-
8/2/2019 Week 7 Lecture 560B on Line Glygoen, Fruct, Gal,
Pentose Most Recent
1/9
Week 7 Lecture
Biochemistry of Nutrition, 560, B
Dr. Charles Saladino
Introduction
This lecture will cover some of the main metabolic pathways
associated with
carbohydrate metabolism. There is a defined and important
inter-relationship between
the pathways with homeostatic balance being maintained through
various types ofregulation, including covalent, allosteric,
hormonal, and second messenger systems.
These pathways will include glycogenesis and glycogenolysis,
gluconeogenersis, glucose
isomer metabolism, and the pentosephosphate pathway, otherwise
known as the
hexosemonophosphate Shunt, and the Krebs cycle. Let us begin
with thepentosephosphate pathway, often given too little time in
introductory biochemistry
courses. The structures of the pathway can be found on pages 449
& 650, Devlin) of
your text. You do not have to memorize the pathway, but you do
need to
understand its significance and the material I will now
cover.
Hexosemonophosphate Shunt or Pentose Phosphate Pathway (PPP)
Purpose of this pathway is two-fold. The first is the production
of NADPH (a reducing
agent and the reduced form of NAD+ used in many processes, such
as fatty acidsynthesis. This can be considered a cosubstrate for
key enzymes. The second
importance of the PPP is that it generates intermediate
metabolites essential for the
synthesis of nucleic acids and nucleotides. Because this pathway
represents analternative to glycolysis, be sure to reacquaint
yourself with the oxidation of glucose to
pyruvate. The PPP occurs in two phases, an oxidative phase and a
non-oxidative phase.Again, I refer you to your text as we start
with the oxidative phase. Here, glucose-6-P isthe starting
metabolite. Remember that this has been formed by the action of
the
phosphorylating enzyme, glucokinase. There are three steps
during this first phase that
result in the generation of the D isomer of ribulose-5-P. Of
importance to note is theformation of NADPH + H+ from NADP+ in
steps 1 and 3.
Notice that in the non-oxidative phase, both isomerization and
condensation reactions of
several sugar molecules occur. I am not happy with the figure
presentation in your text,so I will attempt to clarify it. First,
however, lets look for the most significant
intermediates generated by this non-oxidative phase. These
products are ribose-5-P (used
for nucleic acids), fructose-5-P, and glyceraldehyde-3-P
(G-3-P). (Remember G-3-Pfrom glycolysis?)
Now besides the G-3-P, ribose-5-P, and fructose-6-P, the
non-oxidative phase generatesthe interconversion of 3, 4, 6, and
7-carbon sugars. Some of these enter glycolysis. We
also see that the PPP pathway provides a means for cleaving the
carbon chain, one carbon
at a time. However, a big however, unlike the Krebs cycle, this
pathway does not occur
as a series of consecutive reactions leading from glucose-6-P to
six CO2
molecules.
-
8/2/2019 Week 7 Lecture 560B on Line Glygoen, Fruct, Gal,
Pentose Most Recent
2/9
I now give you a diagram below, where the two, 5-carbon
isomerization products
generated from ribulose-5-P (xyulose-5-P and ribose5-P not shown
below, but shown inyour text) interact with a transketolase enzyme
to form two products. One is the 3-carbon
G-3-P, the other is a 7-carbon sugar (sedoheptulose-7-P), shown
below. The G-3-P
formation is very important, because it can re-enter
glycolysis.
Now, the G-3-P and the seduloheptose, catalyzed by a
transaldolase, join and rearrange to
-
8/2/2019 Week 7 Lecture 560B on Line Glygoen, Fruct, Gal,
Pentose Most Recent
3/9
form two new sugar products, fructose-6-P (6 carbons) and
erythrose-4-P (4 carbons).
(By the way, you should be keeping track of carbons during these
reactions.)
The final piece of the puzzle is in the lower right part of the
above diagram. Notice that
the 4-carbon erythrose-4-P and the 5 carbon xyulose-6-P join and
rearrange (catalyzed by
a transketolase) to form fructose-6-P and G-3-P. Both are
glycolytic intermediates. Youshould also note that transketolases
will transfer 2-carbon units, whereas transaldolases
transfer 3-carbon units. Each of these carbon units is
transiently attached to the enzyme
in the course of the reaction.
Please also note that these reactions of the PPP pathway are all
reversible. That means
that the equilibrium can shift in the direction which provides
the product when it is in
demand. Just by way of one of many examples, take the situation
where more ribose-5-Pis needed than NADPH as in a cell actively
engaged in mitosis. Here is what will
happen: Most of the glucose-6-P is converted into fructose-6-P
and glyceraldehyde-3-P
(G-3-P) by glycolysis. Then the transketolase and transaldolase
convert two molecules of
fructose-6-P and one molecule of G-3-P to yield three ribose-5-P
molecules by reversalof the non-oxidative phase of the PPP pathway
described above. You could summarize
this by the following reaction (Note the 30-carbon balance on
both sides of theequation.):
5 glucose-6-P + ATP -----> 6 ribose-5-P + ADP + H+
So what would be the predominant reaction if NADPH were required
in great amounts,
as would be the case in a demand for fatty acid, cholesterol,
neurotransmitter, and
nucleotide biosynthesis? The answer is the oxidative phase of
the PPP pathway.
Glucose-6-P + 2NADP+ + H2O ribose-6-P + 2NADPH + 2H+ + CO
2
Coordination of Glycolysis and the Pentose Pathway
Obviously by now you realize the glucose-6-P is metabolized by
both glycolysis and the
PPP pathway. So how can we partition this metabolite between the
two pathways? The
cytoplasmic concentration of NADP+ plays a key role in
determining the fate of glucose-6-P. The first reaction in the
oxidative phase of the PPP pathway is essentially
irreversible, and under physiologic conditions serves as a
control site. The most
important regulatory factor is the level of NADP+, because this
is the electron acceptor in
the oxidation of glucose-6-P. Low levels of this key cofactor
are made worse by the factthat NADPH competes with NADPH+ in
binding to the enzyme. The ratio of NADP+ to
NADPH in the cytosol of the liver of a well-fed rat is 0.014.
This low ratio ensures thatNADPH generation is tightly coupled to
its utilization in reductive biosynthesis (ex fatty
acid synthesis). In other words, excess NADP+ will allow NADPH
to go anywhere. This
all translates into the fact that the non-oxidative phase of the
PPP pathway is controlled
by the availability of NADP+.
Some Final Thoughts on the Pentose Pathway
-
8/2/2019 Week 7 Lecture 560B on Line Glygoen, Fruct, Gal,
Pentose Most Recent
4/9
Please take note that by the time theribulose-5-P has been
produced from glucose-6-P,
we have gone from a hexose to a pentose by way of a
decarboxylation at step three of theoxidative phase.
Another aside: Now here is an opportunity for you to have done
some thinking. Youprobably learned in your pre-requisite (I hope)
that when mitochondria are involved,
oxygen is required. The fact that oxidation occurs in the
cytosol tells us that this process
is not dependent upon the mitochondria or upon the Krebs cycle.
Please remember thatfor next week.
You might have heard of the Calvin Cycle in plants. Here, the
cycle begins with CO2
and
goes on to use NADPH in the synthesis of glucose. In the pentose
pathway, we begin
with the oxidation of a glucose-derived carbon to CO2
to yield NADPH. Further, the
Calvin cycle converts 6-carbon and 3-carbon molecules to the
starting material,ribulose-5-P. The pentose pathway converts
5-carbon sugars (ribose-5-P) into 6-carbon
and 3-carbon intermediates of glycolysis. In photosynthetic
organisms, the enzymes arethe same as in mammalian organisms. Some
would call this evolutionary economy.What an interesting stroke of
creative genius.
Glycogen Metabolism
Glycogen can be considered a readily available storage form of
glucose. This glycogen
structure is a very large, branched glucose residue polymerthat
can be degradedto indirectly provide glucose when it is required
for energy production.Although the majority of glucose residues are
linked by -1,4-glycosidic
bonds, branches occur about every ten residues, due to
-1,6-glycosidic bond formation. This bond like that formed in
joining two amino acids is a dehydration synthesis reaction, where
water is removed to form a bond. On yourown, look up the structure
of these two bond types, as well asa -glycosidic linkage, and
please know them for the next exam. Theyare easily found in the
Devlin text or on line. I will note here that -glycosidiclinkages
form open helical polymers, but -glycosidic linkages producealmost
straight chain polymers to form fibrils, as is seen in cellulose,
forexample. Can you suggest for the Discussion Board the
advantage(s) of glycogen
having branches, instead of being a long polymer?
Biochemically, glycogen is not as reduced as are fatty acids.
Then why would we storeglycogen at all? We could just convert
excess energy into fatty acids. Right? Well,
there are several answers to that question. The first is that in
between meals, the
controlled breakdown of glycogen and subsequent release of a
glucose product makes
that sugar available to those organs where it is needed. So we
might think of glycogen asa buffer against non-homeostatic plasma
glucose levels. This is especially important for
the brain, whose only source of energy is glucose
under-non-starvation conditions.
Further, mobilization of glucose stores (as glycogen) is quick
and is a good source of
-
8/2/2019 Week 7 Lecture 560B on Line Glygoen, Fruct, Gal,
Pentose Most Recent
5/9
energy for sudden bursts of activity. Finally, glucose can
supply at least some energy
anaerobically, unlike fatty acids which require the oxygen found
in mitochondria.
The liver and the skeletal muscle are the two major storage
sites of glycogen. However,there is a greater glycogen
concentration in the liver (potentially 7-10%) vs. that of
skeletal muscle (about 2%); but because of its much greater
mass, more glycogen overall
is stored in muscle than in the liver. Microscopically, glycogen
is found in the cytosol asgranules ranging in size from 10 40 nm.
Another point of importance is that the
glycogen stored in muscle is regulated to meet the energy needs
of that itssue, where liver
glycogen degradation and synthesis are balanced so as to
maintain plasma glucose levelsfor the good of the whole
organism.
At this point we need to clarify something. Although I (and many
texts) talk of making
glucose available from glycogen, the breakdown of glycogen does
not yield a glucosemolecule directly. The degradation process is
basically a three-step process:
1) glucose-1-P is released from glycogen; 2) glycogen as a
substrate is remodeled to
allow further degradation, as it is during synthesis; 3)
glucose-1-P is further converted
into glucose-6-P for further metabolism. This overall breakdown
process is calledglycogenolysis. Observe this on page 630-631 of
your text. Then look at Figure15.51 to observe glycogenesis. You
are responsible for the names of the respective
enzymes for the next exam.
The glucose-6-P product of glycogenolysis basically has three
fates: 1) it can continue
into glycolysis; 2) it can enter the now-familiar-to-you pentose
pathway to generateNADPH and ribose derivatives; 3) it can be
converted to glucose to be released into the
blood stream.
Now in both the degradation and the synthesis of glycogen, we
take note of two modified
glucose molecules. These are UDP-glucose and UTP-glucose. The U
stands for
uridine, the DP is diphosphate, and the TP is triphosphate. This
is the same U as is foundin RNA nucleotides. Please note their
respective roles in both process, and if you have
any questions about this, let me know via email or the
Discussion Board, as you are, atleast in general, responsible for
their respective roles, as it pertains to this
discussion. You would, however, expect a phosphate to be
available to help drive the
energy-requiring synthesis of glycogen.
As a point of interest, until not long ago, it was presumed that
blood glucose was the soleprecursor for glycogenesis. However,
under physiological conditions, it is now known
that most the new glycogen is formed from not from plasma
glucose, but from
gluconeogenic (glucose-producing) precursors, especially lactate
and alanine. Both ofthese molecules are easily converted into
glucose in the liver. Please read page 620-621
of your text to find out the relationship of this gluconeogenic
lactate to the Cori
cycle. There is also an alanine cycle which sends alanine from
the muscle cells to theliver.
The final topic for glycogen is its regulation. This is quite
complex. Various enzymes
that participate in glycogen metabolism allosterically respond
to metabolites that signal
-
8/2/2019 Week 7 Lecture 560B on Line Glygoen, Fruct, Gal,
Pentose Most Recent
6/9
the cells energy needs. In other words, enzyme activity is
adjusted to meets the needs of
the cells in which the enzymes are present. Glycogen metabolism
is also hormonally
regulated, in order to allow glycogen metabolism to respond to
the needs of the wholeorganism. My apologies, but you need to know
the main features of Figure 15.54-15.55 in order to appreciate some
of this regulation.
Then email me to explain how the insulin/glucagon ratio directly
affects the events
shown in the figure. I want some mechanism described here,
please.
Feeder Pathways into Glycolysis
The purpose of this section is to account for the fate of two
important monosaccharidesafter they have been digested and
absorbed. These two sugars are galactose and fructose.
The increase in fructose in the American diet (whatever that is)
has been meteoric over
the last couple of decades. This is because of a variety of
reasons, not the least of which
is that it is a very inexpensive commodity. In terms of
metabolic health and efficiency,this huge increase in the use of
fructose is doing none of us any good.
Galactose Metabolism
Please read your text on the Absorption, Transport, and
Distribution of galactose
and fructose. Note that they are not absorbed in the same
manner, with galactose
utilizing the same Na+-dependant carrier system as glucose
does.
Obviously, a major source of dietary galactose is lactose,
especially from milk and milk
products. Other sources include the cell turnover, especially
cell membranes, upon which
lysosomes work to provide glycoproteins and glycolipids from
those membranes.
Like fructose and glucose, galactose must be phosphorylated
before it can be mobilized.
This takes place in the liver. Galactose kinase is the enzyme
responsible for thisphosphorylation, and it costs one ATP. The
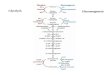
product is galactose-1-P. Refer to the
figure below to follow this pathway, and please pay close
attention to the color scheme,
so that you will more easily understand what is happening.
First, the newly generated galactose-1-P can not enter
glycolysis until it is first converted
to UDP- galactose. This occurs in an exchange reaction. In this
reaction, the enzyme
galactose-1-P uridyl transferase transfers the removal of UMP
(shown in black) fromUDP-glucose and adds it to the galactose-1-P
to form UDP-galactose (shown in red and
black), leaving behind glucose-1-P (shown in blue).
Notice two things that are happening. First, UDP galactose can
under go an
isomerization reaction, using the enzyme UDP-galactose
4-epimerase. This reaction
regenerates UDP-glucose, which is recycled back to start a new
set of reactions. Note the
efficiency of this, as opposed to using a totally different
pathway to generate all the UDP
-
8/2/2019 Week 7 Lecture 560B on Line Glygoen, Fruct, Gal,
Pentose Most Recent
7/9
glucose needed for this reaction sequence which brings us to a
the second point. The
formation of glucose-1-P is now positioned to be converted into
glucose -6-P (by another
isomerization reaction using phosphoglucomutase) or even glucose
by removing thephosphate group using the enzyme
glucose-6-phosphatase. The glucose itself can be
utilized in a variety of ways, including being exported to other
tissues from the liver. On
the other hand, the newly-generated glucose-6-P is a metabolite
that is part of glycolysis,and hence it can be incorporated into
that pathway for oxidation to pyruvate.
There are certainly some side issues here. For example, if
energy is abundant, say from ahigh caloric intake, the glucose-1-P
might be channeled into glycogenesis instead of
glycolysis. This type of common sense thinking is what I hope
you will strive to achieve.
However, the bottom line of what we have shown here is that (and
how) the carbons of
galactose can wind up in glucose for glycolysis, because there
are no catabolic pathwaysto metabolize galactose.
Fructose
In most organisms, like galactose and other hexoses (such as
mannose), fructose can
enter glycolysis after conversion to a phosphorylated
derivative. D-fructose is found in
many fruits, but is unfortunately loaded into our diets in a
variety of ways, from syrups to
-
8/2/2019 Week 7 Lecture 560B on Line Glygoen, Fruct, Gal,
Pentose Most Recent
8/9
candies etc. Whereas galactose and glucose are absorbed into the
intestinal mucosa by
way of an active, energy-requiring transporter with a
countercurrent uptake of sodium,
fructose uptake utilizes a far less efficient,
sodium-independent transporter (GLUT-5) forabsorption. All three
monosaccharides require another transporter (GLUT-2) to then
enter the portal circulation.
Here we must make a clear distinction between the metabolic
events taking place in the
liver vs. those in the muscle. In the liver, the fructose-1-P
pathway is utilized. This is
illustrated in this Figure to the right. The first step
involves
the phosphorylation of fructose to fructose-1-P, using the
enzyme fructokinase and at the
cost of one ATP. This six-carbon product is then split into two,
three-carbon sugars,dihydroxyacetone phosphate (DHAP) and
glyceraldehyde by the enzyme B aldolase.
Note that the DHAP is a glycolytic intermediate. A triose kinase
then converts the
glyceraldehydes to glyceraldehyde-3-P, also a glycolytic
intermediate and at the cost of
one ATP. By this means, fructose can enter glycolysis.
In the liver, fructose can theoretically be converted to
fructose-6-P by a glucokinase
enzyme. However, the affinity of the glucokinase is 20 times
greater for glucose than forfructose. Thus, very little
fructose-6-P is formed in the liver, because glucose is so much
more abundant in this organ. Only a heavy intake of fructose
would activate this
pathway. Besides, in the liver, glucose is the preferred fuel,
as is the case in muscle.
In the muscle, fructose can be converted directly to
fructose-6-P at the cost of an ATP.
This is already an intermediate of glycolysis, although again,
glucose is the preferredfuel.
There has been a ten-fold increase in fructose consumption in
the last 25 years in the
United States, due to the high use of high-fructose corn syrup,
which is sweeter and ischeaper to produce than glucose. Although we
will not discuss carbohydrate regulation
until the next lecture, I will note here that fructose
catabolism in the liver by passes the
phosphofructokinase catalyzed step of glycolysis, wherein
fructose-6-P is converted intofructose-1,6-bisphosphate. This is a
highly regulated metabolic control point. This could
potentially disrupt fuel metabolism, whereby glycolysis would be
directed toward lipid
-
8/2/2019 Week 7 Lecture 560B on Line Glygoen, Fruct, Gal,
Pentose Most Recent
9/9
synthesis, if ATP production were not needed. Importantly,
herein might be a link
between fructose consumption and obesity that is ever-increasing
in the United States.