ANRV322-MI61-25 ARI 6 August 2007 18:18
Morphogenesis in Candidaalbicans∗
Malcolm Whiteway1,2 and Catherine Bachewich3
1National Research Council of Canada, Biotechnology Research Institute, Montreal,Quebec, H4P 2R2, Canada; email: [email protected] Department, McGill University, Montreal, Quebec, H1A 1B1, Canada3Biology Department, Concordia University, Montreal, Quebec, H4B 1R6, Canada;email: [email protected]
Annu. Rev. Microbiol. 2007. 61:529–53
First published online as a Review in Advance onJune 18, 2007
The Annual Review of Microbiology is online atmicro.annualreviews.org
This article’s doi:10.1146/annurev.micro.61.080706.093341
Copyright c© 2007 by Annual Reviews.All rights reserved
0066-4227/07/1013-0529$20.00∗ The Canadian Government has the right toretain a nonexclusive, royalty-free license inand to any copyright covering this paper.
Key Words
hyphae, pseudohyphae, opaque, chlamydospore, budding
AbstractCandida albicans is termed a dimorphic fungus because it proliferatesin either a yeast form or a hyphal form. The switch between theseforms is the result of a complex interplay of external and internal fac-tors and is coordinated in part by polarity-regulating proteins that areconserved among eukaryotic cells. However, yeast and hyphal cellsare not the only morphological states of C. albicans. The opaque formrequired for mating, the pseudohyphal cell, and the chlamydosporerepresent distinct cell types that form in response to specific geneticor environmental conditions. In addition, hyperextended buds canform as a result of various cell cycle–related stresses. Recent studiesare beginning to shed light on some of the molecular controls reg-ulating the various morphogenetic forms of this fascinating humanpathogen.
529
Ann
u. R
ev. M
icro
biol
. 200
7.61
:529
-553
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Con
cord
ia U
nive
rsity
- M
ontr
eal o
n 05
/11/
11. F
or p
erso
nal u
se o
nly.

ANRV322-MI61-25 ARI 6 August 2007 18:18
Contents
INTRODUCTION. . . . . . . . . . . . . . . . . 530THE YEAST-HYPHAL
TRANSITION . . . . . . . . . . . . . . . . . . 532Polarity Determinants . . . . . . . . . . . . 532Spitzenkorper and Hyphal
Development . . . . . . . . . . . . . . . . . 533Nuclear Positioning and
Division . . . . . . . . . . . . . . . . . . . . . . 533Signaling Pathways . . . . . . . . . . . . . . . 534Roles of Cyclins and
Cyclin-Dependent Kinases inMorphological Control . . . . . . . . 537
THE WHITE-OPAQUESWITCH. . . . . . . . . . . . . . . . . . . . . . . . 537Pheromone-Mediated
Morphological Changes . . . . . . . 539CHLAMYDOSPORE
FORMATION . . . . . . . . . . . . . . . . . . . 541OTHER GROWTH MODES . . . . . . 542
Pseudohyphal Growth . . . . . . . . . . . . 542Hyperpolarized Buds . . . . . . . . . . . . . 543
CONCLUSIONS. . . . . . . . . . . . . . . . . . . 545
INTRODUCTION
The fungi are a highly successful and diversegroup of organisms. They exhibit two promi-nent modes of proliferation: the hyphal mode,in which continuous growth at a tip leads toan elongated tube wherein separate cells aregenerally delineated by septa, and the yeastgrowth mode, in which discrete cells elon-gate or bud-off daughter cells that typicallydisassociate from the mother cell. Most fungiare hyphal; even mushrooms are constructedfrom collections of filamentous hyphae. Un-derstanding this mode of growth has been along-standing scientific goal. The yeast formis less frequently encountered in nature butis found in economically important and sci-entifically well-investigated organisms such asSaccharomyces cerevisiae and thus is also a highlystudied growth pattern.
Intriguingly, some fungi are not limitedto a specific growth mode. Depending on
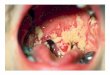
the environmental conditions, these fungiare capable of growing in either the yeast orthe hyphal pattern; such fungi are termeddimorphic. Among the best studied of thesefungi is the human pathogen Candida albicans.Although dimorphic is a bit of a misnomergiven the variety of cellular forms exhibitedby this organism, specific conditions can befound in which essentially all the cells growas yeast, and other conditions can be foundin which most cells grow as hyphae, andthe switch from one mode to the other inresponse to external signals is rapid. Thisability to switch between such distinct modesof proliferation is intriguing, but it has alsodrawn interest because of its apparent rele-vance to the pathogenicity of the organism(120). In addition to this yeast-hyphal tran-sition that defines C. albicans as a dimorphicfungus, a number of other naturally occurringmorphological forms are characteristic ofspecific cellular functions (Figure 1). Thesedistinct morphologies include the opaqueform, characteristic of mating-competentcells (112); the chlamydospore, a currentlyenigmatic thick-walled cell formed typicallyunder suboptimal growth conditions (44); andthe pseudohyphal form, which often coexistswith the yeast and hyphal forms in vegetativecultures and during infections (120).
In this article we provide an overview of de-velopments in our understanding of the mor-phological variation exhibited by C. albicans.The general topic of C. albicans morphogene-sis has been the subject of many recent reviews(21, 72, 80, 120, 130), so this manuscript fo-cuses mainly on recent developments. Thereare also excellent reviews documenting ourunderstanding of the regulation of morpho-genesis in Saccharomyces cerevisiae (31, 58, 70)and of the growth regulation of obligate hy-phal fungi such as Aspergillus nidulans andNeurospora crassa (94, 128). Ultimately our pic-ture of C. albicans morphogenesis involves anunderstanding of both the processes and reg-ulatory circuits unique to Candida, as well asof the general functions characteristic of boththe budding and hyphal growth modes.
530 Whiteway
Ann
u. R
ev. M
icro
biol
. 200
7.61
:529
-553
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Con
cord
ia U
nive
rsity
- M
ontr
eal o
n 05
/11/
11. F
or p
erso
nal u
se o
nly.

ANRV322-MI61-25 ARI 6 August 2007 18:18
Yeast
Pseudohyphae
Polarisome
Spitzenkorper SeptumHyphae
Mating projection Matingprojection
Opaquecell
Suspensor cell
Chlamydospore Chlamydospore
Septin band
Nucleus
Figure 1Distinct morphological forms of C. albicans. In yeast-form growth a blastospore buds off a new cell,resulting in two discrete cells. The separated spindle pole bodies elaborate spindles that separate thechromosomes across the mother-daughter junction defined by a septin band (green). Polarized growth isdefined by a crescent-shaped polarisome at the tip of the growing bud. In pseudohyphal growth thenuclear division also crosses the mother-daughter junction defined by a septin band, and polarizedgrowth is characterized by the polarisome. The cells themselves are more elongated than during yeastgrowth, and the cells remain attached after cytokinesis. Hyphal growth is defined by both a polarisomeand a Spitzenkorper at the tip of the growing hyphae. The nucleus divides within the elongating germtube across the region that defines the eventual septum and then one nucleus migrates back to themother cell and the other moves farther into the elongating germ tube. Opaque-form cells are capable ofresponding to mating pheromone by elongating a mating projection, or shmoo. The nucleus migratesinto the projection and, after fusion of the mating projection to a cell of the opposite mating type,undergoes a Kar3p-mediated fusion with the other nucleus. Chlamydospores are formed at the end ofsuspensor cells. They have a thicker cell wall and are larger than blastospores; the nucleus divides withinthe suspensor cell and then the daughter nucleus migrates into the chlamydospore across a septinstructure. The final chlamydospore has an elaborate septin-derived substructure. See text for details.
www.annualreviews.org • Morphogenesis in C. albicans 531
Ann
u. R
ev. M
icro
biol
. 200
7.61
:529
-553
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Con
cord
ia U
nive
rsity
- M
ontr
eal o
n 05
/11/
11. F
or p
erso
nal u
se o
nly.

ANRV322-MI61-25 ARI 6 August 2007 18:18
GTPase: guanosinetriphosphatebinding/hydrolyzingenzyme
Polarisome:complex of proteinsinvolved in directingpolarized growth
Germ tube: initialelongating structurethat will become ahypha
THE YEAST-HYPHALTRANSITION
Polarity Determinants
The ability to switch rapidly from yeast-formgrowth to hyphal growth is a defining charac-teristic of C. albicans cells, and it is believedthat each form of growth provides criticalfunctions required for the pathogenic lifestyle(120). In both modes the cells exhibit polar-ized growth; the degree of polarity is moreextreme in the hyphal state, because the al-ternation of polarized growth with isotropicgrowth that characterizes the yeast mode ofproliferation is replaced by the essentially per-manently polarized apical growth of the hy-phal cell. In general, polarized growth is animportant characteristic of cells, with intrin-sic cues and extrinsic signals combining todetermine both the shape of individual cellsand their spatial organization in multicellu-lar structures. Eukaryotic cells typically usesmall GTPases of the Rho superfamily, in par-ticular members of the Cdc42/Rac group ofenzymes, to regulate the overall process (47,59); these GTPases appear implicated primar-ily in regulating actin polymerization and thusin controlling polarized growth and secretionthrough modulation of the actin cytoskele-ton (63). Cdc42p is a central regulator of acomplex of proteins required for proper polar-ized growth. This complex, initially identifiedin budding yeast and termed the polarisome(109), contains the formin Bni1p as wellas the proteins Bud6p, Spa2p, and Pea2p(Figure 1).
Members of the polarisome and its GT-Pase regulator are also involved in polarizedgrowth in C. albicans. Deletion (91) or regu-lated shut-off (122) of the C. albicans CDC42homolog blocks cell cycle progression underboth yeast- and hyphal growth conditions andgenerates apolar growth, as did inactivationof the protein in S. cerevisiae. During hyphalgrowth the Cdc42p GTPase remains at the tipof the elongating hyphal tube; this localizationis disrupted by latrunculin A treatment, sug-gesting it is dependent on the actin cytoskele-
ton (52), unlike the localization control duringyeast-form growth. Regulation of the GTPaseactivity of Cdc42p plays an important role incontrolling the formation of hyphae, as reduc-tion in GTPase function through loss of theexchange factor Cdc24p blocks the ability toform hyphae and induce hyphae-specific tran-scripts (14), and an increase in the activatedform by deletion of the GTPase-activatingproteins Rga2p and Bem3p allows cells grow-ing under normally pseudohyphae-inducingconditions to form hyphal-like germ tubes(36).
The polarisome is also implicated in po-larized growth in C. albicans, as deletion ofthe homologs of BNI1 (30, 88), SPA2 (135),and BUD6 (113) results in similar pheno-types. Yeast-growth cells exhibit random bud-ding and delocalized surface growth, leadingto round cells with large bud necks; in hy-phal growth conditions these mutants makeabnormally thick hyphae. Although cells canstill polarize in the absence of polarisomefactors, polar growth cannot be maintained.Unlike S. cerevisiae, but like higher cells,C. albicans also has a homolog of the RacGTPase. This protein, termed Rac1p, playsa role distinct from that of Cdc42p, and isnot needed for budding and proper actin or-ganization, but it is needed for filamentationin matrix-embedded conditions (13). Thus,there may be environment-specific polarity-determining factors in hyphae. Componentsthat directly interact with the actin cytoskele-ton also play important roles in the controlof C. albicans polarity. The type I myosin en-coded by MYO5 is required for hyphal growthbut not for budding; surprisingly, the polar-ized distribution of cortical actin patches canbe disrupted in specific myo5 mutants withoutblocking hyphal development (100). The typeV myosin encoded by the MYO2 gene is alsorequired for development of true hyphae butnot for budding (132), and loss of the Wiskott-Aldrich syndrome protein Wal1 (126) and theSla2 protein (101) also disrupts hyphal devel-opment without these proteins being essentialfor viability during yeast-form growth.
532 Whiteway
Ann
u. R
ev. M
icro
biol
. 200
7.61
:529
-553
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Con
cord
ia U
nive
rsity
- M
ontr
eal o
n 05
/11/
11. F
or p
erso
nal u
se o
nly.

ANRV322-MI61-25 ARI 6 August 2007 18:18
Spitzenkorper and HyphalDevelopment
In inherently filamentous fungi such asAspergillus and Neurospora there is a distinctstructure called a Spitzenkorper (Figure 1)near the tip of the growing cell that isimplicated in polarized growth (125). ThisSpitzenkorper is believed to direct secretionto the growing tip and thus has a functionsimilar to that of the polarisome of the bud-ding yeast (51). However, it does not appearthat the switch from yeast to hyphal growth inC. albicans occurs by the transfer of regulatorycontrol from a polarisome to a Spitzenkorper,as both elements have been identified in hy-phal cells (37) and, as noted, hyphal cells lack-ing polarisome components do not generatenormal hyphae. Thus, the Spitzenkorper ap-pears to be one of the unique characteristics ofthe hyphal growth pattern in C. albicans, and itmay function in conjunction with, instead ofin place of, the polarisome (37). Because theSpitzenkorper is only found in actively grow-ing hyphae of filamentous fungi, care must betaken when using it as a diagnostic feature forthe true hyphal growth mode in C. albicans;the mere absence of the structure in a polar-ized cell may reflect a poor growth rate ornongrowing cell. Individual growth rates andlocalization of Spitzenkorper markers need tobe monitored in the same cells.
Defining the molecular characteristics ofthe Spitzenkorper, and the relationship be-tween this element and the polarisome, willbe an important step in both defining polar-ity control in hyphal cells and specifying thedistinction between hyphal and yeast-formgrowth. Because the Spitzenkorper is identi-fied primarily as a vesicle-rich structure cur-rently visualized in living cells through stain-ing with dyes such as FM4–64, the protein/enzymatic components of the Spitzenkorperare as yet poorly characterized. In C. albicansthe polarisome proteins Spa2p and Bud6p aregenerally not associated with the Spitzenkor-per, whereas Bni1p and, when overexpressed,a fraction of Cdc42p are apparently associated
Spitzenkorper:membraneousstructure at thegrowing tip ofhyphal cells
GFP: greenfluorescent protein
with this structure (37). In filamentous fungi,components of the polarisome that functionat the hyphal tip, but are not involved in theformation or stability of the Spitzenkorper,have been identified (124). The Golgi struc-ture of hyphal C. albicans cells is fundamen-tally relocalized to the growing tip of the cell,and this organellar redistribution is depen-dent on the formin Bni1p (104). It does notappear that this repositioned Golgi representsthe Spitzenkorper, but the ability to identifycellular localization of membranes throughstaining with lipophilic dyes, and the abilityto localize proteins through green fluorescentprotein (GFP) tagging, will permit the iden-tification of genes required for Spitzenkorperformation and positioning and the proteinsthat colocalize with the Spitzenkorper. Theseapproaches applied in both C. albicans andin filamentous fungi should provide rapid in-sights into the organization of the Spitzenkor-per and its relationship to other polarity-related functions in the cell. They will alsoallow a more direct investigation of whetherthe Spitzenkorper-like structure in C. albicansis required for rather than associated with po-larized growth of hyphae. To date, treatmentsthat perturb the integrity of the Spitzenkor-per, including deletion of polarisome genesor exposure to methyl benzimidazol-2-yl-carbamate (37), also disturb polarized growth,but the growth defect may not be due to a di-rect effect on the Spitzenkorper-like structureitself.
Nuclear Positioning and Division
In addition to the presence of the Spitzenkor-per, other characteristics distinguish a C. albi-cans cell initiating a bud from those initiatinga germ tube that elaborate into hyphae. A crit-ical difference is the arrangement of the sep-tum and the division of the chromosomes. Inyeast cells, as in cells of S. cerevisiae, the divid-ing nucleus is positioned to the neck betweenthe mother and daughter cells, and the nucleardivision partitions one chromosomal comple-ment into the mother cell and the other into
www.annualreviews.org • Morphogenesis in C. albicans 533
Ann
u. R
ev. M
icro
biol
. 200
7.61
:529
-553
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Con
cord
ia U
nive
rsity
- M
ontr
eal o
n 05
/11/
11. F
or p
erso
nal u
se o
nly.

ANRV322-MI61-25 ARI 6 August 2007 18:18
Cytokinesis:process of separatingtwo cells aftermitosis hassegregated thegenetic information
APSES (Asm1p,Phd1p, Sok2p,Efg1p, and StuAp):group of structurallyrelated transcriptionfactors
cAMP: cyclic AMP
the daughter cell (Figure 1). This divisiontakes place across the septum structure, whichultimately defines the site of cytokinesis (120).A molecularly similar event occurs in pseudo-hyphal cells—the nuclear division also occursbetween the mother and daughter cell, andalthough the dividing cells are elongated rel-ative to the yeast cells, the behavior of thedividing nucleus and the organization of theseptum are similar.
The cellular behavior of the nucleus is dis-tinct in unbudded yeast cells that are elabo-rating a germ tube while in the G1 phase ofthe cell cycle. In these cells the nuclear di-vision and the position of the initial septumtake place in the elongating germ tube ratherthan at the junction between the mother celland the polarizing structure (46). The nucleusmigrates into the extending germ tube, di-vides across the site of the incipient septum,and then one nucleus migrates back into themother cell and the other moves out towardthe tip of the elongating germ tube (Figure 1).This complex movement of the nucleus dur-ing the initial steps of hyphal development isorchestrated by microtubules (46). The divid-ing cell appears to be committed to elaboratea germ tube or a bud prior to any morpho-logical distinction in the emerging polarizedstructure. However, the cell fate commitmentappears to be reversible. Yeast cells in later cellcycle stages can be induced to form hyphaefrom their established buds (53). In these cells,the first nuclear division occurs across the budneck, where there is also a constriction due tothe initial bud emergence from the yeast cell.Thus, defining the initiation signals and howthose signals are regulated by external con-ditions will be an important goal of the nextperiod of investigation of the yeast-to-hyphaltransition.
Signaling Pathways
Our growing appreciation of the complexmachinery involved in directing polarizedgrowth has been paralleled by an improvedunderstanding about the signaling pathways
involved in transferring information aboutthe environmental state that ultimatelycontrols the yeast or hyphal morphology.These environmental conditions can varyconsiderably, but a standard trigger of hyphaldevelopment is the combined addition ofserum to rich growth medium and theelevation of the growth temperature to 37◦C(130). However, nutrient-poor media suchas Lee’s or more chemically defined inducerssuch as N-acetyl-glucosamine, togetherwith a rise in temperature to 37◦C, are alsosuitable for inducing a high frequency ofhyphal growth. An important consideration isthat improved characterization of the hyphal,yeast, and pseudohyphal states can permit amore exact definition of the role of specificgenes and pathways in cell morphogenesis.Studies in which filamentous growth was de-scribed without specification to the hyphal orpseudohyphal form must be re-evaluated todistinguish functions that are common to bothstates from those common to only one state.
Much of the analysis of the yeast-hyphaltransition has been concerned with transcrip-tional control and the relevant transcriptionfactors (Table 1). The initial observation thatthe combined loss of the Cph1p and Efg1ptranscription regulators blocked the hyphaltransition in most tested conditions, and led toreduced virulence, focused interest on thesetwo transcription factors and their controllingnetworks (81). Subsequently, a large numberof transcription regulators have been reportedto play roles, either positive or negative, inthe yeast-to-hyphal transition. An importantgoal has been to connect these regulators tosignaling pathways transferring informationfrom the external environment and to estab-lish the regulatory relationships among thevarious transcription factors.
The APSES protein Efg1p plays a cen-tral role in the general control of morpho-genesis (42); in the yeast-hyphal transitionthe regulatory circuit leading through Efg1pis linked to cAMP metabolism. The role ofthe cAMP-mediated signaling pathway hasbeen well established in the yeast-to-hyphal
534 Whiteway
Ann
u. R
ev. M
icro
biol
. 200
7.61
:529
-553
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Con
cord
ia U
nive
rsity
- M
ontr
eal o
n 05
/11/
11. F
or p
erso
nal u
se o
nly.

ANRV322-MI61-25 ARI 6 August 2007 18:18
Table 1 Transcriptional regulators of morphogenesis
State Factor Loss-of-function phenotype Reference(s)Opaque state Wor1p Blocks opaque state 136
Efg1p Activates subset of functions 115Tup1p Modulates subset of functions 103
Hyphal state Rim101p Block in alkali-induced hyphae 40Czf1p Block in matrix-induced hyphae 26Flo8p Block in serum-induced hyphae 29
Derepresses embedded hyphae 29Efg1p Block in serum-induced hyphae 42
Derepresses embedded hyphae 49Hap5p Block in hyphae on Spider medium 62Efh1p Synergizes with efg1 42Ace2 Defect in hypoxia-triggered hyphae 95Mcm1p Activates hyphae 106Ash1p Reduces filaments on Spider medium 57Cph2p Moderate inhibition of hyphae 75Tec1p Suppresses serum-induced hyphae 107
Chlamydospores Rim101p Affects proper timing 99Nrg1p Derepresses formation on Staib agar 119Efg1p Blocks chlamydospore formation 114
Pseudohyphal state Tup1p Constitutive pseudohyphae 24Nrg1p Constitutive pseudohyphae 25, 96Fkh2p Constitutive pseudohyphae 19Tcc1p Constitutive pseudohyphae 67Rfg1p Constitutive pseudohyphae 69Ssn6p Moderately pseudohyphal 56Ace2p Inappropriate pseudohyphae 68Rap1p Inappropriate pseudohyphae 22
Mating projections Cph1p Mating defective 33
switch, as a variety of mutations that affectthe pathway influence hyphal development (9,34, 64, 77, 87). In particular, cells defectivein the adenylyl cyclase Cdc35p are compe-tent to grow as yeast cells but are unable toform hyphae under standard inducing condi-tions (105). Transcription profiling suggeststhat Efg1p and Cdc35p control a set of genesimplicated in the yeast-to-hyphal transition,but both control significant nonoverlappingsets of other genes, consistent with each reg-ulator influencing different functions sepa-rately from hyphal development (50). In ad-dition, the Flo8p transcription factor, which
is essential for serum-induced hyphal devel-opment, controls a subset of Efg1p-regulatedgenes (29). The regulation of Efg1p func-tion through this pathway is suggested to bethrough direct phosphorylation by the cAMP-dependent protein kinase (23), and interactionwith other regulators such as Flo8p and Czf1pmay explain the ability of Efg1p to act both asan activator and as a repressor, depending onthe conditions. As yet the direct relationshipsbetween the changes in gene expression andthe changes in cell shape have not been estab-lished, and this will be an important area offuture studies.
www.annualreviews.org • Morphogenesis in C. albicans 535
Ann
u. R
ev. M
icro
biol
. 200
7.61
:529
-553
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Con
cord
ia U
nive
rsity
- M
ontr
eal o
n 05
/11/
11. F
or p
erso
nal u
se o
nly.

ANRV322-MI61-25 ARI 6 August 2007 18:18
MAP kinase:mitogen-activatedprotein kinase
Other pathways that transmit external sig-nals to the transcription machinery, includ-ing a MAP kinase signaling pathway, an exter-nal pH sensing pathway and a matrix-sensingpathway, have also been identified as playingroles in hyphal development under specificconditions. The MAP kinase pathway signalsin opaque-form cells to direct mating, but ini-tial studies established a minor role in yeast-hyphal signaling under specific nutrient con-ditions (38, 71, 76). The target of the MAPkinase cascade is proposed to be the Cph1ptranscription factor on the basis of compar-ison with S. cerevisiae, and Cph1p provideda residual level of virulence in the efg1 mu-tant background (81). The phenotypes of thevarious signaling mutants in limiting hyphaldevelopment under specific conditions weresimilar to those of the transcription factormutant, consistent with a common molecularrole.
The external pH is one of many signalsimportant for regulating the yeast-to-hyphaltransition. Many of the players in a pathwaytransmitting the pH signal to the intracellularmachinery are homologs of the componentsidentified in S. cerevisiae that also regulate re-sponse to pH. The main transcription fac-tor is Rim101p (40), which is proteolyticallyactivated by the Rim13p protease (78); lossof Rim101p function blocks alkaline-inducedhyphal development. Rim20p and Rim8pact upstream of the proteolytic step (40),as do several membrane proteins, includingDfg16p, that may act as pH sensors (12). Theprocessing and activation of Rim101p alsorequire members of the endosomal complexrequired for transport (ESCRT) protein mod-ule (35, 133). A large number of genes are un-der control of the Rim101p factor (20), andconnecting the expression of these genes tothe morphological regulatory system is neces-sary to understand the link between pH sens-ing and morphogenesis.
The regulation of hyphal growth in a ma-trix appears regulated by a complex inter-play of the transcription factors Czf1p, Efg1p,Flo8p, and other signals. Czf1p is encoded
by an autoinhibited gene required for hyphalformation when C. albicans cells are growingin embedded conditions (26). In contrast, theEfg1p/cAMP pathway that is positively re-quired for serum-induced hyphae must be in-hibited to allow hyphal development in theseconditions, because efg1, cdc35, and flo8 mu-tants result in derepression of hyphal develop-ment during matrix-embedded growth (29).The upstream regulatory region of CZF1 islarge, and it appears that transcriptional con-trol integrates a variety of signals to regulateCZF1 expression (123), similar to WOR1 inthe regulation of the opaque state (136). BothCzf1p and Flo8p may physically interact withEfg1p (29, 49), and Czf1p serves to permithyphal growth during embedded growth byrelieving the inhibitory action of Efg1p underthese conditions (123). Separating the rolesof the physical matrix conditions from thechanges in aeration associated with embeddedhyphal development is important to establisha clear picture of this regulatory circuit.
Although the modulation of the transcrip-tion program of the cell is an obvious conse-quence of the transmission of external signalsthat direct morphological changes, these arenot the only outputs of the signaling pathways.For example, kinases are key components ofmany upstream signaling modules that regu-late transcription, and their phosphorylationtargets are not limited to transcription factors.Defining the important substrates of such crit-ical components as the cAMP-dependent pro-tein kinases is essential to fully understandingthe regulation of morphological transitions.Overall, it is critical that researchers establishhow these signaling pathways control the ma-chinery directing polarized growth. Whetherthe polarisome is a direct target of signal-ing pathways, whether Cdc42p activity or lo-calization is regulated through the transmis-sion of external hyphae-inducing conditions,or whether the Spitzenkorper forms in di-rect response to the environment must be ad-dressed. Ultimately the link between signalingand morphogenesis must be made at the levelof the growth pattern itself, not solely at the
536 Whiteway
Ann
u. R
ev. M
icro
biol
. 200
7.61
:529
-553
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Con
cord
ia U
nive
rsity
- M
ontr
eal o
n 05
/11/
11. F
or p
erso
nal u
se o
nly.

ANRV322-MI61-25 ARI 6 August 2007 18:18
spectrum of gene expression correlated withthis pattern.
Roles of Cyclins andCyclin-Dependent Kinasesin Morphological Control
The central cell cycle regulatory kinase, aswell as a group of regulatory cyclin molecules,has been identified in C. albicans, and sev-eral of these molecules appear to have spe-cific roles in morphogenetic control. The ini-tial identification of functional cyclins and thecyclin-dependent kinase was through analy-sis of C. albicans gene functions in S. cere-visiae (108, 129), but genome-sequencing ef-forts have now allowed the comprehensiveidentification of the cyclin/kinase gene fam-ilies in the fungal pathogen (21). As in themodel yeast S. cerevisiae there is a single cyclin-dependent kinase that is the key regulator ofcell cycle progression, and this kinase interactswith a variety of cyclins to control the G1, S,and mitotic transitions, as well as aspects ofmorphogenesis (90).
Ccn1p is a member of the G1 class of cy-clins on the basis of both structural and ex-pression pattern similarity to G1 cyclins ofyeast and is essential for the maintenance,but not initiation, of hyphal growth underspecific nutrient conditions (84). The cyclinHgc1p also shows structural similarity to theG1 cyclin class but was expressed only in hy-phal cells. Hgc1p is required for hyphal devel-opment under a variety of tested conditions(134), and Hgc1p expression is limited to theapical, growing cell of the developing hyphae(127). Another G1 class cyclin, Cln3p, is re-quired for budding but not initiation of hy-phal growth, as regulated shut-off of Cln3pexpression leads to cell enlargement followedby initiation of germ tube extension (8, 32).These data suggest an important regulatoryconnection between the G1 cyclins and mor-phogenetic control, but this connection seemsinconsistent with the observation that initia-tion of germ tube formation was apparentlycell cycle independent (53). Based on expres-
Opaque/White:two morphologicallydistinct cell/colonyforms of C. albicans
sion of Hgc1p, which peaks in the G1 phase(127), perhaps cells are more primed to formhyphae during G1.
B-type cyclins and the cyclin-dependentprotein kinase Cdc28p are also implicated incontrol of morphogenesis, but not directly incontrol of the yeast-to-hyphal transition. Lossof the B-type cyclin Clb4p caused pseudohy-phal growth, whereas shut-off of the essentialcyclin Clb2p resulted in more highly polarizedfilaments called elongated buds (18). Similarfilamentation has been seen in mutants thatlack function in other essential cell cycle com-ponents (7). Although these filaments super-ficially resemble hyphae, they are discussedseparately. Regulated shut-off of CDC28 ex-pression leads to diverse types of filamentation(121), including pseudohyphae, true hyphae,and elongated buds.
Currently the data linking cyclins and cellmorphogenesis have focused primarily on therole of cell cycle regulators in the transitionbetween the yeast and hyphal states. It is im-portant to extend these observations to otheraspects of morphogenesis. For example, cellcycle regulation is coupled to the developmentof the mating projection during mating, andthe control of cyclin function is likely impor-tant for this process. Similarly, cell cycle con-trol in the development of the chlamydosporeis a critical component of the process. Overallthe link between the cyclin-regulated func-tion of the cyclin-dependent protein kinaseand cellular morphogenesis is expected to bestrong, and the challenge is to make this linkin each of the unique morphological forms ofC. albicans and then to establish the details ofthe regulatory circuit.
THE WHITE-OPAQUE SWITCH
Although the dramatic change betweenthe yeast and hyphal state is a well-studiedhallmark of C. albicans, phenotypic switching,in particular the white-opaque transition, hasalso been an area of extensive investigation.The white-opaque switch was initially iden-tified as a cellular and colony morphology
www.annualreviews.org • Morphogenesis in C. albicans 537
Ann
u. R
ev. M
icro
biol
. 200
7.61
:529
-553
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Con
cord
ia U
nive
rsity
- M
ontr
eal o
n 05
/11/
11. F
or p
erso
nal u
se o
nly.

ANRV322-MI61-25 ARI 6 August 2007 18:18
transition that was limited to specific strains.The switch was epigenetic—each state wasstable but capable of transitioning to theother state at a frequency that was higher thanthe standard mutation rate, and the frequencycould be modified by external conditionssuch as temperature (110). White-form cellsare of the classic ovoid shape characteristic ofbudding yeasts such as S. cerevisiae, and theyform domed colonies that are a creamy color.Opaque-form cells are elongated and havea cell wall that has frequent pits (3), unlikethe relatively smooth cell wall surface ofwhite-form cells. Opaque-form cells generatecolonies that are flattened and more graythan the colonies generated by white-formcells (110). The viability of opaque-formcells is reduced relative to white-form cellsunder many growth conditions, and thisallows the opaque-form cells to be distin-guished as forming colonies that can bemore readily stained by vital dyes such asphloxine B.
Recent evidence has established that thiswhite-opaque transition is intimately coupledto the sexual mating process in C. albicans. Theability to switch to the opaque state dependson whether the cells are homozygous for theMTL locus that controls cell type; most C.albicans cells are heterozygous for the MTLlocus (MTLa/MTLα) and thus are unable toswitch (92). The inability of MTL homozy-gous strains to switch is regulated by the a1/α2repressor. This heterodimeric repressor is de-rived from the a1 protein encoded by theMTLa locus, and the α2 protein encoded bythe MTLα locus, and thus can only be gener-ated in heterozygous strains. A key regulatoryrole of this repressor is to control the expres-sion of another transcription factor, the prod-uct of the WOR1 gene (55, 136). The Wor1ptranscription factor (Table 1) appears to bea primary controller of the opaque state; ec-topic expression of Wor1p efficiently inducesthe opaque state in MTL homozygotes, and apseudo-opaque state can be triggered by ac-tivating Wor1p expression even in MTL het-erozygotes (136).
The regulation of Wor1p explains the epi-genetic characteristics of the opaque state.Wor1p is autoregulated; chromatin immuno-precipitation experiments suggest as many asfive binding sites for Wor1p in its own pro-moter region (136). The current data suggestthat once Wor1p expression is established, ittends to remain on, with a positive-feedbackloop keeping the cell in the opaque state.However, once Wor1p levels drop below acritical threshold, this positive-feedback loopis disrupted and the cell establishes a sta-ble white state. It could be expected that thecircuit itself is temperature sensitive becausehigh temperatures lead to the switching ofthe opaque form to the white form. Whetherthis is due to direct temperature sensitivityof the Wor1 protein itself, or to some otheraspect of the regulatory circuit, remains to beestablished.
There are many genes whose expressionswitches between cells in the white formand cells in the opaque form (74). Many ofthese genes are likely to control the distinctphysiologies of the two different cell types:White-form cells express genes characteristicof a fermentative life style, whereas opaque-form cells show characteristics of an oxidativemetabolism (74). However, many genes reg-ulated differently between the two cell typespresumably define the different growth pat-terns and cell surface structures exhibited bythe white- and opaque-form cells. Analysis ofopaque-specific genes for the controllers ofmorphological patterning is an important stepin the analysis of the opaque-phase cells.
The Efg1p transcription regulator that isa critical positive controller of the yeast-to-hyphal transition and the Tup1p negative reg-ulator of pseudohyphal development also playroles in the white-opaque switch (Table 1).Efg1p is highly expressed in white-form cellsbut not in opaque-form cells (114); this ex-pression is driven from a strong, white-phase-specific promoter (73). Loss of EFG1 ex-pression causes otherwise white-form cells totake on some of the morphologies of opaque-form cells, in particular the elongated cell
538 Whiteway
Ann
u. R
ev. M
icro
biol
. 200
7.61
:529
-553
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Con
cord
ia U
nive
rsity
- M
ontr
eal o
n 05
/11/
11. F
or p
erso
nal u
se o
nly.

ANRV322-MI61-25 ARI 6 August 2007 18:18
shape, and to express some opaque-specificgenes (114, 117). Ectopic overproduction ofEfg1p shifts opaque-form cells to the whitemorphology (115), and thus repression ofEFG1 expression appears necessary to es-tablish the opaque state. However, certainstructural characteristics of the opaque state,in particular the surface pimples, are notfound in the efg1-null strains. Therefore itappears that Efg1p may regulate a subset ofthe genes involved in establishing the opaquestate.
The Tup1p transcriptional regulator alsoinfluences the opaque state. Loss of Tup1p,which creates a constitutive pseudohyphae-like state in white-form cells (24), also dramat-ically affects the morphology of opaque-formcells (103). Loss of tup1 deregulates expressionof some phase-specific genes but still permitsthe establishment of a mating-competent celltype. Thus, like loss of Efg1p, loss of Tup1pappears to modify some, but not all, charac-teristics of opaque-form cells. MTL homozy-gous cells deleted for TUP1 have the abilityto switch among a variety of cell and colonymorphologies; whether this pattern of switch-ing is influenced by Wor1p remains to beestablished.
These experiments have identified a num-ber of transcriptional regulators that play crit-ical roles in the establishment of the opaquestate; however, we do not know the detailsof how these regulators control morphol-ogy. Some early experiments characterizedthe actin cytoskeleton in opaque-phase cells(3), but the spatial and temporal regulationof the polarity machinery has not been ex-tensively examined. The ability to form thesurface pimples characteristic of the opaquestate must be under regulatory control, butthis process is currently enigmatic. The abil-ity to manipulate the opaque state throughmodulation of the Wor1p transcription reg-ulator should make the state more amenableto investigation, and so future studies directedat identifying the control elements that es-tablish the unique morphological characteris-tics of opaque-form cells should provide rapid
progress in defining the functional bases forthis distinctive morphological state.
Pheromone-Mediated MorphologicalChanges
In addition to directing the morphology ofthe opaque-form cell itself, the establish-ment of the opaque state is a prerequisitefor the induction of morphological modifica-tions due to the action of mating pheromones.These pheromones are diffusible peptide andlipopeptide molecules that are produced bycells of one mating type and act on cells ofthe other mating type. The action of thepheromones initiates a spectrum of physio-logical, morphological, and cell cycle changesthat prepare the potential mating partners forcell and nuclear fusion (11). The componentscontrolling the signaling process, includingelements regulating both morphological andtranscriptional programs as well as the cellcycle, have been extensively documented inS. cerevisiae, and the processes in C. albicansappear inherently similar (61).
Both C. albicans and S. cerevisiae producestructurally distinct pheromones from eachof the two mating types. MATα cells of thebudding yeast and MTLα cells from C. albi-cans produce simple peptide pheromones (17,83, 97, 102), and the MATa and MTLa cellsfrom the two species produce pheromonesthat are either proven or predicted to be lipid-modified peptides (28, 41). The identificationof the single gene encoding the C. albicansα-factor peptide has allowed for the chemicalsynthesis of the pheromone and the analysisof the behavior of cells in the presence of thisfactor (17, 83, 102). C. albicans cells treatedwith pheromone modify their gene expres-sion profiles (17, 83), their cell cycle progres-sion (102), and their cellular morphology (83).In keeping with the terminology establishedfor the modified morphology developedby pheromone-treated cells in S. cerevisiae,C. albicans MTLa cells treated with α-factorhave been termed shmoos (83), even thoughtheir actual shapes are unlike either the Al
www.annualreviews.org • Morphogenesis in C. albicans 539
Ann
u. R
ev. M
icro
biol
. 200
7.61
:529
-553
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Con
cord
ia U
nive
rsity
- M
ontr
eal o
n 05
/11/
11. F
or p
erso
nal u
se o
nly.

ANRV322-MI61-25 ARI 6 August 2007 18:18
Biofilm: complexassembly of variousmorphological formsof C. albicans cellsembedded in acell-derived matrix
Capp creation or the characteristic forms ofthe budding yeast. Mating projections may bea more accurate terminology for the struc-tures formed in C. albicans in response to mat-ing pheromone, but the term shmoo pro-vides a convenient if physically inaccuratesummary of a complex morphological process(Figure1).
The developing shmoo of a pheromone-treated S. cerevisiae cell uses the machineryof polarized growth characteristic of bud-ding cells, but it modifies, in response tothe pheromone gradient, the direction of po-larized growth by overriding the intrinsicrules for initiating a bud (27). Modificationsalso occur in the arrangement of the septinsat the site of the shmoo evagination rela-tive to septin arrangement in a budding cell(43). Overall, the establishment of the polar-ized growth of the shmoo without the con-striction inherent in the dividing bud of aS. cerevisiae cell has led some researchers toequate aspects of shmoo formation in buddingyeast to the development of a germ tube inC. albicans (37). This analogy cannot be takentoo far, as proteins such as Far1p that are crit-ical for shmoo formation in the budding yeastare not required for germ tube developmentin C. albicans (5). However, the ability to ini-tiate polarized growth from a nonconstrictedpoint of emergence, the ability to migrate thenucleus into this polarized extension, and therequirement for the actin cytoskeleton to di-rect the localization of the Cdc42p GTPase(52) are common characteristics of shmooingyeast cells and C. albicans cells initiating a germtube.
A detailed analysis has been undertaken toestablish the cell biology of the developingmating projections in cells in a mating mixtureconsisting of both mating types (82). Onlymixtures of cells of opposite mating types wereobserved to trigger the formation of matingprojections, and these projections could be ex-tensive. It has been proposed that a complexinteraction between white- and opaque-formcells can lead to biofilm formation that stabi-lizes pheromone gradients and permits MTLa
and MTLα cells to signal over long distances(39). This model implies that rare opaque-form cells can find each other and elaboratemating projections that are many cell diame-ters long. The observation that a subset of thepheromone-induced genes overlap with thehyphae-induced genes (17, 98) is consistentwith the possible involvement of extensivelyelongated structures in the mating process.
Initial studies on the cell biology of themating process suggested that nuclear fusionwas slow or nonfunctional, as the parentalgenomes could be recovered from the matedcells after karyogamy (82). However, more re-cent analysis of the process provides strongevidence for efficient nuclear fusion after zy-gote formation (16). This process occurs withdifferent frequencies in different strains andis highly dependent on the Kar3 motor pro-tein. Thus the typical outcome of the mat-ing process in C. albicans is the formation of astable tetraploid strain, which, because it hasre-established the MTL heterozygous state,switches to the white form for subsequentproliferation.
The ability to form projections in re-sponse to mating pheromones is under ge-netic control. A variety of mutations influencethis process; disruption of the heterotrimericG-protein and MAP kinase-mediated signal-ing pathways (33, 86) prevents morphologi-cal changes by blocking the transmission ofthe signal (D. Dignard & M. Whiteway, un-published data). Further, deletion of a FAR1homolog blocks morphological responses(P. Cote, T. Sulea & M. Whiteway, unpub-lished data); Far1p is not needed for hy-phal growth in the white-form cell phase andthus is not required in general for direct-ing polarized growth in response to externalsignals, but it is necessary for pheromone-mediated mating projections. Overproduc-tion of Far1p also hypersensitizes cells topheromone-mediated cell cycle arrest andto projection formation in the presence ofpheromones. Pheromones also appear to co-ordinate with nutrient signals in the ar-rest of opaque-form cells during mating
540 Whiteway
Ann
u. R
ev. M
icro
biol
. 200
7.61
:529
-553
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Con
cord
ia U
nive
rsity
- M
ontr
eal o
n 05
/11/
11. F
or p
erso
nal u
se o
nly.

ANRV322-MI61-25 ARI 6 August 2007 18:18
(15); therefore developing the link betweenmating-pheromone-induced signaling, nutri-tional status, and cell cycle regulation is neces-sary to fully understand pheromone-mediatedmorphological response.
CHLAMYDOSPOREFORMATION
Chlamydospores of C. albicans superficiallyresemble a stable, resting cell and are dis-tinct from the proliferative forms character-ized by the yeast or hyphal states, but thereis currently no strong evidence that long-term viability is exhibited by these cells. Ini-tial studies on chlamydospores focused pri-marily on their structure and the conditionsthat generated their formation. These inves-tigations established that environments lowin oxygen, light, temperature, and nutrientswere most conducive to the developmentof chlamydospores, and that the cells them-selves were large, with thick walls (60) anda high lipid (93) and carbohydrate (44) con-tent. Chlamydospores have been observed togerminate under certain conditions (60), andprotocols have been developed to purify sig-nificant numbers of these spores (44). Mediaconsisting of rice or cornmeal agar supple-mented with the detergent Tween 80 are stan-dard conditions for the induction of chlamy-dospores. These chlamydospores themselvesform at the ends of branched filaments orsuspensor cells that form under the induc-ing conditions (Figure 1). The formation ofchlamydospores under specific conditions isone of the most effective ways to distinguishC. albicans from its close relative, C. dubliniensis(118).
Current studies are providing a more de-tailed picture of the regulatory circuits con-trolling the production of chlamydosporesin C. albicans. The transcriptional regulatorEfg1p (114) and the MAP kinase Hog1p (1)are required for chlamydospore formation. Inboth cases, filamentation occurred during themicroaerophilic conditions, but production ofthe chlamydospores themselves was blocked.
A more general search for functions requiredfor chlamydospore formation was undertakenwith a collection of insertion mutants. Thisapproach did not demand the preselection ofcandidate genes but rather directly screenedfor defects in chlamydospore formation witha set of 217 genes inactivated through atransposon mutagenesis and mitotic recom-bination strategy (99). This study identifiedthe SUV3, SCH9, and ISW2 genes as essen-tial contributors to the formation of chlamy-dospores, and RIM11, RIM101, and MSD3were needed for proper timing of their pro-duction. The suv3 and sch9 mutants weredefective in the formation of hyphae-like fila-ments that serve as the source of the suspensorcells, and the isw2 mutants formed the sus-pensor filaments but failed to elaborate thechlamydospores, similar to the situation ob-served for the efg1 and hog1 mutants. How-ever, simple genetic investigations of possiblecommon pathways involving Egf1p, Hog1p,and Isw2p did not provide evidence for co-ordinate functions, other than induction ofchlamydospores, in other pathways (99).
The ability of C. albicans to form chlamy-dospores has been used as a clinical diagnos-tic tool for this fungal pathogen. The recentidentification of the closely related speciesC. dubliniensis established that C. albicansand C. dubliniensis could be distinguishedon the basis of chlamydospore formation ina medium, Guizotia abyssinica creatine agaror Staib agar, initially developed for theidentification of Cryptococcus neoformans. Thismedium induces chlamydospores in C. dublin-iensis but not in C. albicans (118). Because C. al-bicans and C. dubliniensis have similar genomes,efforts were made to identify the elements thatgenerated this difference in chlamydosporegeneration (119). A library of C. albicans se-quences was introduced into C. dubliniensisand the resulting transformants screened forrecombinants that were blocked in spore for-mation when cultured on Staib agar. Thisapproach identified the transcriptional reg-ulator Nrg1p (Table 1) as the key elementthat differentiated the behavior of C. albicans
www.annualreviews.org • Morphogenesis in C. albicans 541
Ann
u. R
ev. M
icro
biol
. 200
7.61
:529
-553
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Con
cord
ia U
nive
rsity
- M
ontr
eal o
n 05
/11/
11. F
or p
erso
nal u
se o
nly.

ANRV322-MI61-25 ARI 6 August 2007 18:18
and C. dubliniensis. Increased expression ofNRG1 in C. albicans relative to expression inC. dubliniensis blocked the ability of Staibagar to induce chlamydospore formation(119), and when an nrg1 mutant strain of C.albicans was tested for chlamydospore forma-tion on Staib agar, the mutant strain was in-duced as efficiently as C. dubliniensis. Thisresult establishes the level of expression ofNRG1 as a major determinant for the pro-duction of chlamydospores under the induc-ing conditions established by Staib agar. De-fects in downregulation of NRG1 expressionare predicted to interfere with chlamydosporeformation, and Rim101p, which is necessaryfor the proper timing of chlamydospore for-mation (99), is a repressor of NRG1 expression(85). However, the NRG1 expression level isnot the only determinative factor, because thenull mutant is not constitutive for chlamy-dospore formation, just permissive under thesignaling regime of Staib agar.
A careful analysis of the cell cycle dynam-ics of chlamydospore formation has providedstrong evidence for the uniqueness of thechlamydospore relative to yeast-form cells,pseudohyphal cells, and hyphal cells. The po-sitioning of the dividing nucleus has been animportant marker for the characterization ofthe cellular morphology pattern, with the nu-cleus migrating into the hyphal neck and thenre-entering the mother cell during hyphal di-vision, and dividing across the mother-budjunction in yeast and pseudohyphal cells (120).The chlamydospore shows yet another pat-tern of nuclear division. During the forma-tion of the chlamydospore, the nucleus divideswithin the suspensor cell and then one daugh-ter nucleus migrates into the chlamydospore(89).
This unique patterning of nuclear divisionis paralleled by the arrangement of septinsduring the formation of the chlamydospore.Initially the septins were found at the chlamy-dospore necks, similar to the situation for bud-ding cells, but later in the development of thespores, the septins were localized through-out the plasma membrane. Extensive filamen-
tous structures were formed by the later-stage septins; deletions of the CDC10- orCDC11-encoded septins perturbed the properdevelopment of the chlamydospore and thesuspensor cells (89). These distinctive behav-iors of the chlamydospore confirm that theyare a unique cellular form; however, theirphysiological role is still somewhat obscure.There is no evidence for meiosis occurringwithin the spore (89), although the septin fil-aments are somewhat suggestive of the modi-fied septin structures observed in the develop-ing spores in S. cerevisiae (45). Furthermore,the chlamydospores have not been shown torepresent a stable resting state. The forma-tion of these structures is characteristic ofthe pathogenic C. albicans and C. dublinien-sis; therefore their function may be some-how related to the commensal/opportunisticpathogen lifestyle of these organisms.
OTHER GROWTH MODES
Pseudohyphal Growth
C. albicans cells exhibit several patterns ofgrowth that involve extensive cell elonga-tion but do not involve the formation oftrue hyphae. The pseudohyphal pattern isthe best studied of these filamentous growthmodes. Pseudohyphal growth involves elon-gated cells that remain in chains but are sepa-rated by true constrictions, and not by septa. Ithas been controversial whether pseudohyphalgrowth is an intermediate stage in the yeast-to-hypha transition, but current work sug-gests that pseudohyphal cells represent a dis-crete cell growth pattern (120). The absenceof a Spitzenkorper, and the positioning ofnuclear division across the mother-daughterjunction, suggest that pseudohyphal growth iscloser to yeast growth than to hyphal growth.Major distinctions from the yeast form are theextended period of polarized growth exhib-ited by the pseudohyphal cells and the ten-dency for the cells to remain attached eventhough proper septa have formed betweenthem.
542 Whiteway
Ann
u. R
ev. M
icro
biol
. 200
7.61
:529
-553
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Con
cord
ia U
nive
rsity
- M
ontr
eal o
n 05
/11/
11. F
or p
erso
nal u
se o
nly.

ANRV322-MI61-25 ARI 6 August 2007 18:18
Some environmental conditions, such asmedium rich in phosphate (54) or in alka-nes (116), have been reported to stimulatethe pseudohyphal pattern of division. In addi-tion, a number of mutants lead to constitutivepseudohyphal growth. Several of these muta-tions are in genes involved in cell cycle reg-ulation, implicating this process in control ofthis morphogenetic state. This involvement islogical because the switch from yeast growthto pseudohyphal growth arises from a sub-tle modulation in the length of time the cellspends in the polarized growth mode relativeto the isotropic growth mode. Inactivation ofthe Fhk2p transcription factor (Table1) im-plicated in the regulation of mitotic cyclins,or of the Fkh2p target cyclin Clb4p, gen-erates cells that remain pseudohyphal underboth hyphae-inducing and yeast-growth con-ditions (18). Loss of Grr1p, an F box proteininvolved in the degradation of the G1 cyclinsCcn1p and Cln3p, leads to a pseudohyphalgrowth state (79). Perhaps extension of theG1 phase by stabilizing the G1 cyclins or re-ducing G2 cyclins can trigger the pseudohy-phal state. Another F box protein, Cdc4p, isimplicated in morphogenetic control. Loss ofCdc4p generates cells that are even more hy-phal than those created by loss of Grr1p, sug-gesting that a critical target of the Cdc4p ver-sion of the SCF (Skp1-Cullin-1/Cdc53-F-boxprotein) complex is necessary to block hyphalgrowth (5). In C. albicans the Nim1p-relatedkinases Gin4p and Hsl1p function in the reg-ulation of the pseudohyphal state; mutation ofeither kinase leads to constitutive pseudohy-phae (131).
In addition to cell cycle perturbations thatlead to constitutive pseudohyphae, defects inthe Tup1p regulatory circuit lead to this phe-notype (Table 1). Tup1p and the associatedprotein Ssn6p play important regulatory rolesin S. cerevisiae; they interact with differentDNA binding partners to provide a genericgene repression function (111). For specifictranscription modules the functional conse-quences of loss of Tup1p or Ssn6p can bedifferent; therefore Tup1p and Ssn6p do not
F box protein:component of theubiquitinationmachinery thatserves as part of thesubstrate recognitionsystem
Microarray:collection ofnucleotide probesattached to a solidsupport that allowssimultaneousassessment of all thetranscripts in the cell
provide completely overlapping functions. InC. albicans Tup1p inactivation causes cells toproliferate in a pseudohyphal growth mode(24). However, the ssn6 deletion mutant doesnot trigger equivalent pseudohyphal develop-ment in C. albicans (48, 56), although normalmorphology is perturbed. The global tran-scriptional consequences of the tup1 and ssn6mutations are different (48), suggesting thatthe Ssn6p corepressor is not a critical com-ponent of the Tup1p regulatory circuit thatrepresses the pseudohyphal growth pattern.An alternative tetratricopeptide repeat pro-tein, Tccp1p, may play the corepressor rolewith Tup1p in the repression of pseudohy-phal growth, as loss of Tccp1p creates sim-ilar phenotypes and affects patterns of geneexpression similar to loss of Tup1p (67). Itappears the Nrg1p DNA binding protein tar-gets the Tup1p(Tcc1p) corepressor to a varietyof promoters involved in regulation of mor-phogenesis, as loss of Nrg1p function leadsto constitutive pseudohyphal growth (48), andmicroarray analysis suggests that Tup1p andNrg1p influence distinct but overlapping genesets. A second DNA binding protein, Rfg1p(66), appears to function as a targeting ele-ment for Tup1p(Tcc1p); loss of Rfg1p leadsto a similar constitutive pseudohyphal phe-notype, but the networks of gene expressionmodulated by Nrg1p and Rfg1p are differ-ent. NRG1 is repressed by hyphal inducingconditions, and Rfg1p may be shut off post-translationally (65, 96), and thus inactivationof this negative regulatory circuit is a criti-cal component of morphogenetic control inC. albicans.
Hyperpolarized Buds
A second occasion in which C. albicans cellsexhibit a filamentous growth pattern distinctfrom hyphal growth occurs under certain cel-lular stresses or in the absence of certain geneproducts that influence the cell cycle. Treat-ment of yeast-form cells with the DNA syn-thesis inhibitor hydroxyurea (HU) resulted ina growth mode in which the bud continued
www.annualreviews.org • Morphogenesis in C. albicans 543
Ann
u. R
ev. M
icro
biol
. 200
7.61
:529
-553
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Con
cord
ia U
nive
rsity
- M
ontr
eal o
n 05
/11/
11. F
or p
erso
nal u
se o
nly.

ANRV322-MI61-25 ARI 6 August 2007 18:18
to elongate in the absence of further DNAreplication (7). This is different from the be-havior of S. cerevisiae cells treated with syn-thesis inhibitors, in which the cells arrest withno further growth of the bud. Treatment of C.albicans cells with nocodazole, another chem-ical that blocks cell cycle progression, alsocauses elongation of the bud in the absenceof further DNA replication (10). Several mu-tants generate similar bud elongation; in-activation of the CDC5 gene encoding theC. albicans polo kinase (7) and shutoff ofthe repair gene RAD52 (2) cause extensivebud growth. Blocking mitotic cyclin degra-dation by eliminating the destruction boxesof Clb2p or Clb4p traps cells in mito-sis and also triggers elongated bud growth(18), as does deletion or overproductionof a stabilized form of Sol1p (5). Reg-ulated repression of the cyclin-dependentkinase Cdc28p (121) and the essential B-typecyclin Clb2p (18) also generates extended budelongation. In contrast to deletion of othercell cycle genes that cause typical pseudohy-phal growth, the elongated bud phenotypeseems to be specific for depletion of essentialcell cycle genes.
The elongated buds resemble true hy-phae in that they maintain polarized growthat the tip and do not show periodic con-stricted growth as do pseudohyphae. Theyalso demonstrate nuclear movement out ofthe mother yeast cell and into the filament.Similar to pseudohyphae, a constriction ispresent at the junction between the yeast celland elongated bud, which is consistent withthe bud forming prior to the cell cycle ar-rest and the elongated growth mode. Theelongated buds eventually die, which is notunexpected given that they cannot continuethe cell cycle. However, the cells alter theirtranscription program during elongation. Forexample, time course microarray analysis ofHU-treated and CDC5-depleted cells demon-strated that the elongated buds express manyhyphae-specific genes, but at later stages ofelongation. More hyphae-specific genes were
turned on in the Cdc5p-depleted cells thanin HU-treated cells. The elongated buds maytherefore be capable of changing fate and be-coming more like true hyphae with continuedpolarized growth, suggesting that a feedbackmechanism may exist between the extent ofpolarized growth and activation of the hyphalsignaling pathways (7).
Although phenotypically similar, themolecular mechanisms underlying elongatedbud formation at different cell cycle stagesappear to be different. Transcriptional pro-filing of HU-treated and cdc5 mutant strainsshowed different expression patterns. TheHU-arrested cells require Ras1p for theextended bud growth, and the cdc5-arrestedcells require Bub2p function for the forma-tion of the extended buds (6). The differenttriggers stop the cells at different points inthe cell cycle, and the hyperpolarized buddoes not arise because the cells are trappedat a point corresponding to normal polarizedgrowth. Expression of the C. albicans BUD4(INT1) in S. cerevisiae triggers hyperpolarizedbud growth (4); it will be interesting toestablish if the process in C. albicans is Bud4pdependent.
The physiological relevance of the elon-gated bud morphology remains unclear be-cause it has been reported only in response tocreating cell cycle mutations or upon expo-sure to cell cycle-blocking drugs. However,the full suite of cell morphologies that C.albicans is capable of achieving in vivo is farfrom understood. Because the organism cangrow in such a morphology that eventuallyexpresses hyphal and virulence-specific fac-tors, an act that requires a large amount of re-sources, this growth mode may be importantfor pathogenicity and/or survival in the host.Certain stressful environments in the hostcould feed into the cell cycle and interfere withDNA replication and/or chromosome segre-gation. If such interference were linked withtriggering a highly polarized growth mode,the organism might escape the immediatestressful environment (7).
544 Whiteway
Ann
u. R
ev. M
icro
biol
. 200
7.61
:529
-553
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Con
cord
ia U
nive
rsity
- M
ontr
eal o
n 05
/11/
11. F
or p
erso
nal u
se o
nly.

ANRV322-MI61-25 ARI 6 August 2007 18:18
CONCLUSIONS
Morphological plasticity is a defining char-acteristic of the fungal pathogen C. albicans.A number of naturally occurring morpho-logical states are adapted to specific phys-iological functions of this organism. Themolecular circuits underlying some of thesemorphological states have been investigatedextensively, whereas others have been studiedat the molecular level only recently. Even theregulatory circuits controlling the yeast-to-hyphal transition can be reconsidered in re-spect to the more precise definitions of hyphaland pseudohyphal cells—-accurate interpre-tation of the networks will require a accuratedetermination of the cellular process being
regulated. In addition, the roles of the mor-phological regulators in defining cell formssuch as the opaque-form cell and the chlamy-dospore require more investigation. However,the rapidly developing tools for molecularand genomic studies in C. albicans are provid-ing important insights into the specific con-trols underlying both the unique morpholog-ical states and the interrelationships betweenthese states. Because the ability to regulatemorphology is a critical component of the vir-ulence of C. albicans, a detailed understandingof the roles of the morphological forms andhow they are regulated will provide impor-tant insights into strategies for controlling thispathogen.
SUMMARY POINTS
1. C. albicans has several distinct morphological forms. Improving rules defining thedifferent forms and improving analytical tools are permitting a more sophisticatedanalysis of the molecular bases for this morphological diversity.
2. When growing in the yeast form, C. albicans follows the pattern of growth of obligateyeasts such as S. cerevisiae. When growing in the hyphal form, it follows the pattern ofobligate hyphal organisms such as N. crassa. Understanding the mechanisms throughwhich environmental signals direct the choice of morphology is a central goal ofstudies on C. albicans morphogenesis.
3. The opaque state is controlled by the Wor1p transcription factor and is required forpheromone responsiveness and mating. Morphological changes directed by matingpheromones are a key aspect of the mating process.
4. The chlamydospore forms under adverse conditions through a process of cell divisiondistinct from that of yeast, hyphal, or pseudohyphal growth. The function of thechlamydospore is currently enigmatic; it may represent a resting state but evidencefor this is weak.
5. Pseudohyphae have a morphological appearance that falls between that of yeast-formand hyphae-form cells, but their cell division process resembles that of yeast, as thenucleus divides across the mother-daughter junction, and the cells lack a Spitzen-korper. Thus, it does not appear that pseudohyphae are a transitional state, but ratherrepresent a modification of the yeast form of cell division.
DISCLOSURE STATEMENT
The authors are not aware of any biases that might be perceived as affecting the objectivity ofthis review.
www.annualreviews.org • Morphogenesis in C. albicans 545
Ann
u. R
ev. M
icro
biol
. 200
7.61
:529
-553
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Con
cord
ia U
nive
rsity
- M
ontr
eal o
n 05
/11/
11. F
or p
erso
nal u
se o
nly.

ANRV322-MI61-25 ARI 6 August 2007 18:18
ACKNOWLEDGMENTS
We would like to thank present and past members of the Genetics group at BRI for discussionsand data. We are grateful to Peter Sudbery for providing unpublished information, and to AndreMigneault for graphics. We apologize to all researchers whose work has not been cited becauseof space limitations. Work in the Genetics group was supported by the National ResearchCouncil of Canada, by CIHR grant MOP42516, and by a CIHR team grant. This is NRCCpublication 47550.
LITERATURE CITED
1. Alonso-Monge R, Navarro-Garcia F, Roman E, Negredo AI, Eisman B, et al. 2003. TheHog1 mitogen-activated protein kinase is essential in the oxidative stress response andchlamydospore formation in Candida albicans. Eukaryot. Cell 2:351–61
2. Andaluz E, Ciudad T, Gomez-Raja J, Calderone R, Larriba G. 2006. Rad52 depletion inCandida albicans triggers both the DNA-damage checkpoint and filamentation accompa-nied by but independent of expression of hypha-specific genes. Mol. Microbiol. 59:1452–72
3. Anderson JM, Soll DR. 1987. Unique phenotype of opaque cells in the white-opaquetransition of Candida albicans. J. Bacteriol. 169:5579–88
4. Asleson CM, Bensen ES, Gale CA, Melms AS, Kurischko C, Berman J. 2001. Candidaalbicans INT1-induced filamentation in Saccharomyces cerevisiae depends on Sla2p. Mol.Cell. Biol. 21:1272–84
5. Atir-Lande A, Gildor T, Kornitzer D. 2005. Role for the SCFCDC4 ubiquitin ligase inCandida albicans morphogenesis. Mol. Biol. Cell 16:2772–85
6. Bachewich C, Nantel A, Whiteway M. 2005. Cell cycle arrest during S or M phasegenerates polarized growth via distinct signals in Candida albicans. Mol. Microbiol. 57:942–59
7. Bachewich C, Thomas DY, Whiteway M. 2003. Depletion of a polo-like kinase in Candidaalbicans activates cyclase-dependent hyphal-like growth. Mol. Biol. Cell 14:2163–80
8. Bachewich C, Whiteway M. 2005. Cyclin Cln3p links G1 progression to hyphal andpseudohyphal development in Candida albicans. Eukaryot. Cell 4:95–102
9. Bahn YS, Staab J, Sundstrom P. 2003. Increased high-affinity phosphodiesterase PDE2gene expression in germ tubes counteracts CAP1-dependent synthesis of cyclic AMP, lim-its hypha production and promotes virulence of Candida albicans. Mol. Microbiol. 50:391–409
10. Bai C, Ramanan N, Wang YM, Wang Y. 2002. Spindle assembly checkpoint compo-nent CaMad2p is indispensable for Candida albicans survival and virulence in mice. Mol.Microbiol. 45:31–44
11. Bardwell L. 2005. A walk-through of the yeast mating pheromone response pathway.Peptides 26:339–50
12. Barwell KJ, Boysen JH, Xu W, Mitchell AP. 2005. Relationship of DFG16 to the Rim101ppH response pathway in Saccharomyces cerevisiae and Candida albicans. Eukaryot. Cell 4:890–99
13. Bassilana M, Arkowitz RA. 2006. Rac1 and Cdc42 have different roles in Candida albicansdevelopment. Eukaryot. Cell 5:321–29
14. Bassilana M, Hopkins J, Arkowitz RA. 2005. Regulation of the Cdc42/Cdc24 GTPasemodule during Candida albicans hyphal growth. Eukaryot. Cell 4:588–603
15. Bennett RJ, Johnson AD. 2006. The role of nutrient regulation and the Gpa2 protein inthe mating pheromone response of C. albicans. Mol. Microbiol. 62:100–19
546 Whiteway
Ann
u. R
ev. M
icro
biol
. 200
7.61
:529
-553
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Con
cord
ia U
nive
rsity
- M
ontr
eal o
n 05
/11/
11. F
or p
erso
nal u
se o
nly.

ANRV322-MI61-25 ARI 6 August 2007 18:18
16. Bennett RJ, Miller MG, Chua PR, Maxon ME, Johnson AD. 2005. Nuclear fusion occursduring mating in Candida albicans and is dependent on the KAR3 gene. Mol. Microbiol.55:1046–59
17. Bennett RJ, Uhl MA, Miller MG, Johnson AD. 2003. Identification and characterizationof a Candida albicans mating pheromone. Mol. Cell. Biol. 23:8189–201
18. Bensen ES, Clemente-Blanco A, Finley KR, Correa-Bordes J, Berman J. 2005. Themitotic cyclins Clb2p and Clb4p affect morphogenesis in Candida albicans. Mol. Biol. Cell16:3387–400
19. Bensen ES, Filler SG, Berman J. 2002. A forkhead transcription factor is important fortrue hyphal as well as yeast morphogenesis in Candida albicans. Eukaryot. Cell 1:787–98
20. Bensen ES, Martin SJ, Li M, Berman J, Davis DA. 2004. Transcriptional profiling inCandida albicans reveals new adaptive responses to extracellular pH and functions forRim101p. Mol. Microbiol. 54:1335–51
21. Berman J. 2006. Morphogenesis and cell cycle progression in Candida albicans. Curr. Opin.Microbiol. 9:595–601
22. Biswas K, Rieger KJ, Morschhauser J. 2003. Functional analysis of CaRAP1, encodingthe repressor/activator protein 1 of Candida albicans. Gene 307:151–58
23. Bockmuhl DP, Ernst JF. 2001. A potential phosphorylation site for an A-type kinasein the Efg1 regulator protein contributes to hyphal morphogenesis of Candida albicans.Genetics 157:1523–30
24. Braun BR, Johnson AD. 1997. Control of filament formation in Candida albicans by thetranscriptional repressor TUP1. Science 277:105–9
25. Braun BR, Kadosh D, Johnson AD. 2001. NRG1, a repressor of filamentous growth inC. albicans, is down-regulated during filament induction. EMBO J. 20:4753–61
26. Brown DH Jr, Giusani AD, Chen X, Kumamoto CA. 1999. Filamentous growth of Can-dida albicans in response to physical environmental cues and its regulation by the uniqueCZF1 gene. Mol. Microbiol. 34:651–62
27. Butty AC, Pryciak PM, Huang LS, Herskowitz I, Peter M. 1998. The role of Far1pin linking the heterotrimeric G protein to polarity establishment proteins during yeastmating. Science 282:1511–16
28. Caldwell GA, Naider F, Becker JM. 1995. Fungal lipopeptide mating pheromones: amodel system for the study of protein prenylation. Microbiol. Rev. 59:406–22
29. Cao F, Lane S, Raniga PP, Lu Y, Zhou Z, et al. 2006. The Flo8 transcription factor isessential for hyphal development and virulence in Candida albicans. Mol. Biol. Cell 17:295–307
30. Cao F, Li J, Yan X, Wu Y, Zhang D. 2005. Relationship between host survival and thetype of immune response in different organs during disseminated candidiasis. J. HuazhongUniv. Sci. Technol. Med. Sci. 25:141–384
31. Casamayor A, Snyder M. 2002. Bud-site selection and cell polarity in budding yeast. Curr.Opin. Microbiol. 5:179–86
32. Chapa y Lazo B, Bates S, Sudbery P. 2005. The G1 cyclin Cln3 regulates morphogenesisin Candida albicans. Eukaryot. Cell 4:90–94
33. Chen J, Chen J, Lane S, Liu H. 2002. A conserved mitogen-activated protein kinasepathway is required for mating in Candida albicans. Mol. Microbiol. 46:1335–44
34. Cloutier M, Castilla R, Bolduc N, Zelada A, Martineau P, et al. 2003. The two isoformsof the cAMP-dependent protein kinase catalytic subunit are involved in the control ofdimorphism in the human fungal pathogen Candida albicans. Fungal Genet. Biol. 38:133–41
www.annualreviews.org • Morphogenesis in C. albicans 547
Ann
u. R
ev. M
icro
biol
. 200
7.61
:529
-553
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Con
cord
ia U
nive
rsity
- M
ontr
eal o
n 05
/11/
11. F
or p
erso
nal u
se o
nly.

ANRV322-MI61-25 ARI 6 August 2007 18:18
35. Cornet M, Bidard F, Schwarz P, Da Costa G, Blanchin-Roland S, et al. 2005. Deletionsof endocytic components VPS28 and VPS32 affect growth at alkaline pH and virulencethrough both RIM101-dependent and RIM101-independent pathways in Candida albi-cans. Infect. Immun. 73:7977–87
36. Court H, Sudbery P. 2006. Regulation of Cdc42 GTPase activity in the formation ofhyphae in Candida albicans. Mol. Biol. Cell 18:265–81
37. Identifies forthe first time thebehavior of theSpitzenkorper in C.
albicans.
37. Crampin H, Finley K, Gerami-Nejad M, Court H, Gale C, et al. 2005. Candida
albicans hyphae have a Spitzenkorper that is distinct from the polarisome found inyeast and pseudohyphae. J. Cell Sci. 118:2935–47
38. Csank C, Schroppel K, Leberer E, Harcus D, Mohamed O, et al. 1998. Roles of the Can-dida albicans mitogen-activated protein kinase homolog, Cek1p, in hyphal developmentand systemic candidiasis. Infect. Immun. 66:2713–21
39. Daniels KJ, Srikantha T, Lockhart SR, Pujol C, Soll DR. 2006. Opaque cells signal whitecells to form biofilms in Candida albicans. EMBO J. 25:2240–52
40. Davis D, Wilson RB, Mitchell AP. 2000. RIM101-dependent and-independent pathwaysgovern pH responses in Candida albicans. Mol. Cell. Biol. 20:971–78
41. Dignard D, El-Naggar AL, Logue ME, Butler G, Whiteway M. 2007. Identificationand characterization of MFA1, the gene encoding Candida albicans a-factor pheromone.Eukaryot. Cell 6:487–94
42. Doedt T, Krishnamurthy S, Bockmuhl DP, Tebarth B, Stempel C, et al. 2004. APSES pro-teins regulate morphogenesis and metabolism in Candida albicans. Mol. Biol. Cell 15:3167–80
43. Douglas LM, Alvarez FJ, McCreary C, Konopka JB. 2005. Septin function in yeast modelsystems and pathogenic fungi. Eukaryot. Cell 4:1503–12
44. Fabry W, Schmid EN, Schraps M, Ansorg R. 2003. Isolation and purification of chlamy-dospores of Candida albicans. Med. Mycol. 41:53–58
45. Fares H, Goetsch L, Pringle JR. 1996. Identification of a developmentally regulatedseptin and involvement of the septins in spore formation in Saccharomyces cerevisiae. J. CellBiol. 132:399–411
46. Finley KR, Berman J. 2005. Microtubules in Candida albicans hyphae drive nuclear dy-namics and connect cell cycle progression to morphogenesis. Eukaryot. Cell 4:1697–711
47. Fukata M, Nakagawa M, Kaibuchi K. 2003. Roles of Rho-family GTPases in cell polar-isation and directional migration. Curr. Opin. Cell Biol. 15:590–97
48. This detailedanalysis of thetranscriptional andmorphologicalconsequences ofmutations inTup1p, Nrg1p andSsn6p provides anoverview of thisimportantregulatorynetwork.
48. Garcia-Sanchez S, Mavor AL, Russell CL, Argimon S, Dennison P, et al. 2005.Global roles of Ssn6 in Tup1- and Nrg1-dependent gene regulation in the fungalpathogen, Candida albicans. Mol. Biol. Cell 16:2913–25
49. Giusani AD, Vinces M, Kumamoto CA. 2002. Invasive filamentous growth of Candidaalbicans is promoted by Czf1p-dependent relief of Efg1p-mediated repression. Genetics160:1749–53
50. Harcus D, Nantel A, Marcil A, Rigby T, Whiteway M. 2004. Transcription profiling ofcyclic AMP signaling in Candida albicans. Mol. Biol. Cell 15:4490–99
51. Harris SD, Read ND, Roberson RW, Shaw B, Seiler S, et al. 2005. Polarisome meetsSpitzenkorper: microscopy, genetics, and genomics converge. Eukaryot. Cell 4:225–29
52. Hazan I, Liu H. 2002. Hyphal tip-associated localization of Cdc42 is F-actin dependentin Candida albicans. Eukaryot. Cell 1:856–64
53. Hazan I, Sepulveda-Becerra M, Liu H. 2002. Hyphal elongation is regulated indepen-dently of cell cycle in Candida albicans. Mol. Biol. Cell 13:134–45
548 Whiteway
Ann
u. R
ev. M
icro
biol
. 200
7.61
:529
-553
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Con
cord
ia U
nive
rsity
- M
ontr
eal o
n 05
/11/
11. F
or p
erso
nal u
se o
nly.

ANRV322-MI61-25 ARI 6 August 2007 18:18
54. Hornby JM, Dumitru R, Nickerson KW. 2004. High phosphate (up to 600 mM) inducespseudohyphal development in five wild type Candida albicans. J. Microbiol. Methods 56:119–24
55. Huang G, Wang H, Chou S, Nie X, Chen J, Liu H. 2006. Bistable expression of WOR1,a master regulator of white-opaque switching in Candida albicans. Proc. Natl. Acad. Sci.USA 103:12813–18
56. Hwang CS, Oh JH, Huh WK, Yim HS, Kang SO. 2003. Ssn6, an important factor ofmorphological conversion and virulence in Candida albicans. Mol. Microbiol. 47:1029–43
57. Inglis DO, Johnson AD. 2002. Ash1 protein, an asymmetrically localized transcriptionalregulator, controls filamentous growth and virulence of Candida albicans. Mol. Cell Biol.22:8669–80
58. Irazoqui JE, Lew DJ. 2004. Polarity establishment in yeast. J. Cell Sci. 117:2169–7159. Jaffe AB, Hall A. 2005. Rho GTPases: biochemistry and biology. Annu. Rev. Cell Dev.
Biol. 21:247–6960. Jansons VK, Nickerson WJ. 1970. Induction, morphogenesis, and germination of the
chlamydospore of Candida albicans. J. Bacteriol. 104:910–2161. Johnson A. 2003. The biology of mating in Candida albicans. Nat. Rev. Microbiol. 1:106–1662. Johnson DC, Cano KE, Kroger EC, McNabb DS. 2005. Novel regulatory function for
the CCAAT-binding factor in Candida albicans. Eukaryot. Cell 4:1662–7663. Johnson DI. 1999. Cdc42: an essential Rho-type GTPase controlling eukaryotic cell
polarity. Microbiol. Mol. Biol. Rev. 63:54–10564. Jung WH, Stateva LI. 2003. The cAMP phosphodiesterase encoded by CaPDE2 is re-
quired for hyphal development in Candida albicans. Microbiology 149:2961–7665. Kadosh D, Johnson AD. 2001. Rfg1, a protein related to the Saccharomyces cerevisiae
hypoxic regulator Rox1, controls filamentous growth and virulence in Candida albicans.Mol. Cell. Biol. 21:2496–505
66. Kadosh D, Johnson AD. 2005. Induction of the Candida albicans filamentous growthprogram by relief of transcriptional repression: a genome-wide analysis. Mol. Biol. Cell16:2903–12
67. Kaneko A, Umeyama T, Utena-Abe Y, Yamagoe S, Niimi M, Uehara Y. 2006. Tcc1p,a novel protein containing the tetratricopeptide repeat motif, interacts with Tup1p toregulate morphological transition and virulence in Candida albicans. Eukaryot. Cell 5:1894–905
68. Kelly MT, MacCallum DM, Clancy SD, Odds FC, Brown AJ, Butler G. 2004. TheCandida albicans CaACE2 gene affects morphogenesis, adherence and virulence. Mol.Microbiol. 53:969–83
69. Khalaf RA, Zitomer RS. 2001. The DNA binding protein Rfg1 is a repressor of filamen-tation in Candida albicans. Genetics 157:1503–12
70. Knaus M, Wiget P, Shimada Y, Peter M. 2005. Control of cell polarity in responseto intra- and extracellular signals in budding yeast. Novartis Found. Symp. 269:47–54;discussion 54–58, 223–30
71. Kohler JR, Fink GR. 1996. Candida albicans strains heterozygous and homozygous for mu-tations in mitogen-activated protein kinase signaling components have defects in hyphaldevelopment. Proc. Natl. Acad. Sci. USA 93:13223–28
72. Kumamoto CA, Vinces MD. 2005. Alternative Candida albicans lifestyles: growth on sur-faces. Annu. Rev. Microbiol. 59:113–33
73. Lachke SA, Srikantha T, Soll DR. 2003. The regulation of EFG1 in white-opaque switch-ing in Candida albicans involves overlapping promoters. Mol. Microbiol. 48:523–36
www.annualreviews.org • Morphogenesis in C. albicans 549
Ann
u. R
ev. M
icro
biol
. 200
7.61
:529
-553
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Con
cord
ia U
nive
rsity
- M
ontr
eal o
n 05
/11/
11. F
or p
erso
nal u
se o
nly.

ANRV322-MI61-25 ARI 6 August 2007 18:18
74. Lan CY, Newport G, Murillo LA, Jones T, Scherer S, et al. 2002. Metabolic specializa-tion associated with phenotypic switching in Candida albicans. Proc. Natl. Acad. Sci. USA99:14907–12
75. Lane S, Zhou S, Pan T, Dai Q, Liu H. 2001. The basic helix-loop-helix transcriptionfactor Cph2 regulates hyphal development in Candida albicans partly via TEC1. Mol. Cell.Biol. 21:6418–28
76. Leberer E, Harcus D, Broadbent ID, Clark KL, Dignard D, et al. 1996. Signal trans-duction through homologs of the Ste20p and Ste7p protein kinases can trigger hyphalformation in the pathogenic fungus Candida albicans. Proc. Natl. Acad. Sci. USA 93:13217–22
77. Leberer E, Harcus D, Dignard D, Johnson L, Ushinsky S, et al. 2001. Ras links cellu-lar morphogenesis to virulence by regulation of the MAP kinase and cAMP signallingpathways in the pathogenic fungus Candida albicans. Mol. Microbiol. 42:673–87
78. Li M, Martin SJ, Bruno VM, Mitchell AP, Davis DA. 2004. Candida albicans Rim13p,a protease required for Rim101p processing at acidic and alkaline pHs. Eukaryot. Cell3:741–51
79. Li WJ, Wang YM, Zheng XD, Shi QM, Zhang TT, et al. 2006. The F-box protein Grr1regulates the stability of Ccn1, Cln3 and Hof1 and cell morphogenesis in Candida albicans.Mol. Microbiol. 62:212–26
80. Liu H. 2001. Transcriptional control of dimorphism in Candida albicans. Curr. Opin. Mi-crobiol. 4:728–35
81. Lo HJ, Kohler JR, DiDomenico B, Loebenberg D, Cacciapuoti A, Fink GR. 1997. Non-filamentous C. albicans mutants are avirulent. Cell 90:939–49
82. Thecharacteristics ofthe cellsundergoing themating process aredetailed in thispaper.
82. Lockhart SR, Daniels KJ, Zhao R, Wessels D, Soll DR. 2003. Cell biology of matingin Candida albicans. Eukaryot. Cell 2:49–61
83. Lockhart SR, Zhao R, Daniels KJ, Soll DR. 2003. Alpha-pheromone-induced “shmoo-ing” and gene regulation require white-opaque switching during Candida albicans mating.Eukaryot. Cell 2:847–55
84. Loeb JD, Sepulveda-Becerra M, Hazan I, Liu H. 1999. A G1 cyclin is necessary formaintenance of filamentous growth in Candida albicans. Mol. Cell. Biol. 19:4019–27
85. Lotz H, Sohn K, Brunner H, Muhlschlegel FA, Rupp S. 2004. RBR1, a novel pH-regulated cell wall gene of Candida albicans, is repressed by RIM101 and activated byNRG1. Eukaryot. Cell 3:776–84
86. Magee BB, Legrand M, Alarco AM, Raymond M, Magee PT. 2002. Many of the genesrequired for mating in Saccharomyces cerevisiae are also required for mating in Candidaalbicans. Mol. Microbiol. 46:1345–51
87. Maidan MM, De Rop L, Serneels J, Exler S, Rupp S, et al. 2005. The G protein-coupledreceptor Gpr1 and the Galpha protein Gpa2 act through the cAMP-protein kinase Apathway to induce morphogenesis in Candida albicans. Mol. Biol. Cell 16:1971–86
88. Martin R, Walther A, Wendland J. 2005. Ras1-induced hyphal development in Candidaalbicans requires the formin Bni1. Eukaryot. Cell 4:1712–24
89. This workprovides a carefulexamination of theformation of thechlamydospore anddevelops aframework for theanalysis ofchlamydosporebiology.
89. Martin SW, Douglas LM, Konopka JB. 2005. Cell cycle dynamics and quorumsensing in Candida albicans chlamydospores are distinct from budding and hyphalgrowth. Eukaryot. Cell 4:1191–202
90. Mendenhall MD, Hodge AE. 1998. Regulation of Cdc28 cyclin-dependent protein kinaseactivity during the cell cycle of the yeast Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev.62:1191–243
550 Whiteway
Ann
u. R
ev. M
icro
biol
. 200
7.61
:529
-553
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Con
cord
ia U
nive
rsity
- M
ontr
eal o
n 05
/11/
11. F
or p
erso
nal u
se o
nly.

ANRV322-MI61-25 ARI 6 August 2007 18:18
91. Michel S, Ushinsky S, Klebl B, Leberer E, Thomas D, et al. 2002. Generation of con-ditional lethal Candida albicans mutants by inducible deletion of essential genes. Mol.Microbiol. 46:269–80
92. Miller MG, Johnson AD. 2002. White-opaque switching in Candida albicans is controlledby mating-type locus homeodomain proteins and allows efficient mating. Cell 110:293–302
93. Miller SE, Spurlock BO, Michaels GE. 1974. Electron microscopy of young Candidaalbicans chlamydospores. J. Bacteriol. 119:992–99
94. Momany M. 2002. Polarity in filamentous fungi: establishment, maintenance and newaxes. Curr. Opin. Microbiol. 5:580–55
95. Mulhern SM, Logue ME, Butler G. 2006. Candida albicans transcription factor Ace2regulates metabolism and is required for filamentation in hypoxic conditions. Eukaryot.Cell 5:2001–13
96. Murad AM, Leng P, Straffon M, Wishart J, Macaskill S, et al. 2001. NRG1 represses yeast-hypha morphogenesis and hypha-specific gene expression in Candida albicans. EMBO J.20:4742–52
97. Naider F, Becker JM. 2004. The alpha-factor mating pheromone of Saccharomyces cere-visiae: a model for studying the interaction of peptide hormones and G protein-coupledreceptors. Peptides 25:1441–63
98. Nantel A, Dignard D, Bachewich C, Harcus D, Marcil A, et al. 2002. Transcriptionprofiling of Candida albicans cells undergoing the yeast-to-hyphal transition. Mol. Biol.Cell 13:3452–65
99. Nobile CJ, Bruno VM, Richard ML, Davis DA, Mitchell AP. 2003. Genetic control ofchlamydospore formation in Candida albicans. Microbiology 149:3629–37
100. Oberholzer U, Iouk TL, Thomas DY, Whiteway M. 2004. Functional characterizationof myosin I tail regions in Candida albicans. Eukaryot. Cell 3:1272–86
101. Oberholzer U, Nantel A, Berman J, Whiteway M. 2006. Transcript profiles of Candidaalbicans cortical actin patch mutants reflect their cellular defects: contribution of theHog1p and Mkc1p signaling pathways. Eukaryot. Cell 5:1252–65
102. Panwar SL, Legrand M, Dignard D, Whiteway M, Magee PT. 2003. MFalpha1, the geneencoding the alpha mating pheromone of Candida albicans. Eukaryot. Cell 2:1350–60
103. Park YN, Morschhauser J. 2005. Candida albicans MTLalpha tup1Delta mutants can re-versibly switch to mating-competent, filamentous growth forms. Mol. Microbiol. 58:1288–302
104. Rida PC, Nishikawa A, Won GY, Dean N. 2006. Yeast-to-hyphal transition triggersformin-dependent Golgi localization to the growing tip in Candida albicans. Mol. Biol. Cell17:4364–78
105. Rocha CR, Schroppel K, Harcus D, Marcil A, Dignard D, et al. 2001. Signaling throughadenylyl cyclase is essential for hyphal growth and virulence in the pathogenic fungusCandida albicans. Mol. Biol. Cell 12:3631–43
106. Rottmann M, Dieter S, Brunner H, Rupp S. 2003. A screen in Saccharomyces cerevisiaeidentified CaMCM1, an essential gene in Candida albicans crucial for morphogenesis. Mol.Microbiol. 47:943–59
107. Schweizer A, Rupp S, Taylor BN, Rollinghoff M, Schroppel K. 2000. The TEA/ATTStranscription factor CaTec1p regulates hyphal development and virulence in Candidaalbicans. Mol. Microbiol. 38:435–45
108. Sherlock G, Bahman AM, Mahal A, Shieh JC, Ferreira M, Rosamond J. 1994. Molecularcloning and analysis of CDC28 and cyclin homologues from the human fungal pathogenCandida albicans. Mol. Gen. Genet. 245:716–23
www.annualreviews.org • Morphogenesis in C. albicans 551
Ann
u. R
ev. M
icro
biol
. 200
7.61
:529
-553
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Con
cord
ia U
nive
rsity
- M
ontr
eal o
n 05
/11/
11. F
or p
erso
nal u
se o
nly.

ANRV322-MI61-25 ARI 6 August 2007 18:18
109. Sheu YJ, Barral Y, Snyder M. 2000. Polarized growth controls cell shape and bipolar budsite selection in Saccharomyces cerevisiae. Mol. Cell. Biol. 20:5235–47
110. Slutsky B, Staebell M, Anderson J, Risen L, Pfaller M, Soll DR. 1987. “White-opaquetransition”: a second high-frequency switching system in Candida albicans. J. Bacteriol.169:189–97
111. Smith RL, Johnson AD. 2000. Turning genes off by Ssn6-Tup1: a conserved system oftranscriptional repression in eukaryotes. Trends Biochem. Sci. 25:325–30
112. Soll DR. 2004. Mating-type locus homozygosis, phenotypic switching and mating: aunique sequence of dependencies in Candida albicans. Bioessays 26:10–20
113. Song Y, Kim JY. 2006. Role of CaBud6p in the polarized growth of Candida albicans. J.Microbiol. 44:311–19
114. Sonneborn A, Bockmuhl DP, Ernst JF. 1999. Chlamydospore formation in Candida al-bicans requires the Efg1p morphogenetic regulator. Infect. Immun. 67:5514–17
115. Sonneborn A, Tebarth B, Ernst JF. 1999. Control of white-opaque phenotypic switchingin Candida albicans by the Efg1p morphogenetic regulator. Infect. Immun. 67:4655–60
116. Sorkhoh NA, Ghannoum MA, Ibrahim AS, Stretton RJ, Radwan SS. 1990. Growth ofCandida albicans on hydrocarbons: influence on lipids and sterols. Microbios 64:159–71
117. Srikantha T, Tsai LK, Daniels K, Soll DR. 2000. EFG1 null mutants of Candida albi-cans switch but cannot express the complete phenotype of white-phase budding cells. J.Bacteriol. 182:1580–91
118. Staib P, Morschhauser J. 1999. Chlamydospore formation on Staib agar as a species-specific characteristic of Candida dubliniensis. Mycoses 42:521–24
119. Staib P, Morschhauser J. 2005. Differential expression of the NRG1 repressor controlsspecies-specific regulation of chlamydospore development in Candida albicans and Candidadubliniensis. Mol. Microbiol. 55:637–52
120. Sudbery P, Gow N, Berman J. 2004. The distinct morphogenic states of Candida albicans.Trends Microbiol. 12:317–24
121. Umeyama T, Kaneko A, Niimi M, Uehara Y. 2006. Repression of CDC28 reducesthe expression of the morphology-related transcription factors, Efg1p, Nrg1p, Rbf1p,Rim101p, Fkh2p and Tec1p and induces cell elongation in Candida albicans. Yeast 23:537–52
122. This paperdescribes theconstruction andcharacterization ofconditionalmutants in thepolarity regulatingGTPase Cdc42p. 122. Ushinsky SC, Harcus D, Ash J, Dignard D, Marcil A, et al. 2002. CDC42 is required
for polarized growth in human pathogen Candida albicans. Eukaryot. Cell 1:95–104
123. This workbegins to unravelthe regulatorycircuit controllinghyphaldevelopmentduring embeddedgrowth.
123. Vinces MD, Haas C, Kumamoto CA. 2006. Expression of the Candida albicans mor-phogenesis regulator gene CZF1 and its regulation by Efg1p and Czf1p. Eukaryot.
Cell 5:825–35124. Virag A, Harris SD. 2006. Functional characterization of Aspergillus nidulans homologues
of Saccharomyces cerevisiae Spa2 and Bud6. Eukaryot. Cell 5:881–95125. Virag A, Harris SD. 2006. The Spitzenkorper: a molecular perspective. Mycol. Res. 110:4–
13126. Walther A, Wendland J. 2004. Polarized hyphal growth in Candida albicans requires the
Wiskott-Aldrich Syndrome protein homolog Wal1p. Eukaryot. Cell 3:471–82127. Wang A, Lane S, Tian Z, Sharon A, Hazan I, Liu H. 2007. Temporal and spatial control
of HGC1 expression results in Hgc1 localization to the apical cell of hyphae in Candidaalbicans. Eukaryot. Cell 6:253–61
128. Wendland J. 2001. Comparison of morphogenetic networks of filamentous fungi andyeast. Fungal Genet. Biol. 34:63–82
552 Whiteway
Ann
u. R
ev. M
icro
biol
. 200
7.61
:529
-553
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Con
cord
ia U
nive
rsity
- M
ontr
eal o
n 05
/11/
11. F
or p
erso
nal u
se o
nly.

ANRV322-MI61-25 ARI 6 August 2007 18:18
129. Whiteway M, Dignard D, Thomas DY. 1992. Dominant negative selection of heterolo-gous genes: isolation of Candida albicans genes that interfere with Saccharomyces cerevisiaemating factor-induced cell cycle arrest. Proc. Natl. Acad. Sci. USA 89:9410–14
130. Whiteway M, Oberholzer U. 2004. Candida morphogenesis and host-pathogen interac-tions. Curr. Opin. Microbiol. 7:350–57
131. Wightman R, Bates S, Amornrrattanapan P, Sudbery P. 2004. In Candida albicans, theNim1 kinases Gin4 and Hsl1 negatively regulate pseudohypha formation and Gin4 alsocontrols septin organization. J. Cell Biol. 164:581–91
132. Woo M, Lee K, Song K. 2003. MYO2 is not essential for viability, but is required for po-larized growth and dimorphic switches in Candida albicans. FEMS Microbiol. Lett. 218:195–202
133. Xu W, Smith FJ Jr, Subaran R, Mitchell AP. 2004. Multivesicular body-ESCRT compo-nents function in pH response regulation in Saccharomyces cerevisiae and Candida albicans.Mol. Biol. Cell 15:5528–37
134. Zheng X, Wang Y, Wang Y. 2004. Hgc1, a novel hypha-specific G1 cyclin-related proteinregulates Candida albicans hyphal morphogenesis. EMBO J. 23:1845–56
135. Zheng XD, Wang YM, Wang Y. 2003. CaSPA2 is important for polarity establishmentand maintenance in Candida albicans. Mol. Microbiol. 49:1391–405
136. This paperand Reference 55identify andcharacterize theWor1ptranscription factorthat regulates theopaque state.
136. Zordan RE, Galgoczy DJ, Johnson AD. 2006. Epigenetic properties of white-opaque switching in Candida albicans are based on a self-sustaining transcriptionalfeedback loop. Proc. Natl. Acad. Sci. USA 103:12807–12
RELATED RESOURCES
Li L, Wright SJ, Krystofova S, Park G, Borkovich KA. 2007. Heterotrimeric G protein sig-naling in filamentous fungi. Annu. Rev. Microbiol. 61:423–52
www.annualreviews.org • Morphogenesis in C. albicans 553
Ann
u. R
ev. M
icro
biol
. 200
7.61
:529
-553
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Con
cord
ia U
nive
rsity
- M
ontr
eal o
n 05
/11/
11. F
or p
erso
nal u
se o
nly.
Recommended