Embed Size (px)
Citation preview

11
111 Seminário Internacional sobreBiotecnologia na Agroindústria Cafeeira
111 International Seminar onBiotechnology in the Coffee Agroindustry
24 - 28 de maio May 24 - 28, 1999Londrina - PR - Brasil
Anais/Proceedings
- UFPR

R551a RIEDE. C.R. coord.Anais do 3. seminário internacional sobre biotecnologia naagroindústria cafeeira! coord. por Carlos Roberto Riede e outrosLondrina: IAPAR/IRD. 2000.513p.
l rCafé-Congressos. 2.Agroindústria. 3.Biotecnologia. l.Sera,Turnoru, coord. Il.Soccol. Carlos Ricardo. coord. Ill.Roussos,Sevastianos, coord. IV. Instituto Agronômico do Paraná. Londrina.PRo V.U niversidade Federal do Paraná. Curitiba. PRo Vl.Institut deRecherche pour le Développernent. Montpellier. França. VII.Título.
CDD 633.73AGRIS FOI
E212120

111 Seminário Internacional sobre Biotecnologia na Agroindústria CafeeiraIII Intemational Seminar on Biotechnology in the Coffee Agroindustry
ANAIS DO IH SIBAC
Coordenadores: Carlos Roberto Riede
Tumoru Sera
Carlos Ricardo Soccol
Sevastianos Roussos
Digitação e adequação de textos: Marcos Zorzenon Alteia
Emerson Tadashi Nakamura
Editores: Instituto Agronômico do Paraná - IAPAR
Institut de Recherche pour le Développement - IRD
A responsabilidade da comissão editorial limita-se a adequação dos trabalhos asnormas editoriais estabelecidas.
A ortografia, a correção gramatical e o conteúdo dos trabalhos aqui publicados são deresponsabilidade dos autores.
24 - 28 de Maio, Londrina - PR - Brasil.

111 Seminário Internacional sobre Biotecnologia na Agroindústria Cafeeira111 International Seminar on Biotechnology in the Coffee Agroindustry
Comissão organizadora• Carlos R. Soccol (UNIV. FED. DO PARANÁ - Brasil)• Sevastianos Roussos (IRD -França)• Tumoru Sera (lAPAR - Brasil)
• COMISSÃO EXECUTIVA:• COORDENADOR: Tumoru Sera• SUB-COMISSÃO DE PROGRAMAÇÃO TECNICO-CIENTIFICA: Carlos R. Soccol; Carlos R.
Riede; Luiz F. Pereira; Luiz G. E. Vieira; Sevastianos Roussos; Tumoru Sera.• SUB-COMISSÃO DE CAPTAÇÃO DE RECURSOS: Armando Androcioli fO.; Francisco B. Lima;
Luiz R. Ferrari; Samir Cury.• SUB-COMISSÃO DE MARKETING: Marcos V. F. Martins; Michele Peres.• SUB-COMISSÃO DE IMPRENSA: Oswaldo Petrin.• SUB-COMISSÃO DE POSTERS: Maria C. L. L. Dias; Maria L. Crochemore.• SUB-COMISSÃO DE RELAÇÕES PÚBLICAS: Luiz. F. Kalinowsky.• SUB-COMISSÃO DE INFORMÁTICA: Welfrid Stenzel• SUB-COMISSÃO SERVIÇOS GRÁFICOS: Tadeu Sakiyama.• SUB-COMISSÃO DE INFRAESTRUTURA: Júlio C. D. Chaves; João da Mata Siqueira; Claudomir
A. da Silva.• SUB-COMISSÃO DE CONTACTOS INSTITUCIONAIS: Florindo Dalberto; Marcos Krieger;
Rogério M. L. Cardoso.• SUB-COMISSÃO DE FINANÇAS: José Pereira da Silva; Júlio César Chaves.• SUB-COMISSÃO DE "DIA DE CAMPO": Armando Androcioli fO.; José A. Azevedo; Marcos.
Pavan; Paulo H. Caramori; Rodolfo M. de Carvalho
COMISSÃO TÉCNICO-CIENTÍFICA:Alemar B. Rena (CBP&D CAFÉ/Brasil), Alex Bustillo-Pardey (CENICAFE / Colômbia), Antônio P. Nacif(CBP&DCAFÉ/Brasil), Ashok Pandey (CEFRI/Índia), Christopher Augur (lRD / México), DanielMartinez-Carrera (COL.POST / México), Gerardo Saucedo-Castafieda (UAM / México), German MorenoRuiz (CENICAFE / Colômbia), Gustavo Viniegra-González (UAM / México), Isabelle Gaime-Perraud(IRD / México), Ivonc Ferrao (DG 12/UNIÃO EUROPÉIA), Jaime Zuluaga-Vasco (CENICAFE /Colômbia), Joseph Le Bars (lNRA / França), Marc Berthouly (CIRAD / França), Moacyr Paschoal (UFLA/ Brasil), Maurice Lourd (lRD / Brasil), Ney Sakiyama (UFV / Brasil), Oliveiro Guerreiro (lAC / Brasil),Philippe Lashermes (lRD / França).
SECRETARIA EXECUTIVA:• Secretaria: Emerson Tadashi Nakarnura; Françoise Simoni; Marcos Z. Altéia; Rubens Sacchetto
Fone: (043) 376.2294 Fax: (043) 376.2101 E-mail: [email protected]

SUMÁRIO
Sessão Inicial - PalestraPág.
B10TECHNOLOGY ANO THE FUTURE OF COFFEE PRODUCTION.Gustavo Viniegra González .
Sessão 1 - CLONAGEM E CULTURA DE TECIDOS
A - Palestras
B10TECNOLOGÍAS APLICADAS AL MEJORAMIENTO GENÉTICO DEL CAFETO.Marc Berthouly 9
SOMATlC EMBRYOGENESIS OF COFFEE.Marc Berthouly and Hervé Etienne 23
DESENVOLVIMENTO DE CAFEEIROS GENETICAMENTE MODIFICADOS COMVISTAS À UNIFORMIDADE DE MATURAÇÃO DOS FRUTOS.Luís Felipe Pereira, Luís Gonzaga Vieira e Adilson Kenji Kobayashi . 37
PROGRAMA BRASILEIRO DE BIOTECNOLOGIA APLICADA AO CAFEEIRO.Alemar Braga Rena e Antonio de Pádua Nacif 43
APLlCACIONES DE LA BIOTECNOLOGÍA AL MEJORAMIENTO GENÉTICO DEL CAFÉEN COLOMBlA.Germán Moreno, Myriam de Pefia.Ricardo Acufía, Maria Eugenia Aponte, Juan Carlos Herrera,Diana Molina, Alvaro Gaitán, Marco Cristancho, Pilar Moncada.Carrnenza Góngora y GabrielCadena 47
AVANÇOS NA OBTENÇÃO DE DI-HAPLOIDES EM CULTIVARES DE Coflee arabica L.Maria Filomena Carneiro 55
CUSTO COMPARATIVO DE DIFERENTES TÉCNICAS DE CLONAGEM.Maro R. Sondahl, Clemencia N. Sondahl e Wallace Gonçalves 59
COFFEE (Coflea SP.) GENETlC TRANSFORMATION FOR INSECT RESISTANCE.Magali Dufour, Thierry Leroy, Cathy Carosco-Lacornbe, René Philippe and Catherine Fenouillet........ 507
B - Posters
EFECTO DEL GENOTlPO Y ÉPOCA DE RECOLECCIÓN DEL MATERIAL VEGETAL ENLA EMBRIOGÉNESIS SOMÁTICA DlRECTA DE Coflea arabica VAR. CATURRA XHÍBRIDO DE TIMOR.Molina V., O M.; Aponte A., M E.; Cortina G., H. y Moreno R., L G. 69
DETERMINAÇÃO DO TAMANHO DO BOTÃO FLORAL PARA A CULTURA DEANTERAS NO CAFEEIRO (Coffea arabica L.).Luciana Marques da Cunha Andrade,André Barretto Pereira, Edilene Carvalho Santos e MoacirPasqual 75

INFLUÊNCIA DOS REGULADORES DE CRESCIMENTO NA CULTURA DE EMBRIÕES"INVITRO" DO CULTIVAR CA TUAÍ VERMELHO LCH 2077-2-5-44.Luciana Marques da Cunha Andrade, André Barretto Pereira, Ronilda Juliana de Campos e MoacirPasqual 79
INFLUÊNCIA DE REGULADORES DE CRESCIMENTO NA MICROPROPAGAÇÃO "INVITRO" DA CULTIVAR CA TUAÍ VERMELHO LCH 20 77-2-5-44.Luciana Marques da Cunha Andrade, André Barretto Pereira, Anna Lygia de Rezende Maciel eMoacir Pasqual 83
EV ALUACIÓN MORFOLÓGICA Y AGRONÓMICA DE PLANTAS DE CAFÉREGENERADAS POR EMBRIOGÉNESIS SOMÁTICA.Juan Carlos Herrera - Pinilla y Maria Eugenia Aponte -A. 87
A MODEL FOR THE STUDY OF VARIATION INDUCED BY TISSUE CULTURE ONCOFFEE.Fuentes-Cerda, C. F. J., M. Monforte-González, M. Méndez-Zeel, R. Rojas-Herrera, J. Mijangos-Cortés and V. M. Loyola-Vargas 93
STANDARDIZATION OF ENCAPSULATION TECHNIQUE FOR PRODUCING SYNTHETICSEEDS IN COFFEE.B. Muniswamy and H L. Sreenath 99
DESARROLLO DE LA VARIEDAD PORTA-INJERTOS "NEMAYA" (C canephora)PORMEDIO DE EMBRIOGENESIS SOMATICA EN CENTRO AMERICA.F. Anzueto, A. MoIina , P. Figueroa, H. Etienne y B. Bertrand 103
FUSÃO INTERESPECÍFICA DE PROTOPLASTOS EM Coifea.Cordeiro, AT; Zambolim, L; Pétiard, V eSpiral, J. 109
Sessão 2 - MARCADORES MOLECULARES
A - Palestras
MARCADORES DE DNA PARA MELHORAMENTO DO CAFEEIRO.Ney Sussumo Sakiyama 115
GENETIC DIVERSITY AND MOLECULAR MAPPING OF COFFEE.Lashermes P., Combes M.C., Topart P., Anthony F. 121
MICROSATELLITES IN Coifea arabica CONSTRUCTION AND SELECTION OF TWOGENOMIC LIBRARIES.Vascotto F., Ivanissevich S., Rovelli P., Anthony F. ,Anzueto F., Lashermes P., Graziosi 125
OBTENÇÃO DE MARCADORES MOLECULARES EM Colletotrichum s.p,ISOLADOS DECAFEEIRO(Coifea arabica).Margarida P. Rocheta, T. Bessa, A. Dias, A. Várzea, V. Rodrigues Jr. e C.J.Tenreiro, R. 129
GENETIC FINGERPRINTING OF COFFEE LEAF RUST DIFFERENTIALS WITH RAPDMARKERS.A.Santa Ram and H.L.Sreenath 133

B- Posters
PADRÃO MOLECULAR DE CLONES DE CAFEEIRO DIFERENCIADORES DE Hemileiavastatrix Berk. et Br.Terezinha A. Teixeira, Laércio Zarnbolirn, Ney S. Sakiyama, Antônio A. Pereira e Dalza G. daSilva 141
GENETIC DIVERSITY OF CENICAFE COFFEE GERMPLASM COLLECTION.Marco Cristancho, Hernando Cortina and German Moreno 145
GENETIC ANALYSIS OF BRAZILlAN STRAINS OF Xylella fastidiosa ASSOCIATED WITHCITRUS ANO COFFEE.Rui P. Leite Jr., Angela Mehta, Flávia M. S. Carvalho and Bernardo Ueno 151
BIOCHEMICAL ANO MOLECULAR STUDIES OF THE MA IN PROTEIN IN THE COFFEEENDOSPERM.Ricardo Acufia, Ronald Bassüner, Vadim Beilinson, Hernando Cortina, Gabriel Cadena-Górnez,Virginia Montes and Niels Nielsen 155
USO DE PADRÕES ELETROFORÉTICOS NA DIFERENCIAÇÃO DE ClILTIVARES E DONÍVEL DE DETERIORAÇÃO DE SEMENTES DE Coffea arabica.L. Brandão Jr., Delacyr da Silva; Vieira, Maria das Graças Guimarães Carvalho; Bernadino Filho,José Roberto; Hilhorst, Henk e Guimarães, Rubens José 161
VARIABILlDADE GENÉTICA OBTIDA POR RAPO EM ESPÉCIES E HÍBRIDOS DE Coffea.Paulo M. Ruas, Leandro E. C. Diniz, Claudete F. Ruas e Tumoru Sera 165
POLIMORFISMO GENÉTICO DETECTADO EM 40 VARIEDADES E CULTIV ARES DECoffea arabica UTILIZANDO RAPO ASSOCIADO COM ENZIMAS DE RESTRIÇÃO.Leandro E.C. Diniz, Paulo M. Ruas, Claudete F. Ruas e Tumoru Sera 171
CARACTERIZAÇÃO DE ACESSOS DE Coffea POR MARCADORES RAPO.Terezinha A. Teixeira, Ney S. Sakiyama, Laércio Zambolim, Antônio A. Pereira e Cássia C. H.Sakiyama 177
CARACTERIZACIÓN MOLECULAR DE GENOTIPOS DE CAFÉ.José W. Bustamante P. y Delia Polanco L. 181
MAPEAMENTO DE GENE DE RESISTÊNCIA A FERRUGEM-DO-CAFEEIRO (Hemileiavastatrix Berk. et Br.) COM MARCADOR RAPO.Nara S. Tedesco, Ney S. Sakiyama, Laércio Zambolim, Terezinha A. Teixeira,Antônio A. Pereira eCássia C. H. Sakiyama 185
Sessão 3 - MELHORAMENTO GENÉTICO
A - Palestras
BIOTECNOLOGIA NO DESENVOLVIMENTO DE CULTIVARES NUM MENOR PRAZO NO"MODELO IAPAR DE CAFÉ ADENSADO".Tumoru Sera 191

DESENVOLVIMENTO DE CAFEEIROS RESISTENTES A INSETOS.Oliveiro Guerreiro Filho 205
CENETlC IMPROVEMENT OF COFFEE IN INDlA.C. S. Srinivasan, N. S. Prakash, D. Padma Jyothi, V.B. Sureshkumar and V. Subbalakshmi 211
MELHORAMENTO CLÁSSICO DO CAFEEIRO RELACIONADO COM A BlOTECNOLOGIAMODERNA.L. C. Fazuoli , M. P. Maluf, O. Guerreiro Filho, H. P. Medina Filho e M. B. Silvarolla 217
EL MEJORAMIENTO GENÉTICO EN AMÉRICA CENTRAL.Bertrand B., Etienne H., Santacreo R., Anzueto F. y Anthony F.
B -Posters
231
BlOTECHNOLOGY FOR GENETlC IMPROVEMENT OF INDlAN COFFEE.H.L. Sreenath 247
CUSTO DE PRODUÇÃO MANUAL DE SEMENTES HÍBRIDAS DE CAFÉ.Sergio Fadelli e Tumoru Sera 251
ESTUDO DA CAPACIDADE COMBINATÓRIA E DA HETEROSE EM CRUZAMENTOSENTRE LINHAGENS DE CATUAÍ E HÍBRIDO DE TlMOR, EM CAFÉ.José Roberto M. Fontes, Antônio A. Cardoso, Cosme Damião Cruz Laércio Zambo1im, Ney S.Sakiyama e Antônio A. Pereira 255
RESISTÊNCIA DE CLONES DE C. canephora VAR. CONILLON A Hemileia vastatrix BERK.&BR.Dalza G. da Silva, Laércio Zambolim, Ney S. Sakiyama, Antônio A. Pereira, Aymbiré F. A.Fonseca, Terezinha A. Teixeira e Cássia C. H. Sakiyama 261
Sessão 4 - CONTROLE BIOLÓGICO
A - Palestras
EL CONTROL BIOLÓGICO DE LA BROCA DEL CAFÉ, Hypothenemus hampei, ENCOLOMBlA.Alex E. BustilIo P. 265
CONTROL BlOLOGICO DE PLAGAS DEL CAFETO: PRODUCCION Y SECADO DEBlOPESTlDlDAS EN FERMENTACION EM MEDlO SOLlDO.S. Roussos, C. Bagnis, W. Rodrigues, M. A. Aquihuatl, O. Besnard y R. Duponnois 269
THE NEMATOPHAGOUS FUNGI HELPER BACTERIA (NHB): A NEW DlMENSION FORTHE BIOLOGICAL CONTROL OF ROOT KNOT NEMATODES BY TRAPPING FUNGI.Robin Duponnois, J.L.Chotte, A.M.Bâ and S. Roussos 275
LOS INHIBIDORES DE AMILASAS COMO ALTERNATlVA EN LA PRODUCCION DECAFE RESISTENTE A LA BROCA.Amubio Valencia Jiménez, María T. González y Alex E. Bustillo P. 283

B- Posters
ESTUDOS FISIOLÓGICOS E PRODUÇÃO DE FUNGOS MICORRÍZEOS.Álvaro Alberto de Araújo e Sevastianos Roussos 293
CHARACTERISATION OF Beauveria bassiana AND Metarhizium anisopliae ISOLATES FORPOTENTIAL USE AGAINST THE COFFEE BERRY BORER.Patricia E. Vélez, María T. González, Armando Rivera, Alex E. Bustillo P., María N. Estrada andEsther C. Montoya 299
PRIMEIRA INTRODUÇÃO NO BRASIL DO ENDOPARASITÓIDE Phymastichus co.ffea(LASALLE) (HYMENOPTERA: EULOPHIDAE), PARA O CONTROLE BIOLÓGICO DABROCA-DO-CAFÉ Hypothenemus hampei (FERRARI) (COLEOPTERA: SCOLYTIDAE).Fernando Cantor, Evaldo F. Vilela e Alex E. Bustillo P. 305
EXTRACCION Y EVALUACION DE AMILASAS DIGESTIVAS DE LA BROCA DEL CAFÉ.Hypothenemus hampei (Ferrari) (COLEOPTERA: SCOL YTIDAE).Claudia P. Martínez Díaz, Arnubio Valencia Jiménez, María T. González G., Alex E. BustiJlo P......... 307
ATRATlVIDADE DE FRUTOS À BROCA-DO-CAFÉ, Hypothenemus hampei(FERRARI):ESTÍMULOS VISUAIS E OLFATIVOS.Jorge Rafael Mendoza), José Oscar Gomes de Lima, Evaldo Ferreira Vilela e César José Fanton ...... 313
AVANÇOS NAS TÉCNICAS PARA CRIAÇÃO MASSAL DO PARASITÓIDE Cephalonomiastephanoderis HYMENOPTERA:BETHYLlDAE) SOBRE Hypothenemus hampei(COLEOPTERA:SCOLYTIDAE) MULTIPLICADA EM DIETA ARTIFICIAL.Amador Villacorta e Sônia M. Torrecillas 317
Sessão 5 - VALORIZAÇÃO DOS SUBPRODUTOS DA AGROINDÚSTRIA
A - Palestras
EXPERIÊNCIA BRASILEIRA NA VALORIZAÇÃO BIOTECNOLÓGICA DE SUBPRODUTOSDA AGROINDUSTRIA DO CAFÉ.Carlos Ricardo Soccol, Fan Leifa, Adenise L. Woiciechowski, Debora Brand, Cristina M. M.Machado, Marlene Soares, Pierre Christen e Ashok Pandey 323
EXPERIÊNCIA MEXICANA NA VALORIZAÇÃO BIOTECNOLÓGICA DE SUBPRODUTOSDA AGRINDÚSTRIA DO CAFÉ.Saucedo Castafieda G., Romano Machado J.M., Gutiérrez Sánchez G., Delgado VidaJ F., RamírezRomero G., Perraud Gaime I. 329
LOMBRlCOMPOSTAJE DE LA PULPA DE CAFÉ EN MÉXICO.Eduardo Aranda and Isabelle Barois 335
PROCESAMIENTO DE FRUTOS DE CAFÉ POR VIA HUMEDA Y GENERACION DESUBPRODUCTOS.Jaim Zuluaga Vasco 345

B - Posters
POTENCIALIDADE DE USO DE RESÍDUOS E SUBPRODUTOS DO PROCESSAMENTOAGROINDUSTRIAL DO CAFÉ.Celso Luis Rodrigues Vegro e Flávio Condé de Carvalho 357
EVALUACION DE LA MADUREZ DE LA COMPOSTA DE PULPA DE CAFÉ.Gloria Sánchez, Eugenia J. Olguín, Ricarda Marín y Gabriel Mercado 361
CAFFEINE DEGRADATION IN FRESH ANO ENSILED COFFEE PULP BY SOLID ST ATEFERMENTATION: INFLUENCE OF SUBSTRATE PRETREATMENT ANO INOCULUMLEVEL.Romano-Machado J., Gutiérrez-Sánchez G., Perraud-Gaime 1., Gutiérrez-Rojas M. y Saucedo-Castafíeda G. 365
EV ALUATION OF Penicillium sp. V33A25 CAFFEINASE ACTIVITY IN RELATION TO ITSCONSERVATION METHOD.Gutiérrez-Sánchez G., Perraud-Gaime 1., Augur C., Romano-Machado J.,Saucedo-Castafíeda G. ....... 373
PRODUCTION OF EDIBLE MUSHROOM Lentinus edodes ON THE COFFEE SPENTGROUND.Fan Leifa, Ashok Pandey, Carlos Ricardo Soccol and Radjiskumar Mohan 377
SOLID ST ATE FERMENTATION ANO PRODUCTION OF P/eurotus ostreatus ON THECOFFEE RESIDUES.Fan Leifa, Ashok Pandey, Carlos Ricardo Soccol and Radjiskumar Mohan 381
PRODUÇÃO DE AROMA FRUTAL POR Ceratocystis fimbriata EM RESÍDUO SÓLIDO DEAGROINDÚSTRIAS DE CAFÉ.M. Soares, P. Christen, A. Pandey, M. Raimbault e C.R. Soccol 385
PRODUÇÃO DE ÁCIDO CÍTRICO POR Aspergillus niger LPB 21 EM FERMENTAÇÃO NOESTADO SÓLIDO COM CASCA DE CAFÉ.Luciana P.S. Vanderberghe: Ashok Pandey, Jean-Michel Lebeault e Carlos R. Soccol 389
LA PULPA DE CAFE ES UN SUBPRODUCTO Y NO UN DESECHO.Jose R. Ramirez Martinez 393
CARACTERIZAÇÃO DE ÁGUAS RESIDUÁRIAS DA LAVAGEM E DESPOLPA DE FRUTOSDO CAFEEIRO E POSSIBILIDADES DE SEU USO NA FERTIRRIGAÇÃO.Matos, A.T., Pinto, A.B. e Borges, J.D. 395
SELECTION OF STRAINS OF Vo/variella vo/vacea ANO CHARACTERISTICS ON THEEXTRACT OF COFFEE HUSK.Fan Leifa, Ashok Pandey, Carlos Ricardo Soccol and Radjiskumar Mohan 397
DETOXIFICAÇÃO BIOLÓGICA DA CASCA DE CAFÉ POR FUNGOS FILAMENTOSOS EMFERMENTAÇÃO NO ESTADO SÓLIDO.Debora Brand, Fernando Kawata, Ashok Pandey, Sevastianos Roussos, Maria Carolina Rocha dosSantos e Carlos Ricardo Soccol 401
CASCA DE CAFÉ COMO SUBSTRATO PARA PRODUÇÃO DE GA 3 POR FERMENTAÇÃO.Cristina M. M. Machado, Carlos R. Soccol, Brás H. Oliveira, Carlos Oliveira e Ashok Pandey 405

SCREENING OF FILAMENTOUS FUNGI FOR THE PRODUCTION OFEXTRACELLULARTANNASE IN SOLID-STATE FERMENTATION (SSF).Van de Lagemaat, J., Augur, c., and Pyle, D.L. 409
PRODUCTION OF FUNGAL TANNASE IN A MODEL SYSTEM BY SOLID STATEFERMENTATION.Cristóbal Noé Aguilar, Christopher Augur,Gustavo Viniegra-González , Ernesto Favela-Torres ..... 413
Sessão 6 - QUALIDADE DO PRODUTO
A - Palestras
QUALIDADE DO CAFÉ SOLÚVEL BRASILEIRO.Paulo Pauro de Oliveira 419
INFLUÊNCIA DA VARIEDADE BOTÂNICA SOBRE A QUALIDADE ORGANOLÉPTICADO CAFÉ ESPRESSO.Marino Petraco 421
A QUALIDADE DO CAFÉ PRODUZIDO EM DIFERENTES REGIÕES DO BRASIL.José Guilherme Cortez 427
POTENCIAL DE MERCADO DE CAFÉS ORGÂNICOS E ESPECIAIS.Ivan Caixeta 431
MYCOTOXIGESIS IN GRAINS APLICATION TO MYCOTOXIC PREVENTION IN COFFEE.Joseph Le Bars and P. Le Bars 435
OCRATOXINAS E AFLATOXINAS EM CAFÉ BRASILEIRO.Lúcia Valente Soares 447
B-Posters
RESULTADOS E DESAFIOS DO PROGRAMA DE AUTOFISCALIZAÇÃO DO CAFÉTORRADO E MOÍDO: SELO ABIC.Celso Luis Rodrigues Vegro e Célia Regina R. P.Tavares Ferreira 453
QUALIDADE DE BEBIDA DA CULTIVAR DE CAFÉ IAPAR-59 E OUTRAS.Tumoru Sera e Marcos Aurelio Bacceti 457
EFEITO DA ADUBAÇÃO POTÁSSICA NA QUALIDADE DOS GRÃos BENEFICIADOS DECAFÉ.Enilson De Barros Silva, Francisco Dias Nogueira, Paulo Tácito Gontijo Guimarães, Silvio Júlio DeRezende Chagas e Laerte Costa 459
CONDUTIVIDADE ELÉTRICA DO EXSUDATO DE GRÃos DE CAFÉ (Coffea arabica L.) ESUA RELAÇÃO COM A QUALIDADE DA BEBIDA.Prete, C.E.C. e Abrahão, J.T.M. 463
RELAÇÃO ENTRE O PESO E A CONDUTIVIDADE ELÉTRICA DE GRÃOS DE CAFÉ DEPROGÊNIES DE ICATU.Prete, C.E.C.; Sera, T. e Fonseca, I.C.B. 473

CONDUTIVIDADE ELÉTRICA DE EXSUDATOS DE GRÃOS DE CAFÉ COLHIDOS EMDIFERENTES ESTÁDIOS DE MATURAÇÃO.Prete, C.E.C.; Sera, 1.; Crudi, C.E. e Fonseca, LC.B. 475
AVALIAÇÃO DE PROGÊNIES DE CAFÉ (Coffea arabica L. cv. Icatu) QUANTO ACONDUTIVIDADE ELÉTRICA DO EXSUDATQ.Prete, C.E.C.; Sera, T. e Fonseca, LC.B. 479
MELHORIA DE QUALIDADE EM ÓLEO DE CAFÉ PELO USO DA EXTRAÇÃOACELERADA COM SOLVENTE(ASE).Jair S.S. Pinto, Fernando M. Lanças, Suzana L. Nixdorf e Paulo L. C. Oliveira 483
INCIDÊNCIA DE FUNGOS EM CAFÉ.Marta Hiromi Taniwaki, Andréia Aparecida Banhe e Beatriz Thie Iamanaka 487
MONITORAMENTO DA QUALIDADE DO CAFÉ TORRADO E MOÍDO NO ESTADO DESÃO PAULO.Mori, E. E. M., Anjos, V. D. A . e Bragagnolo, N. 493
AQUISIÇÃO DA TOLERÂNCIA À DESSECAÇÃO NOS DIFERENTES ESTÁDIOSDEMATURAÇÃO DE SEMENTES DE CAFEEIRO.Delacyr da Silva Brandão Jr., Maria das Graças Guimarães Carvalho Vieira,Lilian Padilha, RubensJosé Guimarães, José Roberto Bernardino Filho e Henk Hilhorst 499
EFEITO DE ADUBAÇÕES FOLIARES EM PRÉ E PÓS FLORADA NA PRODUÇÃO EQUALIDADE DAS SEMENTES DE CAFEEIRO.Lima, Dinara Mattioli; Von Pinho, Edila Vilela Resende;Vieira, Maria das Graças GuimarãesCarvalho; Cunha, Rodrigo da; Guimarães, Rubens José e Monteiro, João Vieira 503
BIOTECNOLOGY AND THE FUTURE OF COFFEE PRODUCTION
Gustavo Viniegra-González.
Universidad Autónoma Metropolitana, Iztapalapa, D.F. MEXICO.
INTRODUCTION
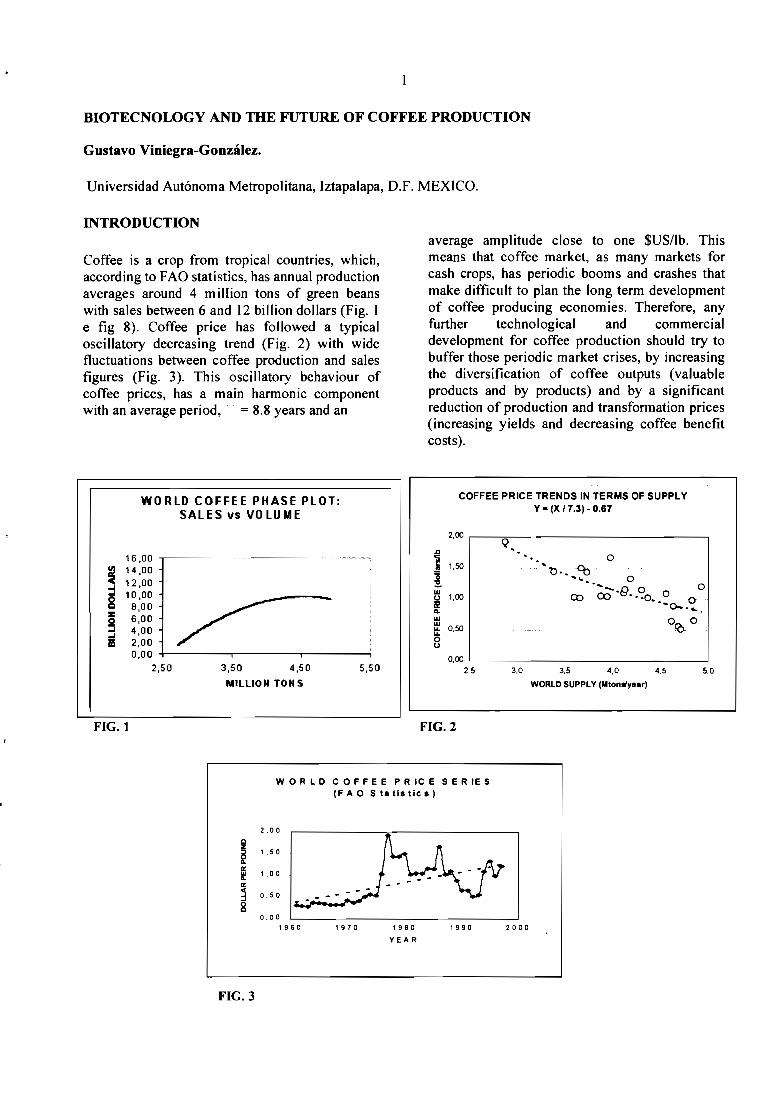
Coffee is a crop from tropical countries, which,according to FAO statistics, has annual productionaverages around 4 m iIIion tons of green beanswith sales between 6 and 12 billion dollars (Fig. Ie fig 8). Coffee price has fol1owed a typicaloscillatory decreasing trend (Fig. 2) with widefluctuations between coffee production and salesfigures (Fig. 3). This oscillatory behaviour ofcoffee prices, has a main harmonic componentwith an average period,· = 8.8 years and an
WORLD COFFEE PHASE PLOT:SALES vs VOLUME
16,00
i 14,00
... 12,00... 10,00ol:l 8,00z 6,00~... 4,00li 2,00 :
0,002,50 3,50 4,50 5,50
MILLlON TON S
FIG.l
average amplitude close to one $US/lb. Thismeans that coffee market, as many markets forcash crops, has periodic booms and crashes thatmake difficult to plan the long term developmentof coffee producing economies. Therefore, anyfurther technological and commercialdevelopment for coffee production should try tobuffer those periodic market crises, by increasingthe diversification of coffee outputs (valuableproducts and by products) and by a significantreduction of production and transformation prices(increasing yields and decreasing coffee benefitcosts).
COFFEE PRICE TRENDS IN TERMS OF SUPPL YY-(X/7.3)-O.67
2,00Q.D
~ olO 1,50 .. ..
'O•• <b! ..... ow -;;;.9. 0 o oU 1,00 co -·0 oli! .....0.... ""lLw 0lõ ow
0,50LLlLoU
0,00
2,5 3,0 3,5 4,0 4,5 5,0
WORLD SUPPLY (Mtona'yeer)
FIG.2
W o R L o C o F F E E P R IC E 5 E R IE 5(F A o 5 ta tia tlc a)
2. ooQZ
~1 ,5 o
a:w 1 ,O o...a:
~ 0,50 ---8 -.
FIG.3
o, oo196 o 1 970 1 980
YEAR1 990 2 ooo

2
THE NATURE DF MARKET OSCILLATIDNSIN CASH CRDP ECONDMIES
A dynamic model for coffee market.
A simple model illustrates some ofthe remarkableproperties of coffee market. This is defined bythree variables, X = the raw material in theproducing country (green beans), Y = theintermediate material transported from theproducing to the consuming countries and Z = thestocks of green beans to be processed andconsumed.
The world trend to protect the environment is amajor concem affecting coffee production andtransformation, because many easy and readytechniques are to be discarded due toenvironmental considerations. For example, pestcontrol using chemical compounds leaving toxicresidues or, disposing of waste waters by thesimple expedient to dump the untreateddischarges in rivers or lakes. But, perhaps theenvironmental challenge may be a blessing indisguise. Specially, if new technologiesintroduced to meet environmental standards are, atthe same time, oriented to increase the quality,diversity and commercial opportunities for coffeederivatives. For example, producing new varietiesof organic coffee beans with better price, orgiving rise to new high value added products fromcoffee pulp. (Table 1).In this paper a brief overview is presented on theopportunities to use biotechnology to meet thechallenge of rising coffee costs in fluctuatingmarkets, with strong emphasis in the notion ofproduct diversification.
(3)
(4)
6,9393,072,515,11,328,258,1259,174,10,753,70,320,053,17160
1250J. Zuluaga, 1989
MoistureDry MaterEthereous Ext.Crude fiberNitrogenTotal protein Nx6,25AshExtract free of NCarbohydratesCaffeineTanninsCalciumPhosphorousPotashSodium (ppm)Iron (ppm)Information from
Dxldt = Vo - k.X
dY/dt = k.X - k2X
COMPONENT
Table 1: APROXIMATE CHEMICALCOMPOSITION OF THE COFFEE PULP
At the end of the market chain there is theconsumption rate, VF, which is a convex functionsaturated by the market inventories, Z, becausethere is a maximal consumption rate, Vc, with asaturation constant Kc, as follows.
There are two other variables, X and Y, toconsider in order to have a complete dynamicsystem which, follow the usual mass balanceconsiderations.
ZYx
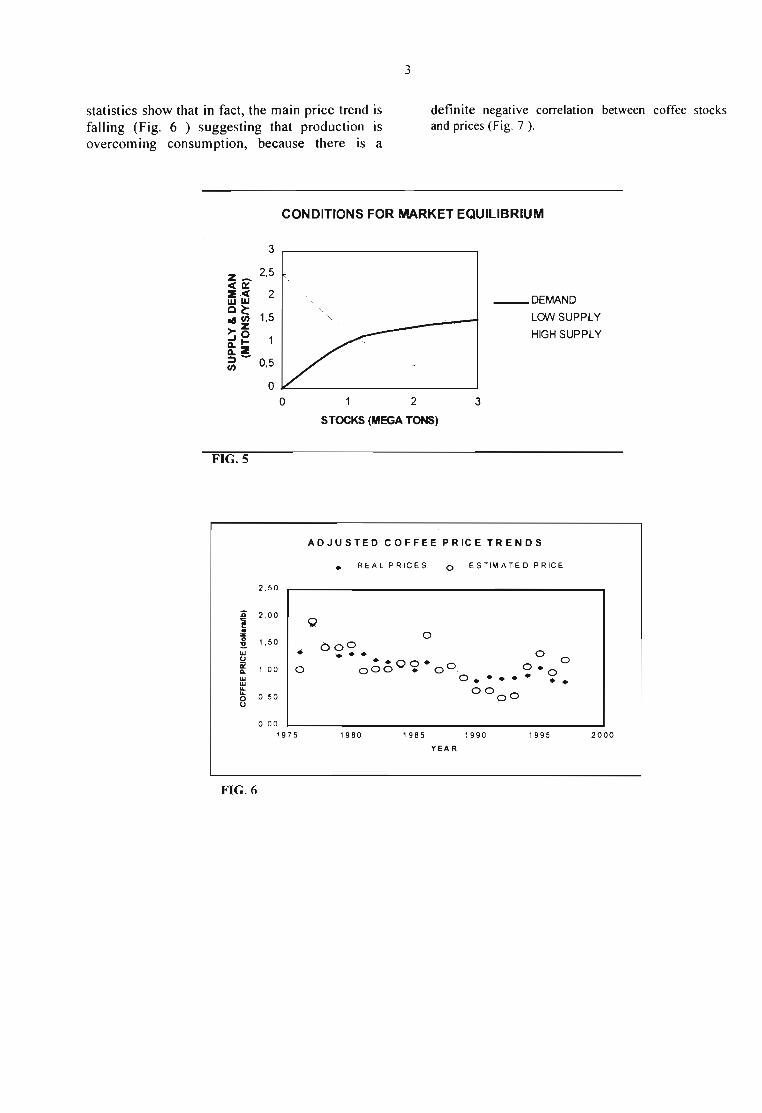
The bottleneck for agricultural ash products. Thismarket will be at equilibrium if each one of therates of change, dXldt, dY Idt, dZ/dt, vanishtogether which lead to the so called steady-stateconditions. This is possible only when Vo (input)= VF (output). Figure 5 shows the graphicalsolution for such a condition. In one case thesaturation levei is high (i. e., K, = 3 Mton) and inother case with K, = 1 Mton (Iow levei).Therefore, the market bottleneck is the final rateof consumption VF because there is a maximumamount of coffee that consumers can buy and alsobecause the rate of growth of consumers is ratherlow as compared to the rate of grow ofproduction. Statistical evidence from FAO
The primary production rate, Vo, should besensitive to the stock leveI, Z, because the price isa decreasing function ofthe stocks
Vo=Vi/[1 +(Z/KdP ] (I)
Where, K, is a saturation constant for thenegative feedback between the stocks and theprimary production. For small values of K,primary production is inhibited more readily thanwith high values of that parameter. Thecoefficient, p> 1, is an indication ofthe delay thatexists between stock accumulation and theinhibition of coffee production.
dZ/dt = k2X - VF (5)
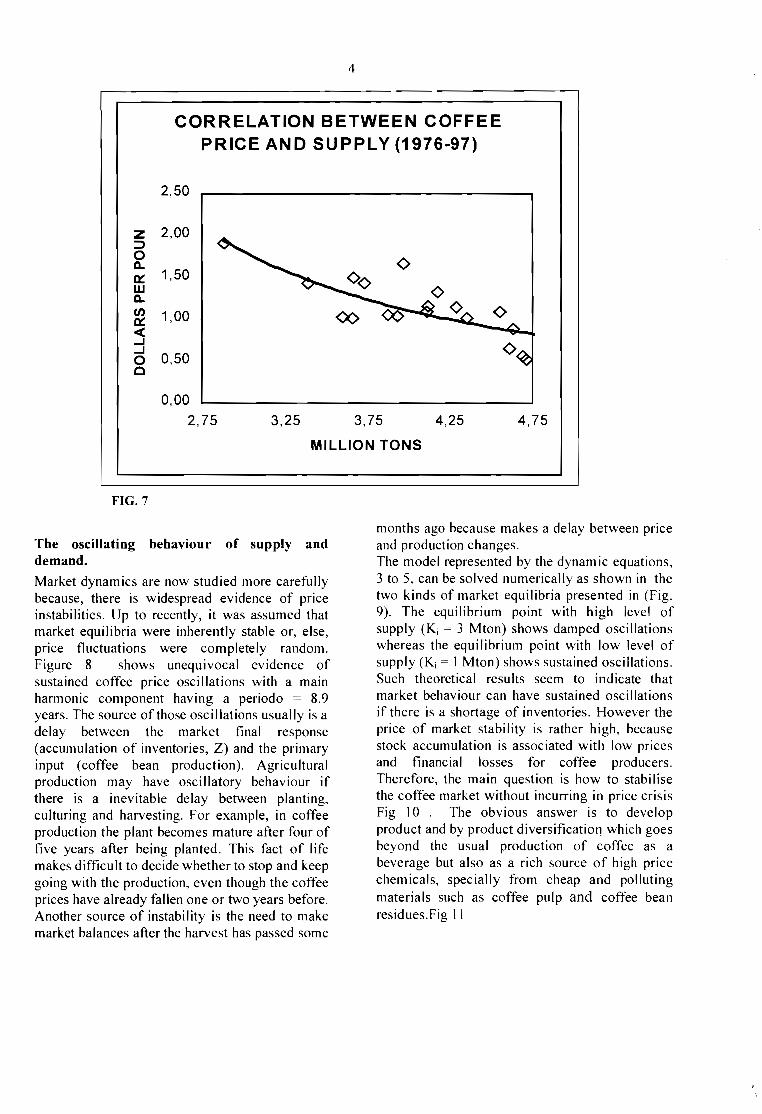
statistics show that in fact, the main price trend isfalling (Fig. 6 ) suggesting that production isovercoming consumption, because there is a
3
definite negative correlation between coffee stocksand prices (Fig. 7 ).
CONDITIONS FOR MARKET EQUILlBRIUM
3
z_ 2.5<o:~ .~ 2
o~1.5 -,
~cn
>-z-lOQ,.I-Q,.:I;:)-
0.5cn
2 3
STOCKS (MEGA TONS)
__DEMAND
LON SUPPLY
HIGH SUPPLY
FIG.S
ADJUSTED COFFEE PRICE TRENDS
• R E A L P R IC E 5 o ESTIMATED PRICE
2 .50
:õ 2 .00
~Õ 1 .50:!!.wu~ 1.00a..ww......
0 .50ou
o
o00 0
••• o•• 00. o
o o o • o o . c;: • oo • • • • • •
0000
FIG.6
0 .00
'975 1980 1985
YEA R
1990 1995 2000
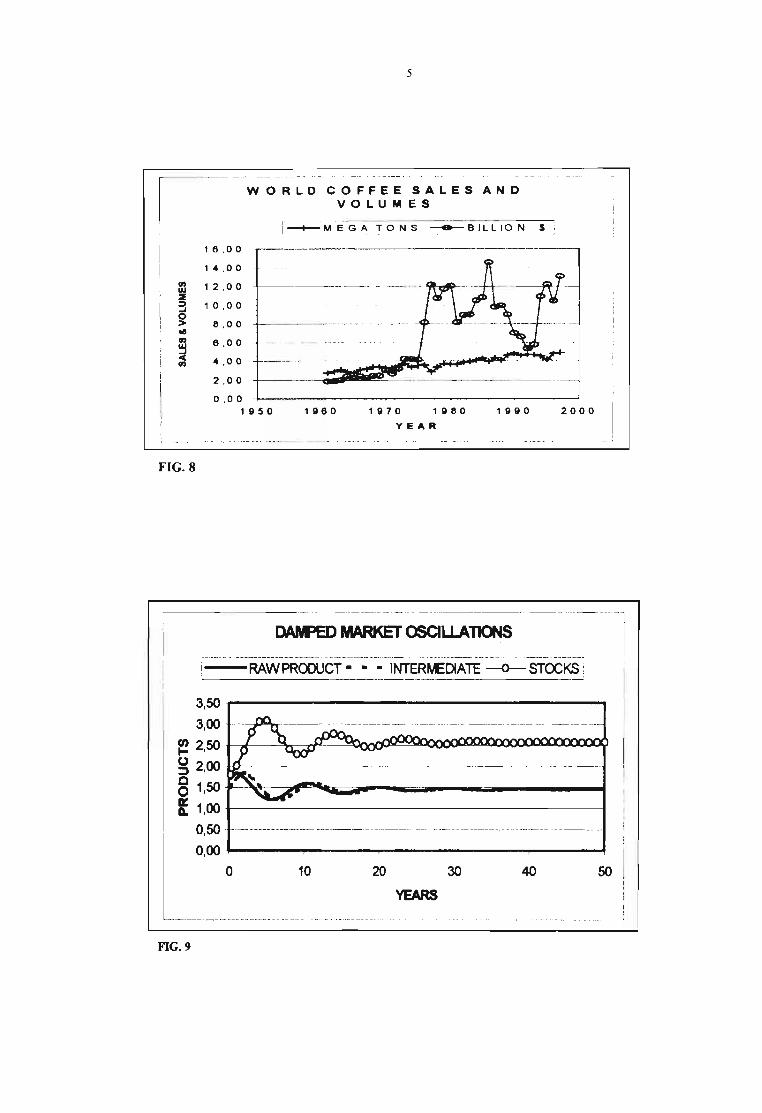
4
CORRELATION BETWEEN COFFEEPRICE AND SU PPLY (1976-97)
2,50
z 2,00=>OQ.
o::: 1,50wQ.rn 1,00 <>o:::«...J <>...J
0,50OC
0,00
2,75 3,25 3,75 4,25 4,75
MILLlON TONS
FIG.7
The oscillating behaviour of supply anddemando
Market dynamics are now studied more carefullybecause, there is widespread evidence of priceinstabilities, Up to recently, it was assumed thatmarket equilibria were inherently stable or, else,price fluctuations were cornpletely random.Figure 8 shows unequivocal evidence ofsustained coffee price osci Ilations with a mainharmonic component having a periodo = 8.9years. The source ofthose oscil1ations usually is adelay between the market final response(accumulation of inventories, Z) and the primaryinput (coffee bean production). Agriculturalproduction may have osci lIatory behaviour ifthere is a inevitable delay between planting,culturing and harvesting. For exarnple, in coffeeproduction the plant becomes mature after four offive years after being planted. This fact of lifemakes difficult to decide whether to stop and keepgoing with the production, even though the coffeeprices have already fallen one or two years before.Another source of instability is the need to makemarket balances after the harvest has passed some
months ago because makes a delay between priceand production changes.The model represented by the dynamic equations,3 to 5, can be solved numerically as shown in thetwo kinds of market equilibria presented in (Fig.9). The equilibriurn point with high level ofsupply (K, = 3 Mton) shows damped oscillationswhereas the equilibrium point with low levei ofsupply (K, = 1 Mton) shows sustained oscillations.Such theoretical results seem to índicate thatmarket behaviour can have sustained oscillationsif there is a shortage of inventories. However theprice of market stability is rather high, becausestock accumulation ís associated with low pricesand financiai losses for coffee producers.Therefore, the main question is how to stabilisethe coffee market without incurring in price crisisFig 10. The obvious answer is to developproduct and by product diversification which goesbeyond the usual production of coffee as abeverage but also as a rich source of high pricechem icals, specially from cheap and pollutingmateriais such as coffee pulp and coffee beanresidues.Fig 11
5
1---- ---~-~--~~~- C voj ~~-~ i :-L E 5 ~ N ~--
i-M-EG-A TONS ~BiLiION$
-------------.,16,0 O
1 .. ,O O
CI) 12,0 Ow::I::l 1 O, O O..JO> 8,0 O011CI) 6,0 O~cc: .. ,O OCI)
2,0 O
O ,O O
1950 1960 1970 1980
YEAR1990 2000
FIG.8
DANFED MARKET OSCIUAllONS--------l
I
I
!
--RAWPROOUCT· •• INTERMEDlATE -o-STOCKS:~---------_.._---
50403020
---------.---------- ----
10
3,50 ~------------------....,
3,00~ 2,50 hr-~~-rfI~~fd.~:>OOOOOc:P.:lOOC)()()OOClOCXxnXXJ«>
g 2,00QO 1,50 t--~~~"""lloI'&;;;""-""iiiiiiiiiiI-- - -I~Q. 1,00 -1------------------------1
0,50 -
0,00+----,.---~---------,.---~O
YEARS
FIG.9
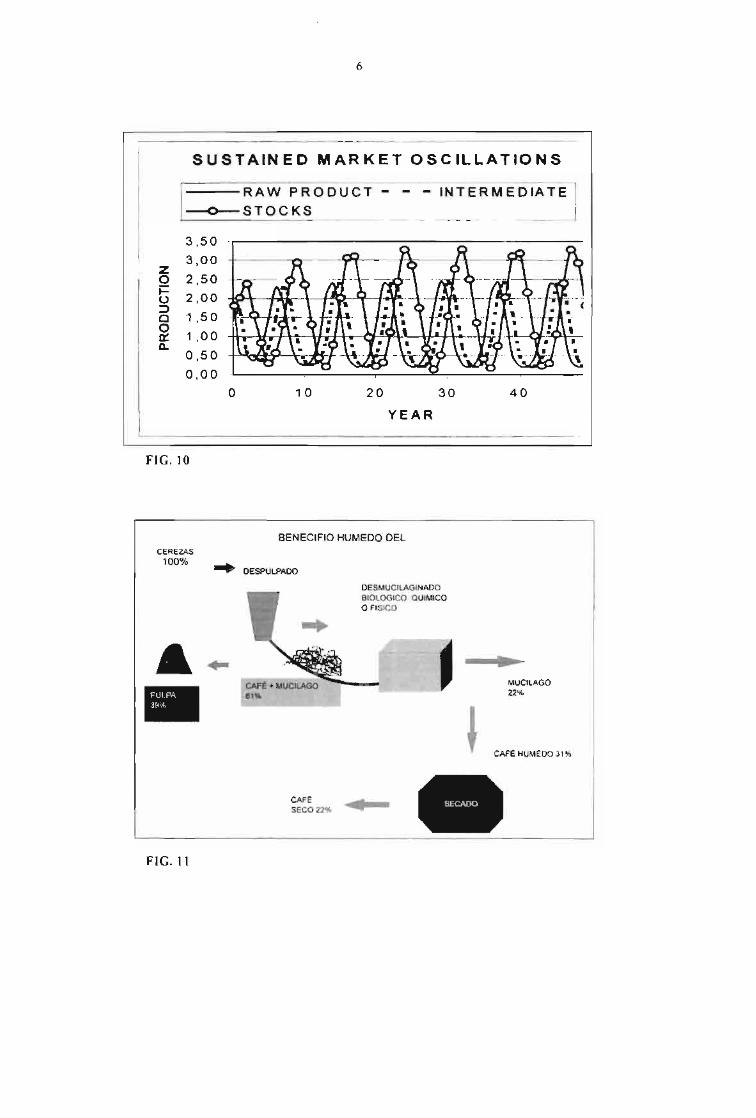
6
SUSTAINED MARKET OSCILLATIONS
I RAW PRODUCT---O--STOCKS
INTERMEDIATE
zoi=o=>aOa::Q.
3,50
3,00
2,50
2,00
1,50
1 .00
0,50
0,00
o 10 20 30 40
L YEAR- - - - - - - - - - - - - --- - - - - ----
FIG.10
BENEC1FIQ HUMEDO OElCEREZAS
100%OESPULPAOO
..•
OESMUCl lAGINiI[) OBIOLOO o OUIMICOo FISJC.i.l
MUCILAGO
22"
CAFEHUMEOO 31%
flG.ll
CAFEseco

7
• THE CHALLENGE OF ECOLOGY ANOCOFFEE PRODUCTlON.
• THE ROLE OF BIOTECHNOLOGY IN THENEW CONTEXT OF COFFEE PRODUCTlONANO MARKETING.
the same market for the traditional or basicproduct, i.e., wine.
d) Waste materiais (grape pomace) can betransformed in new value added products,helping to finance the cost of waste disposal.
It should be stressed that French distilleries werenot developed as an answer to conventionalmarket forces in the conventional sense of privatesupply and demando They were an adaptation to along series of political and social problemsderived from wine market instability at the end ofXIX century and now are becoming a response tothe problem of waste disposal. In other winemaking countries different ways of solving theproblem of grape over production and wasted isposal have been developed, suggesting thatproduct diversification is a problem to be tackledin close relation to the social conditions of eachgiven region.
PRODUCTECOLOGY
• OPPORTUNITIES FORDIVERSIFICAT10N USINGFRIENDLY TECHNOLOGIES.
THE PROBLEM OF PRODUCTDIVERSIFICATION FOR COFFEE DERIVEDMATERIALS.• Diversity of coffee raw materiais in the pulp,
the skin and the green beans.
• The conventional route of decaffeination anddisposal of coffee pulp and torrefactionresidues.
• Coffee pulp as a source of new materiais(antioxidants and antitumoral derivatives).
• Alternatives for wet disposal and storage ofcoffee pulp.
• A rough estimate of cost/benefit analysis forR&D in coffee development.
Product diversification is well developed for cropssuch as cereais and legumes. For example, thelarge surplus of maize from USA. is used toproduce a wide variety of products such as,animal feeds, fructose syrups, alcohol,fermentation products, edible oil, starch, gluten,plastics, etc. In Brazil, sugar cane uses have beendiversified to include the alternative production ofgasohol as a way to cope with increasing fueldemands and saturation of sugar market. Howeverthere are many problems in finding successfulalternative markets for traditional products suchas coffee because oftwo main factors
The importance and the difficulties for productdiversification.
a) The relative low levei of R & D investment intropical countries which hinders the discoveryof new uses for trad itional crops.
b) The complex web of social, cultural, politicaland financiai arrangements that are associatedto traditional crops and which cannot bechanged overnight in order to develop newindustrial concerns for alternative uses ofconventional or waste products.
An interesting example of product diversificationis the case of grape and wine industry in France.The traditional and main product for French grapeyards iswine, but wine is also a product that hasfound market saturation through more than acentury in Europe. To cope with this problern, theFrench government has issued a number of codesand regulations that help to manage winesurpluses and also are oriented to clean theenvironment. This is the network of distilleriesproducing liquor from the grape pomace or marc,as it is called the solid grape residue from winemaking. There, a number of wine co-operativesget together and finance and manage a singledistillery. The main role ofsuch an enterprise is toproduce brandy or "eau de vie" from the residualsugar in the pomace. Surplus amounts of grapescan also be transformed in liquors when the priceof wine faIls down working as alternative outletsfor crop surplus. This scheme has variousadvantages,
a) There is no distraction for pnrnary andtraditional wine making.
b) Special factories are dedicated to newproducts with new ideas derived from R&D.
c) Surplus raw materiais (grape) from primaryproducers can be transformed into newproducts, such as wine pigrnents, grape seedoil and grape sugar syrups, without entering


9
BIOTECNOLOGÍAS APLICADAS AL MEJORAMIENTO GENETlCO DEL CAFETO
Marc Berthouly
CIRAD-AMIS
1. GENERALIDADES
El café es uno do los productos agrícolas másimportantes en el mercado internacional. De estaactividad depende la economía de más de 50países productores. Cultivado en 11,2 x 106 ha, yprodujo, en 1990, 5,5 x 106 toneladas de café oro(Cafe Cacao Thé, 1992).EI café consumido en el mundo estáprincipalmente producido por dos especiescultivadas: Coffea arabica y Coffea canephorarepresentan respectivamente el 75 y el 25 % deimercado mundial.C. arabica está producido en regiones tropicalesde altitud (500 a 2500 m.s.n.m.). Representa laúnica especie tetraploide (2n = 44 cromosomas) yautogama dei género Coffea. C. arabica produceun café de calidad y con reducido contenido decafeína. No obstante, es por la general sensible alas enfermedades como la roya anaranjada(Hemileia vastatrix Berk & Br.) y la antracnosisdo los frutos (Colletotrichum coffeanum (CBD)),principales enfermedades de esta especie.La metodologia de mejoramiento de C. arabica esla aplicada por la general a las espéciesautogamas. Tiene por base la obtención dedescendencias puras por selección genealógicadespués de la recombinación de caracteresaportados par los padres.Recentemente, la existencia de la heterosis fuesefialada en C. arabica en Índia (Srinivasan yVishveshwara, 1978), en Kenia (Van der Vosseny Walyaro, 1981) Y en Etiopia (Ameha, 1983). Setrato aprovechar hibridos FI multiplicados por viavegetativa pero sin éxito, dado que los métodosconvencionales de propagación no permitieronmás que reducidas tasas de multiplicación (Vander Vossen, 1985).La multiplicación vegetativa desempena por lotanto un papel impartante en la selección de loscafetos. Permite la propagación y la explotaciónde las estructuras genéticas heterocigotos.También puede reducir el número de ciclos doselección. En efecto, en el cafeto, cultivo perene,crear una variedad requiere como mínima 35anos. La multiplicación vegetativa puede por lotanto encontrar su aplicación en clonesseleccionados de C. canephora, en híbridosinterespecíficos seleccionados como el Arabusta,o bien en híbridos intraespecíficos de C. arabicatal como el Riuru 11.
La técnica de multiplicación vegetativa másutilizada en el cafeto es el esqueje. Sin embargo.debido ai dimorfismo de los ejes vegetativos. elnúmero de esquejes ortótropos que puede producirun cafeto se halla muy limitado. En el marco deuna multiplicación a gran escala. la pocadisponibilidad de esquejes puede I1evar a plazasconsiderables para la divulgación de un nuevaoclone. Además, el esqueje hortícola exige lainstalación de jardines clonales, lo que implicadiversas limitaciones entre ellas el mantenimientoy las superficies utilizadas (Deus y Descroix,1984).En este contexto, la puesta a punto de técnicas demultiplicación in vitro presenta un gran interéspara incremientar la tasa de multiplicación ypermitir una difusión rápida de los híbridos o delos clones seleccionadas.Desde hace algunos anos. varios autores(Staritsky, 1970; Sondahl y Sharp, 1977; Dublin.1980, 1982; Pierson et ai, 1983; Yasuda a ato1985; De Garcia y Méndez, 1987; Berthouly et ato1987) mostraron que el micro esqueje y laembriogénesis somática pueden aplicarse aicafeto.La embriogénesis somática aparece coma unaalternativa interesante y más económica para lamultiplicación de plantas seleccionadas. Ademásde constituir un modelo interesante para estudiosfundarnentales, su aplicación más promisoria es lamultiplicación de plantas a gran escala(Amrnirato, 1987).Distintos estudios sobre el cafeto permitieronevidenciar la aptitud de esta especie para laembriogénesis somática y para el desarrollo de losembriones en plántulas. No obstante. Ia tasa demultiplicación seguía siendo muy baja para unautilización a gran escala. En efecto, De Garcia yMendez (1987) que trabajan sobre laernbriogénesis somática en media sólidomencionan como producción "récord" 60 a 65embriones par explant después de 14 semanas decultivo.Con el fin de explotar el relevante potencial de laembriogénesis somática y de reducir así el costode producción de las plántulas, su aplicación enmedio liquido resulta necesaria. En efecto, Iaposibilidad de (cultivar suspensiones celularespuede permitir el "scaling-up" y la producciónmasiva de embriones en bio-reactor. En otrasespecies, los resultados son muy promisorios.

Pétiard et a!. (1987) relatan una capacidad deproduccíón de 80.000 embriones de zanahoria porlitro y por dia en un bio-reactor de 10 litros.Hasta ahora no existia un método comercial demultiplicacíón in vitro deI cafeto.
2. ESTRATEGIAS DE MEJORAMIENTOGENÉTICO DE Coffea sp
2.1. Mejoramiento convencional2.1.1. C. canephora2.1.1. 1.Generalidades
Cicanephora es una especie allogama con unaincompatibilidad de tipo gamétofiticamonogénética (Berthaud ,1980);El mejoramiento genético de dicha especie se hizopor dos vias: via vegetativa a via generativa(Bouharmont et Awemo, 1979; Charrier ,1985).EI desarrolló de ténica de multiplicaciónvegetativa ha permitido la valorización degenotipos heterocigotos y en el mismo tiempogrande progresos en el mejoramiento genético dedichos cafetos(Charrier, 1985)
2.1.1.2. Métodos de selección• selección vegetativa: so selección las mejoresplantas con las caracteristicas deseadas. Dichasplantas so siembran en ensayos multi locales. Lasmejores plantas seleccionadas serán utilizadascomo "plantas madres" par ser multiplicadas engran escala por via vegetativa.• Seleccion generativa: desde muchos afies se estarealizando una nueva estrategia de mejoramientoIlamado .. selección recurente reciproca".La prímera fase dei trabajo consíste en la creacíónde poblaciones de dos grupos de dos diferentesorigen: guinea y congo. En estos grupos so hacehibridaciones entre los diferentes individuosdurante varios ciclos hasta obtener plantasseleccionadas (híbridos).La segunda fase consiste en la creación dehíbridos entre los dos grupos mejorados para juntaen una misma planta las mejores característicasgenéticas de los dos grupos (Berthaud, 1985;Charrier et Berthaud .\988)Dichas plantas mejoradas pueden ser utilizadaspor la creación do campo de semillas híbridas.Actualmente un tal programa so esta llevando acabo an Costa de Marfil por el Cirad y lainstitución nacional.
2.1.2.C. arabica2.1.2.1. GeneralidadesLas plantaciones de C. arabica en los paísesproductores de América Central y de Asia del Sury dei Sureste fueron constítuídas oríginalmente a
\0
partir de un número muy limitado de plantas(Coste, 1968). En consecuencia y debido a sucarácter autogamo, los C. arabica cultivadostienen una base genética limitada. Charrier (1980,1985) hizo especíalmente énfasis en el ínterés porprospectar cafetos espontáneos asl como suconservación y su integración en los programas demejoramiento.Las prospecciones de La FAO en 1964 y delORSTOM en 1966, han contribuido a laampliación de la base genética de numerosascolecciones de C. arabica con cafetos de origensubespontáneo deI suroeste etíope (Charrier,1985).La aparición de más de 40 mutantes en lasplantaciones de Brasil contribuyó a la creación denuevas variedades tales como Maragogype, Ceray Caturra (Carvalho, 1988). Entre estos mutantes,varios presentan un interés practico para elmejoramiento de esta especie (Van der Vossen,1985). Por ejemplo, el mutante Caturracaracterizado por un crecimiento compacto fueutilizado en varios programas de mejoramiento.Su tamafio reducido permite cultivarlo en altasdensidades e incrementar asimismo laproductividad.
2.1.2.2. Criterios y métodos de selecciónEn el caso de C. arabica, se enfocaron loscriterios de selección en la adaptación a lascondiciones ecológicas regionales, en elrendimiento y la calidad. En un pasado recente,los trabajos fueron orientados principalmentehacia la creación de variedades resistentes a laroya anaranjada y a la antracnosis de los frutos.Estas dos enfermedades constituyen el problemafitosanitario más serio para C. arabica.La metodologia tradicional seguida paraseleccionar C. arabica fue principalmente lahibridación y luego la seJección genealógica y laseleción por retro cruzamiento (Charrier, 1985;Bettencourt y Rodrigues, 1988). Las variedadesasí logradas son suficientemente homogeneas parapermitir una multiplicación por semilla. Así,prácticamente se instalaron todas las plantacionesde C. arabica con semilla (Van der Vossen,1985). Un descubrimiento interesante quesubrayar, es la existencia de la heterosis en elcafeto C. arabica. Fue referido en Índia(Srinivasan y Vishveshwara, 1978), en Kenia(Van der Vossen y Walyar, 1981) y en Etiopia(Ameha, 1983). La heterosis puede explotarsepara la creación de variedades híbridas FI, lo quemodificaria radicalmente la estrategia clásica deselección (Charrier, 1985). Los ensayos demultiplicación por via vegetativa tradicional dohibridos FI no tuvieron exito. La producción de
,.

hibridos se realiza mediante cruzamiento manualcontrolado de los padres (Van der Vossen, 1985).
2.1.2.3. La selección pa ra la resistencia a lasenfermedadesLos programas de investigación sobre el cafeto enBrasil. en Colornbia, en Índia. en Portugal y enAmérica Latina. se dirigen principalmente hacia elmejoramiento de las variedades cultivadasmediante transferencia de los factores deresistencia Hemileia vastatrix (Bettencourt, 1982).La fuente de resistencia mas utilizada es el híbridodo Tirnor, oriundo de la introgresión natural de C.arabica por los C canephora indonesios. Estehíbrido posee 4 genes mayores de resistenciaprocedente de C. canephora (Bettencourt yRodrigues. 1988).EI cruzamiento deI híbrido de Timor con elmutante enano Caturra Rojo dia la variedadCatirnor, Ia cual se estudia en varias centros deinvestigación deI mundo (Bettencourt, 1982). Elhíbrido de Timor entró en otras combinaciones yllevó a la obtención de las variedades Catimor ySarchimor (Bettencourt, 1983).En el cafeto, el retro cruzamiento se utiliza para laintroducción de caracteres (resistencia a lasenferrnedades, vigor. caracteres del fruto, etc ...)que le faltan a la variedad interesante uti Iizadacomo padre recurrente.EI Catirnor, variedad resistenre a la royaanaranjada, logrado por el "Centro deInvestigação das Ferrugens do Cafeeiro" (CIFC)fue retro cruzado con la variedad Catuai paratransmitirle el carácter de vigor (Carvalho, [988).En efecto, varios investigadores sefialaron la faltade vigor en los Catimor (Bettencaurt, 1982;Zamarripa, 1986).En el transcurso de los últimos 25 anos. elInstituto Agronômico de Campinas en Brasildesarrolló la variedad lcatu, derivada de uncruzamiento artificial entre un C. arabicatetraploide y un C canephora tetraploide logradopor tratamiento con colchicina. EI híbrido FI fueretro cruzado con Mundo Novo y también conotras variedades de C. arabica, tales comoBourbon Amarillo y Catuai. Las poblaciones deIcatu mas eficientes son derivadas de S2 a S3, deBC2 y BC3 (Carvalho. 1982. 1988).La variedad lcatu contiene genes de resistencia ala roya pero también ai CBO y a los nematodos(Carvalho. 1988). Actualmente, Ia selección devarias descendencias de Icatu parece muyadelantada a tal punto que debe exarninarse sudivulgación (Eskes, 1991).Estudios sabre el control genético de la resistenciaindicaron la existencia de por lo menos nuevegenes dominantes de resistencia a Hemileia
11
vastatrix (Bettencourt y Rodrigues, 1988). Laresistencia incompleta de C arabica a H.vastatrix fue objeto de investigaciones. Losresultados logrados en Brasil muestran laexistencia de una resistencia horizontal enpoblaciones de C arábica y de C canephora,variedad Koui1I0u (Eskes, 1983).La antracnosis de los frutos (CBO) es unaenfermedad muy grave de C. arabica cultivado enlas zonas altas. húmedas y frias, de África Centraly Oriental. La CBO no so encuentra en los paísesde América Latina. pera las condicionesclimáticas de altitud son comparables a lasregíones de cultivo dei café de África dei Este(Van der Vassen, 1985). Esta enfermedadpresenta por la tanto un furte peligro potencialpara el cultivo dei café de América.EI programa de selección para la resistencia aiCBO siguió dos estrategias diferentes. En Kenia,el programa de mejoramiento iniciado en 1971.consistió en la introgresión de resistencia ai CBOprocedente dei germo plasma de C arabicaselvaje en variedades regionales tomando laprecaución de mantener la calidad y laproductividad deI café típico de Kenia.Los trabajos lIevadas a cabo en Etiopia sedirigieron hacia la selección y la multiplicación degenotipos resistentes ai CBO seleccionados entrepoblaciones semi-naturales dei centro dediversidad genética do C. arabica. Laproductividad y la calidad tuvieran el segundorango en estos trabajos (Van der Vossen, 1985).Estudios realizados par Van der Vossen yWalyaro (1980) indicaron que la resistencia aiCBO esta controlada por dos genes rnayores de lavariedad Rume Sudan (un gen dominante R y ungen recesive K). y por un gen dei hibrido deTimor (T). de dominancia incompleta.Cabe anotarse que también se verificaron avancesde mejora genética de los cafetos para laresistencia a los nernatodos, Meloidogvne sp y laresistencia a las minadoras de las hojasPerileucoptera coffeella (Eskes. 1991).Desde 1991 se ha iniciado en América Central unprograma de mejoramiento gangetico en base a lacreación de híbridos Fl de Carahica
2.1.2.4. La hibridación inter y intraespecífica:• hibridacián inter especificaLa hibridación inter especifica fue utilizada en elcafeto para la transferencia de caracteresparticulares a las variedades regionales. EI tipo demejoramiento genético en América y en Áfricafue dirigido en el híbrido inter especifico entre Carabica .1' C canephora con dos objetivosprincipales pero diferentes acorde aI pais: mejorarla calidad dei café robusta e introducir el vigor y

estan realmente por la falta de una técnica demultiplicación vegetativa.Desde 10 anos Cirad Promecafe y los paísesCentro americanos un programa de creaciónvarietal intra especifica con variedadescomerciales y silvestres (de Etiopia) Esteprograma pudo ser iniciado porque disponemosactualmente de una técnica de multiplicaciónvegetativa.
2.2.1. Las semilIasEn la naturaleza, el cafeto se reproduce porsemillas. En cultivo, el cafeto puede sermultiplicado medíante técnicas arborícolas talescomo el esqueje y el injerto (Coste, 1968). Para lamultiplicación vegetativa, se pueden utilizar másque tallos ortótropos con motivo del dimorfismovegetativo de los cafetos. Cualquier esqueje deplagiotropo lIeva a un cafeto de porte rastreandosin interés práctico (Coste, 1968).
2.2.2. EI esquejeLa multiplicación vegetativa se práctica sobretodo en C. canephora y el híbrido arabusta debidoa la imposibilidad de reproduccion conforme parvía sexuada de los individuos escogidos. En C.arabica, en cambio, donde la reproducciónconforme puede lograrse por semillas, lamultiplicación vegetativa en condicioneshortícolas se utiliza menos. Además es másprimorosa, por lo menos para el esqueje.Esta técnica conocida en el cafeto desde hacemucho tiempo (Coste, 1968) es actualmente másutilizada (Deuss y Descroix, 1984; Cambrony,1989) en C. Canephora.Se preparan los esquejes a partir de entrenudos dechupones ortótropos tomados en clones dandobuenos resultados. El tallo se corta a los 4-5 empor debajo de cada nudo y justo encima de la basede los pecíolos. Los segmentos comprenden unnudo y 2 hajas cortadas por la mitad o en latercera parte de su largo. Estos segmentos seseccionan luego longitudinalmente para obtenerasimismo 2 esquejes clivados que lIevan cada unouna hoja (Boudrand, 1974). Después detratamiento de la base con un producto auxinico,los esquejes se colocan en tanques de esqueje. EIsubstrato de arraigo puede ser arena, aserrín,parche de arroz.La tasa de enraizamiento varia acarde a los clones,ai estado fisiológico de los esquejes y ai períododei afio (Cambrany, 1989). Así, Boudrand (1974)con clones de C. canephora relata un arraigo
la resistencia a las enfermedades de robusta aC.arabica.En Costa de Marfil, Capot (1972) ernprendió lahibridación entre estas dos especies con elobjetivo de reunir en un nuevo cafeto, rusticidad,productividad y resistencia a la roya de C.canephora y valor organoléptico y bajo contenidode cafeína de C. arabica.Pudo superarse el obstáculo de la esterilidadcausado per los diferentes niveles de ploidia aicruzar genitores C. arabica por genitores C.canephora autotetraploides, lagrados después detratamiento con colchicina. Per selección c1onal,dentro de estos híbridos heterogéneos, seseleceionaron varias formas interesantes ai nivelagronómico y se difundieron bajo el nombre deArabusta.En Brasil, se encuentra otro ejemplo dei use de lahibridación inter especifica, donde se consiguiótransferir a C. arabica caracteres de resistencia ala roya anaranjada y a los nernatodos, procedentede C. canephora. Por selección de los individuosequilibrados, después de 4 a 5 ciclos de backcross con el padre C. arabica, asimismo secrearon cafetos tetraploides fértiles y resistentes,11 amados Icatu Vermelho e Icatu Amarelo(Carvalho, 1988).La hibridación inter especifica oferece un granpotencial de mejoramiento genético relacionadocon numerosas especies de Coffea. Estas poseencaracteres originales e interesantes dearquitectura, de floración y de fructificación, deadaptación pedoclimatica, de comportamientofrente a enfermedades y parasitos y, de valorcualitativo deI café.Hibridos como Congusta resultan dei cruzamientoC. canephora x C. congensis y aparecen como unmaterial interesante por sus caracteristicasagronómicas (Charrier y Berthaud, 1988).Con el objetivo de mejorar la especie C.canephora cultivada. se lleva igualmente a caboun programa de hibridación inter especifica enCosta de Marfil que deja intervenir especies cemoC. congensis, C. eugenioides, C. liberica y C.dewevrei (Louarn, 1987).
• Hibridacion infra especificaLa hibridación intra especifica fue hasta pocosaüos muy poco utilizada debido a la base genéticaestrecha que habla en los países y también a notener una técnica de multiplicación vegetativafiableHace 20 anos Van Der Vossen inicio esteprograma de hibridos FI en Kenya. conselecciones nacionales y catimor colombiano.Existe también un programa semejante enTanzania. En los dos casos dichos selecciones no
12
2.2. TÉCNICASPRODUCCIÓNPROMISORIOS:
CONVENCIONALES DEDE MATERIALES

medio deI 60 ai 90 % de las esquejes puestos entanques. Cambrony (1989) refiere una tasa deéxito deI 60 % después de 8 semanas y dei 80 %después de 12 semanas. También sefiala que en unproyecto de instalación, habida cuenta de lasincertidumbres dei medio ambiente normal de unvivero, el rendimiento medio por contar con seriadel 50 %. La producción anual de un jardin clonalde C canephora es de 150 a 200 esquejes por pié,o sea 2,7 millones a 3,6 millones de esquejes porhectarea y por afio (Cambrony, 1989).
2.2.3. EI injertoMas a menudo se reserva esta técnica a lasmanipulaciones dei materíal vegetal en los centrosde investigaciónes. Couturon y Berthaud (1979)mencionan que en Costa de Marfil, losinvestigadores dei ORSTOM practicancorrientemente el injerto, con miras a mejorar elcrecimiento de certas especies y de certos híbridoso reducir la duración de las gene raciones.No obstante, cabe sefíalarse que en certos paíseslatinoamericanos, tal coma Guatemala, el injertodo C. arabica en C. canephora es una prácticamuy utilizada para controlar los nematodos.Las técnicas de injerto utilizadas son muyclássicas: injerto de púa por rajadura, injerto poraproximación (Coste, 1968; Cambrony, 1989).Couturon y Berthaud (1979) instrumentaron latécnica de injerto de embriones de cafetos, cuyointerés principal es la recuperación de materialquo no se hubiera podido salvar en condicionesnormales de germinación de las semillas. Estaténica permite el desarrollo de certos híbridosentre especies de cafetos genéticamente apartados.
3. LAS BIOTECNOLOGÍAS APLICADAS ACOFFEA SP
3.I.GeneralidadesLa biotecnologla es un conjunto de técnicaaplicadas a un organismo, parte de un organismo,entidades sub-celulares, o proceso biológico".Desde mas de 20 afios dichas técnica son cadadias mas utilizadas en la agricultura, para tratarde responder mas rapidamiente a los problemasque enfrenta dicha agricultura moderna.
3.2.Ténica de micropropagación in vitroPor las técnicas de cultivo in vitro, se conecen dosvías de multiplicación vegetativa deI cafeto:elmicro esqueje y la embriogénesis somática.
3.2.1. Micropropagacción mediantemicroestacasGeneralidadesLa técnica de micro esqueje dei cafeto fue
13
desarrollada por Dublin (1980, 1984) y Custers(1980), el principio de esta técnica tiene por basela deI esqueje hortícola pero real izado in vitrocuyo objetivo es favorecer la caulogénesis.Tiene por finalidad de inducción in vitro de yemasaxilares existentes ai nivel do los nudosortótropos, su desarrollo y su multipl icación enserie. Esta técnica comprende tres fasesesenciales:.Instalacción deI material vegetal in vitro yobtención de micro tallos ortótropos: fase másdelicada y dificil debido a una polulación debacterías y de hongos unos días después de lainstalación de los explantes en el rnedio, y laprodución de los fenoles. Todos estos factoreslIevam a uma muerte más o menos rápida de losexplantes.• Multiplicación em serie de estas plántulasclonales.• Enraizamiento e aclimatación de estas plántulasclonales.
La técnicaInducción in vitro de yemas axilaresLos explantes provienen de tallos ortótropos deplantas en invernadero, en vivero o en el campo.Por 10 general se utilizan los 4 a 5 primeros nudos.
Multiplicación in vitro de los micro tallosoriundos de la inducción de yemas axilaresUnas 12 a 14 semanas después de la puesta encultivo, las yemas axilares desborradasevolucionan en tallos suficientementedesarrollados (3 a 4 pares de hojas), para sertomados y subdivididos en tantos micro esquejes.Se utiliza una sola citokinina, la B.A.P.(Benzilaminopurina en diferentes concentracionesde 0,5 a 3 rng/l.)Se observa que demasiado fuertes dosis decitokinina incrementan el número de micro tallospero reduce el tarnaão de los entrenudos, lo querepresenta un problema para la multiplicación deestas plántulas clonales.Los micro esquejes se siembran por 4 en un frascode 5 em de Oy 20 em de altura. Cada 90 días estasplántulas clonales se subdividen nuevamente: selogra un promedio de 30 a 35 micro esquejes pormaceta, o sea un promedio de 7 a 9 microesquejes por explant.
Aclimatación de las plántulas clonalesUnas 10 semanas después de su puesta en cultivo,las plántulas clonales tienen un desarrollosuficíente para ser tomadas y transferidas en unmedio de arraigo ya sea sólido, ya sea líquido. Enmedio sólido, las raíces no desarrollan pelosabsorbentes. Resulta preferible hacer la inducción

en rnedio líquido. Para esto el medio de base estaconstituido por los elementos minerales deI mediade Murashige y Skoog, adicionados de vitaminasde Morei y de sacarosa a 10 gr/1. Dos auxinas: elA.N.A (ácido naftaleno acética) y el A.I.B. (ácidoindolbutirico) se utilizan en dosis respectivas de50 mg/I a 100 rng/l y de una citokinina (25 mg/I).Esta inducción se hace en medio liquido no estérildurante 24 horas.Después de la inducción radicular en laboratorio,la aclimatación se hace directamente en vivero enun substrato esterilizado a la calor y compuesto de3 volúmenes de tierra, 2 volúmenes de arena, Ivolumen de pulpa de café envejecido u otroproducto químico o rnantillo.Se realiza en general bajo "túnel" plástico duranteI mes y bajo sombra (25 % de la luz). Se lograuna retoma de las p Iántulas clonales de más dei95 %
3.2.2. Micropropagación medianteembriogénesis somáticaGeneralidadesLa célula vegetal posee una característicaimportante, la totipotencia, que demuestra quecada célula contiene todo el potencial genéticopara formar un nuevo individuo genéticamenteidéntico a la célula madre.La embriogénesis somática se define como eldesarrollo de embriones a partir de célulassomáticas. Estos embriones somáticos sedesarrollan aI pasar por las fases (globular,codiforma, torpilla, cotiledonario) idénticas a lasdeI embrión zigótico (Ammirato, 1987). Estosembriones poseen una estructura bipolar(meristemos radicular y caulinar).Este modo de reproducción asexuada, que existeai estado natural en ciertas especies (Tisserat etal.. 1979) fue desde los afies 1960 amplamenteestudiado mediante el cultivo in vitro. Losprimeros ejemplos de embriogénesis somáticafueron sefíalados por Steward et aI. (1958) Y porReinert (1958) a partir de células de Daucuscarota. Treinta afíos después, la embriogénesissomática fue descrita en más de 130 especiesperteneciendo a 30 famillas diferentes (Thorpe,1988).
La técnicaLos primeros trabajos de embriogénesis somáticasobre el cafeto fueron publicados por Staritsky(1970). Consiguió eI desarro110 de cales vigorososa partir de explantes de tallos de C. arabica, C.canephora y C. liberica, pero solos los cales de C.canephora se diferenciaron en embrionessomáticos y plántu Ias.Sharp et aI. (1973). trabajando sobre distintos
14
explantes de C. arabica, relatan que el potencialmorfógeno varia acorde aI explant utilizado. Asi,los cales logrados a partir de tallos plagiotroposno son morfógenos, los oriundos de hojas fonnanraíces, los oriundos de anteras forman proembriones y los logrados a partir de tallosortótropos forman plántulas.Investigacianes realizadas en el transcurso de losúltimos viente afíos pennitieron concluir que laembriogénesis somática puede ser inducida apartir de explantes de origen variado:.tallos (Dublin, 1980; Staritsky y Van Hasselt,1980),.hojas (Herman y Hass, 1975; Sondahl y Sharp,1977; Dublin, 1981; Yasuda et at., 1985),.ovulas (Lanaud, 1981),.anteras (Ascanio y Areia, 1987),.protoplastas (Spiral y Pétiard, 1991; Acuna yPena, 1991)..hojas(Berthouly y Michaux-Ferriere)Estos resultados demuestran la aptitud de estaespecie para la regeneración. No obstante,depende mucho de la naturaleza y deI origen deIexplant. Así, existe una gran diferencia dereacción entre tallo o hoja o entre especies, porejemplo, entre C. canephora o C. arabica(Staritsky y Van Hasselt, 1980).Para conseguir I a diferenciación de los diversostipos de explantes, se utilizó una gran variedad desecuencias de medios nutritivos. De manerageneral, la embriogénesis somática puede lograrseacorde a 2 estrategias:1. en una sola fase por cultivo de los explantes enun media único. En este caso el medio puedecontener ya sea una citokinina sola (Dublin, 1981;Yasuda et aI. 1985), ya sea una auxina y unacitokinina en asociación (Dubl in, 1980; Pierson etaI., 1983).2. en dos fases por cultivo de explantes en unprimer medio de cultivo para iniciar unaproliferación de cales, y luego en un segundo paradesarrollar células embriogenas en embriones(Sõndahl y Sharp, 1977; Dublin, 1984, Berthoulyy Michaux-Ferriere 1996).Sondahl y Sharp (1977) describen dos secuenciasde diferenciación morfogénetica a partir deexplantes foliares de C. arabica que fueron11 amadas "High Frequence SomaticEmbriogénesis' (HFSE) y "Low FrequenceSomatic Enibriogénesis" (LFSE)."LFSE' está caracterizada pOr la aparición deembriones aislados bien constituidos (I a 10 porexplant) observados después de 13-15 semanas enel medio de inducción. En "HFSE", los embrionesse desarrollan a partir de cal friable, este mismooriundo de un cal primario. Los embrionesaparecen más tardiamente que en el caso de LFSE

(16 a 20 semanas). En los cultivos s de tipo"HFSE", a cantidad de embriones varia de 50 a100 embriones (Sôndahl y Sharp, J977).Dublin (1980, 1984) menciona que el proceso deembriogénesis somática en el genero Coffea sedistribuye acorde a 3 fases:• Ia fase de caulogénesis se desarrolla en laobscuridad (4 semanas) en un medio que contienereguladores de crecimiento: una o dos auxinas(2,4 D, A.I.B.) solas o en cornbinación. y unacitokinina (BAP, 2ip).• Ia fase de díferenciación se desarrolla a la luzindirecta en medio que contiene una citokininasola (BAP) o en asociación con una auxina (2,4D). Esta fase dura 8 a 10 semanas.• La fase de desarrollo de los embriones en unmedio que contiene A.I.A. y kinetine ai BAP solo.Esta fase dura 10 semanas. En este medio losembriones se desarrollan hasta la formación deplántulas de 3 a 4 pares de hojas aptas a latransferencia en invernadero.Staritsky y Van Hasselt (1980) publicaron elprirner trabajo sobre la embriogénesis somática enmédio líquido en C canephora. Tiene por base laexplotación de embriogénesis adventiva. En estemétodo, se utiliza e cal logrado en medio semisolido (6-8 semanas) para iniciar el cultivo enmedio líquido. Los primeros embriones (5001000) aparecen 8 a 10 semanas después de latransferencia en rnedio liquido. Las produccionesulteriores de embriones se consiguen entonces porbrote de los embriones y por formación de novo apartir de los agregados celulares. Estos autoresnotan la necesidad de estudiar los aspectoscuantitativos dei procedimiento y los problemasde aclimatación de las plántulas in vitroPoniendo de lado los trabajos anteriores, otrosautores utilizaron el medio líquido para la fase dedi ferenc iación, para mejorar la embriogénesissomática (De Pena, 1983). No obstante, noconsiguieron suspensión celular mantenida portrasplante regular.EI exemplo de suspensiones celularesembriogenas de cafeto fue relatado ya sea con elobjetivo de una producción masiva de embriones(Zamarripa et al., 1991), ya sea como material deinicio para el aislamiento de protoplastas y suregeneración ulterior (Spiral y Pétiard, 1991;Acuna y Pena, 1991).
Factores inportantes para la embriogénesissomáticaLa forrnac ión de embriones somáticos parece sere l resultado de una interacción muy estrecha entrediversos factores, Ast, Rangaswamy (1986) relataque casi 50 substancias perteneciendo a unas 10clases de compuestos orgánicos e inorgânicos.
15
jugos naturales, extracto de plantas y algunosfactores fisicos se describieron como pudiendoinfluenciar la embriogénesis somática.
Medios de cultivoEntre los componentes dei medio de cultivo másestud iados, se puede citar las aux inas. En efecto.el proceso de la embriogénesis se inicia muy amenudo en un medio que contiene auxinas, mascorrientemente 2,4-D (Ammirato, 1983).También se conocen las citokininas paradesempenar un papel en la maduración de losembriones somáticos. Fujimura y Komanine(1975) observaron una especificidad de lasnecesidades en citokinina: la zeatina favorece laembriogénesis de las suspensiones celulares deDaucus carola. mientras que la BAP y la kinetinela inhiben. Por otro lado, la BAP se mencionacomo teniendo influencia en la ernbriogénesissomática de Coffea arabica y dei híbrido Arabusta(Dublin, 1981: Yasudaela/.. 1985).EI ABA desempena un papel en la maduración yla germinación de los enbriones (Arnrnirato,1977). Se puede citar a título de ejemplo el efectofavorable del ABA en la maduración y lagerrninación de los embriones somáticos deGlvcine max (Ranch et al.. 1985) y de Picea abies(Boulay et UI. 1988). En cambio, Tisserat yMurashige (1977a) indican que el ABA inhibe lainducción de las embriones somáticos de Dal/cus
carola.
También se ha descrito a menudo la importanciadei nitrógeno y de certos ácidos aminados. EInitrógeno desempena un papel esencial a la vez enla iniciación y en la maduración de los embriones(Arnmirato. 1983). Los ácidos aminados, yespecialmente la glutamina, parecen desempenarun papel particular (Arnrnirato. 1983). La prolinamejora la embriogénesis en Zea mays (Arrnstrongy Green, 1985). En la soya (Glycine max), Iaglutamina, la alanina y la asparagina son factoresimportantes para el crecimiento de suspensionesembriogenas (FineI' y Nagasawa. 1988).La sacarosa es la fuente de carbono más eficaz(Gautheret, 1959) aunque otros azúcaresmostraron efectos favorables. Por ejernplo. lamaltosa estimula la embriogénesis somática enDal/cus carota (Kinnersley y Henderson, 1988) yGlycine max (Obendorfy Slawinska, 1988).
Medio ambiente gaseoso y fisicoLa diferenciación puede también controlarsemediante la concentración de oxigeno disuelto. EnDaucus carola )' Triticum aestivumconcentraciones reducidas de oxigeno favorecenla embriogénesis somática. AI contrario, loscultivos de Medicago saliva necesitan

concentraciones altas de oxigeno para ladiferenciación embrionar (Stuart et al.. 1987).Se menciona a menudo el papel deI etileno en laembriogénesis (George y Sherrington, 1984).Tisserat y Murashige (1977b) relatan que eletileno inhibe la iniciación de los embriones deCitrus reticulata y de Daucus carota..Se mencionaron hace de eso mucho tiempo losefectos en la embriogénesis somática de otrosfactores dei medio ambiente como la luminosidady la temperatura (Amrnirato. 1983).
Densidad celularLa densidad celular fue sefíalada como unparámetro relevante para el crecimiento de lassuspensiones celulares. Halperin (1967) mencionaque la producción de embriones somáticos dernuestra de perforación esta intluenciada por ladensidad celular. Por debajo de un umbral críticola maduración de cmbriones se halla inhibida.Hari (1980) muestra. para una suspensión dezanahoria. que existe una relación linear entre ladensidad celular a la inoculación y número deernbriones producidos y los cultivos de células. demuy baja densidad o. ai contrario. rnuy alta noexpresan la totalidad de su potencialembriogénico. Este autor rnuestra que elcrccirncnto de ernbriones somáticos por debajo deun umbral mínima de densidad puede mejorarsc aiagregar un medio "acondicionado".Mas recentemente. Ozawa y Komamine (1989)indiean que la densidad de los agregados celulareses uno de los tàctores más importantes para larcgcneración de suspensiones celulares de Orizusat iva L.En el marco de este trabajo hemos observado quela densidad de inoculación es un parámetroprimordial por determinar para una producciónóptirna de embriones somáticos de cateto.
GenotipoLa regeneración de plantas a partir de los cultivoscelulares fue mencionada como dependiente deigenotipo para varias espécies como .\fedicag() sp(Brown ) Atanassov, 1985). UICllne max L.(Parrot c! ai .. 1989). Pisum sativum L. (KyselyJacobsen, 1990). Brassica olcracca (Phippen yOckendon. 1990) y Vitis viniferu (Rajasekaran )Mullins.1983).Meijer y Brown (1985) hacen informe de que de19 genoti pos de 4 subespecies de .\ledicaglisaliva, solamente 8 forrnaron ernbrioncssomáticos. Brown y Atanassov (1985) observanuna variabilidad importante entre 76 cultivares dealfalfa Medicago sp, sometidos a prueba por suaptitud a la ernbriogéncsis somática.La lista de los factorcs citados previamente no es.
16
claro está, exhaustiva pero ref1eja no obstante ladiversidad de los parámetros que intervienen en laembriogénesis somática, diversidad quedemuestra la complejidad de la manifestación dela totipotencia celular.
Multiplicación vegetativa por embriogénesissomática de massa. Principio y estado de latécnicaLa multiplicación de celulas embriogenas ensuspensión permite una propagación de las plantasa gran escala (Arnmirato, 1983).La utilización de la embriogénesis somática enmedio líquido como técnica de multiplicaciónnecesita el dominio de diferentes pasosimplicados en el procedimiento. Esteprocedimiento puede repartirse en 4 fasesprincipales (Pétiard et al., 1992)
• estabelecimiento de las cepas celularesernbriogenas.• multiplicación y conservación de las cepas.• Producción de ernbriones somáticos en bio
reactor,• desarrollo de los ernbrioncs somáticos y
conversión en plántu las.
Estabelecimiento de las suspensiones celularesembriogénicas[I cstabelccirniento de las suspensioncsembriogenas comprende por lo general :2 fases:inducción de cales ernbriogénicos en médio semisol ido e instalación de suspensiones celulares portransferencia de estos cal los en medio líquido. Laprimera eta pa, que consiste en log rar ca1I0s. serealiza por lo general en un medio semi-sol idoque contiene una auxina asociada o no con unacitokinina (Arnmirato. 1983) para permitir laadquisición de la competencia a la cmbriogéncsis.EI genótipo. Ia cornposición dei medio, lascondiciones de cultivo son factores importantespara lograr cultivos embriogénicos (Ammirato.1983: Thorpe. 1988).En la segunda etapa. el cal crnbriogénico setraslada en medio líquido con una densidad deinoculación y un media especifico para cadacultivo. para conseguir una población de tejidosque pueda mantenerse med iante trasplantesregulares. (Van Boxtel y Berthouly. 1996).EI rncdio ) las condiciones de cultivo utilizadospueden intluenciar la morfologia celular yproducir diferentes tipos celulares que cocx istcnen c! cultivo. A titulo de ejcmplo. los cultix osindifcrcnciadas de rnucstra de perforacion cstánconstituídos de :2 tipos celulares. EI primero estáconstituido de células meristámaticas de pequenotamano. de citoplasma denso con granos dealmidón. EI segundo tipo corrcspandc a células

parenquimatosas aisladas, de gran tarnaãofuertemente alargadas y muy vacuolizadas. Soloel prirner tipo celular es ai origen de losembriones.En ciertos casos, el tamafio de los agregadosparece correlado con la capacidad embriogenica.AsL para la muestra de perforación, se inicia laembriogénesis después de selecionar agregados.cuyo tamafio esta incluido entre 50 y 100 um.Esta fracción se conoce por ser mas embriogenica.En cambio. en Ipomoea batatas, los embriones seforman a partir de los agregados de tarnafiosuperior a 180 11m.
Multiplicación y conservación de suspensiones:La multiplicación y la conservación desuspenciones se realizan habitualmente mediantetrasplante regular. La frecuencia de los trasplantesy la densidad de inoculación necesarias aimantenimiento de una cepa se determinan paracada una de ellas, En general. la frecuencia es deaproximadamente una a tres semanas.Una limitación importante para la aplicacióncomercial de este procedimiento es la pérdida deipotencial embriogeno observada en el transcursode los trasplantes sucesivos (Smith y Street,1974). La duración de mantenimiento de estepotencial es diferente acorde a las especies.Para paliar esta limitación, se deben integrarestrategias de conservación (crioconservación,disecación) en el procedimiento (Pétiard et ai ..1992). La técnica de crioconservación fueaplicada para las cepas embriogenas de diferentesespecies coma Picea glauca, Saccharum sp,Daucus carota y CofJea canephora.
Producción de embriones somáticos en mediolíquidoLa multiplicación a gran escala implica que sedomine la produción de embriones somáticos enmedio líquido. La optimización de esta fase debetomar en cuenta la tasa de multiplicación y lacalidad de los embriones producidos que sonfactores importantes desde el punto de vistaaplicada.El desarrollo de embriones somáticos a partir deuna suspensión celular se logra mediantetransferencia de cierta cantidad de tejidos en unmedio de expresión. Este medio se hallageneral mente desprovisto o con muy bajaconcentración de auxina (Ammirato, 1983).El métode de cultivo más común parasuspensiones es en frascos de Erlenmeyer (100 o250 ml ), método muy práctico en el marco de unaoptimización. No obstante. para una producciónde embriones somáticos a gran escala. Iauti Iizac ión de bio-reactor puede resu Itar
17
indispensable. Además, el bio-reactor permitemedir y controlar diferentes parámetros como lasconcentraciones de oxigeno. de dióxido decarbono. el pl-l, y también una automatizaciónmás fácil dei procedimiento (Pétiard et ai .. 1992).La utilización dei bio-reactor para la producciónde embriones somáticos tomó el vuelo en losúltimos afios pero sigue siendo todavia limitada aalgunas especies. Se mencionan ensayos defactibilidad y de "scaling-up' para Daucus carola.para Medicago saliva y para Poinsettia. EI númeroreduc ido de trabajos es comprensible, dado que elcultivo en bio-reactor impl ica en prirner lugar unbuen dominio de la embriogénesis somática enmedio líquido pero también unos conocimientosespecializados en genio bioquímico.
. EI paso dei frasco de Erlenmeyer aI bio-reactorpuede producir una reducción de la eficácia de loscultivos. Así Zamarripa (1989) en un ensayo de"scaling-up" en una suspensión celular do cafeto,relata una producción de embriones en bio-reactor20 veces inferior a la observada en frasco deErlenmeyer. Stuart et al. (1987) logran en bioreactor 1600-6600 embriones de Medicago salivapor gama de cales. mientras que logran 300020000 en frasco de Erlenmeyer.
Desarrollo de los embriones somáticos yconversión en plántulasLa última fase de este procedimiento demultiplicación es el desarrollo de los embrionesen plántulas aptas ai cultivo en vivero o en campo.Esta fase se desarrolla normalmente en mediosemi-sol ido. Redenbaugh (1990) indica. que parala producción de semi lias artificiales, Ia calidad delos embriones y su frecuencia de conversión enp Iántulas deben optimizarse.Stuart et al. (1987) notan que la utilización de bioreactores afecta la tasa de conversión en plántulasde los embriones. Así, Ia conversión de losembriones de Medicago saliva producidos en bioreactor no es más que dei 2 ai 3 % contra el 70 ai90 % de conversión para embriones logrados enmedio semi-sol ido.En caso de modificarlo, el desarrollo deembriones somáticos puede provocaranormalidades ulteriores. Los autores subrayanque una de las dificultades mayores para aplicaresta técnica es el control do la calidad de losembriones.En conclusión. cada etapa presenta problemasdiferente para los cuales deben realizarse unestudio detenido y una optimización adelantadapara tomar el procedimiento cualitativa yeconómicamente v iable.

18
3.3. Mejoramiento no convencional
presentados en este 111 Simposio.
3A.Los marcadores moleculares:En los ultimes anos cl desarollo de nuevastécnicas de biologia moleculares han abiertonuevas oportunidades para el mejoramientogenético dei café la utilizacion de marcadoresmoleculares dcberan assistir v apoyar a la
3.3.I.Producción de haploidesHubo algunos resultados interesantes en Portugal.Colombia y Costa Rica.Esta técnica podria serutilizada para la creación de pantas haploides . ydespues de doblar el número de crornosomas concolchicina, podriarnos obtener plantas dihaploideshomocigotas para las carectiristicas deseadas.Perolastimamente no es una técnica que funcionaactualrnente en café. y por cuensecuencia no estáuti Iizada en los programas de mejoramientogenético.
biotecnologiasestrategia de
3.3.2. Transformación genéticaLos progresos realizados enpermiten pensar en una nuevamejoramiento genetico:
La transformacián genéticaEs la incorporación de "un gene en una célula.Dicha técnica está utilizada por el momento parala resistencia contra los insectos. pero puede serutilizado por otros problemas como la calidad.El desarrollo de una técnica de micropropagaciónvegetativa. como la embriogénesis somática. hapermitido poner mas enfasis en dicha técnica.En 1993 los investigadores de la N ESTLÉ enFranciaobtuvieron por la primera vez plantas decafé transgenicas, utilisando Agrobacteriumrhizogcncs.Dicha bacteria tiene la facultad de transferir en lascélulas vegetales una parte de su ADN. En base:
• estos résultados(Nest Ié)• identificación de genes de Bacillus thurigiensis(trabajo de tesis desarollada en el Cirad por uncstudiante de Brasilj)
• el desarrollo de una técnica de embriogénesissomáticase ha iniciado un programa de cooperación entrela Nestlé y el Cirad desde 1994.EI objetivo de dicho programa es dcsarrollar untécnica de transformación genética para laintroducción de genes de resistencia a diferentesinsectos dei café: minador de la hoja y la broca.Actualmente se logrado obtener mas de 100plantas transgenicas diferentes con genes deresistencia ai minador de la hoja.
lnmersián temporariaEn eiRA!) se ha desarrollado una nueva técnicallamada "inunersion temporária". Dicha técnicautiliza Ull nue vo recipiente producido por elClRAD. y permite utilizar e l medio liquido.Con este sistema se puede producir, en e l rnismoreei pierue. ernbrioncs ac Iimatables como lo hademonstrado el DI' H.Etienne en su trabajorealizado cn cl CA TIF cuyos resultados scrán
Semillas artificialesEI concepto de "semi llas artificiales" implica quese utilicen embriones somáticos cubiertos concapsulas para la propagación a gran escala de losgenotipos interesantes ai nivel agronómico(Redenbaugh et al., 1986). Una de las estrategiasconsiste en envolver los embriones somáticos enun gel de alginato adicionado con elementosnutritivos esenciales para asegurar la germinaciónde los embriones. EI todo estaría protegido de ladisecación por una capa protectora formada de unfilm de resina. soluble en agua (Redenbaugh etal., 1986).Las "sernillas artificiales" permitirian por lo tantoasociar las ventajas prácticas de la semilla(resistencia. conservación) y las relacionadas conla multiplicación vegetativa dei materialse lecc ionado.EI empleo de "sernillas artificiales" para amultiplicación en musa presenta no obstanteciertas limitaciones. Implica necesariamente undominio de la embriogénesis somática quepermite una alta producción de embriones debuena calidad. Ahora bien. a la fecha. rnuchasespecies tienen una baja aptitud ya sea a laformación de ernbriones somáticos. ya sea a suconversión en plántulas.La aplicación de esta tecnologia "semi liasartificiales" implica por lo tanto lainstrumentac ión de las diferentes etapas(regeneración por ernbriogénesis somática."scaling-up", conservación, semillas en píldoras).Para esto. en 1987. se inició un programa deinvestigación que reunia las empresas Limagrain.Rhône Poulenc Agrochimie y Francereco-Nest Ié.las metas eran los híbridos de tomate. los portasemillas de muestra de perforación y el cafeto(Pétiard. cornrn. pers.).Los genotipos interesantes por multiplicarmediante "semillas artificiales" serian los quepresentan un alto \ alor genético aãadido. talescomo híbridos de espccies hortícolas o hortelanas(tomate. melón. apio) o especies perennes (árbolestorestales. trutales o tropicales) o aún individuosdando resultados particularmente buenos oriundosdei cultivo 111 vitro, por ejernplo de latransformación genética (Monnier. 1988).

seleccion.Así y sin entrar en muchos detalles, el progresoreal izado en dicha técnica deberia permitir:• estudiar la variabilidad genética que existe enCoffea sp. abriendo la base genética de Coffeaarabica . y permitiendo una mejor selecion de lospadres utilizados en los cruzamientos• estudiar y selecionar los genes de rnayor
importancia• estudiar en los cruzamientos y retrocruzamientosla transferencia de los genes• realizar una mapa genética deI café• estudiar la variabilidad de los patogenos queatacan el café• estudiar y caracterizar los niveles de resistenciaa enfermedades y plagas
3. 5. Conservación de germoplasma:La conservación de germoplasma puede realizarsede dos manera:• conservación de micro-plantas en medio simplesin hormonas y baja temperatura (20 grados)
• conservación en nitrogeno líquido(crioconservacion)
Estas son las dos técnicas principalmenteutilizadas en la conservación do germo plasma docafé.
4.POTENCIALIDADES Y PERSPECTIVASDE LAS BIOTECNOLOGIAS EN CAFÉ:
4.I.Potencia lidades:I. Acortar el ciclo de selección para C arabicamediante:• La multiplicación conforme in vitro de híbridos(FI. F2. retrocruzarnientos),
• haplométodo (androgénesis).2. Multiplicación rápida de genotipos de Ccanephora, o la multiplicación hortícola planteaproblemas (regiones do media altitud).3. Transformación genética• obtención de resistencias a los insectos(ejemplo: utilización de genes de Bacillus.obtención de plantas masculinas-estériles (paraproducción de semillas FI do C. arabica y• obtención de resistencia a los herbicidas.
4.2.PerspectivasCual es el futuro do las biotecnologias?Esodepende exclusivamente de la decisión de cadapaís. Pero hay que tener en cuenta actualmente .que son técnica importantes para el desarrollop dela agricultura do rnafiana en general. y de lacafeicultura en particular.Por eso hay que saber:• que la bioteonologia una técnica ai servicio dei
19
investigador o del fitomejorador: eso significa quesi no hay cambio en las estrategias domejoramiento no hay interés en las biotecnologias
• que con la transformación genética es un nuevoconcepto del mejoramiento. y que tanto elinvestigador de rnaãana . como el "publico" debeser preparados• que las biotecnologias no van a resolver todoslos problemas de la agricultura• que debido a estas nuevas tecnologias existevárias peguntas tanto aI nível nacional comointernacional:Que tipo de técnica? Que legislaciones o regias?Que tipo de plantas transgenicas?
5. CONCLUSION
Hoy dia. Ias biotecnologías hacen parte entera dela agricultura moderna.AI início dei secundo "rnilenario", hablar deinvestigaciones agronómicas sin hablar debiotecnologia, parece inconcebible, porque escierto que dicha tecnologia abre nuevasexpectativas.En el caso de la cafeicultura moderna. el uso de labiotecnologia deberia permitir:• responder mas rápidamente a los problemasplanteados por los productores acertando. porejernplo, el ciclo de selección de nuevasvariedades (híbridos F I despues 10 anos).• multiplicar masivamente genótiposseleccionados por embriogénesis somática(variedad nernaya),
• abrir nuevas estrategias de mejoramientogenético. por transformación genética (resistenciaai minador de la hoja, o la broca).Pero, es rnuy importante tomar en cuenta:• que dichas "herrarnientas biotecnologicas"pueden ser utilizadas solamente si hay un realcambio de mental idad en la estrategia deinvestigaciones•• que la biotecnologia no va a resolver todos losproblemas. pero viene y debe ser consideradacomo un apoyo a las investigaciones.
6. REFERENCIAS BIBLlOGRAFICAS
ACUNA J. R., DE PENA M., 1991. Plantregeneration from of embryogenic cellsuspensions of CojJea arabica L. cv. Caturra.Plant Cell Reports, 10:345-348.AMEHA M., 1983. Heterosis in crosses ofindigenous coffee selected for yield and resistanceto coffee berry disease. I. First bearing stage. ActaHorlicultural, 140: 155-161.AMMIRATü P. V., 1977. Hormonal control 01'

somatic embryo development from cultured cel\sof caraway : Interactions of abscisic acid, zeatinand gibberellic acid. Plant Physiol., 59:579-586.AMMIRATO P. V., 1983. Embryogenesis. In .Handbook of Plant Cel! Culture: Techniques forpropagation and Breeding. D.A. Evans, W.R.Sharp, P.V. Ammirato el Y. Yamada, eds.,Macmillan Publishing Cornpagny, New York, 1:82-123.AMMIRATO P.V., 1987. Organized eventsduring somatic embryogenesis. In: Plant BiologyPlanl Tissue and Cel! Culture, C.E. Green, D.A.Sorners, W.P. hackett, D,D. Biesboer, ed. NewYork, Alan R. Liss, lnc., 3:57-81.ARMSTRONG C. L., GREEN C. E., 1985.Establishment and maintenance of friable,embryogenic maize cal\us and the involvement ofL-proline. Planta, 164:207-214.ASCANIO E. C. E., ARCIA M. M. A., 1987.Haploids from anther culture in Coffea arabica L.In : International Congress de Plant TissueCulture, Tropical Speeies; (Bogota, 21-25 Sept.)Abstact p.68BERTHAUD l ,\ 985 Propositions pour unenouvelle strategie d'améliorationdes caféiers deI .espécie C.canephora, basée SUl' les résu Itats deI'analyse des populations sylvestres. In: I liêrneColloque Scientifique International SUl' le Café(Lomé, 11-15 février) ASIC:Paris 445-452.BERTHOULY M. Y MICHAUX-FERRIERE,1996.High frequency somatic embryogenesis In.:Coffea eanephora: Induction conditions andhistological evolution. Plant Cell, Tissue andOrgan Culture 44: 169-176BERTHOUL Y M., GUZMAN N., CHATELETP., 1987. Micropropagation in vitro de différentesligneés de Coffea arabica V. catimor. In.: l ZémeColloque Scientifique International sur le Café(Montreux, 29 juin-3 juillet), ASIC, Paris, p. 462467.BETTENCOURT A. X. r., 1982. Variedades decafé arabica resistentes a la roya y perspectivaspara su utilización en la caficultura deI futuro. In :V Simposio Latinoamericano sobre caficultura(San Salvador, El Salvador) ; IICA, traducción deIportugués por J.1. Echeverri, 20 p.BETTENCOURT A. J .. 1983. Característicasagronomicas de seleções derivadas decruzamentos entre híbrido de Timor e asvariedades Caturra. Vil Ia. Sarchi e catuai. In.:Simposio sobre Ferrugens do Cafeeiro (Oeiras,)7-20 Octobre). Centro de Investigação dasFerrugens do Cafeeiro, Oeiras : 353-373.BETTENCOURT A. J., RODRIGUES C. l l R..1988. Príncipes and practice of coffee breedingfor resistance to rust and other diseases. Chapter6. In.: Coffee Agronomy, R.1. Clarke, R Macrae,
20
eds., Elsevier Applied Science Publishers LTD,England, 4: 199-234.BOUHARMONT P., AWEMO r., 1979. Laselection végétative du caféier robusta auCameroun.Café Caeao Thé . 23:227-237.BROWN D. C. W., ATANASSOW A., 1985.Role of genetic background in somaticembryogenesis in Medicago. Plant Cel! TissueOrgan Culture, 4: 11 I. 112.CAMBRONY H. R., 1989. Le caféier. In : LeTeehnicien d 'Agriculture Tropieale. Maisonneuveet Larose et A.C.C.T., 166 p.CAPOT J.. 1972. L' amélioration du caféier enCôte dlvoire : les hybrides 'arabusta'. CaféCacao Thé, 16 (I ):3-18.CAPOT J.,1975. Obtention et perspectives d'unnouvel hybride de caféier en Côte d'lvoireL' Arabusta. In: Teme Colloque SeientifiqueInternational sur le Café (Hambourg, Allemagne,9-14 juin), ASIC, Brême, 449-457.CARVALHO A., 1982. Melhoramento docafeeiro cruzamentos entre C. arabiea e C.eanephora. In.: 10eme Col!oque ScientifiqueInternational sur le Café (Salvador-Bahia, I 114octobre), ASIC, Paris 363-368.C ARVALHO A., 1988. Principies and practice ofcoffee plant breeding for productivity and qualityfactors: Coffea arabiea. Chapter 4. In: Coffee ;Agronomy. R.l Clarke, R. Macrae, eds.. ElsevierApplied Science Publishers LTD England, 4: 129160.CHARRIER A., 1977. La structure génétique dugeme Coffea; ses conséquences pourI'amélioration des caféiers cultivés. In : SémeCol!oque Scientifique International sur le Café(Abidjan, 28 novembre - 3 decernbre), ASIC,Paris, 399-405.CHARRIER A.. 1980. La conservation desressources génétiques du geme Coffea. (CaféCacao Thé, 24 (4):249-256.CHARRIER A., 1985. Progres et perspectives del' amei ioration génétique des caféiers. In.: 11émeColloque Scienfique International sur le Café,(Lomé, 11-15 février), ASIC, Paris 403-425.CHARRIER A., BERTHAUD J., 1988. Principiesand methods in coffee plant breeding: Coffeaeanephora Pierre. Chapter 5. In : Coffee:Agronomy. R.1. Clarke, R. Macrae, eds., ElsevierApplied Selence Publishers LTD, England. 4:167195.COSTE R., 1968. Le caféier. G.P. Maisonneuve etLarose, Paris. 310 p.COUTURON E., BERTHAUD 1.. 1979. Legreffage dembryons de caféiers. Café Caeao Thé,23( 4):267-270.CUSTERS J. B. M., 1980. Clonal propagation ofCoffea arahiea L. by nodal culture. In: geme

Characterization ofHemileia vastatrix inKouillou. Euphytica,
Colloque Scientifique International sur le Café(Londres, 16-20 juin), ASIC, Paris, 589-596.DE GARCÍA E., MENÉNDEZ A., 1987.Embriogénesis somatica a partir de explantesfoliares deI cafeto "catirnor", Café Cacao Thé,31(1):15-22.DE PENA M., 1983. Somatic embryo inductionand plant regeneration from Coffea canephoraand Coffea arabica. In : Simposio sobreFerrugens do Caffeeiro (Oeiras, 17-20 octobre),Centre de Investigação das Ferrugens do Cafeeiro,Oeiras, Portugal, 494-512.DUBLIN P., 1980. Multiplication végétative invitro de larabusta. Café Cacao Thé, 24(4):281289.DUBLIN P., 1982. Culture de tissue etamélioration génétique des caféiers cultivés. In :10eme Colloque Scientifique International sur leCafé (Salvador-Bahia, 11-14 octobre), ASIC,Paris, 433-459.DUBLIN P., 1984. Techniques de reproductionvégétative in vitro et amélioration génétique chezles caféiers cultivés. Café Cacao Thé, 28(4):231244.ESKES A B.,1983.incomplete resistence toCoffea canephora cv.32:632-648.ESKES A. B., 1991. Compte-rendu de mission,Instituto Agronômico Campinas, SP, Brasil (8-18janvier), IRCC/CIRAD, Montpellier, France, 5p.FINER l J., NAGASAWA A., 1988.Development os an embryogenic suspensionculture of soybean (Glcilne max Merill.). PlantCeU Tissue and Organ Culture, 15: 125.136.FUJIMURA T., KOMAMINE A., 1975. Effets ofvarious growth regulators on the embryogenesisin a carrot cell suspension culture. Plant ScienceLetters, 5:359-364.HALPERIN W., 1967. Population density effectson embryogenesis in carrot-cell cultures.Experimental CeU Research, 48: 170-173.HARI V., 1980. Effect of cell density changes andconditioned media on carrot cell embryogenesis.Z.pflanzenphysiol., 96:227.23 I.HERMAN E. 8., HAAS G. l,1975. Clonalpropagation of Coffea arabica L. from callusculture. HortScience, 10(6):588-589.KYSELY W., JACOBSEN H. J., 1990. Somaticembryogenesis from pea embryos and shootapices. Plant CeU Tissue and Organ Culture,20:7-14.LOUARN 1., 1987. Possibilités d'amélioration del'espéce cultivé Coffea canephora Pierre parhybridation inter spécifique avec les caféiersafricains diploides données cytogénétiques sur leshybrides F I. In: lêême Colloque Scientifique
21
Internacional sur le Café (Montreux 29 juin - 3juillet), ASIC, Paris, 441-452.MEIJER E. G. M., BROWN C. W., 1985,Screening of diploid Medicago sativa germplasmfor somatic embryogenesis. Plant CeU Reports,4:285-288.MONNIER M.,1988. Embryogenêse zygotique etsomatique. In Cultures de Cellules, Tissus etOrganes Végétaux, Fondements Théoriques aUtilisations Pratiques, J. P. Zryd, ed., PressesPolytechniques Rornandes, Lausanne, I 19-134.OZAWA K" KOMAMINE A., 1989.Establishment of a system of high-frequencyembryogenesis from long-term cell suspensioncultures of rice (Oryza sativa L.). Theor. AppI.Genet., 77:205-211.PARROT W. A, WILLLAMS E. G.,HILDEBRAND D. F., COLLINS G. B., 1989.Effect of genotype on somatic embryogenesisfrom immature cotyledons of soybean. Plant CeUTissue and Organ Culture, 16: )5-21.PÉTlARD V., COURTOIS O., MASSERET 8.,DELAUNAY P., FLORIN B., 1987. Existinglimitations and potential applications of plant cellcultures. In.: Proceedings 4th European Congresson Biotechnology (Amsterdam), O.M. Neylssl,R.R. Van der Meer, K.C.A.M. Lyuben, eds,Elsevier Science Publisher B.V., 157-169.PHIPPEN c., OCKENDON D. J., 1990.Genotype, plant, bud size and media factorsaffecting anther culture of caul iflowers (Brassicaoleracea varo botrytis). Theor. Appl. Genet.,79:33-38.PIERSON E. S., VAN LAMMEREN A. A M.,SCHEL J. H. N., STARITSKY G., 1983. In vitrodevelopment of embryoids from punched leafdiscs of Coffea canephora. Protoplasma,115:208-216.RAJASEKARAN K., MULLINS M. G., 1983.Influence of genotype and sex-expression onformation of plantlets by cultured anthers ofgrapevines. Agronomy, 3 (3):233-238.RANCH J. P., OGLESBY L., ZIELINSKI A c.,1985. Plant regeneration from embryoderivedcultures of soybeans. In vitro Cellular andDevelopmental Biology, 21 :653-658.RANGASWAMY N. S., 1986. Somaticembryogenesis in angiosperm cell tissue andorgan cultures. Proc. Indían Acad. Sei. (PlantSci.),96(4):247-271.REDENBAUGH K., 1990. Application ofartificial seed to tropical crops. HortScience,25(3):251-255.SHARP W. R., CALDAS L. S., CROCOMO O.r, MONACO L. c, CARVALHO A..1973.Production of Coffea arabica callus ofthreeploidy levels and subsequent morphogenesis.

Phyton, 31(2 ):67-74.SMrTH S. M., STREET H. E., 1974. The declineof embryogenic potential as callus and suspensioncultures of carrot (Daucus carota L.) are seria\lysubcultured. Ann Bot, 38:223-241.SONDAHL M. R., SHARP W. R.. 1977. Highftequency induction of somatic embryos incultured leaf exp1ants of CofJea arabica L.. Z.Pflanzenphysiol. Rd., S. 81 :395-408.SPIRAL J.. PÉTIARD V.. 1991. Protoplastculture and regeneration in Colfea species. In.:J4eme Colloque Scientifique International SUl' leCafé (San Francisco, 14-19 juillet), ASIC, Paris.383-391.SRINIVASAN C S.. VISHVESHWARA S.,1978. Heterosis and stability for yield in arabicacoffee. Indian Journal 0/ Genetics and PlantBreeding, 38(3):416-420.STARITSKY G.. 1970. Embryoid formation incallus tissues of coffee. Acta Bot. Neerl.,19(4):509-5 J4.STARITSKY G.. VAN HASSELT G. A. M..1980. The synchronised mass propagation ofcoffea canephora in vitro. In geme Colloquelnternational SUl' le Café (Londres. 16-20 Juin),ASIC Paris. 597-602.STEWARD F. C. MAPES M. O.. MEARS K,1958. Growth and organized development ofcultured cells. 11. Organization in cultures growthfrom freely suspended ceIls. Am. .J Bot., 45:704708.STUART D. A.. STRICKLAND S. G.. WALKERK. A.. 1987. Bioreactor of alfala sornaticembryos. HortScience, 22(5 ):800-803.THORPE T. A., 1988. In vitro somaticembryogenesis. ISI Atlas of Sciencc Animal andPlant Science, 81-88.
22
TISSERAT 8.. MURASHIGE T., 1977a.Repression af asexual embryogenesis in vitro bysome plant growth regulators. In vitro,13(11 ):799-805.TISSERAT S., ESAN E. S .. MURASHIGE T..1979. Sornatic embryogenesis in Angiosperms. InHorticultural reviews. J. Janick, eds, AVIPublishing. Westport, Conn., I: 1-78.VAN SOXTEL J. y SERTHOULY M. 1996.Highfrequency somatic embryogenesis frorn coffeeJeaves:Factors influenelng embryogenesis, andsubsequent and regeneraction in liquidmedium.Plant Cell. Tissue and organ Culture,44:7-17VAN DER VOSSEN H. A. M.. WALYARO D..1.. 1980. Sreed ing for resistance to coffee berrydisease in Coffea arahica 11. Inheritance of theresistance. Euphytica, 29:777-791.VAN DER VOSSEN H. A. M.. WALYARO D..1.. 1981. The coffee breeding programme inKenya a review of progress made since 1971 andplan of action for the coming years. KenyaCofJee.46(541): 113-130.YASUDA T.. FUJI Y.. YAMAGUCHI T.. 1985.Embryogenic callus induction from Coffeaarabica leaf explants by benzyladenine. PlantCell. Physiol., 26(3):595-597.ZAMARRIPA A..1986. Selección preliminar de13 progenies avanzadas de Coffea arabica L.'Catimor" con resistencia a Hemileia vastatrixBerk & Sr. en Rosario Izapa, Chiapas. Tesis deIngeniero Agrônomo Fitotecnista, UniversidadAutónoma de Chiapas, Villaflores, Chiapas.México. 124 p.

SOMATI C EMBRYOGENESIS OF COFFEE
M. Berthouly And H. Etienne
1. INTRODUCTION
1.1. Importance of cofTee worldwideCoffee is one of the most important agriculturalproducts on the world market, and is the mostwidely grown tropical tree crop. Various LatinAmerican countries such as Colombia, Brazil andCentral America depend heavily on coffeeproduction for foreign currency earnings. Moregenerally, the economies of 50 countries depend onthis crop, which is grown on 11.2 x 106 ha andwhich produced 5.3 x 106 tons of green coffee in1992 (Café Cacao Thé, 1992) and 5,603 metric tonsduring 1995 (FAO 1995).The coffee consumedworldwide primarily comes from two cultivatedspecies: Coffea arabica and Coffea canephora,which account for 75 and 25% ofthe world market ,respectively. The coffees produced by the twospecies are commercially known as "arabica" and"robusta".C. arabica is grown in high-altitude tropical regions(500 to 2500 meters). It is the only tetraploid (2n =
44 chromosomes) and autogamous species of theCoffea genus. C. arabica is well suited to highaltitude regions, and produces high quality coffeewith a Jow caffeine content. However, this speciesis susceptible to main diseases such as leaf rust(Hemileia vastatrix Berk & Br.) and coffee berrydisease (Colletotrichum coffeanum (CBD).C. canephora is a diploid (2n = 22 chrornosornes),allogamous species that produces coffee with ahigher caffeine content, which is poor in quality ascompared to the quality ofC.arabica. It is grown inlow-altitude tropical regions (O - 100 m). Withrespect to genetic improvernent, it is a veryworthwhile source of genes for resistance to rust,CBD and nematodes (Bettencourt and Rodrigues.1988). Given its allogarny, the C. canephoraspecies comprises genetically heterogeneousindividuais. The progenies of cross fertilization,therefore, have a substantial genetic variability.In general, C. arabica is more susceptible to diseasethan C. canephora. This is due to the genetic baseand greater variabi Iity of the latter, given itsallogamy.Two major fungal diseases affect coffee plantswhich are leaf rust caused by Hemileia vastatrixand coffee berry disease (CBD), caused byColletotrichum coffeanum. Other pathogens suchas Rhizoctonia, Fusarium can also affect coffeetrees (Haarer 19 The word "pest" can have a very
23
broad meaning, ranging from insects to nematodesvia weeds (Haarer 1989).In our case, we shall quote just two main pests thatcurrently cause significant damage in plantations:the coffee berry borer (CBB, Hypothenemushampei) and nematodes (root parasites).There seems to be no natural resistance to the CBB,and therefore, possible genetic transforrnation is theonly possible solution.In the case of nematodes, although this approachcould be considered, there is a degree of naturalresistance to certain types, notably in species suchas C. canephora. Nematodes are of deep concernworldwide in C. arabica.
1.2. Conventional protocols for improvementof cofTeeThe C. arabica breeding methodis applied toautogamous species. lt is based on producing purelines by pedigree selection after recombiningparental characters brought by the parents.Heterosis has been reported in C. arabica in India(Srinivasan and Vishveshwara 1978), Kenya (Vander Vossen and Walyaro 1981) and Ethiopia(Ameha 1983), and more recently in Costa Rica(Bertrand et ai. 1997). Trials of vegetativelypropagated FI hybrids proved unsuccessful, as theconventional propagation methods resulted in poormultiplication rates (Van der Vossen 1985).There are generally two ways of breeding C.canephora (Bouharmont and Awemo 1979;Charrier 1985): a) by seeds ; which produceheterogeneous populations, and b) vegetatively,which can be summed up as the choice of twoclones with the desirable characters. The selectedtrees are then vegetatively propagated. Polyclonalplantings are necessary to ensure cross-fertilization.Generally, only C. arabica is distributed or sold inseed form after a relatively long pedigree selectionprocess (almost 30 to 35 years) .This processproduces stable and homozygous lines withdesirable characters. Once the variety is obtained,it is sufficient to establish a seed garden to produceseeds genetically identical to the mother plant,since the plant is autogamous.Until now, varietal propagation ofC.canephora wasdone by vegetative means to benefit from the higherproductivity ofclones in relation to varieties grownfrom seeds (Capot, 1975) and from theheterozygosity of the plant.In recent decades, given the complementary

characteristics of C. arabica and C. canephora( quality of the first one and disease resistance ofthe later), breeding has been greatly appreciated increating interspecific hybrids (Icatu). Thesehybrids are multiplied asexually.Vegetative propagation, therefore, plays animportant role in coffee breeding. It enables thepropagation and exploitation of heterozygousgenetic material. It can also reduce the number ofbreeding cycles. In fact, with coffee, a perennialcrop, it takes at least 35 years to develop a variety.Thus vegetative propagation can be applied toselected C. canephora clones, interspecific hybridssuch as arabusta, or e. arabica intraspecific hybridssuch as Ruim I I.Cuttings are the most commonly used vegetativepropagation procedure in coffee. However, giventhe dimorphism ofthe vegetative axes, the numberoforthotopic cuttings that a coffee tree can produceis extremely limited. For large-scale multiplication,the avaibility of limited number of cuttings can betime consuming. Moreover, horticultural cuttingscall for propagation plots, which involve variousconstraints such as upkeep and required land area(Deuss and Descroix, 1984).
1.3. Need for new mass propagation techniquesOver the past severaI years, the progress made inbiotechnology (micropropagation, molecularbiology) has opened up new prospects for researchprogrammes. In this contexto the development ofinvitro multiplication techniques is extremely usefulfor increasing the multiplication rate enabling therapid distribution of the hybrids and clonesproduced.In the past few years, several authors(Staritsky 1970; Sõndahl and Sharp 1977; Dublin1980. 1982; Pierson et al., 1983; Yasuda et aI.1985; de Garcia and Menendez 1987; Berthouly etaI. 1987) have shown that microcutting and somaticembryogenesis can be applied to coffee.Somatic embryogenesis appears to be aninexpensive alternative for multiplying selectedplants. In addition to providing a worthwhilemodel for fundamental studies, its most promisingapplication is in large-scale plant propagation(Ammirato, 1977). Various studies on coffee haverevealed the suitability of the species for somaticembryogenesis and for embryo development intoplantlets. However, the multiplication rate is stillvery low for its large-scale use. In facto de Garciaand Menendez (1987) working on coffee somaticembryogenesis on a solid mediurn, reported 60 to65 somatic embryos per explant after 14 weeks'culturing as a record figure.To exploit the substantial potential of somatic
24
ernbryogenesis, it is necessary to use a liquidmedium and hence, cost of plantlet production canbe reduced. Indeed, by culturing cell suspensions,mass embryo production can be scaled up in abioreactor. The results obtained with other speciesare highly promising. Pétiard et aI. (1987) reporteda production capacity of80 000 carrot embryos perlitre per day in a ten-litre bioreactor.Currently, thereis no in vitro propagation methods available forcommercial coffee production.
2. MICROPROPAGATION BY SOMATICEMBRYOGENESIS
2.1. Introduction
2.1.1. Backgrouod To Coffee SomaticEm bryogeoesisSomatic embryogenesis has been widely used formany years for micropropagation of bothherbaceous and woody plants(Jain and ai 1995). Ithas been tested on different types oftissue (leaves.stems, ernbryos, etc.). In several Coffea species andgenotypes, somatic embryogenesis is welldocumented (Staritsky 1970; Sõndhal and Sharp1977; Pierson et ai. 1983; Dublin 1984; Yasuda etai. 1985; Berthouly and Michaux-Ferriêre J996)
2.1.2. Different Types OfSomaticEm bryogenesisSince the first work by Staritsky( 1970). two typesof somatic embryogenesis have generally beendescribed by S6ndhal( 1970):a)"low frequency": the somatics embryos areobtained on only one medium without producingcalli. Quite often a small number of somaticembryos (a few to 100 per explant).b) "high frequency using two medium" : atinduction medium for primary callogenesis. and asecondary regeneration medium to produceembryogenic friable calli comprised of solelyembryogenic cells producing several hundredthousand of somatic embryos per gramm of callus,but also the use of a liquid medi um for theregeneration phase.
2.1.3. Prospects For Somatic Embryogenesis loCoffeeThere are three main reasons for using somaticembryogenesis in coffee:a) to shorten the C. arabica breeding cycle bytrue-to-typc micropropagation of hybrids (F 1. 1'1.
backcrosses) :
b) raid multiplication of C canephora, genotypeshaving multiplication problems (medium-altitudcrcgions):

c) as vehicle for genetic transformation.
2.2. Induction of embryogenic calli2.2.1. Material And MethodsLeaves were collected from the selected motherplants in the field. Ali samples were disinfected byimmersion in 10% calcium hypochlorite solutioncontaining 1% Tween 80 for 20 min followed by8% calcium hypochlorite solution for 10 min andrinsed 4 times with sterile water. In this case wepresent information on high frequency somaticembryogenesis which is more viable for masspropagation.Explants (1 cm 2) were cut without the main vein,and placed on a initial basal medium containinghalf-strength Murashige and Skoog (1962) salts,88mM sucrose, 100 mg.l"! casein hydrolysate, 400rngl! malt extract, 10mgl- 1 thiamine, 1mgl- 1
nicotinic acid, 1mgl- I pyridoxine. 1 mgl- 1 glycine,100 mgl- 1 myoinositol, 2.2 f.lM 2,4 0,2,4 f.lM lBAand 9.8 f.lM 2iP, solidified with gelrite (2 gl-I ) inglass tubes or small petri dishes. The pH wasadjusted to 5.6 before autoclaving. The cultureswere incubated at 25°C + 1 in darkness.After 4 weeks, explants with a callus weretransferred to a second medium to promote high afrequency rate of embryogenic callus formation.The second basal medium was unlike the first basalmedium in that it contained 200 mgl-l caseinhydrolysate, 800 mg malt extract, 20 mgl-r l
thiarnine, 20 mgl- 1 glycine and 200 mg.l-lmyoinositol. Pyridoxine and nicotinic acid wereomitted. This medium also contained 40 mgl-lcysteine and 60 mgl-! adenine sulphate. Thecultures were incubated at 26°C + 1 on shelveswithout light,After 10-12 weeks of culture on this secondmedium we obtained yellow friable embryogeniccalli. This calli were transferred to either solid orliquid media to proliferate or regenerate somaticembryos.
2.2.2. Optimum Conditions For HighFrequency Embryogenic CalliOifferent concentrations of plant growth regulatorwere used in the two successive culture media andduring subculture on the first medium. The effectoftemperature was also tested.As shown in table I, the percentages of high
25
frequency embryogenic calli were dependant uponthe age of the culture on the first medium andvaried from O to 100 according to the differentauxin-cytokinin ratios in the medi um.The X2 test at 5% levei showed that the percentagesof high frequency embryogenic calli weresignificantly higher when first culture had grownfor 4 weeks (87 %) as compared to a first culturegrown only 3 weeks (78 %). When the friable calliwere obtained from explants on the second mediumwith only BAP(6Benzyl aminopyrine), theycontained very small globular embryos (-1 OOf.lM indiameter). The absence of 2,4-0(2,4dichlorophenoxy acetic acid) in the medium mayexplain the rapid development ofembryogenic cellsofthe calli into globular embryos. Keeping 2,40 ina maintenance medium is very important tomaintain strong regeneration capacity and inhibitglobular embryo formation.
2.2.4. Genotypic EffectA genotypic effect on embryogenic explantproduction has been seen in many plants, and coffeeis no exception (Berthouly and Michaux-Ferriere1996; Van Boxtel and Berthouly 1996). For Coffeacanephora the percentages of highly embryogeniccalli from different genotypes varied from 21 to 98% on first medi um containing 2,2 f.lM 2,4 O, 4,9f.lM IBA, and 9,8 f.lM 2iP, and from 55 to 100 %when the first culture medium was supplementedwith 2,2 f.lM 2,4 O, 2,4 f.lM IBA, and 9,8 f.lM 2iP(6- y-y-dimethylallyamino purine) (table 2).This variability in obtaining embryogenic callidepends on the genotypes used and has also beenobserved in CofJea arabica, in both varieties andhybrids (table 3)Although genotypic effect is observed in both C.arabica and C. canephora, it is possible to obtainembryogenic calli irrespective of the genotype,more importantly with fairly high response leveis.This characteristic makes coffee a plant that as itrespond well to somatic embryogenesis. Either noor poor response of certain genotypes has beenreported for most species, and this is one of themain obstacles to the routine use ofthis techniquesin breeding or mass production schemes.
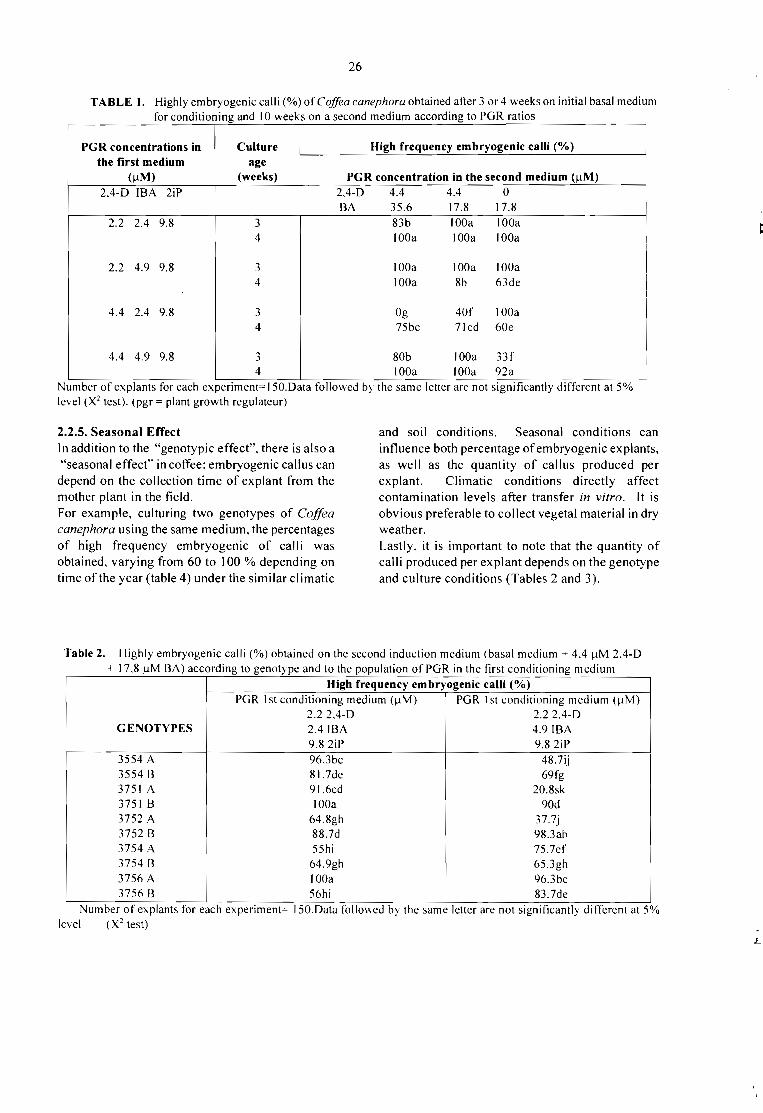
26
TABLE 1. Highly embryogenic calli (%) of Coffea canephora obtained after 3 or 4 weeks on initial basal mediumfor conditioning and 10 weeks on a second medium according to POR ratios
PGR concentrations in Culture High frequency embryogenic calli (%)
the first medium age(11M) (weeks) PGR concentration in the second medium (11M)
2,4-D IBA 2iP 2,4-0 4.4 4.4 OBA 35.6 17.8 17.8
2.2 2.4 9.8 3 83b 100a 100a4 100a 100a 100a
2.2 4.9 9.8 3 100a 100a 100a4 100a 8b 63de
4.4 2.4 9.8 3 Og 40f 100a
I ~494 75bc 71cd 60e
9.8 3 80b 100a 33f4 100a 100a 92a
. ,
Number of explants tor each expenment= 150.0ata followed by the same Jetter are not significantly differcnt at 5%levei (X 2 test). (pgr = plant growth regulateur)
2.2.5. Seasonal EffectIn addition to the "genotypic effect", there is also a"seasonal effect" in coffee: embryogenic callus can
depend on the collection time of explant from themother plant in the field.
For exarnple, culturing two genotypes of Coffeacanephora using the same medium, the percentagesof high frequency embryogenic of calli wasobtained, varying from 60 to 100 % depending ontime ofthe year (table 4) under the similar climatic
and soil conditions. Seasonal conditions canintluence both percentage ofembryogenic explants,as well as the quantity of callus produced perexplant. Climatic conditions directly affectcontamination leveis after transfer in vitro. It is
obvious preferable to collect vegetal material in dryweather.Lastly, it is important to note that the quantity ofcalli produced per explant depends on the genotypeand culture conditions (Tables 2 and 3).
Table 2. Highly embryogenic calli (%) obtained on the second induction medium (basal medium + 4,4 11M 2,4-0+ 17.8 11M BA) according to genotype and to the population of POR in the first conditioning medium
High frequency embryogenic calli (%)
POR 1st conditioning medium (11M) POR 1st conditioning medium (11M) I2.22,4-0 2.2 2,4-0 I
I
GENOTYPES 2.4 IBA 4.9 IBA J9.82iP 9.82iP3554 A 96.3bc 48.7ij3554 B 81.7de 69fg3751 A 91.6cd 20.8sk3751 B 100a 90d3752 A 64.8gh 37.7j3752 B 88.7d 98.3ab3754 A 55hi 75.7ef3754 B 64.9gh 65.3gh
I3756 A 100a
J96.3bc
I3756 B 56hi 83.7de
- . - ... -Number 01 explants tor each expenmenr- 1::lO. Data followed by the sarrie letter are not significantly differcnt at ::l%levei (X2 test)
L
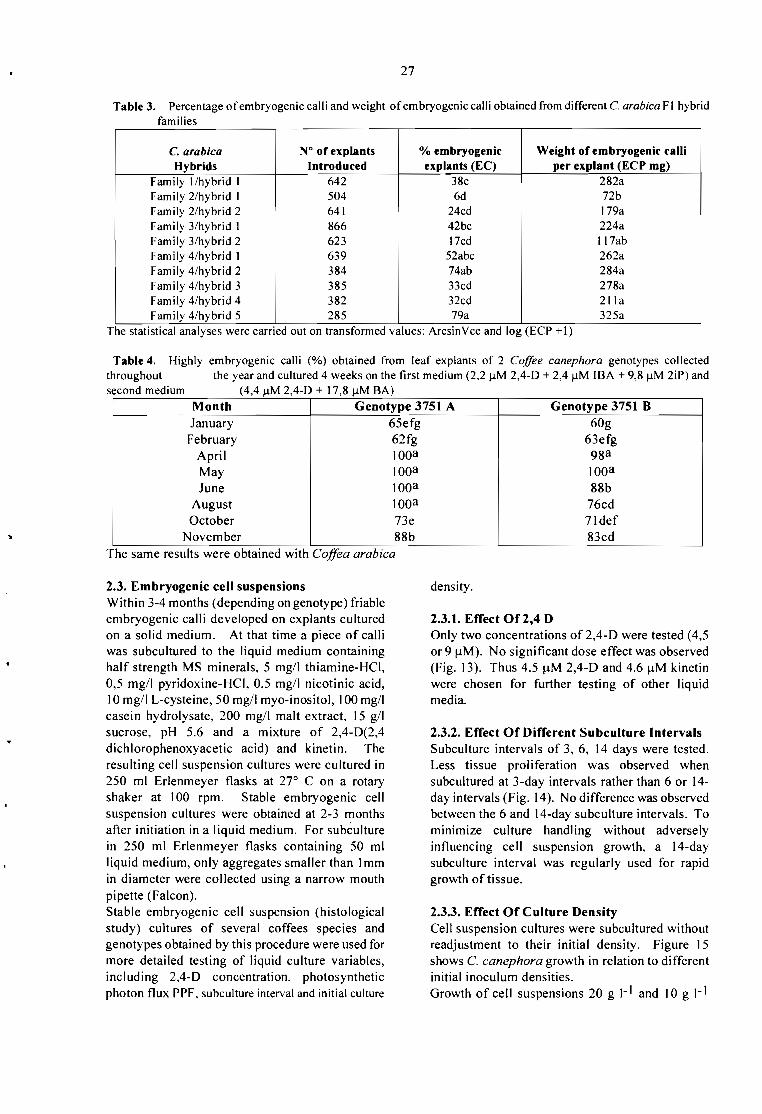
27
Table 3. Percentage ofembryogenic calli and weight ofembryogenic calli obtained frorndifTerent C. arabica F1hybridfamilies
C. arabica N° or explants % embryogenic Weight or embryogenic calliHybrids Introduced explants (EC) per explant (ECP mg)
Family I/hybrid 1 642 38c 282aFamily 2/hybrid I 504 6d 72bFamily 2/hybrid 2 641 24cd 179aFamily 3/hybrid I 866 42bc 224aFamily 3/hybrid 2 623 17cd 117abFamily 4/hybrid 1 639 52abc 262aFamily 4/hybrid 2 384 74ab 284aFamily 4/hybrid 3 385 33cd 278aFamily 4/hybrid 4 382 32cd 211aFamily 4/hybrid 5 285 79a 325a
The statistical analyses were carried out on transformed values: ArcsinVce and log (ECP +I)
Table 4. Highly embryogenic calli (%) obtained frorn leaf explants of 2 Cof/ee canephora genotypes collectedthroughout the year and cultured 4 weeks on the first medium (2,2 JlM 2,4-D + 2,4 IlM IBA + 9,8 IlM 2iP) andsecond medium (4,4 JlM 2,4-D + 17,8 JlM BA)
Month Genotype 3751 A Genotype 3751 BJanuary 65efg 60g
February 62fg 63efgApril looa 98 a
May looa looa
June looa 88bAugust looa 76cdOctober 73e 71def
November 88b 83cdThe same results were obtained with Coffea arabica
2.3. Embryogenic cell suspensionsWithin 3-4 months (depending on genotype) friableembryogenic calli developed on explants culturedon a solid medium. At that time a piece of calliwas subcultured to the liquid medium containinghalf strength MS minerais, 5 mg/l thiamine-HCI,0,5 mg/l pyridoxine-HCI, 0,5 rng/l nicotinic acid,10 mgl/ L-cysteine, 50 mg/l myo-inositol, 100 mg/lcasein hydrolysate, 200 mg/l malt extract, 15 g/lsucrose, pH 5.6 and a mixture of 2,4-0(2,4dichlorophenoxyacetic acid) and kinetin. Theresulting cell suspension cultures were cultured in250 ml Erlenmeyer flasks at 27° C on a rotaryshaker at 100 rpm. Stable embryogenic cellsuspension cultures were obtained at 2-3 monthsafter initiation in a liquid medi um. For subculturein 250 ml Erlenmeyer flasks containing 50 mlliquid médium, only aggregates smaller than 1mmin diameter were collected using a narrow mouth
pipette (Falcon).Stable embryogenic cell suspension (histologicalstudy) cultures of several coffees species andgenotypes obtained by this procedure were used formore detailed testing of liquid culture variables,including 2,4-0 concentration, photosyntheticphoton flux PPF, subculture interval and initial culture
density.
2.3.1. Effect Of 2,4 DOnly two concentrations of 2,4-0 were tested (4,5or 9 !lM). No significant dose effect was observed(Fig. 13). Thus 4.5 !lM 2,4-0 and 4.6 !lM kinetinwere chosen for further testing of other liquidmedia.
2.3.2. Effect Of Different Subculture IntervalsSubculture intervals of 3, 6, 14 days were tested.Less tissue proliferation was observed whensubcultured at 3-day intervals rather than 6 or 14day intervals (Fig. 14). No difference was observedbetween the 6 and 14-day subculture intervals. Tominimize culture handling without adverselyinfluencing cell suspension growth, a 14-daysubculture interval was regularly used for rapidgrowth oftissue.
2.3.3. Effect OfCulture DensityCell suspension cultures were subcultured withoutreadjustment to their initial density. Figure 15shows C. canephora growth in relation to differentinitial inoculum densities.Growth of cell suspensions 20 g I-I and 10 g 1-1

culture density slowed down between 6 and 8weeks. In 0.5 g l-I cuiture density and to a lesserextent in 2 g l-I cell culture density, somaticembryos were observed, no somatic embryosformed in embryogenic cell cultures with celldensity 5, 10 and 20 g l-I. Thus a minimum culturedensity seemed to be necessary to avoid somaticembryo formation. Calculation of themultiplication rate (MR) demonstrated theexponential growth during the first week ofculturefor each density tested. During the second week,the MR of ali cultures decreased to a leve I between1.0 and 1.8( en gr/l en fteh mater), when during thefirst week it was between 3 and 15 in relation withthe density(en gr/l en freh mater).Thus, by culturing small amounts of embryogeniccallus at a density of 5 g I-I, a relatively 10w MRcan be maintained over several weeks. Under theseconditions, subculturing in order to avoid a growthplateau is less often necessary.When cultures had been subcultured weekly withreadjustment to their initial density of 10, 15, or 20g I_I, the MR over 4 week culture period was 2.0,1.8, and 1.5, respectively. The MR of 10 g I_Icultures was significantly higher than the MR of20g I_I cultures. Thus, weekly readjustment ofculture density to 10 g calli I_I can be to usedaccelerate growth of cell suspension cultures.
2.3.4. Nutrient Evolution In The MediumAbsorption of some macro- and micro-salts byembryogenic cell suspensions was monitored byregularly analyzing the used amount of nutrients inthe culture medium. After 12 days of culture, 60 70% of the initial concentration of nutrients wassti II present (Fig. 16).After 3 weeks, macro-salts and nitrogen sourceshad decreased by 50-70%, and in which the amountof available magnesium was the lowest (30% ofinitial concentration). These results suggest thatamount of MS macro- and micro-rninerals at halfstrength in the culture medium do not deplate incoffee embryogenic cell suspension culture for atleast 3 weeks of culture.In trying to optimize long-terrn maintenance ofembryogenic cultures, instead of subculturing in acompletely fresh medium every 2 weeks, a part ofthe embryogenic callus was discarded andconcentrated nutrients were added to theconditioned culture. In ali cases, growth wasnevertheless inhibited after 2 months of culture.Therefore, medium renewal remains necessary atregular time intervals in order to sustain the growthof embryogenic suspensions, presumably byeliminating growth-inhibiting substances present in
28
the conditioned medi um.
2.4. Regeneration in a liquid medium
2.4.1. Systems UsedSomatic embryogenesis, using liquid media,whether for cell suspensions in Ermlenmeyer flasksor in bioreactors, is Iimited to the quantityembryogenic to sornatic embryos produced, andthese embryos have to be transferred to a gelmedium several times to ensure their developmentinto plantlets. However, these operations areextremely costly, as they are labour-intensive interms of both the individual selection/handling ofembryos and gel culture medium production.Using a liquid medium for embryo productionprovide substantial guarantees to improveproductivity. As it enables the automation ofliquidculture methods and reduce the cost as compared tosolid medium.Using a liquid medium for embryo production/orplant procedure and combining the multiplication ofembryogenic aggregates in Erlenmeyer flasks makesuch a technology applicable on an industrial scale.A temporary immersion system has been developedat CIRAD. on different plants, and is now beingapplied on a large scale for coffee. The principiebehind it and the conditions for its use are describedin section Y.However, as seen previously, once embryogeniccalli are obtained and suspension cell cultures areestab lished, regeneration is possible in the liquidmedium according to conventional procedures. Theefficacy of regeneration largely depends on certainparameters such as cytokinin leveis and the initialquantity offresh matter.
2.4.2. Effect Of Ba And Inoculum DensityFigure 17 shows the combined effect ofbenzyladenine (BA) and inoculum density onglobular somatic embryo formation, and theincrease in cal Ii fresh weight in the regenerationmedi um. Globular somatic embryo formation wasinversely related to inoculum density and theearliest abundant somatic embryo formation wasmostly observed with an inoculum density of 1g j.Iand f4.4 !lM or 17.6 !lM BA.Zamarripa (1993) reported the presence of certainsubstances that were inhibiting to somaticembryogenesis in coffee calli cultured at highdensity. Adding of BA in the culture medi umappeared to neutralize the influence of inhibitorysubstances and allowed for somatic embryoformation.

3. DEVELOPMENT OF A NEW CULTURESYSTEM( Temporary immersion system) TOIMPROVE SOMATIC EMBRYOGENESIS
A new temporary immersion systern, known asRITA (Automatic Temporary lmmersion System),has been developed at CIRAO over the past eightyears (Alvard et aI. 1993; Teisson and Alvard 1994;Berthouly et aI. 1995; Etienne et aI. 1997 b) toavoid transfer to a solid medi um during the finalstages of somatic embryogenesis, i.e. embryodeveloprnent, germination and conversion intoacclimatizable plants. The device has alreadyproved its efficacy, i.e. its superiority to semi-solidmedia for producing microcuttings of coffee,banana and Hevea, and for somatic embryogenesison coffee (Berthouly et aI. 1995; Etienne et aI. 1997a), banana (Alvard et aI. 1993), Hevea (Etienne etaI. 1997 b) and citrus species (Teisson and Alvard1994), and lastly for producing potato microtubers.
3.1. OperationIn the beginning, the device used for the initialexperiments was a readily available, autoclavablefilter unit, modified by linking the upper and lowercompartments by a glass tube. The Biotrop in vitroculture laboratory (ClRAO) has now developed asmall, economical device that is easy and above all.flexible to use. The unit is operated in the oppositeway to filtration units in general: the plant materialis placed in the upper part and the liquid medium atthe bottom (Fig. 18).When the lower compartment is pressurized, usinga small laboratory air purnp, the medium is pushedup into the upper compartment. A simple time rswitch is used to control the frequency and durationofpump operation, hence ofimmersion. When thepressure is released, the liquid naturally flows backdown again. The air blown around the containerthroughout the immersion period renews theinternal atmosphere. AlI the air flows are sterilizedby O.21-l hydrophobic vents.Each system is thereforeindependent and can be moved individually.
3.2. Advantages of the systemThis container has the same advantages asbioreactors, without the following two types oflimitations:• Physiologicallimitations: permanent contact withthe culture medium results invitrification andconstant movement of the plant material, thatprevent the establ ishment of polarity and hinderssomatic embryo developrnent, oxygenationproblems.• Technical and financiai limitations: exorbitant
29
bioreactor cost, their lack offlexibility, complicatedtechnology and fragility in the event of power cutswhere the plant material dies within a few hoursdue to lack of oxygen.Currently ali these drawbacks, making it impossibleto use bioreactors for mass production, particularlyin developing countries, are resolved withtemporary immersion systems. Temporaryimmersion enables the development of plantletsfrom embryogenic cell suspension cultures.Moreover, this development "frorn cell to plant"takes place in the same container, without handlingthe plant material; it is sufficient to only change themedium. The explants are in contact with the liquidculture medium for just a few minutes per day, andthe duration of contact can be adjusted to preventany risk ofvitrification. The container is ventilatednaturally and passively by vents, with pulsed airduring immersion to renew ali the air in thecontainer.On a technical leveI, the RITAR is small,autoclavable, easily accessible (US$ 55), portable(it can be transported easi Iy without risk ofcontarnination), and easy to operate. In the event ofpower cuts plant material is not affected .The liquidmedium in the lower section is not in contact withthe cultures but it maintains a high moisture leveI.The plant material can withstand power cuts lastingover 10 days. Another advantage for massproduction is that laboratories can easily havehundreds of RITA units, hence greater flexibilitythan with one or two bioreactors. With RITA units,it will be easy to produce several cultivars at onceand adapt production volumes to demandthroughout the year.
3.2.1 ContextIn somatic embryogenesis procedures aimed atmass production, the use of Iiquid media to producecell suspension cultures in Erlenmeyer flasks or inbioreactors is limited to the multiplication ofembryogenic clumps. These embryos have to betransferred several ( at least three ) times to a ge Imedium in these systems to ensure theirdevelopment into wholly acclimatizable plants.However, these operations are extremely costly dueto the substantial quantities of labour, containers,gel media and laboratory space required.Using liquid media for as much of the process aspossible is a significant guarantee of its efficacy. ltshould contribute substantially to reducing the costper in vitro plantlet, an essential factor forcommercial production. The originality of thesomatic embryogenesis protocol presented here liesin the use ofliquid culture media for the major part
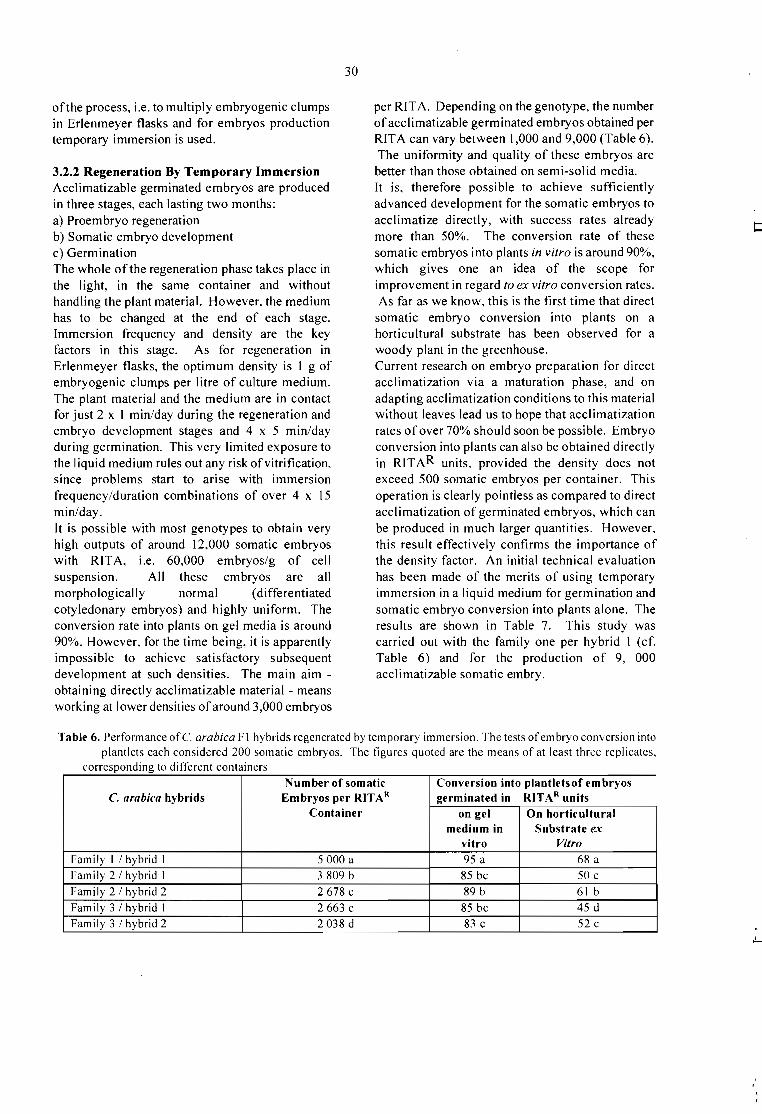
ofthe process, i.e. to multiply embryogenic c1umpsin Erlenmeyer flasks and for embryos productiontemporary immersion is used.
3.2.2 Regeneration By Temporary ImmersionAcclimatizable germinated embryos are producedin three stages, each lasting two months:a) Proembryo regenerationb) Somatic embryo developmentc) GerminationThe whole ofthe regeneration phase takes place inthe iight, in the same container and withouthandling the plant material. However, the mediumhas to be changed at the end of each stage.Immersion frequency and density are the keyfactors in this stage. As for regeneration inErlenmeyer flasks, the optimum density is 1 g ofembryogenic c1umps per litre of culture medium.The plant material and the medium are in contactfor just 2 x I min/day during the regeneration andembryo development stages and 4 x 5 min/day
during germination. This very limited exposure tothe liquid medium mies out any risk of vitrification,since problems start to arise with immersionfrequency/duration combinations of over 4 x 15min/day.
It is possible with most genotypes to obtain veryhigh outputs of around 12,000 somatic embryoswith RITA, i.e. 60,000 embryos/g of cellsuspension. Ali these embryos are alimorphologically normal (differentiatedcotyledonary embryos) and highly uniform. Theconversion rate into plants on gel media is around90%. However, for the time being, it is apparentlyimpossible to achieve satisfactory subsequentdevelopment at such densities. The main aim obtaining directly acclimatizable material - meansworking at lower densities ofaround 3,000 embryos
30
per RIT A. Depending on the genotype, the numberofacclimatizable germinated embryos obtained perRITA can vary between 1,000 and 9,000 (Table 6).The uniformity and quality of these embryos arebetter than those obtained on serni-solid media.It is, therefore possible to achieve sufficientlyadvanced development for the somatic embryos toacclimatize directly, with success rates alreadymore than 50%. The conversion rate of thesesomatic embryos into plants in vitro is around 90%,which gives one an idea of the scope forimprovement in regard to ex vitro conversion rates.As far as we know, this is the first time that direct
somatic embryo conversion into plants on ahorticultural substrate has been observed for awoody plant in the greenhouse.Current research on embryo preparation for directacclimatization via a maturation phase, and onadapting acclimatization conditions to this materialwithout leaves lead us to hope that acclimatizationrates ofover 70% should soon be possible. Embryoconversion into plants can also be obtained directlyin RIT AR units, provided the density does notexceed 500 somatic embryos per container. Thisoperation is clearly pointless as compared to directacclimatization of germinated ernbryos, which canbe produced in much larger quantities. However,this result effectively confirms the importance ofthe density factor. An initial technical evaluationhas been made of the merits of using temporaryimmersion in a liquid medium for germination andsomatic embryo conversion into plants alone. Theresults are shown in Table 7. This study wascarried out with the family one per hybrid 1 (cf.Table 6) and for the production of 9, 000acclimatizable somatic embry.
Table 6. Performance ofC arabica F I hybrids regenerated by temporary immersion. The tests of embryo conversion intoplantlets each considered 200 somatic embryos. The figures quoted are the means of at least three repl icates,
corresponding to different containersNumber of somatic Conversion into plantletsof embryos
C. ara bica hybrids Embryos per RITA R germinated in RITA R unitsContainer on gel On horticulturaI
medium in Substrate exvitro Vitro
Family I / hybrid I 5000 a 95 a 68 aFamily 2 / hybrid I 3809 b 85 bc 50 cFamiiy 2/ hybrid 2 2678 c 89 b 61 bFamily 3 / hybrid 1 2663 c 85 bc 45 dFamily 3 / hybrid 2 2038 d 83 c 52 c
L
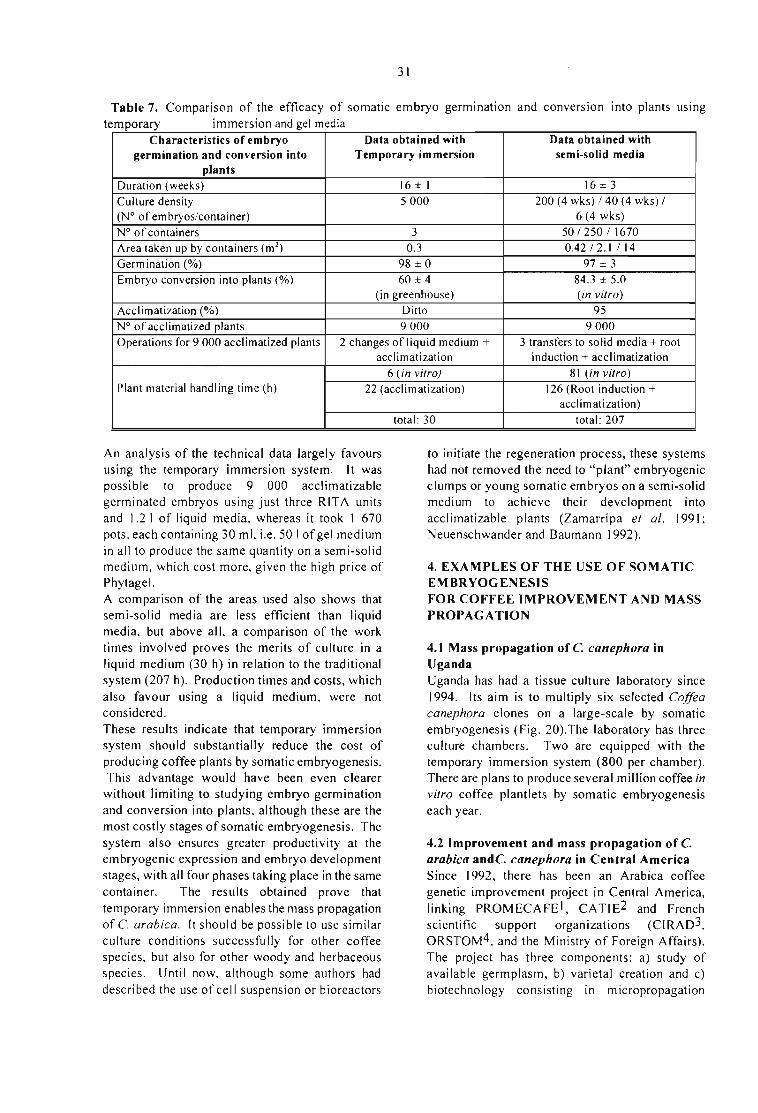
31
Table 7. Comparison of the efticacy of sornatic embryo germination and conversion into plants usingtemporary immersion and gel media
Characteristics of embryo Data obtained with Data obtained withgermination and conversion into Temporary immersion semi-solid media
plantsDuration (weeks) 16± I 16 ± 3Culture density 5000 200 (4 wks) 140 (4 wks)1
(N° of embryos/container) 6 (4 wks)N° of containers 3 50/250/1670Area taken up by containers (rrr') 0.3 0.42/2.1/14
Germination (%) 98 ± O 97 ± 3Embryo conversion into plants (%) 60 ± 4 84.3 ± 5.0
(in greenhouse) (in vitro)Acclimatization (%) Ditto 95N° of acclimatized plants 9000 9000Operations for 9 000 acclimatized plants 2 changesof liquid medium + 3 transfersto solid media+ root
acclimatization induction + acclimatization6 (in vitro) 81 (in vitro)
Plant material handling time(h) 22 (acclimatization) 126(Root induction +acclimatization)
total: 30 total: 207
An analysis of the technical data largely favoursusing the temporary immersion system. It waspossibJe to produce 9 000 acclimatizablegerminated embryos using just three RITA unitsand 1.2 I of liquid media, whereas it took 1 670pots, each containing 30 ml, i.e. 50 1ofgel mediumin ali to produce the same quantity on a semi-solidmedium, which cost more, given the high price ofPhytagel.A comparison of the areas used also shows thatsemi-solid media are less efticient than liquidmedia. but above ali. a comparison of the worktimes involved proves the merits of culture in aliquid medium (30 h) in relation to the traditionalsystem (207 h). Production times and costs, whichalso favour using a liquid medium. were notconsidered.These results indicate that temporary immersionsystem should substantially reduce the cost ofproducing coffee plants by somatic embryogenesis.This advantage would have been even clearer
without limiting to studying embryo germinationand conversion into plants, although these are themost costly stages of somatic embryogenesis. Thesystem also ensures greater productivity at theembryogenic expression and embryo developmentstages, with ali four phases taking place in the samecontainer. The results obtained prove thattemporary immersion enables the mass propagationof C arabica. It should be possible to use similarculture conditions successfully for other coffeespecies, but also for other woody and herbaceousspecies. Until now, although some authors haddescribed the use ofcell suspension or bioreactors
to initiate the regeneration process, these systemshad not removed the need to "plant" embryogenicclumps or young somatic embryos on a semi-solidmedium to achieve their development intoacclimatizable plants (Zamarripa et ai. 1991;Neuenschwander and Baumann 1992).
4. EXAMPLES OF THE USE OF SOMATICEMBRYOGENESISFOR COFFEE IMPROVEMENT AND MASSPROPAGATION
4.1 Mass propagation of e. canephora inUgandaUganda has had a tissue culture laboratory since1994. Its aim is to multiply six selected Coffeacanephora clones on a large-scale by somaticembryogenesis (Fig. 20).The laboratory has threeculture chambers. Two are equipped with thetemporary immersion system (800 per chamber).There are plans to produce several million coffee invitro coffee plantlets by somatic embryogenesiseach year.
4.2 Improvement and mass propagation of e.arabica ande. canephora in Central AmericaSince 1992, there has been an Arabica coffeegenetic improvement project in Central Arnerica,linking PROMECAFEI, CATI E2 and Frenchscientific support organizations (CIRAD3.ORSTOM4, and the Ministry of Foreign Affairs).The project has three components: a) study ofavailable germplasm, b) varietal creation and c)biotechnology consisting in micropropagation

which is of crucial importance, since theimprovement strategies chosen mean using in vitroplantlets.In effect, varietal creation is based on creating andbreeding FI hybrids, for which the only possiblelarge-scale rnultiplication rnethod IS
micropropagation.The aims of micropropagation under this project aretwofold:a) To introduce in vitro, multiply and distribute toproject member countries improved materialproduced by varietal creation;b)To develop a mass propagation method bysomatic embryogenesis and transfer it tolaboratories in improvement project member incountries.
4.2.1. Example Of MultiplicationOf C. Canephora Parents Of The NernayaStock VarietyOn various occasions, a hybrid between two C.canephora has proved highly resistant, and in somecases immune, to different nematodes, for instanceMeloidogyne and Pratylenchus (Anzueto 1989,Bertrand 1995). The hybrid is T3751 (1-2) x T3561(2-1). known as the "Nernaya variety" (F I hybrid)amongst the researchers that described it.Because the demand for this stock is huge, the bestway of producing it on a large scale is to set up seedgardens where the two parents are grown side byside. Each tree will thus produce "Nernaya variety"seeds. As C. canephora is allogamous. vegetativepropagation is required. Horticultural propagationis not possible as large quantities of each parentwould have to be produced and there is only oneindividual in the CA TI E collection. The number oforthotopic axes available for cuttings or buddingsis, therefore, toa small.The decision was therefore, taken to multiply thetwo parents by tissue culture, particularly somaticernbryogenesis, as the region's requirements aresubstantial (for instance, 7 ha ofseed gardens for EISalvador or Guatemala). The ease with which thematerial can be introduced in vitro - a simple squareof leaf that can easily be disinfected - was also adetermining factor in favour of somaticembryogenesis. Thirty thousand plants ofthe twoparents were produced at the CA TIE laboratory in1996-1997 and are now being distributed andacclimatized in lhe countries affected bynematodes. with objective of setting up seedgardens.
32
4.2.2. Example Of Multiplication Of C. ArabicaFI Hybrids or Agronomic WorthThe Arabica coffee genetic improvement programis original in that it is based on hybridizing localvarieties (Caturra, Catuai, Catimor, Sarchimor) withwild origins (Ethiopia, Sudan). The programobjectives are to increase the adaptability andproductivity of the varieties and improve theirdisease and pest resistance, while conserving goodcup quality.ln.sius: micropropagation techniques are the bestway ofrapid multiplication ofselected FI hybrids.Given its high multiplication potential and theposs ibi Iity of eventually producing low-cost plants.it was decided to use somatic embryogenesis forlarge-scale multiplication of the best cultivars.Moreover, using this technique reduces thebreeding cycle from 30 to 10 years, and makes itpossible to offer growers different candidates forthe wide variety of situations in Central America.Then cultivares have already been introduced andmultiplied on a large scale. Clone performancetrials have also been set up. Distribution ofthesehybrids to the countries participating in the projectbegan in 1997. and it is to be stepped up in 1998.Around 40 cultivares will be chosen, multiplied andtested in clone trials under different soil andc1imatic conditions in the six project membercountries. This research could lead to commercialproduction ofthe best hybrids by 2003.
4.3.3. Pilot Project In TanzaniaCoffee research in Tanzania has achieved thecreation ofthousands of hybrids plants establishedin LyamunguReasearch Station . Their average leveI of yield,without any fungicide spraying, in Lyamungu,ismore than doublethe commercial varietiesA rapid multiplication ofthe best individual trees is urgently to make thesehybrids available to the coffee growers.lt cannot bedone through normal cuttins as the te ofmultiplication is toa low.The pilot propagation project that CIRAD isdeveloping in Tanzania is:* to start multiplying some promissing trees (10) toestablish real clones in a wide multisite comparisontrial* to train scientist technicians in tissue culture incoffee propagation* to star a small research laboratory in Lyamungu tomultiply in the future other selected trees* to the prepare the future for commercial masspropagation: contacts have to be take with privatesector

5. F1ELO PERFORMANCE ANOCONFORMITY
5.1. Oefinition of variationWe have known since the work by Larkin andScowcroft in 1981 that tissue culture can causemodifications ofthe genome known as somaclonalvariations. This notion covers three types ofvariation: (I) Heritable, stable, irreversible changes,(2) Unstable, reversible changes and(3) non-heritable, temporary changes known asepigenetic variations, which affect genomeexpression (Karp, 1991). Most ofthis variability iseither due to chromosomal alterations, i.e. cuts,translocations, deletions, aneuploidy, polyploidy,gene amplification, transpositions, somatic crossovers and one-off mutations (Evans and Sharp1986), or, in the case of epigenetic mechanisms, tounder-or overmethylation ofthe DNA. In the lattercase, the variant phenotype is unstable. Forinstance, in oil paIm, 50% ofsterile palms reverttonormality after seven years' flowering (Duval et aI.1997).Somaclonal variations are put down to excessiveuse of phytohormones in the culture medium,particularly auxin, and to over-rapid cellproliferation. In general, somaclonal variationsinvolve a disorganized cell growth phase such ascallus or adventive meristem multiplication (Karp1991). They affect most plant species, in additionto algae and fungi. Lastly, they are cultivardependent (Mohmed and Nabors 1990; Freytag etai. 1989). This has also been demonstrated incoffee (Sõndhal and Lauritis 1992).
5.2. Tbe case of coffeeIn coffee, little data is available on the fieldperformance and conformity ofplants produced bysomatic embryogenesis. The only known work wasby Sõndhal in Brazil (Sõndhal and Bragin 1991;Sõndhal and Lauritis 1992), but Nestlé is alsoknown to have set up large-scale trials in Ecuador.Sõndhal recorded variability of 10% in 12, 176 C.arabica plants evaluated. Among the variants, heobserved modifications affecting leaf and branchmorphology, plant size, fruit colour and shape,increased yields, variable ripening precocity andsusceptibi lity toltolerance of certain diseases.Nestlé, on the other hand, has not attempted true-totype reproduction, but uses somaclonal variation asa source of genetic variability. It is widely acceptedthat in species for which breeding systems are alimiting factor and in "primitive" species that havenot undergone intensive selection - for instancecoffee - somaclonal variation can release a new
33
source of variability (Karp 1991). The percentageof variant plants remains very low - around 1%(personal communication) - and suggests relativelysubstantial genetic variability in coffee. Forinstance, the Arabusta, an interspecific hybrid withan unstable genome, apparently has moresomaclonal variations. These results contradictthose of Sõndhal, since they suggest much lowerrates of variants.
5.3. Work under wayTo remedy this lack of information,PROMECAFE(Programa regional para laproteccion y modernizacion de la cafeicultura enAmerica Centrale) decided to set up an extensivemulti-site network with at least 20 sites in fourCentral American countries and a structurecovering 50 000 plants. Moreover, specific studiesare being carried out on the effect ofcertain cultureconditions, such as suspension age (from O to 2years) or genotype on the conformity ofthe plantsregenerated.t present, 1,100 in vitro plantlets including 650somaplants from embryogenic calli offour hybridsare being evaluated in field trials, with 450microcuttings of the same hybrids as controls, asmicrocuttings are known not to affect conformity.No difference in plant morphology has yet beenobserved, and most are about to flower.One thousand two hundred other somaplants fromhigh frequency embryogenic calli of five otherhybrids are currently in the nursery, and novariations in morphology have yet been observedeither. By the end of 1998, 3,200 somaplantsobtained from calli or embryogenic suspensions ofvarious ages will have been planted in Costa Rica.By the end of 1999, 60,000 C. arabica plantsobtained from somatic embryos wiJl have beenplanted in Central America. These plants were aliproduced by temporary immersion and underclearly defined culture conditions whollycompatible with commercial production.These trials should provide definitive informationabout the conformity of the material produced bysomatic embryogenesis under our cultureconditions. It is important to bear in mind that 5%non-true-to-type plants is a commercially acceptedfigure. However, ifserious fruiting problems wereto arise, it would not be disastrous, as variousstrategies can be adopted to bring the figure below5%, or even to eliminate ali somaclonal variations.For instance, this was done with bananamicropropagation (Côte et al. 1993).

6. CONCLUSION AND PROSPECTS
The results presented here show that somaticembryogenesis procedures are applicable to coffeeon a large scale, irrespective of the species andgenotype (Fig. 21).In particular, the technique can be used to modifythe conventional Arabica breeding strategy. Untilrecent years, development of new varieties of thisspecies was geared towards pedigree breeding toproduce a homozygous variety that could bedistributed as seed. The process is relatively longand takes as long as 30-35 years.The development of a vegetative propagationtechnique such as somatic embryogenesis shouldallow:I.Orientation of genetic improvement programstowards a new strategy: the creation and use of F Ihybrids that can be obtained after a lOto 15-yearselection cycle ; this will enable breeders to shortenthe selection cycle and also to make optimum useof genetic variability, above ali by favouringheterosis. Such a program was launched sevenyears ago in Central America, involving CIRADand ORSTOM;2. Consideration ofmass production, including useof the temporary immersion technique developedby CIRAO. Using such a technique to regenerateplants removes the need for isolation and manualtransfer of somatic embryos required by theconventional in vitro procedure on a gel medium.3. Consideration of new strategies such as genetictransformation. In effect, it is essential therehability of plant regeneration from "cells" if sucha strategy is to be implemented. This is nowpossible as the technique has been fully developed.lt is clear that on the brink ofthe third millennium,thought can now be given to new coffee growingstrategies based on more rational use of existingbiodiversity and on providing a more rapidresponse to the various constraints inherent to thecrop.
7. ACKNOWLEDGEMENTS
On special thanks go to Mrs N. Michaux-Ferriêre(CIRAO) for her help, not only in histologicalstudies of the somatic embryogenesis procedure,but also in writing up this section. We could alsolike to thank Daniel Alvard and Marc Lartaud(CIRAO) for their help and coJlaboration on thiswork.
34
8. REFERENCES
Alvard, D., Côte, F. and Teisson, C (1993)Comparison of methods of liquid medium culturefor banana propagation. Effects of temporaryimmersion of explants. Plant Cell Tissu Org. Cult.32: 55-60.Ameha, M. (1983) Heterosis in crosses ofindigenous coffee selected for yield and resistanceto coffee berry disease. I. First bearing stage. ActaHorticulture, 140: 155-161.Amrnirato, P.V. (1977) Hormonal control ofsomatic embryo development from cultured cells ofcaraway: Interactions of abscisic acid, zeatin andgibberellic acid. Plant Physiol. 59: 579-586.Anzueto F. (1989) Recherche de la résistance auxnématodes dans une collection de Coffeaspp.O.E.A.Univ. de Rennes-ENSAR, 29p.Berthouly, M., Guzman, N. and Chatelet, P. (1987)Micropropagation in vitro de différentes lignées deCoifea arabica v. Catimor. In l Zême ColloqueScientifique International sur le Café (Montreux,29 juin - 3 juillet), ASIC, Paris, p. 462-467.Berthouly, M. and Michaux-Ferriêre, N. (1996)High frequency somatic embryogenesis in Coifeacanephora: induction conditions and histologicalevolution. Plant Cell and Organ Culture, 44: 169
176.Berthouly, M., Oufour, M.. Alvard, O.•Carasco, C,Alemanno, L. and Teisson, C (1995) Coffeemicropropagation in liquid medium usingtemporary immersion technique. ASIC, l ôêrneColloque, Kyoto 9-14 avril, Vol. 11: 514-519.Bertrand, 8., Anzueto, F., Pena, M.X., Anthony, F.,Eskes, AB.( 1995) Genetic imrpovment ofcoffe forresistance te root-knot nematodes (MeJoidogynesp.) in Central America.IN XI CoJ1oqueScientifique International sur le café.A'SfC.Kyoto, Japon, p:630-636Bertrand, B., Santacréo, R., Anthony. F., EtienneH., Charrier A. and Eskes, A.B. (1997) Behaviourof Coifea arabica FI hybrids for resistance.production and technological qualities in CentralAmerica. ASIC, Nairobi 20-25 july.Bettencourt, AJ. and Rodrigues, CJ.J.R. (1988)Principes and practice of coffee breeding forresistance to rust and other diseases. Chapter 6 inCoifee: Agronomy, R.J. Clarke, R. Macrae, eds ..Elsevier Applied Science Publishers LTO, England,4: 199-234.Bouharrnont, P. and Awemo, J. (1979) La sélectionvégétative du caféier robusta au Cameroun, CaféCacao Thé, 23: 227-237.Capot, J (1977) L'amélioration du caféier robusta enCôte d'lvoire. Café Cacao Thé, 21: 233-244.
t

Charrier, A. (1985) Progrês et perspectives del'amélioration génétique des caféiers. In: JlemeCol/oque Scientifique International sur le café,(Lorné, 11-15 février), ASIC, Paris: 403-425.De Garcia, E. and Menendez, A (1987)Embriogénesis somatica a partir de explantesfoliares dei cafeto "catimor". Café Cacao Thé, 31(1): 15-22.Deus, J. and Descroix, F. (1984) Le bouturage ducaféier robusta dans le programme de replantationde la caféiêre au Togo. Café Cacao Thé, 28 (3):165-178.Dublin, P. (1980) Multiplication végétative in vitrode I'arabusta. Café Cacao Thé, 24 (4): 281-289.Dublin, P. (1982) Culture de tissus et améliorationgénétique des caféiers cultivés. In: l Oême Col/oqueScientifique sur le café (Salvador-Bahia, 11-14octobre), ASIC, Paris, 433-459.Dublin, P. (1984) Techniques de reproductionvégétative in vitro et amélioration génétique chezles caféiers cultivés. Café Cacao Thé, 28 (4): 231244.Duval, Y., Arnblard, P., Rival, A, Konan, E.,Gogor, S. and Durand-Gasselin, T. (1997). Progressin oil paim tissue culture and clonal performance inIndonesia and the Côte d'Ivoire. InternationalPlanters Conference, Kuala Lumpur, 21-22 May1997: 291-307.Etienne, H., Bertrand, B., Anthony, F., Côte, F. andBerthouly, M. (1997 a) L'embryogenêse somatique,un outil pour I'amélioration génétique du caféier.ASIC, l ZêrneColloque, Nairobi 21-25 juillet (souspresse).Etienne, H., Lartaud, M., Michaux-Ferriêre, N.,Carron, M.P .. Berthouly, M. and Teisson, C. (1997b) Improvement of somatic embryogenesis inHevea brasilensis (Müll. Arg.) using the temporaryimmersion technique. In vitro cellular andDevelopment Biology.Evans, D.A. and Sharp, W.R. (1986) Somaclonaland gametoclonal variation . Chapter 4. InHandbook of Plant Cell Culture;Tehniques andApplications.. D.A Evans, W.R. Sharp and P.V.Amrnirato, eds, Macmillan Publishing Company,New York, 4: 97-132Freytag, AH., Rao-Arelli, A.H., Anand, S.e.,Wrather, J.A. and Owens, L.D. (1989) Somaclonalvariation in soybean plants regenerated from tissueculture. Plant Cell Rep. 8: 199-202.Haarer, AE. (1969) Produccion moderna de cafe .Instituto dei libro .Edicio revolucionaria R,LaHabana.Karp, A. (1991) On the current understanding ofsomaclonal variation. Oxford surveys of PlantMolecular and Cell Biology. 7: I-58.
35
Larkin, P.J. and Scowcroft, W.R. (1981)Somaclonal variation-a source of variability fromcell cultures improvment.Theo.AppI.Genet. 60: 197214Moharned, AS. and Nabors, M W ( 1990)Somaclonal variant plants and wheat derived frommature embryo explants of three genotypes. PlantCell Rep. 8: 558-560.Monaco, L., Sõndah.l M., Carvalho, A., Crocomo,O. and Sharp, W. (1977) Applications of tissueculture in the improvement of coffee. In: Reinert J.and Bajaj Y. (eds.) Applied and FundamentalAspects ofPlant Cell, Tissue and Organ Culture (pp109-129). Springer-Verlag, Berlin.Murashige, T. and Skoog, F. (1962) A revisedmedium for rapif growth and bioassays withtobacco tissue cultures. Physiol. Plant. 15: 473-497.Neuenschwander, 8. and Baumann, T. (1992) Anovel type of somatic embryognesis in Coffeaarabica. Plant Cell Rep. 10: 608-612.Pétiard, V., Courtois, O., Masseret, 8., Delaunay, P.and Florin, B. (1987) Existing limitations andpotential applications of plant cell cultures. In:Proceedings 4th European Congress onBiotechnology (Arnsterdam), O.M. Neylssl, R.R.Van der Meer, K.e.A.M. Lyuben, eds. ElsevierScience Publisher B.V., 157-169.Pierson, E.S., Van Larnmeren, A.A.M., Schel,J.H.N. and Staritsky, G. (1983) In vitrodevelopment ofembryoids from punched leafdiscsof Coffea canephora. Protoplasma, 115: 208-216.Sõndhal, M.R. and Sharp, Wr. (1977) Highfrequency induction ofsomatic embryos in culturedleaf explants of Coffea arabica L. Z.Ptlanzenphysio. Bd. S. 81: 395-408.Sõndhal, M.R. and Lauritis J.A. (1992) Coffee. In:Biotechnology of perennial fruit crops.Biotechnology in agriculture 8: 401-420.Sõndhal, M.R. and Bragin, A. (1991) Somaclonalvariation as a breeding tool for coffee improvement.In: ASIe. 14eme Colloque ScientifiqueInternational sur le Café. San Francisco 14-19juillet, p 701-710.Srinivasan, e.S. and Vishveshwara, S. (1978)Heterosis and stability for yield in arabica coffee.Indian Genet. and Plant Breed., 38: 416-420.Staritsky. G. (1970) Embryoid formation in callustissues ofcoffee. Acta. BoI. Neerl., 19 (4): 509-514.Teisson, e. and Alvard, D. (1994) A new conceptof plant in vitro cultivation liquid medium:temporary immersion proceedings of the VIII. I.International Congress on Plant Tissue and CellCulture. Florence, Italy 12-17 June: 105-1 09Vasil,V. and Vasil K. (1986) Plant regeneration fromfriable embryogenic callus and cell suspension

cultures of Zea mays L. J Piant Physioi. 124: 399408.Van der Vossen, H.AM. and DJ. Walyaro, D.J.(1981) The coffee breeding programme in Kenya: areview of progress made since 1971 and plan ofaction for the coming years. Kenya Coffee, 46(541): 113-130.Van der Vossen, H.AM. (1985) Coffee selectionand breeding. Chapter 3 in Coffee, botany,biochemistry and production of beans andbeverage,M.N.Clifford, K.C. Willsons,eds.,CromHelm Ltd, Provident House, Burrel Road, p. 48-96.Yasuda, T., Fujii, Y. and Yamaguchi, T. (1985)Embryogenic callus induction from Coffe arabicaleaf explants by benzyladenine. Piant Cell.Physioi., 26 (3): 595-597.
36
Zamarripa, A., Ducos, J.P., Tessereau, H., Bollon,H., Eskes, A.B and Petiard, V. (1991)Developpement d'un procédé de multiplication demasse du caféier par embryogenêse somatique enmilieu liquide.In 14eme Colloque Scientifique Intemational sur le café,ASIC, San Francisco, p: 392402.Zamarripa ,A (1993) Etude et développement del'embryogenêse somatique en milieu liquide ducaféier (Coffea canephora P., Coffea arabica L. ethybride Arabusta). Thêse de doctorat, Ecole NationaleSupérieure Agronomique, Rennes, France, 191 p.

37
DESENVOLVIMENTO DE PLANTAS GENETICAMENTE MODIFICADAS COM VISTAS AUNIFORMIDADE DE MATURAÇÃO DOS FRUTOS DE CAFÉ
Luiz Filipe Pereira, Adilson K. Kobayashi e Luiz Gonzaga Esteves Vieira
IAPAR - Laboratório de Biotecnologia - Londrina - PR - Cx Postal 481 - CEP86000-970
RESUMOA qualidade da bebida do café está diretamente relacionada com a maturação dos frutos. Em regiões com boadistribuição pluviométrica o aparecimento de diferentesfloradas contribui para uma maturação desuniforntcdosfrutos. com efeito direto no preço do produto. As alternativas tradicionais para resolver estes problemas - o
uso de reguladores de crescimento e o emprego de colheita seletiva - apresentam desvantagem de ordemagronômica (queda defolhas efrutos, diminuição da qualidade da bebida) e econômica (aumento do custo deprodução). Em frutos climatéricos, um acúmulo do fitorregulador etileno é observado nas fases iniciais doprocesso de maturação. Uma das alternativas para obter uma maior uniformidade na colheita dosfrutos de catéseria a inibição da síntese de etileno durante os estágios inicias de maturação. Recentemente, com a clonagemdos genes envolvidos na síntese do etileno foi possível a manipulação da produção deste regulador decrescimento emfrutos em processo de maturação. Através da intbição da expressão das enzimas ACC-sintase eACC-oxidase pelo uso da tecnologia do RNA anti-senso, a produção de etileno tem sido bloqueada em frutos devárias espécies, como o tomate e o melão. Este trabalho propõe a transformação de plantas de cufi: com geneanti-senso da enzima ACC-oxidase e seleção de variedades transgênicas apresentando reduzida produção deetileno e inibição da maturação defrutos ainda na planta. 1.1'/0 viria a diminuir a desuniformidade de maturaçãodos frutos, facilitando as operações de colheita e aumentando a qualidade da bebida.
INTRODUÇÃO
o mel horamento genético convencional do cafeeirotem obtido sucesso no atendimento das demandasdo setor produtivo. usando diferentes métodoscomo seleção de populações naturais. seguida decruzamentos e avaliação da progênie.retrocruzamentos e cruzamentos interespecíficos(Sõndahl et al., 1984; Sõndahl e Loh, 1988). Essesesforços levaram a aumentos significativos deprodução através do lançamento de novas cultivaresde cafeeiro. com características genéticassuperiores. que estão sendo introduzidas naslavouras brasileiras. Entretanto. o melhoramentogenético do cafeeiro é limitado por algumascaracterísticas intrínsecas à esta cultura. O longociclo reprodutivo gera uma exposição prolongada adiversos estresses abióticos e bióticos, facilitando oaparecimento de novas doenças e pragas, Também.o longo período juvenil impede que um ciclo deseleção seja completado em menos de 25 anos.dependendo da estratégia de recombinaçãoutilizada.Dessa maneira. a incorporação de técnicasbiotecnológicas poderão reduzir o tempo necessáriopara o desenvolvimento de novos genótipos. commaior produtividade e qualidade. Entre essastécnicas. a transformação genética abre novasperspectivas. permitindo a incorporação. de umamaneira muito mais rápida. de característicasdesejáveis às espécies perenes. sem alterar sua
organização genética global (Manders et al., 1992;Jouanin et al.. 1993; Haines, 1994).Através do uso de técnicas de transformaçãogenética. notoriamente a transformação viaAgrobacterium ou biobalística, a introdução denovos genes em plantas aparece como umaimportante ferramenta que pode ser aplicada emáreas que vão desde os programas de melhoramentoaté estudos básicos de genética. fisiologia e biologiamolecular de plantas. A produção de plantasgeneticamente modificadas (PGMs) tem tambémuma grande importância econômica pelapossibilidade de incorporar novas caracterísiticasagronômicas às plantas cultivadas como resistênciaa herbicidas. insetos ou patógenos e melhora naqualidade dos produtos. No ano de 1998 a áreaglobal cultivada com PGMs foi de cerca de 28milhões de hectares sendo que a previsão do plantio1999 é de cerca de 45 mi Ihões de hectares (James,1998). Atualmente. a soja. o mi lho. a canola e oalgodão representam 98% deste mercado. Nospróximos anos PGMs de batata. tomate. melão.mamão. alfafa, arroz e tabaco. entre outras. devemcomeçar a ser cultivadas em larga escala. Em café(CofJea spp.) PGMs com genes de Bacillusthuringiensis com resistência a insetos estão emfase de testes de campo (Leroy, comunicaçãopessoal). A transformação de café no entanto estálonge de ser um trabalho de rotina. sendo queestudos básicos em cultura de tecidos e.principalmente. em transformação via

Agrobacterium ou biobalística são necessários. Aseguir discutiremos os requisitos básicos para atransformação de plantas de café. assim como aaplicação desta tecnologia na produção de PGMscom maior uniformidade de maturação dos frutos.
CULTURA DE TECIDOS E SELEÇÃO IN
VITRO
Cultura de tecidos de café. A chave para osucesso de qualquer trabalho de transformação deplantas é ter um sistema de regeneração de plantasin vitro eficiente. Os requisitos ideais para tantosão: I) alta capacidade de regeneração no menortempo possível; 2) compatibilidade com sistemasde transformação e seleção in vitro; e 3)independência de resposta entre genótipos. Poucasespécies preenchem esses requisitos e infelizmenteo café não é uma delas. A cultura de tecidos decafé. após os estudos iniciais de Staritsky (1970),teve um grande avanço com os trabal hos deregeneração de embriões somáticos no começo dadécada de 80 (ver Sondahl et aI. 1984. pararevisão). A produção de embriões somáticos a partirde calos provenientes de ex plante foliares foi abase para o desenvolvimento de trabalhos depropagação in vitro em larga escala em meiolíquido (Zamarripa et al., J991; Noriega e Sondahl,1993). A formação de calos. se dá em meio decultura suplementado com os fitorreguladores ácido2A-diclorofenoxiacético (2A-0) ou ácido naftalenoacético (ANA) em conjunto com cinetina (KIN),em concentrações que variam entre diferentesespécies e cultivares. Após a formação de calos otecido é transferido para meio de indução deembrióides com sais inorgânicos reduzidos pelametade. No meio de indução uma maior proporçãode KIN em relação a ANA é utilizada. O tecidoernbriogênico formado pode ser multiplicado emmeio líquido suplementado com 6benzilaminopurina(BAP) ou transferido para meiode desenvolvimento dos embriões sem a presençade fitorreguladores. Alternativamente. aregeneração "direta" de embriões somáticos emmeio com citoquininas (BAP ou 2-isopenteniladenina). sem a passagem por uma fase de calostambém foi demonstrada (Yasuda et ai .. 1985;Hatanaka et al., 1991).
Apesar dos vários estudos já realizados algunsproblemas quanto a cultura de tecido aindapersistem. A regeneração ocorre mais facilmenteem C canephora. encontrando-se uma dificuldademaior para regeneração embriogênica direta ouindireta de C arahica. O tempo para obtenção deembriões é tambem mais demorado para C. arabica
38
(5-8 meses) do que para C canephora (5-8 semanasno caso de embriogênese direta) dificultando otrabalho de transformação e seleção. Além disso. aprodução de embriões secundários até hoje foipouco estudada, mas é uma alternativa para sefacilitar o processo de transformação.
Ldentificação e seleção dos explantes in vitro. Nostrabalhos de transformação genética de plantas énecessário o uso de genes marcadores e de seleçãoalém dos genes de interesse agronômico oucientífico. Os genes marcadores servem paraverificar se o processo de transformação está sendofeito corretamente. identificando células ou tecidostransformados. Um dos genes mais usados comomarcador é o gusA que codifica a enzimaglucuronidase (GUS) e proporciona uma análisehistoquímica ou fluorescente do materialtransformado (Jefferson et ai .. 1987). Na análisehistoquímica o tecido transformado em contato como substrato para a enzima GUS torna-se azul indigoe é facilmente identificado. Outros marcadoresmuito utilizados são os de genes de fluorescênciaverde (GFP), que podem ser detectados in vivo aocontrário das análises com GUS.Dentro de um protocolo de transformação é muitoimportante que o sistema de seleção 111 vitro sejaeficiente, permitindo que o tecido transformado seprolifere com um mínimo de formação de tecido ouembriões não transgênicos. A seleção é feitageralmente com a introdução de genes de seleçãopara resistência a herbicidas ou antibióticos.juntamente com outros genes de interesse. Paratanto, é necessário que se estabeleçam doses doagente seletivo que inibam aproximadamente 95 %da formação de calos ou embriões em tecido nãotransformado. O gene de neornicinafosfotransferase (npt 11) que fornece resistência aoantibiótico canamicina. foi um dos agentes seletivosmais usados nos primeiros trabalhos detransformação. Hoje em dia. os agentes seletivosmais uti Iizados conferem resistência a herbicidas. amaioria com atividade residual baixa. como oglufosinato. o glifosato e o imazapyr que.respectivamente. conferem resistência aosherbicidas Finale®, Round-UP® e Arsenal®. Adose de resposta pode variar de acordo com aespécie e tipo de explante utilizado. Em nossolaboratório foi veri ficado que em explantes foliaresas doses do agente seletivo podem variar entre C.arabica e C canephora (Kobayashi et al., 1998).Ambas espécies apresentaram uma boa inibição naformação calos ou embriões com 0.5 nM deimazapyr. No entanto. quando a seleção foi feitacom glufosinato. a concentração necessária para

inibir a formação de calos em C. arabica foi dezvezes maior do que para C. canephora(TAB. I).
TÉCNICAS DE TRANSFORMAÇÃO
Transformação via agrobacterium.Agrobacterium tumefaciens é uma bactéria gramnegativa que naturalmente tem a capacidade detransferir fragmentos de DNA (T-DNA) para asplantas. Normalmente, a bactéria transfere genesque desregulam o crescimento celular (chamadosoncogenes), ocasionando o aparecimento de galhas,em conjunto com genes de opinas que produzemcarbohidratos e aminoácidos usados pela bactériapara alimentação. Através das técnicas deengenharia genética, é possível substituir os genesda bactéria no T-DNA por genes comcaracterísticas desejáveis. A bactéria serve portantocomo um vetor biológico para transformação. Aeficiência deste processo de transformação dependede vários fatores. O principal é a capacidade dabactéria de infectar a planta, ou seja suaespecificidade, que depende do reconhecimento desinais bioquímicos específicos entre a planta e oAgrobacterium. Essa interação entre a bactéria ehospedeiro é possível na maioria dasdicotiledôneas, mas é limitada entre asmonocotiledôneas. Outros fatores que afetam atransformação são a densidade de Agrobacteriumusada, o tempo de incubação e co-cultivo dosexplantes com a bactéria, pré-tratamento dosexplantes e o uso de culturas auxiliares paratransformação ("feed layers").PGMs de café visando resistência a insetos foramobtidas. tanto para C. canephora como para C.arabica usando Agrobacterium (Berthouly, 1997).A transformação foi muito mais eficiente em C.canephora onde cerca de 100 eventos detransformação independentes foram obtidos,enquanto que em C. arabica houve somente 10eventos de transformação (Leroy, comunicaçãopessoal).
Transformação via biobalística. O uso dabiobalística para transformação vegetal foi criadopor Sanford (1987) e consiste da aceleração departículas de metal conjugadas com o DNA contrao tecido vegetal. Neste processo, o DNA éprecipitado em partículas de metal, que sãoimpulsionadas em alta velocidade, através de umapressão de gás ou descarga elétrica, parapenetrarem no tecido. Vários fatores afetam aeficiência da transformação via biobalística, comopor exemplo a velocidade uti lizada para penetração
39
das partículas, o tamanho das partículas, o précultivo dos ex plantes e o tipo de seleçãoempregado. O processo de biobalística é tambémuma ferramenta importante para a avaliação deconstruções de genes que podem ser analisadostransientemente, ou seja, para verificar a expressãodos genes introduzidos sem que hajanecessariamente uma integração estável destes nogenoma. A introdução de genes via biobalística emcafé foi demonstrada por Boxtel et aI., (1995), ondefoi observada a expressão transiente de GUS emcalos embriogênicos, embriões maduros e folhas.A transformação via biobal ística apresentavantagens e desvantagens em relação atransformação por Agrobacterium. Ela independeda compatibilidade entre a planta e a bactéria equalquer tipo de tecido pode ser usado para atransformação. É uma técnica que vem sendoutilizada não somente em transformação de plantas.mas em fungos, protozoários, insetos e até animais.Entretanto, ela apresenta uma eficiência menor doque a transformação via Agrobacterium e necessitade um eficiente processo de regeneração in vitro(Pereira e Erickson, 1995).
eletroporação de protoplastos. A utilização deprotoplastos (célula vegetal sem a parede celular)foi muito usada em experimentos de transformaçãona década de 70 e começo da de 80. Ela permite umalto grau de transformação, mas apresenta grandesdificuldades na regeneração de plantas. Após aobtenção dos protoplastos, através do emprego deenzimas para digestão da parede celular, osprotoplastos mais o DNA exógeno são submetidos adescargas elétricas que ao abrir poros na membranacelular permite a passagem do DNA para o interiordas células. Apesar de ser um método atualmentepouco utilizado para transformação, ele tem suavai idade principalmente para estudo de construçõesde genes e de expressão transiente. Em café, ocultivo de protoplastos e os primeiros resultados emmanipulação genética destes através deeletroporação foram relatados (Barton et al., 1991).Entretanto. as plantas transformadas não foramcapazes de florescer, demonstrando que o usorotineiro deste protocolos é ainda limitado devidoas dificuldades associadas a de regeneração deplantas a partir da cultura de protoplastos.
CONTROLE DA MATURAÇÃO DOS FRUTOS
A qualidade da bebida do café está diretamenterelacionada com a maturação dos frutos. Regiõescafeicultoras onde há uma boa distribuiçãopluviométrica no final do inverno e começo da
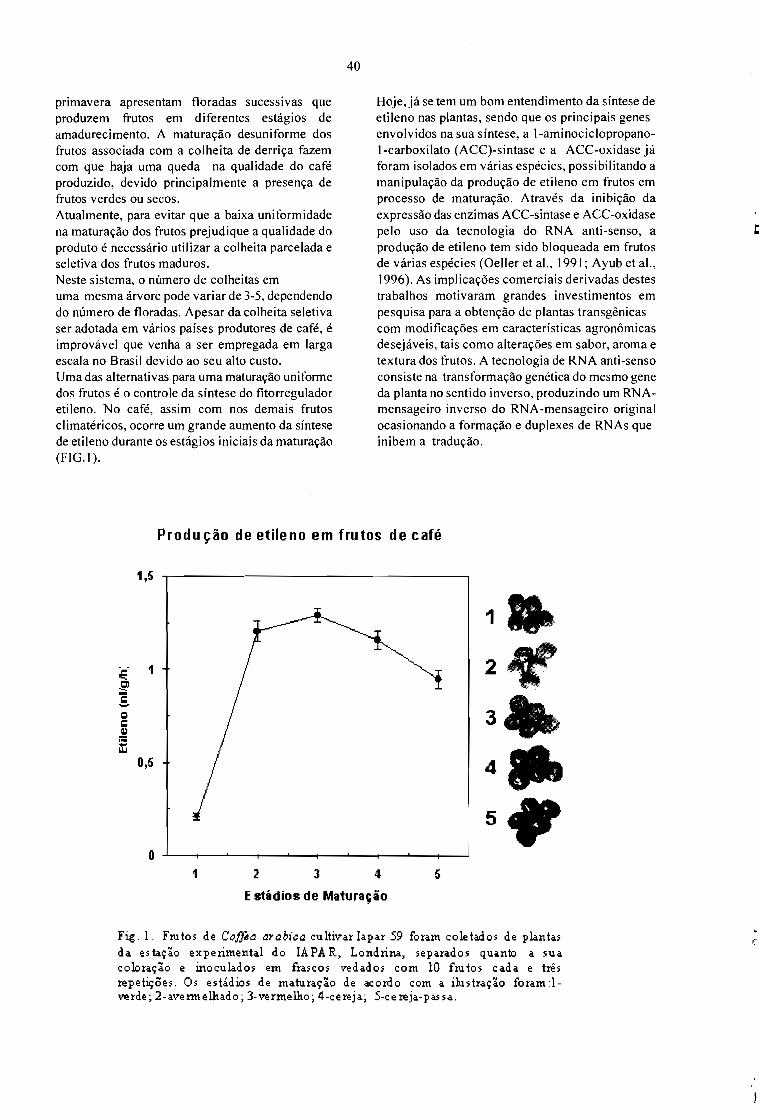
40
primavera apresentam floradas sucessivas queproduzem frutos em diferentes estágios deamadurecimento. A maturação desuni forme dosfrutos associada com a colheita de derriça fazemcom que haja uma queda na qualidade do caféproduzido, devido principalmente a presença defrutos verdes ou secos.Atualmente, para evitar que a baixa uniformidadena maturação dos frutos prejudique a qualidade doproduto é necessário utilizar a colheita parcelada eseletiva dos frutos maduros.Neste sistema, o número de colheitas emuma mesma árvore pode variar de 3-5, dependendodo número de floradas. Apesar da colheita seletivaser adotada em vários países produtores de café, éimprovável que venha a ser empregada em largaescala no Brasil devido ao seu alto custo.Uma das alternativas para uma maturação uniformedos frutos é o controle da síntese do fitorreguladoretileno. No café, assim com nos demais frutosclimatéricos, ocorre um grande aumento da síntesede etileno durante os estágios iniciais da maturação(FIG.I ).
Hoje, já se tem um bom entendimento da síntese deetileno nas plantas, sendo que os principais genesenvolvidos na sua síntese, a l-aminociclopropanoI-carboxilato (ACC)-sintase e a ACC-oxidase jáforam isolados em várias espécies, possibilitando amanipulação da produção de etileno em frutos emprocesso de maturação. Através da inibição daexpressão das enzimas ACC-sintase e ACC-oxidasepelo uso da tecnologia do RNA anti-senso, aprodução de etileno tem sido bloqueada em frutosde várias espécies (Oeller et al., 1991; Ayub et aI.,1996). As implicações comerciais derivadas destestrabalhos motivaram grandes investimentos empesquisa para a obtenção de plantas transgênicascom modificações em características agronômicasdesejáveis, tais como alterações em sabor, aroma etextura dos frutos. A tecnologia de RNA anti-sensoconsiste na transformação genética do mesmo geneda planta no sentido inverso, produzindo um RNAmensageiro inverso do RNA-mensageiro originalocasionando a formação e duplexes de RNAs queinibem a tradução.
Produ ção de etileno em frutos de café
o
1 Da~. 1 2~Qlc -.-Q 3c!~
81-0,5 4
5 ft
1,5
1 2 3 4 5
Estádios de Maturação
Fig. 1. Fm to s de Coffio. o.ro.bú~o. cultivar Iap ar 59 foram coletados de plantasda es tação experimental do IA PA R, Londrina, separados quanto a suacoloração e inoculados em frascos vedados com 10 fru tos cada e trêsrepetições. Os estádios de maturação de acordo com a ihrs traç ão foram:1verde ; 2-ave nn elhad o ; 3-verme lho; 4-cerej a; 5-c e reja. passa.

Em nosso laboratório recentemente foi iniciado umprojeto que propõe a transformação de plantas decafé com gene anti-senso da enzima ACC-oxidase eseleção de variedades transgênicas apresentandoreduzida produção de etileno e inibição damaturação de frutos ainda na planta. Isto viria adiminuir a desuniformidade de maturação dosfrutos, facilitando as operações de colheita eaumentando a qualidade da bebida.
REFERÊNCIAS
Ayub, R., M. Guis, M.B. Amor, L. Gillot, J.P.Roustan, A. Latche, M. Bouzayen, and J.c. Pech.1996. Expression of ACC oxidase antisense geneinhibits ripening ofcantaloupe melon fruits. NatureBiotechnology 14:862-866.Barton, c., T. Adams, and M. Zarowitz. 199 I.Stable transformation of foreign DNA into Coffeaarabica plantlets. ASIC, 141 Coll., SanFrancisco, USA.Berthouly, M. 1997. Biotecnologias y tecnicas dereproduccion de materiales promisorios em Coffeaarabica. Simpósio Latinoamericano de caficulturaXVIII :25-50.Hatanaka, T., O. Arakawa, T. Yasuda, N. Uchida,and T. Yamaguchi. 1991. Effect of plant growthregulators on somatic embryogenesis in leafcultures of Coffea canephora. Plant Cel1 Reports10:179-182.Haines, R. 1994. Biotechnology in forest treeimprovement. FAO, Rome. 230 p.James, C. 1998. Global Review ofCommercializedTransgenic Crops: 1998. ISAAA Briefs No.8.ISAAA: lthaca, NY.Jefferson. R.A., T.A. Kavanagh, and M. W. Bevan .1987. GUS fusions: B-glucuronidase as a sensitivemarker in higher plants. EMBO Journal 6:39013907.Jouanin, L. A.C.M. Brasileiro, lC. Leplé, G. Pilate,and D. Com. 1993. Genetic transformation: a short
41
review of methods and their applications, resultsand perspectives for forest trees. Ann. Sci.For. 50:325-336.Kobayashi, A.K., lC. Bespalhok, L.F. Pereira, andL.G. Vieira. 1998. Effects of imazapyr andglyphosate on cal1us proliferation and somaticembryogenesis in coffee. Genetics and MolecularBiology Supl. 21 :387.Manders, G., M.R. Davey, and r.a Power. 1992.New genes for old trees. J. Exp. Botany 43: 11811190.Noriega, C. and Sondahl, M. 1993. Arabica coffeemicropropagation through somatic embryogenesisvia bioreactors. ASIC, 15:73-8 I.Oeller, P.W., L. Min-Wong, L.P. Taylor, D.A.Pike, and A. Theologis. 1991. Reversible inhibitionof tomato fruit senescence by antisense RNA.Science 254:437-439.Pereira, L.F. and L. Erickson, 1995. Stabletransformation of alfalfa (Medicago sativa L.) byparticle bombardment. Plant cell rep.14:290-293.Sõndahl, M.R.; Nakamura, T.; Medina-Filho, H.P.;Carvalho, A.; Fazuoli, L.c. & Costa, W.M. 1984.In: Ammirato, P.V.; Evans, D.A.; Sharp, W.R. &Yamada, Y. (eds) Handbook ofPlant Cell Culture,vol3. Macmillan Pub. Co., New York, pp 564-590.Sõndahl, M.R. and W.H.T. Loh, 1988. In: Clarke,RJ. & Macrae, R. (eds) Coffee, vol 4. Elsevier,London, pp 235-261.Staritsky, G. 1970. Embryoid formation in calluscultures of coffee. Acta, Bot. Neerdl. 19:509-514.Yasuda, T., Y. Fujii, and T. Yamaguchi. 1985.Embryogenic callus induction from Coffeaarabica leaf explants by benzyladenine. PlantCel1 Physiol. 26:595-597.Zamarripa, A., J.P. Ducos, H. Bollon , M. Dufour,and V. Petiard. 199 I. Production d'embryonssomatiques de cafeier em milieu liquide: effetsdensité dinoculation et renouvellement du milieu.Café Cacao Thé 35:233-244.


43
PROGRAMA BRASILEIRO DE BIOTECNOLOGIA APLICADA AO CAFEEIRO
Alemar B. Rena e Antônio de Pádua Nacif I
I Programa Nacional de Pesquisa e Desenvolvimento do Café, Brasil.
BREVE HISTÓRIA DA BIOTECNOLOGIADO CAFEEIRO NO BRASIL
Provavelmente, o primeiro trabalho debiotecnologia do cafeeiro foi publicado porStaritsky, em 1970. No Brasil, a primeirareferência é a de Sharp et alii, em 1973, masdesde 1970, Sondhal e colaboradores, no IAC, jáestavam pesquisando na área. Os primeirostrabalhos que trouxeram uma contribuiçãoconcreta para a biotecnologia do cafeeiro foram osde Sondhal e Sharp (1977) e Sondhal et alii(1979a, I979b ), quando se demonstrou aembriogênese somática de alta e baixa freqüência.Este início foi, de certa forma, bem revisto porSondhal e Sharp (1979). Infelizmente, em 1984, oDr. Maro Sondhal deixou o Brasil e abiotecnologia do cafeeiro ficou órfã até meados dadécada de noventa, salvo alguns esforçossolitários realizados principalmente por LuizCarlos S. Ramos, no IAC, João Manetti Filho, noIAPAR, Moacir Pasqual, na UFLA e LaércioZambolim, na UFV. Mas, na verdade, muitopouco se acrescentou ao que já se sabiainternacionalmente, em primeiro lugar, pelanatureza lenta intrínseca da própria pesquisa e. emsegundo, pelos recursos financeiros escassos. Oque se fez, basicamente, nesse período, foi adaptarmetodologias às variedades comerciais brasileiras.Assim, por exemplo, pequeno progresso foi feitocom respeito à micropropagação em escalacomercial de germoplasmas de café, conquistaaltamente desejável à cafeicultura.Em 1977, com a constituição do ConsórcioBrasileiro de Pesquisa e Desenvolvimento doCafé (CBP&D/Café), novas perspectivas foramabertas à biotecnologia do cafeeiro. Já sob ainfluência do Programa Nacional de Pesquisa eDesenvolvimento do Café (PNP&D/Café), osprimeiros trabalhos brasileiros em biologiamolecular foram publicados recentemente pelogrupo do BIOAGRO. na UFV.
OBJETIVOS GERAIS DO PROGRAMA DEBIOTECNOLOGIA DO CAFEEIRO NOBRASIL
Ainda que se considere a biotecnologia uma áreamuito jovem da ciência, o Brasil tem sedescuidado da sua aplicação ao cafeeiro. Sãopoucos os grupos brasileiros que a ela se dedicam
e a pesquisa que eles desenvolveram até 1998 foiisolada, bastante incipiente e sem uma política decoordenação nacional e de ajuda financeiraadequada. Os grupos trabalharam quase sempreem laboratórios mal montados e com mão-de-obraespecializada deficiente, além de não estaremperseguindo um objetivo claro e comum. Apesquisa que eles realizaram buscou sempre obterinformações priorizadas pelo grupo, semintegração com outros grupos, o que teriapermitido a concretização de objetivos maisamplos. em menor tempo, a menor custo e queatendessem aos anseios da cafeicultura brasileira.O mérito maior deste programa, que agora seimplanta, é a promoção da integração da pesquisabiotecnológica cafeeira no Brasil e a definição deprioridades, sem perder de vista a cafeiculturacomo um todo. Ainda mais. acena-se aopesquisador com a possibilidade de financiamentoda pesquisa, em geral o seu maior e às vezes únicoproblema.Há grandes diferenças laboratoriais e humanasentre as instituições brasileiras que se propõem arealizar pesquisa de biotecnologia cafeeira.Algumas são mais completas e outras muitodeficientes. Conclui-se que todas necessitam demão-de-obra técnica especializada, principalmentede indivíduos de nível médio, além de recursosfinanceiros específicos para a capacitaçãolaboratorial e de manutenção da capacidade fisicainstalada.As ações do PNP&D/CAFÉ buscam estabelecermetas, de acordo com a demanda da cafeiculturabrasileira. A biotecnologia é uma ferramentafundamental, junto ao melhoramento clássico, naobtenção de híbridos de café portadores decaracterísticas agronômicas e industriais deprocessamento desejáveis. Há cada vez mais anecessidade de se ter cafeeiros que melhoraproveitem os fatores de produção como a luz, aágua e os minerais, que sejam resistentes a pragas,doenças e nernatóides, o que apresenta um forteapelo ecológico. e com características dequalidade de grão desejável à indústria. Medianteo melhoramento clássico, já foram sintetizadosmuito híbridos que apresentam várias destascaracterísticas, mas que, pela sua naturezagenética. não podem ser propagados via sementes.Daí a imperiosa necessidade de se desenvolvertécnicas de micropropagação vegetativas quepermitam a exploração desses híbridos em escala

Os projetos e subprojetos que constituem oprograma de Biotecnologia Aplicada ao Cafeeirosão apresentados a seguir.
Projeto I: Micropropagação e preservação depropágulos,Líder: Luis Carlos S. Ramos. IAC.Objetivos:Este projeto tem prioridade máxima e visa amultiplicação de gemas nodais ou axilares e aembriogênese somática. em biorreatores. Serãodesenvolvidas técnicas de micropropagação emescala comercial de vários germoplasmas(cultivares e híbridos) do cafeeiro. Ao mesmotempo. serão desenvolvidos protocolos para apreservação do material micropropagado, o quefacilitará o seu transporte e comercialização.Subprojetos:Micropropagação do cafeeiro tCoffea sp) viaembriogênese somática e biorreator do tipoimersão temporária e imersão permanente.Responsável: João Batista Teixeira. CENARGEN.Bases preliminares para a micropropagação docafeeiro mediante o cultivo de gemas nodais viabiorreatores.
comercial. Esta é a prioridade número um desteprograma.O processo de obtenção de novos híbridos evariedades deve ser contínuo, mas, como ocafeeiro é um planta perene, pode demandardécadas mediante o melhoramento clássico. Daí anecessidade urgente de se desenvolver técnicas debiologia celular e molecular que permitamacelerar o melhoramento do cafeeiro. Hoje, hágrande interesse no reconhecimento e nacaracterização ao nível de DNA dos grandesgerrnoplasmas, com fins de melhor orientar amanipulação genética no melhoramento via abiotecnologia e de melhor descrever as plantasmelhoradas com vistas a obtenção de patentes eoutras apropriações legais.Também. cada vez mais está-se usando no Brasilo processo de preparo do café por via úmida. Esseprocedimento gera um problema ambiental. que éo grande volume de resíduos do processamento docafé na propriedade agrícola. que deve seradequadamente eliminado, sem a contaminaçãodo solo e dos cursos de água. Problemassemelhantes surgem com os restos da indústria docafé solúvel. A biotecnologia pode proverbioprocessos que permitam transformar essesresíduos em subprodutos inócuos. ou mesmoúteis. ao homem e ao meio ambiente.
PROJETOS EPROGRAMA
SUBPROJETOS DO
44
Responsável: Luis Pedro Barrueto Cid.CENARGEN.Embriogênese somática indireta via culturalíqüida em genótipos de CofJea arabica e Coffeacanephora.Responsável: Antônio Teixeira Cordeiro. UFV.Produção in vitro de café Conilon.Responsável: Marcos Oliveira Athayde,EMCAPA.Propagação de híbridos de cafeeiro usandotécnicas de biotecnologia como suporte aprogramas de melhoramento genético.Responsável:Moacir Pasqual, UFLA.Clonagem de plantas especiais de cafeeiros paraestudos agronômicos.Responsável: Luis Carlos S. Ramos. IAC.
Projeto 11: Genoma e citogenética do cafeeiro.Líder: Ney Sussumu Sakiyama, UFV.Objetivos:Neste projeto serão estabelecidos a variabilidade.o mapeamento genético e. finalmente. o mapagenôrnico do cafeeiro. Primeiramente. este estudofornecerá subsídios moleculares ao melhoramentogenético clássico e ao biotecnológico, permitindoa manipulação mais consciente do germoplasma.Em segundo lugar. constituir-se-á numaferramenta imprescindível na identificação.separação e classificação do germoplasmacafeeiro disponível no Brasil. E. em terceiro lugar.propiciará a oportunidade da legitimação dapropriedade genética. incluindo a obtenção depatentes de plantas mel horadas.Subprojetos:Caracterização de genótipos de Coffea canephorapelo uso de marcadores moleculares RAPO.Responsável: Maria Amélia Gava Ferrão.EMCAPA.Mapeamento molecular e uso de seleção assistidapor marcadores de DNA para ferrugem.nematóide e antracnose do cafeeiro.Responsável: Carlos Roberto Riede. IAPAR.Identificação de cultivares de cafeeiros (Coffeaarabica) por marcadores moleculares.Responsável: Maria Lucia Crochernore, IAPAR.Marcadores moleculares para caracterização deespécies e variedades de CofJea e para genes deinteresse agronômico.Responsável: Paulo Maurício Ruas. UE Londrina.Caracterização de cafeeiros utilizando marcadoresmoleculares: identificação de linhagens evariedades do Instituto Agronômico de Campinas.Responsável: Carlos Augusto Colombo. IAC.Desenvolvimento de marcadores AFLP para aanálise genômica em cafeeiro.Responsável: Messias Gonzaga Pereira. UENF.
'-

CENARGEN.
Projeto V: Fermentação e manejo de resíduosdo café.Líder: Carlos Ricardo Soccol. UFPR.Objetivos: Desenvolver bioprocessos para adetoxificação e valorização de bioresíduos de cafée a produção de esporos de fungos. bactérias etoxinas utilizadas no biocontrole de nematóides.Subprojetos:Desenvolvimento de bio-processos paradetoxificação e valorização de bio-resíduossólidos do café no Estado do Paraná.Responsável: Carlos Ricardo Soccol, UFPR.Desenvolvimento de bio-processos para produçãode esporos de fungos. bactérias e toxinasutilizados no bio-controle de nematóides em café.Responsável: Carlos Ricardo Soccol, UFPR.
No Brasi I. fora as instituições associadas aoCBP&DI Café e as recém-associadas aosubprograma de biotecnologia, há poucapossibilidade de cooperação técnico-científica naárea de biotecnologia aplicada ao cafeeiro. Há. noentanto. vários grupos de biotecnologia aplicada aoutras culturas. especialmente na área molecular.e em menor escala na de rnicropropagação, quepoderiam permitir algum intercâmbio.possibilitando o treinamento do pessoal do café.Mas a contribuição seria pequena. No setorprivado nacional. há a empresa FITOLlNK. depropriedade do Dr. Maro Sondhal, situada emRibeirão Preto. São Paulo. que tem amplaexperiência em micropropagação do cafeeiro. Estatalvez seja a única possibilidade concreta deparceria que poderia ser vantajosa para ospesquisadores do CBP&DI Café. no Brasil.No âmbito internacional. a possibilidade maior eproficua de convênios. com vistas aos estudos debiotecnologia do cafeeiro. seria com a França.Neste país. os grupos do erRAD. em Montpellier.e da Nestlé, em Tours, poderiam contribuir tantona área de micropropagação como na áreamolecular. Mas o forte deles é mesmo amicropropagação, tendo sido obtidos grandesavanços na embriogênese somática. Portanto.convênios como o CIRAD e a Nestlé sãoaltamente desejáveis. Outras possibi Iidades deparcerias menores poderiam também ser buscadasem Cuba e na Costa Rica. onde há algumaexperiência de micropropagação.
Desenvolvimento de marcadores de DNA para omelhoramento genético do cafeeiro.Responsável: Ney Sussumu Sakiyarna, UFV.Citogenética molecular de germoplasma de café.Responsável: Carlos Roberto de Carvalho. UFV.Caracterização citológica de espécies e decultivares de CofJea spp.Responsável: Cecília Alzira Ferreíra PintoMaglio, IAC.
Projeto 111: Mutagênese, hibridação somática eprodução de haplóidesLíder: João Batista Teixeira. CENARGEN.Objetivos:Neste projeto procura-se estimular a variabilidadegenética no cafeeiro, a introgressão de genes deresistência a moléstias e pragas. tanto emcultivares Arábicos como em clones de Canéforae a produção de plantas de café haplóides ecultivares em homozigose.Subprojetos:Cultura de tecidos em CofJea para a produção devariantes somaclonais.Responsável: Antônio Teixeira Cordeiro. UFV.Detecção de variação somaclonal em Coffeacanephora através do uso de marcadores RAPD.Responsável: Maria Amélia Gava Ferrão.EMCAPA.Cultura de anteras de Coffea arabica.Responsável: Moacir Pasqual, UFLA.
Projeto IV: Desenvolvimento de cafeeirostransgênicos.Líder: Luiz Gonzaga E. Vieira. IAPAR.Objetivos:Desenvolver metodologia para obtenção deplantas transformadas de café. No presenteprojeto. serão pesquisadas tanto técnicas básicasde transformação como serão desenvolvidossubprojetos aplicáveis. É um projeto que apenasse inicia. que apresenta elevada complexidade ealto custo. ademais de ser demorado e não seespera obter qualquer planta transgênica emmenos de 10 anos.Subprojetos:Manipulação genética da síntese deetileno no café visando a uniformidade dematuração dos frutos.Responsável: Luiz Gonzaga Esteves Vieira.IAPAR.Transformação de CofJea sp visando aobtenção de plantas resistentes aos herbicidasinibidores da enzima sintase do ácidohidroxiacético (AHAS).Responsável: Luiz Gonzaga Esteves Vieira.IAPAR.lntrodução e expressão de genestransgenes em cafeeiro (Coffea sp) através doprocesso biobalístico e do sistema Agrobacterium.Responsável: Ana Cristina Miranda Brasileiro.
45
POSSIBILIDADE DETÉCNICO-CIENTÍFICAINTERNACIONAL.
COOPERAÇÃONACIONAL E

REFERÊNCIAS
Sharp, W.R., L.S. Caldas, 001 Crócorno, L.C.Mônaco e A. Carvalho. 1973. Production ofCoffea arabica callus of three ploidy leveis andsubsequent morphogenesis. Phyton 31: 67-74.Sondhal, MoRo e W.R. Sharp. 1977. Highfrequency induction of somatic embryos incultured leaf explants of Coffea arabica L. Z.Ptlanzenphysiol. 81: 395-408.Sondhal, M.R. e W.R. Sharp. 1979. Research inCoffea spp, and appl ications of tissue culturemethods. In W.R. Sharp, P.O. Larsen, E.F.Paddok e V. Raghavan (eds) Plant cell and tissueculture - Principies and applications. Ohio StateUniversity Press, Columbus, USA.
46
Sondhal, MoR.. Ll., Salysbury e W.R. Sharp.1979a. SEM characterization of embryogenictissue and globular embryos during highfrequency somatic embryogenesis in coffee calluscells. Z. Pflanzenphysiol. 94: 185-188.Sondhal, M.R., DoA. Spahlinger e WoR. Sharp.1979b. A histological study of high frequency andlowfrequency induction of somatic embryos incultured leaf explants of Coffea arabica L. Z.Ptlanzenphysiol. 94: 10 1-108.Staritsky, G. 19700 Embryoid formation in callustissues of coffee. Acta Bot. Neerl. 19: 509-514.

47
APLICACIONES DE LA BIOTECNOLOGÍA AL MEJORAMIENTO GENÉTICO DEL CAFÉ, ENCOLOMBIA.
Germán Moreno, Myriam de Pena, Ricardo Acuíia, María Eugenia Aponte, Juan Carlos Herrera,Diana Molina, Alvaro Gaitán, Marco Cristancho, Pilar Moncada, Carmenza Góngora y GabrielCadena.
CENICAFE, Chinchiná, Colombia.E-mail: [email protected]
INTRODUCCION:
EI mejoramiento genético dei café en Co lomb iatiene una trayectoria cercana a los 60 afios,durante los cuales se han conseguido resultadossobresalientes con los que se ha dado respuesta alos problemas mas apremiantes de la caficulturanacional (Moreno, 1994). En las ultimas décadasse han producido en el mundo avances muyimportantes en el campo dei cultivo in vitro detejidos y de la biología molecular, los cuales hanmotivado a los investigadores en café a buscarlesaplicaciones en los programas de mejoramientogenético. con el fin de hacerlos mas eficientes(Anthony et ai, 1997; Petiard et ai, 1993).La finalidad de esta exposición es, de una parte,referir las experiencias de CENICAFE aI poner enmarcha un programa de investigación enbiotecnología aplicada ai mejoramiento genéticodel café. y de otra, informar sobre los principalesavances ai respecto en los últimos 10 afies. Me haparecido conveniente referir estas experienciasdesde mi punto de vista de fitomejoradorconvencional, poseedor solamente de una visióngeneral de las bondades y Jimitaciones actuales dela biotecnología.
ALG UNAS CONSIDERACIONESGENERALES SOBRE LA CAFIClILTURACOLOMBIANA.
En Colombia se cultivan unas 900.000 hectáreascon las variedades de C. arabica Caturra, Típica yColornbia, reconocidas por sus buenascaracterísticas agronómicas y excelente calidadde la bebida. La producción media dei país es de
.. 1200 kgr. de café pergamino seco por hectárea yafio. cantidad que es 5 veces inferior ai potencial
productivo de los materiales mejorados, cuandoson cultivados con practicas agronómicasapropiadas.Cerca de 15 patógenos atacan aI café, perosolamente uno, el hongo Hemileia vastatrix,causante de la roya de la hoja, es limitante porquesu efecto reduce la producción en 20 %,aproximadamente. La variedad Colornbia, conresistencia a esta enferrnedad, fue entregada a losagricultores en 1982, un afio antes de la lIegadadel patógeno aI país, y hasta ahora ha sidoutilizada exitosamente para su combate (Castilloy Moreno. 1986).Varias plagas afectan ai café, pero solamente labroca (Hypothenemus hampei) alcanzadimensiones epidémicas que afectan la cantidad ycalidad de la cosecha. Una búsqueda exhaustivade fuentes de resistencia en el pool genéticoprimario de C. arabica, variedades comerciales yalgunas especies diploides, se adelanta enColombia desde hace 7 aüos, pero hasta elmomento no se han encontrado fuentes de interés(Moncada, 1992).
INVESTIGACION EN BIOTECNOLOGIA.
EI objetivo es desarro1Jar técnicas de cultivo invitro y de biologia molecular para incorporariasen el mejoramiento del café. con énfasis en lasolución de problemas en los cuales los métodosconvencionales no han sido exitosos Paracumplir este objetivo, las actividades seconcentran en tres campos principalesrelacionados con técnicas de cultivo in vitro, conla transforrnación genética de variedades de caféy con el desarrollo de marcadores moleculares(figura I).

48
Figura I. Organización deI programa de Biotecnología de CENICAFE.
Biotecnologia para el meJonmiento de variedades de até
Cultivo in vitro.En el café la propagación in vitro tiene variasaplicaciones. pero en el caso de Colombia suutilización principal es para proporcionar tejidoembriogénico para la transformación genética. Delas técnicas de propagación in vitro disponibles,la embriogénesis somática fue escogida por ser lamas eficiente para producir el tejidoembriogénico necesario.Durante los últimos 10 anos CENICAFE hainvestigado sobre los factores que afectan larespuesta embriogénica en cada una de las 4etapas relacionadas con la embriogénesissomática: lnducción. Proliferación. Germinación
y Endurecimiento (Aponte y Molina. 1995;Aponte. 1993). Todos estos factores, y susinteracciones, se han estudiado en progenies deCaturra X Híbrido de Timor, poseedoras deresistencia a la roya, que son los materiales demas interés para el mejoramiento (tabla 1). Soloel 20 % deI material probado tiene una capacidadembriogénica alta. cantidad que es insuficiente.porque la meta es obtener materialestransformados, de diferente origen, paraconformar variedades compuestas. Actualmentese realizan nuevos experimentos con losgenotipos con baja respuesta, aplicandomodificaciones recomendadas para materialesrecalcitrantes.
Tabla I. Factorcs relacionados con la Embriogénesis somática. cstudiados en CENleAFE.
Inducción Proliferación G erminación Endurecim ienlo
Planta dnnant e Sinc r oniz ac ié n M e dio s de cultivo Conce ntr aciones- Genolipo - Separación de - Deshidratac ión de macro y
Fuent e de e xplanr e agregados osmótica m icr onut r ent e s- Est adu fisiológico celulares - 's lve les de sacarosa- Remoción me c ánlca Filtración - Reguladoresde flores y frutos - De nsida d de hormonales
- Fro r sc íón inóculo Condiciones de- Esr r a t os de la planta crecimiento
Condiciones de Fotoperiodnc re c sn ic nr o
Eumpe r iodu
- Te m per a t te- a
- Tie m po de subcult ivo- \1 odalidad de c ult ivn- Se le cc ión dei t ejidu
durante c l subc utnvu
\1 c diu de c ulrive
- Sales orgánicase ino r g ámc as
- Reguladores dec re c en iento
- M arca de agar

Producción de plantas haploides. Lajustificación principal para la obtención deindividuos haploides es disponer, para lostrabajos de mejoramiento, de plantascompletamente homocigotas, en un periodorelativamente corto. Plantas haploides se hanobtenido por métodos in vivo o in vitro endiferentes paises (Couturon, 1982; Carneiro,1997; Duffor et ai. 1995; Ascanio y Areia, 1994).Sin embargo, la poca reproducibilidad y la bajaeficiencia de los métodos han hecho que latecnologia de los haploides sea poco utilizada enprogramas de mejoramiento convencionalEn nuestro laboratorio se han reproducidodiferentes protocolos de cultivo de anteras y sehan podido regenerar varias plantas de Caturra yde otros genótipos, las cuales, sin embargo, nohan sido haploides (Herrera y Moreno, 1995).Dada la baja frecuencia de inducción de tejidoembriogénico, en CENICAFÉ se hizo unseguimiento histológico dei desarrollo de lasanteras en cultivo. comprobándose que el tejidoformado era de origen somático (Herrera, 1998).Ef cultivo de microsporas también se ha intentadoen CEN1CAFE, pero hasta el momento solo hasido posible inducir la formación de pequenasmicrocolonias de células en división, pero confrecuencias muy bajas, inferiores ai 1 %.Actualmente se evalúa la respuesta de lasmicrosporas a tratamientos con colchicina, y losresultados preliminares son promisorios.
49
Estos resultados permiten concluir que losmétodos utilizados para inducir la androgénesisen anteras de café y para el cultivo demicrosporas, todavia son deficientes y por tanto.se requiere explorar nuevas estrategias.
Transformacíón genética de variedades elite decafé.La introducción aI café de genes heterólogos queinhiban el desarrollo de la broca es el objetivoprimordial de las investigaciones adelantadas enCENICAFE en el campo de la transformacióngenética. La ausencia de fuentes de resistencia deinterés dentro dei germoplasma disponible, lasperdidas económicas que ocasiona la plaga y ladificultad para su control, justifican plenamentelos esfuerzos que se hagan en este sentido.EI desarrollo de una variedad mejorada, conresistencia a la broca, depende de los avances quese logren en diferentes etapas que, en su conjunto.constituyen el plan de investigación. Estas etapasse representan esquemáticamente en la figura 2 ya ellas nos referiremos a continuación.
Técnicas de regeneración. Disponer de unsistema de regeneración eficiente es indispensablepara la transformación genética. Como vimos.CENICAFE dispone ya de la metodologiaapropiada
Fig.2 Esquema para desarollo de una variedad de café transgénica, con resistencia a broca
... V ARIEDAD RESISTENTEA LA BROCA
~Iuacionesenel campo

Técnicas de transformación. Gracias a acuerdoscooperativos celebrados con las Universidades deCornell y Purdue, de los Estados Unidos.CENICAFE ha evaluado sistemas directos eindirectos de introducción dei DNA. En el primercaso se ha ensayado la transferencia por medi osquímicos, como el polietilen glícol y por mediosfísicos. como el bombardeo con partículas. obiolística. En el segundo caso se ha utilizado elvector natural Agrobacterium tumefaciens. EIadiestramiento recibido en esas técnicas ha hechoposible que actualmente se ejecuten directamenteen CENICAFE.En todos los sistemas ensayados ha habidoavances importantes. pero parciales. Por ejemplo.CENIC AFE cuenta con la metodología para elcultivo in vitro de protoplastos y para laregeneración de plántulas a partir de estos (Acufíay de Pena. 1991). pero en el caso de latransformación de los protoplastos por medio dePEG. solo se ha podido obtener altos niveles deactividad estable de genes marcadores. comoGUS. en tejido embriogénico y en callos (dePena. 1995). No se han recuperado plántulas apartir de los callos transformados. posiblementepor una baja capacidad de regeneración deigenotipo empleado, después de transformado. talcomo ha ocurrido en otras especies.En el caso dei bombardeo de partículas. losaspectos destacables sono por una parte. el uso deesta metodología para comprobar la expresióntransitoria de promotores de interés. Cuatropromotores se han probado con éxito con estatécnica (35 S. Doble 35 S. PAl y Alfa-tubulina).Hay que destacar también el uso de esta técnicapara demostrar la expresión estable de genes deinterés. Plántulas transformadas con uno de estosgenes están creciendo en medio selectivo parakanamicina, aunque las pruebas definitivas de latransforrnación no se han realizado.En el caso de Agrobacteriun, se ha comprobado elalto poder infectivo de 3 cepas desarmadas.actuando sobre tejido embriogénico. los trabajosen este campo apenas se están iniciando; por seruna técnica de transformación limpia que permitesolo la introducción en el tejido dei segmento deDNA deseado, Ia hacen muy atractiva para elfuturo.
Mecanismos de regulación de genes de interés,En el mercado están disponibles diferentespromotores. útiles para regular la expresión de losgenes heterólogos. Sin embargo. CENICAFEtiene interés en desarrollar sus propiospromotores. principalmente por dos razones. En
50
prirner lugar, porque se necesita que los genes deinterés se expresen en el sitio de la plantafrecuentado por la broca. es decir, en elendosperma dei grano de café. en un momentoque coincida con el ataque dei insecto. Unpromotor que reúna estas características debe serespecifico y como tal. no se encuentra entre losdisponibles. En segundo lugar. por lasrestricciones que impone la legislación sobrepropiedad intelectual. que protegen lospromotores disponibles en e1 mercado y creandependencia de sus poseedores.Cenicafé cuenta con 3 promotores ais lados decafé. EI primero es el promotor que regulatemporal y espacialmente la expresión de laArabicina, Ia principal proteína de reserva deiendospermo de café. descrita por primera vez porAcufía y otros (1999). Se trata de un promotorespecifico para café. cuya acción puede facilitarenormemente el uso de genes heterólogos en elcaso de la resistencia a la broca. Actual mente seevalúa la capacidad de este promotor para regularla expresión transitoria y estable de genesrelacionados con compuestos inhibidores de esteinsecto.EI segundo es el promotor que regula la expresióndei gen constitutivo de Alfa-tubulina en café. Estepromotor ya demostró ser capaz de regular laexpresión dei gen GUS en tejido embriogénico yen plántulas de café provenientes deembriogénesis somática. EI tercero es unpromotor inducible que regula la expresión deFenil alanina arnonioliasa, PAl. enzimainvolucrada en mecanismos de defensa (Gaitán yAldwinkle.1997).los 3 promotores mencionados se encuentranlistos para probar la expresión de genesheterólogos que controlen la formación decompuestos inhibidores de la broca.
Identificación de genes que controlancompuestos inhibidores de la broca. En loreferente a la búsqueda de genes heterólogos queconfieran algún nivel de resistencia a la broca. sehan realizado investigaciones en varios campos.Uno de estos consiste en la evaluación, en dietasartificiales, dei efecto de diferentes extractossobre la broca. los mejores resultados seobtuvieron con extractos de cultivos deStreptornyces con actividad quitinolítica. Despuésde realizar la ingeniería genética de dos de estosgenes que tienen esa actividad, se procedió aintroducirlos en vectores de expresión en plantasde tomate. manzano y café. Hasta el momentosolo las plantas de tomate han sido evaluadas en

-+
I
invernadero, y se ha observado un efectodetrimental sobre el insecto Trichoplusia ni.(Broadway et ai, 1998).También se ha trabajado en la búsqueda yevaluación rápida de fuentes vegetales quecontienen inhibidores de amilasas que afectan labroca, con resultados muy promisorios (Valencia,1996). Finalmente, como parte de un acuerdo conla Universidad de Maryland, se están buscandogenes de interés a partir de hongosentomopatógenos.EI aspecto para destacar es que se está reuniendoun grupo de genes activos contra la broca, queusados con los promotores antes mencionados,facilitaran la generación de poblaciones deplantas con combinaciones de genes deresistencia contra este insecto.
Prueba de los materiales mejoradostransformados. Como se ha visto, se han hechoavances importantes, pero aun no sabemos quetan cerca o lejos estamos de la meta final. Cuandose alcance, la variedad transformada deberá serevaluada rigurosamente por mejoradores,entomólogos y biólogos moleculares, para juzgarsus cualidades respecto a aspectos críticos comolos siguientes :Los genes heterólogos introducidos afectan otrascaracterísticas de interés ? Cómo se comporta ladescendencia de las plantas transgénicas ? Cómose comporta la resistencia a la broca en diferentesambientes ? i,Cómo se afecta la población deiinsecto frente a materiales resistentes? Esperamostener la respuesta cuando lIegue el momento.
Utilización de marcadores moleculares.De las herramientas que ofrece la Biotecnología,los marcadores moleculares son quizás los quemas aplicaciones tienen en un programa demejoramiento genético. En CENICAFE lostrabajos en este campo son muy recientes, perohay algunos resultados. EI objetivo principal esidentificar genes responsables de caracteres deinterés, que sirvan para agilizar la selección, yademás, que puedan ser utilizados eninvestigaciones relacionadas con el conocimientode la diversidad genética y a mas largo plazo, conla construcción de un mapa genético de laespecie C. arabica.Los genes de interés escogidos son los quecontrolan la resistencia a la enfermedad de losfrutos, o CBO, causada (Colletotrichum kahawe) ,y los que gobiernan la resistencia a la roya deicafeto. Los genes responsables de altaproducción, que en varios cultivos se están
51
seleccionando con la metodología conocida comoQTL, no son prioritarios en nuestro caso, porqueun aumento considerable de la producciónnacional puede ser obtenido usando vías masexpeditas, como por ejemplo, aprovechandomejor el potencial productivo de los materialesmejorados.Respecto a los marcadores relacionados con laresistencia ai CBO, en CENICAFE hemosseparado materiales resistentes y susceptibles,inoculados en los laboratorios deI CIFC, enPortugal, usando dos de los marcadores RAPOdesarrollados por Agwanda y Lashermes enFrancia (1997). En otros materiales, tambiéninoculados en el CIFC, los resultados no han sidoigualmente satisfactorios, por lo que ahoraestamos desarrollando marcadores SCAR a partirde los RAPO disponibJes, con el objetivo delograr una metodología que discrimine losmateriales mas eficientemente.En el caso de la roya dei cafeto, estamosinteresados en identificar marcadoresrelacionados con la resistencia incompleta, paralo cual se adelanta un proyecto usando el Análisisde Segregantes Agrupados o BSA (BulkSegregant Analysis), pero todavía no hayresultados.Los primeros trabajos sobre diversidad genéticalos hemos hecho usando marcadores RAPO sobremateriales de C. arabica recolectados en Etiopíapor las misiones de la FAO Y de ORSTOM, ysobre las principales variedades americanas. Oieziniciadores (primers) ensayados han mostradoalgún grado de polimorfismo entre lasintroducciones etíopes. En las variedadescomerciales la variabilidad encontrada es menor(Cristancho, 1998). La colección de germoplasmade CENICAFE, como la de la mayoría de lospaíses occidentales, es rica solamente enintroducciones de la especie C. arabica, quecomo es sabido tiene un grado de diversidadreducido. Sin duda, los trabajos relacionados conel polimorfismo avanzarían mas rápidamente sison ejecutados por los países que poseen lascolecciones mas diversas, como es el caso de lospaíses africanos.Finalmente, Cenicafé tiene como meta, a largoplazo, desarrollar un mapa para la especie C.arabica. Sin embargo, en este caso la falta de unapoblación experimental adecuada es unalimitación importante. En efecto, es necesario quelos parentales utilizados den origen a unasegregación importante para las características deinterés agronómico, y como vimos, elpol imorfismo en los materiales tetraploídes es

escaso. Por otra parte, en café el único mapagenético conocido es el desarrollado por losinvestigadores franceses (Paillard et al, 1996). Sinembargo, es posible que en los materialesutilizados para desarrollar este mapa (dobleshaploides de C. canephora), no estén presentescaracterísticas de interés agronómico útiles en elcaso de C. arabica.Para tratar de solucionar estos problemas, enCENICAFE estamos desarrollando diferentespoblaciones experimentales, y además, estamosproduciendo marcadores de diferente naturaleza.Hasta el momento disponemos de un grupo demarcadores RFLP, RAPO Y Microsatélites, quevan a comenzar a ser utilizados cuando laspoblaciones estén disponibles.
CONCLUSIONES.
La investigación en el campo de la biotecnologíaes costosa, y por tanto, las prioridadesestablecidas deben ajustarse muy bien a lasnecesidades de cada país. Las alianzas entrecentros de investigación pueden contribuir a lareducción de los costos.En CENICAFE ya se encuentran disponiblesvarias métodos de biotecnología para iniciar suaplicación en el mejoramiento genético deI café.La identificación de nuevos genes de interésayudará a los mejoradores en la selección degenotipos superiores.La obtención de una variedad transgénica en cafétodavía esta lejana, aI menos en Colombia. Sinembargo, cuando esto se logre, tal variedaddeberá satisfacer las exigencias de loscaficultores, de los consumidores, del mercadomundial y de la comunidad en general.
BIBLIOGRAFIA
Acufía J. R. and De-Pena, M. 1991. Plantregenration from protoplasts of embryogenic cellsuspensions of Coffea arabica L. cv. Caturra.Plant CelJ Rep. 10: 345-348.Acufía R., R. Bassüner, V. Beilinson, H. Cortina,G. Cadena, V. Montes, and N. C. Nielsen. 1999.Coffee seeds contain 11S storage proteins.Physiol. Planto 105: 122-13 1.Agwanda C. O., P. Lashermes, P. Trouslot, M. C.Combes and A. Charrier. 1997. Identification ofRAPO markers for resistance to coffee berrydisease, Colletotrichum kahawae, in arabicacoffee. Euphytica 97: 241-248.Anthony F., B. Bertrand, P. Lashermes , and A.Charrier. 1997. La biologie moléculaire en appui
52
à l'amélioration génétique du caféier Arabica.Plantations, recherche, développement,Novembre-Décembre: 369-377.Aponte A., M.E. 1993. Somatic embryogenesisinduced by culture on single media in coffeeplants from crosses of Coffea arabica by Timorhybrid. XV Coloquio Científico Internacionalsobre Café. ASIC. p. 82-88. Montpellier, Francia.Aponte A., M.E., and Mofina V., D. 1995.Evaluación de genotipos de café por su capacidadembriogénica "in vitro", IV Congreso SociedadColombiana de Fitomejoramiento y Producciónde Cultivos. Chinchiná, Colombia. p. 120.Ascanio C. E.,and M. A. Areia, 1994. Efecto deIestado de desarrollo de las anteras y de un shocktérmico sobre la androgénesis en Coffea arabicaL. varo Garnica. Café, cacao Thé, 38(2): 75-79.Broadway, M. R.; Gongora, C. ; Sanderson, P. J.;Monroy, AJ.; Bennet, C.K.; Warner, BJ. andHoffrnann, P.M. 1998. Novel Chitinolyticenzymes with Biological Activity againstherviborous insects. Journal of Chemicalecology, 24, 985-998.Carneiro. M.F. 1997. Coffee biotechnology andits application in genetic transformation.Euphytica, 96: 167-172.Castillo Z., J., and Moreno R.. G. 1988. Lavariedad colombia: selección de un cultivarcompuesto resistente a la roya dei cafeto.Cenicafé. Chinchiná, Colombia. 171 p.Couturon E. 1982. Obtention d 'haploidesspontanés de Co.ffea canephora Pierre parI'utilisation du greffage d'embryons. Café,Cacao, Thé Vo XXVI n:3: 155-160.Cristancho, M.A. 1998. Informe Annual deLabores. CENICAFE. Chinchiná, Colombia.Dufour M., M. Jimenez, and D. Duris. 1995.Haplomethods: factors controlling callusobtention on Co.ffea arabica anthers. XVIColoquio Científico Internacional sobre Café.ASIC. p. 765-770. Kyoto, Japón,Gaitán, A.L., and H. Aldwinckle. 1997. Cloningand evaluation of gene promoters in Co.ffeaarabica. Phytopathology 87: S31.Herrera, J.C. 1998. Informe Annual de Labores.CENICAFE, Chinchiná, Colombia.Herrera P., J.C., and Moreno R., G. 1995. Estudiode algunos factores que afectan el desarrollo "invitro" de microsporas aisladas de café (Co.ffeaarabica L.).IV Congreso Sociedad Colombiana deFitomejoramiento y Producción de Cultivos.Chinchiná, Colombia. p. 130.Moncada, M. del P .1992. Búsqueda de fuentes deresistencia genética a la broca Hypothenemus

hampei, en germoplasma de café (ProyectoMEG.OS). Cenicafé. Chinchiná, Colombia.Moreno R, G. 1994. Contribución deimejoramiento genético aI desarro 11o de lacaficu\tura colombiana. Innovación y Ciencia,3(2): 1-6.Paillard M., P. Lashermes, and V. Pétiard. 1996.Construction of a molecular linkage map incoffee. Theor. Appl. Genet., 93: 41-47.Pétiard V., H. Bollon, J.P. Ducos, B. Florin, M.Paillard, J. Spiral, and A. Zamarripa. 1993.Biotechnologies appliquées au caféier. XV
53
Coloquio Científico Internacional sobre Café.ASIC. p. 56-66. Montpellier, Francia.Pena, M. de. 1995. Development of a stabletransformation procedure for protoplast of Coffeaarabica cv. Colombia. Thesis Philosophy Doctor.West, la Fallette, IN. EEUU. 76p.Valencia, A. 1996. Evaluación de inhibidores deamilasas y proteasas y estudio de las enzimasdigestivas de la broca dei café (Hipothenemushampei). Proyecto de investigación ENT-1000,CENICAFE, Chinchiná, Colombia.

55
AVANÇOS NA OBTENÇÃO DE D1-HAPLOÓIDES EM CULTIVARES DE Coffee arabica L.
Maria Filomena Carneiro
Centro de Investigação das ferrugens do Cafeeiro Quinta do Marquês, 2780- Oeiras, Portugal
INTRODUÇÃO
Entre as mais de 70 espécies do gênero Co./feaexistentes, apenas duas, Co./fea arabica e a Co./feacanephora são cultivadas nos diferentes paísesprodutores de café.Coffea arabica são tetraplóide (2x= 44) eautogâmica e todas as outras incluindo a espécieCo./fea canephora são diploídes (2x=22). O ciclode vida do cafeeiro é muito longo e qualquerprograma de melhoramento com o objetivo deobter uma cultivar nova requer longos períodosde tempo para a seleção que podem ir até 30 anos.As técnicas modernas da cultura de células e detecidos vegetais permitem reduzir aquelesperíodos longos de seleção e ultrapassar asdificuldades dum programa de melhoramentotradicional.A androgênese é uma técnica que permite obterlinhas homozigóticas de cultivares novas em curtoespaço de tempo, facio este que pode encurtardrasticamente um programa de melhoramentopossibilitando a introdução e manipulação deespécies novas. Cada antera de cafeeiro tem cercade 2 000 40 000 micrósporos e cada um delespode dar uma nova combinação durante a meiose.A maior vantagem dos haplóides é o fato de elesserem usados para produzir com rapidez linhashomozigóticas. Como por definição os haploídestêm apenas um número básico de cromossomas, aduplicação de cromossomas desses indivíduosressalta numa fixação instantânea da linha. Estefato minimiza o tempo necessário para obtençãode homozigocidade e consequentemente evitaproblemas de amostragem encontrados quando atestagem é feita numa geração precoce.Em cultura, os micrósporos seguem diferentesvias de androgénese que permitem a formação dehaploides quer diretamente por embriogénese ouindiretamente através de formação de calo. Doponto de vista do genetecista a formação deembriões diretamente a partir de grãos de póloensão preferidos a formação indireta de embriões viasolo.Teoricamente. por cultura de anther/micrósporo,pode ser produzido um elevado n" de plantashaplóides.Infelizmente, o processo na cultura de anteras/micrósporo no cafeeiro tem sido lento.
Apesar de referências a cultura de anteralpólen sereportarem a 1973 (simp et aI. 1973), apenas em1987 foram apresentados resultados referindo aregeneração de plantas (Ascanio & Areia 1987;Carneiro, 1987).A nossa intervenção fundamenta-se naapresentação dos resultados obtidos nesta área noCIFC. o trabalho desenvolvido teve comoprincipal objetivos otimizar as espécies deobtenção de plantas di-haplódes em cultivarescomerciais e transferir a tecnologia para ascultivares Catimor e Sachimor a fim de obtercultivares com resistência às principais doençasdo cafeeiro, nomeadamente à ferrugem alaranjadadas folhas, Hemileia vastratix e a antracnose dosfrutos verdes, Co./fetrochium kakawae.Todas as experiências foram realizadas emdiferentes progenie de cvs Catuai e Catimor. Paradeterminar o estádio de desenvolvimento na alturade colocação em cultura e a resposta gametolificados micrósporos durante a cultura foramutilizados dois métodos: Carmim acética a 2% eDAPI (4,6-Diamino-2-phenylindole 2 HCL (Schweizer and Nagi, 1976, Detchepare et aI,1989). Para verificar a viabilidade dosmicrósporos na altura da imaculação e durante acultura, foi utilizado o teste de Diacetato deFluoresceina (FDA)(Widholm, 1972).
ESTÁDIO DE DESENVOLVIMENTO DASANTERAS
Reveste-se de grande importância a seleção debotões florais no estádio de desenvolvimentoóptimo. Contudo estes estádios são difíceis deidentificar em rotina.Embora o estádio óptimo para colocar as anterasou micrósporos isolados seja a I mitose dos grãosde pólen, este estádio depende muito das espécies.Por exemplo, no tabaco é a I mitose do grão depólen enquanto que nos cereais e na Brassica porexemplo é de pouco antes enquanto o miscróporoé uninucleado (Powell 1990). Este fato foi jásal ientado por vários autores, nomeadamente porSunderland (1974). Nitsch (1974 a.b; 1977; 1983).Dunwell (1976) e o Carneiro (1987). Contudo,um pequeno período de tempo a volta da 10mitose parece o momento ideal para induzirdivisão anormal dos micrósporos para seguir a viaesporofitica. O tratamento pelo frio pareceaumentar a divisão anormal dos micrósporos

( Nitsch et Norreel 1973: Picard et de Buyser,1975; Corduan et Spix, 1975). Outros fatores quecondicionam uma ambriogénese é a composiçãodo meio de cultura, diferentes hidratos de carbonoem diferentes concentrações, o tratamento pelocalor, acompanhado ou não de centrifugação.Todos os trabalhos desenvolvidos por nóssuportam estes conceitos prévios.
CULTURA DE MICRÓSPOROS
As primeiras experiências forma realizadas com acv Catuai (CIFe 8223/205) e para a extração demicrósporos foram utilizadas duas técnicas deNistch (1974 a.b; 1977) e a técnica de ' Shedpollen' de Sunderland (1974. 1979).Os micrósporos foram cultivados em diferentesmeios de cultura Nistch( 1974 a.b; 1977) fazendovariar as concentrações de reguladores decrescimento, concentrações de sacarose, vitaminase aminoácidos. L-glutamine e L- serina.As anteras isolodas em nO de 50 correspondendo acerca de 10 botões florais foram colocadas em 5ml de meio basal Nistch ( 1974b; 1977) e forammantidas em condições de escuridão durante O.24. 48. 72 e 96 h. Ao fim destes períodos deindução os micrósporos forma extraídos usando a'técnica de Nistch e foram cultivados em diferentesmeios de cultura acima referidos.Quando da inoculação quase todos os micrósporosse encontravam no estágio de 1 mitoseapresentando dois núcleos. Durante a cultura. osmicrósporos apresentaram comportamentosdiferentes. nomeadamente a-) algunsapresentaram divisões repetidas sem divisão dacélula originando uma estrutura multicelular queacabou por abortar; b-) outros micrósporosseguiram uma divisão nuclear seguida de divisãocelular originando um callus e finalmente algunsmicrósporos apresentaram divisão originandoduas células . as ' células filhas' (célulasandrogénicas) que seguiram uma divisão igual eoriginaram os pró-embriões. Estes pró-embriõesglobulares passaram por outras fases dagerminação dos embriões. i.e, embriões na faseglobular, coração. torpedo e com folhascotiledonares. Este comportamento foi verificadoem todos os meios de cultura ensaiados exceto nomeio que não continha reguladores decrescimento. (Carneiro. 1987).Os sinais de embriogénese desapareceram emamostras onde se utilizaram períodos indutivossuperiores a 72 ou 96 h. Experiências realizadasulteriormente permitiram concluir que os períodosindutivos superiores a 96 h eram letais econduziram à morte das anteras econsequentemente dos micrósporos. Verificou-se
56
também que os meios de cultura tinham um papelimportante no desenvolvimento de embriões e queeste último apenas tinha lugar na presença dosaminoácidos, a vitamina mio-inositol e dosreguladores de crescimento AIA e ZEA~Numa outra experiência, foram cultivados botõesflorais da cv Catuai (CIFC 8223/50) em 7diferentes estádios de desenvolvimento: prémeiose, 1° meiose, 2° meiose, tétrada, tétrada dernicrósporos, micrósporos isolados e grãos depólen maturos em meio MS (1962),suplementados com BAP, AIA, mio-inositol, L Glutarnina, e L - serina.Os micrósporos foram isoladospela técnica deNistch e cultivados em meio liquido estacionário.Neste meio, foi possível induzir a embriogénese eisolar todas as suas fases. O meio de cultura teveas necessárias alterações de acordo com a fase daembriogénese.Dos 7 diferentes estádios de desenvolvimentoensaiados. os melhores resultados foramconseguidos quando as anteras se encontravam nafase pós-meiótica e antes da maturação dos grãosde pólen i.e, nas fases de tétrada, tétrada demicrósporos e micrósporos isolados.No que se refere ao tratamento pelo frio o prétratamento a 4_6° durante 48 h dado aos botõesflorais de cv Catuaí (CIFC 8223/50). aumentou adivisão anormal dos micrósporos.No entanto, as percentagens de micrósporosseguindo isolados a partir dos botões não forammuito diferentes das percentagens obtidas pelosmicrósporos isolados a partir dos botões nãotratados. i.e, colocados a temperatura ambiente .Este fato sugere que a micrósporogénese dosbotões colocados no frio se desenvolvesimultaneamente com a dos botões colocados nascondições normais de ambiente.Os embriões apresentaram um aspecto vegetativo'fraco e a conversão em plântuls foi muito dificil.A composição dos meios de cultura. é outro fatorde importância fundamental. nomeadamente asconcentrações dos reguladores de crescimento. Osaminoacidos L - glutamina e L - serina parecemdesempenhar um papel importante napercentagem dos micrósporos seguindoembriogénese.Foram também estudadas as influências do prétratamento pelo calor (30° C) durante O, 24 e 48 hcombinado com centrifugação dos botões floraisantes da extração dos micrósporos na cv catimor(CIFC 7963/2) Os micrósporos foram cultivadosno meio MS suplementado com BAP e ANA.Verificou-se que os micrósporos resultantes dotratamento dado durante 24 h deram origem aembriões e microcalos ao fim de 13 dias semcultura.

ClILTURA DE ANTERAS
As anteras pertencentes à cv Catuaí (CIFC8223/61-124) foram cultivadas no meio de culturaMS suplementados com tiamina - HCL, mioinositol e diferentes combinações de auxinas: 2. 4-O, ANA, AIB e AIA. A composição do meio decultura foi modificada durante a cultura de acordocom as diferentes fases do desenvolvimento dasanteras. As culturas foram cultivadas no escuro eem condições de luz. Para a proliferação de calliindução de rebentos/embriões, as culturas foram
incubadas à luz. .Tal como na cultura de micrósporos foramensaiados sete diferentes estádios dedesenvolvimento de anteras e o efeito do prétratamento pelo frio, 4_6° durante 0,24, e 48 h.foram avaliadas as percentagens de fenolização,n" de anteras calogénicas, peso fresco do calo e aqualidade do calo.As anteras cultivadas de ação da auxinas; 2,4 - O,ANA, AIB e AIA. A percentagem de oxidaçãofoi baixa nas I° semanas da cultura. Todas asanteras cultivadas no meio MS sem reguladoresde crescimento não originaram calo e acabarampor morrer. A 1° subcultura teve lugar 12 semanasmais tarde, exceto nas anteras crescendo no meiosuplementado com AIB (apenas numa placa) queoriginou um calo embriogénico sem mudar demeio de cultura. Foi observando um caloembriogénico emergindo das anteras apresentandouma bem desenvolvida plântulaForam também avaliadas os pesos frescos doscalos produzidos usando duas concentrações ( 2 e3 mgl - I ) de 2,4 - O e de ANA. Quando seestudou a cv Catuaí (C IFC 8223/61-124)verificou-se que os calos produzidos pelas 4auxinas eram diferentes e variavam e qualidade eem quantidade. Assim, os calos induzidos em 2,4-O eram friáveis e alguns eram embriogénicosoriginando embriões e sua conversão emplântulas. ANA, induziu calos esbranquiçados eesponjosos que eram pequenos que os induzidospor 2,4 - O, IBA, induziu calos brancos enodulares dando origem a um grande nO deembriões.A densidade de calos foi elevada no meiosuplementado com 2,4 - O. Em cada tratamentoquer em cv Catuaí quer em cv Catimor, as 20anteras originaram calos brancos e friáveis. Noque se refere a cv Catimor (CIFC 7963/212)observaram-se diferentes tipos de calos em funçãodas auxinas usadas Estudo da estabilidadecitológica em plantas regeneradas a partir de umaembriogénese repetitiva ou recorrente.Anteras pertencentes a botões florais de cv Catuaí(CIFC 8223/61-124) em 7 diferentes
57
estádios de desenvolvimento foram cultivados nomeio MS suplementado com tiamina-HCL. Mioinositol, cisteina-HCL, sacarose 40 g/I e diferentescombinações e concentrações de 2,4 - O, ANA,AIB, AIA, BAP e 2iP.Anteras crescendo no meio de culturasuplementado com AIB ongmaram um caloembriogénico nodular. Este calo iniciou umaembríogénese secundária ou adventícia e aconversão em plantas teve lugar em 6 meses domeio de conversão. As plantas foram avaliadaspara o nível da plóidia usando a técnica decontagem de cloroplastos nas células guarda doestoma. Foram estudadas 167 plantas. Estas foramdivididas em 4 grupos. O primeiro grupoconstituído por 3 plantas (1,796%) apresentaram8-10 cloroplastos foram consideradas como sendon=2x planta haplóide (di-haplóide) emcomparação com o n'' de cloroplastos observadosnuma polanta haplóide que é de 8,3±1,2cloroplastos que consideramos como sendo umaplanta controlou standard. O segundo grupo deplantas apresentou 14-16 cloroplastos ( 82,030%)e foram considerados como sendo plantastetraplóides em comparação com o n" 13,9±1,8 decloroplastos por um par de células guarda doestoma de uma planta tetraplóides(2n=4x)Os dois outros grupos de plantas apresentaram 5-7(4,191%) e 16-20 (lI ,976%) nO de cloroplastos emcada par de células guarda dos estornas. Estes doisgrupos precisam de um estudo mais detalhadopara se definir o grau de ploidia, provavelmente oI°será um aneuplóide e o 2° um poliplóide.Quer o método de contagem de cloroplastos nascélulas guardando estoma quer o métodoconvencional de contagem de nO de cromossomasnas células meristemáticas são métodos lentos ede baixo grau de acurácia. Qualquer que seja oobjetivo da experiência, as plantas com o desejadograu de ploidia pode ser imediatamenteselecionado após a transferência para o meio deconversão ou logo após a transplantação para osolo.
CONCLUSÕES E PERSPECTIVAS
São conhecidas há muito tempos, no ReinoVegetal, a ocorrência natural de haplóides.Quando se verificou que ou duplos haplóideseram de grande importância para o uso domelhorador, verificou-se que estas plantas não seencontravam disponíveis para a maioria deculturas e além disso não existiam meios paraproduzir.Guha e Maheshwari (1964) conseguiu, comsucesso, obter pela primeira vez embriõescultivando anteras de Datura innoxia. Usando esta

aproximação, foram obtidos embriões quercultivando anteras ou micrósporos num grande n"de espécies e em alguns casos provou-se possívelregenerar plantas a partir destes embriões.Apesar deste sucesso, a obtenção de linhashaplóides continuam difíceis de produzir paraculturas importantes e por esta razão ainvestigação celular e molecular tem-seconcentrados nas primeiras fases da embriogénesedos micrósporos.Os estádios mais precoces da embriogénese naBrassica demostraram envolver uma I divisãopós-meiótica simétrica do que um I divisãoassimétrica do pólen (Pechan and Keller 1988,Zaki and Dickinson 1990) Esta falta de assimetriafoi acompanhada pela formação de uma paredetipo-esporofitica, acumulação de amido ecuriosamente, a síntese de proteínas relacionadascom stress, semelhantes a s encontradas após umchoque por calor (Dickinson, 1992). De acordocom Benito Moreno e colaboradores (1988) hátambém dados na Nicotiana que indicam que odesenvolvimento pode ser dirigido numa viaesporofitica ou gametofitica de acordo dom acomposição do meio de cultura.No que se refere ao cafeeiro, os trabalhos deSharp et al. (1973) e Monaco et aI.referem apenas a formação de calos e a umpequeno crescimento de um micrósporo.Na década de 80, surgem os primeiros trabalhosem que Carneiro (1987) refere a indução deembriogénese em 40-50% de micrósporos de cvCatuai e Ascanio e Areia (1987) também pelaprimeira vez apresentam resultadossurpreendentes de regeneração de plantashaplóides de Coffea arabica cv Garnica. Emsucessivos trabalhos foram referidas indução deembriogénese direta a partir de micrósporos eindução de embriogénese indireta seguida deprodução de plantas' prováveis di-haplóides' porcultura de anteras. (Carneiro, 1991; Carneiro,1992; Carneiro,l993; Carneiro, 1995 a.b;Carneiro, 1997b).Existem várias vias para aumentar a produção dehaplóides , nomeadamente o tratamento pelo frio,adição de carvão ativado e particularmente o usode culturas de micrósporos isolados em meioslíquidos estacionários e com agitação(Sunderland, 1974). Este último procedimentoparece ser uma muito importante via de
58
androgénese que permite a obtenção de elevadosníveis de embriões haplóides. Contudo há outrosimportantes fatores que parecem ter um papeldeterminante na indução e aumento deandrogénese, tais como estádio dedesenvolvimento da antera quando da inoculaçãoou colocação em cultura e acomposição do meio de cultura. Muitos autorestêm salientados estes fatores e as experiênciasrealizadas por nós, suportam estes conceitosprévios. Assim, o estádio uninucleado domicrósporo na altura da inoculação parece sercrucial para mudar a via normal dedesenvolvimento do grão do pólen para uma viaanormal que dá origem a formação de embriõesou calo. A I mitose é um importante momentopara induzir divisão anormal dos micrósporos nosentido de embriogénese .Dos setes estádios de desenvolvimento da anteraestudados, em diferentes progenie de cvs Catuaí eCatimor concluímos que os melhores resultadosforam obtidos quando as anteras se encontravamno estádios pós-meiótico e antes da maturação dogrãos de pólen.O pré-tratamento pelo frio de 48 h a 4_6° C dadoaos botões florais aumentou a divisão anormal dosmicrósporos no sentido da via esporofitica. Noque se refere à importância da composição domeio de cultura, os resultados evidenciam aimportância dos reguladores de crescimento e dasduas concentrações.aminoácidos L - glutamina e L - serina, quandoforam utilizados no meio líquido estacionário nacultura de micrósporos, demonstraram ter umpapel importante no aumento de androgénese.Entre os reguladores de crescimento acombinação de BAP com AIB mostrou ser a maiseficaz quer quando foram utilizados anteras quermicrósporos isolados independentemente dascultivares usadas.Os resultados apresentados são bastantespromissores e constituem um incentivo para aprossecução de estudos relacionados com aandrogénese do cafeeiro, nomeadamente emcultivares Catimor e Sachimor, resistentes àsprincipais doenças do cafeeiro e em algumasprogênies que apresentam simultaneamenteresistência á ferrugem alaranjada e a antracnosedo cafeeiro
L

59
CUSTO COMPARATIVO DE DIFERENTES TÉCNICAS DE CLONAGEM
Maro R. Sondahl l, Clemencia N. Sondahl l e Wallace Gonçalves2
I Fitolink Corp., 6 Edinburgh Lane, Mt. Laurel, NJ 08054 USA. Av.Caramurú, 2100 si 804-ARibeirão Preto, SP 14030-000 Brasil. Tel.-(016) 620-2745 [email protected]
2 Instituto Agronornico, Caixa Postal 28, CEC, Centro de Café, Campinas, SP 13100-000.
INTRODUÇÃO
Sementes é a forma mais comum de propagaçãode plantas, e se aplica sempre que a linhagem éhomozigota ou a espécie permite o uso dehibridização controlada (sementes híbridas). Apropagação vegetativa se aplica quando a espécienão se reproduz por sementes ou não permite aprodução comercial de sementes híbridas,utilizando-se neste caso os métodos deEnraizamento e Enxertia. A Micropropagaçãoé utilizada nos seguintes casos: (a) quando existedificuldade de propagação pelos métodostradicionais; (b) é essencial iniciar a produção apartir de "plantas sadias" (livres de doençasvasculares e vírus); e (c) existe a necessidade deproduzir grandes numeros de mudas em curtotempo. A Micropropagação se baseia nosseguintes métodos de multiplicação in vitro: (I)multiplicação de Gemas Axilares; (2) regeneraçãovia Organogese; e regeneração via Embriogenese.A maioria dos protocolos comerciais demicropropagação utilizam a via de multiplicaçãode gemas axilares preexistentes, em meio semisól ido (Kurtz et aI. 1991). Estes protocolos secaracterizam por três fases, sempre executadassob condições estéreis (in vitro): Inoculação (faseI), Multiplicação (fase 2) e Crescimento/Enraizamento (fase 3). As etapas subsequentesdo processo são realizadas sob condições nãoestéreis (fora do laboratório) e compreendem asseguintes fases realizadas em estufas ou viveiros:Endurecimento (fase 4) e Crescimento Final(fase 5). Planta em fase 5 esta pronta para sertransferida para o local de plantio definitivo.A comercialização de plantas micropropagadaspode ser realizada com mudas na fase 3, 4 ou 5,dependendo das condições do usuário final emcompletar o desenvolvimento das mudas. O preçofinal de mudas fase 3 é dependente dos custos detrabalho sob condições estéreis, a alta demanda demão de obra especializada e dos custo demateriais para preparo dos meios de cultura. Amão de obra é o principal fator de custo na mudafase 3, chegando a representar 60-70% do custofinal desta muda, em processos demicropropagação de gemas axilares em meio
semi-sólido. As mudas da fase 3 são muitodelicadas, pois não tem ainda folhas e raízesfisiologicamente perfeitas. É durante a fase 4(endurecimento) que as mudas diferenciam folhascom estornas e cutículas normais e produzemabundante raízes. Ao fim desta estádio, as mudassão transferidas para recipientes maiores a fim depermitir um maior desenvolvimento durante afase 5. Mudas na fase 4 ou 5 são as maisrecomendadas para serem comercializadas, ouseja, são os tipos de mudas normalmenteentregues ao produtor.A micropropagação via Organogenese não temsido muito utilizada devido a ausência deprotocolos específicos, ou suspeita de elevadoíndice de variação somaclonal. Amicropropagação via Em briogenese vemdespertando grande interesse pela sua alta taxa demultiplicação (menor custo unitário) e pelarelativa estabilidade das mudas produzidas. Tratase de um processo altamente interessante paraplantas tropicais, que possuem altos níveis defenois e taninos (dificeis de enraizar), e paraespécies perenes, quer sejam de clima tropical outemperado, plantas frutíferas ou industriais.Na cultura do café, o método usual de propagaçãoé via sementes. Em caso de café Arabica, espéciede autofecundação, é possível ter plantaçõeshomogeneas (com 98% de homozigose) a partirde seis (6) gerações sucessivas deautofecundação, um processo que exige nomínimo 24 anos de trabalho continuo. EmRobusta no entanto, é impossível ter plantasuniformes via sementes porque trata-se de umaespécie de fecundação cruzada. Portanto, em casode Robusta, grandes ganhos de produção devemser alcançados quando plantas elites sejampropagadas. Existem clones de Robustas em que épossível a propagação por enraizamento, masoutros clones devem ser propagados por enxertiaou embriogenese somática. A propagaçãovegetativa de Arabica se justifica para a imediataexploração comercial de híbridos superiores (altaprodução, resistência a pragas e doenças. e outraspropriedades desejadas). Materiais de Arabicanão permitem a propagação por enraizamento em

escala comercial, portanto devem ser aplicadosmétodos de enxertia ou embriogenese somática.O uso de métodos de micropropagação em caféfoi inicialmente descrito para a multiplicação degemas axilares de ramos ortotrópicos (Custer1980; Sondahl et aI. 1984). Este processo foiutilizado para a micropropagação de clonesselecionados de Catimor em CATI, Costa Rica,dentro do Programa "Promocafe", dando origem auma população de cerca de I0,000 plantas paraensaios de seleção. Este método é mais simplesque embriogenese somática, porém muito poucoeficiente, com baixos rendimentos, e elevadocusto unitário por muda. Portanto, somenterecomendado para certas aplicações de pesquisa,e nunca para propagação comercial de alta escala.O processo de Embriogenese Somática em caféfoi inicialmente descrita para Robusta, usando acultura de ramos ortotrópicos (Staritsky 1970), epara café Arabica a partir de folhas adultas(Sondahl & Sharp 1977). Ligeiras variações nosmétodos originalmente descritos foramapresentados por Dublin (1980) e Yasuda (1985),sempre usando meios semi-sólidos. A produçãode embriões somáticos em café a partir deexplantes de folha ocorre através dois processos:embriogenese de baixa frequência (ou direta) eembriogenese de alta frequencia (ou indireta),qual dá origem a um tecido embriogenico friável(FET). Esta linha de células embriogenicas podeser isolada, multiplicada e/ou preservada sobcongelamento. Acentuado progresso para autilização comercial de embriogenese somáticaem café foram dados pelos protocolos em meioliquido utilizando FET em bioreatores de 5 a 7litros: Zamarripa et ai. (1991) e Noriega &Sondahl (1993). Estes trabalhos mostraram serpossível a produção de 45,000 a 200,000embriões por litro de meio. Um sistemasimplificado e interessante de propagação de caféem meio liquido foi posteriormente apresentadopor Berthouly et ai. (1995), usando recipientes de500 ml de capacidade sob imersão periódica.Com a finalidade de avaliar plantas de caféderivadas do processo de micropropagação, sobcondições de normais de cultivo, foi instalado umensaio em São Sebastião do Paraíso, MG, Brasil,contendo mudas derivadas de embriõessomáticos, provenientes de duas origens: (I)cultivados em meio liquido em bioreatores; e (2)cultivados em meio semi-sólido. Como controle,foram usadas mudas obtidas a partir de sementes.
60
MATERIAL E MÉTODOS
Material Vegetal: Para cultura de tecidos foramusadas folhas adultas de Coffea arabica varoLaurina linhagem #2165. Sementes desta mesmalinhagem foram germinadas para produção dasmudas controle.Cultura de Tecidos: Explantes foliares foramcultivados conforme o método descrito emSondahl & Sharp (1977) e tecidos embriogenicosde alta frequencia (FET) foram isolados paramultiplicação em meio semi-sólido e meioslíquidos. Para a multiplicação em meio liquidoforam utilizados dois modelos de bioreatores, umAplikon de 7 litros e um Ono de 5 litros. Ametodologia usada na multiplicação sob sistemalíquidos seguiu o processo descrito em Noriega &Sondahl (1993).Os embriões somáticos obtidos foram germinadosem condições assépticas e os plantletes obtidos(fase 3) foram transferidos para tubettes comsubstrato artificial e levados para endurecimentoem estufa (New Jersey, USA), com 50% deluminosidade sob nebulização periódica. Asmudas endurecidas (fase 4) foram introduzidas noBrasil via o Serviço de Quarentena doCENARGEN/Embrapa (Brasília, DF). As plantasfase 5 foram levadas para a área experimental emS. S. Paraíso, Minas Gerais para plantio.Simultaneamente, sementes foram germinadas emareia e transferidas para tubettes de café, usandosubstrato artificial, sob luz de 50% no inicio e30% no final do período de crescimento.Área Experimental: As mudas derivadas dacultura liquida foram identificadas como Bl(mod. Aplikon) e B2 (mod. Ono); as mudasobtidas em meio semi-sólido foram marcadascomo TC e as mudas de sementes identificadascomo SE. O plantio foi feito em S.S.Paraiso, MG,no período de 2-4 março de 1995 emespaçamento de 2,0 x 1,0 m, utilizando-se 220plantas de meio liquido, 230 plantas de meiosemi-sólido e 500 plantas de sementes. A áreaexperimental teve um total de 35 parcelas, comum numero de 10-45 plantas/parcela, e o plantiodas parcelas foi feito em delineamentocasualizado.
RESULTADOS E DISCUSSÃO
O processo de micro propagação porEmbriogenese Somática exige inicialmente oisolamento de um "tecido embriogenico" tipoFET, a partir da cultura de folhas das plantasselecionadas. Este tecido FET tem a capacidade

de produzir embriões somáticos, e é multiplicadoinicialmente em meio liquido (cultura emsuspensão), usando-se frascos erlenmeyer de 250ml de volume. Em seguida, este tecidoembriogenico é inoculado em Bioreator paranovos ciclos de multiplicação, e em faseposterior, inicia-se a indução da fase deregeneração de embriões somáticos, e finalmenteentra na fase de maturação dos embriõesproduzidos.A germinação dos embriões somáticos já é feitafora dos Bioreatores, em meio semi-sólido, ououtro substrato qualquer, normalmente ainda sobcondições assépticas. Embriões com cotilédonesplenamente desenvolvidos (fase 3) sãotransferidos para tubettes com substrato artificiale levados para estufa para o endurecimento.Mudas de café derivadas de embriões somáticosque completaram o período de endurecimento(fase 4) foram levadas um local próximo doplantio e deixadas sob condições de viveiro paracompletar o crescimento final (fase 5), antes deserem levadas ao campo. Este fluxo de produçãode mudas de café via a micropropagação.utilizando-se embriões somáticos, em sistemaslíquidos em bioreatores, esta ilustrado na Fig I.processo de estabelecimento do TecidoEmbriogenico tipo FET leva cerca de 6 meses e oestabelecimento de suspensões, requer mais 2meses. As etapas de multiplicação, regeneração ematuração dos embriões dentro de bioreator levaca. de 3 meses. A germinação feita em condiçõesideais de luz e temperatura leva 3 meses. As fasesde Endurecimento e Crescimento final (fases 4 e5) exigem ca. 6 meses. Portanto, a primeiraprodução de mudas micropropagadas de café viaembriogenese somática pelo processo acimadescrito leva um total de ca. 20 meses. Num 20
ciclo de produção, já existe disponível o tecidoembriogenico FET da planta elite selecionada, eportanto a produção de mudas estádio 5, prontaspara plantio, demanda um total de 12 meses.Pela duração das diferentes etapas no fluxo deprodução de mudas via a micropropagação porembriões somáticos apresentado acima, nota-seque a necessidade de um planejamento com I anode antecedência para a produção sequencial demudas, o que é bastante semelhante ao temponormal de produção de mudas via sementes: 7meses da floração ao fruto mais 6 meses paragerminação e crescimento das mudas em viveiro.Qualquer projeto de propagação com estastécnicas vai exigir ainda 8 meses extras, no início
61
do processo, para o estabelecimento das culturasembriogenicas. No caso de propagação porsementes, se necessita de 'um mínimo de 3 anosde crescimento da planta para colher as primeirassementes.A capacidade de multiplicação via embriõessomáticos, em meios líquidos, é muito alta. Porexemplo, se considerarmos uma produção baixade 50,000 embriões por litro de meio, utilizandose bioreatores com dois bioreatores de 10 litros,produz-se um total de I milhão de embriões emca. 3 meses. Considerando-se perdas nas fases degerminação e endurecimento em 50%, estima-seuma produção final de cerca de 500,000mudas/ano, a partir de uma planta híbridaselecionada, ou seja o equivalente para o plantiode 100 ha. Aumentando-se o numero debioreatores é possível atender a uma demandamaior de plantio no ano I. A limitação maior nonumero final a ser propagado não é na fase demultiplicação dentro da Biofábrica, mas sim nasfases de viveiro e plantio.A grande vantagem do processo demicropropagação é a capacidade de produção demudas em alta escala (I - 10 - 50 milhões) apartir de uma única planta, usando pequenossegmentos de folhas. Portanto, é um processo quepermite entrar em produção comercial de "plantaselites" (híbridos) em curto espaço de tempo, epraticamente sem limitações de quantidadesiniciais.Este trabalho apresenta os primeiros dadoscomparativos do método de micropropagação emcafé versus sementes, em plantas semi-adultas (4anos) mantidas em condições normais de cultivo.Foram feitas avaliações anuais quanto aocrescimento vegetativo e aspecto morfológicodestas plantas de café, a fim de estimar qualquerefeito na qualidade da muda (sementes vs.micropropagação) e a presença ou não devariabilidade. Para avaliação morfológica seutilizou medidas de altura da planta, diâmetro daplanta, diâmetro da haste principal e comprimentodos ramos laterais.Não foi possivel detectar nenhuma diferença tantono crescimento vegetativo das plantas quanto naprodução. Os dados da avaliação morfológicarealizada após a 2a colheita (maio/99) sãoapresentados no Quadro I. Usando os valores dasplantas derivadas de sementes como Controle(100%), comparou-se os parâmetros das mudasderivadas de micropropagação.
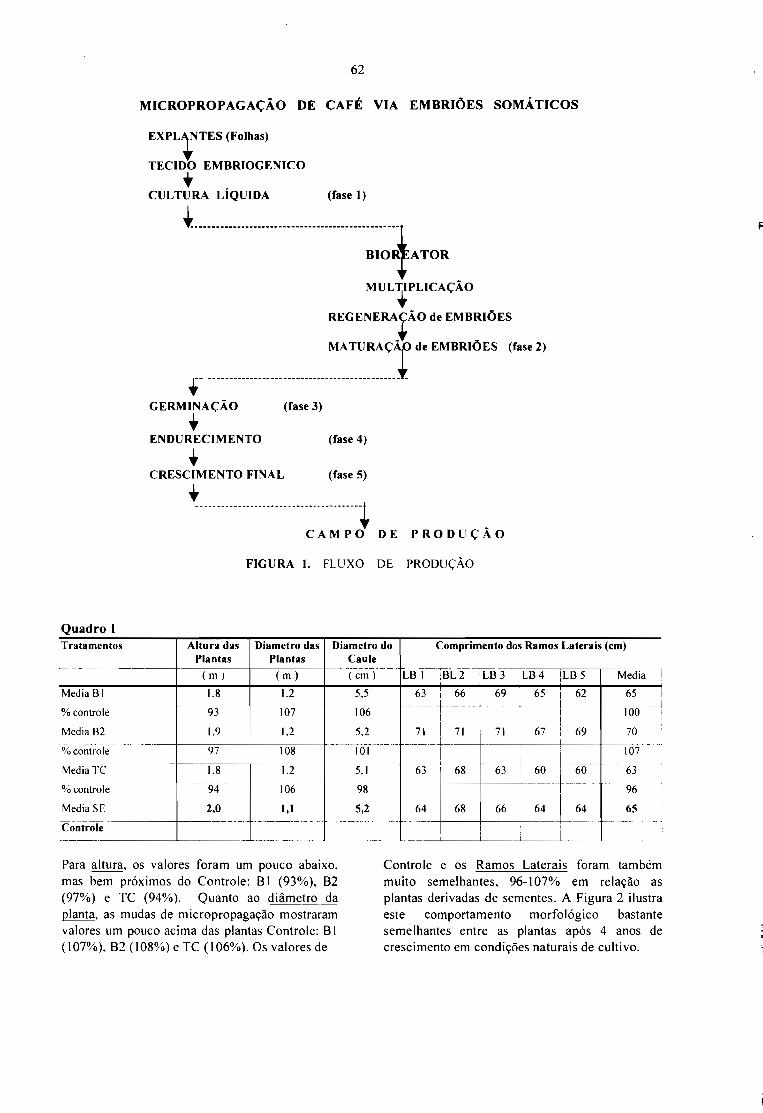
62
MICROPROPAGAÇÃO DE CAFÉ VIA EMBRIÕES SOMÁTICOS
EXPLrTES (Folhas)
TECIDO EMBRIOGENICO• •CULTURA LIQUIDA (fase 1)
*--------------- o 00 00 - ---- ---- --- --- - -- - o-o - -- - --lBIORfATOR
MUL+PLlCAÇÃO
REGENERA~ÃOde EMBRiÕES
MATURAÇ~Ode EMBRiÕES (fase 2)
._ _ 1:
GERMINAÇÃO (fase 3)..ENDURECIMENTO (fase 4)..CRESCIMENTO FINAL (fase 5)..----- -- ---------- --- ---- ------ ----- ----+
CAMPO DE PRODUÇÃO
FIGURA 1. FLUXO DE PRODUÇÃO
Quadro ITratamentos
Media BI
Media B2
Media TC
Altura das Diametro dasPlantas Plantas
(m) (m)
1,8 1.2
93 107
1.9 1.2
97 108
Comprimento dos Ramos Laterais (em)
Para altura, os valores foram um pouco abaixo,mas bem próximos do Controle: 8 I (93%), 82(97%) e TC (94%). Quanto ao diâmetro~
planta, as mudas de micropropagação mostraramvalores um pouco acima das plantas Controle: 8 I(107%),82 (108%) e TC (106%). Os valores de
Controle e os Ramos Laterais foram tambémmuito semelhantes, 96-107% em relação asplantas derivadas de sementes. A Figura 2 ilustraeste comportamento morfológico bastantesemelhantes entre as plantas após 4 anos decrescimento em condições naturais de cultivo.
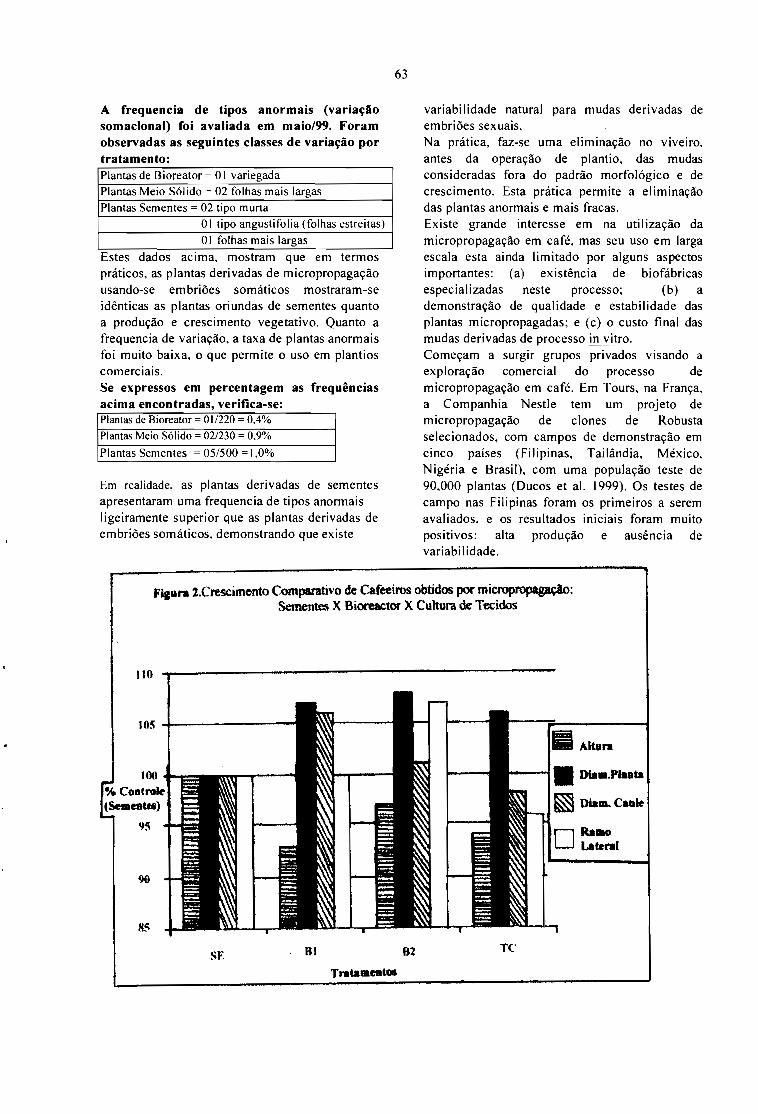
A frequencia de tipos anormais (variaçãosomaclonal) foi avaliada em maio/99. Foramobservadas as seguintes classes de variação portratamento:Plantas de Bioreator = OI variegadaPlantas Meio Sólido = 02 folhas mais largasPlantas Sementes = 02 tipo murta
01 tipo angustifolia (folhas estreitas)OI folhas mais largas
Estes dados acima, mostram que em termospráticos, as plantas derivadas de micropropagaçãousando-se embriões somáticos mostraram-seidênticas as plantas oriundas de sementes quantoa produção e crescimento vegetativo. Quanto afrequencia de variação. a taxa de plantas anormaisfoi muito baixa. o que permite o uso em plantioscomerciars.Se expressos em percentagem as frequênciasacima encon tradas, verifica-se:Plantas deBioreator - 01/220= 0.4%Plantas Meio Sólido=02/230 =0.9%
Plantas Sementes = 05/500 =1,0%
Em realidade. as plantas derivadas de sementesapresentaram uma frequencia de tipos anormaisligeiramente superior que as plantas derivadas deembriões somáticos. demonstrando que existe
63
variabilidade natural para mudas derivadas deembriões sexuais.Na prática, faz-se uma eliminação no viveiro.antes da operação de plantio, das mudasconsideradas fora do padrão morfológico e decrescimento. Esta prática permite a eliminaçãodas plantas anormais e mais fracas.Existe grande interesse em na utilização damicropropagação em café. mas seu uso em largaescala esta ainda limitado por alguns aspectosimportantes: (a) existência de biofábricasespecializadas neste processo; (b) ademonstração de qualidade e estabilidade dasplantas micropropagadas; e (c) o custo final dasmudas derivadas de processo i'l~itrº.
Começam a surgir grupos privados visando aexploração comercial do processo demicropropagação em café. Em Tours, na França.a Companhia Nestle tem um projeto demicropropagação de clones de Robustaselecionados, com campos de demonstração emcinco países (Filipinas, Tailândia, México.Nigéria e Brasil), com uma população teste de90,000 plantas (Ducos et aI. 1999). Os testes decampo nas Filipinas foram os primeiros a seremavaliados. e os resultados iniciais foram muitopositivos: alta produção e ausência devariabilidade.
Fi&lIra 2.Cre5Cimento Comparativo de Cafeeiros obtidos pormicropropagaçlo:Sementes X Bioreactor X Cultura de Tecidos
110
10~ 4---------'
. 81
Trata.ellt.
82
111 Altura--"'f. 1*•.Pla.ta
~ Dia•• Callk
Te

Neste Pais, estão sendo estabelecidos outroscampos de mudas clonadas de Robusta (total70,000), plantados em áreas comerciais de café,em ensaios comparativos com plantas derivadasde sementes e de enxertia. O custo de produçãodestas mudas de Robusta micropropagadas foideclarado ser de US$ 169 o milheiro (ou 16.9centavos de dólares a unidade).Neste mesmo trabalho, os autores declaram umcusto de produção de mudas de Robustaenxertadas igual a US$ 158 o milheiro (Ducos etaI. 1999).Considerando-se uma margem de lucro de 30%,estas plantas micropropagadas poderiam servendidas a US$ 220/milheiro, e as plantasenxertadas a US$ 205/milheiro.Com o aumento da produção, estes custos dasplantas micropropagadas devem baixar, pelaeconomia de escala e racionalização de etapas doprocesso dentro da Biofábrica, No Brasil, existeuma Biofábrica instalada em Ribeirão Preto, SP(BIONOVA) que tem especialistas emmicropropagação de café. Existem tambémlaboratórios em instituições governamentais depesquisa que estão iniciando projectos deprodução de mudas de café micropropagadas e areal ização de testes comparativos em condiçõesde campo.O Quadro 2 apresenta uma comparação de preçosde mudas de café oriundas de diferentesprocessos de propagação. O processo demultiplicação via sementes é o método maisbarato, porem este método não permite apropagação de plantas elites segregantes (ouhíbridos FI), e portanto restrito a plantashomozigotas (variedades). O processo desementes em tubettes permite uma maiortecnificação na qualidade das mudas, ganho de 46 semanas no tempo total de produção das mudas,e possibilidade de plantio mecanizado.O métodode propagação via Enxertia já vem sendo
Quadro 2
64
pratica em escala comercial, com preços queoscilam entre US$ 165-206/ milheiro (R$ 280 350). É um método economico e bastanteeficiente de propagação, mas requer instalações epessoal especializado. Para utilizar Robusta comoporta-enxerto de Arabica, existe um problema detempo de maturação de sementes: Arabica (7meses), Robusta (10 meses). Esta diferença leva auma redução do índice de germinação dassementes de Arabica e a dificuldade decrescimento das mudas enxertadas durante osmeses mais quentes do ano (novembro - março).A localização de viveiros em zonas elevadas, ou ainstalação de estufas c1imatizadas aumenta aeficiência de produção de mudas enxertadas.O método de micropropagação via GemasAxilares é lento, de baixo rendimento emnumeras, e com um preço final bastante elevado:US$ 471/milheiro (R$ 800). Este método somentese aplica para estabelecer jardins clonais para finsde pesquisa e manutenção de germoplasma.O método de micropropagação via EmbriogeneseSomática tem grande potencial de aplicaçãocomercial. podendo competir e até superar asvantagens da propagação por enxertia. O custo demudas de café via embriogenese somática esta nafaixa de US$ 176 - 220 / milheiro (R$ 300 374). A implantação deste sistema de propagaçãodepende da existência de biofábricasespecializadas e da existência de plantas elites ouhíbridos de alto valor comercial (em Arabica). Nocaso de Robusta, a simples seleção de plantasindividuais e a multiplicação destes clones trazuma vantagem produtiva imediata.Se compararmos o custo de uma planta propagadapelo processo de enxertia ou pelo processo deembriogenese somática, verificamos que ospreços de mercado são semelhantes: R$ 0,28 0,35 (enxerto) e R$ 0,30 - 0,37 (embrião). Sãoportanto processos de resultados e custosseme Ihantes.
Valor Estimado por milheiro de mudasrM~todos&P;:-opag~cao : US$ ! Reais $ (.)
L SEMENTES j~--------------------- '-----=-c l-----j.a) Saquinhos i 71 ; 120 Ir.:-;- ---- -------- -+- n_ u - 1---140----~
~~ EI~-~e~t~~IÁ -+ 82 1__ ---------------~
ia)-Tubettes-- -t--165-:zo6--i----28o -350---1·iMICRÕPROPAGA(:AO--i----------+------ ------ ----i
~em-a-s-axilare-s------------ ! 471----t----- 800-----i,---- - --- --- ----------- I --------+-----------------1bj Embriões somáticos 176-220 I 300-374 i-'--- ------ -- ---- I ------j- ----------...,
1(.) Va~o~l~S$= 1.7 __L .:. i

A preferencia por um ou outro método vaidepender das características de cada projeto, apresença (ou não) de instalações preexistentes, e adisponibilidade de pessoal especializado.No preço final de uma muda propagada deverátambém influir o valor intrínseco do materialgenético propagado e as condições de oferta eprocura. Muito importante nesta atividade, é acapacidade de entrega, em numeros e no prazo,das mudas encomendadas pelo usuário final,evitando-se assim prejuízos de execução deprodução em condições de campo.Considerando o exemplo acima de uma produçãoigual a 500,000 plantas/ano, o processo demicropropagação via embriões somáticos permiteum incremento muito rápido de plantasindividuais selecionadas. Este numero por seraumentado ou reduzido, dependendo do numero(e volume) dos bioreatores colocados emprodução. As vantagens da rapidez de propagaçãotraz a oportunidade de negócio para entrar emprodução mais cedo. Alem disto, há queconsiderar o valor intrínseco (genético) superiordo material que esta sendo multiplicado(híbridos), e portanto pelas leis de mercadojustifica-se um custo mais elevado (2 - 3x maior)das mudas micropropagadas, quando comparadascom o preço das mudas de sementes.A síntese de híbridos controlados vai trazer umanova dimensão para a produção de café nomundo, quer para regiões de Arabica ou deRobusta. A micropropagação via embriogenesesomática é o único método que vai poder atendera esta inovação tecnológica da cafeicultura dofuturo.
AG RADECI MENTOS
Os autores agradecem a contribuição financeirada illycaffe s.p.a.. (Trieste, Itália) e Kraft Foods(White Plains. USA) na realização destetrabalho.
65
LITERATURA
Berthouly, M. , Dufour, M., Alvard, D., Carasco,C, Teisson, C. 1995. Coffee Micropropagation.In 16th Intl. Conf. Coffee Science, Kyoto. ASIC.Paris. pp. 514-519.Custer, .I.B.M. 1980. Clonal propagation ofCoffea Arabica L. by nodal culture. In çth Intl.Coffee Science, London. ASIC, Paris, pp. 589596.Dublin. P. 1980. Multiplication vegetative in vitrode l'arabusta. Café Cacao Thé 24(4) 281-289.Ducos, .I.P., Gianforcaro, M., Florin, B.• Petiard,V. And Deshayes, A. 1999. Canaphora (Robusta)clones: in vitro somatic embryogenesis. In 18th
Intl. Conf. Coffee Science, Helsinki. ASIC , Paris.8 p. (in press)Kurtz, S.L., Hartman, R.D. and Chu, I.V.E. 1991.Current methods of commercialmicropropagation. l!1 Scale-up and Automation inPlant Propagation, I.K. Vasil (ed.), AcademicPress. New York, pp. 7-34.Noriega, C. and Sondahl, M.R. 1993. Arabicacoffee micropropagation through somaticembryogenesis via bioreactors. In 15th Intl.Conference Coffe Science, Montpellier. ASIC.Paris, pp. 73-81.Sondahl, M.R., Nakarnura, T., Medina, H.P.,Carvalho. A. Fazuoli, L.c. and Costa. W.M.1994. Coffee. !!1 Handhook ofPlant Cell Culture,Crop Species. Arnrnirato, P.V., Evans, D.A.,Sharp, W.R. & Yamada, V. (eds.), Macmillan.N.Vork. Vol.3, pp.564-590.Sondahl, M.R. and Sharp, W.R. 1977. Highfrequency induction of somatic embryos incultured leaf explants of Coffea Arabica L.Zeitschrift fur Ptlanzenphysiologie 81: 395-408.Staristsky, G. 1970. Embryoid formation in callustissues ofcoffee. Acta Bot. Neerl. 19(4): 509-514.Yasuda, T.. Fujii, V., Yarnaguchi, R. 1985.Embryogenic callus induction from Coffeaarabica leaf explants by benzyladenine. PlantCell. Physio1. 26(3): 595-597.Zarnarripa, A., Ducos. .I.P., Bollon, H., Dufour,M. Petiard, V. 1991. Production d' em bryonssomatiques de caféier en milieu liquide: effetsdensité d'inoculation et renouvellement dumilieu. Café Cacao Thé 34(4): 233-243.

'", liniento • Saúde • Esperança '"
MONSANTO
\
INDIVIDUALMENTE-ELAS SAüM

69
EFECTO DEL GENOTIPO Y ÉPOCA DE RECOLECCIÓN DEL MATERIAL VEGETAL EN LAEMBRIOGÉNESIS SOMÁTICA DlRECTA DE Coffea arabica VAR. CATURRA X HíBRIDO DETIMOR.
Molina V., D M.; Aponte A., M E.; Cortina G., H., Moreno R., L G.
Centro Nacional de Investigaciones de Café (Cenicafé), Chinchiná, Caldas, Colombia. AA. 2427 Manizales.Fax: (968) 504723. E-mail: fchcor@cafedecolombia.
INTRODUCCIÓN
La embriogénesis somática en café como en otroscultivos depende dei estado fisiológico deiexplante (Staristky 1970); tipo de hoja (Rodríguez1985); tiempo de subcultivo (Michaux-Ferriere etai, 1987); estado fisiológico de las plantas y épocadei ano en que se toman los explantes(Purushotham et ai. 1984; Kahia y Owour, 1990);las condiciones dei ambiente (Mroginsky y Roca.1991) y el genotipo (Bieysse et ai, 1993; Boxtel yBerthouly 1996; Berthouly y Michaux-Ferriere,1996).EI efecto dei genotipo en la embriogénesissomática ha sido ampliamente reportado en variasespecies como trigo (Mães et ai, 1996); melón(Yavad et ai. 1990); alfalfa (Chen et ai. 1987);maíz (Close y l.udernan, 1987) y en cítricos(Gmitter y Moore, 1986). En los géneros yespecies de herbáceos y plantas lefiosas algunosgenotipos son recalcitrantes y otros respondenbien a la embriogénesis (Gmitter y Moore, 1986;Chen et al, 1987). En cebada Hanzel et ai (1985).encontraron también diferencias significativas enel porcentaje
embriogénico de los genotipos, lo cual indica quela regeneración esta bajo control genético.EI efecto de la época de recolección dei materialvegetal ha sido reportado en café por Kahia yOwour (1990); Purushotam et ai (1984) Y Boxtely Berthouly et ai (1996); en arroz Ma et ai (1987)Y en avena Rines y McCoy (1981). Los objetivosde este trabajo fueron evaluar la respuesta deEmbriogénesis Somática Directa (ESD) dediferentes genotipos de café y establecer efecto dela época de recolección dei material vegetal en laESD y su interacción con el genotipo.
MATERIALES Y MÉTODOS
Genotipos. Se utilizaron explantes de 13 árboles(genotipos) pertenecientes a las generaciones F3.F4 Y F5 dei cruzamiento entre la variedad Caturray el Híbrido de Timor (Tabla 1).
Manejo de los explantes. Los explantes serecolectaron en diferentes meses de los anos 1993,1994 Y 1996. Se usaron hojas jóvenescompletamente desarrolladas, provenientes deiprimero o segundo par de ramas plagiotrópicas.
Genealogia de los genotipos regeneradosHJBRIDO FI F2 F3 F4 F5
H.3001 CA-CVI x H de T 1343 Ar.1322 PL.1402 A.0192 Nr.066 BK.045
BK.050
I I BK.620 IPr.227
;BI.698,
Sp.092 i BK.635
I
: PL.1372 A.0240 Rs.094 BG.90
IA.0241 Rs.585 BI.302
PL.1406 A.0041 Pr.643 BI.390
Ar.1324 PL.IOOO A.0219
Lb.499 BG.060
Nr.671 BI.720
A.0404 Rs.460 i BK.500
Tabla I.

La desinfección de las hojas se realizó conhipoclorito de sodio comercial (3 y 5%) y tween80. Se tomaron ex plantes de aproximadamente Iem' de la porción media y basal de la láminafoliar, y se colocaron en contacto con el medioprimario (Orozco et ai 1989), en platos de petri a25°C ± 2, en oscuridad.
Diseõo experimental. Se sembraron 24experimentos, en cada experimento se incluyeronde 2 a 4 genotipos. En todos los experimentos seutilizó un disefio completamente aI azar, con 15repeticiones, tomando como unidad experimentaluna caja de petri con 10 explantes. Las variablesevaluadas fueron los tipos de respuestaernbriogénica, así: Embriogénesis somática debaja frecuencia (ESBF) cuando se producen de I a10 embriones por explante, Embriogénesissomática de alta frecuencia (ESAF) en este casose produce tejido embriogénico que da origen aunalto número de embriones por explante yEmbriogénesis somática total (EST), es lasumatoria de la ESBF y la ESBF.
Análisis de la información. Se hizo un análisisde varianza no-ortogonal; como fuente devariación se consideraron: genótipos, meses y suinteracción. Cuando hubo diferencias entre losgenotipos se hicieron comparaciones entre gruposde genotipos, de acuerdo con su genealogía;considerando que estas diferencias tienen unabase genética. Cuando la diferencia fue entremeses se calculó la diferencia mínimasignificativa (DMS). Finalmente, los genotiposfueron separados según su capacidadembriogénica, así: a) Baja. Los genotipos conlimites de confianza inferior (LCI de O a 20; b)Media. Los genotipos con LCI entre 21 a 50 y c)Alta. Los genotipos con LCI mayores de 51 %.
RESULTADOS
Efecto deI genotipo. EI porcentaje embriogénicode los genotipos regenerados estuvo entre 12 a72.7% (Tabla 2), lo que indica que hay granvariación en los genotipos estudiados. Capacidadembriogénica diferente reportó Aponte (1993) endos genotipos dei mismo origen. AdemásMichaux-Ferriere et ai (1989), compararon laevolución histológica de tres genotiposembriogénicos de C. arabica (KF2.1, KF.2.6 Y
70
ET25), Y de uno recalcitrante (ET20.1),encontrando que las diferencias embriogénicasentre genotipos se deben a la interacción deigenotipo y las condiciones fisicoquímicas delcultivo.Seis genotipos fueron clasificados de capacidadembriogénica alta, con excepción dei A.219 todostuvieron un coeficiente de variación menor a38.7% y la desviación estándar fue baja. Adernás,el LCI estuvo entre 50 y 71%, lo que significa quela capacidad de regeneración de embriones y/oTejido embriogénico es repetible. La figura Ipresenta la respuesta embriogénica dei genotipoBK.620 con capacidad embriogénica alta. Losgenotipos que fueron clasificados como decapacidad media y baja tuvieron un coeficiente devariación entre 45 y 86% y el LCI fue menor de42.2%; esto indica que el porcentaje deregeneración no es aceptable. La figura 2 muestrala capacidad embriogénica baja dei genotipoBG.60.AI realizar las comparaciones sefíaladas en latabla 3, se observaron diferencias altamentesignificativas entre las progenies desarrolladas apartir dei Ar.1322 y la dei Ar. 1324, siendo másembriogénicas las descendi entes dei primero.Entre las tres progenies descendi entes dei Ar.1322(PL.1372, PL.1402 Y PL.1406), la PL.1402 fuesignificativamente más embriogénica y sudescendiente el A.0192 dia origen a tres progenies(Nr.66, Pr.227 y Sp.092), siendo las descendientesde la Nr.66 significativamente más embriogénicasy no hubo diferencias entre ellas (BK.50. BK.45 YBK.620); según la escala de clasificación estosgenotipos son de capacidad embriogénica alta.Estos resultados muestran que las di ferencias en lacapacidad embriogénica estuvieron relacionadascon la generación, observándose segregaciónhasta la F3, que disminuyó en las generacionessiguientes y los materiales F4 derivados de unmismo progenitor produjeron respuesta simi lar.indicativo de que la capacidad embriogénicadepende de factores genéticos. Resultadossimi lares reportó Duncan et aI (1985). ai estudiarvarias líneas encontró que las progenies deparentales embriogénicos eran tambiénembriogénicas; esto indica que la proliferación deplantas en maíz esta genéticamente determinada;lo cual tiene utilidad para predecir la capacidad deregeneración de una línea sin probar, por larespuesta de su parenta!.
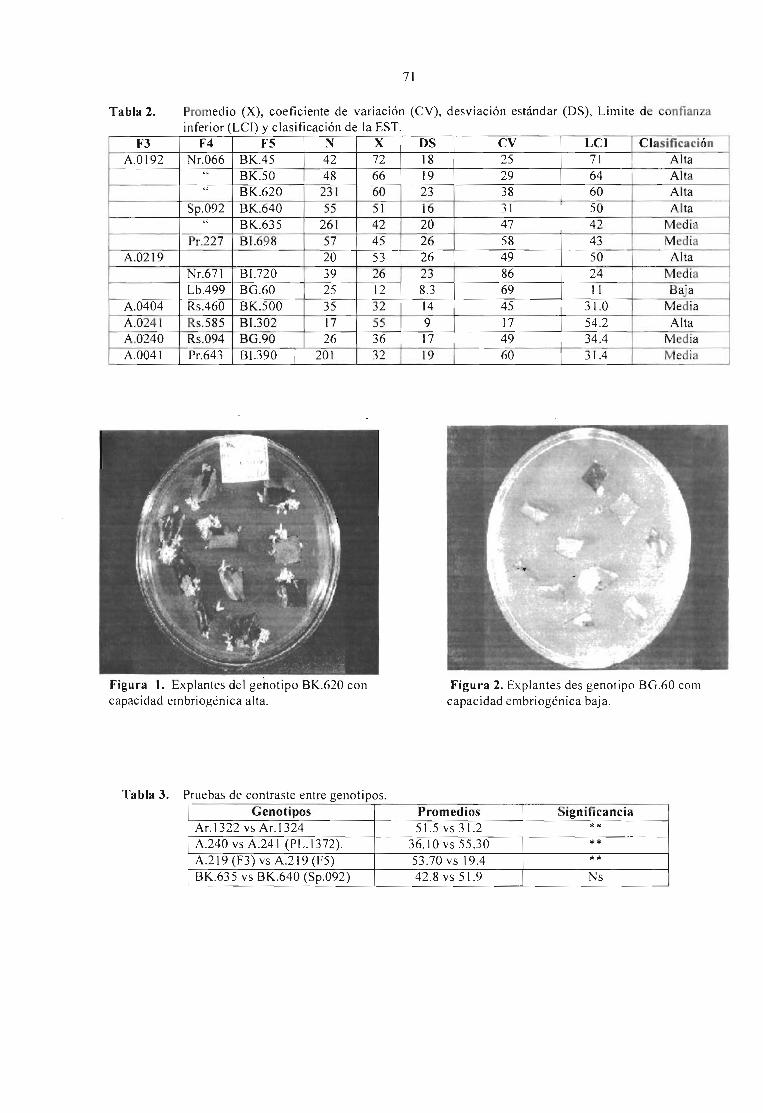
Tabla 2.
71
Prom edio (X), coeficiente de variación (CY), desviación est ándar (DS), Limite de co nfianzainferior (LC[) y clasificación de la EST.
F3 F4 F5 N X DS CV LCI Cl asificaciónA.0192 Nr.066 BK.45 42 72 18 25 71 Alta
" BK.50 48 66 19 29 64 Alta" BK.620 231 60 23 38 60 Alta
Sp.092 BK.640 55 5 1 [6 3 1 50 Alta" BK.635 261 42 20 47 42 Media
Pr.227 BI.6 98 57 45 26 58 43 MediaA.0219 20 53 26 49 50 Alta
Nr.671 BI.720 39 26 23 86 24 MediaLb.499 BG.60 25 [2 8.3 69 11 Baja
A.0404 Rs.460 BK.500 35 32 14 45 31.0 MediaA.024 [ Rs.585 BI.302 17 55 9 17 54 .2 AltaA.0240 Rs.094 BG.90 26 36 17 49 34.4 MediaA.OQ41 Pr.643 BI.390 I 201 32 19 60 31.4 Media
Figura I. Explantes dei genotipo BK.620 concapacidad embriogénica alta.
Tabla 3. Pruebas de contraste entre genotipos.
Figura 2. Explantes des genotipo BG .60 comcapacidad embriogénica baj a.
Genotipos Promedios SignificanciaAr.1322 vs Ar.1324 51.5 vs 31.2 **A.240 vs A.241 (PL.1372). 36.10 vs 55.30 **A.219 (F3) vs A.219 (F5) 53.70 vs 19.4 **BK.635 vs BK.640 (Sp .092) 42.8 vs 51.9 Ns
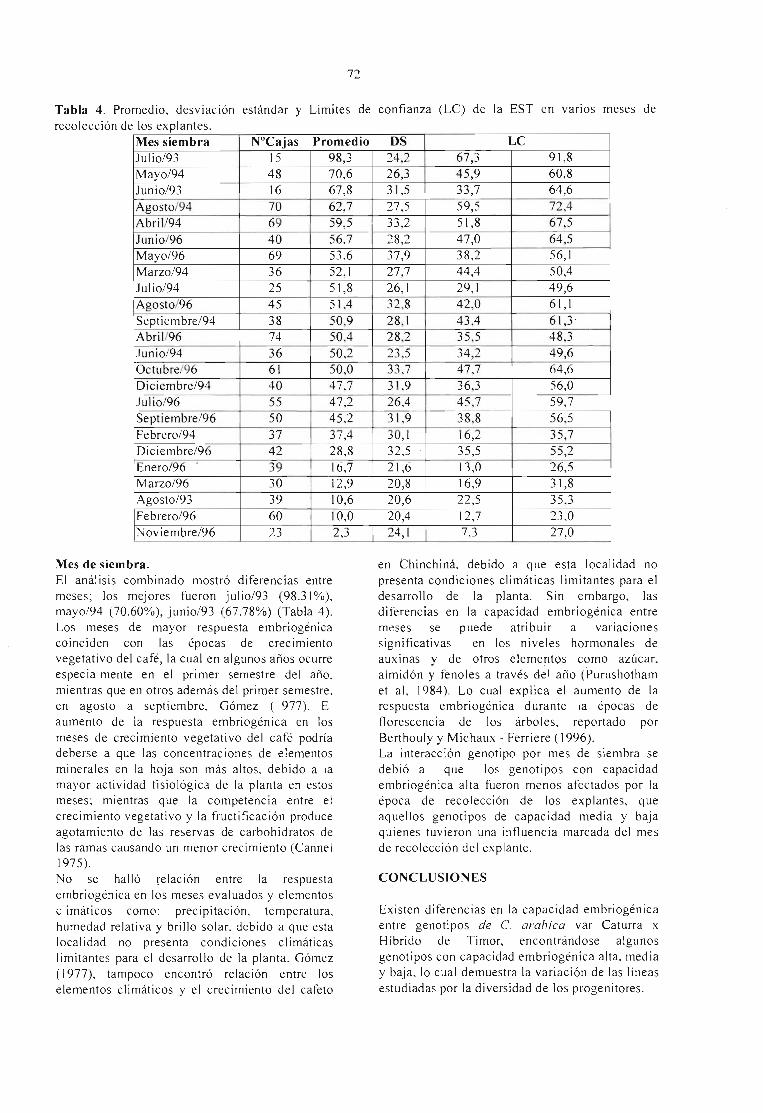
72
Tabla 4. Prorned io. de sviaci ón estándar y Limites de confianza (LC) de la EST en va rios me se s derecol ección de los exp lantes.
Mes siembra N°Cajas Promedio DS LCJulio/93 15 98,3 24.2 67,3 91,8
Mayo/94 48 70,6 26,3 45,9 60 ,8Juni o/93 16 67 ,8 31,5 33,7 64,6
Agosto /94 70 62,7 27.5 59,5 72, 4
Abri l/94 69 59,5 33.2 51,8 67,5
Junio/96 40 56,7 28,2 47,0 64 .5Ma yo/96 69 53 ,6 37 ,9 38,2 56, I
Marzo/94 36 52, I 27,7 44,4 50 ,4
Julio/94 25 5 1,8 26, 1 29, 1 49 ,6
Ago sto/96 4 5 5 1.4 32 .8 42,0 61,1
Septiembre/94 38 50 .9 28 .1 43 .4 6 1.3 'Abr il/96 74 50.4 28.2 35 .5 48 .3Junio/94 36 50 .2 23,5 34,2 49,6
--
Octubre/96 61 50, 0 33,7 47. 7 64,6Dic iembre/94 40 47,7 3 1.9 36,3 56 .0Julio/96 55 47.2 26,4 4 5,7 59.7Septiembre/9 6 50 45 .2 3 1.9 38.8 56.5Fe bre ro/94 37 37,4 30 , 1 16,2 35,7Dici embre/9 6 42 28, 8 32 ,5 35,5 55,2Enero/96 39 16,7 2 1,6 13,0 26,5Mar zo/96 30 12,9 20,8 16.9 31,8Agosto/93 39 10,6 20,6 22,5 35,3Febrero /96 60 10,0 20,4 12.7 23,0Nov iembre/96 23 2,3 24,1 7,3 27,0
Mes de siembra.EI análisi s combinado mostró dife re nc ias ent remeses; los mejores fueron ju lio/93 (98 .3 1%),mayo/94 (70.60%), junio/93 (67 .78%) (Ta bla 4).Los meses de ma yor respuesta embriogéni cacoinciden con las épocas de c rec imientovegetativo dei café, la cual en al gun os an os oc urreespecialmente en el primer seme st re dei ano.mientras que en otros además dei prirner semestre.en agosto a septiernbre, Gó mez (1977). EIaumento de la respuesta embriogén ica en losmeses de crecimiento vegetativo dei café podríadeberse a que las concentraci ones de e leme ntosminerales en la hoja son más a ltos . de bido a lamayor activ idad fisio lógica de la planta en esta sme se s; mientras que la co mpete ncia entre elcrec imiento vegeta tivo y la fruc tificación produceagota miento de las reservas de ca rbohid ratos delas ramas causa ndo un men or creci mie nto (Ca nne l197 5).No se halló re lac i ón entre la respu estae mbriogénica en los meses eva luados y e lementosc limá ticos co mo : precipi taci ón, tem peratura.hum ed ad relativ a y brill o so la r. de bido a qu e estalocal idad no pre senta co ndic iones cl imáticaslimitantes para el de sarroll o de la pla nta . Gó mez( 1977). tampoco en con tró relación entre lose lementos cl imáticos y el crecimi ent o dei cafeto
en Chinchiná, debido a que esta localidad nopresenta condiciones climáticas limitantes para e ldesarrollo de la planta. Sin embargo. Iasdiferencias en la capacidad embriogénica entremeses se puede atribuir a variacionessignificativas en los niveles hormonales deaux inas y de otros elementos como az úcar ,a lmidó n y fen oles a través dei ano ( Purusho tha met al. 1984) . Lo cual expl ica el aumento de lares pues ta embriogénica duran te la épocas deflo resce ncia de los árbo les, repo rtado porBerthou ly y M ich au x - Ferriere ( 1996) .La inte racció n ge no tipo por me s de siem bra sede bió a qu e los geno tipos con ca pac ida demb riogénica a lta fueron menos a fec tados po r laépoca de recol ección de los exp lantes, queaque llos ge no ti pos de ca pac ida d media y baj aqu ien es tuv ie ro n una influenc ia marcad a dei mesde recol ecc ión de i expl ante.
CO NCLUSIONES
Existe n d iferenci as en la capacidad embriogénicaen tre ge no tipos de C arabica var Caturra xHíbrido de Tirnor, encontrándose algunosge noti pos co n capacidad embriogénica alta. mediay baj a. lo c ua l demuestra la variación de las líneasestu diad as por la diversidad de los progenitores.

Las diferencias en la capacidad embriogénica delos genotipos esta relacionada con el origengenético dei material, observándose que hasta lageneración F3 se produce segregación deimaterial, mientras que los genotipos derivados deun misma progenie F4 producen una respuestasimilar, indicando que la F4 da lugar a unasegregación muy baja en su descendencia y la F5se comporta similar a la F4.Hay diferencias en la respuesta embriogénicadependiendo de la época dei afio en que se tomenlos explantes y la interacción genotipo por mes desiembra se debe a que los genotipos concapacidad embriogénica alta fueron menosafectados por la época de recolección de losexplantes, con respecto de los genotipos concapacidad media y baja.No se encontró relación entre la respuestaembriogénica en los meses evaluados y elementosclimáticos ya que esta localidad no presentacondiciones climáticas limitantes para eldesarrollo de la planta.
BIBLIOGRAFIA
Aponte de L, M.E. 1993. Somatic embryogenesisinduced by cultured on single media in coffeeplants from crosses ofCoffea arabica x Híbrido deTimor. In: COLLOQUE ScientifiqueInternational sur le Café, 15. Montpellier(Francia), Juin 6-11. Paris (Francia), ASIC, pp 8288.Berthouly, M., N.M. Michaux-Ferriere. 1996.High frequency somatic embryogenesis in CofJeacanephora. Plant ce11 tissue and organ culture. 44:169-176.Bieysse, O., A. Goftlot, N.M. Michaux-Ferriere.1993. Effect of experimental conditions andgenotypic variability on somatic embryogenesis inCoffea arabica. Canadian Journal Botanic. 71:1496-1502.Boxtel, J.V .. M. Berthouly. 1996. High frequencysomatic embryogenesis from coffee leaves. PlantCell Tissue and Organ Culture (Holland) 44:7-17.Cannell,M.G.R. 1975. Crop physiological aspectsof coffee bean yield: A review. Journal of CoffeeResearch (India) 5 (1-2): 7-20.Close, K.R, and L.A. Ludeman. 1987. The effectof auxin-like plant growth regulators and osmoticregulation on induction of somatic embryogenesisfrom elite maize inbreds. Plant Science 52: 81-89.Chen, T.H.N., 1. Marowitch, and B.G. Thompson.1987. Genotypic effects on somaticembryogenesis and plant regeneration from calluscultures of alfalfa. Plant Cell Tissue and OrganCulture (Holland) 8: 73-81.
73
Duncan, O.R., M.E. Williams, B.E. Zehr, 1.M.Widholm. 1985. The production of callus capableof plant regeneration from immature embryos ofnumerous Zea mays (L.) genotypes. Planta. 165:322-332.Gmitter, F.G., G.A. Moore. 1986. Plantregeneration from undeveloped ovules andembryogenic calli of Citrus: Embryo production,gerrnination, and plant survival. Plant Cell Tissueand Organ Culture (Holland) 6: 139-147.Gómez, G.L. 1977.1nfluencia de los factoresclimáticos sobre la periodicidad de crecimientodei cafeto. Cenicafé (Colombia) 28 (I ):3-17.Hanzel, U., 1.P. Miller. M.A. Brinkman, E.Fendos. 1985. Genotype and media effects oncallus formation and regeneration in barley. CropScience.25.27-31.Kahia, J.W., 1.B.O. Owuor. 1990. In vitropropagation of the disease resistant Coffeaarabica L cultivar-Riuru 11. Kenya Coffee(Kenya) 55(646): 901-905.Ma, H., M. Gu, G.H. Liang. 1987. Plantregeneration from culture immature embryos ofSorghum bicolor (L.) Moench. Theor. Appl.Genet. 73: 389-394.Maês, O.c.. R.N. Chibbar, K. Caswell, N. Leung,K.K. Kartha. 1996. Somatic embryogenesis fromisolated scutella of wheat: effects of physical,physiological and genetic factors. Plant science121: 75-84.Michaux-Ferriere, N, P. Dublin. 1987.Embryogenese somatique chez Coffea arabicainduction et developpement dex cellulesembryogénes. In: COLLOQUE ScientifiqueInternational sur le Café, 12. Montreaux (Suiza).Juin 29-Juillet 3, 1987. Paris (Francia),Association Scientifique International du CaféAsic, pp 418-425.Michaux-Ferriere, N., O. Byesse, D. Alvard, P.Dublin. 1989. Etude histologique deI' embryogenese somatique chez Coffea arábicainduite par culture sur milieux uniques defragments foliaires de génotypes différents. CaféCacao Thé (Francia) 33 (4): 207-217.Mroginski, L.A.. W.M. Roca. 1991.Establecimiento de cultivo de tejidos vegetales invitro. In ROCA. W:M.; MROGINSKI, LACultivo de tejidos en la agricultura: fundamentosy aplicaciones. Cali (Colombia). Centro.Internacional de Agricultura Tropical. pp. 19-41.Orozco, F.J., t.c. Londofío, M.T. González, M.L.Marulanda, B.E. Velez. 1989. Aplicaciones delcultivo de tejidos en el mejoramiento deI cafeto.In: COLLOQUE Scientifique International sur lecafé. 13. Paipa (Colombia) 21-25 de Agosto 1989.Paris (Francia), Association ScientifiqueInternational du Café - ASIC, pp 749-758.

Purushotham, K., U.V. Sulladmath, P.K.Ramalah. 1984. Seasonal changes in biochemicalconstituents and their relation to rooting of(Coffea canephora Pierre) sucker cuttings. JoumalofCoffee Research (India) 14(3): 117-130.Rines, H.W., T.J. Mccoy. 1981. Tissue cultureinitiation and plant regeneration in hexaploidspecies of oats. Crop Science. 21: 837-842.Rodriguez, R.H. 1985. Obtención de embrionessomáticos in vitro de las variedades comercialesde café: Variedad Caturra y Variedad Colombia.Bogotá (Colombia), Universidad Javeriana, 91p.(Tesis Biólogo).
74
Staritsky, G. 1970. Embroid formation in callustissues of Coffea. Acta Botanica Neerlandica(Holland). 19 (4) 509-514.Yavad,R.C., M.T. Saleh,R. Grumet. 1996.Highfrequency shoot regenerationfrom leafexplants ofmuskmelon. Plant Cell Tissue and Organ Culture(Holland)45: 207-214.

75
DETERMINAÇÃO DO TAMANHO DO BOTÃO FLORAL PARA A CULTURA DE ANTERAS NOCAFEEIRO (Cof/ea arabica L.)
Luciana Marques da Cunha Andrade.André Barretto Pereira", Edilene Carvalho Santos", MoacirPasqual"
'Bióloga; 2Pesquisador CEPEC/CEPLAC, Pós-graduação/UFLA; 'Graduanda Agronomia; "Prof. TitularDAG/UFLA, Caixa Postal 37, CEP 372000-000, Lavras-MG.
•
INTRODUÇÃO
Por ser uma cultura perene, os programas demelhoramento do cafeeiro demandam muitotempo. A cultura de anteras é considerada umaferramenta importante no melhoramento, poispossibilita a obtenção rápida de linhashomozigotas. Segundo Moraes Fernandes (1990),a obtenção de plantas haplóides seguida peladuplicação dos cromossomos através da utilizaçãode colchicina resulta em linhas hornozigóticas emcurto espaço de tempo, se comparada com osmétodos convencionais. Além da redução detempo, esta técnica pode ser utilizada em estudosgenéticos, pela obtenção de recessivos e induçãode mutações. A cultura de anteras para a obtençãode plantas homozigotas em gerações segregantesacelera o processo de geração de novos cultivaresem vários anos, além de facilitar a análisegenética, eliminando a complexidade do estadoheterozigoto. As plantas haplóides, possuidoras dametade do número de cromossomos característicoda espécie, ficam livres dos problemas dedominância e recessividade por possuirem apenasum alelo em cada loco gênico. permitindo a rápidaprodução de plantas homozigotas através daduplicação dos cromossomos numa única etapa,substituindo as muitas gerações deautofecundação necessárias no processo usual deobtenção de linhagens.Para a prática de cultura de anteras, é necessárioque se determine o estádio ideal dedesenvolvimento das mesmas, sendo que o estádioutilizado deve conter micrósporos que respondammelhor aos processos androgenéticos. Inúmerostrabalhos indicam que a melhor fase é aquela emque o micrósporo está recém liberado da tétrademeiótica até, no máximo, o estádio binucleado.Nesta fase, o micrósporo ainda possuicaracterísticas esporofiticas que permitem adiferenciação do grão de pólen em embrióides "invitro" (Moraes Fernandes, 1990; Sibi et al., 1979;Luz, 1995; Raghuramulu, 1989).As células "in vitro" são capazes de originarembriões através da ernbriogênese. quandosubmetidas à condições adequadas. antes daprimeira mitose. Após esse período ocorre ainativação dos RNAs ribossômicos e de
transferência e o desenvolvimento segue ocaminho normal para a formação do grão de pólen(Mascarenhas. 1971; Vasil et ai. 1979). O objetivodeste trabalho foi o de relacionar os estádios dedesenvolvimento dos micrósporos com ascaracterísiticas morfológicas do botão floral.determinando o tamanho do botão floral ideal paraobtenção de di-haplóides através da cultura deanteras de cafeeiro.
MATERIAL E MÉTODOS
O presente estudo foi conduzido no Laboratóriode Citologia do Departamento de Biologia daUniversidade Federal de Lavras (UFLA).Foram utilizados 4 cultivares de Coffea arábicaL.: Mundo Novo LCP-37919; Catuaí LCH-20772-5- 44; Rubi MG-1192 e lcatú LC-3282 ..pertencentes ao Banco de Germoplasma daUniversidade Federal de Lavras.Aproximadamente 200 botões florais foramcoletados no período de 8 à 9 horas da manhã, emseguida foram fixados em álcool etílico e ácidoacético na proporção de 3: I durante 24 horas,sendo posteriormente armazenados em álcool70% e mantidos a uma temperatura de 4°C.Os botões florais foram separados de acordo como seu comprimento. em cinco classes distintas, deacordo com os valores mínimos, máximos emédias. a saber: classe I (1,5; 1,72; 2,0 rnrn):classe 2 (2,5; 2,98; 3,5 mm); classe 3 (4,0; 4.48:5,0 rnrn): classe 4 (4.5; 530; 6,0 mrn); classe 5(6,5; 7,20; 8,0 mrn), as quais foramcorrelacionadas com o comprimento do botãofloral, comprimento da antera e as fases dedesenvolvimento do micrósporo.Foi utilizado um paquímetro para a medição docomprimento dos botões florais, bem como docomprimento de suas anteras. Foram analisados 3botões de cada classe para todos os genóti posoO estádio de desenvolvimento dos micrósporosfoi avaliado pela confecção de duas lâminas porbotão, sendo duas anteras por lâmina. cujosmicrósporos foram corados com carminpropiônico I%. Determinou-se a contagem demicrósporos nas fases de tétrade, recém liberadode tétrade, núcleo central. núcleo deslocado e
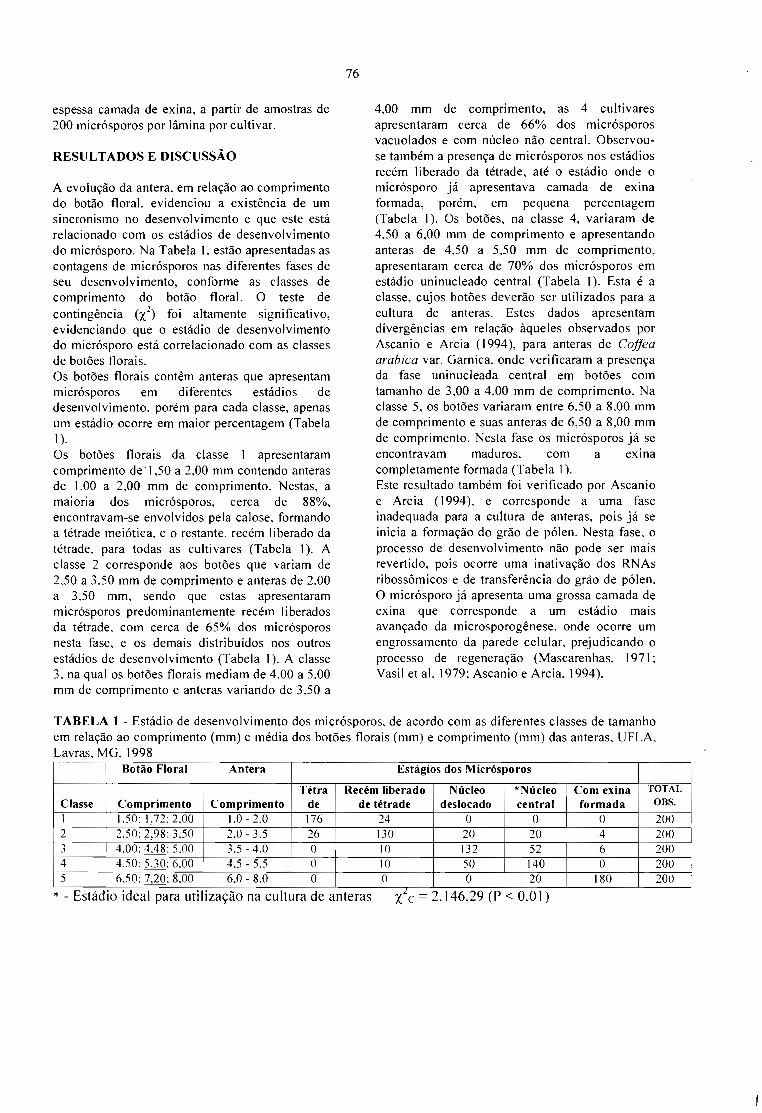
76
espessa camada de exina, a partir de amostras de200 micrósporos por lâmina por cultivar.
RESULTADOS E DISCUSSÃO
A evolução da antera, em relação ao comprimentodo botão floral, evidenciou a existência de umsincronismo no desenvolvimento e que este estárelacionado com os estádios de desenvolvimentodo micrósporo. Na Tabela I, estão apresentadas ascontagens de micrósporos nas diferentes fases deseu desenvolvimento, conforme as classes decomprimento do botão floral. O teste decontingência (l) foi altamente significativo,evidenciando que o estádio de desenvolvimentodo micrósporo está correlacionado com as classesde botões florais.Os botões florais contêm anteras que apresentammicrósporos em diferentes estádios dedesenvolvimento, porém para cada classe, apenasum estádio ocorre em maior percentagem (TabelaI).Os botões florais da classe I apresentaramcomprimento de' 1,50 a 2,00 mm contendo anterasde 1,00 a 2,00 mm de comprimento. Nestas, amaioria dos micrósporos, cerca de 88%,encontravam-se envolvidos pela calose, formandoa tétrade meiótica, e o restante, recém liberado datétrade, para todas as cultivares (Tabela I). Aclasse 2 corresponde aos botões que variam de2,50 a 3,50 mm de comprimento e anteras de 2.00a 3,50 mrn, sendo que estas apresentarammicrósporos predominantemente recém liberadosda tétrade, com cerca de 65% dos micrósporosnesta fase, e os demais distribuídos nos outrosestádios de desenvolvimento (Tabela I). A classe3, na qual os botões florais mediam de 4,00 a 5,00mm de comprimento e anteras variando de 3,50 a
4,00 mm de comprimento, as 4 cultivaresapresentaram cerca de 66% dos micrósporosvacuolados e com núcleo não central. Observouse também a presença de micrósporos nos estádiosrecém liberado da tétrade, até o estádio onde omicrósporo já apresentava camada de exinaformada, porém, em pequena percentagem(Tabela I). Os botões, na classe 4, variaram de4,50 a 6,00 mm de comprimento e apresentandoanteras de 4,50 a 5,50 mm de comprimento,apresentaram cerca de 70% dos micrósporos emestádio uninucleado central (Tabela I). Esta é aclasse, cujos botões deverão ser utilizados para acultura de anteras. Estes dados apresentamdivergências em relação àqueles observados porAscanio e Areia (1994), para anteras de CofJeaarabica varo Garnica, onde verificaram a presençada fase uninucleada central em botões comtamanho de 3,00 a 4,00 mm de comprimento. Naclasse 5, os botões variaram entre 6,50 a 8,00 mmde comprimento e suas anteras de 6,50 a 8,00 mmde comprimento. Nesta fase os micrósporos já seencontravam maduros, com a exinacompletamente formada (Tabela I).Este resultado também foi verificado por Ascanioe Are ia (1994), e corresponde a uma faseinadequada para a cultura de anteras, pois já seinicia a formação do grão de pólen. Nesta fase, oprocesso de desenvolvimento não pode ser maisrevertido, pois ocorre uma inativação dos RNAsribossômicos e de transferência do grão de pólen.O micrósporo já apresenta uma grossa camada deexina que corresponde a um estádio maisavançado da microsporogênese, onde ocorre umengrossamento da parede celular, prejudicando oprocesso de regeneração (Mascarenhas, 1971;Vasil et al, 1979; Ascanio e Areia, 1994).
TABELA 1 - Estádio de desenvolvimento dos rnicrósporos, de acordo com as diferentes classes de tamanhoem relação ao comprimento (mm) e média dos botões florais (mm) e comprimento (mm) das anteras, UFLA,Lavras, MG, 1998
x c = 2.146,29 (P < 0,01)* - Estádio Ideal para utilização na cultura de anteras
Botão Floral Antera Estágios dos Micrósporos
Tétra Recém liberado Núcleo *Núcleo Com exina TOTAL
Classe Comprimento Comprimento de de tétrade deslocado central formada OBS.
1 1.50: 1.72: 2.00 1.0 - 2.0 176 24 O O O 2002 2.50: 2.9!!: 3.50 2.0 - 3.5 26 130 20 20 4 200 i3 4.00: 4.48: 5.00 3.5 - 4.0 O lO 132 52 6 2004 4.50: 5.30: 6.00 4.5 - 5.5 O \0 50 140 O 200 15 6.50: 7.20: 8.00 6.0 - 8.0 O O O 20 180 200
.. - 1

CONCLUSÃO
Nas 4 cultivares de C.arabica avaliadas,verificou-se que a maior percentagem demicrósporos apresentando um núcleo central,ideal para a obtenção de plantas haplóides "invitro", encontra-se em anteras que variam de 4,5 a5,5 mm de comprimento, presentes em botõesflorais que variam de 4,5 a 6,0 mm decomprimento.
77
REFERÊNCIAS BIBLIOGRÁFICAS
ASCANIO, E.C.E; ARCIA, MAM. Efecto deiestado de desarrollo de las anteras y de un shocktérmico sobre la androgénesis en Coffea arábicaL. Varo Garnica. Café Cacao Thé, v:28, n.2 p.7579. 1994.LUZ, J.M.Q. Embriogênese somática "in vitro"em anteras de Pimentão (Capsicum annum L.)MASCARENHAS, lP. RNA and proteinsynthesis during pollen development and tubegrowth. In: Heslop-HARRISSON, J.. ed. Pollendevelloprnent and physiology. London,Butterworth, p.20 1-222, 1971


79
INFLlIÊNCIA DOS REGlILADORES DE CRESCIMENTO NA ClILTlIRA DE EMBRiÕES "INVITRO" DO ClILTIVAR CATlIAí VERMELHO LCH 2077-2-5-44.
Luciana Marques da Cunha Andrade', André Barretto Pereira', JRonilda Juliana de Campos,4Moacir Pasqual
'Biólogar- 2Pesquisador CEPEClCEPLAC. Pós-graduação/UFLA;- 'Graduanda Agronorniac- 4Pro f. TitularDAG/UFLA. Caixa Postal 37. CEP 372000-000. Lavras-MG.
INTRODUÇÃO
A cultura de embriões "in vitro" é uma técnicaimportante pois. tanto possibilita o estudo maisdetalhado dos problemas nutricionais e fisiológicosdo embrião. como também promovem odesenvolvimento de híbridos interespecíficos quenormalmente não sobrevivem; permite realizar aclonagem de plantas selecionadas com o objetivo deantecipar a época de plantio, e também obter plantasa partir de embriões imaturos. ou seja. em estágioprecoce do fruto.O embrião originado de um processo normal defecundação pode ser facilmente separado ecultivado em condições assépticas em meio decultura adequado. Este mantem-se geneticamenteestável. produzindo descendentes idênticos a ele.Para a remoção do embrião. basta desinfetar apenasa superficie externa da semente. devido ao fato deque o embrião esta alojado em região estéri I dasemente. Assim o índice de contaminação "invitroé muito baixo em relação às demais culturas(1IIg. 1985).O meio de cultura adequado. tanto para propagação.quanto para a cultura de embriões deve ser adaptadopara cada espécie. Os primeiros estudos envolvendomeio de cultura para embriões usaram amplamentea solução de Snop (Andreoli, 1985). Emboradiferentes meios sejam capazes de manter osmicrocultivos de embriões. o mais freqüentementeutilizado é o "MS" (Murashige e Skoog, 1962).embora outros meios tenham mostrado grandeeficiência.O cafeeiro. como outras culturas. libera substânciasfenólicas no meio de cultura e por esta razão devese adicionar ao meio de cultura substânciasantioxidantes, tais como: carvão ativado.Polivinilpirrolidone (PVP). ácido ascórbico e ácidocítrico.O objetivo deste trabalho foi determinar asmelhores concentrações dos reguladores decrescimento ANA (ácido naftalenoacético) e BAP(6-benzilaminopurina). utilizados na cultura deembriões "in vitro" da cultivar Catuaí vermelhoLCH 1077-2-5-44.
MATERIAL E MÉTODOS
Os experimentos foram realizados no Laboratóriode Cultura de Tecidos Vegetais do Departamento deAgricultura da Universidade Federal de Lavras UFLA. em Lavras - Minas Gerais.O meio nutritivo básico utilizado foi o "MS"(Murashige e Skoog. 1962). acrescido de 6mg/1 deGA1 sendo que o pH foi ajustado para 5.9. Para asolidificação do meio foi utilizado ágar naproporção de 7g/1.Após o preparo. foram distribuídos cerca de 10 mlde meio por tubo de ensaio (1.5 x 15 em ). os quaisforam vedados com tampa de polipropileno. O meiofoi esterilizado utilizando-se autoclavagem àtemperatura de 121°C. durante 20 minutos e I atmde pressão.O processo de inoculação foi realizadoassepticamente em câmaras de fluxo laminarhorizontal. Após o processo de inoculação. omaterial foi transferido para sala de crescimento
com temperatura igual a 26 ± 1°C. fotoperíodo de16 horas e intensidade luminosa de 2500 lux,suprida por lâmpadas Grow-Iux e Branca-fria. naproporção de 1: I. permanecendo nestas condiçõespor um período de 60 dias. Em todos osexperimentos foram realizadas avaliações visuaisnos primeiros 30 dias após a instalação. e foramavaliadas as variáveis número total de brotos.número total de folhas. comprimento dos brotos(em). peso da matéria fresca e peso da matéria seca(rng),O material vegetal utilizado foi Coffea arábica L.cv. Catuaí Vermelho LCH 2077-2-5-44. sendoutilizados frutos no estádio de cereja para a extraçãode sementes. Após a retirada do pergaminho. assementes permaneceram em água desti lada por 15horas. Após este período elas foram mergulhadasem hipoclorito de sódio por I minuto. eposteriormente em álcool 70% por meia hora. Osembriões foram extraídos sobre placas de petri,sob estereomicroscópio. mantendo-os intactos.Em seguida foram inoculados no meio.colocando-se um embrião por tubo de ensaio.correspondendo à parcela experimental.Os tratamentos foram constituídos de todas ascombinações possíveis entre as concentrações deANA (0.0; 0.01; 0,1 e 1.0 rng/l) e de BAP (0.0:

3,0; 6,0 e 9,0 mg/l), resultando num fatorial 4 x 4,utilizando-se delineamento inteiramentecasualisado (0Ie) com 4 repetições.Para a análise estatística, foram utilizadas duasmaneiras de decompor a variação entretratamentos. A primeira delas correspondeu aodesdobramento em fatores principais e interaçãoentre eles, típico de ensaios fatoriais. A outramaneira fez uso do ajustamento a modelos desuperfície de resposta (Box e Draper, 1987),quando as diferenças entre os tratamentosmostraram-se significativas pelo teste F. Ométodo de 'backward (Draper e Smith, 1981) foiutilizado para a seleção de modelos de regressãomúltipla envolvendo os fatores testados noexperimento, utilizando-se o resíduo da análise devariância para testar a significância doscoeficientes de regressão. Uma vez determinado omodelo. testava-se a significância dos desvios deregressão para verificar a adequabilidade domodelo.
RESULTADOS E DISCUSSÃO
O resumo das análises de variância, para ascaracterísticas aval iadas, estão representadas nasTabela I, considerando a partição da variação entretratamentos segundo o esquema fatorial, e nasvariações devido ao modelo de regressão múltiplaajustado e devido aos desvios de regressão.Observa-se que para as características número totalde brotos, número total de folhas, brotos maioresque I centímetro, peso fresco e peso seco, houveuma interação significativa entre os reguladores decrescimento uti lizados. O desvio de regressão, paratodas as variáveis. não foi significativo,evidenciando que os modelos foram ajustadossatisfatoriamente.
Número de brotaçõesDe acordo com o modelo de superficie de respostaajustado, a utilização da concentração máxima deauxina (ANA=l ,00 mg/l), associada à concentraçãode citocinina BAP de 7,42 mg/l. promoveu o maiornúmero de brotações (valores estimados dentro dointervalo aval iado). O número de brotaçõesesperado para esta combinação seria de 3,00brotações/explante (Figura I). Não houve interaçãoentre os reguladores de crescimento ANA e BAP,através da anál ise fatorial (Tabela I), porém, omesmo não foi observado pelo modelo de regressãomúltipla ajustado (Figura 1), pois neste. ocorre apresença de termos envolvendo os dois fatores, oque demostra a presença de interação. Os desviosde regressão para esta variável, mostraram-se nãosignificativos. evidenciando que os modelosajustados util izados foram satisfatórios (Tabela I).
80
Concentrações elevadas de auxina, associadas àconcentrações elevadas de citocinina, promoveramum aumento no número de brotações. O contráriofoi observado por George (1994), em cujo trabalho,concentrações elevadas de auxina, inibiam odesenvolvimento do embrião. Raghuramulu (1989),também obteve grande êxito em C. arabica, quandoutilizou concentrações de 1,00 mg/I de ambosreguladores de crescimento. Concentrações acimaou abaixo destas não foram satisfatórias. Asconcentrações acima de 1,00 mg/l de auxina foramconsideradas tóxicas, promovendo morte doembrião.
Número de brotos maiores que um centímetroO modelo de superficie de resposta ajustadodemostrou que o número máximo de brotaçõesmaiores que 1,00 em foram obtidas comconcentrações hormonais em sua dosagem máxima(ANA = 1,00 mg/I e BAP = 9,00 mg/l) como podeser observado na Figura 3, em um período de 60dias de cultivo. Para esta característica houveinteração entre os fatores (Tabela I), o que pôde serconfirmado pela presença de termos no modeloenvolvendo os dois fatores, simultaneamente(Figura 2). Inúmeros trabalhos mostram que osucesso na cultura de embriões está associado adiversos fatores. incluindo o comprimento doexplante, que deve ser igualou superior a 1,00 em,permitindo uma maior sobrevivência (Coloria,1972; Krasnyanski et al, 1992; Raghuramulu,1989).
Número total de folhasPara o número total de folhas. ocorreu interaçãoentre os fatores demonstrado, tanto pela análisefatorial (Tabela I). quanto pelo modelo ajustado,sendo confirmado pela existência de termos nomodelo envolvendo os dois fatoressimultaneamente. A não significância dos desviosde regressão demonstra que o modelo utilizado foisatisfatório (Tabela 1). A concentração máxima deauxina (ANA = 1,00 mg/l), juntamente com 7,39mg/I de citocinina (BAP) resultou num maiornúmero de folhas por explante. Segundo o modelode superficie de resposta ajustado para estasconcentrações, espera-se que o número total defolhas seja de 12 folhas por explante.
Peso de matéria fresca e seca da parte aérea dasbrotações
Nestas características as concentraçõesintermediárias de ambos os reguladores decrescimento promovem uma resposta maior.Segundo o modelo de superfície de respostaajustado. a resposta máxima seria obtida com uma
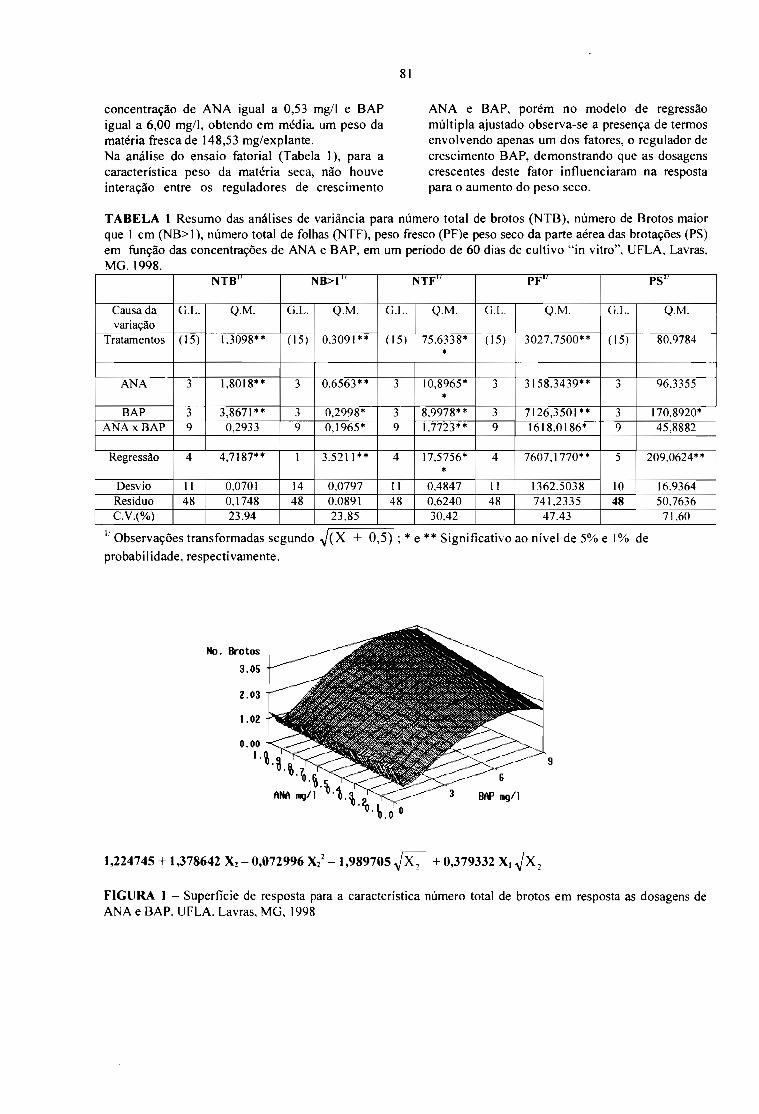
concentração de ANA igual a 0,53 mg/I e BAPigual a 6,00 mg/l, obtendo em média, um peso damatéria fresca de 148,53 mg/explante.Na análise do ensaio fatorial (Tabela I), para acaracterística peso da matéria seca, não houveinteração entre os reguladores de crescimento
81
ANA e BAP, porém no modelo de regressãomúltipla ajustado observa-se a presença de termosenvolvendo apenas um dos fatores, o regulador decrescimento BAP, demonstrando que as dosagenscrescentes deste fator influenciaram na respostapara o aumento do peso seco.
TABELA 1 Resumo das análises de variância para número total de brotos (NTB), número de Brotos maiorque I em (NB> I), número total de folhas (NTF), peso fresco (PF)e peso seco da parte aérea das brotações (PS)em função das concentrações de ANA e BAP, em um período de 60 dias de cultivo "in vitro", UFLA. Lavras.MG.1998.
NTB" NB>I" NTF PF PS
Causada G.L. Q.M. G.L. Q.M. G.L. Q.M. G.L. Q.M. G.L. Q.M.variação
Tratamentos (15) 1,3098** ( 15) 0.3091** (15) 75.6338* ( 15) 3027.7500** (15) 80.9784*
ANA 3 1,8018** 3 0.6563* * 3 10.8965* 3 3158.3439** 3 96.3355*
BAP 3 3,8671 ** 3 0,2998* 3 8.9978** 3 7126,3501** 3 170,8920*ANA x BAP 9 0,2933 9 0,1965* 9 1.7723** 9 1618.0186* 9 45,8882
Regressão 4 4,7187* * 1 3.5211 ** 4 17.5756* 4 7607,1770** 5 209.0624***
Desvio 11 0,0701 14 0,0797 11 0.4847 11 1362.5038 10 16.9364Resíduo 48 0,1748 48 0.0891 48 0.6240 48 741.2335 48 50.7636C.V.(%) 23.94 23.85 30.42 47.43 71,60
1I Observações transformadas segundo ,J(X + 0,5) ; * e ** Significativo ao nível de 5% e 1% de
probabilidade, respectivamente.
No. Brotos
3.05
2.03
1.02
9
1,224745 + 1,378642 Xz - 0,072996 X/ - 1,989705.,JX; + 0,379332 XI.,JX;
FIGURA 1 - Superfície de resposta para a característica número total de brotos em resposta as dosagens deANA e BAP. UFLA. Lavras, MG, 1998
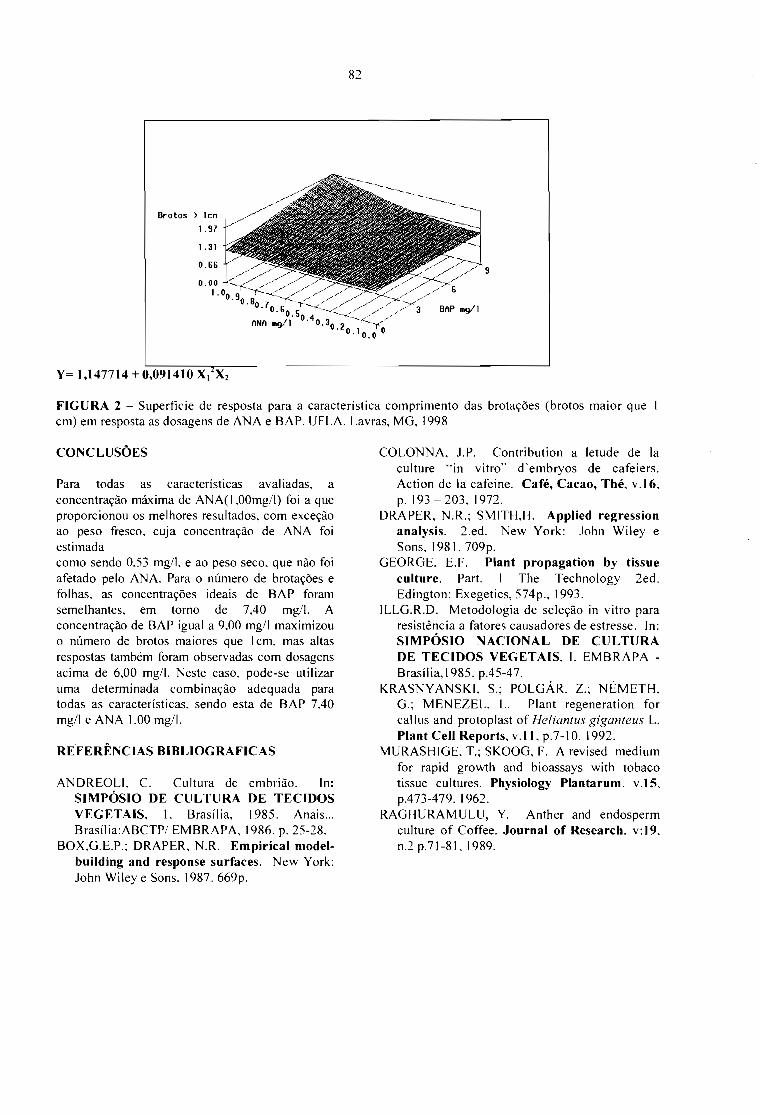
82
Brotos ) lcm
1. 97
1. 31
-----
BAP mgll
y= 1,147714+0,091410X. X2
FIGURA 2 - Superfície de resposta para a característica comprimento das brotações (brotos maior queem) em resposta as dosagens de ANA e BAP. UFLA. Lavras, MG, 1998
CONCLUSÕES
Para todas as características avaliadas. aconcentração máxima de ANA(I .Oümg/l) foi a queproporcionou os melhores resultados. com exceçãoao peso fresco. cuja concentração de ANA foiestimadacomo sendo 0.53 mg/I, e ao peso seco. que não foiafetado pelo ANA. Para o número de brotações efolhas. as concentrações ideais de BAP foramsemelhantes. em torno de 7,40 mg/I. Aconcentração de BAP igual a 9,00 mg/l maximizouo número de brotos maiores que 1em, mas altasrespostas também foram observadas com dosagensacima de 6,00 mg/l. Neste caso, pode-se utilizaruma determinada combinação adequada paratodas as características. sendo esta de BAP 7,40mg/l e ANA 1.00 mg/l.
REFERÊNCIAS BIBLlOGRAFICAS
ANDREOLl. C. Cultura de embrião. In:SIMPÓSlO DE CULTURA DE TECIDOSVEGETAIS. L Brasília. 1985. Anais ...Brasília:ABCTP/ EMBRAPA, 1986. p. 25-28.
BOX.G.E.P.; DRAPER. N.R. Empirical modelbuilding and response surfaces. New York:John Wiley e Sons. 1987. 669p.
COLONNA. LP, Contribution a letude de laculture "in vitro' dembryos de cafeiers.Action de la cafeine. Café, Cacao, Thé, v. I6,p. 193 - 203, 1972.
DRAPER, N.R.; SMITH.H. Applied regressionanalysis. 2.ed. New York: John Wiley eSons. 1981. 709p.
GEORGE. E.F. Plant propagation by tissueculture. Part. I The Technology 2ed.Edington: Exegetics, 574p., 1993.
ILLG,R.D. Metodologia de seleção in vitro pararesistência a fatores causadores de estresse. In:SIMPÓSIO NACIONAL DE CULTURADE TECIDOS VEGETAIS. 1. EMBRAPA Brasília, 1985. p.45-47.
KRASNYANSKI, S.; POLGÁR. Z.; NÉMETH,G.; MENEZEL, L. Plant regeneration forcallus and protoplast of Heliantus giganteus L.Plant Cell Reports, v.I L p.7-10. 1992.
MURASHIGE. T.; SKOOG, F. A revised mediumfor rapid growth and bioassays with tobacotissue cultures. Physiology Plantarum. v.IS,p.473-479. 1962.
RAGHURAMULU, Y. Anther and endosperrnculture of Coffee. Journal of Research. v: I9.n.2 p.71-81, 1989.

83
INFLUÊNCIA DE REGULADORES DE CRESCIMENTO NA MICROPROPAGAÇÃO "INVITRO" DA CULTIVAR CATUAí VERMELHO LCH 20 77-2-5-44
Luciana Marques da Cunha Andrade', André 8arretto Pereira', Anna Lygia de Rezende Maciel",Moacir Pasqual 4
, • 4'Bióloga.>- 2Pesquisador CEPEC/CEPLAc' Pós-graduação/UFLA;-'Bolsista CBP&O/CAFE;- Prof TitularOAG/UFLA, Caixa Postal 37, CEP 372000-000, Lavras-MG.
•
INTRODUÇÃO
A técnica de cultura de tecidos vegetais "in v itro"possibilita a propagação em larga escala e emcurto espaço de tempo, sendo importante para asculturas de ciclo longo. Esta técnica é vantajosaquando aplicada em variedades melhoradas quepossuam pouco material e necessitam serpropagadas rápida e massivarnente, com afinalidade de produção de mudas.A ocorrência natural de determinados hormônios éinsuficiente para suprir as necessidades da planta.portanto é necessário a adição suplementar dessassubstâncias para se obter resultados satisfatórios epromissores em relação à micropropagação(Pasqual e Pinto, 1988).Curster (1980) foi um dos primeiros a trabalhar commicropropagação em Coffea arabica. obtendosucesso com a propagação de brotos ortotrópicos eplagiotrópicos em meio de cultura "MS",concentrações elevadas de auxinas e baixasconcentrações de citocininas, tendo resultadossemelhantes aos obtidos pelos métodosconvencionais "in vivo", sendo que a maiordificuldade correspondeu à contaminação e aoxidação dos explantes utilizados.O objetivo deste trabalho foi aval iar a influência dasconcentrações dos reguladores de crescimentoBAP, GA, e TOZ na micropropagação da cultivarCatuaí Vermelho LCH 2077-2-5-44.
MATERIAL E MÉTODOS
Os experimentos foram realizados no Laboratóriode Cultura de Tecidos Vegetais do Departamento deAgricultura da Universidade Federal de Lavras UFLA, em Lavras - Minas Gerais.O material vegetal utilizado foi Coffea arabica L.cv. Catuaí Vermelho LCH 2077-2-5-44, sendo queas plântulas já se encontravam estabelecidas "invitro", com cerca de seis meses de vida.O meio nutritivo básico utilizado foi o deMurashige e Skoog (1962), sendo que o pH foiajustado para 5,9. Para a solidificação do meio foiutilizado ágar (7g/l).Foram conduzidos dois experimentos, nos quaisforam àvaliadas aos 90 dias as variáveis númerototal de brotos. número de folhas, número de brotos
maiores que 1em, peso de matéria fresca e seca(mg).Os explantes utilizados foram retirados de plântulasisentas de contaminantes, desprezando-se as folhase a gema apical, utilizando-se sempre umamicroestaca por tubo com 2 gemas e estas foraminoculadas verticalmente no meio.
Experimento 1: Multiplicação "in vitro" variandose as influência das concentrações de BAP e GA,no cultivar Catuaí Vermelho LCH 2077-2-5-44.Neste experimento os tratamentos utilizados foramconstituídos de todas as combinações possíveisentre as concentrações de BAP: 0,0; 3,0; 6,0 e 9,0mg/l e de GA3: 0,0; 2,5; 5,0; 10,0 e 20,0 mg/l,resultando em um fatorial 4 x 5, disposto em umdelineamento inteiramente casualizado (OIC), com4 repetições.
Experimento 2: Multiplicação "in vitro" variandose as concentrações de TOZ e GA, no cultivarCatuaí Vermelho LCH 2077-2-5-44.Neste experimento os tratamentos foramconstituídos de todas as combinações possíveisentre as concentrações de TOZ: 0,0; 3.0; 6,0 e 9,0mg/l e de GA, : 0.0; 2,5; 5.0; 10,0 e 20,0 mg/l,resultando em um fatorial 4 x 5, disposto em umdelineamento inteiramente casualizado (OIC), com4 repetições.
Análises estatísticasPara a análise estatística, foram utilizadas duasmaneiras de compor a variação entre tratamentos. Aprimeira delas correspondeu ao desdobramento emfatores principais e interação entre eles, típico deensaios fatoriais. A outra maneira fez uso doajustamento a modelos de superficie de resposta(Box e Draper, 1987) quando as diferenças entre ostratamentos mostraram-se significativas pelo testeF.
RESULTADOS E DISCUSSÃO
Experimento 1O resumo das análises de variância, para ascaracterísticas avaliadas, está apresentado nasTabela 1. Observa-se que nas três primeirascaracterísticas. número total de brotos, número de

brotos maiores que Iem e número total de folhas,houve interação significativa entre os reguladoresde crescimento BAP e GA3 •
Número de brotaçõesO modelo de superficie de resposta ajustado (FiguraI) demonstra que para a maximização no númerode brotações, a dosagem adequada de citocinina(BAP) deve ser igual a 0,99 mg/I em ausência deácido giberélico (GA 3) dentro do intervalo avaliado.O número de brotações esperado para estacombinação é de 2,42 brotações/explante. Nota-setambém que, sob concentrações crescentes de GA3 ,
associadas a uma dosagem constante de BAP iguala 0,99 rng/l, ocorre uma redução no número debrotações. Este fato evidencia que o ácidogiberélico, para esta característica não mostrou-seeficiente.
Número de brotos maiores que 1 emO maior número de brotações maiores que 1,00 emfoi verificado, utilizando concentração máxima deácido giberélico (GA 3 =20,00 mg/l) associado àconcentração máxima de citocinina (BAP = 9,00rng/l), de acordo com o modelo de superficie deresposta ajustado.
Número total de folhasO aumento no número de folhas/explante foiobtido quando a concentração da citocinina BAPera de 0,95mg/1 na ausência do regulador decrescimento GA 3• O número total de folhasestimado seria de 12 folhas/explante. Asconcentrações utilizadas de GA3 associadas àsconcentrações de BAP apresentaram redução nonúmero de folhas e clorose generalizada.
Peso da matéria fresca e seca da parte aérea dasbrotaçõesHouve um aumento no peso fresco das brotaçõesquando o hormônio GA3 estava ausente e o BAP naconcentração de 0,96 mg/l, podendo neste casoatingir o peso fresco médio de 105,79 mg/l. Avariável peso seco apresentou resposta semelhanteao peso fresco, sendo que os melhores resultadosforam obtidos na ausência de GA3 e BAP nadosagem de 1,0 mg/l.
Experimento 2Os resultados obtidos para as característicasavaliadas no experimento 2 estão apresentados na(Tabela 2. Todas as características, exceto pesoseco, foram influenciadas pela interação entre osreguladores de crescimento TDZ e GA3 (Tabela 2).
84
Número de brotaçõesO maior número de brotações estimado, segundo omodelo de superficie de resposta ajustado, é obtidoquando a concentração de GA3 atinge o suadosagem máxima (GA 3 = 20 mg/I) interagindo coma dosagem de 0,94 mg/I de TDZ, sendo o númerode brotações estimado de 2,15 brotações/explante(Figura 2). A interação entre os reguladores decrescimento foi verificada pela análise de variância(Tabela 2). O desvio de regressão para estacaracterística não foi significativo, o que indica queo modelo ajustado foi satisfatório (Tabela 2).Neste experimento houve uma maior influência dohormônio GA 3 . aumentando o número debrotações, ao contrário do ocorrido no experimentoI, no qual, a ausência deste hormônio provocou osmelhores resultados. A citocinina, BAP e o TDZ,em ambos os experimentos, determinaram umamelhor resposta sob semelhantes concentrações,sendo de 0,99 mg/l para o experimento I e 0,94mg/I para o experimento 2, correspondendo aresultados semelhantes.
Número de brotos maiores que 1 cmPara obtenção de maior número de brotações comtamanho superior a 1,00 em, a combinação dasdosagens hormonais seria de 0,98 mg/I de TDZassociado a 20,00 mg/l de GA 3. As concentraçõescrescentes de TDZ em relação à ausência de GA 3
apresentaram comportamento constante, e asdosagens intermediárias entre Oe 3,00 mg/I de TDZassociadas às crescentes dosagens de GA 3
apresentaram respostas satisfatórias.
Número Total de folhasA maximização no número total de folhas/explante,foi obtida na utilização do hormônio GA 3 na suadosagem máxima (GA 3 = 20mg/I), interagindo comTDZ na dosagem de 0,98 mg/l. Neste caso onúmero total de folhas seria de 5 folhas/explante.
Peso fresco e peso seco da parte aérea dasbrotaçõesO aumento no peso fresco das brotações, foi obtidocom concentração de TDZ igual a 0,84 mg/lassociado à dosagem máxima de GA 3 20 mg/l,correspondendo a um peso fresco estimado de80,71 mg.O peso seco da parte aérea das brotações nãoapresentou diferenças significativas entre asconcentrações dos reguladores de crescimento.(Tabelal)
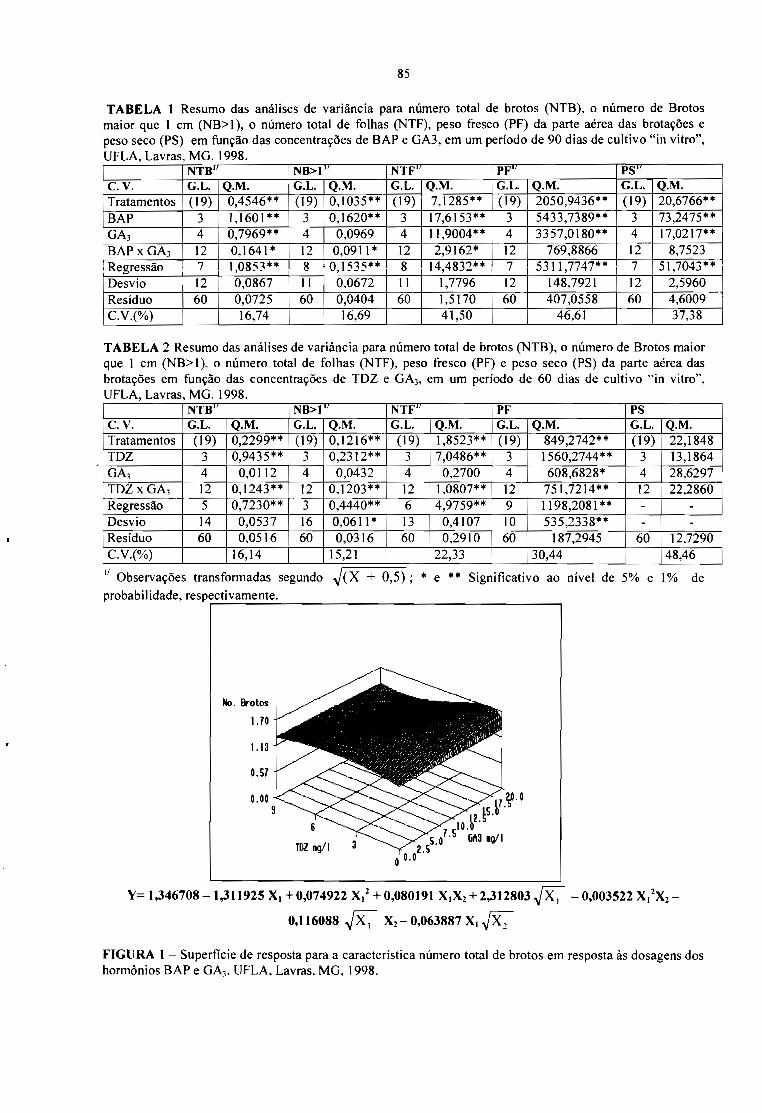
85
TABELA 1 Resumo das análises de variância para número total de brotos (NTB), o número de Brotosmaior que I em (NB> I), o número total de folhas (NTF), peso fresco (PF) da parte aérea das brotações epeso seco (PS) em função das concentrações de BAP e GA3, em um período de 90 dias de cultivo "in vitro",UFLA Lavras, MG. 1998.,
NTU" NU>]" NTF" PF" PS"C. V. G.L. Q.M. G.L. Q.M. G.L. Q.M. G.L. Q.M. G.L. Q.M.Tratamentos (19) 0,4546** (\ 9) 0,1035** (19) 7,1285** (19) 2050,9436** (19) 20,6766**BAP 3 1,1601** 3 0,1620** 3 17,6153** 3 5433,7389** 3 73,2475**GAJ 4 0,7969** 4 0,0969 4 11,9004** 4 3357,0180** 4 17,0217**BAP x GAJ 12 0,1641* 12 0,0911* 12 2,9162* 12 769,8866 12 8,7523Regressão 7 1,0853** 8 0,1535** 8 14,4832** 7 5311,7747"'* 7 51,7043**Desvio 12 0,0867 1I 0,0672 11 1,7796 12 148,7921 12 2,5960Resíduo 60 0,0725 60 0,0404 60 1,5170 60 407,0558 60 4,6009C.V.(%) 16,74 16,69 41,50 46,61 37,38
TABELA 2 Resumo das análises de variância para número total de brotos (NTB), o número de Brotos maiorque 1 em (NB> 1), o número total de folhas (NTF), peso fresco (PF) e peso seco (PS) da parte aérea dasbrotações em função das concentrações de TDZ e GAJ , em um período de 60 dias de cultivo "in vitro",UFLA, Lavras, MG. 1998.
NTU NO>]" NTF PF PSC. V. G.L. Q.M. G.L. Q.M. G.L. Q.M. G.L. Q.M. G.L. Q.M.Tratamentos (\9) 0,2299** (\ 9) 0,1216** (19) 1,8523"'* (19) 849,2742** (19) 22,1848TDZ 3 0,9435** 3 0,2312** 3 7,0486** 3 1560,2744** 3 13,1864GAJ 4 0,01 12 4 0,0432 4 0,2700 4 608,6828* 4 28,6297
. TDZ x GAJ 12 0,1243** 12 0,1203** 12 1,0807** 12 751,7214** 12 22,2860Regressão 5 0,7230** 3 0,4440** 6 4,9759** 9 1198,2081 ** - -Desvio 14 0,0537 16 0,0611 * 13 0,4107 10 535,2338** - -Resíduo 60 0,0516 60 0,0316 60 0,2910 60 187,2945 60 12,7290C.V.(%) 16,14 15,21 22,33 30,44 48,46
11 Observações transformadas segundo .J(X + 0,5); * e ** Significativo ao nível de 5% e 1% deprobabilidade, respectivamente....------------------------,
No. Brotos
1.70
1.13
0.57
y= 1,346708 -1,311925 XI + 0,074922 X/ + 0,080191 XIX2 + 2,312803~ - 0,003522 X/X2
0,116088~ X2 - 0,063887 XI~
FIGURA 1 - Superficie de resposta para a característica número total de brotos em resposta às dosagens doshormônios BAP e GAJ • UFLA, Lavras, MG, 1998.

86
No. Brotos
1.75
1.1&
0.58
17 520.012 5
15.O .10.0 .
5.0 7.5 liA3-o/100.0 2.5
= 1,219206 + 0,180836 XI - 0,019904 X/ - 0,080515 X.X2 + 0,004286 X. 2X2 + 0,126698 IX: X2
FIGURA 2 - Superfície de resposta para a característica número total de brotos em resposta às dosagens doshormonios TOZ e GA3• UFLA. Lavras, MG, 1998.
CONCLUSÃO
As melhores concentrações para as característicasavaliadas, corresponderam à ausência do hormônioGA3 em associação com concentrações mínimas deBAP. As concentrações do hormônio BAP para ascaracterísticas n'' total de folhas, peso fresco, n° totalde brotos e peso seco foram 0,95, 0,96, 0,99 e 1,00mg/I respectivamente. Os melhores resultados, paraa característica n° de brotos maiores que 1,0 em,foram obtidos quando as concentrações máximas deBAP e GA 3 foram utilizadas.Na relação entre os hormônios TOZ e GA3, paratodas as características avaliadas, é recomendada autilização da concentração máxima de GA3
(20,00mg/l) associada às concentrações mínimas deTOZ variando de 0,84 a 0,98 mg/1. Asconcentrações que apresentaram os melhoresresultados para TOZ, nas seguintes características,foram: peso fresco (0,84 mg/l), número debrotações (0,94 mg/I) e número total de folhas ebrotos maiores que 1,0 em (0,98 mg/I).
REFERÊNCIAS BIBLIOGRÁFICAS
building and response surfaces. New York:John Wiley e Sons, 1987. 669p.CAMBRONY,H.O.; SNOECK,J. Hormones etregulateurs de croinance dans les cultures decaféiers et de cacaoyers. Café, Cacao, Thê,Paris, v.27, n.2, p.l13-119, 1983.
CUSTER,J.B.M.; van EE, G; BUIJS, L.C.;Clonal propagation on Coffea arabica L. by nodalculture. In: PROC. IX INTERN. SCIENT.CELL. COFFEE. ASIC., London, p. 588 - 596.1980.ORAPER, N.R.; SMITH,H. Applied regressionanalysis. 2.ed. New York: John Wiley e Sons,1981. 709p.PASQUAL, M.; PINTO, J.E.B. Cultura deembriões. In: NOTICIAS DA ASSOCIAÇÃOBRASILEIRA DE CULTURA DE TECIDOSDE PLANTAS, Brasilia, v.2, p.2-12. 1988.SONDAHL,M.R.; NAKAMURA, T.;MEDINAFilho, H.P.; CARVALHO, A.; FAZUOLI, i.c.,COSTA, W:R: Coffee. In:Ammyrato eMcMillen, eds. Handbook of plant cell culture.New York, p.564-590., 1984.ZOK, S., DUBLIN, P. Multiplication Végétativein vitro par culture D' Apex chez Coffea arabica L.Action de Solutions Minerales et de Regulateursde croissance. Café, Cacao, Thé, v. 14.

87
EVALUACIÓN MORFOLÓGICA Y AGRONÓMICA DE PLANTAS DE CAFÉ REGENERADASPOR EMBRIOGÉNESIS SOMÁTICA
Juan Carlos Herrera - Pinilla, Maria Eugenia Aponte - A
Disciplina Mejoramiento Genético y BiotecnologíaCentro Nacional de Investigaciones de Café CENICAFÉChinchiná, Caldas ColombiaAA. 2427 Manizales. FAX.: (0968) 504723. E- mail:[email protected]
INTRODUCCIÓN
La embriogénesis somática a partir de tejido foliarrepresenta un sistema modelo de regeneración deplantas que resulta particularmente atractivo paraotras especies arbóreas. Desde los prirnerosexperimentos realizados por Sondhal y Sharp(1977), la embriogénesis somática ha mostradotener importantes aplicaciones no solo para lapropagación vegetativa de diferentes cultivares,sino también en la conservación de recursosgenéticos y en la diseminación de indivíduosseleccionados provenientes de cruzasintervarietales o interespecíficas (Dublin, 199 J).Aunque la propagación clonal vía embriogénesissomática se considera como un sistema bastanteestable que permite la obtención de plantasgenéticamente "idénticas", es bien conocida laexistencia de una variación debida al procesomisrno y conocida como variación somaclona1.Esta variación somacJonal tiene dos componentesbásicos: (i) la variabilidad natural presente entodas las células somáticas, y (ii) una variacióndebida a diferentes factores propios de lamanipulación In vitro (Sondhal y Loh, 1985). Lavariación somaclonal se ha planteado como unainteresante vía para obtener variantes en unaespecie de estrecha variabilidad genética como elcafé (Sondahl y Bragin, 199 J), sin embargo, talvariación puede resultar inconveniente cuando setrata de establecer una vía de regeneracióneficiente con miras a la propagación vegetativa deuna variedad, o ai desarrollo de un programa detransformación genética.Desde hace cerca de JO aüos, el laboratorio debiotecnología deI Centro Nacional deInvestigaciones de Café (CENICAFÉ) empezó atrabajar en la optimización dei proceso deregeneración de plantas a partir de explantesfoliares con miras a establecer un protocoloaltamente eficiente y reproducible, con miras a suutilización en un futuro proyecto detransformación genética dei café. Dentro de estetrabajo se consideró importante conocer laestabilidad fenotípica y genotípica de lossomaclones producidos pues de ella dependeráque la(s) característica(s) introducida(s) a las
plantas transgénicas se mantengan y expresenadecuadamente una vez éstas sean lIevadas aicampo. Por tanto, el objetivo dei presente trabajofue el de comparar el desarrollo morfológico y elcomportamiento agronómico de las plantasregeneradas a partir de embriogénesis somáticadirecta (ESD) e indirecta (ESI), con respecto aplantas obtenidas por semi lia y provenientes deimismo árbol donante.
MA TERIALES Y MÉTODOS
Material Vegetal: Se estudiaron dos poblacionesde somaclones obtenidas de árboles de diferentegeneración de selección (F3 y F5) derivados deicruce entre C. arabica var. Caturra y el híbrido deTimor (CIFT 1343). Estos materiales constituyenprogenies elite que están siendo evaluadas comoposibles componentes de la variedad Colombia.
Regeneración de plantas vía embriogénesissomática: Se indujo la regeneración de lossomaclones siguiendo las vía directa ó ESD(embriogénesis somática directa) e indirecta ó ESI(embriogénesis somática indirecta) descritas acontinuación.Para la inducción de la ESD los explantes fueron
sembrados en un medio de inducción denominadoL-3 y compuesto por las sales básicas de M&S(Murashige y Skoog, 1962) diluidas a la mitad ysuplementadas con piridoxina (I mg/l), tiamina(10 mg/l), ácido nicotinico (I mg/I), inositol (100mg/l), L- cisteína (37 mg/l), sacarosa (30 g/l),BAP: bencil amino - purina (3 mg/l) y agarDlFCO (8 g/l) como agente gelificante. EL pH fueajustado a 5.8 antes de esterilizar. Luego de cuatromeses, los explantes se subcultivaron aI medioL1QC-1 cuya cornposición es igual a la descritapara el medio L1QC-3 , con excepción de laconcentración de BAP la cual se redujo a I mg/l.Para la inducción de la ESI se utilizó un mediosimilar ai L-3 pero con las siguientes variaciones:las sales de M&S fueron completas, y comoreguladores de crecimiento se utilizó 2,4-D(2mg/l) y la kinetina (8 mg/l), esta últimaadicionada ai medio luego de la esterilización,usando un filtro de 0.2 micrometros. AI cabo de

un mes en cultivo, el tejido indiferenciado fuetransferido ai medio L-3 para permitir lainducción final deI tejido embriogénico.z aparecieron los primeros embriones (140 díasdespués), éstos se trasladaron a un medio degerminación sin hormonas. Después de dos mesesen oscuridad, fueron transferidos a condiciones deluz (1500 lux, con un fotoperíodo de 12 h luz y 12h oscuridad) para inducir la formación de lasprimeras hojas. Cuatro meses después lasplántulas formadas fueron 1Jevadas a condicionesde invemadero bajo cámara húmeda. Finalmentecada planta fue transferida a bolsa con una mezclade suelo y pulpa (2: I), hasta que estuvieron listaspara lIevar aI campo (aproximadamente 17 mesesdespués de la siembra de los explantes).Paralelamente con este proceso, se colocaron agerminar semilias provenientes deI mismo árboldonante (población SEM), de manera queestuviesen listas para pasar aI campo a una edadsimilar a la de las plantas provenientes deIlaboratorio.Siguiendo los métodos antes descritos fue posibleconformar dos poblaciones: una población Aproveniente de un árbol F3 formada por 50plantas de ESD y 50 de SEM; y una población Bconformada por 62 plantas de ESD, 12 de ESI Y46 provenientes de SEM. Las dos poblacionesfueron sembradas en lotes separados, con unadisposición de plantas ai azar en la estaciónexperimental "Naranjal" ubicada en el municipiode Chinchiná, Caldas (Colombia) a 1400 m.s.n.m.y con una temperatura media de 20.8°C.Variables evaluadas: Cuando las plantasaícanzaron los dos afios y medio de edad, sedeterminaron varios parámetros morfológicostales como: altura, el número y longitud de losentrenudos, el índice foliar (relación largo / anchode la hoja) y el ángulo de inserción de las ramas.Paralelamente se inició la evaluación agronómicade la producción (producción promedio por árboly producción acumulada) de la cual se hancompletado dos ciclos correspondientes a los aüos1996 y 1997. Igualmente, se determinó elporcentaje de granos normales (supremo) ydefectuosos (vanos y caracoles) calculados apartir de dos muestreos realizados durante 1996 y1997. Finalmente, se determinó el grado deincidencia de la roya deI cafeto (Hemileiavastatrix), siguiendo la escala cualitativa
88
desarrol1ada por Eskes y Toma-Braghini (1981)que permite medir la intensidad de ataque deIhongo sobre el fol1aje utilizando una calificaciónque va desde O a 10, siendo O un estado deausencia total deI hongo (resistencia) y 10 elestado de máximo ataque (susceptibilidad)
Análisis estadístico Tanto para la población declones derivados de una planta F3 (población A)como la derivada de una F5 (población B), sereaJizaron análisis de varianza a fin de compararel comportamiento de cada variable (fenotípica oagronómica) entre las plantas de diferente origen(ESD, ESI Y SEM) siempre a un nivel designificancia deI 5 %. Para el análisis deresistencia a la roya se establecieron tres gruposde susceptibilidad de acuerdo ai grado deincidencia (O, 1-4, >4). Utilizando la prueba dechi-cuadrado (aI 5% de significancia), se comparóla proporción de plantas perteneciente a cadagrupo, entre los diferentes tipos de plantasestudiadas para cada población.
RESULTADOS Y DlSCUSIÓN
Evaluación fenotípica: En las tablas I y 2 sepresentan los resultados de la evaluaciónfenotípica de las plantas pertenecientes a las dospoblaciones estudiadas. No se encontrarondiferencias significativas para las variables altura,ángulo de inserción de la rama e índice foliarentre los somaclones (ESD, ESI) y las plantas desemi1Ja pertenecíentes a las poblaciones A y B.Diferencias estrechas pero significantes fuerondetectadas para el número de nudos y la longitudde los entrenudos, sin embargo el comportamientode estas dos variables fue diferente para cadapoblación, mostrando un efecto adicional de otrosfactores diferentes aI tratamiento mismo. Esposible que las diferencias en las épocas desiembra y en el tipo de generación (F3 o F5) entrecada población hayan incidido en las diferenciasya mencionadas.En lo que respecta ai color de los frutos y de los
brotes foliares jóvenes, no se detectaron cambiospara ninguno de los tipos de plantas evaluadas. Entodos los casos, las semi lias fueron de calor rojomientras los brotes mostraron una tonalidadbronce clara.
89
Tabla l.Características morfológicas evaluadas para la población A derivada de una planta F3 proveniente dei cruceentre C arabica varo Caturra X Híbrido de Timor. Comparación de parámetros estadísticos obtenidos para plantasderivadas de semilla (SEM) y de embriogénesis somática directa (ESD).
l/Valores estimados ai nível de 0.05 de probabilidad /.. Diferencias estadísticamente significativas según pruebat-student ai 0.0 I de probabilidad.
VARIABLE Tipo de planta Número de Plantas Promedio Intervalo de onfianza"Altura SEM 49 169.6 138.7 - 200.5(em) ESO 46 168.0 140.3 - 195.8Número de SEM 49 34.5 28.8 - 40.2Nudos ESO 46 36.1 ** 30.9 - 41.3Longitud de los SEM 49 4.9 ** 4.4 - 5.4entrenudos (em) ESO 46 4.6 4.1 - 5.1Angulo de Inserción de SEM 49 58.3 51.5-65.1las ramas (grados) ESO 49 58.4 52.7 - 64.1Indiee Foliar SEM 49 2.5 2.3 - 2.7
ESO 49 2.4 2.2-2.7..
Tabla 2. Características morfológicas evaluadas para la población B derivada de una planta F5 proveniente dei cruceentre C arabica var. Caturra X Hibrido de Timor. Comparación de parámetros estadísticos obtenidos para plantasderivadas de semilla (SEM), de embriogénesis somática directa (ESD) e indirecta (ESI).
11 Valores estimados ai nível de 0.05 de probabilidad21 Promedios con letra distinta son estadísticamente diferentes ai O.OJ de probabilidad
VARIABLE Tipo de planta Número dePlantas Promedio Intervalo de confianza "Altura SEM 46 175.5 a LI 172 - 179(em) ESO 62 172.9 a 170-176
ESI 12 174.5 a 167-182
Número de SEM 46 38.4 a 37.7 - 39.2Nudos ESO 62 36.2 b 35.5 - 36.8
ESl 12 37.5 ab 36.1 - 38.9Longitud de los SEM 46 4.69 b 4.61 - 4.76entrenudos ESO 62 4.85 a 4.78-4.91(em) ESI 12 4.65 ab 4.51 - 4.80
I Angulo de Insereión SEM 46 54.13 a 53.2 - 55.1de las ramas (grados) ESO 62 52.82 a 52.0 - 53.6
ESI 12 53.08 a 51.3-54.9Indiee Foliar SEM 46 2.46 a 2.44-2.49
ESD 62 2.49 a 2.47 - 2.51ESI 12 2.52 a 2.47 - 2.57..
Tabla 3. Producción media de las plantas regeneradas por los diferentes métodos (ESD, ESI Y SEM) y pertenecientes alas dos poblaciones estudiadas. Los datos corresponden a 2 ciclos (anos) de evaluación.
Población Tipo deplanta Número de plantas Producción (Kg de c/árbol) VarianzaPoblaeión A (F3 de SEM 50 5.87 3.539caturra X H. de Timor) ESO 50 5.88 3.264Poblaeión B(F5 de SEM 46 4.45 1.073caturra X H. de Timor) ESD 62 4.30 0.925
ESI 12 4.49 0.481
Evaluación agronómica : Producción: Se evaluó Laproducción promedio por árbol y la producciónpromedio total por tratamiento para lo clones derivadosde un árbol F3 de Caturra X Híbrido de Timor(población A) y los clones derivados de un árbol F5 deCaturra X Híbrido de Timor (población B) y los
respectivos testigos obtenidos por semilla. No sedetectaron diferencias significativas entre los clonesobtenidos por ESD y ESL con respecto a las plantasderivadas de semilla, ai cabo de dos ciclos completosde producción (tabla 3; figuras la y Ib).
90
Tabla 4. Características de grano evaluadas para las plantas regeneradas por los diferentes métodos(ESO, ESI Y SEM) y pertenecientes a las dos poblaciones estudiadas. Los datos corresponden ai promedio dedos muestreos real izados entre 1996 y 1997.
Población Variable Tipo de planta Número de plantas Promedio (%) VarianzaPoblación A Grano vano SEM 48 19.07 132.09(F3 de Caturra Grano ESO 47 31.23 ** 15.96X H. de caracol SEM 48 12.24 2.96Timor) Grano ESO 47 12.88 3.75
supremo SEM 48 68.77 48.00 ,ESO 47 71.20 45.38
Población B Grano vano SEM 44 4.7 4.791(F5 de Caturra Grano ESO 62 4.5 4.967X H. de caracol ESI 12 5.7 62.858Timor) Grano SEM 44 12.6 ** 5.127
supremo ESO 62 10.5 3.61ESI 12 9.2 1.697
SEM 44 79.2 33.017
~ESO 62 79.5 17.925ESl 12 77.3 17.722..** Diferencias estadísticas ai nivel de probabilidad dei 0.01.
Como se esperaba, se encontró una mayorvariación en la producción promedio por árbol delos individuos obtenidos por semi liapertenecientes a la población A (32.04%) respectoa la B (23.3%). Este resultado tiene su explicaciónen la heterozigocidad remanente que presentan lasplantas de la población A ya que derivan de unárbol F3 (hetrozigocidad esperada 12.5 %).mientras que los individuos de la población Bprovienen de semilla F5 (heterozigocidadesperada de 3.1 %).
Caracteristicas de grano: Los datos deIporcentaje de granos normales (supremo) yanormales (vanos y caracoles) para las plantas delas poblaciones A y B se presentan en la tabla 4.No se encontraron diferencias significativas paraporcentaje de grano supremo entre los somaclonesy las plantas de semilla en ninguna de las dospoblaciones estudiadas. AI observar ladistribución dei porcentaje de los diferentes tiposde grano se pudo observar el efecto de laselección a favor de los individuos de grano tiposupremo (figuras 1b y 2b).Las diferencias significativas observadas entre lossomaclones y las plantas de semilla para elporcentaje de granos vanos (población A) yporcentaje de granos caracol (población B).muestran nuevamente un posible efecto deigenotipo y de la época de siembra sobre estasvariables.
Resistencia a roya. La resistencia a roya es unavariable de gran importancia agronómica y portanto cualquier cambio en los genes responsablesde dicho carácter, debido ai proceso de clonación,
podría ser negativo ai momento de utilizar estesistema en la obtención de nuevas variedades. Losdatos obtenidos hasta hoy en las dos poblacionesanalizadas, muestran que la vía de regeneración(ESD o ES1) no afecta de manera alguna laresistencia de estas plantas a la roya y que talcomo se esperaba, los clones muestran igualcomportamiento que las plantas derivadas desemi lia.La población A presentó un porcentaje similar de
plantas ESD y SEM similar (figura 1c) ydistribuido en dos grupos bien diferenciados:plantas parcialmente resistentes (cal ificación 1-4)Y plantas suscepti bles (cali ficación >4). Estadistribución refleja la variación que aun persisteen estas plantas de generación F3. con respecto aesta variable. La población B por su parte mostróun comportamiento diferente. Tanto lossomaclones como las plantas de semilla seagruparon como plantas parcialmente resistentes.lo cual se explica por la selección intensa que seha ejercido sobre estos materiales de generaciónF5 (figura 2c).Uno de los pocos estudios referentes a lavariación somaclonal en café es el publicado porSõndahl y Bragin (1991). En él se reportandiferentes frecuencias de variación somaclonalpara una población de 12.176 plantas regeneradasa partir de embriogénesis somática de alta y bajafrecuencia en distintos genotipos de Coffeaarabica. Según éstos autores. un 10 % de lasplantas presentaron una variación encaracterísticas como la morfología de la hoja, Iaestatura de la planta. el color, tarnafio y forma deifruto. asi como para algunas otras variablesrelativas a la producción. tiempo de maduración

dei fruto y la tolerancia o susceptibilidad a ciertasenfermedades. Las frecuencias de variaciónreportadas en ese trabajo oscilaron entre 0.01 %(para la variable "grano maragogipe") y el 42.3 %(para la mutación de grano amari 110 a grano rojo).La comparación entre estos resultados y losobtenidos en nuestro ensayo, debe tomarse concautela debido a que el tipo y concentración de losreguladores de crecimiento uti lizados por Sõndahly Bragin fueron diferentes a los empleados ennucstro laboratorio. De: hecho, su intención deinducir variabi lidad en los materiales regenerados,se refleja en el uso de concentracionesrelativamente altas de auxina (1,1 mg/l de 2,4-0)y citoquinina (4,3 mg/l de cinetina). Actualmenteexiste suficiente evidencia que muestra que lavariación somaclonal esta influenciada por el tipoy concentración de los reguladores de crecimientoincluidos en el medio. Se sabe que el empleo deeste tipo de sustancias conlleva el riesgo devariaciones a nivel de la estructura y el número delos crornosornas, afectando en forma transitoria opermanente la estabilidad fenotípica y genotípicade las plantas regeneradas. en especial en aquellasespecies perennes como el café. donde el tiempocomprendido entre dos generaciones se hace muylargo (Karp, 1992; Karp, 1995).
CONCLVSIONES
EI estudio morfológico de las dos poblaciones deplantas analizadas, muestra que, siguiendo losprotocolos establecidos en CENICAFÉ, elproceso de regeneracion vía embriogénesissomática directa (ESO) o indirecta (ESI), noinduce cambios significativos en la altura, elángulo de inserción de las ramas o el índice foliar.Asi rnismo, características agronómicasimportantes como la producción media por árbol,el porcentaje de granos normales (tipo supremo)y la resistencia a la roya de los clones regeneradospor embriogénesis somática, tanto directa (ESD)como indirecta (ESI). mostraron ser iguales a lasde plantas obtenidas por semi lia. Estecomportamiento fue similar para las dospoblaciones estudiadas.De otra parte, tanto las características deI granocomo la resistencia a la roya reflejaron lasdiferencias en la intensidad de selección aplicadasobre las dos poblaciones analizadas (F3 y F5)siendo, sin embargo, similar su comportamientopara los clones y para las plantas donantesderivadas de semi lia.Las bajas concentraciones hormonales utilizadasdurante las fases de ind ucción y crecimiento, asícomo la reducción significativa en el tiempo totalde desarrollo In vitro, parecen ser los factores
91
responsables de la mmima variación somaclonalobservada hasta hoy. Esta información será degran importancia para el proyecto detransformación genética en el que, los materialestransformados. deberán ser regenerados por éstavía.
REFERENCIAS BIBLIOGRÁFICAS
Aponte A.. M.E. 1993. Somatic embryogenesisinduced by culture on single media in coffeeplants from crosses of Coffea arabica by Timorhybrid. Memorias XV Coloquio CientíficoInternacional sobre Café, ASIC. p. 82-88.Montpellier, Francia.Dublin P. 1991. Multiplicación vegetativa de café.hevea y cacao. IN: Cultivo de tejidos en laagricultura. Fundamentos y aplicaciones. Roca,W. M.; Mroginski, L. A. (Eds). Capo 26. P.577619. Centro Internacional de Agricultura Tropical,CIAT. Cali, Colombia.Eskes, A. 8.; Torna-Braghini, M. 198 I.Assesment methods for resistance to coffee leafrust (Hemileia vastatrixy. FAO. Plant Prol. BulI..29.56.Karp, A.. 1992. The role of growth regulators insomaclonal variation. British Society for PlantGrowth Regulation Annual Bulletin N° 2. May1992, p. 1-9.Karp, A.. 1995. Somaclonal variation as a tool forcrop improvement. Euphytica 85: 295-302.Sondahl M. R.; Sharp, W.R. 1977. Highfrecuency induction of somatic embryos in cultureleaf explants of Coffea arabica L. Z.Pflanzenphysiol. 81: 395-408.Sondahl M. R.; Loh W. H.-T. 1985. Coffeebiotechnology. IN. Coffee. Clarke R.J.; Macrae R.(Eds). Agronomy, Vol 4. p. 235-262. ElsevierApplied Sciences Publ. London, UK.Sondahl M. R.; Bragin, A. J 991. Somaclonalvariation as a breeding tool for coffeeimprovement. Memorias XIV Coloquio CientificoInternacional sobre Café, ASIC. p. 701-709. SanFrancisco, Estados Unidos.


93
A MODEL FOR THE STUDY OF V ARfATlON fNDUCED BY TlSSUE CULTURE ON COFFEE.
Fuentes-Cerda, C. F. J., M. Monforte-González, M. Méndez-Zeel, R. Rojas-Herrera*, J. MijangosCortés and V. M. Loyola-Vargas.
Centro de Investigación Científica de Yucatán. Calle 43, No. 130, Co\. Chuburná de Hidalgo, Mérida, Yuc.México. C. P. 97200.... Instítuto Nacional de Cíencias Agrícolas. Gaveta Postal I. San José de las Lajas, LaHabana. Cuba.
ABSTRACTCo./fee (Co./fea spp.) is one of the most important agricultural commodities in the world. C arabica and C.
canephora are the principal varieties of the genus. Staritsky (1970) firstly described somatic embryogenesisin C canephora in 1970 and in C arabica by Sonda I & Sharp in 1977. Both systems are based in the use ofgrowth regulators (auxins and citokynines) to induce a de-di./ferentiation stage and the formation of calli.We had modified the protocol descibed by Yasuda et al., (1985) to create a model that al/ow us: 1) to use"in vitro" cultured plants as starting material and thus generate /ines from selected plants, 2) the inducciónof direct somatic embryogenesis avoding the callus stage and 3) a rapid responding system (globularembryos are obtained at day 22) in contrast with other systems where the obtention ofembryos take 2 ar 3months. Histological studies using this system had shown that embryos are originated directly from explantcel/ rather that from cal/uso When we analyzed regenerated plants, we could not find any somaclonalvariation at molecular levei.
fNTRODUCTlON
Coffee (Co./fea spp.) is one ofthe most ímportantagricultural commodíties ofthe world. C arabicaand C canephora are the main commercialspecies of the genus. In vitro somaticembryogenesis of C canephora was first reportedby Staritsky (1970), who descríbed the inductionof callus tissue from orthotropíc internodes; later,Herman and Hass (1975) obtained somaticembryogenesis in C. arabica from callus culturesderived from leaf explants. Sõndahl and Sharp(1977, 1979), developed a two phasesexperimental protocol for somatic embryogenesisfrom leaves of C arabica varo "Bourbon". Dublin( 1981) reported somatic embryogenesis from leafexplants of Arabusta using a medium withcytokíníns but without auxins. Yasuda et al.,(1985) induced embryogenic calli from Carabica varo "Typica" leaf explants using 5 11Mbenzylamino purine (BAP), white and fríable calliwere obtaíned after 16 weeks and somaticembryos were obtained four weeks following calliinitiation.Vigorous research using both C canephora. andC arabica, has allowed the use of differentexplants such as orthotropic stem fragrnents,plagiotropic branches, leaves, ovule coverings forthe development of somatic embryogenesissystems (Dublin, 1980a; 1980b; 1981). Leaveshas proved to be the best explants for thispurpose. The systems proposed by Dublin (1980a;1980b; 1981) and Sõndahl and Sharp
(1977, 1979) are among the most successful; inboth methods indirect somatic embryogenesis inleaves explants from mature trees ispromoted.Our protocol describes a simple onestep system for the induction of direct somaticembryogenesis of C arabica that poses importantadvantages.
MATERfALS AND METHODS
Plant Material. Seeds of C arabica cv. "CaturraRojo" were collected from several productiveplots at Chiapas, Mexico. Seeds were washed andsoaked during 24-48 hours in sterile distilledwater, disinfested with 6% sodium hipochloritefor 20 rninutes and finally rinsed with steri lewater. The culture medium employed for zygoticembryos germination was MS (Murashige andSkoog, 1962), with thiamine (29.6 11M), rnyoinositol (0.555 11M), biotin (0041 11M), L-cysteine(0.15 11M), glucose (166.48 mM), NAA (0.5311M), Kin (2.32 11M) and Gelrite® (0.25%); pH
was adjusted to 5.7 before autoclaving (20 rnin,110°C). Ten embryos were placed on eachMagenta® plastic box. Culture conditíons werephotoperiod (16 h/8 h) and 25°C.
Plantlets CuItu re. Plantlets were grown inMagenta® boxes, containing 40 mL of MSmedi um, with thiamine (11.86 11M), myo-inositol(0.56 11M), L-cysteine (0.16 11M), sucrose (87.64mM), NAA (0.54 11M), Kin (2.32 11M) andGelrite® (0.25%); pH was adjusted to 5.8 before

autoc lav ing (20 rrun, IIO°C). C ulture co nditionswe re photoperiod (16 h/8 h) and 25 °C. Plantletswe re tran sfe rred to fres h medium every 90 days.Somatic Embryogenesis Induction . Lea l'
fragments (0.25 cm2) were cultured in themedium previously descr ibed by Yasuda et ai.,( 1985), us ing two di fferent ge lling agents (Agar0.9% and Gelrite® 0.25 %). C ulture co nd itio nswere photop eriod ( 16 hJ8 h) a t 25 °C.
Histology. Foli ar expl ants we re co llec ted for 60days und er induction condition s, fixed and storedin FEA (formo l, etha no l, acetic ac id) . T hey we redehydrat ed with ethan ol , embedde d in J8-4 res in,sectioned and sta ined wi th tolu idin e blu e.
RESULTS
Model Description . O ur resu lts have a llowed toprop ose a new mod el for so ma tic em bryogenesisin coffee, which is described in figure I.Scarr ing was observed at inj ured edges within thefirst two weeks and bet ween the thi rd and fourth ,glo bular somatic embryos we re obse rve d in theedges 01' explants.
94
Histological s tudies have showed the formation 01'proembryo s d irectly from ce lls rather than fromthe ca llus. A lthough Dubl in ( 1981) had rep orteddirec t somatic em bryogenes is from Arabusta (C.arabica x C. canehora ), no histologica l evi dencesupported his resu lts . As can be seen in figure 2(A , B, C), proembyos originated fro m themesophy ll cel ls and in some rare cases, fromepidermal ce lls (Fig. 2 0). Indi rect em bryoformatio n was als o obse rve d from m icroca lli(F ig. 2 E).G iven the use 01' in vitro ge rm ina ted plantl et s asex plants, our wo rk radi call y di ffers fro m that 01'Yasuda et ai ., ( 1985) whe re adults plant s wereused . Further more the endogenous content 01'growth regulators could di ffer dr ast icall y bet weenad ult plan ts and in vitro p lant lets lead ing to thes pec tac ular res ults .At the mom en t, it wo uld be far-fetched to ass ign abiotech nolog ical application for th is system ( Iowyields) ; but it ca n be an excellent approac h tostudy the biochemical and molecular events thatlead em bryo fo rmation. This mod el also pro videsa unegu aled opportunity to study so mac lona lvariation in coffee embryogenesis .
(
FIC.l

95
FIG 2
Effeet or Genotype And Drigin or TheExplants. Plantlets from zygotic and sornatic
origin were tested and showed a differentialresponse. There was a better response in sornatic
plantlets, as has been previously reported forother species.The position of lhe leaf in lhe plant
and of explant in lhe leaf had a strong effecl onlhe response (Figure 3). The leaf number 3showed lhe besl results lo lhe induction ofsornatic ernbryogenesis, while explants obtainednear lhe apex showed a disminished responsecompared lo those obtained from lhe base of lheleaf.
Nitrogen Souree. Since lhe early 60s. it is knownthat the nitrcgen source has an irnportant effecl onplant differentiation The total nitrogen, lhenitrate/arnrnonium ratio and lhe type of lhenitrogen source have a deep effect on lheresponse of lhe explants lo lhe sornaticembryogenesis
process. However, lhe effecl of nitrogen sourceon sornatic embryogenesis has nol been studied incoffee.As lhe total nitrogen in lhe medium increased, lheresponse lo sornatic embryogenesis decreased(Fig. 4A). A 60 mM nitrogen concentration had aslrong inhibitory effect, while with lhe lowestconcentration, the response was maximal. Thenitrate/ammonium ratio also modified theresponse. The presence of ammonium is essentiallo obtain a response and nitrate has an inhibitoryeffect (Fig. 4B). In fact, both nitrogen sources areessential to induce the phenomenom.When the effecl of the nitrogen source wasmeasured 100 days after the induction lhe effectof the nitrogen source on lhe response, lhe totalnum bel' of embryos and lhe number of embryosper explant was clear (Fig. 5A & B).
96
100
90
8U
~ 70
w no111
~llZo 40O-lIt
3Uwrt
10..
10 .~;
11)1) r--90 l m
~~o E
11170 ;a
I-e
w 50 o(/I
I/)~o mz
o ~X
o- 411 "«
f
,-I/) .")0
I'~ fi~
w "
IIX 10 l;: ..10 ",
:<. \':.O
i_._._
I)(I :1
B) POSITION FROM BASI!I TO TIP
FIG lA nGJB
100
! 00
100ao
~ 18"'00~ 10:$--i'..- T.O.T .5
... 8'10-+- 00,1560
40
lU<li
zo0U>lUIr
wI/lZoo111Wrt
A) TOTAL DF NITRDGEN
FI(; 4A FI(; 48
oo oi> "'! o '"oii <; ... r. e-e- ... .p .... o...
100r---- :~ 10
1_
ao ít!l I'l a I,
l -U8
6 II -t60
~~ o(
•"tr
4 Is II 20
~ i ~
2 ~
I) NlRA11i:AMONl~RELAnOH (II'1II)
0'-------
,140 10II
1J)
8 Il00~
oo~ si~ .!001
40~2-4
20
o O4 8 15 30 60
~ lOTAL OF NI'TROGEN ("'"
o
80
lI 60
I 40
I 20
FIG5A FIG5R

CONCLUSIONS
Dírect somatíc embryogenesis is the formatíon ofsomatíc embryos from the explant whithout theformatíon of an intermedíate calli phase(Raghavan 1986). The production of callus isusually undesirable if embryogenesis is the goalof the research. Unfortunately, in most plantsdírect somatic embryogenesis is dífficult toobtaín. This process in leaves does not happenfrecuently. Conger et a/ .• (1983) and Trigiano etal., (1989) have reported direct somaticembryogenesis from leaves of Dacty/is g/omerataand Duboís et a/.. (1991) in Cichorium. Dublin(1981) reported somatíc embryogenesis from leafexplants of Arabusta using a medíum withcytokinin, but no auxins. To our knowledge this isthe first report of dírect somatic embryogenesisfrom explants of leaves in Coffea arabicasupported by histological evidence. We hadmodified the protocol described by Yasuda et a/o(1985) to create a model that allows us: to use invitro cultured plants as startíng material and thusgenerate lines from selected plants,a fast and responsive system (globular embryosare obtained at day 22) in contrast with othersystems where embryos take 2 or 3 months todevelop.
REFERENCES
Conger, B.V .. Hanning, G.E., Gray, D.1 .. MeDaniel, J.K. Direct embryogenesis frommesophyll cells of orchardgrass. Science 221:850-851, 1983.Dublin, P. Embryogenêse somatique dírecte surfragments de feuilles de caféíer Arabusta. CaféCacao Thé XXV (4):237-242, 1981.
Dublin, P. lnduction de bourgeons néoformés etembryogenése somatique. Deux voíes demultiplícatíon végétative in vitro des caféíerscultivés. Café Cacao Thé XXIV (2): 121-130.1980.
97
Dublin, P. Multiplication végétatíve in vitro deI'Arabusta. Café Cacao Thé XX IV (4):281-290.1980.Dubois, T .. Guedira, M. and Vasseur, J. Directsomatic embryogenesis inleaves of Cichorium. Ahistological and SEM study of early stages.Protoplasma 162: 120-127, 1991.Herman, E.B. and Haas, G.H. Clonalpropagation of Coffea arabica L. from callusculture. HortScience 10(6):588-589, 1975.Raghavan, V. Embryogenesís in angiosperms: adevelopmental and experimental study.Cambridge University Press, Carnbridge, 1986.Sõndahl, M. R. and Sharp, W. R. 1977. Highfrequency inductíon of somatic embryos 10
culture leaf explants of Coffea arabica L. Z.lanzenphysiol. 81 :395-408.Sõndahl. M. R. Y Sharp, W. R. 1979. Research inCoffea spp., and applications of tissue culturemethods. In Sharp, W.R., l.ersen, P.O .. Paddock,E.F., Raghavan, V. (eds.) Plant cell and tissuecultures, principies and applications . Ohio StateUniv. Press. Columbus. pp 527-584Staritsky, G. Embryoid formation in callus tissuesof coffee. Acta Bot.Need. 19:509-514, 1970.Trigiano, R.N., Gray, D.1., Conger, B.V.. MeDaniel, J.K. Origin of direct somaticembryogenesis from cultured leaf segments ofDactylis glomerata. Bot. Gaz. 150: 72-77.Yasuda, T., Fujii, Y., Yamaguchi, T.Embryogenic callus induction from Coffeaarabica leaf explants by benzyladenine. P/ant Ce//Physiol. 26 (3):595-597.1985.
ACKNOWLEDGMENTS
The authors thanks Drs. Teresa Hernández andLourdes Miranda for the revision of the Englishversion of the manuscript. The work of theauthors has been supported by CONACYTMéxico. grant 4123P-N and Consejo Mexicanodei Café.


99
STANDARDlZATION OF ENCAPSULATION TECHNIQUE FOR PRODUCING SYNTHETICSEEOS IN COFFEE
B. Muniswamy And H L. Sreenath
Biotechnology Centre Unit ofCentral Coffee Research Institute Coffee Board, Manasagangothri,Mysore-570 006, INDIA
SUMMARYPlant regeneration from encapsulated zygotic and somatic embryos of coffee is reported. The somaticembryos obtained directly from cultured zygotic embryos of CxR cultivar of robusta coffee on MS mediumsupplemented with NAA and BAP and zygotic embryos of C. arabica were used in the study. They wereencapsulated in six percent sodium alginate in MS salts solution by dropping them into two percent calciumchloride solution. The resulted alginate beads with embryos were cultured both in-vitro on MS medium andex-vitro in potted vermiculite. Fast germination and plantlet development was achieved on half-strength MSmedium supplemented with 0.1 mg/lor 0.5 mg/I of Kn or O.lmg/1 BAP. When the plantlets produced ]-4pairs of leaves with good root system, they were hardened with a good survival rate. The encapsulatedembryos that were sown directly ex-vitro in vermiculite could not be regenerated into plantlets. /mportanceofthe encapsulation technique for producing synthetic seeds in coffee are discussed.Abbreviations: BAP- 6Benzylamino purine, Kn - Kinetin, MS-Murashige and Skoog (1962) NAA - I-Napthalene acetic acid
INTRODUCTION
Seeds are the primary means of propagation inmany crops. The use of genetically uniforrn tissueculture derived plants, would be advantageous inplants like coffee, in which vegetative propagationis used to raise uni forrnly superior clonalplantations. Micropropagation technique isgreatly constrained due to the high cost involvedand inadequate standardization of hardeningprocedures. During transfer of plants fromlaboratory to field, generally high mortal ity ratesresult that further lower the efticacy of resultsachieved in-vitro (Mathur et al, 1989). For thesereasons, efforts have been made for overcomingthe constraints by encapsulating somatic embryos(Kitto and Janick, 1985; Redenbaugh et aI. 1987)in different matrices and attempting to grow themon different media. Studies on somaticembryogenesis and related encapsulation ofsomatic embryos has been reported byRedenbaugh et al. (1993). The first succesfulexamples of synthetic seed technology was inalfalfa and celery. Fujii et aI. ( 1989) could growplants from synthetic seeds of alfalfa planteddirectly in the field at low frequency. KirinBrewery Co Ltd .• Japan and Plant Genetics Inc.USA conducted field trails in Japan with 20,000FI synthetic seeds in 1988 (Sanada et al. 1993).Recently, Rao et al. (1998) reviewed the concept,methods and applications of synthetic seeds.In coffee, somatic embryogenesis has beensuccessfully achieved from different tissues (Sondahl and Loh, 1988; Sreenath and Naidu1997; Muniswamy and Sreenath 1997). However.reports on encapsulation studies and subsequent
plant regeneration for producing synthetic seedsin coffee are very much limited. In ourlaboratory, plant regeneration from encapsulatedzygotic and somatic embryos of coffee isreported. This technique has tremendous use inthe large scale micropropagation and conservingencapsulated zygotic and sornatic embryos inliquid nitrogen (at _196° C) for long termpreservation of coffee germplasm.
MATERIALS ANO METHOOS
Material SourceThe somatic embryos were regenerated directlyfrom the cultured zygotic embryos of CxRcultivar of robusta coffee (an interspecific hybridbetween C. congensis and C. canephora) werecultured on MS medium supplemented withdifferent concentrations of NAA and BAP andzygotic embryos of C. arabica (S.4348 andS.2794). These embryos were cultured on MSmedium with ABA (I mg/l) for 30 days forembryo maturation and then used in theencapsulation study.Preparation of encapsulation matrix Differentconcentrations of sodium alginate and calciumchloride were studied. Sodium alginate (2-6%)was added to MS salts solution, supplementedwith 3% sucrose and with or without growthregulators. pH of the medium was adjustedbetween 5.6 and 5.8 and medium was sterilized ina domestic pressure cooker. For an exchangereaction between Na+ and Ca ++ resulting in theformation of insoluble calcium alginate (i.e ..complexation) two percent of calcium chloridewas prepared.

Formation ofheadsThe embryos mixed with sodium alginatesoJution were dropped through a pasteur pipetteinto two percent calcium chloride soJution in aflask, placed on a magnetic stirrer with continuousstirring. The resulting beads containing entrappedembryos were left as such in the calcium chloridesolution for 30 minutes for complexation. Thebeads were separated from the solution by usingsterilized stainless steel sieve. The beads rangingbetween 4 - 6 mm in diameter were placed ondifferent media for germination and subsequentplantlet development.
Culture mediaThe encapsulated embryos were cultured both invitro on medium and ex-vitro In pottedvermiculite for germination. For in-vitrogermination, MS medium supplemented with 0.1or 0.5 mg/I of Kn or 0.1 mg/I of BAP with 3%sucrose was used. The pH of the medium wasadjusted between 5.6 - 5.8, solidified with 0.8%bacteriological agar and then steri lized in pressurecooker for 20 minutes. The cultures wereincubated at room temperature with 70 - 80%relative humidity under a photo period of 12 h (around 2000 lux) using cool white fluorescenttubes. For ex-vitro germination of theencapsulated ernbryos, finely sieved sterilevermiculite filled in the plastic pots was used.
RESULTS AND DJSCUSSION
In-vitro germinationAmong different concentrations of sodiumalginate and calcium chloride tested, six percentsodium alginate and two percent calcium chloridesolutions gave optirnal, firm and round beads witha complexation period of 30 minutes.Somatic embryos obtained directly from the
zygotic embryos of CxR cultivar of coffee at highfrequency on 0.1 mg/I ofNAA and 1 mg/lofBAPwere used for encapsulation. The nonencapsulated embryos cultured on half-strengthMS medium with 0.1 mg/I of Kn and 0.1 mg/l ofBAP. started germinating and developed intoplantlets with long hypocotyls, greencotyledonary leaves and roots. However, whenthe embryos were encapsulated with half strengthMS salts solution with or without cytokinin andcultured on half-strength MS medíumsupplemented with different concentrations of Kn(0.1 or 0.5 rng/l) and BAP (0.1 mg/l), they startedgerminating with elongation of hypocotyl anddeveloping the cotyledonary leaves after 30-40days of culture period. (Fig. IA). Later, theseplantlets grew further with thick greenish
100
hypocotyl of 7 to 11 mm long and 6-12 mm widewell developed dark green cotyledonary leavesabout 6 - 12 mm wide. Most of them haveproduced first pair of leaves and good roots afterthree months culture. The details of media andcomparison of plant development from nonencapsulated and encapsulated embryos are givenin the Tahle 1.After 2-3 subcultures on half-strength MSmedium supplemented with Kn (O.lor 0.5 mg/I)and BAP (0.1 mg/I) at 30-45 days intervals,plantlets developed further and produced 2-5pairs of additional leaves with long shoot andgood roots (Fig. 1B). At this stage, plantlets weretaken out from culture vessel without damagingthe roots and planted in potted vermiculite forhardening. These plantlets were watered with 1/8strength MS salts on alternate days andestablished with a good survival rate. After twomonths growth in pots, hardened plantlets wereplanted in polybags filled with soil mixture.Ex-vitro germination Encapsulated embryossown directly in potted vermiculite did notgerrninate, they turned brown and died within 2weeks. They were watered with 1/8 strength MSsalts solution on alternate days and sprayed with0.1 % Bavistin once in a week for avoiding fungalgrowth.Though, gerrnination and plantlet deveJopment
was achieved in both encapsulated and nonencapsulated embryos, the encapsulated embryosgerminated faster and developed into good,healthy plantlets with dark green leaves. Therewere c1ear differences in respect of germinationand plantlet development between nonencapsulated and encapsulated embryos. Theencapsulated embryos responded better whenthey were cultured initially on half-strengthMS medium supplemented with 0.5 rng/l ofKn for one month and subcultured subsequentlyon medium of same composition with reducedconcentration of Kn (0.1 mg/l). The time takenfor plantlets to reach hardening stage wasreduced by two to three months in encapsulatedembryos as compared to non-encapsulatedembryos. However, germination and plantletdevelopment could not be achieved from theencapsuJated somatic embryos directly sown exvitro. Encapsulation technique is useful forproducing synthetic seeds in coffee by usingsomatic embryos. This would help in scaling upmicro propagation and for germplasmconservation through cryopreservation.In trees species, development of synthetic seeds isvaluable because of the need for vegetativepropagation and lack of cost effective methods(Redenbaugh et ai, 1988 ). Kamada (1985) also
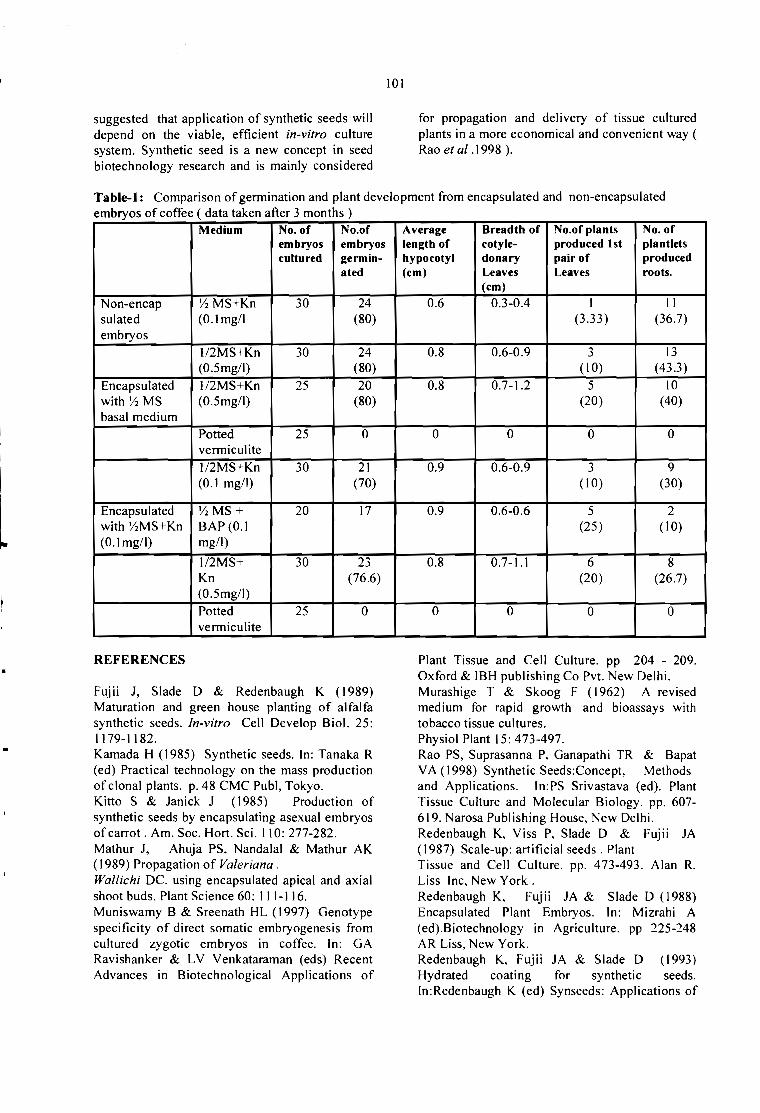
suggested that application of synthetic seeds wíl\depend on the viable, efficient in-vitro culturesystem. Synthetic seed is a new concept in seedbiotechnology research and is mainly considered
101
for propagation and delivery of tissue culturedplants in a more economical and convenient way (Rao et ai .1998 ).
Table-l: Comparison of germination and plant development from encapsulated and non-encapsulatedembryos of coffee ( data taken after 3 months )
Medium No.or No.of Average Breadth of No.of plants No.orembryos embryos length of eotyle- produeed 1st plantletseultured germin- hypoeotyl donary paír or produeed
ated (em) Leaves Leaves roots.(em)
Non-encap Ih MS+Kn 30 24 0.6 0.3-0.4 I 1Isulated (0.1 mg/I (80) (3.33) (36.7)embryos
l/2MS+Kn 30 24 0.8 0.6-0.9 3 13(0.5mg/l) (80) (10) (43.3)
Encapsulated 1/2MS+Kn 25 20 0.8 0.7-1.2 5 10with lf2 MS (0.5mg/l) (80) (20) (40)basal medium
Potted 25 O O O O Overmiculite1/2MS+Kn 30 21 0.9 0.6-0.9 3 9(0.1 mg/I) (70) (10) (30)
Encapsulated Ih MS + 20 17 0.9 0.6-0.6 5 2with If2MS+Kn BAP (0.1 (25) (10)(0.1 mg/l) mg/I)
1/2MS+ 30 23 0.8 0.7-1.1 6 8Kn (76.6) (20) (26.7)(0.5mg/l)Potted 25 O O O O Overmiculite
•
REFERENCES
Fujii r, Slade O & Redenbaugh K (1989)Maturation and green house planting of alfalfasynthetic seeds. In-vitro Cell Oevelop Biol. 25:1179-1182.Kamada H (1985) Synthetic seeds. In: Tanaka R(ed) Practical technology on the mass productionof clonal plants. p. 48 CMC Publ, Tokyo.Kitto S & Janick 1 (1985) Production ofsynthetic seeds by encapsulating asexual embryosofcarrot. Am. Soe. Hort. Sei. 110: 277-282.Mathur J, Ahuja PS, Nandalal & Mathur AK(1989) Propagation of Valeriana .Wallichi De. using encapsulated apical and axialshoot buds. Plant Science 60: 111-116.Muniswamy B & Sreenath HL (1997) Genotypespecificity of direct somatic embryogenesis fromcultured zygotic embryos in coffee. In: GARavishanker & LV Venkataraman (eds) RecentAdvances in Biotechnological Applications of
Plant Tissue and Cell Culture. pp 204 - 209.Oxford & IBH publishing Co Pvt. New Delhi.Murashige T & Skoog F (1962) A revisedmedium for rapid growth and bioassays withtobacco tissue cultures.Physiol Plant 15: 473-497 .Rao PS, Suprasanna P. Ganapathi TR & BapatVA (1998) Synthetic Seeds.Concept, Methodsand Applications. In:PS Srivastava (ed). PlantTissue Culture and Molecular Biology. pp. 6076) 9. Narosa Publishing House, New Delhi.Redenbaugh K, Viss P. Slade O & Fujii lA(1987) Scale-up: artificial seeds . PlantTissue and Cell Culture. pp. 473-493. Alan R.Liss Inc. New York .Redenbaugh K. Fujii lA & Slade O (1988)Encapsulated Plant Embryos. In: Mizrahi A(ed).Biotechnology in Agriculture. pp 225-248AR Liss. New York.Redenbaugh K. Fujii lA & Slade O (1993)Hydrated coating for synthetic seeds.In:Redenbaugh K (ed) Synseeds: Applications of

Synthetic Seeds to Crop Improvement.pp.35-46. CRC Press. Inc. Boca Raton FI. USA.Sanada M, Sakamoto Y, Hayashi M, Mashiko T,Okamoto A & Ohnishi N (1993).Celery and Lettuce. In: Redenbaugh K (ed)Synseeds: Applications of Synthetic Seeds toCrop Improvement. pp 305-327. CRC Press,Boca Raton, USA.
102
Sõndahl MR & Loh WHT (1988). CoffeeBiotechnology. In: Clarke, R.L. and Macrae, R(eds). Coffee volA, Agronomy. pp. 235-262.Elsevier Applied Science, London.Sreenath HL & Naidu R (1997) CoffeeBiotechnology -review. In: GA Ravishankar& LV Ventakaraman (eds) Recent Advance inBiotechnological: Applications of Plant Tissueand Cell Culture. pp. 14-27. Oxford IBHpublishing Co. Pvt. Ltd., New Delhi.

103
DESARROLLO DE LA VARIEDAD PORTA-INJERTOS "NEMAYA" (C. canephora) POR MEDIODE EMBRIOGENESIS SOMATICA EN CENTROAMERICA.
F. Anzueto', A. Molina I, P. Figueroa I, H. Etíenne", B. Bertrand''
'Depto. de Investigaciones, ANACAFE, Guatemala - franciscºA@ª-nacafe.org2CIRAO-CA TIE, Costa Rica - [email protected] RAO - ICAFE, Costa Rica - [email protected]
I. INTRODUCCIÓN
Los nemátodos constituyen uno de los principalesproblemas parasitarios dei cafeto en AméricaCentral. Se reportan básicamente los génerosMeloidogyne spp. y Pratylenchus spp. LosMeloidogyne estan representados por variasespecies, con diferentes niveles de agresividad. EnGuatemala y El Salvador predominan especiesparticularmente "agresivas", mientras que en CostaRica y Honduras, está más difundida Meloidogyneexigua, de menor agresividad en campo (12).Trabajos realizados en Centroarnérica hace cuarentaanos, dieron indicios sobre la tolerancia y/oresistencia dei café "Robusta" a los nemátodos. Estopropició que en Guatemala se desarrollara eI injertohipocotiledonar denominado "Reyna", en referendaa su autor (15). La técnica se practica a escalacomercial con buenos resultados, a pesar que se hatrabajado con Robustas de polinización Iibre, sinselección, Esto podría explicarse por la circustanciaque Pratylenchus es el nemátodo predominante enese país. AI respecto, algunas investigacionesconfinnan que los "Robustas", en general, presentanun buen nivel de tolerancia a este nemátodo (1,4).En investigaciones preliminares se observó quemuchas descendencias de "Robusta" eran resistentesa poblaciones de Meloidogyne exigua yMeloidogyne incognita de Costa Rica (13, 7, 6), sinembargo, las evaluaciones efectuadas conMeloidogyne sp. de Guatemala (fenotipo "F")mostraron altos porcentajes de plantas susceptibles,de 50 a 80 por ciento, en descendencias de Robustade polinización libre provenientes deI CATIE,Costa Rica. En estos estudios también se observóque la descendencia de la planta T3751(1-2), de lamisma colección, presentaba un alto nivel deresistencia a Meloidogyne sp. (I ).La evaluación de un plan de cruzamientoscontrolados entre genotipos de Robusta (6), para laresistencia ai nemátodo Meloidogyne arenaria de EISalvador (14), mostró igualmente el valor de losgenotipos T3751 (1-2) y T3561 (2-1) como padresdestacados. La mejor combinación se obtuvo deIcruce entre ellos, con un porcentaje de plantasresistentes cercano ai 80 por ciento, que sería loesperado para Meloidogyne spp. en Guatemala y EISalvador, y aproximadamente 100 por ciento de
resistencia para los Meloidogynes de Costa Rica,Honduras y Nicaragua.Es importante sefíalar que en una mezcla de variasdescendencias libres -que podría ser el caso deRobustas no seleccionados- se presentan alrededorde 35 por ciento de plantas resistentes a losMeloidogynes de Guatemala y EI Salvador, o seaque habría una ganancia de 45 por ciento por eltrabajo de selección. En pruebas realizadas conPratylenchus también destacaron los genotipos T3751(1-2) y T3561(2-1) con buen nivel detolerancia a este nemátodo (I, 4).Luego de laselección de los genotipos T3561 (2-1) Y T3751(12), el objetivo era su multiplicación vía vegetativa.considerando la naturaleza de alogamia estricta deC. canephora, y luego establecer campos o jardinesbiclonales, para la producción de semi lia híbrida depolinización controlada, destinada a los programasde injertación de los caficultores centroamericanos,A esta variedad portainjertos se le denominóNemaya, por acuerdo de los investigadoresasociados ai proyecto (2).La estrategia y desarrollode las actividades se realizaron en el marco de lacooperación técnica entre PRüMECAFE, ClRAOy CATlE. Para la multiplicación de las plantasmadres se tomó la decisión de utilizar el métodode embriogénesis somática in vitro. Esta técnicaes conocida desde hace varios anos, con diversasapl icaciones en eI cafeto (8, I I, 16. 17). Ia mismaconsiste en tomar partes somáticas de la planta secciones de hoja generalmente-, que sonintroducidos in vitro para la formación de callosernbriogénicos y posteriormente embriones yplántulas.EI proceso in vitro total dura aproximadamente 12meses en laboratorio. La etapa siguiente incluye elenraizamiento y aclimatación de las plantas fueradei laboratorio (ex vitro). Esto se realiza enunidades de aclirnatación, sembrando las plantitasen bolsas con substrato o en camas de arena, yadaptándolas progresivamente desde humedadrelativa saturante (100%) hacia valores más bajosde humedad. Oespués de tres meses, las plantaspueden transferirse ai vivero, donde pasan 6 o 7meses antes de plantarse en campo. Lamultiplicación de callos ernbriogénicos se realizóen el laboratorio de Biotecnología dei CATlE(Costa Rica) y laboratorio de Cultivo de Tejidos

de ANACAFE (Guatemala). Las plantas seaclimataron en Guatemala, dentro de estructurasespecialmente construi das para este fino
2.MATERIALES Y MÉTODOS
2.1 Multiplicación de los padres porembriogénesis somáticaPara la multiplicación de los padres T3751 yT3561 se siguió eI protocolo sugerido por Etienney Berthouly (9, 11), con el siguiente proceso:Para la introducción de los rnateriales, se tomanhojas jóvenes en buen estado, sin lesiones. Lashojas se Javan con agua de chorro, luego sedesinfectan sumergiéndolas en solución de benlateIg/I por 5 minutos, posteriormente se colocan ensolución de hipoclorito de calcio ai 10 % por 20minutos, luego se realizan 3 lavados con aguaestéril. Las hojas quedan preparadas y se cortancuadros de I crrr', que se colocan en el medio decultivo de inducción de callo. Esta etapa sedesarrol1a en la obscuridad por 4 semanas, en unmedio de cultivo constituido por las salesminerales de Murashige et. al., (1962) diluídas50%, suplementado con: tiamina 10 mg, mioinositol 100 mg, ácido nicotínico I rng,pirydoxina I mg, glicina I mg, caseínahidrolizada 100 mg, extracto de malta 400 mg,sacarosa 30g, AIB I mg, 2 ip 2 mg, pH 5.6,Fitagel 2.5 g.Luego de 4 semanas el explante pasa a la etapa deexpresión de callo. Dicha etapa se desarrol1a enluz indirecta durante 6 o 7 meses. en un medioconstitui do por las sales minerales de Murashige,et. al., (1962) diluidas 50%, suplementado con:mio-inositol 200 mg, tiamina 20 mg, caseínahidrolizada 200 rng, extracto de malta 800 mg,cisteína 40 mg, sulfato de adenina 60 mg, glicina20 mg, sacarosa 30g, 2,4 D I rng, BAP 4 mg, pH5.6, Fitagel 2.5 g.Despues de 6 meses en el medio de expresión decallo, la masa callosa pasa a la etapa deregeneración, esta etapa se desarro 11 a en luzdirecta en fotoperíodo de 12 horas luz (2000 lux),
104
12 obscuridad, por 8 semanas, en medio decultivo constituido por las sales minerales deMurashige et. aI. (1962) diluidas 50%,suplementado con: Cisteína 10 mg, tiamina 10mg, ácido nicotínico I mg, glicina I mg, ácidonicotínico I mg, Mio-inositol 200 mg, caseínahidrolizada 400 mg, extracto de malta 400 mg,adenina 40 mg, BAP I mg, sacarosa 40 g, pH 5.6,Fitagel 2.5 g. Luego de 8 semanas, aparecen losembriones somáticos sobre los tejidos queenvejecen. Los embriones que se desarrollan,pasan por todas las etapas clásicas de un embriónformado por sernilla, o sea, globular, torpedo ycotiledonar.Germinación de embriones y formación devitroplantas, ambas etapas se desarrollan en luzdirecta (2000 lux), en fotoperíodo de 12 horas luz,12 obscuridad, durante 3 o 4 meses hasta formarvitroplantas aclimatables. Se utiliza un medio decultivo constituido por las sales minerales deMurashige et ai (1962) completas, suplementadocon: vitaminas de rnorel 2 ml, BAP I mg,sacarosa 40 g, pH 5.6, Fitagel 2.5 g.En todas las etapas el medio de cultivo seesteriliza en autoclave, a 120 grados centígrados.a 15 Ibs de presión por pulgada cuadrada, durante20 minutos. La incubación en todas las etapas fuea 29 ± 2 °C Y 2000 lux de iluminación en losperíodos requeridos.
2.2 Aclimatación de vitroplantasEs eJ proceso de acondicionamiento de lasvitroplantas, ai ambientales externo. Para esta fasese construyó una estructura especial deaclirnatación, con un techo de laminas alternandouna de zinc y una transparente; bajo este techo seestablecieron 4 mini-invernaderos con estructurade tubo PVC de 5 em, como base para la tela depolietileno transparente. Dentro de cada miniinvernadero se colocó un sistema de nebulizaciónautomatizado. que permite la regulación de lahumedad relativa (Figura I).
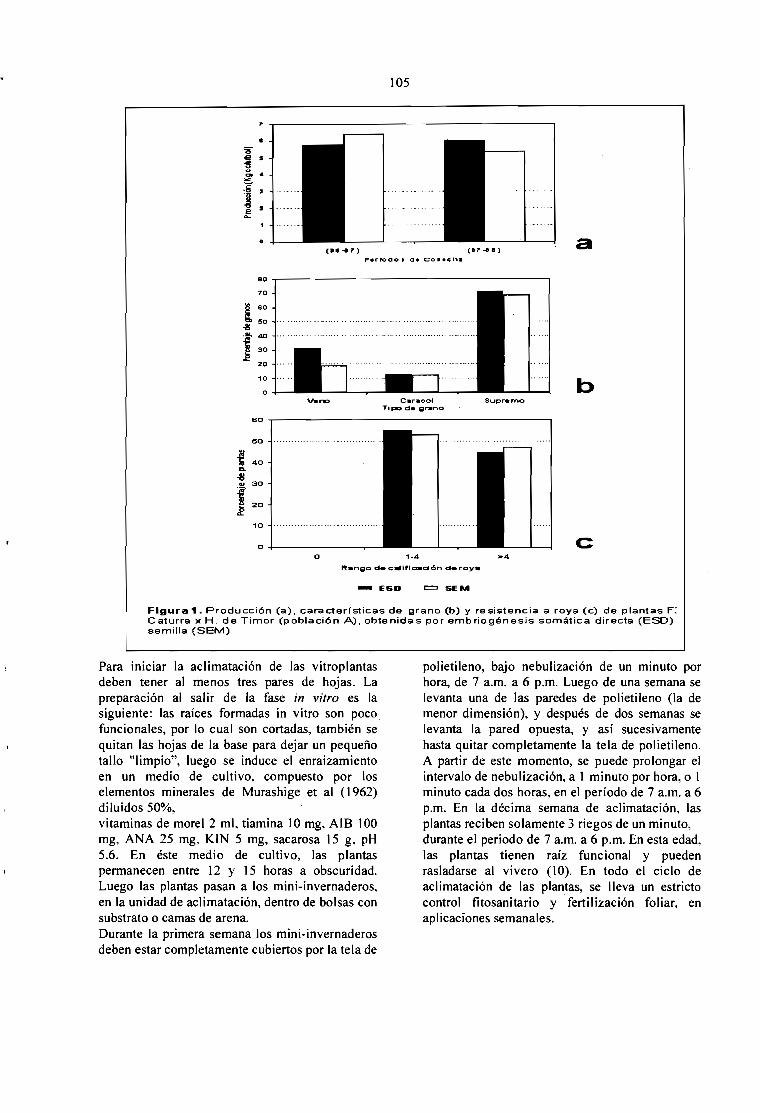
105
~e •~"" .'"-6 •s~
a(8" "") c·, -li.)
Perrodo I O. Co •• o".
80
70
! &0
~ so.",- 40i 30
10
bo\hino Caraool Supremo
Ti J=lO d. gnllno
60
eo
~ 40....~OI 30
f 2015"-
10
O CO 1-4 ~4
R.ngo d. o"ltl c.d 6n d. roy.
- ESO = SEM
FlguI"a 1. Producci6n (a), caracterlsticas de grano (b) y resistencia a roya (c) de plantas F:Cáturra )( H. de Timor (poblaci6n A). obtenidas por embriogénesis somática directa (ESQ)semilla (SEM).
Para iniciar la aclimatación de las vitroplantasdeben tener ai menos tres pares de hojas. Lapreparación ai salir de la fase in vitro es lasiguiente: las raíces formadas in vitro son pocofuncionales, por lo cual son cortadas, también sequitan las hojas de la base para dejar un pequenotallo "limpio", luego se induce el enraizamientoen un medio de cultivo, compuesto por loselementos minerales de Murashige et ai (1962)diluidos 50%,vitaminas de morei 2 ml, tiamina 10 mg, AIB 100mg, ANA 25 mg, KIN 5 mg, sacarosa 15 g, pH5.6. En éste medio de cultivo, las plantaspennanecen entre 12 y 15 horas a obscuridad.Luego las plantas pasan a los mini-invernaderos,en la unidad de aclimatación, dentro de bolsas consubstrato o camas de arena.Durante la primera semana los mini-invernaderosdeben estar completamente cubiertos por la tela de
polietileno, bajo nebulización de un minuto porhora, de 7 a.m. a 6 p.m. Luego de una semana selevanta una de las paredes de polietileno (Ia demenor dimensión), y después de dos semanas selevanta la pared opuesta, y así sucesivamentehasta quitar completamente la tela de polietileno.A partir de este momento, se puede prolongar elintervalo de nebulización, a 1 minuto por hora, o Iminuto cada dos horas, en el período de 7 a.m. a 6p.m. En la décima semana de aclimatación, lasplantas reciben solamente 3 riegos de un minuto,durante el periodo de 7 a.m. a 6 p.m. En esta edad,las plantas tienen raíz funcional y puedenrasladarse ai vivero (10). En todo el ciclo deaclimatación de las plantas, se lleva un estrictocontrol fitosanitario y fertilización foliar, enaplicaciones semanales.

106
5 metros
DI SENO ING AMAURI MOLlNADIBUJO lora.d. J de Cntro UNIOAO DE ACLlMATACION, FINCA SUENA VISTA, ANACAFE
Figura 1. Unidad de Aclimatación con capacidad para 6,000 vitroplantas sobre camas de arena.
2.3 Fase de vivereSe trabaja en un vivero de tipo convencional,como el utilizado para plantas obtenidas porsemi lia. Es importante aI inicio no exponer lasplantas a una radiación solar directa. EI vivero esde bambú con techo de palma, que puederegularse fácilmente para dar mayores entradas deluz sobre las plantas. Las plantas aclimatadas secolocaron en bolsas de polietileno negroconteniendo un substrato con 30% de materiaorgánica.En el vivero las plantas pierden rápidamente sucondición "micro", desarrollando paulatinamentehojas de tamafío normal. Se continuaron lasaplicaciones semanales de fertilizante foliar yfungicidas. Transcurridos dos meses, se efectuóuna aplicación de fertilizante de liberación lentasobre el substrato. Después de 6 o 7 meses envivero, Ias plantas se trasladan a campo.
3.RESU LT ADOS Y DlSCUSIÓN
Para la reproducción vegetativa de ambos clonesde Robusta. Ia embriogénesis somática demostróser un método práctico, permitiendo la difusiónrápida de estos materiales y su establecimiento encampo. EI método convencional que utilizaesquejes o estacas (de brotes ortotrópicos), no eraviable para los objetivos dei proyecto.Los clones "padres" de Nernaya, respondieronpositivamente ai protocolo de desarrollo deembriones somáticos, a pesar de algunosinconveniente por el crecimiento desincronizado
de los embriones in vitro. Los genotipospresentaron una buena tasa de producción deembriones. Bajo condiciones in vitro, el clonT3751 desarrolla más lentamente, y presentamayor brotación basal que el clon T3561. Estacaracterística también se observa en campo, por loque se considera de orden genotípico.En Guatemala, el método de aclimataciónempleado presenta una eficiencia de 90 porciento, en la relación, plantas de viveroútiles/vitroplantas. Con plantas de vivero biendesarrolladas, se han establecido aI momento 2hectáreas como campo semillerista, de un total de8 hectáreas programadas para el afio 1999. Paraesta actividad se cuenta con 15,000 vitroplantasen diferente estado de desarrollo en laboratorio yla unidad de aclimatación. AI iniciarse laproducción comercial de semilla híbrida en estoslotes, la variedad portainjertos Nemaya (C.canephora) estará disponible para que loscaficultores guatemaltecos la utilicen en susprogramas de injertación.
4.REFERENCIAS
ANZUETO, F., 1993. Etude de la resistance ducafeier (Coffea spp.) a Meloidogyne spp. yPratylenchus sp. These doct. Ecole NationaleSuperieure de Rennes, France. 123 p.ANZUETO, F.. BERTRAND, B., DUFOUR, M.1995. "NEMAYA", Desarrollo de una variedadporta-injerto resistente a los principales

nemátodos de América Central. Boletín IICAPROMECAFE, Guatemala. pp 13-15.ANZUETO, F., ESKES, AB., SARAH, J.L.,DECAZY, B. 1993. Résistance de quelquesdescendances de Coffea arabica y ç. canephoravis-à-vis de deux populations de Meloidogynespp., originaires du Guatemala et du Brésil.Association Scientifique Intemational du Cafe,42, rue Scheffer, 75116 Paris, Francia, ASIC n°15, pp: 338-349.ANZUETO, F., VILLAIN, L. & MOLINA, A.1996. Informe grupo técnicoPROMECAFEIANACAFE, Proyecto Regional deResistencia a los Nemátodos en CentroaméricaPROMECAFE-I1CA-CATIE/CIRAD. Guatemala,febrero 1996. 12p.BERTRAND, B., ANZUETO, F., PENA, M.,ANTHONY, F., ESKES, AB. 1995. GeneticImprovemente of Coffee for Resistance to Rootknot Nematodes (Meloidogyne spp.) in CentralAmerica. In: ASIC n? 16, Kyoto, Japon.Association Scientifique Intemational du Café,42, rue SchefTer, 75116 Paris, France.BERTRAND, B.1994. Informe anual deActividades de PROMECAFE-CICAFE-ClRAD.CICAFE, Costa Rica.CALDERON, V.M. 1989. Reacción de diferentesgenotipos de café a Meloidogyne arabicida, Lópezy Salazar (1989), gama de hospedantes y hongosfitopatógenos asociados. Tesis M.Sc. Turrialba,Costa Rica, CATIE. 71p.DUBLIN, P., 1984. Techniques de reproductionvegétative in vitro et amelioration genetique chezles caféiers cultivés. Café Cacao Thé, 28 (4): 231244.ETIENNE H., BERTHOULY, M. 1997.Comunicación personal, visita de asesoría aiLaboratorio de Cultivo de Tejidos de ANACAFE.Guatemala.ETIENNE H., SOLANO W., PEREIRA A,BERTRAND B. & BERTHOULY M. 1996.Protocolo de Aclimatación Para Vitroplantas deCafé. CATIE, CIRAD, PROMECAFE, CICAFE.Laboratorio de Biotecnología, CATIE, CostaRica, octubre 1996.
107
ETIENNE H., SOLANO W., PEREIRA A.,BARRY-ETIENNE O., BERTRAND B.,ANTHONY F., CÔTE F. & BERTHOULY M.1997. Utilización de la embriogénesis somática enmedio líquido para la propagación masal de loshíbridos FI de Coffea arabica. In: Memorias
~----
XVIII Simposio Latinoamericano de Caficultura,Costa Rica 1997, ICAFE, I1CA, PROMECAFE. P253-261.HERNANDEZ, A 1997. Étude de la variabilitéintra et interspécifique des nématodes du gemeMeloidogyne parasites des caféiers en AmériqueCentrale. These doct. Université de Montpellier 11Sciences et Techniques du Languedoc, France.102p.MORERA, N. 1990. 1986. Evaluación de lainteracción entre genotipos de Meloidogyneexigua Goeldi (1887) y Coffea sp. Tesis M.Sc.Turrialba, Costa Rica, CATIE. 59p.PENA, M.X.1994. Evaluación fenotípica y genética para laresistencia aI nemátodo Meloidogyne incognita enhíbridos de Coffea canephora. Tesis M.Sc.Turrialba, Costa Rica, CATIE. 78p.REYNA, E.H. 1966. Un nuevo método deinjertación en café. Boletín técnico # 21. DirecciónGeneral de Investigación y Control Agropecuario,Ministerio de Agricultura, Guatemala. 21 p.SONDAHL M., SHARP W.R., 1977. Highfrequency induction of somatic embryios incultured leaf explants of Coffea arabica L. Z.Ptlazenphysiol. Bd., S. 81: 395-408.ZAMARRIPA A., DUCOS J.P., TESSEREAUH., BOLLON H., ESKES, A.B. & PETIARD V.1991. Mass propagation of coffea spp by somaticembryogenesis in liquid medi um. In: 14th
International Scientific colloquium on coffee. SanFrancisco, E.U. 14-19 July 1991. P. 392-402.


109
FUSÃO INTERESPECÍFICA DE PROTOPLASTOS EM Coffea
Cordeiro, AT 1; Zambolim, e; Pétiard, v' Spiral, JJ
'DBV/UFV. 2DFP/UFV: Av. P.H. Rolfs. s/n - 36.571.000 - Viçosa-MG/Brasil)3CRN - 101, Av. Gustave Eiffel. 37.390 - Notre Dame d'Oé, France)
ABSTRACTlntrogression ofnematode resistancefrom Coffea canephora cv. Apoatã was attempted in diploid (DH1) andtetraploid (Catimor and Catuai Amarelo) C ara bica genotypes by protoplast fusion to evaluate thefeasibility ofinterspecific protoplast fusion as a tool in the parassexual improvement in Coffea. Protoplastswere isolated from embryogenic cell suspensions initiated from leaf explants-derived friable embryogeniccalli. The kinetics of growth and protoplast yield of Apoatã and DH j cell suspensions was evaluated atdifferent intervals of subcultures and conditions of enzymatic digestion of the cell wall of their small cellaggregates. lnvestigations concerning selection ofprotoplast heterofused-derived cell aggregates were alsoaccomplished lmmediately to the subculture. Apoatã and DHj cell suspensions showed a typical sigmoidalgrowth curve. Coincidentally. the lowest protoplast yields were observedfor post-subculture intervals abovethe injlection point. High protoplast yields were achieved when the two pectinases were combined tocellulase, at the highest tested concentrations. The twelve protoplast culture media tested did not allow anydistinction between parental protoplasts Apoatã and DH3• at microcalli growth stages. Alternatively. themetabolic inhibitors iodoacetamide and rhodamine, at the concentrations 2 and 0.9 mM. respectively. wereeffective in inhibiting microcalli formation from cultured parental protoplasts. The inhibitors allowed theaccomplishment of 23 independent interspecific electrofusion assessments, whose products led to thedevelopment ofembryogenic differentiation. Further work wil! be carried out in order to characterize theputative heterofused regenerants and to examine the potential of somatic hybridization as a to01 forparassexual improvement in Coffea.
INTRODUÇÃO
Na cafeicultura. elevadas produtividades comreduzidos custos de produção e preservaçãoambiental requerem material genético queexpresse. além da alta capacidade de produção defrutos com maturação uniforme. resistências àspragas e doenças. As suas inexistências naespécie Coffea arabica têm estimulado a pesquisano sentido das suas transferências. a partir deoutras espécies doadoras de Coffea. paravariedades arábicas. Porem. as instabilidades.genética e cromossômica. e o ciclo de seleçãorelativamente longo têm retardado estasobtenções. que podem ser antecipadas com atécnica de fusão de protoplastos. Sua utilizaçãoentre duas populações distintas de protoplastosproduz parentais não fusionados, homofusões eheterofusões, simétricas (Gaikwad et al., 1996) ouassimétricas (Fahleson et aI.. 1997). Estes últimosprodutos. que representam geralmente menos doque 10% da mistura total fusionada (Waara eGlimelius, 1995). devem ser separados de umapopulação inicial de aproximadamente 5x 105
protoplastos. Para tanto. pode-se fusionarprotoplastos parentais metabolicamenteinativados, cada um com um inibidor de açãodistinta do outro (Bõttcher et al., 1989).permitindo a seleção por complementação
metabólica que libera apenas os heterofusionadosdo impedimento ao desenvolvimento.
MATERIAIS e MÉTODOS
Os protoplastos foram obtidos mediante digestãoenzimática de agregados cel ulares de culturaslíquidas embriogênicas de genótipos canéfora earábicas. O genótipo canéfora foi o cultivarApoatã IAC LC 2258. resistente à Meloidogvneexigua (Fazuoli. 1986) e às raças 2 (Mazzafera etal., 1989) e 3 (Lima et aI.. 1989) de M. incogntta.Os arábicas testados foram o diplóidepartenogenético DH3 e os tetraplóides Catimor eCatuaí Amarelo. À exceção de quando se estudouo rendimento protoplástico (R p) em resposta aotempo de digestão e à concentração das enzimascelulase e pectinases. utilizou-se sempre a soluçãoenzimática para obtenção de protoplastos (SEOPlcitada por Spiral e Pétiard (1991). A pressãoosmótica desta solução. bem como de todasaquelas utilizadas com protoplastos, foi fixada em- 1.537 MPa (Sõndahl et al., 1980), ao perm itir-sevariar apenas a concentração de manitol. Osinibidores metabólicos. rodamina eiodoacetarnida. eram adicionados antes dos 30rnin finais da digestão. Uma vez purificados. osprotoplastos expostos ou não à eletrofusão emcâmara de fusão constituída de um sistemamultieletrodo (Sihachakr et al..1988). eram
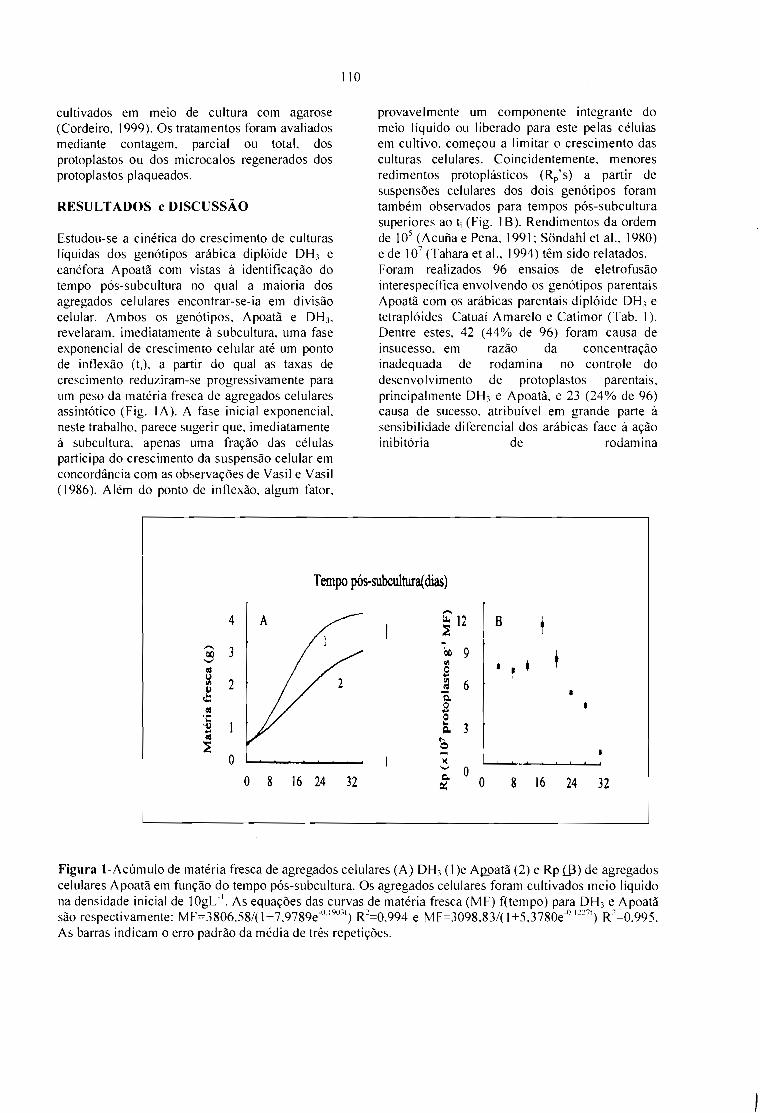
cultivados em meio de cultura com agarose(Cordeiro, 1999). Os tratamentos foram avaliadosmediante contagem, parcial ou total, dosprotoplastos ou dos microcalos regenerados dosprotoplastos plaqueados.
RESULTADOS e DISCUSSÃO
Estudou-se a cinética do crescimento de culturasliquidas dos genótipos arábica diplóide OH, ecanéfora Apoatã com vistas à identificação dotempo pós-subcultura no qual a maioria dosagregados celulares encontrar-se-ia em divisãocelular. Ambos os genótipos, Apoatã e OH"revelaram, imediatamente à subcultura, uma faseexponencial de crescimento celular até um pontode inflexão (ti)' a partir do qual as taxas decrescimento reduziram-se progressivamente paraum peso da matéria fresca de agregados celularesassintótico (Fig. 1A). A fase inicial exponencial.neste trabalho, parece sugerir que, imediatamenteà subcultura, apenas uma fração das célulasparticipa do crescimento da suspensão celular emconcordância com as observações de Vasil e Vasil( 1986). Alérn do ponto de inflexão, algum fator.
110
provavelmente um componente integrante domeio liquido ou liberado para este pelas célulasem cultivo. começou a limitar o crescimento dasculturas celulares. Coincidentemente, menoresredimentos protoplásticos (Rp's) a partir desuspensões celulares dos dois genótipos foramtambém observados para tempos pós-subculturasuperiores ao ti (Fig. 1B). Rendimentos da ordemde 105 (Acufia e Pena, J991; Sõndahl et al., 1980)e de 107(Tahara et al., 1994) têm sido relatados.Foram realizados 96 ensaios de eletrofusãointerespecífica envolvendo os genótipos parentaisApoatã com os arábicas parentais diplóide OH, etetraplóides Catuaí Amarelo e Catimor (Tab. 1),Dentre estes. 42 (44% de 96) foram causa deinsucesso, em razão da concentraçàoinadequada de rodamina no controle dodesenvolvimento de protoplastos parentais,principalmente OH, e Apoatã, e 23 (24% de 96)causa de sucesso. atribuível em grande parte àsensibilidade diferencial dos arábicas face à açãoinibitória de rodamina
Tempo pós-subcultura(dias)
4 ""'~ 12 B~
""'-
C() 3 bo 9~'-' <11 .,.tO 3o
<11 2 <11 6~tO
~ Q,tO o...'I:: e'Qj 3... ÇoGIl r-~ o
o l('-' oo 8 16 24 32 : o 8 16 24 32
Figura l-Acúmulo de matéria fresca de agregados celulares (A) OH, (l)e Apoatã (2) e Rp@) de agregadoscelulares Apoatã em função do tempo pós-subcultura. Os agregados celulares foram cultivados meio líquidona densidade inicial de 10gL'I. As equações das curvas de matéria fresca (MF) f(tempo) para OH, e Apoatãsào respectivamente: MF=3806.58/(1+7.978ge'IJ'<J03t) R2=0.994 e MF=3098.83/(1+5.3780e'OI227t) R2=0.995.
As barras indicam o erro padrão da média de três repetições.
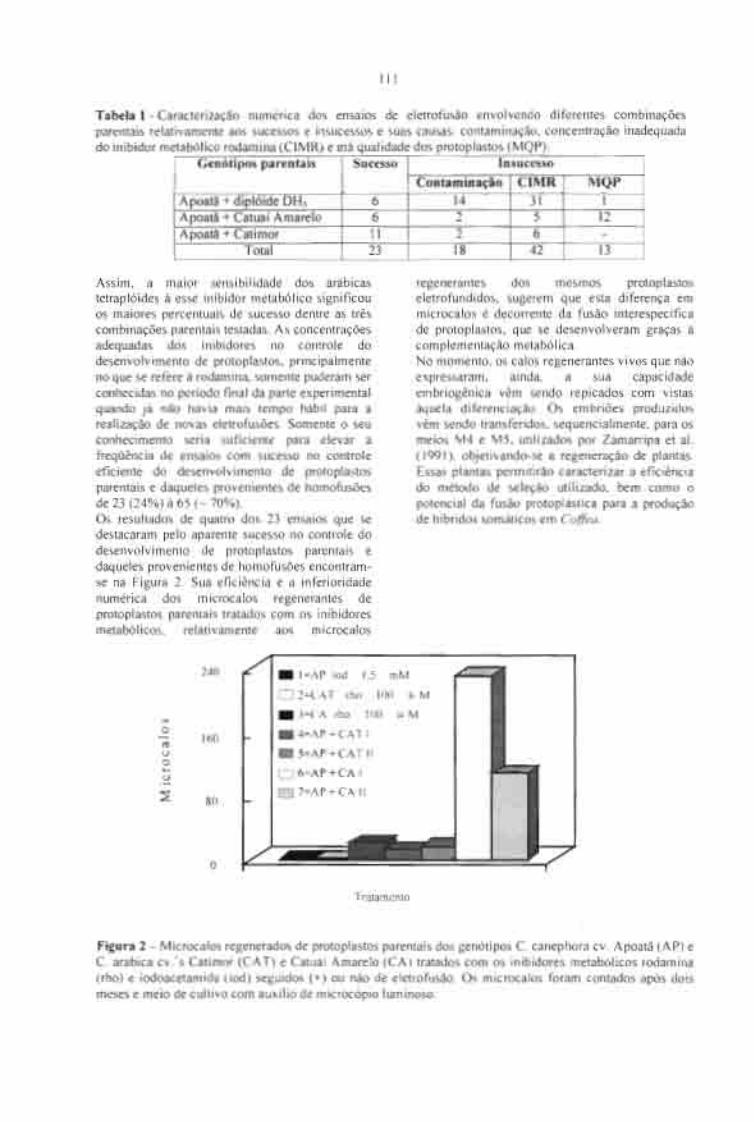
",
............... --....""_....., ........ '" ....."" .""""....
..«"',... , ""'.....- J , ''''''''~" ..... l"" '''''''''''' q.... _."',.."" I·...' •, ........" .., .., ..~.MI • •"" __""" '" ,.•"'_ "'1<""''"'" .,,'" , ,, ,'-"",, .."'" . .... ,..."' ."",,,.,,.mu _ ,..,_ _ ",m .~I..
.. , h _" ..._ --- "'_ ..._"" ,,., "'. __ 1_ .."""l _,,_ _ _l _ , r~_..do __ " .-...- "'.. __ •_,oi.~ ...-.......p.... ,..,......,..._....._.......,~
I II __Lll-----L""im, • m." """".,,,.. __~""'''1'''''''', 'b__" '~ I "o •..,...r",...'" " _ <l< .., "'''''' ....,om_ "'... """" A.<oo<'..' '''''''~ -'"- .., ,,.,- ...........ot>_ ""_"... """, ..._ ......_ _-_....~ _,....,._ ,- ..-...... _--_.-......._.._........ _.-......," _. .._,... _..._ _,-,...... --...-_ _._.~,.......... .~ ......_"' .. "...."... ).),:' , -,, ~ , ,,, ..-.,;o..:1 _ .......1<;"' ,. " " "" -.."' """oi< '"-''',''''''" -.. ~-'... ,"",,U<I<-. """ ,,," "",,..,,.. " ,,,, . ,,,,,,,,•
.. "' f 'l"" : ""' .II.M·"", '""',"'''''''''
.........>C . doi '" """' ''' _,,", 1< ..---~,,-" '" _...--.. ....."- "."""-
-,"
,• ,.••,, .'
: .... .... I'~ ... ... ,, - , ........"',....~......' -,"'<"
llII ,....("
,.
,
. ",... J ..~__...._"... c u •..,...... " ,,_,OPI.e..- ',,'_ ,r~" .1'..... · ....10 ,I'" n_""" '" ".-"_""'" ,,,,,",,, ,,.., __, , _ 1"._"'''''''_ , '''' ,_ ............. _ ..........- ,10......,,"'_ ,..._

112
REFERÊNCIAS
Acufía R & De Pena M 1991. Plant Cell Rep ..10:345-348.
Bõttcher U: Aviv D; Galun E 1989. Plant Sei..63:67-77.
Cordeiro. AT 1999. Tese (Doutorado emFisiologia Vegetal) - Universidade Federal deViçosa. 1999.
Fahleson J: Langercrantz V; Mouras A;Glimelius K 1997. Plant Sei..123:133-142.
Fasuoli LC 1986. In: A.B. Rena et aI. (eds),Cultura do Cafeeiro - fatores que afetam aprodutividade. Piracicaba. SP. AssociaçãoBrasileira Pesq. Potassa Fosfato. p. ~7- 113'3 4
Gaikwad K; Kirti PB; Sharma A: Prakash S;Chopra VL 1996. Plant Breeding. 115:480483.
Gamborg OL; Miller RA: Ojima K 1968. Exp.Cell Res.. 50: 151-158.
Lima MMA; Gonçalves W: Fazuoli LC: TristãoRO 1989. In: Congresso Brasileiro dePesquisas Cafeeiras. 15. Maringá, 1989.Resumos. Rio de Janeiro.COTECIDIPRO/IBC. p. 128-129.
Mazzafera P: Gonçalves W; Fernandes JAR 1989.In: Congresso Brasileiro de PesquisasCafeeiras. 15. Maringá, 1989. Resumos. Riode Janeiro. COTEC/DIPRO/IBC. p. 128-129.
Morei G & Wetmore RH 1951. Am, J. Bot..38:141-143.
Ohgawara J; Kobayashi S; Ohgawara E;Uchimiya H: Ishii S 1985. Theor. Appl.Genet.. 71: 1-4.
Power JB; Berry SF; Freason EM; Cocking EC1977. Plant Sei. Lett.. 10:1-6.
Sidorov VA; Menczel L; Nagy F; Maliga P 1981.Planta. 152:341-345.
Sihachakr D; Haicour R; Serraf I: Barrientos E;Herbreteau C; Ducreux G; Rossignol L;Souvannavong V 1988. Plant Sei.. 57:215223.
Sõndahl MR; Chapman MS: Sharp WR 1980.Turrtalba, 30: 161-165.
Spira] J & Pétiard V 1991. In: ColloqueScientifique International sur le Café. 14.San Francisco. 1991. ASIC (Paris). p.383-390.
Tahara M; Yasuda T; Uchida N; Yamaguchi T1994 .. Hortscience, 29: 172-174.
Vardi A; Frydman-Shani A; Galun W; Gonen P;Bliechman S 1990. Aeta Hort., 280:239-245.
Vasil V & Vasil IK 1986. J. Plant Physiol..124:399-408.
Waara S & Glimelius K 1995. Euphytiea.85:217-233.
Yasuda T: Fujii Y; Yamaguchi T 1985. PlantCell Physiol.. 26:595-597.

~GOVERNO DO
pARANftiSecreta ria da Agricultura
e do Abastecimento
~+~WJR
Instituto Agronómico do Paraná
PROGRAMA CAFÉ• Alaide Ap. Krzyzanowsk . Ncmatulcgia• Alcx Carneiro Leal, Agrossllviculrura
• Amador Villacorta iVJ., Entomologia• Ana Mariil Mencguim, Entomologia
• Arnaldo COIOIZi Filho. Microbiologia de solos• Arma ndo Androciolo Filho. Fitotecnia• CH ios Armênio Khatounian , Fítotecni a• Ca rlos Augusto P. Motta. Propagacâo veuet n!
• Ca rlos Roberto Ricdc , Gen ética• Ce lso J amil Marur, Ecofislolcg! a
• Dimas Soa res Jr.. Sócio -e con omln• Ed rnilson G. Liberal, Difusào de'Tecnologfu• Elcio C. Bat ota, l\ l icrobiologia de solos• Jom ar da Paes Pereira, Fiiorecnia• Júlio C. D. C haves. Manej o de solos e nu tr ição• Luiz Felipe P. Pereira . Biotccn ologia• Luiz Gonzag a E. Vieira . Biotc cnologta
• j\larros A . Pa van . Solos e nuu-lç âo• Murco'S Valcntln. Difusão de tccnoto gi»• Mari a B. Scholz, Bioquirnica r Ter . d e alimentos
• Maria C . L. Dia s. Prop ag ação n'~el al
• Maria Lúcia Crochemore. Biotecn olog}a• Mário Miyazawa . Qu ímica de solos• Nelson Fonseca Jr.• Genética• Paulo H. Curamorl. Agromete oroln gia• Rodclfo Bianco, Eruomoloaia• Rogério J\1.L. Cardoso, Fitopat ologia• Rogerio Teixeira Farias. Eugcnh ar! a ag ríco la• Rubens Siqueira. Engenhartn agrt cola• Rui Gomes Carneiro. Ncmatclop!a• Rui Pereira Leite. Fltopntologl a• Sonia M. Torrecillas, Entomologia
• Turnoru Ser a, Genética


115
MARCADORES DE DNA PARA MELHORAMENTO DO CAFEEIRO
Ney S. Sakiyama
Universidade Federal de Viçosa Departamento de Fitotecnia 36571-000 Viçosa, MG
INTRODUÇÃO
A cafeicultura mundial tem sido grandementebeneficiada pelo sucesso dos programas demelhoramento genético, que têm colocado àdisposição do agricultor cultivares produtivas eadaptadas a condições específicas de cultivo. NoBrasil, por exemplo, as cultivares melhoradas decafé arábica (Co./fea arabiea L.) produzem trêsa quatro vezes mais que as cultivares utilizadasno passado. Entretanto, tal sucesso elevou aprodutividade dessa espécie a um platôaparentemente difícil de ser superado. Tome-se,como exemplo, as produtividades das linhagensmelhoradas de 'Mundo Novo', 'CatuaíVermelho' e 'Catual Amarelo', até hoje as maisprodutivas. Um dos grandes desafios para osmelhoristas é, portanto, superar asprodutividades das cultivares atuais.Em virtude da susceptibilidade da espéciearábica a várias doenças e pragas, outro grandedesafio para os melhoristas é a transferência degenes de resistência e tolerância. Apesar de C.arabiea possuir o dobro do número decromossomos das demais espécies do gênero,algumas hibridações interespecíficas têm sidoexploradas com sucesso pelos pesquisadoresobjetivando essas introgressões. A ocorrênciamundial da ferrugem-do-cafeeiro (Hemileiavastatrix Berk. et Br.), por exemplo, e osprejuízos por ela causados, despertaram ointeresse dos melhoristas na busca de cultivaresde café arábica resistentes, por meio dehibridação natural ou artificial com Co./feaeanephora Pierre.Em café, como em outras plantas, omelhoramento por hibridação pressupõe aidentificação da variabilidade genética quanto àscaracterísticas a serem melhoradas. a escolha dashibridações mais promissoras, e a aplicação demétodos que favoreçam a seleção de híbridos,ou linhagens, ou populações superiores. Nessecontexto, tecnologias de marcadores de DNAtêm sido propostas como instrumentos úteis aosprogramas de pesquisa. Especialmente em café.por tratar-se de uma cultura perene e de períodojuvenil longo, essas tecnologias podembeneficiar o melhoramento genético de formapotencialmente grande.
MARCADORES DE DNA
o termo marcador genético é usado paraidentificar qualquer fator morfológico,fisiológico ou molecular que possibilite adiferenciação entre dois genótipos e que sejaherdável por reprodução sexuada. De modogeral, os marcadores genéticos são expressõesfenotípicas das diferenças genéticas ex istentesentre dois indivíduos. Entre os marcadoresmoleculares, entretanto, há uma classedenominada marcadores de DNA, que resultamda manipulação direta das moléculas de DNA.Foram desenvolvidas várias tecnologias demarcadores de DNA, todas baseadas empolimorfismos de DNA existentes entregenótipos. Dependendo da tecnologia utilizada,o polimorfismo entre genótipos, em um dadoloeus, pode ocorrer de duas formas maiscomuns: na forma de presença de fragmentos deDNA em um indivíduo e ausência no outro, ouna forma de fragmentos de comprimentosdiferentes para os dois indivíduos. Taisfragmentos são separados por eletroforese evisualizados na forma de bandas, por meio detécnicas de coloração ou de hibridação comsondas marcadas. Ao conjunto de bandas dá-se onome de padrão eletroforético.Os marcadores de DNA podem ser qualificados,quanto ao tipo de herança, como marcadoresdominantes ou marcadores codominantes(Figura 1 e 2). Marcadores dominantes sãoaqueles que não possibilitam diferenciar umindivíduo heterozigoto de um indivíduohomozigoto para o alelo dominante, em um dadoloeus em estudo. É o que geralmente ocorre comos marcadores cujos polimorfismos se baseiamna presença (aleI o dominante) ou ausência (alelorecessivo) do fragmento de DNA. Osmarcadores dominantes são visualizados, então,na forma de presença ou ausência de bandas.Com esse tipo de marcador, uma população F]oriunda de autofecundação de indivíduo FIdiplóide heterozigoto apresenta segregação naproporção de 3:1 (3 indivíduos com a bandapresente : I indivíduo com banda ausente).Marcadores codominantes são aqueles quepermitem identificar um indivíduo heterozigotode qualquer indivíduo homozigoto para o loeusem estudo. É o que geralmente ocorre com osmarcadores cujos poli morfismos se baseiam na

Entre os exemplos de marcadores de DNA,podem ser citados: RFLP (Restriction FragmentLength Polymorphism), STR (Short TandemRepeats), SSR (Simple Sequence Repeats),RAPD (Randon Amplified Polymorphic DNA),SPAR (Single Primer Amplification Reaction),AFLP (Amplified Fragment LengthPolymorphism), etc.
Caracterização de padrão molecular (DNAjingerprinting) é a identificação do genótipo pormeio de marcadores de DNA. Os padrõesmoleculares permitem identificar diferenças esimilaridades entre indivíduos ao nível de DNA.O uso de padrões moleculares tem sidofreqüente nos estudos de diversidade genéticadas espécies presentes em bancos degermoplasma e na natureza. Essas informaçõesadicionais sobre a diversidade genética facilitama identificação de novos variantes na natureza aserem introduzidos nos bancos de gerrnoplasma,ou de duplicações a serem eliminadas. Orientam,também, a formação de subamostras dogerrnoplasrna, as chamadas coleções básicas(core collections), as quais devem representar,com o mínimo de redundância, o máximopossível de diversidade presente numa coleçãocompleta. Coleções básicas podem ser maisfacilmente transferidas aos diversos centros depesquisa para atender as necessidades dosmelhoristas. A manutenção de diferentescoleções básicas. em diversas localidades,representa também uma estratégia eficaz eeconômica de preservação da diversidadegenética. Em café, os marcadores de DNA têmsido utilizados para estudos sobre a origem ediversidade genética (Lashermes et al., 1999;1993; Orozco-Castillo et al., 1996; 1994;Teixeira et al., 1999a; 1999b). A Figura 3exemplifica os padrões eletroforéticos obtidoscom marcadores RAPD em café.Padrões moleculares podem ser utilizados comodescritores genéticos, para fins de identificaçãode cultivares. As limitações dos marcadoresmorfológicos utilizados para descrever ascultivares, de forma a dar-lhes adistingüibilidade prevista na Lei de Proteçãode Cultivares (Lei 9.456, de 25 de abril de1997), podem ser diminuídas pela inclusãode marcadores de DNA. Em café. marcadoresRAPD foram utilizados para estimar asdistâncias genéticas entre 16 linhagenscomerciais de C. arabica e. apesar da pequenavariabilidade total, verificaram-se locos
diferença de comprimento de fragmentos deDNA. Os marcadores codominantes sãovisualizados, então, na forma de bandas emposições diferentes; cada banda correspondendoa um alelo alternativo. Com esse marcador, umapopulação F2 oriunda de autofecundação deindivíduo FI diplóide heterozigoto apresentasegregação na proporção de 1:2:1 (I indivíduocom somente a primeira banda presente : 2indivíduos com ambas as bandas presentes: 1indivíduo com somente a segunda bandapresente). Excepcionalmente, um marcadortipicamente dominante toma-se codorninante,quando o heterozigoto é identificável por meioda quantificação do DNA presente na banda.Os marcadores de DNA se destacam peloelevado número em que podem ser obtidos, porapresentarem herança simples e porapresentarem resultados consistentes,independentemente das condições de ambientede cultivo da planta ou do tipo ou idade dotecido amostrado. Essas características são aindamais relevantes para a pesquisa com café, umacultura perene com período juvenil longo. Asavaliações com marcadores de DNA podem, porexemplo, ser realizadas em estádios iniciais dedesenvolvimento da planta, ou até mesmo nasemente, apresentando resultados idênticos aosque seriam obtidos na fase adulta. Os genótiposde café estudados freqüentemente têm idadesdiferentes ou estão localizados em ambientesdistintos e, mesmo assim, os resultados dessesmarcadores podem ser comparados. Eles têmsido utilizados em café para o estudo de padrãomolecular (DNA jingerprint) e ligação gênica(linkage).
Genitor I Genitor 2 Híbrido
I I Banda APolimórfica
I I I Banda BMonomórti
I ca
Figura I: Representação de padrão eletroforético de ummarcador dominante: o híbrido é igual ao genitor I ediferente do genitor 2.
Genitor I Genitor 2 Híbrido
I I Banda APolimórfica
I I Banda BPolimórlica
Figura 2: Representação de padrão eletroforético de ummarcador codorninante: o híbrido é diferente dos doisgenitores.
116
PADRÃO MOLECULARFINGERPRINn
(DNA

117
LIGAÇÃO GÊNICA (LlNKAGE)
Figura 3: Padrão eletroforético de 37 genótiposde café, obtidos por marcador RAPD.Foto: T.A.Teixeira.
Ligação gênica é o fenômeno ou padrão detransmissão de genes ligados . Genes ligados sãoaqueles localizados no mesmo cromossomo, osquais tendem a permanecer unidos durante areprod ução sexuada, ou seja, não apresentamsegregação independente . Marcadoresmoleculares podem ser estudados em função daligação entre si ou entre eles e os genes quecontrolam as características de interesse.Três possibilidades de uso da ligação gêníca sãoexploradas no estudo de marcadoresmoleculares: o mapa molecular, o mapeamentode genes que controlam característicasqualitativas e o mapeamento de regiõescromossômicas responsáveis por característicasquantitativas (Quantitative Trait Loci-QTL).Mapa genético é mapa cromossômico, queinforma sobre a ordem linear dos genes ao longode cada cromossomo e sobre as distâncias entreos genes medidas pelas probabilidades deocorrência de permutas. Os mapas sãoconstruídos com o propósito de local izarfisicamente as informações genéticas. Por meiodo mapa genético podem-se veri ficar as relaçõesexistentes entre as várias características deinteresse agron ômico.corno por exemplo. aocorrência de ligação gênica. pleiotropia,segregação independente das características. Atéo advento de marcadores moleculares, aconstrução de mapas genéticos era dificultadapela necessidade de um número considerável degenes de herança simples, com segregaçãofenotípica visível, e pelo grande número decruzamentos necessários para mapear cada novogene em estudo. Para a construção desses mapasutilizavam-se, com freqüência, genes mutantesem cruzamentos envolvendo séries deaneuplóides ou de trissômicos. Os marcadoresmoleculares apresentam várias vantagens naconstrução de mapas genéticos: comportam-se
li'• . ... . . . I'I •
polimórficos mesmo entre linhagens bastanteaparentadas (Teixeira et al., 1999).A possibilidade do emprego de dados dedivergência genética avaliada por marcadores deDNA para predizer a performance de híbridos FI
tem sido investigada. Essa divergência pode tervalor preditivo especialmente quando medidaentre linhagens melhoradas. Quando se utilizammarcadores tomados ao acaso, nem sempre adivergência máxima observada para um par degenótipos corresponde ao valor máximo deheterobeltiose no FI por eles gerado. A predição,portanto, toma-se mais confiável se os marcadoresutilizados estiverem associados às característicasque se desejam melhorar no hibrido e estiverempresentes de forma complementar no par degenitores.Os padrões moleculares podem ser utilizados naseleção assistida em retrocruzamentos visando arecuperação mais eficiente do genótipo recorrente.A introgressão de genes de interesse, como os quecondicionam resistência a doenças e pragas , éfreqüentemente realizada pelo uso do método deretrocruzamentos. Nesse método, a recuperaçãodo genétipo recorrente ocorre de forma gradual,sendo necessárias várias gerações deretrocruzamento. A seleção de indivíduosfenotipicamente mais semelhantes ao genitorrecorrente, associada à seleção assistida pormarcadores (Marker Assisted Selection-MAS),pode reduzir o número de retrocruzamentosnecessários para atingir o mesmo grau derecuperação desse genótipo. Se o mapa molecularé disponível, recomenda-se a escolha demarcadores uniformemente distribuídos ao longodo genoma. Quando se usam marcadoresdominantes para esse fim, devem-se selecionarmarcadores cujas bandas sejam presentes nogenitor doador e ausentes no recorrente. A cadageração de retrocruzamento, ocorre a fixação devários alelos do recorrente, que pode serobservada por meio de marcadores.Outras formas de utilização de padrõesmoleculares no melhoramento do cafeeiro sãopossíveis, como por exemplo: avaliação de taxa defecundação cruzada, confirmação da obtenção dehíbridos em cruzamentos artificiais; verificação dapureza genética de linhagens e clones, avaliaçãode variações somaclonais, etc.

como fatores mendelianos simples, a segregaçãoalélica independe de segregação fenotípicavisível, são estáveis e obtidos em grandenúmero, e podem gerar um mapa utilizando-seuma população segregante oriunda de um únicocruzamento. Em café, um mapa molecular foiconstruído a partir de uma população de dihaplóides de C. eanephora (Paillard et aI.,1996). Os mapas moleculares podem serutil izados na seleção assistida por marcadores(MAS), no mapeamento e clonagem de genessimples e no mapeamento de QTLs. O maiorimpacto da aplicação de mapas moleculares nomelhoramento de plantas, sem dúvida, advirá domapeamento de QTLs associados acaracterísticas de interesse agronômico.O mapeamento de genes que controlamcaracterísticas qualitativas de importânciaagronômica em cafeeiro pode ser útil aosprogramas de melhoramento pela sua utilizaçãona seleção assistida por marcadores. Esta seleçãoindireta tem aplicação em programas onde sepretende promover, por exemplo, a piramidaçãode genes de resistência a doença, a qual é dificilde ser executada pelos métodos convencionais.A seleção indireta simultânea de diferentescaracterísticas também é possível pela utilizaçãode marcadores. Marcadores de DNA foramidentificados para mapeamento da autoincompatibilidade em C. eanephora (Lashermeset al., 1996), e da resistência a Colletotriehumkahawae (Agwanda, et al., 1997) e a Hemileiavastatrix (Tedesco et al., 1999) em C. arabiea.Nesse último, busca-se o mapeamento de genescom vistas à piramidação de genes dominantesde resistência à ferrugem-do-cafeeiro emlinhagens melhoradas.Característica quantitativa é geralmente referidacomo aquela que é condicionada por váriosgenes de efeitos pequenos e semelhantes. O usode marcadores de DNA, entretanto, têmdemonstrado que as características quantitativaspodem ser condicionadas por loei de efeitosdiferentes. Os loei de efeitos detectáveis navariação de uma característica quantitativa sãochamados QTLs. Por meio do mapa molecular,é possível identificar QTLs que explicam partessignificativas das variações genéticas decaracterísticas quantitativas. Uma aplicaçãoimediata do mapeamento de QTLs emprogramas de melhoramento é a escolha degenitores que apresentem QTLscomplementares, os quais apresentarão maioreschances de manifestação de heterobeltiose noshíbridos e de recombinações favoráveis nasgerações segregantes. Marcadores associados aQTLs podem também ser utilizados em
118
programas de retrocruzamentos onde a seleçãoassistida por marcadores (MAS) é praticada.Uma nova e importante contribuição dos estudosde QTLs se refere à estratégia de avaliação eutil ização de germoplasma. Tanksley eMcCouch (1997) propõem a "procura dosgenes" (novo paradigma) em substituição à"procura do fenótipo" (velho paradigma). Aprocura do fenótipo de interesse agronômico temsido a estratégia tradicional de utilização dogermoplasma no melhoramento. Agora, ogermoplasma pode também ser avaliadodiretamente quanto à presença de genes úteis nosacessos, por meio de marcadores associados aQTLs, mesmo que esses loei não resultem emdiferenças fenotipicamente observáveis.
CONCLUSÃO
Nas palavras do Dr. Alcides Carvalho, "o Brasiljá foi um imenso cafezal oriundo de poucasplantas". As introduções de novos genótipostrouxeram significativos progressos aomelhoramento. As hibridações interespecíficastem sido exploradas no melhoramento, aexemplo das pesquisas visando a obtenção decultivares resistentes à ferrugem-do-cafeeiro eao bicho-mineiro. A utilização do germoplasmade café trará importantes benefícios aomel horamento pela transferência de genes deresistência a doenças e pragas e de QTLsasssociados a essas e outras importantescaracterísticas agronômicas, como aprodutividade. Nesse contexto, os marcadores deDNA deverão ter um papel importante nosprogramas modernos de melhoramento genéticodo cafeeiro.
REFERÊNCIA BIBLIOGRÁFICA
AGWANDA, C.O.; LASHERMES, P.;TROUSLOT, P.; COMBES, M.C.;CHARRIER. A. 1997. Identification ofRAPD markers for resistance to coffeeberry disease, Colletotriehum kahawae. inarabica coffee. Euphytica,97:241-248.
LASHERMES, P.; COMBES, M.C.; ROBERT,J.; TROUSLOT, P.; D'HONT, A.;ANTHONY. F.; CHARR1ER. A. 1999.Molecular characterisation and origin of theCoffea arabiea L.genome.MoI.Gen.Genet., 261 :259-266.
LASHERMES. P.; COUTURON. LMOREAU, N.; PAILLARD. M.;LOUARN,J. 1996. Inheritance and genetic mappingof self-incornpatibility in Coffea eanephoraPierre. Theor.AppI.Genet.,93:458-462.

LASHERMES, P.; CROS,J.; MARMEY, P. &CHARRIER, A 1993. Use ofrandomamplified DNA markers to analysevariability and relationships ofCof/easpecies. Genetic Resources and CropEvolution, 40:91-99.
OROZCO-CASTILLO, c.. CHALMERS, KJ.;WAUGH, R.; POWELL, W. 1994.Detection of genetic diversity and selectivegene introgression in coffe using RAPDmarkers. Theoretical and AppliedGenetics, 87:934-940.
OROZCO-CASTILLO. c. CHALMERS. K.J.;POWELL, W.; WAUGH. R. 1996. RAPDand organelle specific PCR re-affirmstaxonomíc relationships within the genusCo./fea. Plant Cell Reports, 15:337-341.
PAILLARD, M.; LASHERMES, P.; PÉTIARD.V. 1996. Construction of a molecularlinkage map in coffee.Theor.Appl.Genet., 93:41-47.
119
TANKSLEY, S.D. & McCOUCH, S.R. 1997.Seed banks and molecular maps: unlockinggenetic potencial from the wild. Science277: 1063-1066
TEDESCO. N.S.; SAKIYAMA, N.S.;ZAMBOLIM, L.; TEIXEIRA, T.A.;PEREIRA, A.A.; SAKIYAMA, C.C.H.1999. Mapeamento de gene de resistência aferrugem-do-cafeeiro (Hemileia vastatrixBerk. et Br.) com marcador RAPD. Anaisdo IJI SIBAC, Londrina. (em impressão).
TEIXEIRA, T.A.; ZAMBOLIM, L.;SAKIYAMA. N.S.; PEREIRA, A.A.;SILVA, D.G. 1999. Padrão molecular declones de cafeeiro diferenciadores deHemileia vastatrix Berk. et Br. Anais doIJI SIBAC, Londrina. (em impressão).
TEIXEIRA, T.A.; SAKIYAMA, N.S.;ZAMBOLIM, L.; PEREIRA, AA;SAKIYAMA, C.C.H. 1999.Caracterização de acessos de Co./fea pormarcadores RAPD. Anais do In SIBAC,Londrina. (em impressão).

•

121
GENETIC DIVERSITY AND MOLECULAR MAPPING OF COFFEE
Lashermes p.1; Combes M.C.1; Topart P.2
; Anthony F 2
I IRD, GeneTrop, BP 5045, F-34032, Montpellier, France2 CATIE. AP59, 7170 Turrialba, Costa Rica
The large number of named varieties andselections of arabica coffees hides the very narrowgenetic diversity of the base populations fromwhich they were selected (4). Historical evidenceindicates that these base populations ali descendedfrom the few trees that survived various efforts tospread arabica coffee from Southern Arabia, nowYemen, into the main coffee-producing areas. Thecultivars present therefore, a homogeneousagronomic behaviour characterised by a highsusceptibility to many pests and diseases, andvery low adaptability. Enlarging the genetic baseand improvement of arabica varieties havebecome a crucial priority toward an economic andsustainable coffee production. Spontaneousaccessions collected in the primary centre ofdiversity as well as wild relative Coffea speciesconstitute a valuable gene reservoir for breedingpurposes.IRD aims to develop basic knowledges andgenetic engineering strategies, required for anefficient and rational use of genetic resources. Inrecent years, molecular marker techniques havegained widespread applications in many fields ofplant genetics and breeding. Regarding C.arabica, several lines of research are concerned (2).
GENETIC RESOlIRCES
In addition to germplasm collection screening fordesired characteristics (3), molecular markeranalyses are performed (13). These geneticdiversity analyses have already increased ourunderstanding of the relationships among coffeecultivars and between the cultivars and thespontaneous accessions collected in the primarycentre of genetic diversity in Africa. We intend topro pose improved management practices ofcollection and to assist breeding programmes inthe identification of promising genitors.
GENOME CHARACTERISATION
C. arabica is the only polyploid species (2n =4x=44) in the genus. By combining molecularmarkers (6, 10, 12) and DNA in situ hybridisation,we sought to gain insights into its genomeorganisation (8, 1I). An amphidiploid structurewas demonstrated and the diploid progenitorspecies identified. Further work should determinethe degree to which the ancestral genomes haveevolved in the amphidiploid, the chromosomemeiotic pairing control, and the role ofintergenomic interactions in gene expression.
CalurraBourbon
Bourbon type Mbniz: t-----------,c:Jltivars 1.60
3058 ------------1K·7
3099
Typica type Blue Mountam t--------------..1cultivars Typica
EI2Et 16EI5---..1
Et24 1------,Ethiopian EI30subspontaneous EI 59accessions EI12
EI40EI41 --------1EI36 -------------1EI46 -----------.....1
oLinkage Distance
Fig. I Dendrogram 01'Coffea orobico access ions basedon RAPD data and generated by single linkage c1usteranalysis (13)
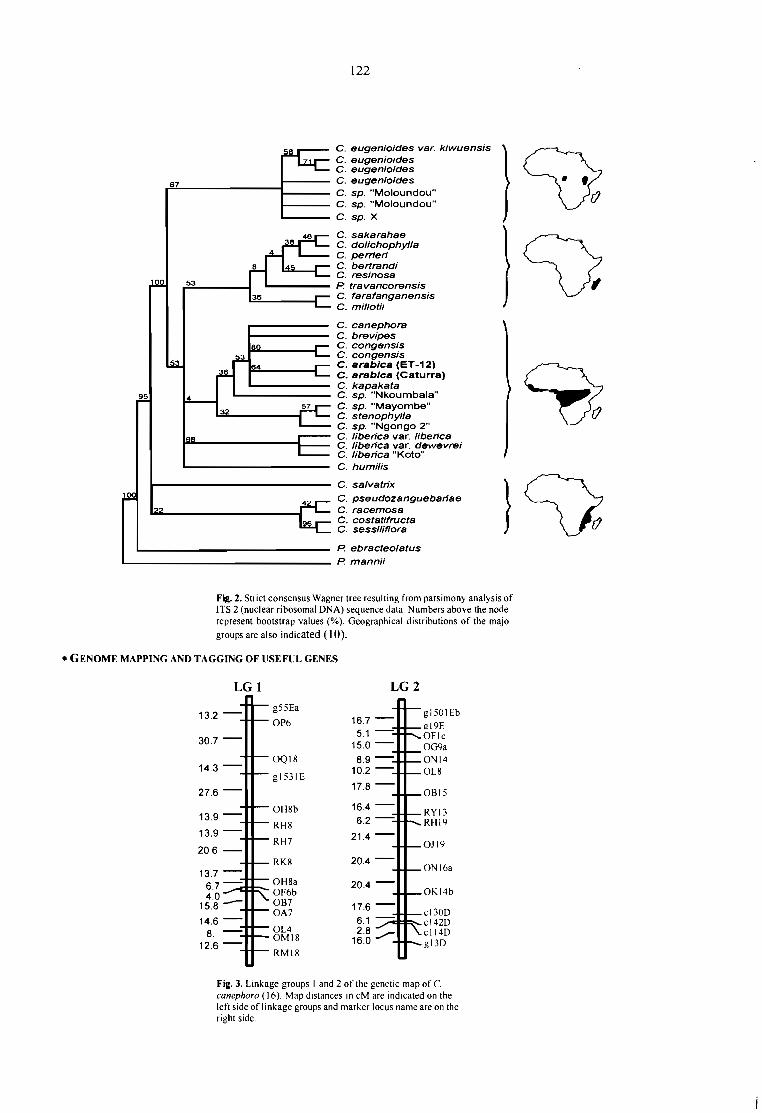
122
C. sekerene«C. doliehophylfaC. perriertC. bertrandiC. resinosa
....---- P. travaneorensisC. farafanganensisC. mittotii
53
67
C. eugenioides ver. kiwuensisC. eugenioidesC. eugenioides
I[i~ {===: C. eugenioidesr C. sp. "Moloundou"t---- C. sp. "Moloundou"L.. C. sp. X
95 4
3257
4
C. eanephoraC. brevipesC. eongensisC. eongensisC. arabica (ET-12)C. arabica (Caturra)C. kapakataC. sp. "Nkoumbala"C. sp. "Mayombe"C. stenophylfaC. sp. "Ngongo 2"C. tiberice var. uberic«C. ttberice var. oewevreiC. tiberice "Koto"C. humilis
C. setvetrix
C.pseudozanguebarlaeC. raeemosaC. eostatifruetaC. sessiliflora
}S)P. ebrecteotetueP. mannii
Fig. 2. Striet eonsensus Wagner tree resulting from parsirnony analysis ofITS 2 (nuclear ribosomal DNA) sequenee data. Numbers above the noderepresent bootstrap values (%). Geographieal distributions of the majo
groups are also indieated (10) .
• G ENOME MAPPING AND TAGGING OF USEFUL GENES
LG 1 LG2
13.2 -g55Ea gl501EbOP6 16.7-
gl9E5.1- OFle30.7- 15.0- OG9a
OQI8 8.9- ONI414.3 - 10.2 - OL8gl531E
27.6-17.8-
0815
OH8b 16.4-RYI313.9- 6.2- RHI9RH8
13.9- 21.4-RH7 OJI9
206 -RK8 20.4-
ONI6a137 -6.7- OH8a 20.4-4.0 OF6b OKI4b
15.8 - 087 17.6-OA7 el30D
14.6- 6.1- el42D8. -- OL4 2.8......- el14D
OM1812.6- 16.0 gl3D
RMI8
Fig. 3. Linkage groups I and 2 of the genetie map of Ccanephora (J 6) Map distanees in eM are indicated on lheleft side of Iinkage groups and marker loeus name are on theright side

A genetic linkage map of C. arabica is beingconstructed based on a F2 population derived froma cross between an improved cultivar and a wildtype. One objective is to determine the genomelocation and genetic contribution of genescontrolling important traits of coffee plant. Genecloning is also considered. Special emphasis isgiven to pest and disease resistances, qual ityrelated traits, and male sterility (7, 9). Monitoringof gene introgression to C. arabica from wilddiploid coffee species especially C. canephoraand Timor Hybrid derivatives, is expected to beconsiderably facilitated by the development ofmolecular marker-assisted selection (14, 15).RAPO markers closely associated with the T geneof resistance to Coffee Berry Oisease have beenalready identified (I).
REFERENCES
I. Agwanda, c.. P. Lashermes, P. Trouslot, M.C.Combes, and A. Charrier. 1997. Identification ofRAPO markers for resistance to Coffee BerryDisease, Colletotrichum kahawae, in Arabicacoffee. Euphytica, 97: 241-2482. Anthony, F., 8. Bertrand, P. Lashermes, and A.Charrier. 1997. La biologie moléculaire en appui àI'amélioration génétique du caféier Arabica.Plantations, recherche, développement, 6: 3693773. Bertrand, 8., G. Aguilar, E. Bompard, A.Rafinon, and F. Anthony. 1997. Comportementagronomique et résistance aux principauxdéprédateurs des Iignées de Sarchimors etCatimors (CofJea arabica L.), évaluées au CostaRica. Plantations, Recherche, Développernent, 5:312-3214. Carvalho, A., 1988. Principies and practices ofcoffee plant breeding for productivity and qualityfactors: CofJea arabica. In: R. J. Clarke and R.Macrae (Eds) Coffee vol 4 : Agronomy, pp 129165. Elsevier Applied Science, London.5. Couturon, E., P. Lashermes, and A. Charrier.1998. First intergeneric hybrids (Psilanthusebracteolatus H. x CofJea arabica L.) in coffeetrees. Cano 1. of Bot., 76: 542-5466. Cros, J., M.C. Combes, P. Trouslot, F.Anthony, S. Hamon, A. Charrier, and P.Lashermes. 1998. Phylogenetic analysis of
123
chloroplast ONA variation in CofJea L. MolecularPhylogenetics and Evolution, 9: 109-1177. Dufour, M., F. Anthony, 8. Bertrand, and A.B.Eskes. 1997. Identification et utilisation decaféiers rnâles-stériles de CofJea arabica enAmérique centrale. Plantations, Recherche,Développernent, 6: 401-4078. l.ashermes. P., M.C. Combes, P. Trouslot, F.Anthony and A. Charrier. 1996. Molecularanalysis of the origin and genetic diversity ofCofJea arabica L.: Implications for coffeeimprovement. Proc. of the EUCARPIA meetingon tropical plants, Montpell ier, 23-299. Lashermes, P., E. Couturon, N. Moreau, M.Paillard, and J. Louarn. 1996. Inheritance andgenetic mapping of self-incompatibility in CofJeacanephora Pierre. Theor. Appl. Genet., 93 : 458462.10. l.ashermes. P., M.C. Combes, P. Trouslot, andA. Charrier. 1997. Phylogenetic relationships ofcoffee-tree species (CofJea L.) as inferred fromITS sequences of nuclear ribosomal ONA. Theor.Appl. Genet., 94: 947-95511. l.asherrnes, P., M.C. Cornbes, J. Robert, P.Trouslot, A. D'hont, F. Anthony, and A. Charrier.1999. Molecular characterisation and origin oftheCoffea arabica L. genome. Molecular & GeneralGenetics, 261 : 259-26612. Lasherrnes, P., J. Cros, M.C. Cornbes, P.Trouslot, F. Anthony, S. Hamon, and A. Charrier.1996. Inheritance and restriction fragment lengthpolymorphism of chloroplast ONA in the genusCofJea L.. Theor. Appl. Genet. 93: 626-63213. Lashermes, P., P. Trouslot, F. Anthony, M.C.Combes, A. Charrier. 1996. Genetic diversity forRAPO markers between cultivated and wildaccessions ofCofJea arabica. Euphytica 87: 59-6414. Lashermes, P., S. Andrzejewski, B. Bertrand,M.C. Combes, S. Oussert, G. Graziosi, P.Trouslot, and F. Anthony. 1999. Molecularanalysis of introgressive breeding in coffee(CofJea arabica L.). Theor. Appl. Genet. In press15. l.ashermes, P., C. Agwanda, F. Anthony, M.C.Cornbes, P. Trouslot, and A. Charrier. 1997.Molecular marked-assisted selection: A powerfulapproach for coffee improvement. 17thconference of ASIC, Nairobi (Kenya), 474-48016. Paillard, M., P. Lashermes, and V. Pétiard.1996. Construction of a molecular linkage map incoffee. Theor. Appl. Genet. 93: 41-47


125
MICROSATELLlTES IN Coffea arabica: CONSTRUCTION AND SELECTION OF TWOGENOMIC LIBRARIES.
Vascotto F., Degli Ivanissevich S!, Rovelli P., Anthony F.2, Anzueto F.J
, Lashermes P.4, Graziosi G.
Dipartimento di Biologia. Università di Trieste, 34143 Trieste, ItalyI DPVTA. Università di Udine, Italy2 PROMECAFE Network, Guatemale City, Guatemala3 CATIE, Turrialba, Costa Rica4 IRD, Montpellier, France
INTRODUCTION.
Almost the entire world production of coffee isobtained by the cultivation of two species: Coffeaarabica and Coffea canephora. The two speciesprovide beans with distinct organoleptic qualitiesand C. arabica is generally preferred for itssmoother taste and reduced caffeine content.Moreover C. arabica accounts for about 70% ofthe total world coffee production (Clifford andWillson, 1985).The commercial importance of C. arabica is metby intense research activities at the agronomiclevel by selection and production of new varietiesand new cultivation techniques as for instance the"Cafe adensado" (Sera et aI., 1994). Neverthelessthe genetic improvement of the varieties as wellas the controlled introgression of valuable genesfrom other species is somewhat hindered by thelow knowledge on the genetics of C. arabica.Although some mendelian inherited loci areknown, as for instance Maragogype (Iarge beamsand leaves) or Caturra (short stature), they aretoa few for building up a genetic map and thelack of a genetic map has obvious effect onbreeding programmes.Polymorphic DNA loci represent the obviouschoice as marker for constructing a genetic mapand indeed a number of polymorphisms areavailable as RAPD (Lashermes et aI., 1996;Orozco-Castillo et al., 1994) and AFLP(Lashermes et aI., 1999). Nevertheless thepolymorphic bands detected by these twomethods show a dominant behaviour: one band iseither present or absent while intensity variationsare generally unreproducible. Therefore thesemethods do not allow for distinguishing betweenthe dominant homozygote and the heterozygote,both characterised by the presence of the samebands.More informative are the polymorphisms basedthe microsatellites: they show codominant bandsand therefore they allow for the identification ofali possible genotypes as well as easy transfer ofmarkers in different crosses.
These characteristics are of great relevance inconstructing a genetic map (Pai lIard et aI., 1996)as well as in marker assisted breedingprogrammes.We undertook this research project in view ofidentifying an characterising polymorphicmicrosatellites.
MATERIAL AND METHODS.
Plant material.The genomic libraries were constructed fromDNA extracted from leaves of a plant of C.arabica varo Caturra which is part of the IRD(Montpellier) collection. The polymorphismswere assessed on a F2 population of 12 plants
obtained from the cross of Caturra x WildEthiopia (EnO) and selfing of the F1.
Unfortunately the parental and the F I plants are
presently unaccessible and they could not betested for heterozygosity.
DNA extraction and purification.The DNA was extracted from young leaves, 2-3em, which were Iyophilised immediately aftercollection. Since the quality of a genomic librarydepends very much on the quality of the startingDNA, particular attention was paid for the initialDNA extraction and purification. We adopted amodification of the extraction technique ofMurray and Thompson (1980) and OrozcoCastillo et aI (1994), as reported in detailed in theprotocol below.- Grind 0.12 g of Iyophilised leaf in a rnortar, add3ml Iysis buffer (4% CTAB, 100mM Tris-HCIpH8.0, IAM NaCI, 20 mM EDTA, io.su: BMercaptoethanol for 15 ml of buffer, to be addedat the time of the extraction)- Following incubation for I h at 65°C, extractwith 2.25 ml CHLlIAA (24: 1) for 5'- Centrifuge 5' at 12000 rpm and transfer thesupernatant into a c lean tube; precipitate with 0.6volumes of isopropilic alcohol 10' at roomtemperature (1.8 ml for 3 ml of buffer)

- Centrifuge 15' at 12000 rpm, drain supernatantand dry the pellet very well (eventually at 65°C)- Resuspend in 250111 of TE and incubate at 65°Cfor 15' with 2.5111 RNAase A (\ mg/ml)Further purification of the extracts was performedas reported below.- Add 45 111 5M NaCI and shake by inversion;add 300 111 2% CT AB, 100 mM Tris pH 8.0, 1,4M NaCI, 20 mM EDT A and shake by inversion- Extract with 500 111 CHLlIAA wait 5' andcentrifuge 5' at 12000 rpm then transfer thesupernatant in a c1ean Eppendorf- Precipitate for I h with the addition of 1.5volumes of 1% CT AB, 50 mM TRIS pH 8.0 atroom temperature and centrifuge at 12000 rpm forlO'- After drying very well the pellet, remove theCTAB from the tube walls and resuspend in 75 1115M NaCI and 300 111 TE (incubate 10' at 56°C)and, if necessary, centri fuge 5' at 12000 rpm toeliminate undissolved particles- Precipitate the DNA with 950 111 ethanol 100%(wait 10' at RT), centrifuge at 12000 rpm for 10';wash the pellet with 500 111 ethanol 70%, wait 5'and centrifuge 5' at 12000 rpm- Dry the pellet at 37°C and dissolve in 50 111 TE(eventually at 56°C)
Construction and enrichment of the genomiclibraries.Two genomic libraries were prepared as describedby Rafalski et aI. (1996), Morgante et al. (1998)with modifications as reported below. One librarywas enriched in (TG)n and the other one was
enriched in (A TC)n. The main steps of the
procedure were the following:Caturra DNA digestion with Tsp5091
DNA fragment selection (250-700 bp fromagarose gels)Fragment ligation to an adapter containing anEcoRI site and adequate primer sequences.Enrichrnent in TG or ATC by DYNABEADSDYNAL conjugated with biotin-strepavidin01 igonuc leotides complementary to TG and ATC;PCR Amplification ofthe two libraries by primingthe adaptersControl of the success of the selection procedureby Southern blot using the TG and ATColigonucleotides as probes.Purification of the two libraries by the WizardPromega kit.Restriction of the genomic fragments by EcoRI.Selective recovery ofthe digested fragments (250700 bp) from agarose gel on DEAE NA45 filters.Fragment cloning in the EcoRI site of -ZAPII(Stratagene, La Jolla, Calif.).
126
Phage packaging and infection of XL l-BlueMRF' strain.Phage count and plating ofthe libraries.Lifting on nylon membranes.Screening of the libraries for the TG/ATCpositives clones.
Sequencing and primer designoThe sequencing of the positive clones wasperformed by Thermo Sequenase dye terminatorcycle sequencing pre-mix kit (AmershamPharmacia Biotec) following the producerind ications.Pairs of primers were designed on the regionsflanking the microsatellites. The on lineprogrammes Primer3 (Whitehead Institute forBiomedical Research, Cambridge, Massachusetts,USA) or Primers.' (Williamstone Enterprises)were used, choosing a Tm of a 58°C .The constanttail KS (5'- TCGAGGTCGACGGTATC-3') wasadded to one ofthe primers for each primer pairo
Amplification and analysis of themicrosatellites.The microsatellites underwent touchdown PCR asreported by Hecker e Roux, 1996. A three primersystem was developed: two primers were thelocus specific prirners, while the third primer wascomplementary to the constant tail of one of theprimers and was tagged by a fluorochrome, either6-FAM (6-carbossifluoresceine) or JOE (2',7'di metossi -4' ,5'-d ic Ioro-ô-carbossi fluoresce ine).The amplification conditions were the following:6 cycles, denaturation 45s at 94°C, elongation45s at 72°C, annealing 45s with decreasingtemperature from 60°C to 55°C; 34 cycles,denaturation at 94°C for 30 s, annealing at 55°Cfor 30 s, elongation at 72°C for 30s (8 mins forlast cycle).The amplified fragment were run on sequencingacrilamide gels in an automatic sequencer ABI373A. The alleles identification was performed bythe GENESCAN 672 (Perkin Elmer) software.
RESULTS
Constructing the genomic libraries.The probability of finding polymorphicmicrosatellite depends very much on theprocedure adopted for the construction of thelibraries as well as on the enrichment strategy.Here we account for some relevant points of thestrategyadopted.As reported in the Material and Method section,we constructed two genomic libraries. The firstlibrary was enriched for the dinucleotidemonomer sequence TG which was chosen on the

basis of the experience of various research groupson barley and other plants. The second library wasenriched for the trinucleotide monomer sequenceATC which was chosen following hybridisationexperiments. The DNA was digested with a fewrestriction enzymes including Tsp509! andhybridised with the following oligonucleotides:(ATC) 10, (ACC) 10, (AAC)IO, (AAG) 10. The
best results were obtained with Tsp509! digestionfor the oligonucleotide (ATC) 10.- in fact the
Southem blot showed a uniform profile where themajority of DNA fragments was below I Kbp.The criticai enrichment step was performed byconjugating the magnetic beads coated bystreptavidin with the biotinilated oligonucleotide(ATC)10 or (TG) 13, the unbound DNA wasdiscarded. The efficiency of enrichment wasevaluated by hybridisation of the -ZAPIIcolonies with the chemiluminescent 5'(DIG)3(ATC)1O or 5'-(DIG)3(TG) 13 oligomers.About 9% of the colonies gave a strong positívesígnal. We picked up only the positive colonies,thus introducing a second degree selectionprocedure.
The sequencing ofthe positive clones.Most of the clones were short enough to besequence in one run. About 70% of the(ATC)IO/(TG) 13 positive clones contained aSSR. Here are reported some examples of themicrosatellite sequences ofthe TG library:GTGTGTGTGTGTGTGAGAGAGAGAGAGAGAGAGAGAGAGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGCACACACACACACACACAAGCTCTCTCTCTCTCTCTCTCTTGTGTGTCTATGTATGTATGTATATATGTGTGTGTGTATATGTGTGTCACACACACACACACACAAGCTCTCTCTCTCTCTCTCTCTBesides the expected TG and the complementaryAC repeats, compound microsatellites wereobtained where the repeats could be: TA, CT orGA. Thus the sequencing to the clones allowedfor the identífication of the monomer sequence ofmicrosate 11 ites which were not speci ficallyselected for. This result could be use in futureprojects for microsatellite production.
Primer designed.Even if about 70% of the sequenced clonescontained a microsatellite sequence, the numberof prímer pairs desígned was much lower. Manyclone sequences could not be used for designingprimers for one or more ofthe following reasons:No repeated sequence could be ídentified;
127
The insert was a chimera, i.e. composed by twoor more DNA fragments of independent originoThe microsatellite covered the entire insert and
there was no space for designing the primers;The microsatellite was toa short, unlikely to bepolymorphic;The Tm ofthe two primers was toa different;
The amplification conditions did not meet therequirements established "a priori", We fixedsome amplification parameters which must becommon to ali primers to allow for thedevelopment of multiplex when analysing largesample population.Because of ali these limitations only 30% of thesequenced clones allowed for primer designed.
Identification of the polymorphicmicrosatellites.The identification of the polymorphic loci wasperformed analysing DNA extracted from thecultivars Mundo Novo and Bourbon. from the 12F2 plants of the segregating population and from
two plants of the same parental lines (ET30 andCaturra) ofthe F2' The identification ofthe alleles
was performed on sequencing gels because wecould expected alleles differing only for 2 basepairs.When using the same amplification conditions forali primers as reported in the Material and Methodsection, about one third of the primer pairs gaveno amplification product. Presumably theseprimers require adjustments for the amplificationconditions and they have been momentarilyabandoned. Many of the remaining primer gavemonomorphic bands. Only 25% of the primerswhich gave an amplification products eventuallygave polymorphic bands either on the varieties oron the F2 population.
Even ifwe analysed a limited number ofvarieties.frequently we obtained multiple alleles, from 2 to5 alleles. Usually each single plant had either oneor two alleles, except the locus E 12-3CTG whogave 3 or 4 bands per plant.
D1SCUSSION.
The construction of the genomic library enrichedin microsatellite sequences proved to be rathercomplex. Nevertheless the results so far obtainedare very promising and there are high probabilitiesof finding a reasonable number of polymorphicmicrosatellites. In facts more than 3% of thesequenced clones allowed for the identification ofpolymorphic loci or, if we refer to the genomiclibraries, we expect that about 0.2% of the clonescontain a polymorphic sequence. We estimated

that our enriched genomic libraries should haveabout 50,000 clones still to be screened andtherefore we could expect about a hundredpolyrnorphisrns to be ídentified.An open question is the redundancy of ourgenomic libraries. Until now, we had very fewclones sharing the same sequence or partialsequence identity. Nevertheless it is reasonable toforesee an increase of clones containingsequences already known. Thus the total numberof potential polymorphisms should be correctedfor the redundancy but, for the moment, wecannot evaluate the correction value.Co./fea arabica is an autogamous plant and areasonable percentage of homozygosity isexpected. Moreover, it has been reported that thegenetic base of the cultivated varieties is rathernarrow (Bertraud and Charrier, 1988). Underthese circumstances we would expect a reducedvariability even at the microsatellite leveI. Ourdata do not allow for a comparison with othervegetal species on theAcknowledgements: The research project hasbeen supported by EU, INCO Contract n.ERBIC 18CT970 181.
REFERENCES.
Bertraud 1.. and A. Charrier. 1988. GeneticResources of Coffea. In "Coffee". Vol. 4.Agronorny, R.J. Clarke and R. Macrae Ed.s,Elsevier Applied Science, London ppl-41Clifford M.N .. and K.C Wil1son. (Editors) 1985.Coffee; botany, biochemistry and production ofbeans and beverage. London, Croom Helm,Hecker K.H .. and K.H. Roux. 1996. High and lowannealing temperatures increase both specificity
128
and yield in touchdown and stepdown PCR.Biotechniques 20: 478-485.Lashermes P., S. Andrzejewski, B. Bertrand .M.C Combes , S. Dussert • G. Graziosi, P.Trouslot, and F. Anthony. 1999 Molecularanalysis of introgressive breeding in coffee(Co./fea arabica L.) Theor. Appl. Genet. (in print).Lasherrnes, P.. P. Trouslot, F. Anthony, M.CCornbes, and A. Charrier. 1996. Genetic diversityfor RAPD markers between cultivated and wildaccessions of Co./fea arabica. Euphytica 87:59-64Morgante, M.. A. Pfeiffer, I. Jurman, G. Paglia ,and A.M. 01ivierí. 1998. Microsatel1ite markers inplants. In: Karp A, Isaac PG. Ingram DS (eds)Molecular tools for screeníng biodiversity. Plantsand animais. Chapman and Hall, pp. 206-208,Murray, M.G., and W.F. Thompson. 1980. Rapidisolation of high molecular weight plant DNA.Nucleic Acids Res. 8: 4321-4325.Orozco-Castillo, C, K.J. Chalrners, R. Waugh,and W. Powell. 1994 Detection of geneticdiversity and se1ective gene introgression incoffee using RAPD markers. Theor. Appl. Genet.87: 934-940.Paillard, M.. P. Lashermes, and V. Pétiard. 1996.Construction of a molecular linkage map incoffee. Theor. Appl. Genet. 93: 41-47.Rafalski, 1.A., 1.M. Vogel, M. Morgante, W.Powell. C Andre, and S.V. Tingey. 1996.Generating and using DNA markers in plants. In:Birren B. Lai E (eds) Nonmammalian genomicanalysis: a practical guide. Academic Press,London New York. pp.75-134.Sera, T.. FAndrocioli, R.M.L. Cardoso. M.CL.L.Dias. A. Guerreiro. and E. Silva. 1994.International Syrnposiurn on High DensityPlanting ofCoffee Tree. p293-294 [APAR
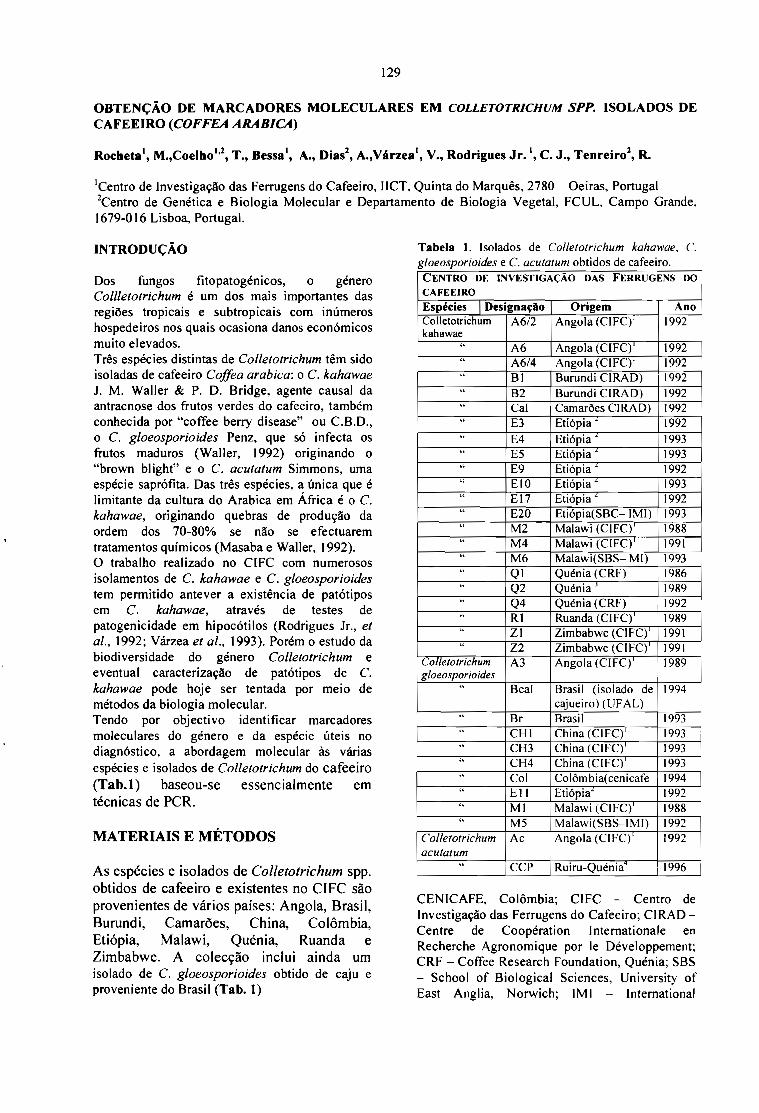
129
OBTENÇÃO DE MARCADORES MOLECULARES EM COLLETOTRICHUM SPP. ISOLADOS DECAFEEIRO (COFFEA ARABICA)
Rocheta', M.,Coelbo··1, T., Bessa·, A., Dias', A.,Várzea·, V., Rodrigues Jr.·, C. J., Tenreiro'', R.
'Centro de Investigação das Ferrugens do Cafeeiro, IICT, Quinta do Marquês, 2780 - Oeiras, Portugal2Centro de Genética e Biologia Molecular e Departamento de Biologia Vegetal, FCUL. Campo Grande.1679-016 Lisboa, Portugal.
INTRODUÇÃO
Dos fungos fitopatogénicos, o géneroCollletotrichum é um dos mais importantes dasregiões tropicais e subtropicais com inúmeroshospedeiros nos quais ocasiona danos económicosmuito elevados.Três espécies distintas de Colletotrichum têm sidoisoladas de cafeeiro Coffea arabica: o C. kahawaeJ. M. Waller & P. D. Bridge, agente causal daantracnose dos frutos verdes do cafeeiro, tambémconhecida por "coffee berry disease" ou C.B.D.,o C. gloeosporioides Penz, que só infecta osfrutos maduros (Waller, 1992) originando o"brown blight" e o C. acutatum Simmons, umaespécie saprófita. Das três espécies, a única que élimitante da cultura do Arabica em África é o C.kahawae, originando quebras de produção daordem dos 70-80% se não se efectuaremtratamentos químicos (Masaba e Waller, 1992).O trabalho realizado no CIFC com numerososisolamentos de C. kahawae e C. gloeosporioidestem permitido antever a existência de patótiposem C. kahawae, através de testes depatogenicidade em hipocótilos (Rodrigues Jr., etaI., 1992; Várzea et al., 1993). Porém o estudo dabiodiversidade do género Colletotrichum eeventual caracterização de patótipos de C.kahawae pode hoje ser tentada por meio demétodos da biologia molecular.Tendo por objectivo identificar marcadoresmoleculares do género e da espécie úteis nodiagnóstico, a abordagem molecular às váriasespécies e isolados de Colletotrichum do cafeeiro(Tab.l) baseou-se essencialmente emtécnicas de PCR.
MATERIAIS E MÉTODOS
As espécies e isolados de Co/letotrichum spp.obtidos de cafeeiro e existentes no CIFC sãoprovenientes de vários países: Angola, Brasil,Burundi, Camarões, China, Colômbia,Etiópia, Malawi, Quénia, Ruanda eZimbabwe. A colecção inclui ainda umisolado de C. gloeosporioides obtido de caju eproveniente do Brasil (Tab. 1)
Tabela I. Isolados de Colletotrichum kahawae, C.gloeosporioides e C. acutatum obtidos de cafeeiro.CENTRO DE INVESTIGAÇÃO DAS FERRUGENS DO
CAFEEIRO
Espécies IDesignação Origem AnoColletotrichum A6/2 Angola (CIFC) 1992kahawae
" A6 Angola (CIFC) 1992" A6/4 Angola (CIFC) 1992.. BI Burundi CIRAD) 11992" B2 Burundi CIRAD) 1992" Cal Camarões CIRAD) 1992" E3 Etiópia" 1992" E4 Etiópia" 1993
I
" E5 Etiópia" 1993" E9 Etiópia" 1992" ElO Etiópia" 1993" EI7 Etiópia " 1992" E20 Etiópia(SBC- IMI) 1993" M2 Malawi (CIFC) 1988" M4 Malawi (CIFC) 1991" M6 Malawi(SBS- MI) 1993" Ql Quénia (CRF) 1986" Q2 Quénia J 1989" Q4 Quénia (CRF) 1992" RI Ruanda (CIFC) 1989" ZI Zimbabwe (CIFC) 1991" Z2 Zimbabwe (CIFC) 1991
Colletotrichum A3 Angola (CIFC)1
1989gloeosporioides
" Bcal Brasil (isolado de 1994cajueiro) (UFAL)
" Br Brasil 1993.. CHI China (CIFC) 1993" CH3 China (CIFC) 1993" CH4 China (CIFC) 1993" Col Colômbia(cenicafe 1994" EII Etiópia" 1992,. MI Malawi (CIFC) 1988" M5 Malawi(SBS-IMI) 1992
Colletotrichum Ac Angola (CIFC) 1992acutatum
" CCP Ruiru-Quénia 1996
CENICAFE, Colômbia; CIFC - Centro deInvestigação das Ferrugens do Cafeeiro; ClRADCentre de Coopération Internationale enRecherche Agronomique por le Développement;CRF - Coffee Research Foundation, Quénia; SBS- School of Biological Sciences, University ofEast Anglia, Norwich; IMI - International

As regioes amplificadas são conservadas nestasespécies não tendo potencial como marcadormolecular diagnosticante do agente da CBO.O facto de não se ter detectado nenhumpolimorfismo de dimensão nestas regiões nãoexclui a hipótese de existirem polimorfismos derestrição. Para testar esta hipótese para cada umdos amplificados de todos os isolados deColletotrichum, está em curso a restrição comvárias enzimas (AR ORA- Amplified RibosomalONA Restriction Analysis).Pelo facto da amplificação de uma região do geneque codifica o rRNA 18S e regiões ITS não terrevelado nenhum polimorfismo de dimensão, nãose exclui a hipótese de poder haver diferenças aonível da composição nucleotídica em cada umadestas regiões, só possível de determinar atravésda sequenciação de cada uma das referidas regiõesem cada um dos isolados.
Marcadores baseados no perfil RAPDAdicionalmente e com o objectivo de detectarpolimorfismos entre indivíduos está a ser aplicadaa técnica RAPO (Random Amplified PolymorphicONA) a todos os isolados (Welsh e McClelland,1990; Williams et al., 1990).A natureza molecular do polimorfismo RAPOpode incluir: alterações nucleotídicas queimpedem a amplificação por nãocomplementaridade num local de ligação aoprimer, delecção do local de ligação, inserçõesque distanciam os locais de lígação de forma a serimpossível a amplificação, inserções ou delecçõesque alteram o tamanho do produto deamplificação.Uma característica fundamental dos marcadoresRAPO é o facto de se comportarem comomarcadores genéticos dominantes herdados deforma mendeliana. Uma banda RAPO só pode serconsiderada um marcador de comportamentomendeliano depois de verificada a sua segregaçãona geração seguinte. De salientar que esteprincípio aplica-se a todo e qualquer marcadormolecular.A aplicação do primer OPC - I] aos isolados dasdiferentes espécies permitiu separar claramente C.kahawae de C. gloeosporioides e C. acuratum(figura não apresentada). Na quase totalidade dosisolados de C. kahawae os fragmentosamplificados situam-se entre 800 e 1800 pb,enquanto nos isolados das outras espécies seobservam fragmentos de dimensões inferiores(300 a 800 pb). A análise numérica dos resultados
Mycological Institute, Londres; UFALUniversidade Federal de Alagoas; (I) - Isolado eidentificado no CIFC; (2) - Isolamento fornecidopelo Or. Biratu; (3) - Fornecido pelo Or. Hindorf;(4) - Coffee Research Fondation, Quénia.
Neste trabalho foram usadas essencialmentetécnicas de PCR utilizando o aparelhoRoboCycler da Stratagene.Para amplificar um fragmento do gene quecodifica o rRNA 18S e para a amplicação deregiões ITS foram utilizados os "prirners", Trl(5'- GTTTCTAGGACCGCCG-3') e Tr2(5'-CTCAAACTTCCATCGAC-3') (Bock et al.,1994) e os "prirners" ITS2, ITS3, ITS4 e ITS5,
respectivamente (White et al., ]990). Atemperatura de "annealing" em ambos os casos foide 56°C, I minuto durante 35 ciclos.Na técnica de RAPO foi utilizado o primer OPCIl (5'- AAAGCTCGGG; Operon Technologies),com um temperatura de "annealing" de 37°C, 2minutos durante 35 ciclos.Para amplificação de sequências demicrosatélites foram utilizados os "primers"referidos na Tabela 2 usando o "touchdown-PCR"(Longato & Bonfate, ]997». A temperaturainicial de "annealing" foi de 65°C variando 0,5°Cdurante 10 ciclos seguindo-se 25 ciclos a 60°C.
Tabela 2. Sequências dos "primers"utilizados na amplificação de microsatélites.
SEQlJÊNCIA 5'- 3'
GACA GACA GACA GACAGTGC GTGC GTGC GTGCGAC GAC GAC GAC GAC
AGCAGCAGCAGCAGCAGCTYYTAGCAGCAGCAGCAGCAGC
RESULTADOS E DISCUSSÃO
MARCADORES BASEADOS NAAMPLIFICAÇÃO DE REGIÕES QUECODIFICAM O rRNA RIBOSSOMALEm todos os isolados de C. kahawae, C.gloeosporioides e C. acutatum, foi ampl ificadoum fragmento do gene que codifica o rRNA 18S.utilizando os "prirners Trl e Tr2. Os resultadosobtidos não permitiram qualquer diferenciaçãoentre as espécies testadas.Nestas espécies foram também ampl ificadassequências intergénicas do rONA (ITS - InternaITranscribed Spacer). Todos os isolados de C.kahawae, C. gloeosporioides e C. acuratumapresentaram um padrão de bandas de tamanhosemelhante para cada uma das regiõesamplificadas, ITS I (280 pb), ITS2 (350 pb) e ITSt(590 pb).
]30
MARCADORESAMPLIFICAÇÃOREPETIDAS
BASEADOS NADE SEQUÊNCIAS
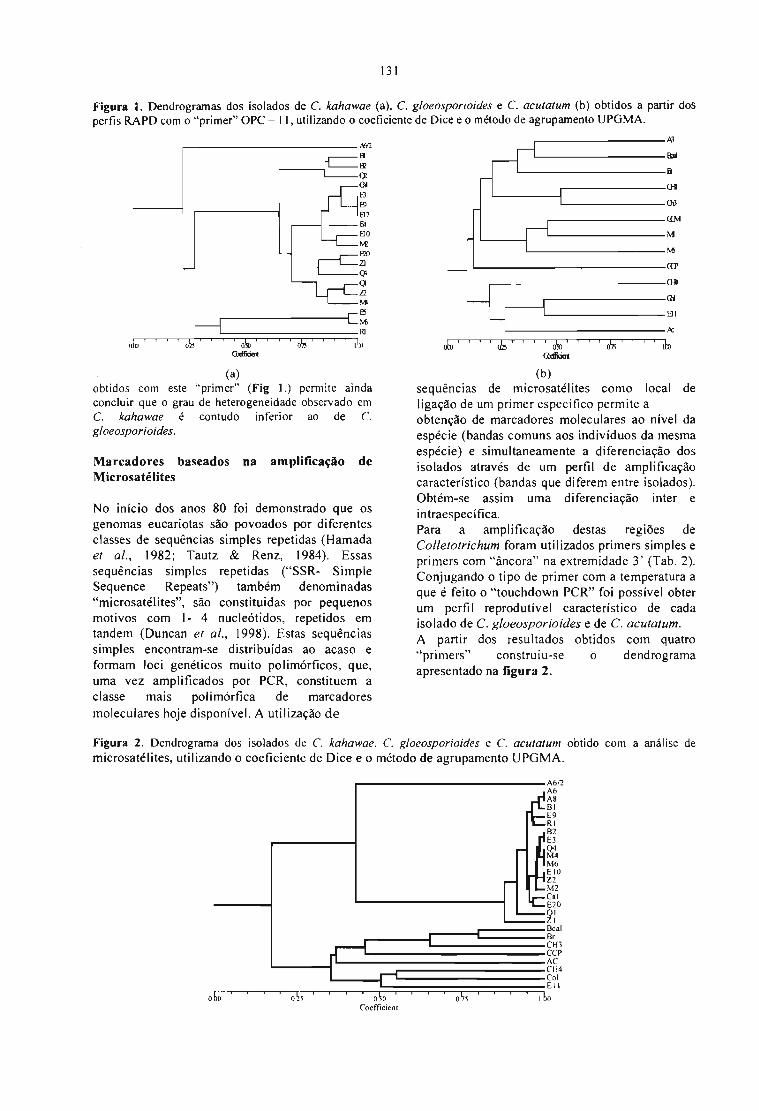
131
Figura 1. Dendrogramas dos isolados de C. kahawae (a), C. gloeo.sporioides e C. acutatum (b) obtidos a partir dosperfis RAPD com o "prirner" OPC - 11, utilizando o coeficiente de Dice e o método de agrupamento UPGMA.
"I" ' JI)' , dí5 ' o\lJ '~
A1
Ibi
8
(]-I
(hJ
([M
M
MS
CO'
CHI
(lj
DI
k,
oÇ., , ,
,lu
(a)obtidos com este "prirner" (Fig 1.) permite aindaconcluir que o grau de heterogeneidade observado emC. kahawae é contudo inferior ao de C.gloeosporioides.
Marcadores baseados na amplificação deMicrosatélites
No início dos anos 80 foi demonstrado que osgenomas eucariotas são povoados por diferentesclasses de sequências simples repetidas (Hamadaet ai., 1982; Tautz & Renz, 1984). Essassequências simples repetidas ("SSR- SimpleSequence Repeats") também denominadas"rnicrosatélites", são constituidas por pequenosmotivos com 1- 4 nucleótidos, repetidos emtandem (Duncan et ai., 1998). Estas sequênciassimples encontram-se distribuídas ao acaso eformam loci genéticos muito polimórficos, que,uma vez amplificados por PCR, constituem aclasse mais polimórfica de marcadores
moleculares hoje disponível. A utilização de
(b)
sequências de microsatélites como local deligação de um primer especifico permite aobtenção de marcadores moleculares ao nível daespécie (bandas comuns aos indivíduos da mesmaespécie) e simultaneamente a diferenciação dosisolados através de um perfil de amplificaçãocaracterístico (bandas que diferem entre isolados).Obtém-se assim uma diferenciação inter eintraespecí fica.Para a amplificação destas regroes deColletotrichum foram utilizados primers simples eprimers com "âncora" na extremidade 3' (Tab. 2).Conjugando o tipo de primer com a temperatura aque é feito o "touchdown PCR" foi possível obterum perfi I reprodutível característico de cadaisolado de C. gloeosporioides e de C. acutatum.A partir dos resultados obtidos com quatro"primers" construiu-se o dendrogramaapresentado na figura 2.
Figura 2. Dendrograma dos isolados de C. kahawae. C. gloeosporioides e C. acutatum obtido com a análise demicrosatélites, utilizando o coeficiente de Dice e o método de agrupamento UPGMA.
A6/2A6ASBI[9RIB2E3
~M6ElOZ2M2C.IE20
~iBcalBrCH3CCPACCH4CalEII
I bo0\0Coefficienr
~ri
r-~
r-~4l-
Lcl.....-
re!I
, I I I I I I i I I i I I I I Iobo

Os resultados obtidos com a amplificação desequências usando "primers" para microsatélitespermite claramente estabelecer um grupo deisolados de C. kahawae e um outro grupo deisolados formado por C. gloeosporioides e C.acutatum. Estes dois grupos apresentam umasemelhança de 17%. Os padrões de amplificaçãodos isolados de C. kahawae revelaram-se muitouniformes comparativamente aos isolados de C.gloeosporioides e C. acutatum, um resultado queestá de acordo com o obtido com o perfil RAPO(Fig.I). Adicionalmente, esta técnica permitiuobter marcadores moleculares para as trêsespécies de Colletrotrichum para além deestabelecer um "fingerprinting" para cadaindivíduo.Este trabalho foi financiado pelo projecto PraxisP/2/2.1/BIA/309/94
REFERÊNCIAS BIBLIOGRÁFICAS
Bock. M., Maiwald, M. Kappe, R., Nickel, P. &Naher, H. (1994). Polymerase chain reactionbased detection of dermatophyte ONA with afungus-specific primer system. Mycoses 37. 7984.Ouncan, 1. M., Cooke, O.. Birch, P. & Toth, R.(1998). Molecular variability in sexuallyreproducing fungai plant pathogens. In: P. O.Bridges, Y. Couteaudier & 1. M. Clarkson (Eds.),Molecular Variability of FungaI Pathogens(pp.19-39) Oxon, UK: C.A.S. International.Harnada, H., Petrino, M. G. & Kakunaga, T.(1982). A novel repeat element with Z-ONAforming potential is widely found inevolutionarily diverse eukaryotic genomes. Proc.NatI. Acad. Sei. USA 79: 6465-6469.Longato, S. & Bonfate, P. (1997). Molecularidentification of mycorrhizal fungi by directamplification of microsatellite regions. MycoI.Res. 4: 425-432.
132
Masaba, O. & Waller, J. M. (1992). Coffee berrydisease; the current status. In: 1. A. Bailey e M. J.Jeger (Eds.), Colletotrichum: biology, pathologyand control (pp. 237-249) Oxon, UK: C. A.B.International.Rodrigues Jr, Várzea, V. & Medeiros. E. (1992).Evidence for the existence of physiological racesof Colletotrichum coffeanum Noack sensuHindorf. Kenya Coffee 57, n" 672: 1477-1420Tautz, O. & Renz, M. (1984). Simple sequencesare ubiquitous repetitive componements ofeukaryotic genomes. Nucleic Acids Res. 12: 41274138.Várzea. V. M. P.. Rodrigues Jr., C. 1., Medeiros,E. F. (1993). Oifferent pathogenicity of CBOisolates on coffee genotypes. In: Proceedings ofthe 15th International Scientific Colloquium onCoffee, Montpellier, 6-11 June 1993- ASIC(Paris) pp. 303-308.Waller, J. M. (1992). Colletotrichum diseases ofperennial and other cash crops. In: 1. A. Bailey eM. 1. Jeger (Eds.), Colletotrichum: biology,pathoIogy and control (pp. 167-185) Oxon, UK:c.x.s. International.Welsh. J. & McClelland. M. (1990). Fingerprinting genomes using PCR with arbitraryprimers. Nucleic Acids Res. 18: 7213-7218.White, T. J.. Bruns, T., Lee, S. & Taylor. J.(1990). Amplification and direct sequencing offungal ribosomal RNA genes for phylogenetics,In: M. A. lnnis, O. H. Gelfand, 1. 1. Sninsky & T.J. White (ed.), PCR Protocols: A Cuide toMethods and Applications (pp. 315-322).Williams, J. G. K.. Kubelik, A. R.. Livak, K. 1.Rafalski, 1. A. & Tingey, S. V. (1990). ONApolymorphisms amplified by arbitrary primers areuseful as genetic markers. Nucleic Acids Res. 18:7213-7218.

133
GENETIC FINGERPRINTlNG OF COFFEE LEAF RUST OIFFERENTIALS WITH RAPOMARKERS
A. Santa Ram & H.L. Sreenath
Tissue Culture & Bíotechnology Centre Coffee Board. Manasagangothri Mysore - 570 006
SUMMARYLeafrust is a major disease ofcoffee known to be causing severe crop tosses. World wide efforts lo combat this diseasebv various means revealed that resistance in lhe coffee plant and virulence in lhe rU.I'1 pathogen t Hemileia vastauix) (JI'('
conditioned bv a gene-for-gene host-pathogen relationship. Thus. known genotypes o] coffee nranites: resistance lospecific sets of roces of lhe rust fungus There are /-1 such differential hO.l'I.I' in lhe gene bank o/TCR/ 711e.l,(, ore.Bourbon. Kents. S. 288. S/2 Kalfa. Geisha, Agam. Ciocce. /IDT-:l. HIJ{-R. Blue Mountain x Cioccie. ...,.. 333 x f)illa";'Alghe (S25"'-I). Dilla & Alghe .r S333 (S25-9) and Blue Mountain .r SI! Kaffa (.\'26661 in Corfea arabica. Twodifferential hosts are drawn from other species ofCoffea. Tltev are C racemosa and C congensis. One genotvpe of;'arabica by name Rume Sudan manifests a high degree offie/d resistance. presumed lO be horizontal reststance lhepresent studv was undertaken in a bid to identify D.IvA markers whicli distinguish each of lhe differentials Preliminarvdata on DNA polvmorphism indicates clear distincuons among lhe differentials Cluster analvsis indicated a/urgedegree ofsunilaritv among lhe ge notvpes ofC. arabica whichfell 11110 /11'0 major clusters.
INTRODUCTION
Perennial plantation crops have a long breedingand selection cycle. Oiscerning segregatíons ín F,generation in these crops is very difficult at thejuvenile stage. Thus, genetic analysis of plantationcrops like coffee by conventional methods provesitself to be a task of considerable dimensions.Molecular markers which are directly derivedfrom ONA are relíable. free frorn environrnentalintluences and can be assayed at any stage ofplant growth with any type of tissue. Recentadvances in the field of plant molecular geneticshave evolved a series of ONA markers. Notableamong them are restriction fragrnent lengthpolyrnorphisrns (RFLPs). (Botstein et ai. 1980)random amplified polyrnorphic ONA (RAPOs)(Williams et ai, 1990; Welsh and McClelland.1990). ampli fied fragment length polyrnorphims(AFLPs) and simple sequence repeats (SSRs) OI'
microsate llites (Weber and May, 1989).Out of the various ONA markers, PCR basedRAPOs are relatively sirnple to generate and areideally suited for a breeders laboratory. In basicPCR. two selected prirners of known nucleotidesequence are used to amplify the specific gene ofinterest (Mullis and Faloona. 1987). Thus, PCR isan effective tool in screening for the presence OI'
absence of a gene of interest in a variety of targetONAs. In contrast, a simple prirner of arbitrarynucleotide sequence is used in RAPO assay.Earlier workon RFLPs and RAPOs of coffee evaluated theiruse as indicators ofphylogeny and geneticdiversity (Berthou et ai .. 1983; Orozco-Castillo etal.. 1994.1996; Lashermes et a! ..1995). Effortswere also devoted
towards developing linkage maps of coffeeusing RFLP and RAPO data (Paillard et al..1996). Thus, it is noted that research work Iinkingthese markers to important agronornic traits suchas disease OI' pest resistance is lacking. However.a lot of back ground information is available onthe genetics of rust disease resistance in coffee(Eskes.1989) and several host types mani festingdifferentíal resistance to the pathogen races areidentified.In the genus Coffea L.. C arabica L. is the onlytetraploid species (2n=4x=44) manifesting anoften-self-pollinating reproductive behaviour. Aliother species ofthe genus are diploid (2n=2x=22)and self-íncompatíble. Arabica is cornmerciallythe most important species and is also the onesusceptible to a variety of pests and diseases. OI'particular importance among the diseases 01'arabica coffee is the leaf rust caused by Hemileiavastatrix B. et Br. Arabica manifests specificresistance against races of the rust fungus. In thegermplasm bank of Central Coffee ResearchInstitute (CCRI ) there are 14 host types of Carabica and 2 host types derived frorn diploidspecies of CofJea. viz. C racemosa Lour. and Ccongensis Pierre. There is also a wi Id C urahicacollection known as Rume Sudan which manifestsnon-specific resistance to the rust fungus, Presentstudy is conducted on these materiais mani festing
a divergent spectrum of reactions to the rust racesand the observations are presented.
MATERIALS AND METHOOS
Materiais of the studyFourteen host physiologic types of CO/JL'U arabicaviz. Bourbon (E). Kents (O). S.288(G). S" Kaffa(W). Geísha (C). Agarot I), Cioccie (J). RUl11c

Sudan (HR). Hibrido- de-Ti mor (HDT-A and R).Blue Mountain x Cioccie (Y). Blue Mountain x 12Kaffa (S.2666)(O). S. 333x Dilla & Alghe(S.2574 )(V). Dilla & Alghe x S.333 (S.2579)(Z)and one each of c'racelflosa (F) and c.congensis(B) and wild arabica Rurne Sudan showing high
134
degree of horizontal resistance (HR) provided thematerial for the present study. Their genotypesand resistance groups are surnmarised in tablel.Host types studied can be classified into fourdistinct groups as follows:
Table-l: Coffee leafrust differentials used in the studvDifferentials Resistance genes Resistance group Reaction
-----------Coffea arabica
! Arábicas
Bourbon SII5 E highly susceptibleI
ID----.-
~ghlysLJscepti ble II Kents S1I2. 5
t= iGeisha SIII.5 C moderately susceptible I----- j------. ----j
SI2 Kaffa SIII. 4. 5 W moderately susceptibleII
! Cioccie S1I4. 5 J moderately susceptible
Agaro S1I4. 5 J moderately susceptible
Rume Sudan ----- HR highly resistant
~Arabicoids-----J
----_..---...~I S.288 S1I3. 5 G moderately resistant: Hibrido-de-Tirnor SH6. 7. 8. 9 A highlyresistant-
l. Hibrido-de-Tirnor S 6 R highly resistantHiI Hybrids
Blue Mountain x Cioccie SI12.4.5 Y moderately resistant
I Blue Mountain x S 12 Kaffa SIII.2.4.5 O moderate Iy res istant
:Dilla & Alghe x S. 333 Sfll.3.5 Z moderate ly resistant
S. 333 x Dilla & Alghe SII1.2.3.5 V moderare I)' resistant..
. Diploid speciesF Coffea racemosa .. F highly susceptible~'ofreacongensis
I.. B moder~tely resist~~t~
Arábicas: These are tetraploids derived from wildarabicas and are considered as pure arabicagenotypes.
Arabicoids: These are also tetraploids derivedfrorn the putative spontaneous hybridization of Carabica with diploid species like C liberica ( S.288. S. 333) ar C canephora (HDT).
Hybrids: These hosts are also tetraploids derivedfrorn artificial hybridization of arabicas and thearabicoids OI' between arabicas.
Diploid species: These host types bclong to thediploid constellation ofthe species oi Coffea.
DNA isolation: High molecular weight genornicDNA was isolated from the young leaves of field
grown plants. About 10-15g of young coffecleaves were ground to a fine powder in LN, in apre-chi Iled mortar and the owder was scoopedinto four Oakridge centrifuge tube containing laml of pre-warrned (60°C) extraction buffer (2%
cetrimide (SRL) (w/v). 1.4M NaCI. 20 mMEDTA. 100 mM TrisHCI pH.80) containing 1%PVP and 0.2% ~-mercaptoethanol which wereadded to the buffer immediately before use. Thisextraction rnixture was incubated in awaterbath at 60°C for one hour with frequentstirring of the contents. To this. 0.6 volume ofchloroform-isoarnylalcohol (24: I) reagent wasadded and the contents of thc tubes thoroughlymixed. Then the tubes were spun at 10.000 rprnin a refrigerated high specd centrifugo (Sigma.Germany) for about 15 mino at 4°C to separate thephases. The upper aqueous phase was transferred

Amplification reactions were carried out in apeltier thermal cycler PTC 200 ONA Engine (MJResearch, USA) with the following programme.
Step-II 45 cyclcs 01' 94° C· 30 seco (Dcnaturuuon j
36° C - I mino (Anncaling )
72° C - 2 min (Chain extcnsion í
ElectrophoresisPCR products were subjected to sub-marineelectrophoresis (Biornetra, Gerrnany) at 10v/cm inan agarose gel of 1.2% strength with 1X TBE astray and gel buffer to visualise RAPO's. Either a100 bp ladder or À ONA digested with EcoRI andHind I" ar Hind 111 restriction enzymes was usedas a standard molecular weight marker. TheRAPO profiles were recorded by visualobservation of ethidium bromide stained gelsbesides being photodocumented in a geldocurnentation system (Bio-Rad. USA).
to fresh pre-cooled oakridge tubes withoutdisturbing the interphase. To this, 0.5 volume ofice-cold isopropanol was added. At this stage, theDNA precipitated as a whitish net work. DNAwas pelleted by spinning the tubes at 12.00014.000 rpm at 4°C for 15-20 minutes. DNA pelletwas washed with 10mM ammonium acetate in70% ethanol. The tubes were again spun at 10.000rpm for 10 minutes and the washing solution wasdecanted. DNA pellet was dried under vaccumfor about 30 minutes. Finally the pellet wasdissolved in 1000fll TE (10 mM Tris HCI pH 8.0)-1 mM EOTA) buffer. These samples werestored at -760 C in an ultra freezer for further use.
PCR Amplification- Components & ConditionsReaction mixture for PCR composed of 2.5 ~t1
standard assay buffer (1 OX). 2 units of Taq DNApolymerase (Genei, lndia), 400 ~lM dNTPs. 25ngof arbitrary prirner (I ü-rner) (Operon Tech. Inc.USA) and 75 ng of template (sample) ONA in atotal volume made upto 25 ul. with stcrilc distillcdwater, The sequenccs 01' the primers used in ihis studyare presented in table 2.
135
Step - I
Step-III
94° C - 3 min (Initial denaturatiom
72° C - 10 min (Final chain cxtension í
TABLE-2: decamer primers used in the studyPrimer Nucleotide Sequence
OPA - OIOPB - 12OPF - 04OPF - 06OPF - 07OPF - 15OPAA - OIOPAA- 12OPAT- OIOPAT - 02OPAT- I1OPAT - 18
5' 3'CAGGCCCTTCCCTTGACCGAGGTGATCAGGGGGAATTCGGCCGATATCCCCCAGTACTCCAGACGGCTCCGGACCTCTTGCAGTGGTTCCCAGGTCTAGGCCAGATCTCCCCAGCTGTGA
AnalysisObserved band patterns were analysed using theUnweighed pairgroup method of analysis(UPGMA) algorithm to understand the similaritiesand relationships of the genotypes studied and theresults are presented in a dendrogram (Fig. 6).

136
Fig.6I ~ ~ .
52579 (Dill. & Alghe x 5.))))
5.2666 (Blue Mounlain x $.12 Kaffa) - ---...,
Blue Mounlain x cro c c te
5.257415.))) x Dfll a & Alghe)
Hibrido de Timor - R
Bourbon
K e n t s
Agaro
Cio c c ie
5.288
Hibrido de r tm or . A
Rume Sudan
512K.II.
C c c n q e n sr s
C r a c e m o s a
~-----------------------------------_._----------
RESliLTS & DISClJSSION
()!\J,i\ 1)01~ 1110rpl1 iSI11 data t'\?J)\?rat\?d OJ) 16 Iwst
l~ p\?S oI' L'llllcc with I::: primcr-. I'C\ calcd t'CIlOlllic
di\crsit~ h~ \\~I~ otdi tfcrcut hund paucrus(Fig 1.2.3.·n A total oI' 1.3::: R:\I'() loci \\e'rc
\ isualiscd Irom thc PCR producis oi ihe:,c
primcrs, OI' thc-,e R~ \\'CrL' trom (. arabu« a/lu -17w erc lrom (' c'!JII,\!.c'l/si\' and (' '''/('(,IIIU.\(/. Withinthe' R/\I'f) bands 01'(' (/rUhIUI\, about 77"" \\CrL'
p(ll~ morphi;: lhis high dcgrcc of pol~ IllOII)lliSlll
could hc ~I 1\?I1cL'li0I1 (11' rhc Cr) puc !,'CIl\?l\L'
\ .ui.n ion-, ,1I11nl1g ucrbic« \ arictie's \\ hich 111,1\
h,l\ C ariscn as ,I I11C,lIh oi' local adapt.uions in ihc
di verse gco!,'raphic locat ious trorn \\ hcrc thL'~
\\L'lC drawn. Thc primcrs OPA·O I and 01':\1·1 Rdislillguishcd l11os1 o l ihc gCllot) PL'S 11) pl(,duL'il1g
L'hal,lc!crist iL' product pro li Ies, (Fig. 2,3) lhc
othcr primcr-, gCl1cr,lted product-. \\ hic h cnn 11L'
uscd to (Iistillgllish one host t) pc (li thc olhL'/
11()\\c\\?!'. withi n cach ofihc grollps ihc gL'll(IllliL'
div L'lsit~ \\~IS kss obvious
!\11 IIl\crcsllllg ol1scr"llloll Ih~II ('dlllC lI) 11t'lll II1
th""lI(l~ IS thc dist incti on hct wccn 111) Ig\?llot~ pc-. Ill)l·A t'e'I\(lt~ 11L' i-, uniquc in ih.u Ii
C~IITic, thc rust rCSiSI~1I1CL' gCllc, '>llh. 7, i'i <Illd <)
,til oI' \\ hich are inlcrrccl to bc dcriv cd 110m
(' '11//('1,11111'" (rkltL'llcnurt c'I "r, I ()l):::) lhu-;it h hCtL'I'(I/\gOlIS ;1\ thc !\\n I,ossihk loc iL'(llllllliolllllg 1'L',is!;IIlL'C Iksilk, thi-.lle'tL'lo/~g(lsil\_ thc 1\\0 loci L';UT)illg dil'll'IL'111 SI:
gL'IIC.S makcs this gCl1ot)pc \L'r) di-une: \\ith \L'r~
durahlc rcsiSI,IIlL'C ( 1{,11ll I C)t)~ l. II[l!'·/\ IlI'\ldtlL'L'S
sL'glegallts \\hich are groupcd imo ~11)()thL'1
rc,ist,IIICC cl;\Ss -R'(IIDr·R). 111 thL' l)re'SL'llt ,tll(l~.
it j, (lbSCI\ cd that S(ll11L' nl' lhe teslL'J prilllLTs
(li,tillgui,lllll) 1·,\ '\IHIIIDl'·R h\ S"IIlL' Ri\I'I)

bands( Fig. 5 ).Besides the above distinctions each of the testedprimers also generated a number of cornrnonbands from ali genotypes of arabicas. arabicoidsand the hybrids. Besides being rnonomorphic,these can be perceived as a part of constitutiveDNA which makes C arabica a species ofdistinction. Severa! of these bands could not beobserved in the RAPD profiles of the diploidspecies C. racemosa and C congensis (Fig. 1,2,3, 4). Among the arabica descendants (Arábicas.Arabicoids and the Hybrids) there are some DNAfragments which appear to be constitutive but areabsent in a few host types. This is a possibleindicator of the dynamic genomic differentiation(Fig. 3,4).The two host types of diploid species C. racemosaand C. congensis could also be distinguished fromali arabica descendants as well as withinthemselves by the RAPD bands generated by alithe primers. Possible homologies of DNA ofthese species with that of C. arabica isrevealed by some primers (Fig. 3). These areindicative of the genomic distinction of Coffeaand the possible interspecific gene flow amongthe species of this genus (Orozco-Castillo et ai.1994).From the dendrogram (Fig.ô), it can be inferredthat the genotypes Bourbon and Kents are veryclosely related. The Ethiopian genotypes SlcKaffa, Agaro, Geisha and Cioccie are similar andmanifest a close sirnilarity with Rume Sudan,Bourbon and Kents reflecting the large sirnilarityof ali Arabicas. lnterestingly, the ArabicoidsS.288 and HDT-A also exhibit a close similaritywith this group indicating the probableinvolvement of Typicoid Arabicas in theirevolution. The hybrids S.2574 (S.333 x Dilla &Alghe ), S.2579 (Dilla & Alghe x S.333). S.2666(Blue Mountain x Slc Kaffa) and Blue Mountain xCioccie share a closely similar band pattern withHDT-R. Geographic origin of S.288 and S.333(one of the parents of S.2574 and S.2579) can betraced back to the Doobla Estate in ChikrnagalurDistrict. Their putative derivation frorn ArábicaLiberica hybridization is reflected in the closesirnilarity of their RAPD pattern. Relativedistinctness of HDT-A and R genotypes isbrought out in this analysis. Inspite of ali the cleardistinctions and 77% of genetic polymorphism,Arabicas. Arabicoids and Hybrids rnanifest aclose clustering of genotypes with geneticdistances ranging frorn O to 5. The species Cracemosa and ('. congensis being diploidsclustered distinctly frorn the tetraploid Arábicas.From the foregoing discussion it can be inferredthat the genus Coffca is of monophyletic origin
137
and draws support from the earlier work (OrozcoCastillo el al., 1996). Also, it can be inferred thatthe genomic differentiation in C arabtca is in adynarnic flux resulting from possiblereorganisation/reorientation of DNA at frequentintervals. Thus there is adequate genetic diversityamong the host differentials which are drawnfrorn the geographically diverse locations as we!las among the land races from the variousprovinces of Ethiopia, the home land of ('arabica.
ACKNOWLEDGEMENTS
Authors wish to express the ir gratefulness toDr.R.Naidu.Director of Research. CCRI for kindencouragement.
REFERENCES
BERTHOU..F.. MATHIEU, C; VEDEL F.1983. Chloroplast and mitochandrialDNA variation as indicator of phylogeneticrelationships in the genus CottcuL.Theor.AppI.Genet. 65: 77-84BETTENCOURT. A.L LOPES. L PALIAS.1992. Factores geneticos que aresistencia as racas de Hemileia vastatrix Berk. etBr dos clones tipe dos gruipos I. 2e 3 dederivados de Hibrido de Timor, Broteria GenéticaXIII (LXXX): 185-194BOTSTEIN. D., WHITE, R.L .. SKOLNICK. M..DAVIS, R.W. 1980. Constructionof a genetic map in man using restriction fragrnentlength ploymorphisms. Arner J.Human Genet.32:314-331ESKES. A.B. 1989. Resistance. In Coffee Rusl:Epidemology, Resistance andManagement (Eds.A.C. Kushalappa and A.B.Eskes) PP. 171-291. CRC Press. Boca Raton.LASHERMES. P.. COMBES. M.C.. CROS.LTROUSLOT. P.. ANTHONY. F..CHARRIER, A. 1995. Origin and geneticdiversity of Coffea arahica L.based on DNAmolecular markers. Proc. XVI InternationalScientific Colloquium on Coffee (Kyoto) pp. 528535.MULLlS. K. B.. FALOONA. F.A .. 1987. Spcc ificsynthesis of DNA in vitro via apolymerase catalysed chain reaction. MethodsEnzyrnol. 155: 335-350OROZCO-CASTILLO, c., CHALMERS. K. .I.WAUGH. R.. POWELL, W.1994.Derection of genetic diversity and selective geneintrogression in coffee using RAPO markcrs,Theor.A ppl.Genet. 87 :934-940.

ORO/CO-(ASTII.1.0. c.. Clli\I.MI:RS. 1\. ..1..POWII.L. W.. \Vi\lIGII. R.199ó.R.'\I)[) and organcllc spccific PCR rc-affirmstuxonornic rclationships within thc genus ('''ff,'u.1)lan1 Ccll Rcports 1~:3 37-3-11P:\III.i\R[). M .. I.ASIILRMI:S. 1'.. PITI:\RD.\'.1996. Construction of a molecularlinkage map In coflcc. Thcor. Appl. Gcnct. 9,'-11 --17R!\~1. A.S. 19l).~. Nc\\ dimcnsions InunderstanJing inhcriiancc o l coffce ru-.trc'si~\~\ncc: i\ mcndclian pcrspcctiv c. Prol'. X\'Ilntcrnnt ional Scicut itic Colloquium on coffcc(Ky oto ). 1'1) :'-IX-.~~3
WI-RLR..1.1 ... \1A Y. I)F 1989. Abundam clas~
othuman DNi\ Poly morphisms\\ hich can bc ty ped using poly mcrase chuinrcaciion. /vmcr.Ll luman Genct -1-1: 3X8-'96WLlSII. .I.. Mc CI.I.I.I.i\N D. ~119l)()
Fingerprillting gcnomcs using PCR witharbitrar, prirners. Nuclcic Acids Res. 1X: 72137218WII.I.IAMS. J.(i.K.. K lIBLl.1 K. A.IL 11VAK.K ..I.. RAI!\ISKI. L\ .. IIN(iIY.S.V 1990. DNA pol vmo rphi-m-, umpli ficd h:arhitrar: primcr-. em: usc lul a, genclic mark cr-.l\uckic Acid« Re~.1 X: ô:" I-ô:,'~

SECRETARIA DA CIENCIA,TECNOLOGIA E ENSINO SUP RI R
Define, coordena e executo políticos e diretrizesnas áreas de ciência, tecnologia e ensino superior, num
processo de contínuo desenvolvimento,em prol do sociedade paronaense
Fone: 55 (4r) 350 rooo www.seti.gov.br
I do por lei estadual, destina 2% da receita tributária doE ado ao desenvolvimento científico e tecnológico do Paranó
Duc:
Prom fomento o pesquisa e capacitação de recursoshumanos nos óreas científica e tecnológico do Estado
UNIVERSIDADE ESTADUAL DE LONDRINA / UEL - UNIVERSIDADEESTADUAL DE MARINGA/ UEM - UNIVERSIDADE ESTADUAL DEPONTAGROSSA/UEPG • UNIVERSIDADE ESTADUAL DO OESTE DO PARANÁ/UNIOESTE/ CAMPUS DE CASCAVEL/CAMPUS DE FOZ DO IGUAÇU!CAM PUS DE MARECHAL CÂNDIDO RONDON/CAMPUS DE TOLEOa • UNIVERSIDADE ESTADUAL DO CENT RO - O ESTE /UN I CEN TRO / CAM PUS DE GUARAPUAVA/CA MP US DE IRATI
FACULDADE DEARTES DO PARANA/FAP - ESCOlA DE MUSlCA E BELAS ARTES DOPARANA/ EM BAP - FACULDADE ESTADUAL DE CIÊNCIAS ECONÔMICAS DE
N'UCARANA FECEA - FACULDADE ESTADUAL DE CIÊNCIAS E LETRAS DE CAMPOMOURÃO/ FEC' LCAM - FACULDADE ESTADUAL DEFILOSOFIA.CIÊNCIAS ELETRAS DECORNEUD PRCXOPIO/ FAF' COP - FACULDADE ESTADUAL DEFILOSOFIA. CIÊNCIASELETRAS DElACAREZINHO/ FAFIJA - FACULDADE ESTADUAL DEEDUCAÇÃO FISICADE JACAREZINHOI FAEFIJA - FACULDADE ESTADUAL DE DIREITO DO NORTEPIONEIRO DE JACAREZINHO, FUN D' N OP' - FACULDADE ESTADUAL DE
FILOSOFIA.C1ÊNClASELETRAS DEPARANAGUA FAF'PAR - FACULDADE ESTADUALDE EDUCAÇÃO. CIÊNCIAS E LETRAS DE PARANAVAí, FA F' PA - FACULDADEESTADUAL DE FILOSOFIA. CIÊNCIAS E LETRAS DE UNIÃO DA VITORIA fAF'UV
Gerencio o Fundo Paranó, visando o desenvolvime nto científico e tecnológico do Estado e fomenta aimplantação de programas e projetos de desenvolvimentotecnológico industrial
TAtua no desenvolvimento sócio -econ ômcoparanaense com soluções tecnol gl casinovadoras e na produção de Imunoblologlcos


141
PADRÃO MOLECULAR DE CLONES DE CAFEEIRO DIFERENCIA DORES DE Hemileiavastatrix Berk. et Br,
Terezinha A. Teixeira l,2, Laércio Zambolim 1,3, Ney S. Sakiyama l,2, Antônio A. Pereir'a'Í, Dalza G.da Silva 1,3
IBIOAGRO/UFV; 2DFT/UFV; 3DFP/UFV; 4EPAM IG. 36571-000 Viçosa.Mfi.
Apoio Financeiro: FAPEMIG. FINEP. CNPq. PNP&DCafé
--INTRODUÇÃO
A ferrugem alaranjada do cafeeiro iHemileiavastatrixl, apresenta grande variabi Iidade. tendoja sido caracterizadas mais de 30 raçasfisiológicas. sendo que no Brasi I já foramidentificadas 12.O estudo genético do comportamento dasprogênies de plantas de Coffea arabica e doshíbridos. mostrou que a inoculação com raçasfisiológicas de H. vastatrix, foi possívelidentificar nove fatores dominantes diferentes.
SH L SH2. SH3. SH4. SH5. SH6. SH7. SH8 eSH9 condicionando a resistência à C. arabica Osfatores SH L SH2 e SH5 estão ligados à espécieC arabica. o fator SH3 é encontrado apenas nosArábicos originários da índia e muitoprovavelmente tenha provindo de C. libcricaatravés de hibridação interespecífica. O fatorSH6. provavelmente é oriundo de C. canephora edeve estar associado a um ou mais fatores não
identificados que conferem resistência a todas asraças. sendo encontrado no Híbrido de Timor
(BETTENCOURT e RODRIGUES. 1988).Clones de C'offea spp têm sido usadas como
diferenciadores de raças fisiológicas de Hcutileiavast atrix no programa de melhoramento daUniversidade Federal de Viçosa. A caracterizaçãomolecular desses clones pode facilitar a
manutenção da identidade genética. que éindispensável para propósitos de identificação dasraças fisiológicas.Este trabalho foi proposto com o objetivo de
caracterizar clones de cafeeiro diferenciadores deHcmileia vastatrix e determinar a divergência
genética entre eles por meio de marcadoresRAPD (WILLlAMS et aI.. 1990).
MATERIAL E MÉTODOS
• Material genéticoO material genético utilizado encontra-se listadona Tabela I.
Tabela 1 - Lista dos 18 clones de cafeeirodiferenciadores de Hcmileia vastatrix.
1 Clones ~C1ones
r~~~~~-'-=jJi~~~~'~O~343/269 RH2.635;3 --==---r 04 4106 13. 134/4I •
105.419/20 14.110;5
106.420/10 15.152/3L_....__ __. . .._.
L22._1~?~_. 16.3211i08.33/1 117.1282.__ . _~9.1532 118.17/12
• Extração de DNADNAs dos 18 genótipos foram extraídos de folhasjovens seguindo o protocolo modificado de Doyle& Doyle (1990) com a adição de PVP-40(solúvel) no tampão de extração. Após a extraçãoo DNA foi quantificado em espectrofotórnetro,
corrido em gel de agarose 0.8 % e armazenado a4°C. Na época da amplificação foi diluido em TE(Tris HCI 10 mM. EDTA I mM. pH 8.0) para
uma concentração final de IO ng/ul.
• Amplificação de DNA e análiseeletroforética dos produtos
Vinte e oito primers de 10 bases da "OperouTechnologies" foram utilizados para amplificar osDNAs dos 18 genótipos. A amplificação foi feita
em termociclador Perk in-elrner 9600 e cadareação contou com um volume total de 25 ul,
contendo os seguintes componentes: 25 ng deDNA genômico. I unidade AmpliTaq DNApolimerase. 0.1 mM de cada dNTP. 0.2 u M deprimeI' 50 mM de KCL 10 mM de Tris Hei e 2
mM de MgCI2 e o volume final completado comágua ultrapura. Foi utilizado o seguinte programa:
um ciclo para desnaturação (95°C por 1 min). 39ciclos para amplificação (15 seg a 94°(', 30 seg a]:";0(', 60 seg a nOC) e para finalizar 7 min a
72°('. Os produtos das reações de amplificaçãoforam separados em geis de agarose 1.4%.
coradoscom brometo de etídio. visualizados em
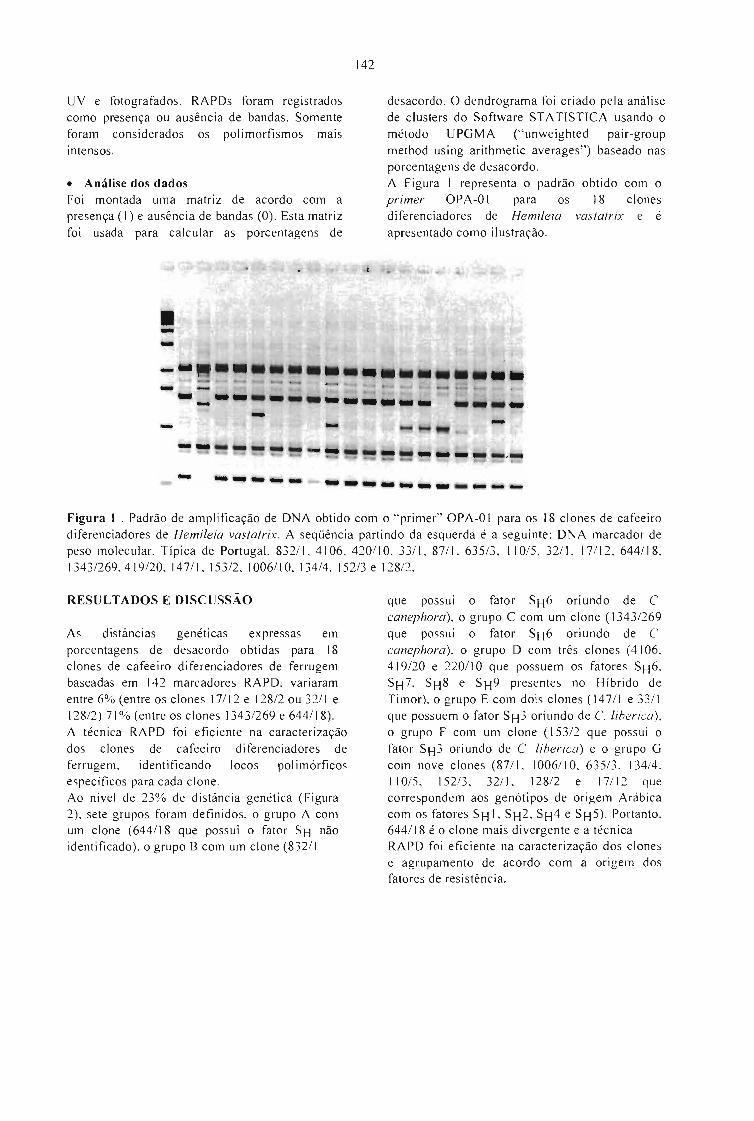
142
UV e foto grafados. RAPDs foram regi stradoscomo presença ou ausência de bandas. Somente
for am co ns iderados os pol im o rfi smos mais
inten so s.
• Análise dos dadosFo i montada uma matri z de acordo com apresen ça ( I) e au sên cia de bandas (O). Es ta matriz
foi usada para ca lc ular as porcentagen s de
desac ord o. O dendrograma foi cri ad o pela análi se
de cluster s do Software ST ATISTI CA usando o
método UPGMA ("unwei g hted pair-groupmethod using arithmeti c averages ") baseado na s
porcentagens de desacord o.A Figura I representa o padrão obtido com oprim er OPA-O I para os 18 clones
d ifer en ciadores de Hemil eia vastatrix e é
aprese ntado co mo ilu stração.
•--..-
----..._- -,~
•
--------.._-_ ..-..--
-----
----
.. ----
Figura I . Padrão de a mplificação de DNA obtido com o "primer" OPA-OI pa ra os 18 clones de cafeeirod iferenciadores de Hemileia vastatrix . A seqüênci a partind o da esquerda é a se guinte: DNA marcador depeso molecular. Tí pica de Portugal. 8.3 2/ I. 4106. 42011 O. .3 .311. 87/1 . 6351.3. I 10/ 5. 32 / I. 1711 2. 644/1 8.1.34 .3 /269. 4 J 9/20. 147/ I. 15.3 /2. 1006/ IO. 1.3 4/4 . 152/3 e 128 /2.
RESULTADOS E DI SCUSSÃO
As distâncias ge néticas expressas em
por centagens de desacordo obtidas para 18c lones de ca fee iro diferenciad ores de ferru gembaseadas em 142 ma rcad ores RAPO. vari aram
entre 6% (entre os clones 17/1 2 e 128/2 ou .3 2/1 e128/2 ) 71% (entre os c lo nes 1.343/269 e 644/ 18).
A técnica RAPO foi efi ciente na caracteri zação
dos clones de cafeeiro diferenciad ores de
ferrugem. identi fic ando locos pol imórficoses pec íficos para cad a c lone.Ao nível de 23 % de di stância genética (F igur a2) . sete grupos fo ram definidos. o grupo A co mum c lone (644/ 18 qu e possui o fat or S H nãoidentificado). o g rupo B co m um c lone (8.32/ 1
que pos sui o fato r SH6 oriund o de Ccanephorai, o g rupo C com um clone ( 1343 /269
que possui o fator SH6 oriundo de Ccanephora ). o grupo D co m três clones (4 106.
4 19/20 e 220/10 que possuem os fatore s SH 6.SH7 . SH8 e S H9 presentes no Híbrid o deTimor). o g rupo E com dois c lo nes (147 /1 e 33/ 1
que possu em o fato r S H.3 oriundo de C. liber icai.o grupo F co m um c lo ne (1 53/2 qu e possu i o
fator SH3 or iundo de C. libericos e o gr upo Gcom nov e c lones (87 /1. 1006/10. 635/3. 134/4.110/5. 152/3. 32/ 1. 128/2 e 17/1 2 quecorrespond em aos genótipos de ori gem Ar ábica
com os fato res S H I. SH2. SH4 e SH5 ). Por tanto .644 /18 é o c lo ne mais divergente e a técn icaRAPO foi e fic iente na caracterização dos c lonese agrupam ento de ac o rdo com a or igem do sfatore s de resi st ên cia .

143
17/12128/232/1
152/3110/513414635/3
1006/10~.ª- 87/1·õ 153/2cO 33/1
14711420/10419/20
410613431269
832/1644118
~t=J-t}-'
D- I
liII
I
OI 03
DIStância Genénca
04 05 06 07
Figura 2 . Dendrograma representando distâncias genéticas estimadas entre 18 clones de cafeeirodiferenciadores de ferrugem baseadas em 142 marcadores RAPO gerados por 33 primers.
BIBLIOGRAFIA
BETTENCOURT, AJ. & RODRIGUES JR., CJ.J988. Princinples e practice of cofffeebreeding for resistance to rust and otherdisease. In: CLARKE, RJ. & MACRAE. R.(eds). Coffee, London, Elsiveir AppliedScience. p. 199-235. v.4:Agronomy.
DOYLE. J. J. & DOYLE. l L. 1990. Isolation ofplant DNA from fresh tissue. Focus, 12.(1):13-15.
WILLlAMS. lG.K.; KUBELlK. A.R.; LlVAK.x.r., RAFALSKI. lA.; TINGEY, S.V.1990.DNA polymorphisms amplified by arbitraryprimers are useful as genetic markers.Nucleic Acids Researchs, 18(22): 65316535.


145
GENETIC DlVERSITY OF CENICAFE COFFEE GERMPLASM COLLECTION
Marco Cristancho, I-Iernando Cortina, And German Moreno.
Disciplina de Mejoramiento Genético y Biotecnologia. CENICAFE. Chinchiná, Caldas. Colombia.
ABSTRACTThe genetic diversity of eENleAFE coffee germplasm collection was studied using the RAPD technique.DNA was extracted from semi-wild Ethiopian accessions of CofJea arabica by a modification of theprotocols of Vroh et al. (1996) and Mettler (1987). Ten random oligonucleotides (10-mer primers) wereused for the ampliflcation of cofJee DNA by the RAPD technique. Ali primers could detect polymorphismbetween the different cofJee introductions tested, showing the ample genetic variability ofthis material andtheir potential in cofJee breeding programs.Additionally, three RAPD markers described by Agwanda et al. (1997) and linked to the T gene that confersresistance to the coffee berry disease (CBD) were tested, for their presence in coffee plants of theeENleAFE germplasm collection. It was possible to reproduce easily two ofthe markers, M62027 and
N18250 although marker N18250 was ofa larger size (380hp) than initially reported by Agwanda (250bp).
Lately, we have had success reproducing the marker M20830 and the primer C03 usedfor the amplification
of this marker also amplified an M20600 marker associated to susceptible genotypes. A noveltyfinding of
this study was the discovery that the T gene was introduced into Hibrido de Timorfrom C. canephora ofthetype Robusta. This is the first report ofa RAPD marker linked to a specific gene to have been introducedinto Hibrido de Timor from one ofits parents.We could notfind a clear association between these RAPD markers and CBD resistant genotypes. MarkersM62027 and N 1258 were present in several CBD resistant and susceptible individuais of FI and F5progenies derived from the cross between Hihrido de Timor plants resistant to CBD and C. arabica Cl'.
Caturra plants susceptible to the disease. In order to have a better understanding of the segregation andligation ofthese RAPD markers. they are being transformed in/o seAR markers.
INTROOUCTION
Coffee (Coffea arahica) semi-wild germplasmhas been very poorly studied and just a few fieldcollections of wild coffee germplasm have beenovertaken mainly in the 60's. among them FAOand French collection missions.In CENICAFE a duplicate of the collection oftheFAO mission to Ethiopia in 1964-1965 (Meyer,1968) is currently being studied for its resistanceto leaf rust and coffee berry borer. These studiesare being also complemented by molecularcharacterization of the di fferent genotypes tested,trying to find markers linked to importantagronomic characteristics. Molecular studies ofcoffee genotypes with the RAPD technique haveconfirmed the small genetic variability present inthe commercial varieties and the relative highgenetic diversity present in serni-wild coffeegenotypes (Orozco-Castillo et aI. 1994:Lashermes et ai. 1996). Those stud ies have shownthe ample diversity of Ethiopian genotypes thatcan be exploited by the breeders.Besides the study of genetic diversity, the RAPDtechnique has been used in coffee to identifymarkers linked to resistance genes. Agwanda et
ai. (1997) could detect RAPD rnarkers linked tothe T gene that confers resistance to CBD. Byusing a large set of random primers they couldidentify three markers linked to this resistancegene. named M62027. M20830. and N 18250' The
markers could be detected in plants of generationsderived from the cross between Hibrido de Timor,the resistant parent, and different susceptiblecultivars.In this research we used the RAPD technique tostudy the genetic diversity present in semi-wildcoffee genotypes of the CENICAFE gerrnplasmcollection. Additionally, we investigated thepresence of RAPD markers linked to CBDresistance in several progenies derived from thecross between Hibrido de Timar and Caturra.
MATERIALS ANO METHOOS
ONA ExtractionCoffee DNA was extracted by a protocol adaptedat CENICAFE from the methods described byMettler (1987) and Vroh et al. (1996). Youngcoffee leaves were collected frorn treesmaintained in the field at CENICAFE mainresearch station. The leaves were washed with tap

water, dried and Iyophilized. Coffee lyophilizedtissue was ground to a powder with liquidnitrogen in the presence of 60mg PVPP, 200mgCT AB, 90 mg DlECA, and 25mg activatedcharcoal. Pre-heated (60°C) extraction buffer(20mM EOTA; 100mM Tris'HCI pH 8.0; 2MNaCI; 5% 2-mercaptoethanol) was added at afinal volume of 7.5mI/0.3g of Iyophilized tissue.Organic extraction was performed with achloroform iso-arnyl alcohol (24: 1) solution andthe ONA precipitated with isopropanol. RNA waseliminated by RNase A treatment.
RAPD AnalysisTen random l O-rner oligonucleotide primers(GIBCO) were used for the amplification ofcoffee ONA by the RAPO technique. The list ofthe primers used is the following: C02(CTGGGCAACT), C03 (AGGTCTTGGG), C04(GGTGAGGTCA), C05 (CTTCCGCAGT), C06(TGCTCTGCCC), C07 (ACTCCTGCGA), C20(ACTTCGCCAC), P05 (CCCCGGTAAC),ENT06 (CCCAAGGTCC), and CC5(TGAGTGGGTG). Primers C02, C03 and C04were also used for the amp Iification of markerslinked to CBO-resistance as described byAgwanda et ai. (1997).For the amplification reaction the mix containedlU ONA Taq polymerase (GIBCO), IX PCRbuffer, 1.8mM MgCI2, 0.2mM dNTP's, 0.2mg/ml
BSA and 15-20ng coffee ONA. The reactionconditions were as follows: 1 cycle at 94°C for 5min.; 40 cycles at 94°C for 1 min., 36°C for 1min., and noc for 2 rnin.: and 1 cycle at nDc for2 mino The amplification products were separatedin 1.5% agarose gels, visualized by EthidiurnBromide staining and analyzed in a PharmaciaImage Master® VOS.
RESULTS AND DlSCUSSION
Diversity of Semi-wild C. arabica AccessionsThe ten random primers used in the studydetected some degree of polimorphysm in thegenotypes assayed as determined by the presence01' absence of at least one band in one of the
146
genotypes. Examples of the variability present inthese genotypes are shown in Figures 1 and 2.We are currently assaying a larger set of primersin order to have a clearer picture of the geneticdiversity of the Ethiopian germplasm of theCENICAFE collection. This collection is beingtested for its resistance to rust and the coffeeberry borer in experiments in the field.
CBD-Resistance Linked MarkersWe attempted to reproduce the three RAPOmarkers described by Agwanda et ai. (1997) aslinked to CBO resistance. We were successful in
reproducing markers M62027' and N 18250
(Figures 3 and 4), and recently we could alsoreproduce rnarker M20830, which we could not
amplify as easily as the other two markers.Curiously with prirner C03 used to amplifymarker M20830 associated to resistant genotypes,
we also amplified a marker linked to susceptiblegenotypes (data not shown). This marker had asize of approxirnately 600bp (designated asM20600) and was not identified by Agwanda
with prirner C03.Marker M62027 was amplified in several Hibrido
de Timor trees, but it was not amplified in others.Ali these trees originated from accession CIFC1343. This suggests that this accession washeterozygous for the T gene (Figure 3). Thismarker was also amplified in the accessionBGB 138 of C. canephora of the type Robusta(Lane 13), but not in the accession 73 of the typeConilon (Lane 14). The band amplified in thesame size region in C. kapakata (Lane 15) is of asmaller size than the marker of 2027bp. MarkerN 18250 showed in our experiments a size of
380bp, larger than the 250bp size reported by
Agwanda (Figure 4). We do not know the reasonfor this difference in size between the two labs.Ali these rnarkers are currently being transformedat CENICAFE into SCAR markers in order tohave a more reproducible test and improve theselection ofCBD-resistance genotypes.
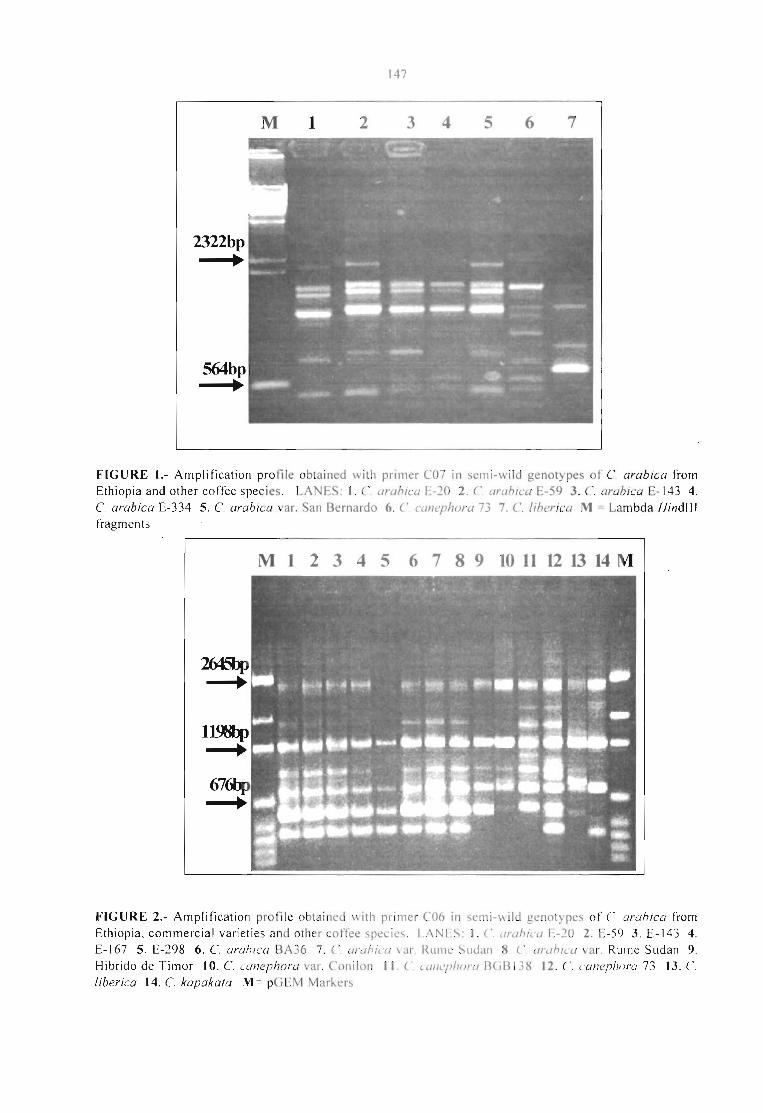
M 1 2
147
3 4 5 6 7
2322bp~
564bp~
FIGURE 1.- Amplification pro file obtained with primer C07 in sem i-wild gen otyp es of C ara bica fromEthiopia and other coffee spec ies. LANES: I . . arahicu 1:-20 2. C arab icu E-59 3. C arabica E- 143 4.C. arabica E-33 4 5. C. arabica var. San Bernardo 6. C cancphoru 73 7. C liher ica M -' Lambda Hindlllfragments
M 1 2 3 4 5 6 7 8 9 10 11 12 13 14 M
FIGURE 2.- Arnplificat ion pro fi!e obtained wit h prirner C06 in scmi-wild ge no ty pes o f C arabica frornEthiopia, commercial vari etie s an d other co rfee spcc ics . I.!\N I :~ : 1. r ' ara b ic:« F-2 0 2. E-59 3. E- 143 4.E-167 5. E-298 6. C. arabtca BA36 7. ('. ara bu:« \ ar . Rumc xudan S. C' arabicu var. Rume Sudan 9.Hibrid o de T imor 10. C. ca nepho ra var. Con ilon 11. C cancphoru BeiB I3X 12. C ca nephora 73 13. Cliberica 14. C. kap akat a M= pG EM Markers
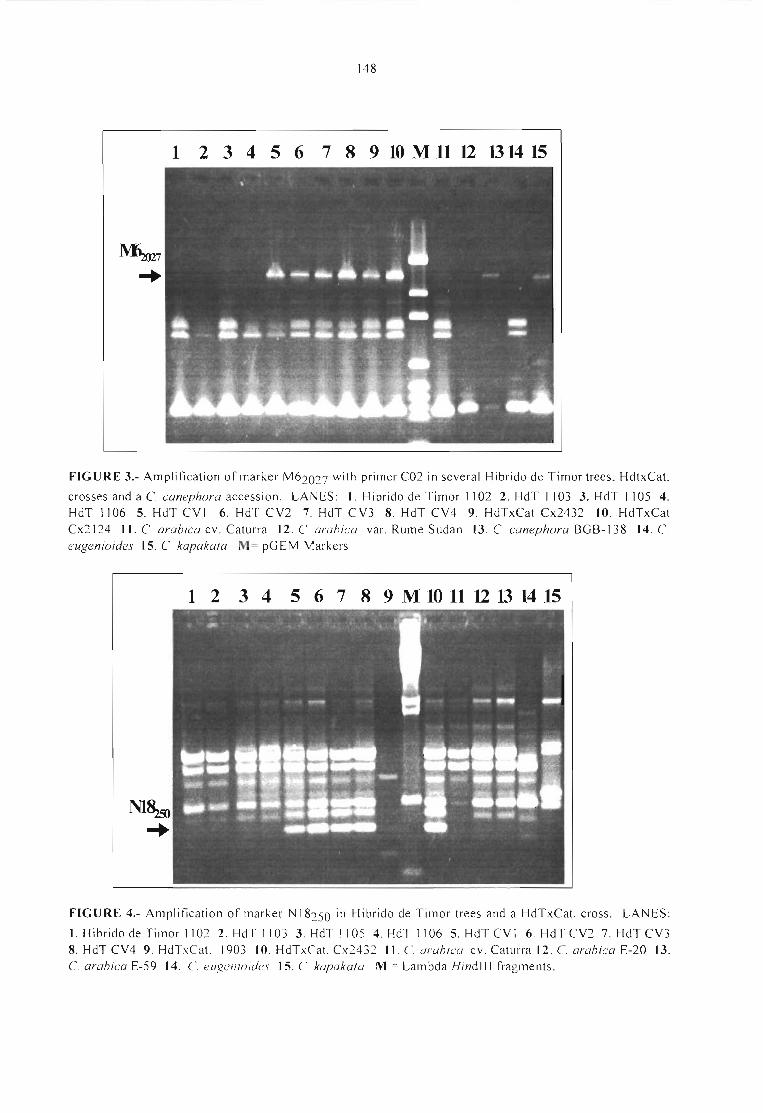
148
1 2 3 4 5 6 7 8 9 10 M 11 12 13 14 15
FIGURE 3.- Ampl ifi cation of marker M62027 with prirner C02 in severaI Híbr ido de Timor trees. HdtxCat.
crosses and a C canephora accession. LAN ES: 1. Híbrido de Timor 1102 2. HdT I 103 3. HdT I 105 4.HdT 1106 5. HdT CV I 6. HdT CV2 7. HdT C V3 8. HdT CV4 9. HdTxCat Cx2432 10. HdTxCatCx2 124 11. C arabica cv . Caturra 12. C arabica var. Rume Sudan 13. C canephora BGB-1 38 14. Ceugenioides 15. C kapaka!a M = pGEM M arkers
1 2 3 4 5 6 7 8 9 M 10 11 12 13 14 15
N1~...FIGURE 4.- A mpl i ficat ion of mark er N 18250 in Hibrido de Tim or trees and a HdTxCat. cross. LANES:
l. Hibrido de T imor 11 02 2. HdT 1103 3. HdT 1105 4. HdT 1106 5. HdT CV I 6. HdT CV2 7. HdT CV38. HdT CV4 9. HdT xCat. 1903 10. HdT xCat. Cx2 432 11 . C orahica cv . Caturra 12. C arab ica E-20 13.C arabica E-59 14. C eugenio ides 15. C kupa kata M = Lambda Hindlll fragments.

CONCLUSION
The initial screening of C. arabica Ethiopian
accessions of the CENICAFE col1ection by the
RAPO technique showed that there is a high
genetic diversity in these semi-wild genotypes.
We are currently using a larger set of random
primers to complete the molecular study of the
Ethiopian collection present at CEN[CAFE. As
expected, C. arabica commercial varieties seem
to have a smaller diversity than the semi-wild
genotypes.We demonstrated in this study that the genes
responsible for CBO resistance (at least the gene
1) in Hibrido de Timor were introduced in this
hybrid from C. canephora, one of its parents
(Moreno 1989; Lashermes et aI. 1993). RA PO
fragments present in C. arabica genotypes and
introduced from C canephora had already beenidentified by Lashermes et aI. (1993) and Orozco
Castillo et al. (1994). However, this is the first
study in that a RAPO fragment introduced from
C. canephora and linked to a specific gene has
been identified in C. arabica.It was not possible to establish a clear association
betweên the M62027 and N 18250 markers and
CBO resistant genotypes since they were
amplified in resistant and susceptible F4
generation plants derived from the cross between
Hibrido de Timor and Caturra (data not shown).
We have not used yet the M20830 marker and the
new identified M20600 marker to establish their
association to CBO-resistant and suscepti ble
genotypes in advanced generations of Hibrido de
Timor x Caturra crosses.
In order to have a better understandi ng of the
segregation and Iigation of these RA PO rnarkers,
they are being transfonned into SCAR rnarkers.
These markers shou Id work as an im proved
systern for the pre-selection of CBO-resistant
individuais that are sent to laboratory tests in
149
Portugal. They can additionally be used as RFLP
markers.
REFERENCES
Agwanda, C.O.. P. Lashennes. P. Trouslot. M.(Cornbes. and A. Charrier. 1997. ldenti fication oi'
RAPO markers for resistance to coffce bcrrv
disease, Colletotrichum kahawae, in arahil';!
coffee, Euphytica 97:241-248.
Lasherrnes, P.. J. Cros, P. Marmev. and i\.
Charrier. 1993. Use of random a 111 pt1fied [) N1\markers to analyze genetic variability aml
relationships of CofJea species. Genet. Res. CropEvol. 40 :91-99.
Lasherrnes, P.. P. Trouslot, F. Anthonv, r-.I.c.Combes, and A. Charrier. 1996. Genetic diver<;it\
for RAPO markers between cultivatcd and \lil:laccessions of Coffea arabica, Euphytica 87:S964.Mettler, I.J. 1987. A simple and rapid mcthcd rorminipreparation of ONA from tissue culturcd
plant cells. Plant MoI. Biol. Rep. 5:346-349.
Moreno. G. 1989. Etude du polyrnorphismc de
lHybride de Timor en vue de lameloriation du
cafeier arabica: variabi Iité enzyrnatiq uc ct
agronomique dans les populations dórigmc:
résistance incornpléte à Hemtleia vastatrix dans
les croisements avec Coffea arahicu. Montpe I1 icr.
Ecole Nationale Supérieure Agronomique lkMontpellier. 153p. (These Doctor-Ingenieur).
Orozco-Castillo, C; KJ. Chalrners. R. Wauuhand W. Powell. 1994. Detection of f.!en;;i~diversity and selective gene introgression in
coffee using RAPO markers. Theor. Appl. Genct87:934-940.
Vroh, 1.. L. Harvengt, A. Chandclier. Ci Mcrucai.and Ou Jardin. 1996. lrnproved R;\PDamplification of recalcitrant plant DNA bv thc
use of activated charcoal during DNi\ extraciion
Plant Breeding 115:205-206.

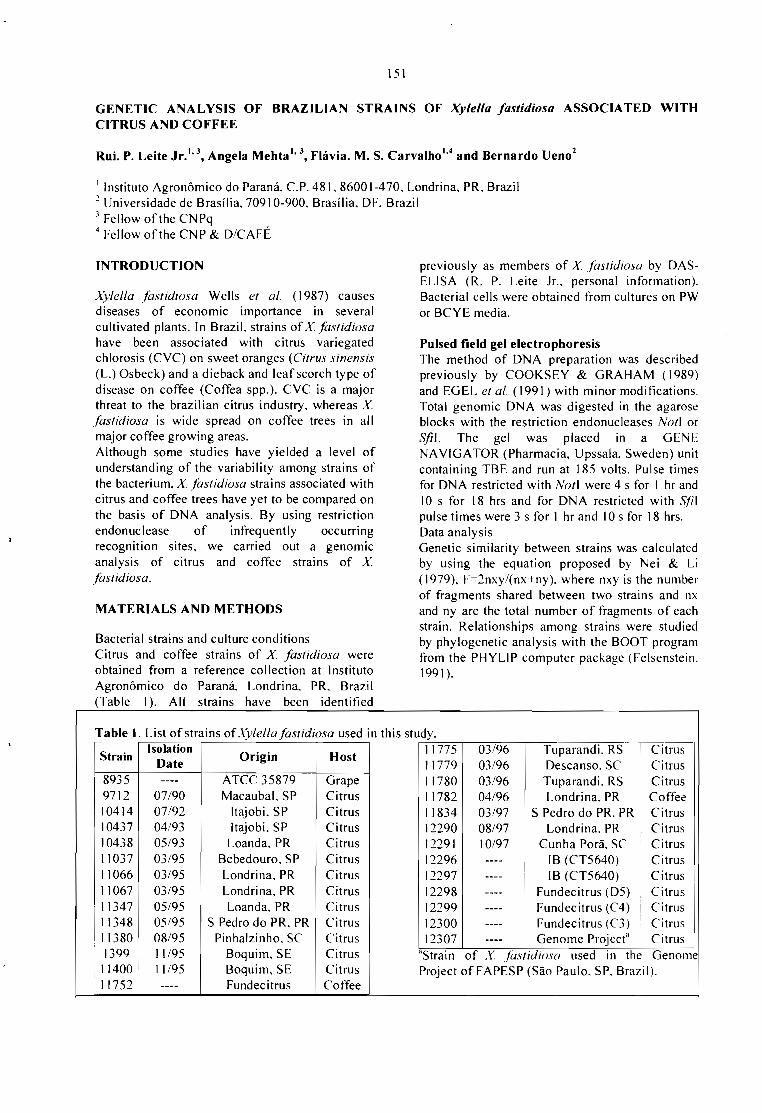
151
GENETIC ANALYSIS OF BRAZILlAN STRAINS OF Xylella fastidiosa ASSOCIATED WITHCITRUS AND COFFEE
Rui. P. Leite Jr:· 3, Angela Mehta l
• 3, Flávia. M. S. Carvalho':" and Bernardo Ueno 2
I Instituto Agronômico do Paraná, CP. 481.86001-470, Londrina. PR, Brazil2 Universidade de Brasília, 70910-900. Brasília, DF. Brazil3 Fellow ofthe CNPq4 Fellow ofthe CNP & D/CAFÉ
INTRODUCTlON
Xylella fastidiosa Wells et 01. (1987) causesdiseases of economic importance in severalcultivated plants. In Brazil, strains ofX fastidiosahave been associated with citrus variegatedchlorosis (CVC) on sweet oranges (Citrus sinensis(L.) Osbeck) and a dieback and leafscorch type ofdisease on coffee (Coffea spp.). CVC is a majorthreat to the brazilian citrus industry, whereas Xfastidiosa is wide spread on coffee trees in alimajor coffee growing areas.Although some studies have yielded a levei ofunderstanding of the variability among strains ofthe bacterium, X fastidiosa strains associated withcitrus and coffee trees have yet to be compared onthe basis of DNA analysis. By using restrictionendonuclease of infrequently occurringrecognition sites, we carried out a genomicanalysis of citrus and coffee strains of Xfastidiosa.
MATERIALS AND METHODS
Bacterial strains and culture conditionsCitrus and coffee strains of X fastidiosa wereobtained from a reference collection at InstitutoAgronômico do Paraná, Londrina, PR, Brazil(Table I) Ali strains have been identified
previously as members of X fastidiosa by DASELISA (R. P. Leite Jr.. personal information).Bacterial cells were obtained from cultures on PWOI' BCYE media.
Pulsed field gel electrophoresisThe method of DNA preparation was describedpreviously by COOKSEY & GRAHAM (1989)and EGEL et 01. (1991) with minor modifications.Total genomic DNA was digested in the agaroseblocks with the restriction endonucleases Notl OI'
Sfil. The gel was placed in a GENENAVIGATOR (Pharmacia, Upssala, Sweden) unitcontaining TBE and run at 185 volts. Pulse timesfor DNA restricted with Notl were 4 s for 1 hr and10 s for 18 hrs and for DNA restricted with Sfi Ipulse times were 3 s for 1 hr and lOs for 18 hrs.Data analysisGenetic similarity between strains was calculatedby using the equation proposed by Nei & Li(1979), F=2nxy/(nx+ny), where nxy is the numberof fragments shared between two strains and nxand ny are the total number of fragments of eachstrain. Relationships among strains were studiedby phylogenetic analysis with the BOOT programfrom the PHYLlP cornputer package (Felsenstein.1991).
Table I. List of strains ofXylella fastidiosa used in this study.
StrainIsolation
Origin Host11775 03/96 Tuparandi, RS ICItrus I
I
CitrusDate 11779 03/96 Descanso. SC8935 ---- ATCC 35879 Grape 11780 03/96 Tuparandi, RS Citrus9712 07/90 Macaubal. SP Citrus 11782 04/96 Londrina. PR Coffee10414 07/92 Itajobi. SP Citrus 11834 03/97 S Pedro do PRo PR Citrus10437 04/93 Itajobi. SP Citrus 12290 08/97 Londrina. PR Citrus10438 05/93 Loanda, PR Citrus 12291 10/97 Cunha Porão SC Citrus11037 03/95 Bebedouro, SP Citrus 12296 ---- IB (CT5640) Citrus11066 03/95 Londrina, PR Citrus 12297 ---- IB (CT5640) Citrus11067 03/95 Londrina, PR Citrus 12298 ---- Fundecitrus (D5) Citrus11347 05/95 Loanda. PR Citrus 12299 ---- Fundecitrus (C4) Citrus11348 05/95 S Pedro do PR, PR Citrus 12300 ---- Fundecitrus (C3) Citrus I
11380 08/95 Pinhalzinho, SC Citrus 12307 ---- Genome Project" 1 Citrus i1399 11/95 Boquim, SE Citrus "Strain of X fastidiosa used In the Genorne
11400 I 11/95 Boquirn, SE Citrus Project ofFAPESP (São Paulo. SP. Brazil).117521 ---- Fundecitrus Coffee

RESULTS
Genomic DNA of the citrus strains of Xfa stid iosa ass ociated with C VC when restr ictedwi th e ithe r Notl or Sfil endonucleases produceddi verse restriction patt erns (F ig . I). Incompariso n to the s tra in 8935 of g ra pev ine(ATCC 35879 ). the c itrus st ra ins had fewcomigrating DNA fragments. In contrast , thecitrus strains had restriction patterns with severa Icom igrating DNA fragments with the coffees trai ns 11752 and I 1782 (Fi g. 1). The genetics im ilarity estimated from th e restriction dataobtained with the combination of the two
152
restncnon endonuclease s ranged from 0.77 to1.00 a mong the citrus s tra ins (Table 2). Incompari son to the gr apevine and the coffeestra ins, the genetic s im ilarity ranged fro m 0 .3 1 to0.45 and 0 .69 to 0 .84. resp ect ivel y (Table 2). Thephyl ogentic tree reconst ructed based o n therestri ct ion fragment dat a sho wed three majorclades with the g rapev ine and the coffee strains asthe so le me mbers of two c lades (Fig. 2). The cladecomprising the c itru s stra ins can be subdi vided intw o subcl ades with one formed with the stra insfro rn the sta te s of São Pau lo and Sergipe . and theother with stra ins from Paraná. Santa Cat arina andRio Grande do Sul states (Fig . 2).
M 23 4 5 6 7 8 9 10 11 12 13 14 15 16
242 .5 -
194.0 -
145. -
970
485-
Fig, I . Restriction endonuclease patterns of strains of x.fa stidiosa restricted with Sfi l. Lanes : M. Lambdamarker; I. 89 35 ; 2 . 11037; 3. 10437; 4 . 10414 ; 5. 123 00: 6. 12299 ; 7. 122 98; 8. 12296 ; 9 . 122 97 ; 10. 12290;I I. I 1779; 12. 12291 ; 13. 11 067 ; 14. 971 2: 15. I 1752: 16. I 1782. Molecu lar sizes are given in k ilobase s .

153
Table 2. Genetic similarity matrix for strains of X fastidiosa based on restriction endonucleose analysis ofgenomic DNA fragments separated by pulsed field gel electrophoresis.
Strain Citrus Coffee Grape
122 122 122 122 117 122 11752 893510437 10414 12300
99 96 97 90 79 91
10437 - 0.87 0.87 0.90 0.97 0.94 0.93 0.77 0.77 0.69 0.3110414 - 0.93 0.93 0.90 0.87 0.93 0.83 0.83 0.73 0.3312300 - 0.96 0.90 0.87 0.93 0.90 0.90 0.73 0.3212299 - 0.93 0.90 0,96 0.86 0.86 0.76 0.3412296 - 0.98 0.90 0.80 0.80 0.71 0.3212297 - 0.87 0.77 0.77 0.69 0.3112290 - 0.83 0.83 0.73 0.3311779 - 1.00 0.84 0.4512291 - 0.84 0.4511752 - 0.50
II8935 -
3
23
43
2
100
___ 12307 - Genom e PrOJ ect
___ 11037 - Bebedouro, SP
___ 11400 - Boquim, SE
L..__ 11399 - Boquim, SE
r--- 09712 - M e ceubal, SP
.....__ 10414 -Itajobi, SP
L..-__ 10437 - Itajobi, SP
r--- 11380 - P inhalzinho , SC
___ 11780 - Tuparandi, RS
L..__ 11779 - Descanso, se
11834 - São Pedro do Paraná,
r---- 11067 - Londrina, PR
10-__ 11066 -Londrina, PR
r---11775 - Tuparandi, RS
r---11347 -Loanda,PR
L..__ 10438 - Loanda, PR
___ 11348 - São P edro do Paraná,
.....__ 11752 - Coffee
......-- 8935 - Grape
Fig. 2. Unrooted phylogenetic tree for strains of X fastidiosa inferred from restriction analysis of DNAfragments, generated by the soor program from the PHYLlP computer package. Numbers in each nodeindicate the bootstrap percentages.

DISCUSSION
The results obtained in this work demonstratedthat strains of X fastidiosa associated with citrusare genetically diverse. The phylogenetic analysissuggested a grouping based on geographic originof the strains. Strains of the bacterium from statesof São Paulo and Sergipe are closely related whilestrains from Paraná, Santa Catarina and RioGrande do Sul states form another clade. Further,the coffee strains of X fastidiosa are geneticallyclosely related to the citrusstrains. In contrast, the citrus and coffee strainsare not closely related to the grapevine strain onthe basis of restriction analysis of DNAfragments. The genetic relationships and diversityof X fastidiosa obtained from citrus and coffeesuggest that the bacterium is endemic to BraziJ.Furthermore, the relatedness of the citrus andcoffee strains of X fastidiosa also indicates thatthose strains rnay have a common geneticbackground.
154
REFERENCES
Cooksey, D.A., and J.H. Graham. 1989. Genomicfingerprinting of two pathovars ofphytopathogenic bacteria by rare-cuttingrestriction enzyrnes and field inversionelectrophoresis. Phytopathology, 79:745-750.Egel, D.S., lH. Graham and R.E. Stall. 1991.Genomic relatedness of Xanthomonas campestrisstrains causing diseases of citrus. Appl. Environ.Microbiol., 57:2724-2730.Felsenstein, l 1991. PHYLlP, PhylogenyInference Package, version 3.4. Departament ofGenetic. University of Washington, Seattle.Nei , M. and W. Li . 1979. Mathematical modeJfor studying genetic variation in terms ofrestriction endonucleases. Proc. Natl, Acad. Sei.76:5269-5273.Wells. lM.. B.C. Raju., H. Hung.. W.G.Weisburg.. L. Mandelco-Paul and D.J. Brenner.1987. Xylellafastidiosa gen. nov., sp: nov.: Gramnegative, xylem-limited fastidious plant bacteriarelated to Xanthomonas spp. Int. J. Syst,Bacteriol..37: 136-143.

155
BIOCHEMICAL AND MOLECULAR STUDIES OI' THE MAIN PROTEIN IN THE COFFEEENDOSPERM
Ricardo Acuüa', Ronald Bassüner", Vadim Beilinsorr', Hernando Cortina" Gabriel Cadena-Gómez',Virginia Montes' and Niels Nlelsen'',
'Centro Nacional de lnvestigaciones dei Café. Chinchiná.Caldas, Colombia."Departamento de Quirnica, Universidad Nacional. Bogotá. Colombia.1 United States Department of Agriculture , Agricultural Research Service and the Departments ofAgronomy and Biochemistry, Purdue Univbersity, West Lafayette, IN 47907. USA.
INTRODUCTION
Coffee bean. the raw material from which alicoffee producs originate, is derived from the seedsof Coffea spp. Despite the inmense economicimportance of coffee bean, little is known aboutthe biochemical and molecular properties of itsseed proteins. Only values for the protein contentof coffee beans have been reported in the range od8.7-12.2% (Macrae, 1985). Work has focused onthe characterization of water soluble proteins indifferent coffee varieties (Centi-Grossi et. aI.1969; Amorim et. aI. 1975) OI' coffee beansprocessed by different methods (Underwood andDeatherage, 1952).The information provided inthese reports describes molecular mass profilesand protein content. Nothing is known about thestructure of genes that encode the proteins, theidentity of the primary gene products, OI' theregulation of their expression.This work describes the identification of aJegumin-like seed storage proteins ofthe 11S sizeclass (Nielsen et. aI. 1997) in coffee. Two cDNAsthat encode the proteins are descri bed, a promoterfor an 1 IS gene is showed. and the subcellularlocalization of the protein in the seed endospermis documented.
MATERIALS AND METHODS
Coffee endosperm were harvested about 150 and
220 days after flowering, and stored at -80°C unti Iuse. FOI' imrnunocytochemistry, the endospermwas harvested about 220 days after flowering andfixed with 3% glutaraldehyde in phosphatebuffer.Extraction and fractionation of total seedprotein was rnade from frozen irnrnature OI'
mature coffee seeds.
Sequential fractionution of coffee seed proteins:seed proteins were extracted sequentially frommature coffee endosperm in "albumin extractionbuffer": pH 4.5. (Sluyterman and Wijdens 1970).phosphate buffer ("Globulin extraction buffer")and the residue remaining was extracted a with70% absolute ethanol and 10 mM DTT and was
considered to be the "Prolamine Fraction". Totalcoffee seed proteins were fractionated in asucrose gradient 6 to 22%) and samples from eachfraction, were analyzed by electrophoresis.
N-Iermina/ sequence ana/ysis Endosperm proteinsamples were electroblotted and the legumin-likecofffee protein was cut from the stained blotmembrane and processed in a model A491 proteinsequencer.
Extraction and purificution of mRNA fromimmature coffee endosperm: Total RNA wasisolated, as described by Schultz (1994) forrecalcitrant plant tissues. from frozen inmaturecoffee endosperm. To prove translatability of theisolated mRNA. an in vitro mRNA cell-freetranslation was carried out in a reticulocyte Iysate.
cDNA cloning: Synthesis of full-Iength cDNAwas made using the TirneSaver" cDNASynthesis Kit (Pharrnacia). An EcoR I/Nol Iadaptor was ligated to the ends of the resultingfragments. A sense oligonucleotide primer wassynthesized on the basis of the N-terminal aminoacid sequence determined for the alpha chain ofthe coffee legumin. A second antisensedegenerate oligonucleotide was synthesized basedon N-terminal amino acid sequence determinedfor the beta. These primers were used to amplifynearly the entire acidic legumin chain. The PCRfragment was cloned into a pCRII vector and bothstrands were sequenced using an ALF automatedDNA sequencer. Based on the nucleotidesequence that was obtained, two additionaloligonucieotide primers were synthesized andused in combination with a short specific primersdesigned on the basis of the EcoR I/Not I adaptorsequence. These were used to amplify the basicchain cDNA and the N-terminal signal peptide.respectively. The overlapping sequences that wereobtained were aligned to yield a full-Iength cDNAsequence.
DNA blot hybridization. Coffee genomic DNAwas digested with various restriction enzyrnes and

the restnction frag ments were separated inaga rose ge ls. A vacuum blott ing sys te m was usedto transfer the DNA fragm ent s from the ge l to amembrane us ing an alk al ine vac uum transferprot ocol. An Eco R 1 DNA fragme nt (-900 bp)that co rresponed to the ac idic chai n of coffeelegumin was purified by agarose ge le lec tro pho resis and lab eled and used forhibrid iza tio n.
Ele clron microscopy immunocytoch emistry.Plant materia l was cut into smal l bloc ks of about Imm length and immed iate ly fixed in 0.1Mphosph ate buffer, pH 7.3. co nta ining 3%glutara ldehyde. Embedding was per formed in LRWhite and in Epon resins. The sec tions wereprehybridized in blocking solut ion, followed byhybridization with purified antibodies aga instcoffee 1IS proteins. Hybrid ization took place forsevera l hours on a rocking platform. Aft erwashing in TBST. the grids were incubated withanti -rabbit IgG-gold conjugatc dilute d 15-fold intoa blocking solution. This was followed by washesin TBST and water. As a final step, the gridswere incubated for two minutes in 1%g luta ra ldehyde, followed by a rinse with water.To enhance contrastothe sections were tre ated in aLKB 2 168 UltraSta iner with a saturated so lut ionof uran yl acetate, pH 4 .0. for 20 min at 26°(, andthis was followed by incubati on for 2 minutes at20 °C with 83mM lead acetate , pH 12. Finally, theth in sec tions we re carbon coated in a vacu umevaporato r (Ladd Research Indu st ries).lnspections under the e lec tron microsco pe (JE M100 CX , Joe l. Japan ) were ca rried o ut at 80kV.
200, 1 6
'7
J1
20
1 4
156
RESULTS
A legu rnin-like seed pro tei n was pu rifi ed from theendosperm of coffee (CojJea arabica L. cv .Co lombia ). In co ntrast to legumes . w he re efficientsto rag e globulin extrac tion requ ires bufferedsa line slolut io ns we ll above the ac id ic pK, of theglobuli ns. coffee legumin is rcadl y extrac ted withacidic aqueo us buffers.. Subun its of coffeeleg umin have a M, of a bo ut 55 kDA afte r onedimensi onal SDS-PAGE ele ctrophoresis in theabs enc e of a redu cing agent. In the pre sence of 2mercaptoethan o l. two po lypeptides appear thathave apparent molecular masses of 33 and 24 kDa(Figure I ). T he coffee legu m in migrates like other11S storage glob ulins in suc rose grad ients (Figure2) . T wo full- Ien gth cDNAs were generated fromrnRNA of de ve loping seeds that were more than98% homcl ogous (Figure 3) . They had openread ing frarnes of 1458 and 1467 bp. Eachenco ded legu m in precur sor s of 486 and 489 aminoacids. rcspective ly ((M r= 54 136 and 54818 ).F~:x amin ati on 01' a 5 ' prornoter region lrom a coffeelegumin gene rev ealed a pu tative legu min -box (figure4). Genorni c ONA fr orn C ar ábica wa s d igested withsix d iíferent restric tion endo nuc lcases. Atter sepuration
0 [' lhe fragments by electropboresis. single discretefragments on DNA blots hybridived strong ty to acO A probe for the acidic chain (figure 5 ). DNA lrornotber species and commercially irnport ant cultivars thatcomprise lhe genus Coffea produccd similar results(Figure 6). lrnmunocytochernical studies revealed ihatsome legumin was detccted in the cytolasm in maturecottee seeds. but that the rnajorit y 01' it was in largestoragc vacuoles that accounted for most 01' lhe cellvolume (Figure 7) .
Fig, I . Proteins extracted from mature and immature coffee endo sperm . One-dimensional SDS-PAGL separationof seed proieins extracted in SOS buffer in prcserice (Ianes 2-5) 01' abscncc (lancs 6-8) 0 1'2-ML Numbers in lhe rightmargin reter to molecular mass in kUa ofprorcin standards, l.ancs I and 9 sizc markc rs, lanc 2 soybeun conglycinin.lanes 3 and 8 total secd protcin frorn faba bcan. lanes 4 and 7 immaturc colfcc sccd procins cxtractcd. lancs 5 and 6 totalsee d prote ins 1'1'0 111 co ífc c .
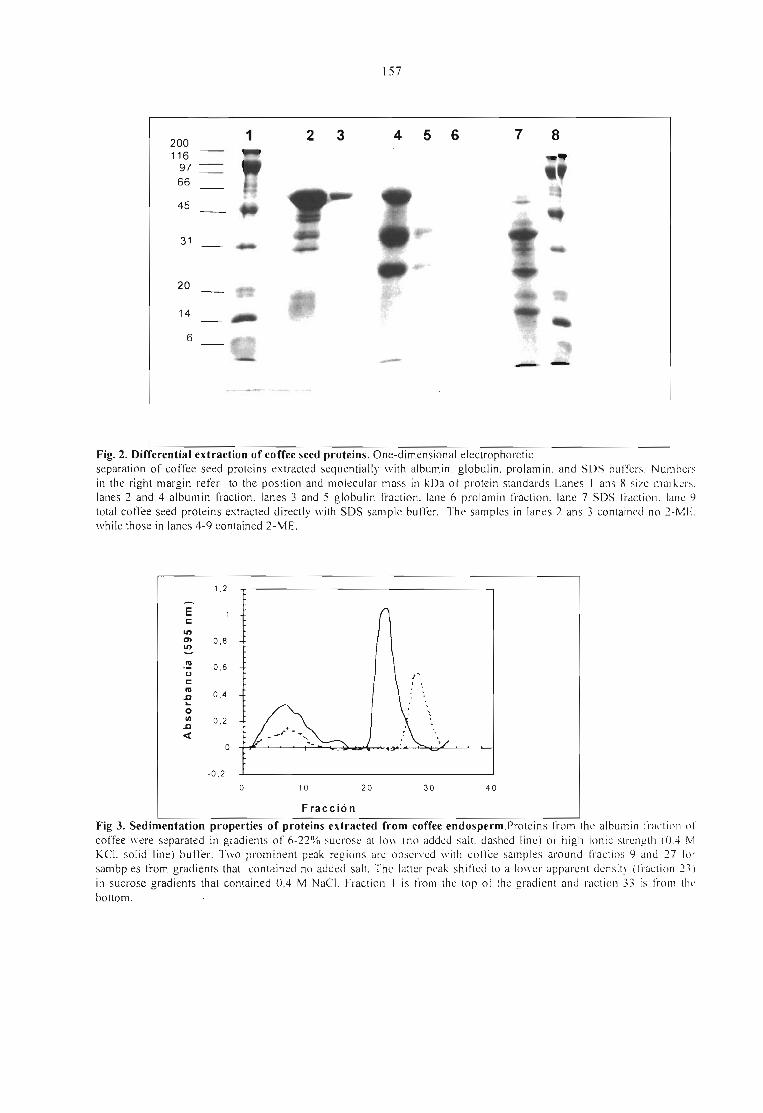
157
200 1 2 3 4 5 6 7 8116
976 6
45
31
20
14
6
- --
Fig. 2. Differential extraction of coffee seed proteins. One-di mensiona l electrophorct icseparation of coffee seed proteins extracted sequcntially with albumin. globul in . prolam in. and SDS bulfcrs. Numbcrsin the right margin reter to the posit ion and mole cul ar rnass in kDa 01' protcin standard s l.anes I ans 8 ,in: murk crxlanes 2 and 4 album in lraction. lanes 3 and 5 g lobul in fraction. lane 6 pr o lamin íraction. lane 7 SOS Iraction. lanc CJ
lo tai coffee seed proteins extracted directly with SDS samplc but fer, The sarn ples in lanes 2 an s 3 co ma incd 11 0 2-MF.while those in lanes 4-9 contain ed 2-M E,
1 ,2
Er::::
li')
(J) 0 ,8li')
1Il 0,6(,J
r::::1Il
0,4.rJ...oli) 0 ,2
.rJ
<l:o
-o,2
o 10 2 0 30 4 0
F racció n
Fig 3. Sedimentation properties of proteins extracted from coffee endosperm .Prot cins from the alburn in íruct ion 0 ['
co tfe e were separated in gradient s o f 6-22% sucrose at 10\\ (n o addcd salt. dashcd line) OI' high ionic strcngth (0 ,4 MKCI. solid line) buffer. '1'11'0 prorninent peak regions are obscrvcd with co ít ce sarn ples around tractins CJ und 27 lorsambp les frorn grad ients that coru ained no added salt, T hc lauer pcak s h iílcd to a lowc r apparcnt dcnsity (Iracti on 23)in sucrose gradients that contained 0.4 M Nael. Fraction I is from lhe 10p 01" lhe gradicru and ract ion 33 is from lhe:bottorn .

180177
158
aacacactacactctcctctgttgtcagagaatggctcactctcatatgatttctctttcctlgtaCgttcttllttgttcctCggctg 90aacacactacactctcctctgttgtcagagaatggctcactctcatatgatttctctttcct gtacgttctt ttgttcctcggctg.87
M A H S H M I S L S S/L Y V L L/- L F L G C 20
tttggctcaactagggagaccalagccaaggctclggggtaaaactcagtgclatattcagaagcttaatgcacaagaaccatccttcaotttggctcaactagggagacca agccaaggctc ggggtaaaactcagtgc atattcagaagcttaatgcacaagaaccatccttcag
L A O L G f P E/O P R L G/R G K T O C N/O I O K L N A O E P S F R 53
gttcccatcagaggctggtttaactgaattctgggattctaataatccagaatttgggtgcgctogtgtggaatttgagcgtaacactgt 270gttcccatcagaggctggtttaactgaattctgggattctaataatccagaatttgggtgcgctggtgtggaatttgagcgtaacactgt 267
F P S E A G L T E F W O S N N P ~ F G C A G V E F E R N T V 83ccaacctaagggccttcgtttgcctcattactctaacgtgcclaaattcgtctacgttgtcgaaggtaccggtgttcaaggcactgtgat 360ccaacctaagggccttcgtttgcctcattactctaacgtgcclaaattcgtctacgttgtcgaaggtaccggtgttcaaggcactgtgat 357
O P K G L R L P H Y S N V P K F V Y V V E G T G V O G T V I 113ccctggttgtgctgaaacatttgaatccCaggglgaatcattttlgggtggtcaggaaCagCCgggCaaagggCaagaagg_ 441ccctggttgtgctgaaacatttgaatcccaggg gaatcatttt gggtggtcaggaacagccgggcaaagggcaagaagg 447
P G C A E T F E S O G E S F S/W G G O E O P G K G O E G -/O-/E-/ 143
IIt t c c a a a g g t g g t c a g g a a g g g c l a a g g c a a a g g t t t c c a g a c c g c c a t c a g a a g c t c a g a a g g t t c c a a a a a g g a g a t g t c c t t a tttccaaaggtggtcaggaagggc aaggcaaaggtttccagaccgccatcagaagctcaoaaggttccaaaaaggagatgtccttat
O-/G S K G G O E G O/R R O R F P O R H O K L R P F O K G O V L Iattgcttcctggtttcactcagtggacatataatgatggagatgttcclcttgtcactgtcgcacttcttgatgttgccaatgaggctaaattgcttcctggtttcactcagtggacatataatgatggagatgttcc cttgtcactgtcgcacttcttgatgttgccaatgaggctaa
L L P G F T O W T Y N O G O V P L V T " A L L O V A N E I'. Ntcagcttgatttgcagtccaggaaatttttcctagccgglaacccgcaacagggtggtggaaaggaaggccatcaaggccagcagcagca
528537
li"618627205708
tcagcttgalllgcaglccaggaaatltttcctagccgglaacccgcaacaggg1991ggaaaggaaggccatcaaggccagcagcagca 717O L O L O S P K F F L A G N P O O G G G K E G H O G O O O O 235
gcatagaaacatcttctcaggatttgatgaccaacttttggclgalgctttcaatgttgacctcaaaataatacagaaattgaagggtccgcatagaaacatcttctcaggatttgatgaccaacttttggc ga gct~tcaatgttgacctcaaaataatacagaaattgaagggtcc
H P N I F S G F O O O L L A E/O A F N V O L K I I O K L K G PgaaagatlaaaggggtagCaCagtcCgagctgaaaaacttcaactgttcctgCCtgaatatagtgagcaagagcaacaaccccaacaacagaaaoat aaaggggtagcacagtccgagctgaaaaacttcaactgttcctgcctgaatatagtgagcaagagcaacaaccccaacaaca
F O V/O P G S T V R A E K L O L F L P E Y S E O E O O P O Q Q
79880726688889"7297
gcagglgcagcaacaacalggtgttggaagaggatggagatccaalggacttgaggaaactttgtgcacggtgaagcttagtgaaaacat 978gcagg gcagcaacaaca ggtgttggaagaggatggagatccaa ggacttgaggaaactttgtgcacggtgaagcttagtgaaaacat 987
O G/E Q O O O/H G V G R G W ~ S N G L E E T L C T V F L S E N 329
tggcctcccccaagaggctgatgtattcaatcctcgtgctggccgcattaccactgttaatagccaaaagattcctatcctcagcagcct 10E8tggcctcccccaagaggctgatgtattcaatcctcgtgctggccgcattaccactgttaatagccaaaagattcctatcctcagcagcct 1077
G L P O E A O V F N P R A G R I T T V N S Q K I P I L S S L 359ccaacttagtgcagaaagaggattcctctacagcaatgccatttttgcaccacactggaatatcaatgcacatagtgccctgtatgtgat 1158ccaacttagtgcagaaagaggattcctctacagcaatgccatttttgcaccacactggaatatcaatgcacatagtgccctgtatgtgat 1167
O L S A E P G F L Y S N A I F A P H W N I N A H S A L Y V I 389tagaggaaatgcaagaattcaggtggtggatcacaaaggaaacaaagtttttgacgatgaagtaaaacagggtcagctaataattgtgcc 1248tagaggaaatgcaagaattcaggtggtggatcacaaaggaaacaaagttt~tgacgatgaagtaaaacagggtcagctaataat~gtgcc1257
R G N A P O V V O H K G N K V F O O E V K Q G Q 1 I I V P 419a~aatar.tttgctgtgatcaagaaagctggaaaClaaggàtttgagtaCqttqCattcaagacgaacgacaatgccatqattaacccact13383caatactttgctgtgatcaagaaagctggaaac aaggatttgagtacgttgcattcaagacgaacgacaatgccatgattaacccact 1347
Q F ~ K K A G N E/Q G F E ., 7 A F K T N D N A M T N P L 450
Ig t t g g a a g a c t t t C g g Ca t tlC ga g C a a t t c c t g a g g a a g t t t ~ g a g g a g c t c t t t c ca a a t t t c c a g cg a g ga a g c t g a gg a a t t gaa 1428gttggaagactttcggcatt cgagcaattcctgaggaagttttgaggagctctttccaaatttccagcgaggaagctgaggaattgaa 1437
G R S A L/F P A P E E V L R S S F Q S S E E A E E L K 48]
gtatggaaqacaggaogctttgcttttgagtgagcagtctcagcagggaaaaagIgaagttgcttgagctaattatgtaaaaataatCgtgtatggaagacagqaggctttgcttttgagtgaqcagtctcagcagggaaaaag gaagttgcttgagctaattatgtaaaaataatcgt
Y G R Q E A L L L S E Q S Q Q G K R E V AatattagtccatgcatgqtctaccaactatatgtgtgaatctaattccaaaataaaatggtcaatggatgtaaagacatggcaatcctagatatcagtccatgCatggtctaccaactatatgt
ccttactactggcgttgattgcgagaagtttgatgtttggtgaccatgagtcaataataaactatgataattaatgt
15181527
16081561
168')
lig. 4. The cDNA nucleotide sequence for two coffee legumins and their deduced amino acid sequences. 'lhc Nterminal amino ucid scqucnccs for ucidic and basic chains that wcrc determines chcrnically are shown h) solidhorivorual tines ahox c thc scqucnccs. Clone [1C;\4 includcs bases 127-\ 023. [1C;\3 extcnds frorn 5" linkcr to base 216.and clone [1C;\9 cxtcnds frorn base 20!\ to thc 3'linker. Thc oligonuclcotides dcnotcd hy thc lcft and right Iacing opcn
aITOI\S pcrrnittcd full coding rcgions of coflcc lcgumin to hc amplificd. Diffcrcnccs hctwccn thc nuclcotide codingregions are indicated in reverso t) pc. and thc dcduccd amino acid scqucnccs are providcd abovc thc nuclcotidc
scqucnccs. The sing lc vertical arrow (,l,) indicares thc position whcrc thc signal pcptidc is clcavcd frorn thc prolcgum inwhilc thc short. doublc vertical arrow ( D ) idcntifics whcrc post-translationa] modification takes placcs to torm thcacidic and hasic chains of mature suhunits. l lppcr scqucnccs corrcsponds to acccssion nurnbcr lI64443 and the lowcrscqucnce to ;\F054895.

159
- - - '" - - - -E - c o " o .c ".. 0.0 ~ : Co " " " Co .c
'" '" '-' Q ;.c: '" Z 7. CJJ ~
I I I I I I I I I I I I I I Ir
i- -II .
23 1 K b -I9 4 Kb-I
66 K b-!,
4 4 K b-1 . - -2 3 K b -I -2 OK] .1 .4 Kb- -1 o Kb-
Fig. 5. Genomic DNA blot analysis of then difTerent species and cultivars from the Coffea family.Genomic DNA blot analysis of ten differnt species and cultivars from Coffea family. Genomic DNA wasdigested with three restriction endonucleases (A Xba I. BEco RV. C Eco R I). DNA fragments after thistreatment were separated in a 0.8% agarose gel, transferred to a nitrocellulose membrane. and probed withthe DNA insert from pCA4 at high stringency. The insert corresponds to the mature acidic chain and Nterminal of the basic chain. Species I. Hibrido de Tirnor: species 2. C. eugenoides: species 3 C liherica:pecies 4 C kapakata: species 5. C. arabica cv. Caturra: specíes 6. C. canephora cv. Conilon: species 7. Ccanephora cv. Robusta: species 8 C. arabica x C.. canephoru: species 9. C. arabica: specíes 10. C arabu:acv. Colombia
-361
-281
-201
-121
-41
caaqatqaatqtqtqtttqatttqqqqtttqattcatcaaaaqccatcqtaqcaqataatqcaccttaccatqccattqc
taaaqtacaaaaatttcatqcaaatacaaacacaaaaqqattqaacaatacatqtcaqaaactcattqccaccaaqqcttlD!I!!a ta9!--Et•.•t~alEtcG*
acacatcatccttqqtqtaaaqaaqtlD!l!!tct~a~ .••a~agmaqccaaqtqtaaaatqaaaat
tttqacqtqtcqattcctcatcttccaattacatq~aaqqaqccatttccaaqctctaatcqccqcatcccctca
ccacaaaaacacactacactctcctctqttqtcaqa~
Fi~ 6. Nucleotidc scqucncc 01'lhe promotor for a coffce lcgumin gene. lhe first 400 bp 01" UNA upstrcarn from the bcginning ofthccoding region for a coffce lcgumin gene is shown. A putativo lcgumin hox is cornparcd with a conscnsus scqucncc derivcd lromlegume genes. Homology is indieated for a nuclcotide identificd o~ reverso printing. Central to the lcgumin box is a degenerareCATGCAT elerncnt (undcrlined). As it is typical of corrcsponding genes [rom legumes. othcr CATCiCAT clcrncnts are prcscnt in
the upstream region. ATATA box clcmcnt (doublc undcrtincd í is also prcscnt.
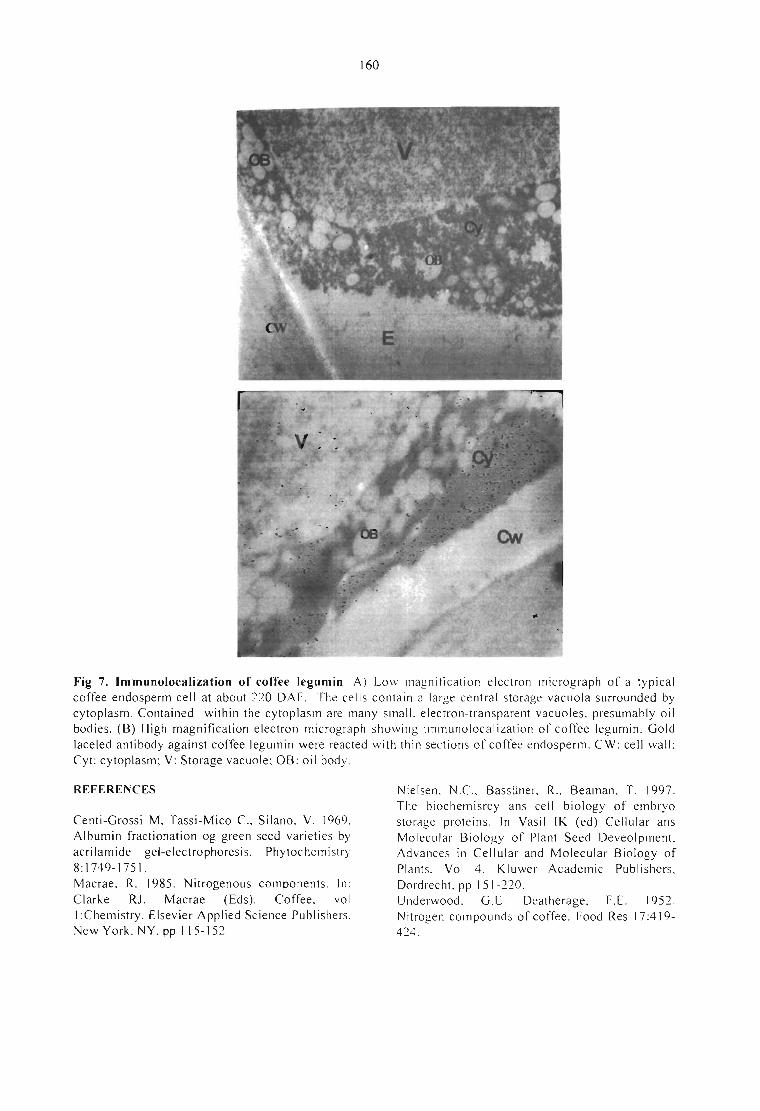
160
Fig 7. Immunolocalization of coffee legumin . A) L ow magni ficat ion electron mic rograph 01' a typ icalcoffee endosperm cel l at about 220 DA F. The cel ls conrain a large central stora ge vac uo la surr ounded bycy to plasm, Contained wit hi n the cytopl asm are many smal l. electron-transparent vacuo les. presurnably oilbodies. (B ) High mag nificati on electron m icrograph sho w ing irn rn uno local izat ion 01' co f fee legumin . Goldlaceled antibody against coffee legumin were reacted with th in sections 01' co f fee endos pe rrn . CW : cell wa l l :Cyt: cy toplasm; V : Storage vacuo le: OB : Gil body :
REFERENCES
Cent i-Grossi M. Tassi-Mico c.. Si lano. V . 1969.A lbum in f ractionation og green seed varieties byacri lam ide gel-e lect rophores is. Phyt och ernistry8: 1749- 1751.M acrae. R. 1985. Nitrogenous componen ts. In:C larke RJ. M acrae (Eds). Coffee. vo lI .Chern istry. Elsev ier A pp lied Science Pub lishers.N ew York. N Y . pp 115-1 52
Nielsen. N.C.. Bass üner. R.. Bearnan, T. 1997.T he biochemis rey ans cel l biol ogy 01' emb ryosto rage pro tein s. In Vasi l IK (ed) Ce l lu lar ansMolecular B io logy 01' Plant Seed Deveo lp men t.A dvances in Cel lular and Molecul ar B iol ogy ofPlants. Vol 4 . Klu wer A cadem ic Publ ishers.Dord recht. pp ] 5 1-220.Underwood . G.E. Deatherage , F.E. 1952.N itrogen compound s 01' coffee. Food Res 17:4 19424.

161
USO DE PADRÕES ELETROFORÉTICOS NA DIFERENCIAÇÃO DE CULTIVARES E DONíVEL DE DETERIORAÇÃO DE SEMENTES DE Coffea arabica L.
Brandão Jr., Delacyr da Silva I • Vieira, Maria das Graças Guimarães Carvalho", Bernadino Filho.José Roberto'' , Hilhorst, Henk ~ , Guimarães, Rubens José'
'Aluno de Doutorado do Curso de Fitotecnia - UFLA: - 2Pro fe ssor Titular do departamento de Agricultura UFLA;- 'Aluno de Graduação do curso de Agronomia - LJFLA: - ~Professor de Fisiologia de Sementes Wageningen Agricultural University - Holanda; - 'Professor Adjunto do Departamento de Agricultura
UFLA.
INTRODUÇÃO
Com a tecnificação da agricultura. novos híbridose cultivares de cafeeiro têm sido desenvolvidosvisando atender as mais diversas exigências emtermos de mecanização. produtividade. resistênciaà pragas e doenças dentre outras. Para assegurar apureza genética de sementes. especialmente doshíbridos. órgãos responsáveis pela certificação desementes têm procurado técnicas eficientes.seguras. rápidas de fácil treinamento. a seremutilizadas na identificação e caracterização decultivares. Tradicionalmente a identificação decultivares tem sido feita por caracteresmorfológicos. o que exige estudos detalhados desementes e plantas em crescimento. envolvendoum número recorde desses descritores. Além deser um processo caro. envolvendo grandeconsumo de tempo e áreas extensivas. muitosdesses descritores sofrem influência ambiental.Vários estudos têm enfatizado a importância e aversatilidade do uso de marcadores molecularesna identificação de cultivares. Dentre as técnicasdisponíveis tem se destacado o uso de eletroforesede isoenzimas e proteínas. que têm sido incluídosnas regras da Internacional Seed TestingAssociation e no manual de pureza varietal daAssociation of Official Seed Analysts. Para aidentificação de cultivares. via eletroforese. énecessário a seleção de sistemas isoenzimáticosque não sofram mudanças durante o processodeteriorativo de sementes. Sabe-se que assementes de cafeeiro em condições normaisperdem a viabilidade rapidamente. O presentetrabalho foi realizado com os objetivos deverificar a possibilidade de diferenciar cultivaresde cafeeiro por eletroforese de isoenzimas. bemcomo selecionar sistemas isoenzimáticos que nãosofram interferência da deterioração de sementes.para fins de diferenciação de cultivares e detectarvariações de perfis eletroforéticos de isoenzimasem sementes de cafeeiro com diferentes níveis dedeterioração. visando selecionar marcadores paraqualidade fisiológica de sementes.
MATERIAL E MÉTODOS
A presente pesquisa foi desenvolvida noLaboratório de Análise e Biotecnologia desementes da Universidade Federal de Lavras(MG). no período de novembro de 1997 a julho de1998. Foram utilizadas sementes das cultivaresRubi (MG -1192). Catuaí Vermelho (CH 2077-25-(9). Acaiá (Cerrado-MG 1474) e Icatu LCG3282. oriundas de lotes com diferentes estágios dedeterioração. As sementes tiveram seu perfildeterminado pelos testes de 1" contagem degerminação. teste de germinação. emergência emsubstrato areia/terra (2:'). índice de velocidade deemergência (Tabela I). Para a avaliação daatividade enzimática. foi uti Iizado bul k desementes inteiras e bulk de embrião. Para tal.foram retirados o pergaminho de duzentassementes. que posteriormente foram Iio f Iizadas.moídas à frio em presença de PVP. Paralelamente.foram retirados embriões de sementes. que foramliofilizados e moídos à frio em presença de PVP.As proteínas. tanto das sementes inteiras. quantodos embriões. foram extraídas ressuspendendo
100 mg do pó em 200~ll de tampão tris Hei 0.2M.
pH 8.0 4 0.1 % de ~ Me. 0.4% PVP. 0,4% PEG cI rriM de EDTA. Após a ressuspensão. a soluçãofoi mantida por I hora em ge ladei ra eposteriormente foi submetida a centrifugação à14.000 xg por 60 minutos à 4)(', A eletroforesefoi desenvolvida em sistema de géis depoliacrilamida a 7.5% (gel de separação) e 4.5°'0(gel concentrador). O sistema tampão gelcletrodoutilizado foi tris-glicina pH 8.9. Após ae letroforese. os géis foram revelados para ossistemas: álcool desidrogenase (ADH). fosfarase
ácida (ACP). catalase (C Ar). esterase ([ST).glutarnato-oxalacetato-transami nase «(JOT) csorbitol desidrogenase (S DH).
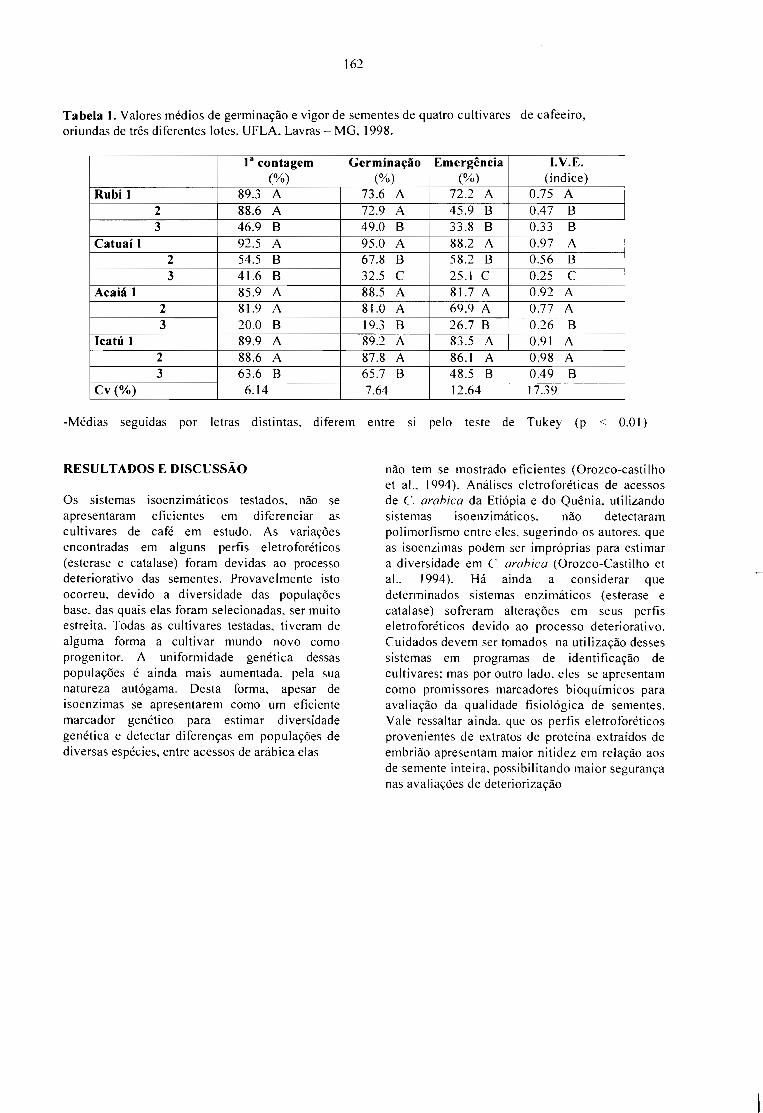
162
Tabela 1. Valores médios de germinação e vigor de sementes de quatro cultivares de cafeeiro,oriundas de três diferentes lotes. UFLA, Lavras - MG, 1998.
I" contagem Germinação Emergência LV.E.(%) (%) (%) (índice)
Rubi 1 89.3 A 73.6 A 72.2 A 0.75 A I2 88.6 A 72.9 A 45.9 8 0.47 83 46.9 B 49.0 8 33.8 8 0.33 B
Catuaí 1 92.5 A 95.0 A 88.2 A 0.97 A2 54.5 B 67.8 8 58.2 8 0.56 83 41.6 B 32.5 C 25.1 C 0.25 C
Acaiá 1 85.9 A 88.5 A 81.7 A 0.92 A2 81.9 A 81.0 A 69.9 A 0.77 A3 20.0 B 19.3 B 26.7 8 0.26 B
Icatú 1 89.9 A 89.2 A 83.5 A 0.91 A2 88.6 A 87.8 A 86.1 A 0.98 A3 63.6 8 65.7 8 48.5 B 0.49 8
._----
Cv(%) 6.14 7.64 12.64 17.39
-Médias seguidas por letras distintas. diferem entre SI pelo teste de Tukey (p < 0.0 I)
RESULTADOS E DISCUSSÃO
Os sistemas isoenzimáticos testados, não seapresentaram eficientes em diferenciar ascultivares de café em estudo. As variaçõesencontradas em alguns perfis eletroforéticos(esterase e catalase) foram devidas ao processodeteriorativo das sementes. Provavelmente istoocorreu, devido a diversidade das populaçõesbase, das quais elas foram selecionadas, ser muitoestreita. Todas as cultivares testadas, tiveram dealguma forma a cultivar mundo novo comoprogenitor. A uniformidade genética dessaspopulações é ainda mais aumentada, pela suanatureza autógama. Desta forma, apesar deisoenzimas se apresentarem como um eficientemarcador genético para estimar diversidadegenética e detectar diferenças em populações dediversas espécies, entre acessos de arábica elas
não tem se mostrado efic ientes (Orozco-casti Ihoet al., 1994). Análises eletroforéticas de acessosde C arabica da Etiópia e do Quênia. utilizandosistemas isoenzimáticos. não detectarampolimorfismo entre eles. sugerindo os autores. queas isoenzimas podem ser impróprias para estimara diversidade em C arabica (Orozco-Castilho etaI.. 1994). Há ainda a considerar quedeterminados sistemas enzimáticos (esterase ecatalase) sofreram alterações em seus perfiseletroforéticos devido ao processo deteriorativo.Cuidados devem ser tomados na uti lização dessessistemas em programas de identificação decultivares: mas por outro lado. eles se apresentamcomo promissores marcadores bioquímicos paraavaliação da qualidade fisiológica de sementes.Vale ressaltar ainda. que os perfis eletroforéticosprovenientes de extratos de proteína extraídos deembrião apresentam maior nitidez em relação aosde semente inteira, possibilitando maior segurançanas aval iações de deteriorização
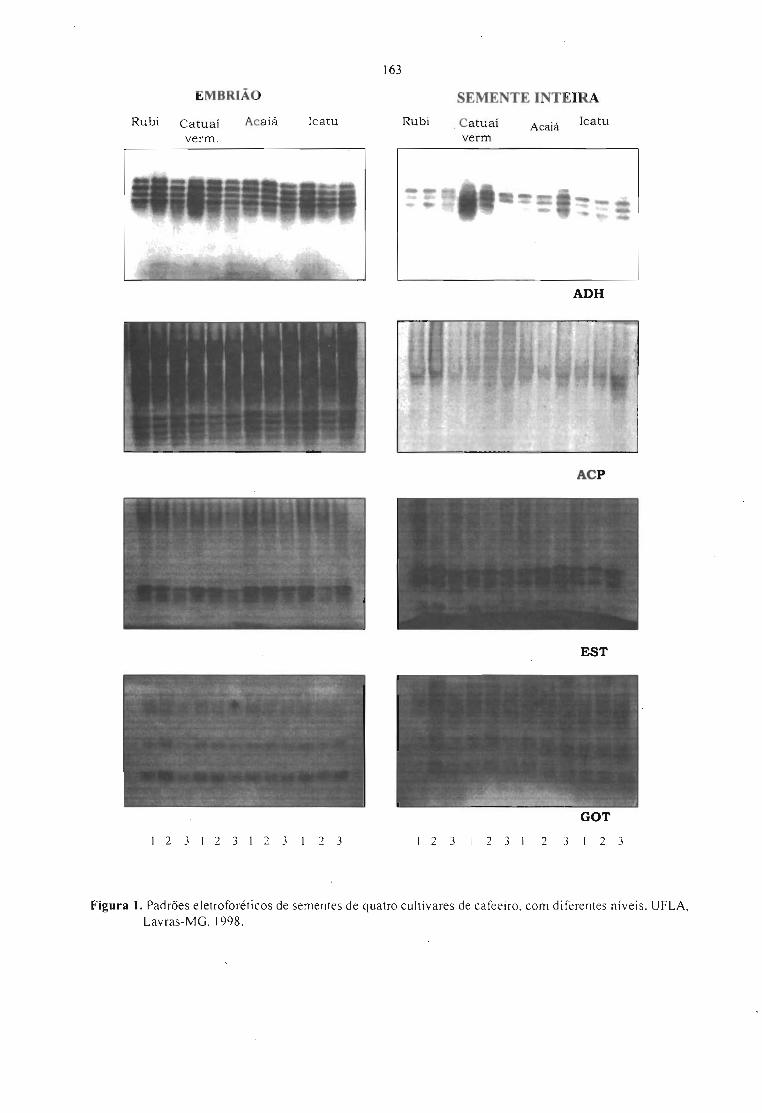
EMBRIÃO
163
SEME TE INTE IRA
Rubi Catuaíverrn .
Aca iá Icatu Rubi , Ca tu aíverm
Aca iá !catu
- ..-...... -.....-.-~ • - ~--:, f ·T-,,
• - .,
~ ... ,l?j ... ~
~."--;: -.,., .. '
~ . r. - " ••'1.. -41"- ,_':':'c. . .,,') 'Jf-.,
F .........
- ..~ ',' .."' .... i..
ADH
ACP
EST
GOT
12 3 123 1 2 3 2 3 1 2 3 23 1 23 1 23
Figura I. Padrões eletroforéticos de semen tes de quatro cultivares de cafeeiro, com di ferentes níveis. UFLA,Lavras -MG, 1998.

CONCLUSÃO
Os sistemas enzimáticos testados não diferenciamas cultivares de café Acaiá, Catuaí, Icatu e Rubi.As enzimas catalase e esterase apresentam-secomo promissores marcadores bioquímicos para aaval iação da qual idade fisiológica de sementes.
REFERÊNCIAS BIBLIOGRÁFICAS
Akkaya, M.S.; Bhagwat. A.A.; Cregan, P.B. 1992Length polyrnorphisms of simple sequencerepeat DNA in soybean Genetics, v.132, p.113 11139.
164
Alfenas, A. C. 1998 Eletroforese de isoenzimas eproteínas afins; fundamentos e aplicações emplantas e microrganismos. Viçosa: UFV, 574p.
Association of Official Seed Analysts. 1991Cultivar purity testing, 371 p.
Lopes, C. R. 1993 Phylogenetic studies of somespecies of the genus Co/Teu 11 - Nurnericalanalysis of sioenzyrnatic data. Revista Brasi leirade Genética. v.16. n.2, p.393-4ü7.

165
VARIABILIDADE GENÉTICA OBTIDA POR RAPD EM ESPÉCIES E HÍBRIDOS DE Coffea
Paulo M. Ruas', Leandro E. C. Dlníz', Claudete F. Ruas' E Tumoru Sera?
I Laboratório de Citogenética e Genética Molecular de Plantas, Departamento de Biologia Geral,Universidade Estadual de Londrina (UEL), 86051-990 Londrina - PR, Brasil.21nstituto Agronômico do Paraná (IAPAR), 86001-970 Londrina - PR, Brasil.e-mail: [email protected]@sercomtel.com.br
INTRODUÇÃO
o gênero Coffea (Rubiaceae) consiste deaproximadamente 80 espécies, as quais sãonativas de regiões tropicais da África. Madagascare ilhas vizinhas (Carvalho et al., 1984). Aprodução internacional do café está concentradoem duas espécies, ColJea arabica e CofJeacanephora. Entretanto, as sementes de cafépodem ser produzidas por muitas outras espéciesde CofJea. Além disso. o pool gênico, útil tantopara as gerações de C. arabica quanto para as deC. canephora, são feitas de todas as espécies deCoffea (Berthaud & Charrier, 1988).As espécies de C. arabica são produzidasprincipalmente na América Latina e representamcerca de 73% da produção mundial de café.CofJea canephora, de outra forma, é produzido naÁfrica Central e do Norte, correspondendo a 80%da produção em todo o continente (OrozcoCasti 110 et al., 1994). Considera-se que C.canephora produz um café com baixa qualidadede bebida, entretanto, esta espécie é umaimportante fonte de genes de resistência paradiversas doença do café. Muitos destes genes temsido transferidos para C. arabica por hibridizaçãoartificial (Smith et al., 1992).
Muitos esforços tem sido direcionados paraexplorar e preservar os recursos genéticos do café,vários métodos tem sido aplicados para descrevera variabilidade presente nas espécies de Coffea.incluindo, características morfológicas eagronômicas. eletroforese da distribuição dealelos variantes entre e dentro de populações, eanálises genéticas das progênies de cruzamentoscontrolados (Charrier & Berthaud, 1985). Padrõesde isozimas foram úteis para estimar a diversidadegenética entre espécies de plantas como arroz(Peng et al., 1988). Chenopodium quinoa(Wi lson. 1988a. 1988b. 1990). Ananas (Dewald etal., 1992: Aradhya et al., 1994) e café (Guedes &Rodrigues, 1974; Payne & Fair-Brothers. 1976;Berthou & Trouslot, 1977; Berthou et al.. 1980).
Entretanto, a análise da eletroforese em acessos deC. arabica oriundas da Etiópia e Quênia. com seissistemas enzimáticos. revelam baixo nível depol imorfismo (Louarn, 1978). Os resultadoscontrastam com o alto nível de variação
morfológica observada no mesmo germoplasmasugerindo que as isozimas não são apropriadaspara descrever a diversidade genética dasvariedades e cultivares de C. arabica.Marcadores moleculares tais como Amplificaçãoao acaso de DNA polimórfico (RAPO) ePolimorfismo no comprimento dos fragmentos derestrição (RFLP) tem sido extensivamente e comsucesso usadas para estudar as relaçõesfilogenéticas em espécies de plantas (Gonzales &Ferrer, 1993; Heun et al.. 1994; Graham &McNicol, 1995). para estudar a diferenciaçãogenética entre subespécies (Zhang et ai.. 1992) epara acessar a diversidade genética inter e intraespécies (Abo-elwafa et al., 1995; Liu, 1996;M'Ribu & Hilu, 1994; Link et al., 1995; Ruas etai.. 1999). A utilidade da técnica de RAPO paraestudar o gênero CofJea foi demonstrado porLashermes et al . (1993), Orozco-Casti 110 et aI.(1994). e Lashermes et al. (1996).O Brasil é um dos principais produtores de cateentre mais de 50 países da América Latina. África eÁsia. O estado do Paraná.. localizado no sul doBrasil. é um importante produtor de café. Neste.estado. o Instituto Agronômico do Paraná (IAPAR)mantém uma coleção de gennoplasma com seteespécies de Coffea. algumas destas espécies temgenes para resistência a seca . geada e doenças.Além disso, a coleção do IAPAR preserva 40variedades botânicas de C. arabica, e diversaspopulações de híbridos interespecíficos. A relaçãofilogenética entre as espécies de Coffea e a origemdos prováveis híbridos mantidos nesta coleção nãoestão claramente definidos. Nesta investigação.procuramos nos ater a determinar as relaçõesfilogenéticas entre as sete espécies e seis híbridosde café utilizando a análise de RAPO.
MATERIAL E MÉTODOS
Material vegetal. As especres e híbridosutilizados neste estudo são parte da coleção degermoplasma do Instituto Agronômico do Paraná(IAPAR). Londrina. Paraná. Brasil. O materialinclui: C. arabica varo arabica. C. canephora varorobusta. C. canephora varo kouillou, C dewevrei.C. eugenioides, C. kapakata. C. racemosa, Cstenophylla e seis híbridos interespecíficos.
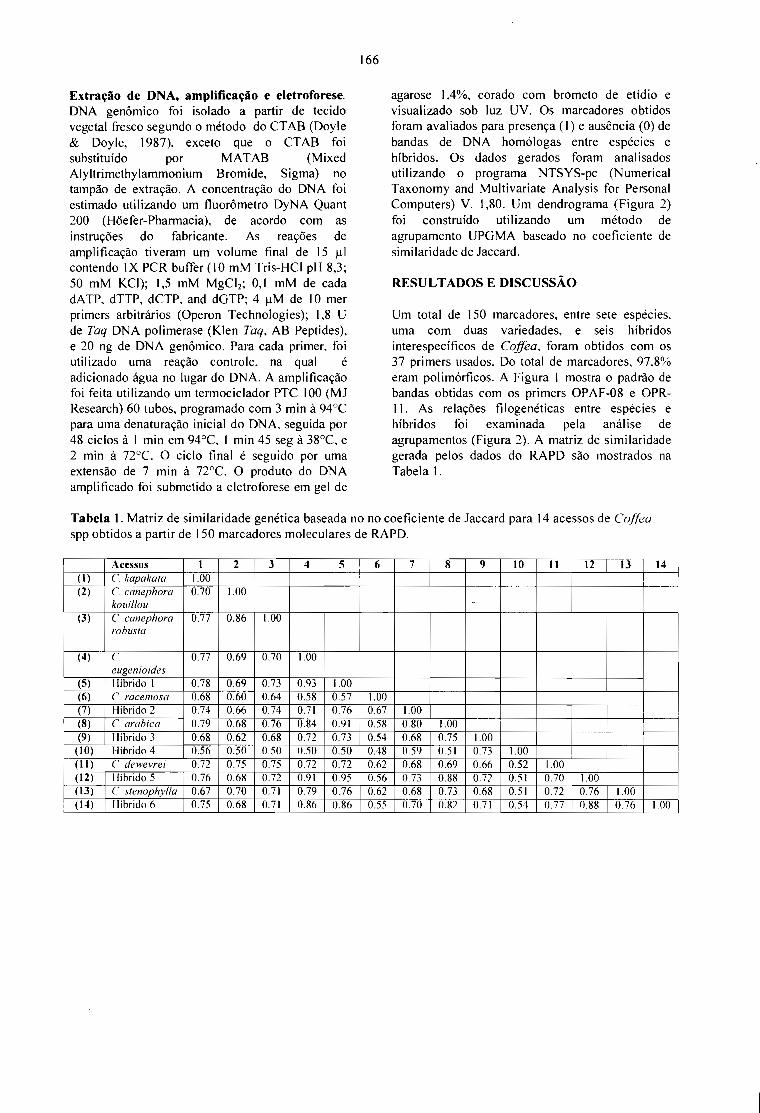
Extração de DNA, amplificação e eletroforese.DNA genômico foi isolado a partir de tecidovegetal fresco segundo o método do CTAS (Doyle& Doyle, 1987), exceto que o CTAS foisubstituído por MATAS (MixedAlyltrimethylammonium Brornide, Sigma) notampão de extração. A concentração do DNA foiestimado utilizando um tluorômetro DyNA Quant200 (Hõefer-Pharmacia), de acordo com asinstruções do fabricante. As reações deamplificação tiveram um volume final de 15 111contendo IX PCR buffer (10 mM Tris-HCI pH 8,3;50 mM KCI); 1,5 mM MgCI2; O, I mM de cadadATP, dTTP, dCTP, and dGTP; 4 11M de 10 merprimers arbitrários (Operon Technologies); 1,8 Ude Taq DNA polimerase (Klen Taq. AS Peptides),e 20 ng de DNA genômico. Para cada prirner, foiutilizado uma reação controle. na qual éadicionado água no lugar do DNA. A amplificaçãofoi feita utilizando um termociclador PTC 100 (MJResearch) 60 tubos. programado com 3 min à 94°Cpara uma denaturação inicial do DNA. seguida por48 ciclos à I min em 94°C, 1 min 45 seg à 38°C, e2 min à n°c. O ciclo final é seguido por umaextensão de 7 min à n°c. O produto do DNAamplificado foi submetido a eletroforese em gel de
166
agarose 1,4%, corado com brometo de etídio evisualizado sob luz UV. Os marcadores obtidosforam aval iados para presença (1) e ausência (O) debandas de DNA homólogas entre espécies ehíbridos. Os dados gerados foram analisadosutilizando o programa NTSYS-pc (NumericalTaxonomy and Multivariate Analysis for PersonalComputers) V. 1,80. Um dendrograma (Figura 2)foi construído utilizando um método deagrupamento UPGMA baseado no coeficiente desimilaridade de Jaccard.
RESU LTADOS E DISCUSSÃO
Um total de 150 marcadores, entre sete espécies,uma com duas variedades, e seis híbridosinterespecíficos de Coffea, foram obtidos com os37 primers usados. Do total de marcadores. 97.8%eram polimórficos. A Figura I mostra o padrão debandas obtidas com os primers OPAF-08 e OPR11. As relações filogenéticas entre espécies ehíbridos foi examinada pela análise deagrupamentos (Figura 2). A matriz de similaridadegerada pelos dados do RAPD são mostrados naTabela 1.
Tabela 1. Matriz de similaridade genética baseada no no coeficiente de Jaccard para 14 acessos de Coffeaspp obtidos a partir de 150 marcadores moleculares de RAPD.
Acessos I 2 3 4 5 6 7 8 9 10 11 12 13 14(I) C kapakata 1.00(2) C canephora 0.70 1.00
kouillou .
(3) C canephora 0.77 0.86 1.00robusta
(4) C 0.77 0.69 0.70 1.00eugenioides
(5) Híbrido I 0.78 0.69 0.73 0.93 1.00(6) C racemosa 0.68 0.60 0.64 0.58 0.57 1.00(7) Híbrido 2 0.74 0.66 0.74 0.71 0.76 0.67 1.00(8) C arabica 0.79 0.68 0.76 0.84 0.91 0.58 0.80 1.00(9) Híbrido 3 0.68 0.62 0.68 0.72 0.73 0.54 0.68 0.75 1.00
(10) Híbrido 4 0.56 0.50 0.50 0.50 0.50 0.48 0.59 0.51 073 1.00---l(11) C dewevrei 0.72 0.75 0.75 0.72 0.72 0.62 0.68 0.69 0.66 0.52 1.00
(12) Híbrido 5 0.76 0.68 0.72 0.91 0.95 0.56 0.73 0.88 0.72 0.51 0.70 1.00(13) C stenophylla 0.67 0.70 0.71 0.79 0.76 0.62 0.68 0.73 0.68 0.51 0.72 0.76 1.00
I (14) Híbrido 6 0.75 0.68 0.71 0.86 0.86 0.55 0.70 0.82 0.71 0.54 0.77 0.88 0.76 100

167
Figura I. Pad rão de bandas obtidas com os prim e iro s O PA F 08 e POR I I em sete espéc ies e seis híbr idosint erespec ificos de Coffe a: (C) reação co ntro le; (I) C. Kapakata; (2) C canephora var , Kouillou, (3)Ccanep hora var. robusta: (4) C eugen ioides ; (5) H íbrido n'' . I; (6) Cracem osa: (7) Híbrido n'' . 2 ; (8)Carahica var. arah ica ; (9) Híbr ido n" . 3; ( 10) Híbrido nO 4 ; (11) C.dewevrei: (12) lI íbrido nO 5;( 13)C. slen ophy lla; ( 14)Hí br ido n". 6 ; (M) Marcado r de peso mo lec ula r pUC-19.
A aná lise dos ag rupa me ntos mostr a um coe ficien tede si mi laridade e nt re es pécies va riando de 0.58 (C.eugenioides e C. racemosa, C. arabica e C.racem osa) a 0 .84 (C. ara bica e C. eugenioidesí;reve lando um a lto nível de var iação ge nética . Osdados do RAPO m ostram que C. arahica tem umassoc iação muito grande com C euge nioides(84%) seg uida por C. kapakata (79 %). Ccan ep hora var. robusta (76%). C stenophylla(73%). C. dewe vre i (69%). C canephora kouillou(68%). e C. racemosa (58%) . Este s resultados sãoco ns isten tes co m Lashermes et aI . ( 1995) eOrozco-Castillo et aI. (1996). os quaisde mon straram q ue C. arabica tem baixa re laçãogenética com C. racem osa e tem mais afini da deco m C. eugenio ides do qu e co m C. caneph ora .Experi me ntos envo lve ndo o c ruza me nto C. arabicavs C. canephora tet rapl óid e prod uziram híbridos FIcom baixo índice de fert ilidad e (Owuor & Van DerVosse n, 1981) . Estimativas da d istância genética
para 3-8 sis temas de enzi ma s (Berthou e t a l. . 198 0 )mostraram que a distân ci a genética entre Ccan ephora e C. eugenioides são co nside ráveis.Estes dad os também são co nfirma dos por Bcrthouet aI. ( 1983) a través de aná lise eletrofor ética defragmentos de ONA mitoco ndríal e c loro plástico .co rtado co m e nz imas de res trição . Na q uelainvestigação. os au tores o bservaram q ue C arabicae C. eugenioides tem uma al ta similari da de e podeter uma origem comum . Dados de cruza mentostambém são concordantes co m os dados de RAPO.Charrier & Berthaud ( 1985) mos traram que C.eugenioides se cru za em gera l mui to melhor comtodos os outros dipl ó ide s do que co m Ccanephora . A es tre ita s im ilaridade ge nética (0. 84 ).reve lado po r marcadores de RAPO entre Carabica e C eugenioides (Tabela I . Figura 2) étambém . co nsiste nte co m trab alhos a nte rio res.sugerindo qu e C cugenioides é um dos a ncestra isde C arabica.

0.55, 0.70,
168
0.85,
r-- -~
-
-1r--
-
FIG.2Raina et aI. (1998) utilizando FISH (Fluorescent in situ hybridization) em espécies de C eugenioides, Ccongensis, C. canephora, C. liberica e C. arabica mostrou que o DNA genômico de C eugenioides hibridizapreferencialmente com os 22 cromossomos de C. arabica. Os outros 22 cromossomos de C. arabicahibridizam com o DNA genômico de C congensis. Entretanto. Lashermes et aI. (1999) aplicando as técnicasde RFLP e GISH sugeriram que C eugenioides e C. canephora são os prováveis progenitores de C arábica,Os resultados sugerem fortemente que C eugenioides é um dos parentais de C. arabica. entretanto. aparticipação de C. canephora não está claramente definida com os dados obtidos por RAPD.
Análise dos híbridos interespecíficos de Co/leu- Os marcadores obtidos por RAPD foramapl icados para deduzir os prováveis parentais deseis híbridos naturais de Coffea. os quaisapresentam características de interesse para osprogramas de melhoramento representando. porexemplo. uma importante fonte de genes pararesistência a muitas doenças de C arabica.
Híbridos n", I e n", 5 agruparam-se com umcoeficiente de similaridade de 0.95 tendo umasimilaridade média de 0.91 e 0.90 com Ceugenioide« e C arabica. respectivamente(Tabela L Figura 2). Híbridos n". 1 e n'', 5 sãotetraplóides altamente férteis e são ambos comoC. arabica no padrão morfológico. Deste modo.como foi sugerido a partir dos caracteresmorfológicos e dos dados obtidos por RAPD. elespodem ser originados por cruzamento entre Ceugenioides e C arabica seguido por sucessivosretrocruzamentos com parentaI arabica.
Híbrido n", 2 tem um coeficiente de similaridadede 0.8 com C. arabica e de 0.67 com C racemosa(Tabela I). Este híbrido é um triplóide estéril quepode ter sido originado a partir de um parenta Idiplóide e outro tetraplóide. O fenótipo do híbrido
n'', 2 é similar àquele de C arabica o qual é umtetraplóide e mostra uma alta associação com ohíbrido n°. 2 através dos marcadores obtidos porRAPD. As folhas do híbrido n'', 2 entretanto. sãoestreitas como as de C. racemosa. Com base nosdados morfológicos e moleculares sugere-se queC. arabicu e C racemosa são os prováveisparentais deste híbrido.
Híbridos n", 3 e n". 4 agruparam-se com umcoeficiente de similaridade de 0.73. O híbrido n".4 associou em 0.51 com C. arabica e em 0.52com C. dewevrei. O híbrido n°. 3. por outro lado.associou em 0.75 com C. arabica e em 0.66 comC. dewevrei. Os híbridos n'", 3 e 4 sãomorfologicamente similares a ambas as espécies.A associação genética detectada com marcadoresmoleculares obtidos com RAPD (Tabela I) entreestes híbridos e o diplóide C dewevrci e otetraplóide C. arabica sugere que as plantashibridas F I (híbrido n", 4) levando gametas comcombinações cromossômicas específicas foramretrocruzadas com o parental recorrente ('.arabica originando o híbrido n'', 3.
Híbrído n", 6 associou muito estreitamente comC eugenioides. C arabica, seguido por C.

dewevrei e C stenophylla (Tabela I, Figura 2). Éconhecido entretanto, que C. stenophylla é oparentaJ feminino do híbrido n°. 6. Ascaracterísticas morfológicas do híbrido n", 6 éintermediária entre as espécies, exceto para foi haslargas e caule roxo. os quais são similares à Cdewevrei. Enquanto os resultados obtidos comRAPD não são conclusivos. o fenótipo das folhase caules sugerem que C dewevrei pode estarenvolvido na origem deste híbrido.
CONCLUSÃO.
Os resultados obtidos neste estudo sugerem que osmarcadores moleculares de RAPD podem seraplicados com sucesso para a análise da relaçãofilogenética entre espécies de Coffea e paradeduzir os prováveis parentais dos híbridosinterespecíficos.
Apoio financeiro: CNPq. CAPES. InstitutoAgronômico do Paraná (IAPAR) e UniversidadeEstadual de Londrina.
BIBLIOGRAFIA
Abo-elwafa, A.. Murai, K. & Shimada, T. (1995).Intra and interspecies variation in Lens revealedby RAPD markers. Theor. Appl. Genet .. 90. 335340Aradhya, M.K.. Zee. F. & Manshardt. R.M.(1994). Isozyme variation in cultivated and wildpineapple. Euphytica. 79, 87-99Berthaud. 1. & Charrier, A. (1988). GeneticResources of Coffea. In- Colfee Agronomy. Vol.4. R.J. Clarke & R. Macrae (eds.). EIsevierApplied Science Publishers Ltd., pp. 1-42Berthou, F. & Trouslot. P. (1977). L'analyse dupolymorphisme enzymatique dans le gemeCoffea: adaptation d'une méthodedélectrophorêse en serie. Premiers résultants ins" International Colloquium on the Chemistry ofCoffee. ASIC. Paris. pp. 373-83Berthou, F.. Mathieu, C. & Vedei. F. (1983).Chloroplast and rnitochondrial DNA variation asindicator of phylogenetic relationship in genusCoffea L. Theor. Appl. Genet., 65. 77-84Berthou, F.. Trouslot. P.. Hamon. S.. Vedei, F. &Ouetier, F. (1980). Analyse en électrophorêse dupolumorphisme biochimique des caféiers:variation enzymatique dans dix-huit populationssauvages: C. canephoru, C eugenioides et C.arabica. Café Cacao tt« 624. 313-326Carvalho. A.. Medina-Filho. H.P. & Fazuoli. L.C(1984). Evolução e melhoramento do cafeeiro. InTópicos de Citogenética e Evolução de Plantas.M.L.R. Aguiar-Perecin, P.S. Martins & G. Bandel
169
(eds.), Sociedade Brasileira de Genética, RibeirãoPreto. S.P., Brazil. pp. 215-235Charrier, A. & Berthaud, l. (1985). Botanicalclassification of coffee. In- Colfee: Botany,Biochemistry and Production of Beans andBeverage. M.M. Clifford & K.C Wilson (eds.).Croom Hei Publ., Beckenham, UK. pp. 13-47Doyle, J.J. & Doyle, l.L. (1987). A Rapid DNAisolation procedure for small quantities of freshleaftissue. Phytochemical Bulletin, 19( 1): 11-15Dewald. M.G., Moore. G.A. & Sherman, W.B.(1992). Isozymes in Ananas (pineapple): geneticsand usefulness in taxonorny, 1. AmeI'. Soe. Hort.Sei.. 117.491-496Gonzales, 1.M. & Ferrer, M. (1983), RandomAmplified Polymorphic DNA analysis In
Hordeum species. Genome. 36, 1029-1031Graham, 1. & McNicol. R.l. (1995), Anexamination of the ability of RAPD markers todetermine the relationships within and betweenRuhus species. Theor Appl. Genet., 90, 11281/32Guedes. M.E.M. & Rodrigues lr, C.J. (1974),Disc electrophoresis patterns of phenoloxidasefrom leaves of Coffee cultivars. Portugaliae AclaBioloica. 13, 169-178Heun. M.. Murphy, l.P. & Phillips, T.D. (1994).A comparison of RAPD and isozyme analysis fordetermining the genetic relationships amongAvena sterilis L. accessions. Theor. Appl. Genet ..87.689-696Lashermes, P.. Cros, 1.. Marrney, P. & Charrier.A. (1993), Use of random amplified DNAmarkers to analyze genetic variability andrelationships of Coffea species. Genetic Resourcesand Crop Evolution. 40. 91-99l.asherrnes, P.. Combes. M.C, Roberts, 1..Trouslot. P., D' Hont, A.. Anthoni, F. & Charrier,A. (1999). Molecular characterization and originof the Coffea arabica L. genome. Molec. Gen.Genel .. 261. 259-266l.ashermes, P.. Trouslot. P., Anthony, F.. Combes.M.C & Charrier, A. (1996). Genetic diversity forRAPD markers between cultivated and wildaccessions of Coffea arabica. Euphytica. 87. 5964Link. W.. Dixkens, C. Singh, M .. Schwall. M. &Melchinger, A.E. (1995). Genetic diversity inEuropean and Mediterranean faba beangermoplasm revealed by RAPD. Theor. Appl.Genet .. 90.27-32Liu, C.J. (1996). Genetic diversity andrelationships among Lablab purpureum genotypesevaluated using RAPD as markers. Euphytica, 90.115-119l.ouam. l. (1978). Diversité cornpararée desdescendances de Coffea arahiea obtenues en

autoféeondation et en féeondation libre auTonkoui. In- Etude de la Structure et de laVariabilité Génétique des Caféiers, A. Charrier(ed.), IFCC Bulletin, n. 14, pp. 75-78Keil, M. & Griffin, A.R. (1994), Use of randomamplified polymorphic DNA (RAPO) markers inthe diserimination and verification of genotypes inEucalyptus. Theor. Appl. Genet.. 89,442-450Michelmore, R.W., Paran, I.E. & Kesseli, R.V.(1991 ), Identifieation of markers linked todisease-resistance genes by bulked segregantanalysis: A rapid method to deteet markers inspeei fie genomie regions by using segregatingpopulations. Proc. Natl. Acad. Sei. USA, 88,9828-9832M'Ribu, H.K. & Hilu, K.W. (1994), Deteetion ofinterspecifie and intraspecifie variation inPanicum mil/ets through random amplifiedpolymorphie DNA. Theor. Appl. Genet.. 88, 412416Orozco-Castillo, C. Chalmers, KJ. & Powell,R.W. (1994), Oeteetion of genetie diversity andselective gene introgression in coffee using RAPOmarkers, Theor. Appl. Genet., 87, 934-940Owuor, J.B.O. & Van Der Vossen. H.A.M.(1981), Interspecific hybridization between CojJeaarábica L. and tetraploid C. canephora P. ex Fr. I.Ferti Iity in F I hybrids and baekcrosses to C.arabica. Euphytica. 30.861-866Payne, R.C & Fair-Brothers. O. E. (1976). Oisceleetrophoresis evidence for heterozygosity111and phenotypic plasticity in selected lines ofCoffea arabica L. Bot. Gazette, 137, 1-6Peng. J.Y., Glaszrnan, J.C & Virrnani, S.S.(1988), Heterosis and isozyme divergence inindica rice. Crops Sei .. 28. 561-563Raina, S.N .. Mukai. Y. & Yamamoto, M. (1998).In situ hybridization identifies the diploid
170
progenitor species of CojJea arabica (Rubiaceae).Theor. Appl. Genet., 97,1204-1209Ram, A.S., Sreenivasan, M.S. & Vishveshwara, S.(1982), Chemotaxonomy of Mozambicoffea. J.CojJee Research (Chikrnagalur), 12.42-6Ruas, P.M., Bonifácio. A., Ruas, CF., Fairbanks,O.J. & Andersen, W.R. (1999), Genetierelationship among 19 accessions of six species ofChenopodium L., by Random AmplifiedPolymorphic DNA fragments (RAPO). Euphytica,105,25-32Ruas, P.M.. Ruas. CF., Fairbanks, DLAndersen, W.R. & Cabral. J.R.S. (1995), Genetierelationship among four varieties of pineapple,Ananas comosus, revealed by random amplifiedpolymorphic ONA (RAPO) analysis. Bras. J.Genet., 18,413-416Srnith. N.J.H .. Wi lliarns, J.R .. Plucknett, O.L. &Talbot, J.P. (1992), Tropical forests and theircrops. Comstock Publishing Associates, íthaca,NY .. pp. 20-37Weising, K.. Nylorn, H.. Wolff, K. & Meyer, W.(1995), ONA fingerprinting in plants and fungi.CRC Press, Boca Raton, Florida., pp. 322Welsh, J. & McClelland. M. (1990),Fingerprinting genomes using PCR with arbitraryprimers. Nucleic Acids Research, 18. 7213-7218Wilson. H.D. (1988a). Quinua biosystematic I:free-living populations, Econ. Botan.. 42. 461-477Wilson, H.O. (1988b). Quinua biosystematic 11:free-living populations. Econ. Botan., 42.478-494Wilson, H.O. (1990), Gene flow in squashspecies. Bioscience, 40,449-455Zhang, Qifa, Saghai Maroof, M.A., Lu. T.Y. &Shen. B.Z. (1992), Genetic diversity anddi fferentiation of indica and japonica rice detectedby R FLP analysis. Theor. Appl. Genet., 83,495499

171
POLIMORFISMO GENÉTICO DETECTADO EM 40 VARIEDADES E CULTIVARES DE Cof/etlarabica UTILIZANDO RAPD ASSOCIADO COM ENZIMAS DE RESTRiÇÃO
Leandro E.C. Diniz", Paulo M. Ruas', Claudete F. Ruas' e Tumoru Sera'
I Departamento de Biologia Geral. Universidade Estadual de Londrina. Londrina. Paraná. Brasil~ Instituto Agronômico do Paraná. Londrina. Paraná. Brasile-mail: [email protected]
ABSTRACTThe genetic variability of40 varieties and cultivars ofCoffea arabica was avaluated using random amplifiedpolymorphic DNA (RAPD) associated with a prior restriction digestion with Eco RI, Haelll, and BamHI. Theacessions include 5 varieties also considered as old c ultivars, lO varieties, 11 recent cultivars, 2 localcultivars, 8 experimental cultivars, 3 mutants, and 1 germoplasm of C arabica, obtained from the Coffeacollection, maintaned at the 1APAR (Instituto Agronômico do Paraná), Londrina. Paraná. Brazil. Thegenetic variability ofthe most important cultivars ofCofJea arabica and the genetic relantioships among alivarieties were determined in a preliminary analysis using 185 differents RAPD primers, no polymorphism OI'
low levei of polimorphic bands were observed. To improve the levei of polymorphism, the DNA of alivarieties were digested with restriction enzymes prior to amplification. Twenty three primers used alone OI'
associated with restriction enzymes, that suggested polymorphism were selected to amplify the DNA of alivarteties and cultivars. A dendogram was constructed using UPGMA. A total ol236 amplification productswere obtained, with 216 polymorphic bands (91.5%). The results showed the usefulness of restrictionenzymes to detect polymorphism in varieties and cultivars ofCoffea.arabica.
1. INTRODUÇÃO
No Brasil, um dos principais estados produtoresde café é o Paraná. Neste estado. o InstitutoAgronômico do Paraná (lAPAR) mantém umbanco de germoplasma com 8 diferentes espéciesde Coffea, sendo algumas destas espécies. fontede resistência à seca. geada e outras doenças queatacam o café. Além destas espécies. a coleção dobanco mantido pelo IAPAR inclui 26 diferentesvariedades e cultivares de C arabica, váriaspopulações de híbridos interespecificos, 144acessos de C arabica da Etiópia. 30 acessos de Ccanephora. 50 progênies de Kattimor, 200progênies de lcatu, 50 progênies de Sarchirnor, 50linhagens de Mundo Novo. 50 linhagens deCatuaí e 20 linhagens de Acaiá.A coleção de Coffea mantida no IAPAR não foiainda caracterizada quanto ao seu potencial. Destemodo. os recursos genéticos do Banco degermoplasma não tem sido amplamenteexplorados. O acesso à variabilidade genética é deextrema importância em qualquer coleção degermoplasma. Usando este conhecimento.genótipos podem ser comparados paradeterminação do grau de similaridade genética
. facilitando o manejo dos recursos genéticosexistentes.
O desenvolvimento da biologia molecular resultouem técnicas alternativas baseadas na anál ise doDNA. Umas das técnicas mais conhecidas é opolimorfismo gerado por fragmentos obtidos comenzimas de restrição (RFLP). esta metodologia.no entanto. é cara elaboriosa. Welsh e McClelland (1990) e Williamset ai. (1990) desenvolveram uma novametodologia baseada na análise de seqüências deDNA amplificadas ao acaso. Esta técnica.conhecida como RAPO (polimorfismo de DNAamplificado ao acaso). faz uso de 10 nucleotídeosde seqüência conhecida e tem sido intensamenteutilizado para detectar polimorfismo em plantas.A informação obtida para o germoplasma de Carabica a partir de marcadores moleculares podefacilitar o acesso e a utilização dos recursosgenéticos mantidos na coleção. aumentando.portanto. a eficiência na caracterização genéticada coleção.
2. MATERIAL E MÉTODOS
O material utilizado neste trabalho faz parte dacoleção de Coffea do Banco de germoplasma doInstituto Agronômico do Paraná (IAPAR). comodescrito na Tabela I.

172
Acessos Designação OrigemAcaiá cultivar atual BrasilArábica variedade e cultivar antigo Brasil/ EtiópiaArábica superprec:JCe germoplasma de Coffea arabica EtiópiaBourbon amarelo variedade e cultivar antigo BrasilBourbon vermelho variedade e cuItivar antigo Ilhas ReuniãoCatuaí amarelo cultivar atual Brasil
I
Catuaí vermelho cultivar atual BrasilCatuaí amarelo erecta mutantc de Catuaí amarelo BrasilCatuaí rubi vermelho cultivar atual BrasilCatuaíamarelo scrnpcrflorens mutante de Catuai amarelo BrasilCatuaí SH,SII, cultivar experimental BrasilCaturra amarelo variedade e cultivar antigo BrasilCaturra vcrnclho variedade e cultivar antigo BrasilCera Variedade BrasilCioicie cultivar local Etiópia
IColômbia amarelo cultivar atual ColômbiaGeisha cultivar local EtiópiaGoiaba Variedade Hrasi IIapar 59 (75.163-22) cultivar atual Brasillapar 75.163-12 cultivar experimental Brasil I
lapar 75.163-21-10 cultivar experimental Brasil II
lapar 77.028 amarelo cultivar experimental Brasillapar 88.480 tupi cultivar atual Brasillcatu amarelo 2944 cuIti\ar atual Brasillcatu precoce 3282 cultivar atual Brasillcatu vermelho 2945 cultivar atual Brasillcatuaí lapar 77.055 cultivar experimental BrasilKattirnor cultivar experimental Brasil/PortugalLaurina Variedade BrasilMaragogipe Variedade HrasiIMokka Variedade BrasilMundo Novo 376-4 cultivar atual BrasilMundo Novo semperllorens mutante de Mundo Novo
IBrasil
Mundo Novo X lapar 59 (F,) cultivar experimental BrasilMundo Novo X lapar 59 (F,) cultivar experimental BrasilPurpurascens Variedade BrasilSan Bernardo Variedade Costa RicaSemi-crecta Variedade BrasilVilla Lobos Variedade Costa RicaVilla Sarchi Variedade Costa Rica
Tabela I Designação e países de origem dos acessos de Cotfea arabica estudados:
2.1. ExtraçãoPara a extração de DNA. foi utilizado a técnicadescrita por Doyle & Doyle ( 1987). com algumasmodificações. O DNA purificado foi quantificado
e diluído à uma concentração final de 10 ng/ul.Foi utilizada a metodologia da "Análise desegregação de bulks" (Michelrnore et al .. 1991).
2.2. Amplificação de DNAAs reações de amplificação para o RAPD tiveram 15
~t1 de reagentes. sendo 7.4 ~t1 de água (HPLC); 1.5 ~t1
de tampão 10X (50 mM Tris-HCI pH 9.1. 16 mM de
sulfato de amônia. 35 mM MgCI" e 150 ug/rnl. BSA
(albumina de soro bovinoj): 1.5 ~t1 de dNTPs (0.1
mM de cada: dA TP. dTTP. dCTP e dGTP); 2.0 ul
de primer (004 ~LM); 2.0 fll de DNA (I O ng/ul) e
0.07 fll (0.7 unidade) de Taq polirnerase da marca
Klen Taq I. As amostras diluídas (I O ng/ul) deDNA genôrnico de cada variedade ou cultivar foramdigeridas com algumas enzimas de restriçãodiferentes. tais como: EcoRI. HaeIlI ou BamHI.
durante 1 hora à 37°C. seguida da reação deamplificação. As amostras foram amplificadas emum ciclador térmico PTC 100 para 60 tubos da MJResearch. O programa utilizado segue o descrito porRuas et a/( 1999). O produto de amplificação obtidofoi submetido a eletroforese em gel de agarose1.4%. corada com brometo de etídio e fotografadosob luz ultravioleta. A análise dos resultados foi feitacom o auxílio do programa NTSYS (""NumericalTaxonomy and Multivariate Analysis") utilizando o

método SIMQUAL ("Similarity for qualitativedata") com o coeficiente de similaridade de Jaccard.
3. RESULTADOS E DISCUSSÃO
A técnica de RAPO combinada com a digestão doONA a ser amplificado utilizando enzimas derestrição (BamHI. EcoRI e HaeIlI) permitiu adetecção de polimorfismo em 40 acessos de C.arabica. Um total de 23 prirners, associados ounão com enzimas de restrição foram usados paraamplificação do ONA de 15 variedades. das quais.cinco também são consideradas cultivares antigos.11 cultivares atuais. oito cultivares experimentais,dois cultivares locais. três mutantes e umgermoplasma de C. arabica introduzidorecentemente na coleção. Um total de 236marcadores. com pesos moleculares variandoentre 300 a 2.200 pb, foram obtidos. Na maioriadas reações submetidas a digestão enzimáticaantes do início do programa de amplificação. nãofoi observado aumento significativo no número debandas em relação às reações amplificadas semdigestão prévia. Entretanto. um padrão diferentede produtos de amplificação foi obtido.melhorando deste modo a eficiência dos primers.O dendograma gerado a partir dos dados obtidosassociou os acessos em quatro grupos distintos(Figura I).No primeiro agrupamento observaram-se trêssubgrupos sendo que o primeiro destes associa asvariedades arábica. bourbon vermelho e bourbonamarelo. As variedades arábica e bourbonvermelho mostraram uma similaridade de 91%.enquanto Bourbon amarelo apresentou 84% desimi laridade com as duas outras variedades.Bourbon vermelho é uma mutação espontâneaocorrida em cultivares da variedade arábica.introduzidos na Ilha Reunião. a partir decultivares da Etiópia. Bourbon amarelo também éum mutante encontrado em progêniesprovenientes do cruzamento Arábica X Bourbonvermelho. No segundo subgrupo o cultivar Acaiáassocia-se ao mutante Mundo Novo semperflorense com o cultivar Mundo Novo 376-4 comcoeficientes de similaridade de 84% e 82%.respectivamente. Mundo Novo originou-se docruzamento natural entre a variedade Sumatra XBourbon vermelho. O cultivar Acaià originou-sede seleção em plantas do cultivar Mundo Novo(Fazuoli, 1986). Mundo Novo semperflorens éuma mutação espontânea ocorrida em MundoNovo. No terceiro subgrupo. as cultivares Catuaíamarelo. Catuaí vermelho e o mutante Catuaíamarelo semperflorens associam-se com urnasimilaridade média de 89%. Ambos. Catuaíamarelo e Catuaí vermelho surgiram por
173
hibridação artificial entre Caturra amarelo XMundo Novo (Fazuo li, 1986). Catuaí amarelosemperflorens é um rnutante de Catuaí amarelo.No segundo agrupamento. aparecem trêssubgrupos. sendo que no primeiro o cultivarIcatuaí lapar 77.055 e a variedade Caturra amarelose associam com 82% de similaridade. A cultivarCatuaí rubi vermelho agrupa-se com Icatuaí lapar77.055 e com Caturra vermelho com 80% e 77%.respectivamente. A cultivar Catuaí rubi vermelhooriginou-se do cruzamento entre Catuaí vermelhoe Mundo Novo. No segundo subgrupo. Icatuprecoce 3282 e Caturra vermelho associam-secom 88% de similaridade. Icatu precoce 3282 foiobtido a partir do cruzamento entre C. canephorae C. arabica var. bourbon vermelho sendo o FIretrocruzado com Mundo Novo. O terceirosubgrupo associou os cultivares lcatu amarelo eIcatu vermelho com similaridade genética de88%. A cultivar Icatu amarelo foi obtido porseleção de plantas resultantes de hibridaçãonatural entre Icatu vermelho X Bourbon amareloou Mundo Novo amarelo (Fazuoli. 1986). Osresultados obtidos com RAPO sugerem queMundo Novo amarelo e Icatu vermelho são osprováveis parentais de Icatu amarelo. Ainda nomesmo subgrupo as variedades Mokka e Laurinaapresentam uma similaridade genética de 86%. Deacordo com Wrigley (1988). Laurina éproveniente da variedade Bourbon e Mokkaapresenta uma relação estreita com Laurina comomostram os resultados obtidos com RAPO.No terceiro agrupamento observam-se doissubgrupos. No primeiro. a variedade Villa Lobosmostra uma similaridade genética de 92% com asvariedades San Bernardo e Villa Sarchi. As trêsvariedades originaram-se de Caturra vermelho. naCosta Rica. Esta relação é confirmada pela estreitaafinidade genética encontrada com os marcadoresmoleculares. No segundo subgrupo encontram-secinco cultivares (Colômbia amarelo, lapar 77028amarelo. lapar 59 (75.163.22). lapar 88.480 tupi elapar 75.163.21-10). Uma similaridade genéticade 90% foi encontrada entre os cultivares lapar88.480 tupi e lapar 75.163.21-10. enquanto lapar59 mostrou-se 89% e 88% similar aos cultivareslapar 75.163.2 J-I O e lapar 88.480 tupi.respectivamente. Os altos índices de similaridadegenética derivam do fato destas cultivares teremuma origem comum, a partir de cruzamentos entreos híbridos de Timor X Caturra ou Villa Sarchi. Acultivar Villa Sarchi mostra uma relação genéticade 85% com as cultivares lapar e pode. portanto.estar envolvida nos cruzamentos que originaramtais cultivares. O quarto grupo. mostra umaassociação entre 11 acessos. divididos em quatrosubgrupos. No primeiro subgrupo. o cultivar lapar

174
75.163-12 associa-se com o híbrido (FI) originadodo cruzamento entre Mundo Novo X lapar 59 ecom o cultivar Kattimor com 86% e 83% desimilaridade, respectivamente. O cultivarKattimor é um germoplasma de Sarchirnor, cujaorigem se deu através através do cruzamento entreVilla Sarchi e o híbrido de Timor. Kattimorassociou-se a Villa Sarchi com 82% desimilaridade. No segundo subgrupo, C. arabicavar. semi-erecta. agrupou-se com o mutanteCatuaí amarelo erecta em 91%. No terceirosubgrupo, as variedades Goiaba, Maragogipe,Purpurascens e Cioicie agruparam-se com 91% desimilaridade genética, enquanto Goiaba ePurpurascens associaram com 90% desimilaridade. a Origem de Maragogipe, Goiaba ePurpurascens se deu a partir de mutaçõesdiferentes ocorridas na variedade Bourbon,
explicando o alto índice de similaridadeobservado entre estas variedades. Cera e Geishaassociam-se com 87% de similaridade.O F2 originado do cruzamento Mundo Novo Xlapar 59, aparece isolado, mostrando uma relaçãode 80% de similaridade com as cultivares lapar59. Iapar 88.480 tupi e lapar 75.163-21-10. Acultivar Arábica superprecoce apresentou baixocoeficiente de similaridade com as outrasamostras, provavelmente devido às muitasdiferenças fenotípicas observada entre estacultivar e as demais variedades e cultivares.Finalmente. o cultivar Catuaí SH,SH, aparececomo um "outgroup", mostrando umasimilaridade média de 60% com os demaisacessos. Isto pode ser facilmente explicado. umavez que Catuaí SH,SH, possui germoplasma deColfea liherica.
Figura 2. Dcndograma de similaridade para as 40 variedades c cultivares de Coffea arabica: 1- arábica. 2-hourhonamarelo: 3-hourhon vermelho: 4-acaia: 6-mundo novo 376-4: 7-mundo novo scrnperflorens: 8-catuai scmperflorcns: 9catuai amarelo: lO-catuai vermelho: I l-catuai ruhi vermelho: 12-icatuai lapar 77055: l I-catuai SIII SI l.: 14-caturravermelho: 15-caturra amarelo: 16-arahica superprccoce: 17-icatu precoce 3282: 18-icatu amarelo 2944: 19-icatuvermelho 2945: 20-mokka: 21-laurina: 23-villa lohos: 24-san bernardo: 25-villa sarchi: 26-colomhia amarelo: 27-lapar77028 amarelo: 28-lapar 59: 29-lapar 88480 tupi: 30-lapar 75163-21-10: 32-lapar 75163- 12: 33-kattim OI': 35-mundonovo X lapar 59 (FJ!: 36-mundo novo X lapar 59 (F,): 38-semierecta: 39-catuai amarelo crecta: 43-goiaha: 44maragogipe: 45-cera: 46-geisha: 47-cioicie; 48-purpurascens.
TreeG0.82 1.00
13
~-------2
~yc'---------;L----------6
.--------8~-----'3
~-----10
,------------ 11~,---------- 12
L- 1 ,:,
~------14
L-------17~-----18
L{ ~6L------21
~---23
~---24L- 25
4=c1.-------- 26.--------- 27
.--------28.------2'3L-----3ü
32,r-1L----- 3533
~----38
"--"----~ as.-------43,--------44L- 48
"------47L-.i,------- 4':,
L------46
'-
0.650.470.30
L- 36L- 16
L- I'::

4.REFERÊNCIAS BIBLIOGRÁFICAS
DOYLE. J.1. & DOYLE. u.. 1987. A Rapid DNAisolation procedure for small quantities of fresh leaftissue. Phytochemical Bulletin, Vol. 19(1): 11-15.FAZUOLL L.c. 1986. Genética e Melhoramento docafeeiro. Cultura do Cafeeiro. Fatores que afetam aprodutividade. Associação Brasileira para Pesquisa DaPotassa e do Fosfato. Piracicaba-SP.MICHELMORE. R.W.: PARAN. I. & KESSELI. R.V.1991. Identification of markers linked to diseaseresistance genes by bulked segregant analysis: A rapidmethod to detect markers in specific genomic regionshy using segregating popu lations. Proe. Natl. Acad.Sei. USA 88: 9828-9832.OROZCO-CASTILHO. c. CHALMERS. K. l &POWELL. R.W. (1994). Detection 01' gcnetic diversityand selective gene introgression in coffee using RAPDmarkers. Theor. Appl. Genet. 87: 934-940.
175
RUAS. P.M.: BONIFACIO. A.: RUAS. C.F.:FAIRBANKS. D.1. & ANDERSEN. W.R. 1999.Genetic relationship among 19 accessions of sixspecies of Chenopodium L.. by Random AmplifiedPolymorphic DNA fragments (RAPD). Euphytica 105:25-32.TAUNA Y. A.E. 1939. História do café no Brasil. Riode Janeiro. Departamento Nacional do Café. 452p.WELSH. r. & McCLELLAND. M. 1990.Fingerprinting genomes using PCR with arbitraryprimers. Nucleie Aeids Res 18: 7213-7218.WELSH. 1.: PETERSEN. C. & McCLELLAND. M.1991. Polymorphisms generated hy arhitrarily primedPCR in the mouse: Application to strain identificationand genetic mapping. Nucleie Acids Research 20: 303306.WI LLlAMS. J.G.K.: JUBELlK. A.R.: LIVAK. K.1. &RAFALSKL lA. 1990. DNA polymorphismsamplified by arbitrary primers are useful as geneticmarkcrs. Nucleie Acids Research 18: 6531-6535.WRIGLEY. G. 1988. Coffee. Tropical AgricultureSeries. Longman Scientific & Technical. New York.USA.


177
CARACTERIZAÇÃO DE ACESSOS DE Coffea POR MARCADORES RAPD
Terezinha A. Teixeira l,2, Ney S. Sakiyama l,2, Laércio Zambolim l,3, Antônio A. Pereira'l,Cássia C. H. Sakiyamal
lBl0AGRO/UFV; 20FT/UFV; 30FP/UFV; 4EPAMIG. 36571-000 Viçosa.MG.Apoio Financeiro: FAPEMIG. FINEP. CNPq. PNP&OCafé
INTRODUÇÃO
o subgênero CofJea compreende cerca de 100espécies distribuídas em diferentes seções. Dasespécies cultivadas CofJea arabica (café arábica)e Coffea canephora (café robusta) são as de maiorimportância econômica, embora, outras espéciestenham uma valíosa reserva de genes paradiferentes propostas do melhoramento genético(LASHERMES et al., 1993).Pesquisas na área do melhoramento genético docafeeiro no Brasil resultaram na obtenção decultivares com potencial produtivo quase 240%superior às cultivares tradicionais. Entretanto.evidências históricas indicam que as populaçõesbásicas para as seleções das cultivares atuaisoriginaram de poucas plantas introduzidas naAmérica Latina vindas do Yemen. A ampliaçãoda base genética tornou-se importante para omelhoramento genético desta cultura.Informações sobre distâncias genéticas entreacessos de um banco de germoplasma baseadas
Tabela 1 - Lista dos 38 genótipos de cafeeiro utilizados.
em marcadores moleculares podem ser úteis naidenti ficação da diversidade genética.Os objetivos deste trabalho foram: a) avaliação dadiversidade genética entre 38 genótipos de cafédo banco de germoplasma da UniversidadeFederal de Viçosa; b) caracterização molecular de16 linhagens comerciais de café arábica usando atécnica de amplificação ao acaso de ONApolimórfico ("'RAPO") (WILLlAMS et al., 1990)
MATERIAL E MÉTODOS
Material genéticoNa Tabela I estão listados os 38 genótipos dobanco de gerrnoplasma avaliados quanto àdiversidade genética com base em marcadoresRAPD.Na Tabela 2 encontram-se listados as 16linhagens comerciais de café arábica utilizadaspara caracterização molecular por marcadoresRAPO.
GENOTIPOS GENOTIPOS01. CofJea racemosa (UFV-545-57) 20. Mundo Novo (UFV-2191)02. Coffea congensis (UFV-3097) 21. Bourbon Amarelo de Portugal (UFV-535)03. CofJea canephora (UFV-3755) 22. Bourbon04. Triplóide 23. Bourbon da China (progênie 5288 " I)05. Híbrido de Timor (UFV-4106) 24. Bourbon da China (progênie 5288112)06. Híbrido de Timor (UFV-376-52) 25. Caturra (UFV-534-4)07. Típica da China 26. Catimor (UFV-395-141)08. Típica de Portugal (UFV-536)
--27. Catimor (UFV-386-19) I
09. Catuaí (UFV-2237) 28. Catirnor (U FV-1310)I
10. Catuaí (UFV-2144) 29. Catimor (UFV-1359)I I. Catuaí (U FV-2147) 30. Catirnor (UFV-2983)12. Catuaí (UFV-2145) 31. Mundo Novo x Híbrido de Tirnor (H-484-2)13. Catuaí (UFV-2154) 32. Catuaí x Híbrido de Tirnor (H-415-3)14. Catuaí (UFV-2155) 33. Mundindu (UFV-315-76)
,--
15. Catuaí (UFV-2156) 34. Cachirnor(UFV-351-13)16. Mundo Novo (UFV-2190) 35. Cavimor (UFV-357-4)17. Mundo Novo (UFV-2164) 36. San Ramon (UFV-3094)18. Mundo Novo (UFV-2150) 37. Airi (UFV-3095)~. Mundo Novo (UFV-2163) 38. Purpuracens (UFV-4072)

178
Tabela 2 - Lista das 16 linhagens comerciais de cafeeiro utilizadas.
GENOTIPOS GENOTIPOSOI. Catuaí (UFV-2237) 09. Mundo Novo (UFV-2190)
02. Catuaí (UFV-2144) 10. Mundo Novo (UFV-2164)
03. Catuaí (UFV-2147) l l . Mundo Novo (UFV-2150)
04. Catuaí (UFV-2145) 12. Mundo Novo (UFV-2163)
05. Catuaí (UFV-2154) 13. Mundo Novo (UFV-2191)
06. Catuaí (UFV-2155) 14. Mundo Novo 12
07. Catuaí (UFV-2156) 15. Topázio (MG 1190)
08. Catuaí (UFV-2143) 16. Acaiá Cerrado (UFV-1150)
Extração de DNAO DNA foi extraído de folhas jovens seguindo oprotocolo modificado de Doyle & Doyle (1990)com a adição de PVP-40 (solúvel) no tampão deextração. Após a extração o DNA foi quantificadoem espectrofotôrnetro, corrido em gel de agarose0,8 % e armazenado a 4°C. Na época daamplificação foi diluido em TE (Tris HCl 10 mM,EDTA I mM, pH 8,0) para uma concentraçãofinal de 10 ng/ul.
Amplificação de DNA e análise eletroforéticados produtosVinte e oito primers de 10 bases da "OperouTechnologies' foram utilizados para amplificar osDNAs dos 38 genótipos. Dezesseis primersforam utilizados para avaliar as 16 linhagenscomerciais de café arábica. A amplificação foi
feita em termociclador Perkin-elmer 9600 e cadareação contou com um volume total de 25 111.contendo os seguintes componentes: 25 ng deDNA genômico, I unidade AmpliTaq DNApolirnerase, 0,1 mM de cada dNTP, 0,2 11M de"prirner", 50 mM de KCI. 10 mM de Tris HCI e 2
mM de MgCI2 e o volume final completado comágua ultrapura. Foi utilizado o seguinte programa:um ciclo para desnaturação (95°C por 1 min),39ciclos para amplificação (15 seg a 94°C, 30 seg a35°C. 60 seg a n°C) e para finalizar 7 min an°c. Os produtos das reações de amplificaçãoforam separados em geis de agarose 1.4%,corados com brometo de etídio, visualizados emUV e fotografados. RAPDs foram registradoscomo presença ou ausência de bandas. Somenteforam consideradas as bandas polimórficas maisnítidas.
Análise dos dadosFoi montada uma matriz de acordo com apresença (I) e ausência de bandas (O). Esta matrizfoi usada para calcu lar as porcentagens dedesacordo. O dendrograma foi criado pela análisede clusters do Software ST ATlST1CA usando ométodo UPGMA ("unweighted pair-groupmethod using arithmetic averages") baseado nasporcentagens de desacordo.
A Figura I representa o padrão obtido com o primer OPC-09 para 37 genótipos e é apresentado comoilustração.
••:=-",~; -">. __'.,.•,-'._ _ ., -:-:: --o +?' ~ ,-i* ,J:'::.: :,.:, r : " ,~'
--··~--··•• t ••~~•••~-~---·~.·r·tl·•• ---~"..~·~'! ..··!~·..•..!····..···!~tJt·tt ..•••••-:•••••:;;;;;;.;;;;;;;.;;;11;1111;••.•..- ..........•..........•....•- .• •
Figura I . Padrão de amplificação de DNA obtido com o "prirner" OPC-09 para 37 genótipos de cafeeiro. Aseqüência partindo da esquerda é a mesma da Tabela I.
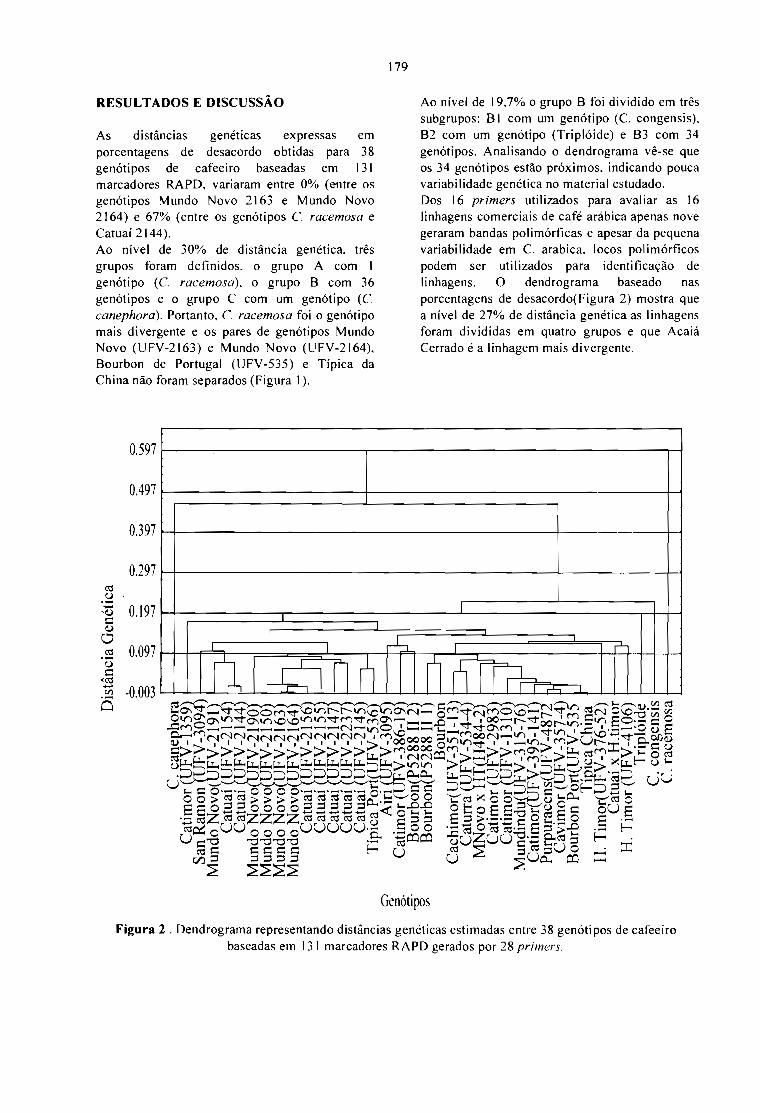
RESULTADOS E DISCUSSÃO
As distâncias genéticas expressas emporcentagens de desacordo obtidas para 38genótipos de cafeeiro baseadas em 131marcadores RAPD, variaram entre 0% (entre osgenótipos Mundo Novo 2163 e Mundo Novo2164) e 67% (entre os genótipos C. racemosa eCatuaí 2144).Ao nível de 30% de distância genética. trêsgrupos foram definidos. o grupo A com 1genótipo (C racemosa), o grupo B com 36genótipos e o grupo C com um genótipo (Ccanephora). Portanto. C racemosa foi o genótipomais divergente e os pares de genótipos MundoNovo (UFV-2163) e Mundo Novo (UFV-2164),Bourbon de Portugal (UFV-535) e Típica daChina não foram separados (Figura 1).
179
Ao nível de 19.7% o grupo B foi dividido em trêssubgrupos: B 1 com um genótipo (C. congensis),B2 com um genótipo (Triplóide) e B3 com 34genótipos. Analisando o dendrograma vê-se queos 34 genótipos estão próximos. indicando poucavariabilidade genética no material estudado.Dos 16 primers utilizados para avaliar as 16linhagens comerciais de café arábica apenas novegeraram bandas polimórficas e apesar da pequenavariabi lidade em C. arabica, locos pol imórficospodem ser utilizados para identificação delinhagens. O dendrograma baseado nasporcentagens de desacordo( Figura 2) mostra quea nível de 27% de distância genética as linhagensforam dividídas em quatro grupos e que AcaiáCerrado é a linhagem mais divergente.
0.597
0.497
0.397
0.297~
.~..... 0.197'Q)c::Q)
O.~ 0.097uc::(~.....
-0.003.~
a
I
I :-S.J. I,.......
Il Qn nn I i-:::L.I I ,...I-,
Genótipos
Figura 2. Dendrograma representando distâncias genéticas estimadas entre 38 genótipos de cafeeirobaseadas em 131 marcadores RAPD gerados por 28 primers.
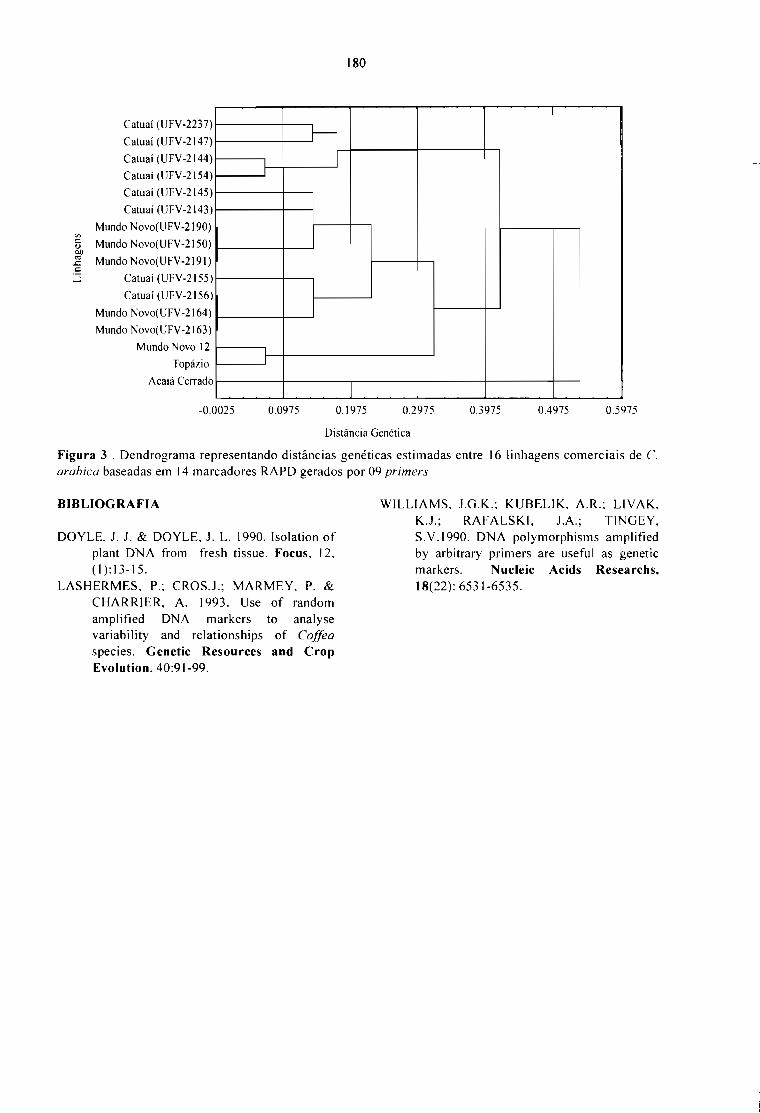
180
I
)
Catuaí (UFV-2237)
Catuaí (UFV-2147)
Catuaí (UFV-2144)
Catuaí (UFV-2154)
Catuaí (UFV-2145)
Catuaí (UFV-2143)
MundoNovo(UFV-2190)V>
i3 MundoNovo(UFV-2150)~
.J::. MundoNovo(UFV-2191)c:
Catuaí (UFV-2155
Catuaí (UFV-2156)
Mundo Novo(UFV-2164)
Mundo Novo(UFV-2163)
Mundo Novo 12
Topázio
Acaiá Cerrado
-0.0025 0.0975 0.1975 0.2975 0.3975 0.4975 0.5975
Distância Genética
Figura 3 . Dendrograma representando distâncias genéticas estimadas entre 16 linhagens comerciais de Carabica baseadas em 14 marcadores RAPD gerados por 09 primers
BIBLIOGRAFIA
DOYLE. J. J. & DOYLE. J. L. 1990. Isolation ofplant DNA from fresh tissue. Focus, 12.(1):13-15.
LASHERMES. P.; CROS.J.; MARMEY. P. &CHARRIER. A. 1993. Use of randomamplified DNA markers to analysevariabi Iity and relationships of Coffeaspecies. Genetic Resources and CropEvolution.40:91-99.
WILLlAMS. J.G.K.; KUBELlK. A.R.; LlVAK.K.J.; RAFALSKI, J.A.; TINGEY.S.V.1990. DNA polymorphisms amplifiedby arbitrary primers are usefu I as geneticmarkers. Nucleic Acids Researchs,18(22): 6531-6535.

181
CARACTERIZACIÓN MOLECULAR DE GENOTlPOS DE CAFÉ.
José W. Bustamante p l y Delia Polanco L.2
I Laboratorio de Biotecnología FONAIAP (Centro de Investigaciones Agropecuarias dei Estado Táchira)Email:[email protected] Apartado 5030. Rubio, Táchira, Venezuela
21nstituto de Agronomia, Facultad de Agronomia, Universidad Central de VenezuelaEmail: [email protected] Apartado 4579. Maracay 2110. Venezuela.
RESUMENLa caracterizaciôn electroforética de genotipos se realiza hoy en forma rutinaria en los hancos de germoplasmupara identificar con precision cada material El polimorfismo electroforético de isoenzimas en geles horizontalesde almidón (SGE: 12%) fue evaluado en veinte cultivares de café (Coffea arabica L. ) pertenecientes ai Bancode Germoplasma dei Centro de Investigaciones Agropecuarias Táchira (FONA1AP-Táchira. Venezuela). Elnúmero de bandas y su distribucion en el gel (movilidad relativa) fue similar en los sistemas enzimáticos paracada uno de los individuas por cultivar y en todos los cultivares. Un individuo perteneciente ai cultivar Bluemountain fue el único que presentô polimorfismo, mostrando una banda adicional en la enzima ACO(frecuencia dei fenotipo electroforético: l/la individuas analizados). Se sehala además una posibleinterpretación genética de fenotipos electroforéticos de los diferentes sistemas enzimáticos evaluados, asi comouna posible teoría sobre "HETEROC/GOS/S FIJADA" en la especie Coffea arabica
INTRODUCCIÓN
EI café (Coffea arabica L.) es un rubro de granimportancia a nivel mundial y es cultivado. no porsu valor alimenticio sino por el alto contenido desustancias aromáticas y estimulantes en el grano.Las especies que comercialmente se siembran anivel mundial son Coffea arabica L. y CofJeacanephora Pierre ex Froehn., Ias cualesrepresentan alrededor de 70% y 30% de laproducción respectivamente. A nivel nacional yen Latinoamérica. Ia especie que se cultiva enmayor escala es Coffea arahica. Ésta secaracteriza por general' una bebida de aroma ysabor suave. mientras que C. canephora ai igualque las demás especies dei género producen unabebida de amargo sabor.EI origen de C. arahica no está aún muy definido;sin embargo. C. arabica y C. eugenioidespresentan idéntico genoma mitocondrial.Presumiblemente ésta última. o un parientecercano, participó como hembra en el posiblecruce que dio origen a C. arahica (Berthou et al.,1983). Por otro lado. el análisis isoenzimáticomostró similitud entre patrones electroforéticos deC. congensis y C. canephora con la especie C.arabica. Es probable que una de estas especiesfuera el macho en el posible cruce (Berthou et al.,1980).La variabilidad conservada en los bancos degermoplasma es necesario caracterizaria eidentificaria y esto se ha realizado mediante el usode características morfológicas. sin embargo. éstaspresentan una serie de dificultades y desventajas:altamente influenciadas por el ambiente.
expresión tardía en etapas feno lógicas. como porejemplo: características de flor. fruto. etc.(especialmente en aquelIas especies de vidajuvenil larga (café. cacao, frutales). Actualmente.existen técnicas mas precisas que complementanla caraterización de genotipos; entre éstas sepueden citar: electroforesis de isoenzirnas, deproteínas y de ADN (RAPD. RFLP). Lasproteínas e isoenzirnas, representan la expresióndirecta de los genes en un ambiente y edad de laplanta determinada. mientras que la electroforesisde ADN mide la variabi lidad directamente en lacadena de ADN.EI banco de germoplasma de café dei CampoExperimental Bramón (FONAIAP- Táchira), es labase de los programas de mejoramiento en estecultivo en el país y cuentan sólo con lacaracterización morfológica de las diferentesentradas. En este sentido. se planteó este trabajocon el objeto de caracterizar mediante marcadoresmoleculares: isoenzimáticos y de proteínastotales, diferentes cultivares de Coffea arabica.
MATERIALES Y MÉTODOS
Se tomaron veinte cultivares existentes en el bancode germoplasma dei campo experimental Brarnón(CIAE - Táchira, Venezuela) como referencia paraeste estudio (cuadro 1). Se cosecharonaleatoriamente 50 semillas de cada una de lasaccesiones, se sembraron en vivero, realizando laslabores culturales recomendadas (FONAIAP. 1988)Y a los cinco meses de edad. se procedió a laextracción y corrida electroforética de proteínastotales e isoenzimas. Para esto se tomaron cuatroplantas por accesión.

182
Cuadro I: Cultivares de Co./fea arabica analizadosGuadalupe Typica Mundo novo Leroy
amariUo amarilloBourbón rojo Semperflorcns Bourbón amarillo CeraVilla sarchis Catimor p-4 Caturraamarillo PacasPurpurascen s Mundo novo H íbrido de ceilán Pache
rojoBlue Híbrido de Híbrido Brarnón I
mountain timor salvadorei'io
La extracción de isoenzimas y corridaelectroforética se hizo de acuerdo a Bustamante y
Polanco (1999), en geles horizontales de almidón(12%) (Shields et al., 1983). En cada gel secolocó un total de veintitrés muestras, de lascuales, tres corresponden ai patrón, colocadas unaa cada extremo y otra aproximadamente en elcentro, con la finalidad de estandarizar la lecturade de los patrones electroforéticos. La tinción sereal izó de acuerdo a Vallejos (1983) Y las bandasse identificaron en función de su movilidadrelativa (m.r), (Laemml i, 1970).
RESULTADOS Y DISCUSIÓN
Las enzimas estudiadas no presentaronpolimorfismo. hecho que imposibilita establecerdiferencias entre los cultivares en estudio a travésde análisis de taxonomía numérica. Sólo unindividuo perteneciente a la variedad BlueMountain presentó pol imorfismo en la enzimaACO, mostrando una banda adicional de las dosque aparecen en el resto de individuos analizados.Aconitasa rACO) t:c. 4.2./ .3La literatura indica la presencia de 1-3 loci quecodifican para un monómero en esta enzima . Sinembargo. los resultados de este trabajo exhiben dosbandas bien definidas en todos los materiales;excepto en un solo individuo de la variedad BlueMountain, el cual presentó polimorfismo (Fig. I).Pudiera sei'íalarse dos posibilidades para explicar lapresencia de las mismas: I.) La existencia de dosloci en los diferentes cultivares. que corresponden auna enzima en el citosol y otra en la mitocondria(Wendel y Weeden . 1989) y 2.) La existencia de unsolo locus con heterocigosis fijada (por la condicióntetraploide y enzima monomérica) y por lo tanto seobservan dos bandas. La tercera banda que seobserva en el individuo de BIlIe Mountain. podríacorresponder a un nuevo alelo dei locus ubicado a0.92 de m.r La baja frecuencia de esta banda 0.1(1II O individuos muestrados) en dicho cultivar.podría ser producto de una mutación a nivel deiindividuo en estudio, que no se encuentra en laplanta madre. ya que en caso contrario se hubieseobservado en más de un individuo . De ser ciertaesta suposici ón. Ia planta en estudio podríaencontrarse en estado heterocigota.
Fosfogluconato lsomerasa (PCI) E.C. 5.3./.9En el patrón electrofo r ético presentado por laenzima PGI. se definen dos rcgi ones bien separadasy de marcada actividad (fig I). Cada una de éstaspodría representar un locus diferente. una isoe nzimaen el citosol . otra en los plastidios ( Wendel yWeeden. 1989). La reg i ón de menor migraciónprcsenta dos bandas bien definidas; aunquc la quese ubiea a 0.66 de nu . parec iera que corresponde ados bandas. De ser asi. este loeus tcndría dos a lelosen estado de heteroeigosis fijada (enzima dim érica),mientras que el otro locus sería homocigota .Figura 1: Patr ón clcctro for ético de las enzimasAconitasa y PG I. co rrcspo ndie nte a las :20 accesionesevaluadas
1 11
11 7 1
lJúú
.\CO \CO 1
* Valores de movilidad relativ a

EI nivel de autogamia que presenta C. arabica,justifica la poca variabilidad tanto en lascaracterísticas morfológicas como a nivel molecularen todos los individuos de un mismo cultivar, peroentre diferentes cultivares no se esperaría el mismocomportamiento. A nivel interespecífico existemayor variabilidad, ya que cada una de éstaspresentan autoincompatibilidad, lo que las hacealógamas obligadas, permitiendo de esta maneraampliar la variabilidad existente dentro de cadapoblación en cada una de las especies, por talmotivo se detecta polimorfismo isoenzimático(Berthou et al., 1980; Montagnon et al., 1992, 1993;Lopes, 1993).Como lo sefialan Berthou et ai (1980), los posibespadres de C. arabica son muy cercanosgeneticamente. En tal sentido, posiblementepresentan algunos loci de isoenzimas idénticos y elhíbrido interespecífico (estéril) presenta un juegocromosómico correspondiente a cada uno de lospadres; aI duplicarse, restaurando de esta manera lafertilidad; sus gametas constarán con el juegocromosómico correspondientes a cada uno de lospadres, lIevando los dos alelos que aportó cada unode los padres, lo que genera patroneselectroforéticos en condición de heterocigosisfijada.
REFERENCIAS
Berthou, F., Manthieu, C. and Vedei, F. (1983).Chloroplast and mitochondrial DNA variation asindicator of phylogenetic relationships in the genusCofJea L. Theor. Appl, Genet. 65: 77-84.Berthou, F., Trouslot, P., Hamon, S., VedeI. F. andQuetier, F. (1980). Analyse en électrophorêse dupolymorphisme biochimique des caféiers. CaféCacao Thé. 24(4): 313-326.
183
Bustamante, J. y Polanco, D. (1999). Extracción ycorrida electroforética de proteínas totales eisoenzimas de café (CofJea arabica L.). Enpublicación.Fondo Nacional de Investigaciones Agropecuarias.(1988). Paquete Tecnológico para la Producción deCafé. Serie de Paquetes Tecnológicos. Maracay,Venezuela. N° 6. 192 pp.Laemrnli, U. (1970). Cleavage of structure proteinsduring assembly of the head of bacteriphage T4.Nature 22: 680-685.Lopes, C. 1993. Phylogenetic studies of somespecies of the genus CofJea. 11- Numerical analysisof isoenzymatic data. Rev. Brasi I. Genet. 16(2):393-407.Montagnon, C; Leroy, T. and Yapo, A. (1992).Diversité génotypique et phénotypique de quelquesgroupes de caféiers (CofJea caephora Pierre) encollection. Café Cacao Thé. 36(3): 187-198.Montagnon, C; l.eroy, T. and Yapo, A. (1993).Caractérisation et eva1uation de caféiers Coffeacanephora prospectés dans des plantations de CoteD'lvoire, Café Cacao Thé. 37(2): 115-119.Shields, C; Orton, T. and Stuber, W. (1983). Anoutline of general resource needs and proceduresfor the electrophoretic separation of acti veenzymes from plant tissue. En S. Tanksley y T.Orton. (Eds.). Isozymes in Plant Genetics andBreeding. Part A. 443-468. Elsevier Sei. Publi. B.V. Amsterdam.Vallejos. E. (1983). Enzyme activity staining. EnS. Tanksley y T. Orton. (Eds.). Isozymes in PlantGenetics and Breeding. Part A. 469-516. ElsevierSci. Pub1i. B. V. Amsterdam.Wendel, J. and N. Weeden. (1989). Visualizationand Interpretation of Plant Isozymes. En: D. Soltis.and P. Soltis (eds.). Isozymes in Plant Biology. 545. Dioscorides Press, Portland.


185
MAPEAMENTO DE GENE DE RESISTÊNCIA A FERRUGEM-DO-CAFEEIRO (Hemileia vastatrixBerk. et Br.) COM MARCADOR RAPD
Nara S. Tedesco l•2
; Ney S. Sakiyama'<; Laércio Zambolím'r', Terezinha A. Telxeira'r"; Antônio A.Pereira"; Cássia C. H. Sakiyama I
IB10AGRO/UFV; 2DFT/UFV; 3DFP/UFV; 4EPAMIG. 36571-000 Viçosa,MG.Apoio Financeiro:FAPEMIG. FINEP, CNPq, PNP&DCafé
INTRODUÇÃO
A ferrugem-do-cafeeiro é um problema que afetamundialmente a cultura do café, resultando numaqueda de cerca de 35% da produção. O fungocausador da ferrugem-do-cafeeiro, Hemileiavastatrix Berk. et Br., é parasita obrigatório,facilmente disseminado. principalmente pelovento. Ao entrar em contato com o hospedeiro,sob condições favoráveis, como temperatura entre21°C e 23°C e na ausência de Iuz, germina,penetra e se multiplica neste. Os sintomas e sinaissão facilmente identificados pois este fungo formauma 'massa' de esporos alaranjada (uredosporos)na face abaxial da folha, enfraquecendo a planta epodendo levá-la à morte.Após a disseminação do fungo, muitos trabalhosjá foram realizados visando obter dados maisaprofundados sobre suas causas, formas decontrole e tentativas para se obter cultivaresresistentes. Acredita-se que o uso de variedadesresistentes é o método mais eficaz e econômico.PEREIRA (1995), estudou a herança daresistência às raças fisiológicas 1\ e XXV deHemileia vastatrix Berk. et Sr. em setecombinações resultantes de hibridações doHíbrido de Timor com cafeeiros das cultivaresCatuaí e Mundo Novo. Os resultados analisadosmostraram que a resistência a este fungo, nosdescendentes de uma das introduções, é conferidapor três genes de segregação independente e comdominância completa, e na descendência da outraintrodução é controlada por um único gene comdominância completa.Diante dos resultados obtidos neste e em outrostrabal hos, ficou destacado o elevado nível deresistência a H. vastatrix nas populaçõesderivadas do Híbrido de Timor. Essa e outrascaracterísticas de interesse, ai iadas à altacapacidade produtiva observada, em condições decampo, para as populações derivadas do Híbridode Tirnor, ressaltam o potencial desse híbrido,para o melhoramento genético do cafeeiro,visando à obtenção de cultivares resistentes àferrugem (PEREIRA, 1995).O presente trabalho teve como objetivo omapeamento de gene de resistência à ferrugemdo-cafeeiro com marcador RAPD tRandom
Amplified Polymorphic DNA) (WILLlAMS et al.,1990).
MATERIAL E MÉTODOS
Material genéticoA população utilizada no presente trabalho, éproveniente do cruzamento interespecífico entreUFV 376-2 (Híbrido de Timor - material recebidodo CIFC, em Portugal) e UFV 2144-32 (CatuaíVermelho - do Campo EP 20.1) do qual surgiu H332-3 (híbrido FI - do campo de seleções dehíbridos da UFV). Após obtenção do híbrido FI, omesmo foi util izado num retrocruzamento comUFV 2144-32, originando a planta H 682-21.Uma população de 261 plantas RC1F1, obtida porautofecundação de H 682-21 foi avaliada emcondições de Câmara de crescimentoapresentando uma proporção 3: 1 em relaçãoresistência: suscetibi Iidade à ferrugem-docafeeiro.
População f 2
UFV 2144-32 X UFV 376-2
+UFV 2144-32 f H 332-3 (FI)
H 682~1 (RC I )
t0População RC IF1 (261 plantas)
Extração de DNAO DNA foi extraído de folhas jovens seguindo oprotocolo modificado de Doyle & Doyle (1990)com a adição de PVP-40 (solúvel) no tampão deextração. Após a extração o DNA foi quantificadoem espectrofotômetro, corrido em gel de agarose0,8 % e armazenado a 4°C. Na época daamplificação foi diluido em TE (Tris HCI 10 mM,EDTA 1 mM, pH 8.0) para uma concentraçãofinal de 10 ng/ul,
Preparo de bulks de DNA para identificação demarcadores RAPDA análise de mistura de segregantes (BSA-BulkSegregam Analysisi, é um método rápido de

186
RESULTADOS E DISCUSSÃO
para eliminar, durante uma hora, o excesso doproduto. O padrão molecular foi visualizado sobluz ultra-violeta e fotodocumentado.
Dos 125 primers testados 14 resultaram embandas pol imórficas entre os bulks R (plantasresistentes) e S (plantas susceptíveis). A Figura Imostra o pol imorfismo apresentado entre os bulkscom a utilização do primer OPC-09 .
Análise de co-segregaçãoPrimers apresentando banda polimórfica entre osbulks S e R foram utilizados para a análise de cosegregação na população segregante para aconfirmação da ligação gênica existente entre osmarcadores e o gene de resistência à ferrugem. ,
,t7l O ... Nq "; '7 ";(,) (,) (,) (,)a. a. Q. Q.O O O O
~oQ.O
~ q(,) (,)a. Q.O O
~ li)q q(,) (,)Q. Q.O O
,.. N ('I)
q q q(,) (,) (,)Q. Q. Q.O O ~
identificação de marcadores ligados a uma regiãoespecífica do genoma (MICHELMORE et al.,1991 ).De uma população segregante F2, 10 plantasresistentes e dez plantas suscetíveis foramselecionadas para respectiva produção de bulk deDNA, sendo assim cada bulk constituído porindivíduos idênticos para a característica deresistência (ou suscetibilidade) à ferrugem. Decada amostra de DNA das plantas escolhidas,foram adicionados em um novo tubo 100 ul.,possuindo cada um dos dois bulks, ao final dopreparo, 1000 ul. de mistura de DNA numaconcentração de 10 ng/mL em TE. Paracomposição do bulk R (resistente) foramselecionadas as plantas que apresentaram maiselevado nível de resistência a H. vastatrix deacordo com avaliação em casa de vegetação e paracompor o hulk S (suscet ível) as plantas queapresentaram mais elevado nível de infecção pelofungo. Os dois bulks de DNA foram usados paraamplificação dos fragmentos com a finalidade dese identi ficar marcadores Jigados aos fatores deresistência do doador, utilizando-se vários primerstomados ao acaso.
Amplificação de fragmentos de DNA paraobtenção de marcadores RAPDCento e vinte e cinco primers de 10 bases, foramutilizados na amplificação dos fragmentos deDNA na tentativa de encontrar marcadorespolimórficos entre os dois bulks. Cada reaçãoobteve os seguintes componentes: 2,5 ul, de KCI50 mM; 2,5 ut. de TRIS-HCI 10 mM; 2,0 ut. deMgCI22 mM; 125 ut. de dNTPs 100 f-lM; 1,0 ul,de primer; 0,2 fiM; 1,0 ul, de Taq DNApolimerase I U e 2,5 ul, de DNA 25 ng, somando
um total de 25 ul. por reação ao completar ovolume com água ultra pura. A amplificação foiefetuada em termociclador Perkin Elmer 9600programado para: I ciclo de I minuto a 95°C. 40ciclos de 15 segundos a 94°C. 30 segundos a 35°Ce I minuto a n°c. e I ciclo de 7 minutos a n°c.
Análise por eletroforese dos fragmentosamplificados e visualizaçãoA separação dos fragmentos amplificados foirealizada em gel de agarose 1,2% a 100 V por 3horas . Após a eletroforese os fragmentosamplificados foram corados em uma solução debrometo de etídeo (10 mg/mL) por 30 minutos e,em seguida, a solução foi substituída por água
Figura \- Marcadores RAPD em bulks de cafeeirosresistentes (R) e bulks susceptíveis (S). A seta indicamarcador polimórfico entre bulk R e S.
Dos 14 primers apresentando bandas polimórficasentre os bulks R e S, quatro foram selecionadospara análise preliminar de co-segregaçãoutilizando-se as 20 plantas individuaispertencentes aos bulks R e S .Da amplificação dos quatro primers selecionadospara análise de co-segregação de 20 plantas. doisprimers apresentaram polimorfi smo : ambosresultaram em dois recombinantes em 20 plantas.A análi se de co-segregação na população de 80plantas foi efetuada com o primer OPC-09 porapresentar melhor nitidez de bandas (Figura 2).

II
!I
187
o objetivo de se encontrar outros marcadores maispróximos ao gene de resistência. Outra populaçãoapresentando resistência condicionada por doisgenes também encontra-se em estudo.
Figura 2- Marcador RAPD obtido pelo primer OPC-09apresentando co-segregação com a reação deresistência ( R ) c susceptibilidade (S) à ferrugem-docafeeiro .
Pela análise de co-segregação. concluiu-se que omarcador RAPD obtido com O prirner OPC-09encontra-se ligado ao gene de resistência àferrugem-do-cafeeiro. com a distância estimada de9.6 eM. Este trabalho continua em execução com
... - .....-". ~
~.. .-- -- .. _- --_... - -
REFERÊNCIAS BIBLIOGRÁFICAS
DOYLE. J. J, & DOYLE, J, L. J 990. Isolation ofplant DNA frorn fresh tissue. Focus, 12.(1) :13-15 .
MICHELMORE, R. W .. PARAN. I. & KESSELLR. V, 1991. Identification of Markers linkedto diasease resistence genes by bulkedsegregant analysis: a rapid rnethod to detectrnarkers in specific genornic regions usingsegregant populations. Proc. Natl Acad. Sei ..88: 9828-32.
PEREIRA, A. A. J 995. Herança da Resistênciaa Hemileia vastatrix Berk et Br emCafeeiros Derivados do Híbrido de Timor.Viçosa. MG . UFV. 66p (Tese M. S .) .
WILLlAMS. J.G .K.; KUBELlK. A.R .; L1VAK.K.J.; RAFALSKL J.A.; TINGEY. S.V, 1990.DNA polyrnorphisrns amplified by arbitrar)'prirners are useful as genetic markers , NucleicAcids Researchs, 18(22): 6531-6535 .

Consórcio Brasileiro de Pesquisa eDesenvolvimento do Café
Coordenação Embrapa
Enibreue Amazônia Oriental
Embrapa Amapá
CATI
IAC
IALIB
IEA
ITAL
Embrapa Acre
Embrapa Rondônia
Embrapa CaféEmbrapa Cerrados
Embrapa Recursos Genéticos e Biotecnologia
SARC/MA
UFLA
UFV
UFUUNIUBE
EMATER-MG
EPAMIG
Instituições Participantespor Unidade da Federação
\\
\EMATER-PRIAPAR
OCEPAR
UELUEM
UFPR
FUNAPE
-. ASCCON
EBDA
UESB
EMATER-RJEmbrapa Agrobiologia
Embrapa Agroindústria de Alimentos
Embrapa Solos
FGV
PESAGRO-RJ
UENF


191
BIOTECNOLOGIA NO DESENVOLVIMENTO DE CULTIVARES NUM MENOR PRAZO NO"MODELO IAPAR DE CAFÉ ADENSADO"
Tumoru Sera
Instituto Agronômico do Paraná - IAPAR/Programa Café. Rod. Celso Garcia Cid, km375, C.PA81 CEP86001-71O, Londrina, Paraná, Brasil
l-INTRODUÇÃO
o melhoramento genético do cafeeiro,especialmente para a produção, não tem sidofácil. Isto se deve à existência do período juvenil,à necessidade de se avaliar vários anos deproduções consecutivas para se conhecer acapacidade produtiva a longo prazo e à existênciada acentuada oscilação anual de produção onde ocafezal é conduzido a pleno sol e sem sombra(SERA, 1987). Apesar disso, os programas demelhoramento executados em vários centros depesquisa tem levado a grandes progressosgenéticos. O programa iniciado em 1933 noInstituto Agronômico de Campinas obteveprogressos de até 240% quando se comparou acultivar Mundo Novo em relação a primeiracultivar plantada no Brasil, Árabica (FONSECA,et aI. 1978). Os métodos de melhoramentonormalmente empregados no Brasil, consistem naseleção de plantas individuais seguida daavaliação de progênies, ou ainda de hibridaçãointra e interespecífica e seleção genealógica. Emadição, o método de retrocruzamento tem sidoaplicado para a transferência de caracteresespecíficos, tais como internódios curtos eresistência a pragas e doenças, presentes emoutras cultivares ou em espécies relacionadas aCoffea arabica L.. A seleção de plantasindividuais pode ser realizada tanto em umaplantação comercial, como em parcelasexperimentais especialmente plantadas com opropósito de melhoramento genético. Nesteúltimo caso, a produção é registrada por váriosanos sucessivos para a identificação de genótipossuperiores (M EDlNA et al., 1984).O conhecimento profundo do ambiente decultivo, processamento, comercialização econsumo é de fundamental importância nodesenvolvimento de cultivares de sucesso. Oambiente de uso destes cultivares em foco,tomado como exemplo na apresentação do tema édentro do "Modelo tecnológico para a produçãode café no Paraná" ("Modelo IAPAR de caféadensado"). Consiste resumidamente do seguinte:• Modelo de produção constituído por maisde 75 tecnologias;
• Espaçamento de plantio adensado entre 8 e12 mil plantas/ha;• Produtividade potencial na propriedadeótima em torno de 80sc.b.lhalano;• Lucro de 50 em 10 ha, pela eficiência(produtividade, qualidade, menor custo/se);• Criação de I emprego a cada ha de café aocusto de US$5000 / ha;• Trabalhadores produtivos e qualificados comosolução;• Reativação econômica em mais de 200municípios cafeeiros do Paraná;• Adequação a micros, pequenas, médias egrandes propriedades;• Retorno inicial de investimento com 2,5 anosdo plantio;• Estabilidade econômica com diversificaçãointegrada e moderada na propriedade;• Plantio parcelado anual (4 a 10 anos),semestral e trimestral no ano;• Adoção gradual e contínua de novastecnologias e cultivares;• Independência em relação a financiamentos;• Adequação a produtores descapitalizados;• Tecnologia ecológica com manejo de solo,pragas e doenças;• Convivência com os nematóides através domanejo integrado de medidas de controle;• Custo menor de adubação/saca, com manejodo solo e da nutrição do cafeeiro;• Convivência com as geadas recuperando-seapós uma safra de descanso;• Manejo varietal regional. local e topoclimática, adaptado a climae solo específicos;• Colheita escalonada usando 4 grupos devariedades, desde precoce até super-tardia;• Café de qualidade a baixo custo, com o uso detecnologias simples;• Café é uma necessidade para o trabalho urbanoe o consumo está em ascensão.O principal objetivo é tomar forte a cafeiculturaparanaense dentro do "PLANO CAFÉ DOPARANÁ" aumentando a RENTABILIDADE ea ESTABILIDADE econorruca dosCAFEICULTORES, através da eficiênciaprodutiva ao nível da propriedade. A eficiênciaprodutiva será alcançada com base em:

1) Aumento da produtividade;2) Redução do custo de produção por saca;3) Melhoria da qualidade do produto;4) Diversificação agrícola integradana propriedade.O melhoramento genético do cafeeiro se darádentro deste ambiente de cultivo selecionando edesenvolvendo cultivares que auxiliem de algumaforma nos objetivos e estratégias do "Plano Cafédo Paraná".
2. NECESSIDADES DE MELHORAMENTOGENÉTICO NO CAFEEIRO
Para alcançar a eficiência na produção de café dequalidade a menor custo há necessidade dedesenvolver vários projetos de pesquisa:• Potencial de produtividade de 80 sc.b.lha/anopara produtores ótimos não está acessível para amaioria, necessitando amenizar vários pontos deestrangulamentos através de variedades e seumanejo adequado;
-. A realização do potencial de produção de caféde qualidade está tendo várias dificuldades.necessitando uniformizar a maturação, escalonara colheita e desenvolver cultivares de grãosmaiores;• Dentre os componentes do custo de produção,a colheita é o componente mais oneroso, tanto deplantio tradicional como de adensado.necessitando escalonar a colheita e desenvolvercultivares de porte pequeno de fruto graúdo;• A vulnerabilidade a oscilações ambientais éalta. necessitando de cultivares menossuscetíveis a geada, a seca e a solo pobre;• Controle de ferrugem está dificil emespaçamentos adensados, onerando o custo edepreciando a qualidade e, com isso, aumentandoa suscetibilidade às geadas, necessitandodesenvolver novas variedades com mais genes deresistência e tolerância, mais adaptados para estesistema de cultivo;• Nematóide, especialmente Meloidogyneincognita com as suas 5 raças fisiológicasconhecidas é limitante na propriedade infestada,necessitando intensificar o desenvolvimento deporta-enxertos resistentes e cultivares pés francosresistentes;• A bacteriose Pseudomonas syringae pv.garcae está se tornando uma doença limitante emregiões de altitude e em faces sul e lestenecessitando desenvolver cultivares resistentes;• Bicho mineiro está se tornando mais danoso ede controle diflcil necessitando intensificar odesenvolvimento de cultivares resistentes;
192
• Alto custo de controle das ervas daninhas nocafé em formação justifica o desenvolvimento decultivares resistentes a herbicidas;• Há necessidade de se precaver da introduçãode antracnose dos frutos e iniciar os trabalhos deaval iação da resistência do germoplasmadisponível;• Controle de broca com incorporação do geneBT é necessária;• Há necessidade de reduzir o tempo gasto nodesenvolvimento de cultivares.
3. REDUÇÃO NO TEMPO GASTO PARAOBTENÇÃO DE CULTIVARES DE CAFÉ
Os métodos de melhoramento genético deplantas utilizados normalmente em espéciesanuais podem ser utilizados em cafeeiro, plantasperene, desde que respeitadas as suascaracterísticas. Contudo, o melhoramento deplantas perenes como o cafeeiro apresentaalgumas particularidades muito importantes,necessitando adotar procedimentos especiaispara não ter um programa de melhoramento debaixa eficiência:• Períodos normalmente mais longos para quese obtenha as flores para a realização doscruzamentos e para a produção de sementes.• Uma importância relativamente maior de seavaliar bem as características agronômicas, nãosomente a produtividade e a qualidade, antes darecomendação, devido ao custo relativamentealto de substituição das cultivares.• Custo alto da avaliação de campo devido anecessidade de maiores áreas e tempo paraexperimentação.• Necessidade de avaliação da precocidade elongevidade produtiva.• Necessidade de avaliação da oscilação anualda produção.• Florescimento e produção anual sobre amesma planta.
4. GANHO GENÉTICO ANUAL NASELEÇÃO EM CAFÉ
Estas características limitam a adoção de algunsmétodos de melhoramento normalmenteutilizados em cultivos anuais de forma clássica.A conseqüência destas características é umamenor eficiência de um programa demelhoramento de uma espécie perene, se adotaros mesmos procedimentos correntes adotadospara as plantas anuais. Esta baixa eficiência, émedido pelo baixo ganho genético da seleção por
IIIJ
I

ano. A seguir será definido e comparado pelaexpressão do ganho genético por ano (G.S.a) até
193
a obtenção de cultivares em plantas anuais eperenes propagadas sexuada ou assexuadamente.
G.S. = (D.S. x h2) 7 (F n x Anos por ciclo)Onde,G.S. = ganho genético anual;D.S. = diferencial de seleção;h2 = Herdabilidade = Variância genética (cr2 G) 7 Variância fenotípica (cr2 F);Fn = número de gerações de seleção necessárias para obtenção de cultivares.Plantas anuais. A h2 depende do esquema seletivo, e geralmente conduzem um ciclo de seleção por anorepetindo várias gerações.
G.S.a.Anuais = D.S. x
Onde.cr2 G = variância genética;cr2 F = variância fenotípica
Cultivar de café tipo clone. Se o cafeeiro for propagada vegetativamente, a cr2 G = cr2 a + cr2 d + cr2 ie então o ganho genético esperado por ano seria:
cr2 a + cr2 d + cr2 iG.S.a.C1onais = D. S. x -------------------------------- 7 Anos por ciclo
cr2 a + cr2 d + cr2 i + cr2 ecr2 a = Variância genética aditiva;cr2 d = Variância genética dominante;cr2 i = Variância genética epistática;cr2 e = Variância ambiental.Ou seja, toda a cr2G poderia ser aproveitada e, a partir da obtenção de um genótipo superior, geralmenteheterozigótica, estas caracteristicas favoráveis seriam transmitidas vegetativamente para as geraçõesposteriores, resultando em G.S. maior do que as cultivares propagadas por semente devido ao h2 maior eFn= I.
Cultivares de café tipo linhagem propagadas por sementes. Já em plantas propagadas por sementes evisando a obteção de genótipos homozigotos na maioria dos loci, o G.S. seria:
cr2 aG.S.a.Linhagem=D.S. x ----------------------------- 7 (Anos por ciclo x Fn)
cr2 a + cr2 d + cr2 i + cr2 e
Ou seja, apenas a cr2a poderia ser aproveitadaneste caso e. necessariamente, teríamos queconduzir as populações obtidas por algumasgerações até obter níveis de homozigose quepertimissem fixar as características favoráveis,resultando em G.S. muito menor que aspropagadas vegetativamente ou a obtida emplantas de ciclo anual.Assim. torna-se evidente que dependendo daforma de propagação do cafeeiro e do tipo decultivar que se pretende obter, pode-se necessitarde maior ou menor tempo para a obtenção degenótipos superiores que possam ser utilizadoscomercialmente pelos produtores. A adoção dealgumas estratégias especiais no melhoramentogenético do cafeeiro pode reduzir muito o tempogasto no desenvolvimento de novas cultivares,
além de possibilitar uma grande redução da áreaexperimental, resultando em grande economia eaumento substancial na eficiência do programade melhoramento.A redução do número de anos e do número degerações. ou seja, a redução no tempo gasto parase chegar a uma nova cultivar é o ponto chavepara aumentar a eficiência ou o progressogenético no melhoramento genético do cafeeiro.
5. REDUÇÃO DO TEMPO GASTO NODESENVOLVIMENTO DE CULTIVARESDE CAFÉ
No desenvolvimento de cultivares, a redução dotempo gasto em uma geração ou ciclo de seleçãoou na obtenção de cultivares é de fundamental

importância para aumentar a eficiência de umprograma de melhoramento, onde é comumcomeçar a florescer somente após 2 ou atémesmo mais anos após a semeadura (SERA,1987). Muito freqüentemente é fator limitanteno desenvolvimento de um projeto demelhoramento genético, num prazo razoável.Além disso, uma das caracteristicas desejáveis nacultura de café é que comece a produzir maiscedo e, ao mesmo tempo, continue produzindopor mais tempo. Para isso, é necessário avaliar aprodução durante a vida útil produtiva da plantaou da progênie para dizer com segurança que talseleção apresenta esta característica. Para oscultivos em que o produto comercial é semente,o problema é maior, pois, antes de iniciar aprodução, existe o período juvenil, préreprodutivo.Após o período juvenil, um uruco ensaio oucampo de seleção exige muitos anos deobservações durante a qual a área experimentalnão pode ser aproveitada para outros fins. Destamaneira, muito espaço, tempo e recursos sãoconsumidos, enquanto que o número de plantasque podem ser mensurados são limitados.Assim, a possibilidade de se conduzir umprograma de melhoramento genético racional decafeeiro envolvendo múltiplos objetivosutilizando, por exemplo, um esquema de retrocruzamento. de seleção recorrente ou de umgenealógico. depende grandemente da reduçãodo número de gerações ou ciclos de seleção e dese efetuar a seleção antecipada para produçãocom menor número de anos por geração. Outrascaracteristicas menos complexas comoresistência a pragas, doenças e adversidadesambientais podem ser avaliados e selecionadosdurante o periodo juvenil (SERA, 1984).Os melhoristas de plantas estão constantementetentando identificar novos procedimentos quepossam aumentar a eficiência nodesenvolvimento de cultivares. A eficiência deestratégias alternativas no melhoramento deplantas perenes pode ser avaliado em termos daquantidade de ganho genético realizado por ano(G.S.a).G.S.a é estimado pela incremento naperformance média de uma progênie que érealizada em cada ciclo. Um ciclo de seleçãoinclui o estabelecimento de uma populaçãosegregante, desenvolvimento de genótipos,aval iação de genótipos. seleção dos genótipossuperiores e utilização dos genótiposselecionados como parental para formar umanova população para um novo ciclo de seleção. O
194
tempo consumido em cada ciclo de seleção podevariar consideravelmente. Na comparação deestratégias alternativas, o G.S. precisa serexpresso em base anual. O melhorista precisatraçar estratégias que utilizem racionalmente osrecursos humanos, espaço, material e tempodisponíveis para propiciar o maior ganhopossível por ano.Para as plantas perenes como cafeeiro existemprocedimentos, métodos e técnicas alternativasem que o tempo gasto e o número de plantas aserem avaliados no campo durante o processo deobtenção de cultivares podem ser reduzidodrasticamente:1. Redução do período juvenil;2. Seleção no período juvenil;3. Seleção antecipada para produção nos anosiniciais;4. Seleção em gerações precoces;5. Métodos que não avaliam a produção
(população, SSD e retro-cruzamento);6. Uso de técnicas biotecnológicas e engenharia
genética;7. Obtenção de cultivares duplo-haplóides;8. Uso da heterose de cultivares híbridas fIf2 e sintéticas;9. Propagação vegetativa para uso de cultivaresdo tipo clone;10. Lançamento antecipado de cultivares.Cada um dos métodos, técnicas e procedimentosdisponíveis para o desenvolvimento de cultivaresapresentam vantagens e desvantagens. Aprobabilidade de obtenção de uma cultivarsuperior pode ser aumentado pela elevação nonúmero de genótipos que podem ser testados acada ano, porém com aumento de custos queprecisam ser minimizados por estratégiasalternativas, usando técnicas e procedimentosespeciais, junto com métodos de melhoramentoapropriados para plantas perenes. Estratégiasapropriadas que possibilitem o rápido avanço degerações como a clonagem, duplo-halóides elançamento antecipado de cultivares podem tergrande impacto na eficiência de um programa demelhoramento de plantas perenes como ocafeeiro.
5.1. Redução do período juvenilEm geral. um projeto de obtenção de cultivaresenvolve várias gerações de seleção.Consequentemente, o comprimento de umageração de reprodução é muito freqüentemente ofator limitante para encerrar um projeto numprazo razoável.
1
JI

a) Seleção para genótipos de florescimentoprecoce. Ocorre no cafeeiro da mesma formacomo ocorre em outras plantas perenes como emmacieira, pereira, videira, figueira ecitrus.(JANICK & MOORE, 1975; SERA,1985).
b) Semeadura imediata após a colheita. Écomum a espera de condições de temperaturanatural ideal para a semeadura. Usando-segerminação em condições controladas e aplasticultura no inverno. pode-se ganhar 6 mesesno caso do cafeeiro. No cafeeiro, se conseguirplantar no campo em outubro pode ganhar umano agrícola do café. Se plantar em abril-maio játerá perdido um ano a mais por ciclo (SERA,1984).
c) Boas condições de cultivo. Tanto no viveirocomo em campo, adubação, irrigação,temperatura, controle de invasoras e parasitas,luminosidade. etc., em condições próximas doideal podem reduzir o período juvenil (JANICK& MOORE, 1975; SERA, 1984), o mesmoocorrendo em cultivo in vitro (SONDHAL et ai,1981) ou em campo (MÔNACO et al.. 1978).
d) Porta enxerto-ananizante. A enxertia porgarfagem de plântulas ou borbulhas de gemasvegetativas de plantas perenes lenhosas em portaenxertos fracos, vel hos ou ananizantes tem sidofreqüentemente indicada. como no caso demacieira, podendo reduzir o período juvenil paramenos da metade do tempo. Enxertia porgarfagem de plântulas de perenes lenhosas emporta enxerto fraco ou ananizante é uma técnicafreqüentemente aplicada como em macieira(JANICK & MOORE. 1975), podendo sertentada em cafeeiro.
e) Indução floral ambiental. A deficiênciahídrica pode induzir a redução do tempo deflorescimento até a maturação. A deficiência denitrogênio durante o enchimento dos grãos leva asementes pequenas e de maturação precoce. Oaumento do teor de fósforo tende a reduzir operíodo de maturação .. A exposição contínua deluz é o meio apropriado de reduzir o tempo deflorescimento de plantas de dias longos. Paraplantas de dias curtos o mais adequando é umamenor exposição a luz mas, o efeito de diascurtos depende do fotoperíodo ótimo. Para asplantas tropicais podem ser mais apropriados odia de 8 horas (WELLENSIEKI. 1962). Atemperatura alta reduz o tempo entre O'
195
florescimento e a maturação dos frutos nocafeeiro (SERA, 1979) permitindo a semeaduraem maio e plantio no campo em outubro.
5.2. Avaliação e seleção juvenil paraprodutividadeA seleção juvenil para produtividade pode serpraticada para as características qual itativas equantitativas de alta herdabilidade, altamentecorrelacionadas com a produção. A seleção podeser feita no estádio de plântulas. mudas noviveiro ou em campo. antes da ocorrência daprimeira produção.
a) Seleção em plântulas. Características comovolume radicular e vigor vegetativo podem serselecionadas desde o estádio de folha cotiledonaraté algumas folhas definitivas, antes dotransplante para o recipiente de mudas, reduzindomuito o custo.
b) Seleção em mudas no viveiro. Característicascomo a resistência a insetos, doenças,nernatóides, seca. alumínio e geada podem serselecionadas junto com outras característicascorrelacionadas com a produção antes de seremlevadas para o plantio em local definitivo,economizando espaço no campo e tempo gastona avaliação e manuseio dos dados, liberandomais espaço. recursos e tempo para materiaismais valiosos.
c) Seleção juvenil no campo. Característicascomo ramificação, diâmetro da planta.resistência ao vento, resistência a temperaturasextremas e número de flores por axila podem serselecionadas nas condições de campo. antes de se 'realizar a primeira colheita. Pode-se plantar 3 oumais plantas por cova e selecionar apenas uma.Com base em avaliações de campo, pode-seeliminar alguns tratamentos por evidenteinferioridade - agronômica, aumentando aeficiência da seleção e economizando tempo erecursos.
5.3. Seleção antecipada para produtividade euso de indice de seleçãoProdutividade é uma característica quantitativadificil de ser trabalhada em programas dedesenvolv imento de cultivares que incorporamnovas características como resistência aosparasitos, a adversidades ambientais e mudançano sistema de cultivo ou de 'exigência de novasqualidades pelos consumidores.A compreensão dos componentes da produção e

suas correlações podem auxiliar na seleçãoantecipada dos genótipos promissores paraprodutividade. São comuns correlações negativasentre número e tamanho dos grãos, entreprodução de folhas e sementes. entre tamanho deplantas e número possível de plantas por área.etc. (GIFFORD & EVANS, 1981).Em plantas perenes. onde uma vez iniciada a
produção. a mesma planta produz por váriosanos. usualmente por 10-20 anos. a produçãoinicial pode não representar a produção total.Assim. podem existir genótipos que apresentamprecocidade de produção. isto é. produção inicialmaior. e menor longevidade de produção, isto é.senescência precoce. enquanto que podem existirgenótipos com produção tardia e longevidade deprodução. O genótipo ideal seria aquele quecomeça a produzir cedo com alta produtividade ese mantêm produtivo por um período maior detempo. Assim. é necessário e possível umsistema de avaliação antecipado da produçãofutura que garanta um avanço rápido em. cadageração de seleção resultando em obtenção maisrápida de um novo cultivar (QUADRO 1) commaiores progressos genéticos.
5.4. índice de seleçãoA identificação antecipada de genótipos maisprodutivos com maior precisão do que aalcançada usando-se apenas a produção dos anosiniciais. é altamente desejável no melhoramentogenético do cafeeiro. Segundo MEDINA et aI.( 1984). cultivares de sucesso devem começar aproduzir cedo e manter produções altas por maisanos. As melhores progênies podem serescolhidos com base nas seis primeirasproduções consecutivas para representar 15 a 20anos de produções (CARVALHO. 1952;FAZUOLL 1977; CARVALHO et ai., 1985).Entretanto, a obtenção de produções num talperíodo de tempo. para uma cultura perenearbustiva que começa a produzir somente apóstrês anos de período vegetativo ainda éconsumidora de tempo. além de ser consumidorade espaço e recursos. O coeficiente dedeterminação (R2) indica o porcentual em que aprodução real é estimada pela equação deregressão múltipla. SERA (1987) usando aregressão múltipla estimou estes coeficientespara um índice de seleção composto porcaracterísticas mensuráveis nos 3 primeiros anosde produção. Ao usar produções deste períodoobteve R2=O,615 para as 3 primeiras produções.R2=0.489 para as 2 primeiras produções eR2=0.070 para a primeira produção. esta última.
196
sendo muito baixo. A seguir são apresentados asestimativas dos melhores índices. incluindo-seoutras características correlacionadas com aprodução. para as características disponíveis atéo terceiro. segundo e primeiro ano de colheitas,bem como sem a produção.Três primeiros anos: R2=0.791 usando-se ascaracterísticas tamanho dos grãos. produção degrãos """ produção de frutos, altura da copa.diâmetro da copa, produção do 1°. biênio.incremento anual da produção nos 3 primeirosanos de produção e oscilação anual da produçãonos 3 primeiros anos.Dois primeiros anos: R2 = 0.717 usando-se ascaracterísticas tamanho dos grãos, produção degrãos """ produção de frutos. altura da copa.diâmetro da copa e produção do primeiro biênio.Primeiro ano: R2 = 0.571 usando-se ascaracterísticas tamanho dos grãos. produção degrãos -i- produção de frutos. altura da copa.diâmetro da copa e la. produção.Sem produção: R2 = 0.454 usando-se apenasaltura e diâmetro da copa.O R2 obtido usando-se produção e caracterescorrelacionados é sempre maior que apenasusando a produção. Usando-se apenas altura ediâmetro da copa obteve-se R2 quase igual aoR2 usando as duas primeiras produções. o queestá concordando com os resultados práticosalcançados por ARAÚJO (1977) e pelostrabalhos de SRINIVASAN( 1982) e WALYARO(1983). SERA (1987) comparando-se asclassificações obtidas pelas progênies. com basena produções totais observadas e as estimadasantecipadamente por índices verificou-se quepara se incluir as sete melhores progênies das 72(i = 10%) seriam necessários selecionar as 13melhores nos três primeiros anos. as 20 melhoresnos dois primeiros anos e as 50 melhores noprimeiro ano de colheita. Portanto. seria viávelantecipar a seleção com segurança razoáveltendo-se dados de produção e caracterescorrelacionados obtidos nos dois primeiros anosde colheitas.Assim. é viável a seleção antecipada na reduçãodo tempo gasto no desenvolvimento de cultivares(QUADRO 1) usando-se índices de produçãocomposto por produções iniciais e característicascorrelacionadas e, para garantir que as progêniesselecionadas antecipadamente incluam asprogênies realmente superiores em produçãototal. é aconselhável utilizar intensidades deseleção mais brandas. tanto entre como dentrodas progênies.
}l
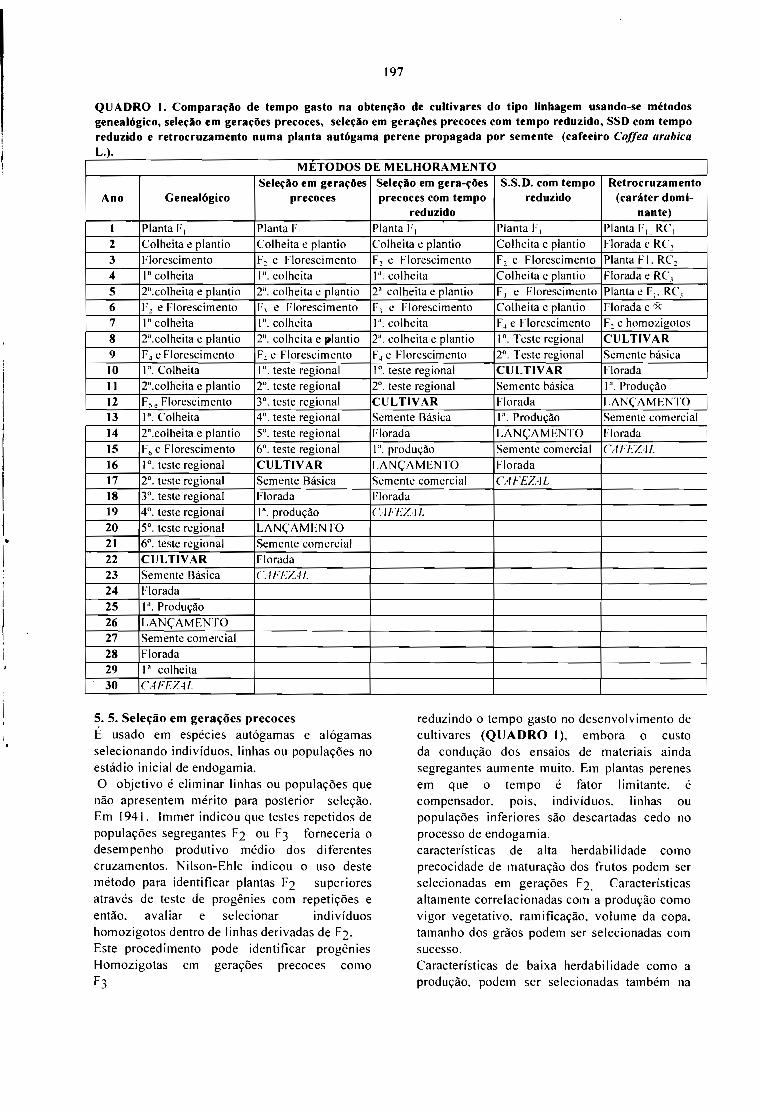
•
I
197
QUADRO I. Comparação de tempo gasto na obtenção de cultivares do tipo linhagem usando-se métodosgenealógico, seleção em gerações precoces, seleção em gerações precoces com tempo reduzido, SSD com temporeduzido e retrocruzamento numa planta autógama perene propagada por semente (cafeeiro Coffea arabicaL.).
METODOS DE MELHORAMENTOSeleção em gerações Seleção em gera-ções S.S.D. com tempo Retrocruzamento
Ano Genealógico precoces precoces com tempo reduzido (caráter domi-reduzido nante)
I Planta FI Planta FI Planta FI Planta FI Planta FI. RC I2 Colheita e plantio Colheita e plantio Colheita e plantio Colheita e plantio Florada e RC2
3 Florescimento 1'2 e Florescimento 1'2 e Florescimento 1'2 c Florescimento Planta F I. RC2
4 )" colheita I". colheita I". colheita Colheita e plantio Florada e RC,
5 2".colheita e plantio 2". colheita e plantio 2' colheita e plantio F] e Florescimento Planta e 1'\. RC,6 F2 e Florescimento F, e Florescimento F, e Florescimento Colheita e plantio Florada c é>:,'
7 ]" colheita I". colheita I". colheita F. e Florescimento F2 e homozigotos
8 2".colheita e plantio 2". colheita e ~Iantio 2". colheita e plantio 10. Teste regional CULTIVAR9 F. e Florescimento F. e Florescimento F. e Florescimento 20. Teste regional Semente básica10 I". Colheita 1°. teste regional 1o. teste regional CULTIVAR Florada
11 2".colheita e plantio 2°. teste regional 20. teste regional Semente básica I". Produção12 F ~ e Florescimento 30. teste regional CULTIVAR Florada LAN(AMENTO13 Ia. Colheita 4°. teste regional Semente Básica la. Produção Semente comercial14 2".eolheita e plantio 5°. teste regional Florada LANÇAMENTO Florada15 F" e Florescimento 6°. teste regional l", produção Semente comercial CAFF.ZAL16 1°. teste regional CULTIVAR LANÇAMENTO Florada17 2°. teste regional Semente Básica Semente comercial CAFEZAL18 3°. teste regional Florada Florada19 4°. teste regional la. produção C.lfEZAL20 5°. teste regional LAN(AMENTO21 6°. teste regional Semente comercial
22 CULTIVAR Florada23 Semente Básica CAFl-;Z4L24 Florada
25 Ia. Produção
26 LANÇAMENTO27 Semente comercial
28 Florada29 I11 colheita30 CAFI~Z4L
5. 5. Seleção em gerações precocesÉ usado em espécies autógamas e alógamas
selecionando indivíduos. linhas ou populações no
estádio inicial de endogamia.
O objetivo é eliminar linhas ou populações que
não apresentem mérito para posterior seleção.
Em 1941. Immer indicou que testes repetidos de
populações segregantes 1'2 ou 1'3 forneceria o
desempenho produtivo médio dos diferentes
cruzamentos. Nilson-Ehle indicou o uso deste
método para identificar plantas 1'2 superiores
através de teste de progênies com repetições e
então. aval iar e selecionar indivíduos
homozigotos dentro de linhas derivadas de 1'2.
Este procedimento pode identificar progênies
Homozigotas em gerações precoces como
1'3
reduzindo o tempo gasto no desenvolvimento de
cultivares (QUADRO I). embora o custo
da condução dos ensaios de materiais ainda
segregantes aumente muito. Em plantas perenes
em que o tempo é fator limitante. é
compensador. pois. indivíduos. linhas ou
populações inferiores são descartadas cedo no
processo de endogamia.
características de alta herdabi Iidade como
precocidade de maturação dos frutos podem ser
selecionadas em gerações 1'2. Características
altamente correlacionadas com a produção como
vigor vegetativo. ramificação. volume da copa.
tamanho dos gràos podem ser selecionadas com
sucesso.
Características de baixa herdabilidade como a
produção. podem ser selecionadas também na

F2, desde que a varibilidade genética dogermoplasma seja alta e o tamanho populacionale o controle ambiental seja rigoroso.
5. 6. Métodos que não avaliam a produção emgerações iniciaisMétodos de melhoramento como descendênciade semente única ou "Single-Seed-Descent"(SSO). população e retrocruzamento, podemlevar a rápido avanço nas gerações segregantesem direção a homozigose ou a ter altasemelhança ao parental recorrente. Permitem oavanço de trabalhos de melhoramento em certaetapa sem realizar a avaliação da produtividade.que são operações caras e demoradas. O métodoSSO será usada como exemplo para ilustrar avantagem de se usar a alternativa de avançar asgerações precoces sem selecionar para aprodução.Ü método genealógico era popular parapopulação endógama de plantas anuais quandosomente uma geração era conduzida por ano.Atualmente. tem sido desbancado por outrosmétodos como o SSO conduzindo duas a trêsgerações num ano usando casa de vegetação.estufa ou viveiro. Uma comparação dos métodosgenealógico. genealógico com seleção emgerações precoces com e sem seleção antecipadapara produtividade. SSO e retrocruzamentoaplicado ao cafeeiro é apresentada no Quadro 1.
5.7. Uso de técnicas biotecnológicasAbrem novas perspectivas para o melhoramentogenético de espécies perenes como o cafeeiro. asquais tem o problema do fator longo tempo gastopara o desenvolvimento de cultivares (ROCA &MROGINSKI, 1991: SÓNOHAL et al., 1981:SÓNOHAL & LÜH. 1988).
5.7.1. Cultura de embriões e meristemas. Astécnicas como. cultura de embriões e cultura demeristema já tem aplicação prática na pesquisavisando aumentar a eficiência dos programas demelhoramento.
5.7.2. Cultura de micrósporos e de óvulos. Acultura de micrósporos ou de óvulos. levando aprodução de uma grande quantidade de plantashaplóides a serem dupl icados e obtidas aslinhagens duplo-haplóides completamentehomozigotas já na geração F2 apresentam umgrande potencial.
5.7.3. Embriogênese somática a partir defolhas. A embriogênese somática direta ou
198
indireta. começa a ser aplicado na redução dotempo gasto no desenvolvimento de cultivares.especialmente no uso de cultivares do tipo clonede indivíduos híbridos simples ou híbridosduplos. agregando várias característicasagronômicas num genótipo com heterose.
5.7.4. Transformação genética ou transgênese.A transformação genética de cultivares porincorporação de ONA de outras espécies.gêneros ou mesmo de espécies muito distantes.introduzidos por vetores como Agrobacterium eoutros e por biobalística por bombardeamento deONA de genes que deseja incorporar no cafeeiroestá to,nando realidade a transferência de genesnão presentes no gênero Coffea, como porexemplo genes de resistência a broca dos frutosHypotenemus hampei e genes de resistência a umherbicida de ação total. muito importante emcultivo intensivo como em plantio emespaçamentos adensados. super-adensados ehiper-adensados. Características de amplo usonas cultivares como os genes da arquitetura daplanta (Ex.: cornpata, erecta e cônica). machoesterilidade. maturação precoce e tardia.resistência a nematóide etc poderão serincorporados rapidamente num futuro próximoem cultivares comerciais de alta produtividade equalidade.
5.7.5. Seleção assistida por marcadores deDNA.O uso de marcadores moleculares para seleçãoasssistida sem dúvida poderá reduzir muito otempo gasto no desenvolvimento de cultivares decafé de várias formas. A seleção na fase juvenilreal izada em laboratório para característicasqualitativas como a arquitetura da planta. cor dofruto. % de frutos com lojas vazias. resistência aparasitos dos frutos e outros determinados nafase juveni I trarão grandes benefícios naeficiência de um programa de melhoramento.Outras técnicas corno a hibridação assexuada viafusão de protoplastos, seleção "in vitro ". "ONAfingerprinting" e a criopreservação, entre outros.auxiliarão muito no aumento da eficiênciaseletiva. com tendência para se incorporardefinitivamente corno ferramenta rotineira dernelhoristas do futuro próximo. O melhoramentogenético que começou de forma empíricaevoluiu para o melhoramento científico. estácaminhando em direção ao melhoramentogenético biotecnológico ou a engenhariagenética.
J
I

5. 6. Uso de cultivares duplo-haplóidesPara as espécies autógamas como o café arábico.onde o produto comercial é um grão. o maiseconômico para os agricultores é o uso decultivares do tipo linhagem propagadas porsementes. O tempo dispendido para odesenvolvimento de linhagens pelo método usualé longo, geralmente em torno de 25 anos(MEDINA et ai .. 1984) e este prazo pode serreduzido para algo como seis anos.Outro tipo de cultivar em que se usa as linhagens.são as cultivares do tipo multi linha. Usado paradar maior estabilidade fenotípica a cultivar. pelamistura mecânica de diversas linhagens emproporção conhecida. É usado também parareduzir o tempo gasto no desenvolvimento decultivares com resistência a diversas raçasfisiológicas de patógenos bem como para darmaior durabilidade a resistência frente àsmutações dos patógenos.Necessita-se normalmente de muito tempo parase chegar a uma linhagem homozigota. Parareduzir este tempo, existe a alternativa de sechegar a linhagens a partir de plantas haplóidesduplicadas (N ITZSCHE & WENZEL.1977).Nos procedimentos usuais de melhoramento. aobtenção de cultivares do tipo linhagem éconseguida através de sucessivas gerações deauto-fecundações. Duplo-haplóides são plantashomozigotas obtidas pela duplicação doscromossomas de células ou tecidos haplóides. Aprodução de linhas diplóides homozigotas porduplicação do complemento cromossômico deindivíduos haplóides é um método alternativo dedesenvolvimento de cultivares num prazo muitocurto. Existem diversos métodos de obtenção deduplo-haplóides, convencionais oubiotecnológicos, e que podem ser utilizados emplantas perenes.Existe a possibilidade de utilizar plantashaplóides obtidas por métodos convencionais.naturais ou induzidas (método bulbosum) noprocesso de obtenção de linhagens homozigotasem cafeeiro.
5.7. Uso da heterose de cultivares híbridasCultivares híbridas são utilizadas em diversasculturas e visam aproveitar a heterosenormalmente observada nas híbridações (ver
capítulo sobre o assunto em outra parte destelivro). Em plantas perenes. o custo da sementehíbrida pode ser amortizado por vários anos deprodução. Se um híbrido. por exemplo. produzir20 % a mais por ano do que uma cultivar do tipolinhagem. em cinco anos ter-se-á um adicional de
199
produção equivalente à uma safra extra. Emplantas perenes pode ser inclusive dispensada aernasculação genética e a busca de autoincompatibilidade. A ernasculação e apolinização manual podem ser viáveis emdiversos cultivos. especialmente em plantas queproduzem muitas sementes por fruto como omaracujazeiro. cacaueiro e anonáceas. Emcafeeiro em que cada flor dá apenas duassementes em média. pode se lançar mão degameticidas e ernasculação térmica. A utilizaçãode híbridos pode. também. representar umagrande economia de tempo. permitindo que omelhorista possa testar diversas combinações delinhagens. selecionar as mais promissoras elançá-Ias como cultivares. Pode-se. inclusivetrabalhar com diferentes tipos de híbridos.
a)Cultivares do tipo híbrido FI. São cultivaresnormalmente obtidas pela híbridação delinhagens homozigóticas ou heterozigóticas,
sendo mais viável para plantas perenes queproduzem muitas sementes por fruto. Adisponibilidade de genes de macho-esterilidade eauto-incompatibilidade. bem como de técnicasquímicas. térmicas e hídricas de indução domacho-esterilidade podem facilitar ainda mais ouso desta alternativa. especialmente em plantasque produzem poucas sementes por fruto.
b)Cultivares do tipo híbrido F2. Caso a vendade sementes híbridas F1 seja antieconômica,pode-se. em alguns casos. cultivarcomercialmente as plantas híbridas F2 .especialmente para plantas perenes que produzpoucas sementes por fruto e que o produto écomercializado a granel (Ex. café). Neste casopode-se aproveitar cerca de metade da heterosedo FI . Algumas características corno a épocade colheita e o tamanho da planta devem seruniformes. porém em alguns cultivos pode-seadm iti r variabi Iidade para esta e outrascaracterísticas agronômicas.
c) Cultivar do tipo sintético. Procura preservar
o vigor de híbrido sem ter que obter as sementeshíbridas toda vez que for plantar. Urnapopulação base de combinações híbridas comheterose de plantas alógamas são usadas paraformar o lote de matrizes para que as sementestiradas a partir daí permitam alcançarprodutividades maiores que as cultivares do tipolinhagem ou população. Pode ser usado em caférobusta (Coffeu canephora).

5. 8. Uso de cultivares do tipo cloneSão cultivares propagadas vegetativamente, ouseja, todos os descendentes são geneticamenteiguais à planta que deu origem às mudas("planta-mãe"). A propagação vegetativa pormeios convencionais como a estaquia e a enxertiaou não convencionais como a micropropagação'in vitro' permitem obter rapidamente cultivaressuperiores. A propagação vegetativa permite autilização de toda a variância genética disponívele a seleção normalmente recai sobre plantasaltamente heterozigóticas.As plantas de grande valor econômico comofrutíferas e ornamentais são no geral propagadasvegetativamente e constituem variedades do tipoclone com alta heterosigosidade.Nas plantas em que ainda não compensaeconomicamente a propagação vegetativa, aspesquisas podem ser direcionadas visandoaperfeiçoar as técnicas para reduzir os custosdeste tipo de propagação.A micropropagação tem sido utilizada emdiversos cultivos (cana-de-açúcar, bananeira.abacaxizeiro, morangueiro. eucalipto. etc) eapresenta como vantagens, além da rápidapropagação, a possibilidade da obtenção demateriais livres de patógenos, especialmentevírus. Por outro lado, durante o processo demicropropagação "in vitro" existe a possibilidadede serem encontradas variações genéticasherdáveis. economicamente importantes nasprogênies (variação somaclonal).A utilização de cultivares do tipo clone ja erealidade no Brasi I através do uso de estaquiaconvencional de café conillon de Coffeacanephora. Embora seja de custo quadruplicado.a maior produtividade e uniformidade aliado aoutras características de cultivares clonados, temmantido o interesse de muitos cafeicultores aimplantarem lavouras comerciais.O uso de cultivares clonados a partir de folhas degenótipos superiores híbridos por embriogênesesomática usando bioreatores está indicandosuficiente estabilidade genética e viabilidadeeconômica. Com isso, vislumbra a possibilidadede aplicação imediata da biotecnologia nomelhoramento genético e na cafeicultura.
5.9. Lançamento antecipado de cultivaresEm culturas perenes. em que se necessita devários anos para avançar uma geração de seleção,é interessante adotar um esquema apropriado deantecipação do lançamento de cultivares paraaumentar a eficiência do programa demelhoramento. Para isso. é necessário gastar
200
mais em termos de área experimental, avaliaçãoe multiplicação de um número maior deseleções, bem como a instalação de uma áreamaior e de um número maior de cultivarespotenciais num campo de semente básica paraganhar tempo. Em café arabica é possívelganhar \O anos pela condução simultânea deavaliação regional, formação de campo desemente básica e campo de semente fiscalizada.desde que protegida a origem por marcadoresgenéticos moleculares ("DNA fingerprinting")de cultivares potenciais.
a) Avaliação regional antecipada de seleções.Todos os materiais promissores devem sertestados regionalmente, simultâneo a seleçãofinal, a fim de ganhar uma etapa do processototal desde o cruzamento até o plantio comercial.Consiste em avaliar um número bem maior deprogênies promissoras para incluir dentre elas ascinco primeiras seleções que poderão se tornarcultivares, ganhando-se algo como quatro anos(SERA, 1987). Por exemplo, se pretendeenglobar no ensaio regional as 5 primeiras e ograu de precisão na discriminação antecipada deseleções superiores for de apenas 15% enecessita de mais 4 anos para ter maiorsegurança na avaliação do potencial produtivofuturo com h2 = 0,35. então, para ganhar estesquatro anos, devemos testar pelo menos 35seleções promissoras ao invés de 15 seleções.
b) Instalação antecipada de campo de sementebásica. Para ganhar pelo menos mais 4 anos.devemos iniciar concomitante a instalação doensaio regional, campo de semente básicaenglobando estes genótipos promissores. Asparcelas das seleções não confirmadas para arecomendação deverão ser el im inadas ecompletadas com materiais recomendados e.após mais dois anos. passará a fornecer sementepara lavouras comerciais junto com ofornecimento de sementes fiscalizadas.
c) Lançamento antecipado de cultivares. Olançamento antecipado de cultivares emgerações precoces. com certa variabi Iidade paracaracterísticas sem muita importânciaagronômica e para produtividade. tamanho dosfruto ou grão. perrn ite lançar versões inacabadascom características de alto valor agronômicocom 4 anos de antecedência como por exemplopara pragas e doenças chaves, qualidade, etc.tem alto retorno econômico-social para osagricultores. O prosseguimento da seleção

permite o lançamento posterior de versõesmelhoradas.
6. CONCLUSÕES
Através da combinação de um conjunto demétodos. técnicas e procedimentos demel horamento genético é possível ganhosconsideráveis na eficiência de um programa demelhoramento de cafeeiro. planta perene onde ofator tempo é a principal limitante para se ter umganho genético razoável por ano até a obtenção eo plantio de cultivares melhoradas. As soluçõesadotadas para outras culturas diversas da que seestá trabalhando podem servir para dar umagrande contribuição tanto em termos deeconomia de recursos como de tempo. tornando otrabalho menos cansativo e mais realizador.Um planejamento estratégico do programa demelhoramento de cafeeiro é vital e decisivo navida do melhorista. É necessário além de umprofundo conhecimento de todos os aspectos dacultura que trabalha. desde a origem e evoluçãoda espécie até a competitividade comercial. paraevitar que o trabalho de uma vida toda resulte empoucos resultados.O êrro de estratégia de um melhorista de café émuito frustrante. pois não tem muito mais tempopara reiniciar ou melhorar o programa. porquefrequentemente gasta-se mais de 20 anos a partirdo cruzamento para se chegar a uma cultivar desucesso. O diagnóstico da cultura com seusproblemas e potencialidades e o conhecimentoprático da cultura para intervenção. mais certeirapossível. atacando prioridades maiores dentro dasaltas prioridades é fundamental. O fator tempopara o desenvolvimento de cultivares pode serminimizado muito. reduzindo de 20 para até 10anos mudando-se alguns procedimentos simplesna estratégia de desenvolvimento de cultivares ede métodos de melhoramento e agregando novasferramentas como as técnicas biotecnológicas,informática e bioestatística. O ganho genético porano pode dar um salto em função da maiordisponibilidade de recursos humanos. materiais.tempo e espaço.
7. REFERÊNCIAS PARA CONSlILTA:
ANTUNES FILHO & CARVALHO. A.Melhoramento do cafeeiro: XII. Variabilidade emIinhas puras de café. Bragantia, Campinas. 16:197-213. 1957.ARAÚJO NETO. K. de. Avaliação preliminarde progênies de Coffea arabica pela estimativa
201
simultânea de vigor vegetativo e carga pendente.In: CONGRESSO BRASILEIRO DEPESQUISAS CAFEEIRAS. 6. Ribeirão Preto.1978. Resumos. Rio de Janeiro. Ministério daIndústria e Comércio -IBC, 1978. p.1 10-113.CANNELL, M. G. R. Crop physiologicalaspects of coffee bean yield. Journal of coffeeReserch, Karnataka, ~(1/2):7-20. 1975.CANNELL. M. G. R. Physiology ofthe coffeecrop. l!l: CLlFFORD, M. D. & WILLSON, K.C. (ed). Coffee, botany, biocemistry andproduction of beans and beverage. Westport,AVI. 1985. p.251-83.CARVALHO. A. Melhoramento do cafeeiro:VI. Estudo e interpretação para fins de seleçãode produções individuais na variedade Bourbon.Bragantia, Campinas. 1l(6): 179-200. abr.-jun ..1952.CARVALHO. A. Evolução nos cultivos de café.o agronômico. Campinas. J7(1):7-11. 1985.CARVALHO. A & MÔNACO. L. CMelhoramento do cafeeiro: XXXI. Ensaio depopulacões F2 de híbridos entre cultivares deCoffea arabica. Bragantia, Campinas 26(5):7992. 1967.CARVALHO. A.; MÔNACO. L. C;ANTUNES-FILHO. H. Melhoramento docafeeiro: XV. Variabilidade observada emprogênies de café. Bragantia. Campinas. ~(26):374-86. 1959.CARVALHO. A.; MÔNACO. L. C;CAMPANA. M. P. Melhoramento do cafeeiro:XXVII. Ensaio de seleções regionais de Jaú.Bragantia, Campinas, ~(13):129-42. 1964.CARVALHO. A.; MÔNACO. L. C; FAZUOLl.L. C Melhoramento do cafeeiro: XL. Estudosde progênies e híbridos de café 'Catuaí'.
Bragantia, Campinas, ~(22):202-16. 1979.CARVALHO. A.; MÔNACO. L. C;FAZUOLl. L. C; COSTA. W. M. da;MEDINA. H. P. Variabilidade na produção emprogentes de cafeeiro . Mundo Novo'.Bragantia, Campinas. :L(32):509-17. 1984.CARVALHO. A.; SCARANARI. H. J.;
ANRUNES-FILHO. H.; MÔNACO. L. C.Melhoramento do cafeeiro: XX11. Resultadosobtidos no ensaio de seleções regionais deCampinas. Bragantia. Campinas. 20 (30):71 140.1961.CASTILLO-ZAPATA, 1. & MORENO-RUIZ,G. Selecion de cruzamentos derivados dei"Híbrido de Timor" en la obtención devariedades mejoradas de café para Colômbia.Cenicafé. Chinchiná, abr.l ian. 37-53 1981.CASTILLO-ZAPATA. J. & QUICENO. G.

Estudio de la producción de seis variedadescomerciales de café. Cenicafé, Chinchiná, 12(1): 18-39. 1968.CHARRIER, A. & BERTHAUD, J. Principies
and methods in coffee plant breeding: Coffeacanephora Pierre. In: CLARKE, R. J. &MACRAE, R. (ed). Coffee Volume 4.' Agronomy.London, Elsevier, 1988. p. 167-198.DHAUWAL, T. S. Correlation between yieldand morphological characters in Puerto Rican andColumnaris varieties of Coffea arabica L..Journal of the Agricultural University of PuertoRico. 52:29-37, 1968.EBERHART, S. A. & RUSSEL, W. A. Stabilityparameters for comparing varieties. CropScience. Madison, §: 36-40, jan./fev. 1966.FAZUOU, L. e. Avaliação de progênies de café'Mundo Novo'( Coffea arabica L.). Piracicaba,
1977. 146 pp (Mestrado - Escola Superior deAgricultura "Luiz de Queiroz" USP).FONSECA. M. A. S. de; ARAÚJO, P. F. e. de;
PEDROSO, I. A. Retorno social aosinvestimentos em pesquisa na cultura do café.São Paulo, BADESP, 1978. 14 p.GIFFORD, M. R. & EVANS, L. T.
Photosynthesis. carbon paritioning, and yield.Ann.Rev. Plant. Physiol. 32:485-509. 1981.MATIELLO, J. B.; SOUZA. D. D. de;ASSUMPÇÃO, J. M.; Á VILES. D. P. Análise dociclo bienal da produção. In: CONGRESSO
BRASILEIRO DE PESQUISA CAFEE1RAS.12., Caxambu, 1985. Resumos. Rio de Janeiro,Ministério da Indústria e Comércio - me. 1985.p.217-219.
MA WARDI. S. & HARTOBUTOYO, S.
Correlation study of yield and branchcharacteristics of robusta coffee (Coffeacanephora Pierre varo rohusta Cheval.) on F I
hybrid population. Menara Perkebunan.Jernber, 49 (5) : 115-20, 1981.
MAWARDI. S.; ISWANTO. A.;HARTOBUTOYO, S. Selection of the F2population of arabica coffee: 11. A preliminary
study for individual selection by applyingselection indexes. Menara Perkebunan, Jernber.
21 (5), 128-32. 1983.MEDINA. H. P.; CARVALHO, A.; SÓNDAHL.M. R.; FAZUOU, L. c., COSTA, W. M. da.
Coffee breeding and related evolutionary aspects.In: JANICK, J .. ed. Plant hreeding reviews.Westport, A VI. 1984. p. 157-94.
MENDES, L. O. T. Poliploidização daseringueira: um novo teste para determinação da
capacidade de produção de seringueiras jovens.Polimeros I(I): 22-30. 1971.
202
MÔNACO, L. e. Efeito das lojas vazias sobreo rendimento de café 'Mundo Novo', Bragantia.Campinas, 12: 179-200, 1952.
MÔNACO:- L. c., MEDINA P H. P.;SÓNDAHL, M. R.; UMA, M. M. A. DE ..Efeito de dias longos no crescimento etlorescimento de cultivares de café. Bragantia,37 (4): 25-30,1978.MÔNACO, L. c. SCAU. M. H.; CARVALHO,
A.; FAZUOLJ, L. e. Variabilidade no sistemaradicular de genótipos de café. l!:!: REUNIÃOANUAL DA SBPC, 25., Ilha do
Fundão/GUANABARA, 1973. Resumos. Ilha do
Fundão, SBPC, 1973. p. 247.MORENO-RUIZ. G.; CASTILLO-ZAPATTA,
J.; OROZCO-GALLEGO, L. Estabilidade de laproduccion de progenies de cruzamientos de'Caturra' por 'H íbrido de Timor'. Cenicafé,Chinchaná, out./dez.:79-90, 1984.NITZSCHE. W. & WENZEL. G. Haploids inplant breeding. In: Advances in Plant Breeding
(Suplements to Journal of Plant Breeding).Berlin, Verlag. 1977. 101 p.
RAMOS, L. e. da S. Desenvolvimento daplântulas de quatro cultivares de café. Bragantia,Campinas, 39:215-8, 1980.RAMOS, L. c., UMA, M. M. ALVES DE &CAR VALHO, A. Crescimento do sistemaradicular e da parte aérea em plantas jovens decafeeiros. Bragantia. Campinas, 41 :93-99, 1982
ROCA. W. M. & MROGINSKI, L. A. Cultivode Tejidos en la Agricultura - Fundamentos yAplicaciones. C1AT, csu. Colômbia, 1991. 969
pp.ROCHA. T. R.; CARVALHO, A.; FAZUOU, L.e. Melhoramento do cafeeiro: XXXVIII.Observações sobre progênies do cultivar MundoNovo de Coffea arahica na EstaçãoExperimental de Mococa. Bragantiu,Campinas. 39 (15): 148-60. 1980.
SERA. T. Zoneamento varietal de café noParaná. In: ANAIS DO ... Congr. Bras. Pesq.Cafeeiras. 1979. Ribeirão Preto.
SERA, T. Estimação dos componentes da
variância e do coeficiente de determinaçãogenotípica da produção de grãos de café (Coffeaarabica L.). Piracicaba. 1980. 62 pp. (Mestrado Escola Superior de Agricultura "Luiz deQueiroz'Yl.Jâ P).
SERA. T. Seleção precoce para produção no
melhoramento genético de plantas perenes.Piracicaba, 1984. 115 pp. (Monografia da
disciplina Tópicos Especiais de Genética Escola Superior de Agricultura "Luiz deQueiroz" / USP).

I.II
SERA. T. Possibilidade de emprego de seleçãonas colheitas iniciais de café tCoffea arahica L.cv. Acaiá). Piracicaba. 1987. 147 p. (Doutorado
Escola Superior de Agricultura "Luiz deQueiroz" / USP).SONDAHL.. M. R.; MÔNACO. L. c.,CARVALHO A.; FAZUOLl. L. C. Efeito dedias longos em progênies de café (Coffeaarabica). tratados com benzi ladenina egiberelina. Ciência e Cultura. 24 (sup.):336.
'1972.SONDAHL.. M. R.; MüNACO. L. c., SHARP.W. R. In vitro methods applied to coffee. ,ir!:THORPE. T. (Ed.). Plant tissue culture. methods and applications in agriculture. NewYork, Academ ic, 1981. p. 325-348.SONDAHL. M. R. & LOH. W. H. -T. Coffee
biotechnology. In: CLARKE. R. J. &MACRAE. R, (ed). Coffee Volume -/: Agronomy.London. Elsevier, 1988. p. 236-262.SRINIVASAN. C. S. Association of some
vegetativa characters with initial fruit yield incoffee (Coffea arabica L.). Journal of CoffeeResearch, Karnataka, ,UL(2):21-7. 1980.
SRINIVASAN. C. S.' Pre-selection for yield incoffee. Indian .Iournal of Genetics, New Delhi.42 ( I) : 15-9. 1982.
SRINIVASAN. C. S. & SUBBALAKSHMI . V.A biometrical study of yield variation in somecoffee progenies. .fournal oi Coffee Research.Karnataka. _I_1(2):26-34. 1981.SRINIVASAN. C. S. & V1SHVESHWARA. S.Variability and breeding value ofsome characters
related to yield in a world collection of arabicacoffee. Indian Coffee. Bangalore, 15 (5): 119-28.1981.
203
STEVENS. W. L. Análise estatística do ensaiode variedades de café. Bragantia. Campinas. 2(5-8):103-123.1949.TORRES. R. índice de Seleção. Piracicaba.1985. 77 p. (Monografia - Escola Superior deAgricultura "Luiz de Queiroz"/USP).VALENCIA A.. G. Relation entre el indice de'area foliar 'y la productividad deI cateto.
Cenicafé. Chinchiná. out./dez:79-89. 1973.VAN DER VOSSEN. H. A. M. Coffee
selection and breeding. In : CLlFFORD. M. N,& WILLSON. K. C. ed.. Coffee; Botany,biochemistry and production oi beans andbeverage. Westport, A VI. 1985. p. 48-96.WALYARO. D. J. & VAN DER VOSSEN. H.
A. M. Early determination of yield potential in'arabica coffee by applying index seíection.Euphytica, Wageningen. 28:465-472. 1979.WELLENSIEK. S. J., Shortening the breedingcycle, Euphytica I 1:5-10. 1962.WALYARO. D. J. ' Considerations in breeding
for improved yield and quality in arabica coffee, (Coffea arahica L.). Wageningen. 1983. 119 p.(Doutorado - Agriculltural University).
WALYARO. D. J. & VOSSEN. H. A. M. VanDer. Early determination of yield potencial inarabica coffee by applying index selection.:Euphvtica. Wagen ingen, 28 :465-72. 1979.

205
DESENVOLVIMENTO DE CAFEEIROS RESISTENTES A INSETOS.
Oliveiro Guerreiro Filho.
Instituto Agronômico deCampinas - Centro de Café e Plantas TropicaisCP 28 - CEP 13.001-970 - Campinas - SP - Brasil
INTRODUÇÃO
A produção de café no Brasil no ano agrícola1998,1999 atingiu cerca de 34 milhões de sacas.sendo 29.5 milhões de sacas oriundas decultivares de (', arabica. Como todos estescultivares são suscetíveis à Pcrileucopteracoffeclla. a principal praga da cultura no país. asperdas de produção na cultura são altamentesignificativas. O Instituto Agronômico deCampinas. vêm há alguns anos desenvolvendocultivares de (' arabica que aliem níveiselevados de produtividade. resistência a esteinseto praga c boa qualidade da bebida. Asprincipais etapas desenvolvidas neste programa.são descritas a seguir.
Identificação de fontes de resistência.Como Coffea arabica é a uruca especietetraplóide (2n 44 cromossomos) do gênero('lIlli:a. a busca de fontes de resistência ao insetoteve início em indivíduos da espécie. sendo quenenhum cultivar. introdução ou forma botânica deC. arubica revelou-se resistente ao bicho mineiro.No entanto. diferentes níveis de resistência foramencontrados em espécies diplóides (2n ")")cromossomos) do gênero ('II/A'a, O mais altonível de resistência ao inseto é verificado em ('stcnophvlla, Nesta espécie. as lagartas morremimediatamente após a eclosão. sendo as lesões nasfolhas dificilmente visíveis a olho nu. Outrasespécies como (' /1 bcrtca. (' cugcuioidc:s. ('kapakat«. C'. salvatrix e (', raCL'IIIO,la apresentamtambém níveis elevados de resistência. masinvariavelmente segregam. em maior ou menorescala. para esta característica. I·.studos real izadospor Guerreiro-Filho (1994). evidenciaramtambém que "irias espécies Malgachcs como ('tctravona, (' bcrtrundi. c 111 i1/1I11 i. (' resinosa.(' tstrununac, (' brntnicri, (' dohcl/IJ/J!I vila e ('jurafú.I!al/L'lISi.l. conhecidas pela ausência decafeína em suas sementes. podem também seremcons ideradas como res istcntes ao bicho mine i1'0.
Apesar do maior nivel de resistência apresentadopor C ,1/CI/O/J/I1'1/u. barreiras gcncticas bastante
importantes. di ficultarn a hihridação destaespécies com C. arábica. Resultados semelhantessão observados com a uti Iização das ~.. spécicsMalgaches. Assim. no programa demelhoramento do cafeeiro \ isando resistência aobicho minei rodesenvolvido pelo Instituto Agronômico deCampinas. (' raCL'I1IIi.l'U foi escolhida C0l110
parcntal doador de genes de resrstôuciu. pnisapresenta intenso florescimento (lan alho S:
Monaco. 1(67) o que facilita as hibridaçôcs comC. arabica. é resistente à seca (Medina-Filho <.'1
al., 1(77) e apresenta maturação precoce dosfrutos (Costc, 1955: Levy ('/ al.. IlJR4). Alémdessas características de inestimável \ aloragronômico. indivíduos pertencentes à segundageração de reirocruzamentos naturais (RC~) entre
(' arabu:a e r ' rUCL'I/IIJ.I'a foram idcnti ficados porMedina-Filho el al. (1977) como sendoresistentes à I' cliffed/u. Fssa descoberta incorreuem uma considerável economia de tempo L'
recursos no programa de seleção. A transferência
da resistência para variedades comerciaissuscetíveis de (' arubica teve início através doscruzamentos dirigidos realizados entre es\cscultiv ares e os indivíduos RC") C I I9S-:,-6-1 c
C 1195-5-6-2. classificados COIllO resistentes.
Metodologia para seleção de plantasresisten tesoA seleção de indiv iduos resistentes L; realizada.em uma primeira etapa. no viveiro, descartandose as plantas suscetíveis de cada progênie obtidapor cruzamentos dirigidos ou uurofccunduçõcs.Posteriormente. as plantas rc- istcntcs ou àquelassuscetíveis ocasioualmcntc nào atacadas. ~ilO
avaliadas individualmente em laboratório
utiIizando-sc a mciodolog ia descrita porGuerreiro-Filho (19lJ4) que consiste na utilivaçaode Ulll suporte onde o pce iolo de folhasdestacadas é mantido hidratado durante o penudode exposição aos insetos. Cerca de -lO plantaspodem ser in testadas diariamente .uril\ és destamcrodolog ia. Discos de folhus são retirado» ilpartir dos locais onde foram leita-, as posturas.sendo mantidos sob espuma em cimal'as úmida,por aprox imadarncntc I:' dias ate o

dcscnvolv intento completo das lesões notratamento controle. A avaliação da resistência éfeita pelo tipo de reação sendo as plantasclassificadas em 4 grupos: I lesões pontuais(Resistentes): 2 lesões til iformes(Moderadamente Resistentes): 3 '~ lesões grandese irregulares (Moderadamente Suscetíveis) e 4lesões grandes e arredondadas. característico daslesões verificadas em C. arabica (Suscetíveis).Apenas as plantas classificadas como resistentes.são utilizadas para instalação de ensaios. Nocampo. a avaliação da resistência ao bichomineiro é realizada através de uma escalaquantitativa (I a 10 pontos), atribuindo-se ospontos em função do nível de infestação dasplantas, Notas mais altas são atribuídas aosII1di\ iduos suscctivcis. tendo os cultiv arescomerciais como referência e notas mais baixas
as plantas com menor nível de ataque.
Criação artificial em insetário.:\ produção massa I de insetos utilizados nostestes de resistência em laboratório é realizadasegundo a metodologia descrita por Katiy ar S:lerrer (1968). em gaiolas de madeira cobertascom tecido. mantidas em insctário com
temperatura de 27 : 20 ('. fotopcriodo de 14h eumidade rclativ a de 70 ' 100
0 . Mudas jovens decafeeiros <uscct íveis. são introduzidas nas gaiolase substituidas na manhã seguinte. O número deplantas. assim como a permanência das mesmasnas gaiolas. varia com o tamanho da população deinsetos. Uma vez infestadas. as mudas sãomantidas em prateleiras nas mesmas condições doinscrário até que os insetos atinjam a tasc decrisálida. Nesta ocasião. as folhas contendocrisálidas são destacadas das plantas erccolocada .... no interior das gaiolas de criação,onde (lCOITe a emergência dos adultos e o
206
consequente fechamento do ciclo biológico dosinsetos. Nas gaiolas. os adultos são alimentadoscom sol ução de Sacarose a 10%. cond ição quesegundo Parra (1985). eleva sobremaneira onúmero de ovos por fêmea.
Expressão da resistência e número de genesenvolvidos.Estudos realizados por Medina-Filho et al(1977). revelaram que o tamanho ou a espessuradas folhas ou o nível de ploidia das plantas. nãotem qualquer relação com a resistência dasplantas. No entanto. Guerreiro-Filho cf al (1999).
evidenciou que em (' raccntosa. assim como noshíbridos entre ('. aruhu:a e r ' tacentosu as tolhas
mais velhas parecem ser mais scnsív eis que asfolhas mais jovens do,", dois primeiros pares.Analises realizadas em progênies obtidas porautofecundacão e por retrocruzarnentoscv idcncinram que a resistência à P ('uttee//a é!lO' crnada pela expressão de dois genescom plcrncntarcs e dom inantes. denorn inados 1.11I1e /.111::. Recentemente, alguns indivíduosuiplóidcs foram obtidos pela hihridaçâo entre ~IS
cspcc res (' arabn:a e C' stcnophvlla. A
resistência desta espccrc ao bicho mineiro,di lcrcntcmcntc de (' ruccmosu. parece ser denatureza recessiva.
Método de melhoramento.O mctodo de melhoramento empregado nodcsenvolv intento de cultivares COI11 resistência aoinseto é o de retrocruzamcntos sucessiv osassociado ao método gencalogico. sendo que na,di\ crsas gerações (Figura I l. são realizadassclcçõc-, para resistência. produção.características agronômicas de interesse equalidade da bebida.
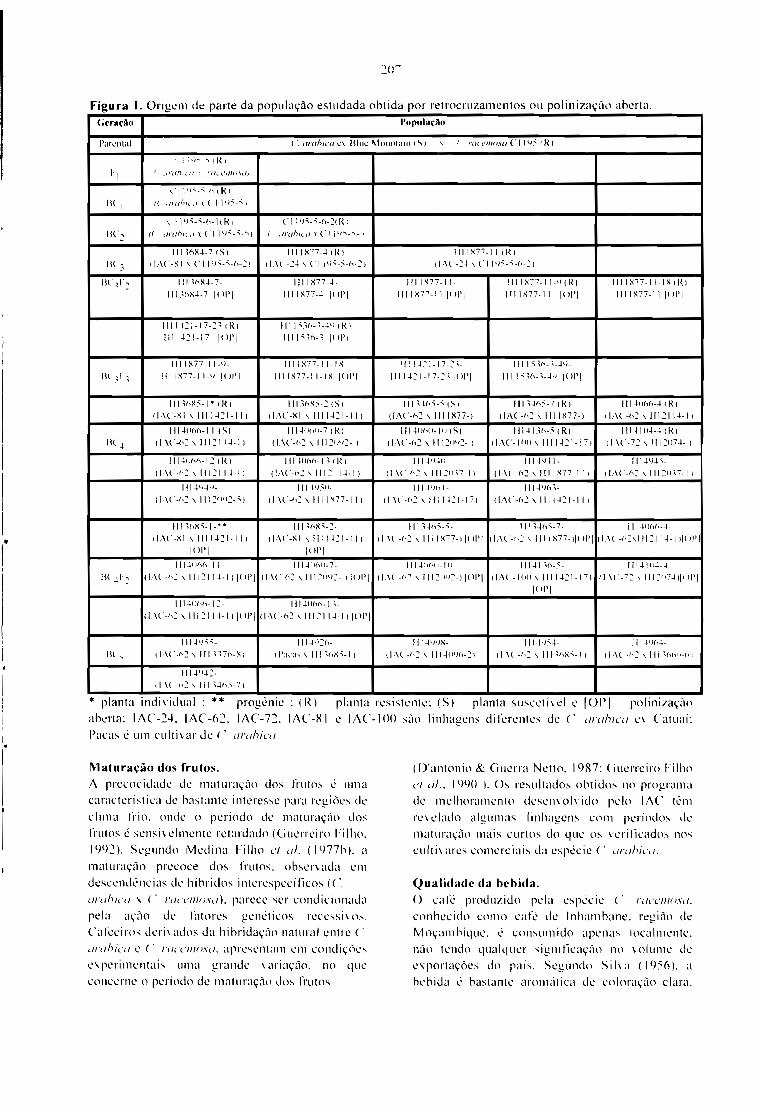
Figura I. Origem de parte da população estudada obtida por retrocruzarnentos ou polinizaçáo aberta.(;Haçj\o I'opulaçj\fl
Parcm.u 1 arabu u,:» Ill"c "10""1'011 I" I \ : 1"/1/ ("UOso (' I ll~:, I J{ I
] i ~/" ." R ,
11 .In,,'l.~,.· . "(I~ t.'",,, \(f/
l': ;q";'." 11 t R I
IH I Ii ,lI"ohltcl \ l 11\1:'-" ,
" 1I'I'·'·h-11 R, C 11l)'·'·I<~1 R:IH, Ii orotnc » v ("11 ~h-".h) .. .nabu « v C! ]q",-",-,.-
1111h~-1·7 IS) 11I1~77·-1 IR I 1I11~77·11 IRIIH 1 II.\('-X I \ ('1 ll)';_",_h_~, II.\C·e-1 \ ClI'I'·'·h';1 (I·\l'-~-l \ CIIl)~-",-h-:)
IlC 11', 1I1 1h~-1·:· 1111 P7--1- 11l1~77·II- 1IIIX77·J 1-'1 IRI IIIIX77·II-IX IR)-1II,1hX-1·7 Illi'l IIIIX77·-1 101' I IIIIXP·11 101' i IIIIX77·11 IllI'l 1IIIX77·11 1111'1
1I11-1;1·17·c' IRI 1111 'v'· '.-1 0 1R'111 l-1e1·1: l°l'j 1I11"h·' II)I'!
1IIIX77·II·'i. IIIIX7"'-II-I,~· 1I11-1cl·17·>· 111 I' ,h· '.-1 'i.III -"-' IIIIX77·11·" [111" IIIIX77·II·IX 1111' 1 1I11-1cl·17·c' IllI'l III ! ."".~(l- ".-Ill 101'1
II1 'h~'·I· I R I 1I11h~'·c ISI 1I1 1-1("" (," I 111,-1/>,·7 I K I 111-111/>1>.-1 1I{ I
II'\C-XI \ 11I1-1cl·11 I II.\('·X I \ 111 l-1e 1·1 I I II.'\l'·he \ 1111 X77·1 IIA(·h2 \ IIIIX77·1 lI.\( ·hc \ 111 c I 1-1·1 I
111-1I1I,h·11 (SI 111-1111,,,-7 IR I I I 1-1I'hll·ll! ISI 111-1 l.1h.,' I RI 111-1111-1 .• I RI
IlC-1 II.\l'·I'c \ 111;11-1·11 I IAl' ·/>e \ 111 Cll'ie· I II.\l' .hc \ 111 c"'ic· I II.\C·IIIII \ 1I11-1el·:" II\C-'c \ IIlcll7-J. I
111-1I1M·1 c I RI 111-1l1h/>·111 R I 111,1"-111. 111 -1'i-1 I . 1I1-1<}-1 <,II.\C·h: \ IIlcll-1·11 II.\l'·he \ IIlell-1·! I 11 \l'-/>c \ 111 ell,17·I) 1I.,\(·he\ 1111X77 11I (1:\(' _11: \ 111211",;. ~ I
111-1<}-1"· 111-1"'''' 111 J"h I. 111-1')h,.
li·\C-h: \ 111:lll~:-';1 11 «.«: \ 1I11,~77·111 11 «.«: \ 1111-1;1·171 II.\C·/>~ \ lIi 1-121·11,
1I11h~'-I·" 1I1~(,X"-~- 11I'-1h'·'- 11' ~ ..U""·,7. 1I1Jllh(,•• ,
1I,,\l'·XI \ 111 l-1e 1- 1II IIA{'·XI \ 111 1,1; 1·1 1I I I \l' -t-; \ 11 i I ~"' - I II 11'1 11.\l'·,'~ -, 1111 X77·111 11'1 11.\( .h:\lIlcl 1-1·1 11111'1
1"1' I 1111'1
111-1"('(" I I· 111-1 111>11.7. III ~11(1(1_111. 111 -1 I '('-,'- 111-1 I 11-1--l
I!C-11 c \I,'\{ -he \ lIicl 1-1.1111)1'1 II.-\l·he \ 111 cll":, IIIIPI 11.-\( _h; \ Illcll"e-lllli'l 11,,\(·1011 \ 1I11-1el·I'1 IIM·ce,III:I17JIII'P!
!oI' I
111-10hh·1 c· 111-1l1hh·1 ;.
11\l'·he \ IIlell-1·1 I 101'1 II\l'·('c \ 111:11-1-/1101'1
IIIJ'J"· 11 (-Il)~() .. /11-1"",· 1I1-1'J'-1· /11,)%-1·IH -; ll·\l'4h~ \ 111.~'7h-Si (P:lr;l,\ \ 11\ ,6,'-:"'-1 ) ti,\( -h'2 \ 111-J111}(~_~ I 11\( .h: \ I11 ;hX'.11 11,\( ,(,~ \ III ;(,(,1,_11:
111-1'l-Jc·ti \1 ·I>e \ 111 '-1h'.71
I.* planta individual : **aberta: I;\C-24. I;\C -ó2.Paras é um cultivar de ('
progeruc : (R) planta resistente: (S) planta suscctiv el e IO!'1 polinizaçãoI;\C-72. I;\C-XI c [;\C-I()() sào linhagens diferente" de C arctbica l'l Catuui:
uruhicu
Maturação dos frutos.;\ precocidade de maturação dos frutos é umacaracteristica de bastante interesse para rcg iõcs declima frio. onde o período de maturação dosfruto" é scnsiv cirncntc retardado (Guerreiro Filho.1(92), Segundo Medina lilho c/ al, (I 1)77 h). amaturação precoce dos frutos. observada emdescendências de híbridos intcrcspccificos (('arabtc« \ r ' I"UCL'/lIIJ.I'o ). pa rccc ser cond icionadapela ação de fatores gcucticos rcccssr, \1'.
Cafeeiro>; derivados da hibridaçào natural entre ('.uobic« e (', I""(L'1/1 IJSU. aprcscntum em condiçõc-,ex pcri mentais uma grande I .uiaçào. no qUL'couccrnc o período de maturação dos frutos
(D'antonio & Guerra Netto. 19X7: Guerreiro lilhoc/ a! .. Il)l)() l. Os resultados obtidos no programade melhoramento desenvolvido pelo 1;\(' têmrevelado algumas linhagens com períodos dematuração mais curto" do que O" verificados no>;cu/til ares comerciais da espécie (' urubic«.
Qualidade da bebida.O café produzido pela cspccic (' r, h 'l'II/II,\ a.
conhecido como café de lnhambanc. regiãn deMoçumhiquc. é consumido apenas localmente,não tendo qualquer >;ignitica~'~10 no volume deexportações do país. Segundo Silla (19:'ó). abebida é bastante aromática de coloraçào clara.
Fi~ura 2. Produção acumulada em :; anos defrutos maduros expressa em kg, resistência a l I.vustatrix (escala de I a 4 pontos). resistência a Pcoffcel!« (escala de I a l Opontos ) e cstág io dematuração dos frutos (escala de I a :; pontos) dealgumas plantas em ensaios de progênies.localizados em Campinas.
Carvalho. A. & Monaco. l..c. Relações genéticasde espécies selecionadas de Coffcu. Cil.-Ilcia eCultura (Süo Paulo). \01. 19. n 1. 19h7. p.I:'I16:'.
Carvalho. A.. Teixeira. A.A .. lazuoli. I..c. S:Gucrreiro l-ilho, O. Qualidade da bebida dealgumas espécies c populações dcriv adas dehíbridos intcrcspecificos de (·n/fel/. Hragantia.Campinas. 49( ~ ):~R 1-~90. 1990.
Costc. R. Catetos ~ Cafés eu el Mundo. IOIlH' I .I.L's Carés. l-d par (i.-I>' Maisonneu-,c 8:. l.arosc(Paris). 19:':' ..~X I p.[)'Antonio. A.M. s. Guerra ~el0. I .c..Comportamento de plantas deri \ udas de scle~<)e,
do cafeeiro C 119:'·:;-6-~ (e.119) COIll resistênciaao bicho rn inci 1'0 Pcritcucoptcr« Cn//l'l'! /"
tGuérin-Mcnév illc. IX4~ I. 14" CongressoHraxilciro de Pesquisas Careeirus. I" Congressol.at ino Americano de lccnolog!a Cafcc ira.Campinas. São Paulo. Brasil. 19X7. IRC (RIo c1eJunci ro l, p.~()~-~O..L
Guerreiro-Filho. O. ('n/Ie'a rUCl'II/(}\U l.our l 'ncRe\ uc. CalL; Cucao lhé.'6(.'): 171-1 X6. 199~.
Guerrciro-lilho. O. ldcntification de gl.-ne'-o de
rcsistance Li ['(,I";/CIIl'II/'h'"'' CII//l'cI/" cn \ uc lILl'amcl ioration de ('n/teu urabtca Potcutic ld'cspecc-. diplordc« du geme ('n!/L''': genes deBacill u» 1/IIII"ill.!.!./d/.\is. Thcsc de I)octora1.INS/\M. Montpclf icr. 173 p.. 199..L
Gucrre iro-Filho. O .. Silvarolla. \1. H. 8:. l.vkcs. ,\.B. l.xprcssion aud modc 01' inhcritaucc of
2
I-1
\hllllnl
"àn 11 n,1''''.1..,
I
I-1
-1-1
12
, 1
I'111.'I1X5cv XI
111,-1115\1\. .'XX-I,
BIBLIOGRAFIA
Ensaio I'rn::ênie Produ Resistên II{eSiSlên
"à" de ria H. cia P.frutos vustatrix t'Ollállll(1..:::)
'--------._. ----.--- ---- ~~--- ----- -47.; 90/2.' "O -' I -1 I
\1'\ 'XX- -'-1 'I <J I
17 i I
III!
I
2-1 i111-111l-1 21 I
I
\' " ,~x- I,
: 17 I IL __~ __ .L ... L __L__
apresenta baixo teor de cafeína. Híbridos entre ('arabica e r ' /"aC(,IIIIi.I"U apresentaram em estudosrealizados em Campinas bebida de boa qualidade.que foi sucessivamente melhorada porretrocruzamentos com (' arabica. Carvalho L'I al ..
(1990) verificaram que a qualidade da bebida doF I triploidc entre C' arabica e C'. raccmosa e dc
cafeeiros pertencentes a segunda geração deretrocruzamentos para c: arahica. eram próximasá qualidade apresentada por (' arahica, utilizadocomo controle dos experimentos. Resultadossemelhantes \ em sendo observados naspopulações mais avançadas do programa demelhoramento desenvolvido em Campinas.
Estado atual do programa.Populações avançadas. pertencentes ú quintageração de rctrocruzamcnros. vêm sendocultivadas em ensaios de campo em EstaçõesExperimentais do IAC. Várias progênies.resistentes ao bicho mineiro. e em alguns casostambém ,1 Hcmiieia vasuurix, têm apresentadosproduções superiores aos cultivares de (' arabu:«atualmente cultiv ados figura ~). Con1(1 aresistência proveniente de C'. ,.UC('lIIn.\U seriadevida a presença de dois genes complementarese dominantes. realizou-se uma geração adicionalde retrocruzamcntos com o progenitor recorrente.o cultivar Catuai Vermelho de C arabu:« (A). crurazão da ausência de seleção por ocasião dos doisprimeiros rctrocruzamentos (csporuáncos j e pelofato que esses retrocruzamcntos não controladosforam realizados com variedades muito poucoprodut ivas. Estima-se que ainda são necessáriasuma geração de autofccundaçào de plantasresistentes supostamente hetcrovigotas para osdois genes (1.111/1111/ e /.11I::/11I::) e uma segundageração de autofccundação das plantas resistentesprov cnicntcs da geração anterior. Os indi , iduos
/.11I//.111/ /'III}/'II/} constituirão a nova variedade.enquanto as plantas /.11I/ !-III/ !-1I1}/III:: e /.111/1111//.11I::/.111:: ainda cn scgreg acão numa proporção de3 resistentes: I suscct Í\ cl e as plantas /.111//11I//.111::/1/1.' (9 resistentes : 7 susccuvcis). serãoeliminadas.

resistance to leafminer . Euphytica, 105 (I) : 715. 1999.Guerreiro-Filho, O.; Medina-Filho. H.P.;Gonçalves, W. & Carvalho, A. Melhoramento docafeeiro. XLIII. Seleção de cafeeiros resistentesao bicho mineiro, Bragantia (Campinas). vo1.49,n° 2,1990, p.291 1-304.Katiyar, K.P. & Ferrer, F. In: InternationalAtomic Energy Agency, Viena, IAEA, 1968. p.165-175.Levy, F.A .. Bogdan, 1.. Ferraz. J.T .. Della Torre.M.A. & Caravalho, A. Precocidade de maturaçãono cafeeiro. 15° Congresso Brasileiro dePesquisas Cafeeiras, Maringá, Paraná. Brasil. 2629 sept. 1989. Instituto Brasileiro do Café (Rio deJaneiro). p.155-157.Medina-Filho. H.P.. Carvalho. A. & Monaco,L.C. Melhoramento do cafeeiro. XXXVII Observações sobre a resistência do cafeeiro aobicho mineiro. Bragantia (Campinas). vol. 36.1977a. p.131-137.
209
Medina-Filho. H.P .. Carvalho. A. & Medina.D.M. Germoplasma de C. raccmosa e seupotencial no melhoramento do cafeeiro. Bragantia(Campinasj.vol, 36. I977b. p.XLlII-XLVI.Parra, J.R.P. Biologia comparada dePerileucoptera coffeella (Guérin - Méneville.1842) (Lepidoptera - Lyoneti idae) visando seuzoneamento ecológico no Estado de São Paulo.Revista Brasileira de Entornologia. 29( I):4576.1985.Silva, H.L. Número de cromossomos em Co/filaracemosa Lour. Bragantia (Campinas). vol. 15.1956, p.XVII-XVIII.


211
GENETIC IMPRDVEMENT DF CDFFEE IN INDIA
C.S.Srinivasan, N.S.Prakash, D.Padma Jyothi, V.B.Sureshkumar and V.Subbalakshmi
Central Coffee Research InstituteCoffee Research Station - 577 117 Chikmagalur Dist., Karnataka, India
Although coffee is not an essential cornrnodity forhuman consurnption, its stimulating properties aretoa well known to ignore its importance in humanhealth, if taken at moderate leveis. In India,eoffee has a place of pride amongplantation erops grown and is traditionallycultivated on the south - western hiJ1 slopessince 150 years, As history indicates. thecommercial eultivation of coffee in India wasstarted during early 19th century by the Europeanenterpreneurs (Anonyrnous, 1995). These earlycoffee plantations have flourished in SouthIndia for nearly a century tiJ1 the leaf rustepidemics were seen and soon the leaf rustbecame the disease of coneern for arabica coffeein view of its high econornic importance.Incidentally the largest number of races as rnanyas 30 has been reeorded from India. Thus, the fateof eoffee industry especially arabica has beenlargely dependent on release of varíetíes resistantto rust with aeceptable yield leveis. beanstandards and cup quality. The Central CoffeeResearch lnstitute established near Balehonnur inChikmagalur Dist. of Karnataka state has renderedyeoman services in breeding and release ofimproved arabica & robusta varieties to thegrowers from time to time since its establishmentin 1925 as "Mysore Coffee Experimental Station",The success aehieved in developing rust resistantvarieties in India during 1940's (Narasirnhaswarny, 1961) are pioneering ones, in the historyof coffee breeding. lt is pertinant to mention thatthe classical work of W.W.Myne on the existanceof physiological races in rust pathogen (Myne,1932. 1936. 1939); supplemented the breedingefforts not only in India but also elsewhere in theworld. This presentation details the objectivesand achievements on genetic improvement ofcoffee in lndia, performance of the varieties andreeent efforts towards further irnprovernent of theproven materiais.
Geographical informationIn India the major coffee tracts are distributed inhi 11 ranges of Southern part of the countrybetween 9° and 14° N latitudes and 500-1500 maltitudes: the arabica in high elevations androbusta in low lands. The annual rainfall variesbetween 1000 - 2500 mm with the temperatureranges between 8° C to 34 o C.
ARABICA IMPRDVEMENT
In lndia, improvement of arabica eoffee wasstarted even before the commeneement oforganised research. Some enterprising plantersmade frantie effo rts to develop superiorcultivars through selection / hybridization, inorder to tackle the rust disease. In this context,hybrids like 'Hamiltons', 'Jacksons','Netrakonda' and seleetions like 'Coorgs', 'Chieks'and 'Kents' were some important ones whichfound place in the pages of coffee breedinghistory, Nevertheless some of these selectionsare still found here and . there in commereialplantations. 'Kents' is one sue h seleetion madeby a private planter which served as the majorsource for planting during the 1920s and is sti I1internationally acclaimed for its liquor quality.Inspite of the best efforts of the plantingcomrnunity, plant materiais with stableresistance could not be developed due to theability of the rust pathogen to mutate intovirulant races. This necessitated the organisedresearch activities for improvement of arabicacoffee in order to evolve superior cultivars withleaf rust resistance. high yielding potential.superior quality and wide adaptability.Organised research towards geneticimprovement :Since the inception of organized research incoffee in India during 1925, development ofimproved strains using conventional breedingtechniques has been found successful. Thesuccess of breeding programme in any cropdepends on the exploitation of the availablegenepool. In lndia, as a first step towardsvarietal irnprovement, initial germplasm withabout 250 diverse collections of arabica coffeefrom dífferent estates were established at CCRIduring 1925-40. In subsequent years, severalexotic wild collections of coffee and cultivatedforms of arabica and robusta were introducedfrom other coffee growing countries toestablish a massive genebank. Presently, thelnstitute's genebank comprises of over 350arabica collections, 15 robusta types and 18different species of Coffea and is one of therecognised germplasm centres for coffee inthe world. The individual collections of thegene bank have been assessed and genesconferring rust resistance. bush formation.

production potential, quality and droughthardiness have been identified and exploitedfrom time to time in developing 12 superiorstrains of arabica for commercial cultivation.
Early Indian Selections (1925-1960)Over the years the simultaneous cultivation ofboth arabica and other diploid species likerobusta and liberica has resulted in the naturalhybridization between these types. The earlylndian selections like S.288 and S. 795 wereevolved frorn some of these natural hybridsidentified during the indigenous germplasmcollection.S. 288 was the first selection released during1936-37. It was superior to other varieties like'Chiks' and 'Kents' under cultivation then . byshowing rcsistance to leaf rust races I and 11.S. 288 strain was developed by pure lineselection from S.26 mother plant a putativenatural hybrid betwen Iiberica and arabica.This selection has the potential to givemoderate yields of about 1000 kg/ha butproduces defective beans to the tune of 2030%. To improve the quality aspects ofS.288. it was crossed with 'Kents' and S.795the most popular strain of lndian arabicas wasdeveloped. Since its release in 1945-46. S.795
attracted the attention of planters and is stilJthe most popular selection occupyinglarge area (60%) under cultivation. The bushes
of this variety are tall, very vigorous. widespreading with profuse growth and high yieldpotential of 2000 kg/ha. This selection iswidely adpatable to different agro-climatic zones.The beans are oblong. bold representing 70% 'A'grade with good cup quality (Narasimhswamy.1960).The SH 3 factor responsible for resistance to
severa I physiologic races ar leaf rust pathogenwas identified in these strains with possibleintroduction from C. liberica. Thus, these twocornmercial arabica cultivars of Indian origindiffer frorn the rest of pure arabicas cultivated inthe Globe elsewhere (Wagner & Bettcncourt,1965). The S.288 and S.795 remained resistantover a period of time till uncommon rust racesV111. X II and XIV became common on thesecultivatrs due to to host preference. Thisnecessitated furiher work to develop resistanceto race VIII the most comrnon and virulantrace affecti ng S.795 populations. As some oflhe exotic introductions frorn Ethiopia, likeCioccie. Agaro and S 12 kaffa were found
resistant to race VIII. Sln. 4 was developedfollowing pureline selection in some of thesecollections. This selection includes three
212
Ethiopian arabica collections viz. Cioccie,Agaro and Tafarikela. Among these, the first twoexhibit serni-erect branching, while Tafarikelashows drooping nature. Ali these are tall typesand moderate yielders. Tafarikela is an earlyripener. Cioccie and Agaro are resistant tocomrnon rust races I. [I and also to raceV1Il. Tafarikela exhi bits general tolerence to leafrust pathogen showing very less rust buildup under field conditions, even thoughsusceptible to race I & II under laboratoryscreerung.
Tetraploid interspecific hybrids (1960-1985)In subsequent years, the spontaneous as wellas synthetic hybrids have been exploited.'Devamachy', a putative robusta x arabica hybridof indigenous origino was exploited to developSelection 5. This selection comprises of twoimportant families derived by crossingDevamachy x Rume Sudan and Devamachy xIndian selection S.333. Devamachy x RumeSudan line is known for its field tolerance to rustand in recent screening at C IFC, reportcd to beresistant to Coffee Berry Disease also. However,small sized beans is the negative aspect in thisfine. The out come of S. 333 x Devamachycross has been found promising and ischaracterized by semi-drooping habit with goodresistance to leaf rust (85% of the population)and bolder bean size (Table I) (Srinivasan andRamachandran, 1998). This variety has shownthe yicld potential of 1300 - 2000 kg lha underditTerent agroclimatic zones (Table 2). In one ofthe trials this line has recorded an average yield of2048 kgs/ha (5 years) maintained without anyfungicidal sprays (Raju et 0/1996).With the experience in exploiting naturalhybrids, systematic approach to developinterspecific hybrids through programmedhybridization between tetraploids vs. diploidsand diploids vs. diploids was also taken up atCCRl in order to develop Sln. 6 and Sln. I I.The Sln. 6 was evolved by crossing robustaand arabica followed by back crossing toarabica. While developing this hybrid the genomeof 2x & 4x occuring in robusta and arabicarespectively were not altered. The resultanttri ploids were back crossed to arabica and theresuliant progeny was subjected to pure linesefection. S, of BC 2 has been released forcommercial cultivation as Sln.6. While advancingthe generations selection was made lo minimisethe percentage of ernpty locules which areotherwise high due to hybridity (Sreenivasan.1983). The plants of Sln.6 are vigorous. widespeading and tend more towards arabica
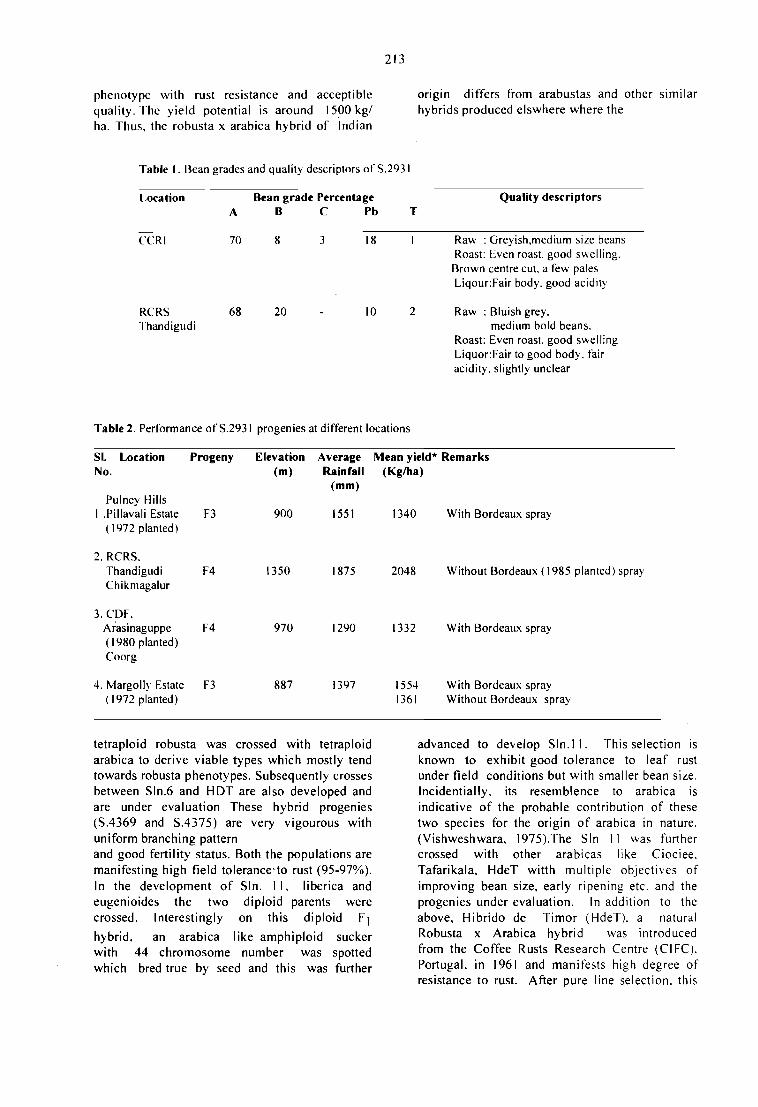
213
phenotype with rust resistance and acceptiblequality. The yield potential is around 1590 kg/ha. Thus, the robusta x arabica hybrid of Indian
Table I. Bean grades and quality descriptors of S.293 I
ongm differs from arabustas and other simi larhybrids produced elswhere where the
Location Bean grade PercentageA B C Pb T
Quality descriptors
CCRI
RCRSThandigudi
70
68
8
20
3 18
10 2
Raw : Greyish.mcdium size beansRoast: Even roast, good swelling.
Brown centre cut, a few palesLiqour:Fair body, good acidity
Raw : Bluish grey.medium bold beans.
Roast: Even roast. good swellingLiquor:Fairto good body. fairacidity. slightly unclear
Table 2. Performance ofS.2931 progeniesat different locations
Pulney HillsI .Pillavali Estare F3
(1972 planted)
Mean yield* Remarks(Kg/ha)
SI. LocationNo.
Progeny Elevation(m)
900
AverageRainfall
(mm)
1551 1340 With Bordeaux spray
2. RCRS.Thandigudi F4Chikmagalur
3. CDF.Arasinaguppe F4(1980 planted)Coorg
4. Margolly Estate F3( 1972 planted)
1350
970
887
1875
1290
1397
2048
1332
15541361
Without Bordeaux (1985 planted)spray
With Bordeaux spray
With Bordeaux sprayWithout Bordeaux spray
tetraploid robusta was crossed with tetraploidarabica to derive viable types which mostly tendtowards robusta phenotypes. Subsequently crossesbetween Sln.6 and HDT are also developed andare under evaluation These hybrid progenies(S.4369 and S.4375) are very vigourous withuniform branching patternand good ferti Iity status. Both the popu lations aremanifesting high field tolerance·to rust (95-97%).In the development of Sln. 11, liberica andeugenioides the two diploid parents werecrossed. Interestingly on this diploid FI
hybrid, an arabica like amphiploid suckerwith 44 chromosome number was spottedwhich bred true by seed and this was further
advanced to develop Sln.l l . This selection isknown to exhibit good tolerance to leaf rustunder field conditions but with smaller bean size.lncidentially, its resemblence to arabica isindicative of the probable contri bution of thesetwo species for the origin of arabica in nature.(Vishweshwara, 1975).The Sln 11 was furthercrossed with other arabicas like Ciociee.Tafarikala, HdeT witth multi pie objectives ofimproving bean size, early ripening etc. and theprogenies under evaluation. In addition to theabove, Hibrido de Timor (Hde T). a naturalRobusta x Arabica hybrid was introducedfrom the Coffee Rusts Researc h Centre (C IFCl.Portugal, in 1961 and manifests high degree ofresistance to rust. After pure line selection. this

was released as Sln. 8. Now, HdeT is a majorsource for rust resistance and widely used inbreeding for rust resistance all over the world.Several F I hybrids between HdeT and other
arabicas were made available by CIFC, Portugalas well as developed in India. AlI thesecombinations manifest a high degree ofresistance
214
and productivity potential (Table 3). Thesuperior combination, HdeT x Tafarikelawas released as Sln.9. The bushes arevigorous, semi-erect to drooping with earlyripening behaviour and comes up well inmarginal areas and supply positions.
Table 3. Performance of F I hybrids (HdeT x other arabieas)
FI hybri Parentage Mean yield (Kg/ha) Rust resistance(6'h to 10th Year Avg.) (%)
S.2790 HdeT x Tafarikala 1760 100.00
S.2792 Tafarikala x HdeT 1258 74.08
S.2794 HdeT x Geisha 1950 98.00
S.2795 HdeT x S.1934 994 87.50
S.2800 HdeT x Bourbon 2162 86.67
S.2803 SI2 Kaffa x IldeT 1733 90.00
Dwarf and Semi- dwarf hybridsTo meet the planters' demand for dwarf and semidwarf varieties suitable for high-density planting,mutants like 'San Ramon' and 'Caturra' havecome in handy. San Ramon, a dwarf mutantsusceptible to several races of leaf-rust, has beenused in a series of crosses with the resistant linesS.795, Agaro and HdeT to produce hybridswhich are found suitable for cultivation in 10wrainfall areas. The cross of Caturra with S 795and Cioccie made available by CIFC is foundpromising and named as Sln.1 ü. The catimorhybrid developed as a international collaborativeprogramme between C [FC and other coffeegrowing countries including India was foundpromising and released by giving popular name .•Cauvery" in 1985. This material becamepopular in a very short time owing to itsprecocious bearing nature, high yield potential(2 to 2.5 tonnes / ha) and good response to theintensive cultivation. Though this material wasresistant to leaf rust in initial years, break downin resistance was observed due to theappearance of five new rust races (Rodrigues etai 1993) which could infect majority of thesepopulations (Prakash et aI 1998). Further,advanced generation of Villasarchi x Hde'T,Catuai x Catimor (both supplied by CIFC) andCatimor x Sarchimor developed in lndia arefound promising with high uniformity andvigour, rust reisistance, bold beans coupledwith good cup quality (Srinivasan. 1996). These
lines are under multilocation evaluation andmay form future rnaterials for cornmercialrelease.
ROBUSTA IMPROVEMENT
The second important and commerciallycultivated species is Coffea canephora('Robusta'). Hence, due importance has alsobeen given for improvement of robusta in lndia.Robusta coffee possesses several usefulcharacters like high tolerance to leaf rustpathogen, white stem borer. nematodeinvasion and potentiality to give consistentyields under irrigation. On the other hand.inability to endure long drought, late cropping aswell as late stabilization of yields and inferiorquality compared to arabica are some of thenegative aspects of robusta coffee. Keepingtliese aspects in view improvement of robustacoffee was undertaken by CC RI .As Robusta is cross pollinated, breedingmethods like mass selection, clonal propagationfollowed by pedigree selection andinterspecific hybridization have been used toevolve three commercial types.The seedling progenies of 12 high yielding motherplants selected from private estates wereevaluated and two materiais S.27Ü andS.274 were found superior and released forcommercial cultivation as Sln. 1 (R). Thesetypes grow into moderately large trees with

215
Table 4. Performance ofC x R hybrid in diflcrent agro-clirnates and bean grades
Zone Yield (Kg/ha) Grade percentages(CxR sibs)
-----------I. Mudigerc 1031 ;\ 56.682. Koppa ISSO B 12.963.Coorg 2080 C 7.80
Ph 19.464. Wynad 1335 BBB 3.10
vigorous growth. The fruits and seeds are bold.borne in large clusters. Liquor is rich in body andthe cup is neutral. S.274 is preferred overS.270 on account of its boider beans and it isperforming well under irrigated conditions.Further, 17 superior plants which haveyielded twice OI' more than the family mean yieldwere identified from the initial single treeprogenies of S.267 and S.278 and named asBalehonnur Robusta (BR selection). Based onthe individual performance of these clones, BR 9.BR 10 and BR I I were found to be superior inproduction and as such seed rnixture of theseclones was issued for commercial cultivation asSln.2 (R). Clones were also issued for theestablishment of bi/polyclonal gardens. Theclones showed high degree of stabi lity for 'A'grade beans The third robusta Iine developedin the series is C x R hybrid. This is similar tothe 'Congusra' hybrid developed in Java earlier(Crarner. 1957). Cicongensls another di ploidspecies known for its compact bush statureand quality was crossed with S.274 rousta in1942. Backcrossing to robusta and sib matingwas followed to achieve compact stature withbold fruits and seeds (Anonymous 1(98). Beansare mediurn, bold, golden brown and gives softJiquor with tlavour nearer to arabica (Table 4).Thus. the continuous efforts of coffeeimprovement group have been wellappreciated by the fanning community and itis noteworthy to mention that the arabicavarieties developed at this Institute havegradually replaced the old cultivars andpresently cover 98% of the total area underarabica coffee. However, due to the longeconomic life. the intake of improved robustastarins is rather slow. Inspite of this, the C xR hybrid is gaining popularity owing to itssuperior quality characteristics and is beingpre ferred over ot her robustas.
NEWER CHALLENGES
The ever increasing racial diversity of theleaf rust pathogen (Hemileia vastatrix B. &Br). sudden appearance of coffee berry borer(Hypothenemus hampei Ferrari) in 1991 andincreased awareness for quality of the producedue to market liberalization pose new challenges,Accordingly, a paradigm shift has beentaking place in coffee research. Achieving thedurability of rust resistance through genepyramiding approach development of highyielding strains with a better response tointensive cultivation through multiple crossingprogrammes. evol ving pest resistant and lowcaffeine I ines through biotech/molecularapproaches are the best considered ernergingtrends of the day.
REFERENCES
Anonymous. 1995. Coffee Guide, Coffee BoardResearch Dept. India. Anonymous. 1998. CoffeeBoard Research Departrnent, CxR Technical BulI.1998.Cambrony. H.R. 1988. Arabusta and otherInterspecific Fertile Hybrids. P.263-291.In:R.J.Clarke and R.Macrae (ed.) Coffee, volume4: Agronomy. Elsevier applied Science PublishersLtd.. 1985. Crown House. Linton Road, Barking,Essex IG 118 JU. England.Cramer, P.G.S. 1957 in F.L.Wellman, Revicw ofLiterature of Coffee Research in lndonesia. SICEditorial Arnerican Institute of Agriculturalciences. Turrialba. Costa Rica.Mayne, W.W., 1932. Physiologic specialization ofHcmilea vastatrix B.& Br. Nature 129:5 10.Mayne. W.W.. 1936. Annual Report ofthe CoffeeScientific Ofticer. 1935-36. Mys. Coffee. Exp.Stn. BuII.14:21.Mayne, W.W.. 1939. Annual report of the CoífccScientific Officer. 1938-39. Mys.Coffee. Exp. Stn.BulI. 19.

Narasimhaswamy, R.L. 1960. Arabica selectionS.795. Its origin and performance. Indian Coffee.24: 197-204.Narasirnhaswamy, R.L. 1961. Coffee leaf rustdisease (Hemileia) in India. Coffee. Turrialba.3:33-39.Raju.T, C.S.Srinivasan and M.Jagadeesan. 1996.Performance of CCRI released arabica selectionsin the Pulneys in Tamilnadu. Indian Coffee.LXV(3): 17-20.Prakash.N.S.. C.S.Srinivasan and R.Naidu. 1998.Races of coffee rust tHemiieia vastatrix} : IsIndia a potential source? PLACROSYM XIII. Inpress.Rodrigues Jr..C.L V.M.P. Varzea, I.L. Godinho,S.Palma and R.C.Raho. 1993. New physiologicraces of Hemileia vastatrix . ASIC. 15th Colloque,Montpellier.1993.318-321.
216
Sreenivasan, M.S. 1983. Cyto-ernbryologicalstudies of coffee hybrids. Ph.D. hesis, Universityof Mysore. rinivasan, C.S. 1996. Current statusand future thrust areas of research on varietalimprovernent and horticultural aspects of coffee.J.Coffee Res. 26( I): 1-16.Srinivasan, C.S. and M.Ramachandran. 1997.Selection 58. S.2931 (S.333 x Devamachyhybrid). An old arabica hybrid rediscovered withpromising features. Indian Coffee. 61 ( 1 I) : 4-6.Vishveswara, S.1975. Coffee: Cornmercialqualities of'sorne newerSelections. Plant. Chron. 70. 122.Wagner. M.. and A. J. Bettencourt, 1965.Inheritance of reaction to Hemileia VastratrixBerk.Br. in Coffea arabica L. In progress report1960-1965 Coffee Rust Research Centre. Oeiras,Portugal.

217
MELHORAMENTO CLÁSSICO DO CAFEEIRO RELACIONADO COM A BIOTECNOLOGIAMODERNA(I)
L. C. Fazuoli (2.3); M. P. Maluf(2.~); O. Guerreiro Filho (2); H. P. Medina Filho(2.3); M. B. Silvarolla (2)
I Com recursos do PNP&D Café/Embrapa.~ Pesquisador Científico - IAC Centro de Café e Plantas Tropicais CP. 28
Campinas.SP -- Brasil., Com Bolsa de Pesquisa do CNPq~ Bolsista FUNAPE
CEP: 13001-970
I. MELHORAMENTO CLÁSSICO.
1.1. Espécies de café e variabilidade.Existem controvérsias no que se refere àtaxonomia das espécies de Coffea. Muitas têmsido descobertas no decorrer das coletas efetuadasnas tlorestas tropicais da África desde 1940.Baseando-se na literatura existente conclui-se queas verdadeiras espécies de Coffea são oriundas daparte central e equatorial da África. deMadagascar e ilhas próximas do Oceano índico.As demais espécies que foram descritas comopertencentes ao gênero Coffea e oriundas deregiões asiáticas já não são consideradas comoespécies verdadeiras de Coffea. No que se refereao número de espécies existentes. Chevalier(1947) listou 65 espécies. onde são incluídas 24que pertencem a outros gêneros. Cramer (1957)sugeriu a existência de pelo menos 100 espécies.Purseglove (1968) c itado por Wrigley (1988)relatou 50 espécies do gênero Coffea, sendo 33da África Tropical. 14 de Madagascar e 3 dasIlhas Mauricio e Reunião. Por outro lado Charrier(1978) listou 56 espécies provenientes deMadagascar. Ao que parece estas espécies estãosendo estudadas. notando-se divergências quantoao número de espécies verdadeiras deMadagascar. Novas espécies de Co/leu tem sidodescritas por Bridson (1982). principalmente daparte oriental africana. Algumas ainda não foramcaracterizadas o suficiente. para serem descritas.Leroy ( 1980) reconheceu três gêneros decafeeiros: (·o//ea. Psilanthus e Nosrolachma.Este último é restrito à Ásia e Indonésia. Omesmo autor distinguiu três subgêneros de Coffeae dois de Psilanthus.A classificação mais recente agrupa os cafeeirosem :2 gêneros: Cofleu c Psilanthus, O gêneroPsilanthus por sua vez é dividido nos subgênerosPsilanthus e Afrocoffca. e o gênero Co/leucompreende os subgêneros: Co/leu e Baracoffea.Enquanto o subgênero Baracoffc« é representadopor 7 espécies. o subgênero Coffca abriga 80espécies sendo aproximadamente 25 oriundas daÁfrica Continental e 55 de Madagascar (Bridson.
1987; Bridson & Verdcourt. 1988: Bridson.1944).As dificuldades de classificação das espécies deCoffea decorrem do fato de não existiremcoleções com essas espécies descritas. o que seriaaltamente desejável. No entanto. a maior partedos países africanos tem imposto sérias limitaçõesà saída de material nativo. como as novasespécies descritas por Bridson (1994) não foramcolocadas em coleção nos países onde foramcoletadas. Novas coletas de materialprincipalmente dessa região oriental se fazemnecessárias. O estabelecimento de Bancos deGennoplasma das espécies conhecidas. e seusderivados a nível mundial e em vários países émuito importante pois entre as formasselvagens. algumas encerram característicasaltamente vantajosas do ponto de vista deresistência a moléstias e pragas. tolerância à secae outras variações de ambiente. bem comodiferentes características das plantas (sistemasradicular. folhas. flores. frutos. etc). Bancos deGerrnoplasma de Coffea existem na Costa doMarfim. Madagascar. Costa Rica. Colômbia.Brasil e outros países. No entanto em todos estespaíses. à exceção de (' arahica e C. canephora.essas coleções contam com poucos exemplaresdas demais espécies de Coffea. No InstitutoAgronômico de Campinas foi possível reunir até opresente. 18 das principais espécies do gêneroCoffea e três do gênero Psilanthus. Algumasespécies possuem muitos representantes como é ocaso de C. arabica e C. canephora e outras.pouquíssimos exemplares. Em muitas coleções.alguns cafeeiros não representam a espécieoriginal. pois foram introduzidos por sementes esão na verdade híbridos entre espécies diferentes.Dentre as espécies conhecidas. C arahica (caféarábica) e C. canephora (café robusta) são as duasmais importantes no mercado internacional.Cerca de 70% do café negociado no mundo éarábica e 30% é do tipo robusta. As outrasespécies de café têm importância para seremuti Iizadas em programas de mel horamento deacordo com as características agronômicas quepossuem. C. arábica é nativa de uma região

estudo não se notou uma tendênciarelacione o maior sucesso dascom a distribuição geográfica das
restrita. marginal às demais espécies. localizadano sudoeste da Etiópia. sudeste do Sudão e nortedo Quênia (Chevalier, 1947: Charrier, 1978:Bridson, 1982: Leroy, 1982). Trata-se de umaespécie nobre. que produz café de boa qualidade.Existem várias variedades de C arabica e mais de40 mutantes já descritos (Carvalho et al., 1991).As características desses mutantes são muitovariáveis. como por exemplo forma ou coloraçãode tolha. crescimento e forma de planta. tipo deflor e florescimento. forma e coloração dos frutose sementes. resistência a moléstias. pragas enernatóides.A outra espécie de interesse comercial (C.canephorai tem distribuição geográfica maisampla. ocorrendo nas regiões ocidental e centraltropical e subtropical de cont inente africano. Essaampla faixa compreende a República da Guiné,Libéria. Sudão e Uganda. Elevada concentraçãode tipos ocorre na República Democrática doCongo (Chevalicr, 1947: Charrier & Berthaud.1985). A variabi Iidade existente dentro dessaespécie é muito grande em relação ao tamanho eforma da planta. folhas. frutos e sementes. Osistema rad icu lar dos representantes de C.cancphoru é bem desenvolvido e a resistência àsprincipais moléstias e pragas é acentuada. O teorde cafeína e de sólidos solúveis em suas sementesé superior aos encontrados no café arábica.As outras espécies de Coffea não são cultivadaseconomicamente. mas ocorrem de forma silvestree possuem variabilidade elevada para muitascaracterísticas. principalmente resistência àsmoléstias. pragas e nernatóides. O grandeinconveniente da maior parte das espéciessilvestres de Co/leu é a quantidade muito baixa deflores. característica esta responsável pela baixaprodução de frutos. Algumas das espécies sãoisentas. ou têm baixo teor de cafeína e sólidossolúveis nas sementes. Outras espécies temmaturação muito tardia ou muito precoce e pelomenos uma. C. racemosa, tem resistência à secae folhas decíduas.
1.2. Biologia da reprodução e relação entreespécies.A espécie C arabica é tetraplóide com 2n=44cromossomos. autocompatível e multiplica-sepredominantemente por autofecundação, queocorre em aproximadamente 90% das flores. Ataxa de fecundação cruzada natural tem sidoavaliada durante anos seguidos em Campinas peloemprego de diferentes mutantes com genótiposrecessivos. O mutante mais apropriado para estadeterminação é o cera. que apresenta endosperrnade coloração amarelada. Os óvulos do cafeeirocura quando polinizados pelo pólen de cafeeiros
218
de endosperrna verde homozigotos dão origem asementes de cor esverdeada devido ao fenômenode xênia, De outra parte. os óvulos do cafeeirocera quando polinizados com pólen da própriaplanta dão origem a sementes com endospenna decor amarela. Desta maneira. mantendo-se oscafeeiros cera rodeados por plantas deendospenna verde e analisando-se as suassementes. poder-se-ão observar algumassementes com endosperma verde. resultantes doscruzamentos naturais. Pelo seu número. avalia-sea porcentagem de fecundação cruzada natural.que é de cerca de 10% em C arabica (Carvalho &Mônaco. 1962). O cafeeiro arábica é portantoautofértil e não tem sido notado efeitodesfavorável das autofecundações sucessivas novigor e na produtividade das plantas. Aferti Iização em C. arabica se dá 24 horas após apolinização. ocorrendo a primeira divisão dacélula do endosperma 21 e 27 dias após afertilização e a primeira divisão do zigoto. 60-70dias após a polinização (Mendes. 1958).A espécie C canephora e as outras espéciesconhecidas do gênero Coffea são diplóides,possuem 2n=22 cromossomos. sãoauto incornpativeis e multipl icam-seexclusivamente por fecundação cruzada(Berthaud. 1980: Conagin & Mendes. 1961.Devreux et al., 1959). Apenas uma exceção éconhecida. A espécie C. sp. Ate/mil/dou édiplóide e autocornpativel. A incompatibilidadeem C. canephora foi determinada como sendo dotipo gametofítico. o que também se verifica emC. dewevrei e C. congensis . Há indicações deque essa incornpatibi Iidade em C. canephora sejacondicionada por uma série alélica S. em umúnico loco (Conagin & Mendes. 1961: Devreuxct al.. 1959), Não se sabe ao certo o número degenes envolvidos na incompatibilidade.No que se refere a espécies do gênero Psilanthus,existem indicações que a espécie Psilanthusbcngalcnsis, embora diplóide. seja compatível.devendo multi pIicar-se predom inanternente porautofecundação, o mesmo ocorrendo com osrepresentantes de P travancorcnsis. (MedinaFi Iho et al.. 1982).No estudo das relações entre espécies de café.hibridações interespecíficas entre 14 espécies deCoffea foram realizadas no IAC por Carvalho etal. (1984a) e os resultados indicaram que osucesso das hibridações entre espécies de umamesma subseção. nem' sempre é maior do queentre as hibridações com cafeeiros de subseçõesdiferentes.No mesmogeral quehibridações

•!
espec ies. o mesmo ocorreu com espécies queapresentam semelhanças morfológicas.Outros estudos efetuados por diferentespesquisadores revelaram que não se pode fazerpredições sobre o sucesso das hibridações,viabilidade. fertilidade dos híbridos. baseando-seem semelhanças bioquímicas ou número ecomportamento dos cromossomos (Longo. 1972:Medina. 1972 e Maglio. 1(83).
1.3. Análises genéticas em Coffea arábica,variabilidade e aproveitamento de algumascaracterísticas no melhoramento.Dentre as plantas perenes. o cafeeiro <- '. arabicaé com certeza a planta melhor estudada no que serefere à herança e relação de dominância defatores genéticos. Para estes estudos. tem-seutilizado o cultivar Arabica (Nacional ou Típica)de C. arabica como padrão.As pesquisas sobre herança das principaiscaracterísticas das variedades de C. arábica têmse mostrado importantes na obtenção de novoscultivares. através de recornbinações genéticas.No decorrer de 67 anos de observaçõesencontraram-se aproximadamente 42 mutaçõesque permitiram efetuar análises genéticas.Apenas os fatores genéticos anômala e anormalisrevelaram ser ligados (Carvalho. 1(60).Encontraram-se ainda anomalias que parecem serde caráter citológico e outras de origemcitoplásmica. Os fatores estudados afetamcaracterísticas da planta como altura. tipo deramificação e das folhas. flores. frutos (Carvalhoet al.. 1(91) e sementes. além de resistência amoléstias e pragas. Vários desses fatoresrevelaram-se dominantes. outros recessivos oucom dominância incompleta. em relação aosalelos do cultivar Arábica (Carvalho. 1958: Krug& Carvalho. 1951 ). Dentre os fatores queafetam a altura das plantas. destacam-se o Caturra(Ct), São Bernardo (Sb). Vila Lobos (VI) e SanRamón (Sr). todos dominantes e independentes.Esses fatores. de grande interesse econômico.contribuem para a redução do comprimento dosinternódios da haste principal e dos ramos laterais.resultando em cafeeiros de menor porte e deaspecto compacto (Carvalho et al.. 1984b). Umexemplo do aproveitamento do fator Caturra foi aobtenção dos cultivares Catuaí Vermelho e CatuaíAmarelo de porte baixo. vigorosos c altamenteprodutivos. largamente plantados no Brasil e noexterior (Carvalho & Mônaco. 1972: Fazuoli.1986: Carvalho & Fazuoli, 1(93). Por suamenor altura. a colheita e os tratos fitossanitáriossão facilitados c menos dispendiosos. Verificouse. também. que vários fatores têm efeitopleiotrópico marcante. como a laurina (Ir). que.
219
além de afetar o porte e a altura de planta.tamanho das folhas. forma e tamanho dos frutos edas sementes. reduz o teor de cafeína nassementes (Krug et al.. 1959: Carvalho et al..1986).O fator genético cera (ce) mostrou-se vai ioso. emvista do fenômeno de xênia que apresenta.Contribuiu para demonstrar. pela primeira vez.que a semente de café é formada por endosperrnaverdadeiro e foi muito utilizado paradeterminação da taxa de fecundação cruzadanatural (Krug & Carvalho. 1939. 1951). Comoapresenta sementes amareladas. serviu tambémpara demonstrar que a cor verde da sementenormal não é devida aos ácidos clorogênicos,pois as sementes cera têm a mesma constituiçãocom relação a esses compostos que as verdes(Carelli et al.. 1974). Resumos de várias dasanál ises genéticas efetuadas em C arabica forampublicados ao longo dos anos (Carvalho & Krug.1949: Carvalho. 1958: Krug & Carvalho.1951: Carvalho et aI. 1991).No que se refere à resistência a moléstias foramdetectadas em C. arabica fontes de resistência aferrugem. a mancha aureolada (Pseudonornassyri ngae pv. garcae), C BD ou antracnose dosfrutos (Coffee Bery Discase, ocasionada por ('coffeanum atualmente designado (' kahawuci(Bettencourt & Carvalho. 1968: Moraes ct al. .1974: Van der Vossen & Walyaro, 1980). Emrelação a nernatóides fontes de resistência têmsido encontradas em materiais procedentes daEtiópia (Fazuo Ii. 1981).Como se pode observar em ('. arabica existe umavariabilidade razoável para algumascaracterísticas. que tem sido utilizada emprogramas de melhoramento. No entanto paracaracterísticas de resistência a moléstias. pragas enematóides, a variabilidade é pequena. Naincorporação em C. arabicu desses genes queconferem estas características deve-se uti Iizarpreferencialmente espécies diplóides.
1.4. Aproveitamento de características deespécies diplóides de Coffea no melhoramentode C arttbica.O modelo de classificação informal das espécies.propostos por Harlan e De Wet (1971) aplicado aogênero Coffea indicou um grande potencial para omelhoramento concentrado na reserva gênicasecundária (Medina Filho ef al., 1(84), Asespécies ('. canephora. c'. congensis, c:racemosa e C. dewevrei. atual mente designadapor C. libcrica var. dewcvrci que pertencem aessa reserva. vêm sendo aproveitadas comoimportantes fontes de resistência a pragas.moléstias. nematóides e a cond ições adversas de

ambiente. Um exemplo é o desenvolvimento nofAC dos cultivares Icatu Vermelho. IcatuAmarelo e Icatu Precoce. provenientes docruzamento de ('. arabicu e C canephora e comresistência a Hetnileia vastatrix (Mônaco el ul ..1974: Fazuoli. 1991: Carvalho & Fazuoli.1(93). Trata-se de um exemplo significativo doaproveitamento de uma espécie diplóide nomelhoramento do cafeeiro C arabica.A utilização da espécie C cancphora também temsido efetuada através do Híbrido dc Tirnor.provavelmente um híbrido natural. entre ('arahica e (' cancpliora (Bettencourt, 1(73). Ouso do Híbrido de Tirnor e derivados é maissimples em relação ao aproveitamento do híbridoArabusta (FI de C. arabica x C. canephorai poiso Híbrido de Tirnor, possui fenótipo de Carabicu. tem 44 cromossomos nas célulassomaticas. é autoférti I e está aparentementeestabilizado (Rijo. 1(74). Os cultivares Obatà(IAC 1669-20). Tupi (IAC 1669-33) e IAPAR59. produtivos. de porte baixo e com elevadaresistência a ferrugem são exemplos doaproveitamento do Híbrido de Timor eindiretamente de C. cancphora (Fazuoli cl al.,1996: IAPAR. 1(93). É interessante ressaltarque determinadas progênies de C. CUI1Cp!lo/"u
constituem também fontes de resistência ,1
Coiletotrichum coffeanum. atualmente conhecidapor C. kaliawae, moléstia designada por C BD(Coffee Berry Disease) que ataca os frutos decafé. provocando a sua queda. Esta moléstia émuito grave e comum em países africanos.(Carvalho et al., 1976: Van der Graaf. 198/ :Van de Vossen e Walyaro. 1(81). A espécie ('cuncphora também apresenta resistência aosnernatóide Mc/oidogl"lle exigua. .\/. tncognu«;.\ I paranacnsis e Pratvlcnchus coffea«. O portaenxerto Apoatã (IAC n.'i8) resistente a váriosnernatóides vem sendo a solução para amanutenção da cafeicultura em várias regiões doBrasi I. onde ocorre a presença de nematóides.Dessa maneira. C. cancphoru é amplamenteutíIizada no programa de melhoramento doInstituto Agronômico de Campinas. como portaenxerto ou como fonte de resistência a estasmoléstias e ncrnatóidcs. (Fazuoli. 1981: Fazuolic! at.. 1984. Carvalho & Fazuoli. 19(3).A espécie ('. rucentosa. pertencente ao gruposecundário. de reserva gênica de Coffea. cruza-sebem com (' arabica produzindo triplóidcsnaturais ou artificiais com facilidade. Tem sidoutilizada como fonte de resistência ao hichomineiro tPentcucoptcro coffccl/u) e à seca.Como o período de maturação dos frutos de ('.ruCCII10I'U. em Campinas - SP. é bastantereduzido (2 a 3 meses). esta característica vem
sendo utilizada no desenvolvimento de cultivaresde C arabica com maturação precoce.Uma outras espécie do grupo secundário dereserva gênica é C dcwcvr«! ou (' lthcrica vurde 1I'l'l'/"e i. que além de ter boa produção. possuiresistência ao agente da ferrugem. ncmatoides ehicho mineiro, Apresenta maturação hem tardia.levando um ano desde o florescimento até amaturação dos seus frutos e tem cndocarpo maisespesso. que pode se constituir em uma barreirafísica que dificultaria a penetração da broca docafé (lhpolcl1CIIIIIS hunipci) em seus frutos,
(' cOl1gclIsil'. também do segundo grupo dereserva gênica vem sendo utilizada em sua formatetraplóide. Esta espécie além de possuir
exuberante sistema radicular e boa produção, éresistente aos nernatóidcs ,\1. exigllu c ,\Iil1cogl1ilu (Fazuoli ct al.. 1(83) e a li 1'U,II<II/'I.l.
Observações atuais efetuada pelos autoresrevelam que os cafeeiros utilizados neste trabalhoeram provavelmente híbridos COI11 C. cuncphora ,A espécie C congensis, segundo Che\ alier( 1(47). em seu habitat natural fica submersa emágua por algum tempo. Este aspecto representauma vantagem. pois o excesso de um idade dosolo é prejudicial aos cafeeiros da espécie ('arabica.Outra espécie que vem sendo utilizada c ('stcnophvlla. pertencente ao grupo terciário dereserva gênica. Esta espécie é praticamente imuneao bicho mineiro. No entanto. o seu uso élimitado pela dificuldade de obtenção de sementesatravés das hihridações com (' arabica(Carvalho & Mônaco. 1(67). Neste caso éindicado o uso de métodos mais sofisticados quepossam auxiliar a utiiizaçáo dessa espécie emprogramas de melhoramento. tais como cultura 11/
\'11m de embriões ou fusão de protoplastos(Sondahl cv o... 1(80).A espécie (', salvutriv: pertencente ao grupoterciário de reserva génicn também está sendoaproveitada no programa de melhoramento doIAC visando resistência a I' coffccl!a. Cafeeirostctraplóides de (' salvotrix foram cruzados comC. arubica com a finalidade de transmitir essaresistência. No entanto. uma atcncào especialdeve ser dispensada a utilização de (', <alvutrtx,
pois a especie é altamente suscetível á saúva(M azza lera. 1991 l,Em todos os projetos em que as espécies silvestressão aproveitadas. há uma necessidade de serealizarem provas de qualidade de bebida parafins de seleção. pois apenas C arabica produzbebida considerada de boa qualidade (Teixeira cl
al.. 1974).Outra característica importante. avaliada noprograma de melhoramento do cafeeiro no IAC é
I

o teor de cafeína. que é um dos componentesquímicos de grande interesse. devido ao efeitoestimulante que apresenta. Reduzir ou aumentar oseu teor é importante para atender distintos nichosde mercados. A redução poderá ser conseguidapelo emprego. nas hibridações, de cafeeirossilvestres de Madagascar sem cafeína. ou entãopela transferência do gene recessivo laurina (Ir) deC arabica para os cultivares comerciais. umavez que esse fator reduz à metade o teor decafeína (Carvalho eI al.. 1965: Carvalho ct al..1983: Charrier & Herthaud, 1975). Odesenvolvimento de cultivares de C' cancphorasem cafeína vem também sendo estudado por Kyt:I ui (1999). Os autores têm utilizado a espécieC pscudozanguebaric nos cruzamentos. pelaausência de cafeína nas sementes. Algumasespécies do gênero Psilanthus, como J'bengalensis e J'. travancorensis. que nãopossuem cafeína em suas sementes podemtambém ser utilizadas em programa similares. Departicular interesse foi a obtenção do híbrido de ('.arubicu com J'. hengulcnsis (Charrier, 1978). Noentanto. 'a utilização de espécies de outrosgêneros no melhoramento do cafeeiro é umatarefa dit1cil e demorada pois as barreirasgenéticas são maiores. Para aumentar o teor decafeína nas sementes pode-se uti Iizar cruzamentoscom a espécie ('. canephora, onde ocorreconsiderável variabilidade para esta característica(Charrier & Berthaud. 1975).Devido ao fato da espécie C arabicu seralotctraplóide, autofértil e autógama e as outrasespécies do gênero Coffea serem diplóides.autoincornpatíveis e de polinização cruzada.ocorrem problemas no aproveitamento dasespécies silvestres. tanto no nível diplóide comotetraplóide. Apesar de haver boas possibilidadesde se utilizarem triplóides, o caminho maisseguro talvez seja o da duplicação do número decromossomos das espécies diplóides paraposterior cruzamento.Na utilização de indivíduos triplóides. emcruzamentos ou retrocruzamentos. estes deverãosempre ser uti Iizados como genitores. Para oaproveitamento no nível tetraplóide. as sementesou plántulas das espécies silvestres deverão sersubmetidas ao tratamento com colchicina para aduplicação do número de cromossomos. Nashibridações os cafeeiros tetraplóides deverão serutilizados como planta masculina. com afinalidade de evitar ou diminuir o aparecimento deindivíduos aneuplóides na descendência.Além dos inúmeros exemplos da utilização comsucesso do gerrnoplasma de Co/teu em programasde melhoramento. a reserva gênica presente emcoleções deverá ainda contribuir intensivamente
221
para o desenvolvimento de novos cultivares comcaracterísticas específicas. principalmente emrelação à resistência aos nematóides ou outrosproblemas fitossanitários, ou ainda. para atendernovas exigências do mercado consumidor.
2, BIOTECNOLOGIA MODERNA
Dentre as técnicas de biotecnologia moderna quepoderiam aj udar no programa de me Ihoramentodo cafeeiro destacam-se a cultura de tecidos. atransformação genética e as de marcadoresmoleculares.
2. L Cultura de TecidosO termo cultura de tecidos corresponde a umconjunto de técnicas que possibilitam o cultivo /11
vitro de tecidos vegetais. a partir de partes deplantas adultas. Culturas de tecidos têm sidoestabelecidas com sucesso para inúmeras espéciescultivadas e selvagens a partir dos mais variadostecidos e órgãos. como é o caso de folhas.embriões. anteras. pólen. A utilização dessastécnicas permitiu práticas diversas. comornicropropagação de genótipos de interesse e dedifícil cultivo. manutenção facilitada de bancos degermoplasrna. produção de haplóides e diplóideshomozigotos. transformação de genótipos. entreoutros. Atualmente. com o desenvolvimento detécnicas cada vez mais avançadas da biologiamolecular. há uma necessidade crescente de seestabelecer metodologias eficientes de cultura detecidos para todos os cultivares comerciaisbeneficiando amplamente os programas demelhoramento genético.As primeiras tentativas de cultivo in vitroconstituíam-se na micropropagação em cultura deex plantes. nodos vegetativos. ou meristernasapicais, Este tipo de cultura é mais eficiente paraprodução de plantas em pequena escala. como porexemplo para multiplicação de genótiposespeciais. ou para o estabelecimento emanutenção de coleções in vitro,O método de cultura deve ser cuidadosamenteescolhido de acordo com o objetivo em vista e dascond içõcs ex peri mentais prev iamcnte testadas eeficazes para cada espécie. Por exemplo. parausos de maior demanda. tais como a propagaçãode cultivares. a metodologia mais utilizada é a.cmbriogênese somática. De maneira geral. acultura pode ser iniciada a partir de um fragmentode tecido ou órgão. que em meio de cultura irá sedesenvolver em uma massa amorfa decrescimento contínuo e ind itcrenciado. Estamassa. denominada calo. pode ser mantida nesteestado indefinidamente através de repicagern, Adiferenciação em tecidos e órgãos. e a regeneração

de plantas adultas é feita através da adição dereguladores de crescimento apropriados(hormônios vegetais) ao meio de cultura. Umavariação dessa metodologia é a cultura deprotoplastos. Neste caso, a cultura é iniciada apartir de células vegetais em suspensão, quetiveram a parede celular removida. Este tipo decélula pode ser utilizada em experimentos detransformação. A diferenciação e regeneração deplantas é posteriormente realizada seguindo osprocedimentos de embriogênese somática.Um outro tipo de cultura pode também seriniciado a partir de anteras intactas ou demicrósporos. A cultura de anteras tem grandeimportância em programas de melhoramento poispossibilita o desenvolvimento de plantashaplóides. Além disso, micrósporos em culturapodem ser duplicados com a utilização decolchicina, que irão regenerar plantas diplóideshomozigotas.Em café, vários esforços têm sido efetuados parao estabelecimento do cultivo in vitro de váriasespécies do gênero Coffea. A primeira tentativabem sucedida de regeneração de plantas em C.arahica foi realizada em 1975, a partir deembriogênese somática direta (Herman & Haas.1975). Desde então vários trabalhos foramreal izados na tentativa de se estabelecer ascondições ideais de cultivo (revisto em vanBoxtel. 1994). Atualmente a regeneração deplantas adultas a partir de calos embriogênicos éobtida com sucesso em C. canephora (van Boxtel,1994). Em C. arahica a freqüência de obtençãode embriões e regeneração de plantas a partir decalos embriogênicos é baixa e depende dogenótipo (van Boxtel & Berthouly, 1996). Estaregeneração, no entanto, não foi possível quando acultura foi iniciada com protoplastos isolados apartir de folhas de C. arahica (van Boxtel. 1994).Recentemente, em outro tipo de experimento, aandrogenesis têm sido estabelecida para avariedade Catimor. Neste caso, a cultura foiiniciada com botões florais, as anteras emicrósporos foram isolados e cultivados emdiferentes meio de cultura. A regeneração deplantas a partir de células de anteras foi alcançadacom sucesso (Carneiro & Antão da Silva, 1999).Esse experimento abre uma porta importante parao melhoramento do cafeeiro, uma vez que aprodução de plantas haplóides representa umaeconomia de tempo e aumenta a eficiência nageração de híbridos.A utilização de técnicas de cultura de tecidos emcafé pode ser uma ferramenta essencial parasolução de problemas encontrados pelosprogramas de melhoramento. Um deles. a baixavariabilidade genética para resistência a moléstias.
222
pragas e nematóides presente em C. arabica, podeser potencialmente contornada através daintrodução de genes de outras espécies, viatransformação genética. Para tal, é fundamental oestabelecimento de uma metodologia eficaz, quepermita a determinação da influência de fatores,tais como tipo e idade do tecido iniciador,composição do meio de cultura, período de tempoentre a embriogênese e a regeneração de plantasadultas, entre outros, para o cultivo in vitro deespécies de Coffea. Outro aspecto que pode serresolvido com esta tecnologia é a propagação econservação de germoplasma em café.Atualmente, a propagação é feita principalmenteatravés de sementes. A utilização daembriogênese somática para a obtenção deembriões que podem ser multiplicados em largaescala em bioreatores, e posteriormentegerminados, a chamada micropropagação. já vêmsendo pesquisada. Em experimentos, plantasoriginadas a partir de embriões somáticosapresentaram desenvolvimento semelhante ao deplantas originadas de plantio por sementes(Sondahl el al.. 1999). A falta de' biofábricasespecializadas em café, o alto custo de produção eausência de dados com relação ao efeito doprocesso sobre a qualidade da bebida representamalguns dos fatores limitantes à utilização damicropropagação em cafeeiros.
2,2. Transformação genéticaEntende-se por transformação genética amodificação de genótipos através da introduçãode genes específicos, visando uma alteraçãoprogramada do indivíduo transformado. No casode transformação de tecidos vegetais. estes genespodem estar relacionados com características deinteresse agronômico, tais como resistência adoenças e pragas. controle da síntese dehormônios e substâncias de interesse comercial.Atualmente. com a constante divulgação deorganismos transformados, ou geneticamentemodi ficados, a compreensão do potencial dasrnetodologias utilizadas para transformação setorna extremamente importante.O método de transformação utilizado em plantasvaria de acordo com as característicasmorfológicas e fisiológicas da espécie em questão.O método mais eficiente e de menor custo é autilização da bactéria Agrobacterium tumefaciens.Esta bactéria é um patógeno natural de plantas,que promove o crescimento desordenado decélulas vegetais infectadas, levando à formação degalhas. Essa alteração é resultante da ação degenes encontrados em um plasmídio da bactéria eque codificam reguladores de crescimento.Durante a infecção, esse genes são transferidos
I!

(
para o genoma da planta, sendo assim ativados.Para sua utilização em experimentos detransformação, os plasmídios de A. tumefacienssão modificados em laboratório, recebendo o genede interesse a ser introduzido na planta, e tambémgenes marcadores para a seleção do materialtransformado. A transformação através destametodologia é normalmente feita em célulasvegetais em cultura, as quais serão depoisselecionadas para a presença do plasmídiobacteriano e regeneradas em plantastransformadas. Em café, plantas têm sidotransformadas através desta metodologia demaneira eficiente. O gene CryIA(c) que confereresistência a P. coffeella, isolado de Bacillusthuringiensis, foi introduzido em C. arabica e C.canephora e as plantas regeneradas estão sendotestadas para a resistência ao bicho-mineiro(Leroy et al., 1999).Uma outra metodologia utilizada com sucessopara transformação de café é a biobalística oubombardeamento de tecidos vegetais. Neste caso,os genes de interesse e os genes marcadores sãointroduzidos em tecidos vegetais através de umaforte pressão, exercida por gás hélio comprimido.Esse bombardeamento é realizado em umequipamento específico, onde o DNA, associadocom micropartículas, é colocado na direção dojato de gás e direcionado sobre o tecido a sertransformado. Além de uma alta eficiência detransformação. outra vantagem deste método éque o tecido transformado não sofre grandesdanos mecânicos. Até o momento, testesrealizados com vários tecidos de C. arabica, taiscomo folhas intactas, células em suspensão ecalos embriogênicos foram promissores (vanBoxtel, 1994). Nestes experimentos foi possívelse detectar a expressão transiente do genemarcador GUS.O impulso para o desenvolvimento demetodologias para transformação veio a partir doconhecimento cada vez mais complexo 'daestrutura molecular do gene. As técnicas debiologia molecular possibilitaram isolamento eclonagem de genes, identificação de regiõesreguladoras e mecanismos de ação gênica. Emcafé, alguns exemplos representam possibilidadesde utilização dessa tecnologia em melhoramento.Em um deles, a resistência ao bicho-mineiro podeser obtida através da utilização de genes isoladosde B. thuringiensis. Estes genes, relacionadoscom a síntese de toxinas com poder inseticida,foram transferidos para células em cultura (Leroyet al.. 1999). Em um outro tipo de experimento, ogene de café que codifica a xantosina- N7metiltransferase. enzima envolvida na biosíntesede cafeína, foi introduzido em células. Porém, o
223
sentido da seqüência foi invertido, de maneira queo RNA produzido será complementar ao RNA dogene normal, e irá portanto se parear com este.Desse modo, não haverá síntese da enzima, econsequentemente a síntese de cafeína pode serbloqueada (Moisyadi et al., 1999).A agricultura atual exige cada vez maisvariedades com uma maior plasticidadeagronômica. As novas técnicas de biologiamolecular, associadas com cultura de tecidos esistema de transformação eficazes, permitem aobtenção de genótipos bem específicos, demaneira controlada, isto é, sem a introdução decaracterísticas indesejáveis. No caso do café, abiotecnologia pode contribuir muito emprogramas de melhoramento genético pois aspesquisas feitas até o momento demonstram aexistência de um potencial para a transferência degenes de interesse, de maneira mais rápida, ondenão seria necessário a realização deretrocruzamentos. No entanto, o sucesso destetipo de abordagem em melhoramento depende deum conhecimento profundo não só dos genótiposa serem avaliados, mas também dos mecanismosde ação e tipo de herança dos genes de interesse.Além disso, um programa de seleção e análise deprogênies é fundamental para que as variedadesobtidas possuam realmente as característicasdesejadas.
2.3, Marcadores MolecularesO desenvolvimento da técnica da eletroforese, quepossibilita a separação e visualização demoléculas diversas, representou a possibilidade dese conhecer e avaliar o genótipo de indivíduos. Aanálise de padrões eletroforéticos permite acaracterização de locos gênicos, com relação aoseu padrão de herança, número e . freqüência dealelos em uma população. Desta maneira, épossível a identificação de marcadoresmoleculares, associados a genótipos específicos.Um marcador molecular pode ser definido comotodo e qualquer fenótipo originado da expressãode um gene ou de um fragmento específico deDNA (Ferreira & Grattapaglia, 1996). Osprimeiros marcadores utilizados em análisesgenéticas foram as isoenzimas (Tanksley &Ortorn, 1983). Atualmente uma gama demarcadores moleculares têm sido identificados.através de técnicas que permitem uma amplacaracterização do polimorfismo genético existenteem diferentes variedades de plantas. Essesmarcadores representam uma ferramentaimportante em programas de melhoramentoassistido e na identificação de variedades. Váriosprogramas de melhoramento de plantas cultivadastêm utilizado os mapa genético-moleculares

gerados por esses marcadores, em conjunto commétodos tradicionais de melhoramento, para aseleção de cultivares de interesse econômico(Rafalski & Tingey, 1993; Lashermes et al.,1997 a).O primeiro marcador de DNA desenvolvido foi oRFLP (Restriction Fragment LengthPolymorphism), cujo polimorfismo está baseado nocomprimento dos fragmentos digeridos porenzimas de restrição (Botstein et aI. 1980). Odesenvolvimento do princípio de reações depolimerização em cadeia (PCR - Polymerisation ofChain Reaction) possibilitou a identificação demarcadores obtidos a partir da amplificação deseqüências aleatórias no genoma. Estas sequênciasamplificadas variam em tamanho e númerodependendo do genótipo analisado. Dessa forma,populações que apresentam grande variabilidadegenética possuem padrões diversificados demarcadores. Já populações com baixa variabilidadegenética apresentam padrões constantes demarcadores. Os métodos de obtenção demarcadores mais utilizados em plantas são oRAPO (Randon Amplified Polymorphic DNA),inicialmente utilizado por Williams et al.(1990) eWelsh & McClelland (1990); o AFLP (AmplifiedFragment Length Polymorphic), desenvolvida eaplicada por Vos et aI. (1995); e os microsatélites(Tautz, 1989; Weber & May, 1989). De maneirageral, todos estes métodos buscam avaliar avariabilidade genética através do mesmo princípio,que é a existência de polimorfismos em regiões doDNA, que são ou não conservados em umadeterminada população. A diferença entre eles estána especificidade e abrangência de detectar essepolimorfismo, e principalmente no custoexperimental. Assim, marcadores de RAPO são osmais simples e baratos de serem obtidos; porém emdeterminadas situações, como é o caso de espéciescom baixa variabilidade genética, não conseguemidentificar polimorfismos que permitam aidentificação de diferentes genótipos. Já osmicrosatélites têm uma boa abrangência, porémcom custo elevado.Vários experimentos têm sido conduzidos natentativa de se estabelecer um mapa genéticomolecular para as espécies de café. No entanto. avariabilidade genética encontrada em umapopulação de C arabica não é muito grande, oque dificulta a identificação de marcadoresgenéticos necessários em programas demelhoramento. Diversos cultivares de C. arabicatêm sido estabelecidos até o momento, através demel horamento genético clássico, resultantes decruzamentos com outras espécies do gênero Coffea(Sondhal & Lauritis, 1992) . Esses cruzamentosinterespecíficos. associados com uma taxa esperada
224
de alogamia levam a acreditar que exista um certonível de polimorfismo molecular entre asvariedades de C. arabica .O estabelecimento de mapas genéticos de cafébaseados em isoenzimas e RFLPs, tanto de DNAtotal quanto de DNA de cloroplasto, não forambem sucedidos devido ao baixo índice depol imorfismo observado (Paillard et aI., 1996;Lashermes et al., I996a). Um outro marcadoranal isado para caracterizar as di ferentes espéciesde café foi a seqüência entre as regiõescodificadoras dos genes 18s e 26s. conhecida comoITS (internai transcribed spacer). No estudo deLashermes et aI. (1997b), várias espécies evariedades de Coffea foram avaliadas quanto aopolimorfismo nas ITS. Os resultados indicam queas ITS podem representar um marcador inter eintraespecífico. uma vez que polimorfismos nestesníveis foram observados. Até o presente a técnicade RAPO foi a que mais permitiu oestabelecimento do nível de diversidade genéticaentre vários cultivares pertencentes a C. arabica eC. canephora. determinando marcadoresespecíficos para as variedades analisadas(Lashermes et al.i, I996b). No entanto. a variaçãointraespecífica não foi estabelecida para C. arabica(Orozco - Castillo et al., 1994). Baseando-se nestasinformações é possível inferir que para aidentificação de polimorfismos em C. arabicadevem ser utilizadas metodologias que permitam arevelação de um número maior de locos, como é ocaso dos AFLPs e microsatélites.Além da sua utilização para identificação deespécies e variedades, os marcadores podemrepresentar uma ferramenta importante emprogramas de melhoramento. Neste caso. aidentificação de marcadores moleculares que cosegreguem com características agronômicasdesejáveis, tais como resistência a doenças epragas, produtividade, qualidade de bebida. podeservir como instrumento de seleção assistida. Estetipo de análise é importante em espécies comciclo de vida longo. como o café, pois aidentificação das progênies desejadas pode serrealizada nos estágios iniciais de desenvolvimentoda planta. Desta forma, não é necessário que todasas progênies sejam plantadas e avaliadas emcampos experimentais, o que representa umagrande economia de tempo, espaço físico erecursos.
3. POSSIBILIDADES DO USO DABIOTECNOLOGIA MODERNA EMCAFEEIROS.
3.1. Exemplos de utilização da cultura detecidos
II
j

3.1.1. Cultura de folhasA cultura de folhas do cafeeiros poderia serutilizada para estudos de variação somaclonal oumicropropagação. No entanto a utilização daembriogênese somática direta através debioreatores de imersão temporária ou permanente,biofábrica, seria interessante para propagação declones de C. canephora livres de moléstias, deplantas especiais geralmente híbridos comresistência múltipla (à ferrugem, aos nematóidese ou ao bicho mineiro), de clones de C.canephora com resistência a várias espécies e ouraças de nematóides.
3.1.2. Cultura de anterasA cultura de anteras seria interessante para aprodução de plantas haplóides. No caso deespécies diplóides poderiam ser obtidasposteriormente plantas diplóides homozigotaspossibilitando análises genéticas nestas espéciesautoincompatíveis e produção de híbridos FI de C.canephora 100% resistentes a nematóides.
3.1.3. Cultura de embriõesEmbriões originados a partir de cruzamentosinterespecíficos ou intergenéricos e queapresentam dificuldades em seu desenvolvimentonatural poderiam crescer e tornarem plantasquando colocados em culturas in vitro. É o caso,por exemplo, de cruzamentos de C. stenophyllaou. P. bengalensis com C. canephora ou C.arabica (Carvalho & Mônaco, 1967; Charrier,1978).
3.1.4. Preservação de células e propágulosA preservação de células ou propágulos relacionase com a conservação in vitro de recursosgenéticos de espécies. híbridos interespecíficos,haplóides, etc, mantendo assim um banco degermoplasma em condições de laboratório.
3.2. Transformação de plantasA transformação uti lizando genes de bactérias,fungos e vírus poderia ser utilizada em caféprincipalmente nos casos da transferência decaracterísticas (genes) não encontradas emespécies selvagens de café. Um exemplo é o casoda resistência à broca do café (H. hampei), poisaté o presente não foram identificadas fontes deresistência a esta praga no germoplasma de café.Há informações de que pesquisadores daColômbia do CENICAFÉ conseguiram clonargenes de Streptomyces sp. que produzemquitinase. Esta deve atuar na quitina do besouro eportanto poderia ser usado no controle da brocado café.
225
Como existem fontes de resistência para outraspragas nas espécies diplóides estes genes estãosendo transferidos para C. arabica nomelhoramento clássico (cruzamentos,autofecundações e retrocruzamentos). Comoexemplo pode-se citar o desenvolvimento decultivares com resistência e tolerância a P.co.ffeella (bicho mineiro) já em fase final deavaliação (Guerreiro Filho et ai., 1999). Apesardisso, alguns pesquisadores estão incorporandogenes Bt em cultivares de café (Leroy et ai. ,1999).Um outro uso de transformação está relacionadocom a obtenção de cafeeiros tolerantes aosherbicídas. Neste caso o interesse seria maiorpara as firmas que vendem seus produtos.É importante ressaltar o uso da transformação semque esta envolva a introdução de genes estranhosao genoma do café como é o caso da manipulaçãogenética da síntese do etileno (Neupane et al..1999) do bloqueio do metabol ismo da síntese decafeína, através da inserção de seqüênciasinvertidas (Moisyadi et ai., 1.999) que neste casoestariam relacionados com a qualidade doproduto.Finalmente o uso da transformação terá umgrande avanço em café quando for possível dessesconhecer o genoma de C. arabica e espéciesdiplóides e houver possibilidades de clonar genesespecíficos de espécies diplóides que poderão sertransferidos para C. arabica. A utilização dessescultivares não representaria maiores riscos comoacontece com a introdução de genes de bactérias.fungos e vírus, cujos danos ao meio ambiente e àsaúde humana não foram ainda suficientementeavaliados.
3.3. Utilização de marcadores molecularesComo exemplos da utilização de marcadoresmoleculares em café pode-se citar:(a) Caracterização de cafeeiros (espécies.híbridos, cultivares, clones, etc.), visandoconhecimento do material e proteção comidentificação mais segura. Na caracterização decafeeiros tem-se utilizado descritores relativo àscaracterísticas morfológicas da planta, folhas,flores, frutos e sementes. Característicasagronômicas, tecnológicas e resistência outolerância a fatores adversos (moléstias, pragas,nematóides e ambientais) também vêm sendoutilizadas. Atualmente o uso de técnicasmodernas de extração de DNA e de marcadoresmoleculares poderá permitir uma melhorcaracterização de cafeeiros mutantes de C.arahica e principalmente das espécies diplóidesde Coffea e outros gêneros.

a) Seleção assistida utilizando marcadoresmoleculares. Neste caso procura-se associarresistência a ferrugem. nernatóides, maturação.propriedades organolépticas, etc comcaracterísticas de melhor definição e mais fácilavaliação.b) Análises genômicas e avaliação da divergênciagenética. Para tal pode-se pensar em estabelecer ogenoma de C arabica e principais espéciesdiplóides ou o genoma de alguma moléstia. pragaou nematóide importante.c) A hibridação somática entre espécies de cafétambém seria interessante e possível com o uso datécnica de marcadores moleculares.
4. CONSIDERAÇÕES FINAIS
Devido ao fato de que os cultivares de cafédisponíveis no mercado são altamente produtivos.pode-se afirmar que as tendências atuais nomelhoramento do cafeeiro estão relacionadas comresistência a moléstias. pragas e nernatóides,resistência e ou tolerância a condições adversas doambiente. melhor sistema radicular. a arquiteturada planta. com a redução do seu porte e umamelhor qualidade da bebida. Na maioria doscasos o melhoramento clássico foi e contínuasendo importante. No entanto a possibilidade douso da biotecnologia moderna é evidente com autilização de culturas de tecidos. de marcadoresmoleculares e de transformação. O uso datransformação em café utilizando genes clonadosde bactérias. fungos e vírus deve ser analisadosob dois aspectos. o científico e o de interesse dacomunidade. O aspecto científico refere-se a umavanço inevitável nas próximas décadas. mas oaspecto de interesse da comunidade requer maisretlexões. pois no momento o ambiente não éfavorável à utilização deste tipo de transformação.Por outro lado a utilização de cultura de tecidos eespecialmente de marcadores molecularesocupará. sem dúvida nenhuma. uma posição dedestaque nos trabalhos de melhoramento docafeeiro e poderá proporcionar grandes avançosem um futuro bem próximo.
5. REFERÊNCIAS BIBLIOGRÁFICAS
Berthaud, J. 1980. L' incompatibilite chez Coffeacanephora methode de test et determinismegenetique. Café. Cacao, Thé 24: 267-274
Bettencourt. A. J. 1973. Considerações geraissobre o híbrido do Timor. Campinas. InstitutoAgronômico. Circo n." 23. 20 p.p.
Bettencourt, A. J. & Carvalho. A. 1968.Melhoramento do cafeeiro visando resistênciaà ferrugem. Bragantia. 27:35-68.
226
Bridson, D. M. 1982. Studies in Coffea inPsilanthus (Rubiaceae subfam.Cinchonoideae) for part 2 of Flora of TropicalEast África: Rubiaceae. Kew Bulletin 36 (4):817-859.
Bridson, D. M. 1994. Additional notes onCoffea (Rubiaceae) from Tropical East África.Kew Bulletin 49 (2): 331-342.
Bridson. D. M 1987. Nomenclatura notes onPsilanthus, including CofJea sect. ParacofJea(Rubiaceae tribe Coffeae). Kew Bulletin LR(2) 453-460.
Bridson D. M. & Verdcourt, B. 1988. "Flora ofTropical East África - Rubiceae (Part 2).Polhill R. M. (eds), 727 pp.
Botstein, D.; White. R.L.; Skolnick, M. &Davis. R. W. 1980. Construction of a geneticlinkage map in man using restriction fragmentlength polymorphisms. Am. J. Hum. Genet.32:314-321.
Care lli, M. L.; Lopes. C. R. & Mônaco. L. C.1974. Chlorogenic acid content in species ofCoffea and selections of C. arábica.Turrialba, 24: 398-40 I.
Carneiro. M. F. & Antão da Silva. C.M. 1999.Androgenesis in some catimor progenies..ut:International Conference on Coffee Science(Helsinki). Paris:ASIC. PA 741.
Carvalho. A. 1958. Advances in coffeeproduction technology. Recent advances inour knowledge of coffee trees. 2 Genetics.Coffee and Tea Industries. New York, ~:30
36Carvalho. A. 1960. Genética de Coffea. XXV.
Ligação genética dos fatores anormais eanômala. Bragantia. Campinas.19:CXCICXCV. (Nota 38).
Carvalho. A. & Fazuoli, L. C. 1993. Café. In:Furlani. A. M. C. & Viégas, G. A. eds. Omelhoramento de plantas no InstitutoAgronômico. Campinas. InstitutoAgronômico. Capo 2. p. 29-76.
Carvalho. A.: Fazuoli, L. C. & Mazzafera, P.1986. Melhoramento do café laurina. In.Reunião Anual da Sociedade Brasileira para oProgresso da Ciências. 38. Curitiba. Anais.p.908.
Carvalho. A. & Krug, C. A. 1949. Genética deCoffea XII. Hereditariedade da cor amarela dasemente. Bragantia. Campinas. 2 :193-202.
Carvalho. A.; Medina 1 ilho, H. P. & Fazuoli, L.C. 1984a. Evolução e melhoramento docafeeiro. In: Aguiar-Parecin, M. L. R. :Martins. P. S.; Bandel. G. Eds. Tópicos deCitogenética e Evolução de Plantas. IColóquio sobre Citogenética e Evolução de

Plantas, Piracicaba, São Paulo. RevistaBrasi leira de Genética. p. 215-234.
Carvalho, A.; Medina Filho. H. P.; Fazuoli, L.C. & Costa. W. M. 1984b. Números de locose ação de fatores para porte pequeno emCoffea arabica.. Bragantia, Campinas. 43(2):421-442.
Carvalho, A.; Medina Filho, H. P.; Fazuoli, L.c.. Guerreiro Filho. O. & Lima, M. M. A1991. Aspectos genéticos do cafeeiro (GeneticAspects of the coffee tree). Rev ista Brasi I.Genet., 14(1): 135-183.
Carvalho, A. & Mônaco. L. C. 1962. Naturalcross pol1ination in Coffea arabica. Proc XVIIntern. Hort. Congr. Brussels ~: 447-449.
Carvalho. A & Mônaco, L. C. 1967. Geneticrelaonships ofselected Coffea species. Ciênciae Cultura. 1.21):161-165.
Carvalho. A. & Mônaco, L. C. 1972.Transferência do fator Caturra para o cultivarMundo Novo de Coffea arabica. Bragantia,Campinas, 31 (31): 379-399.
Carvalho, A.; Mônaco, L. C. & Van der Vossen,H. A. M. 1976. Café Icatu como fonte deresistência a Colletotrichum coffeanum.Bragantia, 35: 343-347.
Carvalho, A; Tango, J. S. & Mônaco, L. C.1965. Genetic control of caffeine content incoffee. Nature, 205: 314.
Carvalho. A; Sondahl, M. R. & Sloman, C1983. Teor de cafeína em seleções de café.Congresso Brasileiro de Pesquisas Cafeeiras,Poços de Caldas. MG .. 10:111-113.
Charrier, A. 1978. La structure genetique descafeiers spontanes de I' region Malgashe(Mascarocoffea). Leur relations avec lescafeiers dorigine africaine (Eucoffea).Memories ORSTOM, Paris. 97. 123 p.p.
Charrier, A. & Berthaud, J. 1975. Variation dela teneur en caffeine dans le geme Coffea,Café. Cacao, Thé 1.2 (4): 251-264.
Charrier, A & Berthaud. J. 1985. Botanicalc1assification of coffee. In: Clifford, M. N.& Willson, K. C. Eds Coffee. Botany.Biochemistry and production of beans andbeverage. Westport, connecticut. The AviPublishing, Inc. p. 13-47.
Chevallier, A. 1947. Lés caféiers du globe. 111.Systhematique des caféiers. Maladies etinsects nuisibles. Encyclopedie biologique 18: 1-256 Fascicule 111. Paul Lechevalier, Paris.
Conagin. C. H. T. M. & Mendes. A J. T. 1961.Pesquisas citológicas e genéticas de trêsespécies de Coffea - autoincompatibilidade emC. canephora. Bragantia. 20: 787-804.
Cramer, P. J. S. 1957. A review of literature ofcoffee research in Indonésia. In: Wellman. F.
227
ed. Muc. Publ. N.o 15. Turrialba, Inter.Am. First of Agric. Sei. P. 1-262.
Devreux, M.; Vallaeys, G.; Pochet, P. &Gi lles, A. 1959. Researches surI 'autosterilite du caféier Robusta (Coffeacanephora Pierres, Publ. INEAC 78, 44 p.p.
Fazuoli, L. C. 1981. Resistance of coffee to theroot-knot nematode species Meloidogyneexigua and M. incognita. Col1oqueInternational sur protection des culturestropicales, Lyon, France. p.57.
Fazuoli, L. C. 1986. Genética e Melhoramentodo Cafeeiro. In: Rena. A B.; Malavolta, E.; Rocha, M.; Yamada, Y. Editores. Culturado Cafeeiro. Piracicaba. Potafos, p. 87-113.
Fazuoli. L. C. 1991. Metodologias, critérios eresultados da seleção em progênies do caféIcatu com resistência a Hemileia vastatrix.Campinas. 322 p. (Tese de Dourtorado-Universidade Estadual de CampinasUNICAMP).
Fazuoli, L. C.; Costa. W. M.; Gonçalves, W. &Fernandes, J. A. R. 1983. Identificação deresistência em Coffea canephora e C.congensis ao nematóide Meloidogyneincognita, em condições de campo. Ciência eCultura, São Paulo, 35 (7):20 (Suplemento).
Fazuoli. L. C.; Costa, W. M.; Gonçalves, M.;Lima, M. M. A. & Fernandes, J. A R. 1984.Café Icatu como fonte de resistência e/outolerância ao nematóide Meloidogyneincognita, In: Congresso Brasileiro dePesquisas Cafeeiras, 11, Londrina. Resumos.Rio de Janeiro. IBC/GERCA P.247-248.
Fazuoli, L. c.. Medina, H. P.; Guerreiro Filho.O., Lima, M. M. A. Silvarolla, M. B..Gallo. P. B. & Costa. W. M. 1996. Obatã(IAC 1669-20 e Tupi (IAC 1669-33).cultivares de café de porte baixo e resistentes aferrugem.Congresso Brasileiro de Pesquisas Cafeeiras,12°. Águas de Lindóia, SP, p. 149-150.
Ferreira. M.E. & Grattapaglia, D. 1996.Introdução ao uso de marcadores molecularesem análise genética. 2".edição. Embrapa. p.p.110.
Guerreiro Filho, O.; Silvarolla, M. B. & Eskes,A. B. 1999. Expression and mode ofinheritance of resistance to leaf miner.Euphytica, I05( I ):7-15.
Harlan, J. R. & De Wet. J. M. J. 1971. Towarda rational c1assification of cultivated plants.Taxon. 20(4): 509-517.
Herman, E. & Haas, G. 1975. Clonal propagationof Coffea arabica L. from callus culture.HortScience 10.588-589.

rAPAR. Café IAPAR 59 1993. Londrina, PR..lAPAR. Folder.
Krug, C A.; Carvalho, A. 1939. Geneticalproof of the existence of Cojjea endosperm.Nature, London, 144: 515.
Krug, C A.; Carvalho. A. 1951. The geneticsof Coffea. Advances in Genetics, New York,4: 127-158.
Krug, C A.; Carvalho. A. & Antunes Filho. H.1959). Genética de Coffea, XXI.Hereditariedade dos caracteres de Coffeaarabica L. var, laurina (Smeathman). DCBragantia. Campinas. U: 247-255.
Ky, C-L.; Lovarn. J.; Akaffou, S.; Guyot, 8.;Chrestin. H.. Charrier, A.; Harnon, S. &Noirot, M. 1999. The use of genetic resourcesfor coffee cup taste improvement. InInternational Seminal' on Biotecnology in thecoffee agroindustry. 3. Londrina, p. 28.(Resumos).
Lashermes, P.; Cros, J.; Cornbes, M.C:Trouslot, P.: Anthony, F.. Hamon, S. &Charrier. A. I996a. Inheritance and restrictionfragment lenght polymorphism of chloroplastDNA in the genus Coffea L. TheQ r, Appl.Genet. 93: 626 -632.
Lasherrnes, P.. Trouslot, P.. Anthony, F..Cornbes, M.C & Charrier, A. 1996b. Geneticdiversity for RAPD markers betweencultivated and wild accessions of Coffeaarabica. Euphytica 87: 59 - 64.
Lashermes, P.: Agwanda, CO.: Anthony, F.;Cornbes, M.C; Trouslot, P. & Charrier. A.1997a. Molecular marker-assisted selection: apowerful approach for coffee improvement.171h International Conference on CoffeeScience (Nairobi). Paris: ASIC 474-480.
Lasherrnes. P.: Combes, M.C; Trouslot, P. &Charrier, A. '997b. Phylogenetic relationshipsof coffee- trees species (Coffea L.) as inferredfrom ITS sequences of nuclear ribossomalDNA. Theor. Appl. Genet. 94: 947 - 955.
Leroy, J. F. 1980. Evolution et taxogenese chezles caféiers. Hypothese SUl' leur origine.Comptes Rendus de Ia Academie des Sciences,Paris. 291 : 593-596.
Leroy. J. F. 1982. L'Origine Kenyane du gemeCoffea L. et la radiation des espécies àMadagascar. Colloque ScientitiqueInternational SUl' le Café. Salvador. Bahia.Brasil. 10:413-442.
l.eroy, T.: Henry. A.M.: Philippe. R.: Royer.M.: Deshayes, A.: Frutos. R.; Duris, D.;Dufour. M.; Tesserreau. S.; Jourdan, 1.;Bossard. G.: Lacombe. C & Fenouillet. C1999. Genetically modified coffee trees forresistance to coffee leaf miner. Analysis of
228
gene expression, insect resistance andagronomic value. 18th International Conferenceon Coffee Science (Helsinki). Paris:ASIC. B209.
Longo. C R. L. 1972. Estudo de pigmentosflavonóides e sua contribuição á filogênia dogênero Coffea. Piracicaba. 91 p. p. (TeseDoutorado-Esc. Sup. Agric. Luiz de Queiroz~ Universidade de São Paulo. Brasil).
Maglio, C A. F. P. 1983). Morfologia doscromossomos nucleolares em fase depaquiteno no gênero Coffea L. Campinas. 92p.p. (Tese de Mestrado ~ UniversidadeEstadual de Campinas - UNICAMP).
Mazzafera. P. 1991. Análises químicas emfolhas de cafeeiros atacados por Atta spp.Revista de Agricultura. Piracicaba. 66( 1):3345.
Medina. D. M. 1972. Caracterizaçãomorfológica de híbridos interespecíficos deCoffea. Piracicaba. I J J p.p. (Tese deDoutorado - Esc. Sup. Agric. Luiz de Queiroz- Universidade de São Paulo.
Medina Filho. H. P.; Carvalho. A.; Fazuoli, L. C1982. Coffea bengalensis and Ctravancorensis: espécies diplóides,autoincompatível e autógamas de Coffea.Ciência e Cultura. Reunião Anual da SBPCCampinas. São Paulo. 34: 713 (Resumo).
Medina Filho. H. P.; Carvalho. A.; Sondahl, M.R.: Fazuoli, L. C & Costa. W. M. 1984.Coffee breeding related evolutionary aspects.In: Plant Breeding Reviews (J. Janick. ed.).Avi Publish. Co. Connecticut, USA V. :2 p.157-193.
Mendes. A. J. T. 1958. Avances in CoffeeProduction Technology: Recent advances inour knowledge of coffee trees. 3 - Cytoiogy,Coffee and Tea Industries 81: 37-41.
Moisyadi, S.: Neupane, K.R.; Stiles, J. I. 1999.Cloning and characterization of xanthosineN7-methyltransferase. the first enzyme of thecaffeine biosynthetic pathway. 181h
lnternational Conference on Cotfee Science(Helsinki). Paris:ASIC B 214.
Mônaco. L. C: Carvalho. A.; Fazuoli, L. C1974. Mel horamento do cafeeiro.Germoplasma do café Icatu e seu potencial nomelhoramento. In: Congresso Brasileiro dePesquisas Cafeeiras. 2. Poços de Caldas.Resumos. Rio de Janeiro. IBClGERCA. p.103.
Moraes. S. A.; Sugirnori, M. H.; Thomaziello.F. M. & Carvalho, P. T. C 1974.Resistência de cafeeiros a Pseudomonasgarcae. In: Congresso Brasileiro de Pesquisas

•Ii
Cafeeiras, 2.. Poços de Caldas - MG. 1974.Resumos... Rio de Janeiro. IBC p. p. 183.
Neupane, K. R.; Moisyadi & Stiles, l I. 1999.Cloning and characterization of fruit-expressedACC synthase and ACC oxidase genes fromcoffee. 18 Ih International Conference onCoffee Science. (Helsinki) Paris : ASIC B213.
Orozco - Castillo, C; Chalmers.Kf: Waugh, R.& Powel, W. 1994. Detection of geneticdiversity and selective gene introgression incoffee using RAPD markers. Theor. Appl.Genet. 93: 934- 940.
Paillard. M.; Lashermes, P. & Pétiard. V. 1996.Construction of a molecular linkage map incoffee. Theor. Appl. Genet. 87: 43 ·47.
Rafalsky, J.A. & Tingey, S.V. 1993. Geneticdiagnostics in plant breeding - RAPDs.microsattelites and machines. Trends Genet. 9:175-280.
Rijo. L. 1974. Observações cariológicas nocafeeiro Híbridos de Timor. Portugaliae ActaBiologica. Lisboa. 13: 157-168
Sondahl. M. R.; Chapman, M. & Sharp, W. R.1980. Protoplast Iiberation, cell wallreconstituition and callus prol i feration inCoffea arabica callus tissues. Turrialba, 30(2): 161-165.
Sondahl. M.R. & Lauritis, J.A. 1992.Coffee. In:Biotechnology of ~rennia~uit_---.fr:-o~
Biotechnolgy in agriculture 8: 40 I -420.Sondahl, M.R .. Sondahl. CN. & Gonçalves. W.
1999. Micropropagação do cafeeiro. !Encontro sobre. Produção de Café comQtJalidade. (Viçosa) Ed. Laércio Zambolin.228 - 240.
Tanksley, S.D. & Orton, LI. 1983. lsozymes inPlant Genetics and Breedi!1-&..._Elsevier.Amsterdam, Part A. pp.516. Part B. pp.472.
Tautz, D. 1989. Hypervariability of sim piesequences as a general source of polymorphicDNA markers. Nucleic Acid Res. 17: 64636471.
Teixeira. A. A.: Carvalho. A.: Fazuoli. L. C &Mônaco. L. C 1974. Avaliação da bebida dealguma espécies e híbridos interespecíficos deCoffea. Ciência e Cultura. 26(7): 565.
229
Van Boxtel, J. 1994. Studies on genetictransformation of coffee by usingelectroporation and the biolistic method. Teseapresentada na Universidade de Wageningen.pp125.
Van Boxtel, J. & Berthouly, M. 1996. Highfrequency somatic ernbryogenesis from coffeeleaves. Plant Cell Tissue ~Organrl!!jul"e 44:7-17.
Van de Graaf, N. A. 1981. Selection of Arábicacoffee types resistant to coffee berry disease inEtiopia. Meded. Landbouwhogeschool.Wageningen. [1: 11.
Van der Vossen, H. A. M. & Walyaro, D. .I.1980. Breeding for resistance to coffee berrydisease in Coffea arabica L. 11. Inheritance ofthe resistance. Euphytica, 29:777-791.
Van der Vossen, H. A. M. & Walyaro, D. J.1981. The coffee breeding programrne inKenia: A review of progress made since 1971and plan of action for the coming years.Kenya Coffee. 46(541)
Vos. P.: Hogers. R.; Bleeker, M.: Reijans, M.:Van de Lee, Hornes, M.; Fritjers, A.; Pot, J.:Pelernan, i., Kuiper, M. & Zabeau, M. 1995.AFLP: a new technique for DNAtingerprinting. I'Jucleiç Acid Res. 23: 44074414.
Weber, J.K. & May. P.E. 1989.Abundant class ofhuman DNA polyrnorphisms which can betyped using the polymerase chain reaction.Am. J. Hum. Genet. 44: 388-397.---------
Welsh. l & McClelland. M. 1990.Fingerprinting genomes using PCR witharbitrary primers. Nucle~_~~~<:U~'f:~._18:7213 7218.
Williams. lG.K.: Kubelif. A.R.: Livak, K..I.:Rafalsky. lA. & Tingey, S.V. 1990. DNApolymorphisms ampl ified by arbirrary primersare usefulas genetic markers, Nuçleic AcidRes. 18: 6531-6535.
Wrigley. G. 1988 Coffee. (Tropical Agricultureseries). Logman Scientitic & Tecnical.USA. 612 p.


231
EL MEJORAMIENTO GENÉTICO EN AMÉRICA CENTRAL
Bertrand BI, Etienne H. I .Santacreo R.', Anzueto F.'. Anthony F.'
'Programa Cooperativo Regional para el Desarrollo Tecnológico y Modernización de la Calicultura en Centroamérica.República Dominicana y Jamaica. CIRi\D/PROMECi\FE/ICi\FE/IICi\. AP 55-2200. Coronado. Costa-Rica.fax:(506)2294689.IHCAFE. Edilicio Banco Atlántico Centro. Tegucilgalpa HondurasANACAFE. Calle O-50. Zona14, Guatemala IRD/CA Tll-. CATIE. Turrialba. Costa Rica.e-rnail: benoiLbertrand{Ú;cirad.fr
I-INTRODUCCIÓN
Del siglo XVII hasta hoy, los objetivos de la seleccióndei café Arábica en América Latina cambiaron paraadaptarse a las exigencias de nuevas prácticasculturales y para com batir nuevas enfermedades. Laxpansión mundial de C arabica. basada en unadiversidad genética reducida favoreció la expansiónparalela de parásitos graves como son la Roya en Asiay América o la antracnosis dei fruto ("eBD·). enprogreso constante en Africa. Paralelamente, Iamonocultura y el abuso de pesticidas han incrementadolos problemas causados por el parasitismo telúrico.Recientemente, Ia tendencia ai aumento dei consumode café de alta calidad obliga a una selección másestricta hacia la calidad dei producto.Nuestro propósito es mostrar cuáles fueron lasestrategias de selección empleadas en América Central,describir las principales variedades obtenidas y hastadonde el mejoramiento genético puede responder a losnuevos desafios de la caficultura del siglo XXI.
2-EXPLOTACIÓN DE LA BASE GENÉTICAINTRODUCIDA EN EL SIGLO XVIII
eran débiles, y casi desaparecian en ausencia defertilizante. Como lo subraya Castillo (1990). "Ianecesidad básica dei cultivo de café era un cambio enlas prácticas culturales, no un cambio de variedad". Apartir de los anos 50. los productores empezaronprogresivamente a adoptar la técnica de plenaexposición solar con altos niveles de fertilizantes(Cowgi!l. 1958; Uribe. 1958). De ahí surgió el interésdeI cambio de material vegetal.A partir de los anos 1930. se habia iniciado unprograma de selección en Brasil dentro de la poblacióndei Borbón (Mendes, 1939). En EI Salvador tarnbién,se seleccionaron líneas de Borbón más productivas(Tekisic). Debido a la poca variabilidad de lapoblación Típica (Castillo, 1984). y a su inferioridadproductiva en cornparación con las \ineasseleccionadas dei Borbón. se abandonóprogresivamente la difusión de este material. Hoy estavariedad todavía se está cultivando en pocos sitios. ensistemas de bajo insumo y con sombra. y por parte deproductores muy pequenos.En cuanto a la variedad Borbón, su siembra semantiene. en sistemas bajo sombra y a veces con unbuen nivel de intensificación como es el caso en EISalvador o Guatemala por ejemplo.
AI Los cultivares originalesDos poblaciones de café de porte alto (o 'norma!') seintrodujeron, el Típica (o Arábigo común) y el Borbón,a partir de rnuy pocas semillas. La base genética y la
variabilidad potencial eran por lo tanto muy reducidas.En el sistema de cultivo que prevalecia hasta la mitadde este siglo. Ias diferencias de productividad entre unmaterial seleccionado y un material poco seleccionado
BI Las variedades mejoradas provenientes de labase tradicional (1950-1980)Con e! uso generalizado de la plena cxposición solar enlos anos 60 y 70 en Colombia y Costa Rica. seevidenció que las densidades de siembra deberían seraumentadas (Campos. 1998). En un sistema de altadensidad (más de 4000 plantas/há). Ias variedades deporte pequeno como el Caturra son las másproductivas (Cuadro I).
* Producción observada en kg. de café oro/ha.
Cuadro I: Producción relativa (%) de tres variedades por tres densidades. Calculada sobre la base de la producciónprornedia de cinco cosechas en Costa Rica. en relación a la producción de la variedad H33. base 100.(adaptados de informes de labores dei TCAFE, Costa Rica)
Producción relativa en relación con la densidad (%)
VARIEDADES 3200 pilhá 4200 pilha 6300 pilhaCATURRA 98..5 123 125
MUNDO NOVO 1T4,,2 112 109H33 100 (2660)* 100 (2620)* 100 (3000)*
"

232
Según Castillo (1990). en Colombia. Ia varicdadCaturra tuvo el importante papel de "catalizador" denuevas prácticas culturales. Esta idea se aplicatambién en el caso de América Central en ciertasregiones favorables, en donde el cultivo de la variedadCaturra (o Pacas) asociado con una densidad desiembra elevada y una fertilización apropiada permitióalcanzar productividades muy elevadas. Sin embargo.en Guatemala. Nicaragua. Honduras y EI Salvador.muchos productores no adoptaron los cultivares deporte pequeno ya sea porque pensaban que se iba adeteriorar la calidad dei café o porque las condicionesedafoclirnáticas o económ icas requerían mantcnermucha sombra. por lo que no se podia valorar las altasdensidades.La genealogia de los pricipales cultivares' se resume
en la figura 1. EI Caturra es actualrncntc la variedadmás sembrada en América Central. EI Pacas, que esuna varicdad rnuy utilizada en EI Salvador y enGuatemala. y cl Villa Sarchi (llarnada también la l.uisao Villalohos Borhón) que se semhraha cn Costa Ricaen los anos 50-60 son geneticamente cornparables aiCaturra. pues todos derivan de la población deiBorhón y tienen el mismo gen Caturra (Carvalho.1991).[n América Central . Ias líneas de Mundo Novointroducidas producen igual que el Borbón y menosque el Caturra en sistemas de altas densidades (cuadro1). por lo que se abandonó. Actualmente pocas áreasestán sem bradas con esta variedad . La varicdad 1133.que fue cultivada en Costa Rica hasta los aüos 1980.provicne probablemenrc de un cruzamiento entreTípica y Borbón y como tal. es geneticamente ccrcanaai Mundo Novo. Finalmente el Catuai da rendim icntossimilares o un poco superiores ai Caturra. En Costa
Rica. el Catuai produce alrededor de 8 a '5% más queel Caturra (informes deI ICAFE. 1973-1989), y serecomendó su cultivo (Campos. 1987). Actualrnente,es una variedad relativamente común en toda AméricaCentral pero su uso no ha introducido un cambio muydrástico en relación con el Caturra.
Cf Ventajas e inconvenientes de las variedadestradicionalesCon la selección y la posterior liberación dei Catuai. seterminó la era de la cxplotación de la base genéticaintroducida en América Central a partir dei sigloXVIII. Con la exccpción dei Catuaí que resulta de unesluerzo real por recorn binar las característicasdescables de dos padres bien conocidos. todas lasdcrnás variedades resultan dei mejoramiento de lapoblación existente (Borbón). dei mejoramiento decruzamientos espontáneos (Mundo Novo). o bien de laselección a partir de una mutación (Caturra. Pacas) .Con la excepción de la variedad Tipica. Ias principalesvariedades descritas anteriormente se caracterizan portenor una alta productividad (González y Gutierrez.1978) ) una buena adaptación a la lum inosidad. EnAmérica Central. las variedades de porte alto (Borbón,Mundo Novo) se rccorniendan para sistemas decultivo baio sombra regulada a densidades de 2000 a3500 plantas/há. mientras cl Caturra o el Catuai secultivan bajo sombra regulada o sin sombra con altasdensidades de siernbra (de 4000 a 7000 plantas/ha).Los defectos dei grano para ambas variedades oscilanentre 5 a /2% para los caracoles. y O a 12 % para losfrutos tlotantes (fruto vano). EI grano dei Típica es máslargo que el grano del Borbón y éste es un poco másgrande que el grano dei Caturra o dei Catuai.
TYPICA BORBON(Sumatra, Arabigo, Blue Mountain) (Híbrido Tiro. Tekisir)
Variedades de porte norma/ (alto) Variedades de porte norma/(alto)
",~ CATURRA
MUNDO NOVO (Pacas, VilIa Sarchí)
Variedades de porte norma/falto) Variedades de porte pequeno
/~CATUAI
Variedades de porte pequeno
Fig I: Genealogia de las principalcs variedades tradicionales2 flechas indicam un cruzamiento.I flecha indica una mutación.

La calidad dei café es muy discutida.Aunque no se ha probado formalmente. muchoscompradores de café indican que el Tipica o el Borbónson superiores a todas las demás variedades en cuantoa la calidad en la taza y presentación flsica de losgranos (Lingle, 1995). En realidad. estas variedades secultivan casi exclusivamente ba]o sombra y muchasveces se han hecho comparaciones con Caturra oCatuai cultivado sin sombra. L1ama la atención elhecho que se considera Típica >Sorbón ~ Caturra ~
Catuai para la calidad. cuando la clasificación para laproductividad en América Central es exactarncnteinversa. Como lo subrayan Guyot et al. (1995). Iasobscrvaciones sobre calidad dcberían tomar en cuentacl estado tisiológico de la planta. En todo caso seconsidera que todas esas variedades tienen un altonivel de calidad y corresponden a un estándarreconocido internacionalmente. Eso tiene comocorolario que cualquier nueva variedad cn AméricaCentral debc alcanzar esc estándar.Debido a la base genética cstrecha, Ia principal
desventaia de las variedades tradicionales es suhornogeneidad genética, Se traduce en particular poruna ausencia de resistencia a las grandesenfermedades y plagas que alectan la caticulturamundial. Ninguna de estas variedades es resistente a laroya y todas son susceptibles a los nemátodos queafcctan el café cn la región y ai CSD .
3-EXPLOTACIÓN DEL HíBRIDO DE TIMOR
Debido ai alto costo que representa el control químicode la roya y a la posiblc contaminación dei medioambiente que su uso conlleva. se contemplo laposibilidad dei uso de la resistcncia genética paracornbatir esta cnfcrmcdad. Los principales logros seobtuvicron utilizando el Híbrido de Timor comoprogenitor.· Después de varios anos de seleccióngenealógica (1972-1995) se obtuvicron diferentesvariedades que cstán descritas brevemente acontinuación. Adernas de tcner varios genes deresistcncia a la roya, el Híbrido de Timor prcsentaresistencia a otras enfcrmedades y plagas. Â partir de1989. se empezaron nuevas sclccciones considerandola posibilidad de obtencr variedades con rcsistenciasimultânea a la roya, a los nemátodos y ai C!ll).
AI Selección para la resistencia a la roya (19721995)1:1 objetivo de la selección cs la obtención de unarcsistencia durable. es decir que pcrmanczca cfcctivapor lapsos prolongados cn grandes áreas de cultivocon ambiente favorable a la enfermedad (Jonhson,1984). En América Central. se pretende introdueir elnúmero más grande posib!e de genes mayores(dominantes) en una variedad homocigota (líncapura). La durabilidad de esta resistcncia se basa en ladi ficultad para eI hongo de dcsarrollarsirnultáncarnente genes de virulcncia a varios loci(Eskes. 1989).
233
A partir de 1972. a través de diferentes intercambloscon el C1FC. Brasil (IAC y lJFV) y Colornbia(CENrCAFE). y luego a partir de 1978 a través dePROMECAFE I. los países de América Centralintrodujeron varias descendcncias de los tres origenesde Híbrido de Timor: CrFC 8321 I. CrFC 832/2. CIFC1343 (fig.2).Varias plantas que derivan deI Híbrido de Timor sonresistentes a todas las razas de roya conocidas por loque se clasifican en un grupo fisiológico lIamado grupoÂ. En América Central. para ascgurar una rcsistcnciadurable. se decidió seleccionar únicamente plantas deeste grupo.Del material genético introducido en 1972 y lucgo apartir de 1978 se seleccionaron varias Iíneas (Echcverriy Fcrnández. 1989). Por lo general. en esta primcruetapa los materiales derivados dei CIFC83211 fueronlos más adaptados a las condiciones de AméricaCentral. Sin embargo. se notó una tcndencia a unagotam lento prematuro de las plantas cuando lasprácticas culturales o las condiciones climáticas nocran óptimas (Echcvcrri y Fernándcz. 1989).Después de 12 aüos de sclección genealógica. se liberóen Honduras la primera variedad Catimor con altaresistencia a la roya, el IflCÂ FE90 (Instituto dei caféde Honduras. 1991). En 1995. Costa Rica libcró otroCatimor, Ia variedad Costa Rica 95 (CR95).Rccicnternentc se liberaron las variedades MIDÂ96 (enPanamá) y l.empira 98 en Honduras. Ias cualcs soncasi idénticas a la CR95.
BI Descripción de las variedades 'Catimores'liberadas en América CentralLa variedad IHCA FE90 (Catimor T5175)En condiciones adecuadas la varicdad produccalrcdedor de 30% más que el Caturra y 20% más que elCatuai en las condiciones de Honduras. En Costa Rica.con altas densidades. Ias diferencias productivas sonrespectivamente de 20 ) 22% cn promedio. (cuadros 2y 3).
La variedad COSTA RICA 95: (MIDA96. Lemplra98)Es una variedad que produce entre 25 y 35 % más que
las variedades Caturra o Catuai según la zona (cuadro3). Para mantcner los altos rcndlmicntos, rcquicre deuna íuertc fcrtilización. si no se agota a partir dei torceraüo de producción. EI grano es más grande (se acercaai Típica) y supera un poco cl tamano dei '1'5175 )significativamente el tamaüo dei Caturrarespectivamente: 67.90% contra 65.17%. 63.30% )sobre un tamiz de 17/64 pulgadas). Su alta rcsistcncia ala roya la hacc rccomendable principalmente parazonas donde existe una rnayor incldencia de royu,Ls importante destacar aqui que tanto Costa Riea 95como rHCAFE90 no prcscntan rcsistcncia ai CBD ni alos ncmátodos (Meloidogyne spp y Pratylenchus J{J{J).
(Rodrígucz, 1992: Anzueto. 1993: Hcrtrand et al..1995).
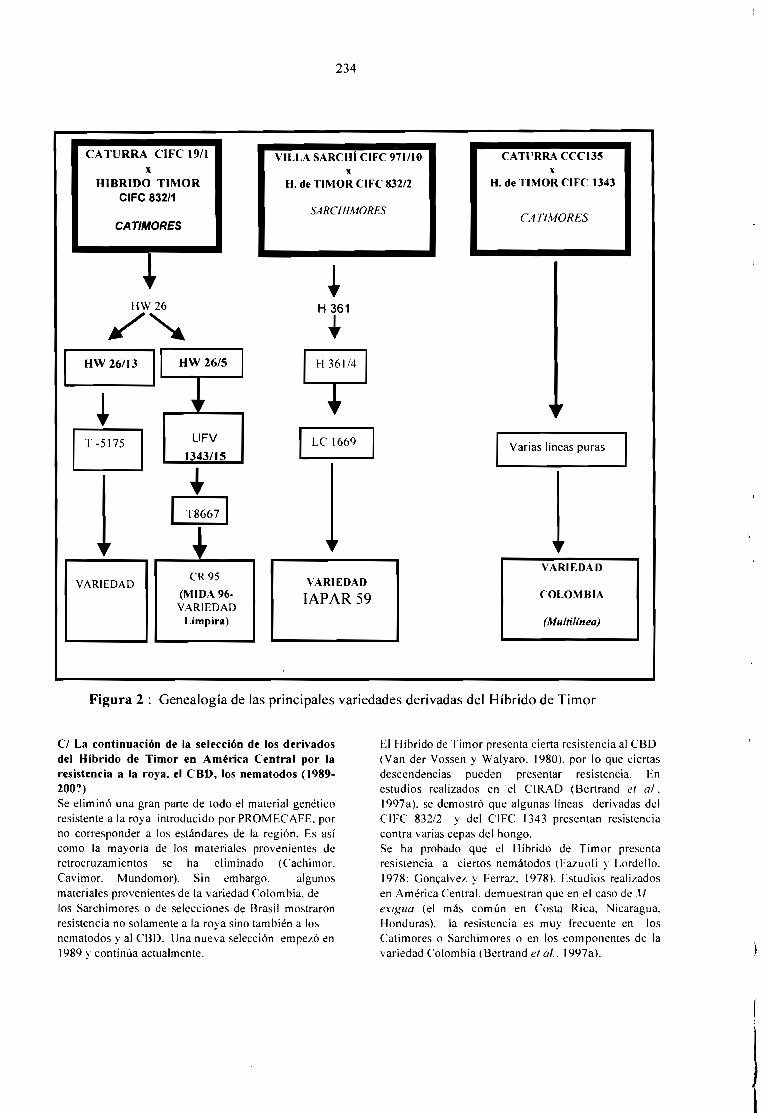
234
CATURRA CIFC 19/1x
HIBRIDO TIMORCIFC 832/1
VILLA SARCHí CIFC 971/10li
H. de TIMOR CIFC 832/2
CATlfRRA CCC13Sli
H. de TlMOR CIFe 1343
CATIMORES
SARCHIMORESCATlMORES
1I Varias lineas puras
"
~H 361
+
CTJI LC 1669 I
HW 26/5
UFV1343/15
HW 26
/"".HW 26/13 11....--.......----'
+Ô
1VARIEDAD
CR 95
(MIDA 96VARIEDAD
Límpira)
VARIEDAD
IAPAR 59
VARIEDAD
COLOMBIA
(Mulfilínea)
Figura 2 : Genealogía de las principales variedades derivadas del Híbrido de Timor
Cf La continuación de la selecciôn de los derivadosdei Híbrido de Timor en América Central por laresistencia a la roya, el CBD, los nematodos (1989200?)Se eliminó una gran parte de todo el material genéticoresistente a la roya introducido por PROMEC AFE. porno corresponder a los estándares de la región. Es asícomo la mayoría de los materiales provenientes deretrocruzamientos se ha eliminado (Cachirnor.Cavimor. Mundomor). Sin embargo. algunos
materiales provenientes de la variedad Colombia. delos Sarchimores o de selccciones de Brasil mostraronresistencia no solarnente a la roya sino tamhién a losnematodos y ai CBD. Una nueva seleceión ernpczó en1989 y continúa actualmente.
EI Híbrido de Timor presenta cierta resistencia aI CBO(Van der Vossen y Walyaro, 1980). por lo que ciertasdescendencias pueden presentar resistencia. Enestudios realizados en el ClRAD (Bertrand et a/o1997a). se demostró que algunas Iíneas derivadas deiCIFC 832/2 y dei CIFC 1343 presentan resistenciacontra varias cepas dei hongo.Se ha probado que el H íbrido de Tirnor presentaresistencia a ciertos nemátodos (Fazuoli y l.ordcllo,1978: Gonçalvez y Ferraz. 1978). Estudios realizadosen América Central. demuestran que en el caso de .Hexigua (el más cornún cn Costa Rica, Nicarágua.Honduras). Ia resistcncia es rnuy trccucntc en losCatimores o Sarchimores o en los componentes de lavariedad Colombia (Bcrtrand et ar. 1997a).
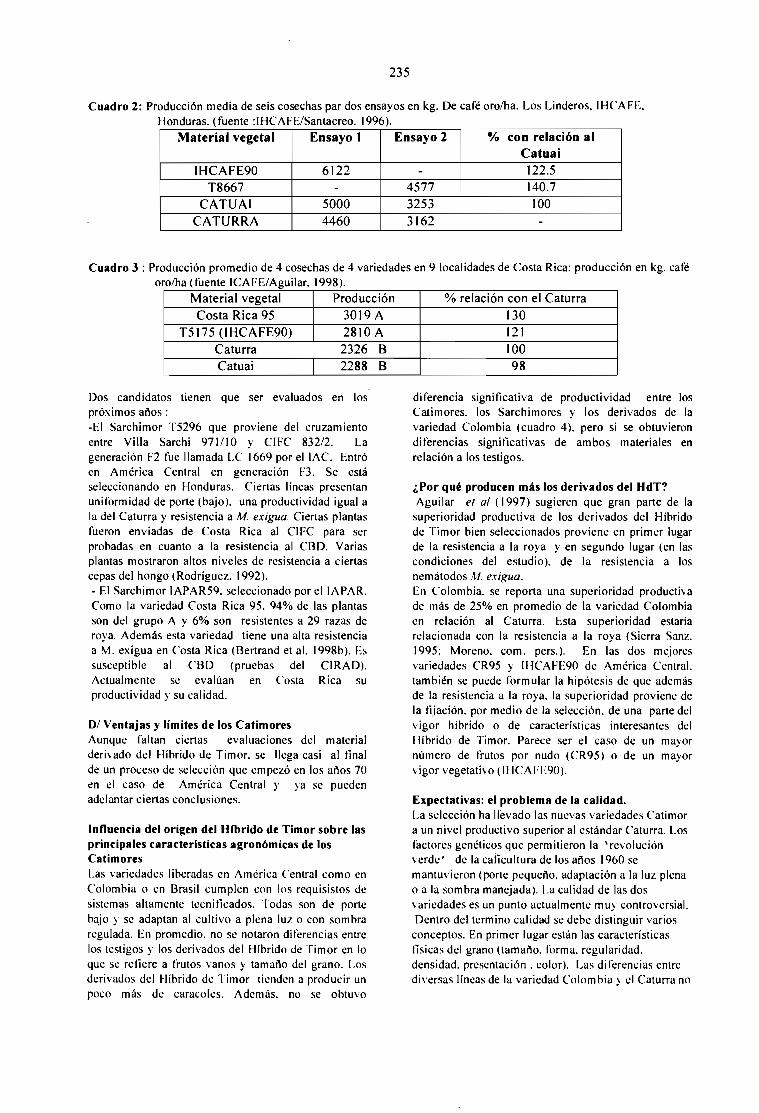
235
Cuadro 2: Producción media de seis cosechas par dos ensayos en kg. De cale oro/ha. Los Linderos. IHCAFE.Honduras (fuente ·IHCAFE/Santacreo. 1996).
Material vegetal Ensayo 1 Ensayo 2 % con relación aiCatuai
IHCAFE90 6122 - 122.5T8667 - 4577 140.7
CATUAI 5000 3253 100CATURRA 4460 3162 -
Cuadro 3 : Producción promedio de 4 cosechas de 4 variedades en 9 localidades de Costa Rica: producción en kg. caleorolha (fuente ICAFE/Aguilar. 1998).
Material vegetal Producción % relación con el CaturraCosta Rica 95 3019A 130
T5175 (IHCAFE90) 2810 A 121Caturra 2326 B 100Catuai 2288 B 98
Dos candidatos tienen que ser evaluados en lospróximos aüos :-EI Sarchimor 1'5296 que proviene dei cruzarnientoentre Villa Sarchi 971/10 y CIFC 832/2. Lageneración F2 fue Ilamada LC 1669 por el IAC. Entróen América Central en generación F3. Se estáseleccionando en Honduras. Ciertas líneas presentanuniformidad de porte (bajo), una productividad igual ala dei Caturra y resistencia a M. exigua. Ciertas plantasfueron enviadas de Costa Rica ai CIFC para serprobadas en cuanto a la resistencia ai CBD. Variasplantas mostraron altos niveles de resistencia a ciertascepas dei hongo (Rodriguez. 1992).- EI Sarchimor 1APAR59. seleccionado por eIIAPAR.Como la variedad Costa Rica 95. 94% de las plantasson dei grupo A y 6% son resistentes a 29 razas deroya, Además esta variedad tiene una alta resistenciaa M. exígua en Costa Rica (Bertrand et aI. 1998h). Essusceptible ai CAD (pruebas dei ClRAD).Actualrnentc se evalúan en Costa Rica suproductividad y su calidad.
DI Ventajas y Iímites de los CatimoresAunque faltan ciertas evaluaciones dei materialderivado deI Híbrido de Tirnor, se lIega casi ai tinalde un proceso de sclccción que empezó en los anos 70en el caso de América Central y ya se puedenadelantar ciertas conclusiones,
lnffuencta dei origen dei Híbrido de Timor sobre lasprincipales características agronómicas de losCatimoresLas variedades liberadas en América Central como enColomhia o en Brasil curnplcn con los requisistos desistemas altamente tccnificados. Todas son de portebajo y se adaptan ai cultivo a plena luz o con sombraregulada. En promedio. no se notaron diferencias entrelos testigos y los derivados dei Híbrido de Tirnor en loque se reficre a frutos vanos y tarnaüo dei grano. Losderivados deI Híbrido de Timor ticndcn a producir unpoco más de caracoles. Además. no se obtuvo
diferencia significativa de productividad entre losCatimores. los Sarchimores y los derivados de lavariedad Colombia (cuadro 4). pero si se obtuvierondiferencias significativas de ambos materiales enrelación a los testigos.
;,Por quê producen más los derivados dei HdT?Aguilar et aI (1997) sugieren que gran parte de la
superioridad productiva de los derivados deI Híbridode Timor bien seleccionados proviene en prirncr lugarde la resistencia a la roya y en segundo lugar (en lascondiciones deI estúdio). de la resistencia a losnemátodos M. exigua.En Colomhia. se reporta una superioridad productivade más de 25% en promedio de la variedad Colornbiaen relación ai Caturra. Esta superioridad estaríarelacionada con la resistencia a la roya (Sierra Sanz.1995: Moreno. com. pers.). En las dos mejoresvariedades CR95 )' IHCAFE90 de América Central.tarnbién se puede formular la hipótesis de que ademásde la resistencía a la roya. la superioridad proviene dela fijación. por medio de la selección, de una parte deivigor híbrido o de características interesantes deiHíbrido de Timor. Parece ser el easo de un mayornúmero de frutos por nudo (CR95) o de un rnayorvigor vegetativo (IHCAFE90).
Expectativas: el problema de la calidad.La selección ha Ilevado las nucvas variedades Catimora un nivel productivo superior ai estándar Caturra. Losfactores genéticos que permitieron la "rcvoluciónverde' de la caficultura de los aãos 1960 sernantuvieron (porte pequeno. adaptación a la luz plenao a la sombra manejada). La calidad de las dosvariedades es un punto actualrnentc rnuy controversial,Dentro dei termino calidad se debe distinguir varios
conceptos. En prirner lugar están las característicasfísicas dei grana (tamaão. forma. regularidad.densidad. prescntación . color), Las diferencias entrediversas líneas de la variedad Colomhia y el Caturra no

parecen ser consistentes según Castillo y Moreno,(1986) y Bertrand et ai., 1997a.En segundo lugar viene la calidad organoléptica (taza).La introgresión de genes dei Híbrido de Timor dentrodel genotipo Caturra (o Villa-Sarchi) puede habertraido ciertos problemas de calidad en la taza. SegúnMoreno et ai. (1995), no hay diferencias entre losCatimores que componen la variedad Colombia y elBorbón o el Típica.Estudios recientes hechos en Costa Rica revelan unasituación más contrastada en cuanto a los Catimoresprobados (fig. 3). En Costa Rica se realizó un estudiosobre la calidad de cuatro variedades: Caturra, Catuai,Costa Rica 95 y T5175 (Astúa y Aguilar, Informeanual ICAFE-1997) y éste estudio se repitió en 1999.La variedad T5175 se distinguió negativamente encuanto ai aroma, ai cuerpo y a el ácidez, Adernás, teníauna sabor a medicina y mostró un tueste disparejo,falta de uniformidad en sus granos y fisura abierta,para los ocho sítios donde fue evaluada. Estas mismascalidades muy bajas se encontraron en otros materialescomo por ejemplo: TI6784 (Sarchimor), T 17126 (1ineade la var. Colombia).Es dificil prever el ritmo de diseminación de losCatimores en América Central. En todos los países, sepiensa limitar los Catimores a las plantacionessem bradas a menos de 1200 msnm. La altaproductividad de Costa Rica 95 es un elemento muyatractivo para eI productor, pero la renovación deicafetal con esta variedad dependerá también:
• dei panorama de los precios en el mercadointernacional,
• de la facilidad de obtención de semillas debuena calidad,
• de la aceptación por parte de los compradoresde café de una calidad de café quizás inferior aiestándar 'Caturra' .de la durabilidad de laresistencia a la roya.
4- LA SELECCIÓN EN EL MARCO DE UNACAFICULTURA SOSTENIBLE
AI Problemas por resolver y criterios de selecciónEI mercado mundial y las características deCentroamérica imponen limitantes a la caficultura queallí se practica. Las condiciones de relieve, la pocadisponibilidad de nuevas tierras, y en ciertos casos laescasez de mano de obra, requieren de una
236
intensificación dei cultivo. Además el consumidor sevuelve más preocupado por la calidad y los grupos depresión conservasionistas exigen un café másecológico. La intensificación aparece como unanecesidad, pero debe ser una intensificación queproteja el medio ambiente y asegure la rentabilidad deicultivo aún en condiciones de precios bajos. Lagenética puede aportar soluciones a una parte de estosproblemas. EI conjunto de los riesgos biológicos(exógenos como endógenos), económicos ycomerciales conduce a fijar los siguientes criterios deselección:
• resistencia a enfermedades y plagas (roya, CBD,nemátodos);
• productividad igual o superior a los mejoresCatimores sin agotamiento precoz y con ciertaestabilidad productiva;
• defectos dei fruto comparables ai estándarCaturra;
• tamai'lo superior aI estándar:• calidad en la taza igual o mejor que la deI
Caturra:• porte pequei'lo y uniformidad de las plantas;
• buena adaptabilidad a condiciones
limitantes.
Para responder a los criterios de selecciónmencionados, se necesita ampliar la base genética enAmérica Central. Existen dos grandes vías paraplantear nuevas alternativas de selección Charrier( 1985).La primera vía es la hibridación interespecífica con laintrogresión de genes de resistencia de otras especiesde Coffea en la especie Coffea arabica por medio depolinizaciones o por medio de la transferencia de genesa través de la ingeniería molecular (plantastransgénicas). Esta última es una tecnologia nueva y suutilización para América Central requiere de una fuerteinversión.La secunda vía es a través de la hibridaciónintraespecífica creando híbridos FI con los cuales seespera tener un vigor vegetativo que se traducirá enuna mejor producción y una mejor adaptabilidad deicultivo en ambientes dificiles. Es la via escogida porPROMECAFE a partir de 1991 y el esquema sedescribe en la figura 4. Las mejores plantas (clones)son multiplicadas por embriogenesis somática
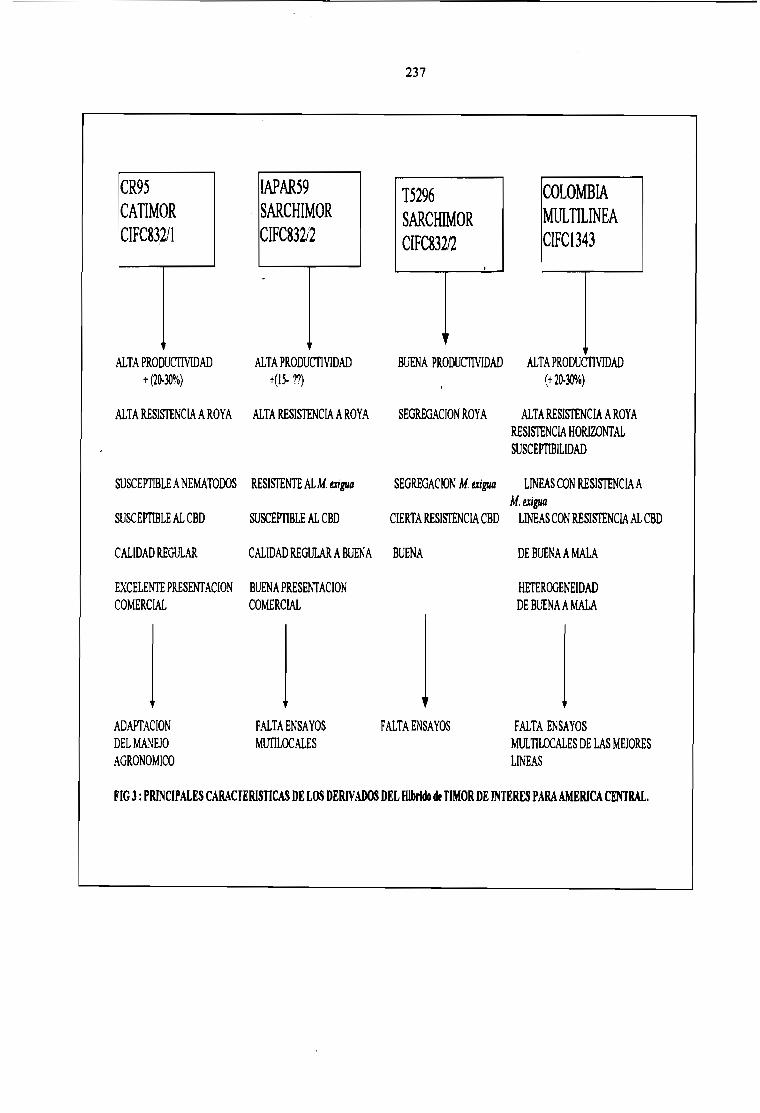
CR95CATIMORCIFC832/1
IAPAR59. SARCHIMOR
CIFC832/2
237
T5296SARCHIMORCIFC83212
COLOMBIAMULTILINEACIFC1343
ALTA PRODUCTIVIDAD ALTA PRODUCTIVIDAD BUENA PRODUCllVIDAD ALTA PRODUCTIVIDAD+(20-30%) +(15- ??) (+ 20-30%)
ALTA RESISTENClA AROYA ALTA RESISTENClA AROYA SEGREGACION ROYA ALTA RESISTENCIA AROYARESISTENClA HORIZONTALSUSCEPTIBILIDAD
SUSCEPTIBLE ANEMATOOOS RESISTENTE ALU exigua SEGREGACIOt' Mexigua LINEAS CON RESISTENCIA AMuigua
SUSCEPTIBLE AL CBD SUSCErnBLE AL CBD CIERTA RESISTENCIA CBD UNEAS CON RESISTENClA AL CBD
CALIDAO REGULAR CALIDAD REGULAR ABUENA BUENA DE BUENA AMALA
EXCELENTE PRESENTACION BUENA PRESENTACION HETEROGENEIDADCOMERCIAL COMERCIAL DE BUENA AMALA
ADAPfACIONDEL MANEJOAGRONOMICO
FALTA ENSAYOSMUflLOCALES
FALTAENSAYOS FALTA ENSAYOSMULTILOCALES DE LAS MEJORESLINEAS
FIG 3:PRlNCIPALES CARACTERISTICAS DE LOS DERIVAOOS DEL Hlbrldo dfTlMOR DE INTERES PARA AMERlCA CENTRAL.
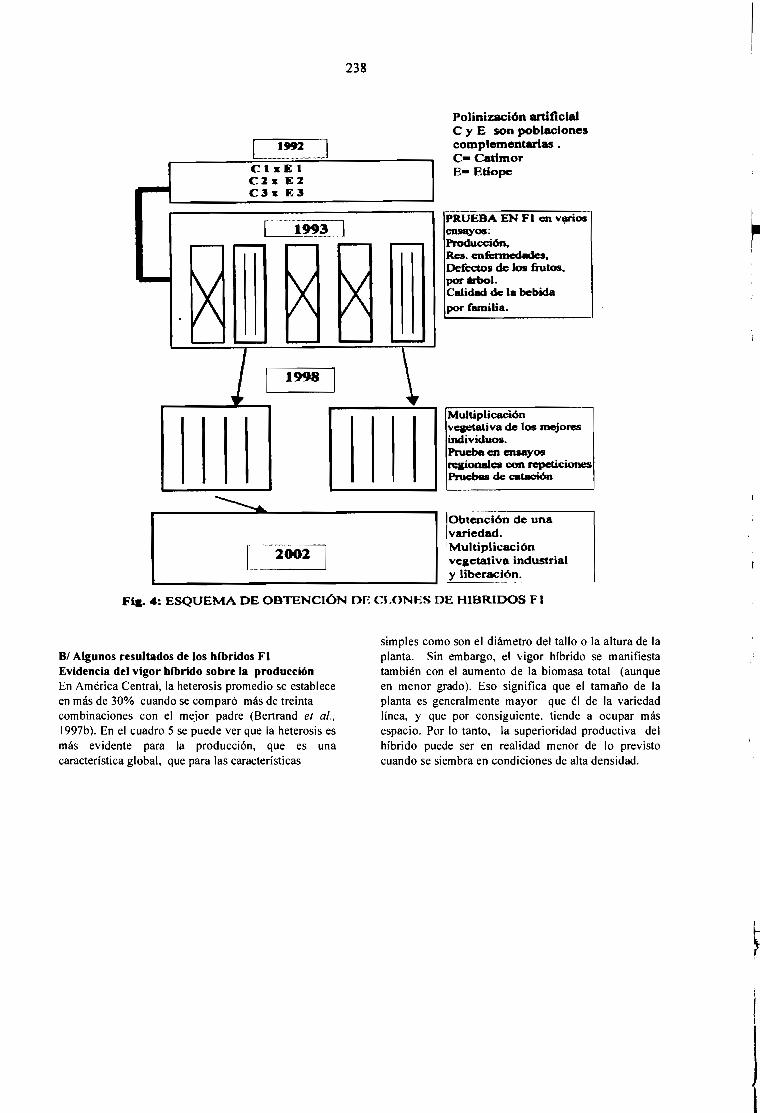
C I. E 1C2. E2C3. E3
238
Polinización artificialC y E 50n poblacionescomplementarias .C-Catfmol'R- Etiopc
PiUEBA EN FI cn v~oseosayos:Produççi6n.Res. cnfcnncd-.tes.Defc:ctos de los flUtos.por irbol.Calidad de la bebidapor (amUia.
I
r
1998
Multiplicaciónvcsetativa de los mejoresindividU05l.PJUebe cn ensayosl"CIPonalcs con rcpet.icionesPruebu de aatación
Obtención de unavariedad.Multiplicaciónvegetativa industrialy liberación.
Fia_ 4: ESQUEMA DE ODTENCIÓN DE CLONES DE HIBRlDOS FI
BI Algunos resultados de los hibridos FIEvidencia dei vigor hibrido sobre la producciónEn América Central, la heterosis promedio se estableceen más de 30% cuando se comparó más de treintacombinaciones con el mejor padre (Bertrand et al.,1997b). En el cuadro 5 se puede ver que la heterosis esmás evidente para la producción, que es unacaracterística global, que para las características
simples como son el diámetro dei taJlo o la altura de laplanta. Sin embargo, el vigor híbrido se manifiestatambién con el aumento de la biomasa total (aunqueen menor grado). Eso significa que el tamai'lo de laplanta es generalmente mayor que él de la variedadlínea, y que por consiguiente, tiende a ocupar másespacio. Por lo tanto. Ia superioridad productiva deihíbrido puede ser en realidad menor de lo previstocuando se siembra en condiciones de alta densidad.
~I
239
Cuadro 4: Comparación de las orígenes dei Híbrido de Timor con los testigos Caturra y Catuai, en dos ensayos, de laproducción acumulada de 4 y 5 cosechas respectivamente, C1CAFE, Heredia, Costa Rica. Producción en
Kg. café orolha (Bertrand et ai, 1997a)Ensayos Origen Nombre eomún Número de lineas Produecién % de produeejõn eoo los
testigosEnsayo I CIFC 832/2 Sarchimor 7 16453 a 124
CIFC 832/1 Catimor 5 15485 a 117TESTIGOS 2 13208 b 100
Ensayo 2 CIFC 1343 Colombia 16 15006 a 123TESTIGOS 2 12 183 b 100
Cuadro 5: Promedio de una población de cuatro Iíneas (Caturra, Catuai, CR95, T5296)en comparación con una población de 17 híbridos FI (Bertrand et ai. 1997b).
Variables Líneas Hibridos % de gananeia
Prod. Promedio, ···,kg. oro/ha 2079 2724 31 I
Peso granos secos (en gramo) 16.13 16.72 3.6Diámetro aI tallo·" (mm) 27.36 30.52 11.5
Altura··· (cm) 102 111.7 9.5Número bandolas" 57.48 61 6.1
Longitud bandolas··· (en em) 61.6 69 12Longitud entrenudos··· (en m) 3.02 3.36 11.25Número de frutos en 6 nudos" 96.9 101.7 4.95
••• estadlsticamente diferentes P<O.OOI, P<O.OI, P<0.05.
Recombínacíõn de genes de resistenda aenfermedadesUna de las grandes ventajas de los FIes que hayacumulación en un genotipo de las resistenciascomplementarias a las enfermedades y plagas como esel caso para los genes dominantes de resistencia a laroya, a los nemátodos y eventualmente ai CBD que seencuentran ya sea dentro dei grupo de los Catimores odentro dei grupo de los silvestres.
Defectos de frutos y calidad en la tazaDe manera general, los F I tienden a producir máscaracoles (+ 5%) y frutos vanos (+ 6%) que lasvariedades Iíneas (Bertrand et ai.. 1997b). En datos deexperimentos en Kenia reportados por Van der Vossen(1985), los híbridos tienen sisternáticamente máscaracoles que sus padres. Sin embargo la seleccióndentro de una familia de las mejores plantas para unamultiplicación clonal debería permitir eliminar en granparte este problema.De las primeras pruebas de catación que se hicieron, senota que la mayoría de los F I igualan o superan elpadre cultivado (CR95, T5296, Caturra, Catuai) (Datossin publicar).
Seleecíõn de Veinte precandidatosUna selección basada sobre los diferentes criterios susmencionados ha Ilevado a ofrecer veinte clonesprecandidatos que estan en curso de multiplicaciónmediante las técnicas de micropropagación. Estosclones producen entre 20 a 70% más que la variedadCaturra, con una calidad de la bebida igualo superior.Ciertos híbridos tienen resistencia a la roya y adiferentes nematodos. De 1999 a 2000, se prevesembrar más de 120 000 plantas directamente en las
fincas de los productores Centroamericanos.Dependiendo dei grado de aceptación por parte de losproductores se seleccionaran los mejores candidatosque serán luego liberados como variedades. La idea eslIegar a tener una oferta de varios clones.
Predueeiõn de variedades FIEI principal cuelJo de botella en el uso de los híbridosera la producción masiva de plantas. Hoy existe laposibilidad de utilizar eI método de multiplicaciónvegetativa por medio de la em briogénesis somática.Los costos actuales de producción de una vítroplantason de 0.10 USD es decir para el productor una plantade 8 meses que cuesta OAOUSD contra 0.30USD enAmérica Central. EI principal logro reside en lasiembra directo dei embrión (Barry-Etienne et ai,1999; Etienne et ai., in press).
Ct Creadón de variedades Porta-injerto: un nuevoconcepto.Para muchos frutales, Ia práctica dei injerto es rnuyusuaJ.Los patrones se utilizan ya sea como fuente deresistencia ai parasitismo dei suelo o a los virus, parareducir el vigor de las plantas (aumento de densidadesde siembra) así como para aumentarlo, o bien para unamejor adaptación a condiciones particulares comoexceso de calcio o de humedad, etc.En café, la injertación (injertación hipocotilodonariaprincipalemente) se utiliza principalmente para lucharcontra el parasitismo telúrico. En Guatemala se utílizadesde hace muchos aüos el patrón Robusta paracontrolar los nemátodos Pratylenchus sp. yMeloidogyne sp. En este país los nemátodos son rnuyagresivos y la práctica dei injerto da resultadosespectaculares en situaciones de fuertes infestaciones

con Pratylenchus , para el cual el patrón Robusta esaltamente tolerante (Anzueto, 1993; Vil1ain et al.,1996). En condiciones de invernadero, Anzueto (1993)ha mostrado que las descendencias de Robustautilizadas en Guatemala no presentan más de 30% deplantas resistentes. Esto indica la necesidad de realizarun trabajo de selección para obtener variedades conmayor resistencia a Meloidogyne spp manteniendo a lavez tolerancia a Pratylenchus.. A partir de 1990, seempezó una selección de nuevas variedades portainjerto que se debería continuar hasta 2003.
La variedad NemayaLa nueva variedad 'Nemaya' (Anzueto et al., 1995;Bertrand et ai., 1995), obtenida en 1995, viene deicruzamiento de dos árboles de la colección dei CATIE,el T3561 (planta 2-1) y el T3751 (planta 1-2). Ladescendencia de este cruzamiento es altamenteresistente a Meloidogyne sp. dei Salvador (Pei'la, 1994),de M. incognita de Guatemala (Anzueto, 1993), de M.exigua de Costa Rica (Bertrand et ai., 1998a) y de M.arabicida de Costa Rica.En relación con los nemátodos de Guatemala y EI
Salvador que son particularmente agresivos, losRobustas no seleccionados presentan alrededor de35% de plantas resistentes cuando la variedad Nemayapresenta cerca de 80% de plantas resistentes. Enrelación a M. exigua y M. arabicida, la variedadNemaya presenta respectivamente 90% y 80% deplantas resistentes. Además cabe notar que el T3751(planta 1-2) - uno de los dos padres de la variedad- hademostrado ser más tolerante a Pratylenchus sp. queotros Robustas (Anzueto, 1993).Complementariamente los Robustas presentan un buennivel de tolerancia a las cochinil1as Dysmicoccuscryptus, netamente superior a la tolerancia de losArábicas (Garcia,1992 ). La multiplicación de lavariedad Nemaya se está realizando en los paísesmiembros de PROMECAFE por medio de laembriogenesis somática.Cabe notar que se debería hacer más investigaciones encuanto a la práctica dei injerto en condiciones debajura «. 800 msnm). En efecto a esas alturaslimitantes para el Arabica, el Robusta confiere unaumento de vigor, tanto en presencia de nemátodoscomo en su ausencia y podría procurar una mejoradaptabilidad en suelos ácidos
Los CatimoresEn situaciones de altura (800-1000 metros), las bajastemperaturas empiezan a causar ciertos disturbiosfisiológicos a los Robusta. A más de 1000 msnm,Bertrand et al., (1998a) estiman que en situación demuy baja infestación con nemátodos, la injertacióncausa una pérdida de 10 a 20% de la producción enrelación a testigos no injertados. Eso significa que enpresencia de un nemátodo como M exigua, que nocausa pérdidas superiores a un 10-20% en condicionesde clima favorable, el injerto sobre patrón de Robustano se justifica. En este caso particular, la resistencia
240
encontrada en los Catimores (variedad IAPAR59 porejemplo), puede ser aprovechada. Un patrón Catimorde igual crecimiento ai Caturra no debería afectar elcrecimiento de la planta. Sin embargo, en situacionesde ataques múltiples de Pratylenchus y M. exigua, estasolución no es recomendable, pues debido a lasusceptibilidad deI Arábica ai Pratylenchus y a lacompetencia entre ambas especies se nota un fuerteincremento de la población de Pratylenchus (Bertrandet al., 1998b)
Haeia la creaccíõn de una variedad porta-injertoArabustaEI híbrido interespecífico Arabusta, creado a partir deuno de los padres de la variedad Nemaya cruzado conun silvestre resistente a M. incognita y M. arabicida,podría ser una valiosa fuente de resistencia heredadadei Robusta y, a la vez, conferir un fuerte vigor aiinjerto en condiciones de más de 800 msnm en dondeel Robusta sufre por las bajas temperaturas. En e)CATlE se esta realizando un programa de duplicacióndei número de cromosomas dei T3751 (1-2) y deiT3561 (2-1), para crear dos tetraploides que luego sepodrán cruzar con uno o varios Arábica silvestreresistente a ciertos nemátodos creando así diferentesvariedades de Arabusta.
5-CONCLUSIÓN
La ampliación de la base genética es el mayor objetivode los programas de mejoramiento genético deAmérica Central. A corto y mediano plazo, lostrabajos de selección permitiran una oferta de otrasvariedades Catimores y de varios clones F I conresistencia a la roya, e/o a los nemátodos y ai CBO ytodos de buena a muy buena calidad. En las regionesen donde los nemátodos son muy agresivos y ensituaciones de bajura, la variedad porta-injerto Nemayaserá una buena solución agronómica. Para lassituaciones de altura en donde el Robusta tieneproblemas de crecimiento, se puede esperar la ofertadentro de 5-6 ai'los de un porta-injerto de tipo'Arabusta' con alta resistencia a los nemátodos y degran vigor.La meta es l1egar ai productor con una oferta dediferentes variedades y patrones las cuales el escogerásegún sus necesidades ecológicas y económicas y susprioridades de comercialización.La diversidad genética presente en América Centralestá en proceso de evaluación por los marcadoresmoleculares RAPO, AFLP (Anthony et al., in press;Lashermes et ai, in press). EI material estudiadoconstituye un amplio muestreo de la totalidad dei áreade origen de la especie C Arabica (individuossilvestres de las recolecciones FAO y ORSTOM enEtiopía y de la prospección IPGRI en Yemen), asícomo por las principales variedades (Típica. Borbón,Caturra, Villasarchi, derivados dei Híbrido de Timor).Los primeros resultados corroboran la existencia deuna base genética limitada en C arabica, revelada por

los estudios preliminares de Orozco-Castillo et aI.(1994) y de Lashermes et aI. (1996c). Esta evaluacióngenotípica se completa con la evaluación fenotípica,basada en la observación en el campo de los caracteresutilizados como criterio de selección en la región, porejemplo los defectos de los frutos y de los granos, o laresistencia a enfennedades y parásitos (roya,nematodos, CBD). Los resultados tendrán fuertesimplicaciones en la conservación y la utilización de losrecursos genéticos. Revelar la estructura de ladiversidad disponible en América Central para elmejoramiento regional conduce a la definición degrupos de individuos genéticamente cercanos. Ladiversidad disponible podrá entonces ser utilizada demanera racional basándose en la estimación de ladistancia genética entre las variedades cultivadas y losdiversos grupos de individuos silvestres, cultivados ointrogresados.En América Central, los híbridos FI creados entre losindividuos silvestres y las variedades Catimor oSarchimor penniten crear poblaciones segregantes paranumerosos caracteres agronómicos, con determinismomonogénico. En 1998 se inició un proyecto financiadopor la UE para identificar marcadores molecularesligados a la resistencia a dos especies de nernatodos,M. exigua de Costa Rica y Meloidogyne sp. deGuatemala. Otros caracteres monogénicos de interéspodrán también ser marcados a corto plazo. Porejemplo, la explotación de la esterilidad masculina,detectada en varios individuos etíopes de la coleccióndei CA T1E (Dufour et aI., 1997). abriría la puerta a unadistribución de híbridos FI por semilla.
6-AG RADECIMIENTOS
Agradecemos la dirección dei ICAFE de Costa Ricapor su apoyo ai programa regional de fitomejoramientode PROMECAFE
7-L1TERATllRA CITADA
Aguilar, G. 1995. Variedad Costa Rica 95. Instituto deiCafé de Costa Rica, Convenio ICAFE-MAG.30 p. ISBN 977-55-014-X.
Aguilar, G, Bertrand B. y F. Anthony. 1997.Comportamiento agronómico y resistencia alas principales plagas de diferentes variedadesderivadas dei Híbrido de Timor. Noticiero deicafé. Revista dei Instituto dei café de CostaRica. W 94-95. ISSN 108-421 X, 8p.
Anzueto, F. 1993. Etude de la résistance du caféier àMeloidogyne sp. et Pratylenchus sp. In Thêsede Docteur Ingénieur, Ecole NationaleSupérieure Agronom ique de Rennes, 17 mars1993.123 p.
Anzueto, F. Bertrand B. y M. Dufour , 1995.Desarrollo de una variedad porta-injertoresistente a los principales nemátodos deAmérica Central. Boletín PROMECAFE,
241
Enero-Junio, I1CA, ap 1815, Guatemala, 1315.
Bertrand B., Anzueto F, Pei'la M, Anthony F. y A.8.Eskes. 1995. Genetic improvment of coffeefor the resistance to root-knot nematodesMeloidogyne sp. In América Central. In 15congreso intemacional sobre el café, Kyoto,mayo de 1995, Japón, ASIC (Paris): 630-636.
Bertrand, B., Aguilar G, Bompard E, Rafinon A, and F.Anthony, 1997a. Comportement agronomiqueet résistance aux principaux déprédateurs deslignées de Sarchimors et Catímors au CostaRica. Plantations Recherche Développement.4 (4): 312-321.
Bertrand, Bi.Aguilar G, Santacreo R, Anthony F.,Etienne H., Eskes A.8.. and A. Charrier.1997b. Comportement d'hybrides F I deCoffia arabica pour la vigueur, la productionet la fertilité en Amérique Centrale. In 17congreso intemacional sobre el café, Nairóbi,julio de 1997, Kenya, ASIC (Vevey, Suiza).
Bertrand, 8., Borbón, O., and G. Aguilar. 1998a.Situación nematoJógica en un cafetal de lameseta Central de Costa Rica y posibilidadesde control. In: ICAFE. San José, Costa Rica(Eds), Memoria III seminario resultados yavances de investigación. p 15-28.
Bertrand, B., Cilas, c., Hervé, G.• Anthony, F..Etienne, H., and L. Villain. I998b. Relationsentre les populations des nématodesMeloidogyne exigua et Pratylenchus sp., dansles racines de CojJea arabica au Costa Rica.Plantations Rech Dev 5:4, 281-286.
Campos, E. 1987. Comparación dei Catuai con elCaturra. Noticiero dei café, Ai'lo 2. N°27, Dec1987. ICAFE, San José, Costa Rica, 3 p..ISSN 0550-1105.
Campos, M. E. 1998. Densidades de siembra ydistancias entre plantas e hileras. In: ICAFE.San José. Costa Rica (Eds). Memoria IIIseminario resultados y avances deinvestigación. pp 95-100.
Carvalho. A.. Medina Filho H.P. Fazuoli L.C, and O.Guerreiro Filho. 1991. Aspectos genéticos docafeeiro. Rev. Brasil. Genet. 14: I, 135-183.
Castillo Zapata, J., 1990. Mejoramiento genético deicafé en Colombia. In: 50 ai'los de CENICAFE1938-1988. Conferencias conmemorativas.Chinchiná (Colornbia). CENICAFE, p. 46-53.CDD -633.730968.
Castillo Zapata, J., and J. Leguizamon Caycedo. 1992.Virulencia de "Hernileia vastatrix"determinada por media de plantasdiferenciales de café en Colombia. Cenicafé(Colombia). 43 (4): 114-124.
Castillo Zapatta, 1.. and G. Moreno. 1986. La variedadColombia: Selección de un cultivarcompuesto resistente a la roya dei cafeto,Federación Nacional de cafeteros deColom bia, Bogotá. 172 p.

Castillo Zapata, J., 1984. Actividades de Colombia eninvestigación en mejoramiento genético deIcafé. In : IV reunión regional de especialistasen mejoramiento genético dei café. Antigua,Guatemala 1-5 Octubre de 1984,PROMECAFE-I1CA, 30 pp.
Charrier, A. 1985. Progrês et perspectives del'amélioration génétique des caféiers. In: 11°Coloquio científico internacional sobre elcafé, Lomé. Vevey, Suiza, ASIC, p 403-425.
CowgilI, W.H. 1958. The Sun-hedge system of coffeegrowing. Coffee and Tea Industries. Advancesin Coffee Production. 81 (11): 87-90.
Dufour, M., Anthony, F., Bertrand, B. and A.B. Eskes.1997. Identification de caféiers mâle-stérilesde Coffea arabica au CATIE, Costa Rica.Plantations, Recherche, Développement 4 (6):401-407
Echeverri, .r.H. and Fernández C.E., 1989. ThePROMECAFE program for Central América.In Coffee Rust: Epidemiology, Resistance andmanagement. Eds Kussalappa A.C., EskesAB., CRC Press, Inc. Boca Raton, Florida, p337. ISBN 0-8493-6899-5.
Etienne, H., Bertrand, B., Anthony, F., Cote, F., andM.Berthouly. 1997. L'embryogenesesomatique, un outil pour l'améliorationgénétique du café. In: ASIC (ed.) 17 th IntSei ColIoquium Coffee. ASIC. Vevey,Switszerland, pp 457-465.
Etienne-Barry, D., Bertrand, B., Vasquez, N., andH.Etienne. 1999. Direct sowing of Cojfeaarabica somatic embryos mass-produced in abioreactor and regeneration of plants. PlantCell reports 18:77.
Eskes, A.B. 1989. Coffee Rust: Epiderniology,Resistance and management. Eds KussalappaAC., Eskes AB., CRC Press, Inc. BocaRaton, Florida, p 337. ISBN 0-8493-6899-5.
Fazuoli, L.C. 1985. Génetica e melhoramento docafeeiro. in Cultura do cafeeiro, Piracicaba,Brasil. Associaçiao Brasileira para pesquisada potasa e do fosfato. 87-1 J3p.
Fazuoli. LiC. and R.R.A. Lordello. 1978 . Resistênciade cafeeiros Híbrido do Timor a Meloidogyneexigua. Ciencia y Cultura, Suplemento,Brasil. 30,3.
Garcia, A. 1992. Les pseudococcidae déprédateurs desracines du caféier iCoffea arabica L.) auGuatemala. Cas particulier de Dysmicoccuscryptus (Hempel, 1918). Thêse doct. Univ.Paul Sabatier Toulouse 111. 122 p.
Gonçalvez, W., and L.C.C.B. Ferraz. 1978. Resistênciado cafeeiro a nematóides: 11 - Testes deprogênies e híbridos para Meloidogyneincognita raça 3. Nemat. brasileira, vol. XI,125-142.
González, G., and Z. Gutiérrez. 1978. Respuesta dealgunos cultivares de Coffea arabica a
242
diferentes densidades de siembra. Agron.Costarr. 2( I): 61-68.
Guyot, B., Gueule, D., Manez, J.c., Perriot, J.1., Giron,J., and L. Vil1ain. 1995. Influence de I'altitudeet de I'ombrage sur la qualité des cafésArabicas. Plantation recherchedéveloppement,3 (4):272-280.
Instituto Hondurei'lo deI café, 1991. Una nuevavariedad de café con resistencia genética a laroya. Boletín de PROMECAFE, n052-53,Junio-Dic., 15-18.
Informes anuales de labores, 1973 a 1989 .Departamento de Investigaciones ytransferencia de tecnología, Instituto dei Café.San José. Costa Rica.
Jonhson, R. 1984. A critical analisis of durableresistance. Anual Review of phytopathology.(EE.UU). 22. 309-330.
Lashermes, P., Trouslot, P., Anthony, F., Combes,M.C.and A Charrier. 1996. Genetic diversityfor RAPD markers between cultivated andwild accessions of Coffea arabica. Euphytica87: 59-64
Lingle, T. 1995. DesarrolIo deI café de especialidades"Gourmet", orgánico y otros. Doc. Nestlé,XVII Simposio de caficultura, 23-27 Octubre.San Salvador, EI Salvador, PROMECAFE,PROCAFE. Actas por aparecer
Mendes, J.E.T.1939. Ensaio de variedades decafeeiros. BoI. Instituto agronomico do estadoem Campinas, Sao Paulo. Tec. (65) 35 p.
Moreno Ruiz, G., Moreno E., and G. Cadena.1995.Bean characteristics and cup quality of theColombia variety (Coffea arabica) as judgedby international tasting pane/s. In: 16°Coloquio científico internacional sobre elcafé, Kyoto ASIC, 16° ColIoque, Kyoto.Vevey, Suiza. ASIC. Acts, p. 574-583.
Orozco-CastilIo, C., Chalmers, K.J., Waugh R., and W.Powell. J994. Detection of genetic diversityand selective gene introgression in coffeeusing RAPD markers. Theor. Appl. Genet. 87:934-940
Peãa, M.X. 1994. Evaluación fenotípica y genéticapara la resistencia aI nernátodo Meloidogyneincognita en Híbridos de Cojfea canephora.Tesis de Master. CATIE. Turrialba, CostaRica. 78p.
Rodríguez, C.1. 1992. Pathology & Improvment ofcoffee for the main deseases. Contract TS20259-P de la CEE. In Fifth scientific report,1st May- 3 J octuber 1992 .
Sierra Sanz, C.A. 1995. Control de la roya deI cafetocon base en niveles de infección y su efectoen la producción. CEN1CAFE, Chinchina,Colombia, Vol. 46, 2, 1I p.
Uribe, A. 1958. EI cultivo deI café aI sol en Colombia.Revista cafetalera de Colombia. 134: 31-34.
Villain, L.. Sarah J.L., Decazy B.• Molina A, andS.Sierra. 1996. Evaluation of grafting on
,
I

Coffea canephora varo Robusta and chemicalstreatment for control of Pratylenchus sp. enC. arabica cropping systems. ThirdInternational nematology congress, abstractsof posters, p.155.
Van der Vossen, A.M. 1985. CotTee selection andbreeding ,In Coffee, Botany, Biochemistry
243
and Production of Beans and Beverage.ClitTord, (M.N.) & Wilson ( K.C.)., Ed., AVI,Westport, CT, 1985, Chap. 3.
Van der Vossen, H.A.M., and D.J.Walyaro. 1980.Breeding for resistance to coffee berrydesease in Co.fJea arabica L. 11. Inheritance ofthe resistance. Euphytica 29: 777-791.



247
BIOTECHNOLOGY FOR GÊNETIC IMPROVEMENT OF INDlAN COFFEE
H.L. Sreenath
Centre. Unit of Central CofTee Research Institute Coffee Board, Manasagangothri, Mysore - 570 006.Karnataka, India e-mail: [email protected]
SllMMARYFor India, a developing country, cojJee earns around 450 million dollars in foreign exchange which is veryimportant for the economy. Due to sustained research efforts Df Central CojJee Research Institute (CCRI),the production and productivity of Indian cojJee has improved steadily during the last five decades.However, further intense research efforts are needed to make the Indian coffee industry more vibrant to facethe challenges of the next millenium. Biotechnology is identified as a thrust area for meeting thesechallenges. In India, research efforts on coffee biotechnology are focussed on micropropagation, antherculture, embryo rescue. endosperm culture, protoplast culture, genetic transformation, marker aidedselection and in-vitro preservation. Research ejJorts at CCRI have resulted in successful plant regenerationthrough somatic embryogenesis in more than 20 selections of C arabica (arabica) and C canephora(robusta). Hardening procedure has been evolved for micropropagated plants. Large scale field trials ofmicropropagated plants is taken up using the 3 selections viz., Cauvery. SIn.9 and CxR and trial plots oftissue cultured plants are established in difJerent agroclimatic zones. Plant regeneration is achieved fromthe apical bud and nodal explants, but further refinement ofthis technique is requiredfor commercialization.For the first time in any plant species, plant regeneration is achieved from integument tissues in the CxRcultivar. Protocols have been optimized for direct and indirect plant regeneration from embryo culture.Plants derived from embryo culture ofinterspeciflc crosses between cultivated C canephora (robusta) and 3indigenous wild species, viz.. C bengalensis, C travancorensis and C wightiana have been established inthe field. Plant regeneration is achieved for the first time from the anther culture in the diploid CxR cultivarofcoffee. Plant regeneration is achieved from endosperm cultures ofC arabica (8. 2803) which is again afirst achievement for any coffee species. Zygotic embryos of C arábica have been preserved for 2 yearsunder slow growth conditions and successful cryopreservation ofzygotic embryos is achieved in two species.Protocols have been developed for protoplast isolation from embryogenic calli in two species. Initialsuccess is achieved in developing DNA markers and genetic transformation. These programmes are aimedat developing molecular markers linked to leaf rust resistance. engineering leaf rust resistance withantifungal genes. isolation of genes for leaf rust resistance, using Bt genes to combat major pests andisolating cajJeine degradation genes for manipulation ofcaffeine content in the beans.
INTRODUCTION
The genus Coffea coming under the familyRubiaceae is described to contain about 70species. However, only two species arecommercially important. C arabica popularlyknown as arabica cofTee accounts for about 70%of the coffee consumed in the world because ofits superior beverage quality. C canephorasupplies the remaining 30%. Cultivated varietiesof this species are generally called robustacofTee. A few other species of cofTee like Cdewevrei, C liberica and C racemosa areconsumed in the local producing areas.CofTee is the most important agricultural exportcommodity in the world and many countries areinvolved in its production, trade andconsurnption. It generates 15 billion USdollarsper year in the green cofTee trade. The averageworld production is estimated to be 93 millionbags (60 kgs each) (about 56 lakh tonnes) frommore than 50 cofTee producing countries in the
tropical regions of Latin America, Africa and Asia.For most of these countries, coffee is the majorsource of hardcurrency earnings. India is among the top tencoffee producing and exporting countries alongwith Brazil, Columbia. Indonesia, Vietnarn,Mexico, Ivory Coast, Ethiopia, Uganda andGuatemala. In India both arabica and robustacofTee are cultivated almost in equal areas. Coffeeproduction in India has reached 2.3 lakh tonnes in1998 from around 19 thousand tonnes in 1950,while the average productivity has increased toover 850 kg/ha from 204 kg/ha during the sameperiod.Major challenges to coffee cultivation in IndiaManaging the existing biodiversity, tackling majorbiotic and abiotic stresses, quality improvementand protection of environment are the majorchallenges to be addressed to make cofTeecultivation more sustainable.The most important disease affecting Indian cofTeeis the leaf rust caused by the fungus Hemileia

vastatrix. The disease first recorded from Indiain 1869. has developed into the most challengingdisease of arabica coffee in India. The majorpests attacking Indian coffee are. the coffee berryborer (Hypothenemus hampei), white stem borer(Xylotrechus quadripes), mealy bugs(Planococcus citri and P. lilacinus). shot holeborer (Xylosondrus compactus) and rootnematode (Pratylenchus colfeae) (Anonymous,1998). Water stress and oxidative stress are themost important abiotic stresses affecting thecoffee plants and responsible for severe yieldlosses. With the expansion of coffee into nontraditional are as. high temperature stress alsoneeds attention. Apart from the biotic andabiotic stresses, increasing stringency ofenvironmental protection is gaining prominence.
Coffee improvement by conventional breedingEmploying conventional plant breeding methodsof individual plant selection followed byprogeny evaluation, hybridization and pedigreeselection, coffee plants with high yield andvigorous plant growth have been developed. Inaddition to these, back cross methods have beenused to transfer specific traits such as. shortinternodes, and disease and pest resistance to Carabica from other cultivars or related species.During the last 60 years, Central CoffeeResearch Institute (CC RI). lndia, has developed12 improved cultivars of arabica and 3 improvedcultivars of robusta. through conventionalbreeding (Anonyrnous, 1996). A few morepromising selections are in the advanced stagesof evaluation. However, inspite of tremendouscontri bution to coffee genetic improvernent,conventional breeding suffers from thelimitations like. the genetic barrier ofchromosome number (diploids vs. tetraploids),autoincompatible alleles (diploid species), lackof genetic understanding and long breedingcycles. As a result of these factors, the transferof genetic traits from wild outbred species of thegenus to the cultivated species is quite difticult.
Biotechnology for coffee improvementBiotechnology can supplement the efforts ofcoffee breeders with additional tools which canovercome the limitations of traditional plantbreeding to a great extent. There is tremendousscope for application of crop biotechnology toolsfor genetic improvement of Indian coffee.Coffee improvement through biotechnology canfocus on three different areas of applications:agronorny, processing industry and consumers.Agronomic benefits should focus on reducingdirect and indirect farming costs. To reduce
248
coffee farming costs, the new technologies canaddress ferti lizer efficiency, disease and pestresistance and crop management aspects that wi 11reduce labour utilization (herbicide tolerance,mechanized harvesting etc.).Various biotechnology approaches likemicropropagation, embryo rescue, anther culture,cell line selection. somaclonal variation, protoplastculture, in-vitro preservation, marker aidedselection and genetic transformation havetremendous potential for genetic improvement ofcoffee. Eversince the first report on tissue cultureregeneration of coffee by Staritsky (1970). therehas been a steady progress in coffee biotechnologyresearch (Sreenath & Naidu, 1997; Sondahl &Lauritis, 1992; Sondahl & Loh, 1988).
COFFEE BIOTECHNOLOGY RESEARCHATCCRI
Biotechnology research is conducted at CCRI withthe short term and long term objectives ofmicropropagation and genetic improvementrespectively. For achieving the short term and longterm objectives, research efforts are focussed onstem, leaf integument. apical bud and nodecultures for micropropagation, anther culture,embryo culture, endosperm culture, protoplastculture, genetic transformation and molecularmarkers for genetic improvement and in-vitropreservation techniques, includingcryopreservation, for conserving the biodiversity.Significant achievements made in these areas ofresearch are summarized here.
MicropropagationIn coffee, micropropagation is possible by microcutting production and somatic embryogenesis.Various experiments conducted to developmicropropagation protocols for Indian cultivars ofcoffee has resulted in plant regeneration throughsomatic embryogenesis from stem and leaf tissuesin more than 20 genotypes/clones of arabica,robusta and hybrid selections (Raghuramulu et ai.,1987; Babu et al..I993: Jayashree et al., 1995;Muniswamy & Sreenath. 1995b; Sreenath, 1998b).Genotypic differences were found in respect tocallus induction, somatic embryogenesis and plantregeneration in arabica coffee (Naidu et aI. 1999).Encapsulation techniques were standardized(Muniswamy & Sreenath, 1995a). Hardeningprotocols are developed for small scale(Muniswamy et al., 1994) and medium scale plantproduction and trial plots of tissue cultured plantsare being established in different agroclimaticzones (Sreenath, 1998b). Three improvedselections viz.. Cauvery, Sln.9 and CxR are being

used for large scale field evaluation ofmicropropagated plants against seedling progeny.Plant regeneration is achieved from the apicalbud and nodal explants (Ganesh & Sreenath,1997), but further refinement ofthe technology isrequired. Effect of TIBA and BAP is tested onintegument cultures of C. canephora (Babu etal., 1997). For the first time in any plantspecies, plant regeneration is ac hieved fromintegument tissues of CxR cultivar (Sreenath etal., 1995).
Genetic improvementWith the objective of using the tissue culturetechnology for genetic improvernent, researchwas done in embryo culture, anther culture andendosperm culture. The research has resulted inplant regeneration through embryo culture bydirect germination as well as somaticembryogenesis (Muniswamy et al.,1993;Sreenath et al., 1989; Muniswamy & Sreenath,1997). Plants produced from the embryo culturehave been established in the field. In addition tothis, plants regenerated from 3 interspecific(Sreenath et al., 1992) and 5 intervarietal crossesthrough embryo rescue methods are establishedin the field and are being evaluated. Plantletshave been regenerated from the anther culture ofCxR cultivar (Muniswamy and Sreenath,communicated). This is the first achievement ina diploid species of coffee. Callus cultures wereestablished from endosperm tissues(Raghurarnulu, 1989) and plantlets areregenerated through embryogenesis from theendosperm calli of S. 2803 (Muniswamy &Sreenath, communicated).Protocols have been optimized for isolation ofprotoplasts from embryogenic calli (Mamatha &Sreenath, 1998). ONA markers are becomingpowerful tools in the hands of coffee breeders(Sreenath 1998a, 1999). Protocols have beendeveloped for isolation of ONA and producingRAPO (Ram & Sreenath, communicated) andAFLP markers (Sreenath et al., communicated).The RAPO markers are being used for evaluatingleaf rust differentials and genetic fingerprinting.Genetic transformation protocols are beingdeveloped with the objective of geneticengineering of coffee. Transient expression ofgus gene is achieved in leaf tissues of CxRcultivar after co-cultivation with Agrobacteriumtumefaciens (Naveen & Sreenath, unpublished).In vitro preservationIn vitro preservation of zygotic embryos upto 2years under slow growth condition has beenachieved in C. arabica (Naidu & Sreenath,communicated). Successful cryopreservation of
249
zygotic embryos is achieved in three species(Krishna & Sreenath, unpublished).
FUTURE AREAS OF RESEARCH
There is great potential for application ofbiotechnology for genetic improvement of Indiancoffee cultivars and growing coffee in an ecofriend Iy way. Resu Its obtai ned so far areencouraging. Now the efforts are being focussedon converting the vast potential of biotechnologyinto reality.Oeveloping molecular markers linked to leaf rustresistance, characterization of coffee germplasmwith ONA markers, engineering leafrust resistancein coffee with anti fungai genes from heterologoussources, isolation of genes of leaf rust resistancefrom resistant cultivars, developing Bt basedbiopesticides to combat major pests, engineeringinsect resistance using Bt and proteinase inhibitorgenes, isolating caffeine degradation genes formanipulation of caffeine conten in the beans, areidentified as thrust areas of coffee biotechnologyresearch in India.
REFERENCES
Anonymous (1996). Coffee Guide, Central CoffeeResearch lnstitute, India.Anonymous (1998). A compendium on pests anddiseases of coffee and their management in India.Central Coffee Research lnstitute, India.Babu KH, Shanta HM, Ganesh OS and SreenathHL (1993). Plant regeneration through highfrequency somatic embryogenesis from leaf tissuesin two cultivars of coffee. J. Plantations Crops 21(supp.): 307 -312.Babu KH, Jayashree G, Naidu MM, Ganesh OSand Sreenath HL (1997). Effect of T1BA andBAP on integument cultures of CxR cultivar ofrobusta coffee (Coffea canephora). In: GARavishankar and LV Venkataraman (Eds).Biotechnological Applications of Plant Tissue andCell Culture. pp 296-300. Oxford and IBHPublishing Co. Pvt. Ltd., New Oelhi.Ganesh OS and Sreenath HL (1997). Shootregeneration from apical bud and node cultures infour clones of robusta coffee (Coffea canephora).In: GA Ravishankar and LV Venkataraman (Eds).Biotechnological Applications of Plant Tissue andCell culture. pp 301-306. Oxford and IBHPublishing Co. Pvt. Ltd.. New Oelhi.Jayashree G, Ganesh DS and Sreenath HL (1995).Effect of kinetin on germination of somaticembryos of CxR cultivar of coffee. J. Coffee Res.25(2): 102-105.

Mamatha HN and Sreenath HL (1998).Optimization of conditions for isolation andculture of coffee protoplasts from embryogeniccalli. Paper presented at the XIII PLACROSYMheld at Coimbatore between 14-18 th December1998.Muniswamy B, Naidu MM and Sreenath HL(1993). Somatic ernbryogenesis and plantregeneration from cultured immature zygoticembryos of Coffea canephora Pierre. J.Plantation Crops 21 (suppl.): 346-350.Muniswamy B, Naidu MM and Sreenath HL(1994). A simple procedure for hardening of invitro raised plantlets of Coffea canephora. J.Coffee Res. 24( 1): 49-53.Muniswamy B and Sreenath HL (1995a).Standardization of encapsulation techniques forproducing synthetic seeds in coffee. J. CoffeeRes.25: 24-29.Muniswamy B and Sreenath HL (1995b). Highfrequency somatic embryogenesisfrom cultured leaf explants of Coffea canephoraon a single medium. J. Coffee Res. 25: 98-10\.Muniswamy B and Sreenath HL (1997).Genotype specificity of direct somaticembryogenesis from cultured embryos in coffee.In: GA Ravishankar and LV Venkataramanan(Eds). Biotechnological Applications of PlantTissue and Cell Culture. pp. 204-209. Oxfordand IBH Publishing Co. Pvt. Ltd., New Delhi.Naidu MM, Ganesh DS, Jayashree G andSreenath HL (1999). Effect of genotype onsomatic embryogenesis and plant regeneration inarabica coffee. In: PB Kavi Kishor (Ed). PlantTissue Culture and Biotechnology- EmergingTrends. pp. 90-95. Universities Press,Hyderabad.Raghuramulu Y (1989). Anther and endopsermcultures of coffee. J. Coffee Res. 19: 71- 81.Raghuramulu Y. Purushotham K. SreenivasanMS and Ramaiah PK ( 1987). In-vitroregeneration of coffee plantlets in India. J.Coffee Res. 17: 57-64.
250
Sondahl MR and Lauritis JA (1992). Coffee. In:RE Por. Hammerschlog and RE Litz (Eds).Biotechnology of Perennial Fruit Crops. pp. 401420.Sondahl MR and Loh WHT (1988). CoffeeBiotechnology In: RJ Clarke and RE Litz (Eds).Coffee Agronomy Vol. 4: pp 236-262 ElevierApplied Science, London.Sreenath HL (1998a). DNA marker techniquescome to the help of coffee breeders. Indian CoffeeLXII (2): 11-13.Sreenath HL (1998 b). Micropropagation ofsuperior coffee genotypes. Indian Coffee LXII (4):17-19.Sreenath HL (1999). Use of molecular markers incoffee. In: PB Kavi Kishor (Ed.) Plant TissueCulture and Biotechnology - Emerging Trends.pp. 289-293. Universities Press (India) Ltd ..Hyderabad.Sreenath HL, Muniswamy B. Raghuramulu YandRamaiah PK (1989). Immature ernbryo culture incoffee. In: XIII Plant Tissue Culture Conferenceheld Shil\ong from 18th-20th Dec. 1989. p.17(Abstracts).Sreenath HL, Muniswamy B, Naidu MM,Dharmaraj PS and Ramaiah PK (1992). Embryoculture of three interspecific crosses in coffee. J.Plantation Crops 20 (suppl.): 243-247.Sreenath HL and Naidu R (1997). Quarter centuryof research in coffee biotechnology - a review. In:GA Ravishankar and LV Venkataraman (Eds).Biotechnological Applications of Plant Tissue andCell Culture. pp. 14-27. Oxford IBH PublishingCo. Pvt. Ltd., New Delhi.Sreenath HL, Shantha HM, Babu KH and NaiduMM (1995). Somatic embryogenesis fromintegument (perisperm) cultures of coffee. PlantCel\ Reports 14: 670-673.Staritsky G (1970). Embryoid formation in calluscultures of coffee. Acta Botanica Neerlandica 19:509-514.
j

251
CUSTO DE PRODUÇÃO MANUAL DE SEMENTES HÍBRIDAS DE CAFÉ I
Sergio Fadelli 2 & Tumoru Sera''
I Trabalho financiado parcialmente pelo Consórcio de P & D Café; 2 Acadêmico de Eng. Agronômica/UEL.bolsista do PlBIC/CNPq/IAPAR; 3 Eng". Agr"., D. Sc., pesquisador do IAPAR
INTRODUÇÃO
Apesar do alto potencial produtivo de caféalcançável palas cultivares de café recomendadaspara o Paraná (> 75 sacas beneficiadas de 60kg/ha/ano, incluindo anos sem produçãodecorrentes da poda) (SERA, 1995) aprodutividade média estadual está muito aquémde uma cafeicultura racional e competitiva (14sc/ha/ano, segundo DEMARCHI, 1998). Paramelhorar esta situação, uma alternativa estuda é aformação de lavoura comercial tipo híbrido F..aproveitando a heterose normalmente observadanas hibridações, e melhorando, desta forma, arentabilidade, principalmente nas pequenas emédias propriedades que representam a grandemaioria da área cafeeira do Estado. Váriostrabalhos tem mostrado a existência de heteroseem Coffea arabica (SRINIVASAN &VISHVESHAVARA, 1978; ARAÚJO & NETO,I980;WALYARO, 1983; AMEHA &BELACHEW, 1985; NETO et aI., 1993;BERTRAND et aI., 1997) com uma médiasuperior a 30% e relato de até 236% em relação aopai superior (WALYARO. 1983). podendo estaser utilizada na obtenção de cultivares tipo híbridoF). Caso a venda da semente híbrida FI seja
. anti econômica, pode-se em alguns casos cultivarcomercialmente as plantas híbridas F2,
aproveitando cerca de metade da heteroseobservado no híbrido FI (SERA & ALVES,1999).A lavoura comercial de híbridos FI pode, ainda.representar para o melhoramento genético umagrande economia de tempo com a redução donúmero de ciclos de seleção necessários paraobtenção de cultivares do tipo linhagem(BERTHOULY. 1997) e permitindo que omelhorista possa testar diversas combinações delinhagens, lançando as mais promissoras comocultivares num período muito menor (SERA &ALVES. 1999).
MATERIAL E MÉTODOS
Na hibridação manual foi usado o cruzamentosimples entre as cultivares Mundo Novo(linhagem 376-4), Catuaí (linhagem H2077-2-581) e Icatu Precoce (linhagem 3282) comoparentais femininos com as cultivares IAPAR 59 eTupi como parentais masculinos. A emasculação e
a polinização foram realizadas em duas floradasocorridas em 1998. A emasculação foi realizadatrês a quatro dias antes da abertura das flores.conforme as condições climáticas atuavam sobre afisiologia do cafeeiro. dando preferência para osramos da metade da saia do lado de exposição aosol pela manhã. A coleta do pólen ocorreu no finalda tarde do dia anterior a abertura das floresquando estas se encontravam no estágio de botãofloral desenvolvido. A polinização ocorreu noperíodo mais quente do dia (a partir das 10:00 atéo final da tarde). O acompanhamento e limpezados cruzamentos para eliminar frutos e floresprovenientes de floradas anteriores e posterioresda florada em que foi realizado o cruzamentoocorreu em todo período que o cafeeiro floresceuem 1998 (agosto a dezembro). quatro a cinco diasantes da abertura das flores. A % de frutificação ede frutos "mocas" foi avaliada em janeiro(aproximadamente quatro meses após apolinização). Quando maduros os frutosprovenientes dos cruzamentos serão colhidos.descascados, degomados e semeados emgerminador de viveiro.O custo de produção manual de sementes híbridasF) em café foi estimado através de duas equaçõespropostas:
a)Determinação do nO de flores a serememasculadas (E) para obter um Kg de semente:
E = S + (J - (b + 2) + (J - (c + 2» + (J - d)
S = nO de sementes /Kg (5.000 para cultivares depeneira 16, 4.500 para cultivares de peneira 17 e4.000 para cultivares de peneira 18); b = taxa defrutos "moca" nos cruzamentos; c = taxa de frutos"chochos" nos cruzamentos e d = taxa de floresque não frutificaram.
b) Determinação do custo/kg de sementehíbrida:
Somando-se o tempo gasto na emasculação, napolinização e na limpeza e dividindo o valorobtido pelo nO de horas efetivamentetrabalhadas/dia/homem (no nosso caso adotamosseis horas/dia/homem) obtém-se o n" de diáriasnecessárias para produção manual de um kg desemente híbrida FI' Multiplicando-se o valor deuma diária pelo n" de diárias necessárias para
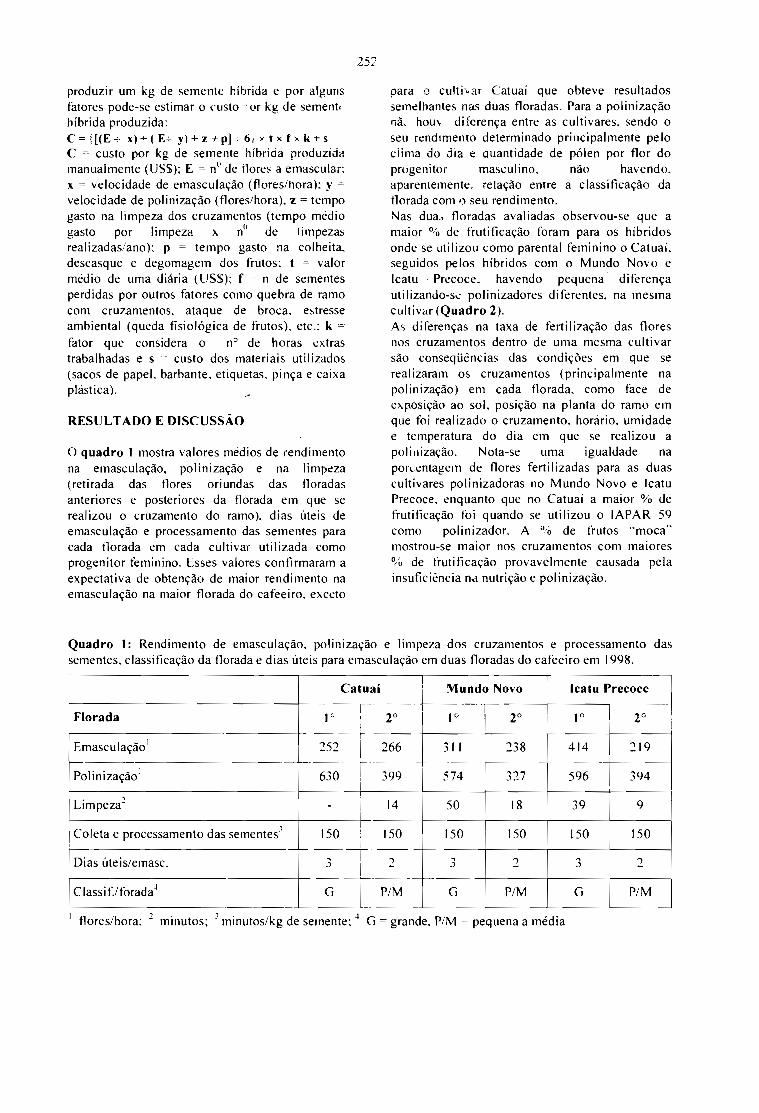
produzir um kgde semente híbrida e por algunsfatores pode-se estimar o custo· OI' kg de sementhíbrida produzida:C = {[(E.;- x) ... ( E.;- YI + z + p1-i- 6 ( " t x f" k + s
C = custo por kg de semente híbrida produzidamanualmente (US$); E = n'' de flores a ernascular:x = velocidade de ernasculação (flores/hora}; y =
velocidade de polinização (flores/hora). z = tempogasto na limpeza dos cruzamentos (tempo médiogasto por limpeza x n'' de limpezasreal izadas/ano): p = tempo gasto na colheita.descasque e degomagem dos frutos; t = valormédio de uma diária (lIS$); f .c n de sementesperdidas por outros fatores como quebra de ramocom cruzamentos. ataque de broca. estresseambiental (queda fisiológica de frutos). etc.: k =fator que considera o na de horas extrastrabalhadas e s = custo dos materiais utilizados(sacos de papel. barbante. etiquetas. pinça e caixaplástica).
RESULTADO E mSCLJSsÁO
() quadro I mostra valores médios de rendimentona emasculação, polinização e na limpeza(retirada das flores oriundas das floradasanteriores e posteriores da florada em que serealizou o cruzamento do ramo). dias úteis deernasculação e processamento das sementes paracada tlorada em cada cultivar utilizada comoprogenitor feminino. Esses valores confirmaram aexpectativa de obtenção de maior rendimento naernasculação na maior florada do cafeeiro. exceto
para o cultivar Catuai que obteve resultadossemelhantes nas duas floradas. Para a polinizaçãonã. houv diferença entre as cultivares. sendo oseu rendimento determinado principalmente peloclima do dia e quantidade de pólen por flor doprogenitor masculino. não havendo.aparentemente. relação entre a classificação datlorada com \) seu rendimento.Nas dua, floradas aval iadas observou-se que amaior % de frutificação foram para os híbridosonde se utilizou como parental feminino o Catuaí,seguidos pelos híbridos com o Mundo Novo elcatu . Precoce. havendo pequena diferençautilizando-SI: polinizadores diferentes. na mesmacultivar (Quadro 2).As diferenças na taxa de fertilização das floresnos cruzamentos dentro de uma mesma cultivarsão conseqüências das condições em que serealizaram os cruzamentos (principalmente napolinização) em cada florada. como face deexposição ao sol. posição na planta do ramo emque foi realizado o cruzamento. horário. umidadee temperatura do dia em que se realizou apolinização. Nota-se uma igualdade naporcentagem de flores fertilizadas para as duascultivares polinizadoras no Mundo Novo e lcatuPrecoce. enquanto que no Catuaí a maior % defrutificação foi quando se utilizou o IAPAR 59como polinizador. A % de frutos "rnoca"mostrou-se maior nos cruzamentos com maiores% de frutificação provavelmente causada pelainsuficiência na nutrição e polinização.
Quadro I: Rendimento de ernasculação, polinização e limpeza dos cruzamentos e processamento dassementes. classificação da florada e dias úteis para ernasculação em duas floradas do cafeeiro em 1998.
I !I
Catuaí
I
Mundo Novo Icatu Precoce
~ IFlorada I c , 2° I v 20 la 2° i
I Emasculação '
I--+ ----lo ~52 -+_ 26~_ 311 ~38 414 219
f~J;n;za','O'__o ----------------
I 630 i 399 574 327 596 394I
I Limpeza I - ! 14 50 18 39~i
IColeta e processamento das sementes' 150 I 150 150 150 150 150 I
Dias úteisiemasc.
Classif'/forada"~----------_._-------
I flores/hora; 2 minutos; J minuros/kg de semente; ~ G = grande. P/M - pequena a média
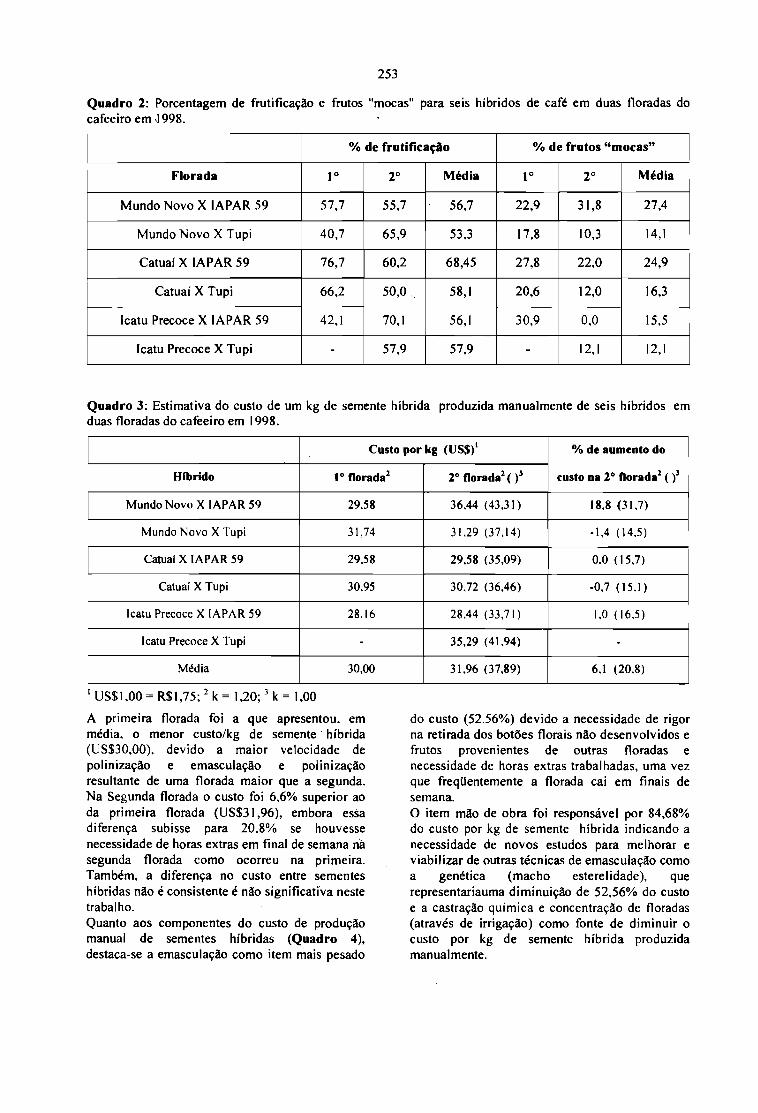
253
Quadro 2: Porcentagem de frutificação e frutos "mocas" para seis híbridos de café em duas floradas docafeeiro em ,1998.
% de frutificação 0/0 de frutos "mocas"
Florada )0 r Média )0 r Média
Mundo Novo X IAPAR 59 57,7 55,7 56,7 22,9 31,8 27,4
Mundo Novo X Tupi 40,7 65,9 53,3 17,8 10,3 14,1
Catuaí X IAPAR 59 76,7 60,2 68,45 27,8 22,0 24,9
Catuaí X Tupi 66,2 50,0. 58,1 20,6 12,0 16,3
Icatu Precoce X IAPAR 59 42,1 70,1 56,1 30,9 0,0 15,5
Icatu Precoce X Tupi - 57,9 57,9 - 12,1 12,1
Quadro 3: Estimativa do custo de um kg de semente híbrida produzida manualmente de seis híbridos emduas floradas do cafeeiro em 1998.
Custo por kg (US$)I % de aumento do I
Hrbrido 1° florada l 20 floradal ( )3 custo na 2° florada l( )3
Mundo Novo X IAPAR 59 29.58 36,44 (43,31) 18,8 (31,7)
Mundo Novo X Tupi 31,74 31,29 (37.14) ·1,4 (14.5)
Catuaí X IAPAR 59 29.58 29,58 (35,09) 0.0 (15.7)
Catuaí X Tupi 30.95 30.72 (36,46) ·0.7 (15.1)
Icatu Precoce X IAPAR 59 28.16 28,44 (33.71) 1.0 (16.5)
Icatu Precoce X Tupi . 35,29 (41,94) -Média 30.00 31,96 (37,89) 6.1 (20.8)
I US$l.OO = R$I,75; 2 k = 1,20; 3 k = 1,00
A primeira florada foi a que apresentou, emmédia, o menor custo/kg de semente híbrida(US$30,OO), devido a maior velocidade depolinização e emasculação e polinizaçãoresultante de uma florada maior que a segunda.Na Segunda florada o custo foi 6,6% superior aoda primeira florada (US$3I,96), embora essadiferença subisse para 20,8% se houvessenecessidade de horas extras em final de semana nasegunda florada como ocorreu na primeira.Também, a diferença no custo entre sementeshíbridas não é consistente é não significativa nestetrabalho.Quanto aos componentes do custo de produçãomanual de sementes híbridas (Quadro 4),destaca-se a ernasculação como item mais pesado
do custo (52.56%) devido a necessidade de rigorna retirada dos botões florais não desenvolvidos efrutos provenientes de outras floradas enecessidade de horas extras trabal hadas, uma vezque freqüentemente a florada cai em finais desemana.O item mão de obra foi responsável por 84,68%do custo por kg de semente híbrida indicando anecessidade de novos estudos para melhorar eviabilizar de outras técnicas de emasculação comoa genética (macho esterelidade), querepresentariauma diminuição de 52,56% do custoe a castração química e concentração de floradas(através de irrigação) como fonte de diminuir ocusto por kg de semente híbrida produzidamanualmente.
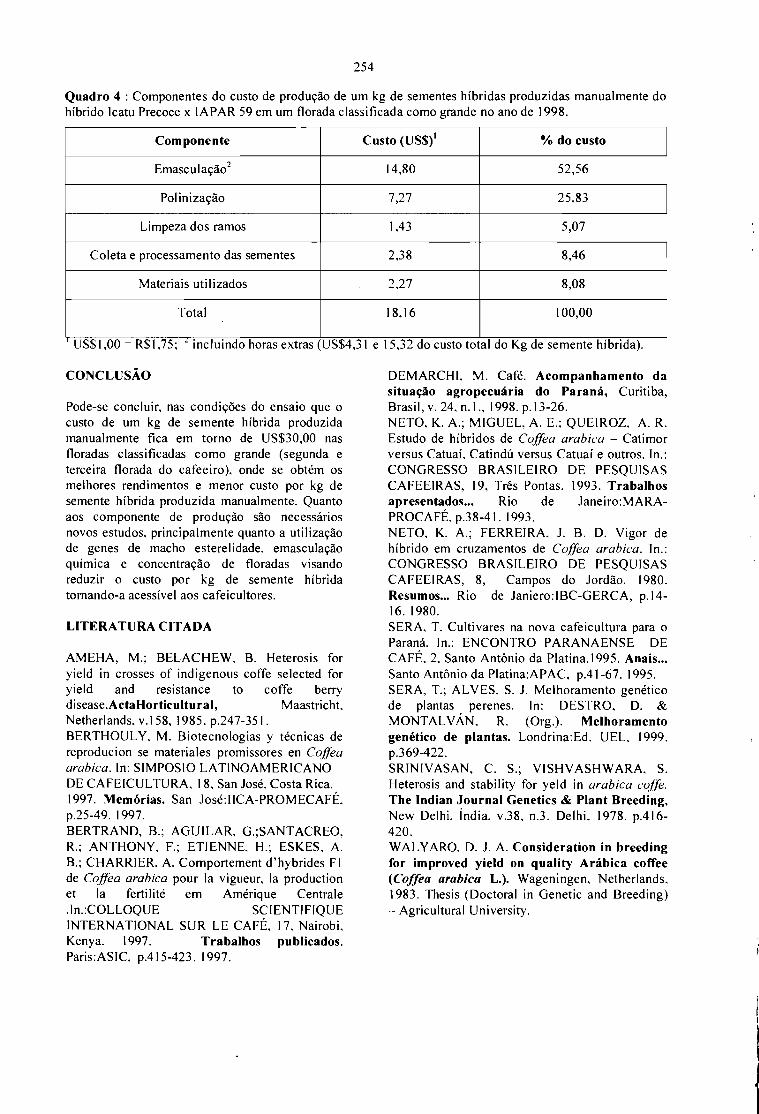
254
Quadro 4 : Componentes do custo de produção de um kg de sementes híbridas produzidas manualmente dohíbrido lcatu Precoce x IAPAR 59 em um florada classificada como grande no ano de 1998.
US$I,OO - R$1,75, incluindo horas extras (US$4,31 e 15,32 do custo total do Kg de semente híbrida).
Componente Custo (USS)\ % do custo
Emasculação/ 14,80 52,56
Polinização 7,27 25,83
Limpeza dos ramos 1,43 5,07
Coleta e processamento das sementes 2,38 8,46
Materiais utilizados 2,27 8,08
Total 18,16 100,00
- -
CONCLUSÃO
Pode-se concluir, nas condições do ensaio que ocusto de um kg de semente híbrida produzidamanualmente fica em torno de US$30,00 nasfloradas classificadas como grande (segunda eterceira florada do cafeeiro), onde se obtém osmelhores rendimentos e menor custo por kg desemente híbrida produzida manualmente. Quantoaos componente de produção são necessáriosnovos estudos, principalmente quanto a utilizaçãode genes de macho esterelidade, emasculaçãoquímica e concentração de floradas visandoreduzir o custo por kg de semente híbridatornando-a acessível aos cafeicultores.
LITERATURA CITADA
AMEHA, M.; BELACHEW, 8. Heterosis foryield in crosses of indigenous coffe selected foryield and resistance to coffe berrydisease.ActaHorticultural, Maastricht,Netherlands, v.158, 1985. p.247-351.BERTHOULY, M. Biotecnologias y técnicas dereproducion se materiales promissores en Coffeaarabica. In: SIMPOSIO LATINOAMERICANODE CAFEICULTURA, 18, San José, Costa Rica.1997. Memórias, San José:IICA-PROMECAFÉ,p.25-49. 1997.BERTRAND, 8.; AGUILAR, G.;SANTACREO,R.; ANTHONY, F.; ETIENNE, H.; ESKES, A.8.; CHARRIER, A. Comportement d'hybrides FIde Coffea arabica pour la vigueur, la productionet la fertilité em Amérique Centrale.ln.:COLLOQUE SCIENTIFIQUEINTERNATIONAL SUR LE CAFÉ, 17, Nairobi,Kenya. 1997. Trabalhos publicados.Paris:ASIC, pAI5-423. 1997.
DEMARCHI, M. Café. Acompanhamento dasituação agropecuária do Paraná, Curitiba,Brasil, v. 24, n.L, 1998. p.13-26.NETO, K. A.; MIGUEL, A. E.; QUEIROZ, A. R.Estudo de híbridos de Coffea arabica - Catimorversus Catuaí, Catindú versus Catuaí e outros. In.:CONGRESSO BRASILEIRO DE PESQUISASCAFEEIRAS, 19, Três Pontas. 1993. Trabalhosapresentados... Rio de Janeiro:MARAPROCAFÉ, p.38-41. 1993.NETO, K. A.; FERREIRA, J. B. D. Vigor dehíbrido em cruzamentos de Coffea arabica. In.:CONGRESSO BRASILEIRO DE PESQUISASCAFEEI RAS, 8, Campos do Jordão. 1980.Resumos... Rio de Janiero:IBC-GERCA, p.1416.1980.SERA, T. Cultivares na nova cafeicultura para oParaná. In.: ENCONTRO PARANAENSE DECAFÉ, 2, Santo Antônio da Platina. 1995. Anais...Santo Antônio da Platina:APAC, pAI-67. 1995.SERA, T.; ALVES, S. J. Melhoramento genéticode plantas perenes. In: DESTRO, D. &MONTALVÁN, R. (Org.). Melhoramentogenético de plantas. Londrina:Ed. UEL, 1999.p.369-422.SRINIVASAN, C. S.; VISHVASHWARA, S.Heterosis and stability for yeld in arabica coffe.The Indian Journal Genetics & Plant Breeding,New Delhi, índia, v.38, n.3. Delhi, 1978. pA16420.WALYARO, D. J. A. Consideration in breedingfor improved yield on quality Arábica cotTee(Coffea arabica L.). Wageningen, Netherlands,1983. Thesis (Doctoral in Genetic and Breeding)- Agricultural University .

255
ESTUDO DA CAPACIDADE COMBINATÓRIA E DA HETEROSE EM CRUZAMENTOS ENTRELINHAGENS DE CATUAÍ E HÍBRIDO DE TIMOR, EM CAFÉ.
José Roberto M. Fontes", Antônio A. Cardoso', Cosme Damião Cruz' Laércio Zambolím", Ney S.Sakiyama', Antônio A. Pereira",
IDFT/UFV; 2DBG/UFV; 3DFP/UFV; 4EPAMIG. 36571-000 - Viçosa-MG.Auxílio financeiro: FINEP,FAPEMIG, CNPq e PNP&DCafé
INTRODUÇÃO
A análise da capacidade combinatória é umametodologia utilizada para identificarprogenitores com capacidade de transmitir seuscaracteres desejáveis a sua descendência,identificar a melhores combinações híbridas eadquirir informações sobre o tipo de ação gênicaque controla os diferentes caracteres agronômicos.Em sua forma mais geral, capacidadecombinatória ou habilidade combinatória refere-seao comportamento de linhagens ou cultivaresquando são usadas em combinações em um ou emvários sentidos, entre si, para obtenção dehíbridos. Sendo a capacidade geral de combinaçãoo comportamento de uma linhagem em sua sériede cruzamentos, tomando por base o valor médiodos híbridos FI nestes cruzamentos. Já odesempenho de um cruzamento particular podedesviar-se da média da capacidade geral decombinação, e este desvio é conhecido comocapacidade específica de combinação (ALLARD,1971).Os termos capacidade geral de combinação(CGC) e capacidade específica de combinação(CEC) foram originalmente definidos por Sprague& Taturn, citados por MELO (1987), utilizandocomo método experimental um sistema decruzamentos dialélicos. Os autores conceituam aCGC como o comportamento médio de umalinhagem em uma série de cruzamentos. Por suavez, a CEC representa o desvio, para melhor oupior, de um determinado cruzamento, tomandopor base a média da CGC dos seus pais. Elesdiferenciam, ainda, os tipos de capacidadecombinatória considerando o modo de açãogênica envolvida. Por conseguinte, a CGC estáassociada a genes de efeitos principalmenteaditivos, além da parte dos efeitos dominantes eepistáticos (aditivo X aditivo). De outro lado, aCEC depende basicamente de genes com efeitodominantes, epistáticos e vários tipos deinterações.Estudos sobre a capacidade específica decombinação de híbridos de C. canephora foramrealizados por FAZUOLi & CARVALHO (1987).Verificou-se que as melhores combinações foramas de Kouillou x Robusta, BP46 x Kouillou,
Uganda x SA 158 e a da progênie de polinizaçãolivre de UgandaI646-4. Os autores mencionaramo descarte de várias combinações híbridas, quenada produziram ou produziram poucas sementes,que germinaram mal; sendo este fato atribuído àauto-incompatibilidade típica da espécie C.canephora. CILAS et alii (1998), tambémtrabalhando com café, mostram que linhagens debaixo rendimento podem apresentar excelenteperformance como progenitores. Por outro lado,excelentes variedades podem ser péssimosprogenitores. Por isso, tanto o estudo da CGCquanto da CEC são essenciais na escolha doscruzamentos a serem realizados num programa demelhoramento.Nos estudos de heterose, maior ênfase tem sidodada predominantemente às espécies alógamas,particularmente à cultura do milho, onde aheterose é explorada comercialmente. Entretanto,têm crescido as atenções dadas às espéciesautógamas, motivada em parte pela possibilidadedo desenvolvimento de sistemas efetivos paraexploração comercial da heterose via híbridos Fiotambém nestas espécies, e pelo potencial dautilização dos dados de rendimento de híbridos FIe outros materiais em gerações iniciais.permitindo ao melhorista dar prioridade aosmelhores cruzamentos (SARAWAT et alii, 1994).O valor da heterose é freqüentemente expresso emporcentagem, eliminando, assim a unidade usadapara a determinação do caráter. Do ponto de vistaprático, tem interesse o vigor do híbrido quandoeste é superior ao pai superior (heterobeltiose).Freqüentemente, um cruzamento pode exibirheterose, por ser mais produtivo do que a médiados pais, porém, se não for superior a um dosprogenitores, pode não ter interesse prático(PATERNIANI, 1975).Há muito, têm sido observado em várias espéciesde plantas que cruzamentos entre progenitoresgeneticamente divergentes exibem maioresheteroses que cruzamentos entre progenitores domesmo grupo (MILLER & MARANI, 1963).Portanto, nem todas as combinações híbridasexibem heterose, o que pode ser facilmenteentendido, já que quanto mais geneticamentedistintos forem os progenitores, maiorprobabilidade de contribuírem com alelos

diferentes, resultando em maior heterozigosidadeno híbrido, e assim obter maior vigor, quer devidoà heterozigosidade por si (genes heteróticos), querdevido a complementação dos genes para vigor(genes dominantes ou parcialmente dominantespara o vigor). De outro lado, progenitoresaparentados terão muitos alelos em comum,reduzindo assim as possibilidades de heterose. Porisso, na escolha de linhagens e ou cultivares parahibridização, devem ser cuidadosamenteescolhidas as que apresentarem divergênciasgenéticas entre si e caracteres altamentedesejáveis (PATERNIANI, 1975; GALVÊAS,1988).BELLACHEW et alii (1993) estudaram seismateriais silvestres de C. arabica, cruzados numdialelo parcial padrão. Os híbridos FI e seusprogenitores foram avaliados para sete caracteresvegetativos para obter estimativas da heterose e dacapacidade combinatória. Os híbridos F I
apresentaram heterose positiva em relação aomelhor progenitor para todas as característicasestudadas, com valores variando de 3 a 18%.Sendo que o cruzamento PI x P4 apresentouheterose de 69% para volume de raiz. Em todos oscaracteres estudados, as maiores heterobeltiosesforam observadas em certos cruzamentos nosquais os progenitores possuíam locais de origem ecaracteres morfológicos distintos.SRINIVASAN & VISHVESHWARA (!978)estudaram a heterose para rendimento de grãos emtreze cruzamentos entre sete progenitores em C.arábica durante seis colheitas. Com relação àmédia das seis colheitas, cinco cruzamentosapresentaram heterose positiva em relação àmédia dos pais, e sete cruzamentos apresentaramheterobeltiose. Os autores encontraram valores deheterose de até 183,3 % em relação a médiaparenta! e de até 100,41 % em relação ao melhorparental, no F I obtido do cruzamento entre Agarox 2045. Tanto o progenitor Agaro quanto oChoeche, que fez parte do cruzamento queapresentou a segunda melhor heterose, foramdiretamente introduzidos da Etiópia. Os híbridosFI mais heteróticos possuíam, também, excelenteestabilidade; com possibilidade de aumento naprodução e aquisição de rendimentos estáveis emcafé arábica.BELLACHEW (1997) estudou aproximadamente2.789 acessos silvestres e exóticos de C. arabicaprovenientes da Etiópia, resultando em 279materiais promissores. O autor relata a existênciade heterose, obtidas em híbridos FI, de 69% paracaracterísticas do grão e de 60% para produção.Este trabalho teve por objetivo estimar acapacidade de combinação geral e específica,entre linhagens de Catuaí e descendentes do
256
Híbrido de Timor, e a heterose e heterobeltiosenos híbridos FI.
MATERIAL E MÉTODOS
Utilizando os cafeeiros resistentes à ferrugemselecionados no germoplasma introduzido naUniversidade Federal de Viçosa, iniciou-se, apartir de 1974, a síntese de novas combinaçõesgenéticas portadoras de resistência à ferrugem docafeeiro. Essas novas combinações são resultantesdo intercruzamento dos cafeeiros resistente ecruzamentos destes com cafeeiros das variedadescomerciais do Catuai, Mundo Novo e Bourbon,visando a síntese de genótipos com resistênciamais completa e transferência dos fatores deresistência, respectivamente.Os cultivares Catuai Vermelho e Catuai Amareloque participaram dos cruzamentos fazem parte doensaio de linhagens, implantado em 1978, emViçosa, onde foram avaliadas 12 progênies deCatuai (6 progênies de Catuai Vermelho e 6progênies de Catuai Amarelo).Essas combinações, após avaliadas quanto aferrugem, através de inoculação artificial. foramplantadas nos chamados Campos de Seleção deHíbridos (CSH) para avaliação do comportamentonas condições de cultivo. Atualmente, estão sendoacompanhadas, nos campos CSH I ao CSH IO,plantas em geração FI; já havendo outros camposem gerações mais avançadas.Procurando realizar uma reorganização destecampos de seleção de híbridos, iniciou-se umtrabalho de estudo dos dados de produção dosdiferentes híbridos existentes. Neste trabalhoforam estudados os híbridos FI provenientes doscruzamentos entre variedades de Catuai comdescendentes do Híbrido de Timor. Este materialfaz parte dos Campos de Seleção de Híbridos, doPrograma de Melhoramento UFV/EPAMIG, naEstação de Pesquisa de São José do Triunfo,distrito de Viçosa, M.G.Foram estudados um total de 74 híbridos,descendentes dos cruzamentos envolvendo 14linhagens de Catuai (Vermelho e Amarelo) e 12progênies descendentes do Híbrido de Timor. Aavaliação dos cafeeiros foi realizada através daprodução média acumulada, em gramas de cafécereja.Foram utilizadas como testemunhas três linhagensdo cultivar Catuaí, UFV2145, UFV21 14 eUFV2154.Foi calculada a heterose através do desvio dageração F I em relação à média dos parentais (H mp;
mp= I00) e a heterobeltiose (H ps; ps= 100),heterose em relação ao pai superior, visandoavaliar melhor o valor prático da heterose.
Ij
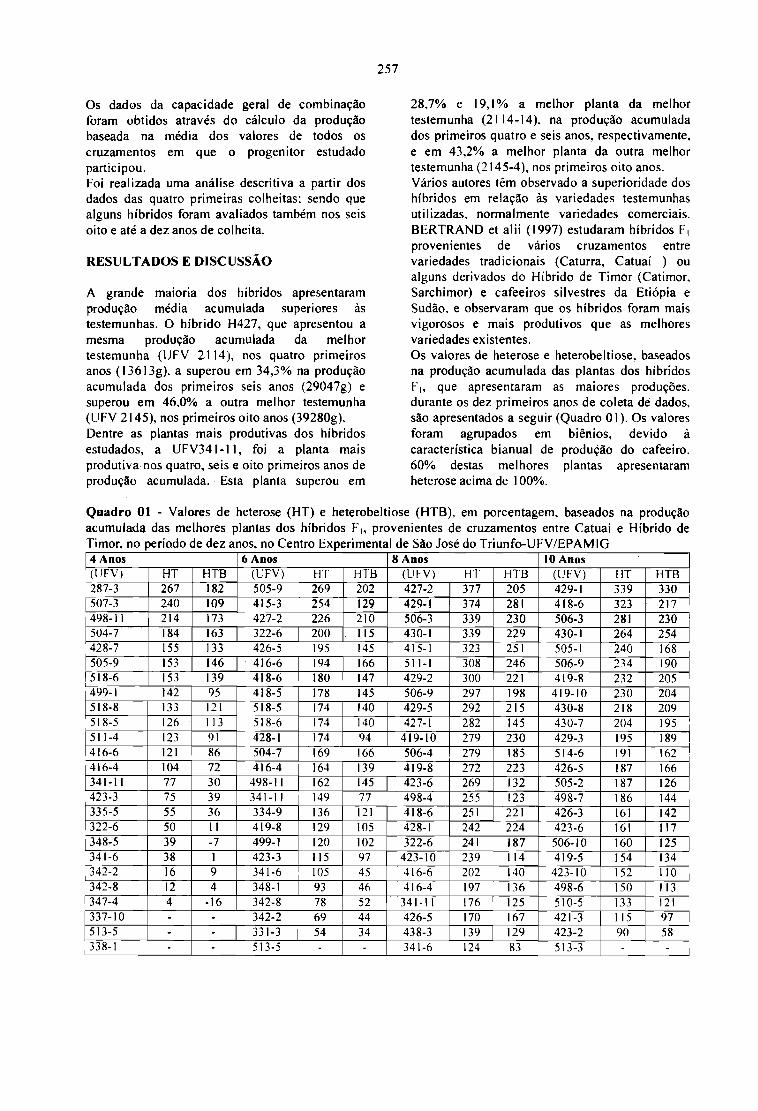
257
Os dados da capacidade geral de combinaçãoforam obtidos através do cálculo da produçãobaseada na média dos valores de todos oscruzamentos em que o progenitor estudadoparticipou.Foi realizada uma análise descritiva a partir dosdados das quatro primeiras colheitas; sendo quealguns híbridos foram avaliados também nos seisoito e até a dez anos de colheita.
RESULTADOS E DISCUSSÃO
A grande maioria dos híbridos apresentaramprodução média acumulada superiores àstestemunhas. O híbrido H427, que apresentou amesma produção acumulada da melhortestemunha (UFV 2114), nos quatro primeirosanos (13613g). a superou em 34,3% na produçãoacumulada dos primeiros seis anos (29047g) esuperou em 46,0% a outra melhor testemunha(UFV 2145), nos primeiros oito anos (39280g).Dentre as plantas mais produtivas dos híbridosestudados, a UFV341-11, foi a planta maisprodutiva nos quatro, seis e oito primeiros anos deprodução acumulada. Esta planta superou em
28,7% e 19,1% a melhor planta da melhortestemunha (21 14-14). na produção acumuladados primeiros quatro e seis anos, respectivamente,e em 43,2% a melhor planta da outra melhortestemunha (2145-4), nos primeiros oito anos.Vários autores têm observado a superioridade doshíbridos em relação às variedades testemunhasutilizadas, normalmente variedades comerciais.BERTRAND et alii (1997) estudaram híbridos FIprovenientes de vários cruzamentos entrevariedades tradicionais (Caturra, Catuaí ) oualguns derivados do Híbrido de Timor (Catirnor,Sarchimor) e cafeeiros silvestres da Etiópia eSudão, e observaram que os híbridos foram maisvigorosos e mais produtivos que as melhoresvariedades existentes.Os valores de heterose e heterobeltiose, baseadosna produção acumulada das plantas dos híbridosFI, que apresentaram as maiores produções.durante os dez primeiros anos de coleta de dados,são apresentados a seguir (Quadro 01). Os valoresforam agrupados em biênios, devido àcaracterística bianual de produção do cafeeiro.60% destas mel hores plantas apresentaramheterose acima de I00%.
Quadro OI - Valores de heterose (HT) e heterobeltiose (HTB), em porcentagem, baseados na produçãoacumulada das melhores plantas dos híbridos F I1 provenientes de cruzamentos entre Catuai e Híbrido deTimor, no período de dez anos, no Centro Experimental de São José do Triunfo-UFV/EPAMIG14 Anos 6 Anos 8 Anos 10 Anos(UFV) HT HTB (UFV) HT HTB (UFV) HT HTB (UFV) HT HTB
1287-3 267 182 505-9 269 202 427-2 377 205 429-1 339 330
I 507-3 240 109 415-3 254 129 429-1 374 281 418-6 323 217
1498-11 214 173 427-2 226 210 506-3 339 230 506-3 281 230 I1504-7 184 163 322-6 200 115 430-1 339 229 430-1 264 254428-7 155 133 426-5 195 145 415-1 323 251 505-1 240 168 I
1505-9 153 146 . 416-6 194 166 511-1 308 246 506-9 234 1901518-6 153 139 418-6 180 147 429-2 300 221 419-8 232 205 I
1499- 1 142 95 418-5 178 145 506-9 297 198 419-10 230 2041518-8 133 121 518-5 174 140 429-5 292 215 430-8 218 209518-5 126 113 518-6 174 140 427-1 282 145 430-7 204 195
1511-4 123 91 428-1 174 94 419-10 279 230 429-3 195 1891416-6 121 86 504-7 169 166 506-4 279 185 514-6 191 162
14 16-4 104 72 416-4 164 139 419-8 272 223 426-5 187 1661341-11 77 30 498-11 162 145 423-6 269 132 505-2 187 126423·3 75 39 341-11 149 77 498-4 255 123 498-7 186 144 I
1335-5 55 36 334-9 136 121 418-6 251 221 426-3 161 142 II322-6 50 11 419-8 129 105 428-1 242 224 423-6 161 117 I
1348-5 39 -7 499-1 120 102 322-6 241 187 506-10 160 125
134 1-6 38 1 423-3 115 97 423-10 239 114 419-5 154 134342-2 16 9 341-6 105 45 416-6 202 140 423-10 152 110 I
1342·8 12 4 348-' 93 46 416-4 197 136 498-6 150 113I 347-4 4 -, 6 342-8 78 52 341-11 176 125 510-5 133 121 I
1337-10 - - 342-2 69 44 426-5 170 167 421-3 115 97 !1513-5 - - 331-3 54 34 438-3 139 129 423-2 90 58338-1 - - 513-5 - - 341-6 124 I 83 513-3 - -
I

VAN DER VOSSEN (1985) mostra ev idênciasconsideráveis do vigor híbrido devido ao efeito degenes epistáticos complementares, pararendimento. particularmente em híbridos Fh
originados de cruzamentos entre variedades deorigens diversas. O autor apresenta dadosacumulados das três primeiras colheitas devariedades e híbridos FI de café arábica em duasdensidades de plantio. Os híbridos FI produziramaté 120% a mais que a média parental (Laurina xHíbrido de Tirnor), para a densidade de 3.333plantas/ha, e até 109% a mais (SL28 x RumeSudan), para a densidade de 6.667 plantas/ha. Asmelhores produções, no entanto, foram obtidaspelos híbridos "Pandang" x "SL34" (15,4% deheterose) e "Pandang" x "Erecta" (38% deheterose), para a menor e a maior densidade deplantio, respectivamente. Entre as variedades,"Pandang" foi a mais produtiva nas duasdensidades estudadas.Das progênies que foram avaliadas em até oitoanos de produção, destacaram-se os híbridos:UFV416-6, UFV416-4, UFV322-6, UFV341-11 eUFV341-6, que apresentaram as melhoresheteroses nos quatro, seis e oito anos de avaliação,UFV418-6 e UFV419-8, que apresentaram asmelhores heteroses nos seis, oito e dez anos deavaliação, e UFV287-3, que apresentou heterosede 267% na avaliação dos quatro anos (períodoem que foi avaliado). Destacaram-se também oshíbridos UFV504-7, UFV505-9, UFV518-6,UFV518-5. UFV342-2 e UFV342-8, queapresentaram heteroses consideráveis. baseadas naprodução acumulada dos quatro e seis anos.O híbrido UFV427-2, apesar de não aparecerentre os melhores nos primeiros quatro anos.apresentou excelente heterose nos dois biêniosseguintes, principalmente nos oito anos com umaheterose de 377% (maior heterose encontrada)
258
Dentre as progênies que apresentaram as melhoresheteroses, encontram-se algumas daquelas queapresentaram também as melhores produções, sãoelas: UFV341-11, UFV341-6, UFV504-7,UFV342-8, UFV518-6, UFV416-6 e UFV416-4.Através do estudo das produções obtidas pelosparentais destas progênies, foi observado que osmesmos apresentaram baixa produção média,indicando que a elevada produção obtida pelasprogênies foi devido ao efeito da heterosemanifestada nas mesmas e não por descenderemde pais com elevada capacidade produtiva.Quando comparamos as produções dos híbridos FIcom o melhor progenitor (heterobeltiose), namaioria das vezes, Catuai, a diferença é reduzida;mostrando que a produção do progenitor Híbridode Timor é que provocava uma redução no valorda média. No entanto, a superioridade de váriasprogênies F" principalmente nos primeiros anosde produção, é confirmada, independente da baixaprodução de um dos progenitores.Os valores de heterobeltiose obtidos nestetrabalho confirmam a superioridade das progêniescitadas anteriormente. mas devemos observar quepossíveis alterações na classificação das mesmasocorrerão se esta classificação for baseada nasmelhores heterobeltioses. Assim, os valores deheterobeltiose também devem ser levados emconsideração, pois, na prática, o que interessa éum híbrido que também supere o parental maisprodutivo (valor prático).MARTINEZ et alii (1988) observaram que aheterose, com relação à média parental, variou de105 a 169%, para uma média geral de 123%. Emrelação ao pai superior, o valor médio da heterosefoi de 102%.Os dados da capacidade geral de combinação,baseados na produção média acumulada, seencontram no Quadro 02.
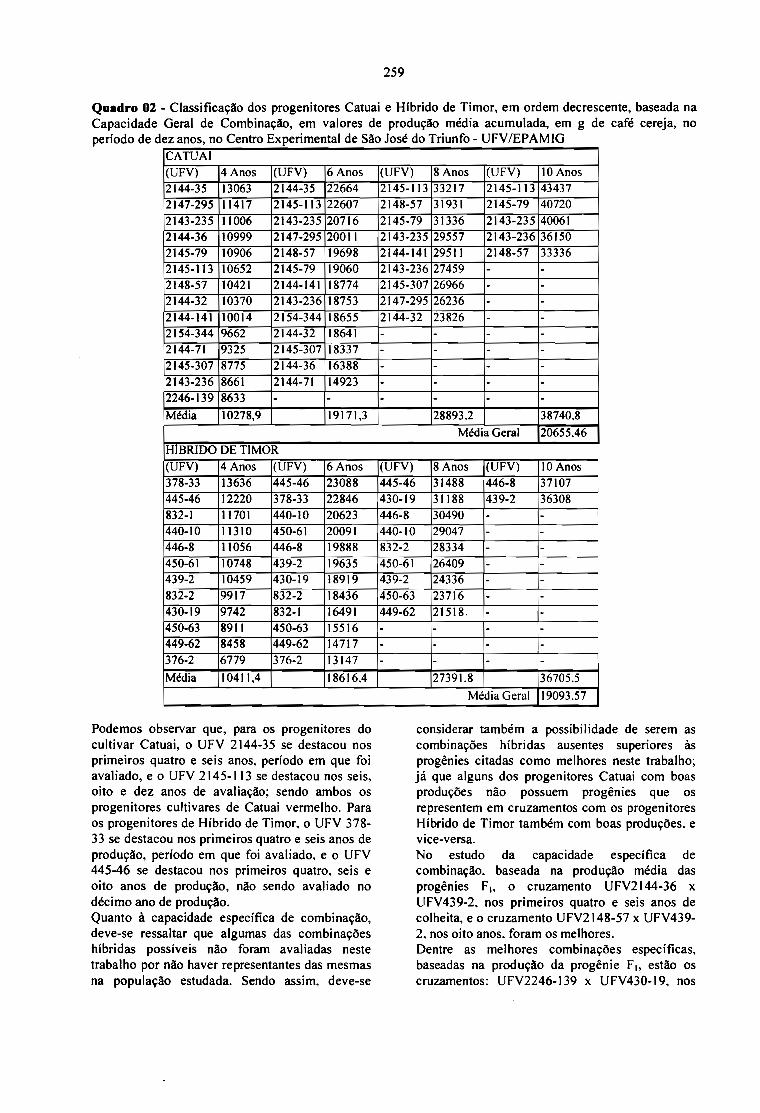
259
Quadro 02 - Classificação dos progenitores Catuai e Híbrido de Timor, em ordem decrescente, baseada naCapacidade Geral de Combinação, em valores de produção média acumulada, em g de café cereja, noperíodo de dez anos, no Centro Experimental de São José do Triunfo - UFVIEPAMIG
CATUAI(UFV) 4 Anos (UFV) 6 Anos (UFV) 8 Anos (UFV) 10 Anos2144-35 13063 2144-35 22664 2145-113 33217 2145-113 434372147-295 11417 2145-113 22607 2148-57 31931 2145-79 407202143-235 11006 2143-235 20716 2145-79 31336 2143-235 400612144-36 10999 2147-295 20011 2143-235 29557 2143-236 361502145-79 10906 2148-57 19698 2144-141 29511 2148-57 333362145-113 10652 2145-79 19060 2143-236 27459 - -2148-57 10421 2144-141 18774 2145-307 26966 - -
I
2144-32 10370 2143-236 18753 2147-295 26236 - -2144-141 10014 2154-344 18655 2144-32 23826 - -2154-344 9662 2144-32 18641 - - - -2144-71 9325 2145-307 18337 - - - -2145-307 8775 2144-36 16388 - - - -2143-236 8661 2144-71 14923 - - - -2246-139 8633 - - - - - -Média 10278,9 19171,3 28893,2 38740,8
MédiaGeral 20655,46HIBRIDO DE TIMOR(UFV) 4 Anos (UFV) 6 Anos (UFV) 8 Anos (UFV) 10Anos378-33 13636 445-46 23088 445-46 31488 446-8 37107445-46 12220 378-33 22846 430-19 31188 439-2 36308832-1 11701 440-10 20623 446-8 30490 - -440-10 11310 450-61 20091 440-10 29047 - -446-8 11056 446-8 19888 832-2 28334 - -450-61 10748 439-2 19635 450-61 26409 - -439-2 10459 430-19 18919 439-2 24336 - -832-2 9917 832-2 18436 450-63 23716 - -430-19 9742 832-1 16491 449-62 21518 - -450-63 8911 450-63 15516 - - - -449-62 8458 449-62 14717 - - - -376-2 6779 376-2 13147 - - - -Média 10411,4 18616.4 27391.8 36705,5
Média Geral 19093,57
Podemos observar que, para os progenitores docultivar Catuai, o UFV 2144-35 se destacou nosprimeiros quatro e seis anos, período em que foiavaliado, e o UFV 2145-113 se destacou nos seis,oito e dez anos de avaliação; sendo ambos osprogenitores cultivares de Catuai vermelho. Paraos progenitores de Híbrido de Tirnor, o UFV 37833 se destacou nos primeiros quatro e seis anos deprodução, período em que foi avaliado, e o UFV445-46 se destacou nos primeiros quatro. seis eoito anos de produção, não sendo avaliado nodécimo ano de produção.Quanto à capacidade específica de combinação,deve-se ressaltar que algumas das combinaçõeshíbridas possíveis não foram avaliadas nestetrabalho por não haver representantes das mesmasna população estudada. Sendo assim, deve-se
considerar também a possibilidade de serem ascombinações híbridas ausentes superiores àsprogênies citadas como melhores neste trabalho;já que alguns dos progenitores Catuai com boasproduções não possuem progênies que osrepresentem em cruzamentos com os progenitoresHíbrido de Timor também com boas produções, evice-versa.No estudo da capacidade específica decombinação. baseada na produção média dasprogênies FI, o cruzamento UFV2144-36 xUFV439-2. nos primeiros quatro e seis anos decolheita, e o cruzamento UFV2148-57 x UFV4392. nos oito anos. foram os melhores.Dentre as melhores combinações específicas.baseadas na produção da progênie FI, estão oscruzamentos: UFV2246-139 x UFV430-19. nos

primeiros quatro anos, UFV2144-36 x UFV4392, UFV2144-35 x UFV445-46 e UFV2144-141 xUFV439-2, nos seis anos, UFV2143-235 xUFV445-46, nos oito anos, e UFV2145-79 xUFV446-8, no dez anos de produção acumulada.Segundo Cruz e Regazzi, citados porCAVALCANTI et alii (1997), o melhor hibrido éaquele que apresenta a maior estimativa dacapacidade especifica de combinação e que pelomenos um dos pais possui alta estimativa dacapacidade geral de combinação. Assim sendo, ohíbrido proveniente do cruzamento UFV2144-35x UFV445-46, que se destacou nos seis anos deprodução acumulada, deve ser considerado omelhor, pois, além de apresentar uma excelenteestimativa da CEC, ambos os seus progenitorespossuíram elevada estimativa da CGc.
BIIBLlOGRAFIA
ALLARD, R. W. Princípios do MelhoramentoGenético das Plantas. Trad. AlmiroBlumenschein, Ernesto Paterniani, José T. doAmaral Gurgel e Roland Vencovsky. AgênciªNorte-Americana para º Desenvolyime!!t0Internacional-USAID ~ Edgard Blücher Ltda. SãoPaulo, 1971. 381p.BELLACHEW, B. Arabica coffee breeding inEthiopia: a review. In: Colloque ScientifiqueInternational sur le Café, 17º, Nairobi, 1997.Resumos... , Paris, Association ScientifiqueInternacionale du Café (ASIC),1997. p.415-423.(CAB Abstracts)BELLACHEW, B.; AMEHA, M.; MEKONNEN,D. Heterosis and combining ability in coffeeiCoffea arabica L.). In: Colloque ScientifiqueInternational surle Cafe, 15º, Montpellier, 1993Resumos..., , Paris, Association ScientifiqueInternacionale du café (ASIC), 1993. p.A9.BERTRAND, B.; AGUILAR, G.; SANTACREO,R.; ANTHONY, F.; ETIENNE, H.; ESKES, A.B.; CHARRIER, A. Comportement d'hybrides FIde Coffea arabica pour la vigueur, la productionet la fertilité en Amérique Centrale. In: Col1oqueScientifique International sur le Café, 17°,Nairobi, 1997. Trabalhos publicados..., Paris,Association Scientifique Internacionale du Café(ASIC), 1997 p.415-423.CAVALCANTI, J. J. V.; PINTO, C. A. B. P.;CRISÓSTOMO, J. R.; FERREI RA, D. F.Capacidade de combinação do cajueiro(Anacardium occidentale L.) em relação àprodução precoce de castanha. Revista Ceres,44(254):466-472,1997.
260
CILAS, c.; BOUHARMONT, P.; BOCCARA,M.; ESKES, A. B.; BARADAT, Ph. Prediction ofgenetic value for coffee production in Coffeaarabica from a half-diallel with !ines and hybrids.Euphytica I04( I ):49-59, 1998.FAZUOLl, L. C. & CARVALHO, A. Capacidadede combinação de híbridos de Coffea canephora..In: CONGRESSO BRASILEIRO DEPESQUISAS CAFEEIRAS, 14, Campinas, 1987.Resumos..., Campinas, IBC, 1987. p.93-95.GAL VÊAS, P. A. O. Características Agronômicasde Sete Cultivares de Pimentão (Capsicum annumL.) e Heterose dos Seus Híbridos F). Viçosa,UFV, 1988. 83p. (Tese M.S.)MARTINEZ, O.; TORREGROZA. M.;RONCALLO, E. J. Heterosis en cruzamientosvarietales de maiz de amplia diversidad genética ygeográfica. Agronomía Colombiana, 5 (1-2):4852, 1988.MELO, P. C. T. de. Heterose e CapacidadeCombinatória em um Cruzamento DialélicoParcial Entre Seis Cultivares de Tomate(Lycopersicon esculentum Mill.), Piracicaba,ESALQ, 1987. 108p. (Tese D. S.)MILLER, P. A. & MARANI, A. Heterosis andcombining ability in dial1el crosses of uplandcotton, Gossypium hirsutum L. Crop Science3(5):441-444,1963.PATERNIANI, E. Estudos recentes sobreheterose - Projeto Nacional de Feijão. FundaçãoCargill, 1975. 36p. (Boletim Técnico)SARAWAT, P.; STODDARD, F. L.;MARSHAL, D. R.; ALI, S. M. Heterosis for yieldand related characters in pea. Euphytica 80:39-48,1994.SRINIVASAN, C. S. & VISHVESHWARA. S.Heterosis and stability for yield in Arabica coffee.Indian Journal of Genetics & Piam Breeding38(3):416-420,1978.VAN DER VOSSEN, H. A. M. Coffee Selectionand Breeding. In: M. N. Clifford & K. C. Wilson(Eds) Coffee, Botany. Biochemistry andProduction of Beans and Beverage. Croom Helm,London & Sydney, 1985. p48-96.

261
RESISTÊNCIA DE CLONES DE C. canephora VAR. CONILLON A Hemileia vastatrix BERK. & BR.
Dalza G. da Sílva'", Laércio Zamboliml,J, Ney S. Sakiyama'", Antônio A. Pereira", Aymbiré F. A.Fenseea'", Terezinha A. Teixeira'", Cássia C. H. Sakiyama',
IBIOAGRO/UFV; 2DFT/UFV; 3DFP/UFV; 4EPAMIG; sEMCAPER.36571-000 Viçosa, MG
Apoio Financeiro: FAPEMIG. FINEP. CNPq. PNP & DCafé
INTRODUÇÃO
No Brasil, a variedade Conillon de Coffeacanephora é a mais cultivada, sendo o estado doEspírito Santo responsável por 80% da produçãonacional (Matiello, 1998 e Costa et ai., 1995).Porém, a produtividade das lavouras capixabasformadas por mudas oriundas de sementes ébaixa, aproximadamente 7 se. benef.lha (Bragançaet al., 1993).A espécie Coffea canephora é diplóide (2n = 22cromossomos), alógama e auto-incompatívelcontribuindo para a formação de lavourasheterogêneas em relação a resistência à ferrugem,época de colheita, uniformidade de maturação dosfrutos, arquitetura das plantas e produtividade,resultando em um produto de baixa qualidade(Bragança et aI., 1993).Com a finalidade de solucionar esses problemas.a Empresa Capixaba de Pesquisa Agropecuárialançou em 1993 as variedades clonais 'EMCAPA8111', 'EMCAPA 8121' e 'EMCAPA 8131', deciclo de maturação precoce, médio e tardio,constituídas de 10, 15 e 14 clonesrespectivamente, obtidos através da seleção emultiplicação vegetativa por estaquia, de plantasmatrizes selecionadas em lavouras comerciais emvários municípios produtores do Norte do Estado.O presente trabalho teve como objetivo, avaliar aresistência desses clones a quatro raçasfisiológicas de Hemileia vastatrix Berk. e Br.
MATERIAL E MÉTODOS
No presente trabalho foram avaliados 33 clonesque compõem as variedades clonais 'EMCAPA8111', 'EMCAPA 8121' e 'EMCAPA 8131', deciclo de maturação precoce, médio e tardio,respectivamente (Quadro 1), obtidos por meio deseleção de plantas matrizes com características dealta produção, resistência a patógenos euniformidade de maturação.
Quadro 1- Composição das variedades clonais deC canephora uti lizados no estudo da resistência aH. vastatrix.
I 'EMCAPA 'EMCAPA 'EMCAPA I8111' 8121' 8131'
02 07 3103 I1 4526 14 4629 16 4936 19 99
104-A 109-A 100104-A IIO-A 106IIO-B 112 116
154 120 139128 143132 153149201
Fonte. Adaptado de Bragança et aI., 1993
Quando as mudas apresentavam 3 a 4 pares defolhas, foram transplantadas para vasos de 3 I decapacidade. contendo substrato na proporção de700 , de terra de camada sub-superficial, 250 I deesterco de curral e 1,5 Kg de superfosfato simples.tratado com brometo de metila. Foram utilizadasas raças fisiológicas I. 11, 111 e XIII de H.vastatrix, pertencente ao banco de raças deDepartamento de Fitopatologia da UFV, mantidasem plantas da variedade Catuaí Vermelho (UFV2144) e em clones diferenciadores. Osurediniosporos coletados foram submetidos aoteste de germinação em ágar 2%. imediatamenteantes de proceder à inoculação nas plantas a seremavaliadas.O delineamento experimental utilizado foi ointeiramente casualizado com 33 tratamentos e 4repetições, utilizando a linhagem UFV-2144como testemunha. A face abaxial do par de folhasjovens e bem desenvolvidas foi inoculado comaproximadamente I mg de urediniosporos,distribuídos com auxílio de pincel 'Tigre' nº 6. Asplantas inoculadas foram mantidas em câmaras denevoeiro a 25°C. 100% de umidade relativa e

ausência de luminosidade, por um período de 72h. e após este período, transferidas para casa-devegetação.A avaliação das interações patógeno-hospedeirofoi realizada semanalmente, a partir do 20º diaapós a inoculação, até 60 dias, empregando aescala de tipos de reações adaptada por Tamayo(1988) das escalas de D'Oliveira (1954), Eskes(1983) e Abreu (1988), onde os valores de 1 a 3 =
reação de resistência e de 4 a 6 = reação desuscetibilidade. Os resultados foram analisadosbaseando-se na estimativa do intervalo deconfiança (IC), do grau médio de doença de cadaclone, na última data de avaliação (60 dias após ainoculação). empregando a fórmula: X
(- S - sJP X-IX""];<J.l<X+IX""]; =I-a
onde.
x = grau médio da doença no clones = desvio-padrão dos graus de doença no clonen = número de plantas aval iadas
t = ta (n - 1)
O nível de significância foi de a = 5%,permitindo inferir que existe uma probabilidadede 95% do IC conter a verdadeira média do graude doença à respectiva raça.A avaliação da variabilidade genética inter eintraclones foi realizada por meio de marcadoresRAPO ("Randomm Amplified PolymorphicONA").de acordo com Will iams et aI. 1990, comalgumas modificações.
RESULTADOS E DISCUSSÃO
Considerando o grau médio de doença, osresultados mostraram que dos nove clones quecompõem a variedade EMCAPA 8111, 100%,50,50%. 22.22% e 10% foram suscetíveis às raças11I. 11, X11I e I, respectivamente. Com relação aos13 clones que compõem a variedade EMCAPA8121,76,90% foram suscetíveis às raças [I e 111;61.53% à raça X11I e 30,77% à raça 1. Dos 11clones que compõem a variedade EMCAPA 8113.encontrou-se que, 81,81 %, 63,63%, 54,55% e10% dos clones foram suscetíveis às raças 111. 11, Ie X11I, respectivamente. Os resultados indicaramque as variedades clonais são compostas porclones resistentes e suscetíveis às raças de Hvastatrix inoculadas. Observou-se variabilidadegenética intraclones quando inoculados com asrespectivas raças fisiológicas de H vastarix,mostrado pela diferenciação nas respostas àinfecção e pelo desvio padrão das notas da escalade avaliação. Esses resultados foram observadosnos clones 02, llO-B e 139, quando inoculados
262
com a raça fisiológica I; clones I04-A. 14 e 153,quando inoculados com a raça 11; clones 03, 29 e104-B quando inoculados com a raça 111; clones14, 19, 31, 49, I lO-A, 116 e 120 quandoinoculados com a raça XIII. Marcadores RAPOutilizados na avaliação de diversidade de clonesdemonstraram ser eficazes no agrupamento deplantas dentro de cada clone. Verificou-setambém, a exemplo dos resultados cominoculação, que alguns indivíduos mostraram-segeneticamente diferentes das demais plantaspertencentes a um mesmo clone, como observadosnos clones 02. 03, 104-A, 139 e 36. Conclui-seque a resistência à ferrugem do cafeeiro em clonesda variedade Conillon deve ser estudada maisdetalhadamente, com vistas à obtenção de clonescom maior nível e durabilidade de resistência a Hvastatrix.
REFERÊNCIAS BIBLIOGRÁFICAS
Abreu, M. S. (1988) Resistência horizontal aHemileia vastatrix Berk. & Br. em cafeeirosdescendentes do Híbrido de Timor. Tese OS,Universidade Federal de Viçosa, Viçosa, 68 p.
Bragança, S. M.; Carvalho, C.H.S.; Fonseca,A.F.A; Ferrão, R.G. & Silveira, J. S. M. (1993)'EMCAPA 8111', EMCAPA 8121'. EMCAPA8131 ': Primeiras variedades clonais de caféconilon lançadas para o Espírito Santo.Comunicado Técnico, nº 68 junho/93 p. 1/2EMCAPA.ISSN 0101 -7683.
Costa. E.B. da (1995) Manual técnico para acultura do café no Estado do Espírito Santo.Vitória, ES: SEAG-ES,. 163 p.i1.
D'Oliveira, B. (1954) As ferrugens do cafeeiro.Oeiras, Junta de Investigações do Ultramar,CIFC, s.d .. 61 p. Separata da Revista do CaféPortuguês. 1: 5-12.
Eskes, A. B. (1983) Incomplete resistance tocoffee leaf rust, Ooctoral thesis, AgriculturalUniversity of Wageningen, The Netherlands,140 p.
Tarnayo, P. 1. (1988) Resistência de progênies decatimor a oito raças de Hemileia vastatrix Berk.& Br., Tese MS, Universidade Federal deViçosa, Viçosa. 64 p.
Zambolim, L.; Vale, F. X. R.; Pereira. A.A. &Chaves, G. M. (1997) CAFÉ. In. Controle dedoenças de plantas. Ed. Vale & Zambolim p. 83179. v.l, 554p.
Williams, 1.G.K.. Kubelik, A.R., Livak, KJ.,Rafaslsk, 1. A., Tingey, S.V. (1990) ONApolymorphisms amplified by arbitrary primersare useful as genetic markers. Nucleic AcidsRes. 18:6531-653 5

Bayer ffiProteção das Plantas


265
EL CONTROL BIOLÓGICO DE LA BROCA DEL CAFÉ, Hypothenemus hampet, EN COLOMBIA.
Alex E. Bustillo P.
EI café es el cultivo de exportación másimportante de CoJombia. Su importancia aumentasi consideramos el aspecto socioeconómico yaque mantiene muchas familias en el campo ygenera millones de empleos directos e indirectos.La plaga más importante de este cultivo es labroca dei café, Hypothenemus hampei (Ferrari).Este insecto produce caída y pérdida en eJ peso delos frutos cosechados, pero más importante es lapérdida que causa en la calidad de la bebida. Ladependencia unilateral en eJ uso deI controlquímico para el combate de este insecto no esaconsejable, por eso se sugiere la implementaciónde un programa de manejo integradofundamentado en el control biológico, para lograruna reducción en los costos de producción,agregando una base ecológica y mejorando así lacalidad deI café producido.La broca dei café es una de las plagas másimportantes dei café en el mundo (Le Pelley,1968). AI continente americano fue introducidadesde Africa a través dei Brasil en 1923 y seencuentra distribuida en casi todos los paísesproductores de café. Hasta el presente no se tieneinformación de su presencia en Costa Rica yPanamá. Las hembras de la broca son las queinician el ataque ai perforar los frutos en la regióndei disco de la corona dei fruto, Ilegando de estaforma aI endospermo formando una cámara dondedepositan los huevos, de los cuales emergen laslarvas que destruyen gran parte de la semilia, poresto causan pérdidas en su peso. EI ataque a frutoscon menos de tres meses de edad se traduce en lacaída de estos. Las perforaciones a los frutosabren una puerta de entrada a agentes patogénicosque demeritan la calidad dei café.EI insecticida más utilizado en el control de brocaha sido el endosulfan, pero el recientedescubrimiento dei desarrol1o de resistencia de labroca a este producto (Brun et al., 1989), laamenaza ai equilibrio biológico y los riesgostoxicológicos en la zona cafetera, han Ilevado amuchos países como México, Guatemala, ElSalvador, Nicaragua, Honduras y Colombia aintroducir parasitoides de Africa para desarrollarprogramas de control biológico.La broca lIegó a Colombia en 1988 y desde esaépoca la Federación Nacional de Cafeteros inicióun programa de investigaciones en controlbiológico con la introducción de parasitoides, eldesarrollo de un micopesticida a base deBeauveria bassiana y la búsqueda de alternativasde insecticidas de menor categoría toxicológica.
USO DE PARASITOIDES
Cuando la broca se introdujo de Africa a América,no vino con sus enemigos naturales quenormalmente ocurren en su lugar de origen. Losparasitoides de la broca hasta el presenteencontrados son originarios de Africa Central,lugar que también es considerado el sitio deorigen de la broca. Estos parasitoides son Proropsnasuta Waterston (Bethylidae), Heterospiluscoffeicola Schmeideknecht (Braconidae) ambosencontrados por Heargraves en 1923 y 1924respectivamente, Cephaionomia stephanoderisBetrem (Bethylidae) fue encontrado en 1960 porTicheler (1963), Y Phymastichus coffea La SaIle(Eulophidae) endoparasitoide encontrado en Togopor Borbón (1989). Entre estos parasitoides losbetilidos C. stephanoderis y P. nasuta son losmás aconsejables para ser incluídos en programasde manejo de la broca, básicamente por ser losque se pueden criar en condiciones de laboratorio.En Colombia la broca se cría masivamente enlaboratorios usando café pergamino con uncontenido de humedad deI 45%. Los granosinfestados se mantienen en bandejas metálicas a80% de HR y 27EC. A los pocos días los granosinfestados muestran actividad alimenticia y aJcabo de unos 20 días presentan suficientes estadosinmaduros para la producción deI parasitoide. Eldesarrollo de C. stephanoderis toma entre 18-21dias a 25EC en un cuarto oscuro. La emergenciade los parasitoides se logra proporcionandotemperaturas más altas y luminosidad. Durante1993 fue posible producir en CoJombia, cerca detres millones de parasitoides y en 1994 laeficiencia se incrementó y se logró producir cercade seis millones de parasitoides por mes en tresunidades de producción. Hasta el momento(septiembre de 1998) se han liberado en diferentesregiones dei país más de 1400 millones deparasitoides en fincas infestadas con broca y enlas que no se hace uso de insecticidas, con el finde asegurar su establecimiento.C stephanoderis se ha establecido en todos loslugares de Colombia donde se ha liberado,después de 5 anos de liberados en cafetales enNarifío el parasitoide aún permanece activo enesta zona (Benavides et ai. 1994, Quintero et ai.1998). Desde 1998 se inició un programa paraliberación masiva de P. nasuta y a partir de 1999el programa se extenderá con P. coffea.

USO DE ENTOMOPATÓGENOS.
EI hongo entomopatógeno Beauveria bassiana(Bb) se encuentra naturalmente infectando labroca en casi todas las regiones de Colombia
. donde la broca hace su aparición. Hasta elmomento se poseen 102 aislamientos de Bb dediferentes países y colectados localmente, de loscuales aproximadamente la mitad han mostradoactividad contra broca.Se estudió y desarrolló una técnica de bioensayopara seleccionar los aislamientos más patogénicos(González et aI., 1993). El ciclo de vida de Bbsobre la broca bajo condiciones de laboratorio, secompleta en promedio en 8,2 días desde lainoculación dei insecto con el hongo hasta eldesprendimiento de las esporas. Estos resultadospueden variar de acuerdo con el aislamiento quese use y las condiciones de temperatura deilaboratorio. Se ha de mostrado también laimportancia de pasar el hongo Bb a través deinsectos para reactivar su patogenicidad. Cuandose cultiva el hongo en medios artificiales por treso más generaciones su patogenicidad se reduceconsiderablemente, y el tiempo promedio paracausar mortalidad en la mitad de la población seincrementa, en comparación con el hongoactivado sobre broca (González et aI., 1993).Los estudios con Metarhizium anisopliae (Bernalet aI., 1994) han mostrado que este hongo puedejugar un papel importante en el control de la brocaen el suelo, ai infectar la poblacion de brocas queemergen de los frutos brocados que han caídopreviamente ai suelo.
PRODUCCION DE B. bassiana
Dos enfoques se han investigado para laproducción de Bb, a nivel industrial y a nivelartesanal. A nivel industrial (Morales et aI., 1991),la tecnología se ha transferido a productoresparticulares para que se encarguen de laproducción dei hongo. En la actualidad existencuatro compaüías con licencia dei ICA, quesuministran hongo formulado para el control de labroca. Durante 1992 se utilizaron cinco toneladasde hongo a una concentración de 1 x 108
esporas/gramo con fines experimentales. Laproducción de Bb para el control de la broca deicafé fue de 60 toneladas en 1993 (Posada, 1993) ypara 1998 se estimó en 300 toneladas de un
266
producto conteniendo aI menos Ix I09 esporas /gramo de producto.También se estudió una metodología paraproducir el hongo a nivel de caficultor en su finca(Antía et aI., 1992). La producción promedia deesporas en estas botellas es de 5 x 1010
esporas/l 00 g de sustrato a 25EC, después de untiempo de desarrollo de 24 días. Una vez que elhongo completa su desarrollo está listo para serusado por el agricultor. La producción de unabotei la es suficiente para asperjar 100 árboles auna dosis de 5 x 108 esporas/árbol. Cenicafé hapuesto a disposición dei cafetero una Unidad deProducción Masiva dei hongo B. bassiana, parasuministrar un inoculo puro para su reproducción.El hongo producido en esta unidas se le hace uncontrol de calidad para asegurar: a) que esté librede contaminantes; b) tenga la concentraciónapropiada; c) viabilidad dei 100%; d)patogenicidad sobre broca en laboratorio superiorai 80% y e) uso de una cepa dei hongo reciénactivada y de mejor comportamiento en el campo.Las forrnulaciones de B. bassiana se hanevaluado bajo condiciones de campo y en todoslos casos el hongo se ha establecido en laspoblaciones de broca. Bb sólo es efectivo cuandola broca entra en contacto con las esporas, aI tratarde penetrar la cereza. Si el insecto ya entró a lacereza es dificil que el hongo lo pueda afectar.La epizootiología de Bb se estudió en una fincainfestada con broca en Ansermanuevo (Valle delCauca) a 1000 msnm en un cafetal de variedadColombia plantado a una densidad de 10.000árboles por hectárea (Bustillo et aI., 1991). Lainfestación de la broca se inició en la parte centralde la parcela en junio de 1990 y en esa época seasperjó con una formulación en polvo de Bb en ladosis de 1 x 108 esporas/árbol. Después de seismeses la broca estaba distribuida en todo elcampo así como el hongo. Se evaluómensualmente la infestación de broca e infecciónpor el hongo sobre 300 ramas tomadasaleatoriamente entre enero y agosto de 1991. Launidad de muestreo fue una rama productiva sobrela cual se contaron todas las cerezas, las cerezasinfestadas y aquellas con infección deI hongo. Losresultados indican que una alta proporción de labroca (> 75%) puede ser infectada por el hongo amedida que el tiempo transcurre causando unareducción en la población de la plaga.

267
Lista de enemigos encontrados atacando o compitiendo con la broca en Colombia
ENEMIGOS NATIVOS ESTADO DE LA ENEMIGOS NATIVOS ESTADO DE LA BROCABROCA AFECTADO AFECTADO
PREDADORESENTOMOPATOGENOS FormicidaeHONGOS Crematogaster sp. Todos
I Hyphomicetes Pheidole sp. TodosBeauveria bassiana larva.pupa.adulto Brachymyrmex sp. TodosBeauveria brongniartii adulto (1ab.) Solenopsis sp. TodosHirsutella eleutheratorum adulto Wasmannia sp. TodosMetarhizium anisopliae adulto ?Prenolepsis sp. Todos
'7iiiecilomyces lilacinus adulto AnthocoridaeCalliodis sp. inrnaduros
-?Scoloposcelis sp inrnadurosBACTERIAS
Bacillaceae
Bacillus sp. larva COMPETIDORESEnterobacteriaceae COMPETIDORESCOMPETENCIA
Serratla sp. larvaINTERESPECIFICA(Por espacio y/o alimento)
PROTOZOARIOS INSECTOSMicrosporidia Hormigas en frutos dei árbol y suelo
cercano a Mattesia adulto Dipteros en frutos dei árbo
Coleópterps en frutos dei árbo
PARASITOIDESBraconidae HONGOS en frutos deI árbo
Cercano a Crytoxilos sp adulto BACTERIAS en frutos dei árbo
NEMATODOS en frutos dei árbol y suelo
ENEMIGOS NATIVOS DE LA BROCA
En los predios en los cuales se ha hecho un usoracional o no se han asperjado insecticidas se hadesarrollado una fauna benéfica que está obrandosobre las poblaciones de broca. En diferenteszonas cafeteras de Colornbia, se ha observado lapresencia de varios enemigos nativos de la broca,especialmente en sitios en los cuales la broca llevamás de dos anos y no se han utilizado insecticidas.Los reconocimientos de la disciplina deEntomología de Cenicafé llevados a cabo endiferentes zonas cafeteras dei país han permitidoelaborar la siguiente lista no sólo de enemigos dela broca sino de otros organismos que compitenpor su nicho.EI anterior listado de factores de mortalidad de labroca, asociado con todos los organismos queestán compitiendo por ocupar su sitio,especialmente en los frutos brocados, sonresponsables de que sus poblaciones se reduzcancon el tiempo, lo cual conduce a disminuir loscostos de control. Las prácticas de control que seusen deben mantener el equilibrio biológico o seala protección de la fauna benéfica. Las épocasII uviosas proporcionan humedad ai suelo,fomentando así la invasión de microorganismos
competidores dentro de los frutos brocados quecaen aI sue lo.EI uso de insecticidas es contra indicado en estascondiciones porque no sólo afectan loscontroladores naturales de la broca, sino queeliminan la fauna benéfica de otros insectos comoel minador, escamas, cochinillas, palomillas ydefoliadores que no se convierten en plagasgracias ai control que ejercen sus enemigosnativos.
MANEJO INTEGRADO DE LA BROCA
Debido a que la broca dei café es una plagaintroducida sin enemigos naturales presentes enColombia, la primera estrategia basada en losresultados de investigación presentados aquí, fueintroducir tanto parasitoides comoentomopatógenos en el ecosistema cafetero que hasido invadido por la broca (Bustillo, 1993) ypreservar y fomentar la fauna benéfica nativa.Luego se estructuró un programa de manejo
integrado que incluye conocimiento en la fincasobre períodos de floraciones, evaluación deinfestaciones, prácticas de cosecha sanitaria ("elRE-RE"), controles en la etapa de beneficio deicafé y los componentes biológicos baseados en

parasitoides como C. stephanoderis, yentomopatógenos como B. bassiana (Bustillo,1990). EI uso de insecticidas se restringe aaquellos casos en que los niveles de infestación loameriten, especialmente en sitios donde la brocase encuentra agregada, para lo cual serecomiendan productos de baja toxicidad(categorías 111 y IV) Y poco impacto ambiental.Las parcelas de manejo integrado que Cenicafémonitorea en fincas de cafeteros han mostrado laviabilidad de las recomendaciones para que elcafetero pueda seguir produciendo café tipo"Federación" en presencia de la broca.
CONCLUSIONES
EI enfoque dei manejo integrado de plagas es lamejor estrategia para el control de la broca delcafé, procurando que el control biológico sea unode los componentes principales. En Colombia sehan introducido y desarrollado agentes benéficospara ser utilizados en el control de esta plaga. EIuso de los parasitoides C. stephanoderis y P.nasuta y deI entomopatógeno B. bassiana, asícomo eI fomento de la fauna nativa, en unprograma de manejo integrado de la broca. esfactible como lo demuestran las investigacionesrealizadas en Colombia.
LITERATURA CITADA
ANTI A, O. P.; POSADA, F. J.; BUSTlLLO, A.E.; GONZÁLEZ, M.T. 1992. Producción en fincadeI hongo Beauveria bassiana para el control dela broca dei café. Cenicafé, Avances Técnicos No.182,8pp.BENAVIDES, P.; BUSTlLLO, A. E.;MONTOYA, E. C 1994. Avances sobre el usodei parasitoide Cephalonomia stephanoderis parael control de la broca dei café, Hypothenemusampei. Rev. Colombiana Ent., 20 (4):247-253.BERNAL, M. G. ; BUSTILLO, A. E.; POSADA,F. J. 1994. Virulencia de aislamientos deMetarhizium anisopliae y su eficacia en camposobre Hypothenemus hampei. Rev. ColombianaEnt. 20 (4):229-233.BORBON, O. 1989. Bioecologie d' un ravageurdes bajes de cafeier Hypothenemus hampei(Ferr.) (Coleoptera: Scolytidae) et des sesparasitoides au togo. These du doctorat deL'Universite Paul-Sabatier de Toulouse Cedex,France. 185p.
268
BRUN. L. O.; MARCILLAUD, C;GAUDICHON, V. ; SUCKLlNG, D.M. 1989.Endosulfan resistance in Hypothenemus hampei(Coleoptera: Scolytidae) in New Caledonia. J.Econ. Entomol. 82 (5): 1312-1316.BUSTILLO. A. E. 1990. Perspectivas de manejointegrado de la broca dei café, Hypothenemushampei en Colombia. Socolen. Medellín,Colombia, Miscelánea No.18. p.l 06-118.BUSTILLO. A. E. 1993. El control biológicocomo un componente en un programa de manejointegrado de la broca dei café, Hypothenemushampei, en Colombia. Memorias XX Congreso deSocolen, Cali,julio 13-16,1993, p.159-164.BUSTlLLO, A. E.; CASTILLO, H.; VILLALBA,O.; MORALES E.; VELEZ, P. E. 1991.Evaluaciones de campo con el hongo Beauveriabassiana para el control de la broca dei café,Hypothenemus hampei en Colombia. In:Colloque Scientifique International sur le café. 14.San Francisco. 14-19 juillet, Paris. ASIC 679686p.GONZALEZ. M. T.; POSADA, F. J.BUSTILLO, A. E. 1993. Desarrollo de unbioensayo para evaluar la patogenicidad deBeauveria bassiana sobre Hypothenemushampei. Revista Cenicafé 44(3): 93-102.LE PELLEY, R. H.1968. Pests of coffee.Longmans, Green and Co. Ltd., London.590 p.MORALES, E.; CRUZ, F.; OCAMPO, A.;RIVERA. G. ; MORALES, B. 1991. Unaaplicación de la biotecnología para el control de labroca dei café. In: Colloque ScientifiqueInternational sur le café, 14. San Francisco. 14-19juillet, Paris, ASIC 521-526p.POSADA, F. J. 1993. Control biológico de labroca dei café, Hypothenemus hampei (Ferrari)con hongos. Memorias XX Congreso Socolen,Calijulio 13-16, 1993, p.137-151.QUINTERO, C, BUSTlLLO A. E.,BENAVIDES, P.• CHÁVES, B. 1998. Evidenciasdei establecimiento de Cepha/onomiastephanoderis y Prorops nasuta, en cafetales deidepartamento de Narifio, Colombia. Manuscritoen prensa en Folia Entomológica Mexicana. 15p.TICHELER, J. H. G. 1963. Estudio analítico de laepidemiología dei escolítido de los granos de café.Stephanoderes hampei Ferr., en Costa deMarfil.(Traducción G. Quiceno). Revista Cenicafé14 (4): 223-294.
jI

269
CONTROL BIOLOGICO DE PLAGAS DEL CAFETO: PRODUCCION Y SECADO DEBIOPESTICIDAS EN FERMENTACION EN MEDlO SOLIDO
S. Roussosl , C. Bagnis l, W. Rodriguez2, M-A. Aquiahuatl2, O. Besnard3, R. Duponnoís'l
ILaboratoire de Microbiologie IRD, Université de Provence, CESB/ESIL, Case 925,163 Avenue deLurniny, 13288 Marseille cedex 9, France : Email: [email protected] Depto Biotecnologia; UAM-Iztapalapa; AP 55-535; CP 09340 Mexico DF ; Mexico3 Société Mycos; Pare Scientifique Agropolis 11; 34397 Montpellier Cedex 5; France4 Laboratoire de Biopedologie IRD; BP 1386; Dakar ; Senegal Biological control of coffee pests : largescale production of fungai biopesticide in solid state fermentation
ABSTRACTOver 400 species of fungi can control specific nematodes or insect pests. The concept of using fungaipathogens to control insects is by no means new; the first reports of mass production appeared in thenineteenth century. During the last 20 years, research has been stimulated by the increasing resistance ofinsects to chemical pesticides and by a greater public awareness of environmental impacts of chemicals.Recent success in using nematophagous and entomophagous fungi as biopesticides demonstrate a highpotential for co./fee production and possible commercial applications. The study of the sporulationphysiology, allowed to define optimum media and environmental conditions to produce large quantities ofconidiospores of Aspergillus niger, Beauveria bassiana, Poecelomyces fumosoroseus, and Trichodermaharzianum in bulk. Pilot plant scale production ofconidiospores was conducted in solid state fermentationusing two methods: disk fermentor and natural solid support (sugarcane bagasse) . In disk fermentor,optimal production of conidiospores was obtained after 6 days of incubation for T.harzianum, with anexceptionnally hihg yield of 3.5xIOI O conidiospores per gram of substrate. In a column type reactor,T.harzianum yielded up to I. IxIOIO spores per gram ofsugarcane bagasse. Conidiospore conservation wastested by refrigeration (4°C), freezing (-4°C) and drying (20 and 40°C). Best results were obtained byrefrigeration and drying at 20°C. The kinetics of viability of refrigerated and dried conidiospores ofTharzianum showed the possibility of a reliable utilization of these strains with a 98% of viability, evenafter two months. We discuss the applicability of these methods to the major groups offungi with actual orexpected potential as biopesticide to control insects and nematodes causing economic damage in co./feeproduction. Information about their physiology, spore production at a commercial scale, equipments andformulations is compiled.
RESUMENEI estudio de la fisiología de esporulación, relacionado con la producción cuantitativa de conidiosporas,permitió definir las condiciones medio ambientales y optimizar la composición de los medios de cultivopara la producción masiva de conidiosporas de hongos filamentosos utilizados en lucha biologica(Beauveria bassiana, Trichoderma harzianum). B.bassiana es un hongo entomopatógeno que se utilizacontra los insectos como la broca dei grano de café (Hypothenemus hampei). T'harzianum se utiliza comobiopesticida en el control biológico de hongos fitopatógenos (Botrytis cinerea, Rhizoctonia solani,Fusarium solani). Se estudió la producción de esporas a nivel laboratorio, a un nivel semi piloto en unesporulador de discos y a un nivel planta piloto en un fermentador estático tipo zymotis. Los rendimientosde esporulación de Lharzianum en Zymotis fueron de 5xl013 esporas, cantidad que nos permite tratar aimenos una hectarea de cultivo. La conservación de las esporas se realizó por secado directo dei productofermentado en el bioreactor. Los mejores resultados se obtuvieron, introduciendo unflujo de Gire seco de 3litros por minuto durante 24 horas a una temperatura de 20°C. En estas condiciones se obtuvo unbiopesticida con 5xl09 esporas por gramo de peso seco, que conservan una viabilidad de 98% despues devarias semanas. La misma técnica de producción y de secado de esporas en Fermentación en Medio Sólido(FMS) podría ser utilizada para otros hongos filamentosos entomopatógenos.
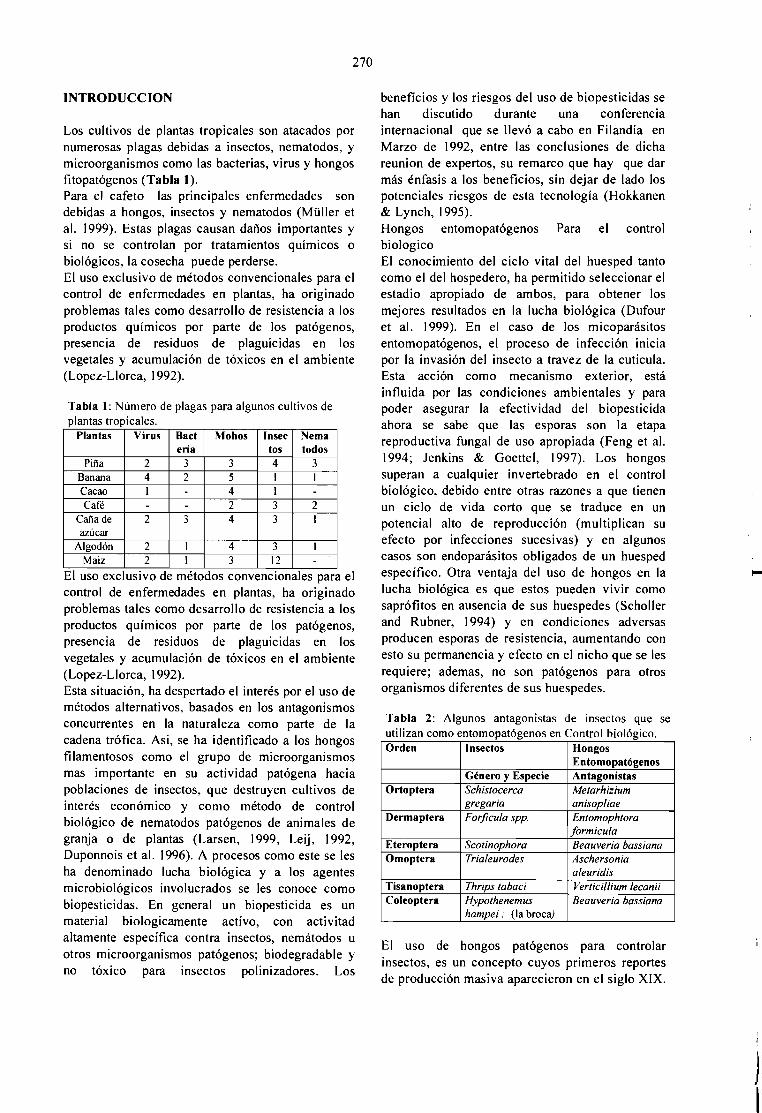
270
INTRODUCCION
Los cultivos de plantas tropicales son atacados pornumerosas plagas debidas a insectos, nematodos, ymicroorganismos como las bacterias, virus y hongosfitopatógenos (Tabla 1).Para el cafeto las principales enfermedades sondebidas a hongos, insectos y nematodos (Müller etaI. 1999). Estas plagas causan danos importantes ysi no se controlan por tratamientos químicos obiológicos, la cosecha puede perderse.EI uso exclusivo de métodos convencionales para elcontrol de enfermedades en plantas, ha originadoproblemas tales como desarrollo de resistencia a losproductos químicos por parte de los patógenos,presencia de residuos de plaguicidas en losvegetales y acumulación de tóxicos en el ambiente(Lopez-Llorca, 1992).
Tabla 1: Número de plagas para algunos cultivos deplantas tropicales.
Plantas Virus Bact Mohos Insec Nemaeria tos todos
Piüa 2 3 3 4 3Banana 4 2 5 I ICacao I - 4 I -Café - - 2 3 2
Caãa de 2 3 4 3 Iazúcar
Algodón 2 1 4 3 1Maiz 2 I 3 12 -
EI uso exclusivo de metodos convencionales para elcontrol de enfermedades en plantas, ha originadoproblemas tales como desarrollo de resistencia a losproductos químicos por parte de los patógenos,presencia de residuos de plaguicidas en losvegetales y acumulación de tóxicos en el ambiente(Lopez-Llorca, 1992).Esta situación, ha despertado el interés por el uso demétodos alternativos, basados en los antagonismosconcurrentes en la naturaleza como parte de lacadena trófica. Asi, se ha identificado a los hongosfilamentosos como el grupo de microorganismosmas importante en su actividad patógena haciapoblaciones de insectos, que destruyen cultivos deinterés económico y como método de controlbiológico de nematodos patógenos de animales degranja o de plantas (Larsen, 1999, Leij, 1992,Duponnois et ai. 1996). A procesos como este se lesha denominado lucha biológica y a los agentesmicrobiológicos involucrados se les conoce comobiopesticidas. En general un biopesticida es unmaterial biologicamente actívo, con activitadaltamente específica contra insectos, nemátodos uotros microorganismos patógenos; biodegradable yno tóxico para insectos polinizadores. Los
beneficios y los riesgos deI uso de biopesticidas sehan discutido durante una conferenciainternacional que se llevó a cabo en Filandia enMarzo de 1992, entre las conclusiones de dichareunion de expertos, su remarco que hay que darmás énfasis a los beneficios, sin dejar de lado lospotenciales riesgos de esta tecnología (Hokkanen& Lynch, 1995).Hongos entomopatógenos Para el controlbiologicoEI conocimiento dei ciclo vital dei huesped tantocomo el deI hospedero, ha permitido seleccionar elestadio apropiado de ambos, para obtener losmejores resultados en la lucha biológica (Dufouret aI. 1999). En el caso de los micoparásitosentomopatógenos, el proceso de infección iniciapor la invasión dei insecto a travez de la cuticula.Esta acción como mecanismo exterior, estáinfluida por las condiciones ambientales y parapoder asegurar la efectividad deI biopesticidaahora se sabe que las esporas son la etapareproductiva fungai de uso apropiada (Feng et aI.1994; Jenkins & Goettel, 1997). Los hongossuperan a cualquier invertebrado en el controlbiológico, debido entre otras razones a que tienenun ciclo de vida corto que se traduce en unpotencial alto de reproducción (multiplican suefecto por infecciones sucesivas) y en algunoscasos son endoparásitos obligados de un huespedespecífico. Otra ventaja deI uso de hongos en lalucha biológica es que estos pueden vivir comosaprófitos en ausencia de sus huespedes (Schollerand Rubner, 1994) y en condiciones adversasproducen esporas de resistencia, aumentando conesto su permanencia y efecto en el nicho que se lesrequiere; adernas, no son patógenos para otrosorganismos diferentes de sus huespedes.
Tabla 2: Algunos antagonistas de insectos que seutilizan como entomopatógenos en Control biológico.Orden Insectos Hongos
EntomopatógenosGénero y Especie Antagonistas
Ortoptera Schistocerca Metarhiziumgregaria anisopliae
Dermaptera Forficula spp. Entomophtoraformicula
Eteroptera Scotinophora Beauveria bassianaOmoptera Trialeurodes Aschersonia
aleuridisTisanoptera Thrips tabaci Verticillium lecaniiColeoptera Hypothenemus Beauveria bassiana
hampei: (la broca)
EI uso de hongos patógenos para controlarinsectos, es un concepto cuyos primeros reportesde producción masiva aparecieron en el siglo XIX.
)l

271
Unos ejemplos de hongos entomopatógenosantagonistas de insectos se reportan en la Tabla 2.Los reportes de los últimos veinte anos sobre el usode hongos entomopatógenos y nematófagos comobiopesticidas ofrece alternativas interesantes de usode estos microorganismos en la agroindústriacafetalera, asi como de su explotación comercial(Busti11os, 1999; Dufour et a!. 1999).Fisiologia de esporulacion de los hongosfilamentososLos hongos filamentosos se reproducen pormultiplicación vegetativa (conidiogénesis)produciendo conidiosporas que se Ilamanconmunemente esporas. Los índices de esporulación(Ie) para un hongo, indican la cantidad de esporasproducidas por gramo de substrato inicialmentepresente en el medio de cultivo (Roussos 1985).Estos indices pueden cambiar según la fuente decarbono y la relación Carbono-Nitrógeno (CtN) delos medios de cultivo. El estudio de la fisiología deesporulación, en relación con la produccióncuantitativa de conidiosporas de 5 cepas de hongosfilamentosos (Tabla 3) permitio definir lascondiciones media ambientales y optimizar lacomposición de los medios de cultivo (Roussos y aI.1989). Se anota que los índices de esporulación sonmuy altos cuando la harina de yuca está utilizadacomo fuente de carbono y alcanzan valores de 7.2 x109 para Beauveria bassiana y IA x 10IO paraTrichoderma harzianum. T. harzianum es un hongofilamentoso antagonístico que se utiliza comoagente de biocontrol contra Pythium ultimum y otrasenfermedades de plantas causadas por hongosfitopatógenos (Jensen & Wolffhechel, 1995).B.bassiana es un hongo entomopatógeno que seutiliza contra la broca de café (Bustillos, 1999;Dufour et aI. 1999).
Tabla 3 : Esporulación de 5 cepas de hongosfilamentosos cultivadas en matraces conteniendo unmedio de cultivo con Papa Dextrosa Agar a 25°C.
Hongos Indlce de Fuente de ClN
Ifilamentosos esporulación carbonoTrichoderma 14.1 x 109 Harina de 14
Iharzianum yucaAspergil1us 5.2 x la' Harina de 24
niger yucaBeauveria 7.2x 109 Harina de 24
I bassiana yucaAspergillus 9.9 x 109 Mclaza 14
terreus
I
Penicillium 7.2 x 109 Melaza
I
24
Iroquefortii
La producción de esporas para uso de laboratorio sehacen en condiciones esteriles utilizando matraces.
Para una producción de esporas en mayorescantitades se utilizan otros tipos de reactores yestratégias de producción de inóculos ya se handesarrollado (Feng et aI. 1994; Jenkins & Goettel1997; Montero et aI. 1989; Roussos et a!. 1992).producción de ESPORAS de hongos en mediaSolidoEI cultivo tradicional de los hongos en mediosólido se practica de manera industrial para laproducción de hongos comestibles (Agaricusbisporus, Pleurotus ostreatus) y para laelaboración de alimentos fermentados como losquesos tipo roquefort o el Ternpe, alimentoobtenido por fermentación de granos cocidos desoya. La Fermentación en Medio Solido (FMS) esun nuevo proceso de cultivo de microorganismossobre substratos naturales (salvado de trigo,bagazo de cana, pulpa de café, harina de yuca,granos de arroz) o sobre soportes sólidospreviamente humidificados con una soluciónnutritiva (Raimbault et aI. 1989; ViniegraGonzalez, 1997).La FMS presenta una multitud de ventajas para laproducción de enzimas, esporas y metabolitos demicroorganismos (Roussos et al. 1997). En elúltimo decenio, no ha habido a nivel mundial undesarrolo de explotación comercial tan evidente dela FMS como el que se ha dado en la investigaciónbásica del mismo proceso. Una de las mayoresrazones para esta situación, es la falta de técnicaseficientes para la obtención de inóculos esporalesactivos, los cuales son requeridos en gran cantidaden los procesos de FMS. La forma, edad yproporción dei inóculo son tambien de importanciacrítica en los sistemas de FMS (Lonsane et al,1992), los cuales requieren una cantidad grande deinóculo (2x 1010 esporas/Kg de substrato seco). Enlos sistemas de FMS, comunmente se prefierecomo inóculo las esporas ai micelio vegetativo,por la facilidad de mezclado de estas, con lossubstratos sólidos humedecidos (Raimbault &Alazard, 1981).
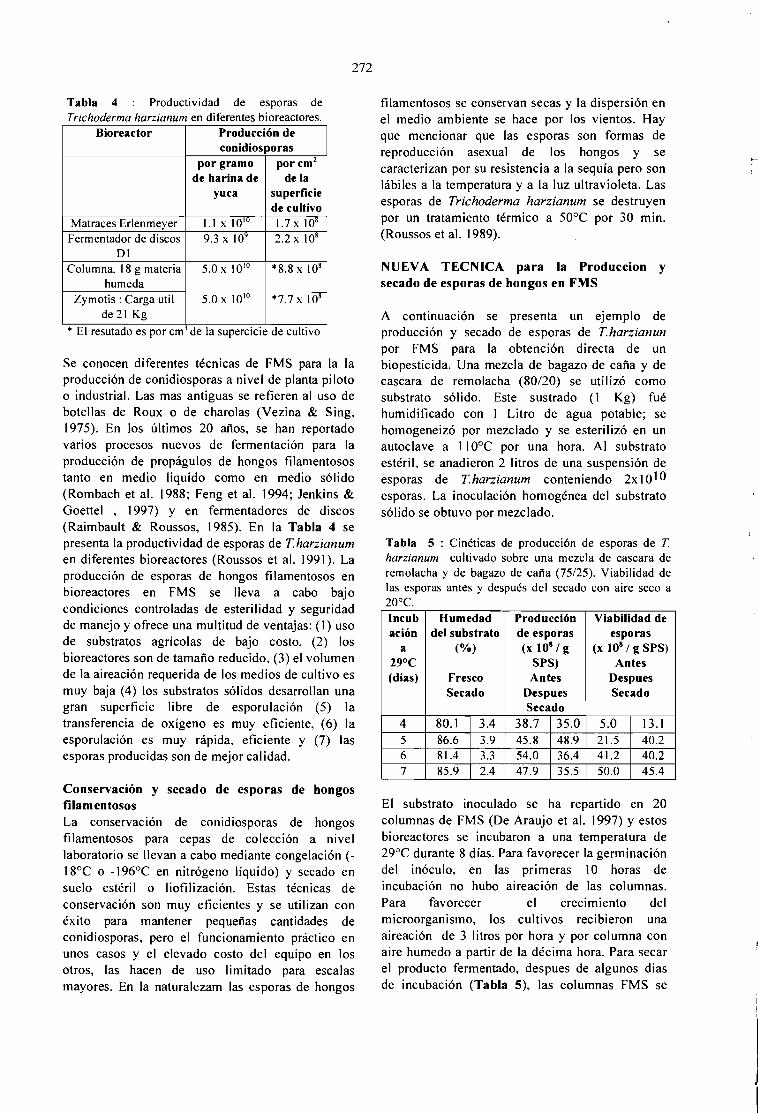
272
Tabla 4 Productividad de esporas deTrichoderma harzianum en diferentes bioreactores.
8ioreactor Producción deconidiosporas
por gramo por em!de harina de de la
yuca superficiede cultivo
Matraces Erlenmeyer 1.1 x 1010 1.7 X 108
Fermentador de discos 9.3 x 109 2.2 x lO"DI
Colurnna, 18 g materia 5.0 x 1010 *8.8 X 108
humedaZymotis : Carga util 5.0 x 1010 *7.7 X 108
de 21 Kg* EI resutado es por crrr'de la supercrcie de cultivo
Se conocen diferentes técnicas de FMS para la laproducción de conidiosporas a nivel de planta pilotoo industrial. Las mas antiguas se refieren aI uso debotellas de Roux o de charolas (Vezina & Sing,1975). En los últimos 20 anos, se han reportadovarios procesos nuevos de fermentación para laproducción de propágulos de hongos filamentosostanto en medio liquido como en medio sólido(Rombach et aI. 1988; Feng et al. 1994; Jenkins &Goettel , 1997) Y en fermentadores de discos(Raimbault & Roussos, 1985). En la Tabla 4 sepresenta la productividad de esporas de Tharzianumen diferentes bioreactores (Roussos et aI. 1991). Laproducción de esporas de hongos filamentosos enbioreactores en FMS se lJeva a cabo bajocondiciones controladas de esterilidad y seguridadde manejo y ofrece una multitud de ventajas: (I) usode substratos agrícolas de bajo costo, (2) losbioreactores son de tamafío reducido, (3) el volumende la aireación requerida de los medios de cultivo esmuy baja (4) los substratos sólidos desarrollan unagran superficie libre de esporulación (5) latransferencia de oxígeno es muy eficiente, (6) laesporulación es muy rápida, eficiente y (7) lasesporas producidas son de mejor calidad.
Conservación y secado de esporas de hongosfilamentososLa conservación de conidiosporas de hongosfilamentosos para cepas de colección a nivellaboratorio se lIevan a cabo mediante congelación (18°e o -196°e en nitrógeno líquido) y secado ensuelo estéril o liofilización. Estas técnicas deconservación son muy eficientes y se utilizan conéxito para mantener pequenas cantidades deconidiosporas, pero el funcionamiento práctico enunos casos y el elevado costo del equipo en losotros, las hacen de uso limitado para escalasmayores. En la naturalezam las esporas de hongos
filamentosos se conservan secas y la dispersión enel medio ambiente se hace por los vientos. Hayque mencionar que las esporas son formas dereproducción asexual de los hongos y secaracterizan por su resistencia a la sequía pero sonlábiles a la temperatura y a la luz ultravioleta. Lasesporas de Trichoderma harzianum se destruyenpor un tratamiento térmico a 500e por 30 rnin,(Roussos et aI. 1989).
NUEVA TECNICA para la Produccion ysecado de esporas de hongos en FMS
A continuación se presenta un ejemplo deproducción y secado de esporas de Tharzianunpor FMS para la obtención directa de unbiopesticida. Una mezcla de bagazo de cana y decascara de remolacha (80/20) se utilizó comosubstrato sólido. Este sustrado (I Kg) fuéhumidificado con I Litro de agua potable; sehomogeneizó por mezclado y se esterilizó en unautoclave a I looe por una hora. AI substratoestéril, se anadieron 2 litros de una suspensión deesporas de Tharzianum conteniendo 2x IOIOesporas. La inoculación homogénea deI substratosólido se obtuvo por mezclado.
Tabla 5: Cinéticas de producción de esporas de T.harzianum cultivado sobre una mezcla de cascara deremolacha y de bagazo de caüa (75/25). Viabilidad delas esporas antes y después dei secado con aire seco a20°C.Incub Humedad Producción Viabilidad deación deI substrato de esporas esporas
a (%) (xlO8/g (x 1081 g SPS)29°C SPS) Antes(dias) Fresco Antes Despues
Secado Despues SecadoSecado
4 80.1 3.4 38.7 35.0 5.0 13.15 86.6 3.9 45.8 48.9 21.5 40.26 81.4 3.3 54.0 36.4 41.2 40.27 85.9 2.4 47.9 35.5 50.0 45.4
EI substrato inoculado se ha repartido en 20columnas de FMS (De Araujo et aI. 1997) y estosbioreactores se incubaron a una temperatura de29°e durante 8 días. Para favorecer la germinacióndei inóculo, en las primeras 10 horas deincubación no hubo aireación de las columnas.Para favorecer el crecimiento deimicroorganismo, los cultivos recibieron unaaireación de 3 litros por hora y por columna conaire humedo a partir de la décima hora. Para secarel producto fermentado, despues de algunos diasde incubación (Tabla 5), las columnas FMS se
r!

273
retiraron de la incubadora y recibieron un f1ujo de 3litros por minuto de aire seco a 20°C durante 24horas. La humedad de producto final fué menor aI4%.Durante todo el tiempo de la incubación de loscultivos en 29°C bajo aireación con aire húrnedo, lahumedad dei producto fermentado se mantuvoarriva deI 80%. Estas condiciones favorecen laesporulación de T.harzianum. Después de 5 dias deincubación el hongo esporuló y la aparición de uncolor verde significa una esporulación abundante yhomogénea. Por conteo ai microscópio, se anotó quela esporulación fué importante y el índice deesporulación alcanzó 4.6x I09 esporas por gramo desubstrato peso seco. Después de 5 días deincubación se cambió eJ flujo, de aire húmedo por elaire seco obtieniendose un producto estabilizadopara conservarlo por un largo plazo (2 meses) sinperdida significativa de viabilidad. Esta mismatécnica se aplicó con diferentes hongosfilementosos (Penicillium, Aspergillus) y condiferentes mezclas de substrato (bagazo de cana,pulpa de café, salvado de trigo) con resultados muysemejantes.
CONCLUSIONES:
Para los hongos filamentosos, las esporas son lasformas de reprodución, de dispersión y desobrevivencia sobre todo en condiciones de bajaactividad de agua. Los hongos filamentosos puedencrecer sobre substratos sólidos y producircantidades elevadas de esporas (109 esporas porgramo de substrato seco). Aprovechando estascaracterísticas de los hongos una técnica deproducción y de secado de esporas en FMS se hadesarrollado para la producción de biopesticidasutilizando diferentes antagonistas. Variosmicroorganismos crecen y esporulan con altosrendimientos sobre un substrato sólido (bagazo decana + cascara de remolacha) en condiciones deFMS. Para obtener una esporulación importante deun hongo antagonista, se debe optimizar tanto unmedio nutritivo como un soporte sol ido que permitatanto el crecimiento y la esporulación como elsecado dei producto final. Esta situación facil itagrandemente la conservación, la viabilidad y eltransporte de los biopesticidas. De tal manera que sepuede centralizar la producción de biopesticidas enun lugar lejano de la aplicación de los mismos,mejorando notablemente el control de la calidad, yla competitividad dei producto.
REFERENCIAS
Bustillos A. 1999. El papel dei control biológicoen un programa de manejo integrado de la brocadei café, Hypothenemus hampei, en Colombia.Proceedings of [H SI BAC , Londrina 24-28 May1999, BrazilDe Araujo, A. A., Lepilleur, C., Delcourt, S.,Colavitti, P. and Roussos S. 1997. Laboratoryscale bioreactors for study fungai physiology andmetabolism in solid state fermentation system. InRoussos, S., Lonsane, B.K., Raimbault, M. andViniegraz-Gonzalez, G. (Eds.), Advances in solidstate fermentation, Kluwer Acad. Publ., Dordrecht,p.93-11I.Dufour B., Barrera J.F., Decazy B. 1999. La Brocade los frutos deI cafeto: La lucha biológica comosolución. In Bertrand B & Rapidel B. (Eds)Desafios de la caficultura en Centroamérica.IJCA.PROMECAFE: CIRAD: IRD:CCCR.FRANCIA, San José, Costa Rica ch.9: 293325Duponnois R., Mateille T., Sene V.!, SawadogoA., Fargette M. 1996. Effect of different WestAfrican species and strains of Arthrobotrysnematophagus fungi on meloidogy.ne species.Entomophaga 41: 475-483.Feng M.G., Poprawski T.J., Khachatourians G.G.1994. Production, formulation and application ofthe entomopathogenic fungus Beauveria bassianafor insect control: current status. BiocontrolScience and Technology 4: 3-34.Hokkanen H.M.T., Lynch J.M. 1995. Biologicalcontrol: Benefits and Risks. Plant and MicrobialBiotechnology Research Series 4, UniversityPress, Carnbridge, 304 p.Jenkins N.E., Goettel M.S. 1997. Methods formass-production of microbial control agents ofgrasshoppers andv locusts. Memoirs of theEntornological Society ofCanada 171: 37-48.Jensen D.F., Wolffhechel H. 1995. The use offungi, particu1ar1y Trichoderma spp. andGliocladium spp., to control root rot and dampingoff diseases. In Hokkanen H.M.T., Lynch J.M.(Eds), Biological control: Benefits and Risks.Plant and Microbial Biotechnology ResearchSeries 4, University Press, Cambridge, p. [77 -189.Larsen, M. 1999. Biological control of helmints.Int. J. Parasitol. 29(1): 139-146.Leij F.D. 1992. The significance of ecology in thedevelopment of Verticillium chlamydosporium as abiological control agent against root-knotnematodes (Meloidogyne spp.) PhD Thesis,Univ.Wageningen, 140 p.

274
Lonsane, B.K., Saucedo-Castaf'ieda, G., Raimbault,M., Roussos, S., Viniegra-Gonzalez, G., Gildyal,N.P., Ramakrishna, M. and Krishnaiah, M.M. 1992.Scale-up strategies for solid state fermentationsystems. Process Biochem. 27: 259-273.Lopez-L1orca L.V. 1992. Los hongos parásitos deinvertebrados y su potencial como agentes decontrol biológico. Revista Iberoamericana deMicología 9: 17-22.Montero, F.A.. Roussos S.. Olmos A. 1989.Producción de biopesticidas a nivel planta pilotopara el uso en cultivo de café. In Roussos, S.,Licona, F.R. and Gutierrez. R.M. (Eds.), I SeminarioInternacional de Biotecnologia en la IndústriaCafetalera .. Memorias 1 SIBAC, Xalapa, Veracruz,p. 199-202.Müller R.. Berry D.. Bieysse D. 1999. Laanthracnosis de los frutos: un grave peligro para lacaficultura Centroamericana. In Bertrand B &Rapidel B. (Eds) Desafios de la caficultura enCentroamérica. IICA.PROMECAFE: CIRAD: IRD:CCCR.FRANCIA. San José. Costa Rica ch.8: 261291.Raimbault, M. and Alazard, D. 1980. Culturemethod to study fungai growth in solidfermentation. Eur. 1. Appl. Microbio!. Biotechnol. 9:199-209.Rairnbault, M. and Roussos, S. 1985. Procédé deproduction de spores de champignons filamenteux.Brevet Français N° 85.08555.Raimbault, M.. Roussos, S.. Oriol, E., Viniegra, G.,Gutierrez. M.. Barrios-Gonzalez, J. 1989. Procédéde culture de microorganismes sur milieu solideconstitué d'un support solide, absorbant,compressible et non fermentable. Brevet FrançaisN° 89. 06558.Rivillas Osorio C.A .. Leguizamon Caycedo 1.E., GilVallejo L.F. 1999. Recomendaciones para el manejode la roya dei cafeto en Colombia. Cenicafé BoletinTécnico 19: 7-36.Rornbach M.C.. Aguda R.M., Roberts D.W. 1988.Production af Beauveria bassiana in different liquid
media and subsequent conidiation of drymycelium. Entomophaga 33: 315-324.Roussos, S. 1985. Croissance de Trichodermaharzianum par fermentation en. milieu solide:Physiologie, sporulation et production decellulases. Thêse d'Etat, Université de Provence,Marseille, 193 p.Roussos, S., Aquiahuatl, M.A., Brizuela, M.A..Olmos, A., Rodriguez, W. and Viniegra, G. 1989.Production, conservation and viability offilamentous fungi inoculum for sol id substratefermentation. Mico!. Neotrop. Apl. 2: 3-17.Roussos, S., Olmos, A.. Raimbault, M.. SaucedoCastafíeda, G. and Lonsane, B.K. 1991. Strategiesfor large scale inoculum development for solidstate fermentation system : Conidiospores ofTrichoderma harzianum. Biotechno!. Tech. 5: 415420Roussos, S., Raimbault. M.. Prebois. J-P. andLonsane, B.K. 1993. Zymotis, A large scale solidstate fermenter : Design and evaluation. AppliedBiochem. Biotechno!. 42: 37-52.Roussos, S., Lonsane, B.K .. Raimbault, M. andViniegra-Gonzalez, G. (Eds.). Advances in solidstate fermentation, Kluwer Acad. Publ., Dordrecht,631 p.Schoeller, M., Rubner, A. 1994. Predaciousactivity of the nematode-destroying fungusArthrobotrys oligospora in dependence of themedi um composition. Microbiol. Res. 149: 145149Vezina, C. and K. Sing, 1975. Transformation oforganic compounds by fungi spores. In: Smith,1.E. and D.R. Berry (Eds.). The filamentous fungi,1. Industrial Mycology. Edward Arnold. Londres.Viniegra-Gonzalez G. 1997. Solid statefermentation: Definition, characteristics,limitations and monitoring. In Roussos, S"Lonsane, B.K.. Raimbault, M. and ViniegraGonzalez, G. (Eds.), Advances in solid statefermentation, Kluwer Acad. Publ., Dordrecht,Ch.2: 5-22.
275
TUE NEMATOPUAGOUS FUNGI HELPER BACTERIA (NUB) : A NEW DIMENSION FOR THEBIOLOGICAL CONTROL OF ROOT KNOT NEMATODES BY TRAPPING FUNGI
R. Duponnois J, J.L. Chotte J, A.M. Bâ 2 and S. Roussos 3
1 IRD. Laboratoire de Biopédologie. BP 1386. Dakar. Sénégal - 2 ISRA/IRD. BP 1386. Dakar. Sénégal -3 IRD. Laboratoire de Mycologie et des FMS (LAMY-FMS). IFR-BAEM; Université de Provence,CESB/ESIL, Case 925; 163 Avenue de Luminy. F-13288, Marseille Cedex 9 FRANCE
SUMMARYThe research work presented in this paper summaries the studies performed in the Biopedology /aboratoryof IRD Dakar on the biological control with nematophagous fungi against the root-knot nematodes. Theresu/ts show that the fungi can act against M incognita and M mayaguensis, the most infective species inWest Africa but a/so frequent/y detected through coffee p/antations. They can a/so contra/ the deve/opmentof Me/oidogyne popu/ations on tomato p/ants. A cultural practice has been developped using compostb/ocks in which the fungus was inocu/ated. In a field experiment, the growth of tomato p/ants has beenincreased while the nematode deve/opment has been decreased. However, all these experiments have beenperformed during a short period. In order to enhance the competitivity of the fungus, the fungus can beassociated with bacterial iso/ates, called NHB (Nematophagous He/per Bacteria), which stimu/ate thefunga/ activity against Me/oidogyne but invo/ve a/so a depressive effect on other p/ant parasitic nematodessuch as Roty/enchu/us reni{ormis.
INTRODUCTION
Many genera and species of plant parasiticnematodes are associated with coffee in the worldbeing therefore responsible for great financiaIlosses to the coffee farmers. Among thesepathogens, root-knot nematodes (Meloidogynespp.) are the most abundant group (Table I) andthe most common species are M. exigua, M.incognita and M. coffeicola. The symptoms ofdamage due to these pathogens are typicalrounded galls on the root systems, white toyellowish brown becoming dark brown in agingroots. The infested coffee plants show foliarchlorosis, leaf fali, general decline. Their growthis generally reduced and sometimes the plantsdied (Hutton et aI., 1982; Lordello, 1984). Thecoffee plantations can be dramatical\y affected bythe nematodes. For example, in Sao Paulo state(Brazil), they have been destroyed by M.
Table 1. Nematophagous fungi used in lhe experiments
incognita with 5-year-old coffee plantations dyingout (Lordel1o, 1984).The control of nematodes is more difficult in aperennial crop than in annual or herbaceouscrops. For example, the rotation schemes,successfully used with annual crops, areimpractible with these long term cultures.Moreover surviving roots of excised plants or oldplants left in the field can provide nutrients fornematodes and consequently maintain thenematodes population in the soil.The control practices used by many farmers isgeneral1y based on (i) the production of seedlingswithout root-knot nematodes (i.e. disinfection ofnursery soils), (ii) the application of nematicidesand/or the culture of resistant or tolerant cultivars.Among these control measures, pesticidecompounds are mostly appl ied to reducenematode multiplication.
Identificadon Code Geographicalorigin AuthorArthrobotrys oligospora S 30 Burkina Faso Sawadogo A. \ I)
Arthrobotrys oligospora S 31 Burkina Faso SawadogoA.Arthrobotrys conoides S 42 Burkina Faso Sawadogo A.
Arthrobotrys sp. BFIO Burkina Faso SawadogoA.Arthrobotrys sp. BF74 Burkina Faso Sawadogo A.Arthrobotrys sp. BF80 Burkina Faso Sawadogo A.Arthrobotrvs sp. SOSU2 Burkina Faso SawadogoA.Arthrobotrvs sp. ORS 18690 S2 Senegal R. Duponnois
Arthrobotrvs oligospora ORS 18692 SS Senegal R. DuponnoisArthrobotrvs oligospora ORS 18692 S7 Senegal R. Duponnois
INERA, Protection des Végétaux, BP 403. Bobo Dioulasso, Burkina Faso

However they are very expensive and very toxicwhen improperly. Thus other control techniquessuch as biological control have been investigated.Besidesvarious microorganisms tested againstMeloidogyne spp. such as arbuscular mycorrhizae(Hussey & Roncadori, 1982), eggs parasitic fungi(Verticillium chlamydosporium)(Kerry, 1990;Bourne et aI., 1994), rhizobacteria (Racke &Sikora, 1986) and fungai endophytes (Schuster etaI., 1995), research focused on nematophagousfungi (Cayrol, 1983; Pelagatti et aI., 1986;Duponnois et aI., 1996). The screening ofefficient fungaI strains against Meloidogyne wasbased on the use of tests performed in axenic orcontrolled conditions O!! vitro tests or glasshouseexperiments with desinfected substrates). Whenthese microorganisms were transferred into thefield, their antagonistic activity was oftenmodulated (generally decreased) by theenvironmental conditions. In fact, the topics ofthis kind of research must integrated therequested qualities of the fungaI strain but also itsrepply from interactions with the soil microbialcommunity in the soil. In the laboratory ofbiopedology (lRD, Dakar), we have developped ascientific program based on (i) the screening ofefficient fungal strains for trapping juveniles ofMeloidogyne, (ii) the determination of potentialeffects of rhizobacteria on i!! vitro growth andnematode-trapping activity of Arthrobotrysoligospora, population deveJopment of plantparasitic nematodes and (iíi) develop a controlmethod with the fungai strain incorporated intocompost blocks. Tobacco and tomato plants, verysusceptible to Meloidogyne, were used in theseexperiments.
MATERIALS & METHOOS
1/ Test of the potential of nematophagous fungiagainst MeloidogyneA collection of nematophagous fungi isolatedfrom several vegetable-producing areas inSenegal and Burkina Faso were maintainedaseptically in the dark at 25°C on the nutrientbroth (8 g.l-I) agar (20 g.l-I) medium (Table I).The trapping activity of each fungai strain wasestimated using technique described byDuponnois et aI. (1996). Populations of differentspecies of Meloidogyne (M. mayaguensis, M.javanica and M. incognita) were reared on tomato(Lycopersicon esculentum Mill.), cv Roma roots.Two months after inoculation, the roots wereharvested, cut into short pieces and placed in amist chamber for one week for egg hatching
276
(Seinhorst, 1950). Fungai agar plugs were takenfrom the margin of two week old colonies andtransferred into Petri dishes filled with distilledwater agar (20 g.l-I). One week later, 100 secondstage juveniles suspended in a water droplet wereplaced on these fungai cultures. After two days,the number of juveniles trapped by the funguswere counted under a dissecting microscope.Each combination Meloidogyne species-fungalisolate was replicated five times. The trappingrate (trapped juveniles / total juveniles) wastransformed by arcsin(sqrt) and treated with a oneway analysis of variance, the mean values beingcompared with the Student's t-test at 0.05probability leveI.The ability of these fungai strains to controlMeloidogyne populations was examined througha glasshouse experiment. Solid fungus inoculawere prepared in 0.5 dm 3 glass tlasks containing0.3 dm3 compost. After autoclaving at 120°C for40 min, the substrate was moistened to fieldcapacity with liquid nutrient broth medium (8 g.I'I),the jars closed with cotton wool and autoclaved asecond time at 120°C for 20 mino After cooling,mycelial plugs, sampling from each fungaicultures on Petri dishes, were put into each glasstlasks. The cultures were incubated for 5 weeks at25°C.The fungai inoculum was mixed to an autoclaved(140°C, 40 min) sandy soil (pH H20 7.1 ; fine silt0.6% ; coarse silt 1.4% ; fine sand 61.6%) at theratel: 100 (v :v). This mixture was divided in 60cm 3 polythene cells (The control treatmentreceived the same quantity of compost butwithout fungus). Then a one week old tomatoseedling was transferred into each cell. After 1week, the tomato plants were inoculated with 5ml suspension of 100 7-day-old second stagejuveniles of M. mayaguensis (water alone for thecontrol). The cells were placed in a glasshouseunder natural climatic conditions (temperaturesfrom 20°C to 35°C; about 15 hours light perday). The treatments were watered daily andarranged in a randomized complete block designwith 14 replicates. One month after the nematodeinoculation, the plants were uprooted and theroots washed. Shoots were dried at 65°C for oneweek and weighed. Galls induced by M.mayaguensis were counted. Then roots were cutinto 1 to 2 em pieces and placed in a mistchamber for 2 weeks to recover hatched juveniles(Seinhorst, 1950). Roots were then oven-driedand weighed. The data were treated with an oneway analysis of variance and the mean valueswere compared with the Student's t test

(P < 0.05). For nematodes, data were previouslytransformed by log (x+ I). 21 Study of thepotential effects of rhizobacteria on theantagonistic activity of Arthrobotrys oligosporaagainst Meloidogyne spp.The basis of this research program have beendescribed in Duponnois & Bâ (1998). The authorshave demonstrated that a large nematophagousfungus population was associated with thepresence of bacteria belonging to the group ofPseudomonas fluorescent. Furtherrnore, the aimsof this research were to test the effect of thesebacteria on the fungai growth (a strain of ~.
oligospora T4 J), on its predatory activíty and toevaluate the bacterial influence on the fungalcontrol of the multiplication of Meloidogynepopulation.Bacterial strains were cultured in Petri dishes on0.3% TSB agar medium at 25°C for two days.The bacterial cultures were suspended in 5 ml ofsterile magnesíum sulfate solution (0.1 M) with abend glass rodo The control treatment wasprepared in the same way from a Petri dishcontaining the same agar medium withoutbactería. One set of experiment was performed bydirect liquid contact between the mycelium andthe bacteria. Fungai plugs sampled as describedabove were dipped in the bacterial suspensions orin the control solution for 1-2 min and transferredinto empty Petri dishes (Duponnois & Garbaye,1990). Another set of expriment was done with noliquid contact. Two-compartment dishes wereused: the fungai plugs were laid on the drybottom of the dish for one compartment while theother was filled by 0.3% TSB agar mediuminoculated or not (control treatment) by thebacterial strains.The gas diffusion from one sídeto the other was permitted because the wallseparating the two compartments did not touchthe lid ofthe dísh.In both experiment types, two dishes, each with 3mycelial plugs, were prepared for each treatment.Then the dishes were incubated at 25°C for fourdays. Observation and numeration were carriedout through the Iid with a stereomicroscope andthe mean radial growth in two perpendiculardirection was calculated. The data werestatistically compared to the control treatmentswithout bacteria with the Student "t" test(P<0.05).The fungai plugs, dipped in the bacterialsuspensions or control treatment as describedabove, were transferred to Petri dishes filled with
distilled water agar (20 g.l_I). There were five
277
replicates per treatment. The Petri dishes wereincubated at 25°C in the dark. Two weeks later,100 7-day-old-second stage J2 of Meloidogynemayaguensis suspended in 100 !lI sterile distilledwater were placed on the fungai cultures.Populations of M. mayaguensis were reared ontomato (Lycopersicon esculentum MiII.), cv.Roma. Two months after inoculation, roots werecut into 2-3 cm pieces and placed in a mistchamber for one week to allow nematode eggs tohatch and the juveniles to be collected (J2)(Seinhorst, 1950). After 48 hours, the number ofthe juveniles trapped by the fungus were counted,The data were statistically treated as describedabove.Tobacco (Nicotiana tabacum L. varo Paraguay xClaro) seedlings were grown in 60 ml polythenepots filled with an autoclaved soil (140°C, 40min) sandy soil (clay : 3.9 %; silt : 2.9%; sand :92.2 %; carbon : 3.7 %; nitrogen : 0.45 %;pH(H20) : 8.3) inoculated with I mg dry weightof fungai biomass and/or 5 ml of each bacterialsuspension (about 10 J2 colony forming unit
(cfuj.ml" I). The fungai strain had been grown inone Iitre glass flask filled with 0.5 litre of 0.3%TSB medi um for 2 weeks at 25°C. The fungaisuspension was then filtered and the myceliumwas collected. The fungai mycelium was threetimes washed in MgS04 0.1 M and finallysuspended in MgS04 0.1 M. The bacterial
isolates were cultured in 3 g.I_1 liquid Difcotryptic soy broth in glass flasks shaken for 8 daysat 25°C. The bacterial suspensions were thencentrifuged (2400 g, 10 min) and the pelletresuspended in MgS04 0.1 M. The control
treatments were performed by injecting in the soi1J ml of MgSO4 0.1 M without either fungus or
bacteria. There were ten replicates per treatment.After 2 month culture, 10 tobacco plants fromeach treatment were transferred in 10 litre potsfilled with the same soil as above but nonautoclaved. The pots were placed in a glasshouseunder natural conditions (temperatures rangedfrom 20°C to 35°C; about 15 hours light per 24 h).After two month culture, the plants were harvestedand the root systems gently washed. The soilfrom each pot was mixed, a 250 g sub-samplewas taken and the nematodes were extracted bythe Seinhorst's (1962) elutriation technique. Theoven-dried weight of shoot (one week at 65°C)was measured. Each root system was then cut into2-3 cm pieces and placed in a mist chamber for.two weeks in order to recover nematodes(Seinhorst, 1950). The nematodes were counted
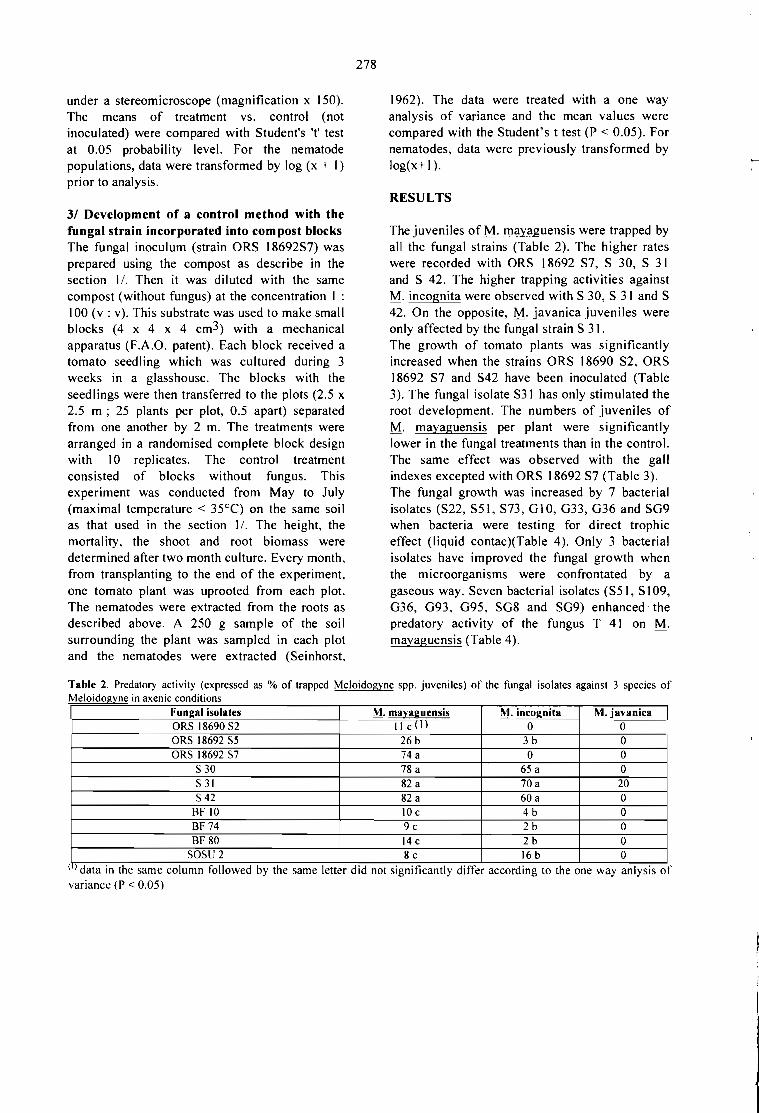
278
under a stereomicroscope (magnification x 150).The means of treatment vs. control (notinoculated) were compared with Student's 't' testat 0.05 probability level, For the nematodepopulations, data were transformed by log (x + I)prior to analysis,
1962). The data were treated with a one wayanalysis of variance and the mean values werecompared with the Student's t test (P < 0.05). Fornematodes, data were previously transformed bylog(x+ I).
RESULTS31 Development of a control method with thefungai strain incorporated into compost blocksThe fungai inoculum (strain ORS 18692S7) wasprepared using the compost as describe in thesection lI. Then it was diluted with the samecompost (without fungus) at the concentration I :100 (v : v). This substrate was used to make smallblocks (4 x 4 x 4 cm3) with a mechanicalapparatus (F.A.O. patent). Each block received atomato seedling which was cultured during 3weeks in a glasshouse. The blocks with theseedl ings were then transferred to the plots (2.5 x2.5 m; 25 plants per plot, 0.5 apart) separatedfrom one another by 2 m. The treatments werearranged in a randomised complete block designwith 10 replicates. The control treatmentconsisted of blocks without fungus. Thisexperiment was conducted from May to July(maximal temperature < 35°C) on the same soilas that used in the section II. The height, themortality, the shoot and root biomass weredetermined after two month culture. Every month,from transplanting to the end of the experiment,one tomato plant was uprooted from each plot.The nematodes were extracted from the roots asdescribed above. A 250 g sarnple of the soilsurrounding the plant was sampled in each plotand the nematodes were extracted (Seinhorst,
The juveniles of M. mayaguensis were trapped byall the fungal strains (Table 2). The higher rateswere recorded with ORS 18692 S7, S 30, S 31and S 42. The higher trapping activities againstM. incognita were observed with S 30, S 31 and S42. On the opposite, M. javanica juveniles wereonly affected by the fungal strain S 31.The growth of tomato plants was significantlyincreased when the strains ORS 18690 S2, ORS18692 S7 and S42 have been inoculated (Table3). The fungai isolate S31 has only stimulated theroot development. The numbers of juveniles ofM. mayaguensis per plant were significantlylower in the fungai treatments than in the control.The same effect was observed with the gallindexes excepted with ORS 18692 S7 (Table 3).The fungai growth was increased by 7 bacterialisolates (S22, S51, S73, GI0, G33, G36 and SG9when bacteria were testing for direct trophiceffect (Iiquid contac)(Table 4). Only 3 bacterialisolates have improved the fungai growth whenthe microorganisms were confrontated by agaseous way. Seven bacterial isolates (S51, SI 09,G36, G93, G95, SG8 and SG9) enhanced thepredatory activity of the fungus T 41 on M.mayaguensis (Table 4).
Table 2. Predatory activity (expressed as % of trapped Meloidogyne spp. juveniles) of lhe fungai isolates against 3 species ofMeloidoavne in axenic conditions
Fungai isolates M. mayaguensis M. incognita M. javanicaOR5 1869052 I1 c \ I ) O OOR5 1869255 26 b 3b OOR5 1869257 74 a O O
530 78 a 65 a O531 82 a 70 a 20542 82 a 60 a O
BF 10 10 c 4b OBF 74 9c 2b OBF 80 14 c 2b O
505U 2 8c 16 b O..
(I) data In the same column followed by the same letter did not significantly differ according to lhe one way anlysis ofvariance (P < 0.05)
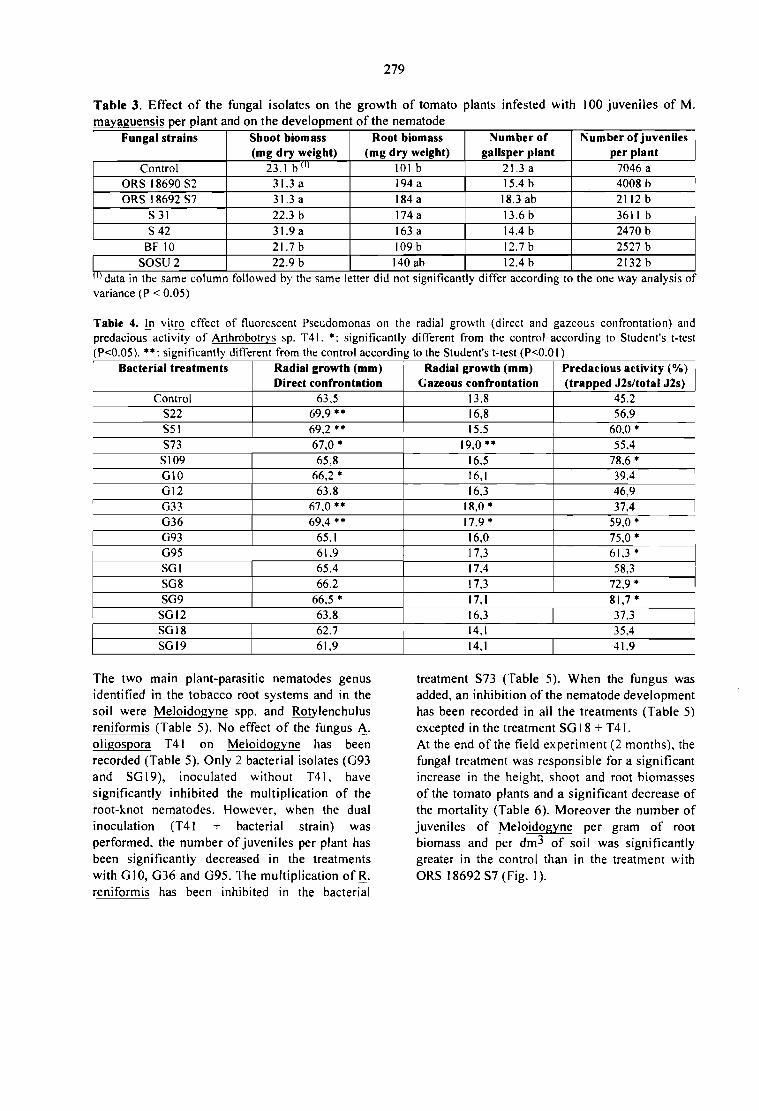
279
Table 3. Effect of the fungai isolates on the growth of tomato plants infested with 100 juveniles of M.rnavaauensis per plant and on the development of the nematode
data In the same column followed by the same letter did not significantly differ according to the one way analysis ofvariance (P < 0.05)
Fungai strains Shoot biomass Root biomass Number of Number of juveníles I
(mg dry weight) (mg dry weight) gallsper plant per plantControl 23.1 b 1'1 101 b 21.3 a 7046 a I
ORS 18690 S2 31.3 a 194 a 15.4 b 4008 bORS 18692 S7 31.3 a 184 a 18.3 ab 2112 b
S 31 22.3 b 174 a 13.6 b 3611 bS 42 31.9 a 163 a 14.4 b 2470 b
BF 10 21.7 b 109 b 12.7 b 2527 bSOSU 2 22.9 b 140 ab 12.4 b 2132 b
(I)
Table 4. In vitra effect of fluorescent Pseudomonas on the radial growth (direct and gazeous confrontation) andpredacious activity of Arthrobotrys sp. 1'41. .: significantly different from the contra] according to Student's t-test(P<0.05).•• : significantly different from the contrai according to the Student's t-test (P<O.O I)
Bacterial treatments Radial growth (mm) Radial growth (mm) Predacious activity (%)
IDirect confrontation Gazeous confrontation (trapped J2s/total J2s)
Contrai 63,5 13.8 45.2S22 69,9·· 16,8 56,9S51 69.2·· 15,5 60,0 •S73 67,0 • 19.0·· 55,4 I
SI09 65,8 16,5 78,6 •I GIO 66,2 • 16,1 39,4
GI2 63,8 16,3 46,9G33 67,0·· 18,0 • 37,4G36 69,4 •• 17,9 • 59,0 •G93 65,1 16,0 75,0 •G95 61,9 17,3 61.3 •SGI 65,4 17,4 58,3SG8 66,2 17,3 72,9 •SG9 66,5 • 17,1 81,7 •
SGI2 63,8 16.3 37,3SGI8 62,7 I 14,1 35,4SGI9 61,9 14,1 41.9
The two main plant-parasitic nematodes genusidentified in the tobacco root systems and in thesoil were Meloidogyne spp. and Rotylenchulusreniformis (Table 5). No effect of the fungus ~.
oligospora T41 on Meloidogyne has beenrecorded (Table 5). Only 2 bacterial isolates (G93and SG 19), inoculated without T41, havesignificantly inhibited the multiplication of theroot-knot nematodes. However, when the dualinoculation (T41 + bacterial strain) wasperformed, the number of juveniles per plant hasbeen significantly decreased in the treatmentswith GIO, G36 and G95. The multiplication of~.
reniformis has been inhibited in the bacterial
treatment S73 (Table 5). When the fungus wasadded, an inhibition of the nematode developmenthas been recorded in ali the treatments (Table 5)excepted in the treatment SG 18 + T41.At the end of the field experiment (2 rnonths), thefungai treatment was responsible for a significantincrease in the height, shoot and root biomassesof the tomato plants and a significant decrease ofthe mortality (Table 6). Moreover the number ofjuveniles of Meloidogyne per gram of rootbiomass and per dm3 of soil was significantlygreater in the control than in the treatment withORS 18692 S7 (Fig. I).
280
Table 5. Effect of lhe bacterial strains on lhe number of juveniles of Meloidogyne spp. and ROlylenchulus reniformisper tocacco plants inoculated or not with lhe nematophagous fungus Arthrobotrys sp. T41. • : significantly differentfrom the control (not inoculated treatment) according to the Student's t test (P < 0.05). ND : Not Determined.
Bacterial strains Meloidogyne spp. Rotylenchulus reniformisBacteria alone Bacteria + T4l Bacteria alone Bacteria + T4l
Control(withollt bacteria) 98944 90080 79452 41950 •S22 186820 75890 54200 • 20780 •S51 33410 35650 118240 NDS73 23674 194411 35790 • 43270 •
S109 71720 104180 70040 29840 •GIO 67010 23000 • 75280 31320 •GI2 48343 49655 98630 21350 •G33 57660 74562 65700 41137 •G36 75370 12740 • 80790 43530 •G93 14806 • 84212 43526 • 39925 •G95 68740 7250 • 86131 47310 •SGI 63700 89990 73160 26010 •SG8 147072 101551 59050 • 29670 •SG9 59640 146770 43730 • 28670 •
SGI2 48138 57910 69826 37400 •SGI8 35140 50140 51890 • 75160SGI9 7640 • 103643 42710· 44070 •
Table 6. Effect of Arthrobotrys oligospo!~ üRS 18692 S7 on height. mortality, average shoot and root biomass of10malo plants in the field experiment after 2 rnonth culture. (I): for each parameter, data in the same Iign followed bylhe same letter did not significantly differ according lhe one way analysis ofvariance (P < 0.05).
Parameters Compost blocks without fungus Compost blocksWith fungus
Height(cm) 23.2 b 31.8 a
Mortality(%) 36.8a 16.4b
Shoot biomass (g dry weight per plant) 7.1 b 12.6 a
Root biomass (g dry weight per plant) 0.7 b 1.8 a
la""",,, I• ,,10(\ I~("J! ,]
Fig. 1. Effects of the nematophagous fungus ORS 18692 S7 on the multiplication of Meloidogyne spp. juveniles intomato roots and in the soil, For each parameter (root and soil), the columns indexed with the same letter are notsignificantly different (P < 0.05). The data of lhe nematode population in lhe soil are expressed as x 100. in lhe roots,lhe numbers are calculated per g of dry weight 01' root biomass and divided by 105
.
DlSCUSSION
The different fungai isolates of Arthrobotrys fromSenegal and Burkina Faso were able to trap thejuveniles of M. mayaguensis and M. incognita.Ali these fungi could be good candidates for
biological control in coffe plantations as M.incognita has occured for rnany years in separateor mixed populations (with M. exigua, M.c9Jfeicola). Moreover the fungaI isolates can actagainst M. mayaguensis. This very damagingMeloidogyne species has spread throughout

vegetable-producing areas in West Africa and isnow present under various agro-climaticconditions (Mateille et aI., 1994).In the glasshouse experiment, the fungi had agreat influence on the growth of tomato plants inspite of a low rate of inoculation (I : 100, v :v).Compared to the concentrations commonly usedin the controlled ectomycorrhization (1: 10,v :v)(Duponnois & Garbaye, 1991), these resultssuggest that the nematophagous fungi have agreat capacity to colonize the soil andconsequently trapped the juveniles beforeinfecting the roots. The decrease of the number ofgalls per plant in the fungal treatments shows thisphysiological advantage. Moreover these fungican also use phytoparasitic nematodes as anenergy source and it can be assessed that morelarger is the nematode population, more importantwi II be the fungai deve lopment.However ali these experiments have beenperformed in controlled conditions and during ashort period (about one month). It is well knownthat the efficacy of these fungi strongly interactwith environmental factors such moisture, pH,temperature and organic matter (Mosse, 1972).Recent researches have shown that the biologicalactivities of soil microorganisms can be enhancedby some rhizosphere bacteria such as fluorescentPseudomonas (Duponnois et al., 1993; Mateille& Duponnois, 1996). The results presented in thispaper focussed on the great interactions betweenthis group of bacteria and the nematophagousfungus. These bacterial isolates have been calledNHB (Nematophagous HelperBacteria)(Duponnois et aI., 1998). From apractical point of view, it is generally assessedthat the production of spores and mycelialbiomass by nematophagous fungi is very limited(Cayrol, 1988). Moreover, the use of NHB candecrease the quantities of fungai inoculum addedto the soil because of their benefical effect on thesaprophytic growth and the predacious activitiesof the fungai strains. These NHB can also enlargethe effect of this practice on other pathogenicmicroorganisms such as Rotyl~.-chulus reniformi~
which is also a widespread plant parasiticnematodes. As we have demonstrated that thecompost blocks can be a good cultural support forthe biological control against nematodes withnematophagous fungi, it is easly possible to addtogether the fungus and a selected bacterial straininside the block. This new concept of thebiological control could integrated the propertiesof the fungus and those from the bacteria (PlantGrowth Promoting Rhizobacteria, etc) and their
281
beneficiai interactions. However, the mechanismsinvolved between these two types ofmicroorganisms must be elucidated in order toincrease the efficiency ofthis dual inoculation.
REFERENCES
Bourne, J.M., Kerry, B.R. & De Leij, F.A.A.M.(1994). Methods for the study of Verticilliumchlamydosporum in the rhizosphere. J. Nematol.,26 : 587-591.Cayrol , J.c. (1983). Lutte biologique contre lesMeloidogyne au moyen d'Arthrobotrysirregularis. Revue Nématol., 6 : 265-273.Cayrol, J.c. (1988). Lutte biologique contre lesMeloidogyne au moyen d' Arthrobotrysirregularis.Bull. OEPP, 18: 73-75.Duponnois, R & Bâ, A.M. (1998). Influence ofthe microbial community of a sahel soil on theinteractions between Meloidogyne javanica andPasteuria penetrans. Nematologica, 44 : 331-343.Duponnois, R & Garbaye, J. ( 1990). Somemechanisms involved in growth stimulation ofectomycorrhizal fungi by bacteria.CanadianJournal ofBotany, 68, 2148-2152.Duponnois, R. & Garbaye, 1. ( 1991).Mycorrhization heiper bacteria associated withthe Douglas fir-Laccaria laccata symbiosis :effects in vitro and in glasshouse conditions.Annale~ des Sciences Forestiêres, 48, 239-251.Duponnois, R., Garbaye, J., Bouchard, D. andChurin, J.L. (1993). The fungus-specificity ofmycorrhization helper bacteria (MHBs) used asan alternative to soil fumigation forectomycorrhizal inoculation of bare-rootDouglas-fir planting stocks with Laccarialaccata.Plant & Soil , 157 : 257-262.Duponnois, R., Sene, V., Sawadogo, A., fargette.M. & Mateille, T. (1996). Effects of differentspecies and strains of the nematophagous fungusArthrobotrys sp. from West Africa onMeloidogyne species. Technology transfer inbiological control : from research to practice.IOBC/OILB. Montpellier.Hussey, R. & Roncadori, R.W. (1982). Vesiculararbuscular mycorrhizae may limit nematodeactivity and improve plant growth. PI. Dis., 6: 914.Hutton, D.G., Eason-Heath, S.A.E. & CoatesBeckford, P.L. (1982). Control of Meloidogyneincognita affecting coffee. In : Proceedings of thethird Research & Conference on Root-KnotNematodes Meloidogyne spp., January 11-15,Region 1: 109-113.

Kerry, B.R. (1990). An assessment of progresstoward microbial control of plant-parasiticnematodes. J. Nernatol., 22: 621-631.Lordello, L.G.E. (1984). Nematoides das plantascultivadas. 8° Ediçao. Livraria Nobel S.A. 314 p.Mateille, T. & Duponnois, R. (1996). Organismesauxiliaires de la lutte contre les nématodesphytoparasites. Applications aux culturesmaraichêres, Séminaire national sur la protectiondes cultures et des stocks dans une agriculturerégénératrice. Thies, Sénégal, 27-29 Mars 1996.Mosse, B. (1972). The influence of soil type andEndogone strain on the growth of mycorrhizalplants in phosphate defficient soi Is. Revue Ecol.Biol. ~oil. 10 : 529-537.Pelagatti, O., Nencetti, V. & Caroppo, S. (1986).Utilizzazione del formulato R35ü a based iArthrobotrys irregularis nel controllo diMeloidogyne incognita. Redia, 89: 276-283.
282
Racke, J. & Sikora, R.A. (1986). Influence ofrhizobacteria on Globodera pallida early rootinfection and Erwinia carotovora tuber soft rot ofpotato. Revue Nemato1., 9 : 305. (Abstract).Schuster, R.P., Amin, N. & Sikora, R.A. (1995).Utilization of fungai endophytes for the biologicalcontrol of Radopholus similis on banana.Nematologica,41 : 342. (Abstract).Seinhorst, J.W. (1950). De betekenis van detoestand van de grond voor het optreden vanaanstasting door het stengelaaltje Ditylenchusdipsaci Kühn Filipjev.- Tijdschrift overPlantenziekten, 56.292-349.Seinhorst, J.W. (1962). Modifications of theelutriation method for extracting nematodes fromsoil. - Nematologica. 8. 117-128.
I
j

283
LOS INHIBIDORES DE AMILASAS COMO ALTERNATIVA EN LA PRODUCCION DE CAFERESISTENTE A LA BROCA,
Arnubio Valencia Jiménez I , Maria T. González 2 y Alex E. Bustillo P 2.
I Departamento de Química, Facultad de Ciencias Exactas y Naturales. Universidad de Caldas-Colombia2 Centro Nacional de Investigaciones de Café, Cenicafé. AA N°2427. Manizales-Colombia.
JUSTIFlCACIÓN
La broca dei café. Hypothenemus hampei (Ferrari)(Coleoptera: Scolytidae), es sin lugar a dudas elproblema fitosanitario de mayor importanciaeconómica para la caficultura colombiana dadoque ataca el fruto desde los primeros estados dedesarrollo hasta estados próximos a la cosecha,ocasionando considerables pérdidas tanto en laproducción como en la calidad dei grano. EIcontrol que se recomienda para esta plaga enColornbia, ha considerado la estrategia de manejointegrado de la broca (MIB) con el fin dedisminuir el dano y los niveles poblacionales deiinsecto. Dentro de las medidas se considera elcontrol químico; el uso de parasitoides como:Prorops nasuta Waterston (Bethylidae) yCephalonomia stephanoderis Betrem(Bethylidae), entomopatógenos como: Beauveriabassiana y Metarhizium anisopliae, el controlcultural a través de recolecciones frecuentes defrutos maduros y el desarrollo de plantasresistentes a esta plaga.
Existen varias alternativas para el desarrol1o devariedades resistentes a una plaga en particular.inicialmente. se deben identificar las fuentes deresistencia ai insecto específico, mediante laexploración dei germoplasma y de los parentalescercanos de la especie. EI conocimiento de labiologia dei insecto, y la influencia de los factoresbióticos y abióticos sobre la biologia de éste,especialmente, hábitos alirnenticios, oviposición,movirniento, parámetros de crecimiento yfecundidad, fundamentan eJ conocimientodetallado de la naturaleza intrínseca de la plaga através de la exploración de las rutas metabólicas yfisiológicas que puedan ser blanco de inhibición(Figura I). Así rnismo, el aparato digestivo de losinsectos y muy especialmente sus enzimasdigestivas como proteasas y amilasas seconstituyen en uno de los puntos más susceptiblesen la fisiología de estos, dado que esta rutametabólica es la única forma deaprovisionamiento energético dei insecto.
Desarrolo de plantasTransgênicas
Figura 1. Estrategia de manejo integrado de Hypothenemus hampei

El problema de la broca, amerita queseconsideren investigaciones tendientes a explorartras vías como el estudio de los mecanismos deresistencia aI ataque de broca y másespecificamente el uso de compuestosanti metabólicos de ocurrencia natural. Algunosde estos compuestos corresponden a losinhibidores de enzimas digestivas dei insecto,tales como inhibidores de proteasas y amilasas loscuales tienen un enorme potencial para serincluidos dentro de programas de mejoramientogenético a través de la biotecnología.El desarrollo de variedades transgénicas de cafécon resistencia a broca, contribuirá de manerasignificativa a la reducción en el uso deagroquímicos los cuales por demás no proveen deuna protección total, a la vez que el usoindiscriminado de estos puede conducir a laaparición de resistencia en el insecto comoconsecuencia de la alta presión de selección. Estametodología adicionalmente conlleva a lareducción de costos en el manejo de la plaga, a laeliminación de residuos tóxicos en los productosalirnenticios, a la conservación de la salud yprotección dei medio ambiente.
ANTECEDENTES
Por varios afíos las investigaciones se han dirigidoai estudio de la regulación y biosíntesis decompuestos específicos asociados con losmecanismos de defensa de plantas ai ataque deinsectos y patógenos. Estos compuestos fueronconsiderados como desecho dei metabolismosecundario de las plantas con funcionesdesconocidas. Sin embargo, actualmente se sabeque muchos de estos compuestos actúan solos oen conjunto jugando un papel en la resistencia deplantas ai ataque de insectos y patógenos (Ryan,1990). Entre esos metabol itos (Tabla I). seincluyen lectinas (Chrispeels y Raikhel, 1991;Huesing et ai., 1991), ceras (Eigenbrode et ai.,1991). fenoles (Summers y Felton, 1994),aminoácidos (Rosenthal y Dahlman,· 1991;Rosenthal et al ..1989), azúcares (Juvik etal.,1994; Liedl et al ..1995), inhibidores deamilasas (Huesing et ai., 1991; Shade et al., 1994)e inhibidores de proteinasas (Burgess et ai .. 1994;Orr et ai .• 1994; Wolfson y Murdock, 1995; Ryan1973,1990; Boulter et al., 1990). Estos compuestosquímicos han proporcionado diversos sistemaspara el estudio de mecanismos de defensainducible en plantas. En muchas plantas puedensintetizarse en respuesta ai ataque de plagas opatógenos generando sefíales intra o intercelularesque activan genes que codifican para su síntesis(Ryan, 1990).
284
La mayoría de los inhibidores se han encontradoen grandes concentraciones en semillas otubérculos de algunas plantas, especialmente engramíneas y leguminosas (Ryan, 1990; Green yRyan, 1972) y se han aislado a partir de frijolmungo (Baumgartner et ai., 1976; Kapur et al.,1989), centeno y trigo (Chang y Tsen, 1981;Gatehouse et al.,1986; Deponte et ai., 1976),guandúl (Godbole et ai., 1994b), papa (Eddy etai.• 1980; Ishikawa et ai., 1994; Orr et ai, 1994),cowpea (Gatehouse et al..1980; Hilder etal.,1989) tabaco (Geoffroy, 1990), amaranto(Rodriguez et al.,1993), tomate (Walker y Ryan,1977), arveja (Domoney et al., 1993), garbanzo(Mulimani et al., 1994) maíz (Blanco e IturbeChiüas, 1981) entre otros. Los inhibidores deproteinasas y amilasas se producen en los tejidosvegetales en diferentes proporciones.Generalmente, son proteínas con pesosmoleculares por debajo de 50Kd y en algunasocasiones por debajo de 20Kd (Boisen y Djurtoft,1981; Chang y Tsen, 1981; Gatehouse et al., 1980;Geoffroy et al.,1990; Godbole et al..1994a;Rodriguez et al.,1993; Domoney et al.,1993;Blanco e Iturbe-Chifias,1981; Weselake etal.,1983; Garcia-carrefío et al., 1993)presentándose en fonna de dímeros o tetrámeros.Muchos inhibidores son producto de familiasmultigénicas y no es muy común encontrarisoinhibidores expresando diferente especificidadhacia proteasas (Ryan, 1990).
Papel fisiológico de los inhibidores de amilasasEI papel fisiológico de los inhibidores de amilasasen las plantas es desconocido (Gatehouse et al..1986). Existen evidencias que indican quepueden actuar como reserva proteica en lassemil1as; sin embargo, los inhibidores no afectangeneral mente amilasas endógenas excepto en muypocos casos. La especificidad de los inhibidoresde amilasas y proteasas sobre las enzimas enestudio son bien conocidas, es así como eIinhibidor de amilasas H-28 mostró actividadinhibitoria muy específica para las amilasas deinsectos y ninguna actividad inhibitoria paraamilasas de fuentes vegetales, fúngicas y animalesincluyendo una amilasa de saliva humana. Seconocen adernás, muchos ejemplos en donde losinhibidores de amilasas y proteinasas,particularmente aquellos aislados de cereales, hanmostrado inhibición de las amilasas dei intestinode insectos (Burgess et al.,1994; Jhonston et ai..1993. Christel1er y Shaw, 1989; Chagolla etal .•1994; Grant et al., 1995; Shade et ai. 1994;Weselake et al.,1983), lo cual ha conducido aproponer que los inhibidores de enzimasdigestivas juegan un papel protectivo contra
Ij

insectos predadores.Durante el ataque de uninsecto a un órgano vegetal se desencadena laproducción de compuestos altamente volátilescomo los jasmonatos y meti I jasmonatos, loscuales se transportan por el aire a otros órganosdentro o fuera de la planta y una vez allí daninicio a una serie de reacciones bioquímicas queconducen a la síntesis de compuestos queconfieren resistencia ai ataque de insectos.Dentro de estos compuestos de ResistenciaSistémica Adquirida (RSA) mediados pormoléculas seüalcomo los jasmonatos y metil jasmonatos; seencuentran los alcaloides y muy especialmente losinhibidores de proteasas (Enyedi et al., 1992).
Efecto de los inhibidores sobre la fisiologiadigestiva de los insectosCuando los inhibidores se encuentran presentes engrandes concentraciones en el alimento. puedenalterar significativamente varios procesosfisiológicos y digestivos interfiriendo con elnormal crecimiento y desarrollo de los insectos.Los inhibidores de amilasas y proteasas no son unproblema directo para los humanos dado que losalimentos que los contienen en grandesproporciones son sometidos a procesos de cocciónutilizando altas temperaturas lo cual conduce a suinactivación (Ryan, 1990).Varios efetos fisiológicos se manifiestan en elinsecto como consecuencia de la ingestión deestos inhibidores (Tabla t), dentro de los cualesse destaca el efecto antinutricional, lahipersecreción de enzimas, la pérdida de apetito yla eventual muerte.Un mejor conocimiento de la regulación de lafisiología digestiva de los insectos y de los efectosreales de los inhibidores de proteasas y amiJasas
285
sobre estos procesos permitirá conocer elpotencial de dichos inhibidores en los sistemas ymecanismos de defensa natural de las plantas.
Evaluaciôn de la activldad biológica deinhibidoresEn general existen varias vías que pueden serusadas para evaluar la actividad biológica deinhibidores contra insectos herbívoros. La formamás directa mide la capacidad de un ínhibídorpara inhibir in vitro la actividad enzimáticaproveniente dei intestino de un organismo blanco(Hatekeyama et al.,1992; Kang y Fuchs, 1973). EIprocedimiento involucra la extracción de lasenzimas del intestino medio dei insecto, suincubación con eJ inhibidor y el análisis de suactividad (Valencia et al., 1994). Esta pruebaconduce a la determinación de la clase de enzimapresente en el intestino medio dei insecto(Dionysius et al.,1993; Houseman y Thie, 1993) yestablece el grado de unión entre la enzima y eJinhibidor. Sin embargo, esta metodología nosiempreindicará que el insecto responda fisiológicamenteai tratamiento (Broadway, 1995a).Otra forma para evidenciar la respuesta de unorganismo aI tratamiento con inhibidores, consisteen incorporar los inhibidores a las dietasartificiales y evaluar la sobrevivencia, crecimientoy desarrollo dei insecto (Broadway, 1995b;Burgess et al., 1994; Johnston et al., 1993; Orr etal.,1994; Gatehouse et al., 1986; Steffens etal.,1978; Lipke et al., 1954). Adicionalmente, sepuede evaluar a nivel de planta la respuesta deIinsecto a la presencia de inhibidores (Broadway,1995c; Green y Ryan, 1972; Hilder et al.,1987;Shade et al., 1994; Schmidt, 1994).
Tabla 1. Efectos fisiológicos causados en los insectos como consecuencia dei consumo de inhibidores.Efectos fisiológicos de los inhibidores de enzimas digestivas sobre los insectos IInhibición de la muda (Gatehouse et al., 1992)
I Hipertrofia pancreática (Liener, 1980)
I Reducción en fecundidad (Spats and Harris, 1984)
I Hiperproducción de enzimas digestivas (Ryan, 1990)I Inhibición de proteolisis (Ryan, 1990)~ Disminución en % de sobrevivencia (Johnston et al., 1993; Broadway, 1995b)

286
Transformoclôn genéticaLa primera transforrnación de una planta con un
gen para inhibidor de proteasas rue desarrolladapor Hi Ider et al.. I98? El gen para el in hibidor de
Tripsina de Cowpea (Vigna unguiculafa) (CpTI)
fue transgénicamente insertado en plantas de
tabaco las cuales mostraron mayor resisrencia aI
ataque de Heliothis virescens que plantas sin la
presencia dei gen que codi ficaba para el inh ibidor(Figura 2).Esta proteína de Cowpea rue considerada como
una fuente valiosa para ser transferida a erras
especies vegerales por medio de la ingenieria
genética por varias razones: Es un efecti voantimetabólito contra un gran número de insectos
plagas de los ordenes Coleoptera, Orthopiera y
Le pido ptera. si n erectos de ieiéreos 50 bre
marniferos y es adernas una protei na pequena con
un peso de 80 Kd (Gaiehouse et 01. , 1992 ).La t ransforrnación genética de plantas a través de
la expres ión de inhibidores de enzimas digestivasde insectos, ha sido usada por otros grupos deinvest igadores en cultivos de importanciaeconórnica de mostrando el potencial de estescomo mecanismo de resistencia ai ataque de
insectos herbívoros (Ta bla 2). Entre elia s. se
incl uyen : arveja (Schrnidt, )994; Shade etQI..1994), alfalfa (Thornas et 01.,)994), tabaco(Hilder et al.,1987) y tomate (Graham et aI.,1986) entre otros,
Figura 2. Plantas de tabaco transg énicas expresando CpTJ (Tomado de Gatehouse et 01., 1992)
-Planta Tipo de inhibidor expresado Referenda
Nico üana tabac um Inhibidor de Tripsina de Cowpea (CpTJ) Hi lder et 01.• 1987
Inhibidor I de tomate Gatehouse et al.. )992
Inhibidor II de lo mate Johnson et ai . 1989
Inhibidor II de papa Gatehouseet al., 1992
Pisum sativum lnhibidor de Alfa amilasa (A I·Pv) Shade,1994
Medicago saliva Inhibidor de Proteinasa Thornas et 01.. 1994
Gossypium hirsutum Inhibidor de Tripsina de Cowpea (CpTI) Woodend, 1994
Ta bl a 2. Plantas t ransgcnicas que expresan inh ibidores de ami lasa v/o proreasas,

287
Tabla 3. Desarrollo dei gorgojo de arveja (Bruchus pisorum ) en semi lias T, de seis plantas dearveja transgénica (F la)'
Plantas No. De sem iIIas
No. de semillas No. de semillas infestadas infestadasI
I
con eme gencla oe
T 5 Control T 5 Control T 5 ControlI 50 38 27 26 O 232 45 44 42 35 O 32
3 50 42 30 35 O 304 48 46 35 30 O 255 O 35 26 27 O 216 46 39 29 28 O 26
Total 279 244 189 181 O 157Fuente: Schroeder et al., 1995
Expresión de estas proteínas en plantas transgénicasha mostrado la perdurabilidad a través de sucesivasgeneraciones de la especie vegetal como en el casode arveja transgénica (Tabla 3) mostrando laestabilidad genética de la planta transformada.La manipulación genética de plantas como parte deun programa de mejoramiento puede tener unacontribución significativa en la producción de plantasresistentes a insectos; transfiriendo únicamente el genseleccionado y acortando el tiempo de producción denuevas variedades. Así mismo, esta metodologíaofrece rnuchas ventajas sobre el uso exclusivo deagroquímicos (Gatehouse et al., 1992) tales como:I. Provee larga protección;2. Los insectos son afectados en su estado más
sensible;3. La protección es independiente de factores comoel clima;4. Protege tejidos vegetales de dificil acceso a los
insecticidas;5. Los efectos están restringidos ai insecto plaga6. No afecta el media ambiente, son biodegradablesy sin toxicidad ai hornbre y animales entre otros.Teniendo en cuenta la especificidad de estosinhibidores (Richardson, 1977), sobre las enzimasdigestivas dei insecto se consideran excelentesmarcadores para incluirlos en programas demejoramiento genético conducente a buscar suexpresión en los tejidos vegetales susceptibles aiataque de plagas o patógenos (Gatehouse et al.,1992).En conclusión las plantas producen diversas proteínascapaces de inhibir una enzima en particular, lo cualles confiere resistencia aI ataque de insectosherbívoros. Esta investigación, pretende anal izarcuidadosamente el potencial de los inhibidores deamilasas y proteasas de insectos como alternativa enel manejo integrado de la broca. Hypothenemushampei. Para ello se hace necesario abordar lapurificación y caracterización tanto de las enzimasdigestivas presentes en el intestino medio deI insectocomo la de inhibidores para estas enzimas. De igual
forma y teniendo en cuenta que los resultados enpruebas enzimáticas in vitro no siempre correspondena los encontrados en pruebas de campo con materialtransgénico, se pretende incluir estos inhibidores enla dieta merídica para la cría de la broca de forma quela información generada a lo largo dei proyectopermita seleccionar inhibidores de enzimas digestivasde la broca que puedan ser utilizados en un futuro nornuy lejano, en la producción de café transgénico.Así rnisrno, se generará un mejor conocimiento de losmecanismos de acción de estos compuestos y elgrado de interacción con las enzimas en estudio.
LITERATURA ClTADA
BAUMGARNERT, B. ; CHRISPEELS, J. M. Partialcharacterization of a protease inhibitor, whichinhibits the major BLANCO-LABRA, A. ; ITURBECH INAS, F. A. Purification and Characterization ofan alfa-amilase inhibitors from mize (Zea maize).Journal ofFood Biochemistry. 5:1-17.1981.BOISEN, S. ; DJURTOFT, R. Trypsin inhibitor fromRye endosperm: Purification and properties. CerealChemistry. 58(3) 194-198.1981BOULTER, D.; GATEHOUSE, J. A.;GATEHOUSE, A. M. R.; HILDER. A. V. Geneticengineering of plants for insect resistance. Endeavour(Inglaterra). New series.14: 85-190. 1990.BROADWAY. M.R. Characterization and ecologicalimplications of midgut proteolytic activ ity in larva IPieris rapae and Trichoplusia ni. Journal ofChemistry Ecology. 15:2101-2113. 1989.BROADWAY, M. R. Potential use of naturalproducts as resistance factors against herbivorousinsects. Syrnposium Cornell University, August1995b (mimeografiado) 3p.BROADWAY. M. R. Dietary proteinase inhibitorsalter complement of midgut proteases. FromEntomological Society of America Conference:Recent advances in the nutritional physiology ofherbivorous insects. 1995a. (Mimeografiado) 29p.

BROADWAY, M. R. Are insects resistant to plantproteinase inhibitors? Journal of Insect Physiology.41(2): 107-116.1995c.BURGESS, E. P. l; MAIN, C. A.; STEVENS, P. S.;CHRISTELLER, l T. GATEHOUSE, A. M. R.;LAING, W. A Effects of protease inhibitorconcentration and combination on the survival,growth and gut enzyme activities of the black fieldcricket, Teleogryllus commodus. Journal InsectPhysiology. 40(9): 803-811. 1994.CHAGOLLA L. A.; BLANCO, L. A.; PATTY, A.,SANCHEZ, R; PONGOR, S. A novel a-amylaseinhibitor from amaranth (Amaranthushypochondriacs) seeds. The Journal of BiologicalChemistry. 269(38): 23675-23680.1994.CHANG, C. R.; TSEN, C. C. Isolation of trypsininhibitors from Rye, Triticale, and Wheat samples.Cereal Chemistry. 58(3) 207-210.1981.CHRISPEELS, l M.; RAIKHEL, V. N. Lectins,lectin genes. and their role in plant defense. The PlantCell. 3,1-9.1991.CHRISTELLER, l T.; SHA W B. D. The interactionof a range of serine proteinase inhibitors with bobinetrypsin and Costelytra zealandica trypsin. InsectBiochemistry. 19(3): 233-241.1989.DEPONTE, R.; PARLAMENTI, R.; PETRUCCI, T.;SILANO, V.; TOMASI, M. Albumin a-amylaseinhibitor families from wheat flour. CerealChemistry.53,805-820. 1976.DIONYSIUS, A D.; HOEK, S. K.; MILNE, M. .I.;SLATTERY, L. S. Trypsin-like enzyme from sandcrab (Portunus pelagicus) purification andcharacterization, Journal of Food Science. 58(4):780-784.1993.DOMONEY,C.;WELHAM,T.; SIDEBOTTOM, C.Purification and characterization of Pisum seedtrypsin inhibitors. Journal of Experimental Botany.44(261): 701-709. J993.EDDY. L. l; DERR, E. J. ; HASS. G. M.Chymotrypsin inhibitor from potatoes interactionwith target enzymes. Phytochemistry. 19:757-761.1980.EIGENBRODE. S. D.; STONER. K. A.; SHELTON,A. M. ; KAIN, W. C. Characteristics of glossy leafwaxes associated with resistance to diamondbackmoth (Lepidoptera :Plutellidae) in Brassica oleracea.Journal of Economic Entomology. 84: 1609-1618.1991.ENYEDI, A. l; YALPANI, N.; SILVERMAN, P.;RASKIN, I. Signal molecules in systemic plantresistance to pathogens and pests. Cell (70): 879-886.1992.GARCIA-CARRENO, F. L.; DIMES, L. E.;HAARD. F. N. Substrate gel electrophoresis forcomposition and molecular weight of proteinases orproteinaceus proteinase inhibitors. AnalyticalBiochemistry. 214: 65-69.1993.
288
GATEHOUSE, A. M. R.; HILDER, V. A.;BOULTER, D. Plant genetic manipulation for cropprotection. Wallingford (England) CABInternational, 1992. 266 p. (Biotechnology In
Agriculture). No 7.GATEHOUSE, A. M. R.; GATEHOUSE, A . .I.;BOU LTER, D. Isolation and characterization oftrypsin inhibitors from cowpea (Vigna unguiculata).Phytochemistry. 19:751-756.1980.GATEHOUSE, A. M. R.; FENTON, A. K.; JEPSON,1..; PAVEY, J. D. The effects of a amylaseinhibitors on insect storage pests: inhibition of aamylase in vitro and effects on development in vivo.Journal of the Science of Food and Agriculture.37:27-734. 1986.GEOFFROY, P.; LEGRAND, M.; FRITlG, B.lsolation and characterization of a proteinaceousinhibitor of microbial proteinases induced during thehypersensitive reaction of tobacco to tobacco mosaicvirus. Molecular Plant-Microbe Interactions. 3(5):327-333. 1990.GODBOLE, A. S.; KRISHNA, G. T.; BHATlA, R.C. Changes in protease inhibitory activity frompigeon pea (Cajanus cajan (L.)(Millsp) during seeddevelopment and germination. Journal ofthe Scienceof Food and Agriculture. 66,497-501. 1994b.GODBOLE, A. S.; KRISHNA, G. T.; BHATIA. R.C. Purification and characterization of proteaseinhibitors from pigeon pea (Cajanus cajan (L.)Millsp) seeds. Joumal of the Science of Food andAgriculture. 64: 87-93. 1994a.GRAHAM. S. i., HALL. G.; PEARCE, G.; RYAN,A.C. Regulation of Synthesis of proteinase inhibitorsI and II mRNAs in leaves of wounded tomato plants.Planta' 69:399-405. 1986.GRANT.G.; EDWARDS. E. .I.; PUSZTAI. A. aamylase inhibitor leveis in seeds generally availablein Europe. Journal of the Science of Food andAgriculture. 67:235-238. 1995.GREEN, R. T.; RYAN, C. A Wound-inducedproteinase inhibitor in plant leaves: A possibledefense mechanism against insects. Science.175:776-777.1972.HATAKEYAMA, T.; KOHZAKI, H.; YAMASAKI,N. A microassay for proteases using succinylcaseinas a substrate. Analytical Biochemistry. 204: 181184. 1992.HILDER, A. V.;GATEHOUSE, A M. R.;SHEERMAN, E. S.; BARKER, F. R.; BOULTER,D. A. Novel mechanism of insect resistanceengineered into tobacco. Nature. 330: 160-163. 1987.HILDER, V. A., BARKER F. R., SAMOUR A. R.GATEHOUSE M. R. A, GATEHOUSE A. J. ;BOULTER D. Protein and cDNA sequences ofBowman-Birk protease inhibitors from the cowpea(Vigna unguiculata Walp.) Plant Molecular Biology.13:701-710.1989.

HOUSEMAN, 1. G.; THIE N. M. R. Difference indigestive proteolysis in the stored maize beetles:Sitophylus zeamais (Coleoptera:Curculionidae) andProstephanus truncatus (Coleoptera: Bostrichidae).Journal of Economic Entomology. 86(4): 10491054.1993.HOUSEMAN, J.G.; DOWNE, A.E.R.;PHILOGENE, BJ. R. Partial characterization ofproteinase activity in the larval midgut of theEuropean com-borer Ostrinia nubilalis Hubner(Lepidoptera:Pyralidae) Canadian JournalZoological. 67(4):864-868. 1989.HUESING, E. 1.; SHADE, E. R.; CHRISPEELS, 1.M.; MURDOCK, L. L. a-Amilase inhibitor, notpyotohemagglutinin explains resistance of commonbean seeds to cowpea weevill. Plant Physiology. 96:993-996.1991 .JOHNSTON, K. A; GATEHOUSE, J. A;ANSTEE, J. H. Effects of soybean protease inhibitorson the growth and development of larval Helicoverpaarmigera. Journal of Insect Physiology. 39(8):657664.1993.JUVIK, 1. A.; SHAPIRO, J. A.; YOUNG, T. E.;MUTSCHLER, M. A. Acylglucoses from wildtomatoes alter behavior and reduce growth andsurvival of Helicoverpa zea and Spodoptera exigua(Lepidoptera: Noctuidae) Journal of EconomicEntomology. 87:482-492.1994.KANG, S.; FUCHS, M. S. An improvement in thehummel chymotrypsin assay. AnalyticalBiochemistry. 54: 262-265. 1973.LIENER, I.E.; KAKADE, M.L. Protease inhibitors.In Toxic constituents of plant foodstuffs, ed. I.E.Liener,pp 7-71. New York: Academic. 1980.LIPKE, H.; FRAENKEL, G. S.; LIENER, I. E.Effect of soybean inhibitors on growth of Triboliumconfusum. Journal of Agricultural and FoodChemistry. 2: 410-415.1954.LIEDL, B. E.; LAWSON, D. M.; WHITE, K. K;SHAPIRO, J, A; COHEN, D. E.; CARSON, W. E.;TRUMBLE, J .T .; MUTSCHLER, M. A Acylsugarsof wild tomato Lycopersicum pennellii alters setlingand reduces oviposition of Bemisia argentifolli(Homoptera: Aleyroedidae) Journal of EconomicEntomology.88:742-748.1995.MULIMANI, V. H.; RUDRAPPA, G.; SUPRIYA,D. Alfa-Amilase Inhibitors in chick pea (Cicerarietinum L). Journal of the Science of Food andAgriculture. 64: 413-415,1994.
289
ORR, L. G.; STRICKLAND, A. J.; WALSH, T. A.Inhibition of Diabrotica larval growth by amulticystatin from potato tubers. Joumal of InsectPhysiology. 40( I0):893-900.1994.RICHARDSON, M. The proteinase inhibitors ofplant and micro-organisms. Phytochemistry 16:159169.1977.RODRIGUEZ, S. V.; SEGURA, N. M.;CHAGOLLA, L. A.; VERVER, A.; CORTINA, V.;MARTINEZ, G. N.; BLANCO L. A Purification,characterization, and complete amino acid sequenceof a trypsin inhibitor from Amaranth (Amaranthushypocondriacus) seeds. Plant Physiology. 103: 14071412.1993.ROSENTHAL, G. A.; DHALMAN, D. L.Incorporation of L-canavanine into proteins and theexpression of its antimetabolic effects, Joumal ofAgricultural and Food Chemistry. 39:987-990.1991.RYAN, C. A Proteolytic enzymes and theirinhibitors in plants. Annual Review of PlantPhysiology. 24:173-196.1973.RYAN, C. A Protease inhibitors in plants: Genes forimproving defenses against insects and pathogens.Annual Review of Phytopatology. 28:425-449. 1990.SCHROEDER, E. H. ;GOLLASCH, S.; MOORE,A.; TABE, L.A; CRAIG, S.; HARDIE.D.C;CHRISPEELS, MJ.; SPENCER, D.; HIGGINS, TJ.Bean alfa amylase inhibitor confers resistance to thepea weevil (Bruchus pisorum) in transgenic peas(Pisum sativum L.) Plant Physiology. 107: 1233-1239.1995.SHADE, E. R.; SCHROEDER, E. H.;PUEYO, 1. 1.;TABE, M. L.; MURDOCK, L. L.; HIGGINS, V. T.J.; CHRISPEELS, 1. M. Transgenic pea seedsexpressing the a-amylase inhibitor of the commonbean are resistant to bruchid beetles.Biotechnology. 12:793-796. 1994.SCHMIDT, K. Genetic engineering yields first pest
resistant seeds. Science. 265: 739. 1994.SPATS, G.E.; HARRIS, R.L. Reduction offecundity, egg hatch and survival in adult horn fliesfed protease inhibitors. Southwest. Entomology.4 :399-413. 1984.STEFFENS, R.; FOX, R. F.; KASSELL, B. Effectoftrypsin inhibitors on growth and metamorphosis ofcorn borer larvae Ostrinia nubilalis (Hubner).Journalof Agricultural and Food Chemistry. 26(1): 170-174.1978.

E51ST
• Produto de alto eficácia no controle do complexo de doenças no café :
Ferrug m, Cerco po a e Ascochyta• Vigor e produtividade promovidos pelo Cyprcconozole.• Maior retençõo faliar devido 00 Cobre.• Compatível com inseticidas e adubos [ol iores (Sulfato de Zinco) no r- oplicoc ôo.
• Dose: 2,5 a 3,0 Kg/ha• Intervalo de aplicação : de 60 a 75 dias.
Visite a nossa homepage:www.novartis.com.br
J1111ft•1ft-t --- . :::.:::-~
tl) NOVARTIS
Erncrg ências : r b nlilo Nl)Y;'lrd<H Horas. Te!.: 10_.24) JS~ . ~58 1.
RI "' I~) I - CI ;\ :;' ~ l fl "',HJ'J pdll MS 4,'mll"
Prf .dUlu pcn~:l lI,.vnt a;1 C I:tS'L'
'f,.xl~:4)lc'.!-t'~ :l Ir (A Ir :l nl-:l1h." T !.i.\ I ( V .
" .1' \0;;1 ," m:'lrd, , ). lo. pd('l ([lr\~ 1 ,\ l,;nrT'll l
prHduc. ~ tJc Cb... ~~ óc· l)cr i ":\ll u'I t.'~tJ("
Atll hil'fl r~l l .!ov' ul., . fl'<:11P.O Ml .
Pr,"Juw rt:~ i $((:,,:lu rm M;\ ~, ) h on": 03."" :< . .\1.Ht. .l (C~ i .... (e.l '.b d. ~(J\·.:l fC i ~ .
fh\i1 ~i .l . ~\UiçJ .•
ATENÇÃOEste produto é perigoso à sáuce
humana. animal e ao meio ambiente.Leia atentamente e siga rigorosamen le
as instruções coruieas no rótulona bula e receua. Utilize sempre os
equipamentos de proreçáo indívicuat,Nunca permita a uuhza çào
do produto por menores de idade .Consulte sempre
um engenheiro agrônomo.Venda sob 'eceiluárlo a9,onélmlco.


293
ESTUDOS FISIOLÓGICOS E PRODUÇÃO DE FUNGOS MICORRÍZEOS
Álvaro Alberto De Araújo] & Sevastianos Roussos-
2IRD (anciennement ORSTOM) - Institut de la Recherche pour le Développement, Laboratoire deMicrobiologie, Université de Provence, CESB/ESIL Case 925. 163, Avenue de Luminy, F-13288 MarseiÍlecedex 9. FRANCE I Bolsista CNPq/Brasília - BRASIL
Palavras chaves: Fungo, crescimento apical, Pisolithus, Suillus, fisiologia, meio agar
RESUMONeste trabalho estudou-se a fisiologia de fungos ectomicorrízeos tais como Suil/us e Pisolithus, cultivadosem meio agar. Os objetivos principais são: produção de pré-inóculo para a micorrização de plantas jovense desenvolvimento de técnicas para o tratamento das amostras possibilitando analisar o consumo deaçúcar, produção de enzimas e biomassa produzida. Numa primeira etapa, verificou-se a influência datemperatura, do pH e de sais de magnésio no crescimento apical dos fungos ectomicorrízeos. Em seguida,desenvolveu-se uma técnica para o tratamento das amostras em meio agar, utilizando uma película decelofane onde pode se analisar separadamente o meio agar da biomassa (fungo que cresceu sobre apelícula de celofane). Esta técnica permite, entre outros, avaliar o consumo de glucose com o crescimentodo fungo, a produção de enzimas e a quantidade de biomasa produzida. Estas técnicas assim como ocultivo/produção defungos micorrízeos podem ser aplicadas para o caso das endomicorrízas do café.Neste trabalho, apresentamos uma técnica para o estudo de parâmetros de crescimento de fungos sobremeio agar. Placas de Petri de 60 mm de diâmetro, contendo 10 ml de meio de cultura sobre o qual sedepositou um disco de papel celofanefim de permitir a separação fácil do micélio após crescimento. Para aanálise da produção de biomassa fúngica, um disco de papel celofane previamente estéril; é depositadosobre a superficie do meio de cultura nas placas de Petri. A inoculação é realizada colocando-se umpedaço de micélio no centro do disco de celofane e as culturas são incubadas à 25°C na obscuridade. Ocrescimento radial da colônia é medido e a biomassa (peso seco) produzida é determinada. Ao meio decultura, 90 ml de água destilada é adicionada e tudo é homogeneizado com a ajuda do Potter (BioblockScientific) para as análises de açúcares redutores e pH O consumo de açúcar, variação do pH e produçãode biomassa após 5, 10 e 15 dias de incubação à 25°C para as linhagens de Suillus collinitus e Pisosithustinctorius em diferentes meios de cultura (PDA, BAF, MNM, MP, CM8, MC) são mostrados neste trabalho.Esta técnica esta bem adaptada às culturas dos fungos e permite avaliar o consumo de açúcar e nttrogênio,os perfis de pH, a produção de biomassa, enzimas e pigmentos.
INTRODUÇÃO
Microorganismos estão presentes em grandequantidade junto às raízes de plantas, e elespossuem um papel importante em inúmerosprocessos fisiológicos. Pelmont (I) afirma queestes processos dinâmicos são mediados porassociações de microorganismos participantes nasatividades das raízes tais como a fixação denitrogênio e a mobilização do fósforo. Mousain(2,5) e Strullu (3) afirmam que as associaçõessimbióticas predominantes nas raízes das plantassão de origem bacterial como Rhizobium ou deorigem fúngica no caso das micorrizas. Marx andCordell (4) definiram as micorrizas como uniãoestável baseada em trocas recíprocas entre asraízes das plantas e os fungos, cada umotimizando seu desenvolvimento graças a essaassociação. Mousain et ai. (5) relataram quefungos ectomicorrízeos tais como Lactarius Pers.,
Pisolithus Alb. & Schwein., e Suillus P. Karst.,são considerados como microorganismosfundamentais para a melhora qualitativa dasárvores nos programas de reflorestamento.De Araújo et ai. (6) estudaram o efeito do meiode cultura, do pH inicial, e da concentração desais no crescimento apical de quatro fungosectomicorrízeos, mas pouca informação sobre aprodução de biomassa fúngica e seus metabólitossão encontrados na literatura. Por causa disto,uma técnica não-destrutível foi desenvolvida afim de avaliar a produção de biomassa e a análisede .rnetabólitos tais como pigmentos, enzimas,açúcares, ácidos orgânicos presentes no meioagar.Baar et aI. (7) determinaram a biomassa fúngicadissolvendo o meio agar em água quente efiltrando as soluções antes de secá-Ias à 100°Cpor 24 h e determinando o peso seco de acordocom Oort (8). Como discutido por Jongbloed &

Borst-Pauwels (9), este método pode causar perdados componentes solúveis na água. A perda depeso fúngico seco de L. bicolor foi estimado porJongbloed & Borst-Pauwels (9) deaproximadamente 19%.Gibson & Oeacon (10) descreveram odesenvolvimento micelial de fungosectomicorrízeos in vitro pelo crescimento radial ebiomassa produzida. Jongbloed & Borst-Pauwels(8) fizeram uma hipótese que o crescimento radialreflete na verdade a exploração dos recursospresentes, sendo por outro lado, a biomassaproduzida uma medida do acúmulo de carbono ede nutrientes.° objetivo deste trabalho é apresentar algumasmodificações na técnica empregada por Baar etaI. (7) e analisada por Jongbloed and BorstPauwels (8) a fim de poder quantificar aconcentração de glucose, o pH e a biomassaproduzida após 5, 10 e 15 dias de incubação à25°C para as linhagens de Suillús collinitus ePisosithus tinctorius sobre diferentes meios decultura (POA, BAF , MNM, MP, GM8, MG).Esta técnica não destrutível está sendo utilizadaagora para avaliar a produção de biomassa eanalisar os metabólitos tais como os pigmentos,as enzimas e ácidos orgânicos presentes no agare/ou na biomassa.
MATERIAIS E MÉTODOS
LinhagemMicélio das especies ectomicorrízicas foramobtidos dos corpos frutíferos nas florestas dePinus. Pisolithus tinctorius (Pers.) Coker etCouch (PF 26) foi isolado na região de Murcia
,(Espanha) em 1991 e Suillus collinitus Fr. (Se24) na região da La Grande-Motte (sudoeste daFrança) em 1994.
Meios de cultura° micélio das linhagens foi cultivado e mantidono meio PDA. A composição dos meios decultura utilizados é dada a seguir.Meios de cultura (g/l) i) Meio PDA: 39,0 (pH5.6). 2) Meio BAF: glucose, 30,0; peptona, 2,0;extrato de levedura, 0,2; KH2P04, 0,5;MgS04.7H20, 0,5; citrato férrico (1%), 0,5 ml;myo inositol, 5 ml; Solução de oligoelementos*,
294
1,0 ml; Solução de vitaminas**, 1,0 ml; agar,15,0 (pH= 6,0). 3) Meio MG: glucose, 10,0; agar,15,0 (pH=6,5). 4) Meio MP: gl ucose, 10,0;peptona, 2,0; KH2P04, 0,5; CaCI2., O, I; FeCI3,0,01; Solução de vitaminas**, 1,0 ml; agar, 15,0(pH=7,5) 5) Meio GM8: glucose, 31,25; peptona,2,5; nitrato de cálcio, 2,0; asparagina, 1,0; tartratode amônia, 1,0; agar, 15,0 (pH=5,0). 6) MeioMNM: glucose; 10,0; peptone, 2.0; extrato demalte, 3,0; CaCI2.2H20, 0,2; KH2P04, 0,5;(NH4)2HP04 , 0,5; MgS04.7H20, 0,15; FeCI3,0,01; Thiamine HCI, 100-l!g; Oligoelementsolution*, 1,0 ml; agar, 15,0 (pH=5,0).* Solução de oligoelementos (g/I): CaCI2, 100,0;MnS04, 5,0; ZnS04.7H20, 1.** Solução de vitaminas (mg/l): Tiamina HC1,500; Biotina, 10; Ácido Fólico, 100.
Inoculação e IncubaçãoUm disco de papel celofane, previamente estéril,é depositado sobre o meio de cultura nas placasde Petri de 60 mm de diâmetro. Foram inoculadascentralmente com um bloco de micélio deaproximadamente (3 x 3 x 2 mm) tomado daextremidade de uma colônia de 15 dias de idadesobre o meio POA.As placas de Petri foram vedadas com "Parafilm",As culturas foram conduzidas à 25°C naobscuridade.
Análise°diâmetro da colônia foi medido com uma réguaa intervalos de tempo regulares, até 15 dias. Osresultados são expressos como uma média de 3triplicadas.° tratamento das amostras é efetuado segundo oesquema da Fig. I. A colônia que cresceu sobre opapel celofane é separada e seu peso édeterminado. Todo o meio agar (10 g) éressuspenso em 90 ml de água destilada ehomogeneizado durante 30 segundos no Potter à 4°C.
A biomassa (peso seco) é determinadacolocando-se o disco de celofane contendo omicélio a secar à ]05 "C por 24 horas. O pesodo disco de celofane foi determinado comouma média de ]5 amostras.O açúcarconsumido foi determinado pelo método deMiller (11).
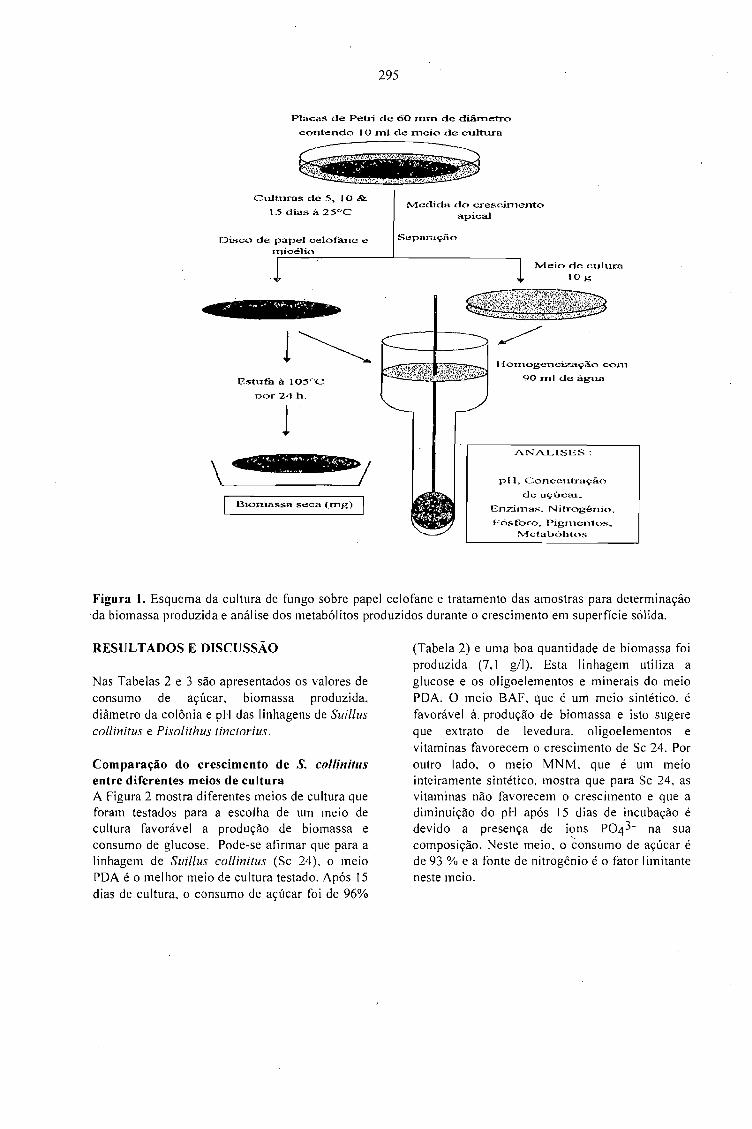
295
Placas de Pett; ele 60 rnrn de diâmetro
contendo 10 1111de Illeio de C'lI.1hua
Cu1t:uras de 5, 10 &
15 di.. :,; à 25"C
IJisco de papel celofane e.Inicélio
"Mcclida do crescj.nlentoapical
SepJlraçOO
l~
Estufa à IOS-'C
Dor 24 h.
1\~
I Bionlassa seca (rng) ,
lvteio de Cllhlra
l'{ornogcTlciz...a.ção 00111
90 rrr I de águ...
AN~"-LISES :
pll, Concellln~çiio
de UÇÚCaI'~
EJ17iI11a.S.. NifT(}gênl0~
Fóstoro,. P'Í,6YTl1cnl<.)s.,."l\,.-feta tu..-:,1í tos
Figura I. Esquema da cultura de fungo sobre papel celofane e tratamento das amostras para determinação'da biomassa produzida e análise dos metabólitos produzidos durante o crescimento em superfície sólida.
RESULTADOS E DISCUSSÃO
Nas Tabelas 2 e 3 são apresentados os valores deconsumo de açúcar. biomassa produzida.diâmetro da colônia e pH das Iinhagens de Suil/uscollinitus e Pisolithus tinctorius.
Com paração do crescimento de S. collinitusentre diferentes meios de culturaA Figura 2 mostra diferentes meios de cultura queforam testados para a escolha de um meio decultura favorável a produção de biomassa e.consumo de glucose. Pode-se afirmar que para alinhagem de Suillus collinitus (Se 24). o meioPDA é o melhor meio de cultura testado. Após 15dias de cultura. o consumo de açúcar foi de 96%
(Tabela 2) e uma boa quantidade de biomassa foiproduzida (7.1 g/l). Esta linhagem utiliza aglucose e os oligoelementos e minerais do meioPDA. O meio BAF. que é um meio sintético. éfavorável à produção de biomassa e isto sugereque extrato de levedura. oligoelementos evitaminas favorecem o crescimento de Se 24. Poroutro lado. o meio MNM. que é um meiointeiramente sintético. mostra que para Se 24. asvitaminas não favorecem o crescimento e que adiminuição do pH após 15 dias de incubação édevido a presença de ions P043- na suacomposição. Neste meio. o consumo de açúcar éde 93 % e a fonte de nitrogênio é o fator limitanteneste meio.
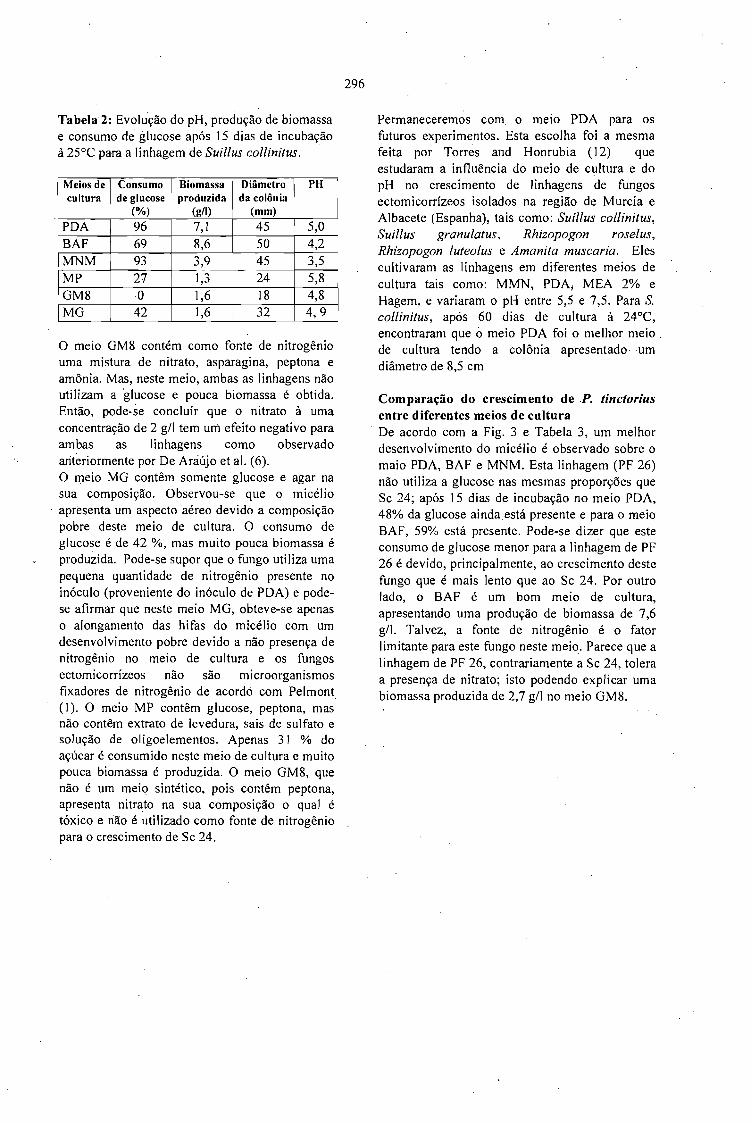
Tabela 2: Evolução do pH, produção de biomassae consumo de glucose após 15 dias de incubaçãoà 25°C para a linhagem de Suillus collinitus.
Meios de Consumo Biomassa Diâmetro PHcultura de glucose produzida da colônia
(%) (g/l) (mm)
PDA 96 7,1 45 5,0BAF 69 8,6 50 4,2MNM 93 3,9 45 3,5MP 27 1,3 24 5,8
IGM8 O 1,6 18 4,8MG 42 1,6 32 4,9
o meio GM8 contém como fonte de nitrogêniouma mistura de nitrato, asparagina, peptona eamônia. Mas, neste meio, ambas as linhagens nãoutilizam a glucose e pouca biomassa é obtida.Então. pode-se concluir que o nitrato à umaconcentração de 2 g/l tem um efeito negativo paraambas as linhagens como observadoanteriormente por De Araújo et al, (6).O meio MG contêm somente glucose e agar nasua composição. Observou-se que o micélíoapresenta um aspecto aéreo devido a composiçãopobre deste meio de cultura. O consumo deglucose é de 42 %, mas muito pouca biomassa éproduzida. Pode-se supor que o fungo utiliza umapequena quantidade de nitrogênio presente noinóculo (proveniente do inóculo de PDA) e podese afirmar que neste meio MG, obteve-se apenaso alongamento das hifas do micélio com umdesenvolvimento pobre devido a não presença denitrogênio no meio de cultura e os fungosectomicorrízeos não são microorganismosfixadores de nitrogênio de acordo com Pelmont(1). O meio MP contêm glucose, peptona, masnão contêm extrato de levedura, sais de sulfato esolução de oligoelementos. Apenas 3 J % doaçúcar é consumido neste meio de cultura e muitopouca biomassa é produzida. O meio GM8, quenão é um meio sintético, pois contêm peptona,apresenta nitrato na sua composição o qual étóxico e não é utilizado como fonte de nitrogêniopara o crescimento de Se 24.
296
Permaneceremos com. o meio PDA para osfuturos experimentos. Esta escolha foi a mesmafeita por Torres and· Honrubia (12) queestudaram a influência do meio de cultura e dopH no crescimento de linhagens de fungosectomicorrízeos isolados na região de Murcia eAlbacete (Espanha), tais como: Suillus collinitus,Suillus granulatus, Rhizopogon roselus,Rhizopogon luteolus e Amanita muscaria. Elescultivaram as linhagens em diferentes meios decultura tais como: MMN, PDA j MEA 2% eHagem, e variaram o pH entre 5,5 e 7,5. Para S.collinitus, após 60 dias de cultura à 24°C,encontraram que 6 meio PDA foi o melhor meio.de cultura tendo a colônia apresentado· -umdiâmetro de 8,5 em
Comparação do crescimento deP. tinctoriusentre diferentes meios de culturaDe acordo com a Fig. 3 e Tabela 3, um melhordesenvolvimento do micélio é observado sobre omaio PDA, BAF e MNM. Esta linhagem (PF 26)não utiliza a glucose nas mesmas proporções queSe 24; após ] 5 dias de incubação no meio PDA,48% da glucose aindaestá presente e para o meioBAF, 59% está presente. Pode-se dizer que esteconsumo de glucose menor para a linhagem de PF26 é devido, principalmente, ao crescimento destefungo que é mais lento que ao Se 24. Por outrolado, o BAF é um bom meio de cultura,apresentando uma produção de biomassa de 7,6g/I, Talvez, a fonte de nitrogênio é o fatorlimitante para este fungo neste meio. Parece que alinhagem de PF 26, contrariamente a Sc 24, toleraa presença de nitrato; isto podendo explicar umabiomassa produzida de 2,7 g/I no meio GM8.
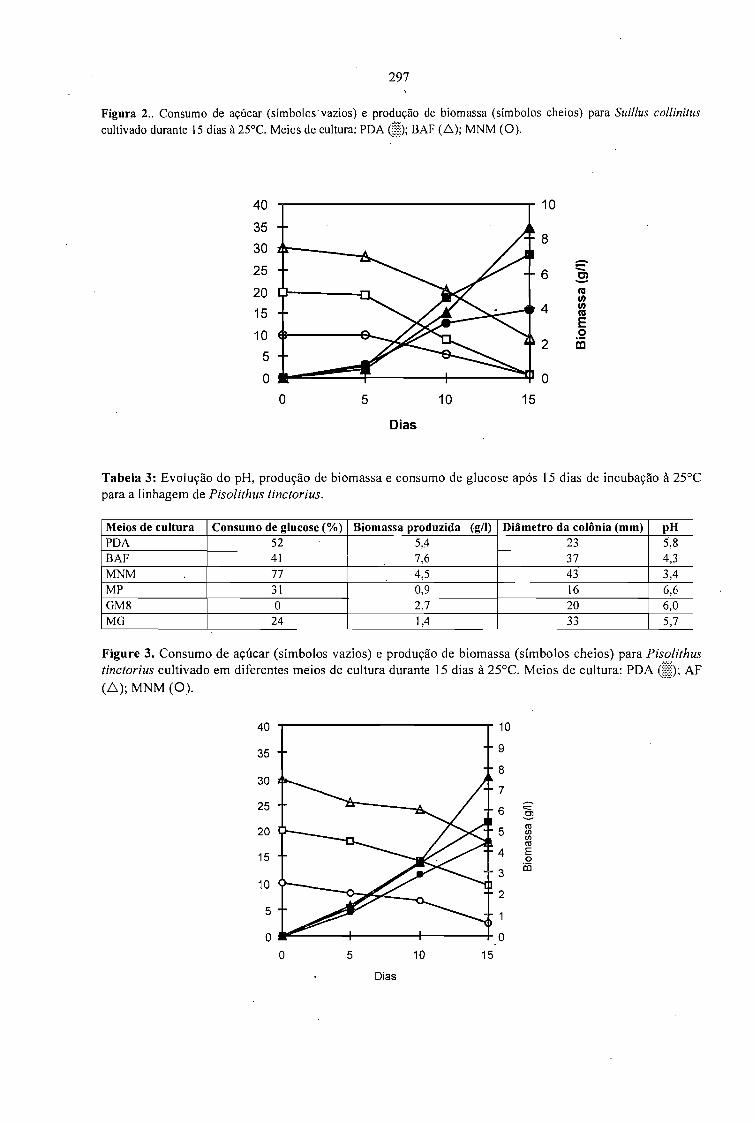
297
Figura 2.. Consumo de açúcar (símbolosvazios) e pr0d..~ção de biomassa (simbolos cheios) para Suillus collinituscultivado durante 15 dias à 25°C. Meios de cultura: PDA <i@); BAF (.0.); MNM (O).
40 10
35
308
-25 6:::S
20 l'lll/l
15 4 l/ll'llE
10 O2 ãi
5
O O
O 5 10 15
Dias
Tabela 3: Evolução do pH, produção de biomassa e consumo de glucose após 15 dias de incubação à 25°Cpara a linhagem de Pisolithus tinctorius.
Meios de cultura Consumo de glucose (%) Biomassa produzida (g/I) Diâmetro da colônia (mm) pHPDA 52 5,4 23 5,8BAF 41 7,6 37 4,3MNM 77 4,5 43 3,4MP 31 0,9 16 6,6GM8 O 2,7 20 6,0MG 24 1,4 33 5,7
Figure 3. Consumo de açúcar (símbolos vazios) e produção de biomassa (símbolos cheios) para Pisolithustinctorius cultivado em diferentes meios de cultura durante 15 dias à 25°C. Meios de cultura: POA GD: AF
(.6.); MNM (O).
40 10
35 9
830
7
25 ~
6 ~20 5 ro
tnsnro
15 4 Eo
3 zs10
25
O O
O 5 10 15
Dias

CONCLUSÃO
Os resultados mostraram que esta técnica para ocrescimento de fungos, análise dos parâmetrostais como pH, consumo de açúcar no meio agarestá bem estabelecida. Os valores obtidos debiomassa produzida, consumo de açúcar e pHpermitem otimizar as condições de cultura eestudar a fisiologia e o metabolismo de fungosmicorrízeos sobre meios de cultura com agar. Agrande vantagem desta técnica está baseada nofato que é um método de tratamento das amostrasnão destrutível, em outras palavras, utilizando-seo homogeneizador e trabalhando à baixastemperaturas, (4°C), todas as enzimas emetabólitos produzidos podem ser detectados equantificados.
AGRADECIMENTOS
Os autores agradecem à Drs. D. Mousain e C.Plasard (Institut National de RechercheAgronomique, Montpel1ier, França) porfornecerem as linhagens de fungosectomicorrízeos. Álvaro A. De Araújo agradeceao Governo Brasileiro (CNPq) por sua bolsa deestudos que foi gerada por SFERE - França(Société Française d'Exportation des ResssourcesEducatives). Esta pesquisa foi financiada pelaComunidade Européia, programa MYCOMEDnOAIR2 - CT 94-1149 e IRD, Dept. DRV ..
REFERENCIAS
I. Pelmont, J. (1993), Bacteries et environnement.1st ed., Presses Universitaires de Grenoble,Grenoble.2. Mousain, D. (1993). Physiologie des arbres etarbustes en zones arides et semi-arides. 1sted.,Editions John Libbey, Eurotext,Montrouge.CEE Project " MYCOMED ",Contract Number AIR 2-CT-94-1149, FinalProgress Report.
298
, .....
3. Strullu, D. G. (1991), Les mycorhizes desarbres et plantes cultivées, 3rd ed., Lavoisier.Paris.4. Marx, D.H. and Corde1J, C.E. (1994), In: J.M.Whipps & R.D. Lumsden (Eds). Biotechnology offungi for improving plant growth. 2nd ed.Cambridge University Press, Cambridge, 1-25.5. Mousain,D., Plassard, C., Roussos, S., Brunel,B., Karkouri, K. E., Fréchet, C.C., Laguette, A.G.,Bouckcim, H., Gobert, A., De Araujo, A.A.(1998) Improvement of the quality of forestseedlings Pinus pinea, Pinus halepensis, Pinusnigra subsp. laricia, Quercus suber) andMEDiterranean reforestation using controlledMYCorrhizal infection (MYCOMED). FinalProgress Raport CEE Contrat na AIR 2-CT-941149.6. De Araujo, A.A., Hannibal, L., P1assard, M.,Mousain,D. and Roussos, S., (l 998), Micol.Neotrop. Apl. 11,23-34.7. Baar, J., Comini, 8., Elferink, M.O., andKuyper, Th. W. (1997) Mycol. Res. 101 (5),523529.8. Oort, A. J. P. (1981), Verhandelingen.Koninklilke Nederlandse akademie vanWetenschappen, Afdeling Natuurwetenschappen,2nd. 76, 1-879. Jongbloed, R.H. and Borst-Pauwels, G. W .F.H.(1990), Acta Bot. Neerl. 39, 349-358.10. Gibson, F. and Deacon, J. W. (1990), Mycol.Res. 94(2),166-172.lI. Miller, G.L. (1959), Anal. Chem. 31,426-428.12. Torres, P. and Honrubia,M. (1991),Cryptogamie Mycol. 12 (3), 183-192.

299
CHARACTERISATION OF Beauveria bassiana AND Metarhizium anisopliae ISOLATES FORPOTENTIAL USE AGAINST THE COFFEE BERRY BORER
Patricia E. Vélez, Maria T. González, Armando Rivera, Alex E. Bustillo, Maria N. Estrada &Esther C. Montoya.
Centra Nacional de Investigaciones de Café, Chinchiná, Caldas, ColombiaInstituto para la Ciencia y Tecnología, Colciencias, Santafé de Bogotá, Colombia.
Key words: Coffee berry borer, Hypothenemus hampei, Beauveria bassiana, Metarhizium anisopliae,Coffee.
ABSTRACTThe e./ficacy of biological control agents depends on the knowledge of their biology, the environment andthe mechanism of interaction between parasite and host. In this study exact number ofmultisporic isolatesof the entomogenous fungus Beauveria bassiana (Bb), were characterised according to: pathogenicityagainst coffee berry borer-CBB, Hypothenemus hampei (Ferrari), LT50, spore production, rate ofmycelial
growth, spore size, germination rate, enzyme reaction and time of enzyme reaction. Similar analysis wascarried out using Metarhizium anisopliae (Ma). In a preliminary analysis, the isolates were classifiedaccording to pathogenicity against CBB, establishing 4 groups. The analysis of variance of the groups(p=0,05) showed significant differences among the isolates in the variables pathogenicity against CBB,LT50' spore production, mycelial growth, germination rate, enzyme reaction & time ofenzyme reaction for
Bb and the variables pathogenicity against CBB and germination rate for Ma; the Tukey test (5%) showeddifferences in Bb & Ma groups, in the variable pathogenicity against CBB. A multivariate principiecomponent analysis was carried out with a group of 17 isolates ofBb in which ali the mentioned variableswere evaluated. Likewise, a dendrogram (descriptive grouping) and a clusterstatistic analysis were done inorder to select groups of isolates according lo the evaluated variables. The components that contributedmost to the total variation were: enzyme reaction, time ofenzyme reaction, pathogenicity against CBB andspore production. In contrast, time of enzyme reaction, LT50, mycelial growth and germination rate,
accounted for a lower degree ofvariation. The dendrogram and the cluster statistical analysis separatedthree groups of isolates. The analysis of variance of the groups classified by cluster analysis showeddifferences between them, the Tukey test (5%) showed that the groups were statistically different in thevariables pathogenicity against CBB, enzyme reaction, spore production, mycelial growth and LT50' The
variable pathogenicity to CBB allowed lhe separation ofgroups of isolates in both type of analysis. Thecharacterisation of the collection isolates in our study has enabled us the selection of those variables thatcontributed most to the separation ofgroups of isolates, to establish relationships between them accordingto their biology and locality, to define their host specificity and their potential to control the CCB and otherinsect pests ofeconomic importance in Colombia and other countries.
INTRODUCTION
The coffee berry borer (CBB), Hypothenemushampei (Ferrari) (Coleoptera: Scolytidae), is themost serious pest of coffee in Colombia and manyother countries. This pest originated in centralAfrica and was accidentally introduced to Brazilat the beginning of the century (Murphy andMoore 1990). Since its arrival in Colombia in1988, the Colombian Coffee Growers Federationand its research centre, CENICAFÉ, havepromoted extensive research into alternativecontrai strategies using an Integrated PestManagement system. Biological contrai withparasitoids and with the entomopathogenic
fungus Beauveria bassiana are promising contraimethods, offering an environmentally benignalternative to chemical pesticides (Cadena 1993).Coleoptera are known to be susceptible to B.bassiana and infection of CBB can occur in twoways. The fungus may have spread with the pestas the latter colonised new geographic areas or itmay have "jumped" from other local insects(Bridge et ai. 1990). lt was of interest to us toassess fungaI isolates from different hosts andgeographic areas to be able to identify variabilityamong them accurately, in order to select the beststrain with adaptive characteristics favouring itssurvival and to optimise its effectiveness as amycoinsecticide against CBB. To accomplish

this, it is necessary to understand its genetics,physiological aspects, virulence factors, etc. Inorder to exploit the potential application of B.bassiana collection against CBB in Colombia, weused morphological and biochemical techniquesto characterise and compare B. bassiana isolatedfrom CBB and other insects in Colombia andother countries.
MATERIALS AND METHODS
Iso/ates tested: We examined 73 isolates of Bband 19 of Ma, from the culture collection held inthe Entomology Oepartment of the CoffeeNational Research Centre in Colombia. Isolateswere grown on Sabouraud Oextrose Agar (SOA)for 15 days at 25 DC and passed once oncomminuted cuticle from adult CBB (Vélez et ai.1997) in order to maintain their infectivity to thetarget host.
Viability: Viability was determined as %germination in culture after 30 days of growth in
SOA. Five aliquots of 5111 (1 x 106 spore/ml)were placed each (n = 10). Plates were incubatedfor 24-48h at 25 DC, one drop of lactophenol bluewas added to each aliquot in order to stop thegermination process and microscopic examinationat 40x of the germinated spore was carried out.The percent germination was computed on thebasis of the number of spore producing germ tubeper 100 counted in each aliquot. A germinatedconidium was defined as one which had produceda germ-tube equal to or greater than the width ofthe conidium (Vélez et ai. 1997).
Pathogenicity: Sixty adult female CBB, dividedinto six groups of ten, were maintainedindividually in vials in a controlled environmentchamber at 27 DC and 80-85 % RH. To determinepathogenicity adult CBB were submerged twominutes in a spore suspension of the fungi (1 x
107 spore/ml sterile distilled H20, I % Tween
80), with constant manual shaking. PretreatedCBB were confined in a 4 ml glass vialcontaining one coffee bean (45 % RH) as theborer substrate, wetted filter paper and cappedwith a piece of sterile cotton and maintained at 23DC and 80 % RH. Additional 60 adults femaleCBB were not infected and were included ascontrol. Over 10 days, mortality and presence ofsporulating fungus (symptoms and signs) on theinsects were recorded using a stereoscopicmicroscope (González et ai. 1993; Vélez et ai.
300
1997). Lethal time fifty (LT50) was measured by
taking into account the time in which theinoculum of the fungus caused 50% of mortalityon the insect population.
Rate of mycelia/ growth: Mycelial growth wasmeasured on colonies growing on SOA. Eachplate was inoculated at the centre with 5111 of
fungal spore suspension (1 x 106 spore/ml) andincubated at room temperature (25 DC) for 5 days.Each treatment was replicated 10 times. Afterincubation, the diameter of the colony up to 30 dwas measured. A mean diameter was calculatedfor each isolate.
Spore Size: Spore diameter was determined usinga compound light microscope fitted with a ocularmicrometer. For each isolate, mean diameter wascalculated based on measurements of 100 spores.
Spore production: We quantified sporeproduction in ali of the isolates by enumeratingspores from 8, 15 21 and 30 day old culturesgrown on SOA. Spores from these cultures weresuspended in 5 ml of 1% Tween 80, diluted(1/10), and spores counted using ahaemocytometer. Ten replications for treatmentand six counts for replication were recorded(Vélez et ai. 1997).
Enzyme reaction: Production and acnvity ofextracelluJar enzymes were screened through thetechnique that involved incorporating particularsubstrates into modified agar growth media;degradation of the substrate was then detectedafter growth either by 'a colour change (pHindicator), a reduction in opacity, or liquefaction.The enzyme systems tested using this methodwere esterase, gelatinase, caseinase, lipase. Thenitrogen sources evaluated were: urea, peptoneand yeast extract and the carbon sources: citrate,starch, lactose, lanolin, N-acetyJ glucosamine andglucosamine (Paterson & Bridge, 1994; Mugnaiet ai. 1989). Six plates were set up for eachtreatment. Biochemical test results were analysedusing the percent of plates that gave some changeof colour. The time at which substrate utilisationwas visible by colour change, reduction in opacityor liquefaction was assessed for each isolate.
Statistica/ ana/ysis: Isolates of Bb and Ma wereclassified according to pathogenicity againstCBB, establishing 4 groups: group I,pathogenicity <25%; group 2, 26% - 50%; group

3, 51% - 80%, and group 4, >80% (Table I).Variance analysis of the groups and Tukey test(p=0,05) were carried out.A multivariate principIe component analysis wascarried out with a group of 17 isolates of Bb inwhich ali the following variables were evaluated:pathogenicity against CBB, LTSO, germination
rate, enzyme reaction, time of enzyme reaction,spore production, mycelial growth and spore size.Likewise, a dendrogram and cluster statisticanalysis were performed in order to select groupsof isolates according to the evaluated variables,variance analysis of the clusters (p=O,OS) andTukey test (S%) were applied to the groups ofisolates.
RESULTS AND DISCUSSION
Analysis 01 Pathogenicity Beauveria bassiana:In group I, were 6 isolates from Colombia, fourof them from Antioquia; one from Thailand andone from Philippines. Among the most viruJentisolates, were isolates from different insectfamilies i. e. from Coleoptera, Scolytidae; fromColeoptera: Curcul ionidae; from Coleoptera:Scarabaeidae and from Homoptera; 2 fromunknown origin and I from a commercialformulation. 17 of these virulent isolates wereobtained from CoJombia, four from Antioquiaand four from Risaralda, two of them of originunknown and the other one from other countries.An analysis of variance of pathogenicity againstCBB, LTSO, spore production, myceJial growth,
germination rate, enzyme reaction and time ofenzyme reaction for Bb and the variablespathogenicity against CBB and germination ratefor Ma, showed differences in Bb & Ma groups inthe variabJe pathogenicity against CBB (Tukeytest p=O.OS) (Tables I & 2)The results show a direct positive relationshipbetween pathogenicity against CBB andgermination rate for the most pathogenic isolatesof Bb (Table I). The group 4 showed lowervalues for LTSO. It is remarkable that there were
no differences for this variable between groups 3& 4, which suggests that isolates withpathogenicities higher than SO%, do not differ inLTSO, while, pathogenicities lower than SO%
(groups 1 & 2), do show strong differences inLTSO (Table I).
The results show a positive relationship betweenenzyme reaction and spore production (group 3).That relationship was maintained in groups 1 &
301
2. The lowest values were found in the isolates ofgroup 4.The most pathogenic group of isolates of Bb(group 4) was composed of 24 isolates, ISbelonging to the Coleoptera. From this group, IIwere isolated from the Scolytidae. Three from theCurculionidae and one from the Scarabaeidae. Inaddition, this large group included five isolatesfrom Lepidoptera, two from the Pyralidae, andone each from the Stenomidae, Geometridae andCossidae. Regarding the geographical origin ofBb isolates, 17 carne from Colombia, five fromother countries: China, Thailand, Ecuador andItaly. Two ofthem were ofunknown originoIn the Bb Colombian isolates of this group werefour from Antioquia and four from Risaralda; thiscould be a reisolation of the same fungaI strainwithin a specific region with similar climaticconditions i.e, from 19.2 to 22.1 °C, from 1200 toIS90 m and 74 to 86% RH; the isolates werefound most frequently in coffee plantations(Posada & Vélez, 1998). Germination rates of theisolates of this group were between 75 & 92%with values higher than 80% in most of theisolates; the enzyme reactions showed averagesbetween 41 & 7S%, most ofthem between SO and60%.On the other hand, isolates of Bb with < 2S%(group 1) pathogenicity for CBB were composedof eight isolates, five from CBB. Additionally, inthis group, isolates from Lepidoptera andHomoptera were found. They were isolated inColombia (6), four in Antioquia (three of themfrom CBB) and one in Risaralda. Two were fromThailand and Philippines. The isolates ofBb fromthis group showed an average pathogenicityof14%, with a minimum value of S and maximumof 2S%. The average germination rate was 71%and ranged between 20 & 89 and the averageenzyme reaction was 75% and ranged between 43and 100%. In this group of Bb, no relationshipwas observed between pathogenicity andgermination rate (Table I).Regarding the weather conditions such astemperature, relative humidity and altitude whereisolates of this group were collected, observedday mean temperatures were between 19.2 to21.6°C, RH between 74 to 86% and altitude,ranged between 1200 and 1S00 m. Additionally,most of them were found in coffee plantations(Posada & Vélez, 1998).With respect to the relationship between host andlocality, the results suggest a certain specificityfor the CBB within the most pathogenic isolates,as 61% of the isolates carne from Coleoptera, and

43% of these carne from CBB. This is inagreement with Prior (1992), bearing in mind thatvirulence may be higher when the isolates comefrorn the same host insect. However, the CBBisolates in the least pathogenic group of isolates(62.5%), did not show any relationship withvirulence against CBB. So, these results do notallow us to infer specificity in those isolates.Another and contrasting possibility is that thepathogenicity of the group 1 isolates could beexplained on coevolution criteria, which suggestsan equilibrium between host and pathogen over along period (Prior, 1992). Thus the most virulentpathogen could be from hosts other than CBB(Hokkanen & Pimentel,1984; Waage, 1990). Inpathogenicity groups 2 & 3, it was observed that30.3% & 62.5% of isolates, respectively carnefrom CBB and they did not show a high virulencefor CBB. Isolates frorn Lepidoptera andHomoptera showed higher leveis of pathogenicityfor CBB (group 4) (Table 1).According to these criteria, it is suggested that thesearch for entomogenous fungi should beconducted in different places frorn that where theinsect pestis found (Hokkanen & Pimentel, 1984;Waage, 1990, Prior, 1992 ; Hajek & St. Leger,1994).
Metarhizium anisopliae: Two pathogenicitygroups were established for Ma. Most of theisolates killed more than 80% in the tests: 9 fromColombia, 3 from Antioquia (Coleoptera)(Table 2).The results showed a direct relationship betweenenzyme reaction and germination rate, so thegroup 3 showed the higher averages; however,this trend was not maintained in group 4 isolates.There was no evident relationship betweenpathogenicity and germination rate in the groupsof Ma (Table 2).The most pathogenic Ma isolates (group 4)consisted of 18 isolates, 9 from Coleoptera,including six from Scarabaeidae, one fromCurculionidae and one from Scolytidae.Additionally, the group contained four isolatesfrom Homoptera and one from Hymenoptera. Maisolates carne from, Colombia (9), USA (1),Australia (3), Brazil (1), Belize (2), New Zealand(1) and one isolate of unknown origino In theColombian isolates, were remarkable three fromAntioquia Department (Coleoptera), one fromNarifio, one from Tolima and two from Caquetá.Since the majority of Ma isolates belonged toColeoptera: Scarabaeidae, it could be assumedcertain specificity toward insects ofthis family.
302
Mu/tivariate analysis: Taking into account thevariables tested in the Bb isolates, themultivariate analysis showed that the componentsthat contributed most to the total variation were:enzyme reaction, time of enzyme reaction,pathogenicity against CBB and spore production,in contrast, time of enzyme reaction, LT50,
myceliaI growth and germination rate, accountedfor a lower degree ofvariation.The dendrogram separated 3 groups of isolates ina descriptive way (Figure 1). For the clusterstatistical analysis the isolates were grouped, sothat those with pathogenicity against CBB rankedbetween 53 & 80% were assigned to cluster 1;isolates with pathogenicity against CBB >80%, incluster 2 and finally, those with pathogenicityagainst CBB· <53,3 % in cluster 3. The analysisshowed three groups of them: six isolates ingroup 1; six isolates in group 2, and five isolatesin group 3. Bb 9305 were not in the same groupof the dendrogram (Table 3). The analysis ofvariance of the groups classified by c1usteranalysis showed differences between them, theTukey test (5%) showed that the groups werestatistically different in ali the evaluated variables(Table 3).From the results, the variable % of pathogenicityto the CBB, used as classification criteria in thepreliminary analysis, was one of the componentswhich had a major contribution to the totalvariation, according to multivariate analysis.Additionally this variable and the mycelialgrowth allowed the separation of differentclusters in the statistical analysis.It was notable that cluster 2, with the highestpathogenicities, had the lowest average of sporeproduction and enzyme reaction. Additionally,that cluster showed the lowest averages of LT50
(Table 3).The highest averages of pathogenicity againstCBB, mycelial growth, germination rate and timeof enzyme reaction were showed by isolates fromcluster 2, evidencing a direct relationship betweenthose variables. However, in this cluster thevariables LT50, spore production and enzyme
reaction presented the lowest averages. Thepathogenicity against CBB and enzyme reactiondid not match with the reports in which enzymeactivity is related to the virulence against the pest(St. Leger et al. 1988; Bidochka andKhachatourians, 1990). Likewise, previousexperiments have shown there is no relationshipbetween pathogenicity against CBB and sporeproduction (Narváez, 1996) (Table 3).
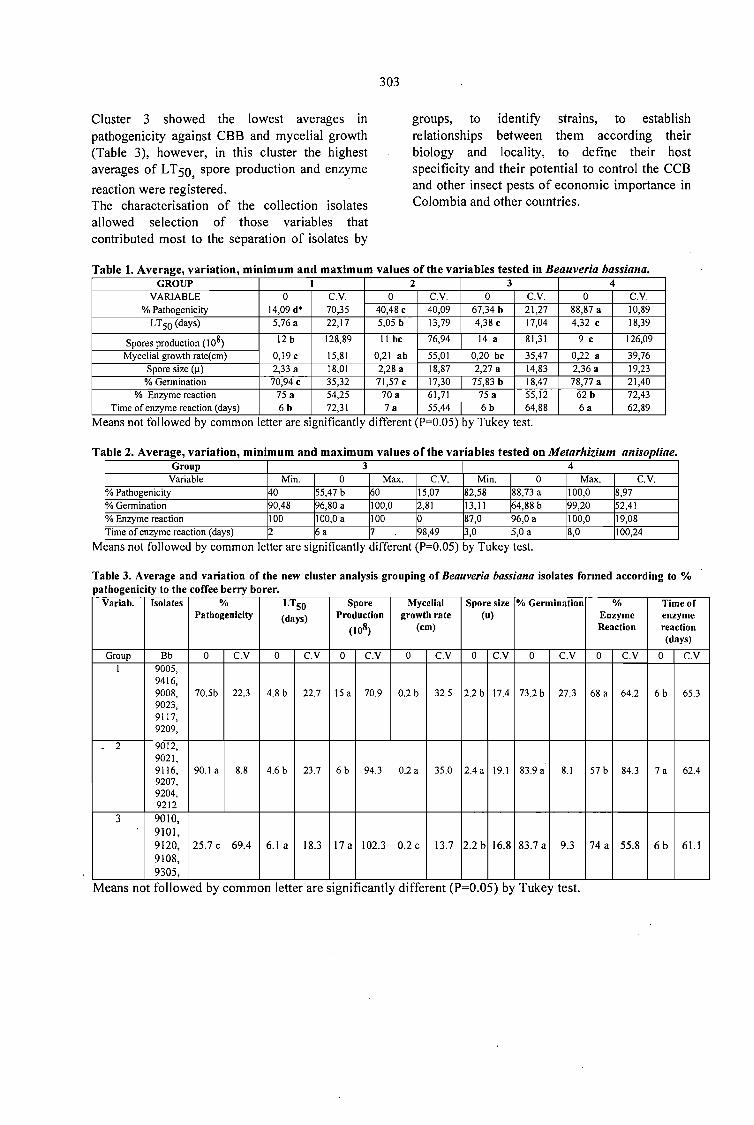
Cluster 3 showed the lowest averages inpathogenicity against CBB and mycelial growth(Table 3), however, in this cluster the highestaverages of LT50 spore production and enzyme, .reaction were registered.The characterisation of the collection isolatesallowed selection of those variables thatcontributed most to the separation of isolates by
303
groups, to identify strains, to establishrelationships between them according theirbiology and locality, to define their hostspecificity and their potential to contrai the CCBand other insect pests of economic importance inColombia and other countries.
Table 1. Average, variation, minimum and maximum values ofthe variables tested in Beauveria bassiana.GROUP 1 2 3 4
VARIABLE ° c.v. ° C.V. ° C.V. ° C.V.% Pathogenieity 14,09 d* 70,35 40,48 e 40,09 67,34 b 21,27 88,87 a 10,89
LT50 (days) 5,76 a 22,17 5,05 b 13,79 4,38 e 17,04 4,32 e 18,39
Spores produetion (108) 12 b 128,89 1i be 76,94 14 a 81,31 9 e 126,09
Myeelial growth rate(em) 0,19 e 15,81 0,21 ab 55,01 0,20 be 35,47 0,22 a 39,76Spore size (u) 2,33 a 18,01 2,28 a 18,87 2,27 a 14,83 2,36 a 19,23% Germination 70,94 e 35,32 71,57 e 17,30 75,83 b 18,47 78,77 a 21,40
% Enzyme reaetion 75 a 54,25 70 a 61,71 75 a 55,12 62 b 72,43Time ofenzyme reaetion (days) 6b 72,31 7a 55,44 6b 64,88 6a 62,89
Means not followed by common letter are significantly different (P=O.OS) by Tukey test.
Table 2. Average, variation, minimum and maximum values ofthe variables tested on Metarhizium anisopliae.Group 3 4
Variable Min. ° Max. C.V. Min. ° Max. c.v.Yo Pathogenieity ~O ~5,47 b ~O 15,07 82,58 88,73 a 100,0 8,97Yo Germination 190,48 f.)6,80 a 100,0 ~,81 13,11 64,88 b 99,20 52,41Yo Enzyme reaetion 100 100,0 a 100 ° 87,0 96,0 a 100,0 19,08Time of enzyrnereaction (days) ~ ~a 7 98,49 3,0 5,0 a 8,0 100,24
Means not followed by common letter are signifícantly different (P=O.OS) by Tukey test.
Table 3. Average and variation of the new c1uster analysis grouping of Beauveria bassiana isolates formed according to %pathogenicity to the coffee berry borer.
Variab. Isolates % LTSO Spore Myeelial Spore size % Germination % TimeofPathogenicity (days) Produetion growth rate (u) Enzyme enzyme
(108) (em) Reaetion reaetion(days)
Group Bb ° C.V ° C.V ° C.V ° C.V ° C.V ° C.V ° C.V ° C.VI 9005,
9416,9008, 70.5b 22.3 4.8 b 22.7 15 a 70.9 0.2 b 32.5 2.2 b 17.4 73.2 b 27.3 68 a 64.2 6b 65.39023,9117,9209,
2 9012,9021,9116, 90.1 a 8.8 4.6 b 23.7 6b 94.3 0.2 a 35.0 2.4 a 19.1 83.9 a 8.1 57 b 84.3 7a 62.49207.9204,9212
3 9010,9101,9120, 25.7 e 69.4 6.1 a 18.3 17 a 102.3 0.2 c 13.7 2.2 b 16.8 83.7 a 9.3 74 a 55.8 6b 61.19108,9305,
Means not followed by common letter are significantly different (P=0.05) by Tukey test.

REFERENCES
BI])OCHKA, M. J. & KHACHATOURIANS; G.G. (1990). Identification of Beauveria bassianaextracellular protease as a virulence factor inpathogenicity toward the migratory grasshopper,Melanoplus sanguinipes, Journal of InvertebratePathology 56,362 - 370.BRIDGE, P. D.; ABRAHAM, Y. J.; CORNISH,M.: C.; PRIOR, C. Se MOORE, D. (1990). Thechemotaxonorny of Beauveria bassiana(Deuteromycotina: Hyphomycetes) isolates fromthe coffee berry borer Hypothenemus hampei(Coleoptera: Scolytidae), Mycopathologia 111,85-90.CADENA G.,G. Políticas de la FederaciónNacional de Cafeteros de Colombia para elcontrol de la broca dei café. In: CONGRESO dela Sociedad Colombiana de Entomología, 20. Cali(Colombia), Julio 13 - 16, 1993. Resúmenes. Cali(Colombia), SOCOLEN, 1993. P. 110. Esp.GONZÁLEZ, M. T; POSADA, F. J., &BUSTILLO, A. E. (1993). Desarrollo de unbioensayo para evaluar la patogenicidad deBeauveria bassiana sobre Hypothenemus hampei,Cenicafé 44, 93-102.HAJEK, A. E.; ST. LEGER, R. J. (1994).Interactions between fungai pathogens and insecthosts, Annual Review of Entomology. 39, 293322.HOKKANEN, H. M. T., PIMENTEL, D. (I 984).New approach for selecting biological control
. agents, Cariadian Entomologisi 116, 1109 - 1121.MUGNAI, L.; BRIDGE, P. D. & EVANS, H. C.(1989). A chemotaxonomic evaluation of thegenus Beauveria, Mycological Research 92, 199209.MURPHY, S.; MOORE, D. (1990). Biologicalcontrol ofthe coffee berry borer, Hypothenemushampei (Ferrari) ( Coleoptera, Scolytidae):previous programmes and possibilities for thefuture. Biocontrol News and Information.11(2):107-117..
304
NARVÁEZ, M dei P. (1996). Estimación de lac.antidad de esporas producidas por losaislamientos patogénicos e los hongos Beauveriabassiana y Metarhizium anisopliae cultivados enarroz y sobre la broca deI café. Manizales(Colombia), Universidad Católica de Manizales.Facultad de Bacteriología, 66 p. 47 Refs. (TesisBacterióloga y Laboratorista Clínica).PATERSON, R. R. M. & BRIDGE, P. D. (1994).Biochemical techniques for jilamentous fungi.IMI Technical Handbooks: No 1. CABInternational. Surrey. Wallingford .
. POSADA, F. J & VÉLEZ, P. E. (1998). Registrode hospedantes y aislamientos de Beauveriabassiana en la colección de hongosentomopatógenos de Cenicafé, Colombia, ManejoIntegrado de Plagas No. 46 p. 50 - 64, 1997.PRIOR, C. (1992). Discovery andcharacterisation of fungal pathogens for locustand grasshopper control. In: Biological Control ofLocusts and Grasshoppers. International Instituteof Biologicai Control. (Lomer, C. J.; Prior, C.Eds.). CAB International, Melksham, UK. p. 230- 238.ST. LEGER, R. J., DURRANDS, P. K.,CHARNLEY, A. K., & COOPER, R. M. (1988).Role of extracellular chymoelastase in thevirulence of Metarhizium anisoplia.e for Manducasexta, Journal ofInvertebrate Pathology 52, 285 292.VELEZ, P. E.; POSADA, F. J.; MARIN, P.;GONZALEZ, M. T.; OSORIO, E.; BUSTILLO,A. E. (1997). Técnicas para el control de calidadde formulaciones de hongos entomopatógenos.Boletín Técnico Cenicafé No.17. 34p.WAAGE, J. (1990). Ecological theory and theselection of biological control agents. In: Criticaiissues in biological controlo (Mackauer, M.,Ehler, L.E: & Roland, J. Eds.). Intercept, UK, p.135 - 157.

305
PRIMEIRA INTRODUÇÃO NO BRASIL DO ENDOPARASITÓIDE Phymastichus coffea(LASALLE) (HYMENOPTERA: EULOPHIDAE), PARA O CONTROLE BIOLÓGICO DABROCA-DO-CAFÉ Hypothenemus hampei (FERRARI) (COLEOPTERA: SCOLYTIDAE).
Fernando Cantor', Evaldo F. Vilela' & Alex E. Bustlllo''
'Depto. Biologia Animal/UFV, Viçosa, MG, 36571-000,2Centro Nacional de Investigaciones dei Café, CENICAFE. Chinchiná(Caldas), Colômbia.
INTRODUÇÃO
São vários os parasitóides que controlam a brocado-café, principalmente na África, de onde doisdeles foram introduzidos no continenteamericano, visando ao controle biológicopopulacional dessa praga. São os casos do Méxicoe da Colômbia, com a introdução da vespa deUganda Prorops nasuta Waterston (Hymenoptera:Bethylidae), e da vespa da Costa do MarfimCephalonomia stephanoderis Betrem(Hymenoptera: Bethylidae).O Brasil iniciou, nos anos 20, um programa decontrole biológico da broca, com a introdução davespa de Uganda, no Estado de São Paulo. Esteprograma, no entanto, foi descontínuado emfunção da popularização dos inseticidas sintéticos,prejudiciais aos inimigos naturais (Bergamin,1943). Na década de oitenta, retomou-se ointeresse pelo controle biológico da broca-docafé, com a nova introdução da vespa da Costa doMarfim, realizada pela antiga Empresa Capixabade Pesquisa Agropecuária - EMCAPA (Linhares,ES). No entanto, como no caso de P. nasuta, osresultados não têm sido contundentes no controleda broca.
IMPORT ÂNCIA
Recentemente, foi constatado pela primeira vez,na África, o parasitóide Phymastichus coffeaLaSalle (Hymenoptera: Eulophidae), identificadocomo um novo gênero e espécie com potencialbiológico para o controle da broca-do-café(LaSalle, 1990).Este parasitóide é originário de regiões próximasao local de origem da praga, e se diferencia dosdemais por atacar fêmeas da broca-do-café emidade adulta e, também, porque seus ovos sãocolocados no interior do inseto hospedeiroComo reportado por Gutierrez et ai. (1998), umcontrole mais eficaz da broca-do-café poderia serconseguido com o uso deste novo parasitóide,obtendo-se, para isto, as informações quepermitirão estabelecer as potencialidades desteinimigo natural, nas diferentes regiões produtoras
de café, considerando, ainda, a eficiência dosdemais parasitóides presentes nos cafezais.Com esta recente descoberta do P. co./fea, abre-seuma nova possibilidade de controle desta pragautilizando inimigos naturais, para todos as regiõesdo mundo onde H hampei constitui-se num sérioproblema.
ACORDO BILATERAL ENTRE CENICAFE(Colômbia) e BIOAGRO/UFV (Brasil)
P. coffea está sendo multiplicado nos laboratóriosdo CENICAFE, na Colômbia e, por meio de umAcordo Bilateral de Cooperação TécnicoCientífico, envolvendo o CENICAFE e aUniversidade Federal de Viçosa (BIOAGRO),está sendo avaliada a introdução deste parasitóideno Brasil. Para isto, estudos de biologia, ecologiae comportamento de P. co./fea estão sendoconduzidos, em laboratório, dentro do Acordo,tendo sido tentada uma introdução do referidoparasitóide, via Laboratório Quarentenário "CostaLima"/Embrapa, porém sem sucesso.
REFERÊNCIAS
Bergamin, J. 1943. Contribuição para oconhecimento da biologia da broca-docafé Hypothenemus hampei (Ferrari,1867) (Col. Ipidae). Arquivos do InstitutoBiológico. 14,31-72
Gutierrez, A.P., A. Villacorta, J.R. Cure, &C.K. Ellis. 1998. Tritrophicanalysis ofthe coffee tCoffea arabica) - Coffee berryborer [Hypothenemus hampei (Ferrari)] [parasitoid system. An. Soe, Entomol.Brasil. 27, 3357-386.
LaSalle, J. 1990. A new genus and species ofTetrastichinae (Hymenoptera:Eulophidae) parasitic on the coffee berryborer, Hypothenemus hampei (Ferrrari)(Coleoptera: Scolytidae). BulI. Entomol.Res. 80, 7-10.


307
EXTRACCION Y EVALUACION DE AMILASAS DIGESTIVAS DE LA BROCA DEL CAFÉHypothenemus hampei (Ferrari) (COLEOPTERA: SCOLYTIDAE)
Claudia P. Martínez Díaz. Arnubio Valencia Jiménez. 2María Teresa González G.IAlex E. Bustillo Pardey. 1
1 Centro Nacional de Investigaciones de Café, Cenicafé. AA N°2427. Manizales-Colombia.2 Departamento de Química, Facultad de Ciencias Exactas y Naturales. Universidad de Caldas-ColombiaE-mail: [email protected]
Palabras claves: Hypothenemus hampei, Broca café, café, enzimas
RESUMOEn el tracto digestivo de los insectos, se encuentran presentes las amilasas, las cuales participanactivamente en la digestión dei almidón; con miras a suministrar la glucosa necesaria para el metabolismoenergético dei insecto. Un conocimiento deta!lado de la actividad amilolítica existente en H. hampei, seconstituye en una necesidad apremiante, en especial cuando se requiere seleccionar inhibidores específicosque puedan ser utilizados para la producción de café transgénico resistente ai insecto. Con miras a conocerel perfil isoenzimático, el pH y temperatura óptima para la actividad de la enzima, se procedió ahomogeneizar 0,5 g de adultos con 2,5 ml de las diferentes soluciones de extracción: H20, NaCI 1% ysoluciones amortiguadoras de pH (3,Oa 8,O). EI pH óptimo se analizó utilizando buflers de pH 3,Oa 9,O. Latemperatura óptima de actividad de la enzima se estimó a temperaturas entre 25 y 70°e. La actividadenzimática se determinó espectrofotométricamente a 580 nm. EI análisis isoenzimático se !levo a cabo porelectroforesis en geles nativos dei 10%. Los resultados encontrados, mostraron que el mejor medio deextracción fue NaCI ai 1%, el pH óptimo de actividad es de 5,0 y la temperatura óptima se presenta a40°e. En H. hampei se encontraron varias bandas de actividad, dos de las cuales ai parecer son enzimasmayoritarias.
INTRODUCCION
En los estudios dirigidos a combati r y proteger loscultivos de los insectos que atacan a las plantas,es importante tener un mejor conocimiento sobrela bioquímica de los insectos y en particular sobrelas enzimas más importantes involucradas en losprocesos digestivos, de forma que conociendoestas enzimas, pueda disefíarse una estrategia quepermita bloquear o interrumpir su función, esdecir, producir una )nhibición dirigidaespecíficamente contra esta enzima (Blanco etal., 1996).El metabolismo digestivo de los insectos seasemeja ai de los vertebrados. Como estos,poseen enzimas adaptadas a la dieta de la que sealimentan. Si viven de una dieta particularmenterica en una sustancia, generalmente produciránabundantes enzimas para su digestión (ej.amilasas para dietas ricas en carbohidratos), y sipor el contrario, la dieta es altamente restringida,las enzimas presentes serán correspondientementelimitadas (Applebaum,1985). La mayor parte delalimento requerido para la nutrición de losinsectos consiste de polímeros como el almidón,celulosa, hemicelulosa y proteínas. EI proceso
digestivo de éstos ocurre en tres fases, durante lafase inicial, se lleva a cabo la disminución en elpeso molecular de compuestos poliméricosmediante la acción de hidrolasas poliméricas(amilasas, celulasas, hemicelulasas yendoproteasas); los oligomeros resultantes sonhidrolizados en la etapa intermedia por hidrolasasoligoméricas, que actúan sobre fragmentosparcialmente degradados. Los produetos de estafase son dímeros o pequefíos oligómeros talescomo mal tosa, celobiosa y dipéptidos. En la etapafinal de la digestión, los dímeros se rompen enmonómeros por hidrolasas como maltasa,celobiasa, dipeptidasas, tripeptidasas,aminopeptidasas y carboxipeptidasas las cuales sehallan generalmente localizadas en la membranaepitelial dei intestino (Terra, 1990). Es muyprobable que las carbohidrasas digestivas de losinsectos hidrolicen poli, oligo y disacaridos a suscorrespondientes monosacáridos para prepararlospara su absorción de manera similar a lascarbohidrasas de vertebrados. En estos últimos,las hidrolasas poliméricas son secretadasprincipalmente por las glándulas salivales y elepitelio dei intestino medio. Se reconocen doscategorías de carbohidrasas, las cuales se

denominan comúnmente como u-amilasas y 13amilasas. Las a-amilasas (1,4-a-D-glucanglucanohidrolasas), se caracterizan por atacarenlaces glicosídicos internos de almidón oglicógeno. Se considera que el ataque (a-l,4)produce rompimientos aI azar que generan unamezcla de dextrinas de longitudes de cadenavariable, aunque algunas evidencias sefíalan quela hidrólisis deI almidón sigue un cierto orden(Bohinski, 1983).Las amilasas de los insectos juegan un papel muyimportante en la digestión deI almidón presenteen la semil1a. EI orden Coleoptera esextremadamente grande, en consecuencia esdifícil hacer generalización acerca de ladistribución de las enzimas digestivas de estos ode un insecto en particular. La digestión deIalmidón por amilasas de insectos ha sidodemostrada en S. granarius (Silano et al., 1975;Baker, 1983); Tribolium castaneum (Krishna ySaxena,1962); Trogoderma spp. (Krishna, 1955);Rhyzopertha dominica (Baker, 1991); Sitophilluszeamais (Baker, 1983; Sandoval-Cardoso, 1991);Sitophillus orizae (L) (Baker, 1987; Yetter et al.,1979); Callosobruchus maculatus (Campos et ai.,1989); Zabrotes subfasciatus (Lemos et al., 1990)y Tenebrio molitor (Applebaum, 1964). Un mejorentendimiento de las enzimas involucradas en lautilización deI alimento por el insecto, y de susprincipales propiedades, permitirá un mayorentendimiento deI papel que desempefían en losmecanismos de resistencia deI insecto frente apotenciales inhibidores. En el presente trabajo sedescriben varias propiedades de amilasasencontradas en adultos de Hyphotenemus hampei.
MATERIALES Y MÉTODOS:
Material Biológico: Las brocas procedían de launidad de cría de parasitoides de Cenicafé(Chinchiná-Caldas), recién emergidas, criadas encafé pergamino seco con una humedad deI 45%, ymantenidas a temperatura de 27°C y humedadrelativa deI 65% - 75%.
Determinación de la actividad amilasa: Laactividad amilasa se determinó por el método deHopkins y Bird, 1954. 50 111 de solución de laenzima se incubó por 15 minutos a 30°C con 650111 de buffer citrato 0.05M pH 5,0 conteniendo 10mM de NaCI y 20 mM de CaCI2 y 500 111 de unasolución 0.125 % de almidón (Almidón soJuble,Sigma Chemical Co. ) en 50 mM de buffer citratopH 5,0. La reacción se detuvo tras la adición de 5
308
ml de solución de Iodo (I: 0.5 % YKI: 5 %). Laabsorbancia de las muestras fue leída a 580 nmutilizando para ello un espectrofotómetro UVNISmarca UNICAM.
Extracción de la enzima: La enzima fueextraída, homogeneizando 0,5 gramos de brocasadultas con 2,5 ml de una solución NaCI aI 1%.EI homogeneizado se centrifugó a 1000 x gdurante 30 minutos y el sobrenadanteresultante fue almacenado a -20°C como fuentede la enzima. Todas las etapas se realizaron a4°C.
Evaluación dei pH óptimo: EI pH óptimo de lasamilasas procedentes de H. hampei, fuedeterminado con soluciones amortiguadoras0.05M conteniendo 20 mM de CaCI2 y 10mM deNaCI . Los buffers utilizados fueron: buffercitrato pH 3,0 Y 5,0; buffer succínico pH 4,0 y6,0; buffer fosfato pH 7,0; buffer tris-HCI pH8,0 Ybuffer borato pH 9,0.
Prueba de estabilidad térmica: La estabilidadtérmica de las amilasas fue determinada,preincubando la solución de la enzima en bufferde actividad durante 5 minutos a 25, 30, 40, 50,60 y 70°C. Se tomaron cinco repeticiones de cadatratamiento.
Perfil isoenzimático: La actividad amilolítica sedetectó en zimogramas in situ empleando elmétodo de Campos et ai. (1989). Muestras de laenzima se corrieron en geles nativos depoliacrilamida (PAGE) ai 10%, utilizando unacelda electrofóretica (Bio-Rad laboratories,Modelo Miniprotean 11). En los geles se adicionóalmidón coopolimerizado a una concentraciónfinal dei 0,5%. La electroforesis fue corrida a 100voltios y a 4°C. Una vez finalizada laelectroforesis, el gel se lavó con agua y se dejó enbuffer de actividad por 2 horas a 30°C. Lasbandas de actividad fueron detectadas porcoloración deI gel durante 15-20 minutos, con unasolución de Iodo. Las zonas de actividad aparecencomo bandas claras sobre un fondo azul oscuro.Finalmente el gel fue fotografiado y la imagendigitalizada.
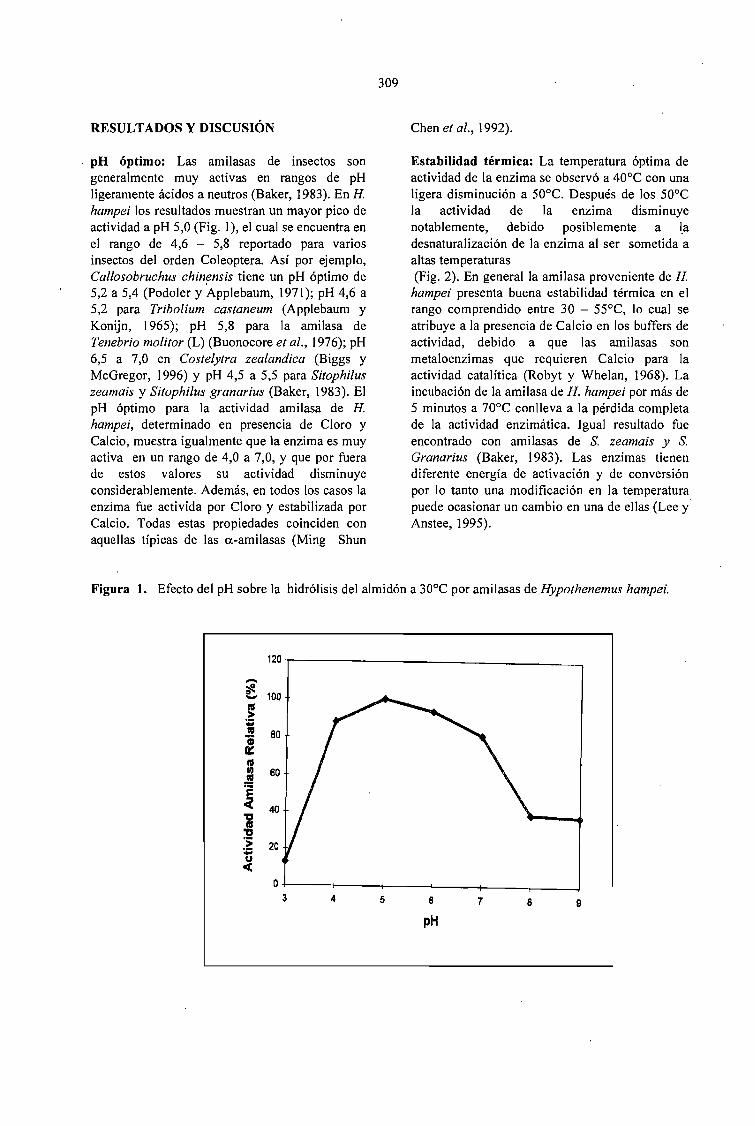
RESULTADOS Y DISCUSIÓN
pU óptimo: Las amilasas de insectos songeneralmente muy activas en rangos de pHligeramente ácidos a neutros (Baker, 1983). En Hhampei los resultados muestran un mayor pico deactividad a pH 5,0 (Fig. I), el cual se encuentra enel rango de 4,6 - 5,8 reportado para variasinsectos deI orden Coleoptera. Así por ejemplo,Callosobruchus chinensis tiene un pH óptimo de5,2 a 5,4 (Podoler y Applebaum, 1971); pH 4,6 a5,2 para Tribolium castaneum (Applebaum yKonijn, 1965); pH 5,8 para la amilasa deTenebrio molitor (L) (Buonocore et ai., 1976); pH6,5 a 7,0 en Costelytra zealandica (Biggs yMcGregor, 1996) y pH 4,5 a 5,5 para Sitophiluszeamais y Sitophilus granarius (Baker, 1983). EIpH óptimo para la actividad amiJasa de Hhampei, determinado en presencia de Cloro yCalcio, muestra igualmente que la enzima es muyactiva en un rango de 4,0 a 7,0, y que por fuerade estos valores su actividad disminuyeconsiderablemente. Además, en todos los casos laenzima fue activida por Cloro y estabilizada porCalcio. Todas estas propiedades coinciden conaquel1as típicas de las o-arnilasas (Ming Shun
309
Chen et al., 1992).
Estabilidad térmica: La temperatura óptima deactividad de la enzima se observó a 40°C con unaligera disminución a SO°C. Después de los 50°Cla actividad de la enzima disminuyenotablemente, debido posiblemente a ladesnaturalización de la enzima aI ser sometida aaltas temperaturas(Fig. 2). En general la amilasa proveniente de H
hampei presenta buena estabilidad térmica en elrango com prendido entre 30 - 55°C, lo cual seatribuye a la presencia de Calcio en los buffers deactividad, debido a que las amilasas sonmetaloenzimas que requieren Calcio para laactividad catalítica (Robyt y Whelan, 1968). Laincubación de la amilasa de H. hampei por más de5 minutos a 70°C conlleva a la pérdida completade la actividad enzimática. Igual resultado fueencontrado con amilasas de S. zeamais y S.Granarius (Baker, 1983). Las enzimas tienendiferente energía de activación y de conversiónpor lo tanto una modificación en la temperaturapuede ocasionar un cambio en una de eIlas (Lee yAnstee, 1995).
Figura 1. Efecto del pH sobre la hidrólisis deI almidón a 30°C por amilasas de Hypothenemus hampei.
120
......'$o
100--~;l1lI ao'iIl:lOCIl 60.!!
~ 40i'O'> 20:;:;u«
o3 4 5 6 7 a 9
pH
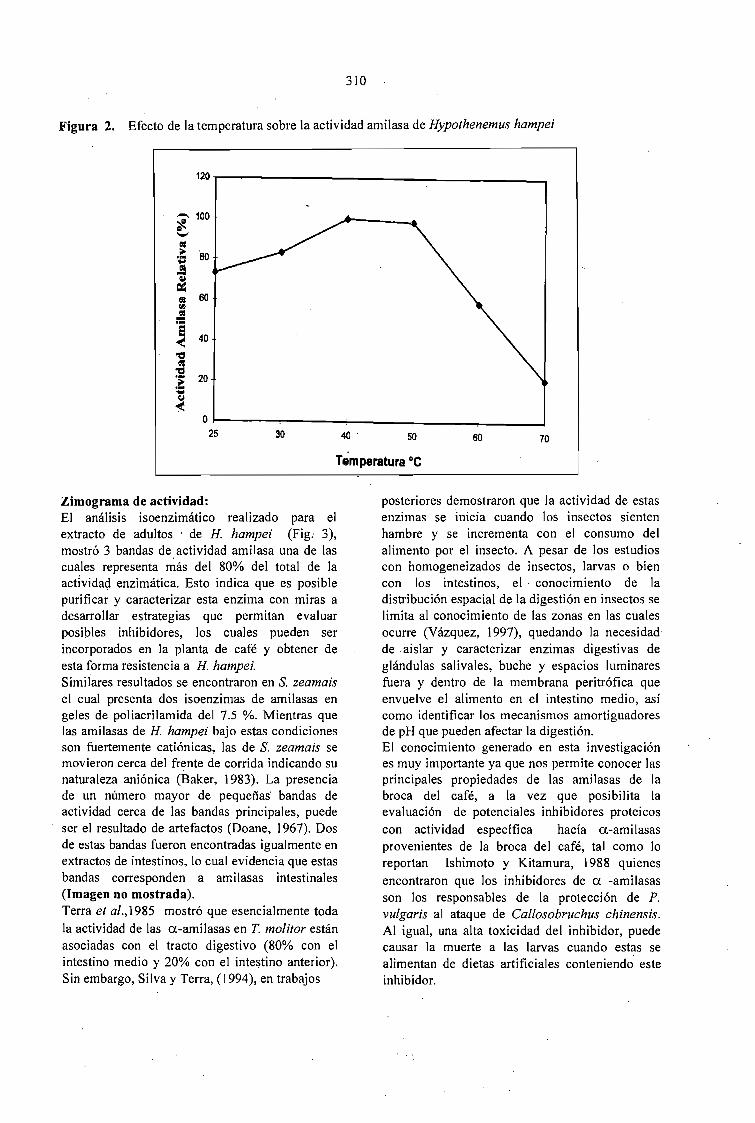
310
Figura 2. Efecto de la temperatura sobre la actividad amilasa de Hypothenemus hampei
120
.-. 100'#......=.i: ee....Jl~=:Cll 60fi)
.!!'i
40<~=~:e 20....u<
O25 30 40 50 60 70
Temperatura -c
Zimograma de actividad:El análisis isoenzimático realizado para elextracto de adultos . de H. hampei (Fig. 3),mostró 3 bandas de actividad amilasa una de lascuales representa más deI 80% deI total de laactividad enzimática. Esto indica que es posiblepurificar y caracterizar esta enzima con miras adesarrollar estrategias que permitan evaluarposibles inhibidores, los cuales pueden serincorporados en la planta de café y obtener deesta forma resistencia a H. hampei.Similares resultados se encontraron en S. zeamaisel cual presenta dos isoenzimas de amilasas engeles de poliacrilamida deI 7.5 %. Mientras quelas amilasas de H. hampei bajo estas condicionesson fuertemente catiónicas, las de S. zeamais semovieron cerca deI frente de corrida indicando sunaturaleza aniónica (Baker, 1983). La presenciade un número mayor de pequefías bandas deactividad cerca de las bandas principales, puedeser el resultado de artefactos (Doane, 1967). Dosde estas bandas fueron encontradas igualmente enextractos de intestinos, lo cual evidencia que estasbandas corresponden a amilasas intestinales(lmagen no mostrada).Terra et al.,1985 mostró que esencialmente todala actividad de las a-amilasas en T molitor estánasociadas con el tracto digestivo (80% con elintestino medio y 20% con el intestino anterior).Sin embargo, Silva y Terra, (1994), en trabajos
posteriores demostraron que la actividad de estasenzimas se inicia cuando los insectos sientenhambre y se incrementa con el consumo deIalimento por el insecto. A pesar de los estudioscon homogeneizados de insectos, larvas o biencon los intestinos, el· conocimiento de ladistribución espacial de la digestión en insectos selimita al conocimiento de las zonas en las cualesocurre (Vázquez, 1997), quedando la necesidadde .aislar y caracterizar enzimas digestivas deglándulas salivales, buche y espacios luminaresfuera y dentro de la membrana peritrófica queenvuelve el alimento en el intestino medi o, asícomo identificar los mecanismos amortiguadoresde pH que pueden afectar la digestión.EI conocimiento generado en esta investigaciónes muy importante ya que nos permite conocer lasprincipales propiedades de las amilasas de labroca deI café, a la vez que posibilita laevaluación de potenciales inhibidores proteicoscon actividad específica hacía cc-amilasasprovenientes de la broca dei café, tal como loreportan Ishimoto y Kitamura, 1988 quienesencontraron que los inhibidores de a -amilasasson los responsables de la protección de P.vulgaris al ataque de Callosobruchus chinensis.AI igual, una alta toxicidad dei inhibidor, puedecausar la muerte a las larvas cuando estas sealimentan de dietas artificiales conteniendo esteinhibidor.

311
Figura 3. Zimograma de actividad amilasa de Hypothenemus hampei en gel de electroforesisnativo dei 10 %. Después de la electroforesis, e1 gel fue incubado en buffer citrato 0.05M. pH 5,0conteniendo 10mM de NaC1 y 20mM de CaC12 por 120 minutos a 30°C. Tinción con solución deIodo. Las zonas claras indican bandas de actividad enzimática.
1
23
AGRADECIMIENTOS
Los autores deI artículo agradecen ampliamente aColciencias, a la Federación Nacional deCafeteros de Colombia y a la Universidad deCaldas, Colombia por el gran apoyo financieropara la ejecución de este trabajo.
LITERATURA CITADA
APPLEBAUM, S.W. 1964. The action partemand physiological role of Tenebrio larvalamylase. Journal of Insect Physiology 10: 897906.APPLEBAUM, S.W.; KONIJN, A.M. 1965. Theutilization of starch by larvae of the flour beetle,Tribolium castaneum. Journal of Nutrition 85:275-282.APPLEBAUM, S.W. 1985. Biochemistry ofdigestion. In: Kerkut, G.A.; Gilbert, L.I. eds.Comprehensive and Insect Physiology,Biochemistry and Pharmacology. OxfordPergamon Press, Vol 7. P. 279-311..BAKBR, J.E. 1983. Properties of amylases frommidguts of larvae of Sitophilus zeamais and
Sitophilus oryzae. Insect Biochemistry 13: 421428.BAKER, J.E. 1987. Purification of isoamylasesfrom the rice weevil, Sitophilus orizae(Coleoptera: Curculionidae), by highperformance liquid chromatography and theirinteraction whith partially-puri fied amylaseinhibitors from wheat. Insect Biochemistry 17(1):37-44.BAKER, J.E. 199 I. Puri fication . and parti alcharacterization of oc- amylase allosymes fromthe leeser grain borer, Rhyzopertha dominica.lnsect Biochemistry 21(3): 303-31 I.BIGGGS D.R.; McGREGOR P. G. 1996. Gut pHand amylase and protease activity in larvae oftheNew Zealand grass grub Costelytra zealandica;Coleoptera: Scarabaeidae as a basis for selectinginhibitors. Insect Biochemistry and MolecularBiology 26(1): 69-75.BLANCO L., A.; MARTINEZ G., A.;SANDOVAL c., A.; DELANO F., J. 1996.Purification and characterization of a digestivecathepsin O proteinase isolated from Triboliumcastaneum (Herbst). Insect Biochemistry andMolecular Biology 26(1): 95-100.

BOHINSKI, R.C. 1983. Modern concepts inbiochemistry. 4. ed. Estados Unidos, Al1en andBacon, 320p.BUONOCORE, V.; POERIO, E.; SILANO, V.;TOMASI, M. 1976. Physical and catalyticproperties of cc-amylase from Tenebrio molitorL. Larvae. Biochemistry Journal 153: 621-625.CAMPOS, F. AP.; XAVIER-FILHO, J.; SILVA,C.P.; ARY, M.B. 1989. Resolution and partialcharacterization of proteinases and cc-amilasesfrom midguts of larvae of the bruchid beetleCallosobruchus maculatus (F). ComparativeBiochemistry and Physiology 92B: 51-57.DOANE W. W. 1967. Quantitation of amylasesin Drosophila separated by acrylamide gelelectrophoresis. Journal of Experimental Zoology164: 363-378.HOPKINS, R.H.; BIRD, R. 1954. Action of alphaamilasa on amylase. Biochemistry 56: 86-96.ISHIMOTO, M.; KITAMURA, K. 1988.Identification of the growth inhibitor on azukibean weevil in kedney bean (Phaseolus vulgarisL.). Japanese Journal ofBreeding 38: 367-370.KRISHNA, S.S. 1955. Physiology of digestion inTrogoderma larva. Journal of the ZoologicalSociety of lndia 7: 170-176.KRlSHNA, S.S; SAXENA, N.K. 1962. Digestionand absorption on food in Tribolium castaneum(Herbst). Physiological Zoology 35: 66-78.LEE, M.J.; ANSTEE, J.H. 1995. Endoproteasesfrom the midgut of larval Spodoptera littoralisinclude a chymotrypsin-like enzyme with andextended binding site. Insect Biochemistry andMolecular Biology 25(1): 49-61.LEMOS, F. J. A.; CAMPOS, F. A P.; SILVA, C.P.; XAVIER FILHO, I. 1990. Proteinases andamylases af larval midgut of Zabrotessubfasciatus (Boh.) (Coleoptera:Bruchidae)reared in Cowpea (Vigna ungiculata (L.) Walp.)seeds. Entomologia Experimentalis et Applicata56: 219-227.MING-SHUN CHEN; GUOHUA FENG; ZEN,K.C.; RICHARDSON, M.; RODRIGUEZ V, S.;REECK R, R.; KRAMER J, K. 1992. a -amylasesfrom three species of stored grain Coleoptera and
312
their inhibition by wheat and corn proteinaceousinhibitors. Insect Biochemistry and MolecularBiology 22(3): 261-268.PODOLER, H.; APPLEBAUM, S. W. 1971. Thea amylase of the beetle Callosobruchuschinensis: properties. Biochemistry Journal 121:321-325.ROBYT J. F.; WHELAN W. J. 1968.The a.amylases. In: RADLEY, J. A. ed. Starch and itsDerivatives. London, Chapman & Hal1, P. 430476SANDOVAL, c., M.L. 1991. Purificación ycaracterización de enzimas larvales de cuatroinsectos que atacan aI maíz durante sualmacenamiento. Irapuato, CIEIAA, Universidadde Guanajuato, 135p. (Tesis Licenciatura)SILANO, V.; FURIA, M.; GIANFREDA, E.;MARCI, A.; PALESCANDOLO, R.; RAB, A.;SCARDI, V.; STELLA, E.; VALFRE, F. 1975.Inhibition of amylases from different origins byalbumins from the wheat kernel. ActaBiochimica et Biophysica 39: 170-178.SILVA, c.r., TERRA, W.R. 1994. Digestive andabsorptive sites along the midgut of the cottonseed sucker bug Dysdercus peruvianus(Hemiptera:Pyrrhocoridae). Insect Biochemistryand Molecular Biology 24(5): 493-505.TERRA, W.R.; FERREIRA,C.; BASTOS, F.1985. Phylogenetic considerations of insectdigestion. Disaccharidases and spatialorganization of digestion in the Tenebrio molitorlarvae. Insect Biochemistry 15: 443-449.TERRA, W.R. 1990. Evolution of digestivesystems of insects.Annual Review of Entomology35: 181-200.VAZQUEZ A, M. 1997. Anatomical, enzymatic,and microbiological studies on the digestivesystern of Prostephanus truncatus (Horn).Leicester, University of Leicester, 150p. Tesis:Doctor ofPhilosophy.YETTER M. A; SAUNDERS R. M.; BOLES H.P.1979. a. amylase inhibitors from wheat kernels asfactors in resistance to postharvest insects. CerealChemistry 56: 243-244.

313
ATRATIVIDADE DE FRUTOS À BROCA-DO-CAFÉ, hypothenemus hampei (FERRARI):ESTÍMULOS VISUAIS E OLFATIVOS.
Jorge Rafael Mendoza (INIAP-EQUADOR), José Oscar Gomes de Lima(UENF-RJ), Evaldo FerreiraVilela(UFV) & César José Fanton(EMCAPA-ES)
Depto. Biologia Animal/ UFV, Viçosa, MG, 36571-000
INTRODUÇÃO
A broca do café, hypothenemus hampei (Ferrari,1867) (Coleoptera: Scolydae) é a praga maisimportante do cafeeiro em diversos paísesprodutores, atacando os frutos em todos osestágios de maturação, causando perdassignificativas no campo e no armazenamento. Osestudos sobre essa praga tem se concentrado emsua biologia, ecologia e controle. Contudo, osfatores de atratividade da broca para os frutos nãoestão completamente elucidados. O presentetrabalho busca esclarecer o papel de estímulosvisuais e olfativos na atração da broca para osfrutos de café.
MATERIAL E MÉTODOS
Bioensaio 1- Preferência da broca-da-café porfrutos em diferentes estágios de maturação Frutos de coloração verde, amarela, vermelha epreta suspensos em alfinetes e fixos em placa deisopor foram confimados com uma fêmea dabroca em um copo plástico transparente de 200ml,e a preferência da broca verificada após 24 horas.Foram testadas 100 repetições e os resultadoscomparados pelo teste do Qui-quadrado.
Bioensaio 2- Preferência da broca-do-café porfrutos simulados de diferentes cores - Os frutossimulados constituíam-se de esferas coloridas deisopor de 20 mm de diâmetro nas cores verde,amarela, vermelha e preta. A avaliação da
preferência e análise dos dados seguiram o mesmoprocedimento do bioensaio I.
Bioensaio 3- Resposta da broca-do-café acomponentes voláteis em olfatômetro - - Paraverificar a resposta da broca a odores provenientesdo fruto, utilizou-se olfatômetro e método descritopor Moeck(1970), em que o ar insulflado passapor um erlenmeyer com água destilada, sendolimpo e umidificado. Passa a seguir por umacâmara que contém o material que se quer testar eserá a fonte de odor para a broca colocada em umaarena. Cada inseto foi testado por três vezesconsecutivas e então descartado. A resposta foiconsiderada positiva sempre que o inseto cessavaa locomoção ao interceptar o fluxo de odor;prosseguindo a seguir em direção à fonte; aresposta era considerada não positiva quando oinseto caminhava em direção à luz oumovimentava-se ao acaso dentro da arena. sresultados foram comparados pelo teste do Quiquadrado.
RESULTADOS
Foi comprovada a preferência da broca-do-cafépelas cores preta e vermelha, tanto em frutoscomo simulados em comparação coma s coresamarela e verde (Tabela 1). A resposta olfativa acomponentes voláteis produzidos por frutos decafé indica que as brocas são mais atraídas porfrutos maduros, tantos sadios com brocados, doque por frutos verdes (Tabela 2).
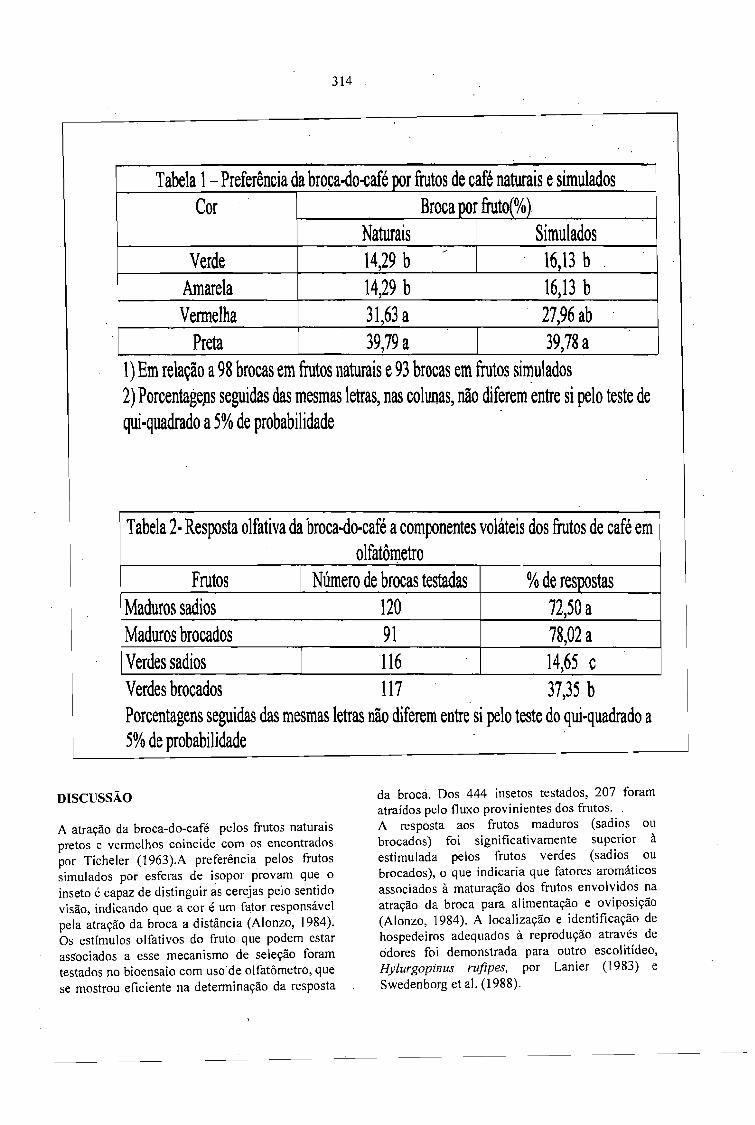
314
Tabela 1-Preferência da broca-do-café por frutos de café naturais esimuladosCor Broca por fruto(%)
Naturais SimuladosVerde 14,29 b - 16,13 b
Amarela 14,29 b 16,13 bVennelha 31,63 a 27,96 ab
Preta 39,79 a 39,78 a1) Em relação a98 brocas em frutos naturais e93 brocas em frutos simulados2) Porcentageps seguidas das mesmas letras, nas colunas, não diferem entre si pelo teste dequi-quadrado a5% de probabilidade .
Tabela 2- Resposta olfativa da broca-do-café acomponentes voláteis dos frutos de café emolfatômetro
Frutos Número de brocas testadas %de respOstasMaduros sadios 120 72,50 aMaduros brocados 91 78,02 aVerdes sadios 116 14,65 cVerdes brocados 117 37,35 bPorcentagens seguidas das mesmas letras não diferem entre' si pelo teste do qui-quadrado a5% de probabilidade "
DISCUSSÃO
A atração da broca-do-café pelos frutos naturaispretos e vermelhos coincide com os encontradospor Ticheler (1963).A preferência pelos frutossimulados por esferas de isopor provam que oinseto é capaz de distinguir as cerejas pelo sentidovisão, indicando que a cor é um fator responsávelpela atração da broca a distância (Alonzo, 1984).Os estímulos olfativos do fruto que podem estarassociados a esse mecanismo de seleção foramtestados no bioensaio com uso 'de olfatômetro, quese mostrou eficiente na determinação da resposta
da brocá. Dos 444 insetos testados, 207 foramatraídos pelo fluxo provinientes dos frutos.A resposta aos frutos maduros (sadios oubrocados) foi significativamente superior àestimulada pelos frutos verdes (sadios oubrocados), o que indicaria que fatores aromáticosassociados à maturação dos frutos envolvidos naatração da broca para alimentação e oviposição(Alonzo, 1984). A localização e identificação dehospedeiros adequados à reprodução através deodores foi demonstrada para outro escolitídeo,Hylurgopinus rufipes, por Lanier (1983) eSwedenborg et aI. (1988).

Comparadas as respostas a frutos sadios (maduros+ verdes) e brocados (maduros + verdes) não seobservou diferença significativa. No entanto, nacomparação das respostas a frutos sadios oubrocados em cada grupo, maduros ou verdes,observou-se aumento da resposta estimulada pelosfrutos brocados, particularmente os verdes, paraas quais a diferença foi significativa.Possivelmente, o dano provocado nos frutos decafé aumenta a liberação de compostos voláteisproduzidos pelo hospedeiro, ou uma mistura desubstância produzidas pelos hospedeiro e peloinseto, aumentando a atratividade para seusconspecíficos, o que explicaria o comportamentode agregação da broca por Baker(1984).
REFERENCIAS BIBLIOGRÁFICAS
Alonzo, F.R. 1984. Aspectos ecológicos de labroca Hypothenemus hampet(Scolytidae:Coleoptera). In: El problema de labroca (Hypothenemus hampei Ferr.)(Col.:Scolytidae) y la caficultura. San Jose, Costa Rica,IlCA- PROMECAFE. p. 7 1-136.
315
Baker, P. S. 1984. Some aspects of the behaviorof the coffee berry borer in relation to its controlin Southern Mexico (Coleoptera, Scolytidae).Folia Entomologica Mexicana, 61:9-24.Lanier, G. N. 1983. Integration of visual stimuli,host odorants, and pheromones by bark beetlesand weevils in locating and colonizing host trees.In: Herbivorous insects. New York, AcademicPress, p. 161-171.Moeck, H. A. 1970. An olfactometer for the bioassay of attactants for scolytids. CanadianEntomologist, I02:792~796.Swedenborg, P. D.; Jones, R L.; Ascerno, M. E.& Landwehr, V. R. 1988.
Hy/urgopinus rufipes (Eichhoff) (Coleoptera:Scolytidae): Attraction to broodwood, 110stcolonization behavior and seasonal activity inCentral Mennesota. Canadian Entomologist,120:1041-1050.Ticheler, J. H. G. 1963. Estudio analitico de laepidemiologia del escolitideo de los granos decafe Stephanoderes hampe/, Ferr., en Costa deMarfil. Cenicafe, 14:223-294.


317
AVANÇOS NAS TÉCNICAS PARA CRIAÇÃO MASSAL DO PARASlTÓIDE Cephalonomiastephanoderls HYMENOPTERA:BETHYLIDAE) SOBRE Hypothenemus hampei(COLEOPTERA:SCOLYTIDAE) MULTIPLICADA EM DIETÀ ARTIFICIAL.
Amador Villacorta & Sônia M. Torrecillas
rapar, Cx. Postal 481. 86.001-970 - Londrina-Pr.
INTRODUÇÃO
A cnaçao massal do parasitóide Cephalonomiastephanoderis (Hymenoptera:Bethylidae) énormalmente feita utilizando seu hospedeiro, abroca do café Hypothenemus hampei(Coleoptera:Scolytidae). Num sistema deprodução é utilizando os frutos de cafébroqueados (Barrera, 1994), e no outro sistema ocafé pergaminho é realizado infestandoartificialmente com broca (Benavides & Portilla,1990, Portilla & Bustillo, 1995). Os problemasbásicos apresentados por ambos os métodos sãoseu alto custo e a sua dependência de frutosbroqueados coletados no campo, o qual não estádisponível todo o ano próximo da unidade decriação. Consequentemente, a criação da brocaem dieta artificial seria aparentemente a soluçãopara este problema. A primeira dieta artificialpara broca foi publ icada por Villacorta (1985), aqual foi modificada para uma dieta comercialpublicada por Villacorta & Barrera, 1993,Villacorta & Barrera, 1996, e uma outra dieta estásendo utilizada para criação da broca para aprodução de ç. stephanoderis na Colômbia(Portilla 1999). Neste artigo descreveu osconceitos e metodologia da técnica de criação de!:!. hampei e seu parasitóide e também como sepretende instalar a planta piloto de produçãomassal de ç. stephanoderis no Brasil, com basenos avanços real izados em outros países daAmérica Latina.
DIETA ARTIFIClAL PARA H. humpei
Basicamente todas as dietas de broca publicadascontem os seguintes ingredientes: agar, caseína,levedo, açúcar, café cru moído e agentesantioxidantes e antimicrobianos. A dieta queusamos no Brasil é a publicada por Villacorta &Barrera (1996) e que originalmente foidenominada ECOBROVILL-160 que tem osseguintes ingredientes: agar 21g, café cru moído100g, caseína 20g, açúcar 14g, levedo de torula20g, nipagin Ig, sorbato de potássio 0.6g, etanol10 ml, formaldeido 2 ml e água 700 ml. Oscustos dos ingredientes da dieta de broca para
produzir um litro de dieta variam grandementede país para país, assim temos os custos de 8dólares no México a I.~ dólares na Colômbia,isto devido ao fato de que alguns dos'ingredientes tem que ser importados. Umingrediente que aumenta grandemente os custos éo Agar mas se seguirmos a sugestão de Portilla(1999) de usar Agar industrial ao invés de Agarbacteriológico os custos podem cair mais de 40%.É necessário mais pesquisas em relação aosrequerimentos nutricionais da broca do café, esobre a relação simbiótica microorganismosbroca do café, tal como foi sugerida a existênciade uma simbiose entre a broca e o fungoTrichoderma sp por Villacorta, (1989), que foireforçada nas publicações de desenvolvimento dedietas de brocas em Villacorta & Barrera, (1996)e Rojas et ai (1999) que consideraram Fusariumsolani como um simbionte da broca.
INFESTAÇÃO DA DIETA
As brocas utilizadas no início da cnaçao dasbrocas em dieta artificial são procedentes defrutos atacados no campo. Sendo que os frutossão esterilizados usando 2% de Formaldeido(37%), seguido de um banho em água destiladaesterilizada. Dos frutos se obtém prepupas, pupase fêmeas ativas. Este material biológico devepassar por uma esterilização de acordo comVillacorta & Barrera (1993). É recomendado usarum macho para cada nove fêmeas para assegurara plena fecundação das fêmeas.
PRODUÇÃO DAS BROCAS
Os tubos de vidro (1.7 x 7 em fundo chato) comdieta infestadas com broca, são mantidos em salascom temperatura de 26 ±2oC e 60±5% U.R. e emcondições de completa escuridão; 80 a 90 diasapós a infestação da dieta os tubos são abertos ecom um toque na base dos tubos os pellets dadieta caem. Todo o conteúdo dos tubos écolocado no topo de um aparelho formado poruma série de peneiras e por agitação mecânica,podem ser separados: os pellets da dieta,diferentes estágios de desenvolvimento da broca,

um pó fino com presença de ovos, larvas,prepupas e pupas. Os adultos ativos, após umaesterilização por superfície são usados parainfestar novos tubos com dieta. Após 85-90 diasde infestação da dieta, os tubos contém entre 250a 300 indivíduos por tubo, contando todos osestágios de desenvolvimento. Consequentemente,de 245 tubos de cada receita da dieta, é possívelproduzir entre 36.250 43.500 brocas.Considerando que cada receita de dieta custa 8dólares no México, o custo para produzir 1.000brocas pode ser estimado entre 0.20 a 0.25dólares.
CRIAÇÃO DE C. STEPHANODERISUSANDO BROCAS MULTIPLICADAS EMDIETA ARTIFICIAL
Aos 85-90 dias após a ínfestação das dietas combrocas, o número de prépupas e pupas de broca, éde 45 a 80 indivíduos aptos para oviposição porparasitóides. Considerando que tubos infestadoscom brocas aos 85-90 dias é o momento idealpara a criação dos parasitóides, é proposto doismétodos (i) criação "in vitro" no qual são usadosas prépupas e pupas da broca separadas da dietadurante o processo de tamizagem (ii) o uso dospellets que contém em seu interior diferentesestágios de desenvolvimento da broca. Ambosmétodos são descritos a seguir.
Criação "in vitro"Utilizando as peneiras é possível separar 50% dasprépupas e pupas das dietas, dando lugar a 28 a40 indivíduos por tubo, os quais podem serusados para a criação dos parasitóides. A criaçãoconsiste na impregnação da superfície de umacartela de papel corrugado (10.5 x 9.5 em) comgoma arábica diluída em água, que em seguida érecoberta por uma cobertura de pó fino desemente de café. Na base da canaleta do papelcorrugado são depositadas as pupas, prépupas,larvas e ovos da broca. E, em seguida sãocolocadas em uma caixa plástica de 11 x 10 x 5crn, com uma abertura na tampa de 5 x 5 em,coberta com uma tela fina. Em cada caixa plásticasão colocados entre 25 a 30 parasitóides emantidos em uma sala com 27±20C e 75±5%U.R. Após 10 dias pode-se observar a formaçãode casulos agrupados a um dos extremos dacanaleta, isto por causa do comportamentomaternal do parasitóide.
318
CRIAÇÃO NOS PELLETS DE DIETA
Os pellets são infestados na proporção de doisparasitóides por pellets. Os pellets de dieta sãocolocados em caixas de plástico redondas comum diâmetro de 22 em e 10 em de altura com umaabertura na tampa, coberta por uma tela fina. Omaterial é colocado em um quarto de criação com27± 2°C e 75 ( 5% U.R. De 17 dias em diante anova geração de parasitóides inicia suaemergência. Usando 140 pellets (um litro dedieta) é possível obter entre 2.800 a 3.000parasitóides .
CONCLUSÕES E RECOMENDAÇÕES
As técnicas descritas neste artigo objetivam amáxima utilização do material biológico da brocaproduzido em dietas artificiais. Para criação emdieta oferece as seguintes vantagens:(i) permite manter uma população de brocas eparasitóides durante o período de entressafra;(ii) tem grande valor para os trabalhos depesquisa para o estudo da biologia ecomportamento da broca e seu parasitóide,(iii) permite produzir um pequeno número deparasitóides para liberações em campo; para (iv)permite manter brocas durante a entressafra osquais podem ser usados na criação de brocas emcafé pergaminho; para (v) abre o caminho parapensar em um processo industrial na criação dosparasitóides.É necessária pesquisa no desenvolvimento demétodos de automação na preparação da dieta, nadistribuição da dieta nos tubos, perfuração dadieta e fechamento dos tubos. Isto pode reduzir oscustos de produção e levamos a uma criaçãocomercial de C. stephanoderis, o qual nospermitiria a integração desta técnica no manejointegrado da broca, com base em liberaçõesinundativas deste parasitóide.
LITERATURA CITADA
Barrera, lF. 1994. Dynamique des populationsdu scolyte des fruits du caféir, Hypothenemushampei (Coleoptera:Scolytidae), et luttebiologique avec la parasitoide Cephalonomiastephanoderis (Hymenopetra:Bethylidae) auChiapas, Mexique. Thesis. Université PaulSabatier, France, 301 pp.
Benavides, G.M. & . M. Portilla. 1990. Uso deicafe pergamino para 'ia cria deHypothenemus hampei y de su parasitoide

Cephalonomia stephanoderis en Colombia,Cenicafe (Colombia) 41: 114-116.
Portilla, M. & A Bustillo. 1995. Nuevasinvestigaciones eIJ la cria masiva deHypothenemus hampei y de sus parasitoidesCephalonomia stephanoderis y Pr.orops nasuta.Revista Colombiana de Entomologia 21(I ):2533.
Portilla, M. 1999. Mass production ofCephalonomia stephanoderis onHypothenemus hampei reared using artificialdiet. Thesis. University of London, England.253 pp.
Rojas, M.G., l.A Morales-Rarnos & T. C.Harrington. 1999. Association betweenHypothenemus hampei (Cleoptera :Scolytidae)and Fusarium solani(Moniliales:Tuberculariacea). Ann. Entomol. Soe. Am.92( 1):98-100.
319
Villacorta, A 1989. Aspectos nutricionales de lacria de la broca deI café (Coleoptera:Scolytidae). III Taller regional de broca,Antigua, Guatemala, Abril 3-7, 1989. nCAPRüMECAFE.181-186.
Villacorta, A. 1985. Dieta meridica paracriação de sucessivas gerações deHypothenemus hampei (Ferrari,I867)(Coleoptera:Scolytidae). An. Soe.Entomol. Brasil. 14:315-319.
Villacorta, A. & l. F. Barrera, 1993. Nova dietameridica para criação de Hypothenemushampei (Ferrari) (Coleoptera:Scolytidae). An.Soe. Entomol. Brasil. 22:405-409.
Villacorta, A. & l.F. Barrera. 1996. Techniquesfor mass rearing of the parasitoidCephalonomia stephanoderis(Hymenoptera:Betylidae) on Hypothenemushampei (Coleoptera:scolytidae) using anartificial diet. Vedalia 3:45-48.


I
.- .- '7"': _ .._ __v,",, .. __ __
... .. ..- _....----


323
EXPERIÊNCIA BRASILEIRA NA VALORIZAÇÃO BIOTECNOLÓGICA DE SUBPRODUTOSDA AGROINDUSTRIA DO CAFÉ
Carlos Ricardo Soccol', Fan Leifat, Adenise L. Woíciechowski', Debora Brandi, Cristina M. M.Machado", Marlene Soares! ,Pierre Christen2 Ashok Pandey',
'Laboratório de Processos Biotecnológicos- Departamento de Engenharia Química, Universidade Federal doParaná (UFPR), CEP 81531-970 Curitiba- PR- Brazil.2Laboratoire de Microbiologie, Université de Provence (CESB/ESIL)-13288, Marseille - France
INTRODUÇÃO
o Brasil, maior produtor, consumidor eexportador mundial de café, com uma produçãoanual de cerca de 30.5 milhões de sacas, das quais26 milhões de café arábica e 4.5 milhões de caférobusta. segundo dados da Associação Brasileirados Exportadores de Café (Abecafé). Dessaprodução, 90% do café produzido atualmenteutiliza a técnica de descasque por via seca. O cafécolhido é lavado, de forma similar à que é feitana via úmida. Depois da lavagem, resultam duasfrações: o café chamado "bóia" e o café chamadomiúdo. As duas frações serão secasseparadamente.O processo de secagem pode ser natural ouartificial. Na secagem natural. a mais utilizada, osgrãos de café são transportados para umasuperficie plana de terra, asfalto ou tijolo de argilae são secos até que a umidade dos grãos fiquemcom cerca de I 1-12%. Pelo processo de secagemartificial, os grãos são secos com convecçãoforçada, até o mesmo teor de umidade.Depois da secagem, o café vai à uma etapa delimpeza, na qual os grãos são separados deimpurezas (pedras,terra, folhas, restos de galhos..etc.) através de peneiras vibratórias.O café limpo sofre então o descasque, que separaas cascas para a parte de fora da máquina. O cafédescascado é selecionado, embalado e enviadopara industrias de torrefação e de fabricação decafé solúvel.Considerando que a casca constitui cerca de 50%do grão seco, grandes volumes são geradosanualmente no país.Outro problema que deveremos enfrentar embreve em nosso país, trata do destino quedeveremos dar aos resíduos líquidos e sólidosgerados no processo por via úmida, que começama ser implantados no país como forma de atendera demanda internacional crescente por cafés finosde alta qualidade. Isso irá levar a uma aumentosignificativo dos problemas ambientaisocasionado por grandes volumes de H20 delavagem e polpa úmida.A técnica da cultura do café adensado práticadaem muitas lavouras no estado do Paraná
como forma de aumentar a produtividade ereduzir as perdas ocasionadas pelas geadas, temlevado a biomassa foliar extremamente importanteo que vem dificultando a colheita das cereja quecaem no solo durante a colheita.Desta forma esse esta palestra busca apresentaralgumas alternativas biotecnológicasdesenvolvidas pelo Laboratório de ProcessosBiotecnológicos da UFPR para o manejo dessesresíduos, pois os mesmos apresentam elevadopotencial para uso na alimentação animal ou parauso como substrato na obtenção de produtos ebiomoléculas de elevado valor agregado (Soccol& Krieger, 1998). No caso do uso desses resíduosagro-industriais na aIimentação animal é defundamental importância que seja estudado oproblema da eliminação dos compostosrecalcitrantes tóxicos (CRT) tais como cafeína,taninos e polifenóis. Atualmente os compostosrecalcitrantes tóxicos representam Uma grandelimitação no uso da polpa, casca do café e mesmofolhas na alimentação animal, embora taisresíduos possuam um valor calórico-protéicoconsiderável (11.3 % proteína bruta ; 2.0% dematéria graxa ; 27.6% de celulose: 20% dematérias pécticas e açucares redutores, (TANGO,1971 : VASCO, 1989).Além da eliminação dos fatores anti-nutricionaiscom vistas a sua utilização na alimentação deruminantes, aves e peixes estaremos tambémrealizados estudos com objetivo de utilizar essessub-produtos como substrato de fermentação paraprodução de fungos comestíveis e biomoléculasde interesse comercial tais como ácidosorgânicos, hormônios vegetais e aromas.
PRODUÇÃO DE FUNGOS COMESTÍVEIS
a) Pleurotus: Estudos foram realizados visandoutilizar cascas, folhas e borra de café na produçãode fungos comestíveis do gênero Pleurotus.Inicialmente 10 diferentes cepas de fungos dogênero Pleurotus pertencentes ao Laboratório deProcessos Biotecnológicos da UFPR foramtestadas em um meio de cultivo a base agarextrato de casca de café. Os estudos de seleção decepas foram realizados em função da velocidade

de crescimento radial e da formação de biomassade acordo com técnica descrita por Soccol(1994).A cepa Pleurotus ostreatus LPB 09 foiselecionada para estudos posteriores defrutificação por apresentar o melhor crescimentoem agar casca de café (9,68 mm/dia e 43,40mg/placa em 9 dias de cultura a 24 De.Quando somente casca de café foi usada comosubstrato o rendimento de produção foi de968g/Kg de casca de café seco, sendo que osprimeiros corpos de frutificação foram colhidosapós 20 de cultura.Rendimentos elevados na produção de fungoscomestívies foram também obtidos utilizandoborra de café como substrato na (903.5g/Kg decasca de café seco). Os resultados demostraramque o uso de folhas de café isoladamente comoúnico substrato não é recomendável para aprodução de fungos comestíveis do gêneroPleurotus.
b) Lentinus edodes: Trabalho semel hante foirealizado com o fungo comestível Lentinusedodes LPB 02 conforme técnica descritaanteriormente. A frutificação foi realizada emsaco plástico utilizando 2 Kg de borra de cafécom 65% de umidade e inoculada com 15% de"spawn" produzido a partir de uma mistura de80% de serragem de eucalipto e 20% de farelo detrigo. Os primeiros corpos de frutificaçãoapareceram 60 dias após inoculação. A eficiênciabiológica após a terceira colheita foi de 886g defungo fresco/Kg de borra de café fresco.
DETOXIFICAÇÃO BIOLÓGICA DA CASCADO CAFÉ
A casca de café apresenta um baixo valornutricional, devido a presença de substânciastóxicas como cafeína (1,2 %), taninos (6,3%) epolifenóis, Uma alternativa para a redução dessassubstâncias tóxicas é a utilização da técnica dafermentação no estado sól ido, empregandomicroorganismos selecionados capazes de utilizara cafeína como fonte de nitrogênio. Afermentação no estado sólido é considerada a maisnatural das fermentações, porque seus processosassemelham-se as condições sob as quais amaioria dos microrganismos crescem na natureza.É definida como o crescimento demicroorganismos em materiais sólidos naausência de água livre, mas o substrato deveconter umidade suficiente, presente na formaabsorvida dentro da matriz sólida.O objetivo do presente trabalho foi incrementar ovalor biológico da casca de café, pela
324
detoxificação de seus componentes tóxicos porfungos filamentosos, utilizando a técnica defermentação no estado sólido.Os microorganismos foram selecionados combase na sua velocidade de crescimento radial ebiomassa produzida em ágar extrato de casca decafé (lOOg/L). Foram utilizadas onze cepas defungos filamentosos do gênero Rhizopus e duas dogênero Phanerochaetae.O crescimento radial em ágar extrato de casca decafé foi realizado por meio de inoculação de umasolução de esporos dos microrganismos estudadosno centro de uma placa de Petri. A biomassa foimedida através da dissolução do ágar e separaçãoda biomassa, que foi então pesada em um papel defiltro previamente tarado.Para o preparo do substrato (casca de café) para adetoxificação. Neste caso a casca de café foiinicialmente seca em estufa com circulação de ar à550 C por 48 horas até atingir uma umidade deaproximadamente 8 %. Em seguida a casca foimoída e penei rada. Uti lizou-se a fração comgranulometrica compreendida entre 0,8 e 2,0 mmpara os estudos de detoxificação. Os trabalhos deseleção em agar café demonstraram que osfungos do gênero Rhizopus são capazes de crescermuito bem neste tipo de substrato, pois avelocidade de crescimento radial das cepasestudadas variaram de 0.94 mm/h à 2.19 mm/h. Acepa Rhizopus arrhizus 16179 foi selecionadapara os estudos posteriores por apresentar a maiorprodução de biomassa e velocidade decrescimento radial (2.03 mm/h e 12.10 mg/placa).A cepa Phanerocaetae chrysosporium BK foitambém selecionada para detoxificação da cascado café.As cepas Rhizopus arrhizus 16179 ePhanerocaetae chrysosporium BK foramselecionadas por serem capazes de degradarparcialmente os compostos tóxicos presentes nacasca de café. Os testes deste estudo foramrealizados sem a adição de nutrientes às cascas decafé, o que significa que os microorganismospodem utilizar somente os componentes presentesno substrato para biotransforma-lo. Os resultadosdemostraram que a cepa de, Rhizopus arrhizus16179 em 6 dias de fermentação foi capaz dereduzir o teor de cafeina presente inicialmente nacasca em 87% e em 65% a concentração detanino. Os melhores resultados alcançados com acepa de Phanerocaetae chrisosporium BK foiuma redução na concentração de cafei na de 71% etanino de 45% em 16 dias de fermentação.

OBTENÇÃO DE SUBSTRATOS PARA ODESENVOLVIMENTO DE PROCESSOSBIOTECNOLÓGICOS ATRAVÉS DAHlDRÓLISE DA CASCA DO CAFÉ.
A técnica da hidrólise ácida, alcalina ouenzimática tem sido mundialmente utilizada naobtenção de substratos fermenteciveis ricos emmonossacarídeos. As hidrólises ácidas (cloridricae sulfurica) e alcalina (NH40H, (NH4)2S04, Uréiae Na2C03) foram realizadas em autoclaveutilizando 200 g de cascas de café por litro desolução. Foram testadas temperaturas de 100, 120e 140°C por tempos de 5, 10. 15, 30 e 60minutos. Obtém-se desse tratamento uma fraçãolíquida e uma sólida, as quais foram separadas porfiltração. Como os açúcares redutores (simples)são solúveis em água, a fase líquida é rica emaçúcares redutores, enquanto a sólida é maispobre. Os açúcares redutores nas fases líquidas esólida foram .determinados pelos métodoscolori métricos de Somogyi-Nelson e D.N.S.Concentrações de açúcares redutores entre 30 e 50g/L foram obtidas na fase líquida. sendo que omelhor resultado foi de 49,5 g/L nas frações emque foram utilizadas somente água e casca de caféna temperatura de 100 DC por 60 minutos. A autohidrólise permitiu obter uma concentração de247.5 gramas de açucares redutores em glicose(ARG) por kg de casca de café seca. Esseresultado representa uma recuperação de 98% dosARG presentes na casca do café, utilizandoapenas água, baixa temperatura (96°) e pressãoatmosférica.
PRODÚÇÃO DE AROMA FRUTAL PORCeratocystis flmbriata EM RESíDUO SÓLIDODA AGROINDÚSTRIA DO CAFÉ
Apesar da maioria dos aromatizantes serematualmente obtidos por síntese química(84%),pesquisas mostram a crescente preferência dosconsumidores por alimentos naturais ou quecontenham aditivos naturais (Janssens et aI.1992). Plantas continuam sendo importantesfontes de óleos essenciais e aromas, mas hágrande dependência em fatores de dificil controle,como clima e doenças. Uma alternativa à síntesede aromas naturais baseia-se nos processosmicrobiológicos, como fermentações (biosíntese)e bioconversões. O custo de produção de taisaromas pode ser questionado; contudo, o preço demercado pago por aromas naturais écompensador. Por exemplo, a 4-decalactonasintética (aroma de impacto em pêssegos) custa150 dólares/ Kg, enquanto seu similar natural
325
custa 6000 dólares/ Kg(4). Fungos do gêneroCeratocystis sintetizam diferentes aromasfrutais/florais (pêssego, abacaxi, banana, cítricos erosa), dependendo da cepa e das condições decultivo, sendo que C. jimbriata é interessantedevido ao seu rápido crescimento, habilidade naprodução de esporos e à variedade de aromassintetizados (Senernaud, 1988). Neste trabalho,procedeu-se a fermentação no estado sólido dacasca de café com o intuito de avaliar a produçãode aromas por C. fimbriata, comparando osresultados com outros mencionados paradiferentes resíduos agroindustriais. A influênciada adição de glicose e o uso de substânciasprecursoras de aromas foram estudados. Oscompostos biosintetizados presentes no headspacedos cultivos foram caracterizados e quantificados.
Procedimentos de fermentação: Os experimentosforam conduzidos em Erlenrneyers de 250mL,contendo 15g de matéria seca (MS) e cobertoscom 8 camadas de gaze. Em todos osexperimentos as condições IniCIaIS foram:temperatura 30'C, pH 6, inóculo I x 10.7 esporos golMS e 70% de água (máxima absorção dosubstrato)(1 ;3). A influência da adição de glicose(20%, 35% e 46%) e da adição de possíveisprecursores (leucina 10 mMol (2), óleo de soja10%(1) e solução salina(2)) foram estudadas paraotimização da produção de voláteis totais (VT).
Procedimentos analíticos: Cada experimento foirealizado em triplicata. O acompanhamento daevolução da produção dos voláteis foi realizadaatravés de avaliações sensoriais paralelamente àanálise por Cromatografia de Gases (CG).Os aromas frutais de banana, abacaxi e macãforam produzidos durante o cultivo deCeratocystis jimbriata em casca de café. Ficoucaracterizado que a biosíntese de ésteres, como:isoamil acetato, etil acetato, isobutil acetatoformam aroma de banana), etil isobutirato e etilbutirato dão origem aos aromas de maçã emorango.A análise por Cromatografia de Gases permitiu aidentificação de diversos compostos, comoindicado na Tabela 1, que apresenta os dadosintegrados para cada composto presente noheadspace dos cultivos. A adição de leucinaaumenta a produção de ésteres derivados dometabolismo deste aminoácido. Todos oscompostos já foram mencionados previamente poroutros autores (Bramorski et aI., 1998 ; Christenet aI., 1991 ; Christen et al., 1997).

326
TABELA 2: Resultados da presença de GA 3 produzidopor fermentação em cromatografia em camadadelgada.
utilização mais extensa, trazendo ótimosbeneficios econômicas à agricultura e indústria.Uma possibilidade de diminuição dos custos estána pesquisa de novas técnicas de fermentação(Kumar e Lonsane, 1987, 1989; Brückner eBlechschmidt, 1991).Recentemente, tem aumentado o interesse decientistas no desenvolvimento de produção degiberelinas por fermentação no estado sólido(FES). Fermentação sobre farelo de trigo édescrita por Kumar e Lonsane (1987), quecompararam o rendimento com a técnicasubmersa. Tal comparação também foi feita porTornasini et aI. (1997), utilizando-se comosubstratos farinha de mandioca, bagaço de canade-açúcar e poliuretano de baixa densidade.Ambos grupos obtiveram maior rendimento com aFES, no caso de Kumar e Lonsane de 1,6 vezes, ecusto cerca de 2,5 vezes menor.Este trabalho teve por objetivo determinar apossibilidade da utilização da casca do café comosubstrato de fermentação para produção de ácidogiberélico. Para tanto foram utilizadas seis cepasde microrganismos produtoras de GA 3 emfermentação no estado sólido e submersa,comparando-se dessa forma, o rendimento dosmétodos.A Tabela 2 mostra os resultados da seleção decepas em casca de café tratada de 3 diferentesmaneiras. O melhor resultado foi obtido com o.resíduo sólido da extração da casca do café. Issose dá, provavelmente, por existirem substânciasinibidoras do metabolismo de produção dasgiberel inas nos resíduos de café (casca e extrato),que com o pré-tratamento (hidrólise) sãoeliminadas.O microrganismo que apresentou melhorprodução foi a cepa LPB-06, para os três sistemasde fermentação. Dessa forma, a quantificação porHPLC foi feita para os fermentados por estemicrorganismo. Para os sistemas em FES, a cepaLPB-O1 e LPB-03 também apresentaram bonsresultados; já para FSm, a cepa
Onde: MaIOr numero de + Significa melhorfi uorescência.
CEPA FSm FES FES resíduo Iextrato casca após extraçãoDe casca de café
LPB-l +++ ++++ ++++LPB-2 ++++ ++ +++ ILPB-3 ++ +++ +++LPB-4 + + ++LPB-5 + + ++LPB-6 ++++ ++++ +++++
" ,~
Ácido giberélico (GA 3) é um hormônio endógenode plantas superiores e metaból ito secundário decertos fungos. É um importante promotor eregulador do crescimento das plantas sendoutilizado na agricultura, viveiros, produção deflores, etc., para uma série de efeitos benéficos,como eliminação da dormência e aceleração degerminação em sementes, melhora no rendimentodas colheitas, promoção da formação do fruto,indução do florescimento e outras. Nos últimosanos, este metabólito tem sido produzidoindustrialmente por fermentação submersa a umalto custo (US$ I a 3/grama, dependendo dapureza e potência), uma vez que o rendimento émuito baixo e sua extração dificil. A redução domesmo levará suas diversas aplicações a uma
PRODUÇÃO DE ÁCIDO GIBERÉLICO(GA3) A PARTIR DE CASCA DE CAFÉ
20: 20% glicose; 35: 35% glicose (controle); 35.L:com leucina: 35.0: com óleo de soja.a expressoem equivalente de etanol moI/ L gMS :b expresso em equivalente de etil acetato moI / LgMS.
o resíduo de casca de café tratada com vaporfluente é considerado um substrato adequado àprodução de aromas por C. fimbriata. A adição deglicose é necessária à esta proposta e aconcentração adequada compreende-se entre 20%e 35%. A adição de leucina aumentou em 58% aprodução de compostos voláteis totais.Comparando os resultados com outros daliteratura percebe-se que a produção total foi duasvezes superior aos melhores resultados obtidoscom bagaço de mandioca, senso que este haviasido adicionado de solução de leucina 15 vezesmais concentrada (3). Contudo, os compostosidentificados foram os mesmos em ambostrabalhos.
Tabela I: Total individual dos voláteis medidos noheadspace dos cultivos de C. jimbriata (dadosintegrados)Compostos 20 35 35.0 35.LAcetaldeídoa 47 57 58 50Etano! a 329 428 607 565Isopropanol a 26 5 - I 1EtiI acetato" 2078 1833 1841 2811Etil isobutirato o 11 10 7 15Isobutil acetato U 19 18 15 35
IEtil butirato U 9 3 I 21Propil acetato o 4 - - -Isoamilacetato U 2 1I - 162- heptanonao - - - 32-octanona a 52 67 3 30Etil 3 hexenoatob - - - 3

LPB-02 teve uma maior produção. As cepas LPB04 e LPB-OS apresentaram uma baixa produção deGA 3 para todos os substratos.Cromatografia Líquida de Alta Eficiência (HPLC)Os resultados da quantificação do GA 3 produzidopor fermentação da cepa LPB-06, obtidos porHPLC estão apresentados na Figura 1, a seguir:Os valores de concentração são dados em mgGA/L substrato para FSm e em mg GAlkgsubstrato para FES.
FIGURA 1; Produção de GA 3 pela cepa LPB-06,em diferentes sistemas de fermentação com cascade café.
- de G\3 emcasca re cate
-lO 40 3Il,r"l-e 35C~ 30-eo 25l~e;- 18,
~ 20.......;:
15~CJ;:
10oU
5",
O /
F&noo, FES= FESre;id
Os resultados acima mostram a possibilidade dautilização da casca de café como substrato paraprodução de GA 3 por fermentação no estadosólido, desde que feito um pré-tratamento dosubstrato.Estudos complementares estão realizados comvistas a otimização das condições de tratamentodo substrato e de fermentação quanto aparâmetros físicos e nutricionais, utilizando-seplanejamento experimental com análise porsuperfície de resposta. Com estas otimizaçõesespera-se aumentar a produção de ácidogiberélico.
CONCLUSÕES
Os resultados apresentados neste trabalhodemonstram as potencialidades para com odesenvolvimento de novas pesquisas que visam oaproveitamento dos residuos sólidos (casca, borra.polpa e fôlhas) da agroindustria do café. Essessub-produtos são ricos emmoléculas orgânicas que poderão sertransformadas em produtos de valor comercialimportante. Ressaltamos também que avalorização dos sub-produtos sólidos e líquidos da
327
agro-industria do café é uma forma recomendávele . moderna de proteção do meio ambiente,podendo propiciar ao produtor,· recursosfinanceiros complementares expressivos.sobretudo em períodos de baixa no preço do café.
AGRADECIMENTO
Carlos Ricardo Soccol agradece ao CNPq (bolsaprodutividade em pesquisa) e a União Européia(projeto INCa DC: N°CT970 185) pejo apoiofinanceiro recebido.
REFERÊNCIAS BlLIOGRÁFICAS
Bramorskl, A. Caracterização do crescimento eprodução de metabólitos voláteis por fungosfilamentosos cultivados sobre substratosagro indústriais. Curitiba, 1997, Dissertação (MScTecnologia Química), UFPr .Brahan, J. E.; Bressani, R. Pulpa de Café composicián, tecnologia y utilizacion. Bogota:Instituto de Nutrición de Centro América yPanama, INCAP, 1978.Brückne, 8.; Blechschmidt, D. The GibberellinFermentation. Criticai Rewiews inBiotechnology, 1991, II (2), 163-192.Christen, P et aI. BiotechnoJogy Letters, 1991,20(4) : 359-362.Christen, P et al. Biotechnology Letters, 1997,101(8): 911-919.Kumar, P. K. R. & Lonsane, 8. K. GibberellicAcid by solid state fermentation: consistent andimprovement yields. Biotechnology andBioengeneering, 1987,30,267-27 J.Janseens, L et aI., Process Biochemistry, 27: 195215,1992.Kurnar, P. K. R. & Lonsane, B. K. MicrobialProduction of Gibberell ins: state of the art.Advances in Applied Microbiology, 1989, 34, 29139.Hesseltine, C. W. Solid State Fermentations.Biotechnology and Bioengineering, 1972. 14.517-532.Pandey, A. Recents process development in solidstate fermentation. Process Biochemistry, 1992, 7,109-117.Senemaud, C. PhD Thesis, France,1998.Soccol, C. R. Contribuição ao estudo defermentação no estado sólido em relação àprodução do ácido fumárico, biotransformaçàode resíduo sólido de mandioca por Rhyzopus ebasidiomicetos do gênero Pleurotus. Curitiba,1994. Tese (Professor titular em Biotecnologia eTecnologia de Alimentos) - UFPr.

Soccol, C. R & Krieger, N. Advances inBuiotechnology, Edited by Educational Publishers& Distributors, New Delhi, June-July, 1998,pag.25-40.Sturza, R. Aproveitamento biotecnológico dosresíduos da extração do suco da maçã por3,203-206.
fermentação no estado sólido. Curitiba, 1995.Dissertação (MSc Tecnologia Química), UFPr .
.Tornasini, A.; Fajardo, C.; Barrios-Gonzáles, J.Gibberellic acid production using different solidstate fermentation systems. Word Journal ofMicrobyology and Biotechnology, 1997,1

329
EXPERIENCIA MEXICANA DE VALORIZACION BIOTECNOLOGICA DE SUBPRODUCTOSDE LA AGROINDUSTRIA DEL CAFE
Saucedo-Castaiieda G., Romano-Machado J.M. 1, Gutiérrez-Sánchez G., Delgado-Vidal F., RamírezRomero G. y Perraud-Gaime I. 2
Universidad Autónoma Metropolitana-lztapalapa. Ave. Michoacán y la Purísima. Iztapalapa, c.P. 09340.A.P. 55535. México O.F. Email:[email protected] ICIOCA. Via Blanca 804. A.P. 4026. Ciudad Habana, Cuba2IRD-México. Cicerón 609. Los Morales, c.r. 11530. México. O.F.
RESUMENLa utilizacion de los subproductos de café representa un reto importante para la integración dei procesohúmedo dei café en México, si son manejados adecuadamente puede representar beneficios econômicosimportantes.En los meses de recolección de la cereza dei café, la atención está orientada hacia el beneficio dei café y elmanejo de subproductos puede ser problemático. En el ciclo de 1997-98 se produjeron en México cerca de4.8 millones de sacos de 60 kg de café verde. La pulpa y el mucilago pueden Ilegal' a representar hasta el50% dei café cereza, de donde se puede concluir que el volumen de subproductos es enorme. Con elobjetivo de estabilizar la pulpa y conservar su valor nutricional, proponemos la realizacián de ensilaje bajocondiciones controladas. En esta etapa se pueden emplear inoculos de bacterias lácticas especializadaspara la pulpa de café, que permita una rápida formacion de ácido láctico )' evite la proliferacion demicroorganismos indeseables que den lugar a la formacion de etanoI )' ácido acético. El destino de la pulpaensilada adecuadamente podrá tener diversas aplicaciones. En este sentido existen alternativas queya hansido ensayadas con éxito como el composteo, lombricultura y produccián de setas comestibles, cZO'OSproductos finales se destinan a la agricultura y ai consumo humano.La pulpa ensilada es un material rico en nutrientes que puede ser utilizado para la alimentacion animal. sinembargo, su empleo presenta limitaciones debido a su alto contenido de cafeina y polifenoles.principalmente. En este respecto, proponemos realizar proceso de fermentacion en medio sólido de la pulpaensilada para disminuir considerahlemente el contenido de cafeína y polifenoles de la pulpa. Esta etapa serealiza con el empleo de hongos filamentosos (Penicillium y Aspergillus) capaces de utilizar la cafeinacomo fuente de nitrágeno y los polifenoles como fuente de carbono. bajo condiciones controladas defermentación. EI producto final de esta fermentación se puede destinar a animales monogástricos,poligástricos y a la acuacultura.La combinacián áptima de uttlizacion de los subproductos dei café deberá realizarse mediante estudlos deinvestigación de operaciones que consideran el desarrollo económico y tecnológico de los procesos.
INTRODUCCIÓN
A producción de café en el mundo es unaactividad comercial importante que da comoconsecuencia una derrama econórnica localizadaprincipalmente en países de América Latina. queen orden de producción son Brasil. Colombia yMéxico. El proceso que se utiliza en México yColombia es el lIamado beneficio húmedo, queutiliza cantidades variables de agua, mientras queen Brasil el proceso que se emplea de manera masextendida es el proceso seco. En el caso particularde México. Ia producción de café verde en 19981999 se estima cercana a 5 millones de sacos (60kg) de café verde. El grano de café representacerca dei 50 % del café cereza en base seca. Laproporción de pulpa en el café cereza puedecambiar de acuerdo ai estado de madurez de' la
cereza, condiciones climáticas y la variedad decafé cultivada. Sin embargo se puede aceptar quela pulpa y el mucilago representan cerca dei 30 y10 % dei café cereza, respectivamente. Estoimplica una cantidad muy importante de residuosa tratar es decir del orden de 1-0.8 millones deToneladas. concentrados en dos estados. Veracruz(23%) y Chiapas (34%). y en un temporada dealrededor de 6 meses.Por otra parte. Ia normatividad en respecto a lasdescargas en los beneficios están cambiandorápidamente. en el caso de México el 1 de Enerode 2000 entrará en vigor una norma que indicaque la descarga de los beneficios deberá tener unnivel de OBO menor a 1500 ppm. Esto haIlevado a los beneficiadores a esforzarse porreducir notablemente el consumo de agua en casiun 90%. Con el uso de las despulpadoras actuales

se llega a consumir 300-800 L por ton de cafécereza. Sin embargo, persiste el gasto de agua enel proceso de fermentación y lavado. Por otraparte. existe un esfuerzo importante de parte delas empresas constructoras de equipo deproporcionar desmucilaginadoras que operan conun consumo muy bajo de agua, generando unresiduo concentrado que es el mucilago, en formade un especie de miel densa y muy concentradaen sólidos solubles y suspendidos.Así mismo la fluctuación de los precios dei cafécereza ha sido importante en los últimos diezafios. con precios tan extremos que varían desde52 a 254 US dólares por saco de 100 libras.Actualmente la cotización dei saco de café verdeen la bolsa de Nueva York oscila alrededor de100 US dólares por saco. Esto puede sugerir laconveniencia de considerar los residuos de lacereza del café como una alternativa paraequilibrar la economía fluctuante de la industriacafetalera (Claridades Agropecuarias, 1999).Las alternativas que se han explorado paraaprovechar los residuos son muy variadas eincluyen la lornbricultura, producción de setascornestibles, alimentación de ganado, secado. etc.algunas de las cuales ya han sido presentadasampliamente en este evento por otrosconferencistas.Estas alternativas van de complejas a muysencillas. sin embargo algunas de estasalternativas no han sido evaluadas auneconómicamente. La solución tecnológica no esúnica. depende del desarrollo tecnológico delbeneficio. de la inversión a realizar y de surecuperación.Una de las alternativas estudiadas es la utilizaciónde la pulpa de café como alimento animal, que seve limitada a causa de su elevado contenido dehumedad, haciendo difícil su manejo. transporte yalmacenamiento; esta humedad, junto con sucontenido rico en nutrientes (azúcares. proteínas.aminoácidos) favorecen el desarrollo de sumicrotlora endógena, que en condicionesincontroladas puede afectar la cal idad delproducto. además de que es difícil suconservación. Además. en la pulpa de café estánpresentes sustancias con efectos anti fisiológicoscomo la cafeína, y antinutricionales como loscompuestos fenólicos que constituyen el principalfactor limitante para su utilización enalimentación animal. La información disponiblesobre los compuestos fenólicos de la pulpa decafé es escasa, más cualitativa que cuantitativa, yobtenida general mente a partir del materialfresco. En cerdos y ganado de carne. alimentados
330
con pulpa de café. se ha observado un aumento enla excresión de orina; niveles mayores de 30% dela pulpa en la ración pueden causar mortalidad enratas y pollos jóvenes. En el ganado se haobservado digestibilidad baja de la proteína.lesiones en la piel, pérdida de pelo y en algunoscasos la muerte cuando hay. consumos altos depulpa.Por otra parte, la Fermentacíón en Medio Sólidose define como el crecimiento demicroorganismos en materiales sólidos enausencia de agua libre. Sin embargo. el sustratopuede contener suficiente humedad parafavorecer el crecimiento microbiano y que existeen forma absorbida dentro de la matriz sólida(Pandey, 1992). EI origen de las fermentacionesen medio sólido no es reciente, el hombre las hautilizado desde hace mucho tiernpo, ya sea para lapreparación de alimentos fermentados o quesos.así como para el ensilaje (Moo-Young y col.,1983; Hesseltine, 1987).La FMS se propone como una alternativa en lastecnologías actualmente empleadas para lauti Iización y la valorización de productosagrícolas primarios. EI objetivo de numerosasfermentaciones sólidas alimentarias tradicionalesno es aumentar la cantidad de proteínas de novoen los alimentos. pero sí mejorar las posibilidadesde conservación. de transformar propiedadesfísicas u organolépticas. Por otra parte. estasfermentaciones permiten eliminar los factoresantinutricionales o antifisilógicos de ciertosproductos vegetales. En el caso de la pulpa decafé. su valor nutritivopuede mejorarse a travésde una fermentación sólida con hongosfi lamentosos empleando la cafeína como fuentede nitrógeno para su crecirniento, y posiblementelos taninos como fuente de carbono (Roussos ycol., 1989).EI objetivo de este trabajo es presentar losavances mas recientes en relación a ladegradación de la cafeína en la pulpa de café através de un proceso de fermentación en mediosólido.
MATERIALES Y METOOOS
Sustratos:Las muestras de pulpa fresca (PF) seobtuvieron un beneficio de café en la zona deJalapa, Veracruz, México durante la cosecha de1997-1998. Una parte de las muestras se congelóa -20 0c hasta su utilización. Un total de 220 kgde pulpa fueron ensilados con una cepa debacteria homoláctica a una concentración de 2.63

- 5.26 x 107 cel/g MH, obteniendo as' la pulpaensilada (PE).
Pasteurización: Las muestras de PF y PE, fueronpasteurizadas en corriente de vapor a presiónatmosférica (95 0c) durante I h. Se analizaroncuentas de UFC y humedad. Posteriormente sedejaron secar a temperatura ambiente por 24 h.
Desarrollo de los inóculos: Se utilizó una cepa dePenicillium com une (V33A25), aislada de la PCperteneciente a la colección IRD (exORSTOM) yUAM y que ha mostrado capacidad dedegradación de cafeína (Aquiahualt y col., 1988;Gaime-Perraud, 1995; Hakil y col., 1997;Roussos y col., 1995). La cepa se subcultivo a 300C durante 7 días. Los niveles de inóculoprobados fue de 104, 105, 106, 107 y 108 esporasg-I de sólido húmedo.
Condiciones' de fermentaciôn: Las experienciasde FMS se realizaron en el sistema descrito porRaimbault y Alazard en 1980, a una densidadaparente de 0.6 g cm-3, 30 0C y con un tlujo deaire húmedo de 0.5 vkgm (dm3 de aire * kg- 1 demedio * mirr l ).
Determinaciones analíticas: Las muestras sólidasfueron moi idas con picador Braun por 20 seg yextraídas con agua en agitación, 20 min, I: 1O(p/p). EI pH se midió en la suspensión resultantecon un potenciómetro. Las suspensiones fuerondecantadas en centrífuga Hettich a 5000x g por 20min y los sobrenadantes se conservaron a -20 0Chasta su análisis. EI contenido de humedad de lasrnuestras sólidas se determinó por gravimetría.La concentración de ácidos grasos volátiles(AGV'S), glucosa y fru~tosa se realizó por HPLCen un cromatógrafo Perkin Elmer con lascondiciones siguientes: columna Rezek OrganicAcid, fase móvil H2S04 30 mM, tlujo 0.6 cm 3
mirr l , temperatura 50 0C, detector de Índice derefracción.La cafeína se extrajo de las muestras sólidas secasy trituradas y tamizadas (malla 35) con unamezcla de cloroformo/ isopropanol (50:50 v/v).La cafeína en los extractos se determinó por
, respectivamente. Con. Ia disminución dehumedad se evitan los riesgos de escurrimientosen la FMS.
331
HPLC, empleando una modificación a la técnica, reportada por Denis en 1996.
Determinación de C02 por cromatografía de
gases: Se empleó un cromatógrafo de gases GowMac 580, equipado con una columna concéntricaAlItech (CTR 1) y un detector de conductividadtérmica de acuerdo a lo reportado por SaucedoCastafieda y co 1.,' 1994.
Conteo de microorganismos: Se utilizó medioAgar para Métodos Estándar (Bioxón) para losconteos de microorganismos totales y PDA(Bioxón) con antibiótico para conteo de hongos ylevaduras. Los conteos se real izaron a las 24 h deincubación a una temperatura de 30 oCo Losresultados se expresaron como UFC g -I demateria seca (Perraud-Gairne, 1995).
RESULTADOS Y DISCUSION
Pasteurización de la PFy PEComo se aprecia en la Figura I la carga inicial demicroorganismos totales y de hongos y levadurases 2 veces superior en la PF que en la PE. Durantela pasterización de la PE y la PF se .obtuvo unareducción completa de los microorganismospresentes a los 20 min de exposición a corrientede vapor directo para ambos sustratos. Estopermitió reducir la carga contaminante de la PCen un corto tiempo y estandarizar las condicionesiniciales para la FMS por esto se decidió hacer lapasteurización por 20 min a los sustratosuti Iizados en las experiencias posteriores.Por otra parte la pasteurización no provocó unaumento apreciable en elcontenido de agua de lossustratos tratados (Figura 2) sino que se mantuvoconstante durante entre 78 y 83 % de humedaddurante todo el proceso de pasteurización. Noobstante los sustratos se mantienen con un altocontenido de agua ai final dei tratamiento térmicolo que puede provocar pérdidas de agua ynutrientes por escurrimiento durante la FMS. Paraevitar este contratiempo se dejaron secar atemperatura ambiente por 24 h y se logró reducirlos niveles de humedad hasta 74.15 y 75.43 % enPE y PF
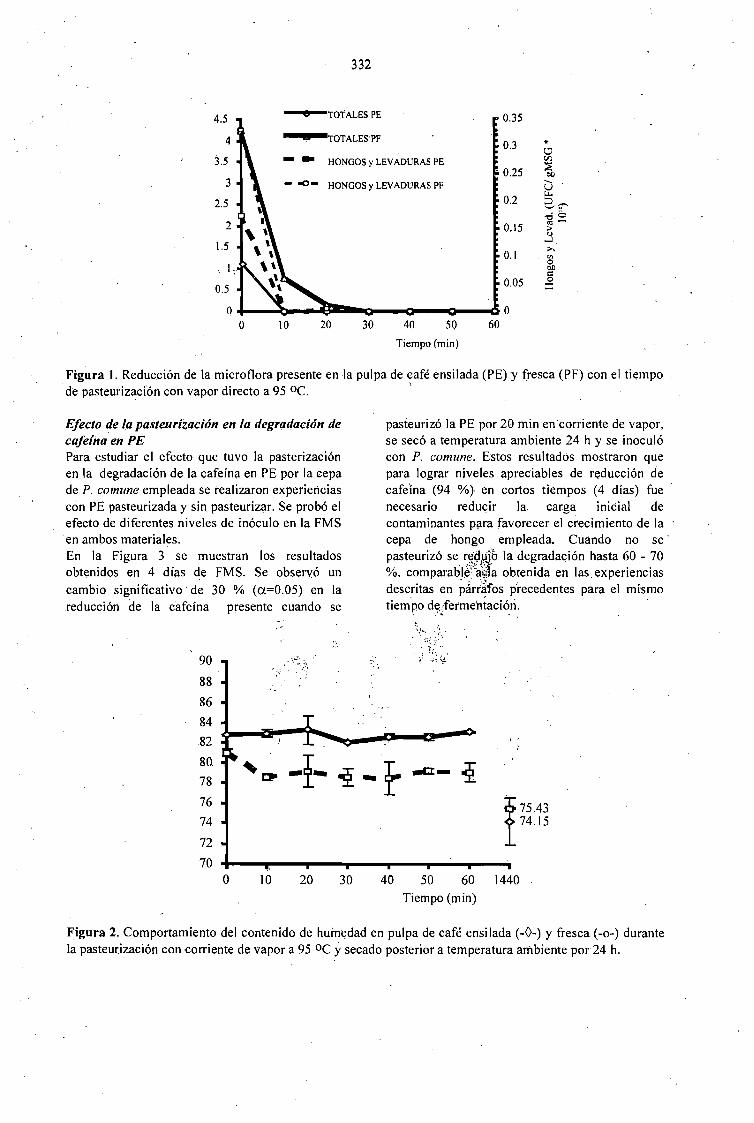
332
0.35
0.3 *oV'l
0.25 :2O/)
<,
u0.2
tJ-
2.~'Õ
0.15"g-;-O.l
...J
0.1>..
'"oO/)c
0.05 o:I:
O60504030
HONGOSyLEVADURASPF
HONGOSyLEVADURASPE
20
e
s
- 00-
-·-TOTALES',PF
--
10
4.5
4
3.5
3
2.5
2
1.5
1.,.
0.5
OO
Tiempo (min)
Figura 1. Reducción de la microflora presente en la pulpa de café ensilada (PE) y fresca (PF) con el tiempo\
de pasteurización con vapor directo a 95 0e.
Efecto de la pasteurizaciân en la degradaciõn decafeína en PEPara estudiar el efecto que tuvo la pasterizaciónen la degradación de la cafeína en PE por la cepade P. comune empleada se realizaron experienciascon PE pasteurizada y sin pasteurizar. Se probó elefecto de diferentes niveles de inóculo en la FMS
'en ambos materiales.En la Figura 3 se muestran los resultadosobtenidos en 4 días de FMS. Se observó un
cambio significativo' de 30 % (0.=0.05) en lareducción de la cafeína presente cuando se
pasteurizó la PE por 20 min encorriente de vapor,se secó a temperatura ambiente 24 h y seinoculócon P. comune. Estos resultados mostraron quepara lograr niveles apreciables de reducción decafelna (94 %) en cortos tiempos (4 dias) fuenecesario reducir la carga inicial decontaminantes para favorecer el crecimiento de la
.cepa de hongo empleada. Cuando no Sepasteurizó se r~i:I~jb la degradación hasta 60 - 70%. comparabl~~Y~~fa obtenida en las. eXperienciasdescritas en páré~tos precedentes para el misrnotiernpo defermentacióri.
. ",''li -
90
88
86
84
82
80
78
76
74
72
70o \O 20 30
f. 75.43174.15
40 50 60 1440
Tiempo (min)
Figura 2. Comportamiento delcontenido de humedad en pulpa de café ensilada (-0-) y fresca (-0-) durantela pasteurización concorriente de vapor a 95 0C y secado posterior a temperatura ambiente por 24 h.
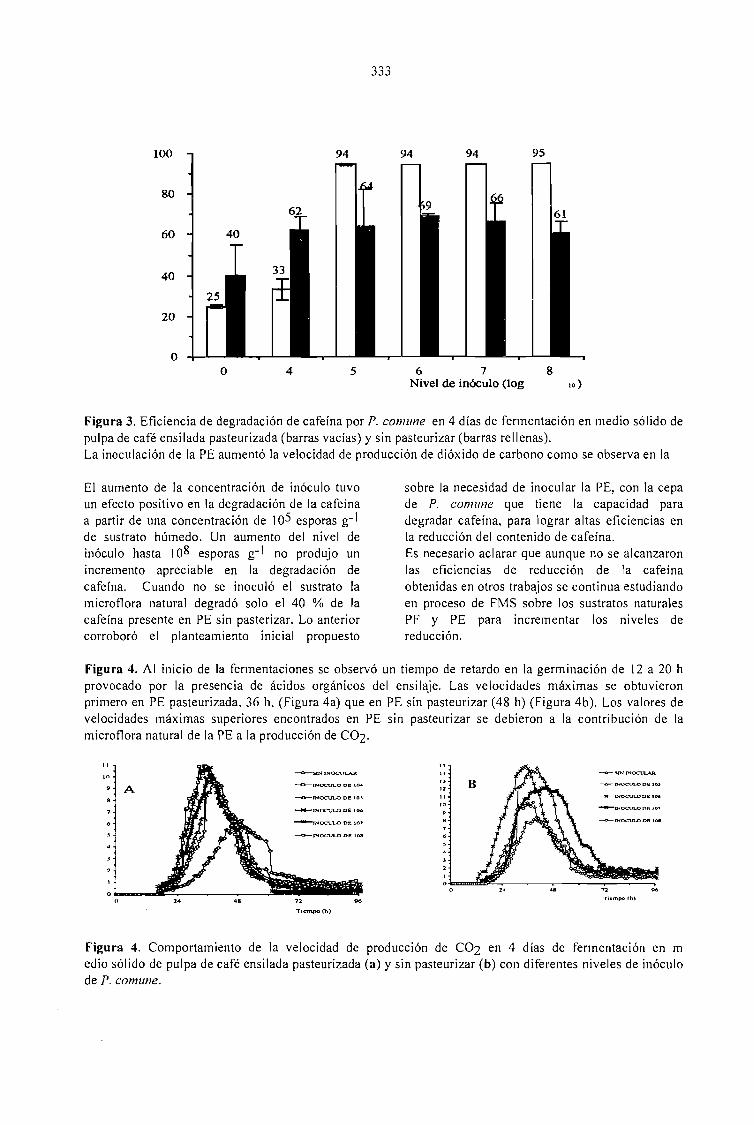
100 94
333
94 94 95
80
60
40
20
oo 4 5 6 7
Nivel de in6culo (log8
10)
Figura 3. Eficiencia de degradación de cafeína por P. comune en 4 días de fermentación en medio sólido depulpa de café ensilada pasteurizada (barras vacías) y sin pasteurizar (barras rellenas).La inoculación de la PE aumentó la velocidad de producción de dióxido de carbono como se observa en la
EI aumento de la concentración de inóculo tuvoun efecto positivo en la degradación de la cafeínaa partir de una concentración de 105 esporas g-lde sustrato húmedo. Un aumento dei nivel deinóculo hasta 108 esporas g-! no produjo unincremento apreciable en la degradación decafeína. Cuando no se inoculó el sustrato lamicroflora natural degradó solo el 40 % de lacafeína presente en PE sin pasterizar. Lo anteriorcorroboró el planteamiento inicial propuesto
sobre la necesidad de inocular la PE, con la cepade P. com une que tiene la capacidad paradegradar cafeína. para lograr altas eficiencias enla reducción dei contenido de cafeína.Es necesario aclarar que aunque no se alcanzaronlas eficiencias de reducción de la cafeínaobtenidas en otros trabajos se continua estudiandoen proceso de FMS sobre los sustratos naturalesPF y PE para incrementar los niveles dereducción.
Figura 4. AI inicio de la fermentaciones se observó un tiempo de retardo en la germinación de 12 a 20 hprovocado por la presencia de ácidos orgánicos dei ensilaje. Las velocidades máximas se obtuvieronprimero en PE pasteurizada. 36 h. (Figura 4a) que en PE sin pasteurizar (48 h) (Figura 4b). Los valores develocidades máximas superiores encontrados en PE sin pasteurizar se debieron a la contribución de lamicroflora natural de la PE a la producción de C02.
11
10
9A
Tiempo (h)
Figura 4. Comportamiento de la velocidad de producción de C02 en 4 días de fermentación en medio sólido de pulpa de café ensilada pasteurizada (a) y sin pasteurizar (b) con diferentes niveles de inóculode P. comune.

CONCLUSIONES
EI ensilado de la PC favoreció la degradación dela cafeína de la pulpa de café por la cepa de P.comune utilizada. La pasteurización mejoró laeficiencia degradativa dei hongo por laeliminaeión de eontaminantes. La inoeulaeión dela PE eon niveles de 106 esporas g-l de sustratohúmedo aumentó significativamente la eficieneiade degradaeión de la cafeína presente.Los resultados obtenidos mostraron la viabilidadde utilizar la FMS para reducir el contenido decafeína de la PC sin alterar las características deIsustrato y con la posibilidad de poder utilizarprimero el ensilado como un método sencillo deconservación para después degradar la cafeína dela PE bajo condiciones controladas de FMS.Además se establecieron estas condiciones lo quepermitirá enfrentar el escalamiento dei proceso.
AGRADECIMIENTOS
Proyecto financiado por la Comunidad Europeacontrato IC 18 * CT 970 J85
REFERENCIAS BIBLIOGRAFICAS
Aquiahualt M.A.. Roussos S. y Trejo M.R.(1988). Coffee pulp detoxification by solid stateferrnentation: Isolation identification andphysiological sudies. Proceedings of the seminal'Solid State Fermentation in Bioconversion ofAgroindustrial Raw Materials, M. Raimbault,ORSTOM (Eds.). Montpellier, France, 13-26.Anónimo (1999). Coffee supply and demandbalance. Edición especial de: ClaridadesAgropecuarias, México, H. Fanghanel y C.Montaãez (Eds.). pp 21-36.
Denis S. (1996). Degradation de la cafeine parAspergillus sp. et Penicillium sp.: Etudephysiologique et biochimique. Thêse de Doctorat,Université Montpellier 11,180 p.Gaime-Perraud I. (1995). Cultures mixtes en
milieu solide de bactéries lactiques et dechampignons filamenteux pour la conservation etla decafeination de la pulpe de cafe. Thêse deDoctorat, Université Montpellier 11.210 p.
334
Hakil M.. Denis S., Viniegra-González G. yAugur C. (1998). Degradation and productanalysis of caffeine and related dimethylxanthinesby filamentous fungi. Enzyme and Microb.Technol.. 22: 355-359.Hesseltine C W. (1987). Solid state fermentation
- An overview. Int. Biodeterioration, 23: 79-89.Moo-Young M. A., Moreira R. y Tengerdy R. P.
(1983) Principies of solid substrate fermentationof the filamentous fungi. En: FungaIBiotechnology, S. E. Srnith, D. R. Berry y B.Vristiunsen (Eds.), Arnold, Londres. Vol.IV: 117174.Pandey A. (1992). Recent process development insolid state fermentation. Process Biochemistry,27:109-117.Raimbault M. y Alazard D. (1980). Culturemethod to study fungai growth in sol idfermentation. Europ. louro of Appl. Microbiol.and Biotechnol, 9: 199-209.Roussos S., Aquiahualt M. A., Cassaigne J.,Favela e., Gutiérrez M., Hannibal L., HuertaS., Nava G., Rairnbault M., Rodríguez W.,Salas J. A., Sánchez R., Trejo-Hernández M.R. y Viniegra-González G. (199).Detoxificación de la pulpa de café porfermentación sólida. Resúmenes I-SIBAc'Jalapa, México, 12-15 Abri I. Parte 11: 121143.Roussos S.. Aquiahualt M. A., Trejo-Hernández
M. R.. Gairne-Perraud 1., Favela E., RamakrishnaM.. Raimbault M. y Viniegra-González G.(1995). Biotechnological management of coffeepulp-isolation, screening, characterization.seleetion of caffeine-degrading fungi and naturalmicroflora present in coffee pulp and husk. Appl.Microhiol. Biotechnol.. 42: 756-762.13 Saucedo-Castafieda G., Trejo-Hernández M.R.. Lonsane B. K., Navarro J. M.. Roussos S. yRaimbault M. (1994). On-line autornatedmonitoring and control system for C02 and 02 inaerobic and anaerobic solid-state fermentations.Process Biochemtstry, 29: J 3-24.

335
LOMBRICOMPOSTAJE DE LA PULPA DE CAFÉ EN MÉXICO
Eduardo Aranda-Delgado, Isabelle Barois
DEPARTAMENTO DE BIOLOGÍA DE SUELOS, INSTITUTO DE ECOLOGÍA, A.C., Apdo. Postal 63.Xalapa, Ver. México. E-Mail: [email protected]
INTRODUCCIÓN
México fué el primer país en proponer y utilizardesde 1985, la Ecotecnología deILombricompostaje para la transformación de lapulpa de café en abono orgánico (Aranda, 1985,1989, 1991a, 1991 b). En este sentido. Ia prácticadeI lombricompostaje podría definirse como lacombinación de procesos biológicos, disefíos ytécnicas utilizadas para cultivar intensiva ysistemáticamente cierto tipo de lombrices epígeas, para acelerar la estabilización de los resíduosorgánicos. Los cuales son ingeridos, moIidos ydigeridos por las lombrices con la ayuda demicroorganismos aeróbicos y de ésta manera seconvierten en un abono granuloso,"Vermicornpost o humus de lombrices"; unmaterial humificado, activo microbiológicamente,con excelente porosidad, aereación y capacidadde retención de agua, rico en nutrientesdisponibles, hormonas, enzimas ymicroorganismos útiles deI suelo. Por éstascaracterísticas, el producto es un medio valioso,superior y comercializable como acondicionadordeI suelo y sustrato deI crecimiento para lasplantas (Edwards and Bohlen, 1996, Aranda et
ai.. 1999; Edwards (en prensa) ).Con la ayuda de las lombrices, la pulpa de café(obtenida por el Beneficiado Húmedo) y muchosotros resíduos orgánicos, pueden sertransformados eficientemente en vermicompostde utilidad para los suelos, la producción agrícolay también para el medio urbano. Los nutrientes enel vermicompost se encuentran fácilrnentedisponibles para las plantas y pueden seradicionados aI suelo agrícola para mejorar suferti lidad y estructura o ser usado como aditivo dealta cal idad comercial como sutrato para macetasy medio de crecimiento intensivo para las plantas.
EL PROBLEMA DE LA PULPA DEL CAFE
La disposición de los resíduos orgánicos es unode los principales problemas en las unidades deprocesamiento de café o "Beneficios Húmedos".En el procesamiento húmedo tradicional, songenerados aproximadamente tres toneladas de
subproductos y son requeridas cuatro toneladas deagua para producir una tonelada de café oro seco(Rolz et al., 1982). La Pulpa dei café es el mayorde estos resíduos de la agroindustria dei café y seproduce cuando las dos semillas son extraidas deifruto fresco por las "despulpadoras", La pulparepresenta alrededor dei 39 % deI peso fresco deIfruto, o el 28.7 % en base seca (Zuluaga.1989)Esta pulpa fué durante mucho tiempo arrojada engrandes pilas en barrancos o cerca de los ríoscausando contaminación dei suelo y agua.La producción anual para México se estima en661,000 toneladas (Consejo Mexicano dei Café,1997) y de acuerdo con los datos proporcionadospor Zuluaga (1989) dei promedio de producciónmundial (85 millones de sacos de 60 kg, 75 %café arábigo), aproximadamente 8.057 millonesde toneladas es producida anualmente en elmundo.Aunque se hicieron grandes esfuerzos para usar lapulpa dei café fresca para la ai imentación deanimales (Braham and Bressani, 1978), loscontenidos de polifenoles, taninos, acidosclorogénicos. ferúlicos y caféicos en la pulpa(Roussos et ai., 1989), obstruyen la viabil idadpara alimentación animal.En muestreos de macrofauna realizados engrandes acumulaciones de pulpa de caféabandonada en campo en México, se encontraronpoblaciones activas de macrofauna de grupostales como Diptera, Acarida, Oligochaeta,Coleoptera, Thysanoptera y Collernbola, entreotras. Las lombrices mas comunmenteencontradas en tales acumulaciones de pulpa decaféfueron Perionyx excavatus (Perrier, 1872) enel estado inicial. o Amynthas gracilis (Kinberg,1867) y Dichogaster sp. en los estados masavanzados, mientras que Eisenia fetida ó E.andrei fueron raramente encontradas (Aranda yArel1ano, 1995).Sin embargo, en todas las acumulacionesobservadas, fué evidente que la proliferación de lamacrofauna solo alcanzó las capas massuperficiales, alrededor de los 5 a 30 emsolamente, mientras que las capas mas internas. Iapulpa se torna amarillenta (cuando es fresca esroja brillante) acida (3-4) y caliente (hasta 65 C ).Sin oxígeno ni macrofauna activa presente

(Gairne-Perraud et al.. 1991 a, 1991 b, GaimePerraud, 1995); este material inactivo,rústicarnente "ensilado"solo reirucra sudescornposición orgamca, hasta que es
fisicamente removido y aereado. Con esta claraevidencia, puede ser concluido que la pulparequiere de un eficiente y efectivo método detratamiento para promover y completar sudescornposición orgánica y favorecer suaprovechamiento. La estabilización de la pulpadeI café con lombrices composteadoras puedecontribuir a resolver un importante problema decontarninación ambiental, modi ficando estosaspectos negativos en materiales útiles yproductivos.
EL PROCESO DE VERMICOMPOSTAJE
A diferencia de otros métodos de compostaje, elVerrni-cornpostaje toma ventaja de las cualidadesbiológicas y fisiológicas de las lornbricescomposteadoras, para estimular la
descomposición rnicrobiana aeróbica de losresíduos orgánicos: de hecho la asirnilación de laslombrices se deriva de los microorganismos queellas estirnulan en la materia orgánica. Lassiguientes son algunas de las propiedades que hansido reconocidas (Van Gansen, 1962; Satchell,1983; Tornati et al., \987; Edwards andNeuhauser, 1988; Edwards and Boh len, 1996;Aranda et al., 1999) y que se resurnen en Aranday Bares (en prensa): 1.- Las lornbrices fabricangalerias en la materia orgánica, 10 que favorece suaereación y ventilación, 2.-Recubren las paredesde las galerias con un muco gelatinoso quefavorece el desarrollo de microorganismos, 3.Con su aparato digestivo las lornbrices trituran yrnuelen la materia orgánica, 4.- Reducen oeliminan microorganismos patógenos de plantas yanimales, 5.- Durante su paso por el tractodigestivo, mezclan los diferentes componentes dela materia orgánica, 6.- Cada turrículo o excreta,se recubre de una membrana gelatinosa,rnucoprotéica, la "membrana peritrófica", que dacohesión y 50porte a la materia orgánica
procesada, 7.- AI colocar sus excretassuperficialmente, cubren la rnateria orgánica másfresca, 10 que abate la generación de olores y laatracción de insectos, 8.- Durante el proceso segeneran importantes cornpuestos orgánicos
reguladores como enzimas, antibióticos,vitaminas. horrnonas y sustancias húmicas. 9.
Los turrículos (heces o casts) conservan en elsuelo su estructura y porosidad, 10 que asegurauna liberación lenta y regulada de los nutrientes,
336
evitando pérdidas por erosión, lavado o lixiviado,10.- Por afiadidura, en cada ciclo se producen máslombrices, lo que permite aumentar la capacidado aprovecharlas como proteínas de alto valorbiológico.
VERMICOMPOSTAJE DE LA PULPA DELCAFÉ
La comunidad científica internacional y losprogramas de investigación han dado a laVerrnicultura un análisis serio crítico y científico,estableciendo el potencial efectivo que laVerrnicultura representa (Bouché.1987; Satchell,1983; Bonvicini-Pagliai y Omodeo,1987;
Edwards y Neuhauser, 1988; Kretzchmar. 1992;Hoerschelrnann y Andrés, 1994; Edwards, 1997).
Para desarrollar un exitoso cultivo delornbrices con pulpa de café y producir uneficiente bioabono para las plantas, sonrequeridos varios componentes: I) especiesadecuadas de lornbrices, 2) Una apropiada calidad
y conservación deI sustrato, c)correctascondiciones ambientales externas e internas aIcultivo y d) un eficiente disefío y operación deicultivo.
Especies de LomhricesLas especies mas com unes de lornbrices usadasen todo el mundo para el manejo de los resíduosorgánicos incluyen a Eisenia andrei (Bouché,1972) ó lornbriz roja y su especie cercana E.
fétida (Savigny, 1826) lornbriz tigre o bandeada,
ambas alguna vez confundidas como E. foetidahasta su aclaración completa (Jaenike, 1982).Ambas son lornbrices resistentes. con arnpliosmárgenes de tolerancia a la temperatura, PH yhumedad, En cultivos mixtos (con otras especies)Eisenia andrei usualmente se Ilega a hacerdominante (Edwards y Boh len, 1996; Salazar etal.. 1995).Muchos sitios de Verrnicompostaje en México(con E. andrei) tienen tarnbién poblacionesinvoluntarias de Perionyx excavatus (Perrier,1872), la Lombriz Oriental de las compostas, que
se iniciaron a partir de poblaciones silvestres.Esta especie de distribución tropical puede serutilizada satisfactoriamente en campo y enalgunas circunstancias puede incluso Ilegar asuperar a las poblaciones inoculadas de E. andrei.Es una especie común en Asia y es usadaextensivamente en Filipinas y Australia (Edwardsy Boh len, 1996; Guerrero, 1983).

Propiedades Biológicas y Demograficas de lasLombricesLas lombrices tienen un ciclo de vida muysim pie. siempre en el interior de la matériaorgánica. Una de las razones por la cual laslornbrices se multiplican tan rápidamente es porser hermafroditas. por lo cual todos los individuosposeen genitales masculinos y femeninos; aún asíse requiere la copulación entre dos individuos.Todos los individuos producen cocoones ocapullos, dentro de los cuales puedendesarrollarse de 1-5 individues, dependiendo dela especie, dei crecimiento de los padres y deItarnafío de los capullos.Aunque en condiciones controladas (Arellano,1997) pueden ser obtenidas densidades de 50.000individuos por m2 (con 3.540 g m-2 ) de Eandrei en pulpa de café. para aspectos prácticosen condiciones de campo, es posible asumirbuenos valores de densidad poblacional enalrededor de 20- 25 mil individuos por m2. EIequivalente de esta cantidad de lombrices es unabiomasa equivalente alrededor de 2,500-3,000 gm-2. es decir un promedio (considerando adultosy juveniles juntos) de aprox. 0.110 g por lombriz.Mientras mas bajo es el valor de densidadpoblacional, mas alto es el valor dei pesopromedio individual de las lombrices; aicontrario, una población mas altamenteconcentrada. su crecimiento individual. su tasa dereproducción y algunos otros aspectosdisminuyen en relación a los ejernplaresindividuales. Si consideramos (solo hasta ciertopunto) que más biomasa puede ser alcanzada conmás indivíduos. (aunque cada uno de ellos estémenos desarrollados), entonces la Capacidad deCarga puede ser entendida como el valor àptimode densidad poblacional en la cual se obtiene unamáxima producción de biomasa en unadeterminada area o volúmen (estimada en 532.4g m-2 en P. excavatus, y 3,371.1 g m-2 en Eandrei, contra un valor de 926 g m-2 de E andreien estiercol de cabal lo (Arellano, 1997). Estarelación de densidad-dependencia tambiénimplica que la mejor manera de prevenircondiciones estancadas de sobrepoblación, esextrayendo sistemáticamente lombrices de lascamas de vermicompostaje para extender elcultivo o como fuente de proteínas para laalimentación de animales (Taboga, 1980;Hartenstein, 1981; Tacon et ai., 1983; Velásquezet al.. 1986; Guerra, 1982). •
337
La Pulpa de Café como SustratoLa pulpa del café tiene una alta capacidad deretención de agua (cercana a 4 veces su propiopeso seco). una buena densidad aparente(alrededor de 0.500 kg 1-1 cuando fresca y 0.700kg l-I cuando es aplicada a las lornbrices), unpotencial de oxido-reducción alrededor de 450mv, pH practicamente neutro (7.4) Y un tarnafíouniforme y manejable. En relación a su contenidode nutrientes. Ia pulpa de café no requiereaditivos ni extracciones o modificaciones previaspara poder ser utilizada para la alimentación delornbrices, es un sustrato "I irnpio", Iibre decontaminantes. Comparada con otros resíduosorgánicos, la pulpa de café puede ser consideradacomo uno de los mejores residuos orgánicos(originados de plantas) para cultivar lombrices.
Factores AmbientalesCondiciones óptimas generales para cultivarlombrices han sido presentadas para E fetidaEandrei por Lofs-Holmin (1985), Edwards andNeuhauser (1988). quienes sugieren que estascondiciones no difieren mucho de aquellaspresentes para otras especies similares:temperatura. 15-20 C (limites 4-30 C); humedad,80-90% (limites 60-90%); aerobicidad, >15 % 02; contenido de C02 <6%; potencial Redox, Eh >100mV; contenido de amonia bajo.<0.5%;contenido de sales bajo, <0.5%; y un pH >5 y <9.Uno de los factores Iimitantes ai proveer resíduosorgánicos a las lornbrices, es la temperatura quese puede generar por los microorganismos si secolocan capas gruesas en las camas deverrnicornpostaje, pues temperaturas >30-35 Cpueden hacerlas escapar del medio o inclusomatarlas. Para evitar tales sobrecalentamientos, esnecesario adicionar capas relativamente delgadas(aprox. 10 em) y permitir que las lombrices lasprocesen en condiciones aeróbicas. Cuando elsustrato está cerca de ser totalmente consumido.otra nueva capa de sustrato debe ser adicionada.evitando dejar a las lombrices sin alimento.Alimentaciones quincenales son el periodo maspracticado, sin embargo siempre verificando porobservación directa la necesidad de aJimentación.
Conversiôn Pulpa- VermicompostSi comparámos el contenido de nutrientes de lapulpa fresca con el producto resultante deiverrnicompostaje, podemos reconocer que ésteúltimo contiene por lo general aproximadamenteel doble de los porcentajes de cada elementoquímico (excepto para el carbono) (Tabla 1).Esto puede ser explicado en términos de la
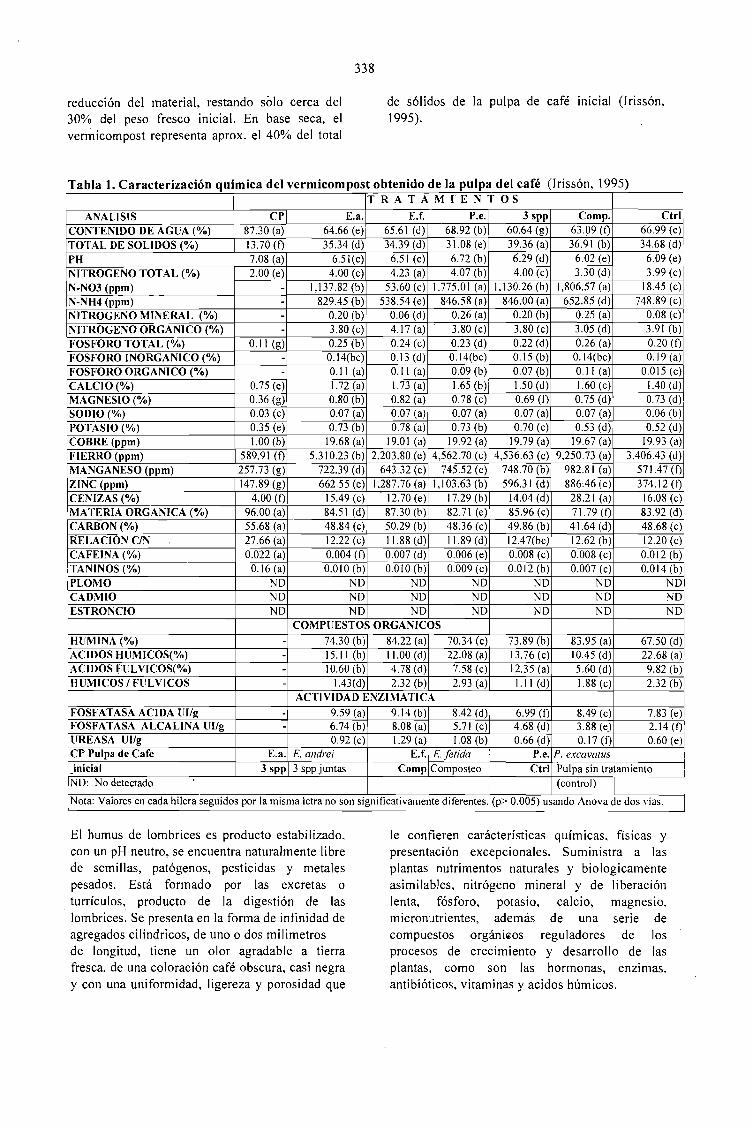
reducción dei material, restando solo cerca deI30% deI peso fresco iniciai. En base seca, elverrnicompost representa aprox. el 40% dei total
338
de sólidos de la pulpa de café inicial (Irissón,1995).
Tabla 1. Caracterización química deI vermícompost obtenido de la pulpa deI café (Irissón, 1995)TRATAMIENTOS
ANALlSIS CP E.a. E.f. P.e. 3 spp Comp. Ctrl
CONTENIDO DE AGlJA (%) 87.30 (a) 64.66 (e) 65.61 (d) 68.92 (b) 60.64 (g) 63.09 (f) 66.99 (c)
TOTAL DE SOLlDOS (%) 13.70 (f) 35.34 (d) 34.39 (d) 31.08 (e) 39.36 (a) 36.91 (b) 34.68 (d)
PH 7.08 (a) 6.51(c) 6.51 (c) 6.72 (b) 6.29 (d) 6.02 (e) 6.09 (e)
NTTROGENO TOTAL (%) 2.00 (e) 4.00 (c) 4.23 (a) 4.07 (b) 4.00 (c) 3.30 (d) 3.99 (c)
N-N03 (ppm) - 1,137.82 (b) 53.60 (c) 1.775.01 (a) 1.130.26 (b) 1,806.57 (a) 18.45 (c)
N-NH4 (ppm) - 829.45 (b) 538.54 (e) 846.58 (a) 846.00 (a) 652.85 (d) 748.89 (c)NTTROGENO MINERAL (%) - 0.20 (b) 0.06 (d) 0.26 (a) 0.20 (b) 0.25 (a) 0.08 (c)'
NITROGENO ORGANICO (%) - 3.80 (c) 4.17 (a) 3.80 (c) 3.80 (c) 3.05 (d) 3.91 (b)FOSFORO TOTAL (%) 0.11 (g) 0.25 (b) 0.24 (c) 0.23 (d) 0.22 (d) 0.26 (a) 0.20 (t)
FOSFORO INORGANICO ("lo) - 0.14(bc) 0.13 (d) 0.14(bc) 0.15 (b) 0.14(bc) 0.19 (a)
FOSFORO ORGANICO (%) - 0.11 (a) 0.11 (a) 0.09 (b) 0.07 (b) O.I! (a) 0.015 (c)CALCIO(%) 0.75 (e) 1.72 (a) 1.73 (a) 1.65 (b)' 1.50 (d) 1.60 (c) 1.40 (d)MAGNESIO (%) 0.36 (g) 0.80 (b) 0.82 (a) 0.78 (c) 0.69 (f) 0.75 (d) 0.73 (d)SODIO (%) 0.03 (c) 0.07 (a) 0.07 (a) 0.07 (a) 0.07 (a) 0.07 (a) 0.06 (b)POTASIO (%) 0.35 (e) 0.73 (b) 0.78 (a) 0.73 (b) 0.70 (c) 0.53 (d) 0.52 (d)1
,COBRE (ppm) 1.00 (b) 19.68 (a) 19.01 (a) 19.92 (a) 19.79 (a) 19.67 (a) 19.93 (a)FlERRO (ppm) 589.91 (f) 5.310.23 (b) 2.203.80 (e) 4,562.70 (c) 4,536.63 (c) 9,250.73 (a) 3.406.43 (d)MANGANESO (ppm) 257.73 (g) 722.39 (d) 643.32 (e) 745.52 (c) 748.70 (b) 982.81 (a) 571.47 (f)
ZINC (ppm) 147.89 (g) 662.55 (e) 1.287.76 (a) 1,103.63 (b) 596.31 (d) 886.46 (c) 374.12(1)CENIZAS (%) 4.00 (f) 15.49 (c) 12.70 (e) 17.29 (b) 14.04 (d) 28.21 (a) 16.08 (c)MATERIA ORGANICA ("lo) 96.00 (a) 84.51 (d) 87.30 (b) 82.71 (e) 85.96 (c) 71.79 (f) 83.92 (d)CARBON (%) 55.68 (a) 48.84 (c) 50.29 (b) 48.36 (c) 49.86 (b) 41.64 (d) 48.68 (c)RELACION CtN 27.66 (a) 12.22 (c) 11.88 (d) 11.89 (d) 12.47(bc) 12.62(b) 12.20 (c)CAFEINA (%) 0.022 (a) 0.004 (f) 0.007 (d) 0.006 (e) 0.008 (c) 0.008 (c) 0.012 (b)iTANINOS (%) 0.16 (a) 0.010 (b) 0.010 (b) 0.009 (c) 0.012 (b) 0.007 (c) 0.014 (b)1PLOMO NO NO NO NO NO NO NOCADMIO NO NO NO NO NO NO NOESTRONCIO NO NO NO NO NO NO NO
COMPlJESTOS ORGANICOS
HllMINA (%) - 74.30 (b) 84.22 (a) 70.34 (c) 73.89 (b) 83.95 (a) 67.50 (d)ACIDOS HlIMICOS(%) - 15.11 (b) 11.00 (d) 22.08 (a) 13.76 (c) 10.45 (d) 22.68 (a)ACIDOS FULVICOS(%) - 10.60(b) 4.78 (d) 7.58 (c) 12.35 (a) 5.60 (d) 9.82 (b)HlJMICOS / FlILVICOS - 1.43(d) 2.32 (b) 2.93 (a) 1.11 (d) 1.88 (c) 2.32 (b)
ACTIVIDAD ENZIMATICA
FOSFATASA ACIDA lJl/g - 9.59 (a) 9.14 (b) 8.42 (d) 6.99 (f) 8.49 (c) 7.83 (e)FOSFATASA ALCALINA lJl/g - 6.74 (b) 8.08 (a) 5.71 (c) 4.68 (d) 3.88 (e) 2.14 (f)UREASA lJl/g 0.92 (c) 1.29 (a) /.08 (b) 0.66 (d) 0.17 (t) 0.60 (e)CP Pulpa de Cafe E.a. E andrei E.f. Efetida P.e. P. excavatus I
-inicial 3 spp 3 spp juntas Comp Composteo i Ctrl Pulpa sin tratarnientoNO: No detectado . (control)
I
INata: Valores en cada hilera seguidos por la misma letra no son significativamente diferentes. (p> 0.005) usando Anova de dos vias.
EI humus de lornbrices es producto estabilizado.con un pH neutro, se encuentra naturalmente librede semillas, patógenos, pesticidas y metalespesados. Está formado por las excretas o
. turrículos, producto de la digestión de laslombrices. Se presenta en la forma de infinidad deagregados cilíndricos, de uno o dos milimetros
de longitud, tiene un olor agradable a tierrafresca, de una coloración café obscura, casi negra
y con una uniformidad, ligereza y porosidad que
le confieren carácterísticas qutrrucas, físicas ypresentación excepcionales. Suministra a lasplantas nutrimentos naturales y biologicamenteasirnilables, nitrógeno mineral y de liberaciónlenta, fósforo, potasio, calei o, magnesio,
micronutrientes, además de una serie decompuestos orgánieos reguladores de losprocesos de crecimiento y desarrollo de lasplantas, como son las hormonas, enzimas,antibióticos, vitaminas y acidos húmicos.

Diseiio y OperaciónEl modelo básico para cultivar lombrices consisteen camellones de sustrato directamente sobre latierra, de aprox. 1-2.5 m de ancho y longitudesadaptables a la configuración dei terreno, bajosombra artificial, o natural como enredaderasinducidas de "chayote' o de arboles comoMacadamia o Ricino (Ricinus sp.). En algunoscasos, algunos productores cubren directamentelas camas de vermicompostaje utilizando puntasde cana, paja o costales viejos de yute,desechados por los propios beneficios de café;con el tiempo, éstos materiales terminanincorporandose ai humus producido.Como se mencionó anteriormente, el métodotradicional de operación consiste en la adiciónsucesiva de capas delgadas de pulpa de café en lasuperficie de las camas; cerca de tres mesesdespués, cuando la cama ha alcanzado una alturaapreciable, se retira temporalmente la capasuperior (en donde la mayoría de las lombrices seencuentran ai seguir el alimento). para permitir lacosecha de las capas inferiores de vermicompost.La capa superior de sustrato con las lombricespuede ser colocada en el mismo sitio inicial obien en una hilera vacía vecina, para reiniciarnuevamente la serie de alimentaciones de pulpade café.Solamente las grandes granjas devermicompostaje utilizan maquinaria para suoperación, mientras que la gran mayoría utilizasolamente equipo tipo jardinería, como carretillas,palas. bieldos, mangueras, etc. La mano de obrarequerida equivale a un aproximado de unapersona por cada 500 m2 de camas deverm icom postaje.Para el caso de la pulpa del café. se ha logradoestablecer un sistema de producción que implicauna estrecha y eficiente interrelación entre losciclos de generación de pulpa, su transformaciónpor las lombrices y su aprovechamientooportunoen las fincas cafetaleras (Aranda et ai, 1999). Estesistema considera el establecimiento de camascon sustrato colocado por camiones de volteo enun solo evento. formando hileras alternas (una síotra no), mismas que a lo largo de un cicloproductivo se transforman por las lombrices. Lapulpa del siguiente ciclo se coloca en los surcosintercalados y durante el inicio de la epoca delluvias, Ias lombrices se mudan por sí mismasdesde las camas con humus ya procesado, hacialas camas con pulpa fresca. Las camas con elhumus (ya sin lombrices) se retiran entonces y seaprovechan principalmente en los programas de
339
viveros y de replantación de cafetales de lamisma finca.
Producciôn de VermicompostCuando el humus es destinado como productocomercial, debe entonces ser tamizado,embolsado y con rangos apropiados de humedad.AI cosecharse, el humus normalmente presenta uncontenido excesivo de agua, (cercano a 80%) quedificulta su manejo y aprovechamiento, por lo quedebe secarse uti lizando los patios asoleaderos delos beneficios. Debido a la propiedad "anfifílica"del humus (Bouché, com. pers.), que siendonormalmente hidrofílico, pero que con secadosexcesivos se convierte en hidrofóbico, el humuspresenta sus mejores características para uso yalmacenamiento en valores cercanos ai 50-60%;durante este secado. una tonelada de humus con80% de húmedad, se llega a convertir en tan solo500 kg de humus germinado (con 60% dehumedad).EI tamizado se realiza, ya bien con la ayuda de.tamizadoras manuales o utilizando sarandasmecánicas movidas con motor, para retirarpiedras, ramas, basuras, partículas de pulpa resecay grumos aglomerados; éstos últimos puedenutilizarse así para fertilizar las propias fincas omolerse mecanicamente para reincorporarsenuevamente a la producción comercial. Lapresentación comercial mas común dei humus delombrices es en forma pura en costales de rafia,en bolsas plásticas impresas, o en mezclaspreparadas como sustrato de crecimiento paramacetas y charolas de invernaderos. EI transportees siempre uno d e los factores limitantes de lacomercialización dei producto a distancia, pero sehacen esfuerzos para expander su mercado yampliar su aceptación.
UTILlZACIÓN DEL VERMICOMPOST DEPULPA DE CAFÉ
EI humus de lombrices de puJpa de café hamostrado un ámplio rango de utilización enprácticamente cualquier suelo y proporción,siempre con resultados favorables y beneficiosospara las plantas y la fertilidad de los suelos. Sinembargo. los costos de elaboración y precios deventa 10 han restringido a cultivos de buenarentabilidad, como hortalizas de exportación(chile, jitomate, brócoli, zanahoria, esparrago,etc.), en semilleros y viveros de invernadero o enplantaciones perennes como cafetos, papaya yflores de exportación, entre otros (García, 1978;lrissón, 1995; Rodríguez et ai, 1995; Aranda y

Barois, en prensa). Los beneficios se manifiestanen mas rápidas emergencias y mayorprendimiento de semillas, mejor y más rápidocrecimiento aereo y radicular y un desarrollo massano y vigoroso general de las plantas. Por otraparte, los suelos en donde se aplica mejoran suestructura, fertilidad y un aprovechamiento máseficiente de la fertilización mineral.
SITIOS DE VERMICOMPOSTAJE ENMÉXICO
En la actualidad, existen cerca de un centenar desitios de vermicompostaje en México, queprocesan de manera comercial o paraautoconsumo pulpa de café, estiercol vacuno,cachaza de cana de azucar, otros estiercolesanimales o resíduos orgánicos de mercados y deorígen urbano. En Términos generales, se estimaque se producen anualmente en México entre5,000 y 7,000 toneladas anuales de humus delombrices que se utilizan para campos agrícolas,hortalizas, semilleros-viveros y jardínería engeneral. Existen solo unos cuantos sitios detamafio grande (mayores a 5,000 m2 de areaefectiva), una veintena de sitios de tamafiomediano (de 200 a 1,000 m2) y el resto se trata desitios pequenos que procesan pequenas cantidadesde sustratos orgánicos, general mente paraautoconsumo.
PERSPECTIVAS FUTURAS
La Lombricultura en los últimos anos estáa1canzando rapidamente popuJaridad en México yse está aplicando crecientemente sobre nuevossustratos orgánicos antes no aprovechados.Creemos que las estrategias actuales debenpromover no solo su establecimiento inicial sinosu consolidación técnica y económica. EIestablecimiento y ejecución de proyectoscomerciales eficientes y bien fundamentados,debe contrarrestar la proliferación excesiva sobrebases poco sustentadas que Ileve -corno hasucedido en otros países-, a la consiguientedesilusión de los cultivadores, a lasobreestimación del producto o incluso aIdescrédito y pérdida de los mercados de consumoya conquistados.Si el humus de lombrices está para ganar lareputación que merecidamente le corresponde, lacreación de una Asociación Nacional deLornbricultores, constituye una tarea prioritariapara promover la aplicación eficiente yproductiva de la tecnologia, el establecimiento de
340
criterios, normas y estándares de cal idad de susproductos, el apoyo científico y sobre todo unoido atento y cercano para escuchar loscomentarios y opinión de los usuarios de susproductos.
CONCLUSIONES
La pulpa de café no es ya considerada un resíduoorgánico indeseable y contaminante, sino unamateria prima de la cual obtener un producto útil,sano y económica y ecológicamente beneficioso.EI Humus de lombrices obtenido con la pulpa decafé, presenta propiedades excepcionales deestructura, retención de humedad, nutrimentos,uniformidad y consistencia en sus resultados, encomparación con otros sustratos orgánicosutiIizados.Los resultados de las investigaciones y promociónrealizadas, junto con la demanda de desarrollo desimilares aplicaciones hacia otros resíduosorgánicos, agroindustriales, pecuarios y dei mediourbano, indican que la potencialidad de estatecnología está empezando a lograrreconocimiento. Nuevas alternativas econórnicas,mejores opciones de reciclamiento yaprovechamiento productivo de los resíduosorgánicos y una rnejoría sustancial en laconservación deI ambiente y la fertilidad de lossueIos, es el objetivo y esencia de la vermiculturamoderna, por lo cual su estudio, desarrollo yconsol idación merece toda nuestroreconocimiento y atención,
AGRADECIMIENTOS
Quiero expresar mi sincero agradecimiento ai Dr.Sevastianos Roussos dei IRD-Francia y ai Dr.Henri Poupon dei IRD-México por su apoyo yfinanciamiento para la asistencia y participacióndei primer autor ai III SIBAC; ai Dr. TumoruSera y aI Dr. Carlos Roberto Soccol, por suamistad y apoyo para la publicación de esteartículo.
LITERATURA CITADA
Aranda, D.E. 1985. Las Lombrices transforman lapulpa de café en abono orgánico. ReporteInterno. Departamento de Entomología,Instituto Mexicano dei Café, Xalapa. Ver.México. 8 p.

Aranda, D.E. 1989. Perspectivas de la utilizaciónde lombrices en la transformación de la pulpa decafé en abono orgánico. Boletin Técnico deCafé, I.M.C, México. 8 p.
Aranda, D.E. 1991 a. EI Vermicompostaje; unanueva alternativa para la transformación de lapulpa de café en Abono Orgánico. en: lICAPrornecafé, Resumenes dei XIV Simposio deCafeticultura Latinoamericana, Panamá. C.A.
120 P
Aranda, D.E. 1991 b EI Vermicompostaje, laalternativa ecológica para transformar la pulpade café en abono orgánico. en: Resumenes dei 11Seminario Internacional de Biotecnológía en laAgroindustria Cafetalera,> Orstom-Cenicafe,Manizales, Colombia.
Aranda, D.E. y P.Arellano, 1995. Factoresambientales que influyen en la transforrnaciónde la pulpa de café abandonada en ai campo. en:Barois, I. and E.Aranda (eds.). Utilización delombrices en la transformación de la pulpa decafé en abono orgánico. Conacyt-Instituto deEcología. Reporte Final. pp. 1-15.
Aranda, D.E. y S.Aguilar. 1995. Tasa deextracción de lornbrices de Eisenia andrei parasu utilización como proteína animal. en: Barois,I. y E. Aranda (eds.). Utilización de lombricesen la transformación de la pulpa de café enabono orgánico. Conacyt-Instituto de Ecología.Reporte Final. pp.47-54.
Aranda, D.E., I.Barois, P.Arellano, S.lrissón,T.Salazar. J.Rodríguez, y J.c.Patrón, 1999.Vermicomposting in the Tropics. en: Lavelle,P., L.Brussaard, y P.Hendrix (eds.) 1999.Earthworm Management in TropicalAgroecosystems. CABI Publishing, U.K. pp.253-287.
Aranda, D.E. y I.Barois, (en prensa).Vermicomposting in México, en: Edwards, C.A.(ed) Manual on Vermicomposting. J&O Press,(Biocycle publisher). Emrnaus, PA.
Arellano, P. 1997. Descomposición de la pulpa decafé por Eisenia andrei (Bouché, 1972) yPerionyx excavatus (Perrier, 1872) (Annelida,Oligochaeta). Tesis de licenciatura, Facultad deBiologia, Universidad Veracruzana. Xalapa,Veracruz, México 84 p.
341
Barois, I. y E.Aranda (eds.). 1995. Utilización delornbrices en la transformación de la pulpa decafé en abono orgánico. Conacyt-Instituto deEcología. (clave: 045-N9108) Reporte Final.104p.
Bonvicini Pag1iai A. M. Y P.Omodeo (eds.), 1987.On earthworms. Se1ected Symposia andMonographs U.Z.! .. Ed. Mucchi, Modena. 562p.
Bouché, M. 1987. Emergence and developrnent ofvermiculture and vermicomposting: from ahobby to an industry, from marketing to abiotechnology, from irrationa1 to crediblepractises. en: Bonvicini Pagliai A. M. y P.Omodeo (eds). On Earthworms. Se1ectedSymposia and Monographs U.Z.!.. Ed. Mucchi,Modena. pp. 519-532.
Braharn, I.E. y R.Bressani, 1978. La Pulpa deCafé. Centro Interamericano para el Desarrollo.Colornbia.Consejo Mexicano del Café, 1997.Estadisticas Cafetaleras, D.F., México. 10 p.
Edwards.C.A. y E.Neuhauser, 1988.Earthwormsin waste and environmental management,Academic Publishing, Netherlands, 392 p.
Edwards C. A.; 1995. Historical overview ofvermicomposting. Biocycle pp.56-58.
Edwards, C. A. y P.Bohlen. 1996. The Biologyand Ecology of Earthworms. 3rd EditionChapman and Hal1, London. 426 p .
Edwards, C.A., 1997. Fifth InternationalSyrnposium on Earthworm Ecology (ISEE 5).Soil Biology and Biochemistry 29(3/4): 215766.
Edwards, C.A. (en prensa). Manual onVermicomposting. J&O press, (BiocyclePublisher) Emrnaus, PA.
Oaime-Perraud, 1., L.Hannibal, M.Trejo yS.Roussos. 1991a. Resultados preliminaressobre e1 ensilaje de la pu1pa de café. Resumen.
IJ Seminario Internacional de Biotecnologia en laAgroindustria Cafetalera (IJ SIBAC),ORSTOM-CENICAFE, Manizales, Colombia.
Oaime-Perraud, 1., J.Zuluaga, O.Saucedo,S.Roussos. 1991b. Microflora natural de café.en: Roussos, S. (ed.). Resumenes dei 11

Seminario Internacional de Biotecnologia en laAgroindustria Cafetalera (JJ SIBAC) ,ORSTOM-CENICAFE, Maniza1es, Colombia.
Gaime-Perraud, I. 1995. Cultures mixtes enmilieu solida de bacteries 1actiques et dechampignons filamenteux pour la conservationet la decafeination de la pulpe de café. These deDoctorat Université Montpellier ll, Sciences etTechniques de Languedoc. 209 p.
García, M.A.B. 1996. Algunos sustratosorgánicos, sus mezclas, caracterización yprocedimientos. (Tesis profesional) IngenieroAgrónomo en Suelos, Universidad AutónomaAgraria Antonio Narro, Saltillo Coahuila,México. 144 p.
Guerra, P.L. 1982. Estudio preliminar sobre lautilización de la lombriz de tierra (Helodrilusfàetidus) en la alirnentación dei bagre de canal(Ictolus punctatus). (Tesis profesional).Universidad Autonoma Agraria, Monterrey,Nuevo León, México.
Guerrero, R.D. 1983. The culture and use ofPerionyx excavatus as a protein source in thePhilippines. en: Satchell, J. (ed.). EarthwormEcology, from Darwin to Ecology. Chapmanand Hall. pp. 309-313.
Hartenstein, R. 1981. Production of earthwormEisenia foetida as a potentiaJy economicalsource of protein. Biotechnoly andBioingenering. 23, 1812-1997.
Hoerschelrnan. H. y H.G.Andres, 1994. WilhelmMichaelsen Gedãchtnis Syrnposion.Mitteilungen aus dem HamburgischenZoologischen Museum und Institut, Hamburg.200 p.
lrissón, N.S. 1995. Calidad deI abono y de lalombriz de tierra resultante dellombricornpostaje de la pulpa de café, (Tesisprofesional). Facultad de QuímicaFarmacéutica Biológica, UniversidadVeracruzana, Xalapa, Veracruz, México.85 p.
Jaenike, J. 1982. "Eisenia foetida" is twobio1ogical species. Megadrilogica 4( 1,2): 6-7
Kretzschmar A. (ed.), 1992. Fourth lnternationalSymposium on Earthworm Ecology (lSEE 4),
342
Soil Biology and Biochemistry 24( 12): 11931774.
Lofs-Holrnin A. 1985. Vermiculture, presentknowledge of the art of earthworm farming.Inst. Ekol. Miljovard, Swedish UniversityAgronomic Sciences 67 p,
Neuhauser, E.F., R.Hartenstein. y D.I.Kaplan,1980. Growth of earthworm Eisenia fetida inrelation to population density and foodrationing. OlKOS, 35( I):93-98
Rodríguez, L, E.Aranda, I.Barois y E.Barrientos.1995. Respuesta de cafetos (Coffea arabica L)en sernillero-vivero a la aplicación de abonoorgánico de pulpa de café y fertilizantequímico. en: Barois, I. y E.Aranda (eds.).
Utilizacion de lombrices en la transformacionde la pulpa de café en abono orgánico.Conacyt-lnstituto de Ecología. Reporte Final.pp.65-73
Rolz, C; J.Menchú, F.Calzada. F.de León,R.García. 1982. Biotechnology in WashedCoffee Processing. Process Biochemistry,MarchiApril 1982. pp. 9-1 J
Roussos, S., A.Aquiahuatl, J.Cassaigne, E.Favela.M.Gutierrez. L.Hannibal, S.Huerta. G.Nava,M.Raimbault, W.Rodríguez. J-A.Sanchez.M.Trejo, G.Viniegra-Gonzalez. 1989.Detoxificación de la Pulpa de Café porFermentación Sólida. en: Roussos, S..R.Licona, M.Gutierrez (eds.) ler SeminarioInternacional sobre Biotecnologia en laAgroindustria Cafetalera. ORSTOM-IMCUAM. 210 p.
Salazar, T., E.Aranda y I.Barois. 1995. Estudiocomparativo dei lombricompostaje de la pulpade café por Eisenia andrei, Eisenia fétida yPerionyx excavatus en condiciones de campo.en: Barois, 1. y E.Aranda (eds). Utilizacián delombrices en la transformación de la pulpa decafé en abono orgánico. CONACYT (clave:045-N9108) Reporte Final. pp. 28-41.
Satchell . .r.E. (ed). 1983. Earthworms Ecology,from Darwin to Vermiculture. Chapman &Hall, Cambridge, Inglaterra. 283-296 pp.
Taboga, L. 1980. The nutricional value ofearthworms for chickens. British PoultryScience. 21 :405-410.

Tacon, A.G., E.Stafford, y C.Edwards. 1983. Aprelirninary investigation of the nutritive valueof three terrestrial lumbricid worms forrainbow trout. Aquaculture. 35: 187-199.
Tomati, V. A., E.Grapelli, Galli, y W.Nizi, 1983.Fertilizer from vermiculture as an option fororganic wastes recovery. Agrochemestry, 27:244-251.
Toscano, S.S. (en prep). Evaluación delcrecimiento de la trucha arcoiris Oncorhynchusmykiss (Billard, 1989) utilizando lombricesEisenia andrei (Bouché, 1972) como sustitutoparcial de proteina en el rancho "EI Roble",Coatepec, Ver. (Tesis profesional), InstitutoTecnológico deI Mar, Boca dei Río, Veracruz,México.
343
Van Gansen, P. 1962. Structures et fonctions dutube digestif du lombricien Eisenia foetidaSavigny. Journal de Microscopie, 1, 1-363.
Velásquez, B.L.. C.Herrera y B.Jbanez, 1986.Harina de Lombriz. I parte: Obtención,composición química, valor nutricional ycalidad bacteriológica.Alimentos, 2(1): 15-20.
Zuluaga, V.J. J989. Utilización integral de lossubproductos dei café. en: Roussos, R., R.Licona, M.Gutierrez (eds.). Memorias del ISeminario Internacional sobre Biotecnologíaen la Agroindustria Cafetalera. ORSTOMUAM-INMECAFE, Xalapa, Ver. México. pp.63-65


345
PROCESAMIENTO DE FRUTOS DE CAFÉ POR VIA HUMEDA Y GENERACION DESUBPRODUCTOS.
Jaime Zuluaga Vasco
Gerente T E C O L-Tecnologías Ecológicas Colombianaserros de la Alhambra # 81. ManizalesCOLOMBIA.Telefax: 68-745790.
RESUMENEn Colombia el proceso de beneficio dei café se ha hecho tradicionalmente por vía húmeda y este es uno delos factores a los cuales se debe la excelente calidad dei café Colombiano. No obstante, en este proceso seconsumen grandes volúmenes de agua limpia y se contaminan cantidades equivalentes. La situación actualen la zona cafetera, caracterizada por una deficiencia de agua para las labores de beneficio, la necesidad deprocesar cada vez volúmenes más grandes y una conciencia creciente sobre los problemas decontaminacion, hacen necesario un replanteamiento dei manejo dei agua y de los subproductos en elproceso de beneficio húmedo de café.La producción de café, durante los últimos anos en Colombia, ha sido dei orden de 12 millones de sacos de60 kilos de café verde, equivalentes a 720.000 toneladas. La pulpa y el mucílago principales subproductosdei proceso de beneficio húmedo de café, generan 350.770 toneladas de materia seca. Si estos subproductosno son utilizados o dispuestos convenientemente se convierten en una fuente de contaminación de! medioambiente.Los conocimientos actuales permiten plantear el desarrollo de un proceso integral para la valorización delos subproductos dei proceso de beneficio húmedo dei café, apoyándose en la racionalizacion dei uso deiagua en e! proceso (abolición dei agua como vehículo de transporte) y en procesos de fermentación enmedio sólido (ensilaje de la pulpa, producción de hongos comestibles sobre pulpa, cultivo de lombriz rojaCaliforniana utilizando la pulpa como sustrato) y en medio líquido (Bíodigestión Anaeróbica, en fasesseparadas, dei mucílago disue!to en las aguas residuales dei lavado dei café). La aplicación de losanteriores desarrollos permite reducir ocho veces e! consumo de agua necesario para el proceso y producirproteína vegetal (hongos comestibles), proteína animal (lombrices), productos mejoradores de laspropiedades fisico-químicas y microbiológicas dei suelo (lombricompuesto), alimento para animales (pulparesidual dei cultivo de los hongos), combustible (biogas) y productos con propiedades fertilizantes (lodos yefluentes de la biodigestión anaerôbica). Los procesos biotecnológicos están brindando cada vez más ymejores oportunidades para el aprovechamiento integral y rentable de los subproductos de los procesosagrícolas, pecuarios y de producción de alimentos; contribuyendo así a mejorar la rentabilidad de losprocesos de producción y evitando la contaminación de! medio ambiente.
1. LA RECOLECCION DEL FRUTO DELCAFE
En la práctica la maduración de los frutos seevidencia por el cambio de color, para el caso deIcafé, este cambio es dei verde ai rojo cereza oamaril1o, dependiendo de la variedad cultivada.-Después de la maduración total los frutos entranen un periodo en el que el cataboJismo predominasobre el anabolismo iniciándose la fase desenescencia. En esta fase hay fermentaciones conproducción de alcoholes y ácidos indeseables,ruptura de la estructura de la pared celular pormodificaciones en las pectinas, celulosas,hemícelulosas y ligninas; oscurecimiento de lapulpa debido a oxidaciones de pigmentos ypudrición. En consecuencia la calidad de losfrutos tiende a disminuir marcadamente.Ha sido ampliamente demostrado que la mejorcalidad de café, tanto dei grano como de la
infusión, se obtiene cuando el fruto es cosechadoen su estado de óptima madurez. En este estado lacomposicíón química, tanto : cualitativa comocuantitativa, se encuentra en su punto óptimopermitiéndole ai fruto Ia expresión de su máximacalidad. GARRUTI y GOMEZ en el Brasil (in I)observaron que los cafés cereza maduros(despulpados y no despulpados) presentaroncal idad de bebida superior (suave) a la de losfrutos cosechados verdes (bebida "rnuy dura").Los frutos verdes presentan algunos compuestosquímicos en niveles diferentes ai ideal. presentadopor los frutos maduros, lo cual se ref1eja en lascaracterísticas organolépticas de la bebida. Demanera similar los frutos secos en el árbol orecogidos deI suelo son materiales que ya han comenzado su fase de senescencia; con laconsecuente ruptura de la membrana celular ypresencia de compuestos químicos derivados de

procesos fermentativos o deI ataque demicroorganismos.En general las condiciones de alta humedad deisuelo, la sobremaduración y el paso deI estado decereza verde a fruto seco pueden provocardegradación de la membrana y favorecer lasfermentaciones y las infecciones microbianas.TEXEIRA et aI (in I) determinaron que mezclasde café normal con porcentajes superiores aI 15%de granos verdes o fermentados así comosuperiores aI 10% de granos negros, perjudicansensiblemente la calidad de la bebida.De acuerdo con CHOUSSY (in I) la intensidad dela alteración en la composición química de losgranos, debida a causas fisiológicas omicrobiológicas, está en relación con lasdiferentes condiciones climáticas de las distintasregiones de cultivo y en consecuencia, para éstasdonde las condiciones ambientales son altamentefavorables a los ataques de microorganismos, losfrutos deben ser cosechados a mano, en su estadode óptima madurez; rápidamente despulpados,fermentados, lavados y secados artificialmente endispositivos mecánicos.Para la obtención de un café de excelente calidades condición indispensable la ejecución de unarecolección racional; entendiéndose por esto elcosechado manual de únicamente cerezas madurasy sanas. EI beneficio de una mezcla de cerezasverdes, amarillo verdosas, anaranjadas, rojas ynegras só lo puede producir una calidadheterogénea tanto en el color de la almendra comoen las propiedades gustativas de la infusión.Durante el despulpado el grano verde se rompe,debido a la ausencia de la propiedad lubricanteque proporciona el mucílago deI café maduro y seorigina el grano partido y/o mordido. Además,como aún no están formados los azucares.. lIafermentación no se lIeva a cabo normalmente y·.seproduce una descomposición que genera maios'olores para la totalidad de la masa de café.Las cerezas de color amarillo verdoso y sobretodo verde dan una almendra de coloracióndefectuosa (grano decolorado) y una bebidaáspera y picante. Las cerezas sobremaduras decalor rojo-vinoso, producen una bebida con sabora fruta ("fruty") e incluso a levadura ("yeasty").
346
Por su parte las cerezas negras, parcialmentedesecadas en el árbol, originan un café pergaminocon parte de la pulpa adherida (grano media cara)y una almendra de color apagado (granodecolorado) asi como una infusión con sabor amadera
2. MORFOLOGIA DEL FRUTO DEL CAFÉ.
EI fruto deI caté tiene el mismo aspecto exteriorque el de una cereza y por esta razón,comúnmente, se le denomina con este nombre. Uncorte longitudinal deI fruto (Esquema I A de laFigura 1) muestra las diferentes capas celulares.Se observa que la pulpa está constituida por elexocarpio (Epidermis) y parte dei mesocarpio. Elcolar de la epidermis depende de la variedad decafé y dei estado de madurez del fruto y varia deverde a rojo; teniendo a veces tonal idades rojooscuro, violeta y aún negro. EI mesocarpio seencuentra protegido por la epidermis y está constituido por una capa gruesa de tejido esponjoso de,aproximadamente 5 mm de espesor, rico en azúcares y mucílagos y a su vez envuelve dos granosopuestos por su cara plana.Los granos están revestidos por una doblemembrana:la primera Ilamada pergamino, decolor amarillo pálido y de consistencia dura yfrágil, la segunda.mucho más fina que la anteriory adherida aI grano (Alburnen), es lIamadapelícula plateada (Tegumento Seminal). En labase de [os granos, sobre la cara interna, seencuentra el germen (Embrión).La observación microscópica dei cortelongitudinal, muestra en detalle cada una de laspartes mencionadas anteriormente (Esquema IB).Este corte permite observar que la pulpa noestá constituida solamente por el exocarpio y elmesocarpio sinó, además, por otras tres capas detejido celular: células en empalizada, célulaslongitudinales del esclerénquima y célulastransversales dei esclerénquima. La películaplateada está constituida a su vez de tres capas:epidermis, capa media y células comprimidas. Enel grana se puede observar el tejido de reservaparenquimatoso y el material graso,
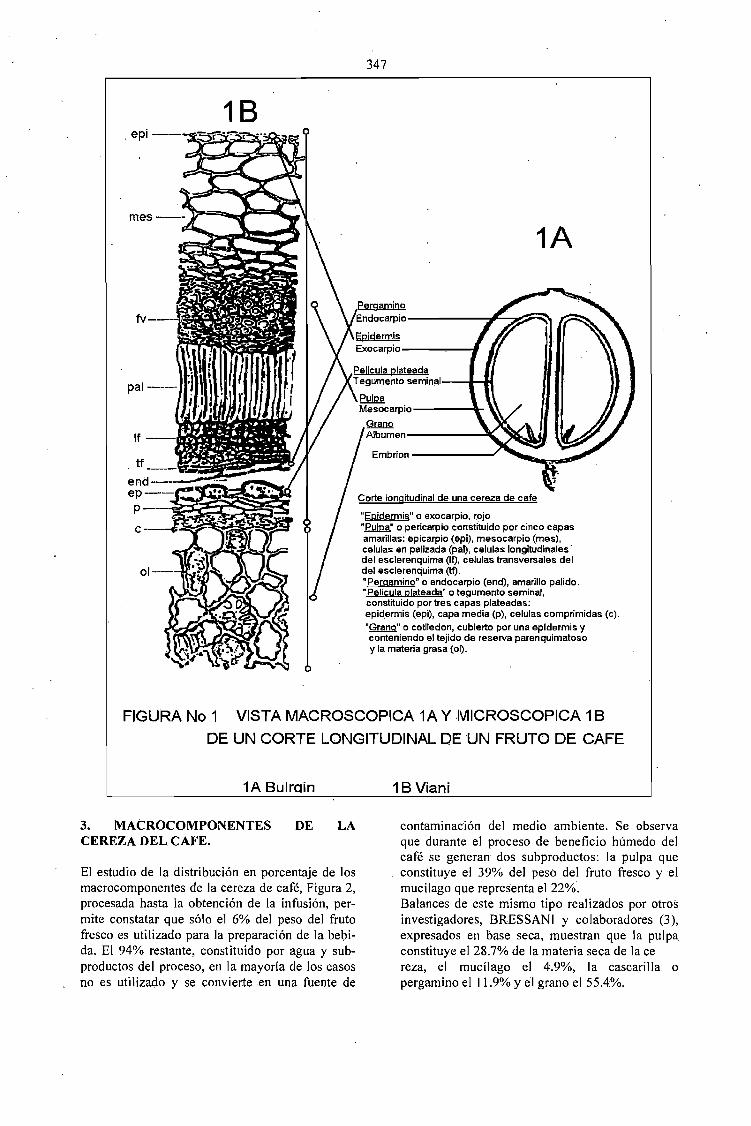
347
18
c-~-~
1A
pergamjnoEndocarpio-----""""'lI~""7~"""
pelicula plateadaTegumento semin;lI-----lHtI
PulpaMesocarpio----+
~Albumen-----~"_
Embrion ------'
Corte longitudinal de una cereza de cafe
"Epjdermis" o exocarpio, rojo"~' o pericarpio constituido por cinco capasamarillas: epicarpio (epi), mesocarpio (mes),celulas en palizada (paQ,celulas longitudinales'dei esclerenquima (lI), celulas transversales deidei esclerenquima (tI)."Pernamlno" o endocarpio (end), amarillo palido."Película plateada" o tegumento seminal,constituido por tres capas plateadas:epidermis (epi), capa media (P), celulas comprimidas (c)."Grano" o cotiledon, cubierto por una epidermis yconteniendo el tejido de reserva parenquimatosoy la materia grasa (01).
FIGURA No 1 VISTA MACROSCOPICA 1A YIVIICROSCOPICA 1B
DE UN CORTE LONGITUDINAL DE UN FRUTO DE CAFE
1A Bulrain 1B Viani
EI estudio de la distribución en porcentaje de losmacrocomponentes de la cereza de café, Figura 2,procesada hasta la obtención de la infusión, permite constatar que sólo el 6% dei peso del frutofresco es utilizado para la preparaciónde la bebida. El 94% restante, constituido por agua y subproductos del proceso, en la mayoría de los casosno es utilizado y se convierteen una fuente de
3. MACROCOMPONENTESCEREZA DEL.CAFE.
DE LA contaminación deI medio ambiente. Se observaque durante el proceso de beneficio húmedo deicafé se generan dos subproductos: la pulpa queconstituye el 39% dei pesá del fruto fresco yelmucilago que representa el 22°/;Balances de este mismo tipo realizados por otrosinvestigadores, BRESSANI y colaboradores (3),expresados en base seca, muestran que la pulpaconstituye el 28.7% de la materia seca de la cereza, el mucílago el 4.9%, la cascarilla opergamino el 11.9% y el grano el 55.4%.
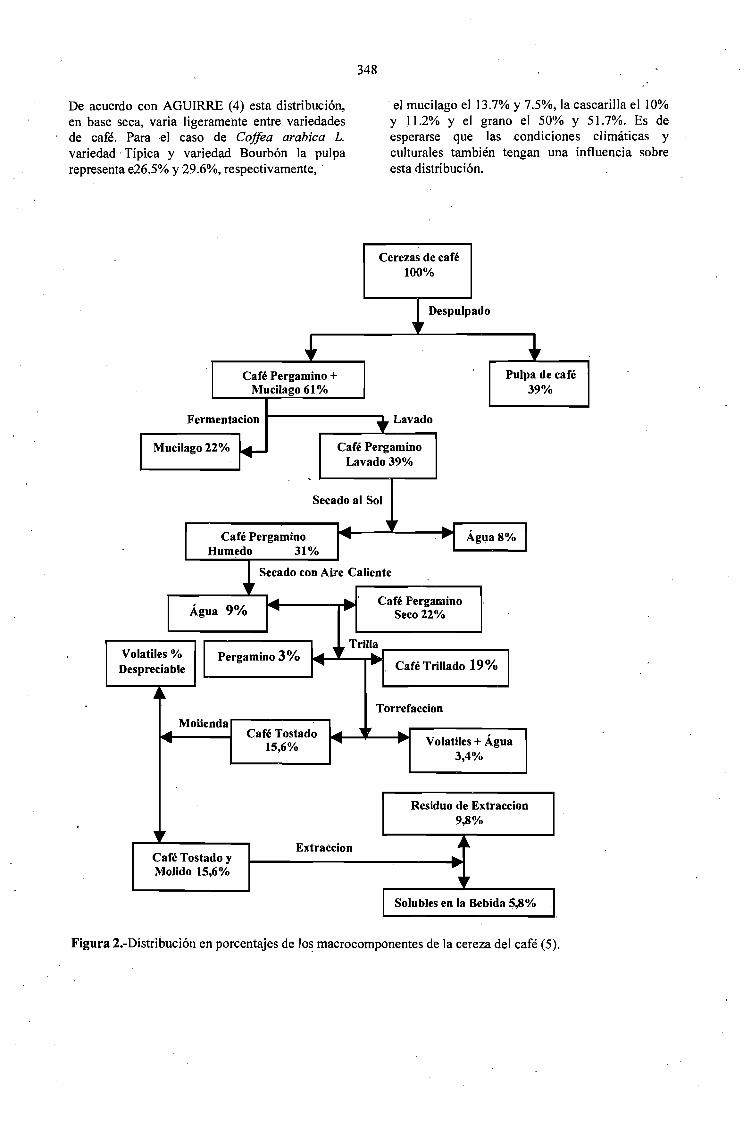
De acuerdo con AGUIRRE (4) esta distribución,en base seca, varia ligeramehte entre variedadesde café. Parael caso de Coffea arabica L.variedad .Típica y variedad Bourbón la pulparepresenta e26.5% y 29.6%, respectivamente, ,
348
. el mucilago el 13.7% y 7.5%, la cascarilla el 10%y 11.2% Y el grano el 50% y 51.7%. Es deesperarse que las condiciones climáticas yculturales también tengan una influencia sobreesta distribución.
Cerezas de café100%
~ Despulpado
~ ~
ICafé Pergamino + Pulpa de café
Mucilago 61% 39%
Fermentacion .. Lavado
I Mucilago 22% j.- Café PergaminoLavado 39%
,
Secado ai Sol
,r
I~ ~
Café Pergamino ....- "I Água 8%Humedo 31%
Secado con Aire Caliente
I..... ... ' Café Pergamino
Água 9% ~ ...Seco 22%
I ' Ir' Trilla
IVolatiles % Pergamino 3% I~ ~Café Trillado 19% lDespreelable
'li>Torrefaccion
Molienda ,Ir.... Café Tostado ... ~
Volatiles + Água....15,6%
~ ...3,4%
Residuo de Extraccion9,8%
,Ir~Il-Extraccion
Café Tostado y "-
Molido 15,6%.., Ir'
Solubles en la Bebida 5,8%
Figura 2.-Distribución en porcentajes de los macrocomponentes de la cereza del café (5).
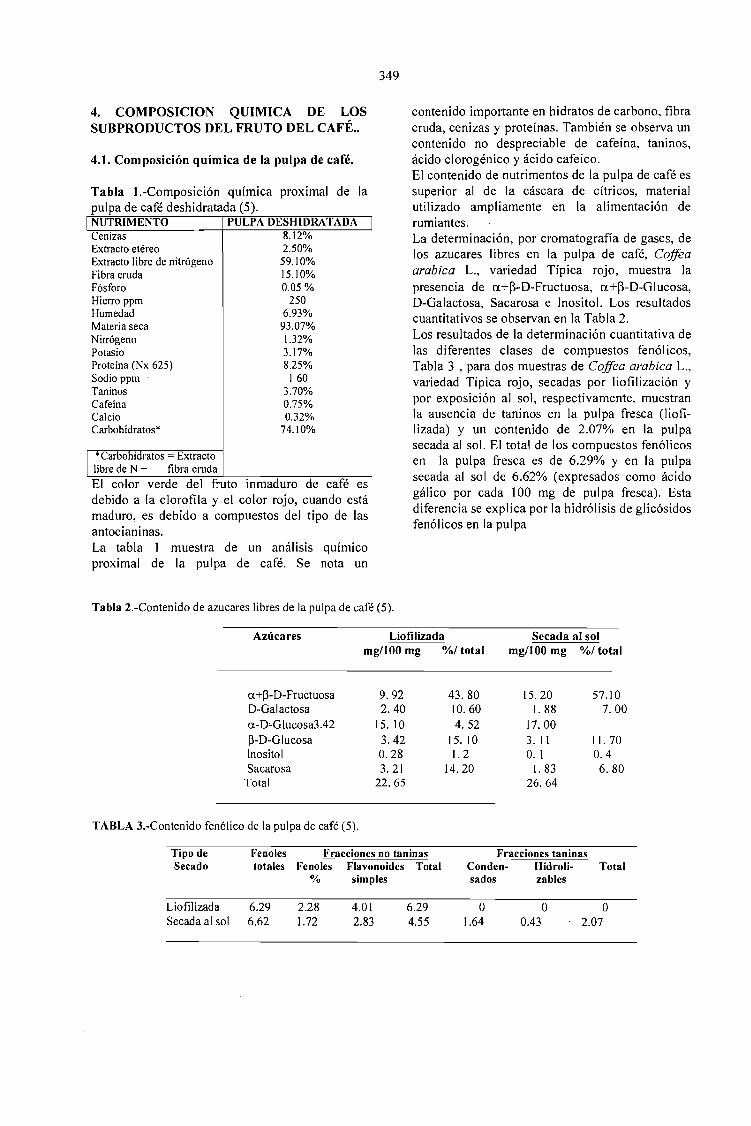
349
4. COMPOSICION QUI MICA DE LOSSUBPRODUCTOS DEL FRUTO DEL CAFÉ..
4.1. Composición química de la pulpa de café.
Tabla 1.-Composición química proximal de lapulpa de café deshidratada (5).NUTRIMENTO PULPA DESHIDRATADACenizas 8.12%Extracto etéreo 2.50%Extracto librede nitrógeno 59.10%Fibracruda 15.10%Fósforo 0.05%Hierro ppm 250Humedad 6.93%Materia seca 93.07%Nitrógeno 1.32%Potasio 3.17%Proteína (Nx 625) 8.25%Sodio ppm 160Taninos 3.70%Cafeina 0.75%Calcio 0.32%Carbohidratos* 74.10%
*Carbohídratos - Extractolibre de N + fibracrudaEI color verde deI fruto inmaduro de cafe esdebido a Ia clorofila y el color rojo, cuando estámaduro, es debido a compuestos deI tipo de lasantocianinas.La tabla I muestra de un análisis químicoproximal de la pulpa de café. Se nota un
Tabla 2.-Contenido de azucares libres de la pulpa de café (5).
contenido importante en hidratos de carbono, fibracruda, cenizas y proteínas. También se observa uncontenido no despreciable de cafeína, taninos,ácido clorogénico y ácido cafeico.EI contenido de nutrimentos de la pulpa de café essuperior aI de la cáscara de cítricos, materialutilizado ampliamente en la alimentación derumiantes.La determinación, por cromatografía de gases, delos azucares libres en la pulpa de café, Coffeaarabica L., variedad Típica rojo, muestra la
presencia de a+~-D-Fructuosa, a+~-D-Glucosa,
D-Galactosa, Sacarosa e Inositol. Los resultadoscuantitativos se observan en la Tabla 2.Los resultados de la determinación cuantitativa delas diferentes clases de compuestos fenólicos,Tabla 3 , para dos muestras de Coffea arabica L.,variedad Típica rojo, secadas por liofilización ypor exposición aI sol, respectivamente, muestranla ausencia de taninos en la pulpa fresca (liofilizada) y un contenido de 2.07% en la pulpasecada aI sol. El total de los compuestos fenólicosen la pulpa fresca es de 6.29% y en la pulpasecada ai sol de 6.62% (expresados como ácidogálico por cada 100 mg de pulpa fresca). Estadiferencia se explica por la hidrólisis de glicósidosfenólicos en la pulpa
Azúcares Liofilizadamg/lOO mg %/ total
Secada aI solmg/lOO mg %/ total
a+p-D-FructuosaD-Galactosaa-DcGlucosa3.42p-D-G1ucosaInositolSacarosa
Total
TABLA 3.-Contenido fenólico de la pu1pade café (5).
9.922.40
15.103.420.283.21
22.65
43.8010.604. 52
15.101.2
14.20
15.20I. 88
17.003.110.1
I. 8326.64
57.107.00
11.700.46.80
Tipo deSecado
Fenoles Fracciones no taninastotales Fenoles Flavonoides Total
% simples
Fracciones taninasConden- Hídroli- Totalsados zables
LiofilizadaSecada ai sol
6.296.62
2.281.72
4.012.83
6.294.55
O1.64
O0.43
O2.07

TabIa 4.-Composición química del rnucilago deI
350
café (1 3)(6).
Componente
Sustancias pécticas totalesAzúcares totalesAzúcares reductoresAzúcares no reductoresCelulosa + cenizasAguaProteínaAcido pécticoCenizas
NadaI
%Base húmeda
4.10
84.208.90·0.910.70
Bressani%
Base seca
35.8045.80
)0.0020.0017.00
Secada ai sol. La diferente distribu'ciónde losfenoles simples y de los flavonoides, en las dosmuestras, explica la formación de los taninoshidrolizables y condensados, respectivamente. Enparticular. la fonnación de los taninos condensados explica el cambio de color de la pulpa, derojo a café oscuro, por polimerización de las antocianidinas: compuestos responsables dei colorrojo de la cereza.
4.2. Composición química dei mucilago.Desde el punto de vista fisico el mucílago es unhidrogel. Quimicamente, el mucílago contieneagua, pectinas, azucares y ácidos orgánicos. Deacuerdo con AGUIRRE (4), el mucilago nocontiene ni cafeína ni taninos. Durante la maduración del fruto dei café. el pectato de calcio ylas protopectinas son convertidos en pectinas poracción de las enzimas pectinolíticas. La Tabla 4muestra la composición química dei mucílagoreportada por varios autores.
5. EL PROCESO DE BENEFICIO HUMEDODE CAFÉ.
5.1. Aspectos genera les.
5.1.1. Antecedentes.El beneficio del café en Colombia se hace, demanera tradicional. desde hace mas de I 50 afios yhan sido muy pocos los carnbios que se le hanincorporado desde entonces.La cantidad y la calidad dei agua disponible, Iafalta de una infraestructura vial adecuada y la carencia de energía eléctrica en la zona cafetera, asícomo. Ia poca conciencia en relación con losproblemas de contaminación generados;determinaron en el pasado que el proceso fuesefundamentalmente hidráulico. En este momento.para un gran porcentaje de las 300.000 famílias
que cultivan el café en Colornbia. todas estascaracterísticas han cambiado y dado el progresoagronômico en el cultivo, Ia producción se ha por10 menos d,uplicado y en consecuencia, se imponeuna nueva orientación para el proceso debeneficio húmedo de café más acordecon lascircunstancias.
5.1.2. Producción de café en Colombia.De acuerdo con e1 censo cafetero 1980-81 (8), lacapacidad potencial de producción del país. encargas de 125 kilogramos de café pergamino seco(cps), es de 8.930.617. Esto equivale a 1.116.237toneladas de cps que a su vez provienen de5.023.472 toneladas de café en cereza (cc)(teniendo en cuenta que 4.5 kilos de cc producen1 kiJo de cps; de acuerdo con URl BE (9)).La producción para el afio cafetero 1990-91 fué de14.400.000 sacos de 60 kilos de café verde (cv)=864.000 toneladas de cv= 1.080.000 toneladas decps= 4.803.840 toneladas de cc.
5. i .3. Consumo de agua en el proceso debeneficio húmedo de café.EI volumen prornedio de agua utilizado en elbeneficiado de un kilogramo de cps es de 40 litros(' O) (aproximadamente 20 Its en el despulpado y20 lts en el lavado). lo que equivale a 8.3 metroscúbicos de agua por tonelada de cc y a 42.754.176metros cúbicos de agua para procesar la cosechacafetera de 1990-91.
5.1.4 Equivalente de contaminación en elbeneficio dei café.DQO prornedio excretada por un habitante/día =67 grs. (I I). DQO total de la pulpa generada porun Kg de cc = 84.8 grs. DQO total dei mucílagogenerado por un Kg de cc = 30.3 grs,DQO total generada en residuos orgánicos por unKg de cc beneficiado= I '5.1 grs.

Equivalente de contaminación por Kg de ccbeneficiado 1.7 habitantes /día = 1718 hab./díapor tonelada de cc beneficiado.Equivalente de contaminación de la cosechacafetera colombiana 1990/1991 8.252.997.120hab./día 22.61 0.951 hab.zaüo,
5.2. EI proceso.EI tipo de proceso aI cual se somete el fruto deicafé después de la recolección depende en granmedida de la especie y variedad de café cultivado.de la disponibilidad de agua y sobre todo dei tipode recolección practicada.Normalmente los cafés arábicos, recolectadosracionalmente (recolección manual de frutos maduros y sanos), son beneficiados por via húmeda ydan origen a las bebidas suaves de más altacalidad pero generan contaminación dei medioambiente.EI 40.6% dei mercado mundial dei cafécorresponde a los cafés suaves lavados producidosen Colombia. Kenia, Tanzania, México. CostaRica. República Dominicana. EI Salvador.Guatemala. Haití, Honduras. Jamaica. Nicaragua,Venezuela. Ecuador, Perú, Ruanda, Burundi,India y Australia (12).En el proceso de beneficio húmedo propiamentedicho, se efectúan básicamente en tres etapas a saber:A.-EI despulpado.B.-La fermentación.C.-EI lavado y clasificación.
Trampa denatas ---+"'r--~---t-<~
Trampa de --+--i:->-9;--f'oo;t~--E::"'"piedras
Criba paracate despulpado---""""","""b~'-F,'
Tanque de segunda ----'~,'\),..cI\J'
351
La operación de secado. Ia cual es común para elbeneficio seco y húmedo, no hace parte realmentedel proceso húmedo (donde se consume agua).Enla Figura 3se observa un esquema de las instalaciones típicas para la realización de las diferentesoperaciones de un proceso de beneficiohúmedo de café
S.2.L-La etapa deI despulpado.En esta etapa se remueve el epicarpio (pulpa) yparte del mesocarpio (mucilago) dei fruto. con elfin de favorecer una aceleración deI proceso dedescomposición dei mucilago y evitar el manchado del café pergamino por difusión de lospigmentos antociánicos presentes en el epicarpiodei fruto.La pulpa constituye aproximadamente el 40% deifruto fresco y el 29% en base seca. presentando uncontenido de humedad dei 77% (5. 13).En la operación de despulpado se pueden originardanos químicos por retardo en la ejecución odanos mecánicos por mal uso de la máquinadespulpadora,Durante el desarrollo dei fruto dei café en el árbol,los azúcares, principalmente la galactosa, seoxidan a ácidos galacturónicos que pordeshidratación producen anhídridos los cuales sepolimerizan dando origen a las sustanciaspécticas. A estas sustancias, depend iendo de supeso molecular. se les conoce como protopectinas,pectinas y ácidos pécticos,
Patios
FIG. 3. Instalaciones típicas para un proceso de beneficio húrnedo de café (7).

La descomposición de estas sustancias pécticas,por hidrólisis enzimática durante la maduracióndei fruto, es lo que da origen ai mucílago y a lasrnieles, las cuales cumplen una función lubricantedurante el despulpado.La ausencia de mucílago en el fruto, por falta demadurez o por resecamiento en el árbol, nopermite una adecuada y completa remoción de iapulpa 10 que da origen ai defecto conocido comograno media cara.Una adecuada y oportuna calibración deI ajuste dela despulpadora, sobre todo aI comienzo y en elgrueso de la cosecha, es necesaria para evitar laaparición dei grano mordido y el grano aplastado.Las heridas causadas en el grano permiten lamezcla y reacción de los constituyentes químicossolubles presentes intra y extracelularmente, asícomo las oxidaciones debidas ai oxígeno deI aire.Lo anterior expl ica la aparición de un color negro,en el borde de las heridas de los granos, poroxidación de cornpuestos principalmente del tipofenólico.De igual manera estas heridas favorecen el ataquede microorganismos (hongos y bacterias) loscuales a su vez producen compuestos químicosextrafíos ai café verde.Los danos físicos causados ai café verde tienen suefecto sobre la calidad dei proceso de torrefacciónai no presentar la almendra una matriz uniformepara los intercambios de calor y los efectos de laspresiones desarrolladas durante la piról isis.Los granos con danos físicos frecuenternente secarbonizan debido a que las reacciones pirolíticasya no ocurren dentro de la celula y en ausencia deaíre sino en contacto directo con esta.Todas estas reacciones carnbian la cornposiciónquímica dei café verde y en consecuencia las propiedades organolépticas de la infusión preparadacon éste.
5.2.2. La etapa de la fermentación.En este proceso se descompone el resto deimesocarpio. mucilago. que no fue retirado en laoperación de despulpado, con el fin de permitir suremoción en la operación de lavado y facilitar losprocesos de secado y almacenamiento dei grano.EI mucilago constituye aproximadamente el 22%dei peso dei fruto tresco y el 5% en base seca.presentando un contenido de humedad del 85 ai90% '(5. 13).De acuerdo aCOSTE (21). Ia descomposición delas sustancias pécticas, de la envoltura mucilaginosa del grano, es atribuída a una accióndiastásica acornpaüada de una fermentaciónláctica; pero sin Ilegar a producir fermentacionessecundarias (acética. propiónica, butirica-pútrida)nocivas para la calidad dei producto.
352
Mediante "pruebas de taza" se ha determinado quela fermentación correctamente efectuada no tieneinfluencia sobre la calidad de la bebida. Sinembargo. se ha observado que el sabor ligeramente ácido de ciertos cafés arábica, muyapreciados por los consumidores, se obtienenmediante una fermentación de cierta duración.WILBAUX (22). observó que la fermentaciónbajo agua mejora ligeramente la calidad de loscafés finos aI producirse una mayor cantidad deácidos; especialmente láctico.Normalmente los danos en la calidad dei café,generados en el proceso de fermentación natural,son debidos a largos perlados defermentaciórusuperiores a 24 horas). En la zonacafetera Colombiana la fermentación deI caférecolectado en su punto óptimo de madurezdemora entre 12 y 20 horas (dependiendo de laaltura sobre el nivel del mar, al cual se encuentrenlas instalaciones de beneficio. lo cual determinaen gran medida la tempertura ambiente).La velocidad de hidrólisis (descomposición) deimucilago, depende de la temperatura de la masade café en fermentación y dei grado de madurezdel fruto; lo cual a su vez determina el contenidode enzimas pectinolíticas y la cantidad deazúcares.EI mucilago se forma en el fruto dei café cuandoéste ha alcanzado un buen nivel de madurez y seencuentra ausente en los frutos verdes y resecosen el árbol.La hidrólisis y ferrnentación de los compuestosque forrnan el mucílago (pectinas y azúcares principalmente) se l1eva a cabo por una accióncombinada de las enzimas naturales dei fruto y delos microorganismos presentes en el café y en eltanque de fermentación. .De acuerdo a PICADO (20). todos losmicroorganismos capaces de atacar las sustanciasorgánicas pueden Ilevar a cabo la transformacióndei mucílago: pero la calidad del café: coloro olory sabor; varian con el tipo de microorganismo queha intervenido en esta transfonnación. Losmicroorganismos pueden ser útiles, perjudicialeso inocuos,Después dei despulpado. el café en baba quedaexpuesto a microorganismos tales como levadurashongos y bacterias que al encontrar condicionesfavorables para su desarrollo infectan el grano.Estos microorganismos producen sus propiasenzimas las cuaJes actúan principalmente sobrelos azúcares oxidándolos y produciendo alcoholesy ácidos. al tiernpo que se eleva la temperatura dela masa de café por el calor generado en estasreacciones. Algunos de los compuestos que seforrnan durante la fermentación dei café. son: el

alcohol etílico, el cual a su vez es.oxidado a ácidoacético y el ácido láctico.FRITZ (23), estudió las reacciones que seproducen en el curso de la fermentación yobservó:I.-Una fermentación alcohólica muy activa, condesprendimiento de C02, entre las 7 u 8 primerashoras.2.-Una fermentación láctica, que comienza hacíala segunda o tercera hora y se mantiene activahasta la 24-ava hora.3.-Una fermentación acética que comienza haciala I I -ava hora, presentándose muy activa entre la12 y I 8-ava hora; para luego disminuir deintensidad.4.-Una fermentación butírica, que comienza entrela 20 a 24-ava hora; cuando la masa mucilaginosase junta en el fondo de la cuva de fermentación.Cuando la fermentación es muy prolongada, lainfección por los microorganismos se tornaimportante y comienzan a aparecer danos en lacalidad, por la formación de compuestosresponsables de sabores y olores indeseablescomo ácidos propiónico y butírico. Lafermentación butírica es la fase pútrida de la etapade fermentación y prosigue su curso en las aguasde lavado, dando orígen a las emanacionesnauseabundas en los beneficiaderos.Todos los danos de calidad causados ai grano porfermentaciones prolongadas (sobrefermentaciones) son de tipo químico y en consecuencia sereflejan directamente en las propiedades organolépticas de la bebida. También hay efectossobre la apariencia dei producto (cambio de colordecoloramiento).La gravedad dei dano depende dei tiempo desobrefermentación lo cual determina la apariciónde toda una gama de granos fermentados conolores y sabores indeseables ("off-flavours"), talescomo: fruta (pifia), cebolJa (ácido propiónico),agrio (vinoso), típico fermento (ácido acético),queso (vinagre penetrante) y "stinker" (podrido).EI grano "stinker" puede considerarse como elúltimo estado deI grano sobrefermentado y se caracteriza por su color tabaco, producción devapores ácidos y expulsión dei embrión. Estedefecto tiene especial importancia por lacapacidad que tienen los granos afectados dedanar grandes cantidades de granos que seencuentren a su alrededor. EI origen de estosgranos y su composición química ha lIamado laatención de varios investigadores; por ejemploBOREL et ai (14) han realizado estudios porCG/EM (cromatografía de gases/espectrometríade masas) lo cual les ha permitido pensar quemuchos de los compuestos presentes en el grano"stinker" provienen de fermentaciones por
353
enterobacterias. Así mismo, CHALOT (15) diceque la aparición de los granos "stinker" parece serla consecuencia de malas condiciones de proceso:almacenamiento prolongado de las cerezas antesdeI despulpado, despulpado con agua sucia,almacenamiento deI café después de un secadoinsuficiente.Todos los danos ocasionados a la calidad deI cafédurante el proceso de fermentación son de importancia económica puesto que provocan rechazodei producto por parte de los compradores. Noobstante, estos problemas pueden ser fácilmenteevitados practicando una recolección selectiva deifruto maduro y sano, despulpandoinmediatamente después de la recolección,determinando oportunamente el "punto de lavado"(fin deI proceso de fermentación) y lavando bienel grano con agua limpia.
5.2.3. La etapa de lavado.En la etapa de lavado se retiran todos losproductos de la hidrólisis y fermentación de]mucílago, con. el fin de evitar la formación desabores y olores indeseables en la etapa de secadoy en el proceso de almacenamiento.Hay dos condiciones que deben cumplirse paraasegurara uri buen lavado deI café: que la etapa defermentación haya sido suficiente (completahidrólisis dei mucílago) y que el agua empleadaen la operación sea limpia. Si la primeracondición no se cumple quedan restos demucilago, sobre todo a nivel de la hendidura delcafé pergamino, el cual continua sudescomposición en la etapa de secado y puedegeneral' café fermentado o servir como sustratopara el desarrollo de microorganismos,principalmente hongos, durante el proceso dealmacenamiento, produciendo café con elpergamino manchado y con sabor y olor a moho,general mente conocidos como cafés sucios. Lautilización de aguas sucias genera una serie dedefectos en el café que dependen de la fuente decontaminación; por ejemplo sí el agua estápolucionada con residuos dei beneficio del café,puede originar el café fermentado y aún los cafés"stinker" pero si está contaminada con tierra seproducen los cafés con sabor terroso, otro tipo decafé sucio. Las aguas ferruginosas comunican aicafé un sabor metálico.Las aguas residuales generadas en el beneficio deun kilogramo de café en cereza poseen una capacidad contaminante equivalente a la de unapersona por día (11), expresada en DemandaQuímica de Oxígeno "DQO". Estos costos decontaminación no están siendo suficientementebien reconocidos, por el mercado internacional, alos países productores de cafés suaves lavados.

5.2.4. La etapa de secado.EI secado dei café es una de las operaciones másdelicadas dei proceso de beneficio que puedetener consecuencias desastrosas sobre la calidaddei producto, si no se tienen los cuidadosnecesarios.Durante el secado se rebaja la humedad dei cafédei 52% b. h. (humedad natural dei café verde) a10-12% b. h. Esto con el fin de poder almacenarel producto en condiciones que permitan conservar su calidad ai disminuir la actividadenzimática y el contenido de agua, lo que impidea su vez el desarrollo de hongos, mohos ybacterias durante el almacenamiento.EI secado puede real izarse naturalmente, secado alsol, ó mecánicamente, mediante dispositivosapropiados para el efecto y provistos de cámarasde combustión y ventiladores para el calentamiento e impulsión dei aire. En los dispositivosmecánicos es indispensable la inclusión de un intercambiador de calor para evitar que los gases decombustión entren en contacto con el café y deterioren su calidad.En algunos países, especialmente en Kenia, se leha dado gran importancia ai secado aI sol y algunos investigadores como' McCLOY (17) yGIBSON (18) atribuyen el. desarrollo del colorverde azuloso, de los cafés de alta calidad, a losefectos de la radiación solar durante el secadonatural.Una mala operación de secado puede intluir nosólo sobre la apariencia dei producto crudo sinosobre el gusto de la infusión. Normalmente losdanos en la calidad dei café, generados en elsecado, son debidos a la utilización de altastemperaturas que inducen cambios en lacomposición química de la almendra. ARCILA(19) constató danos en el embrión (pérdida deipoder germinativo) ai usar temperaturassuperiores a 45°C en el secado de la semilla.EI principal defecto ocasionado por las altastemperaturas de secado, sobre todo cuando elgrano tiene altos contenidos de humedad, es elgrano cristalizado; caracterizado por un color grisazuloso y una consistencia frágil y quebradiza.Muchas de las faltas en el proceso de secado, semanifiestan durante el almacenamiento. deI produeto y son debidas básicamente a un secadoexcesivo o deficiente.Una de las primeras características dei grano quese ve afectada, por un mal proceso de secado, esel color verde-azuloso típico de los granos debuena calidad. La descomposición de lassustancias responsables de esta coloración daorigen ai grano blanqueado o decolorado, el cualpuede también formarse por problemas diferentesa fallas en, el secado. Específicamente por
354
problemas de secado puede formarse el granodecolorado-veteado, debido a rehumedecimientodurante o después deI secado y se caracteriza porpresentar vetas blancas en la superficie de laalmendra.Otro tipo de grano decolorado, formado en elsecado, es el grano decolorado-sobresecado oquemado; el cual es de color ámbar o ligeramenteamarillento y es originado por un excesivo tiempode secado. Por el contrario, una falta de secado daorigen ai grano tlojo el cual se caracteriza porpresentar una almendra de color gris oscuro y deconsistencia blanda. Este último defecto esparticularmente importante por la facil idad conque el grano es utilizado, como sustrato, para elcrecimiento de hongos y bacterias; durante elproceso de almacenamiento.
6. BIBLIOGRAFIA
1. -CARVALHO, V. O. de CHALFOUN, S.M.Aspectos qualitativos do cafe. InformeAgropecuario (Brasil) 11(126):79-92; 1985.2. -VIANI R., Coffee, l-History-Botany-TradeProcessing-Chemistry-Nutritional Value,NESTLE, Lausanne (1979).3.-BRESSANI., R.; ESTRADA., E.; JARQUIN.,R. Pulpa y pergamino de café. L-Composición'química y contenido de aminoácidos de laproteína de la pulpa. Turrialba, 22:299-304. 1972.4.-AGUIRRE, B. F., La utilización industrial deigrano dei café y de sus subproductos. Guatemala,Instituto Centroamericano de Investigación yTecnología Industrial (InvestigacionesTecnológicas dei ICAITI, N° I).5.-ZULUAGA, V.J., Contribution á l'étude de lacomposition chimique de la pulpe de café (Coffeaarabica L). Thése de Doctorat es Sciences.Faculté des Sciences, Université de Neuchatel Suisse. 198 J.6.-NADAL N., G.M. Coffee mucilage, itschemical composition. Coffee and Tea Ind. 82 (N°8): 17-18.J959.7.-ASOCIACION NACIONAL DEL CAFE"ANACAFE". Guatemala. Manual Práctico deBeneficio de Café. Boletín N° 13, 1973, 96 p.8.-FEDERACION NACIONAL DECAFETEROS DE COLOMBIA. División deInvestigaciones Económicas. DepartamentoCenso Cafetero. Censo Cafetero Nacional 198081. pp.24.9.-URIBE H., A. Constantes fisicas y factores deconversión en café. Avances Técnicos.CENICAFE (Colombia). Abril 1977. N° 65.IO.-FEDERACION NACIONAL DECAFETEROS DE COLOMBIA. Manual dei

Cafetero Colombiano. Cuarta Edición 1979. pp.168.II.-ARCILA Ó.,F. Contaminación por residuosdei beneficio húmedo de café. Curso sobreCálculo y Disefío de Beneficiaderos de Café.Federación Nacional de Cafeteros de Colombia.Bogotá, 1986. (Mimeografiado).12.-CHALARCA., J. EI café en la vida deColombia. Federación Nacional de Cafeteros deColombia. Bogotá, 1987.13.-BRESSANI., R. Subproductos dei fruto deIcafé. In: BRAHAM., J. E.; BRESSANI., R. (eds.)Pulpa de café: Composición, tecnologia yutílización. Guatemala, Instituto de Nutrición deCentroamérica y Panamá, INCAP-CIID, I 978.p.9-17.14.-BAREL, M.; CHALLOT., F.; VINCENT.,J.c. Contribution á I'étude de féves de cafédefecteuses. Café, Cacao, Thé (Francia). 20(2): I29-134. 1976.15.-CHALLOT., F. Recherche sur I'origine desfeves puantes chez Coffea arabica. Café, Cacao,Thé. (Francia). 29(3):201-208. 1976.16.-GUYOT., B.; CROS., E.; VINCENT., J.c.Caractérisation et identification des compsés de lafraction volatile d'un café vert arábica sain et d'un
355
café vert arabica puant. Café, Cacao, Thé.(Francia). 26(4);279-288. 1982.17.-McCLOY., J.F. Mechanical drying of arabicacoffee. Kenya Coffee (Kenia) 24(280):1 17-133.1959.18.-GIBSON., A. Phototechnical aspects ofdrying East African arabica coffees. In: ColloqueScientifique Intemational sur le Café, 5. Lisbonne(Portugal) I 4-1 9 Juin 1971. Paris (Francia),ASIC, 1971. p.246_258.
19.-ARCILA P., J. Influencia de la temperatura desecado en la germinación de las semillas de café.CENICAFE (Colombia) 27(2);89-91. 1976.20.-PICADO T., C. Méthodes biologiquesd'amelioration du café. Revue de BotaniqueAppliquée et d' Agriculture Tropicale 16 (177);367-372. 1936.21.-COSTE, R. Beneficiado de cafés verdes. In:Coste, R. Cafetos y cafés en el mundo. Paris, Maisonneuve y Larose, 1954 y. 2, p. 9-10 I.22-WILVAUX, R. Beneficio del café.Organización de las Naciones Unidas para laAgricultura y Ia Alimentación. Roma, 1972. 231p.23.-FRITZ, A. EI beneficio deI café sin fermentar.Revista Agricola (Guatemala) Ii (6);2 85-290.1933.


357
POTENCIALIDADE DE USO DE RESÍDUOS E SUBPRODUTOS DO PROCESSAMENTOAGROINDUSTRIAL DO CAFÉ
Celso Luis Rodrigues Vegro J,Flavio Condé de Carvalho 2
I Eng" Agr" Instituto de Economia Agrícola da Secretaria de Agricultura e Abastecimento do Estado de SP.2 Eng" Agr " DS. Professor Titular da Fundação Armando Alvares Penteado - SP.
nutrientes porprocessamento
Fonte: Elaborado a partir de MALAVOLTA &COURY (1967) e MUZILLI (1982).Tendo como referência o volume total de cascasgerado pela safra de 34,5 milhões de sacas obtidaem 1998/99, as quantidades de nutrientes seriamde: 36 mil t de nitrogênio; 2,9 mil t de fósforo;77,4 mil de t de potássio; 8,7 mil t de cálcio; 2,5'mil t de magnésio e 2 mil t de enxofre. Naprodução de solúvel em 1998, o conteúdo denitrogênio presente na borra foi de 120 mil t; 56mil t de cálcio e 37 mil t de magnésio.Ração para animais: a presença de cafeína etaninos nas cascas requer ensilagem prévia oupeletização, podendo ser fornecida desde que nãoultrapasse 20% no total de ração pois o gostoamargo afeta a palatabilidade, além do elevadoteor de lignina. A utilização fica limitada àsregiões produtoras, devido aos custos detransporte. A borra apresenta baixa capacidade
da casca, requerendo secagem prévia até apresentar33% de umidade (CLAUDE. 1979; ADAMS &DOUGAN, 1987; INTERNATI0NAL, 1995).Adubo orgânico: As cascas são adicionadasdiretamente ao solo ou mediante com postagem(pisoteamento e mistura com esterco em curraisou esparramação como cama nos galpões decriação de aves). Podem, ainda, ser enriquecidascom mistura de minerais e adubos químicos. Aborra do solúvel apresenta alguns inconvenientesnesse uso, devido ao alto índice de. acidez, alémda propensão à impermeabilização do solo(CLAUDE, 1979; TANGO, 1971; BARCELOS,et ai, 1992 e NOGUEIRA & FLORES, 1988).
TABELA 1 - Quantidades detonelada de resíduo doagroindustrial do café(em kg)
Nutriente Casca de café Borra de solúvelIN itrogênio 17,5 32
Fósforo 1,4 ...Potássio 37,4 ...Cálcio 4,2 15Magnésio 1,2 10
Enxofre 1,5 ...
Os tipos de utilização mars freqüentementesugeridos incluem:Combustível: Com poder calorífico de 3.500 kcal/kg,as cascas de café podem permitir autonomia energéticaa uma instalação de secagem de grãos e fornecimentode energia para fins diversos. A transformação decascas em Carvão reduz os custos de transporte.concentrando a energia do resíduo. Briquetes: noQuênia, briquetes de cascas de café sãocomercializadas para uso local e exportação para aEuropa. A borra úmida tem o mesmo poder calorífico
Em decorrência da abertura comercial, cria-seambiente de crescente competitividade e deexigências com preservação do meio ambiente,passando as firmas a se preocupar com oaproveitamento de resíduos e subprodutosgerados pela sua atividade de transformação.O processamento da safra brasileira de café de1998/99 resultou em 2, I milhões de toneladas decascas, 3 10m iI toneladas de borra esgotada decafé solúvel e liberou à atmosfera 561 miltoneladas de monóxido de carbono.O objetivo deste estudo é relacionar e discutir osprincipais usos dos resíduos do processamentoagroindustrial do café.
3 - RESULTADOS
2 - MATERIAL E MÉTODOS
1 - INTRODUÇÃO
A estimativa da produção brasileira de café nasafra 1998/99 baseou-se nos números elaboradospela Empresa Brasi leira de PesquisaAgropecuária (SAFRA, 1999). Para aquantificação dos resíduos e subprodutos, foramusados os seguintes coeficientes: 1kg de cafébeneficiado produz quantidade equivalente decascas; I kg de café solúvel produz 1,5kg deborra esgotada (CLAUDE, 1979 citado porVEGRO & CARVALHO, 1994); Ikg de cafétorrado e moído produz 850mg de monóxido de.carbono (DALLA ROSA, M. et al, 1982).Os principais usos dos subprodutos e resíduosforam pesquisados na literatura, incluindopesquisa na Internet.

nutricional e elevado teor de lignina, o quetambém restringe essa utilização (CLAUDE,1979; TANGO, 1971; ADAMS & DOUGAN,1987; NAZÁRIO et ai 1984; CRUZ, 1983).Os resíduos ainda podem ter usos alternativos queem alguns casos encontram-se em faseexperimental. Assim, tem-se:Meio de Cultura e Pectina: a partir damucilagem da polpa do café é possível cultivarfungos e leveduras comestíveis. O teor de pectinana mucilagem compara-se ao dos citrus,tradicional matéria-prima na obtenção industrialdesse produto para 'fins alimentares efarmacêuticos (ADAMS & DOUGAN, 1987;CLAlIDE, 1979).Aguardente e álcool anidro: é possível obter-seaguardente dentro de padrões comerciais a partirda casca de café cereja na proporção de 3,2 litrospara cada 765 litros de café cereja ou 2, I litrospor saca beneficiada. Na obtenção de J litro deálcool são consumidos 3 litros de aguardente.Com o álcool prepara-se licores e perfumes.Esses rendimentos podem ser significativamentemelhorados caso a casca de café cereja passe poretapa prévia de prensagem (COSTE, 1969;SANTINATO et al, 1981; CALLE, 1951).Gás metano: a colocação em biodigestor demistura de 30 kg de polpa parcialmentedecomposta com 18 litros de água e 2 kg deesterco gera cerca de 670 litros de gás. O gásmetano pode ter diversas utilizações desde amovimentação de motores elétricos até oaquecimento de água (ARCI LA, 1979 e CALLE,1974)Vinagre: o conteúdo de açúcares da polpapermite sua utilização em fermentações acéticasque resultam em um tipo de vinagre claro, comaroma de pêra e paladar que lembra whiskyenvelhecido (HAARER, 1962).Baterias Elétricas: a polpa moída pode seruti Iizada na confecção de baterias elétricas pararádios, telefones e campainhas, não se dispondode maiores informações sobre as característicaselétricas das mesmas (CALLE, 1973) ..Papel: a mistura de papel branco reciclado comcascas e borra de café (como corante) perm ite aobtenção de um tipo de papel para fins maisnobres (agendas, cartões, papéis especiais parapresentes, etc ...) (COSTA RICA, 1999). \Óleo essencial: Obtido a partir do cafébeneficiado, o óleo pode ser utilizado cornoveículo aromatizante, desde que o produto sejaestabilizado e protegido da oxidação, por técnicascorno a microencapsulação (CALLE, 1960 eVARGAS et al., 1996).
358
4 - CONCLUSÃO
A gama de possibilidades de utilização dossubprodutos e resíduos do café é elevada. Muitosdos usos ainda não passaram da faseexperimental. Os usos mais comuns têm sidocomo adubo orgânico e combustível. Usos poucocomuns requerem maior desenvolvimentotecnológico mas, eventualmente, poderãoapresentar-se como excelentes opções derentabilidade. A baixa densidade econômica dossubprodutos mais expressivos limita oaproveitamento às regiões mais próximas daszonas produtoras de café. Não se dispõe deavaliação quantitativa do uso desses materiais noBrasil.
5 - BIBLIOGRAFIA
ADAMS, M. R. & DOUGAN, J. Waste products.In: Clarke, Ronald 1. & MACRAE, R. (ed.).Coffee: tecnology. London, Elsevier, 1987. v.2,p.257-81.ARC1LA O., F. Producción de biogás a base depuJpa de café. In: REUNION Internacional sobrela utilización integral de los subproductos de café,2. Guatemala, diciembre 3-5, 1979. Chinchiná,Colômbia, Cenicafe, J979. 17p.BARCELOS, Adauto F. et aI. Aproveitamento dacasca de café a alimentação de novilhosconfinados. Circular Técnica, Lavras (25): 13,dez.1992.CALLE V., H. Aceites deI café. Cenicafé,Colômbia, 11(9):251-58, )960.CALLE V., N. Baterías eléctricas com pulpa ymucílago de café. Avances Técnicos Cenicafé,
Colômbia, nQ25, 1973.4 p.CALLE, V. Como producir gas combustible compulpa de café. Boletin Técnico Cenicafé,
Colômbia, nQ 37, 1974. 11 p.CALLE, V. Producción de alcohol com losdesperdícios dei café. Cenicafé, Colômbia,2(22):33-34, 1951.CLAUDE, B. Étude bibliographique: utilizationdes sous-produits du café.Café Cacao Thé, Paris, 23(2): 146-152, avril-juin1979.COSTA Rica Natural Papel' Company. 1999.http://www.costaricanatural.comCOSTE, René. EI Café. Ed. Blume, Barcelona,Coleccion Agricultura Tropical. 1969. 285p.CRUZ, Geraldo M. Resíduos de culturas eindústria. Informe Agropecuário, BelsoHorizonte, 9( 108):32-37,dez.1983.

DALLA ROSA, M. et aI. Coffee processing:chemical, physical and technological aspects.Industrie delle Bevande, Italia, 11(59):174-178.1982.HAARER, A. E. Modern coffee productionLondon, Leonard HiJls, 1962. 405p.INTERNATIONAL CO-OPERATIVEINFORMATION CENTRE. Chapter IV -Protecting and Conserving Natural Resorces.http://www.wísc.ed uMALAVOLTA, E & COURY, T. Adubação doCafeeiro. In: Graner, E.A.& Godoy Junior, C.(coord.). Manual do Cafeicultor. São Paulo,1967. Cap.VII, p.153-1 96.MUZJLLI, Osmar. Eficiência agronômica daborra de café solúvel para aplicação em cultura deciclo anual. Londrina, Informe de Pesquisa, anoVI, nA3, jun .1982. 5p.NAZÁRIO, Walter; GOMES, L.C; & RETZ,Luciana. Intoxicação em bovinos pela palha decafé utilizada como cama. In: Revista Brasileirade Medicina Veterinária, SP, 6(6): I92-195,nov.ldez.1984.NOGUEIRA, Luíz A. H. & FLORES, Luíz F. V.Potencialidade energética da borra de café para as
359
indústrias de café solúvel. Pesquisa eDesenvolvimento Tecnológico, Itajubá, p.15-25,jan.1988.SAFRA será de 23,15 milhões de sacas. Jornal
do Café, São Paulo, Ano 8, nº 89, jan.l1999,p.18-19.SANTlNATO, R.; MATIELLO, J.B.; SILVA &SILVA, O.A. de. Aguardente e álcool dacasca de café. In: Congresso Brasileiro dePesquisas Cafeeiras, 9, São Lourenço, MinasGerais 27 a 30 de out./1981.TANGO, João S. Utilização industrial do café edos seus subprodutos. Boletim do Instituto deTecnologia de Alimentos - ITAL, Campinas(28):49-73, dez. 1971.VARGAS Z., A. M.; SÁNCHEZ H.. A. F.;CASTANO c., J. J. Microencapsulación de aceitede café. Cenicafê, Colômbia, 47( 1):32-41, 1996.VEGRO, Celso L. R. & CARVALHO, Flavio C.de. Disponibilidade e utilização de resíduosgerados no processamento agroindustrial do café.Informações Econômicas, IEA, SP, 24(1 ):9-16.1994.

11111111111111111111111111111111111111111111111111111111111111111

361
EVALUACJON DE LA MADUREZ DE LACOMPOSTA DE PULPA DE CAFE
Gloria Sánchez, Eugenia J. Olguín, Ricarda Marín y Gabriel Mercado.
Departamento de Biotecnología Ambiental. Instituto de Ecología, A.C. - Km 2.5 ant.carr. a Coatepec.Xalapa,Ver.91 000. México. Fax: (28) 1878 09. E-mail:[email protected]
INTRODUCCION
Desde hace aproximadamente 10 anos, elDepartamento de Biotecnología Ambiental, havenido desarrollando tecnologías que permitandisminuir los impactos ambientales ocasionadospor las descargas de aguas residuales y por losresiduos sólidos orgánicos de la agroindustriacafetalera. Dichas tecnologías están basadas en laaplicación de procesos y acciones congruentescon los principios de producción más limpia,como lo es la minimización y reciclaje de residuosa través de opciones biotecnológicas tales como elcompostaje. La tecnología desarrollada para elcompostaje acelerado de la pulpa de café, essencilla, rápida y eficaz, la cual ya ha sidotransferida a beneficios ubicados en la región deHuatusco, Ver. México. Esta consiste en acelerarel tiempo de estabilización de la pulpa de café,adicionando otros residuos ricos (cachaza ygallinaza) en compuestos fácilmente disponibles,los cuales permiten acelerar el proceso dedegradación, además de mejorar la calidad finalde la composta en términos de N (4.5%) y P(3%)(Sánchez y col., 1999).Un aspecto importante dentro de la calidad de lacomposta, es su grado de madurez, ya que sólocompostas maduras pueden ser adicionadas aIsuelo, sin causar dano a suelos y plantas. El usode composta madura intluye positivamente encultivos debido a la habi Iidad para abastecerlos denutrientes, además de que ha sido demostrado quesu aplicación mejora la porosidad y capacidad deretención de agua, la capacidad amortiguadora deIsuelo e incrementa el porcentaje de materiaorgánica y la capacidad de intercambio catiónico(lglesias y Pérez, 1991).EI objetivo dei presente trabajo fue evaluar lamadurez de la composta obtenida durante elcompostaje acelerado de pulpa de café con y sinaceleradores orgán icos.
MATERIALES Y METODOS
Se montaron pilas de forma comca con unvolumen de 2m 3
, en las instalaciones deI beneficiode café de la Fam. Marini en Huatusco,Ver.Dichas pilas tenían la siguiente composición: PilaI: 100% pulpa de café, Pila 2: 20% cachaza y12% gallinaza y Pila
3: 26% cachaza y 10% gallinaza. Los aceleradoresfueron obtenidos sin costo en las cercanías deibeneficio. La aireación se llevó a cabo una vez porsemana a través de volteo manual. Las muestrasfueron colectadas periódicamente coincidiendocon el volteo de las pilas. Los días de muestreofueron: 0,5,19,33,46,60,74 y 88 (fasebiooxidativa) y 102,123,137,165 Y 207 (fase demadurez). Cada muestra fue de aproximadamente3 kg con una réplica. Los parámetros a determinarfueron: temperatura, capacidad de intercambiocatiónico (de acuerdo a Harada e lnoko I980a), ysustancias húmicas (extracción de acuerdo aBernal y col., 1996) tales como: relación Carbonode ácidos húmicos/Carbono de ácidos fúlvicos(Cah/Caf), índice de humificación (IH)=(Carbonode ácidos húmicos/Carbono oxidable x 100),contenido de ácidos húmicos (determinación deacuerdo a Iglesias y Pérez, 1991).
RESULTADOS Y DISCUSION
Fig.l muestra el perfil obtenido de latemperatura.La fase mesófila (abajo de los 40° C)duró sólo unas horas, ya que la temperatura seincrementó rápidamente con la primera aireacióny en el día I ya se había establecido la fasetermófila (arriba de los 40° C). Esta fase fuesignificativamente más larga en las pilas conaceleradores (37 días) que en las que solo teníanpulpa (25.5 días). Asirnismo, las pilas conaceleradores permanecieron más tiempo en elperíodo óptimo de descomposición de la materiaorgánica (52-60° C), lo cual pudo deberse a laaportación de carbono y nitrógeno fácilmentedisponible presentes en la cachaza y la gallinaza,además de la gran cantidad de microorganismosmuy activos contenidos en la gallinaza.Losresultados anteriores, son semejantes a losreportados en trabajos anteriores (Sánchez y col.,1999). La fase bio-oxidativa duróaproximadamente 88 días, es decir cuando sealcanzó la temperatura ambiente. Posteriormente,se inició la fase de madurez, la cual escaracterizada por Ilevarse a cabo a temperaturaambiente. Durante este período, se continúa con ladegradación parcial de la materia orgánica,transformándola en compuestos más resistentes ala biodegradación, por lo que el proceso semonitoreó hasta el día 207.

362
Las flechas indican cuandolas pilas fueron aireadas.
80
70
Temp 60
era!ura 50
°C40
30
20
10O 10 20 30 40 50 60 70 80 90 100 110 120 130 140 150 160 170
Tiempo (dias)
1-Temp. ambiente -rI _r2 -r3 1
Figura I. Perfil de temperatura durante el compostaje de pulpa decafé con y sin aceleradores.
Tabla 1. Evolución de la capacidad de intercambio catiónico (CIC) durante el cornpostaje de pulpa de cafécon y sin aceleradores (meq/IOO g).
Día Pl P2 P3
O 87.94 65.70 79.59
5 74.79 68.13 80.96
33 86.76 90.04 82.34
60 86.27 93.77 84.05
88 86.50 97.34 94.23
102 86.05 5.82 93.30
123 90.04 93.21 91.56
137 87.36 96.17 86.99
165 86.21 95.65 90.18
207 88.87 99.51 88.80
En la Tabla 1, se observa la evolución de lacapacidad de intercarnbio catiónico (CIC).parámetro que ha sido utilizado para evaluar elgrado de hurnificación dei suelo, ya que unincremento en éste es una de las princi palesconsecuencias dei proceso de madurez deimaterial orgánico (Levi-Minzi y col., 1986).Durante la fase bio-oxidativa, la CIC de la pulpasola no presentó cambios importantes y en losdias 33. 60 y 88 los valores fueron rnuy similares.terminando con un valor parecido aI inicial (88.87meq/I OOg). Por el contrario. cuando losaceleradores fueron adicionados, se observó unincremento gradual en la capacidad deintercambio catiónico durante la fase dernineralización de la matéria orgánica. Este
incremento fue considerable sobre todo en laspilas que tenían mayor contenido de gallinaza(P2) (48.15%). En la fase de madurez. los valoresde P2 fueron inconstantes, sin embargo aI final deiproceso (dia 207) se registró el valor más alto parala CIC en estas pilas (99.51 meq/IOOg). EIcomportamiento presentado por P3 fue diferenteaI de P2, ya que se registró un ligero decrementodei día 88 ai 207. EI incremento de la CIC a lolargo dei proceso en las pilas con aceleradores fuesignificativamente mayor que en la pulpa sola. Loanterior podría significar que los aceleradoresestarían aportando precursores de moléculas conmayor polimerización que las encontradas en la
pulpa sola y produciendo una mayor cantidad degrupos funcionales como los carboxílicos.

363
Tabla 2. Cambios en los parámetros de humificación durante el compostaje de pulpa de café con y sinaceleradores.
r--;---PI P2 P3DIa
Cah/Caf IH Cah/Caf IH Cah/Caf IH
O 1.95 16.62 1.84 15.31 2.06 20.76
5 1.70 16.13 1.91 17.18 1.88 28.70
19 1.81 18.88 1.92 17.06 2.77 18.53
33 2.84 19.24 3.51 21.99 3.63 23.53
46 2.72 20.40 4.18 27.03 4.05 26.58
60 2.55 20.80 2.90 21.86 3.62 23.63
74 2.74 21.09 3.95 25.78 2.83 18.47
88 2.15 21.10 2.58 22.86 2.49 22.83
102 2.05 12.53 2.94 20.89 2.92 13.79
123 1.90 17.03 2.02 21.85 2.91 21.33
137 1.98 12.84 3.20 21.70 3.22 20.03
165 2.04 15.92 3.32 22.93 3.44 22.20
207 1.96 15.00 3.96 23.59 3.70 21.45
Dichos resultados, concuerdan con loencontrado en los parámetros de hurnificación,ya que como se observa en la Tabla 2. larelación Carbono de ácidos húmicos/Carbono deácidos fúlvicos (Cah/Caf) y el índice dehumificación (IH) también se incrementaron a lolargo dei proceso sobre todo en P2 y P3. Larelación Cah/Caf puede ser considerada como unparámetro válido para establecer el grado demadurez de la composta ya que un incrementoen ésta, indica que la materia orgánica está en unproceso de humificación (Bernal y col., 1996).Durante la fase bio-oxidativa, se observó que enP1 dicha relación se incrementó ligeramentealcanzando un valor de 2.05. Sin embargo en lafase de madurez retomó un valor similar ai deldía O, finalizando con un valor de 1.96. Las pilascon aceleradores mostraron un incremento tantoen la fase bio-oxidativa, como en la fase demadurez. P2 inició con un valor de 1.84 (día O) yfinalizá con un valor de 3.96 (día 207) y P3iniciá en 2.06 y termino en 3.70. Los resultadosobtenidos en las pilas con aceleradores están deacuerdo con lo reportado porotros autores. para el compostaje de desechosmunicipales (Iglesias y Pérez , 1992) y debagazo de sorgo du1ce con excretas de animales(Bernal y col., 1996). Ellos encontraron unincremento gradual en esta relación, de valoresmenores de I hasta valores mayores de 3.Asimisrno. el aumento en dicha relación pudohaber sido el resultado de la degradación de lafracción fúlvica y un incremento gradual en la
velocidad de polimerización progresiva decompuestos orgánicos ocurrida durante elcompostaje.Finalmente, uno de los parámetros más importantespara evaluar el grado de madurez de una compostaes el IH, ya que éste representa la fracción de ácidoshúmicos en relación ai carbono presente en lamuestra (Bernal y col., 1996). Durante los primeros46 días se observó un incremento en este parámetroen todas las pilas (Tabla 2), especialmente en lasque contenían los aceleradores que alcanzaronvalores máximos de 27.03% y 26.58% contra20.40% de la pulpa sola. Durante los siguientesdias, se observó un comportamiento ligeramenteirregular en las pilas con aceleradores. Durante lafase de madurez, el comportamiento del IH se tornómás regular. Sin embargo. posteriormente seregistraron incrementos en las pilas conaceleradores principalmente en P2, a diferencia dela pulpa sola, en la cual se observá un decremento.lo que puede indicar que la pila con mayorcontenido de gall inaza tuvo una mejor humificaciónque la pulpa sola. Los valores finales dei IH de laspilas con aceleradores. fueron significativamentemayores (23.56% y 21.45%) en cornparación al dela pulpa sola (15%).Cah/Caf=relación carbono de ácidoshúmicos/carbono de ácidos fúlvicos IH= índice dehumi ficac iónLos ácidos húmicos son la fracción más importantedentro de las sustancias húmicas e indican el gradode humificación y madurez de la composta (Chefetzy col., 1996). AI inicio del proceso, el porcentaje de

dichos ácidos en la pulpa sola y en las pilas conaceleradores fueron muy similares (66 y 64%).Conforme avanzó el proceso, las pilas concachaza y gallinaza tuvieron un comportamientomuy similar en el contenido de ácidos húmicos,mostrando valores de alrededor de 80.68 ai día46. En la fase de madurez, la pulpa sola nopresentó cambios en el contenido de ácidoshúmicos finalizando con un valor similar aiinicial (66.31%). En las pilas con aceleradores,se observó un comportamiento más o menosconstante, finalizando con porcentajes de79.83% para P2 y 78.25% para P3. EI porcentajede ácidos húmicos fue significativamente mayoren las pilas con aceleradores que en la pulpasola. Los cambiosen los porcentajes de ácidoshúmicos en las pilas con aceleradores, tuvieronun comportamiento similar a lo reportado porChefetz y col. (1996), quienes mencionan quedurante el compostaje, los ácidos húmicos llegana ser la fracción principal dentro de lassustancias húmicas. El aumento en C.h/Caf , en elIH y en el contenido de ácidos húmicos en laspilas conteniendo los aceleradores, sugiere quela gall inaza y la cachaza aportan precursores demoléculas que presentan una mayorpolimerización que las de la pulpa sola.
CONCLUSIONES
La adición de los aceleradores promovió laobtención de compostas con un mayor grado demadurez que las obtenidas con pulpa sola, lo quele da un mayor valor agronómico a la compostaobtenida con esta tecnología. Por otro lado, esimportante mencionar, que los trabajosrealizados por los autores y la vinculación conlos productores han permitido Ilevar a cabo lacreación de la Asociación "Pro-BeneficioLimpio", integrada por beneficiadores de laregiónde Zentla, Ver. (México) y en la cualdestaca la implementación de un Programa quepermita la aplicación de procesos y accionesacordes a los principios de producción máslimpia en el agroecosistema cafetalero. contando
364
para ello con la asesoría dei Departamento deBiotecnología Ambiental dei Instituto de Ecologia,A.C. y del Centro Internacional de Tecnologías MásLimpias y Desarrollo Sustentable, CITELDES,A.C.
AGRADECIMIENTOS
Este trabajo fue financiado por el Sistema deInvestigación deI Golfo de México(SIGOLFO)dependiente dei Consejo Nacional deCiencia y Tecnología (CONACYT) de México.
REFERENCIAS
Bernal, M.P., Navarro, F.A., Roig, A., Cegarra, 1.and Garcia, O., 1996. Carbon and nitrogentransformation during composting of sweetsorghum bagasse. Bio!. Ferti!. Soils, 22: 141148.
Chefetz.B; Hatcher, P. G., Hadar, Y. and Chen.Yona. 1996. Chemical and BiologicalCharacterization of Organic Matter duringComposting of Municipal Solid Waste. 1.Environ. Qua!., 25:776-785.
Harada, Y., and lnoko, A., 1980a. TheMeasurement of the Cation-ExchangeCapacity of Composts for the Estimation ofthe Degree of Maturity. Sai! Sei. Plant Nutr..26: 127-134.
lglesias. E. and Pérez Garcia, V., 1991. Compostingof domestic refuse and sewage sludge.I.Evolution of ternperature, pH, C/N ratio andcation-exchange capacity. Resour. ConservoRecy, 6:45-60.
Iglesias,E. and Pérez,V.1992. Determination ofMaturity Indeces for City Refuse Composts.AgI' Ecosyst Environ., 38:331-343.
Levi-Minzi, R., Riffaldi, R., and Saviozzi. A., 1986.Organic Matter and Nutrients in Fresh andMature Farmyard Manure. AgriculturalWastes, 16:225-236.
Sánchez.G; Olguín,EJ. and Mercado.G, 1999.Accelerated coffee pulp composting.Biodegradation, 1O( I ):35-41.

365
CAFFEINE OEGRADATION IN FRESH ANO ENSILEO COFFEE PULP BY SOLID STATEFERMENTATION: INFLUENCE OF SlIBSTRATE PRETREATMENT ANO INOCULUM LEVEL
Romano-Machado J:,Gutiérrez-Sánchez G:, Perraud-Gaime 1.2, Gutiérrez-Rojas M: Y SaucedoCastafieda G. I
'
'Autonornous Metropolitan University, Ave. Michoacán Y La Purísima, [ztapalapa, 09340, México D.F.Fax: (55) 57 24 47 [2. E-Mail: [email protected] De Recherche Pour Le Développment (Ird-México), Cicerón 609. Los Morales, 1 [530. México D.F.
SUMMARYCa.fJeine degradation in fresh (FP) and ensiled (EP) Co.fJee Pulp (CP) using Solid State Fermentation (SSF)was studied. Thermal treatment and difJerent inoculum leveis were also tested in a SSF processo With SSF a95 % in EP and 55 % in FP of ca.fJeine degradation was reached using a Penicillium comune (V33A25)strain after ten days offermentation without thermal treatment. Presence ofenzymatic activities like tannase(tannin-acyl hydrolase) and exopectinase shown the capacity of strain to utilize tannins and peclinsconstituents of CP. Initial massive inoculation of 10- spores/ g moist substrate only 56 % of caffeine wasdegraded in four days of SSF. Nevertheless the diminishing in natural microflora of CP due to thermaltreatment produced a significative increase in degradation up to 94 % for EP and 100 % for FP in four daysoffermentation. Preservation trough silage and the combination ofproper thermal treatment and the use ofselected strains can reduce contents ofantinutritional and antiphysiological compounds in CP establishingits application potential in animal feed.
RESUMENSe estudió la degradación de cafeína en Pulpa de Café (PC) fresca (PF) y ensilada (PE) utilizando laFermentación en Medio Sólido (FMS). Se probó el tratamiento térmico y diferentes niveles de inàculo enFU.'). Por este método de fermentación fue posible degradar 95 % de la cafeína en PE y 55 % usando PFcon la cepa de Pentcillium com une (V33A25) sin un tratamiento térmico previo y después de 10 días deincubacion. Además la presencia de actividades enzimáticas tanasa (tanin-acil hidrolasa) y exopectinasamostro que el hongo es capaz de utilizar además pectinas y taninos presentes en la Pc. La inoculaciónmasiva inicial permitió degradar 56% de la cafeína. en 4 dias de FMS. con un nivel de inoculo de 10esporas/g de sustrato húmedo. Por otra parte el tratamiento térmico disminuyó significativamente lamicroflora nativa lo queevitá contaminaciones en la FMS y permitió aumentar significativamente ladegradación a 100 % en PF y 94 % en PE en 4 dias de fermentación. La conservación por ensilaje y lacombinación de 1111 tratamiento térmico adecuado J' el uso de microorganismos seleccionados permitereducir el contenido de compuestos anttfisiolágicos y antinutricionales en la PC estableciendo su potencialde aplicación en la alimentacion animal.
INTROOUCTION
Coffee Pulp (CP) is an agroindustrial wasteobtained from coffee fruits benefaction processand represent 40 % of the whole fruit mass(Zuluaga, 1989). CP represents a serious pollutionproblem in tropical areas taking to account thelarge volumes of grain produced each season(Aquiahualt et ai .. 1988; Rosussos et ai .. 1995).Conservation and utilizatíon of CP has beendifficult due to, among other factors, its scatterand seasonal production, high moist content (8085 %). presence of nutrient factors(carbohydrates, proteins and mineraJs) whichfaci1itates its destruction for a wide variety ofmicroorganisms. Alternatives proposed for theutilization of CP include its use as animal food,organic composto substrate for enzymeproduction, biogas, mushroorns, vermiculture
(Gaime-Perraud et al.. 1993). Nevertheless itsutilization as animal food is limited by thepresence of antiphysiological and antinutritionalcompounds as caffeine (I J,7-trimethylxantine),tanníns and polyphenols (Roussos et al.. 1995).Porres et ai. in 1993 found reduction in caffeinecontents up to 63 % in 223 days of silage forconservation of 3 t of CP supplemented with canemolasses. More recent studies for conservation ofCP followed by SSF for reduction of caffeínecontent shown that in anaerobic fermentation withlactic acid bacteria inocula during silage of driedCP did not reduce caffeine content but pennitconservation of the ensiled CP (Gaime-Perraud,1995). SSF studies for caffeine reductiondeveloped include isolation and screening offungistrains able to degrade caffeine in dried fresh CPand complete degradation of caffeine in 45 h offermentation with híghly degrading strains

(Aquiahualt et al., 1988; Rosussos et al., 1995).Experiments with EP shown significative caffeinereductions in longer fermentation times (48 to 120h) due to a retard in germination of spores frominoculum caused by the presence of organic acidsproduced during silage (Gaime-Perraud, 1995).One strategy proposed for valorization of CP isconservation of large volumes of CP producedduring crop season trough silage then use SSF fordetoxification of EP and improve its potential ofuse in animal feed. In present work caffeinedegradation in fresh and ensiled CP was analyzed.Thermal treatment and inoculum Ievel were testedas strategies for improve caffeine degradation andstandardization of SSF conditions for scale-up theprocesso
METHODOLOGY
Strain sourceA Penicillium comune strain (V33A25) isolatedfrom the CP and belonging to the lRD-UAMcollection was used. 1t was considered because ofits high efficiency of caffeine degradationreported previously (Aquiahuatl et al., 1988;Roussos et al., 1995; Gairne-Perraud, 1995; Hakilet al., 1998).
Solid fermentation substrates preparationThe fresh coffee pulp (PF) samples were obtainedfrom a coffee processing plant in Jalapa, Veracruzstate, Mexico during the 1997-1998 coffee season(September 97 - March 98). Samples were frozenat -20°C until their utilization. A batch of 400 kgof PF was inoculated with a lactic acid bacteriumstrain, isolated from the ensiled pulp by GaimePerraud in 1995 and named as 80 I. Inoculationlevel was 2.6-5.2xI07 cellg' wet basiscorresponding to 5 % (v:w). Material was ensiled,in 20 kg plastic containers for its conservation andstored until utilization. Lixiviates from 9containers were taken for enzymaticdetermination of dextrose (D+glucose) and Llactic acid by using a YSI instrument. A 10 emthick samples of 1 kg of FP and EP were treatedin an autoclave at atmospheric pressure (95°C) for1 hour using a direct steam stream. Samples weretaken to analyze UFC and moisture content, afie rtreatment samples were dried at ambienttemperature during 24 hour. Samples from thesame batch without thermal treatment were usedas controls for SSF.
Inoculum development and SSF conditionsStrain was 4 times subculture on CSM(Aquiahuatl et al., 1988): commercial coffee usedwas GrandMére "familial"MR, infusion of 40 gl'l
366
was filtered by Whatman paper No. 41, then saltsand agar were incorporated in followingconcentrations (gl'): sucrose 2.0; KHzP04 1.3;NazHP0402HzO 0.12; MgS0407HzO 0.3;CaCl zo2HzO 0.3; agar 20.0. pH was adjusted to5.6 using KOH, 0.1 M and the volume wasadjusted to 1 liter. The culture medi um wassterilized at 121°C for 15 minutes. Cultures weregrown at 30°C for 7 days using Erlenmeyer flaskor Rusk bottles inoculated with 106 spores cm'3The spores were harvest with Tween 80 solution(I %) and magnetic stirring for 20 minutes. Theywere centrifuged to reach concentrations of104,105,106,107 and 108 spores g' wet solid. SSFwas achieved according to the methodologydescribed by Raimbault and Alazard in 1980,using cylindrical glass laboratory reactorincubated at 30°C and with humid saturated airflow of 0.5 vkgm (dm' of air g'1 of medium min").Samples in duplicate were taken for analyticaldeterminations.
Humidity determination, pH and samplestreatment.Humidity of sol id samples was determined bygravimetry. Wet solid samples were milled andresuspended in water in I: 10 (w/w) ratio and pHwas determined with a Conductronic pH 20potentiometer previously calibrated. Thesupernatant were recovered by centri fugation at5000 rpm for 20 minutes and stored at -20°C untiltheir analysis.
Volatile fatty acids (VFA) determinationThe volatile fatty acids (VFA) concentration,glucose and fructose were determined by HPLCusing a Rezek Organic Acid column and HZS04
30 mM as mobile phase at 0.6 cm'min' flow rate.Analyses were carried out at 50°C using arefraction index detector. A flame ionizationdetector (FIO) gas chromatograph was also usedfor VFA analysis.
Caffeine and metabolic intermediaries analysisDried and milled (mesh 35) solid samples wereextracted with a chloroform/isopropanol mixture(50:50, v/v) doing three successive extractions.Solvent was evaporated by heating and the extractwas resuspended in equivalent Milli-Q watervolume. Caffeine and intermediary molecules wasanalyzed by HPLC. using a modification of lhetechnique reported by Denis. 1996: Spherisorb ODS-2column. acetonitrile/tetrahydrofurane/deionizedwater (5: I :94 v/v/v), 1.5 crrr' min" flow rate, room
ternperature, diodes array detector at À=273 nmwere used.
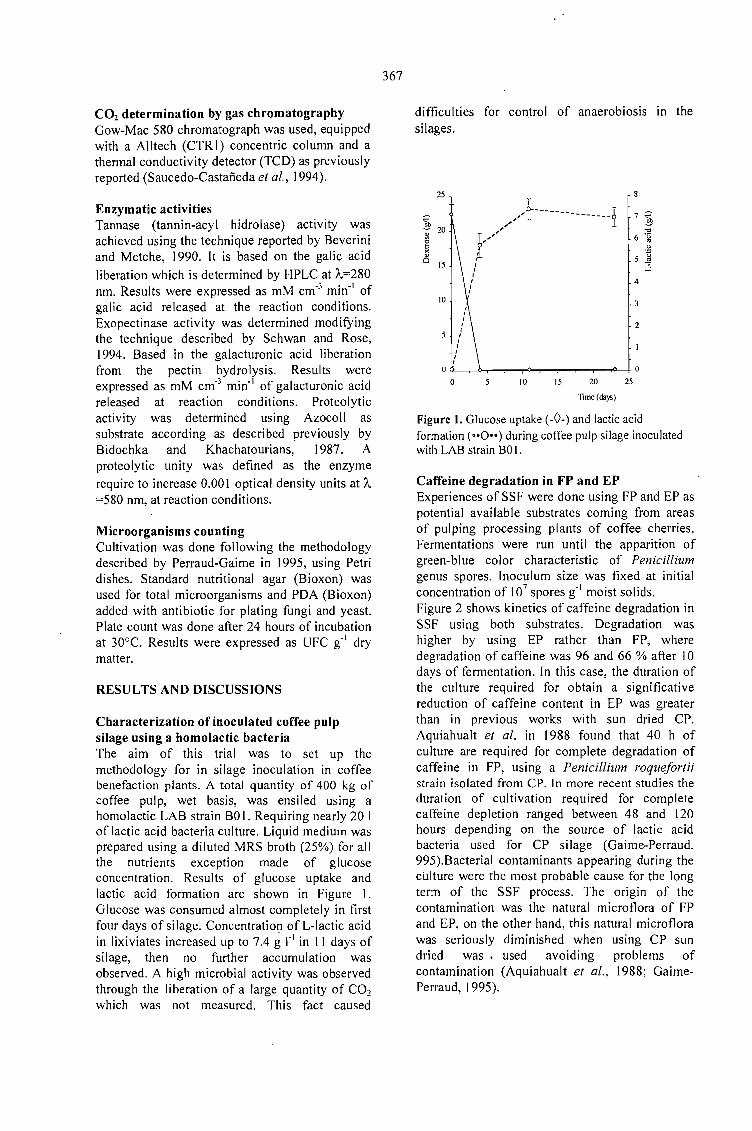
367
CO2 determination by gas chromatographyGow-Mac 580 chromatograph was used, equíppedwith a AlItech (CTR 1) concentric column and athermal conductivity detector (TCO) as previouslyreported (Saucedo-Castafíeda et al., 1994).
difficulties for control of anaerobiosis in thesilages.
4
7 ~'O
6 .~
<.>
5 ~3
20
TIme (da}')
1510
r ~",~ -----------
"r,I,
I,I
I
o<;>---,----<>-r-.....,...---r-o-,.....-~_r__,=__.<>------l-o
o ~
25
10
~';;' 20
glo<
a 15
Figure 1. Glucose uptake (,0-) and lacticacidformation ("00 0
) during coffee pulp silage inoculatedwith LA8 strain 80 I.
Caffeine degradation in FP and EPExperiences of SSF were done using FP and EP aspotential available substrates coming from areasof pulping processing plants of coffee cherries.Fermentations were run until the apparition ofgreen-blue color characteristic of Penicilliumgenus spores. lnoculum size was fixed at initialconcentration of 107 spores g,l moist solids.Figure 2 shows kinetics of caffeine degradation inSSf using both substrates. Degradation washigher by using EP rather than FP, wheredegradation of caffeine was 96 and 66 % after 10days of fermentation. In this case, the duration ofthe culture required for obtain a significativereduction of caffeine content in EP was greaterthan in previous works with sun dried CP.Aquiahualt et ai. in 1988 found that 40 h ofculture are required for complete degradation ofcaffeine in FP, using a Penicillium roquefortiistrain isolated from CP. In more recent studies theduration of cultivation required for completecaffeine depletion ranged between 48 and 120hours depending on the source of lactic acidbacteria used for CP silage (Gaime-Perraud,995).Bacterial contaminants appearing during theculture were the most probable cause for the longterm of the SSF processo The origin of thecontamination was the natural microflora of FPand EP, on the other hand, this natural microflorawas seriously diminished when using CP sundried was , used avoiding problems ofcontamination (Aquiahualt et al., 1988; GaimePerraud, 1995).
Characterization of inoculated coffee pulpsilage using a homolactic bacteriaThe aim of this trial was to set up themethodology for in silage inoculation in coffeebenefaction plants. A total quantity of 400 kg ofcoffee pulp, wet basis, was ensiled using ahomolactic LAB strain BOI. Requiring nearly 20 Iof lactic acid bacteria culture. Liquid medium wasprepared using a diluted MRS broth (25%) for alithe nutrients exception made of glucoseconcentration. Results of glucose uptake andlactic acid formation are shown in Figure 1.Glucose was consumed almost completely in firstfour days of silage. Concentration of L-lactic acidin lixiviates increased up to 7.4 g 1'1 in lI days ofsilage, then no further accumulation wasobserved. A high microbial activity was observedthrough the liberation of a large quantity of CO2
which was not measured. This fact caused
Microorganisms countingCultivation was done following the methodologydescribed by Perraud-Gaime in 1995, using Petridishes. Standard nutritional agar (Bioxon) wasused for total microorganisms and POA (Bioxon)added with antibiotic for plating fungi and yeast.Plate count was done after 24 hours of incubationat 30°e. Results were expressed as UFC s' drymatter.
RESULTS AND D1SCUSSIONS
Enzymatic activitiesTannase (tannin-acyl hidrolase) activrty wasachieved using the technique reported by Beveriniand Metche, 1990. lt is based on the galic acid
liberation which is determined by HPLC at À,=280nm. Results were expressed as mM em') min" ofgalic acid released at the reaction conditions.Exopectinase activity was determined modifyingthe technique described by Schwan and Rose,1994. Based in the galacturonic acid liberationfrom the pectin hydrolysis. Results wereexpressed as mM cm,3 min" of galacturonic acidreleased at reaction conditions. Proteolyticactivity was determined using Azocoll assubstrate according as described previously byBidochka and Khachatourians, 1987. Aproteolytic unity was defined as the enzyme
require to increase 0.001 optical density units at À,=580 nm, at reaction conditions.
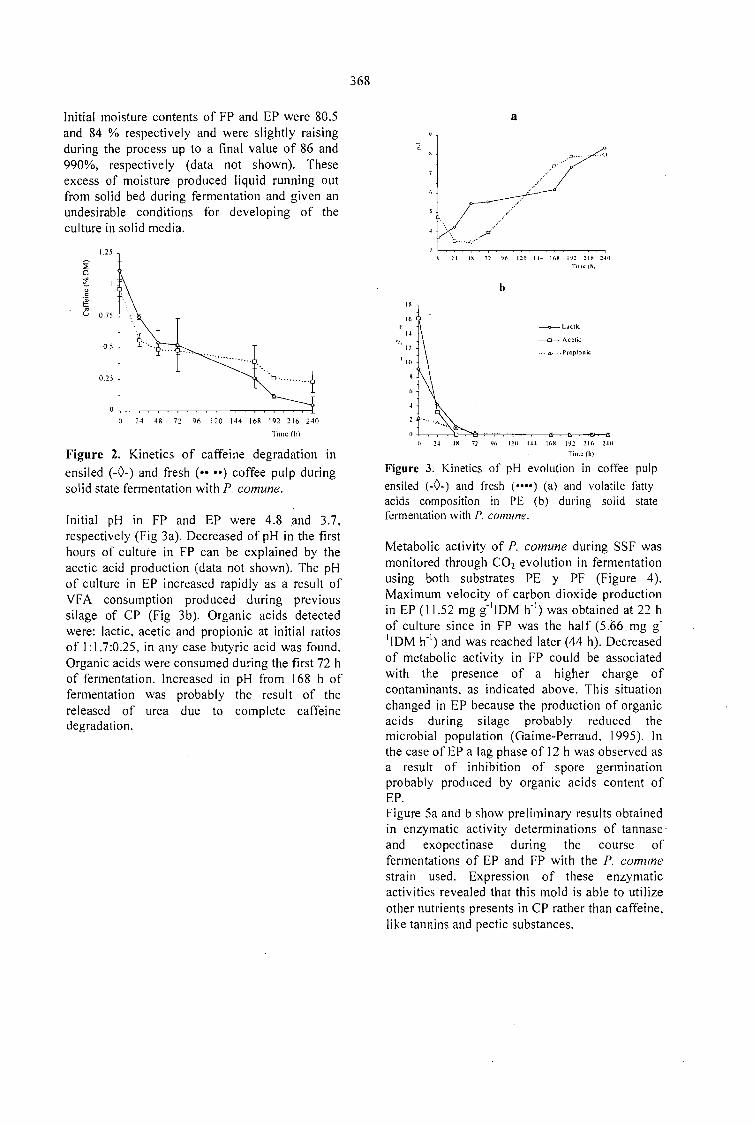
368
Initial moisture contents of FP and EP were 80.5and 84 % respectively and were slightly raísingduring the process up to a final value of 86 and990%, respectively (data not shown). Theseexcess of moisture produced liquid running outfrom solid bed during fermentation and given anundesirable conditions for developing of theculture in solid media.
a
.··0
.....,.'
•••• 1:1G .... .o"
1.25
iCl
e~
c';
~ 00
i:: 0.75"
••• â, ••• Propionic
----o-- Lactlc
---o-- Acerte
110
1(,
F
"'\I"
12
b
lj 1-1 -111 72 96 11ft I·H 1(,11 I'J1 11(. 2-10
TllIle (hl
t -;
o 24 48 72 96 120 144 168 192 216 240
Time (h)
05
0,25
Figure 2. Kinetics of caffeine degradation inensiled (-0-) and fresh (••••) coffee pulp duringsolid state fermentation with P. comune.
Initial pH in FP and EP were 4.8 and 3.7,respectively (Fig 3a). Decreased of pH in the firsthours of culture in FP can be explained by theacetic acid production (data not shown). The pHof culture in EP increased rapidly as a result ofVFA consumption produced during previoussilage of CP (Fig 3b). Organic acids detectedwere: lactic, acetic and propionic at initial ratiosof 1:1.7:0.25, in any case butyric acid was found.Organic acids were consumed during the first 72 hof fermentation. Increased in pH from 168 h offermentation was probably the result of thereleased of urea due to complete caffeinedegradation.
n 2-1 -lI( 72 'J6 120 IH 16K Jl.l2 216 2·10
Time (h)
Figure 3. Kinetics 01' pH evolution in coffee pulpensiled (-0-) and fresh (.... ) (a) and volatile fattyacids composition in PE (b) during solid stateferrnentation with P. comune,
Metabolic activity of P. comune during SSF wasmonitored through CO2 evolution in fermentationusing both substrates PE y PF (Figure 4).Maximum velocity of carbon dioxide productionin EP (11.52 mg g"IDM h'l) was obtained at 22 hof culture since in FP was the half (5.66 mg sIIDM h'l) and was reached later (44 h). Decreasedof metabo\ic activity in FP could be associatedwith the presence of a higher charge ofcontaminants, as indicated above. This situationchanged in EP because the production of organicacids during silage probably reduced themicrobia1 population (Gairne-Perraud, 1995). Inthe case of EP a lag phase of 12 h was observed asa result of inhibition of .spore germinationprobably produced by organic acids content ofEP.Figure 5a and b show preliminary results obtainedin enzymatic activity determinations of tannaseand exopectinase during the course offermentations of EP and FP with the P. comunestrain used. Expression of these enzymaticactivities revealed that this mold is able to utilizeother nutrients presents in CP rather than caffeine,like tannins and pectic substances.
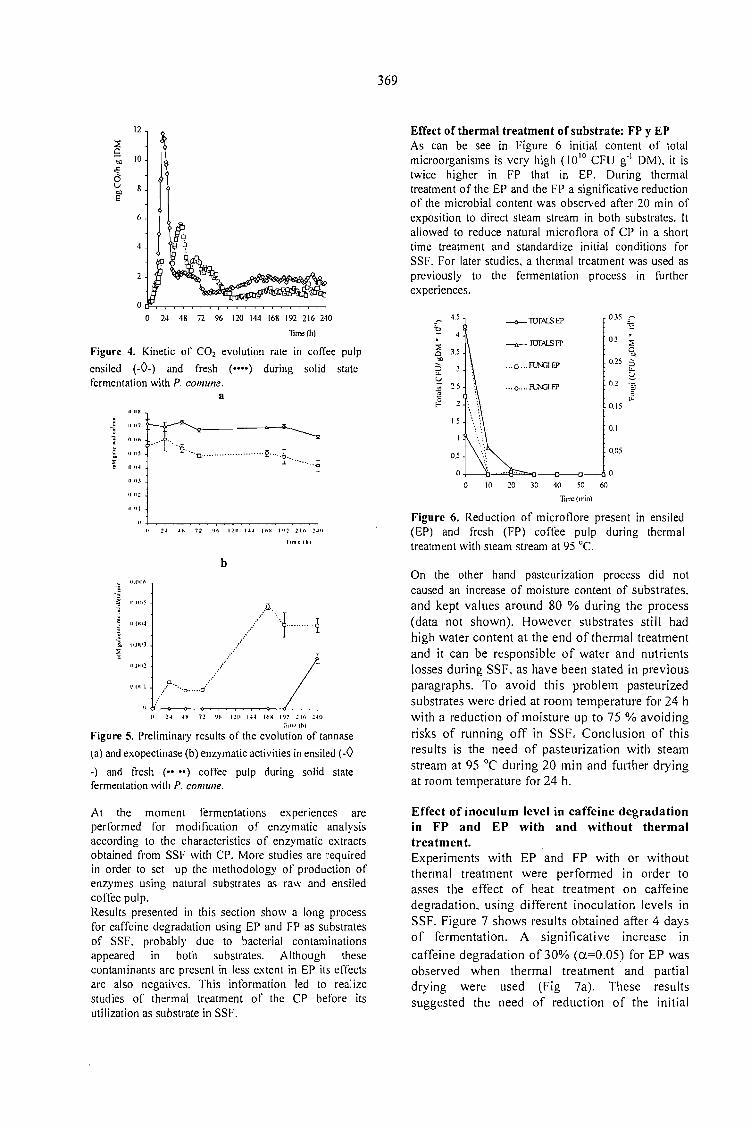
369
~ ."2- ···0······· 2· ~ i
4.5---4- lOTAlS EP 035
~'O
--tr- ·JUrAlSFP 03 z~ 3.5 ClCl oi,eo 0.25 ::,::, c,
u,~
~25 0.2 .~)~
-aÕ u,f-- t 0.15
150.1
0.50.05
+----b.......,."l:!""""',o---{}---o---ó o10 20 30 40 50 60
11m:(minl
Figure 6. Reduction 01' microtlore present in ensiled(EP) and fresh (FP) coffee pulp during thermaltreatment with steam stream at 95°C.
Effect of thermal treatment of substrate: FP y EPAs can be see in Figure 6 initial content 01' totalmicroorganisms is very high (10 10 CFU gol DM). it istwice higher in FP that in EP. During thermaltreatment 01' the EP and the FP a significative reduction01' the microbial content was observed after 20 min ofexposition to direct steam stream in both substrates. ltallowed to reduce natural microtlora 01' CP in a shorttime treatrnent and standardize initial conditions forSSF. For later studies, a thermal treatment was used aspreviously to the fermentation process in furtherexperiences.
I'):! 21(. :!JIl'H. 1211
12
z9
10ec
'"cSu'"E
6
11-DI:)i;J q
Ó
~~ lIU7 t-r~'-...,>--~-~~~
~ o IH.
'[ u.os~
. :::: Ili n.:!
n 01
"f.---.-__~_~~ _
li :!~ oi K 72
O O"'-r---.-~---.-~---.-~---.-~---.-~~~~
O 24 48 72 96 120 144 168 192 216 240
11",::(h)
Figure 4. Kinetic 01' COz evolution rate in coffee pulp
ensiled (-0-) and fresh (....) during solid statefermentation with P. comune.
a
b
(I :!J -IX 7! ')(, 12[1 IH IMI l')~ '::1(, .:;·wlíme Ih.
Figure 5. Preliminary results of the evolution of tannase
(a)and exopectinase (b) enzymatic activities inensiled (-O-) and fresh (•••• ) coffee pulp during solid statefermentation withP. comune.
0.1111(,
~~~ " 1l11~.~
"s ll,l1ll4s~~ 0.1'\1.1:EE
lI,oll1
\1,1111\
.......9.-,
..·l.... ~............... I
:...P-····.o •... ri
On the other hand pasteurization process did notcaused an increase 01' moisture content 01' substrates,and kept values around 80 % during the process(data not shown). However substrates still hadhigh water content at the end of therma1 treatmentand it can be responsible of water and nutrientslosses during SSF. as have been stated in previousparagraphs. To avoid this problem pasteurizedsubstrates were dried at room temperature for 24 hwith a reduction of moisture up to 75 % avoidingrisks of running off in SSF, Conclusion of thisresults is the need of pasteurization with steamstream at 95°C during 20 min and further dryingat room temperature for 24 h.
At the moment fermentations experiences areperformed for modification 01' enzymatic analysisaccording to the characteristics 01' enzymatic extractsobtained from SSF with CP. More studies are requiredin order to set up the methodology 01' produetion ofenzymes using natural substrates as raw and ensiledcoffee pulp.Results presented in this section show a long processfor caffeine degradation using EP and FP as substrates01' SSF. probably due to bacterial contaminationsappeared in both substrates. Although thesecontaminants are present in less extent in EP its effectsare also negativcs. This information led to realizestudies 01' thermal treatrnent of the CP before itsutilizationas substrate in SSf.
Effect of inoculum levei in caffeine degradationin FP and EP with and without thermaltreatment.Experiments with EP and FP with or withoutthermal treatment were performed in order toasses the effect of heat treatment on caffeinedegradation, using different inoculation leveIs inSSF. Figure 7 shows results obtained after 4 daysof fermentation. A significative increase in
caffeine degradation of 30% (a=0.05) for EP wasobserved when therma1 treatment and partialdrying were used (Fig 7a). These resultssuggested the need of reduction of the initial
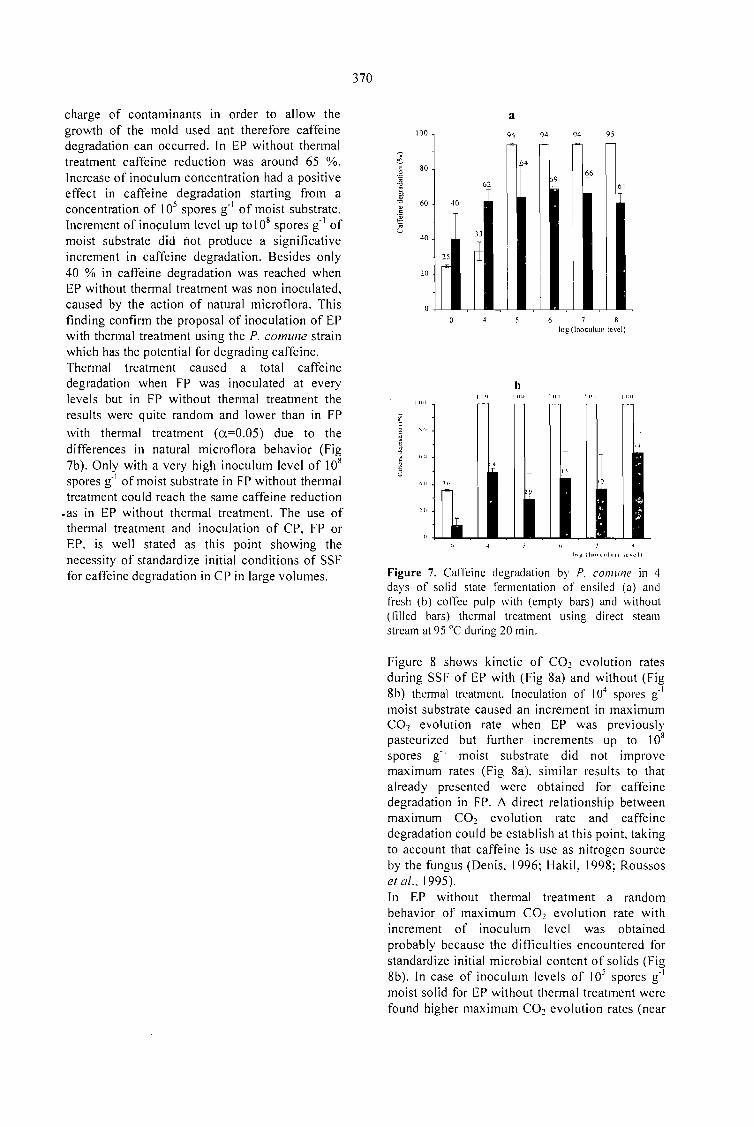
370
a
7 8log (lnocuhun lcvel )
Figure 7. Caffeine degradation by P. comune in 4days of solid siate ferrnentation of ensilcd (a) andfresh (b) coffee pulp with (empty bars) and without(Iilled bars) thermal treatment using direct steamstream at 95 DC during20 mino
j un11'1.1 11 fi
b: IIU111 11
94 94
r= r-e- ~ r-r-
1'466
962 61
40
JJ
25 ~Ir*
II
- c- c- - c-
"
'I ,
",- I
~~ -
·1 li
",
ruo
100
t.~ 80.~
"~'O 60~
~
~v
40
20
charge of contaminants in order to allow thegrowth of the mold used ant therefore caffeinedegradation can occurred. In EP without thermaltreatment caffeine reduction was around 65 %.lncrease of inoculum concentration had a positiveeffect in caffeine degradation starting from aconcentration of 105 spores g'l of moist substrate.Increment of inoculum levei up to I08 spores g'l ofmoist substrate did not produce a significativeincrement in caffeine degradation. Besides only40 % in caffeine degradation was reached whenEP without thermal treatment was non inoculated,caused by the action of natural microflora. Thisfinding confirm the proposal of inoculation of EPwith thermal treatment using the P, comune strainwhich has the potential for degrading caffeine.Thermal treatment caused a total caffeinedegradation when FP was inoculated at everylevels but in FP without thennal treatment theresults were quite random and lower than in FP
with thermal treatment (a=0.05) due to thedifferences in natural microflora behavior (Fig7b). Only with a very high inoculum levei of 108
spores g'l ofmoist substrate in FP without thermaltreatment could reach the same caffeine reduction
.as in EP without thermal treatment. The use ofthermal treatment and inoculation of CP, FP orEP. is well stated as this point showing thenecessity of standardize initial conditions of SSFfor caffeine degradation in CP in large volumes.
Figure 8 shows kinetic of CO} evolution ratesduring SSF of EP with (Fig 8a) and without (Fig8b) thcrmal trcatment. lnoculation of 104 spores golmoist substrate caused an increment in maximumCO} evolution rate when EP was previouslypasteurized but further increments up to 108
spores gol moist substrate did not improvemaximum rates (Fig Sa), similar results to thatalready presented were obtained for caffeinedegradation in FP. A direct relationship betweenmaximum CO} evolution rate and caffeinedegradation could be establish at this point, takingto account that caffeine is use as nitrogen sourceby the fungus (Denis, 1996; Hakil, 1998: Roussoset ai.. 1995).Jn EP without thermal treatment a randombehavior of maximum CO} evolution rate withincrement of inoculum levei was obtainedprobably because the difficulties encountered forstandardize initial microbial content of solids (Fig8b). In case of inoculum leveis of 105 spores g'moist solid for EP without thermal treatment werefound higher maximum CO} evolution rates (near
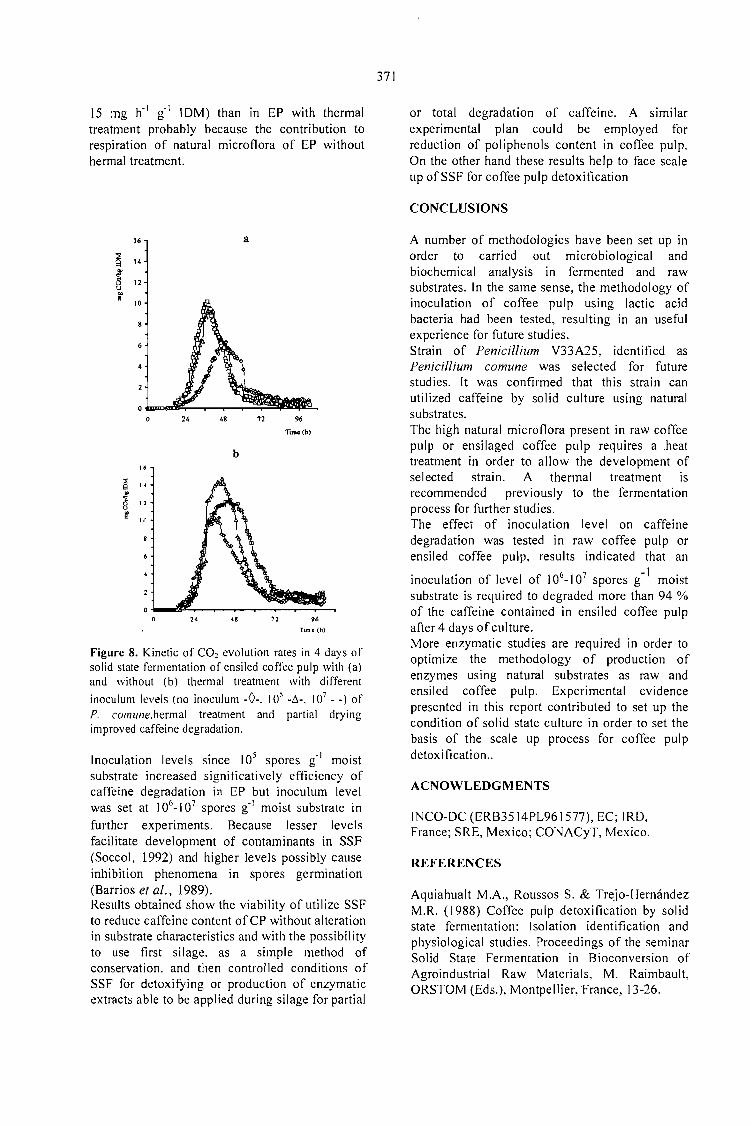
371
15 mg h,l gol 10M) than in EP with thermaltreatment probably because the contribution torespiration of natural microflora of EP withouthermal treatment.
01' total degradation of caffeine. A similarexperimental plan could be employed forreduction of poliphenols content in coffee pulp.On the other hand these results help to face scaleup ofSSF for coffee pulp detoxification
CONCLUSIONS
Figure 8. Kinetic of CO2 evolution rates in 4 days 01'
solid state fermentation of ensiled coffee pulp with (a)and without (b) therrnal treatrnent with different
inoculum leveis (no inoculum -0-. 10; -L'.-. 107- -) 01'
F cOlllune.hermal treatment and partial dryingimproved caffeine degradation.
Inoculation leveis since 105 spores gol moistsubstrate increased significatively efficiency ofcaffeine degradation in EP but inoculum leveiwas set at 106_107 spores gol moist substrate in
further experiments. Because lesser leveisfacilitate development of contaminants in SSf(Soccol, 1992) and higher leveis possibly causeinhibition phenomena in spores germination(Barrios et ai .• 1989).Results obtained show the viability of utilize SSFto reduce caffeine content of CP without alterationin substrate characteristics and with the possibil ityto use first silage, as a simple method ofconservation. and then controlled conditions ofSSF for detoxifying or production of enzymaticextracts able to be applied during silage for partial
REFERENCES
INCO-DC (ERB3514PL961577), EC; IRD.France; SRE, Mexico; CONACyT, Mexico.
ACNOWLEDGMENTS
Aquiahualt M.A., Roussos S. & Trejo-HernándezM.R. (1988) Coffee pulp detoxification by solidstate fermentation: Isolation identification andphysiological studies. Proceedings of the seminal'Solid State Fermentation in Bioconversion ofAgroindustrial Raw Materiais, M. Raimbault,ORSTOM (Eds.), Montpellier, France, 13-26.
A number of methodologies have been set up inorder to carried out microbiological andbiochemical analysis in fermented and rawsubstrates. In the same sense, the methodology ofínoculation of coffee pulp using lactic acidbacteria had been tested, resulting in an usefulexperience for future studies.Strain of Penicillium V33A25, identified asPenicillium comune was selected for futurestudies. lt was confirmed that this strain canutilized caffeine by solid culture using naturalsubstrates.The high natural microflora present in raw coffeepulp 01' ensilaged coffee pulp requires a .heattreatment in order to allow the development ofselected strain. A thermal treatment isrecomrnended previously to the fermentationprocess for further studies.The effect of inoculation levei on caffeinedegradation was tested in raw coffee pulp 01'
ensiled coffee pulp, results indicated that an
inoculation of levei of 106_107 spores gol moistsubstrate is required to degraded more than 94 %of the caffeine contained in ensiled coffee pulpafter 4 days of culture.More enzymatic studies are required in order tooptimize the methodology of production ofenzymes using natural substrates as raw andensiled coffee pulp. Experimental evidencepresented in this report contributed to set up thecondition of solid state culture in order to set thebasis of the scale up process for coffee pulpdetoxification ..
96
Time (h)
96
lime (b)
72
a
b
••
48
2'
24
\6
16
~ l'
~~ 12
i!' tO

Barrios J., Martínez c., Aguilera A. & RaimbaultM. (1989) Germination of concentratedsuspensions of spores from Aspergillus niger.Biotechnol. Lett., 11: 551-554.Beverini. M. & Metche M. (1990) Identification,purification and physicochemical properties oftannase of Aspergillus orizae. Sciences desAliments, 10: 807-816.Bidochka J. & Khachatourians G.G. (1987)Purification and propertíes of an extracellularprotease produced by the entomopathogenicfungus Beauveria bassiana. Appl. and Environm.Microbiol., 53: 1679-1684.Denis S. (1996) Degradation de la caffeine parAspergillus sp. et Penicillium sp.: Etudephysiologique et biochimique. Thêse de Doctorat,Université Montpellierll, 180 p.Gaime-Perraud I. (1995) Cultures mixtes enmilieu solide de bactéries lactiques et dechampignons filamenteux pour la conservation etla decafeination de la pulpe de cafe. Thêse deDoctorat, Université Montpellier 11, 210 p.Gaime-Perraud 1., Roussos S. & Martínez-CarreraD. (1993) Natural microorganisms in the freshcoffee pulp. Micol. Neotrop. Apl.. 6: 95-103.
• Hakil M., Denis S., Viniegra-González G. &Augur C. (1998) Degradation and productanalysis of caffeine and related d imethylxanthinesby filamentous fungi. Enzyme and MicrobialTechnology, 22:355-359.Porres C; Alvarez D. & Calzada J. (1993)Caffeine reduction in coffee pulp through silage.Biotech. Adv.. 1I,Raimbault M. & Alazard D. (1980) Culturemethod to study fungai growth in solidfermentation. Europ. Jour. of Appl. Microbiol.and Biotechnol, 9: 199-209.
372
Roussos S., Aquiahualt M. A., Trejo-HernándezM. R., Gaime-Perraud 1., Favela-Torres ·E.,Ramakrishna M., Raimbault M. & ViniegraGonzález G. (1995) Biotechnologicalmanagement of coffee pulp-isolation, screeníng,characterization, selection of caffeine-degradingfungi and natural microf1ora present in coffee pul pand husk. Appl. Microbiol. Biotechnol., 42: 756762.Saucedo-Castafíeda G., Trejo-Hernández M. R.,Lonsane B. K., Navarro J. M., Roussos S. &Raimbault M. (1994) On-line automatedmonitoring and control systems for CO 2 and O2 inaerobic and anaerobic solid-state fermentations.Process Biochemistry, 29: 13-24Schwan R.F. & Rose A.H. (1994)Polygalacturomase production by Kluyveromycesmarxianus: effect of medium composition . .four.ofAppl. Bacteriol.. 76: 62-67.Soccol C.R. (1992) Physiologie et metabolisme deRhizopus en culture sol ide et submergée enrelation .avec la degradation d'amidon cru et de laproduction d'acide L(+) lactique. Thése deDoctorat. Université de Technologie deCompiegne Francia, 2 J 8p.Zuluaga V.J. (1989) Utilización integral de lossubproductos deI café. In: Roussos S., Licona F.R.y Gutierrez M. (Eds.), Memorias dei I SeminarioInternacional sobre Biotecnología en laAgroindustria Cafetalera (I-SIBAC), México. pp63-76.

373
EVALUATION OF Penicillium sp. V33A25 CAFFEINASE ACTlVITY IN RELATION TO ITSCONSERVATION METHOO
Gutiérrez-Sánchez c.', Perraud-Gaime 1.2, Augur c.', Romano-Machado J.1,and Saucedo-Castaõeda c.'
Departamento de Biotecnología, Universidad Autónoma Metropolitana, MEXICO2IRD-México, Institut de Recherche pour le Dêveloppement, FRANCIA'Corresponding author: Dr. Saucedo-Castafieda G., Departamento de Biotecnología,Universidad Autónoma Metropolitana, Unidad IztapalapaAv. Michoacán y La Purísima SIN, Iztapalapa, A.P. 55-535, México, D.F., c.r. 09340, MEXICOFax + (5)724-47-12, e-rnail: [email protected]
Key words: caffeine, degradation, induction
ABSTRACTCaffeine degradation by Penicillium sp. strain V33A25 (P V33A25) was evaluated using a liquid culturemedium prepared from a coffee infusion with an initial caffeine concentration of 0.5 g/L. After caffeineactivity of the strain P V33A25 was induced, strain conservation was carriedout four methods: I)conservedin a coffee-agar medium, 2) coffee pulp-agar medium, and for the next two methods, the spores wereharvested from 3) potato dextrose agar (PDA) medi um, and 4) coffee-agar. The spores were Iyophilized inthe last two cases. The strain was adapted three times by subculturing on coffee agar medium before beenused for the evaluation ofcaffeine degradation ability. lt was observed that using methods 3 and 4 the fungibegin to degrade caffeine after 60 and 36 hours of culture, respectively. Nevertheless, with method 1, thefungi starts to degrade caffeine 12 hours afier the inoculation time, reaching 92 % ofcaffeine degradation ina 72 hour period; with method 2, 39 % ofdegradation was observed within 72 hours but it was necessary tomake 6 precultures to reach a similar degradation rate with method 2. The results of this study, suggest thatthe enzymatic activity responsiblefor caffeine degradation is inducible and once caffeine degradation abilityis expressed it is important to conserve the strain in a medium with caffeine.
RESUMENSe evaluó la degradación de cafeína por Penicillium sp. strain V33A25 (PV33A25) empleando un media decultivo líquido preparado a partir una irfusián de café con una concentración inicial de cafeína de 0.5 g/L.Una vez inducida la actividad cafeinasa de la cepa P V33A25, se procedió a la conservación de dicha cepausando cuatro métodos: 1) la cepafue conservada en media café agar; 2)en media pulpa de café agar y enlos otros dos métodos las esporas se cosecharon a partir de medio, 3) agar papa dextrosa y 4) café agar, ylas esporas fueron liofilizadas en los estas últimos dos casos. La cepa file resembrada tres veces en mediocafé agar, antes de ser cultivada en el media café líquido para evaluar su capacidad de degradación decafeina . Se observo que con el método de conservación 3 y 4 el hongo empieza a degradar la cafeínadespués de las 60 y 36 horas de cultivo, respectivamente. Sin embargo, con el método de conservación 1, elhongo comienza a degradar la cafeína a las 12 horas, !legando a un 92 % de degradación de cafeína en 72horas; con el método 2, hubo una degradación dei 39 % a las 72 horas, pera es necesario hacer 6resiembras para alcanzar una degradacián similar a la obtenida por el método 2. Los resultados de esteestudio, sugieren que la actividad enzimática responsable de la degradación de la cafeína es inducible y unavez expresada dicha actividad es importante conservar la cepa en un media que contenga cafeína.
INTROOUCTlON
Coffee pulp is one of the major subproductsavailable produced during the pulping operationof coffee cherries. One fraction of this material isused for compost production for coffee plantnurseries and the other part is spilled in rivers orpiled up near them. For the 1997-98 period 14.92milJion tons offresh coffee pulp were produced inthe world (Barreiro, 1999) and for every 2 toncoffee cherry processed, nearly I ton pulp is
generated. Coffee pulp is essentially rich inproteins, minerais and fiber, nevertheless itsutilízation in animal feed is reduced notably dueto the presence of toxic compounds difficult todegrade. such as caffeine, phenolics and tannins(Roussos, J989). These compounds cause adverseeffects on the animals that consume coffee pulp.Silage of coffee pulp for conservation and its foodvalue improvement, is an alternative for latterutilization of the coffee pulp. Silage is ananaerobic and fast processo which involves lactic

acid bacteria. It has been widely used for foragepreservation in regions of moderate cl irnate.Silage enables prevention of forage putrefactionwith minimum degradation of the organicmaterial. The process is faster and improvesforage nutritional quality (Perraud-Gaime, 1995).PefiaJoza in 1985, Gómez in 1985, and Aquiáhualtin 1992, carried out solid state fermentationstudies with coffee pulp with an Aspergillus nigerstrain, in order to evaluate the improvement of thenutritional value. They reported proteicenrichment of the substrate but with no significantcaffeine elimination. In this sense, utilization ofcoffee pulp for feed is constrained byantiphysiologicaJ (caffeine) and antinutritional(polyphenols) compounds.The aim of this work was to study the effect of theexpression of Penicillium sp. strain V33A25caffeine-degrading enzymes after variousconservation methods.
MATERIALS AND METHODS
MicroorganismPenicillium sp. strain V33A25 used in this study,came from the IRD-UAM collection (Roussos,1995) and' was selected because of its ability touse caffeine as nitrogen source (Roussos, 1995;and Denis 1996).
Culture mediaThe culture media composition used in this workare shown in Table I. For CSA and CS,cornrnercial coffee used was GrandMêre"farnilial", The infusion was filtered by Whatmanpapel' No. 41, salts were added, pH was adjustedto 5.5 using KÜH, 0.1 M, agar was added and theend. The volume was adjusted to I L. In the caseof CPA, fresh coffee pulp was milled in a blenderfor 5 minutes, then was heated until boiling, afteraddition of 750 mL of distilled water saltssolution and agar were then incorporated. Theculture medium was sterilized at 121°C for 15minutes.
374
Table 1. Culture media used for strainconservation and spore harvest (Aquiahuatl, 1992;Denis, 1996).
Component Medium (g/L)CPA CSA CS
Saccharose --- 2.0 2.0
Milled coffee --- 40.0 40.0
Fresh coffee pulp 427.7 --- ---KHzP04 1.73 1.3 1.3
NazHP04"2HzO 0.16 0.12 0.12
MgS04"7HzO 0.4 0.3 0.3
CaCl z"2HzO 0.4 0.3 0.3
Agar 26.7 20.0 ---CPA: Coffee pulp agar rnediurn; CSA: Coffeeagar medium;CS: Coffee saccharose medium
Culture conditions and strain conservationFour methods were used for the evaluation ofstrain conservation associated to caffeinedegradation: I) strain was conserved on CSA, and2) on CPA, for the next two methods, the sporeswere harvested from 3) PDA and 4) CSA and inboth cases spores were Iyophilized. The selectedstrain was precultured twice on CSA at 30°C for 6days and caffeine degradation were evaluated onCS medi um according to Denis methodology(Denis, 1996).
Caffeine analysisCaffeine was extracted from liquid sarnples with 1ml, of a chloroform/isopropanol mixture (85: 15;v/v) mixing in a vortex during 30 seconds andrecovering the organic phase until threesuccessive extractions. Solvent was evaporated ina heater and the extract was resuspended in 1.5ml, of deionized water. Caffeine was analyzed byHigh Performance Liquid Chromatography(H PLC), using a modification to the techniquereported by Denis, 1996; Sphersorb üDS-2column, acetonitrile/tetrahidrofurane/deionizedwater (5: I :94; v/v/v), 1.5 rnl-rnin' flow rate,room ternperature, diodes array detector at Â=273nm were used.
RESlILTS AND DlSClISSION
In Figure I, caffeine concentrations related to theconservation methods proposed (seemethodology), are shown. When the strain wasconserved with the method I 01' 2, that isprecultured previously 3 and 6 times on CSA,respectively, the degradation presented a 12 hourdelay phase and degradation rates were near 0.0 Ig'L,I'h,I, in both cases, reaching 92 %
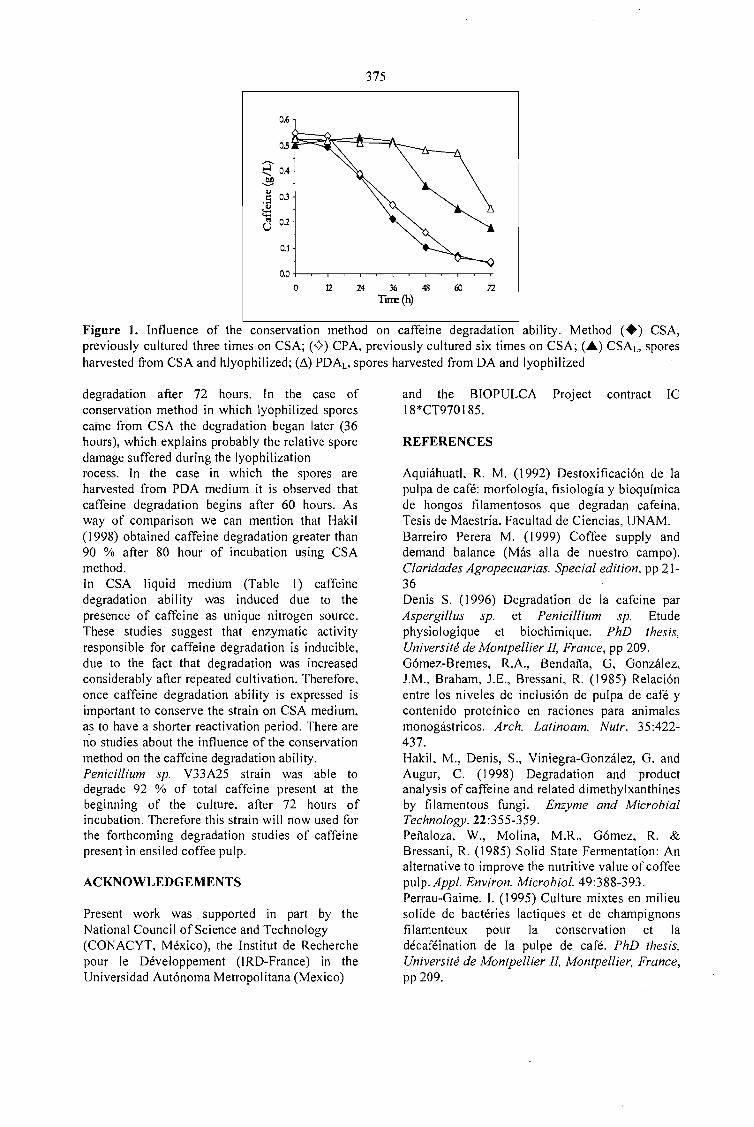
375
0.6
05
2' 0.4~
lU0.3c::.s
'-OI 0.2U
0.1
0.0
O 12 24 36 48 60 72Ture(h)
Figure 1. Influence of the conservation method on caffeine degradation abiJity. Method (+) CSA,previously cultured three times on CSA; (<» CPA, previously cultured six times on CSA; c.") CSAL, sporesharvested from CSA and hlyophilized; (~) PDAL, spores harvested from DA and lyophilized
degradation after 72 hours. In the case ofconservation method in which Iyophilized sporescarne from CSA the degradation began later (36hours), which explains probably the relative sporedamage suffered during the Iyophilizationrocess. In the case in which the spores areharvested from PDA medium it is observed thatcaffeine degradation begins after 60 hours. Asway of comparison we can mention that Hakil(1998) obtained caffeine degradation greater than90 % after 80 hour of incubation using CSAmethod.In CSA liquid medium (Table I) caffeinedegradation ability was induced due to thepresence of caffeine as unique nitrogen source.These studies suggest that enzymatic activityresponsible for caffeine degradation is inducible,due to the fact that degradation was increasedconsiderably after repeated cultivation. Therefore,once caffeine degradation ability is expressed isimportant to conserve the strain on CSA rnedium,as to have a shorter reactivation period. There areno studies about the influence of the conservationmethod on the caffeine degradation ability.Penicillium sp. V33A25 strain was able todegrade 92 % of total caffeine present at thebeginning of the culture, after 72 hours ofincubation. Therefore this strain will now used forthe forthcoming degradation studies of caffeinepresent in ensi led coffee pulp.
ACKNOWLEDGEMENTS
Present work was , supported in part by theNational Council ofScience and Technology(CONACYT. México), the Institut de Recherchepour le Développement (IRD-France) in theUniversidad Autónoma Metropolitana (Mexico)
and the BIOPULCA Project contract IC18*CT970185.
REFERENCES
Aquiáhuatl, R. M. (1992) Destoxificación de lapulpa de café: morfología, fisiología y bioquímicade hongos filamentosos que degradan cafeína.Tesis de Maestria. Facultad de Ciencias, UNAM.Barreiro Perera M. (1999) Coffee supply anddemand balance (Más alia de nuestro campo).Claridades Agropecuarias. Special edition, pp 2136Denis S. (1996) Degradation de la cafeine parAspergillus sp. et Penicillium sp. Etudephysiologique et biochimique. PhD thesis,Université de Montpellier 11, France, pp 209.Górnez-Brernes, R.A., Bendafía, G, González,J.M., Braham, J.E., Bressani, R. (1985) Relaciónentre los niveles de inclusión de pulpa de café ycontenido proteínico en raciones para animalesmonogástricos. Arch. Latinoam. Nutr. 35:422437.Hakil, M., Denis, S., Viniegra-González, G. andAugur, C. (1998) Degradation and productanalysis of caffeine and related dimethylxanthinesby fi lamentous fungi. Enzyme and MicrobialTechnology.22:355-359.Pefialoza, W., Molina, M.R., Gómez, R. &Bressani, R. (1985) Solid State Fermentation: Analternative to improve the nutritive value of coffeepulp. Appl. Environ. Microbiol. 49:388-393.Perrau-Gaime, I. (1995) Culture mixtes en milieusolide de bactéries lactiques et de champignonsfi lamenteux pour la conservation et ladécaféination de la pulpe de café. PhD thesis,Université de Montpellier 11, Montpellier, France,pp 209.

Porres, c, Alvarez, D., Ca1zada, 1. (1993)Caffeine reduction in coffee pulp through silage.Biotech. Adv. 11:519-523.Roussos, S., Aquiáhuatl, M. A., Trejo-Hernández,M. R., Perraud-Gaime, 1., Favela, E.,Ramakrishna, M., Raimbault, M., Viniegra-
376
González, G. (1995) BiotechnologicaJmanagement of coffee pulp: Isolation, screening,characterization, selection of caffeine-degradingfungi and natural microfiora present in coffee pulpand husk. Appl. Microbiol. Biotechnol. 42:756762.

377
PRODUCTlON OF EDlBLE MUSHROOM Lentinus edodes ON THE COFFEE SPENT GROUND
Fan Leifa, Ashok Pandey,Carlos Ricardo Soccol & R. Mohan
Laboratorio de Processos Biotecnologicos, Departamento de Engenharia Quimica, Universidade Federal doParana, CEP 81531-970, Curitiba-PR, Brazi1
INTRODUCTlON
Coffee spent ground is a residue produced involuminous quantity ( about I millions tons yearly )during the processing of instant products and familyconsume (Tango, 1971; Soccol, 1995; ICO, 1998).Brazit is a country producing the largest quantity ofcoffee spent ground in the world. This material wasoften composted as a soil conditioner, but it is alsogiven to the waste depots (Thielke, 1989), resultingin lost of organic material or causing environmentalpollution. Lentinus edodes is one of the mostimportant edible mushrooms cultivated world-wide(Yang, 1985; Fan, 1990). Jt has excellentorganoleptical properties of flavour and aroma, andpossesses good nutritional and therapeutically valuesdue to the presence of alI essentiaI amino acidsrequired for humans. Jt also shows antitumoralactivity, helps in reducing the cholesterol andstrengthening the immunological system (Zhang1989). L. edOdes has been traditionally cultivated inthe trunks of some hard trees (Yang, 1985). But a 10tof reports have appeared on its cultivation on someagricultural residues in the world, especially in themainland China (Yang, 1985; Zhang 1989).The extracted coffee grounds. moistened withsolution of 0,5% yeast extract at a ratio of 1:2(W/V). were used for culture of 1.edodes and thefruiting bodies were obtained (Thielke. 1989). Butthis technique is more expensive and not practicalfor production due to the addition of yeast extract.This work was to explore some techniques morepractical for its cultivation in the Brazilianconditions or for the other developing countries andcheck possible biological efficiency. Water contentand spawning rate are the parameters mostimportant in the cultivation of edible mushroorn,which affects directly the time of colonisation ofmycelia ( vegetative phase ) and initiation offructification. This work objects to determine thosefactors for better appl ication.
MATERIALS AND METHODS
Micro-organism. Ledodes LPB 02, routinelymaintained at 4 DC on PDA, was selected for thisstudy because of its good growth characteristics onthe extract of coffee husk.
Spawn preparation, The sawdust of Eucalyptussp ( 80%) and rice bran ( 20% ) were used for thespawn preparation. The mixture was adjusted atthe moisture of 60% (Yang, 1985 ) and then filIedin the flask. After autoclave and inoculation, theflask were incubated in the dark at 24 DC. Thespawn in the flask was ready for inoculation toobjective substrate after 20 days growth when themixture has turned white total1y.
Substrate preparation. The spent ground carnefrom the solvent coffee factory after sun-dry. Theeffect of water content in the substrate and rate ofinoculation ware conducted in the experiments.The concentration of water is at 45, 50%, 55%,60%,65%,70% and 75% respectively. The rate ofinoculation is at 2,5, 5%, 10%, 15%, 20% and25% respectively. The substrate was moistenedand 4 hours later, was filIed in autoclavableplastic bag (Cai, 1992). The bags were autoclavedat 121 DC for 1,5 hours. When inoculation, thespawn was separated into smalIest particles. Afterinoculation, good mixture between spawn andsubstrate was made so that mycelial growingcould be evaluated. After inoculation, the bagswere incubated in the dark at 24 DC. mycelialdevelopment in the bag was observed each dayafter 5 days.
Fructification. The 100 g of dry spent groundeach bag was used for the fructification. Thehumidity and spawn rate were adopted accordingto the results gained above. At the 30 days, thebags were transferred to a lighted environmentalchamber, cotton plugs Ioosened to allow betteraeration. After total transformation fromwhiteness to brown colour, the plastic wasremoved for fructification.
RESULTS
EFFECT OF WATER CONTENT ON THEMYCELIAL GROWTH OF L. edodes LPB 02IN THE COFFEE SPENT GROUND
The Figure 1 shows that the humidity is veryimportant for the mycelial growth of Lentinus onthe coffee spent ground. It is observed that thehumidity between 60-65% is better than that outof that range. It needs 12-14 days for the
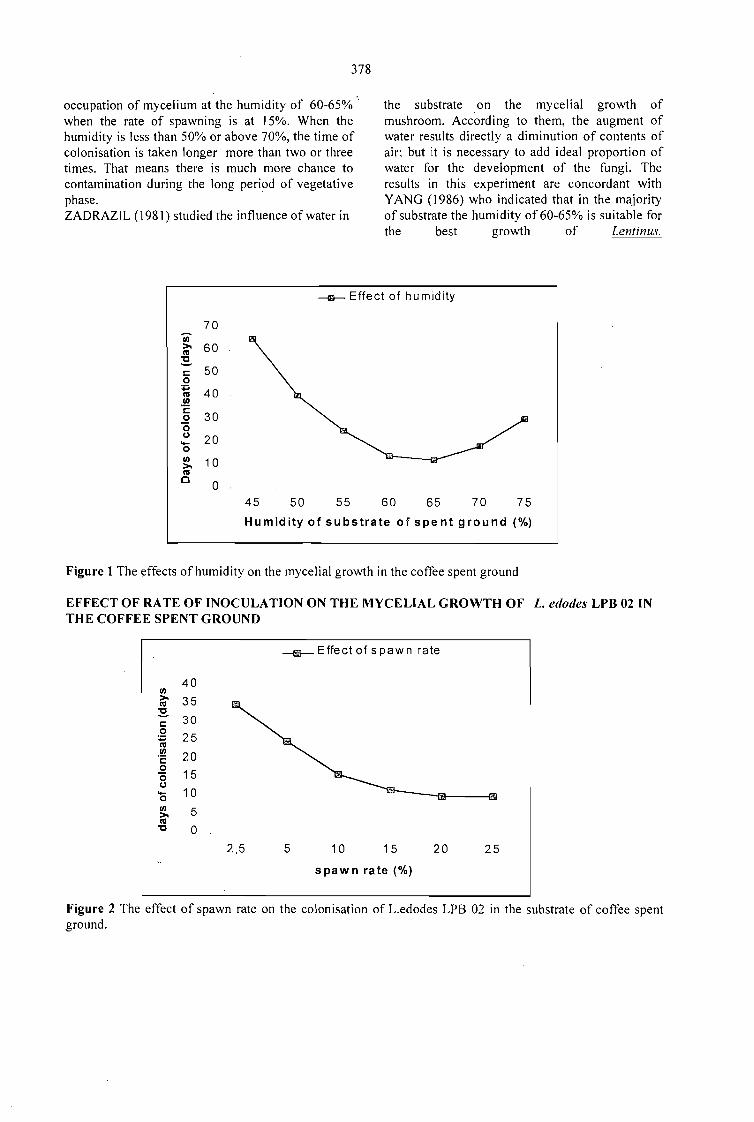
378
occupation of mycelium at the humidity of 60-65% -.when the rate of spawning is at 15%. When thehumidity is less than 50% or above 70%, the time ofcolonisation is taken longer more than two or threetimes. That means there is much more chance tocontamination during the long period of vegetativephase.ZADRAZ1L (1981) studied the intluence ofwater in
the substrate on the mycelial growth ofmushroom. According to them, the augment ofwater results directiy a diminution of contents ofair; but it is necessary to add ideal proportíon ofwater for the development of the fungi. Theresults in this experiment are concordant withYANO (1986) who indicated that in the majorityof substrate the humidity of 60-65% is suitable forthe best growth of Lentinus.
70éil>. 601lI
~l:: 50o
:o:;401lI
.!1ll::
30oÕo 20....oI/l 10>.1lIo O
---iD--- Effe ct o f hum id ity
45 50 55 60 65 70 75
l::o
:o:;1lI
.!1ll::oÕoÕI/l>.1lI'C
Humidity of substrate of spent ground (%)
Figure t The effects ofhumidity on the mycelial growth in the coffee spent ground
EFFECT OF RATE OF INOCULATION ON THE MYCELlAL GROWTH OF L. edodes LPB 02 INTHE COFFEE SPENT GROUND
~ Effect of s paw n rate
40I/l
~ 35~ 30
25
20
1510
5
O
2,5 5 10 15 20 25
spawn rate (%)
Figure 2 The effect of spawn rate on the colonisation of L.edodes LPB 02 in the substrate of coffee spentground.
The Figure 2 demonstrates the effects of thespawn rate on the mycelial growth in the substrateof coffee spent ground by L. edodes LPB 02. It isobserved that with the spawn rate of 15-25%, thetime of colonisation is much less, being 10-12days when humidity is at 60%. With the spawnrate of less than 5%, it needs more than 25 daysforthe occupation ofmycelia.RAJARAT.HNAM e BANO (1987) believe thatquantity of spawn below 10% promotecontamination of substrate. They recommendmore than 20% of spawn rate is better to promotea rapid colonisation of mycelia and morebiological efficiency. Considering econorrty andefficiency, the spawn rate of 15% isrecommended and adopted for the later work onthe substrate of coffee spent ground.
379
FRUCTIFICATION AND BIOLOGICALEFFICIENCY OF Ledodes LPB 02 IN THESUBSTRATE OF COFFEE SPE1~TGROUND
The results above were adopted for theexperiment of fructification, which were at thehumidity of 65% in the substrate and at theexperiment of fructification, which were at thehumidity or" 65% in the substrate and at thespawn rate of 15 %.First primordia appeared 60days after inoculation. There was only one or twoprimordias at one time. The fruiting body lasts 7days till rnaturity. Two or third flushes wereharvested from one bag, but the size of fruitingbody was generally reduced. Biological efficiencyreaches 88,64% onan average.
Figure 3 and 4 some photos of fructification of L.edodes LPB 02 on the coffee spend ground.
Lentinue edodes LPB 02coffee snent ground
DISCUSSION
The studies above showed that coffee ground is agood substrate for the cultivation of thisfungus.This material is plentiful in Brazil andavailable ali year round. The techniques for thecultivation is verysimple, .inexpensive, easi Iyextended to the other region.According to observation of growth of mycelia ofthis fungus, the spent ground also can be used asthe material of the spawn ror inoculation. Toachieve more production, maybe additional wheatbran would have a more positive effect. Forcommercial purposeit must be more effective toreplace the laboratory inoculation by a specialspawning wayor develop a mechanic way. Alsomore large bag should be used .
Lerurrrus edodes LPBcoffee spent ground
REFERENCES
Cai, W. M. and Fang, lL. and Fan~ L.F 1992 Thedisease of soft rotting in Flammulina VelutipesEdible mushroom, Shanghai, China:Chang, S.T. ]9S9 Edible mushroorn and theircultivation CRC Press, Inc. , Florida.Fan, L.F. and Ding, C.K 1990 '.. Handbook ofmushroom cultivation Jiangxi science andtechnology publishing house, Jiangxi, China.lnternational Coffee Organization 1998 Totalproduction of exporting ' membershttp://www.ico.org/proddoc.htmSOCCOL, C.R. 1995 Aplicações da fermentaçãono estado sólido na valorização deresiduosagroindustriais. França-Flash Agricultura..SOCCOL,C.R 1996 . Aplicaciones
. AgroindustriaJes de los Processos deFermentciones em Medio's Sol idos.Biotechnologia y Bioingeneria. Sociedade

Mexicana de Biotechnologia y Bioengeneria,Mexico.Tango. 1. S. 1971 Utilização industrial do café edos seus subprodutos Boletim do Instituto deTecnologia de Alimentos-ITAL São Paulo,Brasil.Thiclke, C. 1989 Cultivation of edible fungi oncoffee grounds Mushroom Science XII (parte 11)Germany.
380
Yang, X. M. 1986 Cultivation of ediblemushroom in China Agriculture printing house,Beijing, China.Zadrazil, F. 1980 Conversion of different plantwaste into feed by basidiomycetes EuropeanJournal of Applied Microbiology andBiotechnology, Berlin.

381
SOLID STATE FERMENTATlON AND PRODUCTlON OF P/etlrottls ostreattls ON THE COFFEERESlDlIES
Fan Leifa, Ashok Pandey, Carlos Ricardo Soccol and Radjiskumar Mohan
Laboratorio de Processos Biotecnologicos, Departamento de Engenharia Quimica, Universidade Federal doParana, CEP 81531-970, Curitiba-PR, Brazil
INTRODlICTION
Coffee is one of the most important beverages ofthe world, produced about I millions tons yearlyin more than 50 countries (lCO, 1998). Atdifferent stages from harvesting to the processingand consurnption, several residues, viz. coffeehusk, leaf and spent ground are generated about200 millions tons yearly (Tango, 1971; Soccol,1995). Brazil is a country producing the largestquantity of coffee sol id residues in the world(lCO, 1998). Due to the presence of compoundslike caffeine, tannins and polyphenols, theseorganic solid residues show toxic nature and thushave not been util ised beneficiaily. This has alsoled the problem of environmental contamination(Adams and Dougan.1981; Bressani, 1979;Soccol, 1996).The edible mushroom Pleurotus sp. , consideredas a good alternative for proteic qualityproduction foods in tropical countries (Bisaria,1983; Yang, 1986; Chang, (989), generallyshows good ability of producing fruiting body andsimultaneously reducing or degrading the toxicsubstances present in the substrate (Yang, 1986;Fan, 1990). The production of Pleurotus on thecoffee pulp has been studied since 1984 (Martineze Quirarte, 1984; Lozano, 1990 e Bermudez,1994), but by that way it produced voluminouswaste water during its washing-method ofpasteurization of the mushroom production andthe humid-processing of the coffee industry,causing environmental pollution. In Brazil. thecoffee husk comes from the dry method. fui! ofthe nutrients, also fuI! with caffeine. tannins,polyphenols, which is very different in thecompositions from that of other countries. Thiswork has objective to make good use of thatresidue, spontaneously resolve potentialenvironmental problem.Coffee leaf generally is not considered as aresidue. but its volume during and after theharvesting difficult the manipulation of the crop,also fac iIitates epidemic of pathogens and pests.The coffee spent ground is another importantresidue, concentrated in the instant coffee productfactory or scattered in the family during the
consume. The spent ground, moistened with asolution of 0,5% yeast extract at a ratio of 1:2(w/v), was used for experiment of Pleurotusostreatus (Thielke, 1989). This work aims toproduce this fungus for human consumption by aneconomic way based on thecoffee residues andexplore the possibilities ofration for animals.
MATERIAL AND METHODS
Micro-organtsm. 10 strains of Pleurotus sp,routinely maintained at 4 -c on PDA, was usedfor selection of the best strain on the extract ofcoffee husk. The procedure was adoptedaccord ing to Soccol (1994).
Spawn preparatíon. The sawdust of EucalyptusspJ 80%) and rice bran ( 20% ) were used for thespawn preparatíon. Th mixture was adjusted at themoisture of60% (Yang, 1986) and then filled inthe flask. After autoclave and inoculation, theflask were incubated in the dark at 24°C. Thespawn in the flask was ready for inoculation toobjective substrate after 20 days growth when themixture has turned white totally.
Substrate preparation. The coffee husk comesfrom the factory by dry method and spent groundcarne from the instant coffee-product factory aftersun-dry. The leaf comes from some friendscollected after the harvesting. The leaf wasgrinded after dry. These substrates werernoistened with water, generally undertaken 4 OI' 5hours before filling the autoclavable plastic bagoThe bags were autoclaved at 121°C for 1.5 hours.When inoculation. the spawn was separated intosmallest particles. After inoculation, good mixturebetween spawn and substrate was made so thatmycel ial development could be rapid and uniform.After inoculation, the bags were incubated in thedark at 24°C. rnycelial development in the bagwas observed and noted each day.
Fructification. After 20 days, the bags weretransferred to a lighted environmental charnber,the plastic was rernoved to allow stimulation ofair, hurnidity and light.

RESULTS
Selection Of Strains Of Pleurotus Sp On TheExtract OfCotTee Husk In The Petri-PlateThe Figure 1 shows the colony characteristics ofthe different strains of pleurotus. sp in thisexperiment, lt was observed that the strain P.ostreatus LPB 09 is the beststrain with morewhite and density of mycelia in this medium, theradial mycelial velocity being 9,68 mrn/day and
'biomass 43,40 mg/plate in 9daysat24 "C. So the. strain P.ostreatus LPB 09 was selected out for the
further studies,
Figure 1 The colony characteristics of Pleurotuson the extract ofcoffee husk
SOLID STATE FERMENTATlON ANDFRUCTIFICATlON OF P. ostreatus LPB 09ON THE COFFEE HUSK.
When coffee husk was used as the substrate, theideal humidity for mycelial growth was workedout at the 55-65%; the best spawn rate at 10-15%was more efficient whi le it took 7 days for fulloccupation of mycelia; first fructification occurredafter 20 days of inoculation; the biologicalefficiency reached at the 96,5%.
Figure 2 Fructification of P. ostreatus LPB 09 onthe coffee husk
SOLID STATE FERMENTATION AND,FRUCTIFICATlON OF P. ostreatus LPB 09ON THE COFFEE LEAF
When coffee leaf was used as the substrate, theideal humidity for mycelial growth was workedout at the 60-70%; the best spawn rate at 10% wasmore efficient and economic while it took 5 days
382
for full occupation of mycelia; but no fruit bodywas obtained in the unique leaf substrate.
SOLID STATE FERMENTATlON .ANDFRUCTlFICATlON OF P. ostreatus LPB 09ON THE COFFEE SPENT GROUND .
Whencoffee spent ground was used as thesubstrate, the ideal humidity for mycelial growthwas worked out at the 50-55%; the best spawnrateat 10% was. moreefficient and economicwhile it took 6days for full occupation ofmycelia; first fructification occurred after 23 daysofinoculation; the biological efficiency reached atthe 90,35%.
P.asr.r~ar.u.s: LPB 0.9 on coffee spent: ground
Figure 3 Fructifícation of P.ostreatus LPB 09 onthe coffee spent ground
DISCUSSION
The studies above showed that coffee husk andspent ground are good substrates for thecultivation of this fungus. This material isplentifulin Braziland available ali year round.The techniques for the cultivation is very simple,inexpensive, easily extended to the other region.As to coffee leaf used in this experiment, no fruitbody was harvested maybe becauseof the loosedtexture or much more caffeine and tannins, Aftercoffee leaf mixtured with coffee husk or spentground at less than 50% de proportion, the fruitbody was obtained and the biologicalefficiencyreached at about 60%.Through the analysis of the fruit body andremains after the fructification, we found that thisfungus has not degraded the caffeine but itabsorbed and accumulated the caffeine into thefruit body. The content of caffeine inthe remainsof coffee husk after the growth of mycelia andfructification was reduced up .to 60,69%, tanninsto 79,17%., The protein content in it wasaugmented while the cellulose was reduced. Alsothe texture of the rerTlains was modified to moresoftly. It is more suitable as ration for animais.According toobservation of growth of mycelia ofthis fungus, the spent ground also can be used asthe material of the spawn for inoculation. Toachieve more production, maybe additional wheat

bran would have a more posttive effect. Forcommercial purpose it must be more effective toreplace the laboratory inoculation by a specialspawning way or develop a mechanic way. Alsomore large bag should be used.
REFERENCES
Adams, M.R. and Dougan, J. 1981 Biologicalmanagement of coffee processing waste Trop.Sci.Bressani, R. 1979 Antiphysiological factors incoffee pulp Coffee pulp: composition, technologyand utilization IGRC, Ottawa.Bermudez, R.C. and Traba, 1. A. 1994 Produccionde Pleurotus sp cfr. Florida sobre residuales de laagroindustria cafetalera en Cuba MicologiaNeotropical Aplicada, Mexico.Bisaria, R. and Madan, M 1983 Mushrooms:potencial protein source from cellulosic residuesEnzyme and microbial technolgy, USA.Chang, S.T. 1989 Edible mushroom and theircultivation CRC Press, Inc. , Florida.International Coffee Organization 1998 Totalproduction of exporting membershttp://www.ico.org/proddoc.htmFan, L.F. and Ding, C.K 1990 Handbook ofmushroom cultivation Jiangxi science andtechnology publishing house, Jiangxi, China.
383
Lozano, 1.C. 1990 Produccion comercial deichampinon (Pleurotus ostreatus) en pulpa decoffee Fitopatologia Colombiana, Colombia.Martinez, D. and Quirarte, M. 1984 Prospects ofgrowing edible mushroom on agro-industrialresidues in Mexico Boletin de la sociedadMexicana de micologia, Mexico.Soccol, C.R. 1994 Contribuição ao estudo dafermentação no estado sólido em relação com aprodução de ácido fumárico. Biotransformação deresíduo sólido de mandioca por Rhyzopus eBasidiomacromicetos do gênero Pleurotus Tesede Livre-Docência. UFPR,Curitiba,1994.228p.Soccol, C.R. 1995 Aplicações da fermentação noestado sólido na valorização deresíduosagroindustriais. França-Flash Agricultura.Soccol, C.R 1996 Aplicaciones Agroindustrialesde los Processos de Fermentciones em MediosSolidos. Biotechnologia y Bioingeneria.Sociedade Mexicana de Biotechnologia yBioengeneria, Mexico.Thiclke, C. 1989 Cultivation of edible fungi oncoffee grounds Mushroom Science XlI (parte lI)Germany.Yang, X. M. 1986 Cultivation of ediblemushroom in China Agriculture printing house,Beijing, China.

j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j

385
PRODUÇÃO DE AROMA FRUTAL POR Ceratocystis fimbriata EM RESíDUO SÓLIDO DEAGROINDÚSTRIAS DE CAFÉ
M. Soares1,2 ; P. Christen3; A.Pandeyl; M. RaimbauJt4; C.R. Soccol 1
I 2
Laboratorio de Processos Biotecnológicos, UFPR, Brasil; CEFET-PR UNED PG, Brasil; 3 Laboratoire deMicrobiologie, IRD, Université de Provence, Marseil1e, França; 4 Laboratoire de Biotechnologie, IRD,Moritpell ier, França. Corresponding author.
INTRODUÇÃO
Apesar da maioria dos aromatizantes serematualmente obtidos por síntese química(84%),pesquisas mostram a crescente preferência dosconsumidores por alimentos naturais ou quecontenham aditivos naturais(4). Plantas continuamsendo importantes fontes de óleos essenciais earomas. mas há grande dependência em fatores dedifícil controle, como clima e doenças. Umaalternativa à síntese de aromas naturais baseia-senos processos microbiológicos, comofermentações (biosíntese) e bioconversões. Ocusto de produção de tais aromas pode serquestionado; contudo. o preço de mercado pagopor aromas naturais é compensador. Por exemplo,a 4-decalactona sintética (aroma de impacto empêssegos) custa 150 dólares/ Kg, enquanto seusimilar natural custa 6000 dólares/ Kg(4). Fungosdo gênero Ceratocystis sintetizam diferentesaromas frutais/florais (pêssego, abacaxi, banana,CÍtricos e rosa), dependendo da cepa e dascondições de cultivo, sendo que C. jimbriata éinteressante devido ao seu rápido crescimento.habilidade na produção de esporos e à variedadede aromas sintetizados(5). Neste trabalho.procedeu-se a fermentação no estado sólido dacasca de café com o intuito de avaliar a produçãode aromas por C. jimbriata, comparando osresultados com outros mencionados paradiferentes resíduos agroindustriais. A influênciada adição de glicose e o uso de substânciasprecursoras de aromas foram estudados. Oscompostos biosintetizados presentes no headspacedos cultivos foram caracterizados e quantificados.
MATERIAL E MÉTODOS
MicroorganismoO microorganismo estudado foi C. jimbriata CBS374-83. A manutenção da cepa e a coleta dosesporos transcorreram conforme descrito emliteratura (I).
Preparo do substrato sólidoDuzentos gramas de casca de café, previamenteseca (8% de umidade) e reduzida em partículas de
diâmetro entre OA-0,8 mm, foram misturados aum litro de água destilada e tratados em vaporfluente por 45 mino Após simples filtração asfrações líquida e sólida foram separadas, sendoque estudos preliminares demonstraram que estamostrou-se mais apropriada à prod ução de aromasdo que a casca e do que· a fração líquida.provavelmente devido à eliminação de sais e/ououtras substâncias.
Procedimentos de fermentaçãoOs experimentos foram conduzidos emErlenmeyers de 250mL, contendo 15g de matériaseca (MS) e cobertos com 8 camadas de gaze. Emtodos os experimentos as condições iniciaisforam: temperatura 30'C. pH 6, inóculo Ix10- 7
esporos s' MS e 70% de água (máxima absorçãodo substrato)( I ;3). A influência da adição deglicose (20%, 35% e 46%) e da adição depossíveis precursores (Ieucina 10 mMol (2), óleode soja 10%( I) e solução salina(2)) foramestudadas para otimização da produção de voláteistotais (VT).
Procedimentos analíticosCada experimento foi realizado em triplicata. Oacompanhamento da evolução da produção dosvoláteis foi realizada através de avaliaçõessensonais paralelamente à análise porCromatografia de Gases (CG). Os compostosforam identificados no headspace dos cultivoscomparando seu tempo de retenção com padrões.Voláteis totais e álcoois foram expressos emequivalente de etanol urnol por Litro de headspacepor grama de matéria seca (umol/L gMS) eésteres, da mesma forma, como equivalente de etilacetato, ambos a partir de curvas padrões (3).
Análise dos dadosOs dados obtidos por cromatografia foramintegrados com o objetivo de calcular os voláteistotais acumulados durante a fermentação. Omodelo logístico de Gompertz foi utilizado paraproceder esta integração e calcular o tempo demáxima produção e a velocidade de produção dosvoláteis(3).
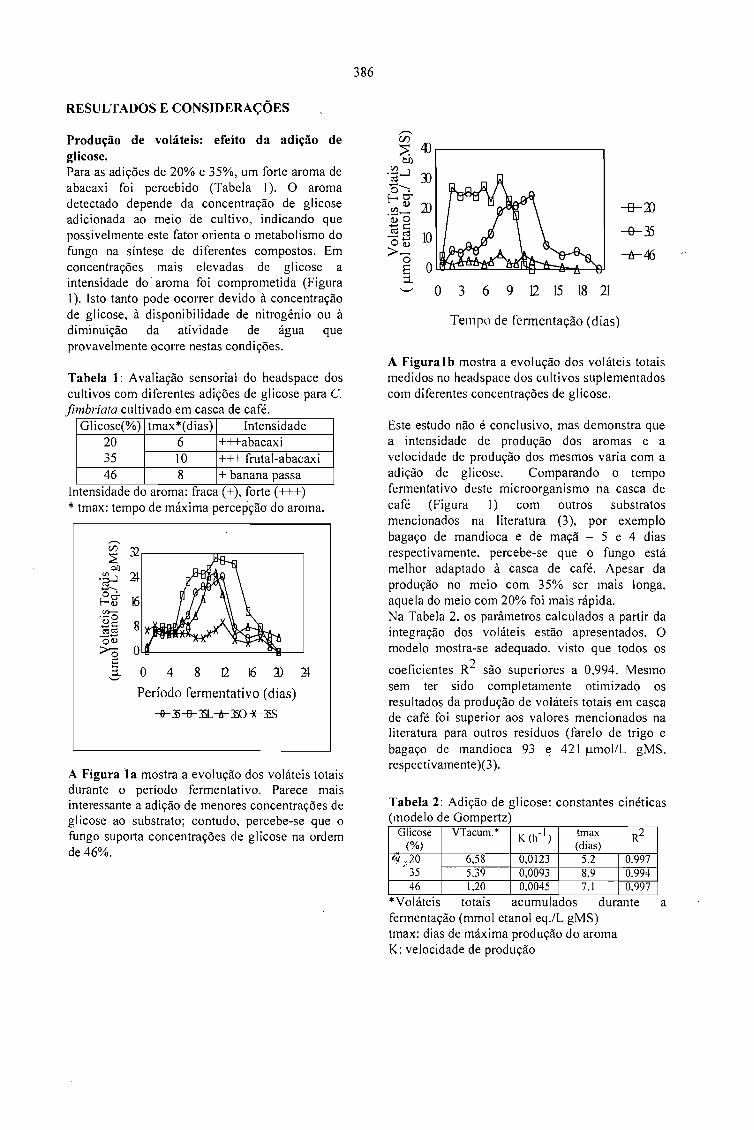
386
RESULTADOS E CONSIDERAÇÕES
3 6 9 12 15 18 21
Tempo de fermentação (dias)
Tabela 2: Adição de glicose: constantes cinéticas(modelo de Gompertz)
-B-::D
--&-35
-fr-46
Glicose VTacum.* K (h-I) trnax R2(%) (dias)
('1.20 6.58 0,0123 5,2 0.99735 5.39 0,0093 8,9 0.99446 1.20 0,0045 7.1 0.997
Este estudo não é conclusivo, mas demonstra quea intensidade de produção dos aromas e avelocidade de produção dos mesmos varia com aadição de glicose. Comparando o tempofermentativo deste microorganismo na casca decafé (Figura 1) com outros substratosmencionados na literatura (3), por exemplobagaço de mandioca e de maçã - 5 e 4 diasrespectivamente, percebe-se que o fungo estámelhor adaptado à casca de café. Apesar daprodução no meio com 35% ser mais longa,aquela do meio com 20% foi mais rápida.Na Tabela 2, os parâmetros calculados a partir daintegração dos voláteis estão apresentados. Omodelo mostra-se adequado, visto que todos os
coeficientes R2 são superiores a 0,994. Mesmosem ter sido completamente otimizado osresultados da produção de voláteis totais em cascade café foi superior aos valores menci-onados naliteratura para outros resíduos (farelo de trigo ebagaço de mandioca 93 e 421 umol/L gMS.respectivamente)(3).
A Figura 1b mostra a evolução dos voláteis totaismedidos no headspace dos cultivos suplementadoscom diferentes concentrações de glicose.
Glicose(%) tmax*(dias) Intensidade20 6 +++abacaxi35 10 +++ frutal-abacaxi46 8 + banana passa
A Figura h mostra a evolução dos voláteis totaisdurante o período fennentativo. Parece maisinteressante a adição de menores concentrações degl icose ao substrato; contudo, percebe-se que ofungo suporta concentrações de glicose na ordemde 46%.
Intensidade do aroma: fraca (+), forte (+++)* tmax: tempo de máxima percepção do aroma.
Período fermentati vo (d ias)
+ 35-B-]iL-Ir-350+ li';
Produção de voláteis: efeito da adição deglicose.Para as adições de 20% e 35%, um forte aroma deabacaxi foi percebido (Tabela I). O aromadetectado depende da concentração de glicoseadicionada ao meio de cultivo, indicando quepossivelmente este fator orienta o metabolismo dofungo na sintese de diferentes compostos. Emconcentrações mais elevadas de glicose aintensidade do aroma foi comprometida (Figura1). Isto tanto pode ocorrer devido à concentraçãode glicose, à disponibilidade de nitrogênio ou àdiminuição da atividade de água queprovavelmente ocorre nestas condições.
Tabela 1: Avaliação sensorial do headspace doscultivos com diferentes adições de glicose para C.jimbriata cultivado em casca de café.
*Volatels totars acumulados durante afermentação (mmol etanol eq./L gMS)tmax: dias de máxima produção do aromaK: velocidade de produção

387
Tabela 3: Avaliação sensorial do headspace doscultivos de C. fimbriata em casca de café (35%glicose)adicionado de determinados compostos.
salina ao resíduo afetou a produção de VT. Esteestudo não permiteconcluir se este efeito énegativo em relação ao crescimento oudiretamente à produção de voláteis.Figura 2: Evolução dos VT medidos no headspacedos cultivos com 35% de glicose (-, controle) esuplementado com leucina (L), solução salina(SS) e óleo de soja (O).A análise por Cromatografia de Gases permitiu aidentificação de diversos compostos. comoindicado na Tabela 5, que apresenta os dadosintegrados para cada composto presente noheadspace dos cultivos. A adição de leucinaaumenta a produção de ésteres derivados dometabolismo deste aminoácido. Todos oscompostos já foram mencionados previamente poroutros autores (1.2.3).
Substância tmax*(dias) IntensidadeSS 10 + bananaO 10 ++ bananaL 10 +++ banana..Intensidade do aroma: fraco (+). médio (++). forte
(+++) tmax: tempo de máxima percepção do aromaSS : solução salina(2): O: I0% de óleo de sojat I): L:IOmMol de leucina(2).
Produção de voláteis: efeito da adição deleucina, óleo de soja e solução salina.Este estudo foi feito com adição de glicose de35%. Os aromas detectados por olfatometria estãopresentes na Tabela3.
CONSIDERAÇÕES FINAIS
Tabela 5: Total individual dos voláteis medidosno headspace dos cultivos de C. jimbriata (dadosintegrados).
20: 20% glicose: 3): 35% glicosc (controle): 3).L:com lcucina:35.0: com óleo de soja.* expresso em equivalente de etanol umol / L gMS :** expresso em equivalente de etil acetato urnol / LgMS.
O resíduo de casca de café tratada com vaporfluente é um substrato adequado à produção dearomas por C. fimbriata. A adição de glicose énecessária à esta proposta e a concentraçãoadequada compreende-se entre 20% e 35%. Aadição de leucina aumentou em 58% a produçãode VT. Comparando os resultados com outros daliteratura percebe-se que a produção total foi duasvezes superior aos melhores resultados obtidoscom bagaço de mandioca. senso que este haviasido adicionado de solução de leucina 15 vezesmais concentrada (3). Contudo, os compostosidentificados foram os mesmos em ambostrabalhos.
Compostos 20 35 35.0 35.LAcetaldeído * 47 57 58 50Etanol * 329 428 607 565lsopropanol * 26 5 - 11Eti I acetato* * 2078 1833 1841 2811Etil isobutirato ** II 10 7 15lsobuti I acetato ** 19 18 15 35Etil butirato ** 9 3 1 21Propil acetato ** 4 - - -lsoamil acetato ** 2 II - 162- heptanona * - - - 32-octanona * 52 67 3 30Eti I 3 hexenoato ** - - - 3
- -
Tabela 4: Adição de leucina, solução salina e óleode soja: constante cinéticas segundo o modelo deGompertz.
Em todos os casos um agradável aroma de ·bananafoi notado. mais intenso quando o meio foiadicionado de leucina. Os parâmetros calculadospelo modelo de Gompertz (Tabela 4) indicam quea adição de óleo de soja ao meio não melhorou aprodução total de voláteis. que foi similar ao meiocontrole. indicando que o fungo não assimila estesubstrato para metabolismo primário. tampouco outiliza como precursor do metabolismo de metilcetonas como outros fungos (Penicilliumroquefortii, por exemplo). A adição de soluçãosal ina diminuiu drasticamente a produção dosvoláteis. mas leucina atuou inversamente (Figura2). Isto indica que o substrato já possui suficienteconcentração de sais e que o fungo degrada aleucina segundo a rota de Erlich a isoamil alcool,o qual é então esterificado em isoamil acetato. oqual é o maior componente de aromas de banana.
o, L e SS como para Tabela 3.*Voláteis Totais acumulados durante afermentação (urnol etanol eq.lL gMS)tmax: tempo de máxima produção do aromaK: velocidade de produção
Após o tratamento da casca com vapor fluenteocorreu lima diminuição na quantidade de sais naordem de 30% e. conforme observadopreviamente. este tratamento aumenta a produçãode VT. Da mesma forma. a adição de uma solução
Substân VTacum. KW) tmax R2Cla * (dias)
- 5.39 0.0093 8.9 0,994O 5.7 0,0085 9.5 0.996L 8.81 0.0098 9,4 0,997
SS 2.8 0.0067 12 0,996

REFERÊNCIAS BIBLIOGRÁFICAS
1-BRAMüRSKI,A. et ai. Biotechnology Letters,20 (4): 359-362,1998.2-CHR1STEN,P et aI. Biotechnology Letters;13 (7): 521-526, 1991.
388
3-CHRISTEN,P et aI. Mycological Research. 101(8):911-919,1997.4-JANSSENS,L et aI. Process Biochemistry.27:195-215.1992.5-SENEMAUD,C. Ph.D. Thesis, France. 1988.

389
PRODUÇÃO DE ÁClDO CÍTRICO POR Aspergil/us niger LPB 21 EM FERMENTAÇÃO NOESTADO SÓLlDO COM CASCA DE CAFÉ
Luciana P.S. Vanderberghe'i'' Ashok Pandey,' Jean-Michel Lebeaulr' and Carlos R. Soccol' '"
'Laboratório de Processos Biotecnológicos, Departamento de Engenharia Quimica, Universidade Federal doParaná, CEP 81531-970 Curitiba-PR, Brazil; 2Laboratoire de Procédés Biotechnologiques, Génie Chimique,Université de Technologie de Compiégne, 60205-Compiegne, Cedex, France
o Brasil é o maior produtor de café no mundo comuma produção anual (1997-1998) estimada em18.86 milhões de sacas. O processamento do cafégera grandes quantidades de resíduo sólido (cascade café ou polpa de café a depender do modo deprocessamento). Atualmente, não existe nenhumaaplicação para a casca de café consistindo em umgrande problema ambiental. Portanto, o uso dessesubstrato em processos para a produção deprodutos de interesse comercial, seria de extremointeresse econômico.Apesar da fermentação no estado sólido (FES)
oferecer inúmeras vantagens em relação àfermentação submersa (FS), a produção do ácidocítrico se dá quase que totalmente por FS,principalmente com meios a base de amido esacarose, utilizando fungos filamentosos (Pandey,1992, Soccol, 1998, Vandenberghe et aI., 1998).O presente trabalho tem como principal objetivo aprodução do ácido cítrico em FES pelo cultivo deuma linhagem de Aspergillus niger em casca decafé. O resíduo foi impregnado com hidrolisado debagaço de mandioca (BM), um resíduo rico emamido (cerca de 60-70%) e também abundante noEstado do Paraná. A hidrólise enzimática do BM,transformação do amido em açúcares (glicose emaltose) foi realizada. As etapas de otimizaçãodeste pré-tratamento foram apresentadas, assimcomo os procedimentos para a fermentação sólida.
MATERIAL E MÉTODOS
Microrganismos: Aspergillus niger LPB 21 foiselecionada para este estudo. A cepa foiconservada em meio Potato Agar Dextrose (PDA),mantida a 4°C e renovada mensalmente.Inóculo: Os esporos de A. niger foram produzidosem PDA (50 mL) distribuído em frascosErlenmeyer de 250 mL e incubados a 30°C durantesete dias. A suspensão de esporos foi preparadacom a adição de 25 ml de água destilada comTween-80 (0.1 %) e conservada a 4° C por nomáximo duas semanas.Hidrólise enzimática: O bagaço de mandioca emuma concentração de 120 g/L de água potável,contendo uma quantidade necessária de cálciopara garantir a boa estabilidade das enzimas, foigelatinizado a 100-1 10°C durante 15 minutos. O
substrato foi então incubado com Termamyl 120 L(u-amilase) e, em seguida, com AMO 200 L(amiloglicosidase) de acordo com o planoexperimental citado abaixo. Após a hidrólise asenzimas foram inativadas a 100-1 04°C durante 10minutos para evitar a permanência de resíduos nohidrolisado. Finalmente, as amostras foramfiltradas e analisadas.Plano fatorial 1: O início dos experimentos foibaseado em um plano fatorial do tipo 24
, compontos centrais, variando as concentrações dasduas enzimas, n-amilase e arniloglucosidase. Osfatores estudados e os níveis correspondentes sãocitados na Tabela I.
Tabela 1. Hidrólise enzimática do bagaço demandioca - Plano fatorial INível Tempo de Tempo de Concentraçã Concentraç
Hidrólise Hidrólise o Termamyl, ão AMG,Termamyl, AMG, IlLl100 g de IlLlIOO g deh h amido amido
-I I 12 100 231O 2 24 200 471
+1 3 36 300 711
Plano fatorial 2: O segundo plano experimentalfoi definido à fim de otimizar as condições dahidról ise enzimática definidas pelo planoexperimental precedente. Sendo assim, osresultados em açúcares foram observados e aanál ise estatística apontou os fatores maissignificativos. A Tabela 2 apresenta os níveis dosfatores selecionados para esta etapa do estudo. Osúnicos fatores significativos foram aquelesrelacionados à amiloglucosidase.Consequentemente. o tempo de hidrólise e aconcentração da enzima of a-amilase formamantidos constantes nos seus níveis baixos,concentração de 100 /lU] 00 g de amido e I horade incubação.
Tabela 2. Hidrólise enzimática do bagaço demandioca - Plano fatorial 2Níveis Tempo de hidrólise Concentração da
AMG AMG, IlLl100 g deh amido
-I 12 471O 18 591
+1 24 711
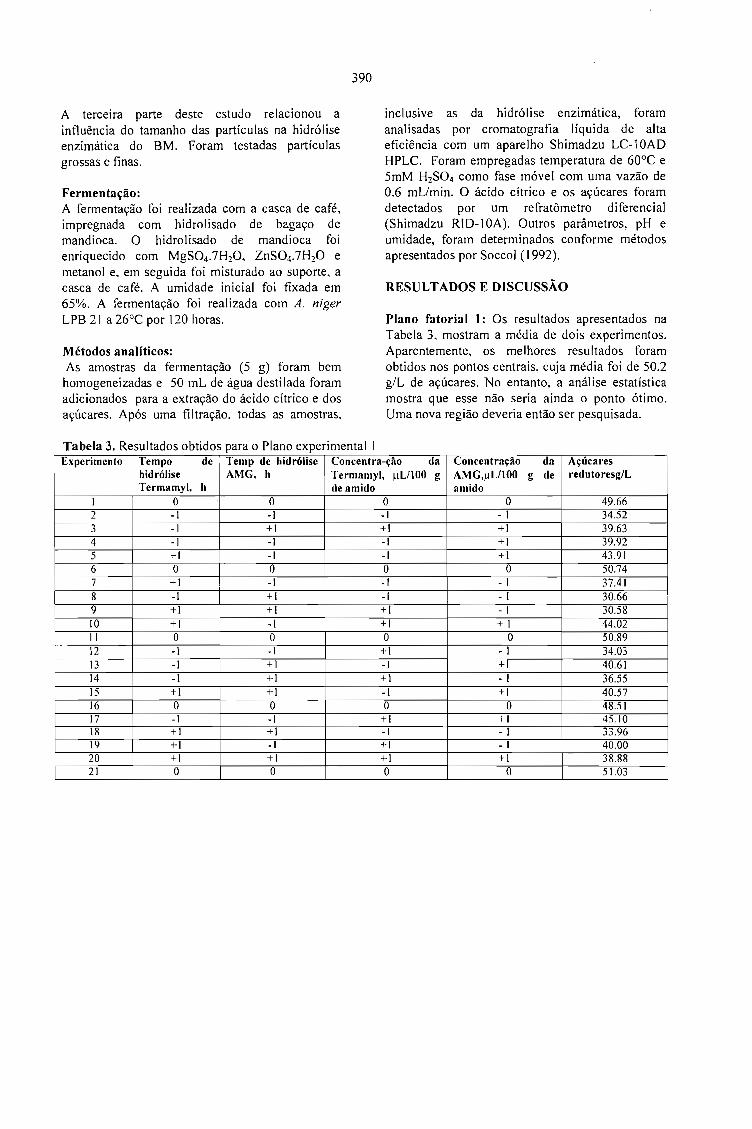
390
A terceira parte deste estudo relacionou ainfluência do tamanho das partículas na hidról iseenzimática do BM. Foram testadas partículasgrossas e finas.
Fermentação:A fermentação foi realizada com a casca de café,impregnada com hidrolisado de bagaço demandioca. ° hidrolisado de mandioca foienriquecido com MgS04.7H20, ZnS04.7H20 emetano! e. em seguida foi misturado ao suporte, acasca de café. A umidade inicial foi fixada em65%. A fermentação foi realizada com A. nigerLPB 21 a 26°C por 120 horas.
Métodos analíticos:As amostras da fermentação (5 g) foram bem
homogeneizadas e 50 mL de água desti lada foramadicionados para a extração do ácido cítrico e dosaçúcares. Após uma filtração, todas as amostras,
Tabela 3. Resultados obtidos para o Plano experimental 1
inclusive as da hidrólise enzimática, foramanal isadas por cromatografia líquida de altaeficiência com um aparelho Shimadzu LC-l0ADHPLC. Foram empregadas temperatura de 60°C e5mM H2S0 4 como fase móvel com uma vazão de0.6 mUmin. ° ácido cítrico e os açúcares foramdetectados por um refratômetro diferencial(Shimadzu R10-1 OA). Outros parâmetros, pH eumidade, foram determinados conforme métodosapresentados por Soccol (1992).
RESllLTADOS E DlSCllSSÃO
Plano fatorial 1: Os resultados apresentados naTabela 3, mostram a média de dois experimentos.Aparentemente, os melhores resultados foramobtidos nos pontos centrais, cuja média foi de 50,2g/L de açúcares. No entanto, a anál ise estatísticamostra que esse não seria ainda o ponto ótimo.Uma nova região deveria então ser pesquisada.
Experimento Tempo de I Temp de hidrólise Concentra-ção da Concentração da Açúcareshidrólise AMG, h Terrnamyl, ~LlIOO g AMG,~Ll100 g de redutoresg/LTermamyl, h de amido amido
I O O O O 49.662 -1 -I -I - 1 34.523 -1 +1 +\ +1 39.634 -1 -1 -\ +1 39.925 +1 -1 -I +1 43.916 O O O O 50.747 +1 -1 - I - I 37.4\8 -I +1 -I - I 30.669 +\ +1 +1 - I 30.5810 +1 -I +1 +1 44.0211 O O O O 50.8912 -1 ·1 +1 - I 34.0313 -I +1 -I +1 40.6114 -I +1 +1 - 1 36.5515 +1 +1 -I +1 40.5716 O O O O 48.5117 -1 -I +\ +1 45.\018 +1 +1 -I - I 33.9619 +1 -I +1 - 1 40.0020 +1 +1 +1 +1 38.8821 O O O O 51.03
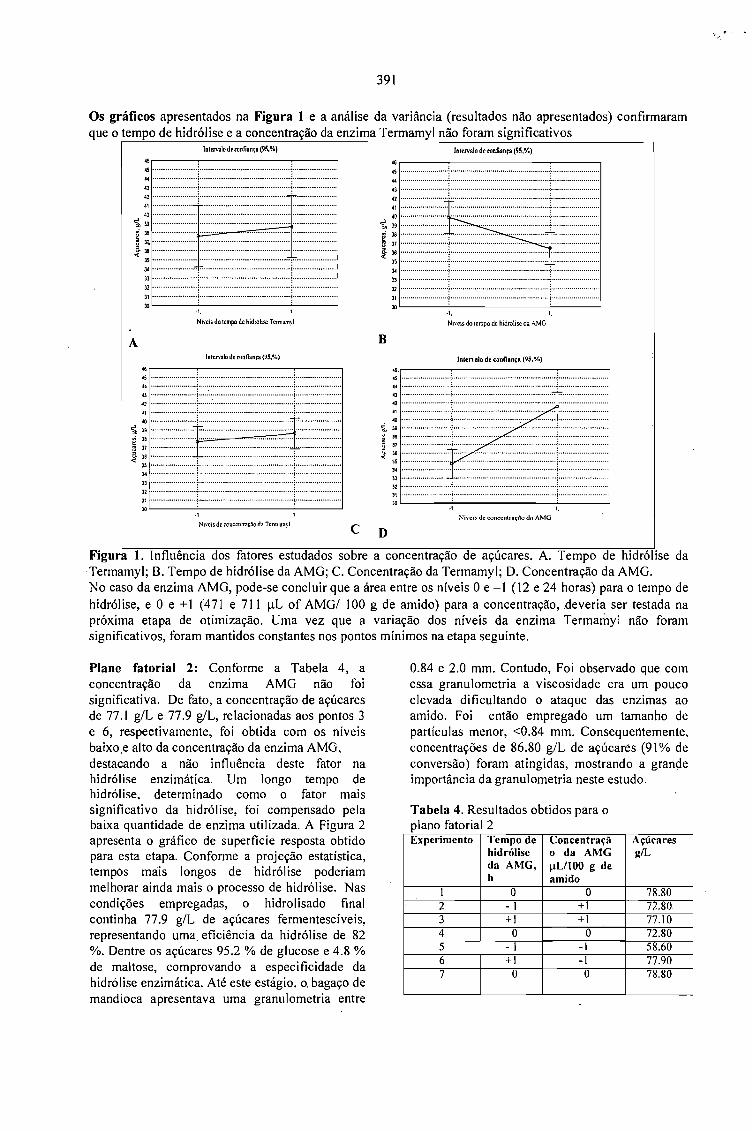
391
Os gráficos apresentados na Figura 1 e a análise da variância (resultados não apresentados) confirmaramque o tempo de hidrólise e a concentração da enzima Termamyl não foram significativos
<.
Inlm'ilodeconfiança (93.'/.)
~. .45 ..••••.•.••••.•••••••...; .••••••.••.•••.••.•.••••••...•.•.••••••••••.••.:•••••••••••••••••••.•••.
44 ••••••••••••••••••••••••! ! .43 ••.••••.•.•••••••••••••• j•••....•.••...•••......••..•..•.•.........•...... j...•...........••.....•.
42 ; ;... .
41 _: ..
4D···························..······..················· .~19 · • ..
~ '" .. ..3 3t .~ 38·············································,..······ .
3S _ : .
34 ·i··· ··· ··..··3] : ; .
32 ~....... ·..·.. ·.. · · i ·..··············..·31 ! j .lO
-1.
Nivcis dotempo de hidrchsc Tcrmamyl
AInten'alode ronliam:a(9.5,'/a)
"r---~------~-----'
:: ::::::::::::::::::::: :::~ ::::: :::~:::::: ::::::::::::::::::::::::::::::: :::~::: :::::::::::::::::::::43 j ·..·····i·····..·················42 •., ; : ······1..····..·..· · ···41 ~ ! .
i1~~~-~~:~-~;!~:35 : ,', •.:', .
34 ! · ·..······ .. ··········.. ,·········,··1·····..··· ..····· ..· .33 ••••••••••••••••,•••..•.: ,•••••••••••••.•••••••
32 ~..... • ,.. , .
31 ··••·· .. ················i..······..··..··········· 'f" , .lO :
".NI\"'Cis deconecntracãcda Tcnnamyl
Intervalo deconfiança (93,'/.)
~r------,---------,-------,
:: ::::::::::::.. ::::::::::~:::::::::::::::::::::::::::::::::::::::::::::::::j::::::::::::::::::::::::43 } ••·.. ··••.. i···············..·······42 ,.. , ;" ; , ..
41 " ! .4D , ; .
~ 39 ~ ..
~ '" : .
I: :::::::::::::::::::::::1::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::: ..J,.t ~ ~ .
3] ; ; .•..•.....••..••
32 ~ I··· ··· i······· .31 .. ••.. • ··•· .. ·.. ··i ··..·..·..·..· ·..· ·..· ·j· ..··· .
·1. 1.
NI\"els do tempode hidrolisc da I\MG
B
Intervalode conflança(95,%)
"r--------------,-------,~5 , , .
44 •.. ·····················i·..···········..················ j .43 j .<12 ~ .
<I, .............•..........~ ,.,............ . .<lO ) .
~ 39 ~ .
~ 36 ~.... • .
~ 37 , , j .
~ ~: :::::::::::::::::::::::: "':::::"'::::::::::::::::::::::::::::::::::::::1:::::::::::::::::::: .34 ····,··· .. ·.·····í···J3 ·.. •.. ·••.. ••••••···· •.. •·••·•· .. •·· .. ·····1..···..· ······ .32 •.••••••..••.•••.•...•..? ,.•.! .]1 ••••••••• j ••••.••.••.••.•.•••.•.•••••••.••••.•••.••..• ·····i········30: :
.1.
Ni"cIS de conccnlroçllo da AMO
C D
Figura 1. Influência dos fatores estudados sobre a concentração de açúcares. A. Tempo de hidrólise daTermamyl; B. Tempo de hidrólise da AMO; C. Concentração da Termamyl; D. Concentração da AMO.No caso da enzima AMO, pode-se concluir que a área entre os níveis Oe -I (12 e 24 horas) para o tempo dehidrólise, e O e +1 (471 e 711 IlL of AMOI 100 g de amido) para a concentração, deveria ser testada napróxima etapa de otimização. Uma vez que a variação dos níveis da enzima Terrnamyl não foramsignificativos, foram mantidos constantes nos pontos mínimos na etapa seguinte.
Plano fatorial 2: Conforme a Tabela 4, aconcentração da enzima AMG não foisignificativa. De fato, a concentração de açúcaresde 77.1 g/L e 77.9 g/L, relacionadas aos pontos 3e 6, respectivamente, foi obtida com os níveisbaixoe alto da concentração da enzima AMO.destacando a não influência deste fator nahidrólise enzimática. Um longo tempo dehidrólise. determinado como o fator maissignificativo da hidrólise, foi compensado pelabaixa quantidade de enzima utilizada. A Figura 2apresenta o gráfico de superficie resposta obtidopara esta etapa. Conforme a projeção estatística,tempos mais longos de hidrólise poderiammelhorar ainda mais o processo de hidró1ise. Nascondições empregadas, o hidrolisado finalcontinha 77.9 g/L de açúcares fermentescíveis,representando uma. eficiência da hidrólise de 82%. Dentre os açúcares 95.2 % de glucose e 4.8 %de maltose, comprovando a especificidade dahidrólise enzimática. Até este estágio. o.bagaço demandioca apresentava uma granulometria entre
0.84 e 2.0 mm. Contudo, Foi observado que comessa granulometria a viscosidade era um poucoelevada dificultando o ataque das enzimas aoamido. Foi então empregado um tamanho departículas menor, <0.84 mm. Consequentemente,concentrações de 86.80 g/L de açúcares (91% deconversão) foram atingidas, mostrando a grandeimportância da granulometria neste estudo.
Tabela 4. Resultados obtidos para oplano fatorial 2Experimento Tempo de Concentraçã Açúcares
hidrólise o da AMG glLda AMG, IlL/I00 g deh amido
1 O O 78.802 - 1 +1 72.803 +1 +1 77.104 O O 72.805 - 1 -I 58.606 +1 -1 77.907 O O 78.80
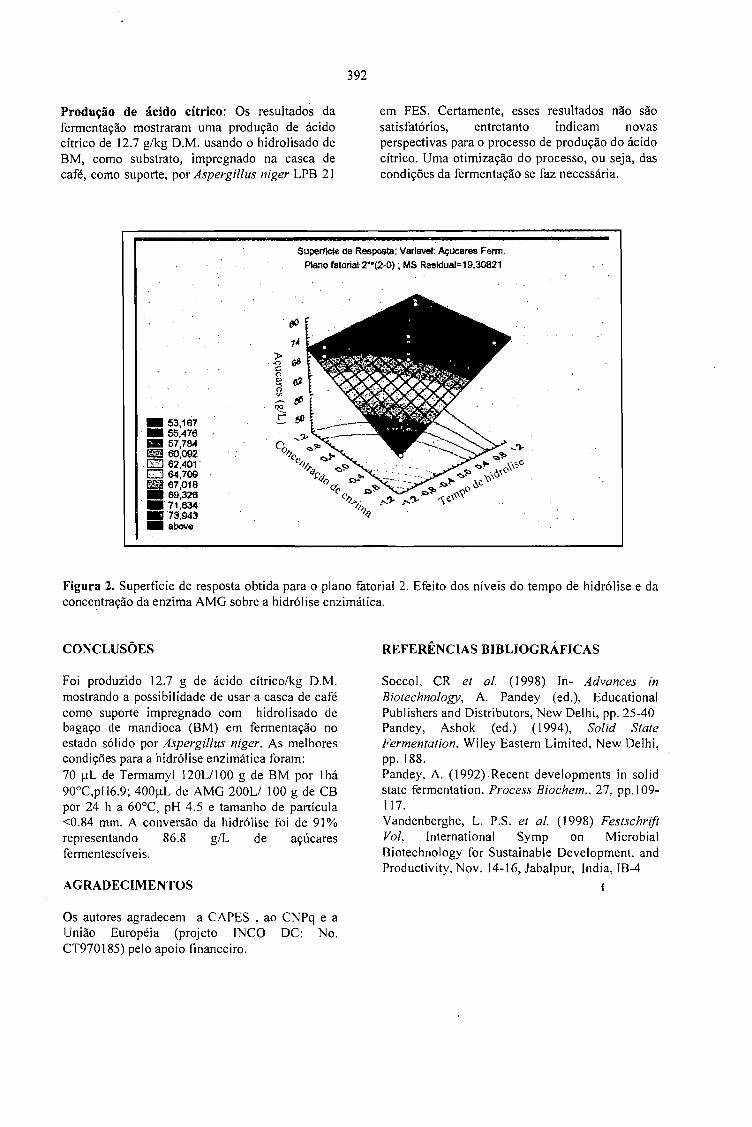
Produção de ácido cítrico: Os resultados dafermentação mostraram uma produção de ácidocítrico de 12.7 g/kg D.M. usando o hidrolisado deBM, como substrato, impregnado na casca decafé, como suporte, por Aspergillus niger LPB 21
392
em FES. Certamente, esses resultados não sãosatisfatórios, entretanto indicam novasperspectivas para o processo de produção do ácidocítrico. Uma otimização do processo, ou seja, dascondições da fermentação se faz necessária.
Superflcie de Re6po&tà; Verlavel: Açlleare6 Ferm.
Plano falorial'2"(2-0) ; MS Residumc19.30821
_53,167_ 55,476_ 57,784_,60,092CD 62,401'064,7.09~ 67,018.69,326
, .71,634'
•73.M3above
Figura 2. Superficie de resposta obtida para o plano fatorial 2. Efeito dos níveis do tempo de hidrólise e daconcentração da enzima AMG sobre a hidrólise enzimática.
CONCLUSÕES
Foi produzido 12.7 g de ácido cítrico/kg D.M.mostrando a possibilidade de usar a casca de cafécomo suporte impregnado com hidrolisado debagaço de mandioca (BM) em fermentação noestado sólido por Aspergillus niger. As melhorescondições para a hidrólise enzimática foram:70 i-LL de Termamyl 120L/l00 g de BM por [há90°C,pH6.9; 400i-LL de AMG 200L/ 100 g de CBpor 24 h a 60°C, pH 4.5 e tamanho de partícula<0.84 mm. A conversão da hidrólise foi de 91%representando 86.8 g/L de açúcaresfermentescíveis.
AGRADECIMENTOS
Os autores agradecem a CAPES. ao CNPq e aUnião Européia (projeto INCa DC: No.CT970185) pelo apoio financeiro.
REFERÊNCIAS BIBLIOGRÁFICAS
Soccol, CR et ai. (1998) In- Advances inBiotechnology, A. Pandey (ed.), EducationalPubJishers and Distributors, New Delhi, pp. 25-40Pandey, Ashok (ed.) (1994), Solid StateFermentation, Wiley Eastern Limited, New Delhi,pp.188.Pandey, A. (1992) Recent developments in solidstate fermentation. Process Biochem., 27, pp.l 09117.Vandenberghe, L. P.S. et al. (1998) FestschriftVol, International Symp on MicrobialBiotechnology for Sustainable Development. andProductivity, Nov. 14-16, Jabalpur, India,l8-4
t

393
LA PULPA DE CAFE ES UN SUBPRODUCTO Y NO UN DESECHO
Jose R. Ramirez Martinez
Universidad Nacional Experimental Del Tachira (Unet)San Cristobal, Estado Tachira, Venezuela
La pulpa de café que se obtiene mediante elbeneficio húmedo de las cerezas de cafégeneral mente se considera como un desecho y losmétodos comunes para deshacerse de estematerial incluyen la descarga en los cursos deagua O el amontonamiento en terrenos agrícolascercanos. En los países productores de café éstoconstituye una fuente muy severa decontaminación y un problema ambiental muyserio. Por esta razón, desde mediados deipresente siglo, se han hecho esfuerzos paradesarrollar métodos para su utilización comomateria prima en la producción de alimentos paraanimales, bebidas, vinagre, biogas, cafeína,pectina, enzimas pécticas, proteína y abonoorgánico. EI uso de la pulpa fresca o procesadaha sido objeto de numerosos estudios los cuales,en general, han conducido a la conclusión de que·se puede usar para sustituir parte de los costososingredientes de la dieta de algunos animales de lafinca. En el Instituto de Nutrición de CentroAmérica y Panamá (lNCAP) de Guatemala selIevó a cabo un intenso programa deinvestigación en los afíos 70 sobre la facti bilidadde la utilización de la pulpa de café (Bressani, R.y Braharn, 1. c., Eds., Pulpa de Café.Composición, tecnología Y utilizacion, CIID,Bogotá, Colombia, ]979). EI énfasis de estosestudios fué sobre su uso potencial como uningrediente. para complementar las raciones delos animales de la finca, lo cual tiene con eltiempo implicaciones económicas para los paísesproductores de café. Los resultados reportadosmostraron que la pulpa de café puede reemplazarhasta un 20% de los concentrados comerciales enla alimentación dei ganado lechero sin efectosadversos y con un ahorro de 30% en los costos.Los resultados en general de los estudios dealimentación de cerdos mostraron que el maízpuede ser reemplazado por pulpa de cafédeshidratada hasta un 16% de la ración total sinefectos detrimentales sobre la ganancia de peso yla conversión alimenticia. Esto significa que aIfinal dei período de engorde cada cerdoproducido ha dejado cerca de 50 kgs. de maízdisponibles para consumo humano u otro usoalternativo.Mas recienternente, un estudio sobre lautilización dela pulpa de café en la alimentación
de ganado lechero se lIevó a efecto en laFacultad de Agronomía de la UniversidadCentral deVenezuela, Maracay (Méndez Medina, 1. B.,1986). Una cantidad conocida de pulpa de caféfresca y alimento concentrado (1 kg. por kg. deleche producida) se les· suministróseparadamente temprano en la rnafiana y pastode corte ad libitum durante el resto deI dia. Seobservó que en general el consumo de pulpa decafé por los bovinos mejoraba la mayoría de susparámetros productivos. En animales jóvenes laganancia de peso fué ligeramente mejor que enaquellos a los cuales no se les suministró pulpade café. Aunque los animales productores deleche tuvieron una ganancia de peso más bajacuando se les alimentó parcialmente con pulpade café, ellos mostraron un ligero aumento enproducción de leche y un aumento significativoen el contenido de grasa.En algunos paises productores de café lacosecha dei cultivo coincide con la estaciónseca; bajo esta circunstancia, la pulpa de cafépuede reemplazar ventajosamente el pasto decorte que se le suministra aI ganado lechero. Lapulpa de café está disponible sólo durante cercade la mitad de] afio y es muy perecedera. Porconsiguiente, requiere un manejo y unapreservación apropiados si se quiere extender elperíodo de su uso. Una manera de manejar ypreservar las grandes cantidades de pulpa decafé que se produce es por medio deI ensilaje.En este proceso el material vegetal a serensilado se - mantiene en un recipienteimpermeable ai aire y ai agua con el fin deproveer condiciones que prevengan sudescomposición. EI material ensilado mantienesu valor nutritivo y puede utilizarse odeshidratarse inmediatamente después de laapertura deI silo. EI silo para hacer el ensilajepuede tener una estructura permanente, pera lossilos de fosa son baratos para construir,requieren poco espacio y proveen un alto gradode protección. EI interior de la fosa se cubre conuna tela de plástico para proteger el silo deIcontacto con el suelo y se provee un drenaje enel fondo. La parte superior se cubre con una telade plástico y encima suelo para proteger lapulpa de café de la Iluvia y mantener lascondiciones anaeróbicas.

394
Recientemente se llevó a cabo un estudio sobre laproducción y evaluación nutricional de la pulpade café ensilada con los esfuerzos conjuntos devarias universidades. Los resultados preliminaresde este estudio fueron presentados en la 17"Conferencia Internacional de Ciencia dei Caféque se realizó en Nairobi, Kenia, dei 20 aI 25 dejulio de 1997. Se preparó un total de 190mini silos cilíndricos, cada uno con una capacidadde 3 kgs. de pulpa de café proveniente deibeneficio húmedo. Se corrieron por cincoperíodos de incubación dos series distintas decuatro tratamientos experimentales los cualesincluyeron diferentes tipos de aditivos y loscorrespondientes controles.AI final de cada período de incubación se midióla temperatura interna de cada minisilo y setomaron muestras para determinar la cantidad debacterias ácido Iácticas, pH, materia seca. ácidosorgánicos de bajo peso molecular, azúcaressolubles, compuestos fenólicos simples, taninoscondensados, y cafeína. Se encontró que lacomposición deI ensilaje dependía en gran partedei origen y manejo de la pulpa de café a serensilada; este hecho explica los resultadoscontroversiales reportados a menudo .Sobre la base de los resultados que se obtuvieroncon los minisilos, se procedió a preparar seis siloscilíndricos grandes (cada uno con una capacidad detres metros cúbicos) para producir la cantidad depulpa de café ensilada (PCE) proveniente deibeneficio húmedo requerida para realizarexperimentos de ai imentación con animales. Lossilos se mantuvieron cerrados por más de 100 díasy se abrieron cuando los experimentos dealimentación animal estaban listos para empezarse.Los experimentos de alimentación se realizaroncon cerdos, peces, pollos, ovejos y conejos. Estosexperimentos incluyeron la determinación deganancia diaria de peso, consumo de materia secadiaria y eficiencia de la conversión alimenticia.Los cerdos alimentados con raciones que conteníanhasta un 15% de pulpa de café ensilada con 5% de
melaza tuvieron igualo mejor ganancia de pesototal que aquellos alimentados con concentradocomerciai. Con relación ai consumo total dealimento no hubo diferencia entre el concentradocomercial y las raciones experimentales quecontenían PCE. Se observó también que éstasdieron una mejor conversión alimenticia que elconcentrado suministrado. Resultados similaresse obtuvieron en experimentos de alimentacióncon híbridos de peces Cachama. EI uso decualquier tipo de PCE para complementar lasraciones de pollos y ovejos no fué satisfactorio.Los conejos que se alimentaron con raciones quecontenían cualquier tipo de PCE toleraron bien supresencia.La pulpa de café que se obtiene dei beneficiohúmedo es sustancialmente diferente de la quese produce manualmente en el laboratorio. Ladiferencia se debe ai lavado de azúcares,cafeína, fenoles y otros compuestos solubles enagua. No hace mucho tiempo se desarrolló unadespulpadora cónica vertical (PenagosHermanos & Co., Bucaramanga, Colombia) queno usa agua. La pulpa de café no lavada que seproduce por este método de procesamiento secoes de esperarse que sea más rica en nutrientes ypudiera Ilegar a ser una materia prima de mejorcalidad para el ensilaje. De todas maneras, sepuede concluir que el ensilaje de la pulpa decafé constituye una alternati va válida paramanejar y almacenar las grandes cantidades depulpa de café que se produce en las plantasbeneficiadoras de cerezas de café alrededor deimundo. La inclusión de PCE en las dietas dealgunos animales de la finca podría contribuir adisminuir los costos de producción de carne yleche, especialmente en los países en vías dedesarrolio.(Traducción ai espafíol dei artículo "CoffeePulp is a by-product, not a waste", publicadoen Tea and Coffee Trade Journal170: 116, 118,120, 122 & 123, 1998)

395
CARACTERIZAÇÃO DE ÁGUAS RESIDUÁRIAS DA LAVAGEM E DESPOLPA DE FRUTOS DOCAFEEIRO E POSSIBILIDADES DE SEU USO NA FERTIRRIGAÇÃO
Matos, A.T.l; Pinto, A.B); Borges, J.D.3
'Prof do Depto. de Eng. AgricolalUFV, 36571-000. Viçosa-MG - 2Mestranda em Eng. Agrícola. Depto. Eng.
AgrícolalUFV - 3 Enga. Agrícola, Depto. Eng. AgrícolalUFV
1 - INTRODUÇÃO
A atividade de lavagem e despolpa de frutos docafeeiro, necessária para obtenção de redução docusto de secagem dos grãos e melhoria naqualidade de bebida. é geradora de grandesvolumes de águas residuárias, ricas em materialorgânico em suspensão e constituintes orgânicos einorgânicos em solução, de grande poderpoluente.A legislação ambiental do Estado de Minas Gerais(Deliberação Normativa COPAM no. 10/86)estabelece que, para o lançamento de águasresiduárias em corpos hídricos. a demanda mg
bioquímica de oxigênio (OB05, 200C) seja de 60
L-I ou a eficiência do sistema de tratamento pararemoção da OBO seja superior a 85% (VonSperling, 1995).Trabalhos recentes indicam que a disposição deáguas residuárias no solo podem proporcionaraumento da produtividade, melhorar a qualidadedos produtos colhidos, promover melhorias emalgumas propriedades fisicas dos solos (Matos et.al., 1996), além de reduzir a poluição ambiental(Matos et al. 1997). Entretanto, a aplicação deáguas residuárias no solo, como forma detratamento ou destinação final de efluentes, deveser feita de forma criteriosa para que não venha acausar prejuízos ao solo. contaminação de águassubterrâneas e superficiais (Matos e Sediyarna.1996). Tendo em vista os problemas expostos,tornou-se objetivo deste trabalho a caracterizaçãodas águas residuárias da lavagem e despolpa dosfrutos do cafeeiro. avaliando-se o seu possívelaproveitamento na fertirrigação de culturasagrícolas.
...2 - MATERIAL E MÉTODOS
Para caracterização da água residuária gerada nalavagem e despolpa de frutos do cafeeiro, na suaforma bruta e após tratamento primário. foiescol hida uma propriedade agrícola, nasproximidades de Viçosa-MG, onde se processa,sazonalmente. este produto agrícola.O sistema de tratamento das águas residuáriasinstalado é constituído de pré-tratamento com
gradeamento simples e, posterior, lançamento doefluente em duas lagoas anaeróbias de contenção,dispostas em sene, sendo a primeira de50mx20mx3m e a segunda de 35mx20mx3m. Estesistema foi utilizado no tratamento da águaresiduária gerada no processamento da safra 98.Em razão da coleta das amostras ter sido feitaapós o período de processamento do fruto. porocasião da coleta das amostras, já não havia águaresiduária bruta para anál ise. Por esta razão, aágua residuária bruta foi produzida emlaboratório, conforme descrito a seguir.Por ser o consumo de água na despolpadora de4.000 L por tonelada de grãos processada, emlaboratório, foi mantida a mesma proporção,misturando-se 1 kg de café em coco para cada 4litros de água. Repetiu-se processo semelhante aoque foram submetidos os frutos para a lavagem edespolpa processados mecanicamente. ou seja. osfrutos ficaram imersos em água por um dia e.posteriormente, foram atritados com as mãos atéque a casca e parte da mucelagem se soltasse dosgrãos. Os grãos e as cascas foram separados dolíquido com uma peneira de malha fina. Dolíquido destituído de material sól ido grosseiro foiretirada uma amostra de 1.6 litros à qual deu-se onome de A 1. Volumes de 1,6 litros das águasresiduárias da primeira (A2) e segunda (A3)lagoas foram coletados a 20 em de profundidade elevados ao laboratório para. juntamente com aamostra A 1. serem submetidas. seguindo-semetodologia da APHA (1995), às seguintesanálises: sólidos sedimentáveis (SP), sólidos totais(ST), condutividade elétrica (CE), pH. nitrogêniototal (N), fósforo total (P). potássio total (K),demanda química de oxigênio (OQO) e demandabioquímica de oxigênio (OBO) .
3 - RESULTADOS E DISCUSSÃO
A grande diferença observada entre o valor de SPencontrado nas amostra A I em relação àsamostras A2 e A3 deve-se à decantação domaterial grosseiro e a conseqüente formação delodo nas lagoas anaeróbias. Pelo mesmo motivo,foram encontradas grandes diferenças nos valoresde ST.Os valores de pH, levemente ácido na águaresiduária bruta, elevou-se nas lagoas anaeróbias.

396
Quadro 1 - Parâmetros de qualidade da água residuáriaproveniente da lavagem e despolpa de frutos do cafeeiro.
5 - REFERÊNCIAS BIBLIOGRÁFICAS
APHA 1995. Standards methods for theexamination or water and wastewater APHA.
A WWA and WPCR. 19a
ed., New York.Ayers, R.S.; D.W. Westcot. 1991. A qualidade daágua na agricultura. Capina Grande, UFPB. 2 J 8 p.(Estudos FAO: Irrigação e Drenagem, 29)Bernardo, S. 1995. Manual de irrigação. 657 p.Malavolta, E.E.; H.J. Kliemann. 1985. Desordensnutricionais no cerrado. Piracicaba. SP.Matos, A.T. e M.A.N. Sediyama. 1996. Riscospotenciais ao ambiente pela aplicação de dejetolíquido de suínos ou compostos orgânicos no solo.In: Freitas. R.T.F. e Viana, C.F.A. 1 Semináriomineiro sobre manejo e utilização de dejetos desuinos. EPAMIG,EMATER,UFV,ASSUVAP.p.45-54.Matos, A.T. e M.A.N. Sediyama, S.M. Vidigal.1996. Características químicas. fisicas emicrobiológicas do solo influenciadas pelaapl icação de compostos orgânicos ou dejetolíquido de suínos. In: Freitas, R.T.F. e Viana.C.F.A. I Seminário mineiro sobre manejo eutilização de dejetos de suínos.EPAMIG.EMATER,UFV,ASSUVAP. p.35-44.Matos, A.T., M.A.N. Sediyama, S. P. Freitas,S.M. Vidigal. N.C.P. Garcia. 1997. Característicasquímicas e microbiológicas dos solo influenciadaspela aplicação de dejeto líquido de suínos. RevistaCeres, 44(254): 399-410.Von Sperling, M.; Costa. A. L. M.; Castro, A.A.1995. Esgotos 'sanitários. In: Barros, RT. V.;Chemicharo, C.A.L.; Hel1er, L.; von Sperling, M.Manual de Saneamento e proteção ambiental paraos municípios. DESA/UFMG. p. 113-160.
nutrientes de interesse agrícola, podem seraproveitadas na fertirrigação de lavouras,entretanto, o potássio, por ser o nutriente presenteem maior concentração na água residuária, deveser utilizado como referencial para o cálculo dasua taxa de aplicação no solo.
mostr pH CE SP ST N P K OQOOBO
~S m-I hnl. L-Ig L-I -------- mg L-I ____ •
I 6.4 ~.863 88,0 10.6 ~62.6 15 1.206 7.999 5.859V\2 7.1 0.985 2.0 0.8 44.9 0.6 154 147 691A3 7.1 1.186 1,9 1.0 38.0 INO /68 539 f410..AI - agua residuâria bruta; A2 - prrmerra lagoaanaeróbia; A3 - segunda lagoa anaeróbia.NO· parâmetronão detectado.
Verificou-se, menor concentração de K, DQO eDBO, na amostra A2 do que na A3. Acredita-seque a lavagem do maquinário e do pátio, após otérmino do período de processamento da safraanual, proporcionou a diluição da água residuáriaarmazenada na primeira lagoa, o que pode terproporcionado diminuição da concentração dessesparâmetros.No que refere-se à condutividade elétrica. osvalores encontrados indicam que o grau derestrição para uso das águas em irrigação,segundo Ayers & Westcot (1991), é consideradosevero para a água A 1 e Iigei 1'0 a moderado paraA2 e A3. Esses resultados indicam que o uso,principalmente da água bruta, para suprimento dasnecessidades hídricas da cultura, por exemplo docafé. que segundo Bernardo (1995) é da rdem de1200 mrn há,l ano' ou 12.000 m' ha' ano", deveser evitado, sob risco de se proporcionar asalinização do solo.Tomando-se por base as recomendações deadubação nitrogenada e potássica para a culturado café que, de acordo com Malavolta e Kliemann(1985), é de 200 kg há" ano" de N e de K10 . osvolumes de água residuária bruta da lavagem edespolpa de frutos do cafeeiro deve ser,respectivamente. de 432 rrr' há' ano" e 138 rrr'há" ano". Para a fertirrigação utilizando água dalagoa I, podem ser usadas, respectivamente, 4.454rrr' há'] ano" e 1.300 rrr' há" ano" e, no caso deuso de água da lagoa 2. de 5.267 m3 há-I ano' I e746 m3 há" ano". Maiores taxas de aplicação sóseriam recomendáveis no caso de monitoramentoquímico do solo, com o intuito de evitar danos aele e à cultura explorada.
4 - CONCLUSÃO
As águas residuárias da lavagem e despolpa defrutos do cafeeiro. em razão da elevada DBO queapresentam. se lançadas em cursos d'água. semtratamento prévio. devem causar poluição degrande impacto ambiental.O tratamento primário em lagoas anaeróbiasmostrou-se eficiente na redução de todos osparâmetros. à exceção do pl-l, avaliados; as águasresiduárias da lavagem e despolpa de frutos docafeeiro, por serem constituídas de muitos

SELECTION OF STRAINS OF VolvariellaEXTRACT OF COFFEE HUSK
397
volvacea ANO CHARACTERISTICS ON THE
Fan Leifa, Ashok Pandey, Carlos Ricardo Soccol and R. Mohan
Laboratorio de Processos Biotecnologicos, Departamento de Engenharia Quimica, Universidade Federal doParana, CEP 81531-970, Curitiba-PR, Brazil
INTROOUCTION
Coffee production is an economically importantagro-industry in more than fifty countries (ICO,1998). While most ofthe industries adopts the wetprocessing method (ICO, 1998), in Brazilmajority of the industries use dry-method andonly about 10% of production comes from thatmethod (SOCCOL, 1995). The wet methodgenerates a great volume of liquid, causingenvironmental problems and resuiting in loss oforganic substance with a lot of sugar and pectin (SOCCOL, 1996).
Volvariella volvacea is an edible mushroom,growing in the tropical and subtropical regions atthe temperature of 25-40 °C ( SHAFFER, 1957;REYNOLDS, 1967). This mushroom is a goodsource of protein, general1y containing 20-40% ofprotein (dry) (CHANG e MILES, 1984; Yang,1986). Mushroom mycelium, like the sporophore,is a good source of amino acids and B-vitamins(Block, 1953). Since first producing of mushroommycelium (HUMFELO, 1948), many works havebeen developed to explore the utilities ofmycelium or produce enzymes, antibiotics andanticancer agents due to its rapid propagation(RAMOS, 1967).The works of V. bakeri and V. bombicina on thecoffee pulp were undertaken ( MARTINEZ, 1984;THIELKE, 1989), but no works have been foundon the liquid residue of coffee processing byusing V. volvacea. This work aims to is to selectsuitable strain of this fungus for the use of coffeehusk which is also in great volume in Brazil andcultivate this fungus as an aliment. The otherobjective is to determine some parameters ofgrowth of this fungus in the uti Iisation of theresidue for later exploitation of the fungusmycelium.
MATERIALS ANO METHOOS
Micro-organism. f. volvacea LPB 59. I I, 23.09, 62, routinely maintained at ambienttemperature on POA, were used for selection ofstrain in this experiment.
Those strains were transferred from tubes to petriplates before the experiments.
Medium preparation, The 40g of dry coffeehusk was used for preparing Ilitre the solution ofthe extract through one hour cocking. After thefiltration, pH of the liquid was adjusted to 7,0.,then added with theAgar. After autoclave, the medium of about 15 mlwas distributed to the each petri-plate (10 em ).The inoeulation was made with a disk of 5 mm indiameter (Soeeol, 1994). Incubation wasundertaken in the dark at 32 "C.
Evaluation of growth. From the third daybeginning measure the growing of mycelium tillthe myeelium firstly reached the margins of theplate. Then weighing the biomass through thedissolving the mediurn, fi Itrating and drying(Soeeol, 1994).
pH and temperature test. The mediumpreparation and evaluation of growing of themycelia was undertaken in the same way as theabove. pH was effected at 4,0, 5,0, 6,0, 7,0, 8,0,9,0 after the autoelave. Temperature was adoptedat8°C,20°C,24°C,28°C,32°C,36°C.
Nitrogen supplements and soybean flour Thebasie medium was prepared in the same wayabove. The source of nitrogen was NaN03 ,
soybean flour, (NH4hHP04 , yeast extract,peptone at the eoneentration of 2 g/L. Theconcentration of soybean flour was used at 2, 4, 6.8, 10 g/L respeetively.
RESULTS
Selection Of Strain Of V. Volvacea In TheExtract OfCoffee Husk.The Figure 1 shows the eolony eharaeteristies ofthe different strains of V. volvacea in thisexperiment. 1t was observed that the strain V.volvacea LPB 09 is the best with more white anddensity of myeelia in this médium, the radialmycelial veloeity being 15,34 mm/day andbiomass 35,29 mg/plate in 5 days at 32 0e. So thestrain V. volvacea LPB 09 was seleeted out for thefurther studies.
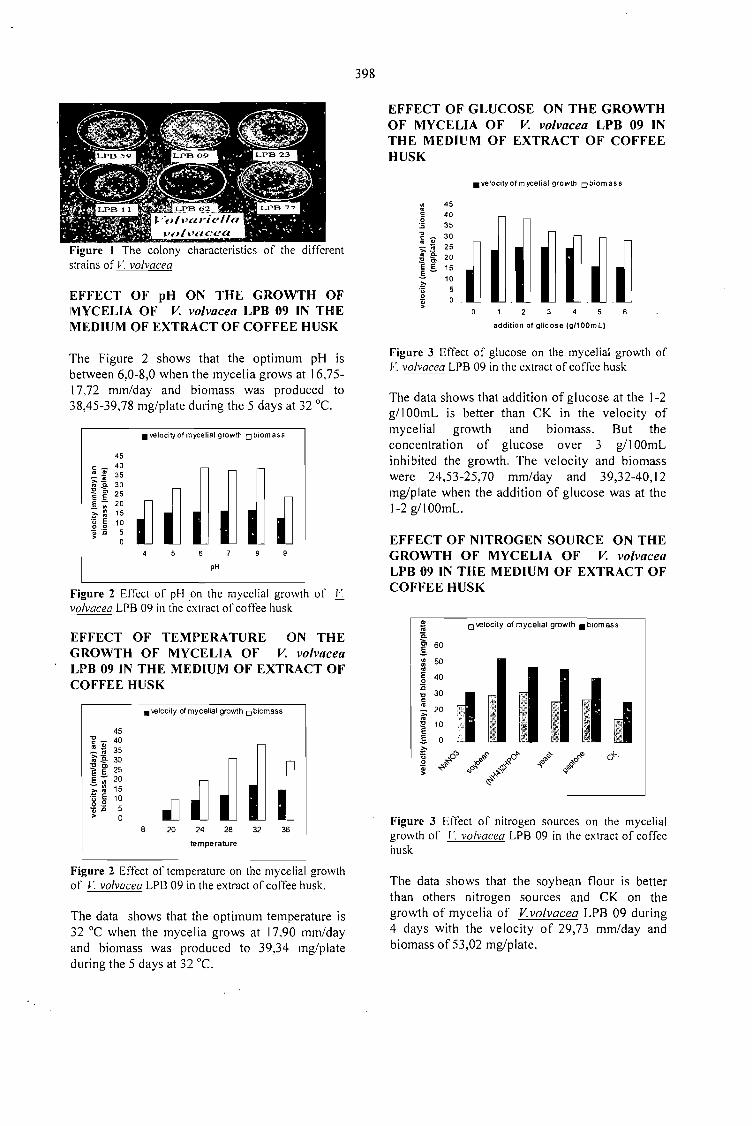
c:"-·
Figure I The eolony eharaeteristies of the differentstrains of V. volvacea
EFFECT OF pH ON THE GROWTH OFMYCELlA OF V. volvacea LPB 09 IN THEMEDlUM OF EXTRACT OF COFFEE HUSK
The Figure 2 shows that the optimum pH isbetween 6,0-8,0 when the myeelia grows at 16,7517,72 mm/day and biomass was produced to38,45-39,78 mg/plate during the 5 days at 32 "C.
• velocity of mycelial growth o biomass
45c: _ 40
~ ~ 35ClI a. 30
~ ~ 25E - 20- ~~ ~ 15'g ~ 10"i:ti 5> o
Figure 2 Effect of pl-l on the myeelial growth of I'.volvacea LPB 09 in the extract 01' eoffee husk
EFFECT OF TEMPERATURE ON THEGROWTH OF MYCELlA OF V. volvaceaLPB 09 IN THE MEDIUM OF EXTRACT OFCOFFEE HUSK
_",Iocily of mycelial growth obiomass
45-g _ 40
~ ~ 351;':e. 30
~ ~ 25E ;; 20;:: ~ 15=ã f5 10~:õ 5> o
8 20 24 28 32 36
temperatura
Figure 2 Effeet of temperaturc on the rnycelial growthof V. volvacea LPB 09 in lhe extract of coffee husk.
The data shows that the optimum temperature is32°C when the mycelia grows at 17,90 mm/dayand biomass was produced to 39,34 mg/plateduring the 5 days at 32 -c.
398
EFFECT OF GLUCOSE ON THE GROWTHOF MYCELlA OF V. volvacea LPB 09 INTHE MEDIUM OF EXTRACT OF COFFEEHUSK
.velocityofmycelial growth obiomass
~ 45E 40o
35:c"C 30
~~~Lc _~ ~
25
.~>:~
~~ 20E E 15§.-
10i>
5'üo o..>
addition of glicose (g/100mL)
Figure 3 Effeet of glueose on the myeelialgrowth ofV. volvacea LPB 09 in the extraet of coffee husk
The data shows that addition of glucose at the 1-2g/100mL is better than CK in the velocity ofmycelial growth and biomass. But theconcentration of glucose over 3 g/100mLinhibíted the growth. The velocity and biomasswere 24,53-25,70 mm/day and 39,32-40,12mg/plate when the addition of glucose was at the1-2 g/l OOmL.
EFFECT OF NITROGEN SOURCE ON THEGROWTH OF MYCELlA OF V. volvaceaLPB 09 IN THE MEDIUM OF EXTRACT OFCOFFEE HUSK
~ o velocity of mycelial growth • biomassã.C, 60§.:il 50..5 40:c
I~ d€ on;,~ ~#>
Figure 3 Effeet of nitrogen sourees on lhe mycelialgrowth of r'. volvacea LPB 09 in the extract of eoffeehusk
The data shows that the soybean flour is betterthan others nitrogen sources and CK on thegrowth of mycel ia of V. volvacea LPB 09 during4 days with the velocity of 29,73 mm/day andbiomass of 53,02 mg/plate.

EFFECT OF ADDlTION OF SOYBEANFLOUR ON THE GROWTH OF MYCELlAOF V. volvacea LPB 09 IN THE MEDlUM OFEXTRACT OF COFFEE HUSK
.Wlocityofm\<'elial growlh obiomass
'"45.. 40E
.2 35<>-g _ 30
~~~ í.. ..~!! 25.. Co:E C, 20E E 15.s-l:' 10'ü 5~> O
O 4 6 8 10
additlon of soybean fiou. (gIL)
Figure 3 Effect of concentration of soybean on themycelial growth of V. vo/vacea LPB 09 in the extractof coffee husk
The data shows that the addition of soybean flourwas a positive effect on the fungai growth until 8g/L. lt gained the radial growing velocity of27,56mm/day and biomass of 42,74 mg/plate at theaddition of soybean flour of 6 g/L during 4 daysat 32 oCO
DlSCUSSION
The studies above have demonstrated that theextract of coffee husk from the dry-methodprocessing of coffee industries serve as a goodbasic medium for Vvolvacea, so that it isconduced that the Iiquid residue of wet processingmethod might also be used as a médium for thisfungus. The optimisation of some parameters forthe mycel ial growth of this fungus on the extractof coffee husk was in agreement with the studies(Fan, 1986).Through the analysis of the extract of coffee husk,we found that the majority of caffeine and tanninsin the coffee husk was removed into the extractafter cooking. Initially, the mycel ium growsslower util some replication of adaptatíon whenthe mycelium grows more rapidly. As for thedegradation of caffeine and tannins, some workswi 11 be undertaken later.
399
REFERENCES
Block, S. S. Mushroom mycelium in experimentswith submerged culture Jour. Agr. Food Chem.USA.Chang, S.T. and Miles, P.G. 1984 A new look atcultivated mushroom Bioscience, USA.Chang, S.T. 1989 Edible mushroom and theircultivation CRC Press, Inc. , Florida.Fan, L.F. and Fang, J.L. 1986 Selection of strainsand cultural techniques of Volvariella volvaceaZhejiang Edible fungi. Zhejiang, China.International Coffee Organization 1998 Totalproduction of exporting membershttp://www.ico.org/proddoc.htmHumfeld, H. 1948 The production of mushroommycelium in submerged culture Science.Martinez, D. and Quirarte, M. 1984 Prospects ofgrowing edible mushroom on agro-industrialresidues in Mexico Boletin de la sociedadMexicana de micologia, Mexico.Ramos, M. A. 1967 A study on submergedculture production of V. volvacea mucelium as asouce of protein, B-vitaminas and food flavor.The Phi lippine Journal of Science.Soccol, C.R. 1994 Contribuição ao estudo dafermentação no estado sólido em relação com aprodução de ácido fumárico. Biotransformação deresíduo sólido de mandioca por Rhyzopus eBasidiomacromicetos do gênero Pleurotus Tesede Livre-Docência. UFPR,Curítiba, 1994. 228p.Soccol, C.R. 1995 Aplicações da fermentação noestado sólido na valorização deresíduosagroindustriais. França-Flash Agricultura.Soccol, C.R 1996 Aplicaciones Agroindustrialesde los Processos de Fermentciones em MediosSolidos. Biotechnologia y Bioingeneria.Sociedade Mexicana de Biotechnologia yBioengeneria, Mexico.Thiclke, C. 1989 Cultivation of edible fungi oncoffee grounds Mushroom Science XII (parte 11)Germany.Yang, X. M. 1986 Cultivation of ediblemushroorn in China Agriculture printing house,Beijing, China.


401
DETOXIFICAÇÃO BIOLÓGICA DA CASCA DE CAFÉ POR FUNGOS FILAMENTOSOS EMFERMENTAÇÃO NO ESTADO SÓLIDO
Debora Brandi, Fernando Kawata', Ashok Pandey', Sevastianos Roussos', Maria Carolina Rocha dosSantos', e Carlos Ricardo. Soccol'
ILaboratorio de Processos Biotecnologicos, Departamento de Engenharia Química.Universidade Federal do Paraná, CEP 81531-970 Curitiba-PR, Brazil2Laboratoire de Microbiologie ORSTOM, Université de Provence, 132288 Marseil1e, France.
INTRODUÇÃO
A transformação tecnológica do café, gera umagrande quantidade de resíduos não aproveitadoscomo: cascas, folhas e borra, que podem serutilizados como substrato de fermentação por seugrande conteúdo nutricional. A casca de caféapresenta um baixo valor nutricional, devido apresença de substâncias tóxicas como cafeí na (1,2%), taninos (6,3%) e polifenóis. Uma alternativapara melhorar este conteúdo é a utilização dafermentação no estado sólido, empregandomicroorganismos capazes de utilizar a cafeínacomo fonte de nitrogênio. A fermentação noestado sólido é considerada a mais natural dasfermentações, porque seus processos assemelhamse as condições sob as quais a maioria dosmicrorganismos crescem na natureza. É definidacomo o crescimento de microorganismos emmateriais sólidos na ausência de água livre, mas osubstrato deve conter umidade suficiente, presentena forma absorvida dentro da matriz sólida.O objetívo do presente trabalho foi incrementar ovalor biológico da casca de café, peladetoxificação de seus componentes tóxicos porfungos filamentosos, util izando a técnica defermentação no estado sólido.
MATERIAL E MÉTODOS
Seleção de MicroorganismosOs microorganísmos foram selecionados combase na sua velocidade de crescimento radial ebiomassa produzida em ágar extrato de casca decafé (lOOg/L) Foram utilizadas onze cepas defungos filamentosos do gênero Rhizopus e duas dogênero Phanerochaete.O crescimento radial em ágar extrato de casca decafé foi realizado por meio de inoculação dosmicroorganismos estudados no centro de umaplaca de Petri. A biomassa foi medida através dadissolução do ágar e separação da biomassa, quefoi então pesada em um papel de filtropreviamente tarado.
Preparo do Ágar extrato de casca de caféPesou-se 100 g de casca de café previamentepulverizada com partículas inferiores à 0,8 mm eadicionou-se 1000 mL de água destilada em umfrasco Erlenmeyer de 2000 mL. Cozinhou-se emautoclave à 100° C, durante I hora. Filtrou-se ecompletou-se o vo Iume para 1000 mL.Adicionou-se então 20 g/L de ágar bacteriológico.Esterilizou-se o meio em autoclave a 121° Cdurante 15 minutos.
FERMENTAÇÃO NO ESTADO SÓLIDO
Preparo do substratoA casca de café inicialmente foi seca em estufa
com circulação de ar à 55° C por 48 horas atéatingir uma umidade de aproximadamente 8 %. Osubstrato foi conduzido a um processo de moagemem moinho manual e posteriormente foi realizadauma classificação do material por meio depeneiras. Utilizou-se a fração granulométricacompreendida entre 0,8 e 2,0 mm.Pesou-se 20 g da casca de café previamentemoída, em frasco Erlenmeyer de 250 mL,adicionou-se 50 % de água à partir da umidadecalculada e procedeu-se a esterilização domaterial. As fermentações foram feitas emduplicata, variando-se as condições iniciais de pHe umidade para cada experimento.
Preparo do inóculoPhanerochete chrysosporium: O fungo foiinoculado em placas com meio POA (ágar batatadextrose), que foi então incubado em estufa a 35°C durante 10 dias. Os esporos foram coletados emCâmara de Fluxo Laminar com o auxílio de umaalça de platina e transferidos para um frascoErlenmeyer de 125 mL contendo água desti lada egotas de tween 80 que foram previamenteesterilizados. Faz-se então a contagem do númerode esporos em câmara de Neubauer.Rhlzopus arrhizus: Utilizou-se a mesma técnicadescrita acima, sendo que a temperatura deincubação é de 32° C.
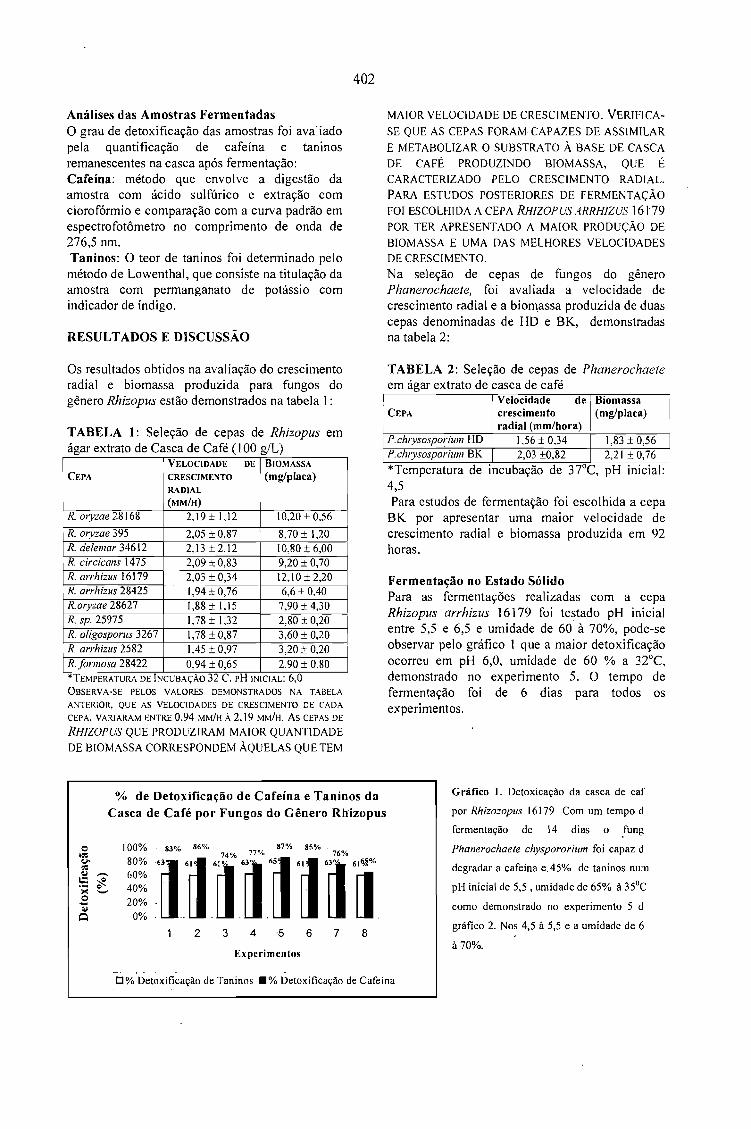
402
Análises das Amostras FermentadasO grau de detoxificação das amostras foi avaliadopela quantificação de cafeína e taninosremanescentes na casca após fermentação:Cafeína: método que envolve a digestão daamostra com ácido sulfúrico e extração comclorofórmio e comparação com a curva padrão emespectro fotômetro no comprimento de onda de276,5 nm.Taninos: O teor de taninos foi determinado pelométodo de Lowenthal, que consiste na titulação daamostra com permanganato de potássio comindicador de índigo.
RESULTADOS E DISCUSSÃO
MAIOR VELOCIDADE DECRESCIMENTO. VERIFICA
SE QUE AS CEPAS FORAM CAPAZES DE ASSIMILARE METABOLIZAR O SUBSTRATO À BASE DE CASCADE CAFÉ PRODUZINDO BIOMASSA, QUE ÉCARACTERIZADO PELO CRESCIMENTO, RADIAL.PARA ESTUDOS POSTERIORES DE FERMENTAÇÃOFOI ESCOLHIDA A CEPA RH1ZOPUSARRH1ZUS16179POR TER APRESENTADO A MAIOR PRODUÇÃO DEBIOMASSA E UMA DAS MELHORES VELOCIDADESDE CRESCIMENTO.
Na seleção de cepas de fungos do gêneroPhanerochaete, foi aval iada a velocidade decrescimento radial e a biomassa produzida de duascepas denominadas de HD e BK, demonstradasna tabela 2:
TABELA 2: Seleção de cepas de Phanerochaeteem ágar extrato de casca de café
*Temperatura de incubação de 37°C, pH inicial:4,5Para estudos de fermentação foi escolhida a cepa
BK por apresentar uma maior velocidade decrescimento radial e biomassa produzida em 92horas.
Fermentação no Estado SólidoPara as fermentações realizadas com a cepaRhizopus arrhizus 16179 foi testado pH inicialentre 5,5 e 6,5 e umidade de 60 à 70%, pode-seobservar pelo gráfico 1 que a maior detoxificaçãoocorreu em pH 6,0, umidade de 60 % a 32°C,demonstrado no experimento 5. O tempo defermentação foi de 6 dias para todos osexperimentos.
Velocidade de II BiomassaCEPA crescimento (mg/placa)
radial (mm/hora)P.cllI:vsosporium HD 1,56 ± 0,34 I 1,83 ± 0,56Píchrysosporium BK 2,03 ±0,82 I 2,21 ± 0,76- ..
TABELA 1: Seleção de cepas de Rhizopus emágar extrato de Casca de Café (100 g/L)
Os resultados obtidos na avaliação do crescimentoradial e biomassa produzida para fungos dogênero Rhizopus estão demonstrados na tabela 1:
*TEMPERATURA DE INCUBAÇAO 32 C. pH INICIAL: 6,0OBSERVA-SE PELOS VALORES DEMONSTRADOS NA TABELA
ANTERIOR, QUE AS VELOCIDADES DE CRESCIMENTO DE CADA
CEPA, VARIARAM ENTRE 0,94 MM/H À 2,19 MM/H. As CEPAS DE
RH1ZOPUSQUE PRODUZ[RAM MAIORQUANTIDADEDE BIOMASSA CORRESPONDEM ÀQUELAS QUETEM
VELOCIDADE DE BIOMASSACEPA CRESCIMENTO (mg/placa)
RADIAL(MM/H)
R. oryzae 28 J68 2,19 ± 1,12 10.20 ± 056
R. oryzae 395 2,05 ± 0,87 8,70 ± \,20R. de/emar 34612 2,13 ±2.12 10,80 ± 6,00
IR. circicans 1475 2,09 ±0,83 9,20 ± 0,70R. arrhizus 16179 2,03 ± 0,34 12,10 ± 2,20R. arrhizus 28425 1,94±0,76 6,6 ± OAOR.oryzae 28627 1,88 ± 1,15 7,90 ± 4,30R. sp. 25975 1.78 ± 1,32 2,80 ± 0,20R. o/igosporus3267 1,78 ± 0,87 3,60 ± 0,20R. arrhizus 2582 1.45 ± 0,97 3,20 ± 0.20R.formosa 28422 0,94 ± 0,65 2.90 ± 0,80
% de Detoxificação de Cafeína e Taninos daCasca de Café por Fungos do Gênero Rhizopus
100%80%60%40%20%
0%
2 3 4 ,5
Experimentos
6 7 8
G rático l. Detoxicação da casca de caf
por Rhizozopus 16179 Com um tempo d
fermentação de 14 dias o ,fung
Phanerochaete chyspororium foi capaz d
degradar a cafeína e,45% de taninos num
pH inicial de 5,5 , umidade de 65% à 35°C
como demonstrado no experimento 5 d
gráfico 2. Nos 4,5 à 5,5 e a umidade de 6
à 70%.
0% Detoxificação de Taninos .% Detoxificação de Cafeína
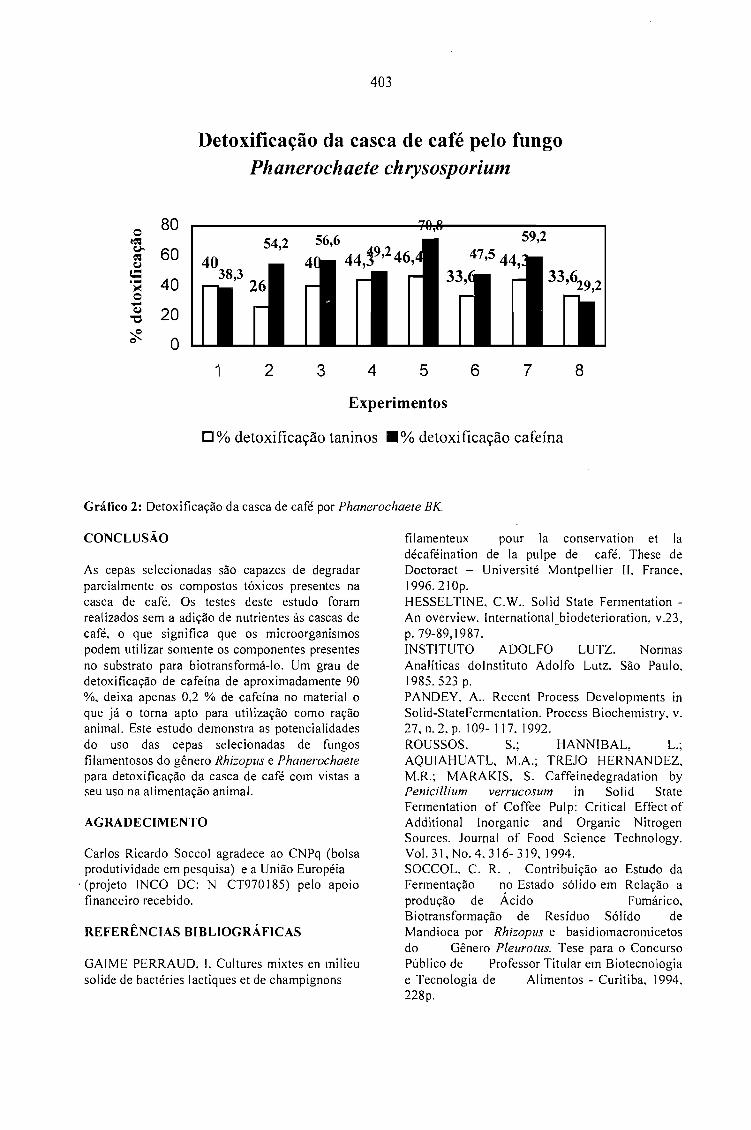
403
Detoxificação da casca de café pelo fungoPhanerochaete chrysosporium
o 80l~ 54,2 56,6e;;-
60 44,19,246,~ 4C.l
'-= 40....~o......~ 20"'O
~Oo
1 2 3 4 5 6 7 8
Experimentos
0% detoxificação taninos • % detoxificação cafeína
Gráfico 2: Detoxificação da casca de café por Phanerochaete BK.
CONCLUSÃO
As cepas selecionadas são capazes de degradarparcialmente os compostos tóxicos presentes nacasca de café. Os testes deste estudo foramreal izados sem a adição de nutrientes às cascas decafé. o que signi fica que os microorganismospodem utilizar somente os componentes presentesno substrato para biotransforrná-lo. Um grau dedetoxificação de cafeína de aproximadamente 90%. deixa apenas 0,2 % de cafeína no material oque já o torna apto para utilização como raçãoanimal. Este estudo demonstra as potencialidadesdo uso das cepas selecionadas de fungosfilamentosos do gênero Rhizopus e Phanerochaetepara detoxificação da casca de café com vistas aseu uso na ai imentação animal.
AGRADECIMENTO
Carlos Ricardo Soccol agradece ao CNPq (bolsaprodutividade em pesquisa) e a União Européia
. (projeto INCO DC: N CT970185) pelo apoiofinanceiro recebido.
REFERÊNCIAS BIBLIOGRÁFICAS
GAIME PERRAUD. I. Cultures mixtes en milieusol ide de bactéries lactiques et de champignons
fi lamenteux pour la conservation et ladécaféination de la pulpe de café. These deDoctoract - Université Montpellier 11. France,1996.210p.HESSELTINE. C.W.. Solid State Fermentation An overview. International_biodeterioration, v.23,p. 79-89,1987.INSTITUTO ADOLFO LUTZ. NormasAnalíticas dolnstituto Adolfo Lutz. São Paulo.1985.523 p.PANDEY. A.. Recent Process Developments inSolid-StateFermentation. Process Biochernistry, v.27. n. 2. p. 109- 117. 1992.ROUSSOS, S.; HANNIBAL, L.;AQUIAHUATL. M.A.; TREJO HERNANDEZ,M.R.; MARAKIS, S. Caffeinedegradation byPenicillium verrucosum in Solid StateFennentation of Coffee Pulp: Criticai Effect ofAdditional Inorganic and Organic NitrogenSources. Journal of Food Science Technology.VoI.31.No.4,316-319.l994.SOCCOL. C. R.. Contribuição ao Estudo daFermentação no Estado sólido em Relação aprodução de Ácido Fumárico,Biotransformação de Resíduo Sól ido deMandioca por Rhizopus e basidiomacromicetosdo Gênero Pleurotus. Tese para o ConcursoPúblico de Professor Titular em Biotecnologiae Tecnologia de Alimentos - Curitiba. 1994.228p.


405
CASCA DE CAFÉ COMO SUBSTRATO PARA PRODUÇÃO DE GA 3 POR FERMENTAÇÃO
Cristina M. M. Machado; Carlos R. Soccol', Brás H. Oliveira,' Carlos Oliveira', Ashok Pandey'
'Laboratório de Processos Biotecnológicos, Departamento de Engenharia Química,Universidade Federal do Paraná (UFPR), CEP 81531-970 Curitiba-PR, Brazil;2Laboratório de Produtos Naturais
INTRODUÇÃO
Ácido giberélico (GA 3) é um hormônio endógenode plantas superiores e metabólito secundário decertos fungos. É um importante promotor eregulador do crescimento das plantas sendoutilizado na agricultura, viveiros, produção deflores, etc., para uma série de efeitos benéficos,como eliminação da dormência e aceleração degerminação em sementes, melhora no rendimentodas colheitas, promoção da formação do fruto,indução do florescimento e outras. Nos últimosanos, este metabólito tem sido produzidoindustrialmente por fermentação submersa a umalto custo (US$.l a 3/grama, dependendo dapureza e potência), uma vez que o rendimento émuito baixo e sua extração difícil. A redução domesmo levará suas diversas aplicações a umautil ização mais extensa, trazendo ótimosbenefícios econômicas à agricultura e indústria.Uma possibilidade de diminuição dos custos estána pesquisa de novas técnicas de fermentação(Kumar e Lonsane, 1987, 1989; Brückner eBlechschmidt, 1991).Recentemente, tem aumentado o interesse decientistas no desenvolvimento de produção degiberelinas por fermentação em estado sólido(FES). Fermentação sobre farelo de trigo édescrita por Kumar e Lonsane (1987), quecompararam o rendimento com a técnicasubmersa. Tal comparação também foi feita porTomasini et ai. (1997), utilizando-se comosubstratos farinha de mandioca, bagaço de canade-açúcar e poliuretano de baixa densidade.Ambos grupos obtiveram maior rendimento com aFES, no caso de Kumar e Lonsane de 1,6 vezes. ecusto cerca de 2,5 vezes menor.O termo fermentação no estado sólido (FES),tradução do inglês "solid state fermentation", éobjeto de numerosas definições. Pandey (1992),define como a fermentação na qual o crescimentodo microrganismo em substratos sólidos ocorre naausência de líquido na forma livre. A água livreindispensável ao crescimento dos microrganismoé adsorvida num suporte sólido ou complexada nointerior de uma matriz sólida. A FES éconsiderada mais natural que outros tipos defermentações, como, por exemplo, a fermentação
submersa, porque seus processos assemelham-seàs condições sob as quais a maioria dosmicrorganismo crescem na natureza (Hesseltine,1972). Diversos resíduos agrícolas ou agroindustriais, celulósicos ou amiláceos têm sidoutilizados em fermentação no estado sólido, taiscomo bagaço de cana-de-açúcar, de mandioca e demaçã, batata refugo, palha de arroz (Soccol, 1994;Sturza, 1995; Ayala, 1996; Bramorski, 1997).Durante o processamento do café, a polpa, acamada mucilaginosa e a casca são removidas.Estes resíduos representam cerca de 50% emmassa do café. Apesar do grande volume deresíduos e alguns esforços isolados na utilizaçãodos resíduos do café, a maior parte destaabundante fonte natural continua sendo dispostacomo resíduo (Molina, et al., 1974; Brahan andBressani, 1978).Com este trabalho pretende-se determinar apossibilidade da utilização da casca do café comomeio de cultivo para produção de ácido giberélicopor fermentação. Para tanto foram utilizadas seiscepas de microrganismos produtores de GA 3 emfermentação no estado sólido e submersa,comparando-se dessa forma, o rendimento dosmétodos.
MATERIAL E MÉTODOS
Microrganismos e SubstratosSeis cepas de Gibberella fujikuroi e Fusariummoniliforme, mantidas em ágar dextrose-batata(PDA) em tubos inclinados foram inoculadas emmeio Czapek Dox em shaker rotatório a 30°C por4 dias para desenvolvimento da solução semente(Bandelier, et al., 1996).O meio para fermentação submersa foi feito apartir do cozimento de 200 g de substrato (cascade café) por uma hora em autoclave a 100° C eposterior filtragem do mesmo. O resíduo sólido,foi seco em estufa e também utilizado comosubstrato para FES. O filtrado teve seu pHajustado e foi esterilizado. Os meios sólidosconsistem na casca de café, e no resíduo daextração, em granulometria entre 0,8-2,0 mm,com pH e umidade ajustados para as condiçõesdeterminadas da fermentação. Estes meios sãoentão esterilizados em autoclave a 121° C por 15minutos.

406
Condições de fermentaçãoPara as fermentações submersas inoculou-sesolução semente à 50 mL de meio líquido extratode café, em frascos erlenmeyer de 250 mL. Nasfermentações em estado sólido inoculou-sesolução semente à 10 g de meio sólido casca decafé, ou resíduo café em frascos erlenmeyer de250 mL.Após a fermentação, os meios foram extraídoscom acetato de etila a pH 2,5. A seguir, asamostras analisadas em cromatografia de camadadelgada (TLC) para confirmação da presença deGA3 . Uma vez confirmada a presença do mesmo,as amostras que apresentaram melhorfluorescência foram purificadas e analisadas emcromatografia líquida de alta eficiência (HPLC).As fermentações foram levadas em condiçõesiguais de temperatura, quantidade de meio, razãode inóculo e tempo de fermentação. Osprocedimentos de extração e purificação dasamostras também foram os mesmos, acimadescritos, val idando, desta forma, a comparaçãodos substratos.As condições das fermentações são mostradas naTabela I a seguir:
trabalho Star versão 4.5 e coluna cromatográficaC 18 de 5 um, 4,5X250 mm.FASE MÓVEL: solução de metano I e água compH ajustado para 3,0 com ácido fosfórico. Volumede injeção de 30 ~L e a detecção em UV em205 nm.
RESULTADOS E DISCUSSÃO
Cromatografia de Camada Delgada (TLC)Os resultados da produção de GA3 por FES e FSmobtidos por TLC estão resumidos na Tabela 2:
TABELA 2: Resultados da presença de GA3
produzido por fermentação em cromatografia emcamada delgadaCEPA FSm FES casca FES resíduo
extrato de café apósde casca extração
LPB-I +++ ++++ ++++LPB-2 ++++ ++ +++LPB-3 ++ +++ +++LPB-4 + + ++LPB-5 + + ++LPB-6 ++++ ++++ +++++
Cromatografia Líquida de Alta Eficiência(HPLC)Os resultados da quantificação do GA3 produzidopor fermentação da cepa LPB-06, obtidos porHPLC estão apresentados na Figura I, a seguir:
Onde: Maior número de "+" significa melhorfluorescência.Considerando-se que uma melhor fluorescência indicamaior produção de GAJ , percebe-se pejos dados daTabela 2 acima, que o melhor substrato foi o resíduosólido da extração da casca do café. Isso se dá,provavelmente, por existirem substâncias inibidoras dometabolismo de produção das giberelinas nos resíduosde café (casca e extrato), que com o pré-tratamento(hidrólise)são eliminadas.O microrganismo que apresentou melhorprodução foi a cepa LPB-06, para os três sistemasde fermentação. Dessa forma, a quantificação porHPLC foi feita para os fermentados por estemicrorganismo. Para os sistemas em FES, a cepaLPB-OI e LPB-03também apresentaram bons resultados; já paraFSm, a cepa LPB-02 teve uma maior produção.As cepas LPB-04 e LPB-05 apresentaram umabaixa produção de GA3 para todos os substratos.
Analise do metabólito
Cromatografia líquida de alta eficiência(HPLC)SISTEMA: da marca Varian constituído de:bomba quaternária modelo 9012Q; detetor defotodiodo modelo 9065 Polychrom; injetorautomático Rainin modelo AI-200; estação de
TABELA 1: Condições de fermentação
Cromatografia de camada delgada (TLC)FASE MÓVEL: clorofórmio, acetato de etila,ácido acético (40:60:5)
Preparação das amostrasApós a extração e evaporação da amostra, amesma foi diluída em acetato de etila e hexano(I :2). A purificação deu-se por passagem emcoluna com sílica empacotada em pipeta dePasteur. A seguir, passou-se 2,5 mL de metanol nacoluna reservando esta última fração para aanálise. O volume foi evaporado e diluído em lOmL de metanol para HPLC.
Condição FSm FESTemperatura (0C) 29 29pH inicial 4.5 4.5
Razão de inóculo 15% (V/V) 15% (Vim)Umidade inicial (%) - 60Rotação (rpm) 220 -Tempo (dias) 7 dias 7 dias
, . , .

407
Bramorskl, A. Caracterização do crescimento eprodução de metabólitos voláteis por fungosfilamentosos cultivados sobre substratosagroindústriais. Curitiba, 1997. Dissertação (MScTecnologia Química), UFPr .Brahan, J. E.; Bressani, R. Pu\pa de Café composición, tecnologia y utilización. Bogota:Instituto de Nutrición de Centro América yPanama,INCAP, 1978.Brückne, 8.; Blechschmidt, D. The GibberellinFermentation. Criticai Rewiews in Biotechnology,1991, 11(2), 163-192. utilizando-se planejamentoexperimental com análise por superfície deresposta. Com estas otimizações espera-seaumentar a produção de ácido giberélico.esseltine, C. W. Solid State Fermentations.Biotechnology and Bioengineering, 1972, 14,517-532.Kumar, P. K. R. & Lonsane, 8. K. GibberellicAcid by solid state fermentation: consistent andimprovement yields. Biotechnology andBioengeneering, 1987,30,267-27 I.Kumar, P. K. R. & Lonsane, B. K. MicrobialProduction of Gibberellins: state of the art.Advances in Applied Microbiology, 1989,34,29139.Pandey, A. Recents process development in solidstate fermentation. Process Biochemistry, 1992, 7,109-117.Soccol, C. R.. Contribuição ao estudo defermentação no estado sólido em relação àprodução do ácido fumárico, biotransformaçào deresíduo sólido de mandioca por Rhyzopus ebasidiomicetos do gênero Pleurotus. Curitiba,1994. Tese (Professor titular em Biotecnologia eTecnologia de Alimentos) - UFPr.Sturza, R. Aproveitamento biotecnológico dosresíduos da extração do suco da maçã porfermentação no estado sól ido. Curitiba, 1995.Dissertação (MSc Tecnologia Química), UFPr .Tomasini, A.; Fajardo, C.; Barrios-Gonzáles, 1.Gibberellic acid production using different solidstate fermentation systems. Word Journal ofMicrobyology and Biotechnology, 1997,13, 203206.
10
Os autores agradecem ao CNPq, à CAPES e àUnião Européia (projeto INCa DC N o CT970 I85) pelo suporte financeiro recebido.
Com os resultados mostrados acima, percebe-se apossibilidade de utilização da casca de café comosubstrato para produção de GA3 por fermentaçãono estado sólido, desde que feito um prétratamento do substrato.Atualmente estão sendo otimizadas as condiçõesde tratamento do substrato e de fermentaçãoquanto a parâmetros físicos e nutricionais,utilizando-se planejamento experimental comanálise por superfície de resposta. Com estasotimizações espera-se aumentar a produção deácido giberélico.
REFERÊNCIAS BILlOGRÁFICAS
AGRADECIMENTOS
Substrato
//O~--...,....---.....------~·
Produção de GAJ em casca de café
FIGURA 1: Produção de GA3 pela cepa LPB-06,em diferentes sistemas de fermentação com cascade café.
*Os valores de concentração são dados em mgGA/L substrato para FSm e em mg GA/kgsubstrato para FES.
Ayala, L. Aproveitamento biotecnológico debatata refugos (Solanum tuberosum sp.) paraprodução de conídios do fungo entomopatogênicoBeauveria bassiana (Bals) will para fermentaçãoem estado sólido. Curitiba, 1996. Dissertação(MSc Tecnologia Química), UFPr.


409
SCREENING OF FILAMENTOUS FUNGI FOR THE PROOUCTlON OF EXTRACELLULARTANNASE IN SOLID-STATE FERMENTATlON (SSF)
1 2 1*Vau de Lagemaat, J ., Augur, C " and Pyle, O.L .
IBiotechnology and Biochemical Engineering, Department of Food Science and Technology, TheUniversity ofReading, PO Box 226, Whiteknights, Reading. RG6 6AP, United Kingdom
2IRD, Institut de Recherche. pour le Développement. IRD-Mexique, Ciceron 609, Los Morales, 11530Mexico D.F. Mexico
*To whom correspondence should be addressed
ABSTRACTSeven fungai strains (Aspergillus, Fusarium and Penicillium) were screened for the production ofextracellular tannase in a solid-state fermentation system containing 1, 5 or 10 % (w/vJ tannic acid. Themajority of the strains produced more tannase in the 5 and 10 % (w/v) tannic acid fermentations comparedto the 1 % (w/v) tannic acidfermentation. Penicillium frequentans showed the highest tannase activity (1.25
umoles.minlintl ) after 120 hours of fermentation in a 10 % (w/v) tannic acid fermentation whileFusarium moniliforme produced none or ver)' small quantities of the enzyme. The tannase activities of theother strains depended very much on the concentration of tannic acid present in the fermentations.Zymograms showed 1-3 bands ofactivity for each ofthe different strains. Penicillium frequentans gave onedistinct band and has been chosenfor further studies.
1 Filamentous fungi (Aspergillus. p e n tcnnu m , Fusanum)
MATERIALS ANO METHOOS
SSF (6 o e v e . 30 °C) o n PUF soaked in mediumc c n t a tnm ç differentconcenlrations t e n n tc acid
~ .J,. ~
L:h' &' lh'111 " n .c; I d n 1\ 'c I ~ "n IC
;\ C IQ ~ C '0 a C .d
Tannase c h a r a c t e rrs an o nAssay t a n n a s ea c tiv ity
Rarnirez-Martinez (1991) only found 1-2.7 %condensed tannins depending onthe coffee species. Future studies are necessary todetermine whethercoffee pu1p is a suitablesubstrate.Little work has been done on the production oftannase in SSF by different fungal strains. Anintensive screening to select a potent strainsuitable for an SSF system is necessary sinceproduct titers in submerged fermentation areusually different from those in SSF(Shankaranand et aI. 1992). Since tannic acidseems to be the best substrate for the enzyme(Yamada et ai. 1968), a liquid medium containingthis component was soaked in polyurethane foam(pU F) and was chosen as the initial SSF system inthis study. In contrast to coffee pulp, PUF enablesa controlled medium composition to be achieved,it offers monitoring of tannase forrnation, andseparation of the enzyme from the inert carrier iseasy.
Tannin acyl hydrolase (E.C. 3.1.1.20), cornrnonlycalled tannase, hydrolyses ester and depsidebonds in hydrolysable tannins and gallic acidesters (Dykerhoff and Ambruster 1933) and ismainly produced by fungai strains belonging tothe Aspergillus and Penicillium genera (Lekhaand Lonsane 1997). It is claimed that tannaseonly acts on hydrolysable tannins and not oncondensed tannins (Dykerhoff and Ambruster1933). However, the hydrolysis of condensedtannins such as (-)epicatechin gallate and ()epigallocatechin-3-galate has been reported (Lckhaand Lonsane 1997).Commercial tannase is currently produced bysubmerged fermentation (Okamura et ai. 1987)and has found practical uses in the chemical,pharmaceutical and food industries. Studiessuggest that fungai tannase production in sol idstate fermentation (SSF) is advantageous overconventional submerged ferrnentation in terms ofproductivity. the extracellular nature and thermo- andpHstability of lhe enzyme (Lekha and Lonsane 1994).lnterest has occurred in tannase production bySSF with coffee pulp as a potential tannin-richand cheap solid substrate. Data for thecomposition of tannins in coffee pulp arecontradictory. Zuluaga (1981) reported thepresence of 2 % total tannins in sun-dried coffeepulp from which 21 % were hydrolysable and 79% were condensed tannins while Clifford and
INTROOUCTION
Figure 2. Outline of material and melhods.
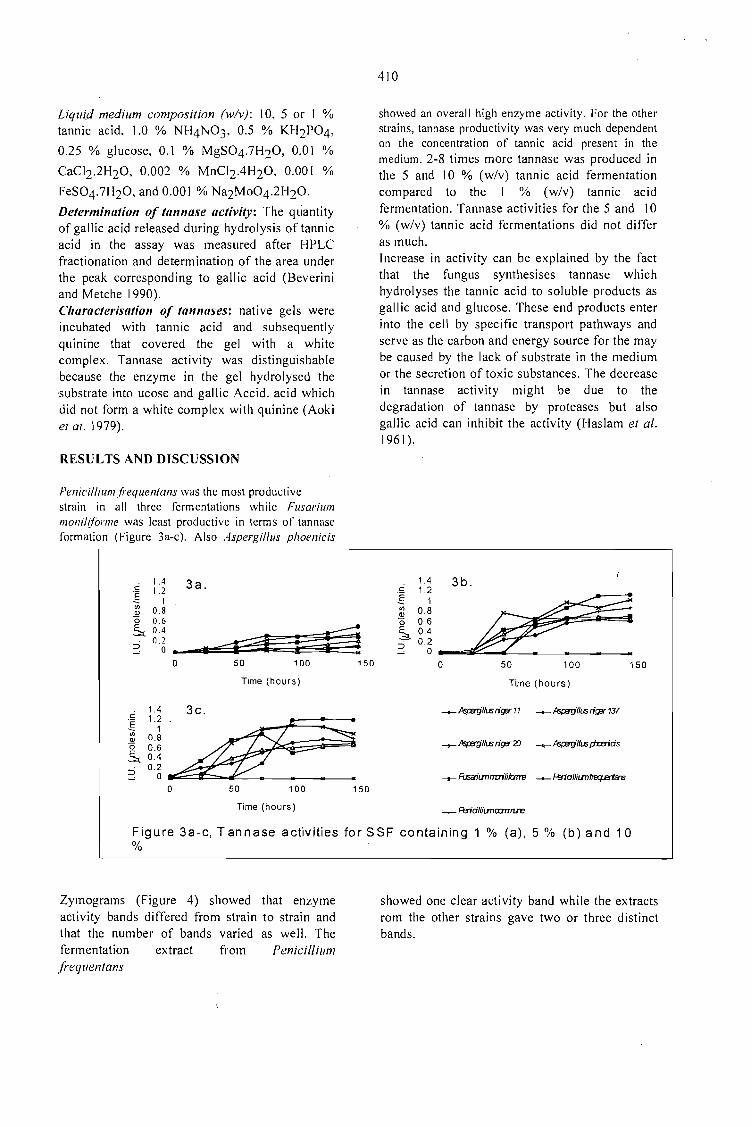
Liquid medi um composition (w/v): la, 5 or 1 %tannic acid, 1.0 % NH4N03, 0.5 % KH2P04,
0.25 % glucose, O.! % MgS04.7H20, 0.01 %
CaCI2.2H20, 0.002 % MnCI2.4H20, 0.00 [ %
FeS04.7H20, and 0.001 % Na2Mo04.2H20.
Determination 01 tannase activity: The quantityof gallic acid re!eased during hydrolysis of tannicacid in the assay was measured after HPLCfractionation and determination of the area underthe peak corresponding to gallic acid (Beveriniand Metche 1990).Characterisation 01 tannases: native gels wereincubated with tannic acid and subsequentlyquinine that covered the gel with a whitecomplex. Tannase activity was distinguishablebecause the enzyme in the gel hydrolysed thesubstrate into ucose and gallic Accid. acid whichdid not form a white complex with quinine (Aokiet aI. 1979).
RESULTS AND DlSCUSSlON
Peniciltiumfrequentans was the most productivestrain in ali thrce fermentaríons while Fusariummoniliforme was least productive in terrns of tannaseforrnation (Figure 3a-c). Also Aspergil/lIS phoenicis
410
showed an overall high enzyme activity. For the otherstrains, tannase productivity was very much dependenton the concentration of tannic acid present in themedium. 2-8 times more tannase was produced inthe 5 and la % (w/v) tannic acid fermentationcompared to the I % (w/v) tannic acidfermentation. Tannase activities for the 5 and 10% (w/v) tannic acid fermentations did not differas much.Increase in activity can be explained by the factthat the fungus synthesises tannase whichhydrolyses the tannic acid to soluble products asgallic acid and glucose. These end products enterinto the cell by specific transport pathways andserve as the carbon and energy source for the maybe caused by the lack of substrate in the medi umOI' the secretion of toxic substances. The decreasein tannase activity might be due to thedegradation of tannase by proteases but alsogaJlic acid can inhibit the activity (Haslam et ai.1961).
1.4
~ 1'7~ 0.8o 0,6!h 0.4
0,2::::J O ..
o
3a.
50 100 150
1.4c 1,2'.ê 1~ 0.8o 0,6:k. 04
::::J 0,2O
O
3b.
50 100 150
Time (hours)
1.4 3 C
}il~O 50 100 150
Time (hours)
Time (hours)
____ !J5{Hçjllus rig3r 11 ~ fJ5{:sg'/Ius rig3r 131
Figure 3a-c, Tannase activities for SSF containing 1 % (a), 5 % (b) and 10%
Zymograms (Figure 4) showed that enzymeactivity bands differed from strain to strain andthat the number of bands varied as well. Thefermentation extract from Penicilliumfrequentans
showed one c1ear activity band while the extractsrorn the other strains gave two or three distinctbands.

41 1
Aniger 137 Aniger 20 P.commune
Anigr 11 I APhOenrS lp.freqUeranS I
Figure 4 , Activity gel with tannases from different fungai strains .
CONCLUSIONS
Penicillium frequ entans produced the highesttiters of extracellul ar tannase. The fermentationextract from th is strain formed a single activityband in the enzyrne characterisation studies whichshould facilirate purifi cation. For those reasonsPenicillium frequentans has been chosen forfurther studies on extrace llular tannase product ionin SSF.
ACKNOWLEDGEMENTS
This study was supported by the European UnionINCO-DC N° IC 18*CT970 185, BIOPULCA
project.
REFERENCES
Aoki, K., T. Tanaka, R. Shink e, and H. Nishira.1979. Detecti on of tannase in polyacrylamidegels. Journal o f Chromatography 170: 446-448 .Beverini. M. and M. Metche. 1990 . Identification.purificat ion and physicochernical propertie s o ftannase of Asperg illus ori zae . Sciences desiments 10: 807-816.Cli fford. M.N. and J.R. Rarnirez-Martinez. 1991.Tannin s in wet-processed coffee beans and coffeepulp. Food Chemi stry 40: 191-200.Dykerhoff, H. and R. Ambru ster. 1933. Tannase.Z. Physiol. Chem. 292 : 38-56.
Haslam, E., R.D. Haworth, K. Jones, and H.J.Rogers. 1961. Gallotannins. Introduction and thefractionation of the tannase. Jou rnal o f theChemical Society 1829-1835.lekha, P.K. and B.K. Lonsane . 1994. Co mparativetitres, location and properti es of tann in acy lhydrolase produced by Asperg illus niger PKL104 in so lid-s tate, liquid sur face and submergedfermentations. Process Biochem istry 29: 497-503 .Lekha, P.K. and B.K. Lonsane. 1997. Product ionand application of tannin acy l hydr olase: State ofthe art. Advances in Applied Microb iology 44 :2 15-260.Okamura, S.. K. Mizusawa, K. Marnor i, K. Takei.Y. lmai. and S. Ito. 1987. Ferrnent ativemanufacture of tannase. Japan patent 62 304 98 1.Shankaranand, V.S., M.V. Ram esh, and B.K.Lonsane. 1992. ldiosyncrasies of solid-statefermentation sys tems in the biosynthe sis ofmetabolit es by some bacteri al and fungai cultures.Process Biochemistry 27 : 33-36 .Yamada. H., O. Adac hi. M. Watanabe, and N.Sato. 1968. Studies on fungai tann ase Part I:Forrna tion. purification and cata lytic propert ies oftannase of Aspergillus fla vus. Agricultura! andBiological Chemistry 32: 1070-1078.Zuluaga. J. 1981. Contribution à I'étude de lacomposition chimique de la pulpe de café (Co ffeaarabica L.) The'se de Doctorat es Scien ce .Université de Neuchatel, Suisse. 93p.


413
PROOUCTION OF FUNGAL TANNASE IN A MOOEL SYSTEM BY SOLlO STATEFERMENTATION
Cristóbal Noé Aguilar,1 Christopher Augur,2 Gustavo Vinlegra-González l & Ernesto FavelaTorres3
1Biotechnology Dept. Universidad Autonoma Metropolitana. Iztapalapa Campus, Mexico. D.F.2l nstitut de Recherche pour le Développement, IRD-Mexico.
Key words: tannase, tannic acid. solid state fertnentation.
ABSTRACTCoffee pulp is considered lhe most abundant waste and pollutant material produced h)' lhe coffee industry.lts utilization as animal feed is limited by presence of antiphysiological factors, such as polyphenoliccompounds, among which, tannic acid present in high concentrations (3-4%). Thus, lhe use of tannaseproducing fungai strains lo degrade tannins might he an alternative lo use coffee pulp as animal feed. Inaddition, tannase production frotn cojJee pulp hy solid state fermentation (SSF) might be an attractivepossibility. The production of tannase by Aspergillus niger Aa-20 lI'as evaluated. Several concentrations oftannic acid (12.5, 25, 50 and 100 g/L) were tested in a mudei systein. Column reactors were packed withpolyurethane foam impregnated with liquid medium and inoculated with spores. Tannase production lI'as
carried out for .:18 h. Tannic acid uptake 1I'as measured hy lhe phenol-sulfuric method and extracellular andintracellular tannase activities were assayed by Hl'Lç-chromatography. Tannase activity increased withincreasing concentration.of tannic acid. Maxintum extracellular and intracellular tannase activitics (11.35and 6.95 U/mL respectively) were recorded with 100 g/L of tannic acid. The substrate uptake was 100% atconcentrations o112. 5. 25 and 50 g/L. while 73.03% 1I'as consumed in lhe presence of I 00 g/L oftannic acidafter 48 h ofculture. These results suggest that high concentrations of tannins can be removed h)' SSF andtannase production can he reached in high titers.
INTROOUCTION
At present, coffee pulp is one of the mostabundant wastes and agro-industrial poIlutantgenerated by the coffee industry. The utilizationof coffee pulp as animal . feed has beeninvestigated for several years (De Rozo el ai ..1985). However. its chemical composition is agreat limiting factor due to the presence ofantiphysiological factors. such as caffeine andpolyphenolic compounds or tannins (Hakil et aI.,1998), among which, tannic acid, present in highconcentrations (3-4%). Tannins are rnoleculesconsidered as water-soluble phenolic cornpoundswith molecular weights ranging frorn 500 to
3000. These are present in several plants wherethey act structurally as pigrnents or as protectorsfrorn the microbial attacks (Lekha and Lonsane,1997). Several proposals to use the coffee pulp asproduct of added value have been made, amongwhich, microbial enzyme production has beenconsidered.Fungai tannase production by SSF using coffeepulp as support might be an attractive possibilitywith two advantages. the remova I in situ ofhydrolizable tannins and enzyme production
itself. Tannase catalyzes the hydrolysis of esterand depside bonds in such hydrolizable tannins astannic acid. Tannase is also used in food andbeverage processing (Coggon et ai .. 1975).A primary characterization of enzyme productionis necessary in order to select better conditionsfor SSF processo In this work, tannase productionand tannic acid degradation by Aspergillus nigerAa-20 was investigated in an SSF based modelsystem. The effects of the tannic acidconcentration on tannase production and itsdegradation were detennined.
MATERIALS ANO METHOOS
Microorganism and culture mediumSpores of the Aspergillus niger Aa-20 strain([RD-UAMI collection) used in this work werestored at -20°C in protect-crioblocks (beadstorage system, Technical Service ConsultantsLimited). Inoculum was prepared by transferingthe spores to PDA médium. incubated at 30°C for5 days, and the spores were suspended into 0.01%Tween 80 solution and counted in a Neubauerchamber.Medium for tannase production-was thesame as reported by Lekha and Lonsane (1994)
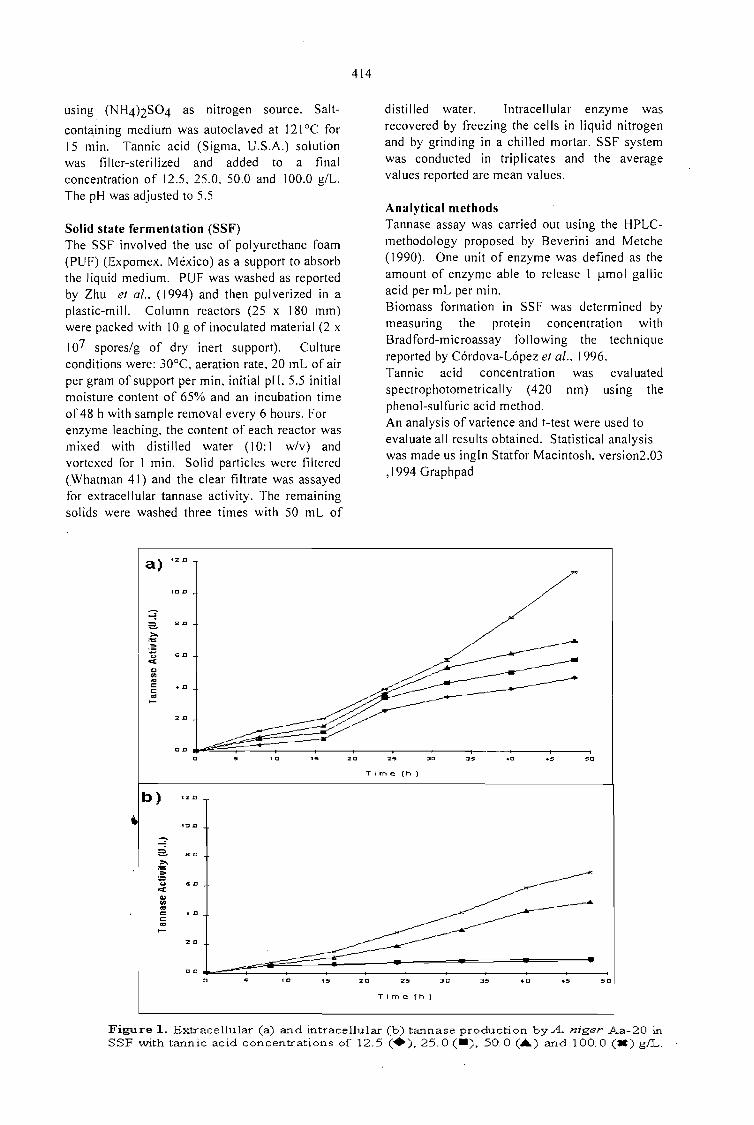
414
using (NH4)2S04 as nitrogen source. Salt
containing mediurn was autoclaved at 121°C for15 mino Tannic acid (Sigma, U.S.A.) solutionwas filter-sterilized and added to a finalconcentration of 12.5, 25.0, 50.0 and 100.0 g/L.The pH was adjusted to 5.5
Solid state fermentation (SSf)The SSF involved the use of polyurethane foam(PUF) (Expomex. México) as a support to absorbthe liquid medium. PUF was washed as reportedby Zhu et al., (1994) and then pulverized in aplastic-mill. Column reactors (25 x 180 mm)were packed with 10 g of inoculated material (2 x
107 spores/g of dry inert support). Culture
conditions were: 30°e, aeration rate, 20 mL of airper gram of support per min, initial pl-I, 5.5 initialmoisture content of 65% and an incubation timeof 48 h with sample remova I every 6 hours. Forenzyme leaching, the content of each reactor wasmixed with distilled water (10:1 w/v) andvortexed for 1 mino Solid particles were filtered(Whatman 41) and the clear filtrate was assayedfor extracellular tannase activity. The remainingsolids were washed three times with 50 ml. of
distilled water. lntracellular enzyrne wasrecovered by freezing the cells in 1iquid nitrogenand by grinding in a chilled rnortar. SSF systernwas conducted in triplicates and the averagevalues reported are mean values.
Analytical methodsTannase assay was carried out using the HPLCmethodology proposed by Beverini and Metche(1990). One unit of enzyrne was defined as the
arnount of enzyme able to release 1 umol gallicacid per mL per minoBiomass formation in SSF was deterrnined byrneasuring the protein concentration withBradford-microassay following the techniquereported by Córdova-López et al.. 1996.Tannic acid concentration was eval uatedspectrophotometrically (420 nm) using thephenol-sulfuric acid method.An analysis of varience and t-test were used toevaluate ali results obtained. Statistical analysiswas made us ingln Statfor Macintosh, version2.03,1994 Graphpad
a) 12 "
10 "
so
.s'lS
'lS:>0
Ti m e (h)
T i me (h)
20 25 30I
'S
,s
'O
."
SO
___________1
O"s
b) '2 "
to10 "
a "">-,..=-:;:;<.> 6"ee......<:: ."<::.......
2"
A" -===O S
Figur~ 1. Extrace llu lar (a) and intracellular Cb) tannase production byA. »ziger: Aa-20 inSSF with tannic acid concentrations of 12.5 C+). 25. O C-), 50. O (Â.) and 100. O C-) glL.

RESllLTS AND DlSCUSSION
Tannase production in SSFTannase production (both intra-cellular andextracellular activity) by A. niger Aa-20 in theSSF process was seriously improved by theincrease of the i initiaI tannic acid concentration.Figure 1a shows the kinetics of extracellulartannase production, while Figure Ib shows theresults obtained by measuring intracellulartannase production. A consistent behavior wasfound, in which the increment in the tannic acidconcentration is proportional to increases intannase activity. In this study a maximum oftannase activity was reached with 100.0 g/L oftannic acid at 48 h. The decrease in tannaseproduction by SSF was not found as reported byLekha and Lonsane (1994). Lekha and Lonsane(1994) and Garcia- Pena (1996) reported thattannase was wholly extracellular during the enti refermentation period as no intracellular enzymewas located in the fungal cells. This studyrevealed that the use of higher tannic acidconcentrations promotes the expression ofintracellular activity; these results might be due tothe cells having a monitoring mechanism of thesubstrate leveis in its environment, therebyinhibiting the excretion process when substrate ispresent in high concentrations. In this case, theextracellular/intracellular ratio was 1.3: I for 50.0g/Land 1.6:1 for 100.Og/L;whilethatto 12.5and25 .0 g/L of tannic acid present the intracellularactivitieswerelow, thus theextracellular/intracellular ratios were superior to6: 1 in both cases.In this study the results of tannase productionobtained in SSF system can be partially explainedby those reported for pectinesterase andpolygalacturonase production by Maldonado andStrasser de Saad (1998). They suggested that thehigh titres of enzymatic production obtained inSSF are due to changes in the composition ofmembrane fatty acids provoked by stressconditions favored by this enzyme productionprocess when the substrate concentration isincreased.
415
Tannic acid degradation in SSFTable I shows a summary of the kineticparameters evaluated in SSF systems. The tannicacid-degrading capacity of A. niger Aa-2ü on SSFprocess was evaluated. This capacity was higherwhen the microorganisms grew on low andmedium concentrations oftannic acid. This aspectis very important in the particular case wheretannase could be used for the removal ofphenoliccompounds OI' tannins from waste water andcoffee pul p.In SSF, a particular behavior of kineticparameters was showed. The product yield (Y p),
specific substrate uptake rate (q s) and specific
product formation rate (qp) were higher to tannic
acid concentration of 100.0 g/L, In addition, thehigher tannase activity (both intracellular andextracellular) and lower uptake substrate percentwas obtained too.The analysis of the values of kinetic parametersobtained in SSF system shows to this process as asystem very attractive to the degradation ofhydrolizable tannins and tannase production.Finally, if a suitable fermentation system isconsidered for the production of tannase, it isimportant to remembel' that some tannaseproduction studies carried out on SmF indicatedthat this system was not meant for the longerfermentation times (Ganga et al., 1978; Pourrat etal., 1982) and the íntracellular nature of theenzyme (Lekha and Lonsane, 1997). Withrespect to tannase production by SSF, resultspresented showed an increase in tannase activitywith respect to tannic acid concentration presentin the medium. In addition, A. niger Aa-20 wasable to degrade high concentrations ofhydrolyzable tannins. These results complernentthose reported by Lekha and Lonsane (1994) andChaterjee et al., (1996), with the testing of amodel systern using polyurethane foam as inertsupport, Thus. SSF should be adopted as tannaseproduction system due to shorter fermentationtime, higher activity titres produced and toleranceto high tannic acid concentrations. The next stepof this work will evaluate the effect oftannase onthe levels of tannins of coffee pulp,
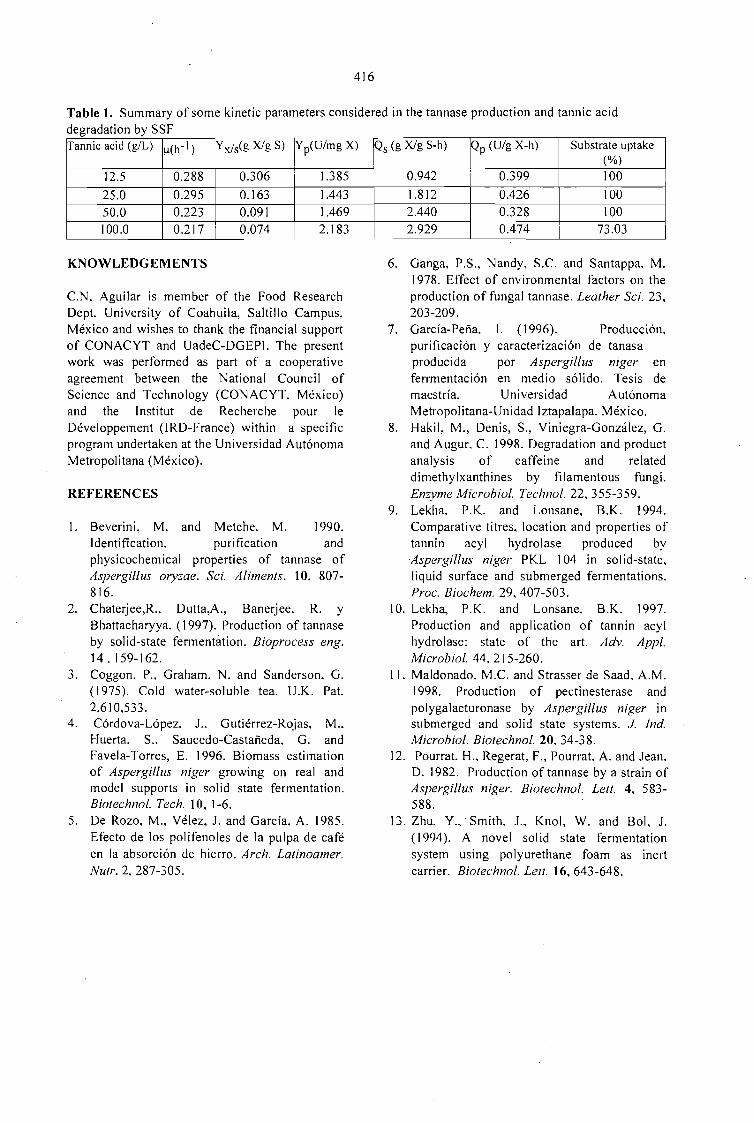
416
Table 1. Summary of some kinetic parameters considered in the tannase production and tannic acid
degradation by SSFTannic acid (g/L) l1(h-l) Yx/s(g X/g S) Yp(U/rng X) IQs (g X/g S-h) Qp (U/g X-h) Substrate uptake
(%)
12.5 0.288 0.306 1.385 0.942 0.399 100
25.0 0.295 0.163 1.443 1.812 0.426 10050.0 0.223 0.091 1.469 2.440 0.328 100100.0 0.217 0.074 2.183 2.929 0.474
I73.03 I
KNOWLEDGEMENTS
e.N. Aguilar is member of the Food ResearchDept. University of Coahuila, Saltillo Campus,
México and wishes to thank the financiai supportof CONACYT and UadeC-DGEPI. The presentwork was performed as part of a cooperative
agreement between the National Council ofScience and Technology (CONACYT. México)
and the Institut de Recherche pour leDéveloppernent (IRD-France) within a specific
program undertaken at the Universidad Autónoma
Metropolitana (México).
REFERENCES
I. Beverini, M. and Metche. M. 1990.Identification. purification andphysicochemical properties of tannase ofAspergillus oryzae. Sei. A/iments. 10, 807816.
2. Chaterjee.R; Dutta,A., Banerjee, R. y
Bhattacharyya. (1997). Production of tannaseby solid-state fermentàtion. Bioprocess eng.14.159-162.
3. Coggon, P., Graharn, N. and Sanderson, G.
(1975). Cold water-soluble tea. U.K. Pato
2.610.533.4. Córdova-López. J.. Gutiérrez-Rojas, M .•
Huerta, S.. Saucedo-Castaüeda, G. andFavela-Torres, E. 1996. Biomass estimationof Aspergillus niger growing on real andmodel supports in solid state fermentation.Biotechnol. Tech. to, 1-6.
5. De Rozo, M.• Vélez, J. and Garcia, A. 1985.Efecto de los polifenoles de la pulpa de café
en la absorción de hierro. Arch. Latinoamer.Nutr. 2. 287-305.
6. Ganga, P.S., Nandy, S.e. and Santappa, M.
1978. Effect of environrnental factors on the
production of fungai tannase. Leather Sei. 23,203-209.
7. Garcia-Pena, 1. (1996). Producción,
purificación y caracterización de tanasaproducida por Aspergillus niger en
ferrmentación en medio sólido. Tesis demaestria, Universidad AutónomaMetropolitana-Unidad Iztapalapa. México.
8. Hakil. M., Denis, S" Viniegra-González, G.and Augur, e. 1998. Degradation and product
analysis of caffeine and related
dimethylxanthines by filamentous fungi.
Enzyme Microbiol. Technol. 22,355-359.9. Lekha, P.K. and Lonsane, B.K. 1994.
Comparative titres, location and properties oftannin acyl hydrolase produced by-Aspergillus niger PKL 104 in solid-state,
liquid surface and submerged fermentations.Proc. Biochem. 29,407-503.
lO. Lekha, P.K. and Lonsane, B.K. 1997.Production and application of tannin acy\hydrolase: state of the art. Adv. Appl.Microbiol. 44, 215-260.
li. Maldonado. M.e. and Strasser de Saad, A.M.
1998. Production of pectinesterase and
polygalacturonase by Aspergillus niger insubmerged and solid state systems. J. Ind.Microbiol. Biotechnol. 20.34-38.
12. Pourrat. H., Regerat, F., Pourrat, A. and Jean,D. 1982. Production of tannase by a strain ofAspergillus niger. Biotechnol. Lelt. 4, 583588.
13. Zhu, Y .• · Smith, 1., Knol, W. and BoI, 1.
(1994). A novel solid state fermentation
system using polyurethane foam as inert
carrier. Biotechnol. Leu. 16. 643-648.



419
QUALIDADE DO CAFÉ SOLÚVEL BRASILEIRO
Paulo Lauro Correa de Oliveira
Empresa: Oliveira &Veiga Consultores AssociadosEndereço: Rua Pistóia, 155- Londrina-86020-450-BrazilE-mail: [email protected] - Phone/Fax:55-43-3223273
l-ESCOPO DA PALESTRA:
Demonstrar a importância do café solúvelbrasileiro no contexto mundial.
2-0BJETIVO DA PALESTRA:
Demonstrar que a qualidade do café solúvelbrasileiro já é reconhecida mundialmente e queessa industria traz ao país benefícios enormes noque se refere à: 1- agregar valor a nossa matériaprima; 2- vender marcas no Exterior( como CaféPelé) o mais conhecido;3-novas tecnologias com conseqüente capacitaçãode mão de obra brasileira para enfrentar a maisrigorosas especificações;4-Introdução demodernas técnicas de Gestão da Qualidade(ISO9001/ HACCP/GMP) e de Gestão Ambiental(ISO1400I) visando ao aumento da competitividadeatravés da redução de desperdícios e retrabalhos.
3-DESENVOLVIMENTO:
A palestra contém 4 etapas a saber:1a -etapa - Introdução ao cenário mundial de
café bem como do cenário do BrasilQuantidades por safra 98/99 e 9912000. Principaisregiões produtivas brasileiras.2a -etapa - Apresentação da Indústria de CafeSolúvel no Brasil. Como e por que começou? Acapacidade industrial instalada..3a -etapa - Os principais mercados destinatáriosdesse produto. Retrospectiva dos últimos 5 anos.Quantidade versus faturamento.4a -etapa - Sistemas de gerenciamento daQualidade adotados pelo Setor do Solúvelbrasileiro como ferramenta de modernização facea um mercado cada vez mais competitivo eglobal.
la-ETAPA:Slidel2- Mostrou que a disponibilidade de caféno mundo durante a safra passada foi deaproximadamente 104 milhões de sacas de 60KG. Mostra o Brasil(33% do total) e demaispaíses produtores de café em todos os
continentes. O Brasil sem dúvida é o maiorprodutor dessa comodity no mundo.Slide13- Mostra que além de maior produtor oBrasil foi também o maior exportador, sempreseguido da Colômbia.Slide 14-Mostra as principais regiões produtorasde café, onde Estado de Minas Gerais contribuiuna safra passada com 57% do volume produzido,ou seja, aproximadamente com 19 milhões desacas , seguido por Espírito Santo, São Paulo eParaná. A safra passada contou comaproximadamente 35 milhões de sacas.Slide15- Mostra que a safra atual terá uma quebraentre 30 a 35% em relação a safra anterior, o quequer dizer aproximadamente 23 milhões de sacas.A distribuição produtiva por estados permaneceráinalterada.
2a-ETAPA:Slidel8- Mostra uma exposição de fatos emotivos pela qual foi criada a indústria e caféSolúvel no Brasil. A capacidade atual instaladaestá ao redor de 100 mil toneladas/ano. Semdúvida somos, também, o maior produtor desolúvel do mundo.Slide 19- Os principais mercados do solúvelbrasileiro- O primeiro foi os EUA, na década de60. O Brasil foi o primeiro país a colocar solúvelna ex- União Soviética, ainda no final da décadade 60. Hoje, os mercados da Rússia, lesteeuropeu e china são os grandes objetivos.Silde20e21- Mostram os principais exportadoresde solúvel do Brasi I nos anos de 1997 e 1998. ACia Cacique de Café Solúvel é o maiorexportador seguida pela Cia Iguaçu de CaféSolúvel e pela Nestlé.
3a-ETAPA:SIides23 e 24 - Mostram os principais destinos equantidades exportadas nos últimos 5 anos peloBrasil. Para a Rússia, EUA, CCE, Japão, etc.Destaque para as quantidades exportadas para aRússia, as quais foram afetadas em 1998, emgrande parte pela crise econômica mundial.Mostra também as quantidades de sacas de caféverde necessários para transformar em solúvel

exportados, em relação ao volume faturado emUSS.
4a- ETAPA:Slide 27 e 28-Além de enfrentar tradicionais efortes concorrentes multinacionais, tarifasalfandegárias diferenciadas, uma política internaque não contempla esse produto e face a ummercado cada vez mais global como manter-secompetitivo? Além dos grandes investimentostecnológicos nem sempre viáveis, comoimplantar e implementar técnicas que visem acapacitação de amo de obra para redução dosdesperdícios? Por onde iniciar esses programasvisando manter esse parque índustrial atualizadocom o mercado internacional?Slide 29 e 30 - A experiência da Oliveira & Veigaconsultores associados, nesse setor é comprovadanão somente pela implementação, mas pelaintegração de técnicas e ferramentas numprograma de Gestão Global entreQualidade(IS0900 I,ISO guide 25, HACCP,GMP, TPM, 5S,Cartão Vermelho, etc.) eAmbiental (ISO 14001, Atuação Responsável,Desenvolvimento Sustentável).executado na Cia Cacique de Café Solúveldurante vários anos.Slide 31, 32 e 33 - Mostra um exemplo de um aplanta de café solúvel onde podemos visualizar,definir, identificar, estabelecer quais técnicas eferramentas a adotar e integrar' visando o"negócio" da empresa, como um todo.Por exemplo, identificamos pontos de GMP(setasem azul) como também pontos de HACCP( emvermelho).Slide 34, 35 e 36 - Mostra o Sistema deGerenciamento Ambiental pela qual o Setor doSolúvel brasileiro já tem muita experiência,inclusive com patente na área de resíduoslíquidos. Destaque para o fundamento da gestãoAmbiental o qual está embasado naIdentificação de Todos os Aspectos e ImpactosAmbientais do negócio da empresa, ou seja, aplanta de café solúvel as quais em sua maioria, sesituam em zonas urbanas de grande densidade.Como resultado de ações já implementadas, a
420
algumas fabricas já conseguem "zero emissões"ao meio ambiente, ou seja:a)Resíduos sólidos - Queima total em caldeiraseconomia de combustível.b)Resíduos líquidos - Infiltração no solo composterior monitoração.c)Resíduos gasosos - Filtros e lavadores de gases,bem como otimização de outras fontesenergéticas, ou seja, ao invés de óleo, gás GLP,ou gás natural para "mitigar" a emissão de SOx eNOx para atmosfera.Slide 37-42 - Mostra que o parque industrial decafé solúvel brasileiro dispõe de laboratóriosequipados com modernos instrumentos paraatender qualquer especificação, ou seja, fisico,química, sensorial e microbiológica.Slide 43 - A capacitação dos ..laboratóriosbrasileiros da industria de café solúvel provémem parte da participação dos mesmos no ComitêTécnico da ISSO o TC 34- SC 15- que é o SubComitê para Café, pois é o Fórum de debates parao estabelecimento de metodologias que facilitamo comércio internacional do café, alinhando aspartes interessadas. Slide 44 - 48 - Desse modo oBrasil, na questão de padronização, demetodologias aplicadas a café verde, torrado esolúvel se mantém atento, através doestabelecimento de parcerias com Universidadese Institutos Oficiais, para a execução de projetoscientíficos e tecnológicos, como é o caso daUnicamp, Instituto de química de São Carlos USP, Embrapa, Instituto Adolfo Lutz. !tal, etc.
4-CONCLUSÃO FINAL:
Qualidade é o atendimentos aos requisitosespecificados. A indústria de café solúvelbrasileiro tem ao longo de seus 35 anos deexistência atingindo esse objetivo. A contínuaimplementação de modernas tecnologias e detécnicas de Gestão (Qualidade e Ambiente) seráum fator diferencial de competição para asempresas brasileiras. Atualmente, podemos dizerque de posse de todos esses conceitosimplementados, no dia a dia, o Brasil já possueum café quase que ecologicamente correto.

421
INFLUÊNCIA DA VARIEDADE BOTÂNICA SOBRE A QUALIDADE ORGANOLÉPTICA DOCAFÉ ESPRESSO
Marino Petracco
II1ycaffe S.p.A. via Flavia 110 - Trieste (Italy)
SUMMARYEspresso is a way to enjoy co./fee that is gaining larger and larger popularity worldwide. The cup producedby this brewing method contains a beverage with a high concentration of sensorially active substances:special care must therefore be devoted to prevent and to eliminate defects from the raw material. Moreover,attention should be paid to the variables tending to a./fect the sensory balance appreciated by consumers.Little is known about their injluence on Espresso beverage quality, and genetic parameters are oftenneglected to the benefit oflocal traditional cultivars.Co./fee improvement programs relying on hybridisation did focus so far essentially on agronomic traits likeincreased yield and resistance to diseases. Strong market demand for quality co./fee has now directedattention to breed for new varieties with enhanced aroma and taste. The experimental work outlined in thispaper describes how a tool for sensory evaluation ofbreeding products must consider each ofthe five humansenses which, with the possible exception ofhearing, are involved in the appreciation ofan Espresso cup.Resulting figures indicate that the botanical origin ofCo./fea arabica plants is indeed important to get thebest industrial outcome in the production ofcoffee beans appropriate for Espresso preparation.
INTRODUÇÃO
o Espresso representa uma das maneiras paratomar uma xícara de café que nos dias atuais estáa ganhar uma grande popularidade no mundointeiro. As raízes desta popularidade podem serencontradas na cultura italiana das comidas ebebidas, onde desenrolou-se um estilo de vidatípico estreitamente ligado à bebida de café [I].As principais características do Espresso são:a) preparação extemporânea, expressamenterequisitada pelo consumidor;b) um método específico de infusão utilizandouma pressão da água muito alta;extração rápida, para que só o material melhorpossa ser introduzido na xícara.A bebida obtida com o acima especificadotratamento, resulta ser muito característicatambém se considerada da perspectiva fisica equímica [2]. Todavia, as particularidades maissignificativas do Espresso são constituídas pelanatureza sensorial delas. Todos os sentidos dohomem, exceto o ouvido, são utilizados nadegustação duma xícara de Espresso:• a visão avalia o aspecto da espuma, examinandoa cor como a consistência e persistência dela;• tato avalia a sensação da bebida no interior daboca conhecida também como "corpo", ou seja apropriedade ligada à densidade e à viscosidade;• gosto estima o balanceamento amargura/acideze a presença de um retrogosto doce-caramelo;• olfato prevê a fragrância, por meio da inalaçãodos vapores originários da xícara,e flavor, ou sejaa percepção nasal das substâncias voláteisdesenvolvidas no interior da boca.
o método de extração do Espresso produz naxícara uma alta concentração de substânciassensorialmente ativas: por isso, é preciso ter muitocuidado no que diz respeito à prevenção e àeliminação dos defeitos provenientes do café cru.Além disso, para puderem oferecer aosconsumidores a qualidade sensorial mais fina, épreciso ter cuidado com as diferentes variáveisque influenciam o equilíbrio sensorial que osconsumidores apreciam.E conhecimento comum que fatores como aregião de produção e a altitude, bem como asituação climática e a técnica de processamento,podem afectar a qualidade do café na globalidadedela, sobretudo respeito ao incremento da acidez edo aroma. Isto foi demonstrado, em ambientesselecionados, pelos dados experimentais [3].Infelizmente, sabe-se pouco em relação àsinfluências deles sobre à bebida Espresso.Os parâmetros genéticos foram durante muitotempo transcurados ao beneficio das tradicionaiscultivares locais e, ainda mais, os programas demelhoramento do café que se fundavam sobre areprodução, tinham o hábito, com poucasexceções [4, 5, 6], de se focalizar principalmentenas características agronômicas tal como pode serum aumento da produtividade e a resistência apragas e doenças.No presente estudo tomamos em consideração aimportância da influência sobre a qualidade doEspresso da variabilidade genética, constituídapelas diferentes características de plantas de caféque se desenvolveram juntas nas mesmascondições geográficas e utilizando sempre omesmo processamento.

422
MATERIAIS E MÉTODOS
I CA: acrônimo por Caturra
resistentes e altamente produtivas, às quais foidado o nome de Icatu.Pelo fato de não termos descoberto durante aprova da xícara preliminar nenhum caráter típicodo café Robusta, concentramos a nossa atenção naseleção de linhagens destas variedadesprometedoras do ponto de vista agronômico, quefossem também produtoras de bebida superior.Os métodos de processamento agrícola queutilizamos foram aqueles típicos do lugar: plantionão sombreado semi-adensado com 1666 plantaspor hectare, e mais algumas operações orientadasà qualidade, tal como uma adequada fertilização,uma regular vaporização de pesticidas, controledo mato, colheita manual, despolpamento,processamento da cereja mediante fermentação.Além das avaliações químicas e agronômicas,cada planta adulta individual foi avaliada pelaqualidade Espresso.A colheita das plantas uma por uma foi levada acabo manualmente, com muito cuidado para evitara presença de cerejas não maduras, madurasdemais ou estragadas. A safra de cada plantaindividual, com cerca de 3 kg de cerejas, foidespolpada mecanicamente. Os grãos resultantes,com o pergaminho mucilaginoso, acabaram oprocesso de fermentação no interior de baldinhosindividuais, pela ajuda de um enzima pectólita(Isozym) em maneira tal de estandardizar operíodo de fermentação entre todas as amostras.Depois da secagem e do descascagem, asamostras' de café ouro foram para o laboratório daillycaffê em Trieste (Itália), onde se desenrolaramas anál ises organolépticas.As amostras foram processadas de acordo com osmétodos ideados para a avaliação de amostrascomerciais [10] no laboratório da lllycaffê:os defeitos visíveis (grãos pretos, quebrados,imaturos) foram eliminados manualmente cadaamostra foi torrada à maneira italiana, utilizando
. 80 g por amostra seguindo condições controladasem equipamento de laboratório para torrefação(Probat) e moagem até à granulosidade idealmediante moinhos profissionais (Faema) cadaamostra, torrada e moída, foi avaliada em trêsxícaras diferentes, segundo três técnicas depreparação específicas:• Espresso: a clássica xícara de Espresso,preparada mediante condições estandardizadas ecuidadosamente controladas [I I]. A bebidaderivante é constituída por uma solução decafeína. açucares e ácidos, na qual está dispersauma emulsão: fase distinta composta de gotinhasmicronicas de lipídios.A concentração total poderia ser tal como 60 g/1.• infusão, uma técnica amplamente utilizada noNorte da Europa e nos E.U.A.: deita-se água
Junto com cultivares tradicionais bem conhecidas,foram escolhidas duas variedades híbridasderivantes de cruzamentos inter-específicos:Catimor e Icatu. Isto foi feito pelo interesse deexplorar a qualidade em xícara de plantas quecontêm algumas características herdadas daespécie Robusta, e que as tornam particularmenteinteressantes aos olhos dos técnicos do café graçasà presença de vantagens agronômicas,introduzidas pelo processo de hibridação, quemerecem atenção.Está amplamente aceitado o fato de o nomeRobusta ser atribuído à variedade mais cultivadade Coffea canephora pela resistência dela emambientes tropicais úmidos. Esta variedadedemonstrou uma pronunciada imunidade no quediz respeito quer à ferrugem das folhas (Hemileiavastatrix) [8] quer aos nematóides das raízes(Meloidogyne spp.) [9].Os produtores de café tentaram, durante muitotempo, o cruzamento da diplóide Robusta com atetraplóide Arábica, obtendo, na maioria doscasos, plantas estéreis.Uma exceção foi alcançada da natureza com oespontâneo híbrido de Timor, o qual pareceu terorigem dum cruzamento na selva pela polinizaçãoentre uma planta Arábica e um raro gameta deRobusta não reduzido. Mais cruzamentos com avariedade Caturra levaram às progênies,resistentes à ferrugem, que têm vários nomesconforme o país: Catimor no Brasil e na AméricaCentral, Colombia em Colombia, Ruiru I I noQuênia.Um distinto experimento foi obtido com sucessono Brasil, onde uma planta de Robusta tetraplóidefoi produzida graças à duplicação doscromossomos induzida com colquicina. Ocruzamento dela com a variedade da ArábicaMundo Novo tem produzido diversas linhagens
Cerca de 20.000 (vinte mil) mudas de Coffeaarabica identificadas individualmente foramplantadas e cultivadas numa boa região deprodução de café, situada no Brasil a cerca de 21°Sul, 1040 rn de altitude, com uma temperaturamédia que varia entre os 20°C e os 26.4°C [7]Uma possível regra de ajuntamento parapudermos descrever a população de café acimamencionada seria de as reunirem segundo aprópria variedade botânica de origem:
CR: " Catimor
CTR: " Catuai Vermelho
CTY: " Catuai Amarelo
IC: " Icatu. .
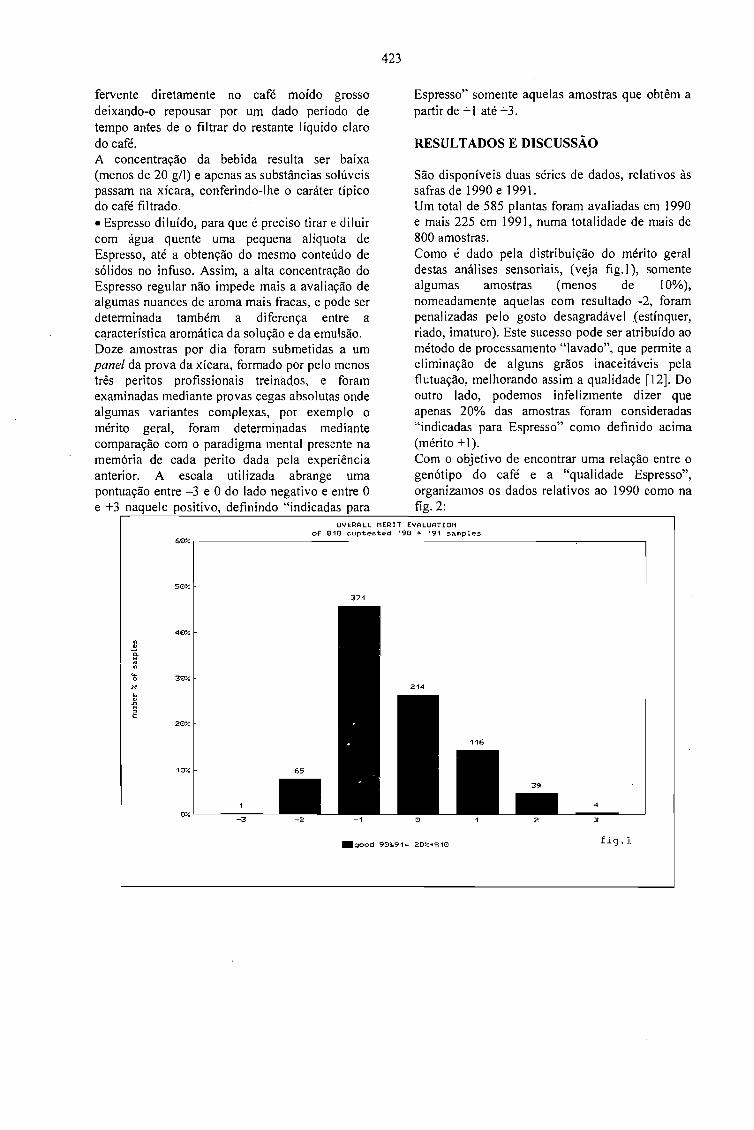
fervente diretamente no café moído grossodeixando-o repousar por um dado período detempo antes de o filtrar do restante líquido clarodo café.A concentração da bebida resulta ser baixa(menos de 20 g/I) e apenas as substâncias solúveispassam na xícara, conferindo-lhe o caráter típicodo café filtrado.• Espresso diluído, para que é preciso tirar e diluircom água quente uma pequena alíquota deEspresso, até a obtenção do mesmo conteúdo desólidos no infuso. Assim, a alta concentração doEspresso regular não impede mais a avaliação dealgumas nuances de aroma mais fracas, e pode serdeterminada também a diferença entre acaracterística aromática da solução e da emulsão.Doze amostras por dia foram submetidas a umpanel da prova da xícara, formado por pelo menostrês peritos profissionais treinados, e foramexaminadas mediante provas cegas absolutas ondealgumas variantes complexas, por exemplo omérito geral, foram determinadas mediantecomparação com o paradigma mental presente namemória de cada perito dada pela experiênciaanterior. A escala utilizada abrange umapontuação entre -3 e Odo lado negativo e entre Oe +3 naquele positivo, definindo "indicadas para
423
Espresso" somente aquelas amostras que obtêm apartir de +1 até +3.
RESULTADOS E DISCUSSÃO
São disponíveis duas séries de dados, relativos àssafras de 1990 e 1991.Um total de 585 plantas foram avaliadas em 1990e mais 225 em 1991, numa total idade de mais de800 amostras.Como é dado pela distribuição do mérito geraldestas anál ises sensoriais, (veja fig.I), somentealgumas amostras (menos de 10%),nomeadamente aquelas com resultado -2, forampenalizadas pelo gosto desagradável (estínquer,riado, imaturo). Este sucesso pode ser atribuído aométodo de processamento "lavado", que permite ael iminação de alguns grãos inaceitáveis pelatlutuação, melhorando assim a qualidade [12]. Dooutro lado, podemos infelizmente dizer queapenas 20% das amostras foram consideradas"indicadas para Espresso" como definido acima(mérito + I).Com o objetivo de encontrar uma relação entre ogenótipo do café e a "qualidade Espresso",organizamos os dados relativos ao 1990 como nafig.2:
OVERALL MERIT EVALUATIONOr 810 cuptested '90 • '91 samples
58%371
48%
'"..õ..e..'"... 30%o
'"~.ce
"c20%
.good 90B.91= 20%"810
4
3
fig.l
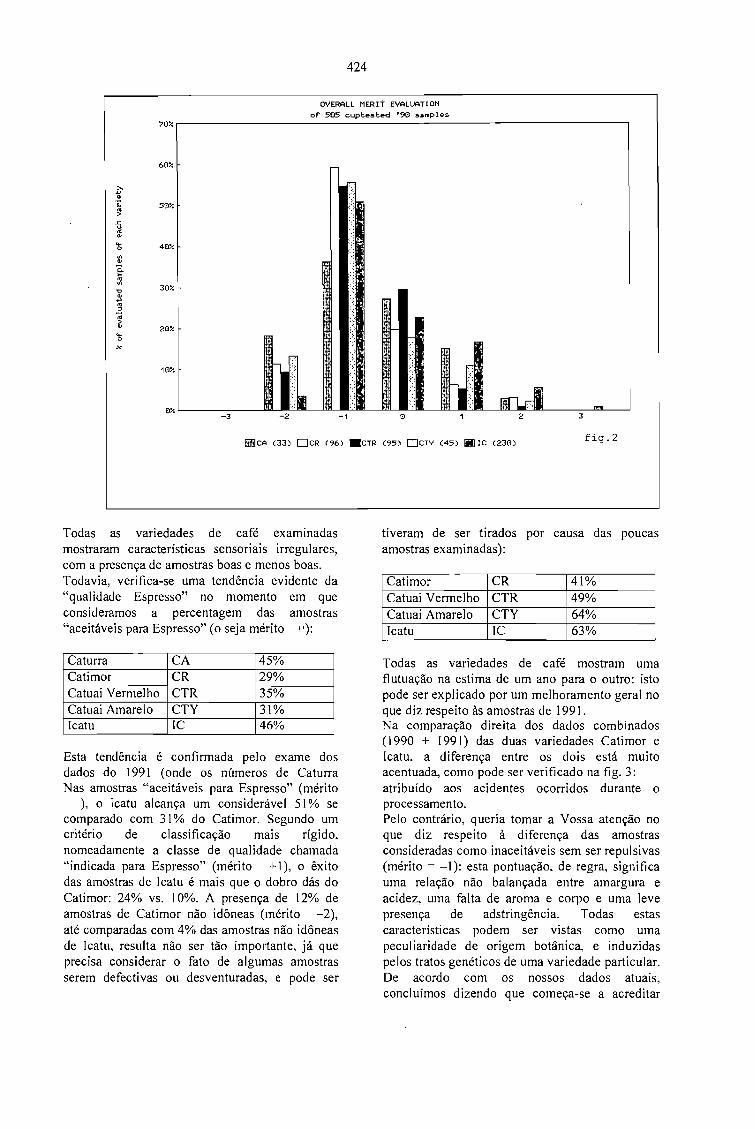
424
OVERALL MERI+ EVALUATIOH
oF 585 cuptested '90 samples70%
50%
48%
30"
20"
10"
,
I I 1 ~-3 -2 -1 2 3
IIll!IlCA (33) OCR (%) .CTR (95) DCTY (45) IiIlIC (230)fig.2
Todas as variedades de café examinadasmostraram características sensoriais irregulares,com a presença de amostras boas e menos boas.Todavia, verifica-se uma tendência evidente da"qualidade Espresso" no momento em queconsideramos a percentagem das amostras"aceitáveis para Espresso" (o seja mérito .-):
Caturra CA 45%Catimor CR 29%Catuai Vermelho CTR 35%Catuai Amarelo CTY 31%Icatu IC 46%
Esta tendência é confirmada pelo exame dosdados do 1991 (onde os números de CaturraNas amostras "aceitáveis para Espresso" (mérito
), o Icatu alcança um considerável 51% secomparado com 31% do Catimor. Segundo umcritério de classificação mais rígido.nomeadamente a classe de qualidade chamada"indicada para Espresso" (mérito +1), o êxitodas amostras de 1catu é mais que o dobro dás doCatimor: 24% vs. 10%. A presença de 12% deamostras de Catimor não idôneas (mérito -2),até comparadas com 4% das amostras não idôneasde Icatu, resulta não ser tão importante, já queprecisa considerar o fato de algumas amostrasserem defectivas ou desventuradas, e pode ser
tiveram de ser tirados por causa das poucasamostras examinadas):
Catimor CR 41%Catuai Vermelho CTR 49%Catuai Amarelo CTY 64%Icatu IC 63%
Todas as variedades de café mostram umaflutuação na estima de um ano para o outro: istopode ser explicado por um melhoramento geral noque diz respeito às amostras de 1991.Na comparação direita dos dados combinados(1990 + 1991) das duas variedades Catimor elcatu, a diferença entre os dois está muitoacentuada, como pode ser verificado na figo 3:atribuído aos acidentes ocorridos durante oprocessamento.Pelo contrário, queria tomar a Vossa atenção noque diz respeito à diferença das amostrasconsideradas como inaceitáveis sem ser repulsivas(mérito = -\): esta pontuação. de regra, significauma relação não balançada entre amargura eacidez. uma falta de aroma e corpo e uma levepresença de adstringência. Todas estascaracterísticas podem ser vistas como umapeculiaridade de origem botânica, e induzidaspelos tratos genéticos de uma variedade particular.De acordo com os nossos dados atuais,concluímos dizendo que começa-se a acreditar
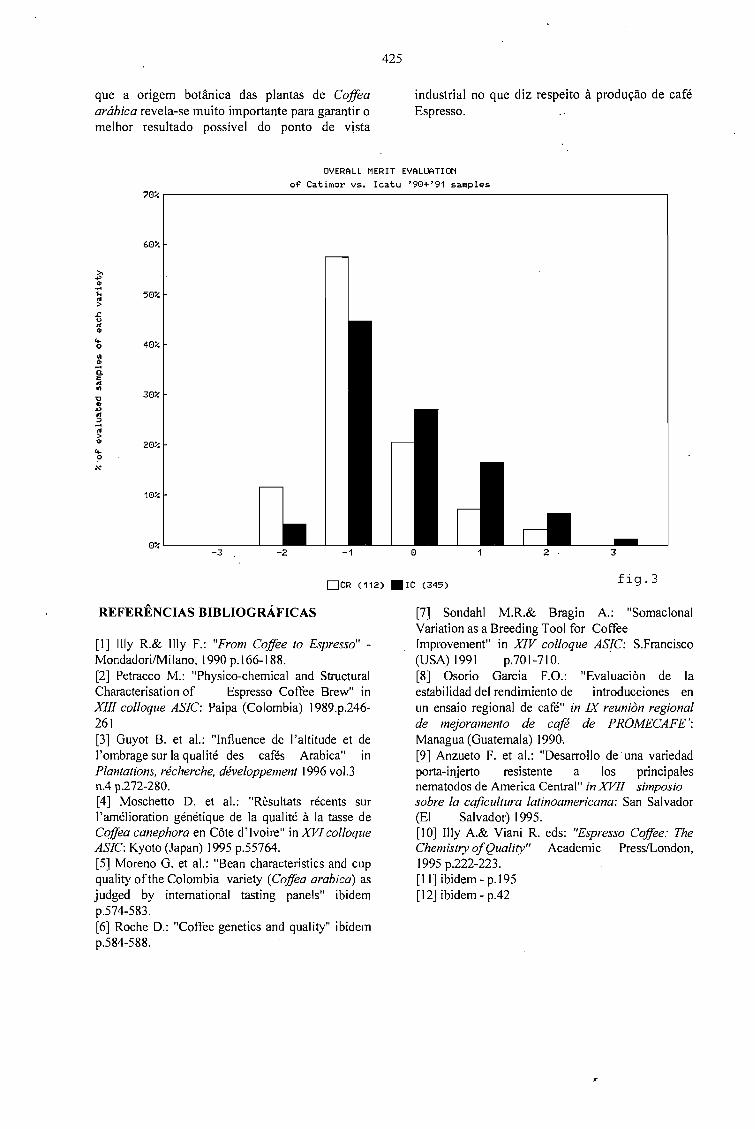
que a origem botânica das plantas de Coffeaarábica revela-se muito importante para garantir omelhor resultado possível do ponto de vista
425
industrial no que diz respeito à produção de caféEspresso.
OVERALL MERIT EVALUATIONof Catimor vs. Icatu '98+'91 samples
78%,...---------------------------------------,
8% L- ---'--_
68%
>-...,QI.. 58%'":>
J::o
'"QI
4- 48%ovlQI
a.E
'"vl
u 38%QI...,'";)'":>QI
28%4-o.'<
18%
-3 -2 -1 8 3
OCR (112) • IC (345) fig.3
REFERÊNCIAS BIBLIOGRÁFICAS
[I] Hly R.& 111y F.: "From Coffee to Espresso" MondadorilMi lano, 1990 p.166-188.[2] Petracco M.: "Physico-chernical and StructuralCharacterisation of Espresso Coffee Brew" inXIII colloque ASIC: Paipa (Colombia) 1989.p.246261[3] Guyot B. et aI.: "Influence de I'altitude et del'ombrage sur la qualité des cafés Arabica" inPlantations, récherche, développement 1996 vol.3nA p.272-280.[4] Moschetto D. et al.: "Rêsultats récents surl'amélioration génétique de la qualité à la tasse deCoffea canephora en Côte d'lvoire" in XVI colloqueASIC: Kyoto (Japan) 1995 p.55764.[5] Moreno G. et al.: "Bean characteristics and cupquality of'the Colombia variety (Coffea arabica) asjudged by intemational tasting panels'' ibidemp.574-583.[6] Roche D.: "Coffee genetics and quality" ibidemp.584-588.
[7] Sondahl M.R.& Bragin A.: "SomaclonalVariation as a Breeding Tool for CoffeeImprovement" in XIV colloque ASIC: S.Francisco(USA) 1991 p.701-710.[8] Osorio Garcia F.O.: "Evaluaciõn de laestabilidad dei rendimiento de introducciones enun ensaio regional de café" in IX reuniõn regionalde mejoramento de café de PROMECAFE':Managua (Guatemala) 1990,[9] Anzueto F. et aI.: "Desarrollo de una variedadporta-injerto resistente a los principalesnematodos de America Central" in XVII simposiosobre la cajicultura latinoamericana: San Salvador(EI Salvador) 1995.[10] Il1y A.& Viani R. eds: "Espresso Coffee: TheChemistry ofQuality" Academic Press/London,1995 p.222-223.[11] ibidem-p.195[12] ibidem - p.42
,.


427
A QUALIDADE DO CAFÉ PRODUZIDO EM DIVERSAS REGIÕES DO BRASIL
José Guilherme Cortez
Divisão de Desenvolvimento Rural Ministério da Agricultura Av. Heitor Penteado, 214513087-000
CAMPINAS- SP
INTRODUÇÃO
A cafeicultura brasileira apresentou uma sensívelrenovação a partir da década de 60, através daintrodução de variedades mais produtivas daSeção de Genética do Instituto Agronômico deCampinas (IAC), ao lado das adubações químicase do apoio creditício e orientações técnicas doInstituto Brasileiro do Café. Embora a maioratenção da nova cafeicultura brasileira fossedirecionada à maior produtividade, asrecomendações sobre a qualidade traduziam-sepelos maiores cuidados com a colheita, como acolheita no pano, a separação dos cafés devarrição e ao uso do sistema de preparo viadespolpamento, em regiões com problemas debebida.A abertura de novas áreas para a cafeicultura, sobcritérios de expansão geográfica ou para fugir dasgeadas, trouxe à tona alguns novos conceitossobre a qualidade do café brasileiro,especialmente aquele produzido em regiões declima seco e frio durante a fase de colheita eprocessamento, como no Cerrado Mineiro(regiões do Triângulo Mineiro / Alto Paranaíba).Sob estas condições de clima, os ciclosfenológicos do cafeeiro eram mais lentos eregulares, a colheita mecânica era favorecida, osprocessos ferrnentativos ' não eram estimuladosdurante a secagem dos grãos e as característicasorganoléticas do produto eram bastante indicadaspara modernos sistemas de preparo da bebida,como o "café expresso".Os processos fermentativos foram definidos comoa ação prolongada de microrganismos, comobactérias, fungos e leveduras dos gênerosKlebsiella Aspergillus, Fusarium eSaccharomyces spp. Onde as condições de climaou tecnologia são favoráveis para odesenvolvimento microbiano, é promovida umadegradação da mucilagem e a produção de álcooise ácidos; se o processo fermentativo forinterrompido após o consumo da camadamucilaginosa, o grão não é atacado e a qualidadeda bebida é preservada; isto é observado quandose retira o café dos tanques de fermentação
(produção de café despolpado) ou quando aumidade do grão é levada a um ponto em que nãoocorre desenvolvimento microbiano (secagem dogrão). Ao contrário, quando o processofermentativo não é interrompido, as camadascelulares da semente são degradadas, diversoscompostos químicos são alterados e observa-seuma alteração da bebida (formação do gosto Rio)ou a ocorrência de grãos defeituosos (defeitospretos e ardidos).A melhor compreensão dos processosfermentativos que o grão de café está sujeito,durante as fases de maturação, colheita eprocessamento, levou ao desenvolvimento de umnovo sistema de preparo do café, conhecido como"cereja-descascado". Nesse processo, combina-sea colheita por sistema de derriça (mecanizada oumanual) com a separação das diversas fases dematuração do café colhido (verdes ou imaturos,cerejas ou maduros e secos ou sobremaduros);após a retirada da casca (exocarpo) dos grãosmaduros, estes são colocados para secar emcamadas finas e as diferentes fases são secadas ecomercializadas isoladamente. Com isso,diminui-se a ocorrência de grãos imperfeitos oufermentados e observa-se uma nítida mel hora naqualidade do café produzido, mesmo em regiõesde clima desfavorável para a bebida.A generalização dos conceitos de qualidade docafé brasileiro, em função do clima, da tecnologiaapl icada e das características sensoriais obtidas,leva agora ao estabelecimento de "marcas" oucertificados de origem, sustentando umacampanha publicitária eficiente e atendendo osdiversos sistemas de preparo de bebida e desejosdos consumidores. Nesse contexto, são incluídostambém a ampliação e diversificação do mercadointerno brasileiro e a utilização mais difundidados cafés robustas produzidos no Estado doEspírito Santo, Sul da Bahia e Rondônia.Nesse trabalho, são apresentados os resultados deanálises físicas e sensoriais de amostrasrepresentativas das diferentes regiões e sistemas deprodução e processamento agrícola dos cafés doBrasil, com comentários sobre suas possíveisindicações para formas de consumo, dentro de umaintegração mais ampla dentro do agronegócio café.
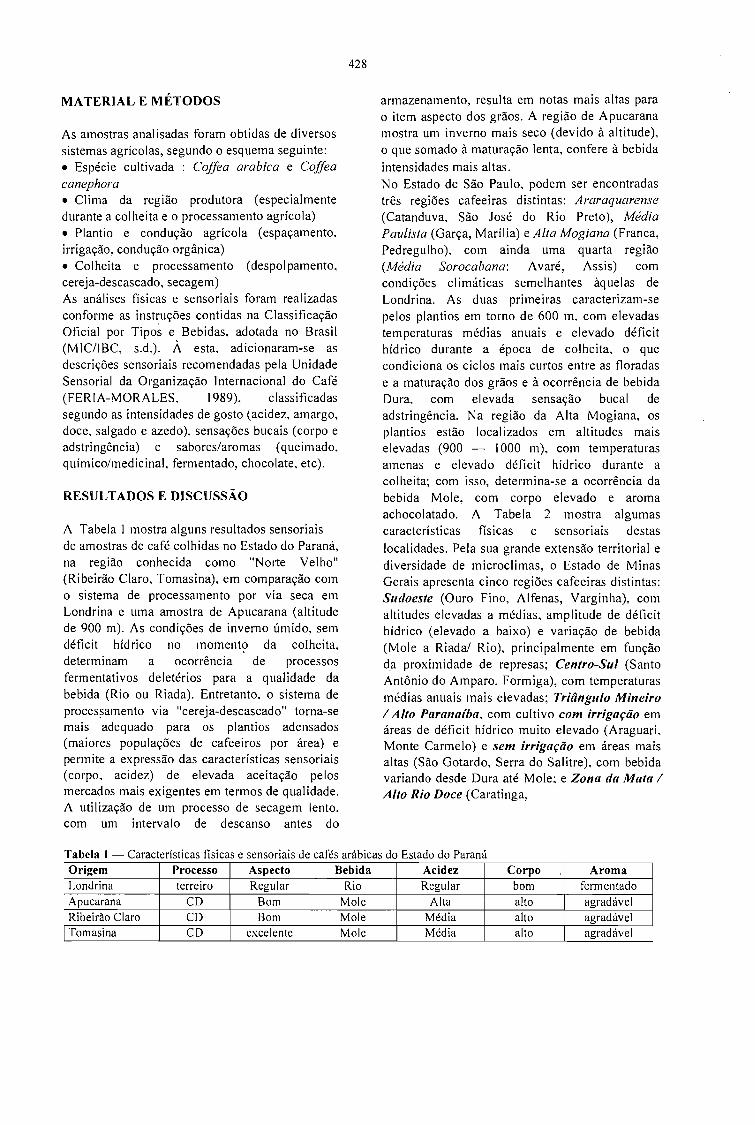
MATERIAL E MÉTODOS
As amostras analisadas foram obtidas de diversossistemas agrícolas, segundo o esquema seguinte:• Espécie cultivada : CojJea arabica e CojJeacanephora• CIima da região produtora (especialmentedurante a colheita e o processamento agrícola)• Plantio e condução agrícola (espaçamento.irrigação, condução orgânica)• Colheita e processamento (despol parnento,cereja-descascado, secagem)As análises físicas e sensoriais foram realizadasconforme as instruções contidas na ClassificaçãoOficial por Tipos e Bebidas, adotada no Brasil(MIC/IBC, s.d.). À esta, adicionaram-se asdescrições sensoriais recomendadas pela UnidadeSensorial da Organização Internacional do Café(FERIA-MORALES. 1989), classificadassegundo as intensidades de gosto (acidez, amargo,doce, salgado e azedo). sensações bucais (corpo eadstringência) e sabores/aromas (queimado.químico/medicinal, fermentado, chocolate, etc).
RESULTADOS E DISCUSSÃO
A Tabela I mostra alguns resultados sensoriaisde amostras de café colhidas no Estado do Paraná,na regiao conhecida como "Norte Velho"(Ribeirão Claro, Tomasina), em comparação como sistema de processamento por via seca emLondrina e uma amostra de Apucarana (altitudede 900 m). As condições de inverno úmido, semdéficit hídrico no momento da colheita,determinam a ocorrência de processosfermentativos deletérios para a qualidade dabebida (Rio ou Riada). Entretanto, o sistema deprocessamento via "cereja-descascado" torna-semais adequado para os plantios adensados(maiores populações de cafeeiros por área) epermite a expressão das características sensoriais(corpo, acidez) de elevada aceitação pelosmercados mais exigentes em termos de qualidade.A utilização de um processo de secagem lento,com um intervalo de descanso antes do
428
armazenamento, resulta em notas mais altas parao item aspecto dos grãos. A região de Apucaranamostra um inverno mais seco (devido à altitude),o que somado à maturação lenta, confere à bebida
intensidades mais altas.No Estado de São Paulo, podem ser encontradastrês regiões cafeeiras distintas: Araraquarense(Catanduva, São José do Rio Preto), MédiaPaulista (Garça, Marília) e Alta Mogiana (Franca,Pedregulho), com ainda uma quarta região(Média Sorocabana: Avaré, Assis) comcondições climáticas semelhantes àquelas deLondrina. As duas primeiras caracterizam-sepelos plantios em torno de 600 m, com elevadastemperaturas médias anuais e elevado déficithídrico durante a época de colheita, o quecondiciona os ciclos mais curtos entre as floradase a maturação dos grãos e à ocorrência de bebidaDura, com elevada sensação bucal deadstringência. Na região da Alta Mogiana, osplantios estão localizados em altitudes maiselevadas (900 - 1000 m), com temperaturasamenas e elevado déficit hídrico durante acolheita; com isso, determina-se a ocorrência dabebida Mole, com corpo elevado e aromaachocolatado. A Tabela 2 mostra algumascaracterísticas físicas e sensoriais destas
localidades. Pela sua grande extensão territorial e
diversidade de microclirnas, o Estado de MinasGerais apresenta cinco regiões cafeeiras distintas:Sudoeste (Ouro Fino, Alfenas, Varginha), comaltitudes elevadas a médias, amplitude de déficithídrico (elevado a baixo) e variação de bebida(Mole a RiadaJ Rio), principalmente em funçãoda proximidade de represas; Centro-Sul (SantoAntônio do Amparo, Formiga), com temperaturasmédias anuais mais elevadas; Triângulo MineiroI Alto Paranaíba. com cultivo com irrigação emáreas de déficit hídrico muito elevado (Araguari,Monte Carmelo) e sem irrigação em áreas maisaltas (São Gotardo, Serra do Salitre), com bebidavariando desde Dura até Mole; e Zona da Mata IAlto Rio Doce (Caratinga,
Tabela 1 - Características fisicas e sensoriais de cafés arábicas do Estado do ParanáOrigem Processo Aspecto Bebida Acidez Corpo AromaLondrina terreiro Regular Rio Regular bom fermentadoApucarana CO Bom Mole Alta alto agradávelRibeirão Claro CO Bom Mole Média alto agradávelTomasina CD excelente Mole Média alto agradável
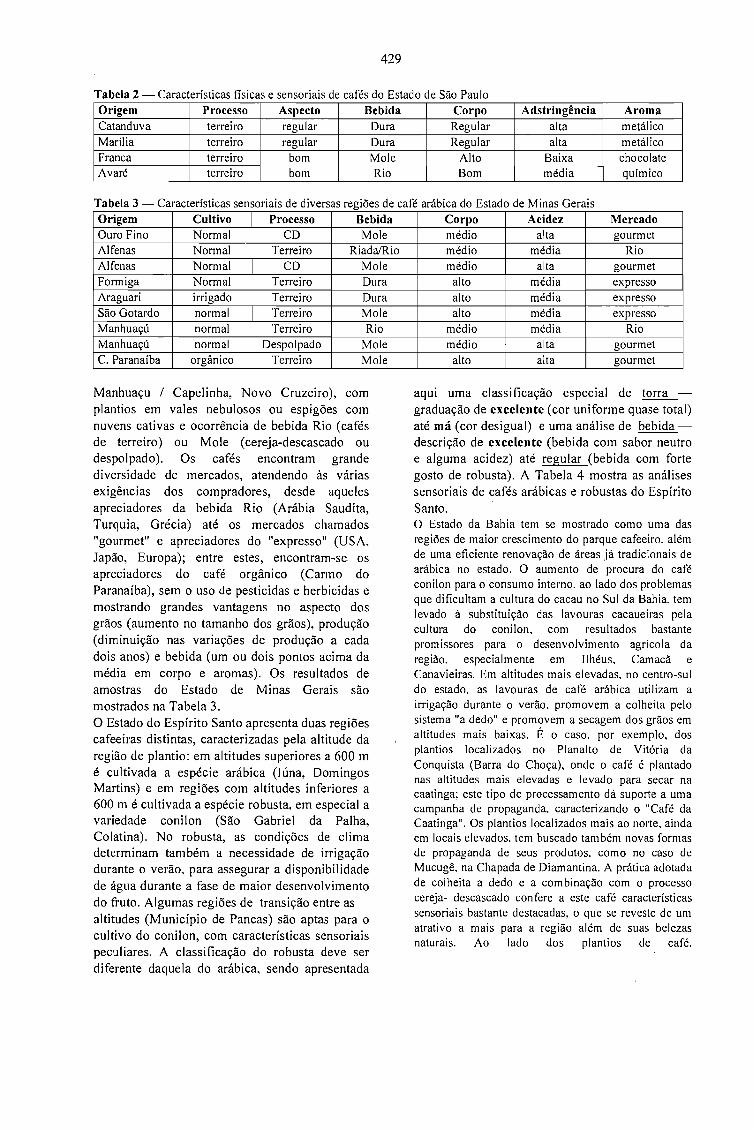
429
Tabela 2 - Características fisicas e sensoriais de cafés do Estado de São PauloOrigem Processo Aspecto Bebida Corpo Adstringência AromaCatanduva terreiro regular Dura Regular alta metálicoMarília terreiro regular Dura Regular alta metálicoFranca terreiro bom Mole Alto Baixa chocolateAvaré terreiro bom Rio Bom média químico
Tabela 3 - Características sensoriais de diversas regiões de café arábica do Estado de Minas GeraisOrigem Cultivo Processo Bebida Corpo Acidez MercadoOuro Fino Normal CO Mole médio alta gourmetAlfenas Normal Terreiro RiadalRio médio média RioAlfenas Normal CO Mole médio alta gourmetFormiga Normal Terreiro Dura alto média expressoAraguari irrigado Terreiro Dura alto média expressoSão Gotardo normal Terreiro Mole alto média expressoManhuaçú normal Terreiro Rio médio média RioManhuaçú normal Despolpado Mole médio alta gourmetC. Paranaíba orgânico Terreiro Mole alto alta gourmet
Manhuaçu / Capelinha, Novo Cruzeiro), complantios em vales nebulosos ou espigões comnuvens cativas e ocorrência de bebida Rio (cafésde terreiro) ou Mole (cereja-descascado oudespolpado). Os cafés encontram grandediversidade de mercados, atendendo às váriasexigências dos compradores, desde aqueles
apreciadores da bebida Rio (Arábia Saudita,Turquia, Grécia) até os mercados chamados"gourmet" e apreciadores do "expresso" (USA,Japão, Europa); entre estes, encontram-se osapreciadores do café orgânico (Carmo doParanaíba), sem o uso de pesticidas e herbicidas emostrando grandes vantagens no aspecto dosgrãos (aumento no tamanho dos grãos), produção(diminuição nas variações de produção a cadadois anos) e bebida (um ou dois pontos acima damédia em corpo e aromas). Os resultados deamostras do Estado de Minas Gerais sãomostrados na Tabela 3.
O Estado do Espírito Santo apresenta duas regiõescafeeiras distintas, caracterizadas pela altitude daregião de plantio: em altitudes superiores a 600 mé cultivada a espécie arábica (Júna, DomingosMartins) e em regiões com altitudes inferiores a600 m é cultivada a espécie robusta, em especial avariedade conilon (São Gabriel da Palha,Colatina). No robusta, as condições de climadeterminam também a necessidade de irrigaçãodurante o verão, para assegurar a disponibilidadede água durante a fase de maior desenvolvimentodo fruto. Algumas regiões de transição entre as
altitudes (Município de Pancas) são aptas para ocultivo do conilon, com características sensoriaispeculiares. A classificação do robusta deve serdiferente daquela do arábica, sendo apresentada
aqui uma classificação especial de torra graduação de excelente (cor uniforme quase total)até má (cor desigual) e uma análise de bebidadescrição de excelente (bebida com sabor neutroe alguma acidez) até regular (bebida com forte
gosto de robusta). A Tabela 4 mostra as análisessensoriais de cafés arábicas e robustas do Espírito
Santo.O Estado da Bahia tem se mostrado como uma dasregiões de maior crescimento do parque cafeeiro, alémde uma eficiente renovação de áreas já tradicionais dearábica no estado. O aumento de procura do caféconilon para o consumo interno. ao lado dos problemasque dificultam a cultura do cacau no Sul da Bahia. temlevado à substituição das lavouras cacaueiras pelacultura do conilon, com resultados bastantepromissores para o desenvolvimento agrícola daregião, especialmente em Ilhéus, Camacã eCanavieiras. Em altitudes mais elevadas. no centro-suldo estado, as lavouras de café arábica utilizam airrigação durante o verão. promovem a colheita pelosistema "a dedo" e promovem a secagem dos grãos emaltitudes mais baixas. É o caso. por exemplo, dosplantios localizados no Planalto de Vitória daConquista (Barra do Choça), onde o café é plantadonas altitudes mais elevadas e levado para secar nacaatinga; este tipo de processamento dá suporte a umacampanha de propaganda. caracterizando o "Café daCaatinga". Os plantios localizados mais ao norte. aindaem locais elevados. tem buscado também novas formasde propaganda de seus produtos. como no caso deMucugê, na Chapada de Diamantina. A prática adotadade colheita a dedo e a combinação com o processocereja- descascado confere a este café característicassensoriais bastante destacadas, o que se reveste de umatrativo a mais para a região além de suas belezasnaturais. Ao lado dos plantios de café.
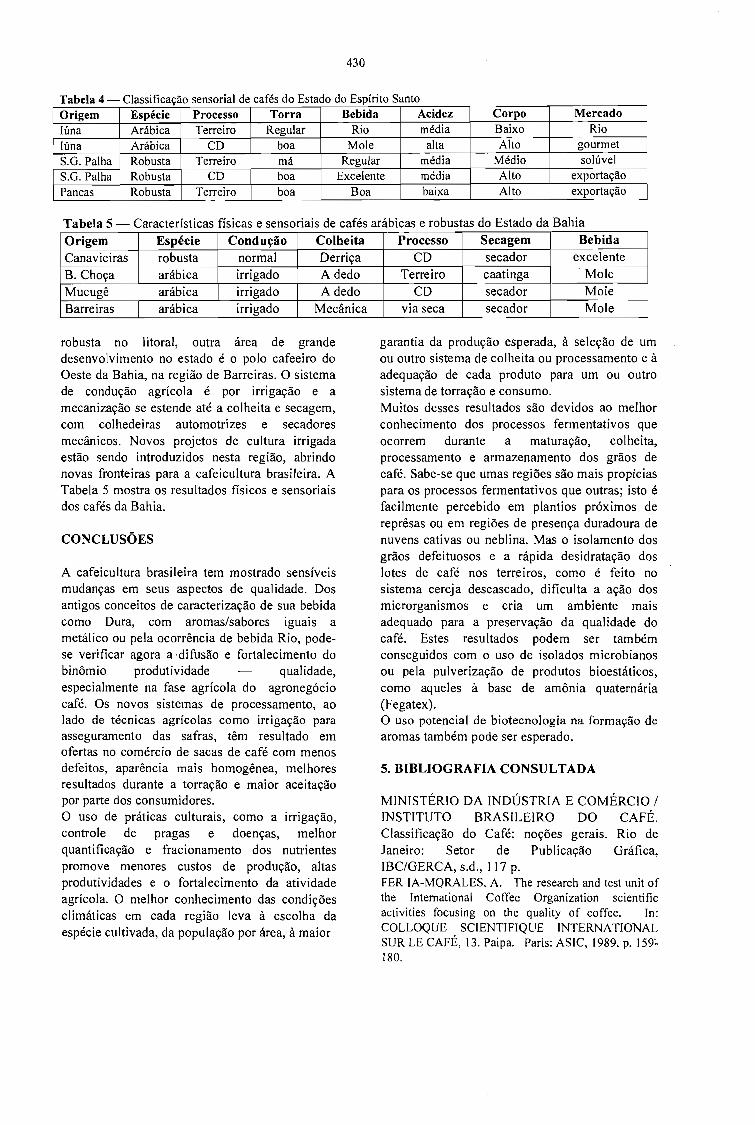
430
Tabela 4 - Classificação sensorial de cafés do Estado do Espírito SantoOrigem Espécie Processo Torra Bebida Acidez Corpo Mercadolúna Arábica Terreiro Regular Rio média Baixo Riolúna Arábica CD boa Mole alta Alto gourmets.o. Palha Robusta Terreiro má Regular média Médio solúvels.a. Palha Robusta CD boa Excelente média Alto exportaçãoPancas Robusta Terreiro boa Boa baixa Alto exportação
Tabela 5 - Caracteristicas físicas e sensoriais de cafés arábicas e robustas do Estado da BahiaOrigem Espécie Condução Colheita Processo Secagem BebidaCanavieiras robusta normal Derriça CO secador excelenteB. Choça arábica irrigado A dedo Terreiro caatinga . Mole
Mucugê arábica irrigado A dedo CO secador MoleBarreiras arábica irrigado Mecânica via seca secador Mole
robusta no litoral, outra área de grandedesenvolvimento no estado é o polo cafeeiro doOeste da Bahia, na região de Barreiras. O sistemade condução agrícola é por irrigação e amecanização se estende até a colheita e secagem,com colhedeiras auto motrizes e secadoresmecânicos. Novos projetos de cultura irrigadaestão sendo introduzidos nesta região, abrindonovas fronteiras para a cafeicultura brasileira. ATabela 5 mostra os resultados físicos e sensoriaisdos cafés da Bahia.
CONCLUSÕES
A cafeicultura brasileira tem mostrado sensíveismudanças em seus aspectos de qualidade. Dosantigos conceítos de caracterização de sua bebidacomo Dura, com aromas/sabores iguais ametálico ou pela ocorrência de bebida Rio, podese verificar agora a ·difusão e fortalecimento dobinômio produtividade qualidade,especialmente na fase agrícola do agronegóciocafé. Os novos sistemas de processamento, aolado de técnicas agrícolas como irrigação paraasseguramento das safras, têm resultado emofertas no comércio de sacas de café com menosdefeitos, aparência mais homogênea, melhoresresultados durante a torração e maior aceitaçãopor parte dos consumidores.O uso de práticas culturais, como a irrigação,controle de pragas e doenças, mel horquantificação e fracionamento dos nutrientespromove menores custos de produção, altasprodutividades e o fortalecimento da atividadeagrícola. O melhor conhecimento das condiçõesclimáticas em cada região leva à escolha daespécie cultivada, da população por área, à maior
garantia da produção esperada, à seleção de umou outro sistema de colheita ou processamento e àadequação de cada produto para um ou outrosistema de torração e consumo.Muitos desses resultados são devidos ao melhorconhecimento dos processos fermentativos queocorrem durante a maturação, colheita,processamento e armazenamento dos grãos decafé. Sabe-se que umas regiões são mais propíciaspara os processos fermentativos que outras; isto éfacilmente percebido em plantios próximos dereprêsas ou em regiões de presença duradoura denuvens cativas ou neblina. Mas o isolamento dosgrãos defeituosos e a rápida desidratação doslotes de café nos terreiros, como é feito nosistema cereja descascado, dificulta a ação dosmicrorganismos e cria um ambiente maisadequado para a preservação da qualidade docafé. Estes resultados podem ser tambémconseguidos com o uso de isolados mícrobianosou pela pulverização de produtos bioestáticos,como aqueles à base de amônia quaternária(Fegatex).O uso potencial de biotecnologia na formação dearomas também pode ser esperado.
5. BIBLIOGRAFIA CONSULTADA
MINISTÉRIO DA INDÚSTRIA E COMÉRCIO /INSTITUTO BRASILEIRO DO CAFÉ.Classificação do Café: noções gerais. Rio deJaneiro: Setor de Publicação Gráfica,IBC/GERCA, s.d., 117 p.FER IA-MQRALES. A. The research and test unit ofthe International Coffee Organization scientificactivities focusing on the quality of coffee. In:COLLOQUE SCIENTIFIQUE INTERNATlONALSUR LE CAFÉ, 13. Paipa. Paris: ASIC, 1989. p. 159:180.

431
POTENCIAL DE MERCADO DE CAFÉS ORGÂNICO E ESPECIAL.
Ivan Franco Caixeta
Produtor de café orgânico, Eng. Agrônomo M.Sc., Professor da Esacma Machado -MG
Fonte: Boletim do lBD Botucatu I SP
França Incitação na compra de produtos em %
GARANTIAS ECOLÓGICAS INFLUENCIAMBASTANTE
Europa os principais atributos dos produtosalimentícios em %
46,673,126,7
9, I64,8
Países do NorteAtributos Países do SulNatural 73, ISaudável 48,6Nutritivo 35,0Barato 4,7Sem Agrotóxicos 57,6Vitaminas e saisminerais 28, I 38,9
- Mais de 70% da população sabe que consomeprodutos com agrotóxicos e também de seu efeitonocivo à saúde,- Mais da metade da população (52%) já temconhecimento da existência de produtos semagrotóxicos,- Sua imensa maioria (90%) sentem-se inseguras egostaria de dispor de um certificado de garantia daqualidade e procedência dos produtos.Esses parâmetros mostram o imenso potencialpara o mercado de produtos sem agrotóxicos entreos consumidores brasileiros.No mercado internacional esta realidade ainda émais expressiva Tem se hoje algo em torno de17.000 produtores certificados ,distribuídos naEuropa (10.000), nos EUA (5.000) e nos outrospaíses2.000). Para o Brasil estima-se 1.000produtores certificáveis.Alguns dados sobre a pesquisa de mercado emoutros países: O preço é o de menos.
Características pesquisadas PercentualGarantiasde higiene e segurança 84O preço é competitivo 81O produto é produzido na França 74O produto porta sinal de qualidade 71A marca inspira confiança 68O produto tem garantias ecológicas 64O produto é fabricado na minha região 59O fabricante defende causas humanitárias 51O produto é fabricado na Europa 49
E ainda sobre a influência de garantias ecológicassobre o desejo de compra:
INTRODUÇÃO
Em franca expansão a agricultura orgânica estáganhando terreno em todo o mundo. Na Europa enos Estados Unidos da América a área cultivadasem agrotóxicos esta crescendo 30% ao ano e noBrasil, o aumento anual chega a 10%. Aagricultura orgânica tem sido tema de amplosdebates recentes sobre os rumos da produção e éhoje uma realidade nacional e internacional.Folha de São Paulo, 20/10/98).A agricultura orgânica reaparece como umaalternativa para fazer sua parte em tentarreequilibrar os exageros e os desequilíbrioscausados pela agricultura química que apesar dealtas tecnologias e da chamada revolução verdenão resolveu a falta de alimentos no mundo, mastrouxe grandes danos ao nosso ecossistema e aqualidade de vida do consumidor. Hoje a pressãopor parte de organizações internacionais e dopróprio consumidor, quanto aos aspectos depreservação e sustentabilidade do meio ambientetem levado os órgãos públicos a adotaremmedidas restritivas quanto ao uso dos recursosnaturais. Exigindo uma legislação própria para aadequação de tecnologias visando a preservação ea qualidade ambiental.A comercialização dos produtos limpos ouorgânicos, vem se deslocando de um mercadoonde os consumidores se preocupavam sobretudocom a saúde, para um mercado onde eles tem emmente questões de caráter ambientale social. O consumidor tem consciência de que amaior proporção do prêmio que ele paga a maisirá:
• favorecer o meio ambiente,
• chegar diretamente aos produtores rurais,
• contribuir para uma maior equidade sócioeconômica (Fairtrade). Pesquisa do InstitutoGallup, publicada no jornal da AgriculturaOrgânica, informativo da AAO-SP, Associação deAgricultura Orgânica de São Paulo, ressaltaalguns resultados interessantes, sobre o potencialde mercado de produtos orgânicos na capitalpaulista:- Cerca de 72% da população da capital paulistaaceita pagar 20 a 30% a mais por produtoscomprovadamente sem agrotóxicos,
Fonte: IGIA Boletimdo IBD Botucatu I SP

POTENCIAL DE MERCADO DE CAFÉSORGÂNICO E ESPECIAl.
o café é a segunda maior commodity. Os cafésespeciais, entre eles o orgânico são os únicosprodutos que estão com crescimento expressivoem todos os principais consumidores mundiaiscomo é o caso de toda a Europa, América doNorte e Japão. Em todos eles e nos principaispaíses produtores a cafeicultura orgânica éconsiderada um fi lão de expressivo valorambiental, social e econômico. Não existembarreiras para os cafés produzidos semagrotóxicos e sem afetar o meio.Para o Brasil é uma ótima oportunidadecompetitiva de melhorar sua imagem de cafés dequalidade no mercado internacional, oferecendoum produto saudável, que preserva o meioambiente e de qualidade. ? Há condições de, emum curto período de tempo, oferecer quantidadesexpressivas de cafés de qualidade com garantiasde origem e produtos com certificação para asdiversas, exigências do consumidor final.O mercado interno tem condições de absorverquantidades expressivas destes cafésdiferenciados, pois atualmente os valores sãomuito pequenos diante de um mercadoconsumidor de 12 milhões de sacas.Os principais países produtores dos dois tipos de
cafés mencionados se encontram na AméricaCentral, México, Colômbia, Peru e Equador,sendo que o Brasil ocupa uma posição de poucodestaque, mas com grande potencial decrescimento. ? Os principais países consumidoresdestes cafés diferenciados continuam entre os quemais consomem o café tradicional como, EUA,Alemanha, Japão, Países Baixos, Suíça, Itália,França, Áustria, Espanha, Canadá, entre outros.A grande dificuldade de comercialização destescafés diferenciados é a necessidade de busca dealianças estratégicas, ou seja, encontrar oscaminhos de vendas diretas, que são difíceis etrabalhosos. O produtor precisa ter bons contatos,quantidades e qualidades uniformes, grandeconfiabilidade e ética profissional para atendercom presteza os exigentes compradores.Outra grande dificuldade encontrada é o famoso
Custo Brasil, que onera nossa capacidade deexportar. Se for reduzido melhoram-se ascondições de concorrer com outros produtoresdestes cafés.
432
METAS DOS GRUPOS ENVOLVIDOS NOMERCADO DESTES CAFÉS
Agricultura orgânica não é uma volta ao passado,como muitos afirmam e nem um grupo querepudia a pesquisa tecnológica, ao contrário,precisa-se somar informações e novas tecnologiasque visem uma produção mais limpa e projetosque integrem todo ecossistema. Prova disto é ogrande passo que já foi dado com a tecnologia doplantio direto; até bem pouco tempo o sistemaconvencional de plantio era um dogma, e hoje, éuma realidade inclusive para o plantio de café emgrandes áreas.
Principais metas:- conseguir apoio dos órgãos de pesquisa,universidades e entidades para seremdesenvolvidos projetos amplos e seguros quedêem suporte e segurança ao produtor, trabalhadorrural e consumidor. Isto sem reduzirprodutividade e qualidade e que tenham custo deprodução acessíveis e renda digna ao produtor.
buscar o comércio justo, onde o pequenoprodutor tenha seus ganhos assegurados, mesmocom pequena produção. É tarefa dos grupos depesquisa, das associações e do próprio governocriar estratégias que encontrem soluçõesmercadológicas e estimulem os grupos depequenos produtores a ter mais acesso acomercialização de café orgânico e especial. E porhaver também maior preocupação com meioambiente neste grupo de agricultores.- o aprimoramento da certificação e sua
adequação às regiões tropicais com suaspeculiaridades diferentes das de clima frio é desuma importância para sua aplicabilidade. Pois oconsumidor final e o produtor devem ter atranqüilidade de saber de onde vem e com quegarantias ele recebe aquele produto em sua mesa.Não somente garantias de qualidade, mas sociais,da forma de produção e de proteção ao meioambiente.- fortalecer as associações tanto depequenos produtores como do produto café. Istofortalece o setor, aumenta o poder decomercialização e de marketing. Como exemplo,um ano após a fundação da ACOB (Associação deCafeicultura Orgânico do Brasil) , obteve-seresu ltados promissores em termos dereconhecimento tanto político quanto comercial,com um pequeno número de associados.- a filantropia com fins sociais e ecológicos deveser uma meta importante dos grupos envolvidoscom os cafés especial e orgânico, buscandoresgatar desastres que danificaram pessoas e omeio ambiente em qualquer parte do nossoplaneta.

No Brasil, tanto a produção quanto o consumodestes cafés são muito pequenas, mas com umcrescimento expressivo. No caso específico docafé orgânico os produtores ligados a ACOB, temuma produção que representa pouco mais de0,15% da produção brasi leira conformeestimativas de 1999. Mas este crescimento, desdede 1992 é de mais de 100% ao ano. Do ano de 98para 99 passamos de 22 mil sacas para umaestimativa de 40 mil sacas e de 6 produtores em97 para 22 em 99.Apesar deste grande crescimento temos dito:- O que temos feito em termos de preservaçãoambiental, de sustentabilidade e de produção decafé orgânico é apenas uma gota de água nooceano porém o prazer de se fazer algo pelonosso ecossistema é imenso e grandementerecompensador.
CONCLUSÃO
Os cafés especial e orgânico não são um modismoque passará, como dizem muitos agourentos epessimistas, mas sim novos produtos que vierampara ficar, e que ganham mercado a passos largos,onde existem muitas pessoas sérias envolvidas enão somente ecologistas radicais. O mercadoconsumidor se fortalece em todo o mundo,principalmente nos países desenvolvidos, o quenão deixa de ser uma forma de transferir riquezasdestes países para os menos desenvolvidos, poisos cafés orgânico e especial conseguem umprêmio na sua comercialização. Por ser umproduto nobre, não só por suas qualidades, mastambém por sua preocupação com o meioambiente, com o trabalhador rural e com aremuneração justa do produtor, estimulaconsumidores também preocupados com estasquestões sumamente importantes para o nossobem estar social e ecológico.
433
CONSULTA BffiLIOGRAFICA
AAO, Jornal da Agricultura Orgânica,Informativo da Associação de AgriculturaOrgânica €AAO, diversos. São Paulo - Brasil.ACOB News, Boletim Informativo da Associaçãode Cafeicultura Orgânica do Brasil € ACOB,diversos. São Paulo - BrasilBOLETM AgroEcológico, Informativo daLivraria e Editora AgroEcológica, diversos.Botucatu ,SP - BrasilPASCHOAL, A . D. Produção Orgânica deAlimentos: agricultura sustentável para os séculosXX E XXI 1 ed.. Piracicaba, SP: ESALQ/ USP,1994. 191p.PRIMAVESI, A. M. O Manejo ecológico dosolo: agricultura em regiões tropicais. 4 ed. SãoPaulo, SP: Nobel, 1982. 541 p.TEODORO, V.C.de A. & CAIXETA, LF.Boletim técnico de café orgânico. 1 ed. Lavras,MG: UFLA/ Faepe, 1999.69 p.


435
MYCOTOXIGENESIS IN GRAINS APPLlCATION TO MYCOTOXIC PREVENTION IN COFFEE
J. Le Bars & P. Le Bars
Laboratoire de Pharmacologie - Toxicologie I.N.R.A. BP 3 .31931 TOULOUSE Cedex
ABSTRACTMycotoxins are loxic secondary metabolites produced by fungi. These loxins are low molecular weightcompounds derived from polyketo - acids, lerpenes or amino-acids. A loxin can be synthesized by a diversityof fungai species. The term "toxigenic fungai species" means a species for which certain strains canproduce, at least under cerlain conditions, one 01' several toxic metabolites. Depending on lhe biology ofthefungus and ecological parameters ( substrate, hydro-thermic conditions, ...), toxigenesis can occur in lhe field(i.e. alkaloids from endophytic 01' parasitic fungi) OI' during periharvest period (i.e. fumonisins fromFusarium moniliforme), or in storage (i.e. aflatoxins from Aspergillus flavus). Moreover, lhe contaminatingloxin can be stable (i.e. aflatoxins, ochratoxins A....) 01' instable in certain substrate and / or with thermaltreatment (i.e. patulin, citrinin....). So many parameters have lo be considered in lhe mechanism of amycototoxic contamination ofa commodity. Ochratoxin A, lhe main reported mycotoxin in green, roasted andsoluble coffee, will be discussed ali along lhe chainfrom thefield lo lhe final product, in order lo determine,in collaboralion with agricultura! and technological experts, lhe moment and lhe fale of actualcontamination and, as a consequence, effective measuresfor prevention and obtaining end products withoutmycotoxin.
INTRODUCTlON
Numerous fungai species may develop ali alongthe food chain, from the plant in the field, duringprocessing steps and storage, down to consumer'splate or cup. FungaI development in alimentarysubstrates can lead to several detriment effects: (I)alterations of aspect and technological properties,(11) modification of nutritive value, (1/1)impairment of organoleptic features. (IV)production of joxic compounds. The presentobjective is, first. to remind the general mainmechanisms and conditions conducive tomycotoxic contamination in order to contribute toa logical prevention. Second. on the basis of dataon occasional contamination of coffee. toxinorigino its fate during processing and mostprobable criticai points are discussed.
1. MYCOTOXINS AND MYCOTOXIGENESIS:
I-I. Toxic products resulting from fungaIdeveloped in vegetal products .Occurrence of toxic metabolites in foods andfeeds resulting from fungai development on plantsubstrates may result from any of four generalmechanisms. Firstly. a saprophyte mould growingon a favorable substrate can directly produce toxicmetabol ites (e.g. atlatoxi ns by Aspergillus flavus).Second ly, the association of an endophyte with aliving plant can lead to toxicosis in animais (e.g.fescue foot disease in cattle with Neotyphodium
coenophialumi. Third ly, in response to theinvasion of phythopathogenic fungi. a living plantcan produce diverse rnetabolites, some ofthem atleveis that are toxic (e.g. ipomeamarone inIpomea pala/as). Fourthly. atoxigenic species maycontribute by bioconversion to transform atoxiccompounds to toxic ones (i.e. dicoumarol inmouldy Melilotus forage).(Le Bars and Le Bars,1996). Only the two first situations correspond tomycotoxins sensu stricto, which may be definedas zootoxic metabolits directly produced bymicromycetes. They are structural cornplex, theyvary from simple C4-compounds (e.g.tremorgenic mycotoxins) (Steyn. 1980).
1-2. Origin of mycotoxinsMycotoxins are principally characterized assecondary metabolites. The secondary metabolismcomprises biochemical pathways whose terminalsynthesis products have no pertinent role for thebiology ofthe organismo In contrast to the prirnarymetabolismo which is fundarnentally similaramong different living organisrns, the secondarymetabolism qualitatively depends on the fungaispecies and quantitatively is often characteristic ofeach individual strain. In comparison to otherorganisms, secondary metabolism is verydeveloped in micromycetes and leads to a vastarray of molecules (Turner 1971. Turner andAlridge 1983), some of which are toxic to otherorganisms: bacteria (antibiotics), animaIs(mycotoxins).

Mycotoxins are elaborated through differentmetabolic pathways (Steyn, 1980). Polyketidesand terpenes are two largest groups (Table 1,Steyn 1998).Finally, secondary metabolites often aresynthesized as families of chemically relatedproducts. The type and quantity of moleculesproduced being largely dependent on thecharacteristics of the individual strain as well asenvironmental conditions (nutrients, physic chemical parameters, biochemical and biologicalstatus ofthe substrate).So, the term "toxigenic fungaI species" generallymeans a fungai species for which certain strainscan produce, at least under certain conditions, oneor more toxic metabolites (Le Bars, 1988).Main mycotoxins and their major fungal sourcesare presented in the Table 2.
1-3. Factors influencing toxigenesis (Le Bars1988,1998)Conditions permissive to toxigenesis are morelimited than those permitting fungal growth.Among the factors leading to the presence ofmycotoxins in foods, there are two maincategories: intrinsic factors which are dependenton the fungai species or strain and extrinsicfactors which depend on environmentalconditions.
3.1 Intrinsic factorsOne mycotoxin is rarely produced by only onespecies or closely related ones, as it is the case for
436
atlatoxins by Aspergillus flavus or A. parasiticus,fumonisins by Fusarium moniliforme (and relatedspecies), sporidesmijns by Pithomyces chartarum(Table 2). More frequently, the same toxin can besynthesized by a diversity of fungai speciessometimes belonging to different genera: forexample, cyclopiazonic acid by A. flavus and P.camemberti, ochratoxin A (OTA), patulin andpenicillic acid by strains of different species ofAspergillus and Penicillium. Moreover, onefungal species and strain can produce more or less'simultaneously several mycotoxins: OTA andcitrinin by P. verrucosum, OTA and penicillicacid by A. ochraceus, atlatoxins andcyc1opiazonic acid by A. flavus, PR toxin,roquefortine, mycophenolic acid and patu1cin byP. roqueforti.In contrast, within a particular species reputed tobe toxigenic, every strain does not possess thisproperty; the frequency of toxigenic strains beingdependent on the species considered and in somecases the region or substrate from which the strainwas isolated. Of greater concern than thisfrequency is the difference in toxigenic potentialwithin a particular species: among isolates fromthe sames species toxin production can vary by asmuch as 1000 - fold.As an overview on the dozen of mold mycotoxinpair we studied, in each case on the basis of 100to 200 freshly isolated in optimal conditions foreach toxigenesis, the four schemes of toxigenicpotential distribuition (TPD) were observed:
Biosynthetlc Category Representatlve mycotoxlns
Polykelides Di- MonillforminTetra- Patulln, penieillic aeidPenta- Citrinin, diplosporin. ochratoxtnsHexa- Maltoryzine,.Hepla- Rugulosin. viomellein. vtrldttoxín. xanlhomegninocta- Ergoehromes. luteoskirinNona- Cítreoviridin, cytocnalasm. fumonisins. zearalenoneOeea- Atlatoxíns, austocystins. erythroskirine
Telramic aeids Cyelopiazonie aeid, tenuazontc aeidDiketopiperazines Simpie Aspergillie acld. eehinulins
Modified Brevianamides. lumitremorgens, oxaline,roquefortine. sporldesmins, verrucutotoxín
Peptides Ergolamine. phomopsins , lryptoquivaline
Terpenes Mono- VlrtdicaturntoxlnSesqui. TrieholheeenesDi- Aflatrem, oenitrems
Table 1: Mycotoxins as representative of tne Main Biosyntnetic Categorias ofsecondary metabolites (Steyn, 1998).

Main mycotoxins
AflatoxinsCyclopiazonic acidOchratoxin A
Patulin
Sterigmaloc:ystinCitrininFumonisins, moniliformine,
fusarenon X
ZearalenoneTrichothecens:T2 toxin, otacetoxyscupenol. ....Deoxynivalenol ....
SporidesminsPhomopsinsErqot alkatotds
437
Major toxlgenlc species
Aspergillus tisvus, A. psrssittcusA. ttevus, Penicillium cyclopiumA. ocnteceus, P. verrucosum (viridicatum)
A. ctevetus, P. expansum, P. granulatum.8yssochlamys nivee, Paecilomyces vetioti. ...
A. verslcolor, A. nidulansP. citrinum, P. verrucosum.... Monascus purpureus
Fusarium moniliforme
F. graminearum, F. culmorum
F. spototncnoides. F. poaeF. culmorum, F. graminearum
Pithomyces chartarumPhomopsis /eptostromiformisC/avieeps, Neotyphodium coenopnielum •
• Endophyte in Festuea arundinaeea.
Table 2: Main myeotoxins (natural/y occurring aeeording to súbstrete, cttmeticand tecnnaoçtceteonditions) and their major fungaI sources.
1- Almost 100% toxigenic strains; TPO is close toa normal distribution. inasmuch potential isexpressed by the logarithm of the maximalobserved concentration of toxin: sterigmatocystin/ A. versicolor, penicilJic acid / P. verrucosumcyclopiurn, cyclopiazonic acid / P. camemberti,fumonisin B l/F. moniliforme.2- A noticeable proportion of non-toxigenicstrains (20 to 75%); the other ones presenting aTPO similar as the precending one: aflatoxins andcyclopiazonic acid / A. flavus, OTA / A.ochraceus, deoxynivalenol and zearalenone í F.graminearum.3- Two clear - cut groups of strains, non toxigenicand highly toxigenic ones: satratoxins /Stachybotrys atra, sporidesmins / Pithomyceschartarum.4- Very rare producing strains: fusarochromanone(TOPI) / F. roseum (only one strain out onOO).Specific biological properties of the fungus maydetermine the moment and the nature of substratecontamination. Certain fungi considered aspathogens develop within the living plantsometimes symptomless and can lead tomycotoxic contamination of end product, i.e.Fusarium spp in cereaIs and their mycotoxins,Other species are occasionally pathogenic on
specific substrate, like P. expansum on appIesconducive to patulin contamination. Other onesare endophytes, strictly associated to a vegetalspecies, non pathogenic but producingrnycotoxins: Neotyphodium coenophialum inFestuca arundinacea. Other ones present in anopportunist weak pathogenicity: i.e. A. jlavuspenetraing the flower of com. or invading peanutpod and grains in the field. Lastly, rnost of theknown mycotoxigenic species are onlysaprophytes. When investigations are undertakenon a toxigenic species or on a mycotoxiccontarnination, it is important to know (andsometimes to verify) what is such biologicalsituation. Thus, if the mycotoxigenic species istransmited by the seed (i.e. F. moniliformei, thefirst step to lirnit fumonisin contarnination in comis to use upstream Fusarium - free seeds.
3.2 Extrinsic factorsWater availubitity: water activity (aw) influencestoxin production considerably, particularly in lowhydrated products. Toxigenesis takes place onlyfor aw slightly higher (i.e. + 0,02) than aw limitfor growth, which is particular for each fungaIspecies (0,80 for A. flavus, 0,76-0,78 for A.ochraceus, 0.85 for numerous Penicillia, 0.90 for
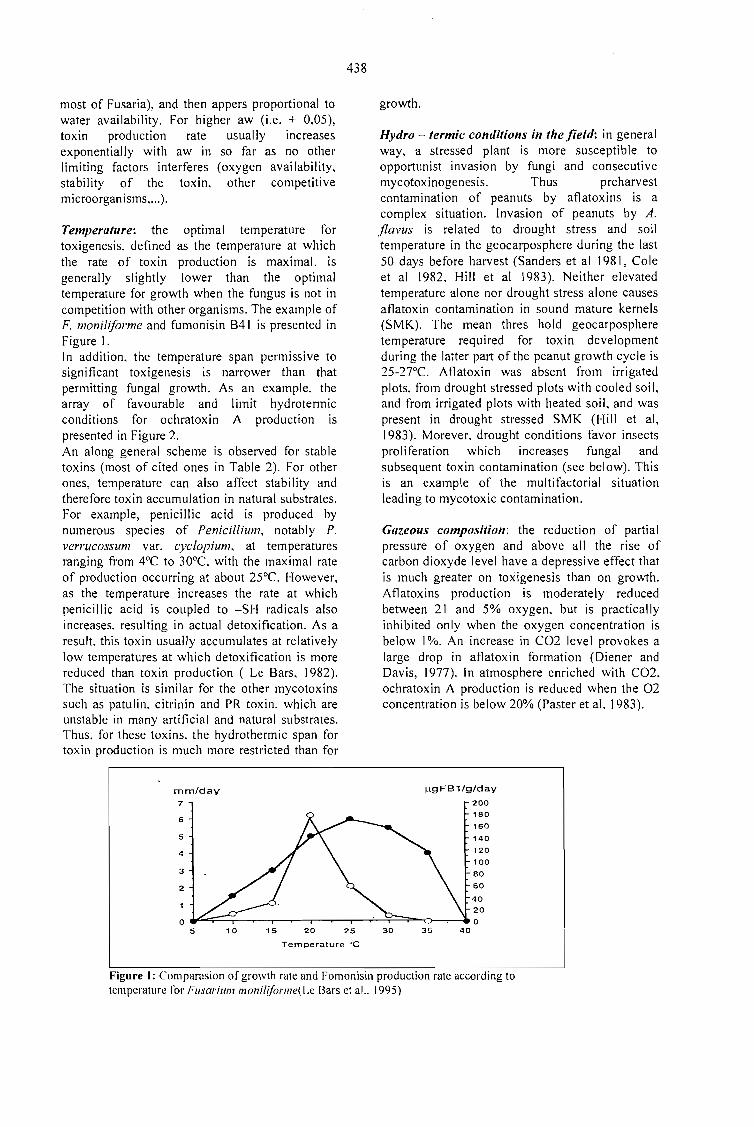
438
most of Fusaria), and then appers proportional towater availability. For higher aw (i.e. + 0,05),toxin production rate usually increasesexponentially with aw in so far as no otherlimiting factors interferes (oxygen availability,stability of the toxin, other competi tivemicroorganisms....).
Temperature: the optimal temperature fortoxigenesis, defined as the temperature at whichthe rate of toxin production is maximal. isgenerally slightly lower than the optimaltemperature for growth when the fungus is not incompetition with other organisms. The example ofF. moniliforme and fumonisin B41 is presented inFigure I.In addition, the temperature span permissive tosignificant toxigenesis is narrower than thatpermitting fungai growth. As an example, thearray of favourable and limit hydrotermicconditions for ochratoxin A production ispresented in Figure 2.An along general scheme is observed for stabletoxins (most of cited ones in Table 2). For otherones, temperature can also affect stabil ity andtherefore toxin accumuJation in natural substrates.For exarnple, penicillic acid is produced bynumerous species of Penicillium, notably P.verrucossum var. cyclopium, at temperaturesranging from 4°C to 30°C. with the maximal rateof production occurring at about 25°C. However,as the temperature increases the rate at whichpenicillic acid is coupled to -SH radicais alsoincreases, resulting in actual detoxification. As aresult, this toxin usually accumulates at relativelylow temperatures at which detoxification is morereduced than toxin production ( Le Bars, 1982).The situation is similar for the other mycotoxinssuch as patulin, citrinin and PR toxin, which areunstable in many artificial and natural substrates.Thus, for these toxins, the hydrothermic span fortoxin production is much more restricted than for
growth.
Hydro - termic conditions in the fleld: in generalway, a stressed plant is more susceptible toopportunist invasion by fungi and consecutivemycotoxinogenesis. Thus preharvestcontamination of peanuts by aflatoxins is acomplex situation. Invasion of peanuts by A.flavus is related to drought stress and soiltemperature in the geocarposphere during the last50 days before harvest (Sanders et ai 1981, Coleet ai 1982, Hi11 et ai 1983). Neither elevatedtemperature alone nor drought stress alone causesaflatoxin contamination in sound mature kernels(SMK). The mean thres hold geocarpospheretemperature required for toxin developmentduring the latter part ofthe peanut growth cycle is25-27°C. Aflatoxin was absent from irrigatedplots, from drought stressed plots with cooled soil,and from irrigated plots with heated soil, and waspresent in drought stressed SMK (Hill et ai.1983). Morever. drought conditions favor insectsproliferation which increases fungal andsubsequent toxin contamination (see below). Thisis an example of the multifactorial situationleading to mycotoxic contamination.
Gazeous composition: the reduction of partialpressure of oxygen and above ali the rise ofcarbon dioxyde levei have a depressive effect thatis much greater on toxigenesis than on growth.Aflatoxins production is moderately reducedbetween 21 and 5% oxygen. but is practicallyinhibited only when the oxygen concentration isbelow 1%. An increase in C02 level provokes alarge drop in aflatoxin formation (Diener andDavis, 1977). In atmosphere enriched with C02.ochratoxin A production is reduced when the 02concentration is below 20% (Paster et al, 1983).
3530252015105
2
3
6
4
~lgFB1/g/day
200
160
160
140
120
100
60
60
4020
o..,:;.oe;:::=-T-~---'r-~---.--~.,...---,-'--T==::::;=--<:>-~""'o
40
5
mm/day
7
Temperature 'c
Figure 1: Cornparasion of growth rale and Fornonisin production rale accordíng toternperature ror Fusarium moniliformeç l.e Bars et ai.. 1995)
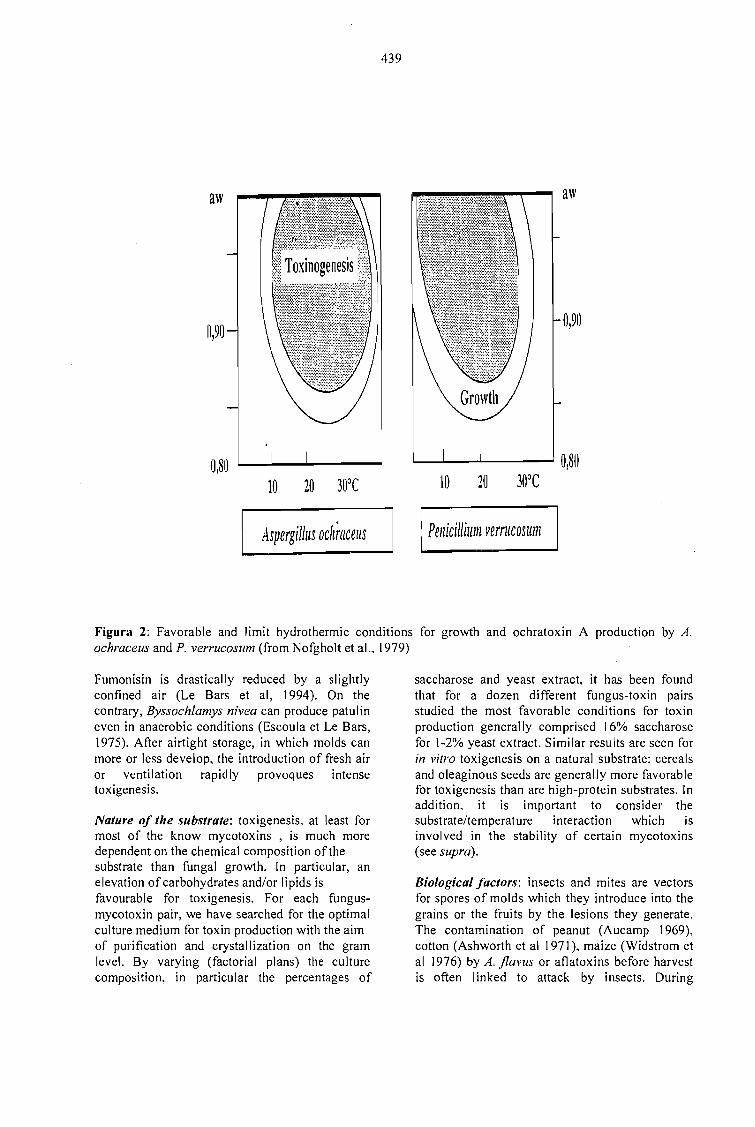
439
aw
n90,
0,80
Aspergillus oc/!racells Pellicillilllll verrllCOS1l11l
Figura 2: Favorable and limit hydrothermic conditions for growth and ochratoxin A production by A.ochraceus and P. verrucosum (from Nofgholt et al., 1979)
Fumonisin is drastically reduced by a slightlyconfined air (Le Bars et ai, 1994). On thecontrary, Byssochlamys nivea can produce patulineven in anaerobic conditions (Escoula et Le Bars,1975). After airtight storage, in which molds canmore or less develop, the introduction of fresh airor ventilation rapidly provoques intensetoxigenesis.
Nature of the substrate: toxigenesis, at least formost of the know mycotoxins , is much moredependent on the chemical composition ofthesubstrate than fungai growth. In particular, anelevation ofcarbohydrates and/or lipids isfavourable for toxigenesis. For each fungusmycotoxin pai r, we have searched for the optimalculture medi um for toxin production with the aimof puri fication and crystall ization on the gramleveI. By varying (factorial plans) the culturecornposition, in particular the percentages of
saccharose and yeast extract, it has been foundthat for a dozen different fungus-toxin pairsstudied the most favorable conditions for toxinproduction generally comprised 16% saccharosefor 1-2% yeast extract. Similar results are seen forin vitro toxigenesis on a natural substrate: cereaisand oleaginous seeds are generalJy more favorablefor toxigenesis than are high-protein substrates. Inaddition, it is important to consider thesubstrate/temperature interaction which isinvolved in the stability of certain mycotoxins(see supra).
Biological factors: insects and mites are vectorsfor spores of molds which they introduce into thegrains or the fruits by the lesions they generate.The contamination of peanut (Aucamp 1969),cotton (Ashworth et ai 1971), maize (Widstrom etai 1976) by A. flavus or af!atoxins before harvestis often linked to attack by insects. During
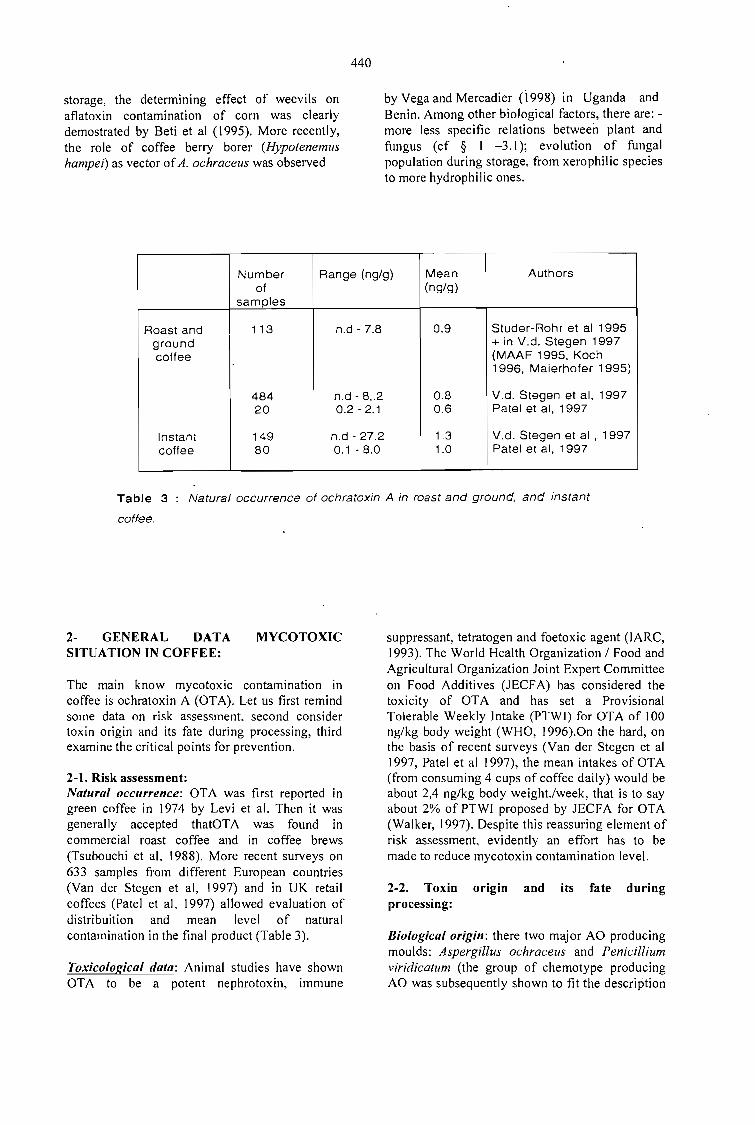
storage, the determining effect of weevils onaflatoxin contamination of com was clearlydemostrated by Beti et al (1995). More recently,the role of coffee berry borer (Hypotenemushampei) as vector ofA. ochraceus was observed
440
by Vega and Mercadier (1998) in Uganda andBenin. Among other bioJogical factors, there are: more less specific relations between plant andfungus (cf § I -3.\); evolution of fungaipopulation during storage, from xerophilic speciesto more hydrophilic ones.
Number Range (ng/g) Mean Authorsof (ng/g)
samples
Roast and 113 n.d - 7.8 0.9 Studer-Rohr et ai 1995ground + in V.d. Stegen 1997coffee (MAAF 1995. Koch
1996. Maierhofer 1995)
484 n.d - 8,.2 0.8 V.d. Stegen et ai, 199720 0.2 - 2.1 0.6 Patel et ai, 1997
Instant 149 n.d - 27.2 1.3 V.d. Stegen et ai , 1997coffee 80 0.1 - 8.0 1.0 Patel et ai, 1997
Table 3 Natural occurrence oi ochratoxin A in roast and ground, and instant
coffee.
Toxicological data: Animal studies have shownOTA to be a potent nephrotoxin, immune
The main know mycotoxic contamination incoffee is ochratoxin A (OTA). Let us first remindsome data on risk assessment. second considertoxin origin and its fate during processing, thirdexamine the criticai points for prevention.
2-1. Risk assessment:Natural occurrence: OTA was first reported ingreen coffee in 1974 by Levi et aI. Then it wasgenerally accepted thatOTA was found incommercial roast coffee and in coffee brews(Tsubouchi et aI. 1988). More recent surveys on633 samples from different European countries(Van der Stegen et ai, 1997) and in UK retailcoffees (Patel et ai, 1997) allowed evaluation ofdistribuition and mean levei of naturalcontamination in the final product (Table 3).
2- GENERAL DATASITUATlON IN COFFEE:
MYCOTOXIC suppressant, tetratogen and foetoxic agent (IARC,1993). The World Health Organization / Food andAgricultural Organization Joint Expert Committeeon Food Additives (JECFA) has considered the
. toxicity of OT A and has set a ProvisionalTolerable Weekly Intake (PTWI) for OTA of \00ng/kg body weight (WHO, 1996).On the hard, onthe basis of recent surveys (Van der Stegen et ai1997, Patel et ai 1997), the mean intakes of OTA(frorn consuming 4 cups of coffee daily) would beabout 2,4 ng/kg body weight./week, that is to sayabout 2% of PTWI proposed by JECFA for OTA(Walker, 1997). Despite this reassuring element ofrisk assessment. evidently an effort has to bemade to reduce mycotoxin contamination leveI.
2-2. Toxin origin and its fate duringprocessing:
Biological origin: there two major AO producingmoulds: Aspergillus ochraceus and Penicilliumviridicatum (the group of chemotype producingAO was subsequently shown to fit the description

of P. verrucosum; Frisvad 1984, Pitt 1987, Moss1996). They differ somewhat in their hydrotermicrequirements (Northolt et al 1979, Ramos et ai1998). The minimal Aw for growth of A.ochraceous Ís 0,76 and 0,81 for P. verrucossum,but the minimal Aw for growth of A. ochraceousis 0,76 and 0,81 for P. verrucosum, but theminimum for AO production is 0,85 for bothorganisms. The temperature range is lO-37°C(optimum 25-30°C) for A. ochraceus and 4-30° forP. verrucosum (Figure 2). Such respectiveecological characteres explains, at least partially,the fact that the former one is associated with OAcontamination in warm countries, and the secondone in cooler ones (Danmark, ...).
Fate during processing and brewing: inconsistentresults have been published regarding theinf1uence of roasting on the OTA content inroasted beans and the transfer into the coffeebrew. This would be due to the highinhomogeneity of the OTA contamination withinone sample (Studer-Rohr et aI 1995, Blanc et aI1998). a general problem encountered withmycotoxins in grains and fruits.Studer-Rohr et aI (1995) observed that in naturallycontaminated samples no statistically significantdifference between OTA content of the green androasted beans was found. However, in artificiallyinoculated sarnples, much more homogeneouslycontaminated, loss of OTA was estimated from 2to 29%. More recently, after analysis of OTAdistribuition in a naturally contaminated greencoffee lot. Blanc et al (1998) obtained a slighteliminatÍon oftoxin during green coffee cleaning.But the roast and ground coffee contained only16% of OTA originaJly present: this reduction ismostly due to thermal degradation (Iess than 20%accounted by the chaff). Such a result. obtainedwith a roasting cycle of 14 mn and a finaltemperature 223°C. is consistent with thetherrnostability data of OTA in a dry substrate:half-Iife times of 12 and 6 min respectively at 200and 250°C (Boudra et al, ]995).In the brewing methods frequently used, 70% toalmost complete extraction of OTA occus(Studer-Rohr et ai 1995. Viani 1996. Van derSregen et ai 1997, Blanc et ai 1998).Taking into account this partial stability (andtransfer to the brew) during processing, decreaseof contamination frequency and levei in the endproduct is based on prevention ofochratoxigenesis upstream.
2.3. Most probable criticaI poiuts:Taking in account the preceding data, let usexamine the three major possible periods for
441
fungaI infection and toxigenesis: in the orchard,during harvesting processing drying, and storage.
The orchard: the ochratoxigenic fungaI speciesare not phytopathogens nor endophytes, so directinternai infection of the fruit from the tree is notpossible. They are opportunist saprophytes. Sogeneral prevention, ali along the chain, would belogically based oh: avoiding fungaI sporessources, Iimiting open entries into the fruit,reducing, after ripeness, water availability as soonas possible.In a orchard, clearing away decaying organicmaterial, particularly fallen damaged fruits,actually contributes to reduce fungaI sporessources.
Healthy plant predispose to healthy fruits.On tree infection occurs due to inoculated sporesby insects or airbone spores penetrating into thepulp or seed through skin lesions such droughtlacerations, insect damages or splitting berríesconsecutively to rain during fruit ripening stage,simiJarly as for A. flavus infection in figs (Boudraet ai, 1994). Actually, partial studies on sound,over ripe, sun scorched and split fruits on theplant indícated that Aspergillus and Penicilliumpopulations were comparatively high in split fruits(Naidu et ai, 1997). Unfortunatelycompleteidentification of species nor mycotoxin analysiswere not reported. Moreover, in Uganda andBenin, 5 to 17% of adults coffee berry borderemerging from the beans were infected by A.ochraceous (Veja and Mercadier, 1998). Theseauthors consider that its potential to serve as avector for this cosmopolitan fungus is high.Jt would be very important to examine when andhow the fungus colonizes the pulp and the beansby mycological and histological techniques(passive or active penetration?) and the delay andlocalization of toxigenesis (in the beans ordiffusion of the toxin from the pulp?), because wehave found different mechanisms in eachcommodity (figs, walnuts, grains ...) leading topractical different means for prevention.
Harvest-Processing-Drying: as a resuIt of morethan thirty years studies on mechanisms. ofmycotoxic contamination of different grains,fruits, forages and their prevention, we considerthat this "pre-harvest period" (Le Bars et al, 1994)comprises frequently the most critical steps orpoints.Maturity state at harvest, particularly for fruits,determines greatly the risk of physical lesions andconsecutive fungai contamination (Bouda et ai,1994). The best compromise results from a close

cooperation with the specialists ofthe commodity.Freshly fallen berries, insect damaged and splitfruits have to be collected separately and driedwithout delay. May be, as for other commodities,mats under the tress would reduce fungai infectionof the fruits from the soi I.For most of grains and fruits, the intermediarypost-harvest period before drying (01' otherstabilization procedures) has to be as short aspossible. Sometimes, when drying process isdelayed 01' transport is long, volatile fungistaticsmay be used to prevent fungai development easilydetected by self heating). The effects of mainparameters, along wet 01' dry processing of coffee,on molds and mycotoxins contamination have tobe studied. Such research is difficult beca usemany parameters, variable in time and space,interfer. Usually fruitfull investigations comprisetwo approaches: na analytical one on models, asynthetic one on pilot scale. They require a veryclose collaboration between complementarypartners (mycologists, technologists, analysts, ...).At each step, it is beneficent to sort out foreignmaterial and damaged berries 01' grains.Drying should be done as soon as possible tominimize the three main parameters (duration,free water, temperature) effects. For certaincommodities, when drying equipment istemporarily insufficient, it is sometimesperformed in two steps: a first one to decreasewithout delay moisture content to about 20%,followed by a slightly delayed one to obtain awater content allowing safe storage.
Storage: during storage, a commodity issomewhat in equilibrium with interparticularwater vapor. So the notion of EquilibriumRelative Humidity (ERH%) 01' water activity(Aw) may be used as a determinant parameter for
442
fungai development initiation. The Aw Iimit forinitial growth of A ochraceus is 0,76, whichcorresponds to 14,2% moisture content (Me) onwet weight basis (w.b.) for green coffee beans(Figure 3).They are respectively 0,81 and 15,5% MC (w.b.)for P. verrucosum. But, for several monthsstorage, to avoid any funga! evolution (starting byxerophilous species as A. glaucus groupconducing with time to more hydrophilous onesincluding toxigenic spacies), it is recommended tostore grains at a maximal 0,65 Aw, correspondingto 11,8% MC (w.b.) for green coffee (Figure3).During experimentation onlong term storage (8months) of green coffee under topical climate andin different industrial conditions, the average dataon MC and Aw fit the predictive model relativelywell (Bucheli et al, 1998). The storage conditionswas 13,5% MC (dry basis, ca 11,8 MC w.b.) and71% ERH: in no cases, increase in OTA amountwas observed (average OTA of 3,9 ug/kg at thebeginning of storage). Microbiological evaluationand OTA data during different industrial storageconditions in this study favor the idea that OTAcontamination, as frequently for numerous othermycotoxins in grains and fruits, occurs in thecourse of steps before storage.For long term storage of a lot, homogeneity in thephysico-chemical characteristics of thecommodity and in the hydro-therrnic conditionsare highly recommended. When storage of ainsufficient dry product goes wrong it is typicallydue to heterogeneous conditions: local differencesin temperature in a lot (i.e. because sunny side)leads to vapor migratíon and initiation of fungaIdevelopment (for example, for a 10° differencebetween si lo sides, the safe storage period ofcompound feeds for anirnals is divided by two).
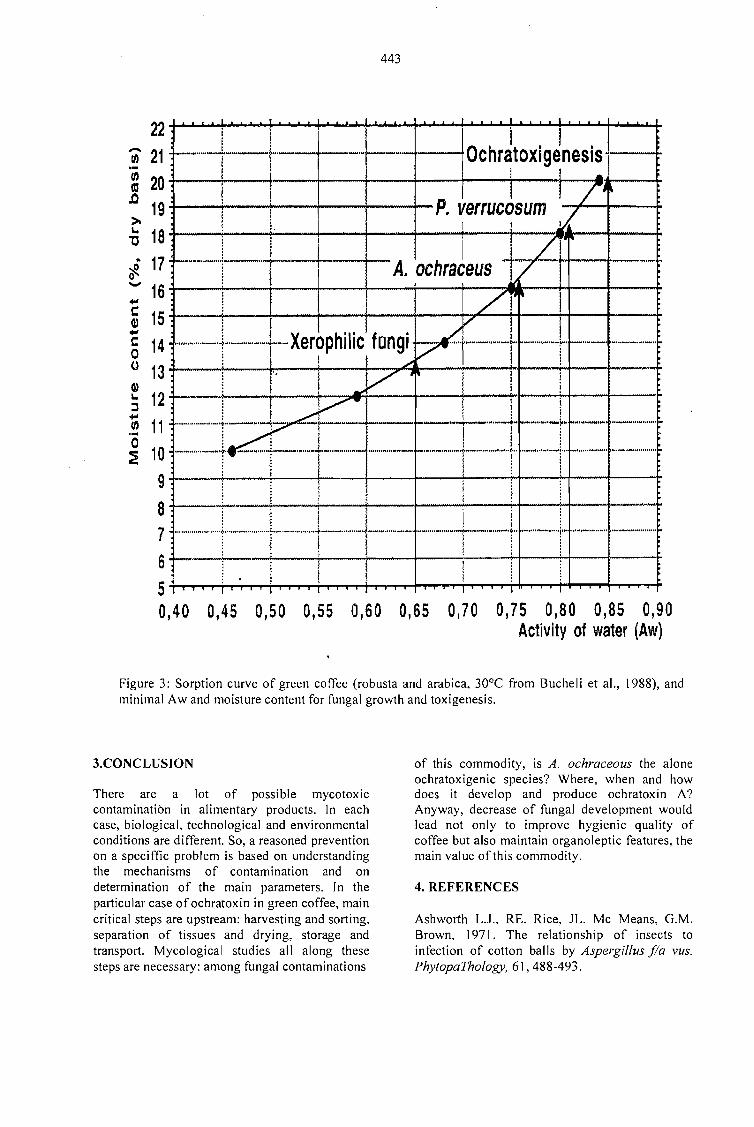
443
I
I
,I
I
0,75 0,80 0,85 0,90Activity of water (Aw)
I
0,700,65I
0,60I
0,55I
0,50I
0,45
8 T:: j 1 '[.: i l! . i7 ·..· ·..·····..··+·..· · ··(..··..· ·..r·· ···· ·1..· ·..·..··..· · · ·· r·..·_·..····..·..~·· .
6~._~.~-----+---+--t----:-----.-t---H---t---ri!5~'I""T"'I-i-r"'T"'T""....-i-T...,...,..,...;-,-rT""T""l-r-T""'T~"'T"'T""P"'T""'i'...,.-'T"T""T"'Í...,....,....,.-'T"'t""I'-t...,......'Í"""T""r-T""'T-r
0,40
22 I , I
~ 21 f-._..~.....+---~..--l--..~.._...-+-----t ochr~toxig~neSiS"i1---I-.~ 20 ! I I ... 1
.o 1 I I /h>- 19 j L I P. ~erruc~sum IJ/-C 18 +---i--~--+---t---+----'l~-"'Í""""'-"".'''''--t---+
j i I I 1 /T"i 17 f-·· ..••........·i-····-_··T\c....--+-- - A. ochraceus l'7-'''rHI --t----:: 16 :. '~, I
cQl 15 +-----+---t----4---+-----t""'7/~-!'+-_M---t----to:! I b v 1 I.., ~ s • • • I ,
c 14 _..+ _..'1' Xerophlllc frmgl' ..,. ..·..t-·_..·..-+·t-· ······..r .~ ~. ~ _~ -..o ~! I I ./, . 1 !o 13 +----!'---+.:---t---t-~ .........--r---t;---H--r--"'t
1 j 1./ ,~ i !I' lOJ }!..Á'" l ''" 12 +--~-_+_-_+_~._+_-_+--+-~-t--H---r---t'
:~)~L. ': I~ : l: !! i..... :; I:;rn 11 ·..······· ·_..-·····l··_· .-r----' -I--•••..,..-_.•...- I"-_ _~.I" - _ ...• ;'. li! lo i: I I l:2: 1O 1' •·•··• 1"· ··· ·+1··· ·..··_· on __•••• _ f"_ _ ! ·..·..·..·..i..· ..
~ ~ ! ; ;
Figure 3: Sorption curve of green coffee (robusta and arabica, 30°C from Bucheli et aI., 1988), andminimal Aw and moisture content for fungai growth and toxigenesis.
3.CONCLUSION
There are a 10t of possible mycotoxiccontamination in alimentary products. In eachcase, biological, technological and environmentalconditions are different. So, a reasoned preventionon a speciffic problem is based on understandingthe mechanisms of contamination and ondetermination of the main parameters. In theparticular case ofochratoxin in green coffee, maincriticai steps are upstream: harvesting and sorting,separation of tissues and drying, storage andtransporto Mycological studies all along thesesteps are necessary: among fungai contaminations
of this cornmodity, is A. ochraceous the aloneochratoxigenic species? Where, when and howdoes it develop and produce ochratoxin A?Anyway, decrease of fungaI development wouldlead not only to improve hygienic quality ofcoffee but also maintain organoleptic features, themain value ofthis commodity.
4. REFERENCES
Ashworth L.J., RE. Rice, JL. Mc Means, G.M.Brown, 1971. The relationship of insects toinfection of cotton balls by Aspergillus fia vus.Phytopa'Ihology, 61,488-493.

Aucamp 1.L., 1969. The role ofmite vectors in thedevelopment of atlatoxin in groundnuts. J StoredProd. Res., 5,245-249.Beti 1.A. Phillips T.W .. Smalley EU., 1995.Effects of maize weevi1s on production ofatlatoxin B I by Aspergillus jlavus in stored corn.J Econ. Entomol, 88, 1776-1782.Blanc M., A Pittet, R. Munoz-box, R. Viani,1998. Behavior of ochratoxin A during greencofTee roasting and soluble coffee manufacture. JAgrie. Food Chem., 46,673-675.Boudra H., 1. Le Ears, P. Le Ears, 1. Dupuy, 1994.Time ofAspergillus jlavus infection and aftatoxinformation in ropening of figs. Myeopathologia,127.29-33.Boudra H., P. Le Bars, .1. Le Bars, 1995.Thermostability of ochratoxin A in wheat undertwo moisture conditions.. Appl. Environ.Microbiol., 61, 1156-1156.Bucheli P., I. Meyer, A. Pittet, G. Vuataz, H.Vian-, 1998. Industrial storage of green robustacoffee under tropical conditions and its impact onraw material quality and ochratoxin A contento JAgrie. Food Chem., 46,4507-4511.Cole RJ.. PA. Hill, PD. Blankenship, T.H.Sanders, K.H. Garren, 1982. Intluence ofirrigation and drought stress on invasion byAspergillus f/a vus of corn kernels and peanutpods. OevIneA Mierotiot, 23, 229-236.Diener UL., N.D. Davis, 1977. Atlatoxinformation in peanuts by Aspergillus jlavus. Agrie.Exp. Sta., Auburn Univ., BulI. n0493.Escoula L., 'I. Le Bars, 1975. Production depatuline par Byssoehlamys nivea sur un modeledensilage. Ann. Reeh. Vet., 6. 219-226.Frisvad 1.C., 1984. Expression of secondarymetabolism as fondamental characters inPenieillium taxonomy. In Toxigenic Fungi, ed. H.Kurata and Y. Ueno, Elsevier, Amsterdam, 98106.Hill RA., PD. Blankenshíp, RJ. Cole, Tb.Sanders, 1983. The effects of soíl rnoisture andtemperature on preharvest invasíon of peanuts byAspergilfus jlavus and subsequent atlatoxindevelopment. Appt Environ. Mierobiol., 45, 628633.ARC, 1993. Ochratoxin A. Monograph on theevaluation of carcinogenic risks to humans (Lyon:Internat. Agency Research Cancer), 489-521.Le Bars 1., 1982. Facteurs de 'accumulationdacide p--nicillique dans les denrdes doriginevdg6tale. SoL Ailments, 2 (h.s 11),29-33.Le Bars 1., 1988.Toxigenesis as a fonction of theecological conditions of the grain/microorganismssystem. In "Preservation and storage of grains,seeds and their by-products", J.LMULTON,Lavoisier pub. New York, Paris, 347-366.
444
Le Bars U., 1990. Contribution to a practicalstrategy for preventing atlatoxin contamination ofdried figs. Mierobiologie- Ailment -Nutrition, 8,265-270.Le Bars 1.. P. Le Bars. J. Dupuy, H. Boudra, 1994.Biotic and abiotic factors in Fumonisin Biproduction and stability. JA.o.A.C InternaL, 77,517-52LLe Bars P., 1. Le Bars, 1995. Ecotoxinog6n6se duFusarium mon//itorme. Enveloppe des risques desFumonisines. Crytogamie- %4yeofoe. 'ie. 16, 5964.Le Bars J., P. Le Bars, 1996. Recent acute andsubacute mycotoxicoses recognized in France.Veto Res.. 27,383-394.Le 8ars U" P. Le Bars, 1998. strategy for safe useof fungi and fungaI derivatives in food processing.Vet, Res., 149,493-500.Levi C., H.L. Trenk. H.K. Mohr, 1974, Study ofthe occurrence of ochratoxin A in green coffeebeans. JA. CA. c. 57,866-870.Moss MO.. 1996. Mode of formation ofochratoxin A Food Add. Coot, 13, (suppl). 5-9.Naidu R., S Daivasikamani J.S. Nagaraj, 1997.Current status and strategies for management ofmycotoxins in 1ndia coffeesln 17th Col I. Scientif.sur le caf6, ASIC (ed), Nairobi. 20-25 107/1 997,p.69-78.Northolt M.D., H.P. Van Egmond, WE. Paulsch,1979; Ochratoxin A production by some fungaispecies in relation to water activityandtemperature. JL Food Proteetion ,42,485-490.Faster N., N. Lisker, I. Chei, 1983. Ochratoxin Aproduction by Aspergillus oehraeeus grown undercontrolled atmospheres. Appi. Environ. Mierobiot45, 1136-1139.Patel S., CM. HazeJ, A.C.M Winterton, AE.Gleadle, 1997. Survey of ochratoxin A in UKretai I coffees. Food Add. Cont,14,217-222.Pitt. J.I.. 1937. Penicillium viridica turn, F.verrucosum, and the production ofochratoxin A. AppL Environ. Mierobiot, 53, 266269.28. Ramos AJ., N. Labernia, S. Mann, V.Sanchis, N. Magan, 1998. Effect of water activityand temperature on growth and ochratoxinproduction by three strains of Aspergillusoehraeeus on a barley extract medium and onbarley grains. Int. J Food mierobioL, 44, 133140.29. Sanders T.H., R.A. Hill, RJ. Cole. P.D.Blankenship, 1981. Effect of drought onoccurrence of Aspergillus jlavus in maturingpeanuts. J Am. GIL Cem. Soe.. 58,9661 -9670.30. Steyn P.S. 1980.The biosynthesis ofmycotoxins , 432 pages, Acad. Press.

31. Steyn P.S., 1998. The biosynthesis ofmycotoxins. Rev. Med. v«, 149,469-478.32 Studer-Rohr J , D R. Dretrich, 1. Schlatter, e.Schlatter, 1995. The occurrence ofochratoxin A incoffee. Fd. Chem. Toxicol., 33, 341-355.33. Tsubouchi H., H. Terada. K. Yamamoto. K.Hisada, Y. Salabe, 1938. Ochratoxin A found incommercial roast coffee. I Agric. Food Chem., 36,540-542.34. Turner W.B. 1971. Fungai metabobtes. 426pages, Aced. Press, London.35. Tumer WE. And Alridge D.e. 1983. Fungaimetabolites lI, 631 pages, Acad. Press, London.36. Van der Stegen G., U. Jorissen, A. Pittet, M.Saccon, W. Steiner, M. Vincenzi. M. Vinkler, J.Zapp, e. Schiatter, 1997. Screning of Europeencoffee final products for occurrence of ochratoxinA. Food Add.Cont, 14,211-216.
445
37. Vega FE., 2. Mercadier, 1998. Insects, coffeeand ochnatoxin A. Florida Entomologist, 81, 543544.38. Viani R., 1996. Fate of ochnatoxin A duringprocessing of coffee. Food Add. Coot. 13, (suppI),29-33.39. Walker P., 1997. Quality and safety of coffee.17 th Coi!. Scient,cafd. ASIC Intemat. sur le (Nairobi 1997), .51-60.40. Widstrom NW., EB. Lillehoj, AN. Sparks,WE. Kwolek. 1976. Com damage and aflatoxinon com ears protected with insecticide. I Loon.Entornot. 69,677-679.41. World Health Organisation, 1996. Evaluationof certain food additives and contaminants. 44thReport of JECFA. WHO Technical Report seriesn.o859(Geneva).

j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j

447
OCRATOXINAS E AFLATOXINAS EM CAFÉ BRASILEIRO
Lucia Valente Soares
Professora Livre Docente da Universidade Estadual de Campinas, Faculdade de Engenharia de AI imentos,Caixa Postal 6121, 13081-970 Campinas, S.P.
INTRODUÇÃO
Fungos ou bolores tem sido amplamente usadospela humanidade na produção e preservação dealimentos, produção de matéria prima emedicamentos. Porém, como uma conseqüênciada luta pela sobrevivência dos fungos em seumicroarnbiente, substâncias tóxicas são por elesproduzidas e algumas delas altamente tóxicas aohomem e aos animais. São as micotoxinas.Dentre estas, a ocratoxina A e as aflatoxinas temsido encontradas em café (Levi, 1980; Nakajimaet ai, 1997; Micco et al., 1989). Organismosinternacionais como o Codex Alimentarius temdiscutido a possibilidade de sugerir limites paraocratoxina A em café (Codex Alimentarius,1998). A Itália adotou, desde 1996, o limite de 4ng/g de ocratoxina A para café verde (Ministeriodelle Finanze, Itália, 1996).O Brasil como grande produtor de café éespecialmente afetado por estas descobertas, tantoa nível de saúde pública pelo alto consumo dabebida dentre sua população, como a nível decomércio exterior no qual o café representa umítem importante. Torna-se assim importante umaanálise do assunto para que possamos nos situar evisualizar os caminhos a tomar diante doproblema.
UM POUCO SOBRE AS TOXINAS
As ocratoxinas são produzidas principalmente porcepas de Penicillium verrucosum e de váriasespécies de Aspergillus, entre estas A. ochraceus.As evidências atuais indicam que P. verrucosum éencontrado em climas temperados e A. ochraceusem climas quentes. Estas toxinas compreendemuma família de 7 compostos porém, apenas aocratoxina A tem sido encontrada comocontamínante importante em alimentos (Moss,1996).A ocratoxina A é agudamente tóxica parapássaros, mamíferos e peixes. Seu alvo principalsão os rins e secundariamente. o fígado. Étambém imunotóxica e teratogênica (Busby eWogan, 1981). A "International Agency forResearch on Cancer" classificou a ocratoxina Ano grupo 2B de substâncias, o que signi fica quehá evidência suficiente de sua carcinogenicidade
em animais. permitindo classifica-Ia comopossivelmente carcinogênica para humanos(JARC, 1993). Ainda em humanos, a ocratoxinaA tem sido correlacionada com a nefropatiaendêmica dos Balcãs, doença degenerativa dosrins observada na população desta região e queem casos extremos leva à morte (Plestina, 1996).Mais recentemente, nefropatias tem sido relatadasna Algéria e na Tunísia e a toxina tem sidoencontrada em fluídos corpóreos da populaçãoafetada (Creppy et al., 1993). A meia vida daocratoxina A no organismo humano foi estimadaem 35 dias (Schlatter, 1996). Na União Europeiae na América do Norte há indicações que osalimentos mais frequentemente contaminadoscom ocratoxina A são alguns cereais e produtos àbase de carne e vísceras de suínos. Atualmentevinho. cerveja e café estão sendo examinadoscomo possível fonte de contaminação (KuiperGoodman. 1996). Dentre os cereais. a cevada é omais frequentemente contaminado, seguida porcenteio e aveia (Poh land et al., 1992).As aflatoxinas B I, B], G 1 e G] são rnetabólitos dosfungos Aspergillus flavus e A. parasiticus. Sãoaltamente tóxicas para mamíferos. primatas nãohumanos, aves e peixes. Seu alvo principal é ofígado. São também teratogênicas, rnutagênicas ea atlatox ina B I está entre os compostoscarcinogênicos mais potentes conhecidos (Busbye Wogan. 1981). A "International Agency forResearch on Cancer" classificou a aflatoxina B I
como um carcinógeno humano (Grupo 1) (IARC.1993). As aflatoxinas tem sido encontradas emdiversos tipos de alimentos. principalmenteamendoim, alguns tipos de nozes e castanhas.semente de algodão. milho e figos. Não há dadosem humanos quanto à persistência da aflatoxinano organismo. Em macacos a maior parte datoxina é eliminada em 2 - 3 dias via urina e fezes.O fígado porém retém cerca de I% e resíduos sãoainda detectados na urina 5 semanas após aingestão da toxina (Busby e Wogan. 1981).
PRODUÇÃO E ESTABILIDADE DAOCRATOXINA A E DA AFLATOXINA B,
A presença de A. ochraceus e ocratoxina A emcafé foi primeiro relatada em 1974 (Levi et al.,1974). Mais adiante, a microbiota fúngica de 944amostras de café verde importada pelos EUA foi

examinada e os autores do trabalho relataram umainvasão interna de 80,5% das amostras asiáticas eafricanas contra 49,4% das amostras das AméricasCentral e do Sul. O gênero Aspergilluspredominou e incluiu espécies toxigênicas cornoA. ochraceus, A. jlavus e A. versicolor. O gêneroAspergillus foi também encontrado e incluiuespécies toxingênicas como P. cyclopium, P.citrinum e P. expansum (Mislivec et al., 1983).Estes estudos alertaram para a possibi lidade docafé conter micotoxinas,Em meios sintéticos. o A. ochraceus apresenta
uma temperatura ótima de produção de 25 De. Afaixa de temperaturas em que ocorre produção é12 - 37 De. A atividade de água ótima paraprodução da toxina é acima de 0,97 e a mínima é0.79 (Moss, J996). Porém. os dados atualmenteexistentes sobre as condições para produção deocratoxina A em café são extremamente pobres.Em um trabalho recente. realizado na Tailândia.acompanhou-se por 8 meses o armazenamento decafé em si los com aeração. sem aeração e com arcondicionado e ainda em sacas em armazéns. Foiconstatado que não houve produção da toxinadurante o armazenamento. Parte do café jácontinha ocratoxina A e os níveis não aumentaramdurante os meses de estocagem. Concluíram osautores que a produção da toxina ocorreu antes doarmazenamento (Bucheli et al., 1998). Apassagem da maior parte da toxina presente nocafé torrado para a bebida durante sua preparação(Tsubouchi et al., 1987; Blanc et al., 1998) temsido motivo de preocupação. Olhou-se então paraos efeitos da torrefação corno uma possibilidadede redução do teor da toxina. A terrnoestabilidadeda ocratoxina A foi estudada em trigo a diferentestemperaturas. A 200DC houve uma destruição de50% com 12 minutos em trigo seco e 19 minutosem trigo molhado. A 250 DC e em trigo seco.conseguiram os autores a eliminação de 50% da
448
toxina em 6 minutos. A toxicidade dos produtosde degradação não foi verificada (Boudra et aI.,1995). São resultados que apontam umaestabi Iidade térmica relativamente alta para aocratoxina A. Em café, existem controvérsiassobre o efeito da torrefação como possível meiode eliminação da toxina. Por um lado, hátrabalhos que apontam destruição da maior parteda toxina, de 80% ou mais. e outros quedescrevem a torrefação como pouco eficiente.capaz de reduzir a toxina em apenas 12% oumenos ( Levi et al, 1974; Cantàfora et aI., 1983;Tsubouchi et al., 1987; Studer-Rohr et aI., 1995;Blanc et al., 1998).Não há relatos sobre condições para produção deaflatoxinas em café. A.jlavus pode produziraflatoxinas entre 12 - 42 De. As temperaturasótimas estão no entanto entre 28 e 30De. Emamendoim a produção de aflatoxinas atinge omáximo entre 15 e 35 % de umidade (Rustorn,1997). Não há também estudos que verifiquemse as aflatoxinas passam para a bebida durante oseu preparo. Apenas dois trabal hos tentaramdeterminar a destruição da aflatoxina B I com atorrefação. Ambos afirmam haver uma destruiçãode 80 - 99% da toxina (Levi, 1974; Micco et al.,1992).
INCIDÊNCIA DE OCRATOXINA A EAFLATOXINAS EM CAFÉ
Levantamentos sobre a incidência de ocratoxina Aem café na literatura científica. são relativamentepoucos e na sua maioria recentes. Na Tabela 1temos um resumo dos levantamentos realizadosem diferentes países em amostras de café verde.torrado e solúvel. Levantamentos sobre aincidência de aflatoxinas em café são ainda maisraros(Tabela 2.)
Tabela I. Níveis de ocratoxinaA encontrados por países importadores em café procedente de várias partes do mundo.
Tipo de café Procedência Amostras Faixa Média Referência
Total Contaminadas (~g/kg) (~g/kg)
Verde Diversa 29 17 0.2 - 15.0 3.1 Micco et al., 1989.
Verde Diversa 47 14 0.2 - 1704 2.6 Nakajirna et ai.. 1997
Verde Am. do Sul 19 9 0,1 - 4.6 IA Trucksess et al..1999
Torrado Diversa 40 14 004- 4.2 1.2 Studer-Rohr et aI.. 1995.
Torrado Diversa 11 II s 3.2 0.51 Jorgensen et al., 1997
Torrado Diversa 484 s 8.2 0.8 Stegen et ai.. J997
Torrado Diversa 20 17 s 2.1 0.6 Patel et ai.. 1997
Torrado Am. do Sul 13 9 0,1 - 1.2 004 Trucksess et ai.. 1999
Solúvel Diversa 101 75 ~ 6.5 1.0 Pittet et al., 1996.
Solúvel Diversa 80 64 ~ 8.0 1.0 Patel et al., 1997
Solúvel Diversa 149 ~ 7.8 1.3 Stegen et ai.. 1997

449
Tabela 2. Níveis de aflatoxina B I encontrados por países importadores em café procedente de várias partes do mundo.
Tipo de café Procedência Amostras Faixa Média Referência
Total Contaminadas (~g/kg) (ug/kg)
Verde Diversa 47 14 2,0- 32,9 7.8 Nakaiima et al., 1997.
Verde Diversa 1365 2 3 - 12 Levi, 1980.
Quando na faixa de contaminação é indicado umvalor inferior, significa que a média refere-seapenas às amostras contaminadas. Quando não háindicação de valor inferior, a média foi calculadaincluindo todas as amostras (contaminadas e nãocontaminadas).
OCRATOXINA A E AFLATOXINA B. EMCAFÉ BRASILEIRO
Os dados, ainda que parcos, sobre café verdebrasileiro analisado pelos países compradores,mostram uma situação bastante favorável, compoucas amostras contaminadas e teores baixos deocratoxina A (Tabela 3). O mesmo ocorre comrelação à aflatoxina B I (Tabela 4).Os dados colhidos na UNICAMP (Tabela 5)mostram uma contaminação média de 2,4 ug/kgde ocratoxina A em 142 amostras de café verde.Este valor está dentro dos encontrados por
levantamentos realizados por países importadoresem amostras de café provenientes de diversaspartes do mundo (Tabela I). A incidência deamostras contaminadas no levantamento daUNICAMP foi de 21 % e compara favoravelmentecom os dados da Tabela I para café verde, ondeincidências de 30%, 47% e 59% são relatadas(Nakajima et al., 1997; Trucksess et aI., 1999;Micco et aI., 1989).Trabalho realizado por Furlani (1998) e Leoni(1999) através de cooperação entre a UNICAMP ea Cia. Cacique Café Solúvel. As amostrascorrespondem a café ofertado no comércio e nãotem relação com café adquirido pela Cacique parasua produção de café solúvel. As amostras de C.arabica procediam dos estados de Minas Gerais,Paraná, Espirito Santo, São Paulo e Bahia e asamostras de C. canephora, dos estados de EspíritoSanto, Bahia e Rondônia.
Tabela 3. Níveis de ocratoxina A encontrados em café procedente do Brasil por países importadoresTipo de café Amostras Faixa Média Referência
Total Contaminadas (ug/kg) (ug/kg)
Verde 8 O Cantáfora et aI., 1983
Torrado 9 O Tsubouchi et aI., 1988
Verde 14 10 0.2 - 5,5 2.0 Micco et aI. 1989
Verde 10 O Nakajima et aI.. 1997
Tabela 4. Níveis de allatoxina BI encontrados em café procedente do Brasil por países importadoresTipo de café Amostras Presença de A. flavus Referência
Total Contaminadas
Verde 9 O 1 Nakajima et aI.. 1997.
Tabela 5. Níveis de ocratoxina A em amostras de café verde brasileiro oferecido no mercado interno nos anos de 1997e 1998.
Amostras Média (ug/kg)
Espécie Total Contaminadas Faixa (ug/kg) do total dascontaminadas
C. arabica 119 27 ~47.8 1.9 8,4
C. canephora 23 3 ~114.2 5.3 40.4
Total 142 30 ~I 14.2 2.4 11,6

IMPACTO DO CAFÉ NA INGESTÃO DEOCRATOXINA A ATRAVÉS DA DIETA
Em 1995, o JECFA (Joint Expert FAO/WHOCommittee on Food Addtives) avaliou aocratoxina A e estabeleceu um PTWI (ProvisionalTolerable Weekly Intake = ingestão semanaltolerável provisória) de 100 ng de ocratoxina Apor kg de peso corpóreo (ng/kg p.c.). Apósestudos, este comitê de especial istas concluiu queesta PTWI pode ser adotada provisoriamente poisdiante dos dados atuais este nível parece nãoapresentar riscos de causar nefropatias emhumanos (Codex Alimentarius, 1999). Este valorcorresponde a um PDTI (Provisional DailyTolerable Intake = ingestão provisória diáriatolerável) de 14 ng/kg p.c. Note-se que o comitênão levou em consideração o papel da toxinacomo carcinógeno.No Canadá, uma avaliação de risco com relação àocratoxina A sugeriu que o valor da PDTI ficasseentre 1,2 e 5,7 ng/kg p.c.. O Conselho deMinistros dos Países Nórdicos, por sua vez,escolheu uma TDI (Tolerable Daily Intakeingestão diária tolerável) de 5 ng/kg p.c. e oComitê Científico da Comissão Europeia paraAlimentos sugeriu que a TDI ficasse abaixo de 5ng/kg p.c. levando em consideração a atividadecancerígena da toxina (Kuiper-Goodman, 1996).Na União Européia, 8 países analisaram amostrasde café recolhidas em lojas e mercados. Combase nos resultados e na estimativa de que oeuropeu consome em média 4 xícaras de café pordia (o que eqüivaleria a 24 g de café torrado ou 8g de café instantâneo) calcularam que a ingestãode café seria responsável apenas por 2% do PTWI(Stegen et aI., 1997). Na Suíça, considerandouma ingestão de 3 a 4 xícaras de café por dia euma contaminação média do café de I ug/kg,calculou-se a ingestão média de ocratoxina Aatravés do café seria de 0,4 ng/kg p.c. por dia,portanto correspondendo a 2,5 % do PTWI(Studer-Rohr et al., 1995). Uma análise relataque um dinamarquês médio ingere cerca de 821mL de café por dia e que para cada xícara de 170mL gasta-se 6 g de café torrado. O dinamarquêsadulto médio de 70 kg consumiria 0,2 ng/kg p.c.por dia. Este valor corresponde a 12,5% do PTWI(Jorgensen, 1998). Na Inglaterra, o consumidormédio consome 4,5 g café por dia no preparo dabebida e o grande consumidor 19,3 g ,correspondendo a 0,4 ng/kg e 1,9 ng/kg p.c. porsemana, respectivamente (Pate1 et al, 1997).Portanto, o consumidor inglês em média estariaexposto a ingerir 0,4% e o grande consumidor1,9% do PTWI para ocratoxina A através do café.
450
Levantamento realizado na regrao de Campinas,Estado de São Paulo, mostrou que a parcela dapopulação adulta que consume café, ingere emmédia 284 mL por dia. Isto corresponde a cercade 3 a 4 cafezinhos, ou seja um total de 25 g decafé torrado e moído e preparado ao gostobrasileiro (Camargo, 1996). Assumindo que estamédia seja válida para o resto do país econsiderando um adulto hipotético de 70 kg ,podemos calcular a ingestão média semanal .Com base no dado que atualmente dispomos sobreo café verde vendido internamente no Brasil(Tabela 5) de que a contaminação média comocratoxina A é 2,4 ug/kg teríamos uma ingestãosemanal da toxina de 6 ng/kg p.c. através doconsumo de café. Isto assumindo ainda o casoextremo de que não ocorram perdas durante oprocessamento, o que diga-se de passagem, não éprovável. Este adulto estaria consumindo 6% doPTWI.O JECFA diante do problema de ocratoxina A emcafé, questiona se a ingestão de ocratoxina A emcafé é significativa para os consumidores a pontode merecer limites específicos. Lembra ainda asdiscrepâncias de resultados sobre perdas deocratoxina A durante a torrefação (CodexAlimentarius, 1999), assunto este que aindademanda estudos mais completos.
UMA AVALIAÇÃO DA SITUAÇÃO DOCAFÉ BRASILEIRO
Podemos resumir reconhecendo que o problemade contaminação do café brasileiro comocratoxina A não pode ser ignorado. Os dadosatuais indicam, no entanto, que não se mostracrítico para a população interna consumidora dabebida, caso o café seja a única ou a maisimportante fonte de ocratoxina A na dietanacional. A ingestão, calculada com base nacontaminação média em café verde e assumindoque não haja perdas durante o processamento,corresponderia a 6% da ingestão semanaltolerável provisória (PTWI) estabelecida pelaFAO para ocratoxina A. O aspecto de saúdepública para nossa população não pode, noentanto, ser descurado por estarmos tratando comtoxinas cancerígenas. Assim sendo. fazem-senecessários levantamentos mais extensos queforneçam um quadro mais completo da situaçãodo café produzido no país, inclusive da variedadeC. canephora, da qual poucas amostras foramexaminadas até agora.Do ponto de vista de exportação, o café nacionalevidentemente não apresenta problemas maioresquando comparado com o produzido em outraspartes do mundo. Os dados atuais indicam

inclusive uma situação ligeiramente favorável aocafé brasileiro. No entanto, o café exportado estásujeito às regulamentações dos paísesimportadores. Alguns países adotaramrecentemente limites para ocratoxina A em caféverde. Em 1996, a Itália estabeleceu, a título de
recomendação, o valor de 4 ug/kg como limitepara ocratoxina A em café verde (Ministerio delle
Finanze, 1996). Já a Grécia adota 20 ug/kg comolimite para café verde. A República Checaregulamenta em 20 ug/kg e a Romênia em 5
ug/kg , o limite para ocratoxina A em todos osalimentos (FAO, 1996). Seria arriscado ignoraresta tendência dos países importadores emestabelecer limites e as possíveis barreirasalfandegárias que daí adviriam.O assunto como um todo merece estudos maisdetalhados que preparem este segmento produtivopara o problema. Medidas preventivas são asmais recomendáveis em se tratando decontaminantes. Estudos que revelem em queetapa a contaminação ocorre e que fatores podemcontrola-Ia são necessários. O efeito da torrefaçãoe de outras etapas do processamento precisamtambém ser melhor estudados.
BIBLIOGRAFIA
Blanc, M., Pittet, A., Munoz-Box, R. e Viani R.1998. Behavior of ochratoxin A during greencoffee roasting and soluble coffee manufacture.J. Agric. Food Chem. 46: 673 - 675.Boudra, H., Le Bars, P. e Le Bars, J. 1995.Thermoestability of ochratoxin A in wheat undertwo moisture conditions. Appl. Environ.Microbiol. 61 :1156 - 1158.Bucheli, P., Meyer, I.. Pittet, A., Vuataz, G. eViani, R. 1998. Industrial storage of green robustacoffee under tropical conditions and its impact onraw material quality and ochratoxin A content. 1.Agric. Food Chem. 46: 4507 - 451 I.Busby Jr., W.F. e Wogan, G.N. 1981.Ochratoxins. In Shank, R.C. (ed.) Mycotoxins andN-Nitroso Compounds: Environmental Risks.Vol 11., CRC Press, Boca Raton, FL.Camargo. M.C.R. 1996. Avaliação da ingestãopotencial de cafeína pela população de Campinas.Tese de mestrado UNICAMP.Cantàfora, A" Gossi, M., Miraglia, L. e Benelli,L. 1983. Determination of ochratoxin A in coffeebeans using reversed- phase hígh performanceliquid chromatography. Riv. Soe. Ital. Sei. Alim.12: 103 - 108.Codex Alirnentarius. 1999. Positionpapel' on ochratoxin A. Codex Comittee on FoodAdditives and Contaminants, Roma, Itália.
451
Creppy, E., Castegnaro, M. e Dirheimer, G. 1993.Human ochratoxicosis and its pathologies, JohnLibbey Eurotext, França.FAO. 1996. Worldwide regulations formycotoxins. FAO Food and Nutrition Papel',Food and Agriculture Organization, Roma,Itália.Furlani, R.P.z. 1998. Ocratoxina A em cafébrasileiro, tese de mestrado, UNICAMP.Jorgensen, K. 1998. Survey of pork, poultry,coffee, beer and pulses for ochratoxin A. FoodAdd. Contam. 15: 550-554.IARe. 1993. Some naturally occurringsubstances; food items and constituents;hetrocyclic aromatic amines and mycotoxins,International Agency for Reseach on Cancer,Geneva, Suíça. Kuiper-Goodman, T. 1996. Riskassessment o ochratoxin A: an update. FoodAdd. Contam. 13 (Supplernent): 53-57. Leoni,L.A.B. 1999 (Comunicação pessoal).Levi, C. 1980. Mycotoxins in coffee. J. Assoe.Off. Anal. Chem. 63: 1282-1285.Levi, c., Trenk, H.L. e Mohr, H.K. 1974. Studyof the occurrence of ochratoxin A in green coffeebeans. J. A.O.A.C. 57: 866 - 871.Micco, C; Grossi, M., Miraglia, M. e Brera, C.1989. A study ofthe contamination by ochratoxinA of green and roasted coffee beans. Food Add.Contam. 3:333-339.Micco, C; Miraglia, M., Brera, C, Desiderio C. eMasci, V. 1992. The effect of roasting on the fateof af1atoxin A in artificially contaminated greencoffee beans. Mycotoxin Res. 8: 93 - 97.Mislivec, P.B., Bruce, V.R. e Gibson, R. 1983.Incidence oftoxigenic and ogher molds in greencoffee beans. J. Food Protec. 46: 969 - 973.Ministerio delle finanze, 1996. DípartamentoDogane, Circolare no. 76, Roma, Itália.Moss, M.O. 1996. Mode of formation ofochratoxin A. Food Add. Contam. 13(Supplement): 5-9.Nakajima, M., Tsubouchi, H., Miyabe, M. eUeno, Y. 1997. Survey of af1atoxin BJ andochratoxin A in commercial green coffee beansby high-performance liquid chromatographylinked with immunoaffinity chromatography.Food Agric. Immun.9: 77 - 83.Patel, S., Hazel, C.M., Winterton, A.G.M. eGleadle, A.E. 1997. Food Add. Contam 3: 217 222.Plestina, R. 1996. Nephrotoxicity of ochratoxin A.Food Add. Contam. 13 (Supplement): 49-50.Pohland, A.E.; Nesheim, S. e Friedrnan, L. 1992.Ochratoxin A: a review. Pure Appl. Chem. 64:1029 - 1046.

Rustom, l.Y.S. 1997. Aflatoxim in food and feed:occurrence, legislation and inactivation byphysical methods. Food Chem. 59: 57- 67.Schlatter, c., Studer-Rohr, 1. e Rasonyi, T. 1996.Carcinogenicity and kinetic aspects of ochratoxinA. Food Add. Contam. 13 (Supplement): 4344.Stegen, O.v.d., Jorissen, U., Pittet, A., Saccon, M.,Steiner, W., Vincenzi, M., Winkler, M., Zapp, J. eSch1atter, C. 1997. Screening of Europeancoffee final products for ocurrence of ochratoxinA. Food Add. Contam. 14: 211-216.Studer-Rohr, 1., Dietrich, D.R. Schlatter, J. eSchlater, C. 1995. The occurrence of ochratoxinA in coffee. Fd. Chem. Toxic. 33:341- 355.
452
Tsubouchi, H., Yamamoto, K., Hisada, K.,Sakabe, Y. e Udagawa, S. 1987. Effect ofroasting on ochratoxin A levei in green coffeebeans inoculated with Aspergillus ochraceus.Mycopathology 97: I1I - 115.Tsubouchi, H., Terada, H., Yamamoto, K.,Hisada, K. e Sakabe, Y. 1988. Ochratoxin Afound in commercial roast coffee. J. Agric.Food Chem. 36: 540 - 542.Trucksess, M.W., Giler, 1. e Young, K., White,K.D. e Page, S. 1999. Determination and surveyof ochratoxin A in wheat, barley, and coffee 1997. 1. AOAC Int. 82: 85 - 89.

453
RESULTADOS E DESAFIOS DO PROGRAMA DE AUTO FISCALIZAÇÃO DO CAFÉ TORRADOE MoíDO: SELO ABIC
Celso Luis Rodrigues Vegro', Célia Regina R. P.Tavares Ferreira 2
I EngOAgr" MS. Pesquisador Cientifico de Economia Agrícola da Secretaria de Agricultura eAbastecimento do Estado de SP.2Enga Agr" MS. Pesquisadora Cientifica de Economia Agrícola da Secretaria de Agricultura e
Abastecimento do Estado de SP.
Os resultados das coletas nacionais liberados paradivulgação pública formam o materialfundamental de análise. Foram separadas para esteestudo quatro coletas da ABIC para verificação dapureza: 1996 (duas), 1997 e 1998, publicada emCONTROLE (1996), CAI (1997), MAIOR (1997)e PROGRAMA (1998), respectivamente. Oprocedimento de coleta adotado baseia-se emrotina estabelecida em ABIC 1993, sendorealizada por meio de empresa de auditoriaindependente.
Entre os gêneros alimentícios, o T&M é um dosque não oferece grandes dificuldades para afraude.As "vantagens" da adulteração do produto,são inúmeras, como por exemplo: a) melhorrentabilidade, uma vez que, as impurezas emisturas mais comuns apresentam preços menoresquando comparados com os do café verde (mesmode tipo inferior); b) reduz o número de xícaras,obrigando assim, o comprador adquirir maioresquantidades" (CAMARGO e TELLES, 1953), e c)a adição de açúcar ao grão torrado reduzir oconsumo de energia e a quebra de peso.O café apresenta, historicamente, ciclos de altas ebaixas cotações no mercado. Nos ciclos de alta,tornou-se comum nos países importadoreschancelar as fraudes visando baratear o produto,sendo mantido o critério nas baixas.As torrefadoras que dispõe de equipamentosdefasados ou modernos, porém mal ajustados, aoadquirirem matéria-prima de tipo inferior, nãoconseguem produzir T&M com margensinferiores a 1% de impurezas (cascas, paus,resíduos celulósicos e terra). Dessa forma, seuproduto é considerado adulterado pela legislação3. Essa dificuldade já foi reconhecida pela ABICque elevou seu patamar para verificação da purezaao nível de 2%. Noutras situações, a adição deimpurezas ao café verde é estratégia para baratear
INTRODUÇÃO
o Programa de Autofiscalização do Café,patrocinado pela Associação Brasileira daIndústria do Café (ABlC), foi lançado em 1989,tendo por objetivo melhorar a imagem do T&Matravés da garantia de sua pureza aosconsumidores brasileiros. O elemento chave desseesforço foi a inclusão nas diferentes marcascomerciais fiscalizadas de um selo padrãodenominado: "Selo de Pureza ABIC"(ASSOCIAÇÃO,s.d.).A autofiscalização surtiu resultadosinflexionando a curva do consumo de T&M pois,enquanto em 1989 consumiram-se 6,4 milhõessacas de café verde, estima-se que em 1998consumiu-se 12 milhões.O programa já analisou. no Instituto Adolfo Lutz(IAL), 37 mil amostras de café. Na última coleta,divulgada em setembro de 1998, foram analisadas2.819 amostras. Como resultado tem-se que 95%das marcas de café comercializadas no País sãopuras; e as 5% impuras representam apenas I% docafé consumido no mercado inteiro. Há dez anos,30% das marcas eram impuras (PROGRAMA,1998).
OBJETIVO
O objetivo deste estudo é verificar os resultadosdo "Programa de Autofiscalização de Café daABIC - Selo de Pureza ABIC", a partir de sériehistórica de coletas de amostras de café T&Mpara o período 1996 a 1998 em nível nacional eregional.
MATERIAL E MÉTODO
Chama-se de impurezas toda substância que não écafé, resultante do mau beneficiamento doproduto, como cascas, paus. areia, etc., enquantomisturas são os produtos adicionados ao café parafraudá-lo, como milho, cevada, caramelo, etc. Sãotambém consideradas substâncias estranhasnaturais as bactérias, os fungos, os ácaros e
.fragmentos de insetos (lAL, 1997).
POSSIBILIDADES EFRAUDES NO T&M
PRÁTICA DE
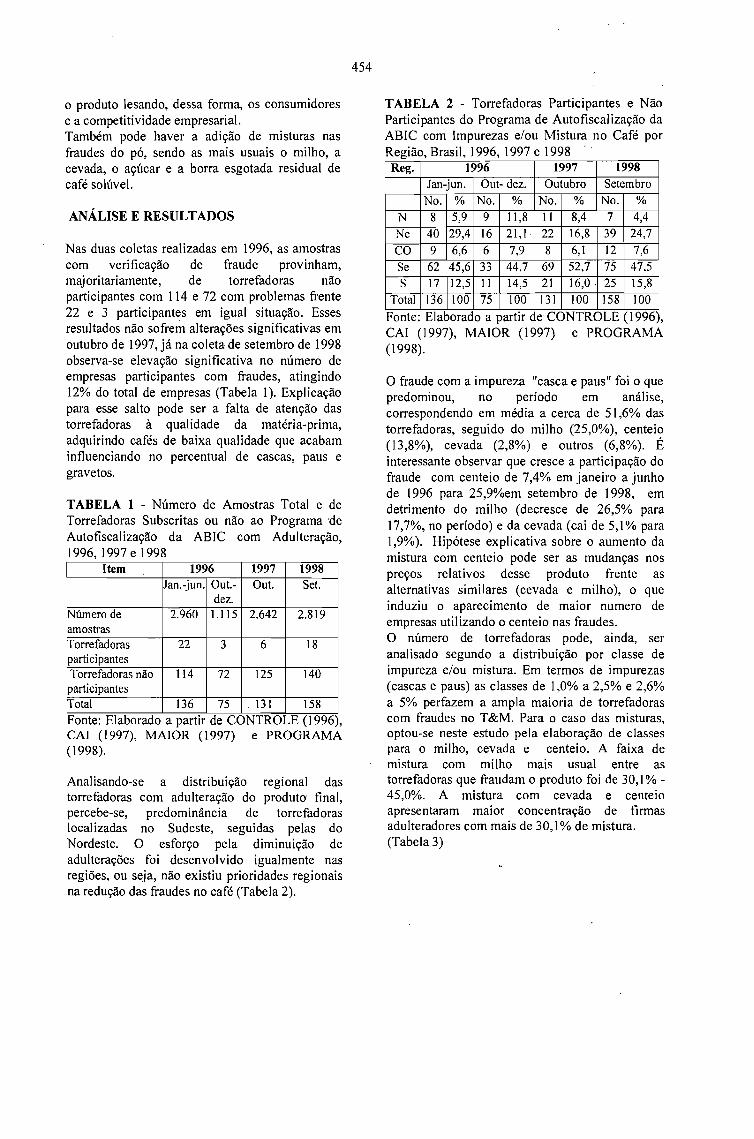
o produto lesando, dessa forma, os consumidorese a competitividade empresarial.Também pode haver a adição de misturas nasfraudes do pó, sendo as mais usuais o milho, acevada, o açúcar e a borra esgotada residual decafé solúvel.
ANÁLISE E RESULTADOS
Nas duas coletas realizadas em 1996, as amostrascom verificação de fraude provinham,majoritariamente, de torrefadoras nãoparticipantes com 114 e 72 com problemas frente22 e 3 participantes em igual situação. Essesresultados não sofrem alterações significativas emoutubro de 1997, já na coleta de setembro de 1998observa-se elevação significativa no número deempresas participantes com fraudes, atingindo12% do total de empresas (Tabela I). Explicaçãopara esse salto pode ser a falta de atenção dastorrefadoras à qualidade da matéria-prima,adquirindo cafés de baixa qualidade que acabaminfluenciando no percentual de cascas, paus egravetos.
TABELA 1 - Número de Amostras Total e deTorrefadoras Subscritas ou não ao Programa deAutofiscalização da ABIC com Adulteração,1996, 1997 e 1998
Item 1996 1997 1998Jan.-jun. Out.- Out. Set.
dez.Número de 2.960 1.115 2.642 2.819amostrasrrorrefadoras 22 3 6 18participantesTorrefadoras não 114 72 125 140
participantesrrotal 136 75 131 158Fonte: Elaborado a partir de CONTROLE (1996),CAI (1997), MAIOR (1997) e PROGRAMA(1998).
Analisando-se a distribuição regional dastorrefadoras com adulteração do produto' final,percebe-se, predominância de torrefadoraslocalizadas no Sudeste, seguidas pelas doNordeste. O esforço pela diminuição deadulterações foi desenvolvido igualmente nasregiões, ou seja, não existiu prioridades regionaisna redução das fraudes no café (Tabela 2).
454
TABELA 2 - Torrefadoras Participantes e NãoParticipantes do Programa de Autofiscalização daABIC com Impurezas e/ou Mistura no Café porRegião, Brasil, 1996, 1997 e 1998Reg. 1996 1997 1998
Jan-jun. Out- dez. Outubro SetembroNo. % No. % No. % No. %
N 8 5,9 9 11,8 11 8,4 7 4,4Ne 40 29,4 16 21,1· 22 16,8 39 24,7CO 9 6,6 6 7,9 8 6,1 12 7,6Se 62 45,6 33 44,7 69 52,7 75 47,5S 17 12,5 11 14,5 21 16,0. 25 15,8
Total 136 100 75 100 131 100 158 100Fonte: Elaborado a partir de CONTROLE (1996),CAI (1997), MAIOR (1997) e PROGRAMA(1998).
O fraude com a impureza "casca e paus" foi o quepredominou, no período em análise,correspondendo em média a cerca de 51,6% dastorrefadoras, seguido do milho (25,0%), centeio(13,8%), cevada (2,8%) e outros (6,8%). Éinteressante observar que cresce a participação dofraude com centeio de 7,4% em janeiro a junhode 1996 para 25,9%em setembro de 1998, emdetrimento do milho (decresce de 26,5% para17,7%, no período) e da cevada (cai de 5, I% para1,9%). Hipótese explicativa sobre o aumento damistura com centeio pode ser as mudanças nospreços relativos desse produto frente asalternativas similares (cevada e milho), o queinduziu o aparecimento de maior numero deempresas utilizando o centeio nas fraudes.O número de torrefadoras pode, ainda, seranalisado segundo a distribuição por classe deimpureza e/ou mistura. Em termos de impurezas(cascas e paus) as classes de 1,0% a 2,5% e 2,6%a 5% perfazem a ampla maioria de torrefadorascom fraudes no T&M. Para o caso das misturas,optou-se neste estudo pela elaboração de classespara o milho, cevada e centeio. A faixa demistura com milho mais usual entre astorrefadoras que fraudam o produto foi de 30, I% 45,0%. A mistura com cevada e centeioapresentaram maior concentração de firmasadulteradores com mais de 30, I% de mistura.(Tabela 3)
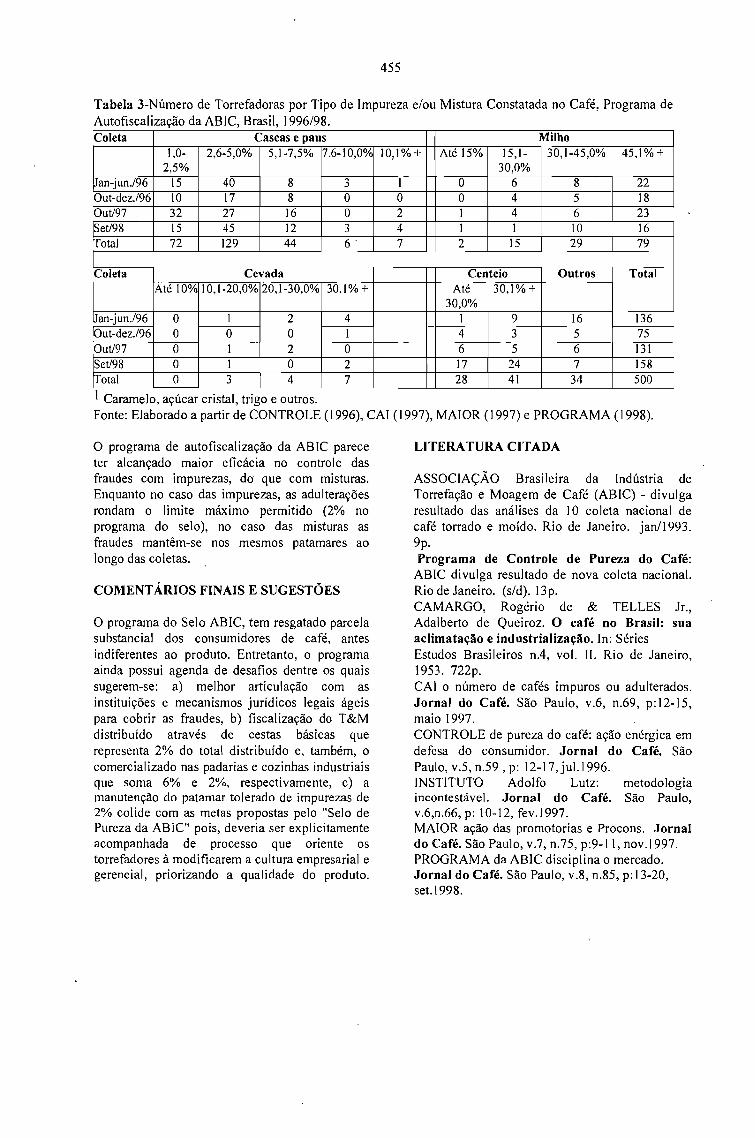
455
Tabela 3-Número de Torrefadoras por Tipo de Impureza e/ou Mistura Constatada no Café, Programa deAutofiscalização da ABlC, Brasil, 1996/98.Coleta Cascas e paus Milho
1,0- 2,6-5,0% 5,1-7,5% 7.6-10,0% 10,1%+ Até 15% 15,1- 30,1-45,0% 45,1%+2,5% 30,0%
Jan-jun.l96 15 40 8 3 I O 6 8 22Out-dez.l96 10 17 8 O O O 4 5 18Out/97 32 27 16 O 2 1 4 6 23Set/98 15 45 12 3 4 1 I 10 16[Total 72 129 44 6' 7 2 15 29 79
Coleta Cevada Centeio Outros TotalAté 10% 10,1-20,0%20,1-30,0% 30.1 % + Até 30,1%+
30,0%an-jun.l96 O 1 2 4 1 9 16 136
Out-dez.l96 O O O 1 4 3 5 75Out/97 O 1 2 O 6 5 6 131Set/98 O 1 O 2 17 24 7 158[Total O 3 4 7 28 41 34 500
I Caramelo, açúcar cristal, trigo e outros.Fonte: Elaborado a partir de CONTROLE (1996), CAI (1997), MAIOR (1997) e PROGRAMA (1998).
O programa de autofisca\ização da ABIC pareceter alcançado maior eficácia no controle dasfraudes com impurezas, do que com misturas.Enquanto no caso das impurezas, as adulteraçõesrondam o limite máximo permitido (2% noprograma do selo), no caso das misturas asfraudes mantêm-se nos mesmos patamares aolongo das coletas.
COMENTÁRIOS FINAIS E SUGESTÕES
O programa do Selo ABIC, tem resgatado parcelasubstancial dos consumidores de café, antesindiferentes ao produto. Entretanto, o programaainda possui agenda de desafios dentre os quaissugerem-se: a) melhor articulação com asinstituições e mecanismos jurídicos legais ágeispara cobrir as fraudes, b) fiscal ização do T&Mdistribuído através de cestas básicas querepresenta 2% do total distribuído e, também, ocomercializado nas padarias e cozinhas industriaisque soma 6% e 2%, respectivamente, c) amanutenção do patamar tolerado de impurezas de2% colide com as metas propostas peJo "Selo dePureza da ABIC" pois, deveria ser explicitamenteacompanhada de processo que oriente ostorrefadores à modificarem a cultura empresarial egerencial, priorizando a qualidade do produto.
LITERATURA CITADA
ASSOCIAÇÃO Brasileira da Indústria deTorrefação e Moagem de Café (ABIC) - divulgaresultado das análises da 10 coleta nacional decafé torrado e moído. Rio de Janeiro. jan!l 993.9p.Programa de Controle de Pureza do Café:
ABIC divulga resultado de nova coleta nacional.Rio de Janeiro. (s/d).13p.CAMARGO, Rogério de & TELLES Jr.,Adalberto de Queiroz. O café no Brasil: suaaclimatação e industrialização. In: SériesEstudos Brasileiros nA, vol. 11. Rio de Janeiro,1953. 722p.CAI o número de cafés impuros ou adulterados.Jornal do Café. São Paulo, v.6, n.69, p: 12-15,maio 1997.CONTROLE de pureza do café: ação enérgica emdefesa do consumidor. Jornal do Café. SãoPaulo, v.5, n.59 , p: 12-17,juI.1996.INSTITUTO Adolfo Lutz: metodologiaincontestável. Jornal do Café. São Paulo,v.6,n.66, p: 10-12, fev.1997.MAIOR ação das promotorias e Procons. Jornaldo Café. São Paulo, v.7, n.75, p:9-11, nov.1997.PROGRAMA da ABIC disciplina o mercado.Jornal do Café. São Paulo, v.8, n.85, p: 13-20,set.1998.


457
QUALIDADE DE BEBIDA DO CULTIVAR DE CAFÉ IAPAR-59 E OUTRAS
Tumoru Sera \ Marcos Aurelio Bacceti 2
'Instituto Agronômico do Paraná - [APAR, Londrina, Paraná, Brasil; - 2 Corretora Bacceti, Centro de2 Comércio de Café do Norte do Paraná - CCCNP, Londrina, Paraná.
INTRODUÇÃO
o 'IAPAR-59' é uma nova cultivar do tipolinhagem resistente a ferrugem derivada deseleção efetuada na descendência do cruzamentoentre Coffea arabica varo Vi lia Sarchi 971/1 O e o"Híbrido de Timor 832/2", realizado no CI FCCentro de Investigação das Ferrugens do Cafeeiro,em Portugal, onde recebeu a denominação deH361 (IAPAR, 1994). A geração F2 (H361-4) foirecebida pelo IAC - Instituto Agronômico deCampinas, que a denominou LC 1669. Em 1975 oIAPAR introduziu a geração F3 (LC 1669 Ep.127c.506), que passou a ser denominada IAPAR75163.Nos estudos realizados pelo IAPAR, a progênie75163-22 destacou-se pelas característicasagronômicas e resistências à ferrugem .Seudesempenho nas regiões de Londrina, Loanda eCarlópolis demonstrou condições para seulançamento como cultivar, passando a receber adenominação de IAPAR-59.Na geração F I do H361 anal isada no CIFC, todasas plantas pertenciam ao grupo fisiológico A(plantas que apresentam resistência às 30 raças deH vastratrix conhecidas no mundo). Assim, amaioria (94%) da população do IAPAR 59 temresistência às 30 raças conhecidas de Hvastratrix.Apesar dos testes de qualidade realizados antes doseu lançamento como cultivar, existemquestionamentos quanto a qualidade da sua bebidacomparativamente a outras cultivares. O objetivodeste trabalho foi o de avaliar a qualidade dabebida e outras características relacionadas aqualidade do 'lAPAR-59' e outros cultivaresexperimentais junto com os cultivares padrõesCatuaí, Mundo Novo e Bourbon Vermelho.
MATERIAIS E MÉTODOS
Amostras de frutos colhidos em estádio cereja dos10 cultivares foram descascados, degomadosmicrobianamente e secados a sombra. Testaramse os seguintes cultivares:1. IAC2077-2-5-8l (Catuaí Vermel ho):2. lcatu Precoce IAC 3282:3. [AC376-4 (Mundo Novo);4. IAPAR-59 (Sarchimor);5. F2 IAPAR-59 X Mundo Novo;
6. F2 de IAPAR-59 X Jcatuaí;7. IAPARLF77028 (Sarchimor);8. Catuaí Sh2Sh3 (cultivar experimental);9. Maragogipe;10. [APAR LF77054-21 (lcatu x Catuaí, cultivar
experimental).A análise sensorial através da prova em xícaraforam realizados no Centro de Comércio de Cafédo Norte do Paraná - CCCNP por 3 provadorespara cada cultivar.
RESULTADOS OBTIDOS E DISCUSSÃO
Obtiveram-se os resultados apresentados noQUADRO I quanto a tamanho dos grãos e o tipodebebida aval iada por 3 provadores. Em confronto
com as características de bebida das cultivaresrecomendadas para o estado do Paraná (SERA,1995) podemos tirar algumas conclusões.A testemunha "Catuaí Vermelho" 'IAC 2077-2-581' apresentou as bebidas dura ácida, dura e duralevemente ácida e com peneira média dos grãosde 16,3.A cultivar IAC 3282 de "Icatu Precoce" apresentoubebidas dura. dura e dura com peneira 16,). portanto.com tamanho dos grãos e bebida semelhantes ao'Catuai'. A cultivar lAC 376-4 de "Mundo Novo"apresentou bebidas dura para melhor, dura limpae duracom peneira 16,6, portanto melhor que "Catuaí".Repetindo os testes anteriores (SERA, 1996), acultivar lAPAR-59 mostrou-se melhores que"Catuaí" e "Mundo Novo", apresentando bebidasdura para melhor, dura limpa e dura para melhorcom peneira 16,6. Assim, •IAPAR-59' recebendonível de tratos culturais semelhantes, superam ascultivares comerciais mais cultivadas quanto abebida e igual a "Mundo Novo" em tamanho dosgrãos.Ao obter cultivar híbrida F2 IAPAR-59 x MundoNovo, as bebidas dura, dura e dura superior compeneira 16,7, grãos ligeiramente maiores que o"Mundo Novo". A bebida foi ligeiramente inferiorque o "Mundo Novo" mas assim mesmo entre osmelhores mas com vantagem de grãos um poucomaiores do que qualquer um dos parentais o queindica que apresenta vigor de híbrido como defato apresenta.A bebida predominante foi de "Catuaí" na cultivarhíbrida F2 IAPAR-59 x (Icatu x Catuaí)apresentando as bebidas dura ácida, dura e dura
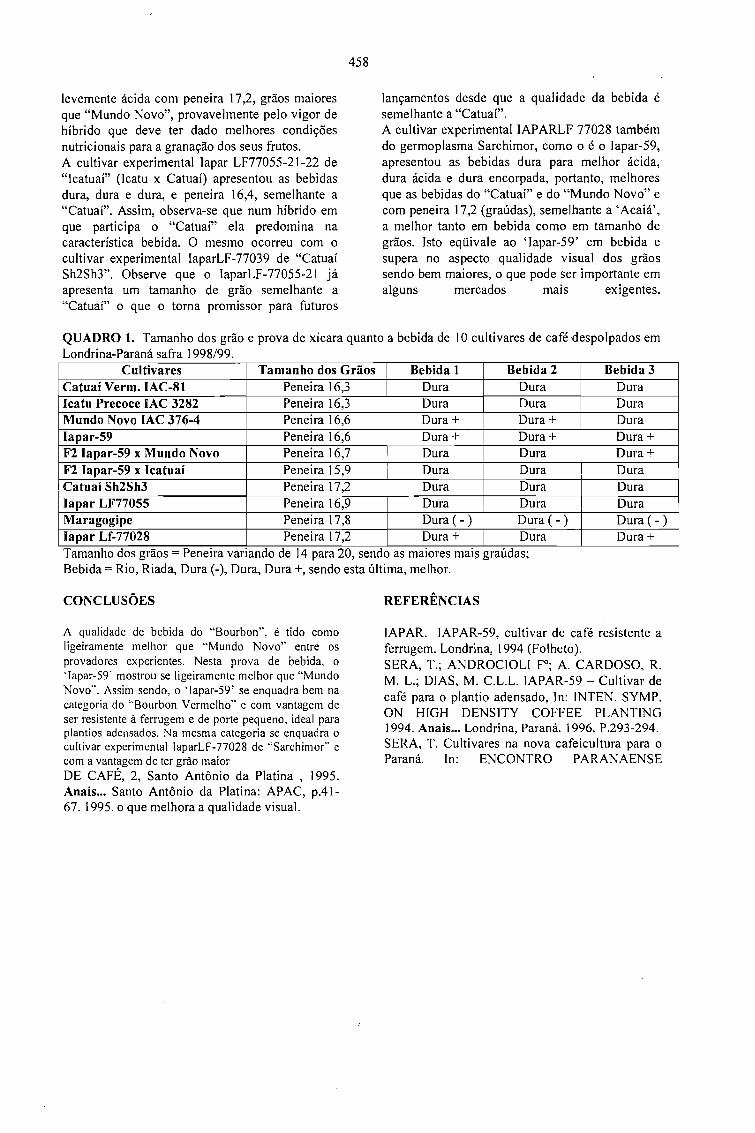
458
levemente ácida com peneira 17,2, grãos maioresque "Mundo Novo", provavelmente pelo vigor dehíbrido que deve ter dado melhores condiçõesnutricionais para a granação dos seus frutos.A cultivar experimental Iapar LF77055-21-22 de"Icatuaí" (lcatu x Catuaí) apresentou as bebidasdura, dura e dura, e peneira 16,4, semelhante a"Catuaí". Assim, observa-se que num híbrido emque participa o "Catuaí" ela predomina nacaracterística bebida. O mesmo ocorreu com ocultivar experimental IaparLF-77039 de "CatuaíSh2Sh3". Observe que o IaparLF-77055-21 jáapresenta um tamanho de grão semelhante a"Catuaí" o que o torna promissor para futuros
lançamentos desde que a qualidade da bebida ésemelhante a "Catuaí".A cultivar experimentalIAPARLF 77028 tambémdo germoplasma Sarchimor, como o é o Iapar-59,apresentou as bebidas dura para melhor ácida,dura ácida e dura encorpada, portanto, melhoresque as bebidas do "Catuaí" e do "Mundo Novo" ecom peneira 17,2 (graúdas), semelhante a 'Acaiá',a melhor tanto em bebida como em tamanho degrãos. Isto eqüivale ao 'Iapar-59' em bebida esupera no aspecto qualidade visual dos grãossendo bem maiores, o que pode ser importante emalguns mercados mais exigentes.
QUADRO 1. Tamanho dos grão e prova de xícara quanto a bebida de 10 cultivares de café despolpados emLondrina-Paraná safra 1998/99.
Tamanho dos graos = Peneira vanando de 14 para 20, sendo as rnarores mais graudas;Bebida = Rio, Riada, Dura (-), Dura, Dura +, sendo esta última, melhor.
Cultivares Tamanho dos Grãos Bebida I Bebida 2 Bebida 3
Catuaí Vermo IAC-8I Peneira 16,3 Dura Dura DuraIcatu Precoce IAC 3282 Peneira 16,3 Dura Dura DuraMundo Novo IAC 376-4 Peneira 16,6 Dura + Dura + DuraIapar-59 Peneira 16,6 Dura + Dura + Dura +F2 Iapar-59 x Mundo Novo Peneira 16,7 Dura Dura Dura +
F2 Iapar-59 x Icatuaí Peneira 15,9 Dura Dura DuraCatuaí Sh2Sh3 Peneira 17,2 Dura Dura DuraIapar LF77055 Peneira 16,9 Dura Dura DuraMaragogipe Peneira 17,8 Dura (- ) Dura ( - ) Dura (-)Iapar Lf-77028 Penei ra 17,2 Dura + Dura Dura+
-
CONCLUSÕES REFERÊNCIAS
A qualidade de bebida do "Bourbon", é tido comoligeiramente melhor que "Mundo Novo" entre osprovadores experientes. Nesta prova de bebida, o'Iapar-59' mostrou se ligeiramente melhor que "MundoNovo". Assim sendo, o 'Iapar-59' se enquadra bem nacategoria do "Bourbon Vermelho" e com vantagem deser resistente à ferrugem e de porte pequeno. ideal paraplantios adensados. Na mesma categoria se enquadra ocultivar experimental laparLF-77028 de "Sarchimor" ecom a vantagem de ter grão maiorDE CAFÉ, 2, Santo Antônio da Platina, 1995.Anais... Santo Antônio da Platina: APAC, pAI67. 1995. o que melhora a qualidade visual.
IAPAR. IAPAR-59, cultivar de café resistente aferrugem. Londrina, 1994 (Folheto).SERA, T.; ANDROCIOLI P; A. CARDOSO. R.M. L.; DIAS, M. C.L.L. lAPAR-59 - Cultivar decafé para o plantio adensado, In: INTEN. SYMP.ON HIGH DENSITY COFFEE PLANTING1994. Anais... Londrina, Paraná. 1996. P.293-294.SERA, T. Cultivares na nova cafeicultura para oParaná. In: ENCONTRO PARANAENSE

459
EFEITO DA ADUBAÇÃO POTÁSSICA NA QUALIDADE DOS GRÃOS BENEFICIADOS DECAFÉI
Enilson De Barros Silva, Francisco Dias Nogueira, Paulo Tácito Gontijo Guimarães, Silvio Júlio DeRezende Chagas & Laerte Costa
INTRODUÇÃO
Para uma maior competitividade do cafébrasileiro através da melhoria do produto, tornase necessário, apoiar-se numa pesquisamultidisciplinar, com características voltadas paraa qualidade da bebida, que é o interesse final.
A influência do local de cultivo na qualidade dabebida tem sido intensivamente documentada,porém o mecanismo destes efeitos é poucoestudado. No Estado de Minas Gerais tem sido
muito discutida a diferença existente entrequalidades de cafés produzidos na Zona da Mata,Sul de Minas e Região do Triângulo Mineiro e
Alto Paranaíba, sem nenhuma referência nesteslocais do efeito da adubação potássica naqualidade do café.°potássio tem sido. há muito tempo, consideradoo "elemento da qualidade" em nutrição de
plantas. Apesar desta reputação qualitativa e deaumentar a produção agrícola, normalmenteignora-se a extensão dos benefícios da adubaçãopotássica. A qualidade dos produtos agrícolas nãoé facilmente definida ou medida como se faz para
produção. ° padrão de qualidade depende depropósitos, pelos quais. a planta ou parte dela éutilizada (Mengel e Kirkby, 1987). No caso do
café, o mais importante é o reflexo da adubaçãona produção, além dos efeitos sobre os compostosquímicos responsáveis pela qualidade da bebida(Silva. 1995).A atividade da enzima polifenoloxidase está
correlacionada positivamente com a qualidade dabebida do café arabica, no Brasil (Amorim e
Silva, 1968) foi confirmada posteriormente porRotemberg e Iachan (1972), também no Brasil epor Sanint e Valência (1972) na Colômbia.Dentro desta linha de pesquisa, Arcila-Pulgarin eValência-Aristizábal (1975) estudaram o efeito dafertilização potássica sobre a atividade dapol ifenoloxidase e não encontraram diferençasestatísticas entre os seguintes tratamentos:testemunha, cloreto de potássio (KCI). sulfato de
potássio (K2S04), KCI + N + P + Mg e K2S04 +N + P + Mg; a fertilização potássica aumentou osteores de K nos grãos, sem correlacionar com aatividade da enzima, mas observou-se uma maior
atividade da enzima polifenoloxidase, quando seaplicou o sulfato de potássio.
Carvalho et aI. (1994) registraram haver variaçõesda atividade da polifenoloxidase, que permitem
separar as classes de bebida com base nasatividades destas enzimas, mostrando para o caféde bebida "riado e rio" atividades inferiores a55,99 U mim-I g-I de amostra; nos cafés de
bebida "dura" atividades entre 55,99 e 62,99 Umim-I g-l de amostra; nos cafés de bebida
"mole" atividades entre 62,99 e 67,66 U rniml g
I de amostra e nos cafés de bebida "estritamentemole" atividades superiores a 67,66 U mim-I s'de amostra, constatando assim um aumentosignificativo na atividade da polifenoloxidase àmedida que o café apresenta-se de melhor
qualidade.Santinato et al. (1996), no Brasil, utilizaram dosescrescentes de O, 114, 145 e 208 kg de K.ha- I para
variedade Acaiá em Latossolo Vermelho Amarelomas não encontraram melhoria da bebida.avaliada sensorialmente pela "prova da xícara"entre as doses de K na forma de sal itre potássico.Malavolta (1986) estudou o efeito das fontes de S
(sulfato de amônio, K2S04MgS04 e fosfogesso)e doses de S (O, 20 e 40 kg de S.ha- 1) na
produção e qualidade da bebida pela "prova daxícara" mas não encontrou diferença na qualidadeda bebida na média de quatro safras (1981/82 a
1984/85) no município de Olímpia (SP) emPodzólico Vermelho Amarelo. As dosesutilizadas pelo autor correspondern. na forma desulfato de potássio (15% de S). em O, 133 e 266kg de sulfato de potássio.haÍ.Em trabal ho recente, Si Iva et aI. (1999) estudaramo efeito de fontes e doses de K na composição
físico-química e química dos grãos de cafébeneficiados da região do Sul de Minas (SãoSebastião do Paraíso) observaram uma maioratividade da polifenoloxidase, índice de coloraçãoe açúcares totais. e baixa acidez titulável total emcafés adubados com a fonte sulfato de potássio,confirmando que esta fonte proporcionava umamelhor qualidade do café. Os autoresencontraram uma redução do teor de açúcar nãoredutor com aumento das doses de K na forma deKCI e o inverso com a fonte sulfato de potássio, emesma tendência ocorreu para os açúcares totais.Com relação aos íons influenciando a atividadeenzimática da polifenoloxidase, Fox (1991)descreve os efeitos de solutos sobre a atividade

enzimática enfocando a inativação de enzimaspelo KCI e pelo NaCI atribuindo-se ao íonhalogêneo (Cl") esta ação inibitória, tendo
mencionado ainda alterações no sabor dosprodutos o que é de suma importância naavaliação da qualidade do café. Quimicamente,uma explicação poderia considerar que, asenzimas são ativada por íons metálicos, mas um
excesso de íon halogênio poderia ocasionar umareação de precipitação. Conhece-se também umpoderoso efeito de celtos ânions na desnaturaçãode proteína pela sua habilidade em promover aruptura da cadeia orgânica. os quais são, emordem decrescente: SCN- > C104- :;:: 1- > Cl- >
acetato> S04-2. No caso da polifenoloxidase
esperar-se-ia uma interação ou reação deprecipitação entre Cu+2 e Cl- (Robinson e Eskin,
1991 ).
Por outro lado. plantas que recebem elevadasquantidades de CI aumentam o conteúdo de água
(Gouny.1973). e isto seria um efeito da adubaçãopotássica na forma de KCI. Um maior teor deumidade nos frutos favorece a proliferação demicroorganismos. e. segundo Leite (1991),qualquer condição adversa aos grãos de café
beneficiados. a atividade da polifenoloxidase atuasobre os compostos fenólicos, diminuindo suaação antioxidante sobre os aldeídos. facilitando
consequenternente, a oxidação dos mesmos.° objetivo deste trabalho é verificar oefeito de fontes e doses de K nas característicasfísicas. físico-químicas e químicas dos grãos decafé beneficiados em dois locais de cultivo.
MATERIAL E MÉTODOS
Foram instalados dois experimentos. sendo um nomunicípio de São Sebastião do Paraíso emLatossolo Roxo (LR) (pH=6,O; P=7,O, K=70 e SS04-2=19.5 mg.dm-3; Ca=3,8. Mg=0,9 e AI=O.1cmolc.dm-3; V=65% e matéria orgânica=2,6dag.kg<l ) e outro em Patrocínio em LatossoloVermelho Amarelo (LV) (pH=5.9; P=3,O. K=63 eS-S04-2=17,5 mg.drrr-i: Ca=3.4, Mg=1,3 eAI=O.I cmol c.dm-3; V=69% e matériaorgânica-Ll dag.kg" l ), cujas análises de solo na
profundidade de ° a 20 em, encontram-se naTabela I. Em ambos os locais. os experimentosforam dei ineados em blocos casualizados emesquema de parcelas subdivididas. utilizando-setrês fontes de K: cloreto de potássio (KC1),
sulfato de potássio (K2S04) e nitrato de potássio(KN03) nas parcelas e quatro doses de K (O, 100.200 e 400 kg de K.ha- I) aplicadas nassubparcelas com quatro repetições, A parcela
460
'experimental foi constituída de três linhas de oitocovas, formando um total de 24 covas porparcela, sendo a parcela útil as seis covas, centrais.
Usou-se nos experimentos a espécie CofJeaarabica L., cultivar Catuaí Vermelho noespaçamento 3,5 x 0,7 m, com uma planta porcova, em ambos os locais de cultivo.A atividade enzimática da polifenoloxidase foideterminada em amostras de grãos de cafébeneficiados moídos em moinho tipo CrotonMod.TE-580, em peneira de 30 mesh, referente acada tratamento aplicado em cada ano agrícolapor cada local dé cultivo. A metodologia paraavaliar a atividade enzimática da polifenoloxidasefoi adaptado para maior rendimento nas análises
de laboratório. conforme foi descrita por Draetta e
Lima (1916). Foram pesados 5 g da amostra decafé previamente moída e adicionaram-se 40 ml,da solução de fosfato de potássio 0,1 mol.Lr l àpH 6.0. Em seguida, foi agitado por 5 minutos.
Todo material utilizado foi mantido gelado. Apósa agitação, fez-se a fi ltragem, uti lizando-se papelde fi Itro Whatman nQ I. A atividade da
polifenoloxidase foi determinada pelo métododescrito por Ponting e Joslyng (1948). Util izou-seo extrato da amostra sem DOPA (3,4dihidroxifenil-alanina) como branco expressa emU.mim-I.g- l (Unidade de atividade enzimática
por unidade de tempo em minutos por gramas degrãos beneficiados).Os dados das variáveis qualitativas foramsubmetidos à anál ise de variância e regressãopol inomial. Uti Iizou-se o programa SAS paraanálise estatística. estabelecendo-se os níveis designificância de 5% e 1%.
RESULTADOS E DISCUSSÃO
Os valores médios da atividade enzimática dapolifenoloxidase em dois locais de cultivo sobre oefeito de fontes e doses de K encontram-se naTabela \. Houve interação signifícativa entrefontes e doses de K na atividade enzimática dapo Iifeno lox idase,Em relação aos dados médios obtidos (Tabela 1),verifica-se que as atividades enzimáticas dapolifenoloxidase, em Patrocínio (Cerrado), foramelevadas em relação aos obtidos em SãoSebastião do Paraíso (Sul de Minas). quando secomparam os valores de atividade enzimática nostratamentos que não receberam de adubaçãopotássica (testemunha= O kg de K.ha- I). Verifica
se que nas testemunhas representativas de cadafonte de K. em São Sebastião do Paraíso que osgrãos de café beneficiados foram classificados,
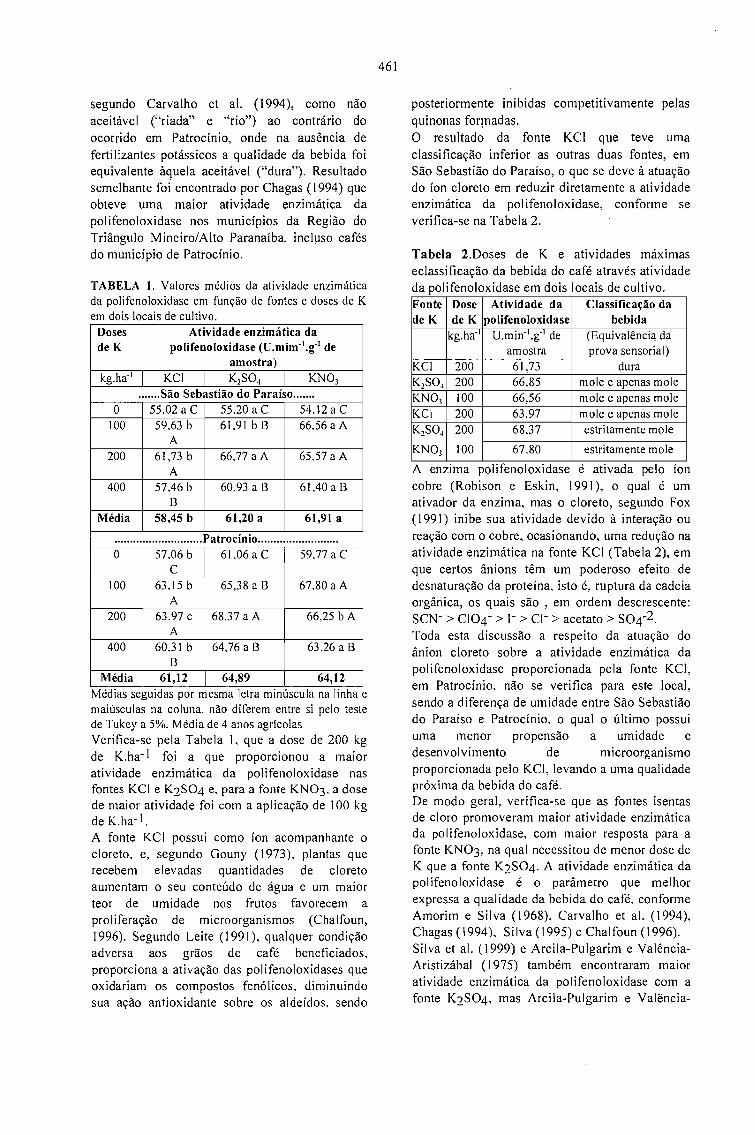
461
TABELA l. Valores médios da atividade enzimáticada polifenoloxidase em função de fontes e doses de Kem dois locais de cultivo.
segundo Carvalho et a!. (1994), como nãoaceitável ("riada" e "rio") ao contrário do
ocorrido em Patrocínio, onde na ausência defertilizantes potássicos a qualidade da bebida foiequivalente àquela aceitável ("dura"). Resultadosemelhante foi encontrado por Chagas (1994) queobteve uma maior atividade enzimática dapo1ifenoloxidase nos municípios da Região doTriângulo Mineiro/Alto Paranaíba, incluso cafésdo município de Patrocínio.
A enzima polifenoloxidase é ativada pelo íoncobre (Robison e Eskin, 1991), o qual é umativador da enzima, mas o cloreto, segundo Fox(1991) inibe sua atividade devido à interação oureação com o cobre, ocasionando, uma redução naatividade enzimática na fonte KCI (Tabela 2), emque certos ânions têm um poderoso efeito dedesnaturação da proteína, isto é, ruptura da cadeiaorgânica, os quais são, em ordem descrescente:SCN- > CI04- > 1- > CI- > acetato> S04-2.Toda esta discussão a respeito da atuação doânion cloreto sobre a atividade enzimática dapolifenoloxidase proporcionada pela fonte KCI,em Patrocínio, não se verifica para este local,
sendo a diferença de umidade entre São Sebastiãodo Paraíso e Patrocínio, o qual o último possuiuma menor propensão a umidade edesenvolvimento de microorganismoproporcionada pelo KCI, levando a uma qualidadepróxima da bebida do café.De modo geral, verifica-se que as fontes isentasde cloro promoveram maior atividade enzimáticada polifenoloxidase, com maior resposta para afonte KN03, na qual necessitou de menor dose deK que a fonte K2S04. A atividade enzimática dapolifenoloxidase é o parâmetro que melhorexpressa a qualidade da bebida do café. conformeArnorirn e Silva (1968), Carvalho et aI. (1994),Chagas (1994), Si Iva (1995) e Chalfoun (1996).Silva et aI. (1999) e Arcila-Pulgarim e ValênciaAristizábal (1975) também encontraram maioratividade enzimática da polifenoloxidase com afonte K2S04, mas Arcila-Pulgarirn e Valencia-
Tabela 2.Doses de K e atividades maximaseclassificação da bebida do café através atividadeda polifenoloxidase em dois locais de cultivoFonte Dose Atividade da Classificação dade K de K polifenoloxidase bebida
kg.ha' ILrninl.g" de (Equivalência daamostra prova sensorial)
KC1 200 61.73 duraK2S04 200 66.85 mole e apenas moleKN03 100 66,56 mole e apenas moleKCI 200 63,97 mole e apenas moleK2S04 200 68,37 estritamente mole
KN03 100 67.80 estritamente mole
posteriormente inibidas competitivamente pelas
qui nonas formadas.° resultado da fonte KCI que teve umaclassificação inferior as outras duas fontes, emSão Sebastião do Paraíso, o que se deve à atuaçãodo íon cloreto em reduzir diretamente a atividadeenzimática da polifenoloxidase, conforme severifica-se na Tabela 2.
Médias seguidas por mesma letra minúscula na linha emaiúsculas na coluna. não diferem entre si pelo testede Tukey a 5%. Média de 4 anos agrícolasVerifica-se pela Tabela I, que a dose de 200 kgde K.ha- I foi a que proporcionou a maioratividade enzimática da polifenoloxidase nasfontes KCI e K2S04 e, para a fonte KN03. a dosede maior atividade foi com a apl icação de 100 kgde K.ha- I,
A fonte KCI possui como íon acompanhante ocloreto, e, segundo Gouny (1973), plantas querecebem elevadas quantidades de cloretoaumentam o seu conteúdo de água e um maiorteor de umidade nos frutos favorecem aproliferação de microorganismos (Chalfoun,1996). Segundo Leite (1991). qualquer condiçãoadversa aos grãos de café beneficiados,proporciona a ativação das polifenoloxidases queoxidariam os compostos fenólicos, diminuindosua ação antioxidante sobre os aldeídos. sendo
Doses Atividade enzimática dade K polifenoloxidase (Ll.miml.g' de
amostra)kg.ha KCI K2S04 KN03
.......São Sebastião do Paraíso.......O 55.02 a C 55.20 a C 54.12aC
100 59.63 b 61.91 b B 66.56 a AA
200 61,73 b 66.77 a A 65.57 a AA
400 57.46 b 60.93 a B 61.40 a BB
Média 58,45 b 61.20 a 61,91 a
............................Patrocínio..........................O 57.06 b 61.06 a C I 59.77 a C
C100 63.15 b 65,38 a B 67.80 a A
A200 63.97 c 68.37 a A 66,25 b A
A400 60.31 b 64.76aB 63.26 a B
BMédia 61,12 64,89 64,12..

Aristizábal (1975) não encontraram resposta aadubação potássica, embora observaram umatendência de maior atividade da enzimapolifenoloxidase, quando se aplicou sulfato depotássio. Malavolta (1986) estudou o efeito defontes e doses de S na qual idade da bebida docafé mas não encontrou diferença na qualidadeavaliada pela "prova da xícara". A avaliação daqualidade, praticada comercialmente através deprovadores, conhecida como "prova da xícara" émuito subjetiva (Chagas, 1994) em relação aatividade da polifenoloxidase ("BebidaQuímica"), dificultando detectar diferenças entreas amostras de grãos de café beneficiados quanto.à adubação. O mesmo ocorreu com Santinato etaJ. (1996) não encontraram melhoria da qualidadeC'prova da xícara") com a aplicação de dosescrescentes de K na forma de salitre de potássio
(K N03)'
CONCLUSÕES
Pelos resultados obtidos conclui-se que:Fontes isentas de cloreto _ proporcionaram amelhor qualidade da bebida do café, com melhorresposta em São Sebastião do Paraíso;As doses de 200 kg de Kiha" I na forma de KCI eK2S04 e 100 kg de K.ha- 1 na forma de KN03foram os que proporcionaram os melhoresresultados qualitativos.
REFERÊNCIAS
Amorirn, H.V. e Silva. D.M. 1968. Relationshipbetween the polyphenol oxidase activity ofcoffee beans and quality of the beverage.Nature. 219(27):38 I-382.
Argila-Pulgarin, J. e Valência-Aristizábal, G.1975. Relacion entre la actividade de lapolifenol oxidasa (P.F.O.) y las pruebas decatacion como medidas de la calidad de labebida dei cafe. Cenicafé. 26(2):55-7 I.
Carvalho, V.D., Chagas, S.J.R., Chalfoun, S.M.,Botrel, N. e Juste Junior, E.S.G. 1994. Relaçãoentre a composição físico-química e químicado grão beneficiadoe a qualidade de bebida docafé. Pesquisa Agropecuária Brasi leira.29(3):449-454.
Chagas, S.J .R. 1994. Caracterização qurrmca equalitativa de cafés de alguns municípios detrês regiões produtoras de Minas Gerais.ESAL. 95. Dissertação de Mestrado
462
Chalfoun, S.M.S. 1996. O café (Coffea arabicaL.) na Região Sul de Minas Gerais-relação daqualidade com fatores ambientais. estruturais etecnológicos. UFLA. 171. Tese de Doutorado.
Draetta, I.S. e Lima, D.C. 1976. Isolamentos ecaracterização das polifenoloxidases do café.Coletânea do Instituto de Tecnologia deAlimentos. 7:3-28.
Fox, P.F. J991. Food enzymology. London,Elsevier Applied Science, 378.
Gouny, P. 1973. Observacones sobre elcomportamento dei vegetal en presencia deions de cloro. Revista de la Potassa. 45(5): 114.
Leite. I.P. 1991. Influência do local de cultivo edo tipo de colheita nas características físicas,composição química do grão e qualidade docafé (Coffea arabica, L.). ESAL. 131.Dissertação de Mestrado.
Malavolta, E. 1986. Efeitos de doses e fontes deenxofre em culturas de interesse econômico.IV - Café. São Paulo. 41. Boletim Técnico 4.
Ponting, J.D. e Joslyng, M.A. 1948. Ascorbic acidoxidation and browning in apple tissueextracts. Achives of Biochemistry. 19:47-63.
Robinson, D.S. e Eskin, N.A.M. 1991. Oxidativeenzymes in foods. New York, ElsevierApplied Science, 314p.
Rotenberg, B. e lachan, A. 1972. Contribuição aoestudo enzimático do grão de café. I.Tirosinase e lacase. Revista Brasileira deTecnologia. 3: 155-159.
Sanint, 0.8. e Valéncia, A. 1972. Actividadeenzimática en el grano de café en reiación conla cal idade de la bebida. I. Duración de lafermentación. Cenicafé. 23:59-7 I.
Santinato, R., Oliveira, L.H. e Pereira, E.M.1996. Efeitos do uso de sal itre de potássiocomo fonte de nitrogênio e potássio naadubação química do cafeeiro - Carmo doParanaíba/MG - 1992/1996. In. CongressoBrasileiro de Pesquisas Cafeeiras. Águas deLindóia. Anais. 22: 180-184.
Silva. E.B. 1995. Potássio para cafeeiro: efeito defontes. doses e determinação de cloreto.UFLA. 87p. Dissertação de Mestrado.
Silva. E.8.. Nogueira, F.D., Guimarães, P.T.G ..Chagas. S.J.R. e COSTA. L. 1999. Fontes edoses de potássio na produção e qualidade dogrão de café beneficiado. PesquisaAgropecuária Brasileira (No prelo)

463
CONDUTIVIDADE ELÉTRICA DO EXSUDATO DE GRÃOS DE CAFÉ (Coffea arabica L.) E SUARELAÇÃO COM A QUALIDADE DA BEBIDA
Prete l, C.E.C. e Abrahão", J.T.M.
lprof'. Assossiado.do Departamento de Agronomia da Universidade Est. de Londrina, Londrina - PR, Brasil.2Prof'. Titular Aposentado da Escola Superior de Agricultura Luiz de Queiróz, Piracicaba - SP, Brasil.
SUMMARYWith the objective ofenlarging the possibilites to determinate the cofJee quality, the relation between electricconductivity of the exudate from crude cofJee grains and the type quality of the beverage was studied. Themethodology to evaluate the electric conductivity was developed at the Seed Laboratory of the AgricultureDepartment of ESALQ in Piracicaba-SP, during the period of 1990-1992. This initial exploratory testshowed the best methodology to be the use ofsamples with 50 cofJee grains, with known weight, immersedin 75 ml of destilled water, at 25°C during 3,5 hours, followed by shaking. After that the electricconductivity was measured using a conductivimeter and expressed in llS.cm·J.g·J for sample. The resultsindicate that: the most lixiviated ion from the cofJee grains is potassium. wich is direct correlated (r = 99%)with the electric conductivity; the electric conductivity evolution follows the speed of the potassiumlixiviation and of the water absorption by the grains; the electric conductivity is remarkable afJected byimperfection in the cofJee grains (stinkers, black bean, sours, unripes and boreds), because this imperfectionsequence correspond to the order of importance in the degradation of the membrane system; also it wasverified the injluence of the humidity content, of the genotype and of the grain size; the increment ofgreenfruits harvested depreciate the cofJee quality, moreover when they are submitted to a drying temperaturehigher than 45°C. In this experimental conditions the eletric conductivity and the potassium lixiviation testswere more sensible to detect differences between treatments than usual procedures. The hulling of the fruitspropitiate the obtainment of better cofJee quality (better beverage, less imperfections and smaller electricconductivity) in ten harvest locations (Maringa, Londrina, Piracicaba, Campinas. Garca, Pindorama.Mococa, Patrocinio, Alfenas e Machado). 1t was concluded that occurs an inversed relation betweencofJee quality and electric conductivity, or lhe better the beverage quality the smaller is the electricconductivity ofthe exudates from the crude grain ofcofJee.
INTRODUÇÃO
A competência dos produtores de café depende doempenho em controlar custos. aumentar aprodução por hectare e elevar a qualidade dabebida.Os atuais procedimentos de avaliação comercialda qualidade do café estão baseados emparâmetros empíricos e subjetivos pois dependemde sensações (visão. olfato e paladar) ehabilidades pessoais, adquiridas com muitos anosde experiência. Assim a complementação dosprocedimentos em uso com a adoção de métodosfisicos e químicos tornaria mais real e objetiva adeterminação da qualidade do café.Os trabalhos de Arnorim (1978), relacionandoaspectos bioquímicos e histoquímicos do grão decafé verde com a deterioração da qualidade. muitocontribuíram para realçar a hipótese de que aperda da permeabilidade e estrutura dasmembranas celulares conduz à deterioração docafé.Inúmeros trabal hos citados em Cl iffo rd (1985).indicam que os piores cafés. em termos dequalidade de bebida. possuem: menos proteínassolúveis, mais aminoácidos livres. mais ácido
clorogênico, menos fenóis hidrolisáveis, menosácido ascórbico, baixo teor em carboidratos emaior teor de ácido graxos livres com diminuiçãodo conteúdo de lipídeos. Estes resultados indicamque ocorrem importantes reações oxidativasdurante o processo de deterioração dos grãos decafé sugerindo intensa peroxidação de lipídeos.Dentre os compostos orgânicos estudado,s.especial atenção foi dada a enzimapolifenoloxidase (PPO) a qual estavacorrelacionada positivamente com a qualidade dabebida do café como demonstraram Amorim eSilva (1968). In vivo. a enzima polifenoloxidaseencontra-se ligada às membranas e é ativadasomente quando liberada destas. Arnorirn (1978)sugere que danos ocorridos nas membranasliberam. e portanto ativam. a polífenoloxidase(PPO). que por sua vez oxida os ácidosclorogênicos à quinonas. A PPO é inibida pelasquinonas formadas. diminuindo sua atividade.Segundo Arnorirn (1978) qualquer fator ambienteque altere a estrutura da membrana. por exemploataque de insetos. infecções por microrganismos.alterações fisiológicas e danos mecânicos,provocam uma rápida deterioração dos grãos decafé. Uma vez rompida a membrana celular ocorre

um maior contacto entre as enzimas e oscompostos químicos presentes intra e extracelularno grão. Isto ocasiona reações químicas quemodificam a composição química original do caféverde e em conseqüência as propriedadesorganolépticas da infusão preparada com este.Esta seqüência sugere. na verdade, que aqualidade inferior dos cafés com baixa atividadeda PPO está relacionada com os danos àsmembranas celulares. sendo então a diminuiçãoda PPO uma conseqüência disto.Apesar da qualidade e aroma da bebida do caféserem determinantes no estabelecimento de seupreço. as causas de variação da qualidade doproduto somente agora estão sendo esclarecidas.A descoberta do composto 2,4-6 tricloroanisole(TCA) presentes em amostras de café Rio quesofreram ação de fungos Aspergillus spp estarelacionada com a má qualidade da bebida. Semdúvida a ordem das reações químicas que ocorremdurante a formaçào do grão de café culmina nacomposição química final destes. e esta pode serum importante fator na determinação da qualidadeda bebida (Amo rim e Melo. 1991).Há. certamente. mecanismos tão complexos einterdependentes no processo de deterioração quenenhuma teoria simples poderia fornecerexplicações definitivas e permitir afirmaçõestaxativas sobre o que é, na realidade.conseqüência de uma somatória de eventos. Poroutro lado, existe uma concordância de que adegeneração das membranas celulares esubsequente perda de controle da permeabilidade,seja um dos primeiros eventos que caracterizam adeterioração.Testes baseados na perda de integridade dasmembranas foram desenvolvidos. estudados eutilizados por Mathews e Bradnock (1968).Absul-Baki e Anderson (1972), McDonald Jr eWilson (1979). Yaklich et aI. (1979). Steere et ai.(1981), Powell (1986).Nestes trabalhos as sementes são imersas em águae durante o processo de embebição, de acordocom o grau de integridade de suas membranas.lixiviam solutos citoplásmáticos no meio líquido.Os solutos, com propriedades eletrolíticas.possuem cargas elétricas que podem ser medidascom um condutivímetro. Assim sementes combaixo vigor liberam grande quantidade deeletrólitos na solução. resultando em alto valor decondutividade elétrica. ou em elevadasconcentrações de determinados íons,principalmente potássio.Amorim (1978) estudando aspectos bioquímicos ehistoquímicos do grão de café verde erelacionando os resultados com a deterioração daqualidade, verificou haver maior lixiviação de
464
potássio nos grãos dos piores cafés. Este trabalhovisa a confirmação da hipótese de que existe umarelação entre o teste de condutividade elétrica doexsudato de grãos de café com a sua qualidade,avaliada pelo tipo e bebida.Pretende-se com isso obter um teste rápido. debaixo custo, fácil e seguro que possacomplementar a avaliaçào da qualidade de grãosde café, favorecendo assim a obtenção de cafés demelhor qualidade.
MATERIAL E MÉTODOS
Metodologia para Análise:Condutividade elétricaUtilizou-se metodologia proposta por Loeffler etal. (1988) que consistiu em utilizar quatroamostras de 50 grãos de cada parcela, sem escolhados grãos defeituosos (pretos, brocados. verdes eardidos). pesadas (precisão de O, Ig) e imersas em75 ml de água destilada (no interior de copos deplástico de 180 ml de capacidade) e colocadas emcâmara à 25°C.Após o período de embebição de 3,5 horas.procedeu-se à leitura da condutividade elétrica dasolução. em aparelho Digimed CD-20. Osresultados obtidos foram expressos em flS.cm'l.g'l
Lixiviação de potássioApós a leitura da condutividade elétrica, a soluçãosem os grãos foi vertida para recipientes de vidro,dos quais eram removidas ai íquotas paradeterminação da quantidade de potássio lixiviado.A análise do potássio foi realizada em fotômetrode chama Digimed NK-2002, sem prévia digestãoe com os dados obtidos foi calculado o lixiviadode potássio expresso em ppm/g de amostra.
Teor de água dos grãos de caféAvaliado pelo método da estufa a 105 ± s-e,durante 24 horas, de acordo com as Regras paraAnálise de Sementes (Brasil. 1976).
Determinação da qualidade da bebidaNos laboratórios de classificação de qualidade docafé da UNICAFÉ - Londrina - PRo da FederaçãoMeridional de Cooperativas Agropecuárias(FEMECAP) - Campinas - SP, e da Bolsa deMercadorias e Futuros (BMF) - São Paulo - SP,foi realizada a determinação da qualidade dabebida pela prova de xícara.
Prova de XícaraDe cada amostra foram tomados 100 gramas decafé beneficiado, que foram torrados. até os grãosatingirem cor achocolatada (ponto de estalo,torração americana). Após a torração o café foi

moído em moinhos especiais com granulaçãomédia apropriada (tamis de 14 fios por em), Aseguir foi preparada a infusão colocando-se la gde pó para 100 ml de água em ponto de fervura.Para cada parcela foram preparadas cinco xícaras,que foram provadas por degustadores de elevadacompetência nos diferentes laboratórios declassificação.Os resultados da degustação de cada xícara foramreduzidos a valores numéricos, segundo Tabela I,proposta por Garruti e Conagin (1961), tambémutilizada por Miya et aI. (1973/74).
Tabela]. Características organolépticas e grau depontuação dos tipos de bebida padrão em café.Classificaçã Característica Pontoso da Bebida Organolépticasritamente Bebida de sabor suavisíssimoMole e adocicado 24
Mole Bebida de sabor suave.acentuado e adocicado 18
Apenas Mole Bebida de sabor suave. porémcom leve adstringência 13
Dura Bebida com saboradstringente. gosto áspero 1I
Riada Bebida com leve sabor deiodofórrnio ou ácido fênico 7
Rio Bebida com sabor forte edesagradável lembrando Iiodofórmio ou ácido fênico
Classificação por TiposNo laboratório de classificação de qualidade docafé da Bolsa de Mercadorias e Futuros - SãoPaulo - SP, foi realizada a classificação por tipos,onde cada amostra foi apreciada sobre cartolinapreta, segundo as normas estabelecidas na "TabelaOficial Brasileira de Classificação" (Teixeira etal., 1974).
Metodologia para obtenção dos tratamentos:Efeito dos grãos defeituosos sobre acondutividade elétrica do exsudadoForam obtidas amostras de café cultivar MundoNovo, safra 199 J/92, do Departamento deAgricultura da ESALQ - Piracicaba e separadosos grãos com defeitos grão verde. grão brocado,grão ardido, grão preto e grão preto-verde ou"stinker" .Foram separadas 4 repetições de IO grãos de cadatipo de defeito para nelas determinar o peso dosgrãos, em balança de precisão de 0,00 I g; acondutividade elétrica, após imersão dos grãos em100 ml de água destilada (no interior de coposplásticos) a 25°C por 3,5 horas; a lixiviação depotássio na solução após 3,5 horas de embebição eo teor de umidade dos grãos.
465
Para a instalação deste experimento foi obedecidoo delineamento inteiramente casualizado comcinco repetições.
Efeito da temperatura de secagem de frutos decafé colhidos nos estádios de maturação cerejae verde sobre a condutividade elétrica dosgrãos.Em lavoura de café Mundo Novo linhagem LCP388-17 pertencente ao Departamento deAgricultura da ESALQ. em Piracicaba foi colhidauma mistura de frutos verdes e cerejas de café aosvinte e sete dias do mês de maio de 1991.Desta mistura foram separados manualmente osfrutos cerejas dos verdes e em seguida de cadaestádio de maturação foram compostas dezesseisparcelas de 600 gramas cada uma distribuídas nointerior de bandejas. Cada quatro parcelas defrutos cerejas e verdes foram postas a secarsimultaneamente à diferentes temperaturasconstituindo os seguintes tratamentos:l . Secagem natural à temperatura do ambiente delaboratório. mantida à 20°C,2. Secagem artificial em estuda de circulaçãoforçada de ar à 30°C,3. Secagem artificial em estuda de circulaçãoforçada de ar à 45°C,4. Secagem artificial em estuda de circulaçãoforçada de ar à 60°C.O grau de umidade inicial dos frutos cerejas everdes antes de iniciar a secagem eramrespectivamente de 69.7% e 71,5%. Atemperatura da massa de frutos durante toda aduração do experimento foi monitorada com acolocação de termômetros dentro das parcelas, aqual correspondeu perfeitamente as diferentestemperaturas selecionadas. Tomando por base oteor de água inicial e o peso de cada parcela.calculou-se o peso final de cada parcela quando osfrutos atingiam 12% de umidade final.A partir disto, periodicamente as parcelas forampesadas e finalmente retiradas da estufa aoatingirem 12% de umidade. A seguir as parcelasficaram armazenadas em ambiente de laboratóriopor três semanas, para uniformização do grau deumidade. Vencido este período de tempo cadaparcela foi descascada em máquina para retiradade amostras marca Pinhalense.De cada parcela foram separados e pesados osgrãos normais, verdes, verde-escuros. pretos epreto-verdes brilhantes. A seguir foi calculada aporcentagem de grãos classificados em cadacoloração. A seguir retirou-se de cada parcelaquatro amostras de 50 sementes as quais, apóspesadas e imersas em 75 ml de água destilada nointerior de copos de plástico a 25°C durante 3.5horas foram submetidas a análise da

condutividade elétrica em condutivímetroDigimed-CD-20. No exsudato de cada parcela foideterminada a quantidade de potássio emfotômetro de chama e os resultados expressos emppm/grama.De cada parcela foram determinados o tipo e aqualidade da bebida no laboratório declassificação de café da Bolsa de Mercadorias eFuturos em São Paulo - SP.A instalação deste experimento obedeceu adelineamento inteiramente casualizado emesquema fatorial 2 (Estádios de Maturação) x 4(Temperaturas de Secagem) com quatrorepetições.
Efeito de locais e métodos de colheita sobre acondutividade elétrica de exsudatos de grãos decaféEm dez locais, realizou-se a colheita de frutos decafé no estádio de maturação cereja, seguido dedespolpamento; frutos de café em diferentesestádios de maturação derriçados no pano,constituindo assim uma mistura: e frutos de caféno estádio de maturação seco recolhidos do chão,denominado café de varrição. Cada porção eraconstituída de aproximadamente 12 kilogramas decafé em coco.Os locais, com a latitude, longitude e altituderespectivas, de coleta dos lotes de cafédespolpado, mistura e varrição durante operíodode junho a setembro de 1991, foram:Piracicaba - SP 22° 45'S 47° 40'W 540 mPatrocínio - MG 18° 57'S 47° OO'W 933 m
Campinas - SP 22° 53'S 47° 06'W 562 mAlfenas - MG 21°28'S 45° 58'W 889 mMachado - MG 21° 40'S 45° 55'W 873 mMococa- MG 21°28'S 47° 01'W 665 mPindorama - SP 21° 13'S 48° 56'W 562 mGarça - SP 22° l4'S 49° 36'W 663 mLondrina - PR 2]0 23'S 51° 11'W 566 mMaringá - PR 23° 26'S 52° 02'W 355mApos a colheita os frutos de cafe foram secos ate12% de umidade e permaneceram armazenadosaté novembro de 1991, no interior de sacos dejuta, em ambiente apropriado na sala de sementesdo D.A.H. / ESALQ, Piracicaba.Em novembro de 1991, os diferentes lotes foramsubdivididos em cinco amostras de dois litroscada uma. Estas foram beneficiadas em máquina,marca Pinhalense, para retirada de amostra. Emseguida cada amostra foi subdividida em três subamostras de 100 gramas, as quais foramembaladas em sacos plásticos devidamenteidentificados com numeração de I a 150,constituindo assim três grupos de 150 amostras.Cada grupo de 150 amostras foi enviado aoslocais para determinação da qualidade da bebida.
466
Na porção restante de cada amostra foramseparadas quatro repetições de 50 grãos semescolha, as quais foram pesadas e imersas em 75rnl de água destilada, no interior de copos deplástico de 180 ml de capacidade, e colocados emcâmara a 25°C por 3,5 horas. Após este períodode embebição, procedeu-se a leitura dacondutividade elétrica da solução emcondutivímetro Digimed-CD-20. Os valoresobtidos em cada local de colheita foramsubmetidos a análise estatística. Foram analisadasas correlações entre as variáveis estudadas e osintervalos de confiança entre qualidade da bebidae condutividade elétrica.
RESllLTADOS E DISCUSSÃO
Efeito dos grãos defeituosos sobre acondutividade elétrica do exsudatoSubmeteu-se a anál ise de variância os dados depeso médio do grão, condutividade elétrica,lixiviação de potássio e teor de água apósembebição, obtendo-se para todos os parâmetrosaval iados valores significativos para efeito dostratamentos.Os grãos com dimensões semelhantes (tamanho)apresentaram pesos médios diferentes, sendo queos grãos normais ou sem defeitos apresentaram omaior peso médio. Os grãos defeituososapresentaram a seguinte ordem decrescente depeso: grãos verdes, ardidos, brocados, pretos epreto-verdes.Os valores de condutividade elétrica, lixiviação depotássio e teor de água após embebição foramsignificativamente menores nos grãos de cafénormais em relação aos grãos defeituosos, edentre os grãos com defeitos, os preto-verdesapresentaram os maiores valores.Considerando os dados da tabela 2, observa-se aseguinte ordem crescente de lixiviação de íons econdutividade elétrica: grãos normais, brocadossemelhantes aos verdes, ardidos, pretos e pretoverdes.Após a embebição, o teor de água ou grau deumidade dos grãos foram determinados eobedeceu a seguinte ordem crescente: grãosnormais, brocados, verdes, ard idos semelhantesaos pretos e preto verdes.Illy et aI. (1982) realizou um estudo sobre ascaracterísticas de reflectância dos grãosdefeituosos visando sua eliminação dos lotes decafé através de seleção eletrônica pela cor. Nesteestudo os autores além da refleciância estudaramtambém, através de microscopia eletrônica, ascaracterísticas da superfície e das células dosgrãos defeituosos. Verificaram que adesorganização celular aumenta dos grãos verdes
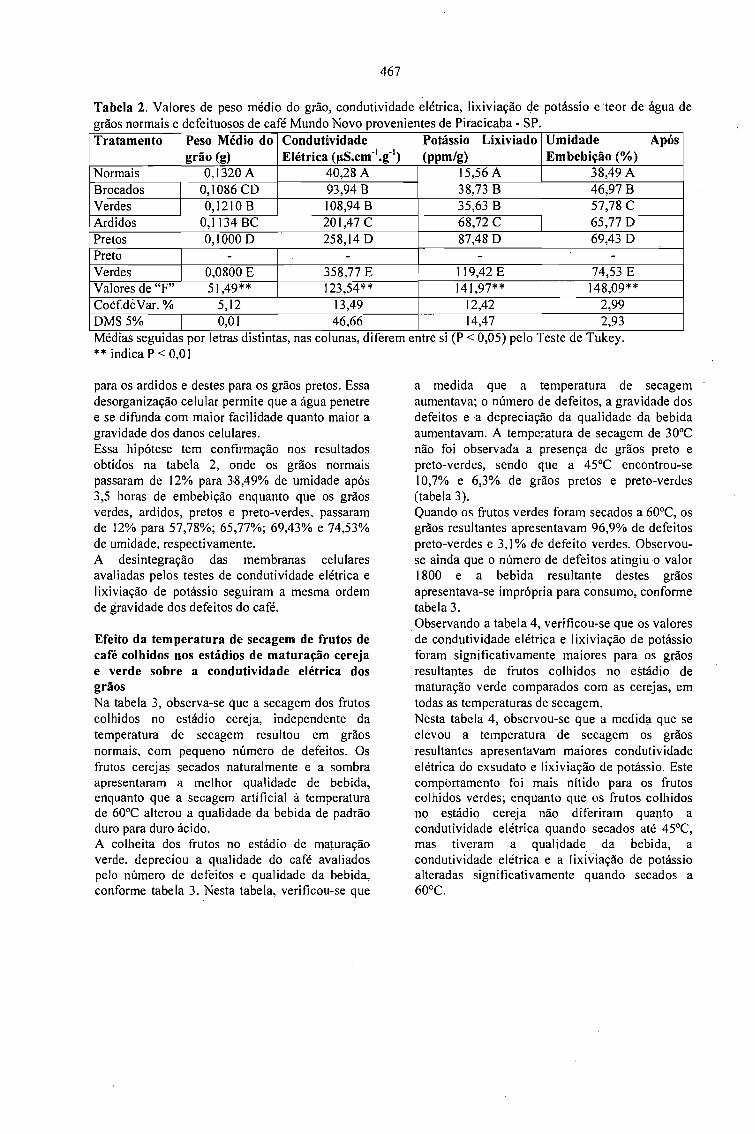
467
Tabela 2. Valores de peso médio do grão, condutividade elétrica, lixiviação de potássio e teor de água degrãos normais e defeituosos de café Mundo Novo provenientes de Piracicaba - SP.
Medias seguidas por letras distintas, nas colunas, diferem entre SI (P < 0,05) pelo Teste de Tukey.** indica P < 0,01
Tratamento Peso Médio do Condutividade Potássio Lixiviado Umidade Apósgrão (g) Elétrica (IJ.S.cm·l.g'l) (ppm/g) Em bebição (%)
Normais 0,1320 A 40,28 A ]5,56 A 38,49 A
Brocados 0,1086 CD 93,94 B 38,73 B 46;97 BVerdes 0,1210 B 108,94 B 35,63 B 57,78 CArdidos 0,1 ]34 BC 201,47 C 68,72 C 65,77 DPretos 0,1000 D 258,14 D 87,48 D 69,43 DPreto - - - -Verdes 0,0800 E 358,77 E 119,42 E 74,53 EValores de "F" 51,49** 123,54** 141,97** 148,09**Coef.deVar. % 5,12 13,49 12,42 2,99DMS5% 0,01 46,66 14,47 2,93. .
para os ardidos e destes para os grãos pretos. Essadesorganização celular permite que a água penetree se difunda com maior facilidade quanto maior agravidade dos danos celulares.Essa hipótese tem confirmação nos resultadosobtidos na tabela 2, onde os grãos normaispassaram de 12% para 38,49% de umidade após3,5 horas de embebição enquanto que os grãosverdes, ardidos, pretos e preto-verdes, passaramde 12% para 57,78%; 65,77%; 69,43% e 74,53%de umidade. respectivamente.A desintegração das membranas celularesavaliadas pelos testes de condutividade elétrica elixiviação de potássio seguiram a mesma ordemde gravidade dos defeitos do café.
Efeito da temperatura de secagem de frutos decafé colhidos nos estádios de maturação cerejae verde sobre a condutívídade elétrica dosgrãosNa tabela 3, observa-se que a secagem dos frutoscolhidos no estádio cereja, independente datemperatura de secagem resultou em grãosnormais, com pequeno número de defeitos. Osfrutos cerejas secados naturalmente e a sombraapresentaram a melhor qualidade de bebida,enquanto que a secagem artificial à temperaturade 60°C alterou a qualidade da bebida de padrãoduro para duro ácido.A colheita dos frutos no estádio de maturaçãoverde. depreciou a qualidade do café avaliadospelo número de defeitos e qualidade da bebida,conforme tabela 3. Nesta tabela, verificou-se que
a medida que a temperatura de secagemaumentava; o número de defeitos, a gravidade dosdefeitos ea depreciação da qualidade da bebidaaumentavam. A temperatura de secagem de 30°Cnão foi observada a presença de grãos preto epreto-verdes, sendo que a 45°C encontrou-se10,7% e 6,3% de grãos pretos e preto-verdes(tabela 3).Quando os frutos verdes foram secados a 60°C, osgrãos resultantes apresentavam 96,9% de defeitospreto-verdes e 3, I% de defeito verdes. Observouse ainda que o número de defeitos atingiu o valor1800 e a bebida resultante destes grãosapresentava-se imprópria para consumo, conformetabela 3.
,Observando a tabela 4, verificou-se que os valoresde condutividade elétrica e lixiviação de potássioforam significativamente maiores para os grãosresultantes de frutos colhidos no estádio dematuração verde comparados com as cerejas, emtodas as temperaturas de secagem.Nesta tabela 4, observou-se que a medida que seelevou a temperatura de secagem os grãosresultantes apresentavam maiores condutividadeelétrica do exsudato e lixiviação de potássio. Estecomportamento foi mais nítido para os frutoscolhidos verdes; enquanto que os frutos colhidosno estádio cereja não diferiram quanto acondutividade elétrica quando secados até 45°C,mas tiveram a qualidade da bebida, acondutividade elétrica e a lixiviação de potássioalteradas significativamente quando secados a60°C.
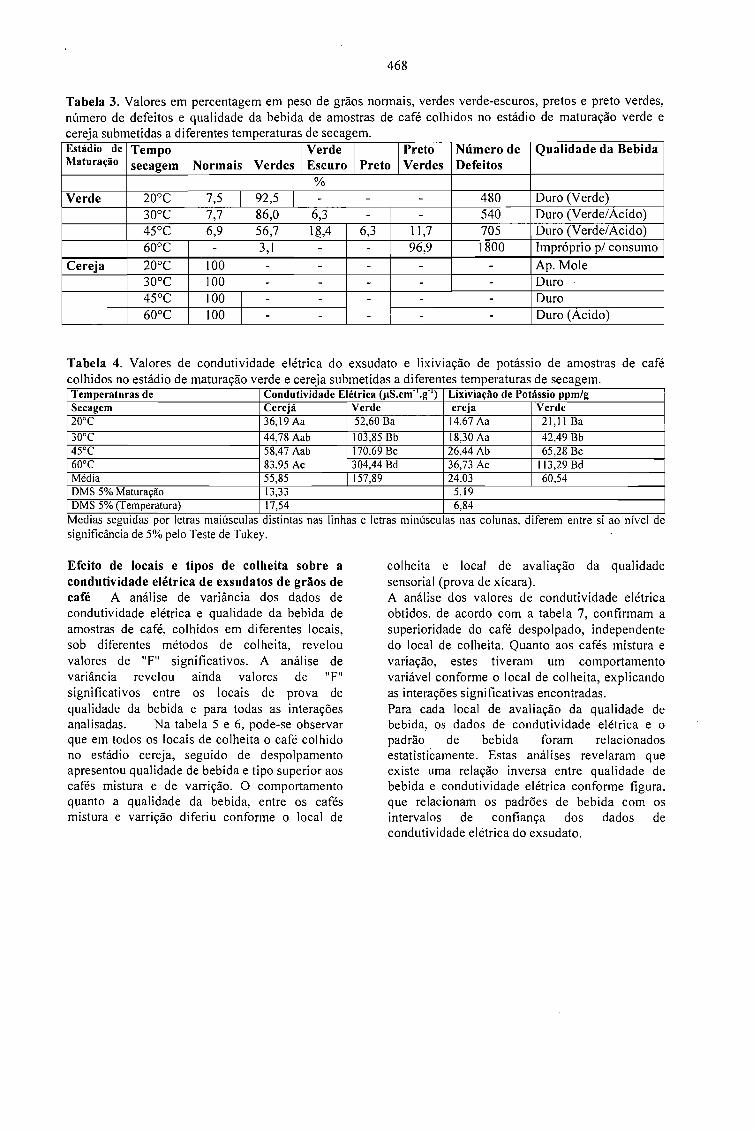
468
Tabela 3. Valores em percentagem em peso de grãos normais, verdes verde-escuros, pretos e preto verdes,número de defeitos e qualidade da bebida de amostras de café colhidos no estádio de maturação verde ecereja submetidas a diferentes temperaturas de secagem.Estádio de Tempo I IVerde I IPreto Número de Qualidade da BebidaMaturação secagem Normais Verdes Escuro Preto Verdes Defeitos
%Verde 20°C 7,5 92,5 - - - 480 Duro (Verde)
30°C 7,7 86,0 6,3 - - 540 Duro (Verde/Acido)45°C 6,9 56,7 1~,4 6,3 11,7 705 Duro (Verde/Acido)
60°C - 3, I - - 96,9 1800 Impróprio p/ consumo
Cereja 20°C 100 - - - - - Ap. Mole30°C 100 - - - - - Duro45°C 100 - - - - - Duro60°C 100 - - - - - Duro (Acido)
Tabela 4. Valores de condutividade elétrica do exsudato e lixiviação de potássio de amostras de cafécolhidos no estádio de maturação verde e cereja submetidas a diferentes temperaturas de secagem.
Medias seguidas por letras maiúsculas distintas nas linhas e letras minúsculas nas colunas. diferem entre SI ao nível designificância de 5% pelo Teste de Tukey.
Temperaturas de Condutividade Elétrica (,..S.cm" .g" ) Lixiviação de Potássio ppm/g,
Secagem Cerejá Verde ereja Verde200 e 36,19Aa 52.60Ba 14.67 Aa 21,11 Ba300 e 44.78 Aab 103,85 Bb 18,30 Aa 42,49Bb45°e 58,47 Aab 170.69 Bc 26,44Ab 65,28Bc600 e 83.95 Ac 304,44 Bd 36,73 Ac 113,29 BdMédia 55,85 157,89 24.03 60,54DMS 5%Maturação 13.33 5,19DMS 5%(Temperatura) 17,54 6,84
"
Efeito de locais e tipos de colheita sobre acondutividade elétrica de exsudatos de grãos decafé A análise de variância dos dados decondutividade elétrica e qualidade da bebida deamostras de café, colhidos em diferentes locais,sob diferentes métodos de colheita, revelouvalores de "F" significativos. A análise devariância revelou ainda valores de "F"significativos entre os locais de prova dequalidade da bebida e para todas as interaçõesanalisadas. Na tabela 5 e 6, pode-se observarque em todos os locais de colheita o café colhidono estádio cereja, seguido de despolpamentoapresentou qualidade de bebida e tipo superior aoscafés mistura e de varrição. O comportamentoquanto a qualidade da bebida, entre os cafésmistura e varrição diferiu conforme o local de
colheita e local de avaliação da qualidadesensorial (prova de xícara).A análise dos valores de condutividade elétricaobtidos. de acordo com a tabela 7, confirmam asuperioridade do café despolpado, independentedo local de colheita. Quanto aos cafés mistura evariação, estes tiveram um comportamentovariável conforme o local de colheita, explicandoas interações significativas encontradas.Para cada local de avaliação da qualidade debebida, os dados de condutividade elétrica e opadrão de bebida foram relacionadosestatisticamente. Estas análises revelaram queexiste uma relação inversa entre qualidade debebida e condutividade elétrica conforme figura,que relacionam os padrões de bebida com osintervalos de confiança dos dados decondutividade elétrica do exsudato.
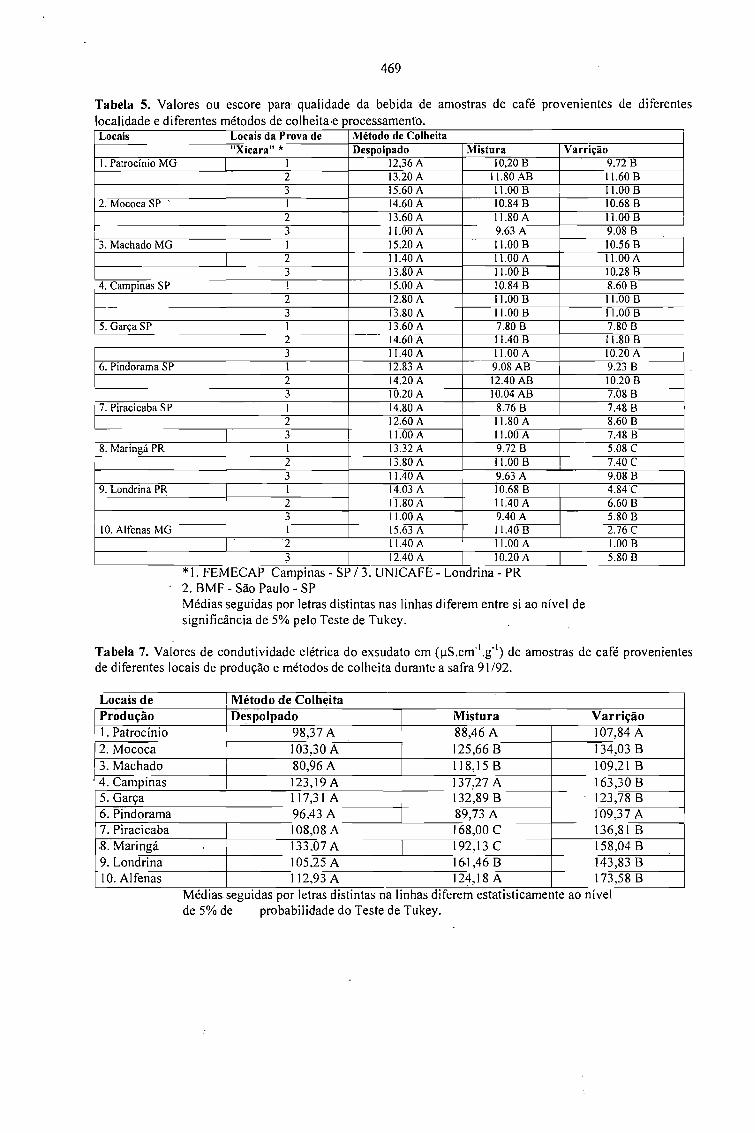
469
Tabela 5. Valores ou escore para qualidade da bebida de amostras de café provenientes de diferenteslocalidade e diferentes métodos de colheitae processamento.Locais Locais da Prova de Método de Colheita
"Xícara" * Despolpado Mistura Varrição1. Patrocínio MG I 12,36 A 10,20 B 9.72 B
2 13.20 A 11.80 AB 11.60 B3 15.60 A 11.00 B 11.00 B
2. Mococa SP , 1 14.6DA 10.84 B 10.68 B2 13.60 A 11.80 A 11.0083 11.00 A 9,63 A 9,088
3. Machado MG I 15.20 A 11.008 10,56 B2 11.40 A 11.00 A 11.00 A3 13.80 A 11.00 B 10.28 B
4, Campinas SP ! 15.00 A 10.84 B 8.60 B2 12.80 A 11.00 B 11.00 B3 13.80 A 11.008 11.00 B
5. Garça SP 1 13.60 A 7.80 B 7.80 B2 14.60 A 11.408 11.80 B3 11.40 A 11.00 A 10.20 A
6. Pindorama SP I 12.83 A 9.08 AB 9.23 B2 14.20 A 12.40 AB 10.20 B3 10.20 A 10.04 AB 7.08 B
7. Piracicaba SP I 14.80 A 8.76 B 7.48 B I
2 12.60 A 11.80 A 8.60 B3 11.00 A 11.00 A 7.48 B
8. Maringá PR I 13.32 A 9.72 B 5.08 C2 13.80 A 11.00 B 7.40 C3 11.40 A 9;63 A 9.08 B
9. Londrina PR I 14.03 A 10.68 B 4.84C2 11.80 A 11.40 A 6.6083 11.00 A 9.40 A 5.80 B
10. Alfenas MG 1 15.63 A 11.40 B 2.76 C2 11.40 A 11.00 A 1.00 B3 12.40 A 10.20A 5.80 B
*I. FEMECAP Campinas - SP 13. UNICAFE - Londrina - PR2. BMF - São Paulo - SPMédias seguidas por letras distintas nas linhas diferem entre si ao nível designificância de 5% pelo Teste de Tukey.
Tabela 7. Valores de condutividade elétrica do exsudato em (u.S.cm'l.g") de amostras de café provenientesde diferentes locais de produção e métodos de colheita durante a safra 91/92.
Medias seguidas por letras distintas na Iinhas diferem estatisticamente ao nívelde 5% de probabilidade do Teste de Tukey.
Locais de IMétodo de ColheitaProdução Despolpado Mistura VarriçãoI. Patrocínio 98,37 A 88,46 A 107,84 A2. Mococa J03,30 A 125,66 B J34,03 B3. Machado 80,96 A 118,15 B 109,21 B
14. Campinas ]23,19 A 137,27 A 163,30 B5. Garça 117,31 A 132,89 B 123,78 B6. Pindorama 96,43 A 89,73 A 109,37 A7. Piracicaba 108,08 A 168,00 C 136,81 B8. Maringá 133,07 A 192,13 C 158,04 B9. Londrina 105,25 A 161,46 B 143,83 B10. Alfenas 112,93 A 124,18 A 173,58 B..
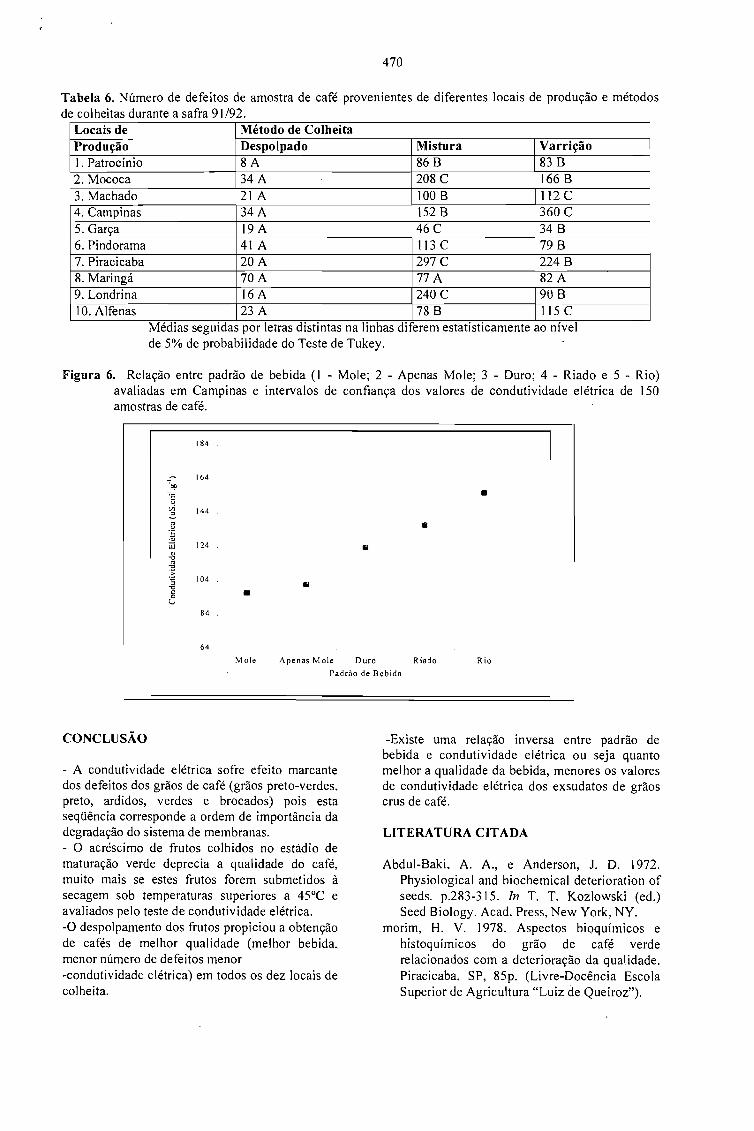
470
Tabela 6. Número de defeitos de amostra de café provenientes de diferentes locais de produção e métodosde col heitas durante a safra 91/92.
Locais de Método de ColheitaProdução Despolpado Mistura Varrição1. Patrocínio 8A 86 B 83 B2. Mococa 34 A 208 C 166 B3. Machado 21 A 100 B 112 C4. Campinas 34A 152 B 360 C5. Garça 19A 46 C 34 B6. Pindorama 41 A 113 C 79 B7. Piracicaba 20A 297 C 224 B8. Maringá 70A 77A 82 A9. Londrina 16A 240 C 90 BlO. Alfenas 23 A 78B 115 C..Médias seguidas por letras distintas na linhas diferem estatisticamente ao nível
de 5% de probabilidade do Teste de Tukey.
Figura 6. Relação entre padrão de bebida (I - Mole; 2 - Apenas Mole; 3 - Duro; 4 - Riado e 5 - Rio)avaliadas em Campinas e intervalos de confiança dos valores de condutividade elétrica de 150amostras de café.
184
IM"7O!J
•'1:Li
<li )442-"" •.g
.<;u:i 124 •"-e""-e:~
104'5 •-e" •o
U
84
64
Mole Apenas Mole Duro Riado Rio
Padrão de Bebida
CONCLUSÃO
- A condutividade elétrica sofre efeito marcantedos defeitos dos grãos de café (grãos preto-verdes.preto, ardidos, verdes e brocados) pois estaseqüência corresponde a ordem de importância dadegradação do sistema de membranas.- O acréscimo de frutos colhidos no estádio dematuração verde deprecia a qualidade do café,muito mais se estes frutos forem submetidos àsecagem sob temperaturas superiores a 45°C eaval iados pelo teste de condutividade elétrica.-o despolpamento dos frutos propiciou a obtençãode cafés de melhor qualidade (melhor bebida.menor número de defeitos menor-condutividade elétrica) em todos os dez locais decolheita.
-Existe uma relação inversa entre padrão debebida e condutividade elétrica ou seja quantomelhor a qualidade da bebida, menores os valoresde condutividade elétrica dos exsudatos de grãoscrus de café.
LITERATURA CITADA
Abdul-Baki, A. A., e Anderson, J. D. 1972.Physiological and biochemical deterioration ofseeds. p.283-315. In T. T. Kozlowski (ed.)Seed Biology. Acad. Press, New York, NY.
morim, H. V. 1978. Aspectos bioquímicos ehistoquímicos do grão de café verderelacionados com a deterioração da qualidade.Piracicaba. SP, 85p. (Livre-Docência EscolaSuperior de Agricultura "Luiz de Queiroz").

Amorim, H. V., e Mello, M. 1992. Significance ofEnzymes in Nonalcoholic Coffee Beverage.p.189-209 In P. F. Fox (ed.) FoodEnzymology. Elsevier , London.
Amorim, H. V., e Silva, D. M. 1968. Relationshipbetween the polyphenol oxidase activity ofcoffee beans and the qual ity of the beverage.Nature.219:381-382.
Ministério da Agricultura. 1976. Regras paraanálise de sementes. Dept", Nac. Veg. OISEM.188p.
Clifford, M. N. 1985. Chemical and PhysicalAspects of Green Coffee and Coffee Products.p.305-359. In M. N. Clifford e K. C. Wilson(ed.) Coffee, Botany, Brochemsitry androduction of Beans and Beverage. Croomhelm, London.
Garruti, R. S., e Conagin, A. 1961. Escala devalores para avaliação da qualidade de bebidade café. Bragantia 20:557-562.
IlIy, E., Brumen, G., Mastropasqua, L., eMaughan, W. 1982. Study on thecharacteristics and the industrial sorting ofdefective beans in green coffee lots, p.99-128.In Coloquio Científico Internacional sobre oCafé, 10. Salvador, BA.
Loeftler, T. M., Tekrony, D. M., e Egli, D.B.1988. The bulk conductivity test as anindicator of soybean seed quality. Journal ofSeed Technology 12:37-53.
Mathews, . S., e Bradnock, W. T. 1968.Relationship between seed exudation and fieldemergence in peas and french beans. Hort.Res.8:89-93.
471
Miya, E. E., Garrut, R. S., Chaib, M. A.,Angelucci, E., Figuereido, r., e Shirose, r.1973/4. Defeitos do café e qualidade dabebida. Co!. Int, Tecn. Alim. 5:417-432.
McDonald Jr., M. 8., e Wilson, D. O. 1979. Anassessment of the standadization and theabi Iity of the ASA-610 to rapidly predictpotential soybean germination. Journal of SeedTechology 4: 01-11.
Ministério da Agricultura. 1976. Regras paraanálise de sementes. Dept", Nac. Veg. OISEM.188p.
Powell, A. A. 1986. Cell membranes and seedleachate conductivity in relation to the qualityof seed for sowing. Journal of SeedTechnology 10:81-100.
Steere, W. c., Leveengood, W. C., e Bondie, 1.M. 1981. An eletronic for eval uating seedgerminaion and vigor. Seed Science andTechnology 9:567-576.
Teixeira, A. A., Pereira, L. S. P., e Pinto, J. C. A.1974. Classificação de café. Noções Gerais.IBC/GERCA - RJ, 88P.
Yaklich, R. W., Kulik, M. M., e Anderson, 1. D.1979. Evaluation of vigor tests in soybeanseeds: relationship of ATP, conductibity andradioactive trace r multi pIe interia laboratorytests to field performance. Crop Science19:806-810.


473
RELAÇÃO ENTRE O PESO E A CONDUTIVIDADE ELÉTRICA DE GRÃOS DE CAFÉ DEPROGÊNIES DE ICATU.
Pretel , c.E.c.; Sera2, T. e Fonseca l , I.C.B.
IProfessores do Departamento de Agronomia/Universidade Estadual de Londrina, Londrina - PR, Brasil.2Pesquisador do Instituto Agronômico do Paraná, Londrina - PR, Brasil.
SUMMARYRELATIONSHlP BETWEEN THE WEIGHT AND THE ELECTRIC CONDUCTIVITY OFCOFFEE BEANS OF PROGENIES OF ICATU.In this study, 500 weight and electric conductivity analysis of23 co./fee progenies cv !catu were correlated.The samples were obtained of competition assay conducted in the Instituto Agronômico do Paraná,Londrina, in july 1993. After the harvest the fruits were peeled for retreat of the pulp and drying untilhumidity degrees dose of 11% w.b. Each portion was handled in sample mills, and 4 repetitions of 50grains of each sample were picked, and accurately weighed in a scale of two decimais. After weighting,each sample was immersed in 75 ml of deionized water at 25°C for 3.5 hours, followed by electriccondutivity determination expressed in j.1S.cm-1.g-l. The correlation was negative between the weight ofgrains and electric conductivity variables, which was significant at the levei of5% ofprobability; but withlittle intensity (r = -0,28). The result conjirm the tendency that as larger and heavier the co./feebeans higherthe probabilitiy of obtaining a good quality beverage. However this relationship not reach the sufficientintensity to predict the quality, suggesting that a use ofthis hypothesis in programs of improvement ofco./feeis not recommended.
INTRODUÇÃO
Popularmente é aceito que grãos de café detamanhos grandes são indicadores de boaqualidade de bebida. Evidentemente grãos de cafébem formados, com pleno desenvolvimento, como máximo de acúmulo de nutrientes e matériaseca proporcionarão um lote com menor númerode defeitos que grãos miúdos, podendo estacaracterística refletir na qualidade da bebida.Lazarini e Moraes (1958) e Teixeira et ai (1974)encontraram uma maior concentração de defeitosa medida que a classe de tamanho dos grãos ou,peneiras, do café diminuía, pois a probabilidadede encontrarmos defeitos verdes. pretos e ardidos,é maior nas porções de cafés mal formados ou demenores peneiras.Roche (1995) avaliou 15 cultivares de caféproduzidos no Hawaii e não encontrou correlaçãoentre o tamanho dos grãos e qualidade da bebida,concluindo que o tamanho dos grãos não era umbom indicador da qualidade da bebida quandocomparava cultivares de uma única área deprodução.O objetivo deste trabalho foi verificar a hipótesede que quanto maior o peso dos grãos melhor é aqualidade do café, avaliada indiretamente peloteste de condutividade elétrica.
MATERIAL E MÉTODOS
Foram correlacionadas 500 observações entre opeso de grãos e a condutividade elétrica de 23progênies de café cv. Icatu. As amostras paradeterminação dos dados foram obtidas de umensaio de competição de progênies das geraçõesF3, F4, F5 e F6 de retrocruzamento 2, dos ensaiosde Icatu conduzidas pelo IAPAR. Os frutoscerejas foram coletados na primeira quinzena dejunho de 1993. E a seguir foram descascados pararetirada do endocarpo, obtendo assim "cerejasdescascadas" que foram secos em terreiro atéatingirem grau de umidade próximo de 11%.Cada parcela obtida foi beneficiada em moinhopara amostras e os grãos de café obtidos forampesados em balança com precisão de duas casasdecimais.Após a pesagem de cada amostra de 50 grãos foiimersa em 75 rnl de água deionizada, à 25°C por3,5 horas de embebição seguida de leitura emcondutivímetro elétrico DIGIMED CD-21expressando os resultados em flS.cm-1.g-1.A análise de correlação foi feita utilizando-se oprograma "Statistica for Windows" (1993),versão 4.3.
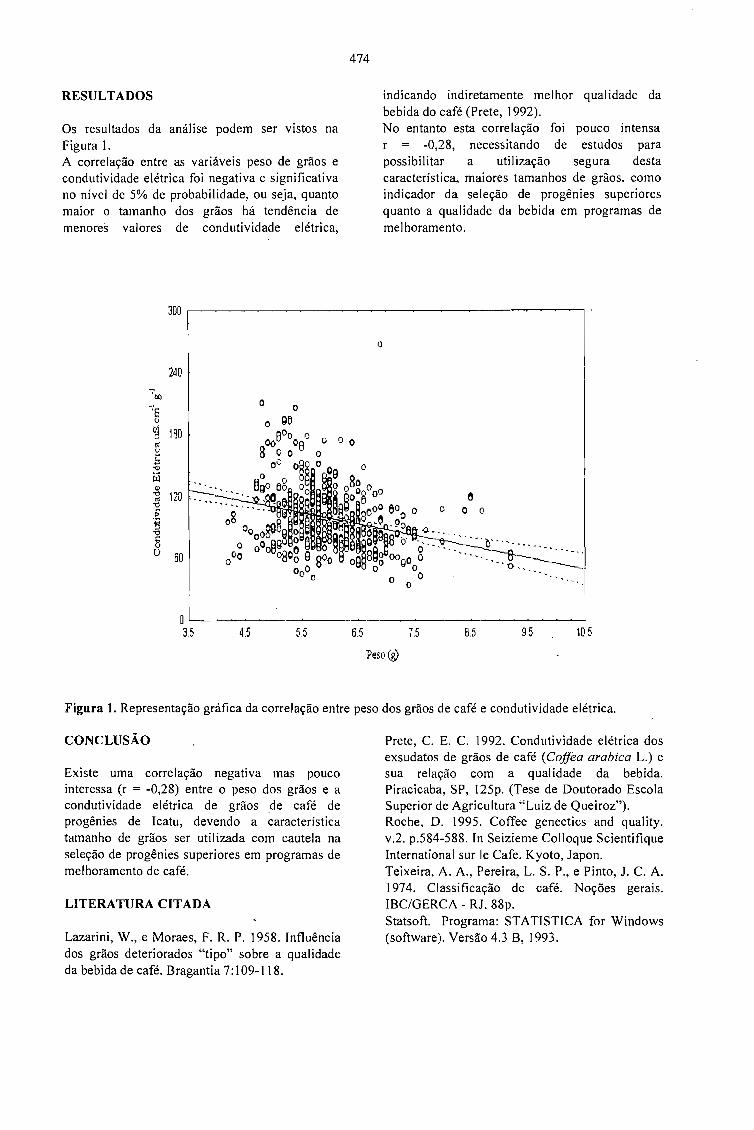
RESULTADOS
Os resultados da análise podem ser vistos naFigura 1.A correlação entre as variáveis peso de grãos econdutividade elétrica foi negativa e significativano nivel de 5% de probabilidade, ou seja, quantomaior o tamanho dos grãos há tendência demenores valores de condutividade elétrica,
474
indicando indiretamente melhor qualidade dabebida do café (Prete, 1992).No entanto esta correlação foi pouco intensar -0,28, necessitando de estudos parapossibilitar a utilização segura destacaracterística, maiores tamanhos de grãos. comoindicador da seleção de progênies superioresquanto a qualidade da bebida em programas demelhoramento.
300 I
Io
240 I"70() i"7E
o
"~ 1BO o ü o"vli~
iii'"-e 120 e"-o.:;§-ecoo 60
' ..o
o35 4.5 5.5 6.5 7.5 8.5 95 105
Peso (g)
Figura 1. Representação gráfica da correlação entre peso dos grãos de café e condutividade elétrica.
CONCLUSÃO
Existe uma correlação negativa mas poucointeressa (r = -0,28) entre o peso dos grãos e acondutividade elétrica de grãos de café deprogênies de Icatu, devendo a característicatamanho de grãos ser utilizada com cautela naseleção de progênies superiores em programas demelhoramento de café.
LITERATURA CITADA
Lazarini, W., e Moraes, F. R. P. 1958. Influênciados grãos deteriorados "tipo" sobre a qualidadeda bebida de café. Bragantia 7:109-118.
Prete, C. E. C. 1992. Condutividade elétrica dosexsudatos de grãos de café (Coffea arabica L.) esua relação com a qualidade da bebida.Piracicaba, SP, 125p. (Tese de Doutorado EscolaSuperior de Agricultura "Luiz de Queiroz").Roche, D. 1995. Coffee genectics and quality.v.2. p.584-588. In Seizieme Colloque ScientifiqueInternational sur le Cafe. Kyoto, Japon.Teixeira, A. A., Pereira, L. S. P., e Pinto, J. C. A.1974. Classificação de café. Noções gerais.IBC/GERCA - RJ. 88p.Statsoft. Programa: STA TISTICA for Windows(software). Versão 4.3 B, 1993.

475
CONDUTIVIDADE ELÉTRICA DE EXSUDATOS DE GRÃOS DE CAFÉ COLHIDOS EMDIFERENTES ESTÁDIOS DE MATURAÇÃO.
Pretel , C.E.C.; Sera 2, T.; Crudi3, C.E. e Fonseca l, I.C.B.
I Professores do Departamento de Agronomia/Universidade Estadual de Londrina, Londrina - PR, Brasil.2 Pesquisador do Instituto Agronômico do Paraná, Londrina - PR, Brasil.3 Estagiário do Curso de Agronomia/Universidade Estadual de Londrina.
SUMMARYELECTRIC CONDUCTIVITY OF EXSUDATES OF COFFEE BEANS HARVESTED INDIFFERENT STAGES OF MATURATION.This work was developed with the objective to verify the effect ofdifferent stages ofcoffee maturation on thevalues of electric conductivity, which causes inverse relation on the beverage quality. The fruits of coffeecv. Red Catuaí were harvested in different stage of maturation: green, green-cane, cherry (ripe), raisinstage and dry stage in the Agronomic Institute ofParaná/Londrina during July, J993. Four replications of2liters offruits were picked for each maturation stage. The fruits were placed to dry on sieves with periodicshaking until reach JJ% of humidity. Soon after the samples were hulled and each portion separated infour samples of 50 grains were weighted, immersed in 75 ml of distilled water at 25°C for 3.5 hours,followed by electric condutivity determination, expressed in pS.cm-J.g-J. For each maturation stage the
following medium values of electric conductivity were obtained: green, J49.24 pS.cm-J.g-J; green-cane,
J05.72 pS.cm-J.g-J; cherry, 76.62 pS.cm-l.g-J; raisin, 79.60 pS.cm-J.g-J; and dry, 76.04 pS.cm-J.g-J. Thestatistical analysis of the data indicates that the fruits harvested in the green and green-cane stage hadsignificantly inferior quality, once as larger the values of electric conductivity, worst were the qualities ofthe beverage. The results conjirm the need to begin the crop harvest with the smallest possible percentage ofgreenfruits, with intention to preserve the quality ofthe coffee beans.
INTRODUÇÃO
o aumento na procura de cafés de qual idadesuperior fez com que o Brasil perdesse umaparcela significativa do mercado externo. Para aobtenção de um café de boa qualidade énecessário a aplicação de técnicas adequadas decolheitas e preparo do café, facilitando dessaforma sua comercial ização e obtendo maioresretornos econômicos. Sendo assim a melhorépoca de realizar a colheita é quando o caféapresenta-se em seu ponto ótimo de maturação(maturação fisiológica), isto é, nos estádios decereja. Caso a colheita seja realizadaprecocemente, nos estádios verde e verde-cana,aparecerá um grande número de grãos verdes oque depreciará a qualidade do café. O café porapresentar mais de uma floração, proporcionanuma mesma planta. frutos em diferentes fases dematuração, sendo assim importante efetuar acolheita no momento em que a maioria dessesfrutos se encontrarem no ponto ideal dematuração que é o estádio de cereja.Ao trabalharem com cafés em vários estádios dematuração como verde. verde-cana, cereja, passae seco, Freire e Miguel (1985), demonstraram quea máxima qualidade do fruto se dá no estádio
cereja; ja o café colhido precocemente comgrande percentual de verde, além de proporcionarprejuízo no tipo e bebida, poderá também atingirum índice de 20% de perdas em relação aorendimento final. Para Teixeira (1984), o cafécolhido no estádio de maturação verde apresentaaspecto de torração de pior qualidade, comconseqüente inferioridade na qualidade de bebida,além de apresentarem menor peso e tamanho dosgrãos.As transformações bioquímicas indesejáveis queocorrem no grão de fruto verde, antes e após acolheita, e que conduzem a formação de umabebida inferior, são principalmente de naturezaenzimática, envolvendo a polifenoloxidase,gl icosidase, lipase e protease. Algumas dessastransformações bioquímicas degradam as paredese membranas celulares. outras podem mudar acolaboração do grão e da película prateada,estando a qualidade de bebida sensivelmentealterada por estas modificações (Amorim eTeixeira, 1975).Em sua tese de mestrado, Pimenta (1995),demonstrou através de análises físico-químicasque entre grãos no estádio cereja, passa e seco asdiferenças dos teores de acidez titulável, sólidossolúveis, açúcares totais, compostos fenólicos,

476
RESULTADOS
Estádios de MaturaçãoVerde Verde Passa Maduro Seca
Cana149,24a 105,73 b 79,60 c 76.63 c 76.04 c
(I)
.. -
Quadro I. Condutividade elétrica média (~S.cm-I.g-I)
de exsudatos de grãos de café colhidos nos diferentesestádios de maturação.
No Quadro I encontram-se os resultados do testede condutividade elétrica, onde pode serconstatado que os grãos no estádio cereja, passa eseco obtiveram os melhores resultados, ou seja,menores valores de condutividade elétrica, e nãodiferiram entre si significativamente. Já os frutosno estádio verde e verde-cana apresentaramcondutividade elétrica elevada refletindo ummaior dano ou degradação do sistema demembranas (desorganização do sistema demembranas), o que ocasiona uma maiorlixiviação de Íons..
(I) Medias seguidas de mesma letra nao diferem entresi no nível de 5% de significância pelo teste de Tukey.DMS = 21,17 (uSrcml.g')
próxima a Ii %. Após o beneficiamento dosgrãos, realizou-se o teste de condutividadeelétrica do exsutados desenvolvido por Prete(1992), que consiste na utilização de quatroamostras de 50 grãos de café, sem escolha,pesadas com precisão de O, Ig, imersas em 75 mlde água destilada (no interios de copos plásticosde 180 ml de capacidade), e colocadas emambientes à 25°C por 3,5 horas, seguida deagitação e leitura em condutivírnetro elétrico,expressando os resultados em IlS.cm-l.g-lO estudo do efeito de diferentes estádios dematuração sobre os valores de condutividadeelétrica foi realizado através da análise devariância complementada pelo teste decomparação múltipla de Tukey, no nível de 5% designificância.
polifenoloxidase e qualidade de bebida nãoapresentaram diferenças significativa.Amorim (1978) constatou que cafés de piorqualidade influenciados pela presença do defeitoverde apresentam valores mais elevados delixiviação de potássio. Prete (1992), confirmou asobservações de Amorim (1978) verificando quecafés com maiores quantidades de defeito verdeapresentam maiores valores de condutividadeelétrica.O teor de acidez titulável em grãos de café podevariar de acordo com os níveis de fermentaçãoque ocorrem nos grãos e também com osdiferentes estádios de maturação dos mesmos.podendo também servir como suporte paraauxiliar na avaliação da qualidade de bebida docafé. Alguns autores como Arcila - Pulgarin eValência - Aristizabal (1975), verificaram em seutrabal ho que os frutos de café no estádio dematuração verde possuem menores teores deacidez titulável, que aumenta a medida que seintensifica o processo de maturação dos frutos.Com relação aos prejuízos causados ao tipo docafé, Nobre et aI. (1980) através de experimentoconstatou que o melhor tipo ocorreu para ostratamentos cereja e passa. Notável tendência dedesmerecimento do tipo do café (inferior a 8) foiobservado para o tratamento verde, ficandointermediário o tipo no tratamento verde-cana.Em todos os cultivares testados, o tratamentoverde foi o que apresentou menor porcentagem degrãos graúdos, diferindo estatisticamente dosdemais tratamentos. O tratamento verde-canaapresentou comportamento intermediário e ostratamentos cereja e passa apresentaram melhordesempenho.Com base no conhecimento da influência doestádio de maturação dos frutos na qualidade docafé, o presente trabalho tem por objetivoverificar o efeito de diferentes estádios dematuração do café sobre os valores decondutividade elétrica, a fim de avaliar o melhorestádio para realizar a colheita.
MATERIAL E MÉTODOS
O experimento foi realizado no InstitutoAgronômico do Paraná de Londrina em 7 de julhode 1993, onde foram colhidos no pano, frutos decafé cv. Catuaí Vermelho nos estádios dematuração: verde, verde-cana, cereja (maduro),passa e seco, sendo 4 repetições de 2 litros defrutos em cada estádio de maturação. Os frutosforam colocados para secar sobre peneiras comrevolvimento periódico até atingirem umidade
O melhor desempenho do grão no estádio seco foidecorrente das condições cl imáticas secas, semchuva, desfavorável ao desenvolvimento depatógenos que poderiam comprometer aqualidade dos frutos secos na planta.
CONCLUSÃO
Através dos resultados obtidos conclui-se que acolheita deve ser iniciada quando houver omínimo de grãos verdes na planta, ainda que isso

implique no aumento do número de grãos secosna planta.
LITERATURA CITADA
Amorim, H. V. 1978. Aspectos bioquímicos ehistoquímicos do grão de café verderelacionados com a deterioração da qualidade.Piracicaba, SP, 85p. (Livre-Docência EscolaSuperior de Agricultura "Luiz de Queiroz").
Amorim, H. V., e Teixeira, A. A. 1975.Transformações bioquímicas, químicas efísicas dos grãos de café verde e a qualidadeda bebida. In Congresso Brasileiro dePesquisas Cafeeiras, Curitiba, PRo p.21
Arcila-Pulgarin, J., e Valência-Aristizabal, G.1975. Relaciõn entre la actividad de lapolifenoloxidase (PFO) y las pruebas decataciõn como medidas de la bebida de café.Cenicafé 26: 55-71.
Freire, A. C. F., e Miguel, A. C. 1985.Rendimento e qualidade do café colhido nosdiversos estádios de maturação em VarginhaMG. In Congresso Brasileiro de PesquisasCafeeiras, 12. Caxambu, MG. p.21 0-214
477
Nobre, G, W., Teixeira, R. A. F., e Carvalho, C.H. S. 1980. Rendimento e qualidade do caféem frutos colhidos em .diferentes estádios dematuração. p.417-419. In Congresso Brasileirode Pesquisas Cafeeiras, 8. Campos do Jordão,SP.
Pimenta, C. J. 1995. Qualidade do café (Co./feaarabica L.) originado de frutos colhidos emquatro estádios de maturação (Dissertação deMestrado Escola Superior de Agricultura"Luiz de Queiroz").
Prete, C. E. C. 1992. Condutividade elétrica dosexsudatos de grãos de café (Co./fea arabica L.)e sua relação com a qualidade da bebida.Piracicaba, SP, 125p. (Tese de DoutoradoEscola Superior de Agricultura "Luiz deQueiroz").
Teixeira, A. A. 1984. Observações sobre váriascaracterísticas do café colhido verde e maduro.In Congresso Brasileiro de PesquisasCafeeiras, 11. Londrina, PRo p.227-228


479
AVALIAÇÃO DE PROGÊNIES DE CAFÉ (Coffea arabica L. cv. Icatu) QUANTO ACONDUTIVIDADE ELÉTRICA DO EXSUDATO.
Pretel , C.E.C.; Sera2, T. e Fonsecal , LeB.
IProfessores do Departamento de Agronomia/Universidade Estadual de Londrina, Londrina - PR, Brasil.2 Pesquisador do Instituto Agronômico do Paraná, Londrina - PR, Brasil.
SUMMARYEVALUATION OF COFFEE Coffea Arabic L. cv, iCATU PROGENIES BY ELECTRICCONDUCTlV1TY OF EXSUDATEWith the objective to select promising strains, the quality of coffee cv. Icatú was evaluated indirectly byelectric conductivity. The samples were obtained by an assay schedule using competition of 23 coffee cv.Icatú progenies cultivated in at Agronomic Institute of Paraná/Londrina, which were harvested during themonth of July, 1993. As the quality pattern reference, the coffee cv. Red Bourbon was used. The sampleswere picked in a sieve, peeled to retreat the pulp, obtaining like this peeled cherries, that were dry in yarduntil humidity of 11% w.b. Each portion was handled in sample mills, and the obtained coffee beansweighted, immersed in 75 ml of deionized water (plastic glasses of 180 ml) and placed in a atmosphere of25°C for 3.5 hours. After this time the electric conductivity oflhe samples were determined, and the resultsexpressed in uS.cm-l.g-l. Except one strain, ali ofstrains showed no difJerence each other when comparedto the reference strain Red Bourbon. The strain Icatú no. 20 showed the lowest electric conductivity value,deserving more detailed studies to confirm its superior characteristics.
INTRODUÇÃO
Todos os atributos do café, incluindo o aroma esabor da bebida são influenciados pelo ambiente eseus limites são determinados pelo genótipo.Carvalho (1982) descreve a origem de cafeeirosdo germoplasma Icatú, que em língua tupisignifica bonança (Monaco et al., 1974). Foiobtido a partir do cruzamento inicial de C.arabica cv. Bourbon Vermelho com o C.canephora cv. Robusta em 1950 e de doisretrocruzamento para C. arabica cv. MundoNovo, com seleções subsequentes praticadas nasgerações F2, F3 e F4 de polinizações livres dasmelhores plantas. Portanto, Icatu é umgermoplasma que tem cerca de 87,5% dos genesde C. arabica e 12,5% de genes do C. canephora.Apesar de ter genes de C. canephora, o Icatu temapresentado qualidade de bebida semelhante a C.arabica, tanto em materiais com um ou com doisretrocruzamentos, levando a conclusão de que osfatores determinantes de qualidade de bebidadevem ser dominates em relação aos de C.canephora (Fazuol i et aI., 1977)O café da variedade lcatu apresenta grandevariabilidade entre e dentro progênies quanto aprodutividade, vigor vegetativo, resistência aferrugem, precocidade e uniformidade dematuração dos frutos e constituintes químicos dosgrãos como a cafeína, ácido clorogênico e sólidossolúveis entre outros. Esta variabilidade édecorrente de sua origem híbrida entre C. arabica
e C. canephora. Entretanto, pouco se sabe sobresua variabilidade quanto a qualidade da bebida.Para dificultar esta separação entre progênies, osatuais procedimentos de avaliação sensorial daqualidade do café estão baseados em parâmetrosempíricos e subjetivos, pois dependem desensações e habilidades pessoais, adquiridas commuitos anos de experiência.Visando ampliar os procedimentos para avaliaçãoda qualidade do café, Prete (1992) desenvolveu oteste de condutividade elétrica do exsudato degrãos de café, baseado em métodos físicos equímicos.Comparando diferentes genótipos de café, Prete( 1992) veri ficou que o cv. Icatu apresentouvalores inferiores de qualidade em relação aosdemais genótipos pelo teste de condutividadeelétrica e pela "prova de xícara".Devido a sua origem, o Icatu apresenta grandevariabilidade, recombinando as diversascaracterísticas de C. canephora e C. arabica(Cambrony, 1988), tais com produtividade(Fazuoli et al., 1981, 1983); vigor vegetativo(Ramos e Lima, 1980; Costa et aI.. 1981),resistência à ferrugem (Costa, 1978; Eskes eCosta, 1980) e na composição química (Carvalho,1982).Estas características podem estar associadas coma qualidade da bebida de forma direta e/ouindireta e muitos aspectos ainda estão para serelucidados em benefício da cafeicultura.

480
Assim este trabalho visa verificar, quantificar eselecionar as progênies de café cv. Icatu queapresentem melhores qualidades da bebida.
Quadro 1. Condutividade elétrica média(IlS.cm-l.g-l) dos exsudatos das progênies docafé Icatu e cultivares Bourbon Vermelho eIAPAR-59.
LITERATURA CITADA
CONCLUSÃO
(1) Médias seguidas de mesma letra não diferementre si no nível de 5% de significância peloTeste de Tukey.DMS = 62,125 (IlS.cm-l.g-l)CV = 25,86%
Cambrony, H. R. 1988. Arabusta and otherinterspecific fertile hybrids. p.263-291. In R.T. Clarke, e R. Macrae, Coffee. v.4Agronomy.
Carvalho, A. 1982. Melhoramento do cafeeiro:Cruzamento entre C. arabica e C. canephora.p.363-368. In Dixieme Colloque ScientifiqueInternacional sur le café. Salvador, BA.
Os progênies de Icatu testadas possuemcaracterísticas semelhantes ao BourbonVermelho. sendo que a progênie lcatu 20apresentou menor valor de condutividade elétrica,merecendo maiores estudos para confirmar suaspossíveis características superiores.
Progênie CondutividadeICATU 20 64,37 bICATU io 70,27 abICATU 5 77,76 ablCA TU AMARELO 6 81,19 ab
:ICATU I 83,58 ab'ICATU 8 91,39 ab
ICATU li 92,01 abICATU 15 95,12abICATU 13 97,39 abICATU VERMELHO 24 97,77 abICATU 14 108,27 abICATU VERMELH019 109,54 ablCATU 23 112,29 abICATU AMARELO 30 112,58abICATU VERMELHO 7 115,42 abICATU TARDIO 115,98abAMARELO 4ICATU AMARELO 28 122,14 abICATU VERMELHO 17 122,16 abICATU VERMELHO 22 124,33 abICATU 2 128,75 aBOURBON VERM. 9 74,72 abIAPAR-5929 87,07 ab..
Os resultados da análise estatística dos dadospodem ser vistos no Quadro 1. A maior parte dasprogênies não diferiram entre si e das.testemunhas Bourbon Vermelho e IAPAR-59,sendo que a linhagem Icatu 20 apresentou menorvalor de condutividade elétrica sendo diferenteapenas do Icatu 2 que apresentou o maior valor decondutividade.
RESULTADOS
MATERIAL E MÉTODOS
Foram avaliadas as melhores progênies F3, F4, F5e F6 de retrocruzamento 2 dos ensaios de lcatuconduzidas pelo IAPAR, já em produção.Foram escolhidas as seis melhores plantas decada uma das 23 melhores progênies pelaprodutividade e outras característicasagronômicas.Como testemunhas foram utilizadas as cultivaresIAPAR-59 e Boubon Vermelho.Os frutos cereja foram coletados numa única datapara todas as plantas de todas as progênies etestemunhas na primeira quinzena de junho de1993. A quantidade de frutos coletado foi de umlitro que foi descascado e secado a sol. sem adegomagem e depois testados pelo método dacondutivida elétrica.A determinação de condutividade elétrica doexsudato foi realizada conforme metodologiadesenvolvida por Prete (1992) que consiste nautilização de quatro amostras de 50 grãos de café,sem escolha, pesados com precisão de 0,1 g,imersas em 75 ml de água destilada (no interiorde copos de plásticos de 180 ml de capacidade) ecolocadas em ambiente a 25°C por 3,5 horas,seguida de agitação e leitura em condutivímetroelétrico, expressando os resultados em /lS.cm1.g: 1 de amostra.A avaliação dos progênies quanto a condutividadeelétrica do exsudato foi feita através de análise devariância complementada pelo teste decomparação de médias de Tukey paradelineamento inteiramente casualizado com 20progênies Icatu e 2 cultivares testemunhas,Bourbon Vermelho e IAPAR-59.

Costa, W. M. 1978. Relação entre grau deresistência a H. vastatrix e a produtividade docafé Icatu. Bragantia 37: 1-9.
Costa, W. M., Carvalho, A., e Fazuoli, L.c. 1981.Estudo de caracteres dentro de progênies docafeeiro lcatu visando a sua seleção. p.169-73In Congresso Brasileiro de PesquisasCafeeiras, 9. São Lourenço, RJ.
Eskes, A. 8., e Costa, W. M. 1980.Characterization of incomplete resistance toHemileia vastatrix in the Icatu coffepopulation. Euphytica 32:649-57.
Fazuoli, L. C., Carvalho, A., Monaco, L. c., eTeixeira, A. A. 1977. Qualidade da bebida docafé Jcatu. Bragantia 36: 165-72.
Fazuoli, L. C., Carvalho, A., Costa, W. M., Nery,c., Laun, C. R. P., e Santiago, A. 1983.Avaliação de progênies e seleção no cafeeiroIcatu. Bragantia 42: 179-89.
481
Fazuoli, L. C., Gallo, P. 8., Carvalho, A., Costa,W. M., e Rocha, T. R. 1981. Seleção do cafélcatu em Mococa. p.174-179. In CongressoBrasileiro de Pesquisas Cafeeiras, 9. SãoLourenço, RJ.
Monaco, L. c., Carvalho, A., e Fazuoli, L. C.1974. Melhoramento do cafeeiroGermoplasma do café Icatu e seu potencial nomelhoramento. p.103. In Congresso dePesquisas Cafeeiras, 2. Poços de Caldas, MG.
Prete, C. E. C. 1992. Condutividade elétrica doexsudato de grãos de café (Coffea arabica L.) esua relação com qualidade da bebida.Piracicaba, SP, 125p. (Tese de Doutorado Escola Superior de Agricultura "Luiz deQueiroz".
Ramos, L. C. S e Lima, M. M. A. 1980.Avaliação da superfície relativa do sistemaradicular de cafeeiros. Bragantia 39: 1-5.


483
MELHORIA DE QUALIDADE EM ÓLEO DE CAFÉ PELO USO DA EXTRAÇÃO ACELERADACOM SOLVENTE(ASE)
Jair S.S. Pinto I , Fernando M. Lanças', Suzana L. Nixdorf", Paulo L. C. Oliveira'
'Laboratório de Cromatografia, Instituto de Química de São Carlos (IQSC), Universidade de SãoPaulo(USP), Cx. Postal 780. 13560-970 São Carlos-SP. Brasil; 2Companhia Cacique de Café Solúvel SA.Cx. Postal 824.86001-970 Londrina-Pk, Brasil.
Palavras-chaves: Óleo de café, extração acelerada por solvente (ASE}, matéria graxa, dererminação detem" de óleo, acides:
RESUMOComparativos na determinação de teores de óleo do café utilizando-se o sistema ASE e o extrator soxhlet(metodologia da AOAC) demonstraram resultados equivalentes, com um menor consumo de solvente (20mL)e uma redução no tempo análise (25 minutos) para o ASE, permitindo desta forma a melhoria nos processosindustriais em função dos resultados analíticos mais rápidos fornecidos pelo laboratório.
INTRODUÇÃO
o óleo de café. obtido através da prensagemmecânica dos grãos de café torrado. é um produtonatural com alto valor comercial agregado. Suaprincipal aplicação na indústria cosmética ealimentícia se deve a sua fragrância sendoutilizado para intensificar o odor de café em seusprodutos. Como um produto de alta qualidade seuconsumo e sua exportação dependem documprimento de certas especi ficações. Adeterminação do índice de acidez consiste em umadestas exigências', A acidez. é a medidaquantitativa do conteúdo dos ácidos graxos livrespresentes no óleo. um importante indicador doestado de deterioração do produto revelando seuestado de conservação. A decomposição dosglicerídeos é acelerada por aquecimento e pela luzsendo que a rancidez é quase sempreacompanhada pela formação destes ácidos graxoslivres. O teor de acidez do óleo de cafécomercializado depende desde da escol ha dosgrãos de· café utilizados em sua produção. os quaispodem variar de acordo com a variedade de caféselecionada, com a região e o solo onde o mesmofoi cultivado. com as condições climáticas a quefoi submetido além do modo com que foimanuseado em sua colheita entre outros fatores.sendo que este índice pode sofrer elevaçãodurante o processamento ou ainda na estocagemdo óleo após sua extração. Assim. a matéria-primadeve ser primeiramente analisada. a fim de severificar se a mesma é adequada para a obtençãode um produto final com baixa acidez. Paraefetuar a análise do índice de acidez a extraçãoprévia da matéria graxa ou teor de óleo érequerido a partir dos grãos de café cru. Portanto.este teor de óleo não é só necessário para ocumprimento de uma especificação descrita tendoque ser feito como pré-requisito para a anál ise de
acidez. mas também é fundamental por fornecerparâmetros que permitam prever e monitorar orendimento de extração do óleo de café noprocesso industrial. Como o setor produtivodepende dos resultados fornecidos pelolaboratório para a escolha da matéria prima a fimde iniciar o processo de extração. quanto maisrápido ocorrer o procedimento analítico com orepasse destas informações. tanto melhor. Aextração acelerada com solvente CASE) vem deencontro a esta necessidade.', A metodologiaoficial atualmente adotada para a determinação dematéria graxa do café cru descrita pela AOACutiliza soxhlet por 16 horas com 150mL de éter depetróleo como solvente. O ASE é um novoprocedimento de extração que utiliza solventesorgânicos a alta pressão (1500-2000psi) etemperatura acima do ponto de ebulição (50200°C)~ para extrair rapidamente componentes deamostras sólidas ou semi-sólidas com menorconsumo de solventes quando comparado àstécnicas convencionais. Isto acontece pela reduçãona interação soluto-matriz com a quebra doequilíbrio superficial. ocorrendo umamaximização nas taxas de transferência de massae solubilidade com redução da viscosidade e datensão superficial do solvente. Este procedimentose insere no contexto atual que visa promover aminimização do emprego de solventes orgânicos.reduzindo a quantidade de resíduos descartadoscom uma menor exposição do analista a estessolventes. e ainda promovendo a redução dotempo de análise. Com esta finalidade um sistemade ASE foi projetado e construído. o qual primapela simplicidade e funcionalidade 'com baixocusto. agregando valor ao trabalho dedesenvolvimento de técnicas que facilitam opreparo de amostras. sendo o mesmo aplicado àextração da matéria graxa do café cru visandomelhorar a qual idade do processo industrial.

MATERIAIS E MÉTODOS
Sistema ASEO sistema ASE construído (Figura I) consiste deuma bomba pneumática, onde o solvente écolocado e pressurizado diretamente comnitrogênio (ou qualquer outro gás inerte ); ce la deextração de aço de diferentes tamanho s fechadacom filtros de aço inox que evitam a passagem depart ículas; válvulas que visam direci onar osolvente extrator ou o gás de purga antes e após acela de extração; um sistema de aquecimento : umcoletor e algumas conexões. O solvent epressurizado percola a amostra disposta na celade extração promovendo a extração do material deinteresse.
Figura I - Diagrama cxqucrnatico do sistema de evtracãacelerada com solvente (ASE) I Nitrog énro . 2. Vuspressuri/ad or: (1+2 = Bomba pneumatica): 3. Válvulas 4Sistema de aquecimento: 5 Cela de extração com filtros. 6Coletor. 7 Manômetro
Preparação das amostrasCinqüenta gramas de grãos de café Arábica cruforam secos em estufa a 100"C por 5 horas.resfriados. triturados e passados em peneira malha0.42 mm. As amostras foram cedidas pela CiaCacique de Café Solú vel SA.
Extração em soxhlet J
Oito gramas de amostra de café previamentepreparadas foram extraídas em soxhlet com 150mL de éter de petróleo em chapa (30-60 "C) por 8horas. O solvente foi evaporado até completasecagem do resíduo em estufa (105°C) poraprox imadamente 1 hora . O teor de óleo oumatéria graxa foi determinado gravimetricamenteapós resfriamento.
484
Extração ASEO vaso pressurizador foi preench ido com éter depetróleo e pressurizado a 1500 psi com nitrogênio.Oito gramas de amostra de café previamentepreparadas foram coloc adas na cela de extração(0.8 x 30 em) a qual foi transferida para o sistemade aquecimento mantido a 80°C. Inicialmente acela foi preenchida com éter de petróleoaguardando-se por 5 minutos para que o equil íbriotérmico fosse atingido. Uma extração estát ica foifeita durante !O minutos sendo que mais 10minutos foram necessários para efetuar a extraçãodinâmica a um fluxo de aproximadamente2mLlmin. Após o término da extração o sistemafoi purgado com nitrogênio para promover aeliminação do solvente da cela com sua completatransferência para o frasco co letor. O solvente daamostra extraída foi evaporado em rota-vaporsendo totalmente eliminado pela passagem denitrogênio. O teor de óleo foi determinadogravimetricamente .
RESULTADOS E DISCUSSÃO
Os dados obtidos na Tabela 1 demonstram aviabilidade do emprego do sistema proposto deASE como altern ativa ao método tradicionalmenteutilizado de extração via soxhlet. pelo qual seobtém uma extração com uma eficiência de 95%em relação ao valor do método oficial. Setemperaturas mais altas forem empregadas valoresmais próximo s com um menor tempo de análisecertamente serão obtidos. Comparaçõe s tambémdemon stram lima redução no consumo de sol venteem cerca de 7 vezes, bem como uma redução notempo de análise em aproximadamente 20 vezes.cuja extraç ão agora leva cerca de 25 min para serrealizada. Esta redução no tempo analítico permiteque se efetue uma otimização nos parâmetrosanalíticos permitindo a validação dasmetodologias adotadas pelo laboratório, abrindocaminho para um monitoramento mais completoem todas as etapas do processo produtivopermitindo a investigação não somente damatéria-prima, mas também possibilitando umamelhoria na qualidade do óleo pelo conhecimentoe controle das variá veis de processo. Uma outravantagem é que o sistema pode ser automatizadopelo desenvolvimento de hardware e software quepermitam o controle dos solventes dispensadosatrav és do acionamento das válvulas.

485
REFERÊNCIAS BIBLIOGRÁFICAS
I. A.O.A.e. Association of Official AnalyticalChemists Methods of Analysis of ThirteenthEdition. 1980. Washington De. 15.033, p. 235.2. Heernken, O. P.; Theobald, N. andWenclawiak, B. W. 1997. Comparison of ASEand SFE, Sonication, and MethanolicSaponification Extractions for the Oeterminationof Organic Micropollutants in Marine ParticulateMatter. Anal. Chem. 69( 1 I): 2171-2180.3. Normas Analíticas do Instituto Adolfo Lutz.Métodos químicos e físicos para análise dealimentos. 1976. Vol. I. i ed., SP, p. 36 e 189.4. Richter, B.E.; Jones, B.A.; Ezzel, J.L. andPorte r, N.L. 1996. Accelerated SolventExtraction: A Technique for Sample Preparation.Anal. Chem. 68(6): 1033-1039. Correspondingauthor: Jair S.S. Pinto Laboratório deCromatografia Instituto de Química de São Carlos(IQSC)
Pesquisa do Estado de São Paulo (FAPESP), e aCAPES/PICO pelo suporte financeiro.
Os autores agradecem ao Conselho Nacional dePesquisas (CNPq). a Fundação de Amparo a
AGRADECIMENTOS
CONCLUSÕES
Tabela I - Extração de óleo de café
o sistema desenvolvido permite uma economia desolvente e de tempo analítico com resultadosequivalentes a metodologia oficialmente adotadapara o teor de óleo de café. As novas condiçõesanalíticas empregadas permitem a validação demétodos e o monitoramento do processo industrialde produção possibilitando ainda a automatizaçãodos dispositivos instrumentais utilizados.
Méto 501- Volu Tempo Teor Desviodo vente me de de óleo padrão
(ml.) extração ("lo) (0=2)(mio) (mIm)
Soxhlet Eter dePetróleo 150 480 16.23 -
ASE Eter dePetróleo 20 25 15.27 0.15

Ij
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j

487
INCIDÊNCIA DE FUNGOS EM CAFÉ
Marta Hiromi TANIWAKI 1, Andréia Aparecida BANHEI, Beatriz Thie IAMANAKA I
I Instituto de Tecnologia de Alimentos - Av. Brasil, 2880 - CEP 13.072-00 I, Campinas - SP, Brasil
1. INTRODUÇÃO
o café é um produto agrícola de extremaimportância econômica e social para o Brasil. Opaís vem ocupando a posição de maior produtormundial, não significando, que o excelentedesempenho em termos quantitativos sejaacompanhado no aspecto qualitativo. O café passapor vários processos até chegar a ser consumidocomo bebida. Vários fatores contribuem para aqualidade final do café, entre eles a populaçãomicrobiana presente e as condições de quefizeram uma ou outra espécie prevalecer. Existeuma associação de microrganismos presentescomo patógenos de plantas, contaminantes dasuperficie do fruto, no solo e no suco da polpa. Aecologia microbiana é complexa e envolve umasucessão de crescimento de bolores, leveduras ebactérias.Vários fungos ocorrem sobre os frutos do cafeeirono terreiro, .acelerando o processo de fermentaçãodos frutos, entre eles espécies de Fusarium,Cladosporium, Colletotrichum, Penicillium,Aspergillus, Trichoderma e Gliocladium (Carvalho et aI., 1997) tem sido encontrados.É um fato conhecido que muitas espécies defungos. principalmente dos gêneros Aspergillus,Penicillium, Fusarium são capazes de produzirmicotoxinas, metabólitos tóxicos com efeitosmutagênicos, teratogênicos e carcinogênicos. Aocratoxina A, com potencial carcinogênico enefrotóxico tem sido encontrada em algumasamostras de café provenientes do Brasil e essefato tem despertado muito interesse,especialmente, pelos países que importam café doBrasil. A ocratoxina é produzida por fungos dogênero Aspergillus e Penicillium, quenormalmente contaminam o café e outros grãos(Levi et al., 1974; Micco et aI., 1987). Dentre asespécies de Aspergillus destacam-se A. ochraceuse as espécies relacionadas A. alliaceus, A. meleuse A. suphureus. A espécie A. ochraceus foichamada de A. allutaceus por Samson & Gams(1985), contudo mais, recentemente. houve umacordo de que o nome mais apropriado para estaespécie é A. ochraceus (Pitt & Samson, 1993).Outras espécies de Aspergillus produtoras deocratoxina incluem: A. niger, A. carbonarius e A.terreus (Heenan et aI., 1998). Das espécies dePenicillium, P. verrucosum é o maior produtor deocratoxina em climas temperados. Originalmente,
o fungo produtor de ocratoxina foi identificadocomo P. viridicatum, contudo Pitt (1987)demonstrou que a espécie de Penicilliumprodutora de ocratoxina é P. verrucosum.
2. MATERIAL E MÉTODOS
2.1. Coleta das amostrasAmostras de café, variedade arábica, comaproximadamente 500 g, foram coletadas em umafazenda de Patrocínio - MG, nas fases de: árvore,colheita com pano, colheita de varreção, café bóia,no lavador, no terreiro com vários dias desecagem e na tulha. As amostras foram embaladasem saco plástico estéril e transportadas em isoporcom gelo até o Laboratório de Microbiologia doInstituto de Tecnologia de Alimentos (lT AL).
2.2 Plaqueamento das amostrasOs grãos de café descascado e a casca do cafépassaram por uma desinfecção superficial comhipoclorito de sódio 0,4%, por 2 mino Asamostras foram plaqueadas em seguida, no meioDicloran Rosa de Bengala Cloranfenicol (ORBe)e Dicloran 18% Glicerol (DG 18) de acordo com atécnica de Pitt & Hocking (1997). As placasforam incubadas a 25°C por 5 dias.
2.3 Identificação dos fungosPara a identificação dos fungos as chaves seguidasforam: gênero Penicillium (Pitt, 1991; Sarnson etaI., 1996); Aspergillus (KI ich & Pitt, 1988);Fusarium (Burgess et al., 1994); e outros fungos(Domsch et aI., 1993; Sarnson et ai., 1996; Pitt &Hocking, 1997).
2.4 Teste de produção de ocratoxinaO teste de produção de ocratoxina pelo fungo A.ochraceus isolado no presente trabalho, seguiu atécnica de Davis et al., 1966. O fungo foicultivado em meio YES e extraído comclorofórmio. O extrato foi analisado emcromatografia de camada delgada sob luzultravioleta, usando como solvente dedesenvolvimento tolueno:acetato de etila:ácidofórmico (90%) (5:4:1 v/v) conforme Scott et al.,1970).
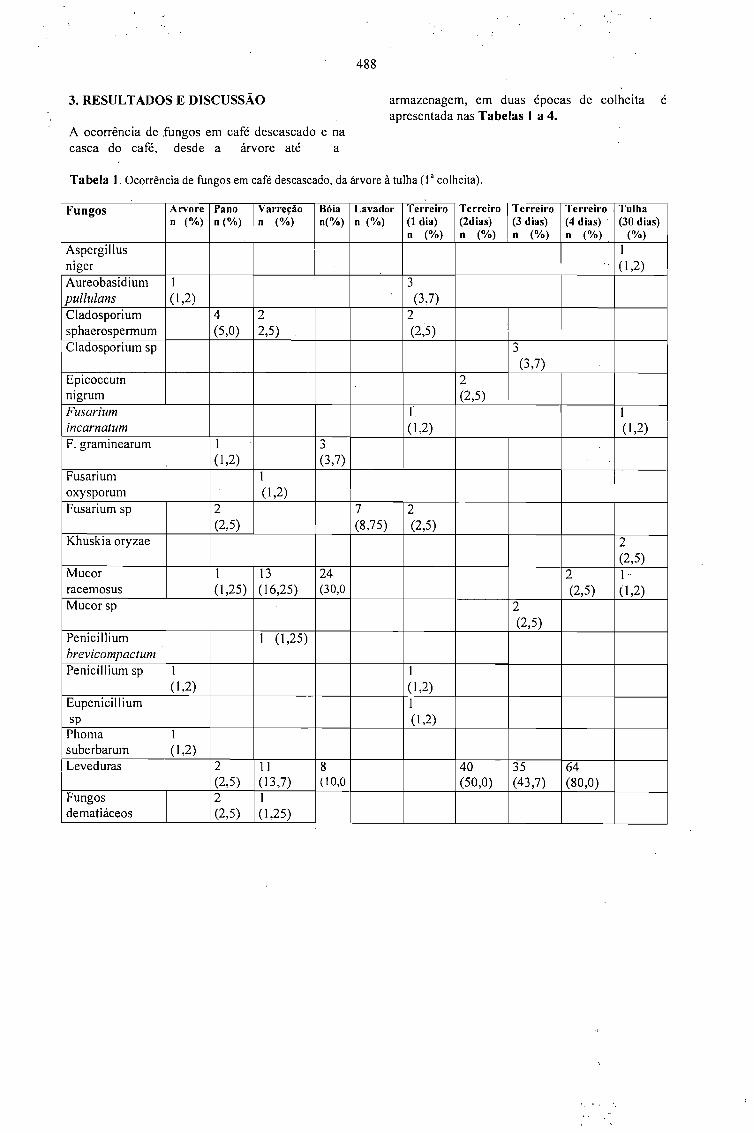
3. RESULTADOS E mscussxo
A ocorrência de fungos em café descascado e nacasca do café, desde a árvore até a
488
armazenagem, em duas épocas de colheita éapresentada nas Tabelas 1 a 4.
Tabela 1. Ocorrência de fungos em café descascado, da árvore à tulha (I a colheita).
Fungos Arvore Pano Varreção Bóia Lavador Terreiro Terreiro Terreiro Terreiro Tulhan (%) n(%) n (%) n(%) n (%) (1 dia) (2dias) (3 dias) (4 dias) . (30 dias)
n (%) n (%) n (%) n (%) (%)
Aspergill us 1niger (1,2)Aureobasidium I 3pullulans (1,2) (3,7)Cladosporium 4 2 2sphaerospermum (5,0) 2,5) (2,5) I
Cladospori um sp 3(3,7)
Epicoccum 2nigrum (2,5)Fusarium I Iincarnatum (1,2) (1,2)F. graminearum I 3
(1,2) (3,7)Fusarium 1oxysporum (1,2)Fusarium sp 2 7 2
(2,5) (8,75) (2,5)Khuskia oryzae 2
(2,5)Mucor 1 13 24 2 I·racemosus (1,25) (16,25) (30,0 (2,5) (1,2)Mucor sp 2
(2,5)Penicillium 1 (1,25)brevicompactumPenicillium sp 1 1
(1,2) (1,2)Eupenicillium Isp (1,2)Phoma 1suberbarum (1,2)Leveduras 2 11 8 40 35 64
(2,5) (13,7) (10,0 (50,0) (43,7) (80,0)Fungos 2 1dematiáceos (2,5) (1,25)
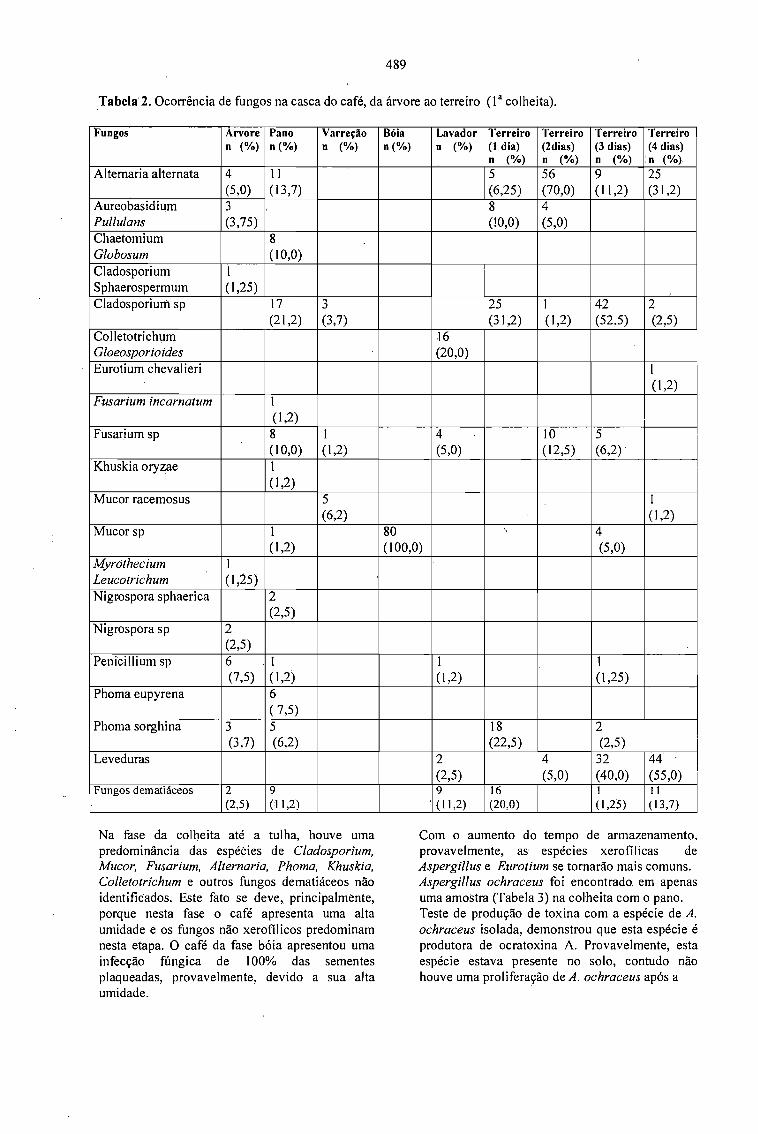
489
Tabela' 2. Ocorrência de fungos na casca do café, da árvore ao terreiro (l a colheita).
Fungos Arvore Pano Varreção Bóia Lavador Terreiro Terreiro Terreiro Terreiron (%) n (%) n (%) n (%) n (%) (1 dia) (2dias) (3 dias) (4 dias)
n (%) n (%) n (%) .n (%)Alternaria alternata 4 11 5 56 9 25
(5,0) (13,7) (6,25) (70,0) (11,2) (31,2)Aureobasidium 3 8 4Pullulans (3,75) (lO,O) (5,0)Chaetomium 8Globosum (10,0)CIadosporium ISphaerospermum (1,25)Cladosporium sp 17 3 25 I 42 2
(21,2) (3,7) (31,2) (1,2) (52,5) (2,5)Colletotrichum 16Gloeosporioides (20,0)Eurotium chevalieri I
(1,2)Fusarium incarnatum I i
(1,2) IFusariurn sp 8 I 4 10 5
(10,0) (1,2) (5,0) (12,5) (6,2) .
Khuskia oryzae I(1,2)
Mucor racemosus 5 I(6,2) (1,2)
Mucor sp I 80 -, 4(1,2) (100,0) (5,0)
Myrothecium ILeucotrichum (1,25)Nigrospora sphaerica 2
(2,5)Nigrospora sp 2
(2,5)Penicillium sp 6 ;J I 1
(7,5) (1,2) (1,2) (1,25)Phoma eupyrena 6
( 7,5)Phoma sorghina 3 5 18 2
(3,7) (6,2) (22,5) (2,5)Leveduras 2 4 32 44
(2,5) (5,0) (40.0) (55,0)I
Fungos dematiáceos 2 9 9 16 I 11(2,5) (11,2) . (11,2) (20,0) (1,25) (13,7)
Na fase da colheita até a tulha, houve umapredominância das espécies de Cladosporium,Mucor, Fusarium, Alternaria, Phoma, Khuskia,Colletotrichum e outros fungos dematiáceos nãoidentificados. Este fato se deve, principalmente,porque nesta fase o café apresenta uma altaumidade e os fungos não xerofilicos predominamnesta etapa. O café da fase bóia apresentou umainfecção fúngica de 100% das sementesplaqueadas, provavelmente, devido a sua altaumidade.
Com o aumento do tempo de armazenamento.provavelmente, as espécies xerofilicas deAspergillus e Eurotium se tornarão mais comuns.Aspergillus ochraceus foi encontrado em apenasuma amostra (Tabela 3) na colheita com o pano.Teste de produção de toxina com a espécie de A.ochraceus isolada, demonstrou que esta espécie éprodutora de ocratoxina A. Provavelmente, estaespécie estava presente no solo, contudo nãohouve uma prol iferação de A. ochraceus após a
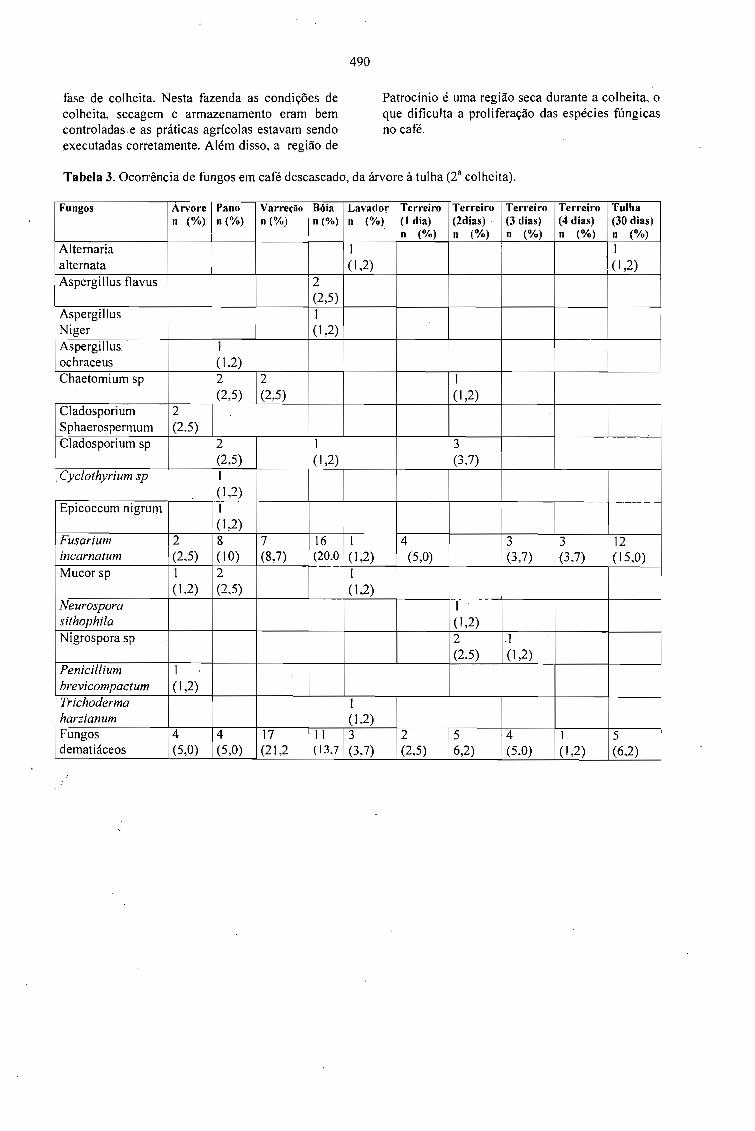
fase de colheita. Nesta fazenda as condições decolheita, secagem e armazenamento eram bemcontroladas.e as práticas agrícolas estavam sendoexecutadas corretamente. Além disso, a região de
490
Patrocínio é uma região seca durante a colheita, oque dificulta a proliferação das espécies fúngicasno café.
Tabela 3. Ocorrência de fungos em café descascado, da árvore à tulha (2a colheita).
Fungos Arvore Pano Varreção Bóia Lavador Terreiro Terreiro Terreiro Terreiro Tulhan (%) n (%) n (%) n (%) n (%) (1 dia) (2dias) (3 dias) (4 dias) (30 dias)
n (%) n (%) n (%) n (%) n (%)Alternaria I Ialternata (1,2) (1,2)
I Aspergi lIus flavus 2(2,5)
Aspergillus INiger (1,2)Aspergillus J
ochraceus (1.2)IChaetomium sp 2 2 1
I(2,5) (2,5) (1,2) I
Cladosporium 2Sphaerospermum (2.5)Cladosporium sp 2 I 3
(2,5) (1,2) (3.7).Cyclothyrium sp I
(1,2)Epicoccum nigrum I
(1,2)Fusarium 2 8 7 16 I 4 3 3 12incarnatum 1(2,5) (10) (8.7) 1(20.0 (1,2) (5,0) (3,7) (3,7) (15,0)Mucor sp I 2 I
(1,2) (2.5) (1,2) I
Neurospora Isithophila (1,2)Nigrospora sp 2 .1
(2,5) (1,2)Penicillium 1brevicompactum (1,2)Trichoderma Iharzianum (1.2)Fungos 4 4 17 11 3 2 5 14 I 5dematiáceos (5,0) (5,0) (21,2 ( 13.7 (3,7) (2.5) 6,2) i (5.0) (1,2) (6,2)
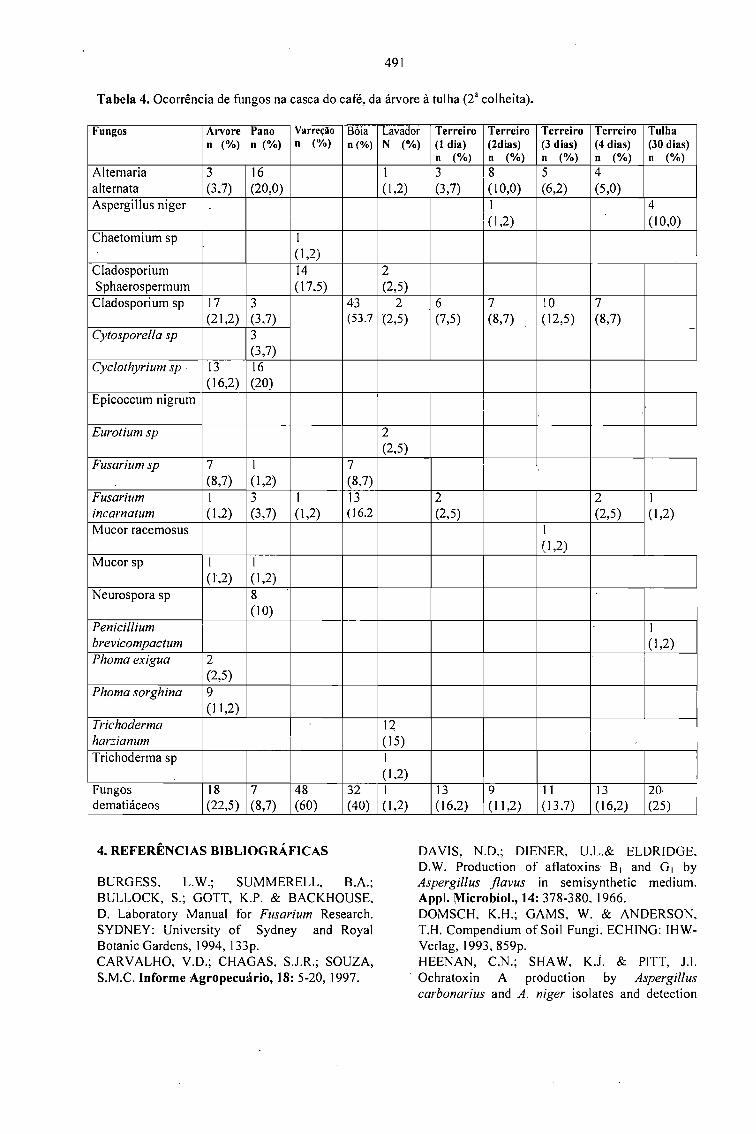
491
Tabela 4. Ocorrência de fungos na casca do café. da árvore à tulha (2a colheita).
Fungos Arvore Pano Varreção Bóia Lavador Terreiro Terreiro Terreiro Terreiro Tulhan (%) n (%) n (%) n(%) N (%) (I dia) (2dias) (3 dias) (4 dias) (30 dias)
n (%) n (%) n (%) n (%) n (%)
Alternaria 3 16 I 3 8 5 4alternata (3.7) (20,0) (1,2) (3,7) (10,0) (6,2) (5,0)Aspergillus niger - I 4
! (1,2) (10,0)Chaetomium sp I
(1,2)Cladosporium 14 2Sphaerospermum (17,5) (2,5)
Cladosporium sp 17 3 43 2 6 7 10 7(21,2) (3,7) (53.7 (2,5) (7,5) (8,7) (12,5) (8,7)
Cytosporella sp 3(3,7)
Cyclothyrium sp . 13 16(16,2) (20)
Epicoccum nigrum I
Eurotium sp 2(2,5)
Fusariumsp 7 I 7(8,7) (1,2) (8,7)
Fusarium I 3 1 13 2 2 1incarnatum (1,2) (3,7) (1,2) ( 16.2 (2,5) (2,5) (1,2)Mucor racemosus I
(1,2)Mucor sp I I
(1,2) (1,2)Neurospora sp 8
(1O)Penicillium 1brevicompactum (1,2)Phoma exigua 2
(2,5)Phoma sorghina 9
(11,2)Trichoderma 12harzianum (15)Trichoderma sp I
I(1.2) I
Fungos 18 7 48 32 I 13 9 11 13 20·dematiáceos (22,5) (8,7) (60) (40) (1,2) (16.2) (11,2) (13.7) (16,2) (25)
4. REFERÊNCIAS BIBLIOGRÁFICAS
BURGESS, L.W.; SUMMERELL, B.A.;BULLOCK, S.; GOTT, K.P. & BACKHOUSE.D. Laboratory Manual for Fusarium Research.SYDNEY: University of Sydney and RoyalBotanic Gardens, 1994, 133p.CARVALHO, V.D.; CHAGAS, S.J.R.; SOUZA,S.M.C.lnforme Agropecuário, 18: 5-20, 1997.
DAVlS, N.D.; DIENER, U.L.& ELDRIDGE.D.W. Production of aflatoxins B) and G 1 byAspergillus flavus in semisynthetic medi um.Appl, Microbiol., 14: 378-380. 1966.DOMSCf-L K.H.; GAMS, W. & ANDERSON.T.H. Compendium of Soil Fungi. ECHING: IHWVerlag, 1993, 859p.HEENAN, C.N.; SHAW, K.l & PITT, 1.1.
. Ochratoxin A production by Aspergilluscarbonarius and A. niger isolates and detection

al. The occurrence ofFd. Chem. Toxic., 33:
1995.
using coconut cream agar. J. Food Mycol., 1: 6772,1998.KLICH. MA & P1TT, Ll. A Laboratory Guide toCommon Aspergillus Species and TheirTeleomorphs. SYDNEY: CommonwealthScientific and Industrial Research Organization.1988, 115p.EVI, cr., TRENK, H.L. & MOHR. H.K. Studyof the occurrence of ochratoxin A in green coffeebeans. J. A.O.A.C., 57: 866-870, 1974.MICCO. c. GROSS1, M., MIRAGLIA. M.;BRERA, C. A study on the contamination byochratoxin A in green and roasted coffee beans.ASIC, 1ZO Colloque, p. 392-396, 1987.PITT, Ll, Penicillium viridicatum, Penicilliumverrucosum and production of ochratoxin A.Appl. Environm. Microbiol., 53: 266-269, 1987.PITT, 1.1. A Laboratory Guideto CommonPenicillium Species. SYDNEY: CommonwealthScientific and Industrial Research Organization,1991, 187p.
492
PITT, lI. & HOCKING, A.D. Fungi and FoodSpoilage. London: Blackie Academic &Professional, 1997, 593p.PITT, i.r. & SAMSON, R.A. Species names in
. current use in the Trichocomaceae (Fungi,Eurotiales). In: Greuter, W. ed. "Names in currentuse in the fami Iies Trichocomaceae,Cladoniaceae, Pinaceae andLemnaceae, regnumVeggetabile 128: 13-57, Konigstei n Germany:KoeltzScientific Books, 1993.SAMSON, R.A.; HOEKSTRA, E.S.; FRISVAD,r.c. & FILTENBORG, O. lntroduction to FoodBorne Fungi. BAARN: Centraalbureau voorSchirnmelcultures, 1996. 322p.SCOTT. P.M.; LAWRENCE, l W. & vanWALBEEK, W. Detection of mycotoxins by thinlayer chromatography: application to screening offungai extracts. Appl. Microbiol., 20: 839-842,1970.STUDER-ROHR, I. etochratoxin A in coffee:341-355.

493
MONITORAMENTO DA QUALIDADE DO CAFÉ TORRADO E MOÍDO NO ESTADO DE SÃOPAULO
Mori, E. E. M.; Anjos, V. D. A .; Bragagnolo, N.
Instituto de Tecnologia de Alimentos-ITAL Av. Brasil 2880 . C. Postal: 139CEP: 13073-001-Campinas-SP
INTRODUÇÃO
o mercado interno de café é o segundo maior domundo e tem apresentado um consumo crescenteos últimos cinco anos.Em 1997. o consumo doméstico de café alcançouo patamar de 11,5 milhões de sacas conformeSAES et al.(1997) distribuídos em torrado emoído(88%), solúvel(12%) e expresso(5%).Verifica-se entretanto que o café consumido nomercado interno não apresenta qualidadesatisfatória e que muitos fatores contribuem paraisso. principalmente a oferta de cafés de pioresqualidades para as indústrias de torrefação. Oconceito de qual idade do café está diretamenteassociado ao paladar do consumidor.Os fatores que influenciam as características dequalidade da bebida do café são: local de origem(latitude. altitude, microclima...). espéciebotânicarArábica, Robusta). estágio euniformidade de maturação dos grãos. métodos decolheita, . processamento. armazenamento.torração. moagem, embalagem do café torrado emoído e preparo da bebida. As variações detorração alteram a percepção do sabor, do"aftertaste", da acidez e do corpo. A manutençãodo ponto de torra é crítica para avaliaçãocomparativa.Por meio da anál ise sensorial o torrefador podedeterminar qual é a característica do café que maisagrada o consumidor.A análise sensorial permite avaliar através dassensações de olfação, gustação e sensação na bocaas características de aroma, gosto e corpo dabebida do caféA análise sensorial da bebida do café é umaferramenta que permite produzir "blends"homogêneos com características consistentes desabor. "Blend" é o grande segredo industrial atéporque os consumidores são fiéis ao sabor dedeterminada marca. Quando o sabor de um"blend"varia. a análise sensorial pode ser utilizada paraajustar as suas características de sabor.Avaliar a qualidade do café torrado e moídovisando a melhoria dos processos industriais.desde a seleção das matérias-pri mas à corretaescolha dos equipamentos e padrões deindustrialização é o objetivo do projeto que vem
sendo desenvolvido entre o Sindicato da Indústriade Café do Estado de São Paulo e o Instituto deTecnologia de Alimentos sediado em CampinasSP (ABIC, 1998).
MATERIAIS E MÉTODOS
Cerca de 100 amostras de café torrado e/ou moídode indústrias paulistas e algumas de outrosestados. foram analisadas quanto a umidade,granulometria, cor e anál ise sensorial da bebida..O teor de umidade foi determinado pelo métodode estufa (HELRICH, 1990) a granulometria emagitador de peneiras utilizando-se peneiras denúmero 16/20/30/40/50 e fundo. a cor emespectrofotômetro Color Eye 2020 e sistema dediscos AGTRON/SCAA para classificação datorração variando de torração muito escura(disco25). escura(disco 35). moderadamenteescura(disco 45), média(disco 55). médiaclara(disco 65). moderadamente clara(disco 75).clara(disco 85) e torração muito clara(disco 95)(LlNGLE,1992).A análise sensorial foi realizada com equipe dejulgadores treinados e sistema computadorizadode análise sensorial Compusense utilizandoanálise descritiva quantitativa para avaliação dascaracterísticas de fragrância. aroma. defeitos,acidez. amargor. sabor. sabor residual. corpo equalidade global. por meio de uma escala linearnão estruturada de 10 em. conforme apresentadana Figura l , (HOWELL, 1998).

ANÁLISE SENSORIALTORRADO E MOÍDO
DE CAFÉ
494
pode lembrar ao odor de chocolate. Para o café detorração escura pode lembrar ao odor de resina,remédio, especiarias, queimado e cinzas:
DEFEITOS: odor e sabor de terra. mofo. batatacrua, rançoso, borracha, tabaco. queimado,madeira, azedo (vinagre), fermentado, conferidospelos grãos defeituosos como ardido. preto, verdee impurezas como terra, areia, paus e cascas. Caféde excelente qual idade é aquele isento de defeitos'.
ACIDEZ: propriedade sensorial desubstâncias como ácidos clorogênico, cítrico,málico e tartárico que produzem gosto ácidopercebido nos lados .posteriores da língua. Écaracterístico de café arábica particularmente devariedades cultivadas em altitude alta. Quando énatural e desejável é chamado de ácido e quandonatural e indesejável é chamado de azedo:
AMARGOR: propriedade sensorial desubstâncias como cafeína,' trigonelina, ácidoscafeico e quínico e outros compostos fenólicosque produzem o gosto amargo percebido no fundoda Iíngua. Este gosto no café é consideradodesejável até certo ponto. É afetado pelo grau detorração e pelo método de preparo da bebida.Quanto mais escuro, mais amargo é o café.Quanto mais fino o pó e portanto, demorado oprocesso de extração mais acentuado o amargor.
9.Qualidade Global
______-----;- ------,-__-----JLmuito ruim excelente
Figura 1 - Ficha de Avaliação Sensorial
Foi apresentado aos provadores a definição dostermos presentes na ficha de avaliação do caféconforme descrito a seguir:
FRAGRÂNCIA: é a percepção olfativa dopó de café torrado e moído. A intensidade dafragrância revela o frescor da amostra.
AROMA: é a percepção olfativa da bebidado café inalando os vapores que exalam da bebidaainda quente e, por via retronasal durante suadegustação. Para o café de torração clara, o aromapode lembrar ao odor de nozes, amêndoas,castanhas frescas e cereal, malte, pão torrado. Parao café de torração média clara pode lembrar aoodor de caramelo, para o café de torração média
SABOR: é a combinação das sensações degosto doce, salgado, amargo e ácido com osaromas de chocolate, caramelo ou cereal torradoformando o sabor característico desejável.Quando apresenta sabores estranhos lembrando asabor de terra. herbáceo, especiaria, queimado,resina, rançoso, madeira etc é consideradoindesejável.
SABOR RESIDUAL: é a sensação de gostopercebida após a ingestão da bebida. O saborresidual é desejável quando deixa um sabor quelembra ao chocolate e indesejável quando lembraao cigarro, produto queimado, resina. químico(remédio), madeira ou outro sabor estranho.
CORPO: é a sensação tátil de oleosidade ede viscosidade na boca. Encorpado écaracterístico de bebida forte e concentrada.produzindo uma sensação agradável na boca.
ADSTRINGÊNCIA: é a sensação deixada naboca criando uma sensação de secura que éindesejável no café.
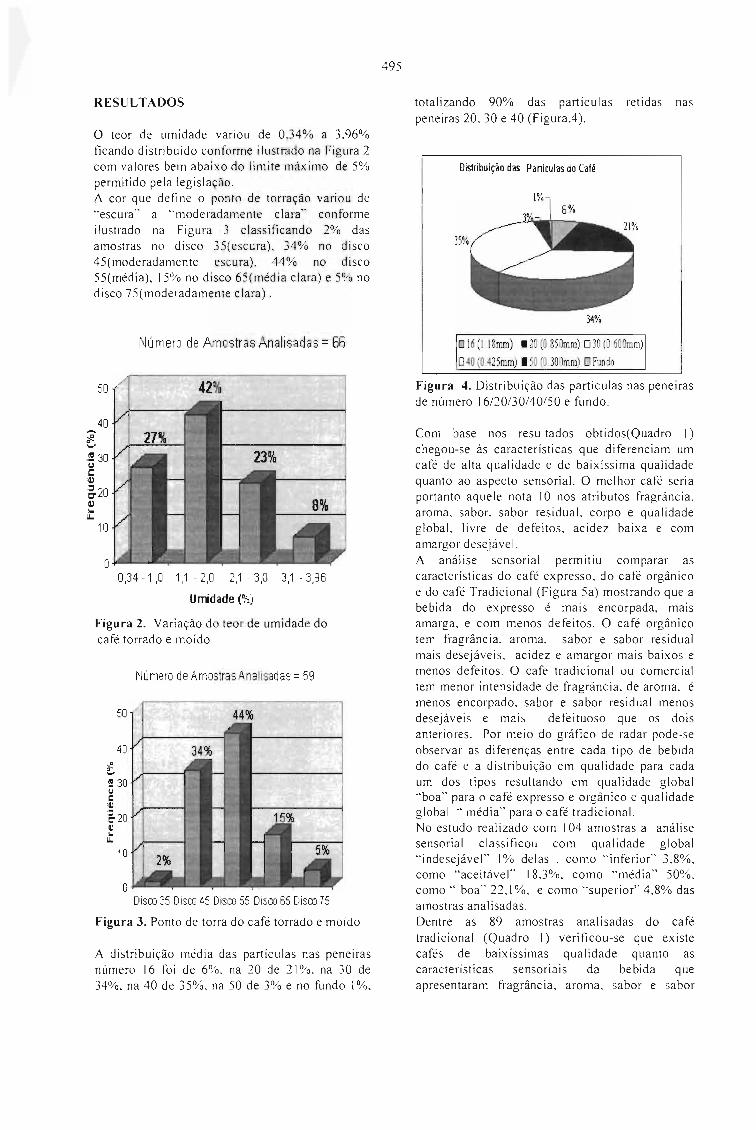
495
RESULTADOS
o teor de umidade variou de 0.34% a 3.96%ficando distribuído con forme ilustrado na Figura 2com valores bem abaixo do limite m áximo de 5%permitido pela legislação.A cor que define o ponto de torração vario u de"esc ura" a "moderadamente clara" conformeilustrado na Figura 3 classificando 2% dasamostras no disco 35(esc ura). 34% no disco45(moderadamente escura). 44% no disco55(média). 15% no disco 65(média clara) e 5 '~· O nodisco 75( modera da mente c lara) .
tota lizando 90% das partícu las ret idas naspene iras 20. 30 e 40 (FiguraA ).
Distribuição das Partículas do Café
j5%
34%
NúmerodeAmostrasAnalisadas=59
Número deAmostrasAnalisadas =66
Figura 4. Distr ibuição das partículas nas penei rasde número 16/20/30/40/50 e fundo.
Com base nos resultados ob tidos(Quadro I)chegou-se às caracteristicas que diferenciam umcafé de alta qualidade e de bai xíssirna qual idadequanto ao aspecto sensor ial. O melhor café seriaportanto aquele nota lO nas atributos fragrância .aroma. sabor. sabor residu al. co rpo e qualidadeglobal. livre de defeitos. ac idez baixa e comamargor desejável.A análise sensorial permiti u comparar ascaracterísticas do café expr esso. do café orgâ nicoe do café Tradicional (Figura 5a) mostrando que abebida do expre sso é ma is encorpada. maisamarga. e com menos de feitos. O café orgânicotem fragrância. aroma. sabor e sabor residualmais desejáveis. ac idez e amargor mais baixos emenos defeit os. O café tradiciona l ou comercialtem menor intensidade de fragrânc ia. de aroma. émenos encorpado. sa bor e sabor residual menosdesejáveis e mais defei tuoso que os doisanteriores. Por meio do gráfico de radar pode-seobservar as diferenças entre cada tipo de beb idado café e a distribuição em qua lidade para cadaum dos tipos resultando em qualidade globa l"boa" para o ca fé expre sso e orgâníco e qua lidadeg lobal .. média" para o café tradic ional .No estudo realizado com 104 amostras a aná lisesensoria l c lassificou com qual idade globa l"indesejável" I% delas. como "i nferior" 3.8%.como "aceitável" 183 %. como " média" 50%.como ., boa" 22.1%. e como "superior" 4.8% dasamostras analisadas.Dentre as 89 amostras ana lisadas do ca fétradicional (Quadro 1) verificou-se que ex istecafés de baixíssimas qualidade quanto ascaracterísticas sensoriais da bebida queapresentaram fragrância, aroma . sabor e sabor
44%50 '
A distr ibuição média das part ículas nas pene irasnúmero 16 foi de 6%. na 20 de 2 1%. na 30 de34%. na 40 de 35%. na 50 de 3% e no fundo 1%.
DisOJ 35 Discü 45 DisOJ 55 DisOJ 65 DisOJ 75
Figura 3. Ponto de torra do café torrado e moído
0,34 - 1 ,o 1,1 - 2,0 2,1 - 3,0 3,1 · 3,96
Umidade (%)
o
40
50
Figura 2. Variação do teor de umidade docafé torrado e moído
,~ 30(J
c'Q)
&20Q)lu..
10
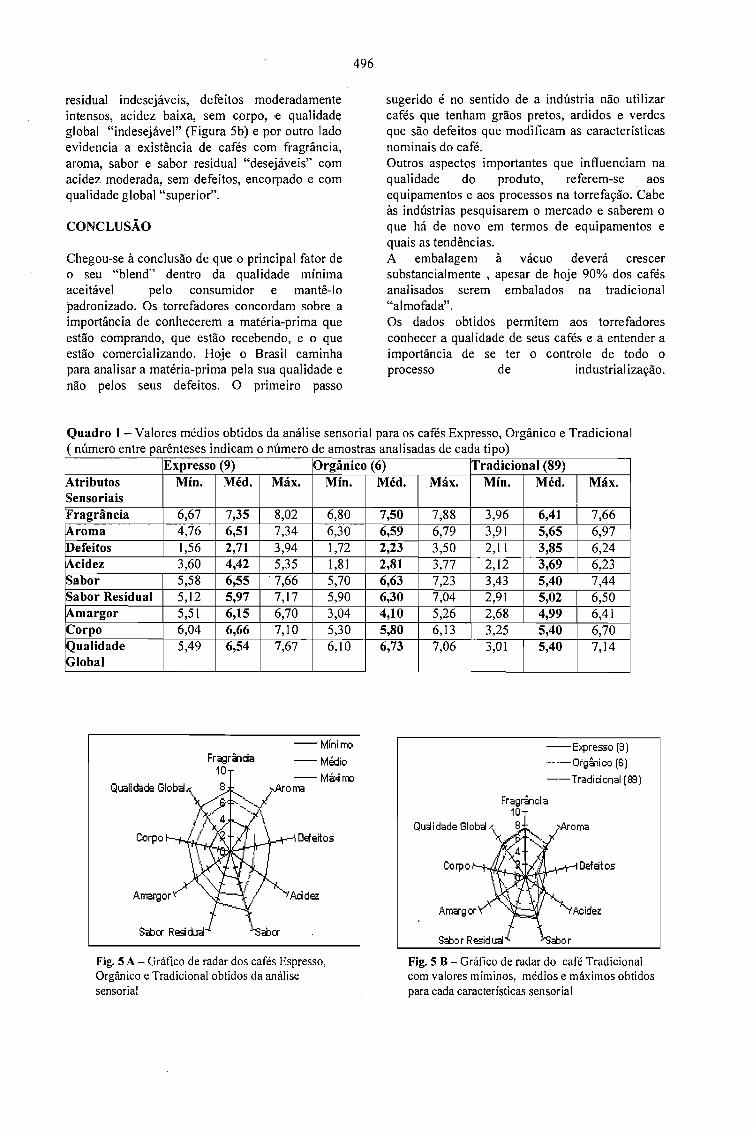
496
residual indesejáveis, defeitos moderadamenteintensos, acidez baixa, sem corpo, e qualidadeglobal "indesejável" (Figura 5b) e por outro ladoevidencia a existência de cafés com fragrância,aroma, sabor e sabor residual "desejáveis" comacidez moderada, sem defeitos, encorpado e comqualidade global "superior".
CONCLUSÃO
Chegou-se à conclusão de que o principal fator deo seu "blend" dentro da qualidade mínimaaceitável pelo consumidor e mantê-lopadronizado. Os torrefadores concordam sobre aimportância de conhecerem a matéria-prima queestão comprando, que estão recebendo, e o queestão comercializando. Hoje o Brasil caminhapara analisar a matéria-prima pela sua qualidade enão pelos seus defeitos. O primeiro passo
sugerido é no sentido de a indústria não utilizarcafés que tenham grãos pretos, ardidos e verdesque são defeitos que modificam as característicasnominais do café.Outros aspectos importantes que influenciam naqualidade do produto, referem-se aosequipamentos e aos processos na torrefação. Cabeàs indústrias pesquisarem o mercado e saberem oque há de novo em termos de equipamentos equais as tendências.A embalagem à vácuo deverá crescersubstancialmente , apesar de hoje 90% dos cafésanalisados serem embalados na tradicional"almofada".Os dados obtidos permitem aos torrefadoresconhecer a qual idade de seus cafés e a entender aimportância de se ter o controle de todo oprocesso de industrial ização.
Quadro 1 - Valores médios obtidos da análise sensorial para os cafés Expresso, Orgânico e Tradicional(número entre parênteses indicam o número de amostras analisadas de cada tipo)
Expresso (9) Orgânico (6) [I'radicional (89)!Atributos Mín. Méd. Máx. Mín. Méd. Máx. Mín. Méd. Máx.SensoriaisFragrância 6,67 7,35 8,02 6,80 7,50 7,88 3,96 6,41 7,66Aroma 4,76 6,51 7,34 6,30 6,59 6,79 3,91 5,65 6,97Defeitos 1,56 2,71 3,94 1,72 2,23 3,50 2,11 3,85 6,24Acidez 3,60 4,42 5,35 1,81 2,81 3,77 2,12 3,69 6,23Sabor 5,58 6,55 ·7,66 5,70 6,63 7,23 3,43 5,40 7,44Sabor Residual 5,12 5,97 7,17 5,90 6,30 7,04 2,91 5,02 6,50Amargor 5,51 6,15 6,70 3,04 4,10 5,26 2,68 4,99 6,41Corpo 6,04 6,66 7,10 5,30 5,80 6,13 3,25 5,40 6,70Qualidade 5,49 6,54 7,67 6,10 6,73 7,06 3,01 5,40 7,14Global
--E><presso (8)
--Orgâlico (6)
--Tradicional (B3)
Corpo Defeitos
~oma
Fragrâ-lci a10
Quaidade GIOba\ 8
/": 4Deteilos
--Míniroo
--Médio
-- Má>eiroo...Aroma
Fr<l;lrâ-lda10
8
Corpo
Fig.5 A - Gráfico de radardos cafés Espresso,Orgânico e Tradicional obtidosda análisesensorial
Fig.5 B- Gráficode radardo café Tradicionalcomvalores míminos, médiose máximosobtidosparacadacaracterísticas sensorial

REFERÊNCIAS BIBLIOGRÁFICAS
ABIC- Jornal do Café. Seminário de Qualidadedo Café, p. 12-15, Ano VIlI- Novembro/98- N.87.
HELRICH, K. (ed) Official Methods of Analysisofthe Association of Official Analytical ChemistsA.O.A.C. 15 (ed) Arlington, Virgínia, 1990.
HOWELL, G. SCAA Cupping form & How to
use it Presentation Handouts at J Oth Annualconference & Exhibition "PEAK ofPERFECTION", Denver, Colorado-USA 1721,1998.
497
LINGLE, T. R. The coffee cupper's handbook.Systematic Guide to the sensory evaluation ofcoffee's flavor. Second Edition. Specilaty CoffeeAssociation of America, 57 p.. J992.
SAES,M. S. M.; JAVO,M.; NAKAZONE,D.;SILVEIRA.R. L.F. Competitividade do sistemaAgroindustrial do Café. In: Competitividade noAgrobusiness Brasileiro, Volume IV, coordenaçãode Farina, E. M. M. Q.; Zylbersztajn,PENSA/FIA/FENUSP, São Paulo, J998.


499
AQUISIÇÃO DA TOLERÂNCIA À DESSECAÇÃO NOS DIFERENTES ESTÁDIOS DEMATURAÇÃO DE SEMENTES DE CAFEEIRO.
Delacyr da Silva Brandão Jr I; Maria das Graças Guimarães Carvalho Vieira 2; Lilian PadilhaRubens José Guimarães ~; José Roberto Bernardino Filho 5; Henk Hilhorst 6.
'Aluno de Doutorado do Curso de Fitotecnia - UFLA; 2 Professora Titular do Departamento de AgriculturaUFLA; ~ Aluna de Doutorado do Curso de Fitotecnia - UFLA; 4 Professor Adjunto do Departamento deAgricultura - UFLA; 5Aluno de Graduação do Curso de Agronomia - UFLA; "Professor de Fisiologia deSementes - Wageningen Agricultural University - Holanda.
INTRODUÇÃO
As sementes de cafeeiro, em condições normais.mantém-se viáveis por um período relativamentecurto. o que dificulta seu LISO a longo prazo. Outroaspecto a ser considerado é a conservação derecursos genéticos em bancos de gerrnoplasma,extremamente dificultada em função da baixalongevidade das sementes de cafeeiro. Esta baixalongevidade pode estar relacionada a capacidadedas sementes em resistir ou não a dessecação. Aliteratura tem apresentado controversa em relaçãoa este tópico. Vossen (1979) concluiu que assementes de café podiam manter sua viabilidadequando. armazenadas com 10% e 40% deumidade. King e Roberts (1979) classificaram assemente de cafeeiro como recalcitrantes. mas apóscinco anos foi sugerido que estas poderiam serortodoxas (Roberts et al. 1984). Segundo Ellis etaI. (1990). as sementes de Coffeea arabica podemser classificadas em uma categoria intermediáriapois a germinação não é prejudicada quando estassementes são secas até cerca de 10% de umidade.ocorrendo o inverso quando armazenadas àstemperaturas de 00 e -20oe. O estádio dedesenvolvimento do fruto do cafeeiro influencia acapacidade das sementes em tolerar o processo dedessecação, sendo que sementes de cultivares docafé arábica oriundas de frutos de maturidadeintermediária foram hábeis para tolerar uma maiordessecação (4-5% de umidade) quandocomparadas com as de frutos maduros ou verdes(Ellis et aI. 190 I) existe a necessidade de seconhecer mais profundamente os mecanismosenvolvidos na perda da viabilidade de sementes decafé. Este trabalho teve como objetivo caracterizaras diferenças de tolerância à dessecação dassementes de cafeeiro provenientes de diferentesestádios de maturação. Além disto. selecionarsistemas enzimáticos que permitam verificarvariações de perfis eletroforéticos nas sementescom diferentes níveis de tolerância a dessecação.
MATERIAIS E MÉTODOS
Sementes de cafeeiro da cultivar catuaí vermelhoforam colhidas em diferentes estádios dedesemvolvimento do fruto: verde (chumbão),verde cana e cereja. Os frutos foram despolpadosmanualmente e a degomagem foi realizada atravésda fermentação em água por 24 horas a 30° e. Asecagem foi efetuada em estufa de circulaçãoforçada regulada à temperatura constante de 30°C,até as sementes atingirem teores de água em tornode 50% ("sementes úmidas") ed 15%("sementessecas"). Estas sementes foram embaladas emsacaria de polietileno e armazenadas por trêsmeses em câmara fria a io-c. A qualidade dassementes foi avaliada no Laboratório de Análisede Sementes da Universidade Federal de LavrasMG. através da determinação do teor de água dassementes pelo método da estufa 1050C/24h
(Brasi I. 1992) e dos testes de primeira contagem econtagem final da germinação, aval iação dapresença de raízes secundárias e anál iseeletroforética de isoenzimas. O teste degerminação foi realizado de acordo com oscritérios estabelecidos pelas Regras para Análisede Sementes (Brasil, 1992). utilizando-se porém.4 subamostas de 50 sementes. para cada parcelaexperimental. O substrato utilizado foi o papeltoalha do tipo "Germitest" e os rolos foramcolocados em germinador mantido a 30oe. Aprimeira contagem foi realizada após 15 dias dainstalação. e a contagem final aos 30 dias. Nestaúltima. foi ainda computada a percentagem deplântulas que apresentavam as raízes secundáriasdesenvolvidas. A atividade de isoenzimas foiavaliada pela técnica de eletroforese segundoBrune et al. (1998), onde foram utilizadassementes inteiras para a extração das proteínas.Os géis foram revelados para as enzimas esterase(EST). álcool desidrogenase (ADH), catalase(CAT). glutamato oxaloacetato transaminase(GOT). malato desidrogenase (MDH) e superóxido dismutase (SOD). O delineamento utilizadofoi o inteiramente casualizado em esquemafatorial constituído por 3 níveis do fator estádio de
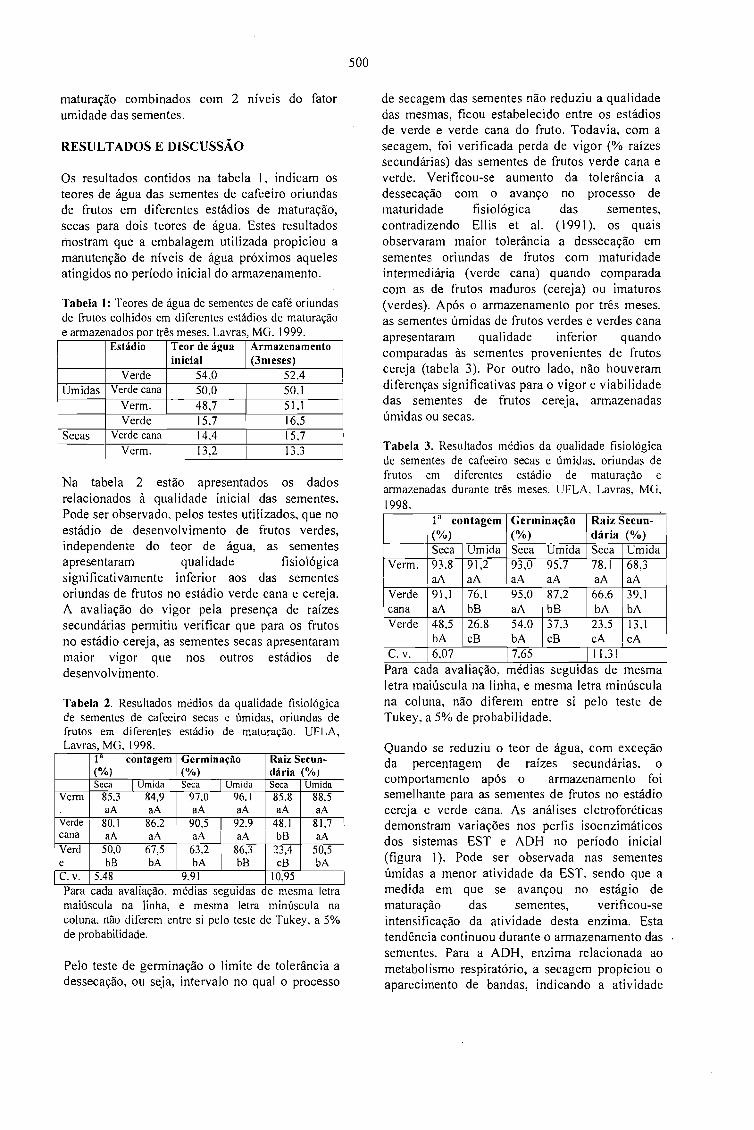
500
RESULTADOS E DISCUSSÃO
maturação combinados com 2 níveis do fatorumidade das sementes.
Pelo teste de germinação o limite de tolerância adessecação, ou seja, intervalo no qual o processo
Tabela I: Teores de água de sementes de café oriundasde frutos colhidos em diferentes estádios de maturaçãoe armazenados por três meses. Lavras, MG. 1999.
la contagem Germinação Raiz Secun-(%) (%) dária (%)Seca Umida Seca Umida Seca Umida
Vermo 93,8 91,2 93,0 95.7 78,1 68.3aA aA aA aA aA aA
Verde 91,1 76,1 95,0 87,2 66,6 39.1cana aA bB aA bB bA bAVerde 48,5 26,8 54,0 37,3 23,5 13.1
bA cB bA cB cA cAC. V. 6.07 17.65 11.31- , .
Tabela 3. Resultados médios da qualidade fisiológicade sementes de cafeeiro secas e úmidas. oriundas defrutos em diferentes estádio de maturação earmazenadas durante três meses. UFLA. Lavras, MG,1998.
Para cada avaliação, medias seguidas de mesmaletra maiúscula na linha, e mesma letra minúsculana coluna, não diferem entre si pelo teste deTukey, a 5% de probabilidade.
Quando se reduziu o teor de água, com exceçãoda percentagem de raízes secundárias, ocomportamento após o armazenamento foisemelhante para as sementes de frutos no estádiocereja e verde cana. As análises eletroforéticasdemonstram variações nos perfis isoenzimáticosdos sistemas EST e ADH no período inicial(figura 1). Pode ser observada nas sementesúmidas a menor atividade da EST, sendo que amedida em que se avançou no estágio dematuração das sementes, verificou-seintensificação da atividade desta enzima. Estatendência continuou durante o armazenamento dassementes. Para a ADH, enzima relacionada aometabolismo respiratório, a secagem propiciou oaparecimento de bandas, indicando a atividade
de secagem das sementes não reduziu a qualidadedas mesmas, ficou estabelecido entre os estádiosde verde e verde cana do fruto. Todavia, com asecagem, foi verificada perda de vigor (% raízessecundárias) das sementes de frutos verde cana everde. Verificou-se aumento da tolerância adessecação com o avanço no processo dematuridade fisiológica das sementes,contradizendo EIIis et aI. (1991), os quaisobservaram maior tolerância a dessecação emsementes oriundas de frutos com maturidadeintermediária (verde cana) quando comparadacom as de frutos maduros (cereja) ou imaturos(verdes). Após o armazenamento por três meses,as sementes úmidas de frutos verdes e verdes canaapresentaram qualidade inferior quandocomparadas às sementes provenientes de frutoscereja (tabela 3). Por outro lado, não houveramdiferenças significativas para o vigor e viabilidadedas sementes de frutos cereja, armazenadasúmidas ou secas.
Tabela 2. Resultados médios da qualidade fisiológicade sementes de cafeeiro secas e úmidas, oriundas defrutos em diferentes estádio de maturação. UFLA,Lavras, MG, 1998.
Na tabela 2 estão apresentados os dadosrelacionados à qualidade inicial das sementes.Pode ser observado, pelos testes utilizados, que noestádio de desenvolvimento de frutos verdes,independente do teor de água, as sementesapresentaram qual idade fisiológicasignificativamente inferior aos das sementesoriundas de frutos no estádio verde cana e cereja.A avaliação do vigor pela presença de raízessecundárias permitiu verificar que para os frutosno estádio cereja, as sementes secas apresentarammaior vigor que nos outros estádios dedesenvolvimento.
Para cada avaliação, medias seguidas de mesma letramaiúscula na linha, e mesma letra minúscula nacoluna. não diferem entre si pejo teste de Tukey, a 5%de probabilidade.
Os resultados contidos na tabela I, indicam osteores de água das sementes de cafeeiro oriundasde frutos em diferentes estádios de maturação,secas para dois teores de água. Estes resultadosmostram que a embalagem utilizada propiciou amanutenção de níveis de água próximos aquelesatingidos no período inicial do armazenamento.
Estádio Teor de água Armazenamentoinicial (3meses)
Verde 54,0 52.4Umidas Verde cana 50,0 50,1
Vermo 48,7 51.!Verde 15,7 16,5
Secas Verde cana 14.4 15,7Vermo 13,2 13.3
la contagem Germinação RaizSccun-(%) (%) dária (%)Seca Umida Seca Umida Seca Umida
Venn 85.3 84,9 97.0 96,1 85.8 88.5aA aA aA aA aA aA
Verde 80.1 86.2 90,S 92.9 48,1 81,7cana aA aA aA aA bB aAVerd 50,0 67,5 63.2 86,3 23,4 50,5e bB bA bA bB cB bAC. v. 5.48 9.91 10.95
- , .

desta enz ima nestas sementes. arma zenamentodas sementes implicou em altera ões o perfi I debandas da ADH. onde pôde er ob rvadoausência de banda present er íodo a terior aoarmazenamento. A s enzimas MDH. GOT e 'OD(dados não mostrados) das sementes armazenadas.não apresentaram ahera çõe: em seus perfis
quando submetidas aos diferentes tratamentos.Por outro lado. a CAT aprese ntou menor atividadepara as sementes armaze nadas úmidas. quandocomparadas às sementes secas. sendo que para assementes oriundas de frutos verde cana . nãohouve o aparec imento de banda .
A t\ L1 S .S ELETRO f O RÉT ICAS
# PERíODO INICIEST ADH
- -2 3 4 5 6
#- 3 MESES DI::: ARMAZENAMENTOEST ADH
Ar
Figura I. Padrões e letr for éticos de ernentes de cafeeiro oriundas de frutos de di ferentes estádio dematuração com alto e baixo grau de umidade. UFL . Lavras. MG . 1998.
I. Semente de cafee iro verde seca 4. Semente de cafee iro verde cana úmida2. Semente de cafeeiro verde úmida 5. Semente de cafeeiro cereja seca3. Semente de cafeeiro verde cana seca 6. Semente de cafeeiro cereja úmida
CONCLUSÕES
As sementes ori undas de fruto. no está íio cerejapossuem maior habilidade para tol rar adessecação quand o comparada com as de frutos
no estádio verde e verde cana. Os sistemasenzimáticos EST. ADH e CAT perm item verificarvariações de perfi s e letroforéticos nas sementescom diferentes níveis de to lerânc ia a dessecação.

REFERÊNCIAS
Brune, W.; Alfenas, A.C. and Junghans, T.G.1998. Identificações específicas de enzimas emgéis. pol0 1-328 In: Alfenas, A.C. (Ed.)Eletroforese de isoenzimas e proteínas afins:fundamentos e aplicações em plantas emicrorganismos. Ed. UFV, Viçosa.Brasil, Ministério da Agricultura. SecretariaNacional de defesa Agropecuária. 1992. 365p.Regras para Análise de Sementes. Brasília.Ellis, R. H.; Hong, T. D.; Roberts, E. H. 1990. Anintermediate category of seed storage behaviour?I. Coffee. Journal of Experimental Botany,
502
Oxfort. Societt for Experimental Biology,41: 1167-1174.Ellis, R. H.; Hong, T. D.; Roberts, E. H. 1991. Anintermediate category of seed storage behaviour?[I. Effects of provenance, immaturity, andimbibition on desiccation-tolerance in coffee.Journal of Experimental Botany, Oxfort. Societtfor Experimental Biology 42:653-657.King, M. W. and Roberts, E. H., 1979. Thestorage of recalcitrant seeds: achievements andpossible approaches. International Board for PlantGenetc Resources.Vossen, H.A.M.V. 1979. Methods of preservingthe viability of coffee seed in storage. SeedScíence and TechnoJogy. 7:65-74.

503
EFEITO DE ADUBAÇÕES FOLIARES EM PRÉ E PÓS FLORADA NA PRODUÇÃO EQUALIDADE DAS SEMENTES DE CAFEEIRO.
LIMA, Dinara Mattioli l. , VON PINHO, Edila Vilela Resende"; 3VIEIRA,
Guimarães Carvalho''; CUNHA, Rodrigo da"; GUIMARÃES, Rubens José5. ,
Vieira"
Maria das GraçasMONTEIRO, João
Ipesquisadora, bolsista DTI do CBP & D -UFLA; 2Professor Adjunto do Departamento de AgriculturaUFLA; - 3professor Titular do Departamento de Agricultura-UFLA; - "Pesquisador da EPAMIG; sprofessor Adjunto do Departamento de Agricultura-UFLA - 6Aluno de Graduação do Curso de AgronomiaUFLA - Lavras - MG.
INTRODUÇÃO
o uso de sementes de alta qualidade é um dosfatores primordiais para o sucesso da produção e,entre outros, a adubação é um dos fatores queafeta esta característica. A adubação é um dosfatores que afeta tamanho, peso e vigor dassementes, sendo que em muitas situações essesefeitos podem estar ligados à permeabilidade eintegridade das membranas dos tecidos dassementes, uma vez que diversos nutrientes atuamcomo ativadores enzimáticos ou comocomponentes dessas membranas (Sá e Buzzeti,1994).A absorção de nutrientes varia em funçãoda espécie, das condições ambientais, bem comodo estádio de desenvolvimento da planta. Nocafeeiro esta absorção se intensifica nos estádiospré e pós floração (Carvajal et al.,I969).Santinato et aI. (1991), avaliando os efeitosde P, Ca e B via foliar no pegamento de floradase frutificação do cafeeiro, observaram que aaplicação de Boro e Cálcio no período de pré epós florada promoveram maior retenção de frutoscom consequente aumento da produtividade daordem de 20 a 34%. A aplicação de Fósforoisoladamente não beneficiou a retenção de frutose quando aplicado juntamente ao Boro, ou aoCálcio inibiu a ação destes. Observa-se umacarência de pesquisas visando verificar asrelaçõea existentes entre nutrientes fornecidos aocafeeiro e qualidade das sementes. Sendo assim,o presente trabalho teve como objetivo avaliar ainfluência de nutrientes aplicados em pré e pósflorada na produção e qualidade das sementes decafeeiro.
MATERIAL E MÉTODOS
O presente trabalho foi instalado em uma lavourada cultivar Acaiá, no Departamento deAgricultura da Universidade Federal de Lavras. Odelineamento experimental utilizado foi emblocos casual izados com 15 tratamentos e4 repetições, totaJizando 60 parcelas. Cadaparcela constou de 3 fileiras de 6 plantas cada,
sendo avaliadas as 4 plantas centrais. Ostratamentos com 2 pulverizações, constaram de 1aplicação foliar antes da florada principal e 1aplicação pós florada; e os tratamentos com Ipulverização, receberam apenas uma aplicaçãofoliar após a florada principal.Os tratamentosforam:1-2g bórax/planta2-ác. bórico(0,25%)*3-ác. bórico(0,25%) + KCI(O,1%)*4-KCI(0, I%)*5-ác. bórico(0,25%) + Ca Cl 2 (0,05%)*6-ác. bórico(0,25%) + Ca Ch (0,05%) +KCI(O,l%)*7-Ca Ch (0,05%)*8-c.bórico(0,25%)+KCI(0, I%)+açúcar(O,1 %)
+úreia(O, I%)+sulfato de Zn(0,3%)9-trat,8+MAP(0, I%)10-trat,9+sulfato de CuCO, I%)+sulfato deMn(O,1%)+Mo(O,OI%)ll-trat.l 0+CaCI 2(0,05%)*
12-açúcar(0, I%)+úreia(O, 1%)+sulfatoZn(0,3%)+sulfato Mg(O,I%)+MAP(O, 1%)13-ác.bórico(0,25%)+açúcar(0, I%)+ ureia(O,I%)+sulfato de Zn (0,3%)+sulfato Mg(O,I%)+ MAP(O,I%)*14-trat.13+ Ca CI2 (0,05%)*15-Testemunha sem pulverização* Foram feitas 2 pulverizações.A qualidade fisiológica das sementes foi avaliadapelos testes de germinação(Germ), índice develocidade de germinação(IVG), T so, teste decondutividade elétrica e eletroforese de proteínatotal e isoenzimas. Os quais foram revelados paraos sistemas: Catalase(CAT) . e Esterase(EST).Para a determinação da produção, os dados foramtransformados em sacos de 60 Kg de cafébeneficiado/ha.
RESULTADOS E DISCUSSÃO
Nas condições do ensaio, foi verificado que aaplicação de Boro e Cálcio em pré e pós floradafoi o tratamento que apresentou melhor resultadoa produção em relação a testemunha.
De uma maneira gera l (I tra tament os q uereceberam as d uas adubações rol iarc: 'In pré>pós florada resu ltaram em 11la 101' ' . produç õesqu ando co mpa rad as co m lh tratame ntos q uerece beram ape nas uma pulverização foliar en pósflorad a (Ta bela l ). l ão foi constatadr e feito do :
Fig.
Fig :2
",: ',"I ,.,;~~:tiol:' ".•.~ .. r~J
t'!1 ~; ." " 4_
I"ig. J
Fig...\
50..\
tra tamento" na porce ntage m de ger m ina ção dasseme ntes . índ ice de ve loc idade de ge rm inaçã o.T50 c condu ti v idade e lét rica. Os marcadoresbioq uirnicos se le io nad os nào detecta ramdife re nça entre () ' trata me ntos (Figu ra I).
I ig. :'
' ·ig .7
Fig.X
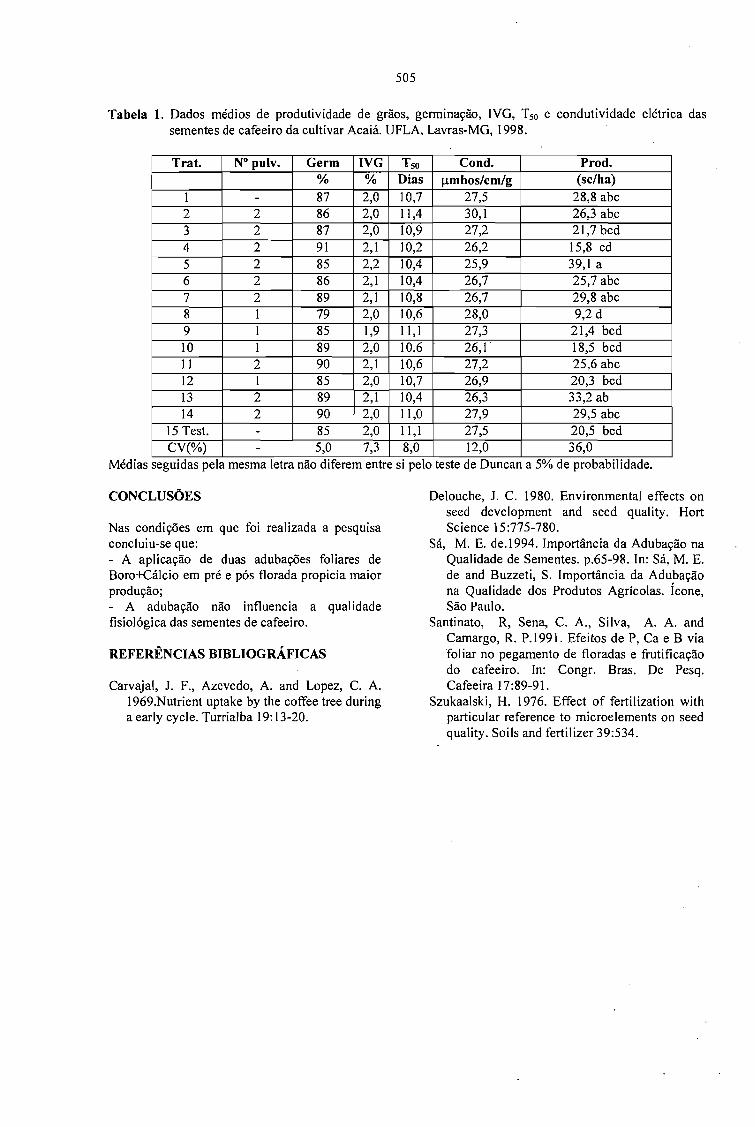
505
Tabela 1. Dados médios de produtividade de grãos, germinação, IVO, Tso e condutividade elétrica dassementes de cafeeiro da cultivar Acaiá. UFLA, Lavras-MO, 1998.
Trat. N° pulv, Germ IVG Tsü Cond. Prod.
I% % Dias umhos/cm/g (sc/ha)
1 .- 87 2,0 10,7 27,5 28,8 abc2 2 86 2,0 11,4 30,1 26,3 abc3 2 87 2,0 10,9 27,2 21,7 bcd4 2 91 2,1 10,2 26,2 15,8 cd5 2 85 2,2 10,4 25,9 39,1 a6 2 86 2,1 10,4 26,7 25,7 abc7 2 89 2,1 10,8 26,7 29,8 abc8 I 79 2,0 10,6 28,0 9,2 d9 I 85 1,9 I I, I 27,3 21,4 bcd10 I 89 2,0 10,6 26,1" 18,5 bcd11 2 90 2,1 10,6 27,2 25,6 abc12 I 85 2,0 10,7 26,9 20,3 bcd13 2 89 2,1 10,4 26,3 33,2 ab14 2 90 2,0 11,0 27,9 29,5 abc
15 Test. - 85 2,0 11,1 27,5 20,5 bcdCV(%) - 5,0 7,3 8,0 12,0 36,0 . .
Médias seguidas pela mesma letra não diferem entre SI pelo teste de Duncan a 5% de probabilidade.
CONCLUSÕES
Nas condições em que foi realizada a pesquisaconcluiu-se que:- A aplicação de duas adubações foliares deBoro+Cálcio em pré e pós florada propicia maiorprodução;- A adubação não influencia a qualidadefisiológica das sementes de cafeeiro.
REFERÊNCIAS BIBLIOGRÁFICAS
Carvajal, J. F., Azevedo, A. and Lopez, C. A.1969.Nutrient uptake by the coffee tree duringa early cycle. Turrialba 19:13-20.
Delouche, J. C. 1980. EnvironmentaJ effects onseed development and seed quality. HortScience 15:775-780.
Sá, M. E. de. I994. Importância da Adubação naQualidade de Sementes. p.65-98. In: Sá, M. E.de and Buzzeti, S. Importância da Adubaçãona Qualidade dos Produtos Agrícolas. Ícone,São Paulo.
Santinato, R, Sena, C. A., Silva, A. A. andCamargo, R. P.1991. Efeitos de P, Ca e B viafoliar no pegamento de floradas e frutificaçãodo cafeeiro. In: Congr. Bras. De Pesq.Cafeeira 17:89-91.
Szukaalski, H. 1976. Effect of fertilization withparticular reference to microelements on seedquality. Soils and fertilizer 39:534.

Ij
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j! j
j

507
COFFEE (Cof/ea 8P.) GENETIC TRAN8FORMATION FOR IN8ECT RE8I8TANCE
Magali Dufour 1, Thierry Leroy 2, Cathy Carasco-Lacombe 1, René Philippe 1and Catherine Fenouillet 1
1CIRAD-CP, B. P. 5035, 34032 Montpellier Cedex 1, France; 2Centre de Recherches Nestlé,B. P. 9716, 37097 Tours Cedex 2, France
Running title- Coffee genetic transformation
1. INTRODUCTION
Coffee, as a woody species, has a long biologicalcycle and it takes between four and five yearsfrom seed to seed. Therefore, classical breedingprogrammes generally spread over 20 to 30 years.Unconventiona1 breeding techniques would be ofutmost importance for quick genetic progressoOne ofthese techniques is genetic transformation.Using this process, it could be possible to insertselected traits in coffee without changing thewhole genome. Genetic transformation isbasically the introduction of foreign DNA intoplant cells. Two main techniques are used forplant transformation: i) direct transformation,through biolistics ( McCabe et ai., 1988), DNAuptake (Zhang and Wu, 1988), or protoplastelectroporation (Fromm et ai., 1985), and, ii)indirect transformation using viruses orAgrobacterium sp (Bevan et ai., 1983).Agrobacteria are natural soil-bom bacteria thatinfect the majority of dicotyledons and a fewmonocotyledons. Their pathogenecity is due tothe plasmid Ti-plasmid (tumour inducing) for A.tumefaciens, or Ri-plasmid (root inducing) for A.rhizogenes. Portions of the plasmid known as TDNA are transferred into the nuclear genome ofthe plant. The attachment of the bacteria on theplant cells as well as the transfer itself is madepossible by functions encoded by the vir region ofthe plasmid (Zupan and Zambryski, 1995). Thetransferred genes stably incorporate into the plantgenome. The genes responsible for tumourformation can be removed (the plasmid is thencalled disarmed) and replaced with virtually anygene, These genes can, therefore, be transferredinto plants. The efficiency of transformation isdependent on four factors:a) ! Ability of Agrobacterium to efficientlytransform cells in the target tissue (controlled bygenetic and physiological factors ofboth the plantand Agrobacterium). The' virulence of theAgrobacterium strain has to be tested first.b) Efficient selection of all the transformed cells.Selectable and scorable (i.e. reporter) genes are
needed.c) Efficient regeneration from the transformedcells. It is the bottleneck in the process, especiallyfor perennials. However in coffee, variousregeneration pathways can be used likeregeneration from protoplast, regeneration fromleaf explants through organogenesis or somaticembryogenesis (Yasuda et ai., 1985; Zamarripa etai., 1991; Berthouly and Michaux-Ferriêre, 1996).d) Stability of the incorporated DNA into thegenome of the plant. In annual plants, this cari bedetermined easily by selfing the plant in order todetermine if the gene is inheritable. Withperennials, Southem blot analysis must be used toidentify different segments of the inserted TDNA.Genetic transformation was described for the firsttime in coffee by Barton et ai. (1991) whoworked on protoplast electroporation of Coffeaarabica. However, no transgenic plant wasregenerated from these experiments. Later on,Feng et ai. (1992), using C. arabica cotyledonsand somatic embryos integrated a foreign geneconferring resistance to Kanamycin (anantibiotic). Again, no plant was obtained. It wasonly in 1993 when transgenic plants were createdby Spiral et ai. using a co-culture of C. canephorasomatic embryos with Agrobacterium rhizogenes.The regenerated plants were scored to haveseveral DNA inserts. Unfortunately, most of theseplants expressed a "hairy root" pattern, Sincethen, genetic transformation has been extended toother genotypes and other explant sources (Spiraland Pétiard, 1993; Sugiyama et ai., 1995). Leroyet ai., (1997) successfully used A. tumefaciens.Protection against attacks by detrimental pestsrepresents a major challenge for crop productionin agricuiture. This is especially true for coffee,for which a large percentage of the potentialharvest yield ofBrazil and East Africa is lost eachyear due to insects, in particular to the leaf miner(Perileucoptera sp.) (Guerreiro et aI., 1990).Since coffee is a perennial crop, it is not possibleto use crop rotation as seen with annuals. AIso,there are no breeding programmes using natural

sources of resistance for that .insect.Presently, control of pest development is based onapplication of protective agro-chemicals. For thefarmers, the products are expensive and theirimpact is often harmful for the environment..Bacillus thuringiensis (B. t.) is an entomocidalbacterium. For more than 35 years, crystalline 0endotoxins, which accumulate during sporulationof this bacterium, have been available as analtemative to synthetic insecticides in controllingharvest losses. The crystalline inclusionscontaining the endotoxins are activated in themidgut of the insects and bind to specific highaffinity receptors on cell membranes. Withinminutes, the midgut cells are paralysed anddisrupted. B. t. toxins are diverse and have beenfound active against many insects within theorder of Lepidoptera, Diptera and Coleoptera.The use of B. t. genes to transform plants is afeasible strategy to fight insects efficiently(Estruch et ai., 1997; Schuler et ai., 1998).Several B.t. endotoxins have been tested againstPerileucoptera coffeella Guérin Méneville andone, CryIA(c), was shown as very efficient(Guerreiro et ai., 1993, 1998). The genecorresponding to this toxin has been isolated,cloned, artificially synthesised, and included in avector construction containing also a selectableand a marker genes. Several transgenic coffeeplants actually resistant to the leaf miner havebeen obtained (Leroy et ai., 1999,2000). Here, wedescribe genetic transformation of four differentcoffee genotypes with the cry/A(c) gene and theverification of the gene expression throughWestem blot. The accuracy of the correlationwith insect bioassays is also discussed.
2. EXPERIMENTAL
2.1. Plant materialTwo C. canephora and two C. arabica genotypeswere used (clones number 126 and 197, Catimor(8661-4), and Et29 x Ca5 FI hybrid,respectively). They were propagated in vitromicro-cutting on MS-based medium (Murashigeand Skoog, 1962) containing 4.44 JlM BAP.
2.2.Transformation protocolLBA4404 disarmed strain of A. tumefaciens wasused with the pBin19 plasmid (Bevan, 1984),engineered with three genes: uidA, coding for ~
glucuronidase and containing an additionalintron; the csr1-1 gene conferring resistance tochlorsulfuron used as a selection agent; and a
508
synthetic cry1A(c) gene (Sardana et ai., 1996),driven by the Ef lu- promoter (Curie et ai., 1991).Leaf explants were cultured on specific semisolid media and embryogenic cells and somaticembryos were obtained. Between 1,000 and10,000 somatic embryos, or 10 to 50 gramsembryogenic cells were used for transformation.These were soaked with bacteria (O.D'600nm:0.3-0.5) for two hours and co-cultivated for threedays in hormone-free MS medium. They werethen cultivated in presence of cefotaxime (400mg.Ll ) for 21 to 28 days, and then transferred toselective MS medium with cefotaxime (400 mg.l:1) and chlorsulfuron (80 Jlg.l-1). After three to sixmonths, regeneration occurred and the somaticembryos germinated on MS mediumsupplemented with Morei vitamins, 1JlM BAPand 0.03-M sucrose.
2.3. Rooting and hardeningOnce the somatic embryos germinated andexhibited one, or two pairs of leaves, their basewas sectioned with a scalpel and soaked for 12hours in a sterile MS/2 medium containing IBAand NAA (50 mg.l-l each). They were thentransferred to the greenhouse in peat pellets underhigh humidity conditions for at least three weeks.Relative humidity was then loweredprogressively. Rooting occurred within a month.
2.4. Histochemical GUS assay~- glucuronidase was assayed by ovemightincubation in the c1assical medium (Jefferson,1987). In order to reduce endogenous activity ofnon-integrated uidA gene, 20% v/v methanol wasadded. (Kosugi et ai., 1990).
2.S. Molecular analysisOnly the GUS positive plants were used for totalDNA extraction. The protocol was described byEdwards et ai. (1991) and modified by theaddition of 100-mM sodium bisulphite. Southemblotting analysis was done as described by Leroyet ai. (1999) and Spiral et ai. (1999).
2.6. BioassaysRegenerated plants of different ages were put inpresence of adult Tanzania leaf miners(Leucoptera caffeina) for 24 hours. Two weekslater, an overall score was affected to the plantand number ofpups was recorded.
2.7. Western blotsLeaves were ground 10 liquid nitrogen and

proteins were extracted from about one gram offresh leaves in a standard buffer containing Tris HCI pH8, 87% glycerol (57.5 ml.l- l), EDTA,DTT (155 mg.lJ), PMSF (175 mg.IrJ) and Triton1ml.ll. The extracts were then concentratedusing Millipore Ultrafree-4® (biomax-50membranes) filtration units (30-50 minocentrifugation at 4500 rpm) and immediatelychecked for protein content (Bradford, 1976). Theextracts were then aliquoted and denatured(Laemmli, 1970). A 10% SDS-Page gels wereused and migration of the proteins was carried outfor four hours at ca. 100 V. Gels were thentransferred to nitro-cellulose membranes usingthe Bio-Rad transblot system in Towbin buffer(Towbin et aI., 1979) for one hour at 100 V.Blocking occurred ovemight in 0.5% non fat milkin TBS buffer supplemented with 0.1% (v/v)Tween-20. A polyclonal rabbit antiserum raisedagainst the purified Cry1A(c) protein was used(1/2500 v/v) and detected by a goat anti-rabbitantiserum alkaline phosphatase (AP)-conjugate(Promega) at a dilution of 113300 (v/v).
3. RESULTS AND DISCUSSION
For the C. canephora genotypes, transformationefficiency was about 1%, which wasapproximately 1% of the explants submitted tothe pracedure produced embryogenic calli onselective medium. This result slightly increased infurther experiments (Leroy et aI., 1999). Notevery embryogenic callus gave rise to somaticembryo. About 30% of calli produced embryos,of which ~O% germinated properly. More than200 independent primary transformants wereobtained using somatic embryos, or embryogeniccalli for clone J26, and only 30 for clone 197.Three copies each of 120 primary tansformants ofclone 126 were transferred to the greenhouse. Thetwo arabica genotypes behaved differently. Onselective medium, O to 5% of the primaryCatimor explants produced directly somaticembryos, depending on the experimento Less than0.1% embryogenic cal1i appeared on the FIhybrid, 10% of which produced somatic embryoswith a germination rate similar to that of C.canephora. Obviously, there was a wide genotypeeffect for response to the genetic transformationprocedure. C. canephora is a diploid species incontrast with C. arabica, which is tetraploid.Such behaviour differences have already beennoted for the aptitude to liquid medium somaticembryogenesis (Zamarripa et aI., 1991).Chlorsulfuran is a herbicide and seems to be an
509
adequate agent for selection of coffee transformedtissue. However, a certain rate of escape, up to50% has been observed. Consequently, GUSassay was also usedas a screen (Leroy et aI.,1999). Former experiments had shown that theantibiotic Kanamycin was not as efficient (Spiralet aI., 1993). Two types of analysis were carriedout on the transformed plants growing in thegreenhouse for at least two months. PCR andSouthem blotting analysis established the :presence of foreign genes. The qualityofintegration of the T-DNA and the number ofcopies integrated was documented by Southemanalysis. Among 51 transformation events fromC. canephora, the majority (69%) contained onecopy of the foreign T-DNA. Up to five copieshave been observed in other events ( Leroy et aI.,1999). The presence of an expressed crylA(c)gene was established by Westem blottingperformed on 21 independent transformationevents (clone 126) (Fig. 1). Among these plants,the majority appeared rather resistant whensubmitted to the bioassay (Table 1) with threeplants being very resistant (note :o:; 1 and O pupa)and eight intermediate-resistant (1<notesz andnumber of pupae :o:; 2). All these plants hadCryIA(c) protein activity detected by Westemblotting., which confirmed the bioassay results.Five plants were susceptible; two of them werecontrais, two others did not have any CryIA(c)protein detected and one was Westem positive.That plant was probably in a bad physiologicalstate, which made it susceptible to the leaf miner,although the CryIA(c) protein was expressed.AIso, the levei of expression was perhaps low.Besides these two categories, five plants were"intermediate-susceptible", their note beingbetween 1.2-1.8 with a number of pupae from 39. Three were Westem positive, but two did nothave any CryIA(c) protein detected. The cryIA(c)gene was detected by PCR and Southem blot.These occurrences have already been discussed(Leroy et aI., 1999) and have been confirmed byrecent Westem blots. It could be likely that thetoxin was expressed in a small amount (below0.1% of total pratein), rendering the plantssomehow susceptible.
4. CONCLUSIONS
The transformation technique described here hasbeen efficient on four different coffee genotypesalthough some differences have been observed inresponse according to genotype. A large numberof plants was transferred to the greenhouse and
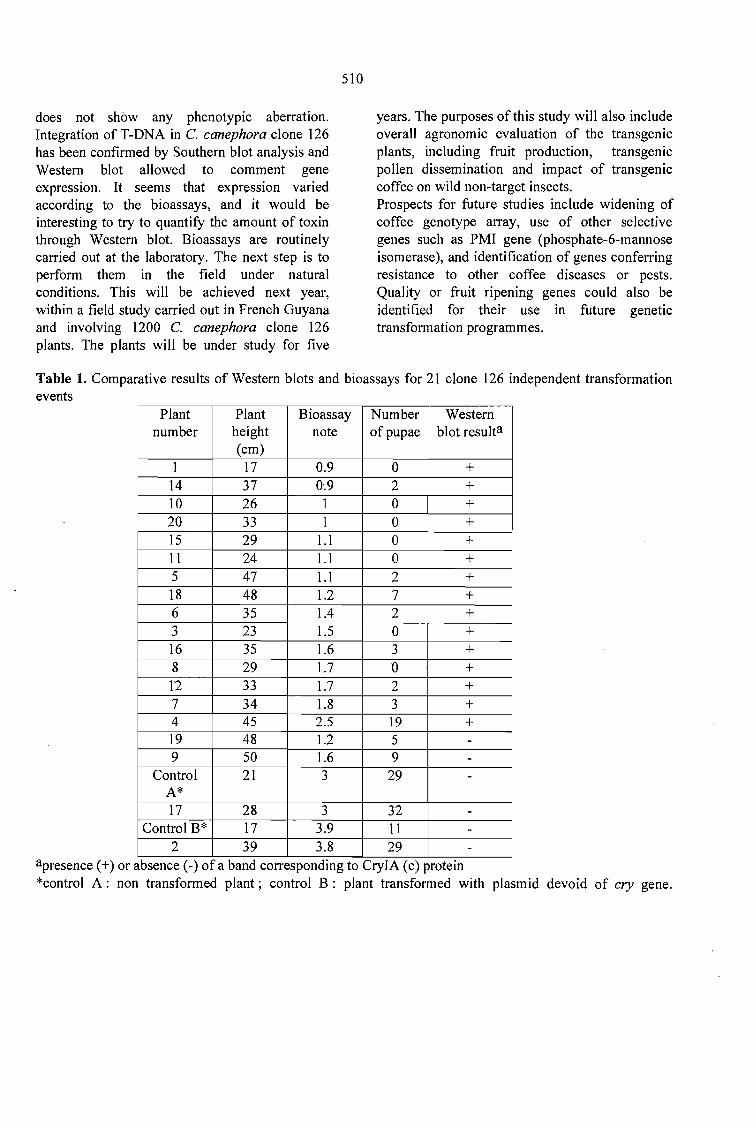
does not show any phenotypic aberration.Integration of T-DNA in C. canephora clone 126has been confirrned by Southern blot analysis andWestern blot allowed to comment geneexpression. It seems that expression variedaccording to the bioassays, and it would beinteresting to try to quantify the amount of toxinthrough Western blot. Bioassays are routinelycarried out at the laboratory. The next step is toperforrn them in the field under naturalconditions. This will be achieved next year,within a field study carried out in French Guyanaand involving 1200 C. canephora clone 126plants. The plants will be under study for five
510
years. The purposes of this study will also includeoverall agronomic evaluation of the transgenicplants, including fruit production, transgenicpollen dissemination and impact of transgeniccoffee on wild non-target insects.Prospects for future studies include widening ofcoffee genotype array, use of other selectivegenes such as PMI gene (phosphate-6-mannoseisomerase), and identification of genes conferringresistance to other coffee diseases or pests.Quality or fruit ripening genes could also beidentified for their use in future genetictransformation programmes.
Table 1. Comparative results of Western blots and bioassays for 21 clone 126 independent transformationevents
Plant Plant Bioassay Number Westernnumber height note ofpupae blot resulta
(em)1 17 0.9 O +
14 37 0.9 2 +10 26 1 O +20 33 1 O +15 29 1.1 O +11 24 1.1 O +5 47 1.1 2 +18 48 1.2 7 +6 35 1.4 2 +3 23 1.5 O +16 35 1.6 3 +8 29 1.7 O +12 33 1.7 2 +7 34 1.8 3 +4 45 2.5 ]9 +19 48 1.2 5 -9 50 1.6 9 -
Control 21 3 29 -A*17 28 3 32 -
Control B* 17 3.9 11 -2 39 3.8 29 -
apresence (+) or absence (-) of a band corresponding to CryIA (c) protein*control A: non transformed plant; control B: plant transformed with plasmid devoid of cry gene.

511
Figure 1 : Westem blot membrane showing different plant profiles. C = control, non transgenic plant. Tx =purified CryIA(c) toxin
~...
:.";..;:,;;.::':,:,::,~:
.;.," :'-:.
8 129538Tx· '.~ cL2..:~--="::2.-----=- ._
5. REFERENCES
Barton, C.R., Adams, T.L. and Zarowitz, M. A.(1991), Stable transformation of foreignDNA into Coffea arabica plants. In Asic:14eme colloque scientifique sur le café,San Francisco, pp. 460-464 .
Berthouly, M. and Michaux-Ferriêre, N. (1996),High frequency somatic embryogenesis inCoffea canephora (var. Robusta): inductionconditions and histological evolution.Plant Cell, Tissue Organ Culture, 44: 169176
Bevan, M. (1984), Binary Agrobacterium vectorsfor plant transforrnation. Nucleic AcidRes., 12: 8711-8721
Bevan, M., Flavell, R. and Chilton, M.-D. (1983),A chimaeric antibiotic resistance gene as aselectable marker for plant celltransforrnation. Nature, 304: 184-187
Bradford, M.D. (1976), A rapid and sensitivemethod for the quantitation of microgramquantities of protein utilizing the principieof protein dye binding. Anal. Biochem., 72:248-254
Curie, C., Liboz, T., Bardet, C., Gander, E.,Medale, C., Axelos, M. and Lescure, B.(1991), Cis and trans-acting elementsinvolved in the activation of Arabidopsisthaliana A1 gene encoding the translationelongation factors EF-l alpha. NucleicAcid Res., 19: 1305-1310
Edwards, K., Johnston, C. and Thompson, C.(1991), A simple method for PCR analysis.Nucleic Acid Res., 19: 1349
Estruch U., Carrozzi, N.B., Desai, N., Duck,N.B., Warren, G.W. and Koziel, M.G.
(1997), Transgenic plants : an emerging approachto pest control. Nature Biotechnol., 15:137-141
Feng, A. V., Yang M.Z., Zheng X.S., Pan N. S.and Chen Z. L. (1992), Agrobacteriummediated transformation of coffee (Coffeaarabica L.), Chinese J Biotechnol., 8 (3) :255-260
Fromm, M., Taylor, M. and Walbot, V. (1985).Expression of genes transferred intomonocot and dicot plant cells byelectroporation.. Proc. Natl. Acad. Sci.,USA, 82: 5824-5828
Guerreiro F. O., Penna M.F., Gonçalves W. andCarvalho A. (1990), Melhoramento docafeiiro: XLIII Selecao do cafeeirosresistentes ao bicho-miniero. Bragantia 49(2): 291-304
Guerreiro, F. O., Denolf, P., Frutos, R., Peferoen,M. and Eskes, A. B. (1993),L' identification de toxines de Bacillusthuringiensis actives contre Perileucopteracoffeella. Asic, l Sême colloque scientifiquesur le café, Montpel1ier, France, pp. 329335
Guerreiro, F. O., Denolf, P., Peferoen, M.,Decazy, B., Eskes, A. B. and Frutos, R.(1998), Susceptibility of the coffee leafminer (Perileucoptera spp) to Bacillusthuringiensis ô-endotoxins: a model fortransgenic perennial crops resistant toendocarpic insects. Current Microbiol. 36:175-179
Jefferson, R. (1987), Assaying chimeric genes inplants: the GUS gene fusion system. PlantMole. Biol. Reporter 5 (4): 387-405
Kosugi, S., Ohashi, Y., Nakayama, K. and Arai,Y. (J990), An improved assay for pglucuronidase in transformed cellsmethanol almost completely suppresses a

putative endogenous ~-glucuronidase
activity. Plant Science 70: 133-140.Laemmli, U.K. (1970), Cleavage of structural
proteins during the assembly of the head ofbacteriophage T4. Nature 227: 680-685
Leroy, T., Royer, M., Paillard, M., Berthouly, M.,Spiral, J., Tessereau, S., Legavre, T. andAltosaar, I. (1997), Introduction de genesd'intérêt agronomique dans l'espêce Coffeacanephora Pierre par transformation avecAgrobacterium sp. In Asic, l Zêrne colloquescientifique sur le café, Nairobi,Kenya, pp.439-446
Leroy, T., Philippe, R., Royer, M., Frutos, R.,Duris, R., Dufou,r M., Jourdan, 1.,Lacombe, C. and Fenouillet, C. (1999),Genetically modified coffee trees forresistance to coffee Jeaf miner. Analysis ofgene expression, resistance to insects andagronomic value. In Asic l Sêrne colloquescientifique sur le café, Helsinki, Finland
Leroy, T., Henry, A-M ., Royer, M., Altosaar, 1.,Frutos,R., Duris, R. and Philippe, R.(2000), Genetically modified coffee plantsexpressing the Bacillus thuringiensiscrylAc gene for resistance to leaf miner.Plant Cell Reports, 19: 382-389
McCabe, D., Swain, W., Martinelli, B. andChristou, P. (1988), Stable transformationof soybean (Glycine max) by particleacceleration. Bio/Technology 6: 923-926
Murashige, T. and Skoog, F. (1962), A revisedmedium for rapid growth and bioassayswith tobacco tissue culture. Physiol. Plant.,15: 473-497
Sardana, R., Dukiandjiev, S., Giband, M., Cheng,X.Y., Cowan, K., Sauder, C. and Altosaar,I. (1996), Construction and rapid testing ofsynthetic and modified toxin genesequences CrylA (b&c) by expression inmaize endosperm culture. Plant CellReports, 15 : 677-681
Schuler, T.H., Poppy, G. M., Kerry B. R. andDenholm I. (1998), Insect resistanttransgenic plants. Tib Tech., 16: 168-175
Spiral, J. and Pétiard, V. (1993), Développementd'une méthode de transformation génétiqueappliquée à différentes espêces de caféierset régénération de plantules transgéniques.In Asic, l Sêrne colloque scientifique sur lecafé, Montpellier, France, pp. 115-122
512
Spiral, J., Thierry, C., Paillard, M. and Pétiard, V.(1993), Obtention de plantules de Coffeacanephora Pierre (Robusta) transforrnéespar Agrobacterium rhizogenes. C.R. Acad.Sei. Paris 316 (série 11I): 1-6
Spiral, J., Leroy, T., Paillard, M. and Petiard, V.(1999), Transgenic coffee (Coffea species).In-: Biotechnology in Agriculture andForestry, Vol. 44, Y.P.S. Bajaj (ed.),Springer-Verlag, Berlin, Germany, pp 5576
Sugiyama, M., Matsuoka, C. and Takagi T.(1995), Transforrnation of coffee withAgrobacterium rhizogenes. In Asic, l ôêmecolloque scientifique sur le café, Kyoto,Japan, pp. 853-859
Towbin, H., Staehelin, T. and Gordon, J. (1979),Electrophoretic transfer of proteins frompolyacrylamide gels to nitocellulosesheets: Procedure and some applications.Proc. Natl. Acad. Sei., USA, 76: 4350-4354
Yasuda, T., Fujii, Y. and Yamaguchi T. (1985),Embryogenic callus induction frorn Coffeaarabica leaf explants by benzyladenine.Plant Cell Physiol., 26: 595-597
Zamarripa, A, Ducos, l.P., Tessereau, H., Bollon,H., Eskes, A and Pétiard, V. (1991),Production d'embryons somatiques decaféier en milieu liquide : effet de ladensité d'inoculation et du renouvellementdu milieu. Café Cacao Thé, XXXV (4):233-244
Zhang, W. and Wu, R. (1988), Efficientregeneration of transgenic plants from riceprotoplast and correctiy regulatedexpression ofthe foreign gene in the plants.Theor. Appl. Gen.r 76: 835-840
Zupan, J.R. and Zambryski, P. (1995), Transfer ofT-DNA frorn Agrobacterium to the plantcell. Plant Physiol. 107: 1041-1047
,•..J

513
A comissão Organizadora do m SIBAC agradece o apoio prestado pelas seguintes empresas:
1- UBY AGROQUÍMICA Ltda.
2- Hokko do Brasil Ltda.
3- Hoesht Shering AgrEvo do Brasil Ltda.
4- PREMOLTEC - Indústria de Pré-moldado Ltda.
5- URSO BRANCO - Indústria de Máquinas e Equipamentos Ltda.
6-' Máquinas Agrícolas Jacto S.A.
7- Milenia Agrociência Ltda.
8- Prefeitura Municipal de Londrina: COMURB.
9- SERCOMTEL.
10- Sociedade Rural do Paraná.
Agradecimentos Especiais:
l-UNIÃO EUROPÉIA - EUROPEAN UNION (EU).
2- CIRAD - Centre de Coopération Internationale em Recherche Agronomique pour le Développement.





























