Embed Size (px)
Citation preview

t~rBER S T O R U N G E N DES K E R N T E I L U N G S R H Y T H M U S . ZUGLEICH: UBER DEN EINFLUSS DER RONTGENSTRAHLEN AUF
DIE ZELLTEILUNG. HI. MITTEILUNG.
Von
G. POLITZER.
(Aus dem embryologischen Institute der Wiener Universit~t, Vorstand: A. FISCHEL.)
Mit 28 Textabbildungen. (Eingegangen am 22. Juli 1925.)
Gliederung. seite I. Einleitung, Material und Methode . . . . . . . . . . . . . . . 61
IL Ergebnisse . . . . . . . . . . . . . . . . . . . . . . . . . . 65 A. Der Einflu~ des Neutralrots auf den Kernteilungsrhythmus . . 65 B. Der EinfluB der RSntgenstrahlen auf den Kernteilungsrhythmus 67 C. Der Eiuflul3 des Brillantkresylblaus auf den Kernteflungsrhythmus 72 D. Der EinfluB der Hitze auf den Kernteilungsrhythmus . . . . 76
I I I . Allgemeiner Teil . . . . . . . . . . . . . . . . . . . . . . . 78 A. Die Wachstumsgeschwindigkeit und ihre Komponenten . . . . 78 B. t~ber unvollst~ndige Teilungsvorgange . . . . . . . . . . . . 80 C. ]~ber die sogenannte uuspecifische Wirkung der RSntgenstrahlen 83
IV. Zusammenfassung . . . . . . . . . . . . . . . . . . . . . . . 86 V. IAteraturverzeichnis . . . . . . . . . . . . . . . . . . . . . 89
I. Einleituug, Material und Methode.
Das W a e h s t u m - und daher auch jeder En twick lungsvorgang - - b e r u h t un te r anderem auf dem Vorgang der Zel l te i lung. W~hre nd fiber den formalen Ablauf der Caryokinese zahlreiche Unte r suchungen vor- liegen, beg innt m a n ers t in neuerer Zei t zielbewul]t nach den urs~ch- l ichen Bedingungen der Zel l te i lung zu forsehen. G~RWITSCH und seine Sehule k a m e n zu dem Ergebnis , dab die Ze]l tei lung durch zwei von- e inander unabh~ngig var i le rende F a k t o r e n geregel t sei. Auch die F r a g e der Beeinflul~barkeit de r Caryokinese durch ~uBere Mit te l verschieden- s te r A r t wurde neuer l ieh gepriif t . So h a t KORNFELD in unserem I n s t i t u t den EinfluB der F t i t t e rung auf den Te i lungs rhy thmus b e s t i m m t (1922). Es lag nahe auch die Wirkungsweise andere r F a k t o r e n a m gleichen Mater ia le wie KORNFELD ZU prtifen. - - KORNFELD (1925) un te r such te die W i r k u n g zahlre icher Chemil~alien (Salzs~ure, Kal i lauge , Subl imat , Alkohol , )~ther u. a. m.), w~hrend ALBERT1 und ~ch den EinfluB der RSntgens t rah len , des u l t r av io l e t t en Lichtes, der Fu lgu ra t i on und end-

62 G. Politzer:
lich der Vitalfarbstoffe festzustellen suchfCn, l~ber die formalen Veri~nderungen der Caryokinesen unter dem Einflusse dieser Reiz- mittel ist bereits beriehtet worden (ALBE~TI-POLITZER 1923, 1924, POLITZER-ALBERTI 1924, POLITZER 1924, 1925). In der vorliegenden Untersuchung sollen die StSrungen des Kernteilungsrhythmus be- sprochen werden.
Als Versuchsmaterial dienten Larven von Salamandra maculosa. Naeh der Entnahme aus dem Uterus wurden die Larven erst mehrere Tage gefiittert; dann wurde eine drei- bis vierti~gige Hungerperiode eingeschaltet, worauf neuerdings t/~glich gefiittert wurde. 4 5 Tage nach AbschluI~ der Hungerperiode wurden die Versuehe begonnen. Zu verschiedenen Zeitpunkten nach der Bestrahlung bzw. Vitalf~rbung wurden die Larven in BouINschem Gemisch fixiert und dann in 80 proz. Alkohol iibertragen. Zur Untersuchung gelangten Stiick- pr/~parate von Hornhaut, Kiemenpli~ttchen, Mundh6hlenboden u. a. m., die mit EHRLmHS H~matoxylin gef~rbt und in Canadabalsam ein- gesehlossen wurden. Hierauf wurde der Umril~ der I-Iornh~ute mit dem ZEIsssehell Projektionszeichenapparat bei schwacherVergr6Berung gezeichnet. ])ann wurden bei mittlerer Vergr6Berung die einze]nen Caryokinesen projek$ionsgereeht eingetragen. Hierzu wurde nur bei Pr/s mit sehr zahlreiehen Mitosen die Zeichenkreismethode von KORNFELD (siehe dor t )angewendet . In den meisten F/fllen ge- niigte eine sorgf/~ltige Durchmusterung der Hornhaut mit Hilfe des Objektfiihrapparates. Es wurde dies insbesondere dadurch erleichtert, dab ja in den zu sehildernden Versuchen nicht nur der ZeUteilungs- rhythmus ver/indert war, sondern aueh die einze]nen Caryokinesen sehr mannigfaltige Abnormit/s aufwiesen. Diese konnten durch einige Worte in der Besehreibung der einzelnen Ho~nh/~ute gekenn- zeichnet werden, so daf~ eine Verweehslung bzw. doppelte Z/~hlung vermieden wurde. Kontrollz/~hlungen ergaben, dab die Fehler bei dieser Methode nur sehr kleine sind. Die Z/~hlungen erstreckten sich jedoch nicht nur auf die Ermittlung der Gesamtzahl der Caryokinesen, sondern es wurde auch die H/~ufigkeit der einzelnen Kernteilungsphasen in jeder Cornea festgestellt. Hierbei bediente ich mieh der KOR~FELDschen Einteilung der Caryokinese, die hier im Wortlaut wiedergegeben wer- den soll.
,,Als ,a' bezeichnete ich (KoRN~'ELD) die Vorbereitung zur Mitose yon dem Zeitpunkte an, in dem sich der Zellkern dureh Anordnung seines Chromatins deutlieh yon einem ruhenden Kern unterscheidet und das dichte Spiremstadium, als ,b' den lockeren KnKuel, solange die Chromosomen noch nicht v611ig voneinander gesondert sind. Das Aster- stadium wurde als ,c', die Metakinese - - solange noch wenigstens ein Paar yon Toehterehromosomen eine Beriihrung zeigte - - als ,d', das

l~ber StSrungen des Kernteilungsrhythmus. 63
Diasterstadium als ,e ' bezeichnet; ,f' diente als Bezeichnung ffir das Di- spiremstadium, ,g' ffir die Endphase der Teilung, in der die betreffenden Toehterkerne sich yon ruhenden Kernen noch dureh auffallende Fs barkeit und charakteristische Form- und LageverhMtnisse auszeich- neten". - - PETER (1924) hat seither diese Einteilung erweitert. Er zer- legt das Stadium ,a' yon KORNFELD in die Stadien ,A' und ,B', wobei ,A' das Vorbereitungsstadium, ,B' den diehten Kn~uel darstellt. In ~hnlieher Weise grenzt er das zeffallene Spirem von der ~quatorial- platte ab, w~hrend diese beiden bei KORNFELI) unter der Bezeiehnung ,c' vereinigt sind. Ieh habe nun diese weiteren Unterteilungen nicht angenommen und zwar aus dem Grunde, weil ffir die Unterscheidung von ,A' und ,B' (PETER) kein objektives Zeiehen vorhanden ist, so dab die yon einem Uatersucher gewonnenen Zahlen mit denen anderer nicht verglichen werden kSnnen. Die Unterseheidung der beiden Asterphasen von PETER ist sehr schwierig, wenn die Zellen nicht in polarer oder seitlicher, sondern in schr~ger Richtung gesehen werden. Die Unter- teilung der ,A'-Phase ist jedoch wie in Abschnitt B des allgemeinen Teiles n/~her ausgeffihrv wird, yon einer gewissen Bedeutung, weswegen in man- chen F/~llen fiber die H/iufigkeit der An- fangsstadien berichtet werden wird.
Die gefundenen Werte ffir die einzelnen Phasen wurden in Tabellenform 1) gebracht und zur Erleichterung der Vorstellung graphisch dargestellt.
30
Zg
r
s I a b c d e f g
Abb. i,
Ehe wir nun zur Besprechung der Versuchsergebnisse fibergehen, sei einiges fiber die normale Phasenverteilung berichtet. In der KOR~- FELDSchen Untersuchung wurden insgesamt 7357 Mitosen gez~hlt. Davon entfallen 28 vH. auf ,a', 7 vH. auf ,b', 26 vH. auf ,c', 3 vH. auf ,d', 14 vH. auf ,e', 12 vH. auf ,f' und 10 vi i . auf ,g'. Tragen wir in ein Koordinatensystem die Phasen als Abszissen, die H~ufigkeit der einzelnen Phasen in Prozenten als Ordinaten auf, so erhalten wir die Kurve Abb. 1. Naeh KORNFELD herrscht in der Hornhaut normale Phasenverteilung, wenn ,a' und ,c' Maxima, ,b' und ,d' Minima, ,e', ,f' und ,g' Mittelwerte darstellen. Ich babe die Phasenverteilung in den von mir untersuchten Kontrolltieren bestimmt und komme, obwohl ich fiber eine wesentlich geringere Zahl ungeseh~digter Tiere (Kontroll- tiere) verffige als KORNFELD dennoch zu ann/ihernd gleichen Durch- schnittswerten wie er. Nur ist die Phase ,a' etwas weniger stark vertreten, so dab das Maximum der Verteilung in ,c', nicht in ,a' wie bei KORNFELD
1) Eine solche Tabelle ist bei der Brillantkresylblauwirkung abgebfldet. Die Wiedergabe der anderen wurde zwecks Raumersparnis unterlassen,

64 G. Politzer:
liegt. Dieser Unterschied ist meines Erachtens nicht auf die zu geringe Zahl von eigenen Untersuchungen zuriiekzufiihren; ich glaube vielmehr, daB KORNFELD Kerne, welche ich noch fiir Ruhekerne hare , als im Anfangsstadium der Teilung befindlich betrachtete. Diese Verschieden. heit in der Wertung einzelner Kernteilungsbilder ftihrt naturgem~B bei verschiedenen Untersuchern zu verschiedenen Z~hlungsergebnissen, be. sonders dann, wenn sichere objektive Unterscheidungsmerkmale zweier Phasen fehlen.
Ffir die in dieser Arbeit wiedergegebenen Kurven wurden mit Aus- nahme der Abb. 1 und der nach WASSERMA~ berechneten Kurven (Abb. 26--28) stets die absoluten Zahlen verwendet. Ferner wurden die Werte in beiden Hornh~uten addiert. Die ~bereinstimmung der Er- gebnisse in beiden Corneen war meist eine sehr gute. Mitunter waren die absolutee Werte in einer Hornhaut hSher als in der anderen, die Phasenverteilung war jedoch die gleiche. GrSfiere Untersehiede bestanden nur sehr selten und in diesen Fhllen meist im Beginne hochgradiger RhythmusstSrungen, so dab es den Anschein hatte, als versp~te sich eine Hornbaut ein wenig in ihrer Reaktion auf die Sch~digung. Diese Ergebnisse stimmen sehr gut mit den Beobaehtungen KORNFELDS (1922) iiberein.
Der grSBte Teil der yon KORNFELI) und mir untersuchten Reiz- mittel erwies sich nun fiir das Studium der StSrungen des Zellteilungs- rhythmus als ungeeignet: Dies lag einerseits in der Beschaffenheit der Versuchstiere, andererseits in der Natur des Reizmittels. Vor allem lie[~en sich nur solehe Wiirfe verwenden, bei denen die Mitosenzahl in der t tornhaut hinreiehend groI~ war. Eine entsprechende Steigerung der Caryokinesenzahl gelang oftmals aueh nicht mit Hilfe der KORNFELD- schen Fiitterungsweise. In keinem meiner Versuche konnte ich bei den KontroUtieren so hohe Mitosenzahlen erzielen wie KORNFELD. Auch PETER machte ~hnliehe Erfahrungen. Viele Versuche mui~ten deshalb unverwertet bleiben, weil neben den Seh~digungen des Kernteilungs- rhythmus und der Caryokinesen hochgradige Sch~digungen der Ruhe- kerne eintraten. Dies l~l~t sich unsehwer aus der ri~umlichen Verteilung der Mitosen in der Hornhaut verstehen. Die Zellteilungen liegen nahezu ausschlie~lich in der basalen Schichte, w~hrend die oberfl~ichliche mito- senfrei ist. Reizmittel, die von auBen her auf die Hornhaut einwirken, miissen erst die oberfl~chliche, dann die tiefe Schichte treffen. Die ZeUen der oberfl~chlichen werden daher wohl starker geseh~digt als die der tiefen. Da jedoch die Mito~en bereits Sch~idigungen erkennen lassen, welehe sich bei Ruhekernen unserer Beobachtung entziehen, linden wit mitunter ausschliel~lieh StSrung der Kernteilungsti~tigkeit, w~hrend die Ruhekerne normal aussehen. Dies t~uscht in vielen F~llen eine st~rkere Empfindlichkeit der Mitosen vor, obgleich es sieh in

Uber StSrungen des Kernteilungsrhythmus. 65
Wahrheit nur um die/riihzeitige Erkennung einer sonst latent bleibenden Sch5digung handelt. Hiermit soll jedoch keineswegs eine wirkliche hShere Empfindlichkeit der Mitosen gegen manche Reizmittel in Abrede gestellt werden. Diese scheinbare (oder wirkliche) h(ihere Empfindlich- keit der Caryokinese maehte unsere Versuche iiberhaupt erst m6glich, d a selbst bei durchdringenden Reizmitteln die oberfl~chliche (mitosen- fide) Schicht stgrker geschgdig~ werden mul3. Die meisten der ange- wendeten Reizmittel zeigen nun einen so starken Wirkungsabfall beim Durchtritt dutch die Hornhaut, dab trotz der geschilderten Empfind- hchkeitsunterschiede die oberfl~chliche Schicht bereits necrotisch wird, ehe die Caryokinesen in der Tiefe irgendwie yon der Norm abweichen. In dieser Art wirken die meisten Chemlkalien (S~iuren, Laugen, manche Vitalfarbstoffe, z. B. Methylvioletr ferner die ultravioletten Strahlen, die photodynamische Wirkung des Lichtes auf Tiere in fluorescierenden FarblSsungen usw. Von den untersuchten Reizmitteln entsprach nur die Wirlmng der RSntgenstrahlen und mancher Vitalfarbstoffe {Neutral- rot, Nilblau, Auramin, Brillantkresylblau) der Bedingtmg, Mitosenver- gnderungen ohne hochgradige Sch~digungen der Oberfl~chenschicht hervorzurufen. Eine groBe Schwierlgkeit bot die Art der Dosierung. In den meisten Fgllen liegt die toxische Dosis dicht an der letalen. Es gelingt deshalb nur schwer, jene Dosis zu finden, die bereits Ver~inde- rungen an den Caryokinesen auslSst, ohne jedoch die Tiere schon abzutSten.
In folgendem sollen nun vier verschiedene Typen yon Rhythmus- stSrungen gezeigt werden, yon denen drei an meinem Versuchsmaterial festgestellt wurden, wghrend die vierte aus den Tabellen einer Unter- suchung WASSER~AN~S berechnet wurde.
II. E r g e b n i s s e .
A. Der Einflufl des Neutralrot~ au! den Kernteilungsrhythmus.
Dem Neutralrot kommt eine charakteristische Giftwirkung zu, welche ich bereits beschrieben habe (1924): Im Anschlusse an eine mehrstiindige F~rbung mit einer LSsung yon etwa 1 : 100 000 treten in der Hornhaut Pseudoamitosen aufl). Nach etwa 10 Stunden ist die Hornhaut mitosenfrei. Each etwa 2 Tagen erscheinen neuerdings Caryokinesen, die voUkommen normales Aussehen zeigen und nur au Zahl hinter der Norm zurfickbleiben. Die Z~hlung der Caryokinesen in einer der Neutralrotserien ergab nun das in den Kurven Abb. 2--5 dargestellte Ergebnis. Hierzu ist noch folgendes zu bemerken. Die
1) Die Entstehung und die verschiedenen Ablaufsarten der Pseudoamitosen wurde in friiheren Arbeiten eingehend geschildert (ALBERTI-POLITZER 1923, POLITZER 1924).
Z. f. Zellforschung u. mikr. Anatomie Bd. 3. 5
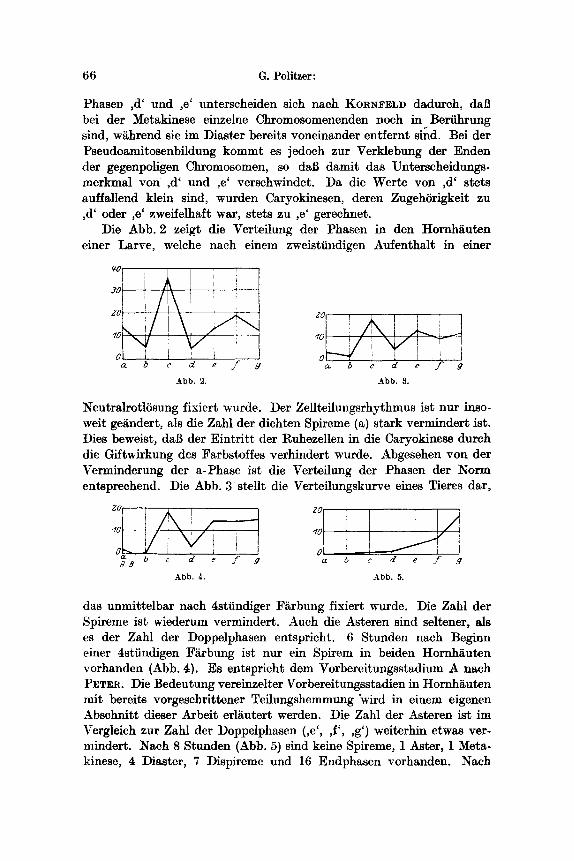
66 G. Politzer:
Phase~ ,d' und ,e' unterscheiden sich nach KORNFELD dadurch, dab bei der Metakinese einzelne Chromosomenenden noch in Beriihrung sind, w~hrend sie im Diaster bereits voneinander entfernt sind. Bei der Pseudoamitosenbildung kommt es jedoeh zur Verklebung der Enden der gegenpoligen Chromosomen, so dab damit das Unterscheidungs- merkmal yon ,d' und ,e' verschwindet. I)a die Werte yon ,d' stets auffallend klein sind, wurden Caryokinesen, deren ZugehSrigkeit zu ,d' oder ,e' zweifelhaft war, stets zu ,e' gerechnet.
Die Abb. 2 zeigt die Verteilung der Phasen in den Hornh~uten einer Larve, welehe nach einem zweistiindigen Aufenthalt in einer
q5
3 5
5 a b c d e 3 " g a b c d e f g
A b b . 2. A b b . 3.
NeutralrotlSsung fixiert wurde. Der ZeUteilungsrhythmus ist nur inso- welt getindert, als die Zahl der dichten Spireme (a) stark vermindert ist. Dies beweist, dab der Eintrit t der Ruheze]len in die Caryokinese dutch die Giftwirkung des Farbstoffes verhindert wurde. Abgesehen yon der Verminderung der a-Phase ist die Verteilung der Phasen der Norm entspreehend. Die Abb, 3 stellt die Verteilungskurve eines Tieres dar,
a I 0 ' i ~ '
R B b c d e f g ~ b c ,~ e f 3
A b b . 4. A b b . 5.
das unmittelbar nach 4stiindiger F~rbung fixiert wurde. Die Zahl der Spireme ist wiederum vermindert. Auch die Asteren sind seltener, als es der Zahl der Doppelphasen entspricht. 6 Stunden nach Beginn einer 4stiindigen F~rbung ist nur ein Spirem in beiden Hornh~uten vorhanden (Abb. 4). Es entspricht den* Vorbereitungsstadium A nach PETV.a. Die Bedeutung vereinzelter Vorbereitungsstadien in Hornh~uten mit bereits vorgeschrittener Teflungshemmung "wird in einem eigenen Abschnitt dieser Arbeit erl~utert werden. Die Zahl der Asteren ist im Vergleich zur Zahl der Doppelphasen (,e', ,f', ,g') weiterhin etwas ver- mindert. Nach 8 Stunden (Abb. 5) sind keine Spireme, 1 Aster, 1 Meta- kinese, 4 ]:)iaster, 7 ])ispireme und 16 Endphasen vorhanden. Nach

~ber StSrungen des Kernteilungsrhythmus. 67
weiteren 3 Stunden (11 Stunden} ist die Zellteilungst~ttigkeit voll- kommen erloschen.
I)er Neutralrottypus der RhythmusstSrung ist dadurch gekenn- zeichnet, dab die vorhandenen Mitosen ablaufen, ohne dab neue gebildet werden. Auch bei starker ErhShung der Konzentration der Neutral. rotl6sung bleibt die Wirkungsweise die gleiche. Auffallend ist die lange Dauer des Ablaufes der vorhandenen $fitosen. Da diese Verlangsamung der Caryokinese bei der RSntgenwirkung deutlicher in Erscheinung tritt, sol[ sie erst dort ausfiihrlicher besprochen werden.
B. Der Ein/lufl der RSntgenstrahlen au] den Kernteilungsrhythmus.
Als zweite Wirkungsweise sei die der R6ntgenstrahlen besproehen. ALB~RTI und ich (1923, 1924) konnten zeigen, dab im Ansehlul3 an ein RSntgenbestrahlung die Zahl der Caryokinesen unter Auftreten abnormer Teilungsfiguren rasch abnimmt. 6--10 Stunden nach der Bestrahlung ist die Cornea mitosenfrei. Nach einigen Tagen treten neuerdings Caryokinesen auf, welehe jedoch (im Gegensatz zur Neutral- rotwirkung) abnorm sind. Diese drei Abselmitte der RSntgenwirkung haben wit a|s ,,Prim&reffekt", ,,mitosenfreie Zwischenzeit" und ,,Se- cund~reffekt" bezeichnet. Der Primk'reffekt zeigt Pyknose der Asteren und Pseudoamitosen. Seine Dauer ist unabh&ngig vonder angewendeten Dosis. Trotzdem lieB er Sehwankungen in seiner Dauer bei Versuehen, die zu versehiedenen Jahreszeiten unternommen wurden, erkennen, Sehwankungen, welehe wir uns bisher nieht erkl~ren konnten. Die mitosenfreie Zwischenzeit ist in ihrer Dauer yon der RSntgendosis ab- h~ngig. Sie betrug (1924) bei 1/.~ H wenige Stunden, bei 1 H einen Tag, bei 3--4 H 3 Tage, bei 12 H 5 und bei 20 H 8 Tage. Der Secundiir- effekt dauert Wochen hindurch an und ist durch Chromosomenablen- klmg, Teilkernbildung, Deviation der Tochtersterne u a.m. gekenn- zeichnet.
Fiir die vorliegende Untersuehung wurden zwei neuerliche Versuchs- reihen angefertigt. Da mir auffiel, da~ der Prim~reffekt im Sp~therbst l~nger dauerte als im Frfihling, lag der Gedanke nahe, daft sich hinter diesen Unregelmiifligkeiten ein EinfluB der Temperatur auf die RSntgen- effekte verberge. Deshalb wurde eine Reihe yon Tieren bei 10 ~ (d. h. in fliel]endem Leitungswasser gekiihlt), die zweite bei 26 ~ C (in Thermo- staten) gehalten. Zur Bestrahlung wurden die Tiere aus dem Thermo- staten bzw. Kfihlapparat entnommen, so daft w&hrend der Bestrahlung keine Konstanz der Temperatur bestand. Die Bestrahlung wurde im Central-RSntgenlaboratorium des allgemeinen Krankenhauses in Wien (Vorstand: Prof. G. HOLZK~ECHT) an einem Veifa Neo-Intensiv-Refom- apparat mit Coolidger(ihre durchgeffihrt. Die Stromst~rke hn Secund~r- strom war 2 MA, die Spannung 65 KV, die Objektfokusdistanz 25 cm,
5*
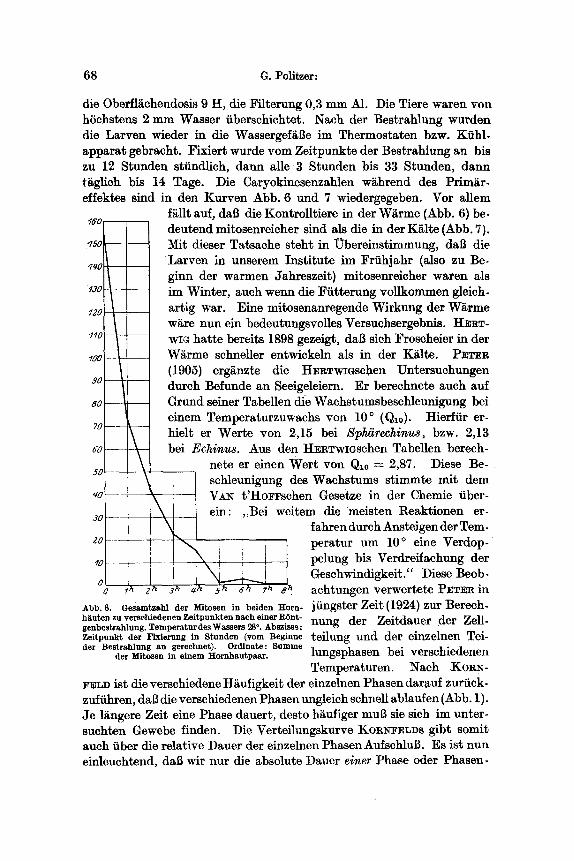
68 G. Politzer:
die Oberfl~ehendosis 9 H, die Filterung 0,3 m m A1. Die Tiere waren yon hSehs~ens 2 mm Wasser iibersehichtet. ~Iaeh der Bestrahlung wurden die Larven wieder in die Wassergef~Be im Thermostaten bzw. Ktihl- appara t gebraeht. Fixiert wurde vom Zeitpunkte der Bestrahlung an bis zu 12 Stunden stiindlieh, dann alle 3 Stunden bis 33 Stunden, dann tiiglich bis 14 Tage. Die Caryokinesenzahlen wi~hrend des Primiir- effektes Mind in den Kurven Abb. 6 und 7 wiedergegeben. Vor allem
f~llt auf, dab die Kontrolltiere in der Wiirme (Abb. 6) be- deutend mitosenreieher sind als die in der K~lte (Abb. 7}.
15o Mit dieser Tatsaehe steht in ~bereinst immung, dab die Larven in unserem Inst i tute im Friihjahr (also zu Be-
~q0 ginn der warmen Jahreszeit) mitosenreicher waren als
~3o im Winter, auch wenn die Fiit terung voUkommen gleich- ~2o artig war. Eine mitosenanregende Wirkung der Wi~rme
ware nun ein bedeutungsvolles Versuehsergebnis. HERT- I~0 -- WIG hat te bereits 1898 gezeigt, dab sieh Froseheier in der ~aa- W~rme schneller entwickeln als in der Kiflte. PETER
{1905) ergiinzte die HERTWmsehen Untersuchungen so dutch Befunde an Seeigeleiern. Er bereehnete auch auf so - Grund seiner Tabellen die Waehstumsbesehleunigung bei
einem Temperaturzuwaehs yon 10 ~ (Qlo). Hierfiir er- 7 o -
hitelt er Werte yon 2,15 bei Splffirechinus, bzw. 2,13 ~o - bei Echinus. Aus den HERTWmsehen Tabellen bereeh-
nete er einen Wef t yon Qlo = 2,87. Diese Be- 55 schleunigung des Waehstums s t immte mit dem
~o!- \ V~-~ t 'HoFFsehen Gesetze in der Chemie iiber- \ _ ein: ,,Bei weitem die meisten Reaktionen er-
30 \ fahren durch Ansteigen der Tem- zo- ~ peratur um 10 ~ eine Verdop- ~o-- - - \ ! pelung bis Verdreifachung der
Gesehwindigkeit." Diese Beob- a 0 ~ e~ 3~ ~ ~ 8~ achtungen verwertete PETER in
Abb. 6. Gesamtzahl der Mitosen in beiden Horn- ]iingster Zeit (1924)zur Berech- h~uten zu verschiedenen Zeitpunkten nach einer R6nt- h u n g der Zeitdauer der Zell- genbestrahlung. Temperatur des Wassers ~26 ~ Absziss9: Zeitpunkt der Fixierung in Stunden (yore Beginne t e i l u n g und der einzelnen Tei- der Bestrahlung an gerechnet). Ordinate: Somme
der Mltosen in einem Hornhautpaar . hingsphasen bei verschiedenen Temperaturen. Nach KOR~-
F ~ D ist die verschiedene H~ufigkeit der einzelnen Phasen dar~uf zuriick- zufiihren, dab die versehiedenen Phasen ungleich sehnell ablaufen (Abb. 1). Je l~ngere Zeit eine Phase dauert, desto h~ufiger mu~ sie sieb im unter- suchten Gewebe finden. Die Verteilungskurve KORNFELDS gibt somit auch fiber die relatiVe Dauer der einzelnen Phasen AufschluB. Es ist nun einleuchtend, dab wit nur die absolute Dauer einer Phase oder Phasen-
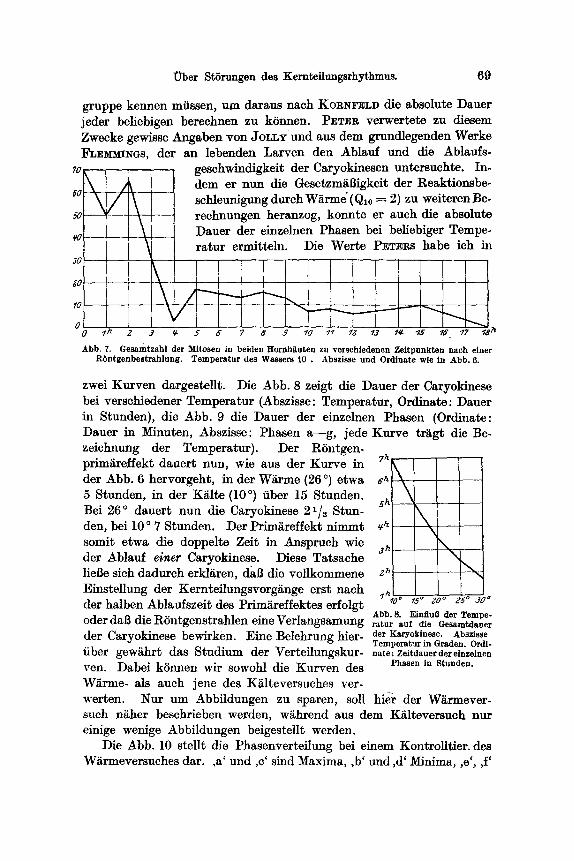
Uber St6rungen des Kemteilungsrhythmus. 69
ii gO
gruppe kennen miissen, um daraus nach KORNFELD die absolute Dauer jeder beliebigen berechnen zu kSnnen. PETER verwertete zu diesem Zwecke gewisse Angaben yon JOLLy und aus dem grundlegenden Werke FLEI~MI~GS, der an lebenden Larven den Ablauf und die Ablaufs-
geschwindigkeit der Caryokinesen untersuchte. In- ~ ~ ' dem er nun die Gesetzm~Bigkeit der Reaktionsbe-
/ schleunigung durch W~rme" (Qlo -~ 2) zu weiteren Be- reehnungen heranzog, konnte er such die absolute Dauer der einze]nen Phasen bei beliebiger Tempe-
~ ratur ermitteln. Die Werte PETEP~S habe ich h
0 I 0 "/ g 3 ~ 5 6" 7 8 .9 ~0 ~ IZ "/3 ~'r "/5 18 ~7 ":8 ~
Abb. 7. Gesamtzahl der Mltosen in beiden Hornh~uten zu verschiedenen Zeitpunkten nach einer R6ntgenbestrahlung. Temperatur des Wassers l0 . Abszisse und Ozdinate wie in Abb. 6.
zwei Kurven dargestellt. Die Abb. 8 zeigt die Dauer der Caryokinese bei versehiedener Temperatur (Abszisse: Temperatur, Ordinate: Dauer in Stunden), die Abb. 9 die Dauer der einzelnen Phasen (Ordinate: ])auer in Minuten, Abszisse: Phasen a--g, jede Kurve tr~g~ die Be- zeichnung der Temperatur). Der R6ntgen- prim~reffekt dauert nun, wie aus der Kurve in der Abb. 6 hervorgeht, in der W~rme (26 ~ etwa 5 Stunden, in der K~ilte (10 ~ fiber 15 Stunden. Bei 26 ~ dauert nun die Caryokinese 21/2 Stun- den, bei 10 o 7 Stunden. I)er Prim~effekt nimmt somit etwa die doppelte Zeit in Anspruch wie der Ablauf einer Caryokinese. Diese Tatsache lieBe sich dadurch erkl~ren, dab die vollkommene Einstellung der Kernteilungsvorg~nge erst nach der halben Ablaufszeit des Prim~reffektes effolgt oder dab die R6ntgenstrahlen eine Verlangsamung der Caryokinese bewirken. Eine Belehrung hier- fiber gew~hrt das Studium der Verteilungskur- yen. Dabei kSnnen wir sowohl die Kurven des W~rme- als auch jene des K~lteversuehes ver- werten. Nur um Abbildungen zu sparen, soll hie-r der W~mever- such n~her beschrieben werden, w~hrend aus dem K~lteversuch nur einige wenige Abbildungen beigeste]lt werden.
Die Abb. 10 steUt die Phasenverteilung bei einem Kontrolltier. des W~irmeversuches dar. ,a' und ,c' sind Maxima, ,b' und ,d' Minima, ,e', ,f'
7 ~
6.h
\
I~0o IB ~ ZO ~ Z5 ~ 30 e
Abb. 8. Einflu~ der Tempe- ratur auf die Gesamtdauer der Karyokinese. Abszisse Temperatur in Graden. Ordi- nate: Zeitdauer der einzelnen
Phasen in Stunden.
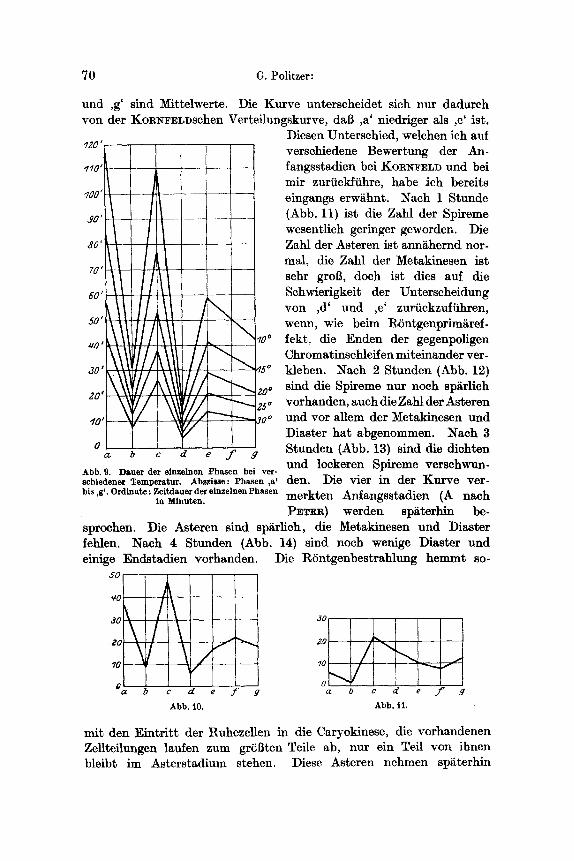
70 G. Politzer:
und ,g' sind Mittelwerte. Die Kurve unterscheidet sich nur dadurch yon der KOR~FELDschen Verteilungskurve, dab ,a' niedriger als ,c' ist.
"120'
�9 1 1 0 j
30'
80'
~ //11 ::, // II /\
IIl\ll // \,oo -"\i;ll//\ \ ,o. :II, N I/i x-._ ,o. ,~,// ,~.///--~.o
, ,// 0
,z b c d e f ,9"
Abb. 9. Dauer der elnzelnon Phasen bei ver- schiedener Temperatur, Abszisse : Phasen ,a ~ bis ,g'. O r d i n a t e : Zeitdauer der einzelnen Phasen
in Minuten.
Diesen Unterschied, welchen ich auf verschiedene Bewertung der An- fangsstadien bet KORS~ELD und bet mir zuriickfiihre, habe Jch bereits eingangs erw/~hnt. Nach 1 Stunde (Abb. 11) ist die Zahl der Spireme wesentlich geringer geworden. Die Zahl der Asteren ist ann~hemd nor- mal, die Zahl der Metakinesen ist sehr groB, doch ist dies auf die Schwierigkeit der Unterscheidung yon ,d' und ,e' zuriickzuf/ihren, wenn, wie beim RSntgenprim&ref- fekt, die Enden der gegenpoligen Chromatinschleifen miteinander ver- kleben. Nach 2 Stunden (Abb. 12) sind die Spireme nur noch sp~rlich vorhanden, auch die Zahl der Asteren und vor allem der Metakinesen und Diaster hat abgenommen. Nach 3 Stunden (Abb. 13) sind die dichten und lockeren Spireme verschwun- den. Die vier in der Kurve ver- merkten Anfangsstadien (A nach P~.TER) werden sp~terhin be-
sprochen. Die Asteren sind sp~rlich, die Metakinesen und Diaster fehlen. Nach 4 Stunden (Abb. 14) sind noch wenige Diaster und einige Endstadien vorhanden.
so I
/\ "\/\ ,o'~/ \ / 0
a b c d e f 9
Abb. i0 .
Die RSntgenbestrahlung hemmt so-
JO I
c~ b c d e f y
Abb. t L
mit den Eintr i t t der Ruhezellen in die Caryokinese, die vorhandenen Zellteilungen laufen zum grSBten Teile ab, nut ein Teil von ihnen bleibt im Asterstadium stehen. Diese Asteren nehmen sp~iterhin

~ber StSrungen des Kernteilungsrhythmus. 71
gleichfalls an Zahl ab, nicht aber weft sie welter gefiihrt werden - - sonst miiBten ja die Phasen ,d' und ,e' auftreten - - sondern weil sie auf andere Weise ausseheiden. Diese Asteren sind n~mlieh durchwegs hochgradig pyknotisch und es lieg~ nahe, anzunehmen, dab sie zu- grunde gehen und ausgestoBen werden. DaB zur Zeit des Prim~reffektes Nekrosen auftreten, wurde bereits in friiheren Untersuchungen be- schrieben (PoLrrZER-ALBERTI 1924). Der K~lteversuch lieferte ein iibereinstimmendes Ergebnis. So ist zwei Stunden nach der Bestrahlung
k I ton, 1 - ~ ~ ~ d e f 3 b c d e f 9' ~ 8
Abb. i2. Abb. 13.
(Abb. 15) die Phase ,a' nur mehr in geringem MaBe vertreten. Naeh 3 Stunden sind keine Spireme mehr auffindbar, naeh 5 Stunden (Abb. 16) tritt wieder die typische zweigipfelige Kurve auf. Der erste Gipfel entspricht den nekrotisierenden Asteren, der zweite den sp~ten Doppel- phasen der trotz der Bestrahlung ablaufenden Caryokinesen.
Die Ergebnisse dieser Versuche gestatten eine Erkl~rung der relativ langen Dauer des Prim~reffektes. Wfirde die Caryokinese im K~lte- versuch normal lang dauern, so miiBten die letzten Spireme noch naeh
2O
~ / b c d e 3" 9
k b b . I~.
2~
0 5 c d e f g ~z b c d e y q
Abb, 15. Abb. 16.
8 Stunden naehweisbar sein. Dann wiirde der Ablauf dieser Formen his zum Versehwinden der Mitosen iiberhaupt, so ~Se es den PETER- schen Angaben fiir diese Temperatur entspricht, 7 Stunden dauern. Die Spireme sind jedoch bereits nach 3 Stunden v~llig verschwunden. Die Asteren, die in diesen HornhKuten vorhanden sind, brauchen somit zu ihrem Ablaufe 12 Stunden, w~hrend in der Norm die Entwicklung von ,e' bis ,g' (siehe Abb. 9) weniger als 5 Stunden dauert. Daraus geht hervor, dab die RSntgenstrahlen den Ablauf der Caryokinese wesentlich verlangsamen. Auf diese Verlangsamung der Caryokinese ist aueh die auffallend lange Dauer des Prim~ireffektes zuriickzuffihren.

72 G. Politzer:
Der Einflul3 der RSntgenstrahlen auf den Zellteflungsrhythmus ist somit folgender:
1. Die Dauer des Prim~reffektes ist von der Temperatur des Wassers, in dem die Versuchstiere gehalten werden, abh~ngig.
2. I)er Prlmii~ffekt dauert etwa doppelt so lange wie der normale Ablauf einer Caryokinese bei der Temperatur des Versuches.
3. Die RSntgenstrahlen bewirken eine Hemmung des Zellteilungs- vorganges.
a) Die Ruhezellen werden am Eintreteu in die Caryokinese gehindert. b) Die zur Zeit der Bestrahlung vorhandenen Zellteflungen werden
zum groBen Tefl ~ wenn auch wesentlich langsamer als es der Tempe- ratur entspricht ~ zu Ende gefiihrt.
c) Ein Tell der Asteren wird jedoch pyknotisch, die betreffenden Zellen gehen zugrunde und werden aus der Hornhaut ausgestoBen.
C. Der I~in/lu~ des Brillant~esylblau8 au] den Kernteilung~rhythmus. Brillantkresylblau und Auramin bewirken hochgradige StSrungen
des ZeUteflungsrhythmus. Auch die Mitosen selbst sind abnorm. Die Chromosomen in den Asteren liegen dicht geballt, aber nieht unter- einander verbacken (pyknotisch) im Centrum der Zelle. Die Chromo- somen sind kiirzer und dicker als in der Norm, jedoch untereinander yon gleicher GrSBe. Sie entsprechen den yon KOR~YELD jiingst be- schriebenen Maulbeerformen.
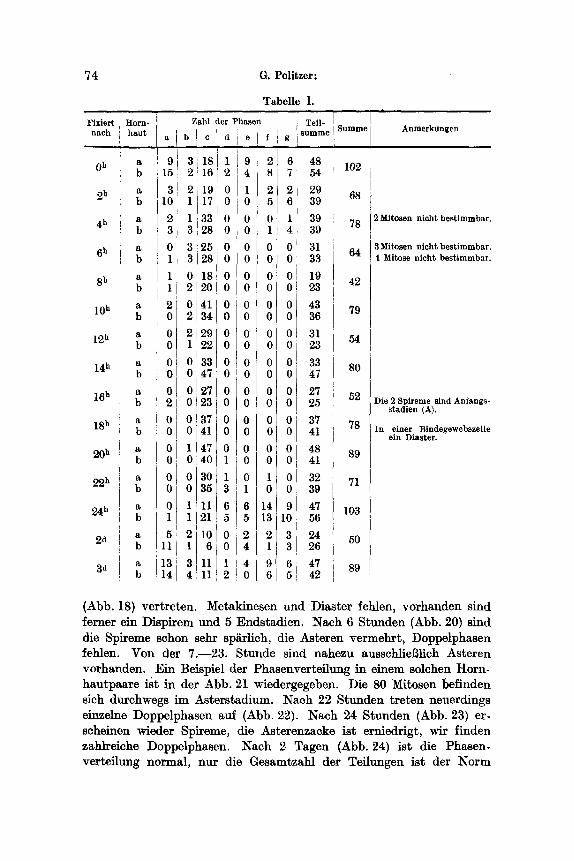
Fiir die nachfolgenden Auaffihrungen lag eine Versuchsreihe yon Tieren vor, die dutch 12 Stunden mit BriUantkresylblau in einer Kon- zentra.tion yon 1 : 20 000 gef~rbt worden war. Die Fixierungen wurden 2stiindig yore Beginn der F~rbung bis zu einem Tag vorgenommen, 1 Tier wurde nach 2, 1 nach 3 Tagen fixiert. Tabelle I enth~ilt eine Zusammenstellung der Z~hlungsergebnisse. Die Gesamtzahl der Caryo- kinesen in beiden Corneen bei s~mtllchen Versuchstieren der Reihe wurde in der Kurve der Abb. 17 graphisch dargestellt. Die Abszisse gibt den Zeitpunkt der Fixierung, die Ordinate die Summe der Caryo- kinesen in beiden Hornh~uten des Versuchst, ieres an. Die Kurve zeigt unregelm~Bige Zacken. Zwei Gipfel sind deutlich erkennbar. Sie be- treffen das KontroUtier (102 Mitosen) und das nach einem Tag fixierte (103 Mitoaen). Die iibrigen Schwankungen sind jedoch geringe. Sie k6nnen ebensogut in Kurven vorkommen, die yon normalen, unbeein- fluBten Tieren stammen, da sich auch bei diesen Schwankungen ~hn- lichen AusmaBes linden. Aus dieser Kurve ist nicht erkennbar, daB, wie aus den Verteilungskurven Abb. 18--25 hervorgehen wird, eine 16 Stunden w~hrende voUkommene Hemmung der Caryokinese vor- handen ist. Diese Tatsache zeigt auch die Unzul~nglichkeit der Summen-
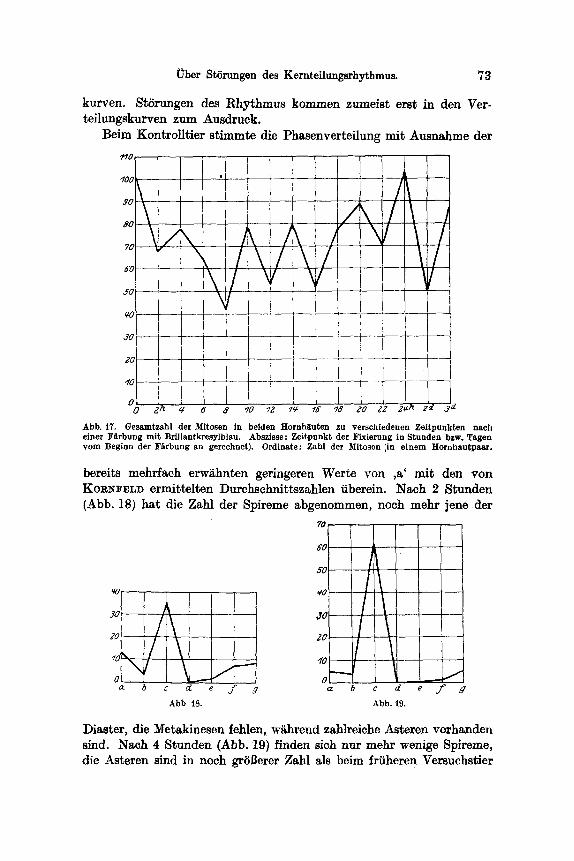
~ber St6rungen des Kernteilungsrhythmus. 73
kurven. St6"rungen des Rhythmus kommen zumeist erst in den Ver- teihmgskmwen zum Ausdruck.
Beim Kontrolltier sthnmte die Phasenverteilung mit Ausnahme der
\ J~ IX i ! \ 1 "</ I \ 1 \ I \ 1 JO
1o i i o 1-7-7- 1
0 2 ~ q 6 8 10 r "/~ r ~8 go g,Z gr
Abb. t7. Gesamtzahl der Mitosen in beiden Hornh~uten zu versehfedenen Zel tpunkten nach einer F~rbung m i t Bril lantkresylblau. Abszisse : Zei tpunkt der Fixierung in Stunden bzw. Tagen yore Beginn der F~rbung an gerechnet). Ordinate: Zahl der ~Iito~en ',in e inem Hornhau tpaa r .
bereits mehrfach erw~hnten geringeren Werte yon ,a' mit den yon KORNFELD ermitteltmn Durchschnlttszahlen iiberein. Nach 2 Stunden (Abb. 18) hat die Zahl der Spireme abgenommen, noch mehr jene der
6"O
5O
qO
jo
2o
o. J ,z b c aT. e , f .q
Abb t8.
~ L
/ \
t t 1
0 a a e ~ e f g
Abb. i9.
Diaster, die Metakinesen iehlen, w~hrend zahlreiche Asteren vorhanden shad. Nach 4 Stunden (Abb. 19) finden sich nur mehr wenige Spireme, die Asteren sind in noch gr6Berer Zahl als beim frfiheren Versuchstier
74 G. Politzer:
Fixier t nach
0 h
2h
4 h
6 h
8 h
10 ~ i
12h !
14h I
18 h
20 h
22 h
24 h
2a
3d
Horn- h a u t
a
b &
b a
b
, b
a
b a
b a
b a
b a
b a
b
b a
b a
b a
b &
b
Zahl der
I c ] d ~ b I , i
9 3j lS 15 16
0 47
0 0 27[ 2i 0 23i
, 0 1 4 1
0 i 1 [47 O, 0 4 0
i 0 30 0135 1!11
11 1121
, 1 1 6 3111 4 1 1 1
Tabelle 1.
Phasen Teil-
e i f g l s u m m e i
8 7 54
5 39
oool 1 39
0 0 0 31 0 0 0 33
0 0 0 19 0 0 0 23
0 0 0 0 43 0 0 0 0 36 0 0 0 0 31 0 0 0 0 23
0 0 0 0 33 0 0 0 0 47
0 0 0 0 27 0 0 0 0 I 25
i
0 0 0 0 ~ 37 0 0 0 0 41
0 0 0 0 48 1 0 0 0 41
1 0 1 0] 32 3 1 0 01 39
6 6 14 91 47 5 5 13 10! 56
0 2 2 3 !
4 9 47 0 6 5! 42
S u m m e
102
68
78
64
42
79
54
80
52
78
89
71
103
50
89
i
Anmerkungen
2 Mitosen nicht bes t immbar .
3Mitosen n ich t b e s t i m m b a r . 1 Mitose n ich t b e s t i m m b a r .
Die 2 Spi reme sind Anfangs- s tadien (A).
I n einer Bindegewebszel le e i n Diaster .
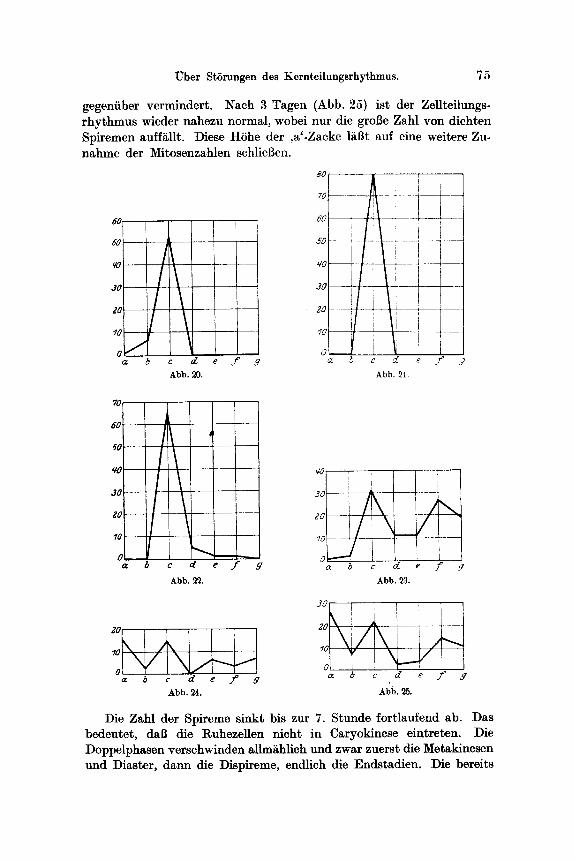
(Abb. 18) vertreten. Metakinesen und Diaster fehlen, vorhanden sind ferner ein Dispirem und 5 Endsta~lien. Nach 6 Stunden (Abb. 20) sind die Spireme schon sehr sp~rlich, die Asteren vermehrt , Doppelphasen fehlen. Von der 7.--23. Stunde sind nahezu ausschliei~lich Asteren vorhanden. Ein Beispiel der Phasenverteflung in einem solchen Horn- hautpaare ist in der Abb. 21 wiedergegeben. Die 80 Mitosen befinden sich durchwegs im Asterstadium. Naeh 22 Stunden t re ten neuerdings einzelne Doppelphasen auf (Abb. 22). Nach 24 Stunden (Abb. 23) er- scheinen wieder Spireme, die Asterenzacke ist erniedrigt, wit l inden zahlreiche Doppelphasen. Nach 2 Tagen (Abb. 24) ist die Phasen- verteflung normal, nur die Gesamtzahl der Teflungen ist der N o r m
~ber StSrungen des Kernteilungsrhythmus. 7 5
gegeniiber vermindert. Nach 3 Tagen (Abb. 25) ist der ZeUteilungs- rhythmus wieder nahezu normal, wobei nur die groi3e Zahl yon dichten Spiremen aufft~Ut. Diese H6he der ,a'-Zacke ltii3t auf eine weitere Zu- nahme der Mitosenzahlen schliel~en.
50
.o / \
r
,z b c d e
Abb. 20.
8O
70
80
SO
~0
30
2O
~0
0 CL b d
Abb. '2.1
f g J e 3"
7C
56
TO
qO
30
gO
I0
/\
b c d e
Abb. ~2.
f g
J
26
75 i - - - i I
o - - - - - I i ~z b c oL e f g
Abb. 23.
\ J 0
b c d e 2" 9
Abb. ~24.
3 0
~z b c d e f oa
Abb. 25.
Die Zahl der Spireme sinkt bis zur 7. Stunde fortlaufend ab. Das bedeutet, dal3 die Ruhezellen nicht in Caryokinese eintreten. Die Doppelphasen verschwinden allm~hlich und zwar zuerst die Metakinesen und Diaster, dann die Dispireme, endlich die Endstadien. Die bereits

7 6 G. Politzer:
fiber den Aster hinaus vorgeschrittenen Caryokinesen werden somit zu Ende gefiihrt. Die Asteren sind vermehrt. Dies ist darauf zuriiek- zuffihren, dal3 die Asteren sich nieht weiber entwiekel~n und dab die zur Zeit des F~rbungsbeginnes im Spirem befindlichen Mitosen in Asteren umgewandelt wurden. Diese Asterer~ gehen nun keineswegs zugrunde, da sonst die Gesamtzahl der Caryokinesen abnehmen miiBte. Die Wiederherstellung des normalen Kernteilungsrhythmus effolgt nun dadurch, da~ die Asteren sieh welter entwickeln und neuerdings Ruhe- zellen in das Sloirem eintreten. Hierbei sind in verschiedenen Reihen kleine Unterschiede erkennbar. Manchmal linden wir bereits Doppel- phasen, bevor noch Spireme auftreten (Abb. 23), manchmal geht die Bildun.g yon Spiremen der Fortffihrung der Asteren um 1--2 Stunden voraus ~ ).
Brill.antkresylblau und Auramin hemmen den Zellteilungsablau] in zwei Stadien, ngmlich in der Zeit zwischen Ruhestadium und Spirem, sowie zwischen Aster und l]letakinese. Nun wird uns aueh das Aus- sehen der Summenkurve (Abb. 17) verst~ndlieh. Der Abfall in den ersten 2 Versuchsstunden ist darauf zuriiekzufiihren, dal3 zahlreiche Doppelphasen zu Ruhezellen wurden, ohne dal~ ein neuer Nachschub yon Spiremen erfolgte.
D. Der Ein/lufl der ~Hitze au/ der~ Kernteilungsrhythmus.
Ein vierter Typus der St5rung des Kernteilungsrhythmus wurde yon WASSE~mAW.W besehrieben. W A S S E R ~ setzte Zwiebelwurzeln dutch 15--60 Minuten der Temperatur yon 36--37 ~ aus. Die zu verschiedenen Zeiten fixierten Wurzeln warden geschnitten und gef~rbt und dann die Zahl der Caryokinesen auf je 1000 Zellen bestimmt. Hierbei warden drei Phasen bei der Z~hlung unterschieden: Die Prophase bis zur AuflSsung der Kernmembran (,a' KOR~FELD, ,A' ,B' P~TW~), die Mesophase bis zum Zusammensehlu• der Toehterehromosomen (,b' bis ,e' KORI~FELD, ,C' bis ,G' PETER) und die Telophase (,f',g' KORN- rELD, ,H' ,I' P~.T~R). Ich habe nun die aus den Tabellen der WASS~.R- MA~schen Arbeit ersichtlichen Werte graphisch dargesteUt. Hierbei wurden als Abszissen die Phasen (P, M, T) als Ordinaten die Anzahl der entsprechenden Phasen in 1000 Zellen eingetragen. Waren mehrere Pr~parate zur gleichen Zeit fixiert worden, so wurde das arithmetisehe Mittel der Z~hlungen bestimmt. Die reehts oben yon jeder Kurve in Klammer stehende Ziffer gibt die Anzahl der Praparate an. Links oben
1) Der zweite Typus, der hier geschildert wurde (Erscheinen der Spireme vor der Fortfiihrung der Asteren), war merkwiirdigerweise bei meinen Auramin- reihen die Regel. Trotzdem glaube ich dieses gegens~tzliche Verhalten zwischen Auramin und Brillantkresylblauwirkung nur als Zufallsbefund auffassen zu miissen, da die Zahl der Versuchsreihen eine zu geringe ist.
Uber St6rungen des Kernteilungsrhythmus. 77
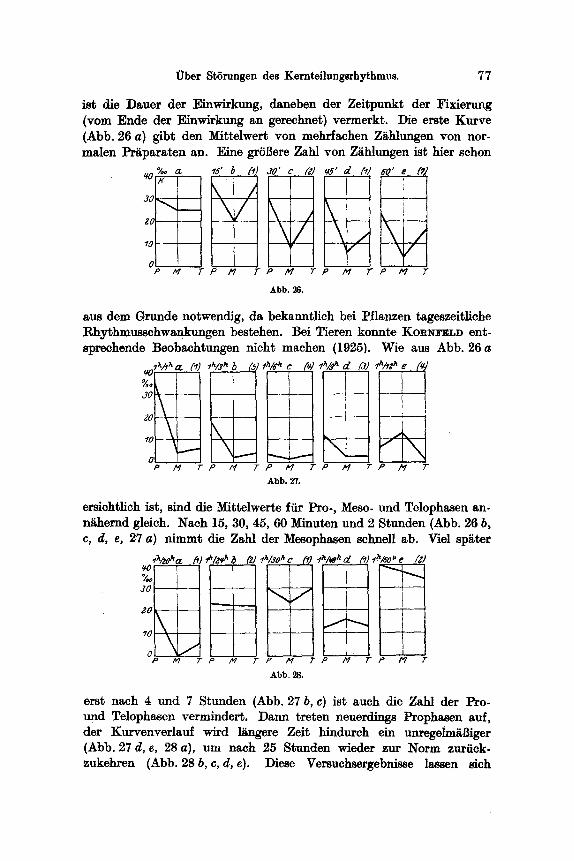
ist die Dsuer der Einwirkung, daneben der Zeitpunkt der Fixierung (yore Ende der Einwirkung an gerechnet) vermerkt. Die erste Kurve (Abb. 26 a) gibt den Mittelwert yon mehrfachen Ziihlungen yon nor- malen Pr~iparaten an. Eine grSflere Zahl yon ZRhlungen ist bier schon
%o ,z qs' b (z) ~" d fql 60' e
i " \ / \ , !
, o \ /
0. ,o ,v I T P It/ T P PI 7- p M T P PI T
A b b . 26.
sus dem Grunde notwendig, da bekanntlich bei Pflanzen tageszeitliche Rhythmusschwankungen bestehen. Bei Tieren konnte KORNFELD ent- sprechende Beobachtungen nicht machen (1925}.
30 \
O . . . P
Wie aus Abb. 26 a
P / T P PI T P Iv/ T P PI 7" p ~ I T
Abb. 27 .
ersichtlich ist, sind die Mittelwerte ffir Pro-, Meso- und Telophasen an- n~hernd gleich. Nach 15, 30, 45, 60 Minuten und 2 Stunden (Abb. 26 b, c, d, e, 27 a) nimmt die Zahl der Mesophasen schnell ab. Viel sparer
r"lz~ h b (e) ~alJoh c .P,l~ h e~ (q) r e (Z/
2 0 ~ I I
o / P I't/ T P PI T P M T P M r P ~ T
A b b . 28.
erst nach 4 und 7 Stunden (Abb. 27 b, c) ist such die Zahl der Pro- und Telophasen vermindert. ])ann treten neuerdlngs Prophasen auf, der Kurvenverlauf wird l~ngere Zeit hindurch ein unregelm~Biger (Abb. 27 d, e, 28 a), um nach 25 Stunden wieder zur Norm zuriick- zukehren (Abb. 28 b, c, d, e). Diese Versuchsergebnisse lassen sich

78 G. Politzer:
folgendermaBen deuten. Die auffallend geringe Zahl der Mesophasen ist darauf zuriickzufiihren, dab sich die Prophasen nicht zu Asteren weiterentwiekeln und dab die zur Zeit der Erw~rmung vorhandenen Mesophasen in Telophasen umgewandelt werden. Die Telophasen laufen ab, anscheinend jedoeh etwas verlangsamt. Das sphere Ver- schwinden der Prophasen fiihrt nun WASSE~A~ auf eine Riick- bildung zu Ruhezellen zuriick. Hierauf wird sp~ter noeh eingegangen werden.
Die Hernmung des ZelUeilungsablau/es 8etz~ also wie beim BriUant- kresylblautypus auch bei der W~rmewirkung in zwei Stadien ein. So wird der Eintritt der Ruhezdlen in die Prophase verhindert; w~hrend jedoch das zweite Stadium der Kernteflungshemmung beim Brillant- kresylblauversuche zwischen Aster und Metakinese (,c' und ,d') liegt, ist es bei den WAss~R~r~sehen W~rmeversuchen zwischen Pro- und MesoThase (also etwa zwischen ,a' und ,b') zu verlegen.
III. Allgemeiner Tefl.
A. Die Wachstumsgeschwindigkeit und ihre Koml~onenten.
Das Wachstum effolgt dutch Zellwachstum (,,Streckung", GUR- W~TSCH) und dureh Zellvermehrung. Die Geschwindigkeit der Zell- vermehrung h~ngt ab yon der Zahl der jeweils vorhandenen Caryo- kinesen (,,caryokinetischer Index", DE I~AB~S et FOReSTrY) und yon der Geschwindigkeit ihres Ablaufes. W~hrend wit den caryokinetisehen Index unschwer Ieststellen kSnnen, ist die Ablaufsdauer der Mitosen schwer bestimmbar. Die Beobachtung der Caryokinese am lebenden Objekt ist bei hSheren Tieren schwierig. Man ist daher beider Be- stimmung der Ablaufsdauer auf indirekte Methoden angewiesen, die naturgem~] zahlreiche Fehlerquellen in sieh schlieBen. Die Bereclmung der Ablaufsdauer nach P~TER wurde bereits erw~hnt. Sio hat nur einen bedingten Wert, da die Zahl der direkten Messungen, auf Grund weleher die Ablaufsdauer der Caryokinese in Gewebszellen ermittelt wurde, nur goring ist. Sie basiert ferner auf der Annahme, dab die Zellteflungs- dauer in allen Geweben die gleiche ist. Die ermittelte Wachstumsbe. selfleunigung (Q10 - 2) ist in den Versuehen HF~RTWIGS und PETE~ aussehlie]lieh auf eine Beschleunigung der Zellteilung bezogen worden, obzwar doeh eine Beschleunigung des Teilungstempos sowohl dutch die Vermehrung als auch durch die Beschleunigung der Caryokinesen mSglich ist. Eine zweite Annahme PETV.RS ist darum, dab der caryokine- tisehe Index der Gewebe unabh~ngig yon der Temperatur ist. Hierfiir gibt es jedoeh keine Belege. Ich habe bereits hervorgehoben, dab ich fiber --bisher allerdings nieht geniigend viele - - Befunde verfiige, die eine ZeUteilungsanregung dureh W~rme wahrseheinlich machen, die

~ber StSrungen des Kernteilungsrhythmus. 79
also gegen eine Unabh~ngigkeit des Caryokinesenindex vonder Tem- peratur sprechen. Trotzdem glaube ich, dai~ die P~.TEP~schen Werte fiir die Besehleunigung der Caryokinese keine zu hohen sind. Denn P~.Tv.R bestimmt den Wert von Qlo = 2, obwohl seine Beobachtungen an Echinus einen Weft yon 2,13 an Sphaerechinus 2,15~ bei Rana/usca sogar yon 2,87 ergaben. Es ist nun wohl mSglich, dab die willkiirliche Herabsetzung des Q10 ein Ansteigen des caryokinetischen Index aus- zugleiehen vermag. Aus diesen Erw~gungen ergibt sich, dal~ die PET~.R- sche Methode nicht zur Ermittlung genauer Werte fiihren kann und dab ferner der earyokinetische Index allein, der h~ufig zur Kennzeich- nung der Waehstumsgesehwindigkeit z. B. yon Carcinomen herangezogen wird, nut eine Komponente der Wachstumsgeschwindigkeit angibt. Da nun insbesondere in bSsartigen Geschwiilsten alle erdenklichen abnormen Caryokinesen anzutreffen sind, diirfen wir wolff auch StS- rungen des Teilungsrhythmus annehmen. Wenn wit uns nun vergegen. w~rtigen, wie wenig die Summenkurven (z. B. Abb. 17) aussagen, wenn Rhythmusst6rungen vorhanden sind, dann wird uns die geringe theo- retische Bedeutung des caryokinetischen Index klar. Seine praktiselle Bedeutung hat ja Rv.GAUD bereits mit Recht in Abrede gestellt.
Ein anderer Weg zur Bestimmung der Ablaufsdauer besteht darin, dab wir dutch irgendeine Sch~digung den Eintritt neuer Ruhezellen ins Spirem verhindern. Dann l~Bt sieh sowohl bestimmen, wann die letzten Spireme auftreten, als aueh warm die letzten Phasen iiberhaupt versehwinden. Die Zeitspanne zwisehen diesen Beobachtungen ent. spricht der Dauer des Zellteflungsablaufes. Ein Beispiel m5ge diese Ausfiihr~angen erg~nzen. Wird der Hoden eines S~ugetieres mit einer nicht allzuhohen RSntgendosis bestrahlt, so schwindet in typiseher Weise nahezu das gesamte Samenepithel, so dab zwisehen den erhaltenen SERTOLIschen Zellen nut wenige Spermiogonien zuriickbleiben. Dieser Vorgang wurde yon einem seiner Beobachter uls ,,ddpopulation" (R~- GAUD) bezeiehnet. SCHINZ und SLOTOPOLSKY unterscheiden acht Sta- dien dieses Vorganges, welehe in Tabelle 2 wiedergegeben sind. Aus der Tabelle ist ersichtlich, dal~ dureh die Bestrahlung unmittelbar nur die Spermatogonien gesch~digt wurden. Sie gehen zum Teil zugrunde, zum Teil biiBen sie ihre Teilungsfhhigkeit voriibergehend ein. Die iibrigen Zellen des Samenepithels teilen sich fort. I)~ die reifen Sper- mien in die Lichtung der Kan~lehen ausgestoi3en werden und den Hoden verlassen, kommt es zur fortschreitenden Leerung der Kan~l- chen. Fiir diese acht Stadien bringen SCHINZ und SLOTOPOLSKY iiber- zeugende Abbildungen. Es lag nun nahe die Zeitspanne zwisehen Bestrahlung und vollkommenen Verschwindens der Spermiocyten, Sper- miden und Spermien der Dauer der normalen Spermiogenese gleichzu- setzen. ~hnliehe Versuchsergebnisse wurden auch in der tt~matologie

8 0 G. Politzer:
Stadium der $ch~tdigung
I.
II.
HI.
IV.
V.
VI.
VIL
VIII.
Sertolische Zellen
+
+
+
+
+
§
§
§
Tabelle 21).
Spermlogonien
+ m germger Zahl
+ m germger Zahl
+ m germger Zahl
+ m germger Zahl
+ m germger Zahl
+ m germger Zahl
+ m germger Zahl
+ m geringer Zahl
Spermiocyten
A-
A-
in T: lun
Spermiden [ Spermien
+ I : -
+ + - ]
i I
zur Bestimmung der Lebensdauer der weiBen BlutkSrperchen ange- wendet. Bereits SCHI~Z und SLOTOPOLSKY warnten aber vor einer der- artigen Verwertung ihrer Versuchsergebnisse. Wie berechtigt diese Warnung war, geht klar aus dem in dieser Arbeit erbrachten Nachweis hervor, dab die RSntgenstrahlen - - selbst unter Beriicksichtigung der Fehlerquellen der P~.T~P~chen Berechnungen - - zweifellos den Ablauf der bestrahlten Caryokinesen verlangsamen. Es wird bei der erwi~hnten Methode nicht die Dauer der normalen Spermiogenese, sondern jene der Spermiogenese nach RSntgenbestrahlung berechnet, die ebenso verlangsamt sein kSnnte, wie die Zellteflungen bei meinen Versuchen. Aus diesen Untersuchungen ging zwar hervor, da~ die Verlangsamung anscheinend eine gesetzm/~Bige ist; so dauert der Prhn~reffekt doppelt so lang wie eine normale Caryokinese. Doch miiBte diese Beobachtung erst an einem grSBeren Materiale besti~tigt werden. Wichtig w~re es, die Dauer des Prim/~reffektes in verschiedenen Geweben des gleichen Tieres zu untersuchen. Erg~ben sich hierbei Unterschiede, so wiirden sie fiir eine verschiedene Dauer der Caryokinese in verschiedenen Ge- weben sprechen.
B. •ber unvollstSndige Teilungsvorggnge. Aus zweien unserer Kurven (Abb. 4, 13 ) l~B t sich eine Tatsache
ersehen, welche auch in mehreren bier nicht abgebildeten F~llen deut-
1) Erkl~rung zu Tabelle 2: Acht Stadien der Wirkung der R~ntgenstrahlen auf das Samenepi~hel des S~ugetierhodens (nach ScHI~z und Sr,OTOPOLS]KY). Die Pfeile zeigen die Umwandlung einer Zellart in eine andere an.

Uber St6rungen des Kernteflungsrhythmus. 81
lich in Erscheinung trat. Es finden sick ngmlich noch vereinzelte Stadien ,a' vor, obwohl bereits lockere Spireme und oft auch die Asteren geschwunden sind. Da die Phasen ,b' und ,c' auch in spgter fixierten Pr~paraten der Versuchsreihen nicht mehr auftreten, muB man an- nehmen, dab diese friihen Spireme sich nicht weiterentwickeln. Da sie ferner bei spgter fixierten Tieren nicht mehr vorhanden sind und An- zeichen ffir ein Zugrundegehen dieser Zellen fehlen, ist nur die An- nahme m6glich, dab sich diese ZeUen in Ruhezellen umgewandelt haben. Betrachten wir diese Spireme ngher, so f/~llt uns auf, dab sie durchwegs dem friihesten Stadium des dichten Spirems entsprechen, dem Vorberei- tungs- oder An~angsstadium A nach PETER. Zur Erklgrung dieser Umwandlung kann man wohl kaum annehmen, dab die physikalisch- chemischen Vorggnge, welche der Umbildung der RuhezeUen in Spireme zugrunde liegen, an sich umkehrbar sind. Im normalen Zelleben gehen nun Vorggnge vor sich, die dieser ,,R/ickbildung" vergleichbar sind und zwar bei der Umbildung yon Telophasen in Ruhezellen. Wir werden daher diese Umwandlung besser als abortiven Verlauf auffassen, indem bei dieser ,,Teilung" die Meta- und Anaphasen ausgefallen sind. Diese eigenartige Hemmung der Caryokinese wurde bereits yon friiheren Untersuchern erw~hnt. WASSERMAN~ deutet in/~hnlicher Weise seine in den Kurven Abb. 26--28 dargestellten Versuchsergebnlsse. Er meint, da6 die langsam erfolgende Abnahme der Prophasen nicht auf einer Weiterbildung zu Metaphasen beruhe - - sonst miiBte ja die Mesophasen- zahl ansteigen - - sondern nimmt an, dab sich die Spireme zu Ruhezeilen umbilden. LYDIA G U R W I T S C H 1) berichtet fiber den EinfluB der Chloral- narkose auf die Zellteilung in Zwiebelwurzeln. Bereits 20 Minuten nach Eintauehen der Wurzelspitzen in eine ChloraU6sung macht sieh ein waehstumshemmender EinfluB bemerkbar. Nach 2 Stunden ist das Wachstum nur mehr sehr gering, bleibt jedoch bis zu 10--12 Stunden naehweisbar. Dann ist vollkommener WachstumsstiUstand eingetreten. Die mikroskopische Untersuchung ergab: ,,Eine gewisse Alteration der achromatischen Figur macht sich schon nach etwa 30 Minuten geltend. Es kommt nun innerhalb der ersten Stunden zu einer v611igen Verun- staltung auch der ehromatischen Figur, namentlich der Metaphase und Anaphase, wo die Chromosomen zu formlosen Haufen verklumpen. Spireme erwcisen sich am stabilsten. Nach etwa 15 Stunden sind keine deutliehen Residuen yon Mitosen mehr erkennbar. Es kann wohl kaum bezweifelt werden, dab es sich nicht um wirklich abgelaufene Zell- teilungen handelt. Es sieht vielmehr danach aus, als ob die mitotischen Figuren eine Art regressive Metamorphose durchgemacht hgtten, die
1) LYDIA GURWITSCH: Untersuchungen fiber die mitogenetischen Strahlen. Arch. f. Entwicklungsmech. 103. 1924.
Z. f. Zellforschung u. mikr. Anatomie Bd. 3. 6 a

82 G. Politzer:
auf den Ruhestand der Kerne ffihren." Diese abgekfirzten Teflungs- vorg~nge scheinen auch in Carcinomen h~ufig aufzutreten.
PALUGYAY berichtet fiber eine auffallende H~ufung von Prophasen in manehen Carcinomformen. So sind im Uterusearcinom niederer Reifea) 58vH. der ZeUen Ruhezellen, 40vH. Prophasen und nur 1,5 vH. Mitosen. (PALuGYAY sollte richtiger ,,die fibrigen Phasen der Caryokinese" sagen, da wir ja die Prophase gleichfalls zur Caryokinese rechnen.) PALVGYAu sucht diese H~ufigkeit der Prophasen zu erkl~ren. Es kSnnte die Prophase besonders lange Zeit in Anspruch nehmen. Dies erscheint ihm jedoch unwahrseheinlich, da die Prophasen in Carcinomen mittlerer und hoher Reife seltener sind und man daher zur Annahme einer versehieden langen Dauer der Prophasen bei ver- schiedenem Reifegrad des Carcinoms gezwungen w~re. Eine zweite MSglichkeit ist nach PALUGYAY darin gegeben, dab die Zellen nach Abschlul3 der Teilung nicht immer in ein Ruhestadium eintreten, viel- mehr manche gleich in das Stadium der Prophase iibergehen und so bis zur n~chsten Teilung verharren. Dann f~hrt PALUGYAy fort: ,,Viel wahrscheinlicher erscheint mir die Ursache ffir die tiberwiegende Zahl der Prophasenformen in Carcinomen niederer Reife darin zu liegen, daB es bei der rasehen iiberstiirzten Zellteilung in diesen Carcinomen, in vielen F~llen gar nieht bis zum Stadium der Mitose kommt, sondern daI~ es sich in diesen F~tlen um einen abgekiirzten Teilungsvorgang handelt, bei welchem sich die Kernteilungsvorg~nge innerhalb der nicht zur LSsung gelangten Kernmembran abspielen." PAL~G~AY bringt nun Abbildungen, die veranschauliehen sollen, dab innerhalb der ge- schlossenen Kernmembrap~ Chromosomen auftreten, die sieh in mannig- faltiger Weise anordnen und Asteren und Diaster bilden. Die Caryo- kinese innerhalb der geschlossenen Kernmembran ist nun hSehst un- wahrscheinlieh. Es wirken ja bei der mitotischen Teilung auch Zellteile mit, die im Plasma liegen wie z. B. die Centrosomen. Diese mfiBten bei den von PXLVC~u besehriebenen Teilungen im Kerne neugebildet werden oder gar die Kernmembran durehwandern. Es ist .~ueh un- wahrseheinlieh, dab viele dieser Prophasen iiberhaupt zu einer Teilung des Kernes oder der ZeUe ffihren. Wenn wir eine ZeUteilungsdauer yon einer halben Stunde, wie sie P~T~R fiir S~ugetiere bereehnet und einen earyokinetisehen Index von 41x/2 vH. annehmen, dann miiBte man ja die Careinome mit freiem Auge waehsen sehen! lch glaube vielmehr, da~ die meisten Prophasen in Carcinoraen niederer Rei]e, wie sie PALUOYAY untersuehte, gar nicht zur Teilung ]iihren, sondern wieder zu Ruhezellen werden.
1) FRANKL unterscheidet bei Uteruscarcinomen je nach dem Differen- zierungsgrad der ZeUen Carcinome hoher, mittlerer und niederer Reife.

Uber St~rungen des Kernteilungsrhythmus. 83
C. ~ber die sogenannte unspecl]iache Wirlcung der RSntilenstrahlen. Je eingehender in jiingster geit die RSntgeneffekte untersueht wur-
den, desto deutlicher traten die Unterschiede in der Wirkung der R6ntgenstrahlen und der Wirkung anderer Reizmittel zutage. Vor kurzem wurde nun der Versueh gemaeht, eine specifische Wirkung der R6ntgenstrahlen zu leugnen und die Verschiedenheiten in der Wirkungs- weise der R6ntgenstrahlen und anderer Reizmittel einzig auf deren verschiedene physikalische Eigenschaften zu beziehen.
ScmNz und SLOTOPOLSKY ste]]ten, wie bereits frfiher erwghnt wurde, Untersuchungen fiber den EinfluB der R6ntgenstrahlen auf das Hoden- gewebe an. Bei Verwendung sehr starker Dosen wurde das gesamte Hodenepithel necrotisch. Mittlere bzw. kleine Dosen deckten eine ver- schiedene Empfindlichkeit der einzelnen Elemente des Hodens auf. Kurz nach der Bestrahlung ist nur eine Verminderung der Zahl der Spermio- gonien naehweisbar. Dann versehwinden die Spermioeyten, hierauf die Spermiden, endlich die Spermien, so dab die geleerten Hodenkan~Ichen nur mehr SERTOLISche ZeUen und vereinzelte Spermiogonien enthalten. SCHINZ und SLOTOPOLSKY erklgren den Vorgang in folgender Weise: Dureh die Bestrahhmg werden vorwiegend nut die Spermiogonien ge- sch~tigt; ein Teil yon ihnen geht zugrunde, ein anderer verliert vor- fibergehend oder dauernd seine Teilungsfghigkeit. Die Spermiocyten hingegen vollenden die Spermiogenese. I)a ein Naehschub yon Spermio- eyten infolge der Teilungshemmung der Spermiogonien unterbleibt, kommt es zur fortsehreitenden Leerung der Hodenkanglchen.
Grunds~tzlich anders sind die Vergnderungen bei der Hodenatrophie nach Verlagerung des Organes, nach Vasectomie und bei Ern~hrungs- st6rungen. Hier sehwinden zuerst die Spermien, hierauf die Spermiden, dann die Spermiocyten und zuletzt erst die Spermiogonien. Es scheint also, dab die Spermiogonien im Gegensatz zu ihrer hohen R6ntgen- empfindliehkeit bei der ,,gew6hnliehen Hodenatrophie" (ScHINZ) am widerstandsfghigsten sind. Bedenkt man jedoch, da6 die Spermiogonien den die Hodenkan~lchen umspinnenden Gef~flen am ngchsten liegen, so ist es klar, dab die Spermiogonien aus diesem Grunde und nicht, weft sie widerstandsfghiger sind als die anderen Zellen der Hoden- kanglchen, zuletzt gesch~dig~ werden. Bei der R6ntgenwirkung ist die Verteilung des Reizmittels eine so gleichmgBige, dab die Empfindlich- keitsuntersehiede der einzelnen Zellen zutage treten.
SCHI~Z geht jedoeh in der Deutung seiner Versuehe noeh weiter. Die R6ntgenstrahlen zeigen nicht nur die wahre R6ntgenempfindlichkeit, sondern die wahre aUgemeine Empfindliehkeit auf, da ,,aUgemeine Sen. sibili~t und RSntqensenaibilitg~ lebender Strulaur sich decken". Diese Annahme sucht nun Scr~INZ dutch eine Reihe yon Angaben aus der Literatur zu stfitzen, die fibereinstimmende Vergnderung yon Zellen
6*

84 G. Politzer:
und G eweben nach R6ntgenbestrahlmtg und andersartigen Sehiixiigungen betreffen.
Unsere Versuche ergaben Unterschiede in der Wirkungsweise der R6ntgenstrahlen und anderer Reizmittel, die nochmals erw~hnt werden sollen. Es wird sich auch hierbei empfehlen, die abnorme~ Caryokinesen und die Rhythmusst6rungen gesondert zu besprechen. Abnorme Mitosen finden sich nach RSntgenbestrahlung sowohl im Primer- als auch im Secund~reffekt. Der Prim~reffekt ist gekenn- zeichnet durch Pseudoamitosen und dutch Pyknose der Asteren, der Seeund~reffekt dutch Rhexis und Versfreuung der Chromosomen und ~hnliche Abnormit~en. Diese Ver~nderungen an sich sind fiir das angewendete Reizmittel nicht kennzeiehnend: Pyknose der Asteren ist ein h~ufiges Vorkommnis und wurde bereits bei zahlreichen Sch~di- gungen beobachtet (GAL~OTTI, KORNFELD 1925), Pseudoamitosen wur- den bei Tieren (HAECKER, BURY, SCHILLER, ALVERDES) und bei Pflanzen (NATHANSON, NEMEC, WASIELEWSKY, MAINX} mitunter gefunden. Ihr regelmitl~iges Vorkommen nach Nilblau- oder Neutralrotf~bung babe ich vor kurzem beschrieben. Diese Versuche ergaben auch, dab die Pseudoamitosen nach R6ntgenbestrahlung und Vitalf~rbung (Neutral- rot, Nilblau) in gleicher Weise entstehen und auch gleiehe wohl gekenn- zeiehnete Ablaufsformen zeigen. Noch h~ufiger wurden Chromosomen ablenknng, Briickenbildung, Deviation der Toehtersterne und die anderen den Secund~reffekt kennzeichnenden Teilungsanomalien be- obachtet. Die h~ufige Wiederkehr ~hnlicher Ver~nderungen der Caryo- kinese erweckt den Anschein, als ob der Zelle nur eine beschr~nkte Anzahl yon I~eaktionsmSglichkeiten zur Verfiigung stehe. Trotzdem sind fiir jedes Reizmittel kennzeichnende Wirkungsweisen m6glich, da die eine oder andere yon diesen StSrungsformen in Erseheinung treten kann. Ziehen wit ferner in Betracht, dab auch die einzelnen Typen der abnormen Mitosen zeitlich verschieden auftreten kSnnen, so ist die Zatd der Kombinationen eine sehr groBe. Bei Beriicksichtigung des ge- samten Ablaufs der Sch~digung l~l~t sich nun feststellen, dab nut hin- siehtheh weniger Punkte eine ~bereinstimmung zwischen der RSntgen- und Neutralrotwirkung besteht. Die R6ntgenstrahlen ffihren zur Pyknose der Asteren, die Neutralrotvergiftung nie. Gewisse Ablaufs- fomen der Pseudoamitose, wie sie sieh naeh Neutralrotft~rbung finden, treten nach RSntgenbestrahlung nicht oder nur sehr selten auf. Vor a.llem jedoch ist nach Neutralrot/Srbung nlemal8 ein SecundSre/[e~ nach- weisbar. Die naeh der mitosenfreien Zwisehenzeit erscheinenden Caryo- kinesen sind vSllig normal.
Versucht man die Kurzfristigkeit der Neutralrotwirkung zu erkl~ren, so l ~ t sich vor allem eine GewShnung der Zelle ausschliel~en, da nach einer neuerlichen F~rbung stets wieder Pseudoamitosen auftreten. Viel-

Uber St6rungen des Kernteilungsrhythmus. 85
leicht ist folgender Erkl~rungsversuch m6glich: Die F~rbung mit Neutralrot (oder Nflblau) ist zuerst eine Diffusf~rbung des gesamten Zellplasmas, dann erst geht aus ihr eine Granulaf~rbung hervor. Mit dieser ,,r~umlichen Isolation" des G~tes wird dieses fiir die Zelle un- wirksam. Wollten wir mit dieser Annahme auch den R6ntgensecund~r- effekt erkl~ren, mii~ten wir uns vorstellen, dab die R~ntgenstrahlen Giftstofie in der Zelle erzeugen, deren Beseitigung dem Plasma unmSg- lich ist. l~ese Annahme entspricht wold kaum unseren gegenw~rtigen Anschauungen iiber die Wirkungsweise der R6nt~enstrahlen.
Ein anderer Erkl~rungsversuch k~nnte yon der Tatsache ausgehen, da~ die Vitalfarbstoffe stets nur das Plasma, nie den Kern f~rbenl). Man k6nntc nun annehmen, da~ ein Secund~reffekt deshalb nicht auf- tritt, weft der Kern durch Vitalfarbstoffe wenig oder gar nicht ge. sch~digt wird, der Secund~reffekt jedoch ein Ausdruck der Kernschg~li- gung ist. Der Prim~reffekt muB nicht ausschlieBlich durch Plasma~ sch~figung bedingt sein, da ja bei den zur Zeit der F~rbung vorhandenen Meta- und Anaphasen die Kernmembran aufgel6st ist, daher eine Tren- nung zwischen Kern und Plasma nicht besteht. Hingegen mii~te die mitosenfreie Zwischenzeit auf eine Plasmasch~Jigung zuriickgeffihrt werden, da die Zellen, welche sp~terhin flare Teflungsf~higkeit einbii~en, im Zeitpunkt der Diffusf~rbung eine Kemmembran besaBen. Fiir die R6ntgeneffekte wiirde diese Annahme bedeuten, da~ die mitosenfreie Zwischenzeit und wahrscheinlich auch der Prim~reffekt die Folge einer P ~ c ~ d i ~ , der Secund~reffekt die Folge einer Kernsch~digung darste]lt. Auch dieses Ergebnis unserer ~berlegung entspricht wohl kaum unseren gegenw~rtigen Anschauungen iiber die Wirkungsweise der R6ntgenstrahlen. Nehmen wir hingegen an, dab die Giftwirkung des Neutralrotes in jedem Falle auch den Kern betrifft, dann ist die gegen- s~tzliche Wirkung der R6ntgenstrahlen und des Neutralrots ~ eine unspecifische Wirkung vorausgesetzt - - wieder nicht erkl~rbar. Die Annahme, da~ zwar die unmittelbare Wirkung verschiedener Sch~i~ gungen die gleiche und nur die Erholungsfghigkeit der Zelle eine ver- schiedene ist, ver|egt nur die specifische Wirkung auf einen sp~teren Zeitpunkt der Zellreaktion. Die ~Virkung des Brillantkresylblaus end- |ich ~hnelt weder der des Neutralrots noch jener der R~ntgenstrahlen.
Dem RSntgens~u~reffekte ghnliche Ver~nderungen werden auch dutch andere Reizmittel hervorgebracht. Doch rufen alle diese ~hnlich wirkenden Gifte diese formalen Anomalien der Caryokinese un~it~elbar nach ihrer Einwirkung hervor, nicht abet wie die R6ntgenwirkung ers~ Ta~e, Wochen, ~a ~elb~ Monat~ ~t~i~er. Diese Unterschiede in der Wir-
x) Damit ist nat~rlich nicht gesa~, dab die Vitalfarbstoffe nicht in den Kern eindr~gen; sie sind nut nicht innerha|b des Kernes sichtbar.
Z. f. ~llfor~chung u. m ~ r . Anat~mde Bd. 8. 6b

86 G. Politzer:
kung sind nun physikalisch unerkl~rlich, da vielen dieser Reizmittel (Vitalfarbstoffe, RSntgenstrahlen) eine starke Durchdringungsf~higkeit gemeinsam ist. Hierfiir spricht die Tatsache, dab nur die Caryokinesen in der tiefen Hornhautepithelschichte gesch~igt werden, w~hrend die Zellen der oberfl~chlichen Schichte unversehrt zu sein scheinen, eine Tatsache, deren Bedeutung bereits eingangs gewiirdigt wurde.
Noch gr6flere Verschiedenheiten als die formalen Veriinderungen der Caryokinese selbst zeigen die RhythmusstSrungen nach verschieden- artigen Sch~digungen. Beim Neutralrot wird nur der Eintritt der Ruhezellen in das Spirem verhindert. Bei der RSntgenwirkung tritt eine teilweise Hemmung des l~berganges der Asteren in die Metakinese hinzu. Diese ,,gehemmten" Asteren sterben ab und werden dann ab- gestogen. Es liel3e sich denken, dab die fiir Neutralrot kennzeichnende Wirkungsweise nur ein geringerer Grad der R6ntgensch~digung, also nur quantitativ verschieden sei. Diese Ansicht ist abzulehnen. Bei starker Verminderung der RSntgendosen ist zwar zuletzt gar keine Wirkung mehr nachzuweisen, eine der Neutralrotwirkung ~hnliche konnte ich bisher nicht erzielen. Doch reichen hierzu meine Versuche vielleicht nicht aus. Hingegen ergab die Steigerung der Nflblauwirkung 1) zwar bei starken Dosen Nekrose des gesamten Hornhautepithels, jedoch keine Pyknose der Asteren. Die Wirkung des Brillantkresylblaus (Hem- mung der Asteren ohne Nekrose) und die von WASSERMANN beschriebene Hitzewirkung (Hemmung der Spireme) kSnnen mit der RSntgenwirkung iiberhaupt nicht in Einklang gebracht werden. Wir sehen, daft auch die kritlsche Phase~) bei verschiedenen Reizmitteln eine verschiedene ist und miissen, da eine,,physikalische" Erkl~rung dieser Verschiedenheit derzeit nicht m6glich ist, entweder annehmen, dab alle bisher verwen- deten Reizmittel specifisch und nur die R6ntgenstrahlen unspecifisch wirken oder folgerichtig auch diesen eine specifische Wirkung zuerkennen.
Zusammenfassung.
Die Z~hlungen der Caryokinesen in verschieden beeinfluflten Horn- h~uten yon Urodelenlarven ergab:
1. Das Neutralrot hemmt den Eintritt der Ruhezellen in die Caryo- kinese. Die vorhandenen Zellteilungen laufen - - unter Auftreten yon Pseudoamitosen - - ab. Dadurch wird die Hornhaut voriibergehend mitosenfrei.
2. Der Einflul3 der R6ntgenstrahlen auf den Zellteilungsrhythmus ist folgender:
z) Nilblau und Neutralrot rufen die gleichen Ver~nderungen hervor. Nil- blau ist jedoeh glftiger.
2) Das heiBt das Stadium, in dem die Ver~nderungen in Erscheinung treten.
~ber St6rungen des Kernteilungsrhythmus. 87
a) Die Dauer des Prim~reffektes ist yon der Temperatur des Wassers, in welchem die Versuchstiere gehalten werden, abh~ngig.
b) Der Prim~reffekt dauert etwa doppelt so lange wie der norma|e Ab]atff einer Caryokinese bei der Temperatur des Versuches.
c) Die R6ntgenstrahlen bewirken eine Hemmung des Zellteilungs- vorganges: Die Ruhezellen werden am Eintr i t t in die Caryokinese ge- hindert. Die zur Zeit der Bestrahlung vorhandenen Zellteilungen werden zum groBen Teil, wenn auch wesentlich langsamer, als es der Tempe-
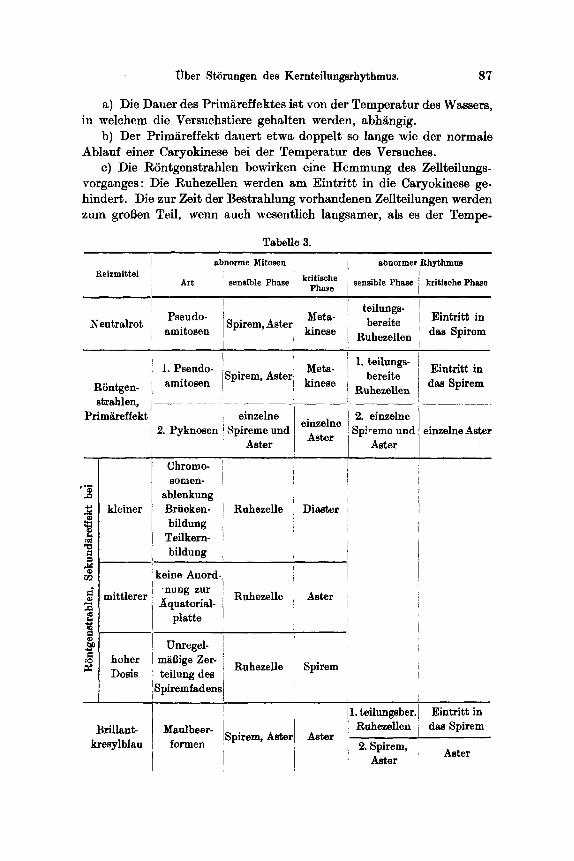
Tabelle 3.
R e i z m i t t e l
Neutralrot
RSntgen- strahlen,
Prim~reffekt
kleiner
mittlerer
A r t
Pseudo- amitosen
1. Pseudo- amitosen
2. Pyknosen
Chromo- somen-
ablenkung Briieken- bildung
Teilkern- bildung
keine Anord- 'nung zur Aquatorial-
platte
:~ hoher Unregel- m~13ige Zer-
Dosis teilung des Spiremfadensl
Brillant- kresylblau
abnorme Mitosen
sensible Phase
Spirem, Aster
Spirem, Astez
einze]ne Spireme und
Aster
Ruhezelle
Maulbeer- formen
RuhezeHe
I i
Ruhezelle
Aster Spirem,
i
k r i t i s c h e P h a ~ e
Meta- kinese
Meta- kinese
einzelne Aster
Diaster
Aster
Spirem
Aster
sensible Phase
ahnormer Rhythmus
kritische Phase
teilungs- bereite
Ruhezellen
1. teilungs- bereite
Ruhezellen
Eintritt in das Spirem
Eintritt in das Spirem
2. Spirem, Aster Aster
1. teilungsber. Ruhezellen
Eintritt in das Spirem
2. einzelne 1Sp],~eme und I einzelne Aster ! As ~ I
88 G. Politzer:
ra tur entspricht, zu Ende gefiihrt. Ein Tel| der Asteren wird jedoeh pyknotisch, die betreffenden Zellen gehen zugrunde und werden aus der Hornhaut ausgestol3en.
3. Brillantkresylblau hemmt den Zellteilungsablauf wghrend zweier Teilungsstadien, zwischen Ruhestadium u n d Spirem, sowie zwischen As~er und Metakinese.
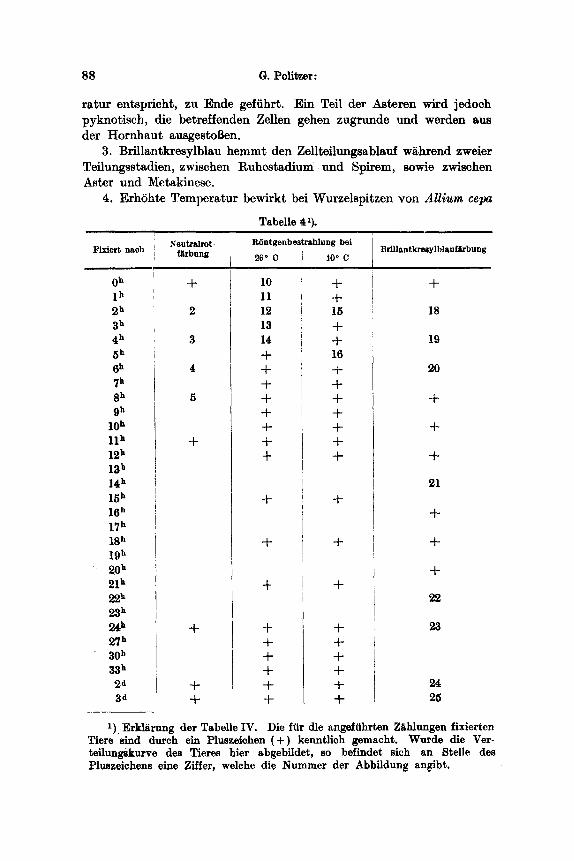
4. Erh6hte Temperatur bewirkt bei Wurzelspitzen yon AUium cepa
Tabelle 41).
R6ntgenbestrahlung bei Fixier t nach Brillantkresylblauf~bung
O h
2 h 3 h 4 h 5 h 6 h 7 h 8 h 9 h
l0 h 11 h 12 h 13 h 14 h 15 h 16 h 17 h 18 h 19 h 20 h 21 h 22 ~ 23 ~ 24h 27 h 30 h 33 h
2~ 3d
Neutralrot- farbung
+ 10 11
2 12 13
3 14 +
4 + +
5 + + §
+ + +
+
+
+
+ + + + +
+ +
~6 ~ C JO ~ C
+ 15 + + 16 + + + + + -p -t-
+
+
+
+ + + + § +
+
18
19
2O
+
+
+
21
+
+
§
22
23
24 25
1) Erklgrung der Tabelle IV. Die fiir die angeffihrten Zghlungen fixierten Tiere sind dutch eln Pluszeichen ( + ) kenntlich gemacht. Wurde die Ver- teilungskurve des Tieres hier abgebildet, so befindet sich an Stelle de~ Pluszeichens vine Ziffer, welche die Nummer der Abbildung angibt,

Ober St6rungen des Kernteilungsrhythmus. 89
gleichfalls eine Hemmung des Zellteilungsablaufes wAhrend zweier Sta- dien, zwischen Ruhestadium und Spirem, sowie zwischen Pro. lind Metaphase (Sehlui~folgerungen abgeleite~ aus den Versuchsergebnissen yon WASS~RMA~I~).
5. Die in der ,,sensiblen" Phase gesetzten Sch~digungen werden erst in der ,,kritischen" Phase erkennbar (Latenz). Die kritische und sensible Phase der Caryokinese fallen bei verschiedenen Reizmitteln in ver. schiedene Teilungsstadien. In der Tabelle III wurde der Einflull des Neutralrots, der RSntgenstrahlen und des Brillantkresylblaus auf die Zellteilungen selbst und auf den Zellteilungsrhythmus zusammengefaBt.
6. Bei der Einwirkung verschiedenartiger Reizmittel kommt es bei bereits vorgeschrittener Zellteilungshemmung mitunter zum Auftreten einzelner Anfangsformen der Caryokinese, die sich jedoeh nicht in Spireme umwandeln, sondern wieder zu Ruhestadien werden.
7. ScHINz fiihrt die verschiedenartige Wirkung der RSntgenstrahlen und anderer Reizmittel nur auf die verschiedenen physikalischen Be- dingungen zuriick, unter denen sie einwirken. Im Gegensatze hierzu wird auf Grund der in Tabelle I I I dargesteUten verschiedenartigen Wirkungsweisen verschiedener Reizmittel eine specifisehe Wirkung der Rfntgenstrahlen angenommen.
Literaturverze ichni s . Aibet~ W. u. Polltzer, G.: Uber den Einflufl der R6ntgcnstrahlcn auf die
Zellteilung. Arch. f. ]~ntwicklungsmech. d. Organlsmen 100. 1923 und 108. 1924. - - Mverdes, F.. Das Verhalten des Kernes der mit Radium behan- delten Spermatozoen yon Cyclops nach der Bcffuchtung. Arch. f. Entwicklungs- mech. d. Organismen 47. 1921. - - Bury, J.: Experimentelle Untersuchung fiber die Einwirkung der Temperatur 0 ~ suf die Entwicklung des Echinidencies. ArCh. f. Entwicklungsmech. 38. 1913. - - De Nsbtas, S.u. Forestier, J.: Note sur le traitement curieth6rapique des 6pith~liomas malpighiens. Cpt. rend. des s~ances de l& soc. de biol. 20. I. 1923. - - Flemming, W.: Zellsubstanz, Kern and Zell- teflung. 1882. - - Galeottl, G.: Ober experimcntelle Erzeugung yon Unregclmt~Big- keiten des caryokinetischen Prozesses. Zieglers Beitr. z. pathol. Anat. 14.1893. Haeeker, V.: Mitosen im Gefolge amitosen~hnlicher Vorg~nge. Anat. Anz. 17. 1900. - - Jolly: Zitiert naeh Peter. - - Jusellus: Experimentelle Untersuehungen iiber die Regeneration des Epithcls der Cornea unter normalen Verht~ltnissen und untcr therapeutischen MaBnahmcn. Arch. f. Ophthalmolog. 75. 1910. - - Kornfeld, W.: ~ber den Zellteilungsrhythmus und seine Regelung. Arch. f. Entwicklungsmech. 50. 1922. - - Ders.: Experimentelle Untersuchungen fiber StSrungen der Zellteilungst~tigkeit, Zellwandernngen, Pigmentverschiebungen und Epithelwucherungen bei Urodelenlarven. Zeitschr. f. Zellforsch. u. mikro- skop. Anat. 2. 1925. - - Malnx, F.: ~ber kiinstliche Beeinfiussung des Kern- teilungsvorgangs. Ber. d. dtsch, bot. Ges. 41. 1923. - - Nathanson, A.: Phy- siologisehe Untersuchungen fiber amitotische Kernteitung. Jahrb.f. wiss. Bot. 35. 1900. ~ Nemee: ]~ber die Einwirkung des Chloralhydrats au/die Kern- und Zcll- teilung. Jahrb. f. wiss. Bot. 39. 1904. - - Pslugyay, J.: Reifestadium des Carcinoms

90 G. Politzer: Uber StSrungen des Kernteilungsrhythmus.
und Zellteilung. Zeitsehr. f. Krebsforseh. 22. 1925. - - Peter, K.: Der Grad der Be- schleunigung tierischer Entwicklung durch erhShte Temperatur. Arch. f. Entwick- lungsmech, d. 0rganismen 20. 1905. - - Peter, K.: ~ber die Dauer der indirekten Kernteilung bei Amphibien. Zeitsehr. f. Morphol. u. Anthropol. 24. 1924. - - Ders.: t~ber Zellteilungsprobleme. Klin. Wochenschr. Jg. I I I , Nr. 48. 1924, - - Ders.: Zellteilung und Zellt~tigkeit. Zeitscbr. f. Anat. u. Entwieklungsgesch. 72. 1924 und 7~. 1925. - - Politzer, G. u. Alberti, W.: ~be r die Einwirkung des ultra- violetten Liehtes auf tierisches Gewebe. Zeitschr. f. Zellen- und Gewebslehre. I. 1924. - - Politzer, G.: Versuche fiber den Einflufl des Neutralrots auf die Zellteilung (Mitose, Amitose, Pseudoamitose). Zeitschr. f. Zellen- u. Gewebs- lehre 1. 1924. - - Ders.: Histologische Untersuchungen fiber die Giftwirkung der sogenannten Vitalfarbstoffe. Sitzungsber. d. Wien. zool.-bot. Ges. Jg. 1924 --25. - - Regauds zahlreiche Arbeiten siehe bei Schinz u. Slotopolsky, RSntgen- hoden usw. - - Schiller, J . : ~be r kfinstliehe Erzeugung primitiver Kernteilungs- figuren bei Cyclops. Arch, f. Entwieklungsmeeh. d. Organismen 27. 1909. - - Sehinz, H . R . : Grundfragen der Strahlenbiologie. Kiln. Wochensehr. 3. Jg . , Nr. 51, 52. 1924. - - Ders. u. Slotopolsky, B.: Der RSntgenhoden. Ergebn. d. reed. Strahlenforsch. 1. 1925. - - Dies. : Beitr~ge zur experimenteUen Pathologie des tIodens und zur Histologie and Histogenese des normalen Hodens, der Hodenatrophie und der Hodennekrose. Denkschr. d. Sehweizer Natltrforscher- Ges. 1924, 61, Abh. 2. - - Wasielewski, W . v . : Theoretische und experlmentelle Bcitr~ge zur Kenntnis der Amitose. Jahrb. f. wiss Bot. 88. 1903 und 39. 1904. - - Wassermann, F. : ~be r den Einflufl erh~hter Temperatur auf die Zellen des Wurzelmeristems yon Allium ce~ , ein Beitrag zur Analyse de ~ Kernteilungsvor~anges. Ber. d. dtsch, anat. Ges. 1922.








![Störungen des Sozia] - Dr. Christian Kienbacher · kommende psychische Störungen, wie die Essstörungen (Anorexia nervosa und Bulimia nervosa) oder die Hyperaktivitäts-und Aufmerksamkeitsstörungen](https://img.pdfslide.net/doc/110x75/5e02ff29d9e2ea2f2041461b/strungen-des-sozia-dr-christian-kommende-psychische-strungen-wie-die-essstrungen.jpg)




















