Embed Size (px)
Citation preview

PART 11. PARTIAL REACTIONS A N D REACTION MECHANISM
SYNTHESIS OF ADENOSINE TRIPHOSPHATE BY NA', K+-ATPAsE *
Robert L. Post, Kazuya Taniguchi,? and Gotaro TodaS Department of Physiology
Vanderbilt University Medical School Nashville, Tennessee 37232
Introduction
The kinetics of phosphorylation and dephosphorylation of Na', K+-ATPase provide a pathway t o investigate the mechanism of this ion transport enzyme. Heretofore this pathway has been explored in the direction of the forward reac- tion. The forward reaction involves transfer of the terminal phosphate group of ATP t o the enzyme in the presence of Mg2+ and Na+ and subsequent stimulation of hydrolysis of the phosphoenzyme by K+. A working hypothesis'>2 may be summarized for present purposes as a series of the following steps:
N a . E l + A T P + M g 2 + - N a * M g * E l % P + A D P (1)
Na * Mg E l % P t--) Mg Ei-P + Na+ (2)
Mg Ei-P + K++ K * Mg * E2-P ( 3 )
K . M g * E 2 - P o K * E2 + M g 2 + + P i (4)
E2 + Na+ * Na * E l + K+ (5 ) K
In these equations E represents the enzyme. E l indicates forms that can bind and accept phosphate from ATP. Correspondingly El 2, P indicates forms that can transfer their phosphate group t o ADP. E2 designates forms that can accept phosphate from inorganic phosphate, Pi, t o make a phosphoenzyme, E2-P.3 The superscript s indicates a form of E2-P that is sensitive t o K+ as shown by an in- crease of the rate of splitting of the enzyme-phosphate bond in the presence of K+. (Previously E2-P was defined entirely with respect t o its sensitivity t o K+ and insensitivity t o ADP4). Our current experiments indicate a t least four varieties of E2-P according t o this new definition. In addition t o the classical Ei-P (which can be formed from ATP but once it is formed then exchanges its phosphate group with Pi) and the familiar complex of the enzyme with the inhibitor ouabain, ouabain * E2-P, there is also a complex of the enzyme with K+, namely K * E2-P (Equations 3 and 4 above), and an insensitive phospho- enzyme, E$-P, which responds very little either t o K+ or t o ADP.
*This work was supported by grants from the National Institutes of Health, the National Heart and Lung Institute (No. 5R01 HL-01974), and the National Institute of Arthritis and Metabolic Diseases (No. 5PO1 AM-07462).
?On leave from the Department of Pharmacology, School of Dentistry, Hokkaido Uni- versity, Sapporo, Japan.
$Present address: First Department of Internal Medicine, Tokyo University Medical School, 7-3-1 Hongo Bunkyoku, Tokyo, Japan.
80

Post et al. : Synthesis of ATP by Na', K'-ATPase 81
It is the insensitive Ei-P that is the starting point of the series of experiments presented here on reversal of the ATPase reaction, that is, synthesis of ATP from ADP and Pi. These reactions may be outlined as follows:
Mg * Ei-P + Na+ t) (Mg * E;-P) ts Na Mg * E l 'L P
(- 1) Na - Mg * E l 'L P + ADP c) Na * E l + ATP + Mg2+ (7) In these experiments the synthesis of ATP was rendered irreversible by including one further reaction:
Mg2+ + CDTA + Mg - CDTA (8)
CDTA is cyclohexylenedinitrilotetraacetic acid, a powerful chelator of divalent metal ions.
The important features of this sequence of reactions are (1) that the enzyme accepts phosphate from inorganic phosphate to form a covalent enzyme-phos- phate bond in the absence of metallic monovalent cations, and (2) that this phosphoenzyme can be activated sufficiently by addition of a high concentra- tion of Na+ to permit synthesis of ATP. This is strictly a two-stage reaction. Na' inhibits phosphorylation by Pi and we have not found any significant steady- state tracer exchange between Pi and ATP. The amount of ATP formed has always been less than the amount of enzyme present. K E2-P is unsuitable as a starting material and K+ inhibits synthesis of ATP, as will be seen later, These reactions tend t o support a hypothesis that the functional energy level of the enzyme-phosphate bond in this enzyme is controlled at least partly by the selective binding of monovalent cations t o the e n ~ y m e . ~
Methods
The enzyme used in these experiments was obtained by differential centri- fugation of a homogenate of guinea pig kidney and selective collection of a fluffy layer of membranes on top of the mitochondria1 fraction.' These crude membranes usually were treated with concentrated sodium iodide by the method of Nakao and coworkers6 t o remove or inactivate interfering enzymes. To re- move residual Na' from the preparation, the membranes were subsequently washed in 1 mM MgC12 buffered t o pH 7 with imidazole and histidine hydro- chloride. The enzyme was phosphorylated with 32Pi and the phosphorylation reactions were stopped with trichloroacetic acid. In order t o estimate the amount of phosphoenzyme, the denatured phosphorylated membranes were centrifuged down, washed, and digested with pepsin. The peptic digests were subjected t o paper electrophoresis t o separate the radioactive [ 32 PI phospho- peptides overlapping the active site from 32Pi.7 The spots on the paper contain- ing the peptides were cut out and counted. This step was essential since Pi binds erratically t o denatured membranes8 In order t o estimate the amount of radio- active ATP synthesized, the supernatants from the acid denaturation were chromatographed on a Dowex column (FIGURE 1).
Results
When the enzyme was incubated with 32 Pi and Mg 2 + , a [ 32 PI phosphoenzyme appeared. A chase of excess unlabeled Pi showed that the phosphate group on the
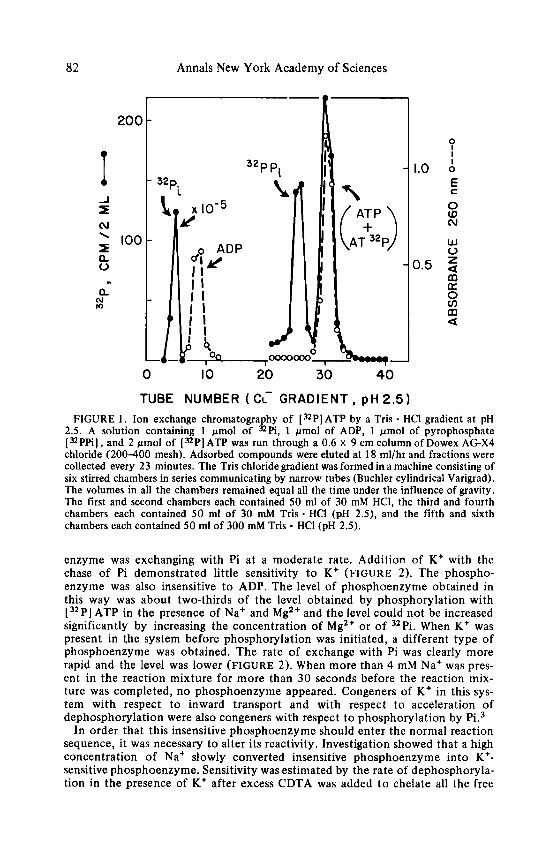
82 Annals New York Academy of Sciences
200
I A I (u
I a 0
a
' 100
c
(u rr)
--r: 1 (
AT 32P
1.0
3.5
P
E
0 (D cu w u z U €K
I 0
C
m
m 5: Q
0 10 20 30 40
TUBE NUMBER (CC GRADIENT, p H 2.5) FIGURE 1. Ion exchange chromatography of ["PI ATP by a Tris * HCI gradient at pH
2.5. A solution containing 1 pmol of "Pi, 1 pmol of ADP, 1 pmol of pyrophosphate [ "PPi] , and 2 Wmol of [=PI ATP was run through a 0.6 X 9 cm column of Dowex AG-X4 chloride (200-400 mesh). Adsorbed compounds were eluted at 18 ml/hr and fractions were collected every 23 minutes. The Tris chloride gradient was formed in a machine consisting of six stirred chambers in series communicating by narrow tubes (Buchler cylindrical Varigrad). The volumes in all the chambers remained equal all the time under the influence of gravity. The first and second chambers each contained 50 ml of 30 mM HCl, the third and fourth chambers each contained 50 ml of 30 mM Tris - HCI (pH 2.5), and the fifth and sixth chambers each contained 50 ml of 300 mM Tris - HCI (pH 2.5).
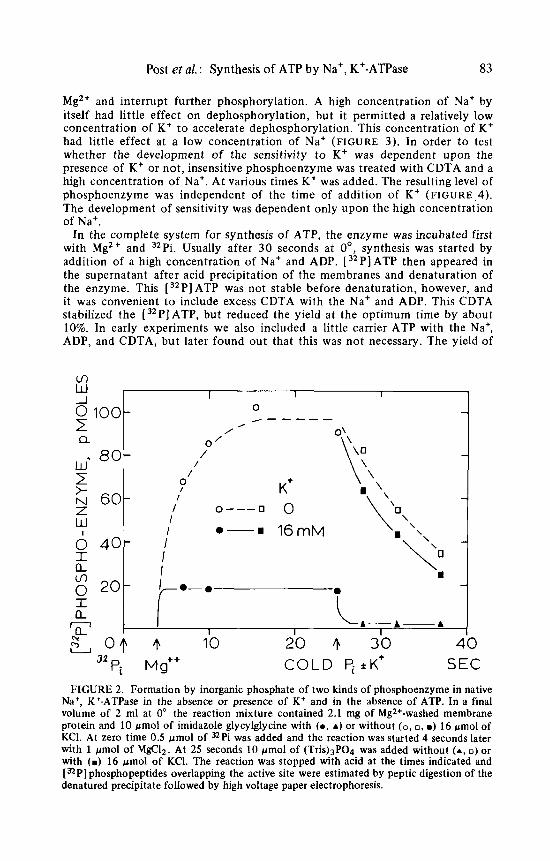
enzyme was exchanging with Pi a t a moderate rate. Addition of K + with the chase of Pi demonstrated little sensitivity t o K+ (FIGURE 2). The phospho- enzyme was also insensitive t o ADP. The level of phosphoenzyme obtained in this way was about two-thirds of the level obtained by phosphorylation with [ 32P] ATP in the presence of Na+ and Mg2+ and the level could not be increased significantly by increasing the concentration of Mg2+ or of 32Pi. When K + was present in the system before phosphorylation was initiated, a different type of phosphoenzyme was obtained. The rate of exchange with Pi was clearly more rapid and the level was lower (FIGURE 2). When more than 4 mM Na+ was pres- ent in the reaction mixture for more than 30 seconds before the reaction mix- ture was completed, n o phosphoenzyme appeared. Congeners of K* in this sys- tem with respect t o inward transport and with respect t o acceleration of dephosphorylation were also congeners with respect t o phosphorylation by Pi.3
In order that this insensitive phosphoenzyme should enter the normal reaction sequence, it was necessary t o alter its reactivity. Investigation showed that a high concentration of Na+ slowly converted insensitive phosphoenzyme into K+- sensitive phosphoenzyme. Sensitivity was estimated by the rate of dephosphoryla- tion in the presence of K+ after excess CDTA was added t o chelate all the free
Post et al. : Synthesis of ATP by Na', K+-ATPase 83
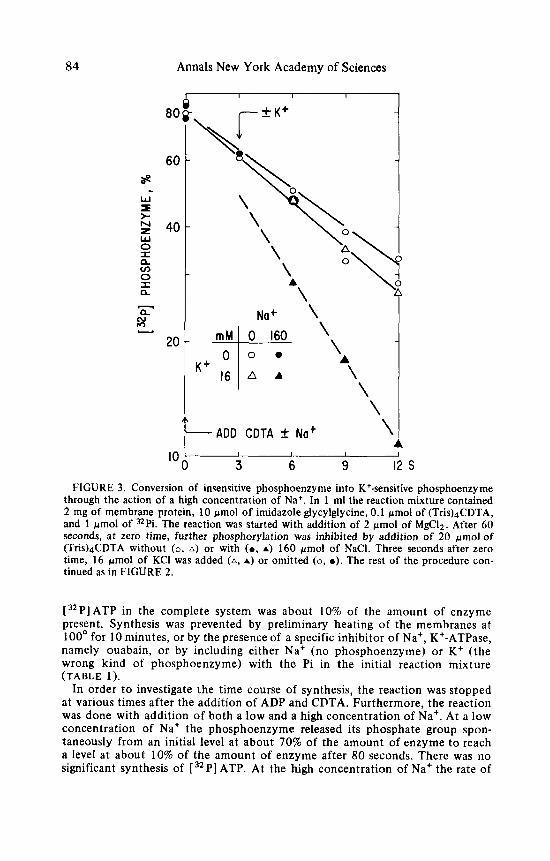
Mg2+ and interrupt further phosphorylation. A high concentration of Na+ by itself had little effect on dephosphorylation, but it permitted a relatively low concentration of K + t o accelerate dephosphorylation. This concentration of K + had little effect at a low concentration of Na+ (FIGURE 3) . In order t o test whether the development of the sensitivity t o K + was dependent upon the presence of K + or not , insensitive phosphoenzyme was treated with CDTA and a high concentration of Na+. At various times K + was added. The resulting level of phosphoenzyme was independent of the time of addition of K + ( F I G U R E . 4 ) . The development of sensitivity was dependent only upon the high concentration of Na+.
In the complete system for synthesis of ATP, the enzyme was incubated first with Mg2+ and 32Pi. Usually after 30 seconds at Oo, synthesis was started by addition of a high concentration of Na+ and ADP. [ 32P] ATP then appeared in the supernatant after acid precipitation of the membranes and denaturation of the enzyme. This [ 32P] ATP was not stable before denaturation, however, and it was convenient t o include excess CDTA with the Na+ and ADP. This CDTA stabilized the [ 32P] ATP, but reduced the yield at the optimum time by about 10%. In early experiments we also included a little carrier ATP with the Na+, ADP, and CDTA, but later found out that this was not necessary. The yield of
80
60
w- 2
Z W
I
0 40 r a g 2 0 I a 'n'
I I I 0 /----- .
/ 0' 0' /
/ /
7 K+ I
I o----n 0 I \
I 0 - 8 16mM I I I 8
A I
2 0 3 3 0 - 32 Pi Mg" COLD PitK+ SEC
FIGURE 2. Formation by inorganic phosphate of two kinds of phosphoenzyme in native Na+, K*-ATPase in the absence or presence of K + and in the absence of ATP. In a final volume of 2 ml at 0" the reaction mixture contained 2.1 mg of Mgz+-washed membrane protein and 10 gmol of imidazole glycylglycine with ( 0 , A) or without (0 , O , B) 16 pmol of KCl. At zero time 0 .5 pmol of =Pi was added and the reaction was started 4 seconds later with 1 gmol of MgClz. At 25 seconds 10 gmol of (Tris)3P04 was added without (A, 0) or with (B) 16 pmol of KCI. The reaction was stopped with acid at the times indicated and [BPI phosphopeptides overlapping the active site were estimated by peptic digestion of the denatured precipitate followed by high voltage paper electrophoresis.
84 Annals New York Academy of Sciences
I I I
Na+ \
\ \
- ADD CDTA k Na' \
-~ through the action of a high concentration of Na+. i n 1 ml the reaction mixture contained 2 mg of membrane protein, 10 pmol of imidazole glycylglycine, 0.1 pmol of (Tris)4CDTA, and 1 rmol of =Pi. The reaction was started with addition of 2 pmol of MgClz . After 60 seconds, at zero time, further phosphorylation was inhibited by addition of 20 gmol of (Tris)4CDTA without (0 , A) or with (0 , A) 160 pmol of NaCl. Three seconds after zero time, 16 rmol of KCl was added (A, A) or omitted (0, 0 ) . The rest of the procedure con- tinued as in FIGURE 2.
["PIATP in the complete system was about 10% of the amount of enzyme present. Synthesis was prevented by preliminary heating of the membranes at 100' for 10 minutes, or by the presence of a specific inhibitor of Na', K+-ATPase, namely ouabain, or by including either Na+ (no phosphoenzyme) or K+ ( the wrong kind of phosphoenzyme) with the Pi in the initial reaction mixture (TABLE 1).
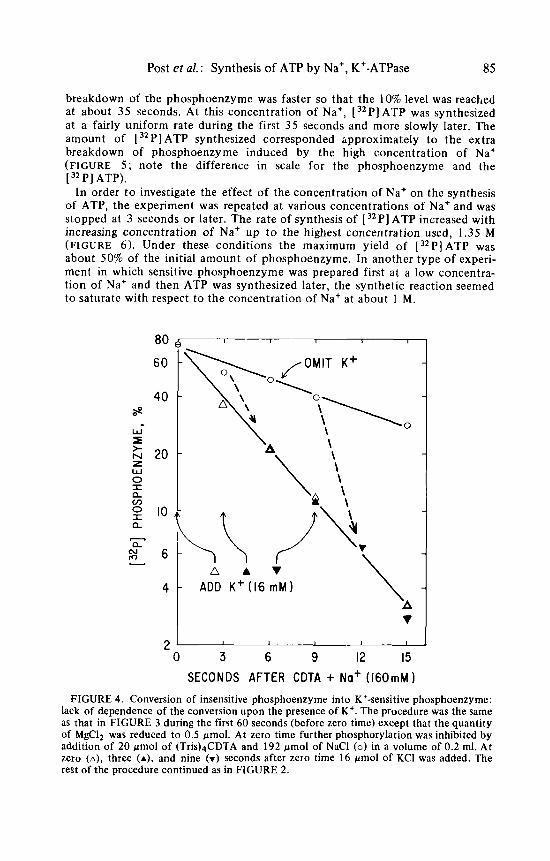
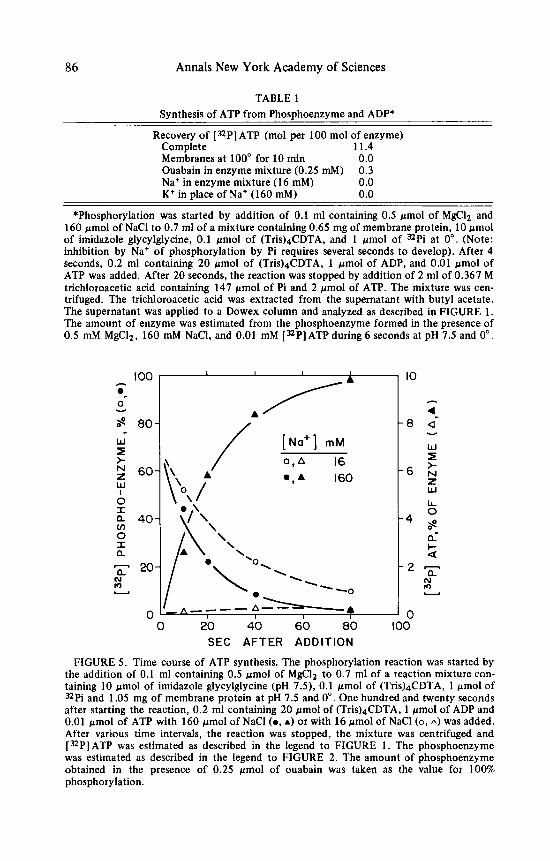
In order t o investigate the time course of synthesis, the reaction was stopped at various times after the addition of ADP and CDTA. Furthermore, the reaction was done with addition of both a low and a high concentration of Na'. At a low concentration of Na' the phosphoenzyme released its phosphate group spon- taneously from an initial level a t about 70% of the amount of enzyme t o reach a level a t about 10% of the amount of enzyme after 80 seconds. There was no significant synthesis of [ 32P] ATP. At the high concentration of Na+ the rate of
Post et al. : Synthesis of ATP by Na+, K+-ATPase 85
breakdown of the phosphoenzyme was faster so that the 10% level was reached at about 35 seconds. At this concentration of Na', [32P]ATP was synthesized at a fairly uniform rate during the first 35 seconds and more slowly later. The amount of [ 32P] ATP synthesized corresponded hpproximately t o the extra breakdown of phosphoenzyme induced by the high concentration of Na+ (FIGURE 5 ; note the difference in scale for the phosphoenzyme and the [ 32 PI ATP).
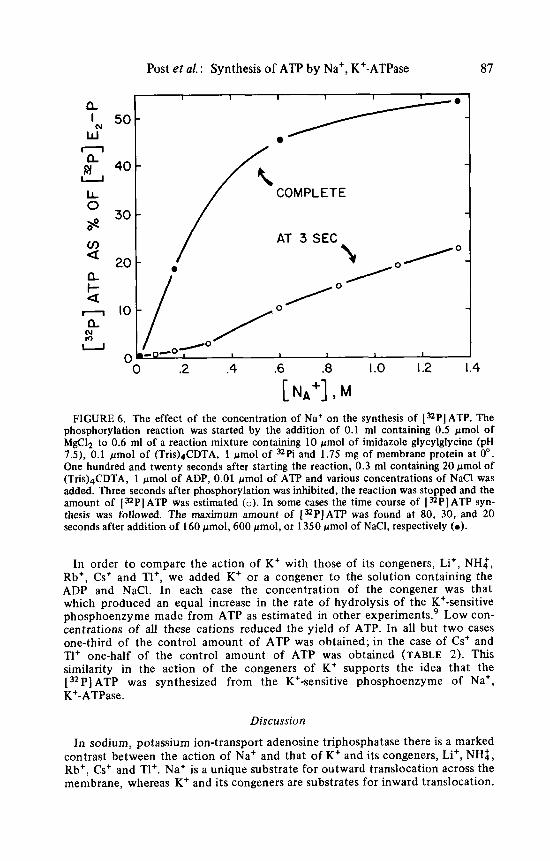
In order t o investigate the effect of the concentration of Na+ on the synthesis of ATP, the experiment was repeated at various concentrations of Na+ and was stopped at 3 seconds or later. The rate of synthesis of [ 32P] ATP increased with increasing concentration of Na+ up t o the highest concentration used, 1.35 M (FIGURE 6). Under these conditions the maximum yield of [32P]ATP was about 50% of the initial amount of phosphoenzyme. In another type of experi- ment in which sensitive phosphoenzyme was prepared first at a low concentra- tion of Na' and then ATP was synthesized later, the synthetic reaction seemed to saturate with respect to the concentration of Na+ at about 1 M.
80 60
40 s w I L 20 z w 0 I a m g 10 a
a m 6
4
- c\I
Y \l A A
- ADD K f (
2 ' I I 1 I I
0 3 6 9 12 15 SECONDS AFTER CDTA + Na' (160mM I
FIGURE 4 . Conversion of insensitive phosphoenzyme into K'-sensitive phosphoenzyme: lack of dependence of the conversion upon the presence of K+. The procedure was the same as that in FIGURE 3 during the first 60 seconds (before zero time) except that the quantity of MgC12 was reduced to 0.5 pmol. At zero time further phosphorylation was inhibited by addition of 20 pmol of (Tris)4CDTA and 192 pmol of NaCl ( 0 ) in a volume of 0.2 ml. At zero (a), three (A), and nine (v) seconds after zero time 16 pmol of KCI was added. The rest of the procedure continued as in FIGURE 2.
86 Annals New York Academy of Sciences
TABLE 1 Synthesis of ATP from Phosphoenzyme and ADP*
Recovery of ["PI ATP (mol per 100 mol of enzyme) Complete 11.4 Membranes at 100" for 10 min 0.0 Ouabain in enzyme mixture (0.25 mM) 0.3 Na+ in enzyme mixture (1 6 mM) 0.0 K+ in place of Na+ (160 mM) 0.0
*Phosphorylation was started by addition of 0.1 ml containing 0.5 pmol of MgClz and 160 pmol of NaCl to 0.7 ml of a mixture containing 0.65 mg of membrane protein, 10 pmol of imidazole glycylglycine, 0.1 pmol of (TriQ4CDTA, and 1 pmol of "Pi at 0". (Note: inhibition by Na+ of phosphorylation by Pi requires several seconds to develop). After 4 seconds, 0.2 ml containing 20 pmol of ( T ~ ~ s ) ~ C D T A , 1 pmol of ADP, and 0.01 pmol of ATP was added. After 20 seconds, the reaction was stopped by addition of 2 ml of 0.367 M trichloroacetic acid containing 147 pmol of Pi and 2 pmol of ATP. The mixture was cen- trifuged. The trichloroacetic acid was extracted from the supernatant with butyl acetate. The supernatant was applied to a Dowex column and analyzed as described in FIGURE 1. The amount of enzyme was estimated from the phosphoenzyme formed in the presence of 0.5 mM MgClz, 160 mM NaCl, and 0.01 mM [=P]ATP during 6 seconds at pH 7.5 and 0".
I00 - 0-
ae 80
0 Y
W 5 > 2 60 w
I 0 I
0 I
40
a ii- 20 (u m u
0
/ [No ' ] mM
n W
0 20 40 60 80 I00 SEC AFTER ADDIT ION
FIGURE 5 . Time course of ATP synthesis. The phosphorylation reaction was started by the addition of 0.1 ml containing 0.5 pmol of MgClz to 0.7 ml of a reaction mixture con- taining 10 pmol of imidazole glycylglycine (pH 7.5), 0.1 pmol of (Tris)&DTA, 1 pmol of "Pi and 1.05 mg of membrane protein at pH 7.5 and 0". One hundred and twenty seconds after starting the reaction, 0.2 ml containing 20 pmol of ( T ~ ~ s ) ~ C D T A , 1 pmol of ADP and 0.01 pmol of ATP with 160 pmol of NaCl (m, A) or with 16 pmol of NaCl ( 0 , a) was added. After various time intervals, the reaction was stopped, the mixture was centrifuged and ["PI ATP was estimated as described in the legend to FIGURE 1. The phosphoenzyme was estimated as described in the legend to FIGURE 2. The amount of phosphoenzyme obtained in the presence of 0.25 pmol of ouabain was taken as the value for 100% phosphor ylation.
a
Wi I 50
n $ 40 u LL 0
30 8 cn a 20 a a l-
a U
n 10 i rr)
0
Post et al. : Synthesis of ATP by Na', K+-ATPase
I I
COMPLETE
AT 3 SEC
.2 .4 .6 .8 1.0 1.2 1.4
87
FIGURE 6 . The effect of the concentration of Na+ on the synthesis of [%PI ATP. The phosphorylation reaction was started by the addition of 0.1 ml containing 0.5 pmol of MgCl? to 0.6 ml of a reaction mixture containing 10 pmol of imidazole glycylglycine (pH 7.9, 0.1 pmol of (Tris)&DTA, 1 pmol of "Pi and 1.75 mg of membrane protein at 0". One hundred and twenty seconds after starting the reaction, 0.3 ml containing 20 pmol of (Tris)4CDTA, 1 pmol of ADP, 0.01 pmol of ATP and various concentrations of NaCl was added. Three seconds after phosphorylation was inhibited, the reaction was stopped and the amount of [ XP] ATP was estimated (0). In some cases the time course of [=PI ATP syn- thesis was followed. The maximum amount of ["PJATP was found at 80, 30, and 20 seconds after addition of 160 pmol, 600 pmol, or 1350 pmol of NaCI, respectively (0 ) .
In order t o compare the action of K+ with those of its congeners, Li+, NH;, Rb', Cs+ and Tl', we added K+ or a congener t o the solution containing the ADP and NaCl. In each case the concentration of the congener was that which produced an equal increase in the rate of hydrolysis of the K+-sensitive phosphoenzyme made from ATP as estimated in other experiments.' Low con- centrations of all these cations reduced the yield of ATP. In all but two cases one-third of the control amount of ATP was obtained; in the case of Cs+ and T1+ one-half of the control amount of ATP was obtained (TABLE 2 ) . This similarity in the action of the congeners of K+ supports the idea that the [ 32P] ATP was synthesized from the K+-sensitive phosphoenzyme of Na', K+-ATPase.
Discussion
In sodium, potassium ion-transport adenosine triphosphatase there is a marked contrast between the action of Na+ and that of K+ and its congeners, Li', NH;, Rb', Cs+ and Tl". Na+ is a unique substrate for outward translocation across the membrane, whereas K+ and its congeners are substrates for inward translocation.

88 Annals New York Academy of Sciences
TABLE 2 Inhibition of ATP Synthesis by K + and Its Congeners*
mM ATF', % EP Complete system
Additions Na + 160. 23
Li+ 12. 8
K + 0.16 6 Rb+ 0.1 7 CS' 0.6 11 TI+ 0.05 12
NH4 1. 8
*The phosphorylation reaction was started by the addition of 0.1 ml containing 0.5 pmol of MgC12 to 0.7 ml of a reaction mixture containing 10 pmol of imidazole glycylglycine (pH 7.5), 0.1 pmol of (Tris)4CDTA, 1 pmol of %Pi and 1.12 mg of membrane protein at 0". Thirty seconds after the reaction was started, 0.2 ml containing 20 pmol of (Tris)4CDTA, 160 pmol of NaCI, and K' or congener was zdded to produce the concentration indicated. The reaction was stopped 10 seconds later and the amount of [%P)ATP was estimated as described in TABLE 1. The amount of phosphoenzyme, EP, formed in the presence of 0.5 mM MgCl,, 160 mM NaCl, and 0.01 mM [BPI ATP during 6 seconds was taken as the 100% level.
With respect t o the dephosphoenzyme, Na' uniquely inhibits phosphorylation by Pi and accelerates phosphorylation by ATP. Simply in the binding of ATP t o the dephosphoenzyme the dichotomy is almost as marked; K+ and all of its congeners except Li' antagonize binding of ATP, and Na' uniquely opposes this antagonism.1° Li+ has n o influence on the binding of ATP. With respect t o the phosphoenzyme, K+ and its congeners accelerate dephosphorylation and Na' opposes this action, although it has a small stimulatory effect of its own at high concentrations. The present experiments have contributed t o this picture by showing that high concentrations of Na' accelerate transphosphorylation from the enzyme t o ADP and that K+ and its congeners oppose this action (TABLE 2). The different effects of K+ and Na' on the phosphoenzyme correlate well with the effects of phosphate compounds on the exchange of intracellular and extracellular ions across the intact plasma membrane of erythrocytes" and squid axons." ADP catalyzes an exchange of intracellular Na' with extracellular Na' and Pi catalyzes an exchange of intracellular K+ with extracellular K'. These ex- change reactions are consistent with the idea that phosphyrylation of E l and de- phosphorylation of El -P are linked t o translocation of Na across the membrane. In a similar way phosphorylation of E2 and dephosphorylation of E2-P can be linked t o translocation of K+ across the membrane. This hypothesis has been presented in more detail e l ~ e w h e r e . ~
In the exchange of intracellular and extracellular Na+ it is interesting t o find a requirement for a relatively high concentration of extracellular Na+.13 In relation to the present experiments this requirement suggests that the high concentration of Na+ that stimulated synthesis of ATP was combining with the transport sys- tem at a binding site in communication with the solution on the extracellular face of the membrane, or what was the extracellular face before the cells were broken. The membrane preparation used in these experiments probably contains more-or-less resealed vesicles made from the original plasma membranes, but it is not possibie for us to know the homogeneity, leakiness, o r sidedness of these

Post et al. : Synthesis of ATP by Na’, K+-ATPase 89
membranes. Consequently it is not possible t a specify the necessity for a gradient of Na’ concentration across these membranes t o permit synthesis of ATP. It seems reasonable to suppose that a high concentration of Na’ is required t o put Na’ on the enzyme from the solution in contact with the extracellular face of the membrane, but it is not possible to specify whether or not there is also a requirement that Na+ should also dissociate from the enzyme into the solution in contact with the intracellular face of the membrane in order t o permit synthesis of ATP.
The active site of phosphorylation has been characterized as probably the beta carboxyl group of an aspartic acid residue in the larger protein found in the purest available preparations. 14,15 According to radioactive fingerprints of phosphopeptides overlapping the active site of phosphorylation, the active site appears t o be the same in all the reactive forms of the phosphoenzymep$16 so far examined, namely, El -P in the poisoned enzyme, ouabain * Ez-P, sensitive Ez-P, insensiiive E2-P, and K * Ez-P. Since the acyl phosphate bond can be formed from Pi in the absence of Na’, one might be inclined t o refer t o it as a “low-energy’’ acyl phosphate and since the phosphate group can subsequently be transferred t o ADP in the presence of Na’ t o form a high-energy terminal pyrophosphate bond in ATP, one might be inclined to speak of binding of Na+ to the enzyme as raising the energy level of the “low-energy” bond. However, such a line of thought is an oversimplification in that it ignores the noncovalent bonds that the phosphate group surely makes with the active center of this enzyme. Specific investigation of these bonds appears t o be out of reach at present.
Summary
In the absence of monovalent inorganic cations sodium, potassium ion trans- port adenosine triphosphatase accepted a phosphate group from inorganic phos- phate in the presence of magnesium ion with formation of a phosphoenzyme. The phosphate group was probably bound covalently t o the beta carboxyl group or an aspartic acid residue in the enzyme protein. Treatment of this phospho- enzyme with a high concentration of sodium ion, adenosine diphosphate and a chelator of magnesium ion produced adenosine triphosphate. The maximum yield was 50% of the amount of phosplioenzyme. Addition of a low concentra- tion of potassium ion with the sodium ion inhibited the reaction. It is suggested that binding of sodium ion t o a low-affinity site on the enzyme is necessary for reversal of hydrolysis of adenosine triphosphate by this enzyme. This binding site may communicate with the extracellular solution when the enzyme is functioning as an active transport system in an intact cell.
A ckno wledgmen t s
The authors are indebted t o Jean Sorrells, Anita Kemp, Marianne Talbert, and Jackie Waynick for help with the experiments.
References
1. FAHN, S., G . J . KOVAL & R. W. ALBERS. 1966. J. Biol. Chem. 241: 1882-1889. 2. WHITTAM, R. & K . P. WHEELER. 1970. Annu. Rev. Physiol. 32: 21-60. 3. POST, R. L., S. KUME & F. N. ROGERS. 1973. In Mechanisms in Bioenergetics. G . F.
Azzone, L. Emster, S. Papa, E. QuagliarieUo & N. Siliprandi, Eds.: 203-218. Aca- demic Press. New York. N .Y.

90 Annals New York Academy of Sciences
4. POST, R. L., S. KUME, T. TOBIN, B. ORCUTT & A. K. SEN. 1969. J. Gen. Physiol.
5. POST, R. L. & A. K. SEN. 1967. In Methods of Enzymology. R. W. Estabrook & M. E.
6. NAKAO, T., K. NAGANO, K. ADACHI & M. NAKAO. 1963. Biochem. Biophys. Res.
7. BOND, G. H., H. BADER & R. L. POST. 1971. Biochim. Biophys. Acta 241: 57-67. 8. BOYER, P. D. & L. L. BIEBER. 1967. In Methods in Enzymology. R. W. Estabrook &
9. POST, R. L., C. HEGYVARY & S. KUME. 1972. J. Biol. Chem. 247: 6530-6540.
54: 306s-326s.
Pullman, Eds. Vol. 10: 762-768. Academic Press. New York, N.Y.
Commun. 13: 444-448.
M. E. Pullman, Eds. Vol. 10: 768-773. Academic Press. New York, N.Y.
10. HEGYVARY, C. & R. L. POST. 1971. J. Biol. Chem. 246: 5234-5240. 11. GLYNN, 1. M., J . F. HOFFMAN & V. L. LEW. 1971. Phil. Trans. Roy. SOC. Lond. B.
12. DE WEER, P. 1970. J. Gen. Physiol. 56: 583-620. 13. GARAY, R. P. & P. J. GARRAHAN. 1973. J. Physiol. 231: 297-325. 14. HOKIN, L. E. 1974. Ann. N.Y. Acad. Sci. This monograph. 15. POST, R. L. & S. KUME. 1973. J. Biol. Chem. 248: 6993-7000. 16. SIEGEL, G. J., G. J. KOVAL & R. W. ALBERS. 1969. J. Biol. Chem. 244: 3264-3269.
Discussion
DR. BRODSKY: Is it really determined that there is truly a net synthesis of ATP starting from Pi in those experiments, or is it an exchange of the isotope? In other words, is more of the cold ATP breaking down than hot ATP is being synthesized from Pi?
DR. POST: The high concentration of CDTA prevents an exchange reaction. Furthermore, some of these experiments were conducted in the absence of added ATP, u p t o the time that the acid was added t o stop the reaction. I t has been our practice t o put cold ATP in that acid. There is also a tiny contamination of the ADP by a little bit of ATP (less than 5 pM final concentration). A specific experiment at 1 0 pM ADP excluded participation of this contaminating ATP. The presence or absence of supplemental cold ATP has not affected the results.
DR. MINKOFF (SUNY, Downstate Medical Center, Brooklyn, N . Y . ) : Dr. Post, in February 1973 in the Biophysical Journal I published a comparison of the energy available from hydrolysis of ATP with that required by summation of the activity of many solute pumps in E. coli, and I found a tremendous dis- crepancy between the turnover of ATP necessary for the bacteria t o perform this transport and the available supply. Now in your model you illustrate almost a one t o one correlation between ATP hydrolysis and sodium-potassium exchange. And I was wondering if you would comment on the possibility that the amount of ATP split could be much less than the sodium-potassium exchange.
DR. POST: Your discrepancy showed incorrectness of at least one of the assumptions that you made in setting up your calculation, but did not identify which assumption was incorrect. The available experiments done with mamma- lian membranes are fairly clear and consistent. As far as I know there are only the papers of Bonting on presence of ( N a + + K+)-ATPase in E. coli, and the amount that he found was not very great and was not easily distinguished from the background ATPase. I would like t o find further confirmation of the pres- ence of (Na+ + K+)-ATPase in E. coli.
DR. GLYNN: There were two striking things about your experiments on the synthesis of ATP. One was the slowness of the reaction. It seemed t o take several seconds t o reach the maximum. The other was the very high sodium concentra- tions. Do you think that what is happening is that you have got simply a chem-
262: 91-102.







![Increased Rate of Adenosine Triphosphate …...(CANCER RESEARCH 55, 4352-4360, October 1, 1995] Increased Rate of Adenosine Triphosphate-dependent Etoposide (VP-16) Efflux in a Murine](https://img.pdfslide.net/doc/110x75/5e7e8d68c5d0407f2447f2a9/increased-rate-of-adenosine-triphosphate-cancer-research-55-4352-4360-october.jpg)





















