Embed Size (px)
Citation preview

THE PHOSPHORUS METABOLISM OF INVERTEBRATE NERVE
BY GEORGE L. ENGEL AND R. W. GERARD
(From the Marine Biological Laboratory, Woods Hole)
(Received for publication, August 26, 1935)
Previous work (Gerard and Wallen, 1929; Gerard and Tupikow, 1931; Tang, Tang, and Gerard, 1932) has indicated that the solu- ble phosphorus compounds of vertebrate and invertebrate nerve play a role in the important chemical sequence of activity. There are grounds (Gerard, 1932, 1934) for supposing that in nerve, as in muscle, phosphocreatine breaks down in the most intimate relation with the physiological response, energy for its resynthesis being derived next immediately from the succeeding breakdown of adenylpyrophosphate (adenosinetriphosphate). Recent experi- ments (Lohmann, 1934) with muscle brei suggest that in muscle this order may be reversed. In frog nerve, but not in crab, there is a burst of heat production as an early recovery phase after con- duction. The present experiments started from the consideration that if this heat phase represented the adenylpyrophosphate link of the chain, then the nucleotide compound would be absent or unchangeable in crustacean nerve, though active in frog nerve metabolism (see Hill, 1934; Gerard, 1934). The results demon- strate that adenylpyrophosphate plays an intimate r81e in the metabolism of these nerves, probably quite analogous to that in frog nerve. In addition, they supply further evidence for the sequence phosphagen-adenylpyrophosphate in breakdown and resynthesis.
Method
Analytical-Phosphates were determined by the Fiske and Sub- barow (1925) method, as employed by Lohmann and Jendrassik (1926). To obtain sufficient color for reliable readings (in four fractions each of a control and two experimental portions) 0.5 to
379
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

380 P Metabolism of Invertebrate Nerve
0.8 gm. of tissue (four to seven lobsters) was used and occasionally standard P (up to 0.02 mg.) added to a fraction. Cooled nerves were ground in 6 cc. of iced 10 per cent trichloroacetic acid and the suspension filtered in 10 to 15 minutes. The filtrate was neutralized with NaOH until pink to phenolphthalein, adjusted to 0.05 N HCl by addition of N HCl, and fractionated as follows. (The original 5 cc. of filtrate became 7 cc. after treatment. It is assumed that soluble P is equally distributed between filtrate and residue in calculating the results.)
Fraction I-A 2 cc. aliquot was analyzed immediately for reac- tive P. This represents essentially inorganic P.
Fraction II-A second aliquot of 2 cc. was hydrolyzed for 20 hours at room temperature before analysis. The increase in active P (Fraction II-I) is a measure of arginine P (Meyerhof, 1930).
Fraction III-Another aliquot of 1.5 cc. was brought to 1 N with HCl, hydrolyzed at 100” for 7 minutes, and the reactive P de- termined. The increase above Fraction II is taken as measuring pyrophosphate P (Lohmann, 1928) ; and Fraction III itself repre- sents the total of the labile fractions.
Fraction IV-A final 1 cc. aliquot was ashed with 1.5 cc. of 10 N H&JO4 plus a few drops of concentrated HNO, (Teorell, 1931). The total acid-soluble P was now measured ; the increase over Frac- tiori III represents the stable fraction.
This fractionation is crude. Fraction I, though predominantly inorganic phosphate, may include some phosphorus of dihy- droxyacetone phosphoric acid, which breaks down rapidly in alka- line solution. In addition, during the 15 minutes testing in 0.7 N acid and molybdate, a possible hydrolysis of some nucleotide polyphosphates and lactic acid intermediaries is not excluded. However, since no significant hydrolysis of the phosphoarginine occurs within this time, and since the color remains bon&ant for at least 10 minutes longer, any error would be negligible.
The difference between Fractions II and I probably represents arginine phosphate fairly accurately, and the present values check ver.y closely with those obtained by Tang, Tang, and Gerard (1932) by the Ca precipitation method.’
1 In an earlier series of results (1933), in which Fraction I wasdetermined on the original trichloroacetic acidextract before neutralization and in which
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

G. L. Engel and R. W. Gerard
The proportions and kinds of phosphate compounds which are hydrolyzed in Fraction III cannot be accurately stated, but in all likelihood adenylpyrophosphate (adenosinetriphosphate) is mainly involved. By the procedure no distinction can be made between adenylpyrophosphate and free pyrophosphate ion, and there would be no change if adenylpyrophosphate breaks down to adenylic acid and pyrophosphate rather than orthophos- phate, as reported for muscle in some conditions (Ferdmann and Feinschmidt, 1935). Pyrophosphate will be taken to mean adenylpyrophosphate in the further treatment. Other compounds that would be partially broken down in this fraction include hexosediphosphoric acid (30 per cent hydrolyzable), hydroxyace- tone phosphoric acid (50 per cent), glyceric aldehyde phosphoric acid (50 per cent), and phosphopyruvic acid (50 per cent). It is not certain that these compounds occur in invertebrate nerve, but in any event, since they serve as intermediates in lactic acid forma- tion, their content at any time would be small, except possibly for the hexosediphosphate.
The stable fraction determined by the increase after wet ashing represents the compounds wholly resistant to 7 minutes acid hy- drolysis as well as the unhydrolyzed portions of the others just considered. The stable compounds include hexosemonophos- phoric acid, phosphoglyceric acid, glycerophosphoric acid, and the nucleotides minus any pyrophosphate. The acid ashing method, though criticized, gave very consistent results.
Nerve Prepwation--Nerves of the large claw and the walking legs and the abdominal ganglion chain of the lobster (Homarus americanus) were utilized. The animals were placed on ice 1 to 2 hours before use, and the nerves dissected out into dry, iced mortars, stretch, as in pulling through joints, being carefully avoided. The tissue was gently blotted with filter paper, weighed
single animals were dissected in $ to 4 hour, values for inorganic P agreed accurately with present ones, but the arginine phosphate was twice as high. Since the 1933 data were less regular and complete and in disagreement with results by the Ca method, which had been fully controlled, we have used these only in summary fashion. All general findings were, however, entirely similar in the two sets. No indications of a creatine phosphate fraction was obtained, although this should have been obvious in hydrolysis curves. (See also Needham, Needham, Baldwin, and Yudkin (1933).)
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

382 P Metabolism of Invertebrate Nerve
rapidly on a torsion balance with a glass hook, and either ground at once or placed under the desired conditions. No drying occurred, water condensing rather from the moist air onto the cold tissue. When exposed at room temperature, however, nerves lost 5 per cent of their weight in 10 minutes; 12 per cent in an hour. Tissue from different animals was distributed equitably among the samples.
Nerves were exposed to oxygen or hydrogen on the walls of a 50 cc. dropping funnel closed with a stopper containing a straight tube stop-cock, and containing a few cc. of artificial sea water (NaCl, KCl, CaCL (Garrey, 1915) ). Gas was bubbled in through the water for 10 minutes after placing the nerves, and flushed through subsequently every hour. The temperature varied from 20-25” in different experiments.
Results
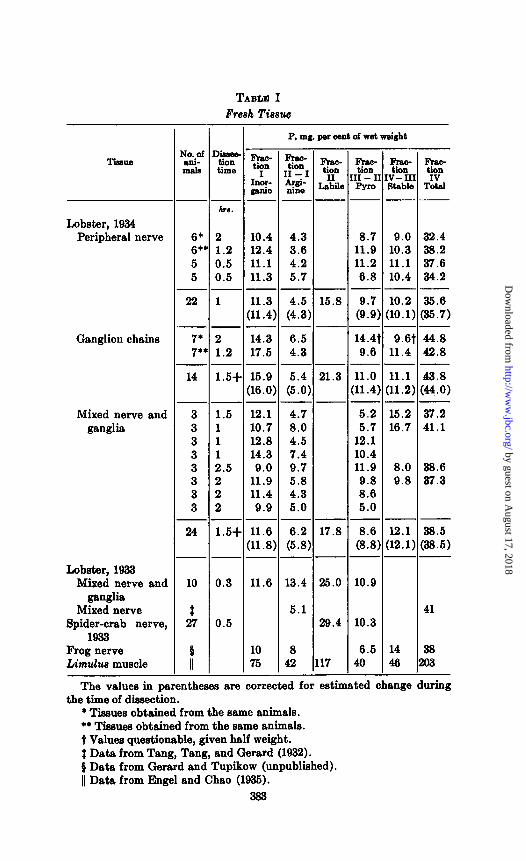
Fresh Nerve-The phosphate distribution in freshly dissected nerves is shown in Table I. The difference in different types of tissue is due in part to variable dissection time, for the same changes take place as during rest in oxygen, although slower, even when the nerves as dissected are kept at about 5’ and in air. Arginine phosphate, for example, is generally higher when 14 hours elapse between dissection of the first nerve and of the last than when .this time is only 1 hour. A rough correction for changes during dissection is obtained by taking half the dissection time, correcting for temperature difference (times 0.2), and assuming changes in 02 are linear with time. A 10 per cent lower arginine P and a 10 per cent higher pyrophosphate result. However, there is a real difference in the P values for claw nerves and ganglia of thesame animals (two experiments) ; although calculated per unit of dry weight (nerves 21 per cent solid, ganglia 24 per cent (Chang, 1931)) even this difference is within experimental error. The variation from batch to batch is inherent in the material, duplicate determinations on the same animals agreeing within 5 per cent. Earlier data showed Fractions II and III (the only ones deter- mined) to be 10 per cent greater in spider-crab nerve than in mixed nerve and ganglia of lobster. Data on frog nerve and Limulus muscle are included for comparison.
Rest in Oxygen-In Table II are summarized the changes during
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from
TABLP~ I Fresh Tissue
T Tissue
P. ma. per omt of wet wei&
“tit2 time %z IA- glmic be.
2 10.4 4.3 1.2 12.4 3.6 0.5 11.1 4.2 0.5 11.3 5.7
11.3 :11.4:
14.3 17.5
(i:::
6.5 4.3
15.9 ;lS.O:
1.5 12.1 1 10.7 1 12.8 1 14.3 2.5 9.0 2 11.9 2 11.4 2 9.9
;:t:
4.7 8.0 4.5 7.4 9.7 5.8 4.3 5.0
11.6 :11.8:
11.6 13.4
5.1
10 8 75 42
32.4 38.2 37.6 34.2
44.8 42.8
do. of i2l.i
6* 6” 5 5
!cticiK II - II IV- III Pyre Stable
--
8.7 9.0 11.9 10.3 11.2 11.1 6.8 10.4
-- 9.7 10.2
(9.9) (10.1) -- 14.4t s.si 9.6 11.4
-.- 11.0 11.1 :11.4) (11.2) --
5.2 15.2 5.7 16.7
12.1 10.4 11.9 8.0 9.8 9.8 8.6 5.0
-- 8.6 12.1
(8.8) (12.1) --
10.9
10.3
6.5 14 40 46
_
r
. .
.
I
. .
. .
. _
-
Lobster, 1934 Periphersl nerve
22 1 15.8
7* 71’
14
2 1.2
Ganglion chains
1.51 21.3 43.8 N?.O)
37.2 41.1
38.6 37.3
1.51
Mixed nerve and ganglia
17.8 24
Lobster, 1933 Mixed nerve and
ganglia Mixed nerve
Spider-crab nerve, 1933
Frog nerve Limulus muscle
10
t 27
0.3
0.5
25.0
29.4
117
41
38 to3
The values in parentheses are corrected for estimated change during the time of dissection.
* Tissue8 obtained from the same animals. ** Tissues obtained from the same animals. t Values questionable, given half weight. 2 Data from Tang, Tang, and Gerard (1932). 0 Data from Gerard and Tupikow (unpublished). 11 Data from Engel and Chao (1935).
383
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Cond
ition
Initia
l va
lues
Chan
ges
durin
g 1
hr.
in
01
Chan
ges
durin
g to
tal
time
in O
1 (8
hrs
., 19
34;
5 hr
s.,
1933
)
Chan
ges
in
01
afte
r 1s
t hr
.
- _-
_-
--
_-
-
Tissu
e*
Lobs
ter,
1934
G
angli
on
(14)
N
erve
(2
2)
Mixe
d (1
2)
Lobs
ter,
1933
(10
) Cr
ab,
1933
(4)
Lobs
ter,
1934
G
angli
on
Ner
ve
Mixe
d Lo
bste
r, 19
33
Crab
, 19
33
Lobs
ter,
1934
G
angli
on
Ner
ve
Mixe
d Lo
bste
r, 19
33
Crab
, 19
33
Lobs
ter,
1934
G
angli
on
Ner
ve
Mixe
d Lo
bste
r, 19
33
Crab
, 19
33
- - _-
.- . -
--
-
TABL
E II
Rest
in
Oxyg
en
IIKCg
IlniC
Ar
ginine
Py
re St
able
15.9
5.
4 11
.0
11.3
4.
5 9.
7 12
.5
6.1
8.5
11.9
13
.4
11.3
31
.0
9.2
11.1
10
.2
12.0
-0.5
+2
.2(2
.6)
-0.3
+1
.8(2
.0)
-0.6
+0
.9(1
.3)
+1.1
+3
.3
+3.9
-3.1
(3.5
) -1
.8(2
.0)
-0.6
(0.8
) -1
.7
-3.2
-0.3
(0.4
) +0
.3(0
.4)
-O.l(
O.l)
+0.4
+0
.9
+1.4
+l
.S
+2.0
-0
.7
+3.6
+3
.8
+1.4
-1.3
. -1
.0
+2.7
+0
.9
-2.2
-0.7
-2
.7
-3.4
+0.9
-1
.3
+1.7
-0
.2
+2.6
-1
.6
+2.5
+0
.5
-2.5
+1.s
+o
.s
+3.3
+1
.2
+1.0
-0.4
-3
.0
-3.3
P, m
g.
per
oentt
* Th
e va
lues
fo
r tis
sue
give
n id
pa
rent
hese
s re
pres
ent
the
num
ber
of a
nim
als
aver
aged
. t
The
valu
es
for
P gi
ven
in
pare
nthe
ses
are
corre
cted
fo
r es
timat
ed
chan
ge
durin
g di
ssec
tion.
- --
--
--
--
-
T&J
E 43
.8
35.7
39
.1
g g
-1.7
0.
0 3’
-0.4
o m
7 is 1
-0.5
g e
-0.7
3
+0.6
$ 3
+1.2
-0
.7
+1.0
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

G. L. Engel and R. W. Gerard
rest in oxygen, following the unavoidable stimulation of dissec- tion. All data are obtained by direct difference between com- panion samples. Since similar changes occur in the control nerves during the dissection period, the total change after dissection is greater than shown (see estimated correction). The data for ten experiments, with forty-eight animals, are averaged for nerve, ganglion chain, and both.
During short rest in oxygen (2 to 1 hour plus 1 to 12 hours dis- section) there is a slight decrease (5 per cent) in inorganic phos- phate and possibly in the total phosphate, with no change in the stable fraction. More striking is a regular and distinct increase (by one-third) of arginine phosphate which is balanced by a numerically equal loss of pyrophosphate (one-fourth of the total). In four experiments with rapid dissection and short rest in oxygen this balance was exact, as it was also in the average for all experi- ments with peripheral nerve (arginine phosphate, +1.8 mg. per cent; pyrophosphate, - 1.8). The averages for ganglion (+2.2 and -3.1) and for mixed nerve and ganglion (+0.9 and - 0.6) showed less perfect agreement, but in these experiments longer dissection and exposure times were involved, and one ganglion experiment was unreliable. Earlier results on crab and lobster were less regular but in agreement.
During further rest in oxygen (8 hours) these changes are partly reversed. Inorganic phosphate increases definitely (15 per cent) ; total phosphate, possibly; and the stable fraction decreases in all cases (20 per cent average). Arginine phosphate again decreases somewhat (15 per cent), while the pyrophosphate lost during the 1st hour is restored. The final balance after a long period in oxygen shows, therefore, no change from the initial values for total phosphate or pyrophosphatc and a dubious increase of arginine phosphate, while a definite shift of phosphate from the stable to the inorganic fraction has occurred. After only 1 hour, conversely, the only clear change is from pyrophosphate to arginine phosphate and this is subsequently reversed.
Rest .in Hydrogen-Anoxia leads to marked and rapid changes which have reached completion or equilibrium in 5 hours (Table III). Arginine phosphate is completely lost in nerve, three-fifths in mixed ganglion and nerve; and three-fourths or more of the pyrophosphate of nerve, somewhat less with ganglia present,
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

386 P Metabolism of Invertebrate Nerve
breaks down. These changes are complete by 2 hours. Inorganic phosphate increases for 5 hours, when it has doubled, at the ex- pense of the above fractions and also of the acid-stable one which decreases two-fifths in 5 hours.
In all cases the total acid-soluble P showed an unexpected decrease, greater after 5 hours than after 2, and contrary to earlier
Condition Tissue’
Initial values
Change during2.hrs. in Hz
Change during 5-9 hrs. in Ha
Change in HZ after first 2 hrs.
TABLE III Anoxia
Inor- gania
Argi- nine Pyre Stable Total
-----
Lobster, 1934 Nerve (20) Mixed(l2)
Lobster, 1933(5) Crab, 1933(3)
Lobster, 1934 Nerve Mixed
Lobster, 1933 Crab, 1933
1st 6t lot 15.4 44.3 10.5 6.2 8.8 8.9 34.4 12.6 14 13.9
31.4 10.5 ---
I j-10.5 -5.8 -6.8 -2.8 -5.0 +6.6 -3.3 -5.5 +0.7$ -1.5 j-12.8 -4.7 -7.8
+12.1 -8.6 ---
Lobster, 1934 Nerve Mixed
-6.6 -6.8 tlO.6 -3.4 -5.6 -O.S$ -1.21 -----
Lobster, 1934 Nerve Mixed +4.01-0.11 -o.l~~qG::t
* The values in parentheses represent the number of animals averaged. t Values estimated for this series. $ Values on mixed tissue are not all from complete parallel experiments,
therefore not in full agreement. Those marked are least certain. Proba- bly the stable fraction change at 5 hours should be considerably greater.
&dings on the frog and crustacean nerve (Gerard and Wallen, 1929; Tang, Tang, and Gerard, 1932). It is unlikely that a syn- thesis of insoluble compounds occurs under these conditions, and the previous results indicated the reverse change. More prob- ably, anoxia enhanced a loss of inorganic phosphate into fluid
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Cond
ition
Valu
es
afte
r 3
hrs.
in
HZ
Chan
ge
durin
g 1
hr.
in
0,
Chan
ge
durin
g 4-
6 hr
s.
in
On
Tim
lo’
Lobs
ter
nerv
e,
1934
(14)
Cr
ab,
1933
(3)
Lobs
ter
nerv
e,
1934
Lobs
ter
nerv
e,
1934
Cr
ab,
1933
TABL
E IV
R
ecov
ery
from
An
oxia
IUO~
ganiC
Ar
giniIl0
23.5
(+10
.5)
0.2(
-5.8
)
33.7
(+7.
5)
-4.5
1
+2.5
-7.4
(+3.
1)
+4.2
(-1.6
)
-1.9
(+5.
6)
P, m
g. p
er ce
ntt
MO
3.2(
-6.8
)
3.5(
-6.8
)
+2.8
+3.3
(-3.5
)
+2.6
(-3.8
)
Stab
le
13.4
(-2.6
)
-2.7
-4.6
(-7.2
)
* Th
e va
lues
fo
r tis
sue
give
n in
pa
rent
hese
s re
pres
ent
the
num
ber
of a
nim
ale
aver
aged
. t
The
valu
es
for
P gi
ven
in
pare
nthe
ses
give
th
e ch
ange
fro
m
the
initia
l va
lues
.
- --
_-
--
-
TOta
l
40.3
(-4.7
)
-1.9
-4.5
(-9.2
)
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

388 P Metabolism of Invertebrate Nerve
condensed or sprayed onto the tissue, and which unfortunately was not analyzed. In oxygen-lack the diffusible fractions are increased, and nerve permeability may also be greater (Hill, 1934; Gerard, 1934). Eggleton (1933) has observed an out-diffusion of phosphate when its concentration in muscle cells is increased relative to that in the interspaces.
Rest in Oxygen jollowing Hydrogen-Nerves (no ganglia were used) placed in oxygen after a period of anoxia show a marked recovery (Table IV). Arginine phosphate regenerates strongly and returns from zero to the level for fresh nerve in 6 hours. Py- rophosphate is restored from less than one-third of the original concentration to two-thirds after an hour in oxygen, but shows little further increase in another 5 hours. Inorganic phosphate falls from twice the initial value to 40 per cent more. The stable fraction, however, decreases further, to half the initial value, and the total soluble phosphate continues to fall from the value at the end of anoxia by an amount equal to the loss of the stable fraction.
Stimulation-In the 1933 series, in eleven experiments on lobster and crab, stimulation in oxygen with 10 to 35 shocks a second* caused a loss of 1.2 mg. per cent of arginine phosphate and 0.5 of pyrophosphate, compared to the resting controls. Since, on the one hand, stimulation was probably far from the maximal and, on the other, local electrode effects were not excluded, these results are only suggestive.
DISCUSSION
The breakdown of adenylpyrophosphate in conjunction with a synthesis of phosphoarginine is especially interesting. Since some injury and excitation during dissection are unavoidable, the subse- quent changes may be considered as a recovery process. This is further evidenced by the early synthesis of arginine phosphate during rest in oxygen. The results strongly indicate that the following reaction occurs during aerobic recovery of invertebrate nerve. Adenylpyrophosphate + 2 arginine + 2 arginine phosphate + adenylic acid
In at least half the experiments, those of short duration, the bal- ance was quantitatively exact. When longer times were used,
* We are indebted to Dr. H. Davis for the loan of a neon stimulator for these experiments.
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

G. L. Engel and R. W. Gerard
the arginine phosphate synthesis was greater than the apparent adenylpyrophosphate breakdown. This is undoubtedly the result of the observed resynthesis of adenylpyrophosphate at a later stage, after arginine phosphate is rebuilt. The main reaction in question is probably complete in 1 hour.
Lohmann (1934) concluded from experiments on muscle brei that the reverse reaction is associated with muscle activity.
(1) Adenylpyrophosphste + adenylic acid + 2 phosphoric acid (2) Adenylic acid + 2 creatinephosphoric acid + adenylpyrophosphate
+ 2 creatine
Any involvement of pyrophosphate has been challenged by Lundsgaard (1934), studying intact iodoacetate muscle, and in fact Meyerhof and Lohmann (1931) earlier observed that muscle extracts did not form phosphagen unless adenylpyrophosphate was present and that the nucleotide broke down as creatine phosphate formed. More recently Needham and van Heyningen (1935) have been able to obtain this reaction in either direction in muscle brei.3
On aerobic recovery following anoxia all the arginine phosphate is reformed in 5 hours, while two-thirds of the adenylpyrophos- phate is restored in 1 hour with no further significant increase. The stable fraction decreases by one-third during this period, probably with the formation of some compounds that appear partially in the pyrophosphate fraction. Presumably a balance is established in an hour between adenylpyrophosphate breakdown and its resynthesis from other phosphate sources.
A loss of 4.2 mg. per cent of adenylpyrophosphate P would be re- quired for the resynthesis of arginine phosphate; actually a gain of 3.3 mg. per cent is observed, so that a total synthesis of 7.5 mg. per cent of P as nucleotide is required. The phosphate for this could come from the stable fraction (maximum possible, 4.6 mg. per cent) and the inorganic one. In muscle brei, adenylic acid can be phosphorylated by phosphocreatine and by phosphopyruvic acid, probably acting via the phosphagen (Parnas, Ostern, and Mann, 1935; Needham and van Heyningen, 1935). The present evidence is against arginine serving as a donator in nerve. (This could
*An excellent statement of the case for muscle, indicating this rever- sibility and its functional significance, has been published by Parnas and Mann (1935).
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

390 P Metabolism of Invertebrate Nerve
possibly occur during the later stages of rest in oxygen.) Some of the lactic acid intermediates in the stable fraction are probably involved, although they cannot be the sole source of phosphate.
The extensive breakdown of both arginine and pyrophosphates during continued anoxia is in accord with previous results on frog and lobster nerve (see Gerard and Tupikow, 1931). To maintain even the resting nerve in its physiological state of responsiveness requires a continuous supply of energy (Gerard, 1932), derived ultimately from the oxidation of foodstuffs. During anoxia the necessary energy may be partly released by hydrolysis of the phos- phorus compounds, which are later restored, when oxygen is again available, with the aid of oxidative energy. The cycle of break- down and resynthesis presumably proceeds uninterruptedly under normal conditions and in any case there is a definite correlation between oxidation and phosphorylation. (See also Runnstrom, Lennerstand, and Borei (1934), Runnstrijm and Michaelis (1935), and Lipmann (1934). Contrast the view of Sacks and Sacks (1935), that, in muscle, acidity controls phosphorylation.)
Besides anoxia, long standing in oxygen leads to hydrolysis of arginine phosphate. Whether this results from depletion of sub- strate and reduced oxidative energy, or is a consequence of bac- terial action, cannot be stated. COZ, easily formed by bacteria, leads to phosphagen breakdown, even in 5 per cent gas concentra- tion (Gerard (1932) p. 499) and bacterial effects may be manifest in dog&h (Root, 1934) or mammalian (Schaffer, Chang, and Gerard, 1935) nerve in 7 hours or less, although mainly at higher temperatures.
The stable fraction tends to decrease, irreversibly, under all conditions studied except short rest in oxygen. It diminishes more rapidly under anoxic conditions than during prolonged aerobic rest, and after anoxia continues to fall even in oxygen. If this represents a loss of phosphorylated carbohydrate intermediates, these are presumably being destroyed more rapidly than they are reformed from glycogen, etc., of which a plentiful store exists (Holmes, 1929). Pending more specific measurement, this does not merit further discussion.
The total soluble phosphate remained unchanged in oxygen but diminished in oxygen-lack and continued to fall when oxygen was again admitted. The anoxic loss is most probably due to diffu-
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

G. L. Engel and R. W. Gerard 391
sional losses, as discussed, and the subsequent loss in oxygen may be similar. In the latter case, however, a synthesis of insoluble phosphates may have occurred, and it is interesting that the loss of total phosphate is equal to the loss in the stable fraction.
SUMMARY
Inorganic P, arginine P, adenylpyrophosphate P, stable P, and total acid-soluble P fractions have been determined in lobster claw nerves and ganglia and crab nerves.
During rest in oxygen, the phosphoarginine decomposed during dissection (and attendant stimulation and injury) is rebuilt, apparently by direct reaction with adenylpyrophosphate, which decomposes. This, in turn, is subsequently resynthesized, the lactic acid intermediates probably serving in part as phosphate donators. During anoxia there is an extensive hydrolysis of both phosphoarginine (up to 100 per cent) and adenylpyrophosphate, with an attendent rise in the inorganic fraction. Subsequent exposure to oxygen results in full resynthesis of arginine phos- phate in 5 hours; and of two-thirds of the adenylpyrophosphate within an hour, with no further change. Both substances are probably decreased by stimulation.
The stable fraction (hexosephosphate, etc.) regularly shows a loss of bound phosphorus after several hours, greater during or following the absence of oxygen. There is no evidence that the phosphorus of any acid-insoluble compounds (phosphoproteins, phospholipids, nucleoproteins, etc.) is involved in the aerobic metabolism of these nerves.
BIBLIOGRAPHY
Chang, T. H., Proc. Sot. Esp. Biol. and Med., 28, 954 (1931). Eggleton, M. G., J. Physiol., 79,31 (1933). Engel, G. L., and Chao, I., J. Biol. Chem., 108,389 (1935). Ferdmann, D., and Feinschmidt, O., Biochem. Z., 277,203 (1935). Fiske, C., and Subbarow, Y., J. Biol. Chem., 66,375 (1925). Garrey, W., Biol. Bull., 28, 77 (1915). Gerard, R. W., Physiol. Rev., 12,469 (1932); Science, 79, suppl., W (1934). Gerard, R. W., and Tupikow, N., Am. J. Physiol., 97,523 (1931). Gerard, R. W., and Wallen, J., Am. J. Physiol., 89, 163 (1929). Hill, A. V., Science, 79, suppl., 9 (1934). Holmes, E. G., Biochem. J., 23,1132 (1929). Lipmann, F., Biochem. Z., 274, 412 (1934).
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

392 P Metabolism of Invertebrate Nerve
Lohmann, K., Biochem. Z., 292,466 (1928); 271,264 (1934). Lohmann, K., and Jendrassik, L., Biochem. Z., 178,419 (1926). Lundsgsard, E., Biochem. Z., 289,308 (1934). Meyerhof, O., Die chemischen Vorgiinge im Muskel, Berlin (1930). Meyerhof, O., and Lohmann, K., Biochem. Z., 237,473 (1931). Needham, D. M., and van Heyningen, W. E., Nature, 136,585 (1935). Needham, D. M., Needham, J., Baldwin, E., and Yudkin, J., Z. physiol.
Chem., 222,63 (1933). Parnas, J. K., and Mann, T., Biochem. Z., 279, 94 (1935). Parnas, J. K., Ostern, P., and Mann, T., Biochem. Z., 276, 163 (1935). Root, W. S., J. Cell. and Comp. Physiol., 6, 141 (1934). Runnstriim, J., and Michaelis, L., J. Gen. Physiol., 18,717 (1935). Runnstr6m, J., Lennerstand, A., and Borei, H., Biochem. Z., 271,15 (1934). Sacks, J., and Sacks, W. C., Am. J. Physiol., 112,116 (1935). Schaffer, M., Chang, T. H., and Gerard, R. W., Am. J. Physiol., 111, 697
(1935). Tang, V., Tang, P. S., and Gerard, R. W., Proc. Sot. Exp. Biol. and Med.,
29,545 (1932). Teorell, T., Biochem. Z., 230, 1 (1931).
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

George L. Engel and R. W. GerardINVERTEBRATE NERVE
THE PHOSPHORUS METABOLISM OF
1935, 112:379-392.J. Biol. Chem.
http://www.jbc.org/content/112/1/379.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
alerts to choose from all of JBC's e-mailClick here
tml#ref-list-1
http://www.jbc.org/content/112/1/379.citation.full.haccessed free atThis article cites 0 references, 0 of which can be
by guest on August 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from







![Changes in composition of epigeic invertebrate communities (with accent on spiders) in Carpathian alpine meadows influenced by added nitrogen and phosphorus [Peter Gajdos]](https://img.pdfslide.net/doc/110x75/54c0656d4a79596e0b8b45ea/changes-in-composition-of-epigeic-invertebrate-communities-with-accent-on-spiders-in-carpathian-alpine-meadows-influenced-by-added-nitrogen-and-phosphorus-peter-gajdos.jpg)