971). Analgesia from eleetrieal stimulation in the brainatem of the rat. Science, 174, 1351-1354.
Maurs, G., Roge, R. &~zars. Y. (1960). p,esultats de la stimulation du faisceau sp' o-thalamique et leu'r incidence sur la physio))athologie de le dou ur, Revue Neurologique, 103, No. 2, 136-138. ' '
___ i
Maurs, G., Heriennet) L., Cioloca, C. (l974). Traitments de certains types de douleurs par des stimulateurs thalamiques implanta,bles. Neurochirurgie, 12. 1,17-124.
Mazara, G. J., Merienne, L. & Cioloea, G.'{l976). Contributions of thalamic stimulattotl to th"! physiopalhology of pain. ln J. J. Boniea & D. Albe-Fessard (I~ds. ).Advanees in pain research and therapy, (Vol. 1, pp. 483-485). Raven Press: New York.
Maurs, G. J., Merienne', L~ ~ Cioloea, C. (l979). Comparative study of electrieal stimulation of po'ater\.or thalamie nuc1ei, peri aqueci!letal gray, and other midline '"lDeaencephalic 8truc~ures in man. 1 n Bonica, J. J., Liebeskind. J. C,. & Albe-Fessard, D. G. (Eds.>.. Advances in pain researeh and therapy, (Vol. 3, pp. 541-546), Ravén Press: New York.
Melzack, R. & Melinkoff, D.F. (974). Analgesia produced by brain stimulation: evidence of a prolonged onset period. Experimenta 1 Neurology, 43, 369-374.
Helzack, R., Stotler, W. A. and Livingstone, W. K. of diserete brainstem lesions in cats on perception of stimulation. Journal of Neurophysiology. l!; 353-367. . .
(1958). Effeets noxious ·0
Munkvad. L, Pakkenberg. H. & Randrup. A. (-1968). Aminergic systems in basal gang lia' associated with stereotyped hyperact ive behavior and catalepsy. Brain, Behavior and Evolution, 1, 89-100.
Hyslobodsky, M. S. & Mintz, H. (981). Post idal behavior.l Arrest in the rat: "catalepsy" or "eatatonia". Life Sciences, 1!, 2287-2293.
• Na~~Q, Y. (1983). Experimental and clinical studies on pain reduction by electrical stimulation of the internai capsule, Part 1. Acta Medica, Okayama (-1apan),. ll. 1209 .. 1226.
4 '
.. '

<
o
----- -------
\. ~,
Neba, S., Nakao, Y., MatlumotO, Y., Ohmqto, T. & Ni.himoto, A. (1984). Electric stimplation of the posteri6r limb of the internaI caplu1e for the treatment of thalamic pain. Applied Neuro~hysiology,ll, 137-148.
,Nishimoto , A., Namba, S./Makao, Y., Hatsumoto, Y. & Ohmoto, T.' (1984). Inhibition of nociceptive neurones by internaI capsule ~t imulation. Applied Neurophysiology, 47, 117-127.
Oliveras, J. l.., Bessofi, J. M., Guilb!J,ld, G. & Liebeskind, J. C • . ' ,(1974). Beha'vioural and electrophysiological evidence of pain
inhibition from midbrain stimulation in the cat. Experimènta1 Brain Research, 20,. 32-44.
.,.,. Paxfihos, G. & Watson, C. (1982). The rat brain in stereotoxic
coordinates, A subsidiary of Karcourt, Brace, & Jovanovich: Publishers. Sydney, New York, London, San Diego, San F~ansisco, Sao Pau 10, Tokyo, Toronto. .
88
~ Pert, A., Simantov, R. & Snyder, S. M. (1977). A morphi ne-like factor in mammalian brain: analgesic activity in rats. Brain Research, 136, 523-534.
Pert, A.J 6 Walter, H. (1976). Compari3-son between naloxone ~ reversaI of morphine and electrical stimulation induced analgesia in the rat mesenceehalon. Life Sciences, .!2., 1023-1032.
Plotkin, R. (1982). Results in 60 cases of 4eep brain st imulation for chronic intractable pain. Applied Neurophysiology, 45, 173-178.
Prado, W. A. & Robert s, M. H. T. (1985). An assessmènt of the antinociceptive and aversive effects of stimulating identified sites in rat brain. Brain Research, 340, 219-228.
~ -Racine, R. J. (1972). Modification of seizure 'activity by
electrical stimulat ion: Il Motor seizure. Elect roencephalography and Clinical NeurophysioUky, ]1, 281-294.
Racine, R., Livingstone, K. & Joaquin, A. (1975). Effec~s of procaine hydroChloride, diazepam, and diphenylhydantoin on seizure development in cortical and subcortical structures in rats. Electroencephalography and Clinical· Neurophysiolo8Y, 38, 355-365.
~
, Reynolds, D. v. (1969). Surgery in the rat dudng electrical analgeaia induced by focal brain. _stimulat ion. Science, 164, 444-445.
Reynolds, D. v. (1970). Reduced response to aversive stimuli during focal brain stimulation: E1ectrical analge'BÏa and electrical anae.the,aia. I~ D. V. Reynolds. & A; E. Sjoberg (Eds.). Neuroe1ectric Re.earch, Thomas, Springfield, 11Hnois, 151-167.
Rhode., D. L. & Liebeak.ind, .J. C. (1918). AnalgesLa from rostral brain .tem .ti.ulatiOll in thé rat. Brain Research, ill, 521-523.
- ~- --

•
o
o
• 89 , Richardson, D. E: (1970). Auto inhibition in the sensor'y system of
cats. Sur&ery Forum, ~, 447-449. ,
_ Richardson, D. E. (1982). Analgesia produced by stimulat ion of various sites in the human ,beta-endorphin '1ys~ Applied Neurophysiology, 45., 116-122. '0 '\
• Richardson, D. Eoi (1985). IJèi,ep brain st imulat ion for pain re 1 ief. --Neurosurgery, Vol. 3, Hc Graw-Hill, 2421-2426.
Richardson, D. E. & Akil," h. (1977). Pain reduction by electrical ... stimulation in man. Part 1. Acute administration in periaqueductal and periventricular sites. Journal of Neurosurgery, 47, 178-183.
Richardson, D. E. & Akil, H. (1977). Pain reduction'by electrical stimulation in man: Part 2. Chronic self administration in periventricular gray matter. Journal of Ne\Jrosurgèry, .!!l, 184-1'1/ ••
,.
R~ndeau, D. B., Turcotte, H., 'Îoung, L. & Herbert, ô. <J982). 'Similari t ies of the ca laleptic state induced by beta endorphin and morphine. Pharmacology. Biochemistry & Bellavior, .!2. ' 703-707.
Ruda, M., Hayes, R. L., ~rice, D. D., Hu, J. W. & Dubner, R. (1976). Inhibition of noc'iceptive reflexes in the primate tiy electrical stimulation or narcotic microinjection at medial mesencephalaic and diencephalic sites: Bebavioural and electro-physiological analyses. Proceedings of the Society for Neuro'science, 2, 952.
-+
Samenin, R. & Valzelli, L. 09,71). analgesia by stimulation of the nucleus Journal of Pharmacology, .!2., 298-302.
Increase of morphi ne-induced raphe" dors~ 1 i s. European
SeigEried, J. (1982). Monopolar electrical stimulation of nucleus ntroposteriormedialis thalami for post herpetic facial pain. ApplÏed uro h siolo , 45, 143-155. •
Sessle, B. J., Dubner, R., Greenwood, L. G. & Lucier, G. E. (1975). Descending influences of per\.aqueductal gray matter and somatosensory cerebral cortex on neurons in trigeminal brain stem nuclei. Canadian Journal of Pharmacology and Physiology, 54, .6(,-'e9.
t • ____
Soper, W. Y. (1976). Effects of analgesic 1 mid-brain st imulat ion on reflex withdrawal and' thermal escape in the rat. Journa 1 of Comparative Physiological Psychology, 90, 91-101 •
• Teschemach~r, H., Blasig, J. & Kromer, W. (976). p,tcine: - pituitary peptides with opiate-like .activity: Partial puri ficalion and effects in the rat after intraventricular injection. Naunyn Schmiedebel'gs: Archives of' Pharmacology, 294, 293-295.
.Thorn-Gray, B. & Levitt, R.- A~·(1983). _Rat brain sites responaive to etorphine: Analgesia and catatonia. Behavioral NeurOlC iencel. !!.. ~'8-778. .
/ /
• -t
r -

,
c ~ ~;;",',
"
/
Thorn-:Gray, B., Levitt, R. A., Hill, J. & Ward, K. (1981). A neuroanatolJlical study of analgesia and catatonïa indueed by etorphine in the rat. Neuropharmaeology, 20, 763-767.
Tsubokowa, T., Yamamoto, T., Katayama, Y. & Horiy8'8u, N. (1982). Clinieal resuIts and phYslologieal buis of thalamic relay nueleùs. stimu lation for relief of int raeta.b le, pain with morphine toleraià:~. Applied' Neurophysiology, 45, 143-155.
Turnbu 11, 1. H. syndrome. Canadian
Ttnbull, 1. H.
/li
(1982). Capsulàr stimulation for' the thalamie Jsiurnal of 'Neurologieal Sciences, ,i, 271.
"Helzack, R. (Eds.), Textb k·of ain, Churc~ill Edinbrugh, London, Melbourne and New York •
ln Wall, P. D. & Livingstone:
..
90
, Turnbull, 1. M., Schulman, R.
l !ltimulat ion for neuropathie pain. 486-493.
& Woodhurst, W. B. (980). Thalamie Journal of Neuroàurgery, 52,
_ Urca, G., Yitzhaky, J. ~ Frenk, H. (1981). Different opioid 'systems may participate in Post electroeonvul~ive shoek (ECS)
\ analgestà and eatalepsy. Brain Researeh, 219, 385-396.
, Willis, W. D., Ge~hart, K1 D., Wi'ueoekson, W~ S., Yezierski, R. P.,.rWilcox, T. K. & Cargill, '0.. L. (1984). Primate raphe- and reÜculospinal neurons: Effects"[,f stimulation in periaqueduetal gray or VPLc thalamic nucleus. Journal of Neurophy'siology, Vol. 51, No. 3, 467-480.
Woolfe, G., & Mac Donald, A. D. (1944). analgetic action of pethidine hydroehloride Pharmaeology and Experimental Therapeutics,
The evalua"ion of the (Demerol). ~ournal of 80, 300-307. - .
Yeung, J. C., Yaksh, r.. L. & Rudy, T. A. (1977). Concurrent mapping of brain sites' for sensitivity of the directypplication of morphine and focal electrieal stimulation in the prot"uetion of 1
antinoeieeption in the rat. Pain, 4,23-40. . .
York, 'J. L. & Maynert, E. W. (1978). Alterations in morphine analgeaia produeed by chronic deficits of braln catecholamines or serotonin: Role of analgesimetric procedure. Psychopharmacology, 56, 119-125.
Yitzhaky, J., Frenk, M. & Urea, G. (1982). KindHng-induced changea in morphine analgesia and eatalepsy: Evidence for independen~
. opio id .ystems. Brain Research, m, 193-201. ..
- ---
o '
.....





























