Embed Size (px)
Citation preview

lable at ScienceDirect
Journal of Human Evolution 84 (2015) 42e61
Contents lists avai
Journal of Human Evolution
journal homepage wwwelsevier comlocate jhevol
A systematic revision of Proconsul with the description of a new genusof early Miocene hominoid
Kieran P McNulty a David R Begun b Jay Kelley c d Fredrick K Manthi eEmma N Mbua e
a Evolutionary Anthropology Laboratory University of Minnesota 395 Hubert H Humphrey Center 301 19th Avenue S Minneapolis MN 55455 USAb Department of Anthropology University of Toronto Toronto ON M5S 2S2 Canadac Institute of Human Origins and School of Human Evolution and Social Change Arizona State University Tempe AZ 85287 USAd Department of Paleobiology National Museum of Natural History Smithsonian Institution Washington DC 20013 USAe Department of Earth Sciences National Museums of Kenya Nairobi 40658-00100 Kenya
a r t i c l e i n f o
Article historyReceived 8 July 2014Received in revised form10 February 2015Accepted 24 March 2015Available online 9 May 2015
KeywordsEkemboUgandapithecusFossil apeTaxonomyTinderetRusinga
Corresponding authorE-mail address kmcnultyumnedu (KP McNulty
httpdxdoiorg101016jjhevol2015030090047-2484copy 2015 Elsevier Ltd All rights reserved
a b s t r a c t
For more than 80 years Proconsul has held a pivotal position in interpretations of catarrhine evolutionand hominoid diversification in East Africa The majority of what we lsquoknowrsquo about Proconsul howeverderives from abundant younger fossils found at the Kisingiri localities on Rusinga and Mfangano Islandsrather than from the smaller samples found at Korudthe locality of the type species Proconsul africa-nusdand other Tinderet deposits One outcome of this is seen in recent attempts to expand the genusldquoUgandapithecusrdquo (considered here a junior subjective synonym of Proconsul) wherein much of theTinderet sample was referred to that genus based primarily on differentiating it from the Kisingirispecimens rather than from the type species P africanus This and other recent taxonomic revisions toProconsul prompted us to undertake a systematic review of dentognathic specimens attributed to thistaxon Results of our study underscore and extend the substantive distinction of Tinderet and UgandanProconsul (ie Proconsul sensu stricto) from the Kisingiri fossils the latter recognized here as a newgenus Specimens of the new genus are readily distinguished from Proconsul sensu stricto by morphologypreserved in the P africanus holotype but also in I1s lower incisors upper and lower canines andespecially mandibular characteristics A number of these differences are more advanced among Kisingirispecimens in the direction of crown hominoids Proconsul sensu stricto is characterized by a suite ofunique features that strongly unite the included species as a clade There have been decades ofcontentious debate over the phylogenetic placement of Proconsul (sensu lato) due in part to there being amixture of primitive and more advanced morphology within the single genus By recognizing twodistinct clades that in large part segregate these character states we believe that better phylogeneticresolution can be achieved
copy 2015 Elsevier Ltd All rights reserved
Introduction
The genus Proconsul (Hopwood 1933) has been a mainstay instudies of hominoid evolution and diversification since its discov-ery in the early 20th century Initially reported from fragmentaryremains found inwestern Kenya seven decades of subsequent fieldwork particularly discoveries from the Kisingiri localities onRusinga and Mfangano Islands have made Proconsul the best
)
documented of all fossil catarrhines Nearly every element of theProconsul skeleton is now known often with variation recorded inmultiple specimens
This wealth of information has not ended debates over thetaxonomy phylogeny functional-adaptive morphology or ecolog-ical preferences of this taxon rather one might argue that theexpansive evidence has provided grist for milling out new ideasand arguments One persistent debate regards whether Proconsulcan be considered a true hominoid or is best placed among stemcatarrhines Following an early period when Proconsul species werelinked more closely to modern great ape lineages (Hopwood 1933Le Gros Clark and Leakey 1951 Simons and Pilbeam 1965 but see
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 43
Leakey 1943) the consensus view emerged of this genus as a basalhominoid (eg Andrews 1985) Around this time Harrison (19821987) raised the possibility that Proconsul and other East Africannon-cercopithecoid catarrhines from the early Miocene might bebetter placed among stem catarrhines phylogenetically precedingthe divergence of the hominoid and cercopithecoid lineagesWalkers (Walker and Pickford 1983 Walker and Teaford 1989Walker 1997 see also Rae 1993 1999) resurrection of the homi-nid (frac14 great apethorn human) status for Proconsulwas ultimately shortlived with most researchers maintaining Proconsul as a basalmember of the Hominoidea
Taxonomic history of Proconsul
Germane to the phylogenetic debate is the taxonomic frame-work within which these fossils are analyzed Proconsul (typespecies Proconsul africanus) was erected for fossil primate speci-mens discovered from 1926 to 1931 at Koru a Western Kenya lo-cality situated on the periphery of the extinct Tinderet volcano(Hopwood 1933 Le Gros Clark and Leakey 1951) In that firstpublication Hopwood (1933460) made oblique but favorablecomparison to recently discovered but unpublished specimensfrom Rusinga Island Yet LSB Leakeys initial presentation of theRusinga material characterized it as being quite different fromHopwoods Proconsul leading Sir Arthur Keith to liken it to theyounger hominoid Dryopithecus (see Myres 1932 Pickford 1986)Leakeys formal publication however included both Koru andRusinga specimens in a single genus which was thought torepresent the base of the human-ape clade (Leakey 1943)
At this time early Miocene primate localities in Western Kenyawere all geographically clustered around either the Kisingiri vol-cano (Rusinga Island Mfangano Island Karungu Uyoma) or theTinderet volcano (Koru Songhor Legetet Hill and others Fig 1)Continued collecting in both regions by the East African Archaeo-logical Expedition and British-Kenya Miocene Expedition graduallyrevealed substantial size variation in the type species Proconsulafricanus and both the Kisingiri and Tinderet localities were re-interpreted as having a smaller and a larger Proconsul species
Figure 1 Map depicting the major Proconsul localities Early Miocene localities clus-tering around the Tinderet volcano include Koru Legetet Hill Chamtwara MeswaBridge Songhor Kapurtay and Mteitei Valley Those associated with Kisingiri areRusinga Island Mfangano Island Karungu and the Uyoma peninsula The middleMiocene type locality of P gitongai is not shown here
This prompted Le Gros Clark and Leakey (1950) to erect two newspecies Proconsul major for the large specimens from Tinderet andProconsul nyanzae for the large Kisingiri specimens The smallerspecies P africanus continued to be recognized in both areas bymost workers Large-bodied catarrhines discovered in Ugandaexpanded the geographic range of P major (Bishop 1964) althoughsome of this sample was later reassigned to a new taxon Moroto-pithecus bishopi by Gebo et al (1997)
This basic taxonomic arrangement remained relatively stable forthree decades although individual specimen identifications did notalways adhere to these simple geographic distinctions (egAndrews 1978) Subsequent studies by Bosler (1981) Harrison(1982 1987) Kelley (1986) and others (Pickford 1986 Teafordet al 1988 Walker and Teaford 1988) further standardized thetaxonomic attributions of specimens and thereby reduced thenumber of Proconsul species recognized at each site P nyanzae and(for some) P africanus associated with Kisingiri localities P majorand P africanus associated with Tinderet localities Ugandan Pro-consul restricted to P major
Fieldwork on Rusinga in the 1980s generated sufficient newevidence to confirm that the smaller Kisingiri Proconsul specimensdiffer morphologically from P africanus (Kelley 1986) leadingWalker et al (1993) to assign this material to a new species Pro-consul heseloni The presence of two species of Proconsul on Rusingaand Mfangano was not universally accepted however Kelley forexample referred all Kisingiri specimens to P nyanzae voicingconcerns over the unlikely distribution of sexes (cf Kelley 1986)and the disregard of likely body size variation within the twospecies (Kelley 1993) Likewise Pickford (1986) recognized onlyP nyanzae from Rusinga and Mfangano notably highlighting thedistinct morphology of Kisingiri Proconsul when compared to Tin-deret and Ugandan specimens Despite these and other objectionsthe presence of P nyanzae and P heseloni at the Kisingiri localitieshas come to be universally accepted (Begun et al 1994 Leakeyet al 1995 Rafferty et al 1995 Ward et al 1995 MacLatchy andBossert 1996 Harrison 2002 2010 Smith et al 2003 Ishidaet al 2004 Nakatsukasa et al 2007 Deane 2009 Harrison andAndrews 2009 Peppe et al 2009 Pickford et al 2009 Michelet al 2014)
The naming of P heseloni after 30 years of relative taxonomicstability heralded a new period of systematic revision First Senutet al (2000) transferred P major into a new genus ldquoUgandapithe-cusrdquo a genus whose validity is debated within the paleoanthro-pological community (see Begun 2007 Harrison and Andrews2009 Harrison 2010) and which we consider to be a junior syn-onym of Proconsul (see below) Pickford and Kunimatsu (2005)subsequently allocated a few isolated teeth from Kipsaraman tothis genus under the species name ldquoUgandapithecusrdquo gitongai Inaddition the proconsulid material from Meswa Bridgedlongrecognized as a distinct species (Andrews et al 1981)dwas giventhe name Proconsul meswae by Harrison and Andrews (2009) Inthat same year however Pickford et al (2009) published acomprehensive taxonomic revision of Proconsul (including ldquoUgan-dapithecusrdquo) based on cranial mandibular and dental material Indoing so they identified a number of similarities between ldquoUgan-dapithecusrdquo major and the smaller Tinderet Proconsul specimensparticularly with regard to mandibular morphology In thatmanner specimens that have been historically referred toP africanus plus the smallest specimens referred to P major (cfPilbeam 1969 Bosler 1981) were transferred to a new speciesldquoUgandapithecusrdquo legetetensis (Pickford et al 2009) Likewise themandible KNM-SO 1112dregarded as P africanus by otherworkersdwas referred to P meswae and the latter species trans-ferred to ldquoUgandapithecusrdquo (Pickford et al 2009) As a result of thissystematic revision P africanus was left almost entirely without
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6144
mandibular representation whereas ldquoUrdquo legetetensis was repre-sented mostly by mandibles and lower teeth (Pickford et al 2009)
Table 1 summarizes the relevant taxonomic allocations used bydifferent authors according to the geographic distribution of lo-calities In its broadest conception (eg Harrison and Andrews2009) the genus Proconsul is quite speciose relative to otherMiocene hominoid genera including as many as six distinct spe-cies This number is substantially reduced under the taxonomicscheme proposed by Pickford et al (2009) wherein half of thesespecies are referred to ldquoUgandapithecusrdquo Those preferring tomaintain all of this material in Proconsul however have noted thatthe greatest distinction within that sample is between the Kisingiriassemblages on the one hand and those from Tinderet and Ugandaon the other (Walker et al 1993 Begun 2001 2007MacLatchy andRossie 2005 Harrison and Andrews 2009 Harrison 2010)daninterpretation presaged in some ways by the taxonomy proposedby Kelley (1986) and Pickford (1986) and ultimately echoing theoriginal presentation by LSB Leakey (Myres 1932)
In accordancewith other researchers (eg Harrison 2002 2010MacLatchy 2004 MacLatchy and Rossie 2005 Suwa et al 2007Harrison and Andrews 2009 McNulty 2010 Hill et al 2013) wedo not recognize the generic distinction of ldquoUgandapithecusrdquo fromProconsul This is due in part to our findings detailed below of adistinct clade uniting P africanus with species assigned to ldquoUgan-dapithecusrdquo In addition the most recent characterization of
Table 1Identifications of Proconsul (sensu lato) species by different authors listed byregiona
Authors KisingiriLocalities
TinderetLocalities
UgandanLocalities
Hopwood 1933 e P africanusb e
MacInnes 1943 P africanus P africanus e
Le Gros Clark andLeakey 1950
P africanus P africanus e
P nyanzaeb P majorb
Andrews 1978 P africanus P africanus P majorP nyanzae P majorP major
Bosler 1981 P africanus P africanus P majorP nyanzae P nyanzaeP major P major
Kelley 1986 P nyanzae P africanus P majorP major
Pickford 1986 P nyanzae P africanus P majorP major
Walker et al 1993 P heselonib P africanus P majorP nyanzae P major
Senut et al 2000 P nyanzae P africanus U majorU majorc
Harrison andAndrews 2009d
P heseloni P africanus P majorP nyanzae P major
P meswaeb
Pickford et al 2009 P heseloni P africanus U majorP nyanzae U major U legetetensis
U legetetensisb U gitongaiU meswaec
This paperd E heselonic P africanus P majorE nyanzaec P major
P meswae
a Very small samples from other regions (eg Buluk) that have been attributed toProconsul (sensu lato) are not included here as they are not critical to understandingthe placement of Kisingiri specimens
b New genus andor species named in publicationc Existing species transferred to a new genusd Proconsul gitongai from the middle Miocene Muruyur Formation (Kipsaraman)
is also recognized but not at the early Miocene localities assessed in this paper
ldquoUgandapithecusrdquo which removed most Tinderet specimens fromProconsul and distributed them among new and existing species ofldquoUgandapithecusrdquo was based largely on contrasts with Rusingaspecimens rather than with the type species of Proconsul (Pickfordet al 2009) This is particularly true for mandibles and the lowerdentitiondspecimens less securely tied to P africanus because thetype specimen M 14084 is a partial maxilla (eg Bosler 1981) Weargue here that if specimens are sorted without preconceived taxo-nomic assumptions the purported diagnostic features of ldquoUganda-pithecusrdquo (sensu Pickford et al 2009) serve instead to differentiateall specimens in the Tinderet and Uganda samples of Proconsul(Proconsul sensu stricto) from those found at Kisingiri localities (cfKelley 1986 Walker et al 1993 Begun 2001 2007 MacLatchy andRossie 2005 Harrison and Andrews 2009 Harrison 2010)
Geochronology of Proconsul localities
The chronological framework of Proconsul localities provides animportant context for understanding historical and current in-terpretations of species distributions and relationships Early re-searchers while acknowledging the limitations of theirstratigraphic and faunal correlations were nevertheless able tocorrectly position the Western Kenyan localities within the lowerMiocene associating some of the deposits with the Burdigalian astage defined by marine sediments in Europe and dated betweenabout 16 and 205 Ma (Kent 1942 MacInnes 1943) The applicationof radiometric dating techniques helped refine these estimateswith some of the earliest KeAr studies conducted on East Africanlocalities associated with Proconsul (Evernden et al 1964 Bishopet al 1969 Van Couvering and Miller 1969) These datesconfirmed an early Miocene age for most localities with estimatesranging between 23 and 16 Ma However the Kisingiri depositsyielded discrepant results due to post-depositional alteration of thedatable biotites (Drake et al 1988) Re-sampling and further ana-lyses yielded more satisfactory and repeatable age estimates of179e178 Ma but notably these were based on only five samplesconfined to a small segment of the Rusinga geological section(Drake et al 1988)
Radiometric dates in conjunction with studies of the associatedfaunal communities (eg Pickford 1981) created a geochronolog-ical framework for Proconsul wherein the Tinderet localities areoldest dating to perhaps 225 Ma at Meswa Bridge (see review inHarrison and Andrews 2009) with Napak in Uganda beingapproximately contemporaneous with Songhor and Koru (Bishopet al 1969 Pickford and Andrews 1981) and the Kisingiri local-ities preserving the youngest sediments (Pickford1981 Drake et al1988) However themost recent re-dating of Kisingiri sites presentsa more complex scenario Rather than being younger than all of theTinderet sites the Kisingiri stratigraphic sequence is substantiallylonger than previously thought with the oldest fossil stratacontemporaneous with those from Koru and Legetet Hill atapproximately 20e19 Ma (McCollum et al 2013) The youngestMiocene fauna from Rusinga is not associated with radiometricdates butmagnetostratigraphic analysis suggests an age near 17Maor younger (Peppe et al 2009) Hence differences between speciesof Proconsul and in particular between assemblages from TinderetandKisingiri cannot be simply explained bychronology Indeed thegreat length of the Kisingiri sequence suggests that taxonomicuniformity throughout this timeperiodmight require re-evaluation
Aims of this study
Motivated by new specimens analyses and interpretations ofProconsul thathave appeared in the last severalyearsweundertookathorough review of the craniodental and mandibular specimens
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 45
attributed to this genus and here provide an alternate taxonomicinterpretation to those in current use Our primary objective was toevaluate the morphological homogeneity of the genus Proconsul astraditionally constituted (Proconsul sensu lato) and explore thepossibility that theKisingiri specimens formadistinct group (Walkeret al1993 Begun 2001 2007MacLatchyandRossie 2005Harrisonand Andrews 2009) While we agree with Pickford et al (2009) thatthe material traditionally referred to Proconsul exceeds what can becomfortably accommodated within a single genus our resultsconform to the suggestion proposed by Begun (2001 2007) and laterby MacLatchy and Rossie (2005) and Harrison and Andrews (2009)that the Kisingiri specimens from Rusinga and Mfangano Islandsconstitute a distinct cladewith respect to Proconsul from theTinderetand Ugandan localities Our assessment of the taxonomic andphyletic distinction of the Kisingiri material necessitates genericdifferentiation of this sample from Proconsul
Systematics
Order Primates Linnaeus 1758Infraorder Catarrhini E Geoffroy Saint-Hilaire 1812Superfamily Hominoidea Gray 1825Family incertae sedisEkembo gen novurn lsidzoobankorgactB11CECF0-9DD1-4265-9C06-A60E4BEE974E
Synonymy
1950 Proconsul Hopwood Le Gros Clark and Leakey 2601965 Dryopithecus (Proconsul) (Hopwood) Simons and Pilbeam
1201978 Proconsul Hopwood Andrews 901986 Proconsul Hopwood Kelley 4791993 Proconsul Hopwood Walker et al 512010 Proconsul Hopwood Harrison 437
Type species
Ekembo nyanzae comb nov (Le Gros Clark and Leakey 1950)
Included species
Ekembo nyanzae (Le Gros Clark and Leakey 1950) Ekemboheseloni comb nov (Walker et al 1993)
Etymology
Ekembo means ldquoaperdquo or ldquomonkeyrdquo in the Suba language whichwas historically spoken by peoples in western Kenya includingthose who settled Rusinga and Mfangano Islands (Jack Wanyendepers comm)
Diagnosis
Medium- to large-bodied and sexually dimorphic hominoidcharacterized by the following combination of features Frontalbone moderately wide at the coronal suture narrowing anteri-orly toward gracile supraorbital rims which are not demarcatedby a post-toral sulcus Large frontal sinuses behind a prominentglabella Minor postorbital constriction Temporal lines promi-nent and situated high on frontal and parietal bones convergingposteriorly without forming a sagittal crest (in female
specimens) Nasal bones long and thin Broad interorbital regionflat rather than projecting Upper face more vertically orientedrelative to the mid- and lower face Nasolacrimal duct positionedwithin a broad orbit Rostrum broad with premaxilla extendingup to the nasal bones Canine jugum prominent in both sexes butcanine fossa developed only in males Nasal aperture broad withwidest portion inferior and sometimes near the base Inferioraperture margin concave or incised in the midline Nasoalveolarclivus intermediate in height between hylobatids and hominidsClivus does not overlap palatine process of the maxilla pre-senting typical mammalian incisive fenestration ante-roposteriorly positioned at or posterior to mid-canine Maxillarysinus extensive reaching anteriorly to P3P4 junction Zygomati-coalveolar crest arises low on the face near M2 Nearly verticalmalar region Broad anterior palate A single palatine foramenpositioned near the M2M3 junction Broad mandibular ramusangled slightly posteriorly from vertical leads to a graduallyrounded and gracile gonial region Mandibular corpora rangefrom short and gracile to taller and moderately robust Sym-physis characterized by moderately developed superior trans-verse torus and variably developed inferior transverse torus andgenioglossal pit Symphyseal planum and subplanum regionvertically or nearly vertically oriented A single mental foramenpositioned near the P3P4 junction approximately halfway up thecorpus or higher
Upper central and lateral incisors heteromorphic Centralincisor characterized by a narrow lingual tubercle often consist-ing of two or three narrow ridges and distinct but thin marginalridges that are continuous with the lingual cingulum with themesial marginal ridge joining the cingulum at a sharp angle incontrast to a more continuously curving transition from cingulumto distal marginal ridge resulting in an asymmetric ridge andcingulum morphology Lateral incisors have well-developedincisal edges and are asymmetric with a moderately developedlingual cingulum Narrow lower incisors with central and lateralincisors having similar morphology Upper and lower caninesstrongly sexually dimorphic with only slight bilateral compres-sion and moderately low-crowned in relation to basal crown di-mensions with apices terminating at a simple point Canine rootssmooth and conical Cheek tooth cusps low and broad and con-nected by inflated intercuspal crests Upper premolars morpho-logically similar with well-developed mesial foveae onlymoderate cusp heteromorphy and cusps positioned at theapproximate mesiodistal midpoint of the crown resulting innearly symmetric buccal profiles Molars with moderate bucco-lingual flare and reduced cingulum development Upper first andsecond molars approximately square with nearly straight distalmargins and buccally oriented posthypocone cristae Lower firstand second molars rectangular with M1 substantially narrowerthan M2 Upper and lower third molars vary in degree of distalcusp development
Forelimbs and hind limbs of similar length Long vertebral col-umn with six lumbar vertebrae characterized by transverse pro-cesses that arise from the dorsal margin of the vertebral centrumand prominent accessory processes for erector spinae musclesPossessing a coccyx rather than a tail Thorax long narrow anddorsoventrally deep Broad flat sternebrae Pelvis long and narrowwith sagittally oriented ilia an elongate ischium lacking osseousevidence for the presence of callosities and a narrow sacrumwith asmall sacro-iliac joint Scapula positioned laterally on thorax with anarrow ovoid and ventrally oriented glenoid fossa Humerus witha posteriorly directed head retroflexed shaft with little torsion anda distal articular surface lacking a well-differentiated trochlea andcapitulum and having a narrow zona conoidea and shallowtrochlear notch but lacking an entepicondylar foramen Radius
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6146
with a small ovoid head and flat distal articular surface that mainlycontacts the scaphoid Ulna displays a large olecranon processnarrow semilunar notch with only a slight keel anteriorly posi-tioned radial notch and a long distal styloid process that retainsarticulation with triquetrum and pisiform Carpals relatively nar-row and include an unfused os centrale a hamate with a smallhamulus and a flat medially oriented triquetral surface Handcharacterized by straight metacarpals with narrow bases andheads Pollex well developed Manual and pedal phalanges gener-ally similar to each other but less so than in Old World monkeysFemur slender with a small head long neck with a high neck-shaftangle centrally placed fovea capitis reduced greater trochanterand a broad distal end with the medial condyle slightly larger thanthe lateral Broad flat patella Tibia long and slender but fibularobust Tarsals and metatarsals elongate with a robust hallux
Differential diagnosis
Ekembo can be readily distinguished from all other medium-and large-bodied catarrhine genera from the early Miocene Cra-nial mandibular and dental differences between Ekembo andProconsul are described in detail below and summarized inTable 2 Ekembo also differs from Proconsul in having a rounderradial head and a femur with a shorter greater trochanter poste-riorly projecting lesser trochanter less extensive inter-trochantericcrest more robust phalanges with more strongly developedfibrous flexor sheath ridges and broader proximal and interme-diate phalangeal distal joint surfaces (condyles) with widertrochlear grooves and less sharply defined trochlear ridges(Harrison 1982 Andrews et al 1997 Senut et al 2000 Pickfordet al 2009)
Ekembo differs from Afropithecus in having a more inflatedglabellar region no frontal trigon less strongly developed anteriortemporal ridges less postorbital constriction a stronger mandib-ular superior transverse torus less posterior shallowing of themandibular corpus higher-crowned lower canines in relation tomesiodistal length buccolingually narrower upper premolarsrelative to molars upper premolars with cusps positioned closer tothe mesiodistal midline reduced P3 cusp heteromorphy a moresymmetric I2 and substantially less buccolingual flare in the cheekteeth (Leakey et al 1988 Rossie and MacLatchy 2013)
Ekembo differs from Heliopithecus in having buccolinguallynarrower premolars relative to molars premolar cusps positionedcloser to the mesiodistal midline and reduced P3 cusp hetero-morphy (Harrison 2010)
Ekembo differs from Equatorius in having a more gracilemandible a larger superior but smaller inferior mandibular trans-verse torus labiolingually narrower lower incisors with straightermesial and distal margins higher-crowned lower canines in rela-tion to mesiodistal length greater molar cingulum development aless posteriorly directed humeral medial epicondyle a more ante-riorly directed radioulnar joint and narrower metacarpal heads(Ward et al 1999)
Ekembo differs from Nacholapithecus in having a shorter sub-nasal clivus a more expansive incisive foramen a broader nasalaperture at the base a minimally developed mandibular inferiortransverse torus a narrower lingual tubercle on I1 higher-crownedlower canines in relation to mesiodistal length a larger humeralradial fossa a shallower coronoid fossa a less well-developed zonaconoidea a narrower and distally slanted ulnar coronoid process alonger and lower femoral neck a more posteriorly positioned lessertrochanter and a smaller posteromedial tubercle on the talus(Ishida et al 2004)
Ekembo differs from Nyanzapithecus in having more anteriorlyextended maxillary sinuses a relatively taller mandibular corpus
less buccolingually compressed upper and lower canines with lesscurved crowns lower and more rounded cusps with more inflatedocclusal crests on the cheek teeth P3 and P4 that more closelyresemble each other and have much less cingular development abroader P4 relative to M1 M1e2 occlusal outline more closelyapproximating a square and lower molars that are broader relativeto mesiodistal length with a more mesially placed entoconid rela-tive to the hypoconid (Harrison 1986 2010)
Ekembo differs from Rangwapithecus in having a broad andexpansive palatal fenestration a wider palate relative to length amore robust mandible with a well-defined superior transversetorus broader upper and lower incisors relative to height lessbuccolingually compressed canines without scimitar-shapedcrowns upper premolars with reduced cingulum and cusps posi-tioned near the mesiodistal midpoint resulting in expanded mesialfoveae and symmetric buccal profiles a broader P4 relative to M1upper and lower molars that are buccolingually broader relative tolength and have more rounded cusps and inflated occlusal crestsmore squared upper molars with substantially less cingulumdevelopment and lower molars with a more mesially placedentoconid (Andrews 1974 Harrison 2002 2010 Cote et al 2013Hill et al 2013)
Ekembo differs from Turkanapithecus in having an expandedglabellar region less postorbital constriction a narrower temporalfossa with less flaring zygomatic arches the lacrimal fossa withinthe orbital margin a relatively broader palate with non-convergingtooth rows narrower nasal bones and interorbital region a distinctmandibular superior transverse torus a more robust mandibularcorpus taller mandibular ramus upper canines more rounded incross-section a reduced cingulum on cheek teeth awell-developedhypocone on M1e2 a more squared M2 with a protocone subequalto paracone and broader lower molars relative to length (Leakeyand Leakey 1986 Harrison 2002 2010)
Ekembo differs from Mabokopithecus in having a relativelybroader M3 crown without distal recurvature of the crown andlacking a buccal concavity a hypoconulid that is in-line with thebuccal cusps more bunodont cusps a well-defined protocristidand a broad open talonid without an accessory cuspid (Harrison2002 2010)
Ekembo differs from Xenopithecus in having reduced uppermolar flare a substantially reduced cingulum no prehypoconecrista molar occlusal profiles more closely approximating a squareand less constricted trigons (Hopwood 1933 Le Gros Clark andLeakey 1951 Harrison 2002 2010)
Ekembo differs from Morotopithecus in having a broader inter-orbital region a better developed canine fossa in males premolarssmaller relative to molars greatly reduced or absent buccalcingula on upper molars lumbar vertebrae with transverse pro-cesses positioned more ventrally on the centrum and a narrowerdistal femur with a deeper patellar groove and similarly-sizedmedial and lateral femoral condyles (Gebo et al 1997MacLatchy 2004)
Recently Stevens et al (2013) likened the new genus Rukwapi-thecus represented by a mandible fragment with well-preservedP4-M3 to nyanzapithecine species Ekembo can be distinguishedfrom this genus in having a more gradually rounded gonial angle aless obliquely oriented P4 with subequal protoconid and metaconidand a broader mesial fovea less mesially positioned metaconidreduced buccal cingula and smaller mesial foveae on the molarssmaller hypoconulids broader M1e2 relative to length a singularrather than twinned metaconid no accessory cuspules in thelingual notch and a well-defined crest between entoconid andhypoconulid
Family Proconsulidae (Leakey 1963)
TabDen
Fe
G
U
U
U
U
U
M
Lo
Lo
Lo
a
b
c
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 47
Genus Proconsul Hopwood 1933
Type species
Proconsul africanus Hopwood 1933
Included species
Proconsul africanus (Hopwood 1933) Proconsul major (Le GrosClark and Leakey 1950) Proconsul gitongai comb nov (Pickfordand Kunimatsu 2005) Proconsul meswae (Harrison and Andrews2009)
Diagnosis (replaces the emended diagnosis of Harrison 2010)
Proconsul species are medium- to large-bodied sexually dimor-phic catarrhines that can be distinguished from other proconsulidgenera by the following combination of features Robust mandib-ular corpus and symphysis characterized by a long shallowlysloping planum alveolare terminating at a large superior transversetorus and a subplanum surface oriented posteroinferiorly Upper
le 2tognathic features that distinguish Ekembo from Proconsula
ature Ekembo
eneralCusp morphology More bunodont occupying more of theMolar crests Inflated incorporated into cuspsCingulum Reducedpper central incisorLingual pillar Narrow generally low reliefMarginal ridges and cingulum Asymmetric with sharply angled cingulu
ridge transitionpper canineTip PointRoot fluting AbsentRoot striations Rare and weakly developedpper third premolarb
Mesiodistal cusp position Crown midpointMesial fovea Well developedBuccal profile SymmetricMesial crown margin Straight or slightly curvedDistal crown margin Slight to moderately curvedCusp heteromorphy ReducedCrown shape Converges on P4 shapeMesiobuccal flange Nopper fourth premolarParacone height Taller relative to crown lengthpper second molarc
Occlusal crown shape SquarishDistal margin Straight to slightly curvedBuccal cingulum Absent or rudimentaryDistolingual cingulum Merges into hypoconePosthypocone crista Distobuccally to buccally orientedBuccolingual flare ReducedandiblePlanum alveolare Short vertically inclinedSuperior transverse torus Moderate to well-developedOrientation of subplanum area More vertically orientedCorpus robusticity More gracilewer incisorsCrown shape Narrow relative to heightwer canineTip Pointwer molarsBuccolingual flare ReducedM1 M2 breadth M1 narrower relative to M2
Although presented here as discrete characteristics for descriptive purposes some oNearly all of these features tend to distinguish P4s as well but in that tooth the diffeThese features tend to distinguish M1s as well but in that tooth the differences are
incisors heteromorphic with I1 exhibiting an inflated lingual tu-bercle and a near symmetrical configuration of lingual marginalridges and cingulum with a curving transition from cingulum tothe mesial marginal ridge Lower incisors broad relative to crownheight Upper and lower canines with distinct blade-like or burin-like tips in both deciduous and adult teeth Upper canine rootswith broad longitudinal grooves or fluting defining two or threedistinct columns plus numerous small longitudinal striationsextending cervically from the root apex Upper P3s morphologicallydissimilar from P4s P3 has substantial cusp heteromorphy (para-cone much taller than protocone) with mesially positioned cuspsand a notched mesial border that together limit or eliminate amesial fovea A long steeply angled preparacrista contributes to ahighly asymmetric buccal profile before being interrupted near thetooth cervix by a mesially projecting flange The distal margin isstrongly curved P4 has a tall paracone relative to crown lengthMolars characterized by extensive cingular development individ-uated conical cusps connected by sharp occlusal crests and sub-stantial buccolingual flare Upper molars rhomboid in shaperesulting from a broad trigon and a distally expanded hypocone M2
has a distinct buccal shelf between paracone and metacone a dis-tolingual cingulum that wraps fully around the hypocone rather
Proconsul
occlusal surface area More distinct individuated conesSharper more distinct from cuspsExtensive
Inflatedbulbousm-mesial More symmetric with curving cingulum-mesial
ridge transition
Blade-likePresentVery common and well developed
MesialAbsent or reducedAsymmetricNotchedStrongly curvedStrongDistinct from P4
Yes
Shorter relative to crown length
RhomboidStrongly curvedDistinct cingular shelfDistinct from hypoconeDistally to distobuccally orientedPronounced
Long more horizontally orientedStrongly developedMore posteriorly inclinedMore robust
Broad relative to height
Blade-like
PronouncedM1 more similar to M2
f these features are undoubtedly correlated aspects of single evolutionary changesrences are not as pronouncednot as pronounced
Figure 2 Upper central incisors of Proconsul and Ekembo Lingual views of Ekembo(KNM-RU 1685 cast) left and Proconsul (KNM-CA 1300) right Note the differences inthe transition between the lingual cingulum and the mesial marginal ridge which ismore rounded in Proconsul and more angled in Ekembo The central pillar is also morebulbous or inflated and without ridging in Proconsul Additional specimens illustratingthe Proconsul morphology are shown in Pickford et al (2009 Fig 17 p 197) butreferred in that manuscript to Ugandapithecus
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6148
thanmerging into the hypocone on its lingual surface and a distallyoriented posthypocone crista Lower first and second molars nearlyequal in buccolingual breadth
Comparative morphology of Ekembo and Proconsul
Samples and comparative methods
The above diagnoses are based on extensive survey and analysisof the original fossil material Specimens housed in the NationalMuseums of Kenya (NMK) and the Natural History Museum (Lon-don) were analyzed by the authors during several separate and twojoint (NMK) research visits Fossils in the Uganda National Museumwere assessed by one of us (KPM) during two visits All measure-ments were collected by KPM Dental terminology follows Szalayand Delson (1979)
The following results are organized with respect to themorphology of the Kisingiri sample (attributed to E nyanzae andE heseloni) versus that of Proconsul from Tinderet and Napak(traditionally referred to P africanus P major and more recentlyP meswae) Specimens from additional localities that have at onetime been attributed to Proconsul (eg Andrews1978 Pickford andKunimatsu 2005 [cf Harrison 2010]) are for the most part notconsidered here most are middle Miocene in age and have beentransferred to other genera (Harrison 2002 2010) Of those that arefrom early Miocene sites there is growing evidence that P majorcan also be found at Moroto and differentiated fromMorotopithecus(Pickford et al 2009) an issue of some interest but ultimately notcritical to the taxonomic questions being considered here Whereappropriate however we do provide observations on a few earlyMiocene specimens attributed to ldquoUrdquo gitongai (Pickford andKunimatsu 2005 Pickford et al 2009 see Harrison 2010) Yetwhile we consider the middle Miocene holotype likely to representa distinct species here referred to Proconsul we concur withHarrison (2010) that many specimens placed in this species byPickford et al (2009) cannot be distinguished from P major
Sample sizes particularly from Tinderet and Ugandan localitiestend to be small but where possible standard univariate (eg t-tests) and bivariate (regression ANCOVA) statistical analyses wereperformed A single eigendecomposition of the covariance matrixof symphyseal measurements was done to characterize the multi-variate shape of the mandibular symphysis after accounting for sizedifferences (sensu Jolicouer 1963) Quantitative results are illus-trated as box-and-whisker plots of shape indices or bivariate plotsof the measurements For samples of fewer than five specimensshape index values are plotted in lieu of box-and-whiskersummaries
Morphological comparisons
Upper incisors Two features of the Ekembo upper central incisorsdistinguish them from those of Proconsul including specimenswidely attributed to the type species P africanus The lingual tu-bercle in Ekembo is quite narrow generallywith low relief and oftenscored by two or more irregular ridges in contrast to the muchmore inflated or bulbous tubercle in Proconsul I1s (Fig 2) Ekembospecimens also have a characteristic morphology in the transitionfrom the lingual cingulum to the mesial marginal ridge (Fig 2)from the mesiodistal mid-point of the crown the cingulumextends either directly mesially or even angles slightly cervicallyto intersect the mesial marginal ridge at a fairly sharp angle Incontrast in I1s of Proconsul the cingulum angles incisally from themid-point and transitions into the mesial marginal ridge in agradual curve The expression of these traits varies somewhat inthe smaller Tinderet specimens attributed to P africanus
compared with those of P major (cf Pickford et al 2009)However these variations are relatively minor with respect to thedifferences between the two genera Of 25 I1s known fromKisingiri localities only one specimen KNM-MW 562 somewhatresembles the Tinderet sample in both of these featuresdhavinga somewhat less angled transition from the mesial marginal ridgeto the cingulum and a more bulbous lingual tubercle This isparticularly interesting as the specimen was discovered in theMakira Beds now dated to older than 190 Ma and contemporarywith the Tinderet localities (McCollum et al 2013) Neverthelessits overall morphology is more similar to other Ekembospecimens than to Proconsul incisors Two other Ekembospecimens KNM-RU 1677 and RU 1933 have somewhat moreinflated lingual tubercles but otherwise resemble other Ekembo I1s
No consistent differences were found in upper lateral incisors
Upper canines Upper canine crowns of Ekembo are typical of thoseof most catarrhines in terminating in an evenly tapering point Thisdiffers from the Proconsul specimens which have a blade-like(Kelley 1986 Senut et al 2000 Harrison 2010) or ldquoburin-likerdquo(Pickford et al 2009) canine tip (Fig 3) While blade-like apicalmorphology has long been recognized as a distinctivecharacteristic of P major we confirm the report by Pickford et al(2009) that this feature is also present in the P africanus typespecimen (Fig 3a) although somewhat understated as expectedin the more slender and higher-crowned P africanus caninesUnfortunately other canines attributed to P africanus (eg M44837 KNM SO 419 LG 921 CA 1910 SO 5353 Kelley 1986) havebroken apices and therefore cannot be evaluated for this featureA blade-like tip is also present in the deciduous upper canines ofP major (eg KNM-SO 371 MV 10 NAP IV UMP 62-20) andP meswae (KNM ME 11) Hence the blade-like upper canine tipunites all Tinderet and Napak Proconsul species but is neverexpressed in Ekembo specimens from Rusinga and Mfangano
Two features of the canine roots also differentiate these generalongitudinal grooving or fluting along the length of the root (cfKelley 1986) and the development of numerous finer longitudinal
Figure 3 Canines of Proconsul and Ekembo a) Upper canine of the P africanus holotype (M 14084) lingual view b) Upper canine of P major (KNM-CA 2127) lingual view c) Lowercanine of P major buccal view d) lower canine of Ekembo cf nyanzae (KNM-RU 1676) buccal view and e) Upper canine of P major (KNM-SO 584) distal view Note the blade-like orldquoburin-likerdquo tip of the unworn Proconsul canines including on the nearly unworn canine tip of the P africanus type specimen (a) Ekembo canines have more typical pointed tipsProconsul canines also have longitudinal grooves and fine striations on the root (e) Ekembo lacks these features Scale bars frac14 1 cm
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 49
striations near the root apex that extend to varying degrees towardthe cervix of the tooth Longitudinal ldquoflutingrdquo consists of broadgrooves that segment the root into distinct sections (cf Pickfordet al 2009 Fig 3e) Most often the root in cross-section has theappearance of a cylinder that has been sectioned longitudinallywith the two halves offset from each other but in some cases thereare three segments and the root cross-section is more triangular inappearance (Kelley 1986) This characteristic was noted by Kelley(1986) in the P africanus canines but not in those of P majorHere we affirm that fluting is present on all of the visible uppercanine roots attributed to both P africanus and P major rootmorphology could not be evaluated in the deciduous upper caninefrom Meswa Bridge since it is implanted in the maxillary alveolusNone of the Ekembo specimens display this morphology havinginstead a simpler round to ovoid cross-section along the length ofthe root
The fine longitudinal striations on Proconsul upper canine rootsvary in the degree to which they extend toward the cervix (Fig 3e)but they are visible in all specimens attributed to P africanus(KNM-SO 419 521 921 5353 KNM-X 502) and five of six uppercanines of P major (excepting KNM-SO 1614) This morphology isalso present in UMP 62-12 which was included as part of theMorotopithecus bishopi hypodigm (Gebo et al 1997) Fine striationsalong the upper canine root are almost entirely absent in EkemboTwenty of twenty-three specimens lack this feature three speci-mens (KNM-RU 1763 1845 1891) exhibit incipient to slight
striations but these are never developed to the extent seen inProconsul canines
Upper premolars Upper third premolars display some of thegreatest differences between Ekembo and Proconsul P4s tend todiffer in similar ways but the differences are not nearly as markedEkembo P3s have cusps positioned near the mesiodistal midpoint ofthe crown well-developed mesial foveae pre- and postparacristaeof similar length and slope resulting in symmetric or nearsymmetric buccal profiles a straight or slightly curved mesialmargin and only moderately curved distal margin (Fig 4)Proconsul P3s are more similar in appearance to those of small-bodied catarrhines from the early Miocene with cusps positionedmesially on the crown greatly reducing or eliminating the mesialfovea They also have a steep preparacrista and more shallowlysloped postparacrista resulting in a highly asymmetric buccalprofile and an inwardly angled or notched mesial margin but astrongly curved distal margin This morphology is bestcharacterized in the P africanus holotype (M 14084) but can alsobe seen in other specimens attributed to this species as well asthose referred to P major (Fig 4d e) Also noteworthy is adistinctive mesiobuccal flange that mesially extends the buccalportion of the crown but only near the cervical line of the toothPickford et al (2009) describe this feature in some detail forldquoUgandapithecusrdquo but in fact this morphology is clearly expressedin the P africanus holotype as well (Fig 4a d) A single adult P3
Figure 4 Upper premolars of Proconsul and Ekembo occlusal (aec) and lingual (def) views a) P3e4 of the P africanus holotype (M 14084) b) P3 of P major (M 14331) c) P3e4 of theE heseloni holotype (KNM-RU 2036) d) P3 of the P africanus holotype (M 14084) e) P3 of P major (M 14331) f) P3 of the E heseloni holotype (KNM-RU 2036) Note the mesiallypositioned cusps lack of mesial fovea and strongly asymmetric crown in Proconsul compared to Ekembo (see text for further comparisons) Image of M 14331 reversed for easiercomparison Photo 4a courtesy of Rutger Jansma Scale bars frac14 1 cm
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6150
germ and two dP3s attributed to P meswae look remarkably similarto those of other Proconsul species including the presence of thisflange In contrast the Ekembo P3 has a well-developed mesialfovea that terminates at a mesiobuccal shoulder positionedapproximately halfway between the paracone tip and the cervixFrom this shoulder the mesial margin angles slightly distallydown to the cervix (Fig 4f)
The degree of cusp height heteromorphy in P3 also differentiatesEkembo from Proconsul (contra Pickford et al 2009) Only four non-Kisingiri specimens were sufficiently unworn to take measure-ments but more worn specimens also appear to express this dif-ferential in cusp height Figure 5a shows the ratio of protoconeparacone height measured from the tooth cervix and with only oneexception Ekembo P3s have substantially less cusp heteromorphyThis exception KNM-RU 1677 does not in fact have greater heter-omorphy but instead has an abnormal extension of the enamel ontothe buccal (paracone) root which skews the measurement Hencecusp relief relative to the occlusal surface in RU 1677 is not nearly asheteromoprhic as in Proconsul and when this is taken into accountthe difference between genera is significant (t-test dffrac14 11 t frac14 283p frac14 00165) The adult P3 germ from Meswa Bridge is incompletelyformed and therefore could not be measured However the differ-ence between paracone and protocone heights with respect to theocclusal surface appears to be as great as that in the P africanusholotype It does not appear that sex is an important determinant ofthis feature since well-established male (eg M16647 KNM-RU16000) and female (KNM-RU 2036 7290) specimens do not segre-gate across the Kisingiri range of variation
Upper fourth premolars of Ekembo are broadly similar inmorphology to Ekembo P3s in fact the similarity between P3 and P4
is substantially greater in Ekembo than in Proconsul (Figs 4a c)Likewise most of the morphological differences that discriminatethe P3s of these genera also tend to differentiate the P4s but less
clearly Cusp heteromorphy does not distinguish P4s of thesegenera but the overall height of the P4 paracone in Ekembo issignificantly greater relative to crown length than in Proconsul P4s(t-test df frac14 14 t frac14 316 p frac14 00069 Fig 5b)
Upper molars Cusps of Ekembo upper molars are uniformly morebunodont than those of Proconsul occupying more of the occlusalsurface Intercuspal crests are more fully incorporated into thecusps in Ekembo molars giving them a more inflated appearanceThis is particularly evident in the crista obliqua which is very broadin Ekembo but thin and knife-like in Proconsul (Fig 6) Cusps ofProconsul molars stand more as individual cones Pickford et al(2009) have further differentiated the cusp morphology ofsmaller and larger Tinderet specimens arguing that the latterhave more bulbous cusps While accurate this difference is minorcompared with the morphological difference between Ekemboand Proconsul and we regard the within-Proconsul cuspalvariation as size-related In fact the intermediately sizedP meswae has cusp morphology that is appropriatelyintermediate between that of P africanus and the larger P major
Other differences between Ekembo and Proconsul upper molarsare most evident in M2 We concur with previous assessments thatthe Kisingiri upper molars have less developed cingula than non-Kisingiri molars (eg Senut et al 2000 Harrison 2002 2010Harrison and Andrews 2009) More specifically Ekembo M2s lackor have only rudimentary buccal cingula between the paracone andmetacone Likewise the lingual cingulum is discontinuous merg-ing into the hypocone lingually and then re-emerging as a distalmarginal ridge (Fig 6) Proconsul M2s display substantially morecingular development overall and in particular exhibit a prominentbuccal shelf between the paracone and metacone and a distinctdistolingual cingulum that is continuous around the hypocone towhere it merges with the distal marginal ridge
Figure 5 Box-and-whisker plots a) P3 cusp heteromorphy index (protocone heightparacone height) Results of a t-test show this difference to be significant (p frac14 00165) Theexcluded Ekembo outlier KNM-RU 1677 only appears to have extreme heteromorphy because of extensive expansion of the buccal enamel onto the anterior root (see text) b) P4
relative paracone height (paracone heightmesiodistal crown length) Results of a t-test show this difference to be significant (p frac14 00069)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 51
The occlusal profiles of Ekembo M2s are squarish in shape withrelatively straight and equal sides with a posthypocone crista thatis distobucally to buccally directed In contrast the occlusal profilesof Proconsul M2s have a more rhomboid shape with substantialbasal flare around the mesial cusps and a distally expanded hypo-cone with a distally directed posthypocone crista resulting in astrongly curved distal margin (Fig 6) Even in specimens with lesscurved distal margins the posthypocone crista is still more distallydirected compared to the condition in Ekembo These features arecharacteristic not only of P africanus P meswae and P major butare also expressed in the isolatedM2 fromMoroto Uganda (MOR IIb2098) attributed by Pickford et al (2009) to ldquoUrdquo gitongai butregarded here as belonging to P major
As noted by others (eg Harrison and Andrews 2009) the de-gree of buccolingual molar flare differs among these samplesQuantified as the distance between the tips of the mesial cuspsdivided by maximum crown breadth the degree of molar flare in
Ekembo M2s is significantly less than in Proconsul M2s (t-testdf frac14 29 t frac14 396 p frac14 00004) In a bivariate plot of these twomeasures the distributions of Ekembo and Proconsul teeth barelyoverlap with only KNM-RU 1677 encroaching into Proconsul ter-ritory (Fig 7)
These differences in M2 morphology also tend to distinguishEkembo from Proconsul M1s but not always as clearly Whereas M2smore consistently express distinct morphotypes the M1s in bothgenera show more variation in these features
The degree of upper third molar development (ie whether M3
has four distinct cusps or the distal cusps are reduced or absent) haslong been cited as taxonomically informative for these species orgroups of these species (eg Hopwood 1933 Le Gros Clark andLeakey 1951 Andrews 1978 Pickford et al 2009) However wefound this tooth to be unreliable for taxonomic purposes Withinprimates M3s are typically the most variable molars in both sizeand shape (Blankenship et al 2007 Kavanagh et al 2007) and this
Figure 6 Occlusal view of the M2s of Proconsul and Ekembo Left P africanus holotype(M 14084) Right E heseloni holotype (KNM-RU 2036) Note in Proconsul the overalldifference in crown shape as well as the more conical individuated cusps and nar-rower cristae strongly developed lingual cingulum that continues to the distal aspectof the crown broad buccal cingular shelf and more distally directed posthypoconecrista In Ekembo the cusps are more bunodont and expanded the cristae are broaderwith better developed accessory tubercles the lingual cingulum merges into thehypocone there is no buccal shelf and the posthypocone crista is more buccallydirected
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6152
is observable even in small samples of Proconsul species TheP africanus holotype has a substantially reduced M3 whereas inKNM-CA 2250dattributed by most authors to this species(Andrews 1978 Pickford et al 2009)dit is fully developed Simi-larly the hypodigm of P major includes both fully developed M3s(eg KNM-CA 389 LG 1815) and specimens with reduced ormissing hypocones (KNM-CA 397 1299) Ekembo displays compa-rable ranges of variation from highly reduced to fully developedM3s in both species Thus these species and genera cannot bedistinguished based on M3 development and we did not find anyother distinguishing characteristics between Ekembo and Proconsulin the upper third molars
Figure 7 Scatter plot depicting the de
Mandible Pickford et al (2009) enumerated several traits thatreadily distinguish Kisingiri from non-Kisingiri mandibles andthese distinctions are entirely consistent with the distinction ofEkembo from Proconsul Our analyses specifically underscoredifferences in the symphyseal region The planum alveolare inboth male and female specimens of Ekembo is short and morevertically oriented even the most elongate of these (KNM-RU47805 McNulty et al 2007) is substantially more verticalcompared to specimens of Proconsul In contrast the planumalveolare in Proconsul is conspicuously long and nearlyhorizontally oriented Well-preserved mandibles attributed toP major (eg KNM-SO 396 and 404) exhibit this morphology inextreme fashion (Fig 8) smaller specimens (KNM-SO 1112P africanus) show less extreme expression but are still readilydistinguishable from Ekembo mandibles For its size KNM-LG 452has a surprisingly short planum alveolare though not to thedegree seen in Rusinga mandibles Correlated to the shortplanum alveolare in Ekembo is a reduced superior transversetorus which while well-developed does not approach therobusticity expressed in Proconsul mandibles This difference isreflected in an index of symphyseal robusticity (thicknessperpendicular to the long axis of the symphysislength of thelong axis) which shows Ekembo and Proconsul to be significantlydifferent (t-test df frac14 14 t frac14 430 p frac14 00007)
Development of the inferior transverse torusdoften used todifferentiate among species of Proconsul (eg Pickford et al 2009Harrison 2010)dwas found to be highly variable and inconsis-tently expressed within these samples However the overallorientation of the subplanum region (as defined by a line con-necting the posterior-most midline points of the superior andinferior transverse tori or the posterior-most midline point of theinferior margin of the symphysis in the absence of the latter) doesdistinguish Ekembo from Proconsul mandibles The orientation ofthis line in relation to the alveolar margin in the vicinity of thepostcanine teeth was assessed qualitatively by placing a small rod
gree of buccolingual flare in M2
Figure 8 Mandibles of Proconsul and Ekembo Occlusal views of P major a) KNM-SO 396 b) KNM-SO 404 c) KNM-LG 452 Occlusal views of Ekembo d) KNM-RU 2087 e) KNM-RU1674 f) KNM-RU 7290 Occlusal views of P africanus g) KNM-SO 372 h) KNM-SO 1112 i) Lingual view of P africanus (KNM-SO 1112) Note the long more horizontally orientedplanum alveolare in Proconsul resulting in a pronounced superior transverse torus The planum alveolare is shorter and more vertical in Ekembo such that one can see the inferiormandibular border from occlusal view
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 53
(eg a dental pick) in themidline internally along the two referencepoints and recording the point in lingual perspective at which therod intersected the alveolar margin (Fig 9) In Ekembo the sub-planum region is more vertically oriented so that the rod intersectsthe alveolar margin most often in the premolar field Not surpris-ingly there is a difference between sexes with females having areduced superior transverse torus and therefore a more verticallyoriented subplanum region compared to males In contrast withEkembo Proconsul mandibles have much more inclined subplanumsurfaces with the rod intersecting the lingual alveolar margin inthe vicinity of the M1 (females) or M2 (males Fig 9) When sex istaken into account therefore the distributions of Ekembo andProconsul in this feature are discrete Note that male and femalemandibles attributed to both P africanus and P major are includedin this assessment and are surprisingly similar despite dramaticsize differences
We also quantified internal symphyseal morphology using threemeasures symphyseal height (infradentaleegnathion) planumalveolare length (posterior-most point on the superior transversetoruseinfradentale) and subplanum length (posterior-most pointon the superior transverse torusegnathion) a combination thatincorporates not only the lengths of these intervals but also theangular relationships among them To compare symphyseal shapeacross a range of sizes we chose a simple principal componentsapproach to partition size onto the first eigenvector (cf Jolicouer1963) While this is not the preferred method for partitioning sizein most datasets (Jungers et al 1995) for this particular sample andset of measurements Mosimann shape variables were still
significantly correlatedwith size (see discussion in Coleman 2008)Hence we assessed differences in symphyseal morphology be-tween Ekembo and Proconsul using the second and third principalcomponents of the three variables Distributions of the two generaare distinct in this shape space (Fig 10 Supplementary OnlineMaterial [SOM] Table 1)
Robusticity of the mandibular corpus also differs between thesesamples though it is more difficult to test due to differentialpreservation among specimens Corpus thickness and depth underP4 can be measured in the greatest number of specimens andcomparison of these measures suggests a difference betweengroups in the scaling of corpus robusticity (Fig 11) Among largepresumably male specimens Ekembo mandibles are substantiallyless robust than those of Proconsul However among smallermandibles only a single Proconsul specimen (KNM-SO 372) could beincluded and it is similar in robusticity to Ekembomandibles of thissize Considered together the impression is that corpus robusticitymay scale differently in Ekembo than it does in Proconsul and a testfor differences in slopes using an ANCOVA design of the loggedvariables was significant (df frac14 2 F frac14 2481 p lt 00001) Never-theless additional specimens are needed to properly evaluate theinfluence of taxonomy sex and size on corpus robusticity areasonable alternative interpretation of Figure 11 is that it dem-onstrates a significant difference between P major and all otherspecies of both Proconsul and Ekembo
Lower incisors Ekembo specimens are uniformly narrower inmesiodistal length than incisors attributed to P africanus P major
Figure 10 Scatter plot of principal component scores based on an eigenanalysis of threeposterior point on the superior transverse toruseinfradentale) and subplanum length (mprimarily captures size differences among specimens (Jolicouer 1963 see text and SOM Trespectively) in the symphyseal variables
Figure 9 Line drawing of a hemimandible depicting different orientations of the sub-planum area in Ekembo and Proconsul Females are denoted using shorter vectors maleswith longer vectors Vector is estimated in M 14086 which is broken at the midlineEkembo specimens have a more vertical subplanum compared to those from Songhorand Legetet but within both groups females have more vertically oriented regions thanmales Hence even the most ventrally rotated subplanum in the largest Ekembo malespecimen (KNM-RU 47805) groups with the Tinderet females in this feature
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6154
and P meswae both absolutely and in relation to crown height(Fig 12) This is true for both I1 and I2 A t-test adjusted for non-independent observations (eg I1 and I2 from the same mandible)confirmed the difference as significant (t-test df frac14 12 t frac14 258p frac14 00242) Moreover while the measurable Proconsul samplewas small this same pattern is observable in other specimenswhose tips were slightly to moderately worn and therefore notmeasured
Lower canines The only lower canine trait that discretely sepa-rates these genera is the pointed crown tip in Ekembo (Fig 3d)contrasted with a blade-like (or ldquoburinrdquo) tip in Proconsul(Fig 3c) This feature is well established for P major but is alsoexpressed in KNM-CA 1772 which was referred to P africanus byKelley (1986) and is too small to be either P major or P meswaeTwo other specimens commonly attributed to P africanus KNM-CA 2149 and KNM-SO 1112 both have broken canine tipsLikewise a new specimen (Kor 65004) attributed to P africanusby Pickford et al (2009) also has a broken tip and cannot beevaluated (see Pickford et al 2009 Fig 23) As with the uppercanines the blade-like tip is also present in the deciduous lowercanines of P major (eg KNM-CA 361 S0 5352) and P meswae(KNM-ME 1)
Lower premolars Like most previous authors (eg Andrews 1978Harrison 2002 2010 Pickford et al 2009) we found neitherqualitative nor quantitative characteristics of the lower premolarswith any taxonomic value
Lower molars Differences in lower molar morphology are few andrelatively subtle but in some cases mirror those seen in the uppermolars Cusps on the lower molars of Ekembo are more bunodontthan those in Proconsul and occupy more of the occlusal surfaceBroad inter-cuspal crests are incorporated into the Ekembo cuspsmaking them appear more inflated Molar cusps of Proconsul are
variables symphyseal height (infradentaleegnathion) planum alveolare length (mostost posterior point on the superior transverse torusegnathion) The first componentable 1) and thus PC 2 and PC 3 illustrate the residual shape variance (72 and 28
Figure 11 Scatter plot showing mandibular corpus robusticity under P4 Trend lines were computed using ordinary least squares (OLS) regression Similar relationships areobservable at other parts of the corpus but differential preservation results in smaller samples at those positions Slopes are significantly different (p lt 00001 tested on loggedvariables)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 55
more distinct from one another and are linked by narrower sharperinter-cuspal crests The lower molars of Ekembo also have asubstantially less developed buccal cingulum than those ofProconsul (Harrison 2002 2010 Harrison and Andrews 2009)
Figure 12 Scatter plot of lower incisor crown proportions Differenc
Lastly and again as seen in the upper molars the two genera canbe distinguished by the significantly greater amount of lowermolar flare in Proconsul (Fig 13 t-test df frac14 59 t frac14 713p lt 00001 see also Harrison and Andrews 2009)
es between Ekembo and Proconsul are significant (p frac14 00242)
Figure 13 Box-and-whisker plot of buccolingual flare in lower molars measured asthe average distance between the mesial buccal and lingual cusps divided by overallbuccolingual crown breadth Differences between Ekembo and Proconsul are highlysignificant (p lt 00001)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6156
Relative molar size has been used to distinguish species ofProconsul but we observed the primary difference to be in relativebuccolingual breadths of M1 and M2 In Ekembo M1 is substantiallynarrower than M2 than is the case in Proconsul (Fig 14 Harrison2002 2010) Although none of the smaller Proconsul mandiblespreserve complete crowns for both M1 and M2 KNM-SO 1112 hasan intact M2 and at least preserves the cervix for M1 However evenif the cervical measurement is used for its M1 breadthdthusunderestimating the true crown breadthdKNM-SO 1112 still has abroader first molar in relation to M2 than would be expected inEkembo If crown breadth is estimated from the ratio of cervixbreadth to maximum crown breadth in other Proconsul fossils therange of likely breadths for the SO 1112 M1 clearly distinguishes it
from the Ekembo sample (Fig 14) Testing this relationship onlogged variables using the mean estimate for the SO 1112 M1(Fig 14) reveals a significant difference between the two groups(ANCOVA df frac14 1 F frac14 2424 p frac14 00002)
As with upper third molars we find a great deal of variation inM3 morphology and mistrust the taxonomic significance some-times placed on characteristics of this toothdat least with regard tothese taxa Harrison (2002 2010) cited the mesiodistal position ofthe M3 entoconid as differentiating Kisingiri third molars fromthose attributed to P africanus We concur with his assessment butfound the entoconid position to vary in specimens attributed toP major Thus this character does not differentiate Ekembo from allProconsul species but may distinguish it from the type speciesP africanus
Discussion
The distinctiveness of the Kisingiri large catarrhine materialfrom that at Tinderet and Ugandan localities has been noted pre-viously by several authors (Pickford 1986 Walker et al 1993Begun 2001 2007 MacLatchy and Rossie 2005 Harrison andAndrews 2009 Harrison 2010) What had not been fully docu-mented was the nearly complete lack of overlap between thesesamples in a number of features despite very large assemblagesfrom Rusinga and Mfangano On descriptive criteria alone (Table 2)many isolated teeth (I1 C1 P3 M2 C1) and all of the mandibles canbe assigned to either Proconsul or Ekembowith very little chance ofmisattribution That some of the most diagnostic features occur inthe upper dentitiondthe only anatomy preserved in theP africanus type specimendadds weight to the taxonomicarrangement proposed here The morphology of the upper teeth inall Proconsul species compellingly unites them into a single cladedistinct from E heseloni and E nyanzae This is reinforced bynumerous features of the mandible and mandibular dentition thatfurther distinguish these samples The number and variety of thesefeatures are such that Proconsul and Ekembo are more differentfrom one another than are many closely-related modern catarrhinegenera
Derived morphology of Ekembo and Proconsul
It is premature to formally assign primitive and derived status tothe individual features of Ekembo and Proconsul before completinga formal phylogenetic analysis This is especially true given theabundance of evidence available for Ekembo and hence the poten-tial impact that our generic revision could have on phylogeneticinterpretations Nevertheless in comparison with both extanthominoids and the small-bodied catarrhines from the earlyMiocene it is informative to differentiate the evolutionary grades ofthese genera Many characteristics of Ekembo are more advanced inthe direction of modern apes compared to those of Proconsul(Table 2) General trends such as expanded and more bunodontcusps broader molar crests reduced cingulum and morphologicalsimilarity between P3 and P4 all represent morphologies thatpresage those of later fossil and extant hominoids The moreadvanced morphologies of P3 and M2 are particularly conspicuouscompared not only with those of Proconsul but to most earlyMiocene catarrhines In contrast the mesially positioned stronglyheteromorphic cusps on the P3 of Proconsul plus the correspondingasymmetric buccal profile notched mesial margin and missingmesial fovea are characteristic of other early Miocene catarrhinessuch as Dendropithecus Rangwapithecus and Limnopithecus andare likely primitive among East African catarrhines The moresymmetric P3 of Ekembo with midline positioned cusps andreduced cusp heteromorphy is reminiscent of later hominoids and
Figure 14 Relative buccolingual dimensions of M1 and M2 KNM-SO 1112 has a preserved M2 but only the cervix of M1 Its M1 breadth was estimated based on the ratio to cervixbreadth to maximum crown breadth in other Tinderet fossils Even the cervical breadth of SO 1112 is broader than would be predicted for the crown breadth of a Kisingiri M1 Trendlines computed using OLS regression and using the average estimate of SO 1112 M1 breadth Group differences were significant (p frac14 00002)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 57
even hominid taxa such as Rudapithecus (eg Kordos and Begun2001)
Proconsul is likewise united by a suite of features that are for themost part characteristic only of this lineage (Table 2) Themorphology of both upper and lower canines is unique to thisgenus and shared by all three well-known species plus the poorlyrepresented middle Miocene P gitongai (Pickford and Kunimatsu2005) The mandibular symphysis is also distinctive in Proconsulcompared with other catarrhines with its very long and shallowlysloping planum alveolare and steeply inclined subplanum surfaceIndeed even the small edentulous anterior mandibular fragmentKNM-SO 372 is readily placed within Proconsul and excluded fromother similarly sized catarrhine taxa such as Rangwapithecus andEkembo based on this unique symphyseal morphology (Figs 8 and9)
Other differences between Ekembo and Proconsul are of uncer-tain evolutionary polarity (eg orientation of the posthypoconecrista) or may be of taxonomic relevance without broader phylo-genetic significance (eg degree of molar flare Table 2) Werecognize that there are morphological differences within Procon-sul particularly between the better known species P africanus andP major However many of these differences are likely related tosize and represent variations on a single generic theme that aremarkedly different from the morphology of Ekembo The recentdescription (Harrison and Andrews 2009) of the intermediatelysized P meswae underscores the broad morphological unity withinProconsul and further substantiates a well-defined Proconsul cladecomprising Tinderet and Ugandan species
Based on the abundant morphological and morphometric evi-dence supporting genus-level distinction between Proconsul andEkembo we are currently conducting a more detailed phylogeneticanalysis of all early Miocene catarrhines in order to better under-stand the distribution and evolutionary significance of these andother morphological features We anticipate that the debates sur-rounding the phylogenetic relationships of an historically consti-tuted sample of Proconsul can be more satisfactorily resolved nowthat the Kisingiri material is appropriately recognized as a distinctgenus
The generic disposition of ldquoUgandapithecusrdquo species
A great many features said to characterize ldquoUgandapithecusrdquowere added to the original diagnosis of the genus by Pickford et al(2009) in their systematic revision of Proconsul (sensu lato) As partof that study they also erected a new species ldquoUrdquo legetetensis andtransferred P meswae to ldquoUgandapithecusrdquo Evaluating the same setof features across the entire available sample however we wereunable to find support for this more expansive conception ofldquoUgandapithecusrdquo for several reasons First some diagnostic fea-tures cited in Pickford et al (2009) were quite variable within theldquoUgandapithecusrdquo sample As an example marked reduction of theM3 entoconid to a bead on the lingual cingulum is not shared byseveral ldquoUrdquo major specimens such as KNM-CA 393 NAP I 49000and Kor 253004 (see Pickford et al 2009 Fig 33e) and was likelynot a feature of a few of the more worn specimens eg BUMP 600and BUMP 601 or even the holotype of P major M 16648 Second
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6158
some diagnostic features of ldquoUgandapithecusrdquo were found to bevariably present in Ekembo arguing against their utility for dis-tinguishing ldquoUgandapithecusrdquo from Pickford et als (2009) com-bined P africanus thorn Kisingiri conception of Proconsul For examplesignificantly reduced curvature of the I1 distal margin in ldquoUganda-pithecusrdquo is also found in some Kisingiri specimens including KNM-RU 1685 1712 1714 1831 1846 and 1951 several more Rusingaspecimens have moderately reduced curvature Third some fea-tures that purportedly distinguish ldquoUgandapithecusrdquo fromP africanus are likely related to size For example the somewhatmore central position of the P3 paracone in ldquoUrdquomajor is not seen inthe smaller ldquoUrdquo meswae whose paracone is positioned similarly tothat of the holotype of P africanus (see Harrison and Andrews2009 Fig 6d)
However the major part of the evidence for recognizingldquoUgandapithecusrdquo (sensu Pickford et al 2009) was assembled by re-assigning specimens traditionally attributed to P africanus into newspecies of ldquoUgandapithecusrdquo The P africanus holotype was left inassociation with the Rusinga sample despite having morphologyoverwhelmingly characteristic of other Tinderet specimens Theresulting specimen allocations left P africanus morphologicallyimpoverisheddstripped of much of its hypodigmdso that Pro-consul for Pickford et al (2009) came to be characterized primarilyby the Rusinga sample (see similar comments in Harrison andAndrews 2009) Within that conceptual framework any spec-imen that differs from those at Rusinga could not be Proconsul butmust instead belong to ldquoUgandapithecusrdquo Thus specimens such asthe KNM-SO 1112 mandible were erroneously assigned to ldquoUgan-dapithecusrdquo because they were (correctly) perceived as differingfrom their Rusinga counterparts (Pickford et al 2009)
Here we restore the P africanus hypodigm and associate it withthe other Tinderet fossils according to their shared morphologySpecimens assigned to ldquoUgandapithecusrdquo revert to Proconsul andmany of the features cited to support a speciose ldquoUgandapithecusrdquo(Pickford et al 2009) instead become diagnostic features of Pro-consul This correction serves to highlight the distinctiveness of theKisingiri remains and the need to recognize this distinctiveness atthe generic level
As demonstrated above specimens from Kisingiri do not displaythe morphology of Proconsul as represented by the holotype ofP africanus M 14084 Given that this specimen preserves only theupper canine and cheek teeth similarities with or departures fromthis morphology represent the most compelling arguments forassessing inclusivity in the genus However for the mandible andlower dentition if one simply sorts the specimens by elementwithout relying on taxonomic preconceptions the overwhelmingdistinction within the traditionally constituted Proconsul sample isbetween Kisingiri material on the one hand and that from Tinderetand Uganda on the other (cf Walker et al 1993 Begun 2001 2007MacLatchy and Rossie 2005 Harrison and Andrews 2009Harrison 2010) Additionally with the Kisingiri specimensremoved to Ekembo the postcranial differences used to partlyjustify a distinct ldquoUgandapithecusrdquo lose their relevance (seeHarrison and Andrews 2009)
We have differed among ourselves in our past treatment ofP major as generically distinct from the smaller species of Pro-consul (Begun 2007 McNulty 2010) Morphologically many ofthe unique features of P major can be interpreted as size-relatedvariants on the smaller P africanus morphology Even if so thequestion is whether these are sufficiently different and numerousto warrant generic distinction that is do they suggest thatP major belongs in a separate clade from P africanus andP meswae As we have demonstrated differences among thesethree species are certainly fewer and show less morphologicaldistinction than those between Ekembo and any of the three
Nevertheless while we ultimately agree with Harrison andAndrews (2009 see also Begun 2007) that the features dis-tinguishing ldquoUrdquo major from P africanus are consistent with thosethat distinguish congeneric species and are at least partly due todifferences in size we concede the possibility that the largestProconsul species could belong in a separate genus (Senut et al2000 Pickford and Kunimatsu 2005) However whether or notone recognizes a separate genus for P major the P africanus ma-terial does not sort morphologically with the Kisingiri materialTherefore the taxonomic alternatives are either two generaProconsul and Ekembo or three Proconsul Ugandapithecus andEkembo with Proconsul and Ugandapithecus united in a cladeexclusive of Ekembo Thus while we strongly favor the firstalternative recognizing Ugandapithecus does not change thefundamental argument concerning the need to refer the Kisingirispecies to a generically and cladistically distinct taxon from thespecies represented at Tinderet and the Ugandan sites
Specimens of interest
Despite the substantial dental and gnathic differences betweenProconsul and Ekembo there are a few specimens whose taxonomicaffinities require additional scrutiny
KNM-RU 1676-77 This set of associated upper and lower teeth is aconsistent outlier among E heseloni and E nyanzae specimens Insome ways this specimen more closely resembles Tinderet Pro-consul in the features that distinguish it from Ekembo Molar cuspmorphology and cingulum development I1 lingual tubercledevelopment and M2 shape and posthypocone crista orientationare more consistent with specimens from the Tinderet sites On theother hand it otherwise resembles Ekembo and lacks the strongestdiagnostic features that unite species of Proconsul such as thedistinctive canine and P3 morphology Overall this specimen doesnot fit unambiguously into either genus in fact the degree ofbuccolingual flare cusp morphology enamel wrinkling and espe-cially the long extension of enamel onto the anterior root in P3 arein some ways reminiscent of Afropithecus although it clearly differsfrom that genus as well (cf Rossie and MacLatchy 2013)
Were these characteristics individually distributed among thelarge Kisingiri sample one might attribute this to intra-genericvariation but when found together in a single specimen from alarge sample it suggests a unique individual One of us (KPM)thinks it likely that the specimen was not actually found at aKisingiri locality based on its unique preservation compared toother Rusinga fossils and on other examples of mis-proveniencedspecimens (Leakey 1967 Andrews and Molleson 1979 Pickford1986 Kelley 1986) However we recognize that relying toofrequently on this explanation allows almost any scenario to bejustified and suggest some alternative possibilities One is that thespecimen was found on Rusinga but in the much older WayandoFormation strata instead of the Hiwegi Formation The differencesin morphology in that case might reflect more than a million yearsof temporal change (McCollum et al 2013) with KNM-RU 1676-77preserving certain more primitive Proconsul-like features As analternative it is reasonable to assume that fossil assemblages asvast as those from Rusingawill include not only abundant taxa butalso rare or infrequent inhabitants of the local biome There aremany examples of this from Rusingas mammalian communityeg ldquoHemicyonrdquo Kulutherium Tadarida cf Hyainailouros cfArchaeobeledon Kelba (see relevant chapters in Werdelin andSanders 2010) As such KNM-RU 1676-77 may simply representa rare and well-preserved specimen of some other catarrhinetaxon that lived among or visited a primate community dominatedby Ekembo
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 59
KNM-SO 1112 This specimen was initially referred to Rangwapi-thecus but soon after was transferred to Proconsul afrianus (Bosler1981 Martin 1981)da designation that most subsequent authorshave followed (Pickford 1986 Nengo and Rae 1992 Kelley 19861995 Walker et al 1993 MacLatchy and Rossie 2005 Hill et al2013) Pickford et al (2009) more recently referred this specimento ldquoUrdquo meswae citing two primary reasons First they reiteratedBoslers (1981) concern that its P3 differs from others attributedto P africanus However at that time the majority of what wasconsidered to be P africanus would have been from RusingaIsland In fact while noting its contrasting morphology Boslersuggested that differences between KNM-SO 1112 and other P3smight reasonably be attributed to sexual dimorphism notingthat this specimen provides ldquoa not implausible match for the Pafricanus holotype maxilla M 14084rdquo (1981152) Thatassessment is borne out in this study Using the much largersample of Ekembo P3s as a model variation can be seen toencompass specimens with well-developed mesial beaks andthose with more bilaterally compressed crowns and parallelsides The latter morphology is predominantly found amongmale specimens (KNM-RU 1674 1924 and KPS I and III) thussupporting Boslers (1981) suggestion that this variation isrelated to sexual dimorphism and hence that KNM-SO 1112 is amale P africanus
The second feature cited by Pickford et al (2009) in referringthis specimen to ldquoUrdquo meswae was posterior shallowing of themandibular corpus However because they redefined ldquoUgandapi-thecusrdquo to include all of the well-preserved Tinderet mandibles(Pickford et al 2009)dleaving Proconsul to be represented only byRusinga specimensdtheir only alternative within this scheme for aspecimen the size of SO 1112 was to refer it to ldquoUrdquomeswae It seemsodd though to assign this specimen to a species from a (perhapsmuch) older site when the Proconsul africanus holotype is ofappropriate size and similar age and as there are no other adultmandibles from either species with which to compare it In anycase mandibular shallowing is variable in both Ekembo and Pro-consul with some Kisingiri specimens exhibiting considerableshallowing (eg KNM-RU 2087) and some ldquoUgandapithecusrdquo spec-imens having very little (eg M 14086)
We therefore see no morphological grounds to assign KNM-SO1112 to P meswae but do find some morphological evidence toassign it as a male P africanus which occurs at Songhor
KNM-LG 452 This very complete mandible was originally recog-nized as a female of P major based on the size of the dentition androbusticity of the mandible (Martin 1981) Bosler (1981) howeverleft this specimen unassigned noting similarities to P major in P3-M1 size but having distinctly smaller M2e3 In this she likenedKNM-LG 452 to a few other specimens (notably M 14086) thatare smaller in their distal molars in a manner more consistentwith taxonomic rather than sex differences (Bosler 1981 see alsoPilbeam 1969) Kelley (1986 1995) also assigned this specimen asa female of P major based on canine size and morphology butnoted that the canine lacked the shallowly-sloping mesial ridgetypical of most female anthropoid canines This is also truehowever of two other canines assigned as female P major KO 9and SO 373 (Kelley 1986) and so might simply be an unusualcharacteristic of P major female lower canines In the end Kelley(1995) concluded that considering all of the criteria fordistinguishing male and female canines the evidence that KNM-LG 452 is female remains questionable
Pickford et al (2009) further developed the idea that two spe-cies are represented within the P major sample and made KNM-LG452 the holotype of a new species ldquoUrdquo legetetensis We concur thatthis specimen is unique among the well-preserved Tinderet
mandibles in some important features Whereas male and femalemandibles of P major have long internal symphyseal planesexpansive extra- and retromolar sulci and rami that cross theocclusal plane at distal M3 KNM-LG 452 has a shorter symphysealplane (though not to the extent seen in Ekembo) a moderateextramolar sulcus but no retromolar sulcus and a ramus thatcrosses the occlusal plane more mesially If KNM-SO 396 and SO404 represent males and females respectively of P major then themorphology of KNM-LG 452 appears distinct
At present however we do not think there is sufficient evidenceto differentiate this specimen from either P major or P meswaePickford et al (2009) argue that KNM-LG 452 cannot belong to thelatter species because of its larger size The size differences how-ever are well within the range of variation seen in P majorE heseloni and E nyanzae Moreover whereas the Legetet mandibleis larger than most of the P meswae hypodigm an adult P4 referredto this species (KNM-ME 25) substantially exceeds its counterpartin KNM-LG 452 Should the Legetetmandible ultimately prove to bea male specimen size alone would not rule out its attribution toP meswae On the other hand if it is female then it becomes harderto justify its inclusion in that sample of mostly smaller specimens Iffemale therefore it could represent a separate species for whichthe binomial Proconsul legetetensis comb nov is available Recentlycollected fossils from the SonghorKapurtay area provide someadditional support for the possibility that three species of Proconsulexisted at these localities (McNulty 2014ab) However toconvincingly demonstrate this and determine whether a thirdspecies can be differentiated from P meswae will require a moredetailed analysis of variation among Proconsul species a corre-sponding study of variation in modern taxa anddalmost certain-lydadditional fossil specimens For now we regard ldquoUrdquo legetetensisas a subjective junior synonym of P major
Conclusions
The genus Proconsul has been central to studies of ape and hu-man evolution since its first description by Hopwood (1933) As oneof the best known fossil catarrhines represented by nearly everyskeletal element our knowledge of Proconsuls functional anatomypositional behavior and overall morphology has made it thefoundation for paleoanthropological research in the early Mioceneas well as studies on the origin and early diversification of homi-noids With respect to systematics however Proconsul is less wellunderstood the type species P africanus is known from a rela-tively small number of specimens from sites in the Tinderet areaand no postcranial bones can be unambiguously assigned to itRather it is the copious and well-preserved specimens from theKisingiri sites of Rusinga and Mfangano Islands that have formedthe basis for the common image of Proconsul
We argue here that the Kisingiri species traditionally referredto ProconsuldP heseloni and P nyanzaedare generically distinctfrom P africanus and hereby transfer them into a new genusEkembo based on a substantial number of morphological differ-ences (Table 2) Considering only those traits preserved in theP africanus holotype the entire sample from Rusinga and Mfan-gano differs markedly in the morphology of the upper canine P3P4 and M2 Additional differences in the I1 lower incisors andcanines molar cusp and occlusal crest morphology degree of flarein all molars and especially mandibular features clearly distin-guish the Kisingiri from non-Kisingiri samples in a way that isincompatible with congeneric species With the transfer ofE heseloni and E nyanzae Proconsul becomes a well-defined cladecomprising at least three speciesdP africanus P major andP meswaedunited by several features that are unique for thegenus (Table 2)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6160
We found no support in this study for the genus ldquoUgandapi-thecusrdquo as constructed by Pickford et al (2009) In that paper theytransferred the vast majority of Proconsul fossils from the Tinderetand Napak localities into ldquoUgandapithecusrdquo leaving P africanusrepresented only by its holotype and a small number of isolatedteeth and aligning it with the material from Rusinga and Mfan-ganoWhen the P africanus hypodigm is reconstituted and properlyaffiliated with the other Tinderet species according to itsmorphology much of the evidence marshaled by Pickford et al(2009) for ldquoUgandapithecusrdquo actually supports the distinction be-tween the Tinderet and Kisingiri samples ie the generic distinc-tion of Ekembo In addition some of the traits enumerated byPickford and colleagues in support of ldquoUgandapithecusrdquo wererevealed in our analysis to be variable within one or more speciesand therefore of questionable taxonomic utility We do not rejectout of hand the original conception of ldquoUgandapithecusrdquo (Senutet al 2000) as either a monotypic genus distinguishing ldquoUrdquo ma-jor or perhaps including a closely allied middle Miocene ldquoUrdquogitongai as well (Pickford and Kunimatsu 2005) but we have fornow retained the early Miocene specimens of both in Proconsulpending further study However even if Ugandapithecus is a taxo-nomically valid genus comprising just the latter two species theirmorphology indicates that theywould still be united in a cladewithProconsul and they lack several more advanced features that partlydefine Ekembo Better understanding of the role that body size playsinmediating differences within Proconsulwill help to further clarifythe relationships among these species
Despite the existence of one well-represented and several lesscomplete skeletons of what was long assumed to be Proconsul thephylogenetic status of this genus has defied consensus This islargely due in our view to Proconsul having been an admixture oftwo distinct genera one more advanced in the direction of extantapes than the other As we demonstrate here the material fromRusinga and Mfangano traditionally referred to Proconsul andwhich includes all of the more complete skeletal material isdistinct from the Tinderet and Napak material and belongs in aseparate genus Ekembo By disentangling these distinct clades weanticipate being able to achieve better phylogenetic resolution forboth genera thereby improving our knowledge of early hominoidevolution and diversification
Acknowledgments
We thank many people and agencies that assisted with thedevelopment and completion of this project This research wasinitiated as a result of funding from the Wenner-Gren Foundationand the Johns Hopkins School of Medicine to DR Begun in1987e1989 and was renewed following research and travelgenerously funded by multiple Leakey Foundation grants to KMcNulty National Science Foundation grants to K McNulty(0852609 1241807) the McKnight Foundation the University ofMinnesota the Lloyd A Wilford Endowment the LeverhulmeFoundation and the Turkana Basin Institute J Kelley gratefullyacknowledges support from the Institute of Human Origins Ari-zona State University DR Begun also wishes to acknowledge theAlexander Humboldt Stiftung and NSERC for funding support Weare grateful to the National Museums of Kenya the UgandaMuseum the Natural History Museum and to Rose Mwanja EzraMusiime Tom Mukhuyu Samuel Muteti James Yatich and MaryMuungu for access to and assistance with specimens in their careMany people provided assistance comments criticisms insightsand suggestions on various aspects of this work and we especiallythank Holly Dunsworth Will Harcourt-Smith Tom Lehmann DanPeppe Nicole Garrett Kirsten Jenkins Rutger Jansma Susy CoteIsaiah Nengo Andrew Hill and Brenda Benefit K McNulty
acknowledges the evolutionary anthropology faculty and studentsat Durham University for their helpful comments on an earlyversion of this manuscript and for their gracious hospitality Weare likewise grateful to the anonymous reviewers associate editorand editor whose comments strengthened the paper This manu-script is publication 1 supporting Research on East AfricanCatarrhine and Hominoid Evolution (REACHE)
Appendix A Supplementary data
Supplementary data related to this article can be found at httpdxdoiorg101016jjhevol201503009
References
Andrews P 1974 New species of Dryopithecus from Kenya Nature 249 188e190Andrews P 1978 A revision of the Miocene Hominoidea of East Africa Bull Brit
Mus (Nat Hist) Geol 30 85e225Andrews P 1985 Family group systematics and evolution among catarrhine pri-
mates In Delson E (Ed) Ancestors The Hard Evidence Liss New Yorkpp 14e22
Andrews P Harrison T Martin L Pickford M 1981 Hominoid primates from anew Miocene locality named Meswa Bridge in Kenya J Hum Evol 10123e128
Andrews P Begun D Zylstra M 1997 Interrelationships between functionalmorphology and paleoenvironments in Miocene hominoids In Begun DWard C Rose M (Eds) Function Phylogeny and Fossils Miocene HominoidEvolution and Adaptations Plenum Press New York pp 29e58
Andrews P Molleson T 1979 The provenance of Sivapithecus africanus Bull BritMus (Nat Hist) Geol 32 19e23
Begun DR 2001 African and Eurasian Miocene hominoids and the origins of theHominidae In de Bonis L Koufos GD Andrews P (Eds) Hominoid Evolutionand Climatic Change in Europe Phylogeny of the Neogene Hominoid Primatesof Eurasia Cambridge University Press London pp 231e253
Begun DR 2007 Fossil record of Miocene hominoids In Henke W Tattersall I(Eds) Handbook of Paleoanthropology Springer Berlin pp 921e977
Begun DR Teaford MF Walker AC 1994 Comparative and functional anatomyof Proconsul phalanges from the Kaswanga primate site Rusinga Island KenyaJ Hum Evol 26 89e165
Bishop WW 1964 More fossil primates and other Miocene mammals from North-East Uganda Nature 203 1327e1331
Bishop WW Miller JA Fitch FJ 1969 New potassium-argon age determinationsrelevant to the Miocene fossil mammal sequence in East Africa Am J Sci 267669e699
Blankenship JA Mincer HH Anderson KM Woods MA Burton EL 2007Third molar development in the estimation of chronologic age in Americanblacks as compared with whites J Forensic Sci 52 428e433
Bosler W 1981 Species groupings of early Miocene dryopithecine teeth from EastAfrica J Hum Evol 10 151e158
Coleman MN 2008 What does geometric mean mean geometrically Assessingthe utility of geometric mean and other size variables in studies of skullallometry Am J Phys Anthropol 135 404e415
Cote S Malit N Nengo I 2013 Additional mandibles of Rangwapithecus gordonian early Miocene catarrhine from the Tinderet localities of Western KenyaJ Hum Evol 153 341e352
Deane AS 2009 Early Miocene catarrhine dietary behavior the influence of theRed Queen Effect on incisor shape and curvature J Hum Evol 56 275e285
Drake R Van Couvering JA Pickford M Curtis G Harris JA 1988 New chro-nology for the Early Miocene mammalian faunas of Kisingiri Western KenyaJ Geol Soc Lond 145 479e491
Evernden JF Savage DE Curtis GH James GT 1964 Potassium-argon datesand the Cenozoic mammalian chronology of North America Am J Sci 262145e198
Gebo DL MacLatchy L Kityo R Deino A Kingston J Pilbeam D 1997A hominoid genus from the Early Miocene of Uganda Science 276 401e404
Geoffroy Saint-Hilaire E 1812 Tableau des quadrumanes 1 Ord QuadrumanesAnnls Mus Hist Nat Paris 19 85e122
Gray JE 1825 Outline of an attempt at the disposition of the Mammalia into tribesand families with a list of the genera apparently appertaining to each tribeAnnls Philos 10 337e344
Harrison T 1982 Small-bodied apes from the Miocene of East Africa PhDDissertation University of London
Harrison T 1986 New fossil anthropoids from the middle Miocene of East Africaand their bearing on the origin of the Oreopithecidae Am J Phys Anthropol 71265e284
Harrison T 1987 The phylogenetic relationships of the early catarrhine primates areview of the current evidence J Hum Evol 16 41e80
Harrison T 2002 Late Oligocene to middle Miocene catarrhines from Afro-ArabiaIn Hartwig WC (Ed) The Primate Fossil Record Cambridge University PressCambridge pp 311e338
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 61
Harrison T 2010 Dendropithecoidea Proconsuloidea and Hominoidea (CatarrhiniPrimates) In Werdelin L Sanders WJ (Eds) Cenozoic Mammals of AfricaUniversity of California Press Berkeley pp 429e469
Harrison T Andrews P 2009 The anatomy and systematic position of a newspecies of early Miocene proconsulid from Meswa Bridge Kenya J Hum Evol56 479e496
Hill A Nengo IO Rossie JB 2013 A Rangwapithecus gordoni mandible from theearly Miocene site of Songhor Kenya J Hum Evol 65 490e500
Hopwood AT 1933 Miocene primates from Kenya J Linn Soc Zool 38 437e464Ishida H Kunimatsu Y Takano T Nakano Y Nakatsukasa M 2004 Nacholapi-
thecus skeleton from the Middle Miocene of Kenya J Hum Evol 46 69e103Jolicouer P 1963 The multivariate generalization of the allometry equation Bio-
metrics 19 497e499Jungers WL Falsetti AB Wall CE 1995 Shape relative size and size-
adjustments in morphometrics Am J Phys Anthropol 38 (S2) 137e161Kavanagh KD Evans AR Jernvall J 2007 Predicting evolutionary patterns of
mammalian teeth from development Nature 449 427e433Kelley J 1986 Species recognition and sexual dimorphism in Proconsul and
Rangwapithecus J Hum Evol 15 461e495Kelley J 1995 Sex determination in Miocene catarrhine primates Am J Phys
Anthropol 96 390e417Kelley J 1993 Taxonomic implications of sexual dimorphism in Lufengpithecus In
Kimbel WH Martin LB (Eds) Species Species Concepts and Primate Evo-lution Springer New York pp 429e458
Kent PE 1942 The country round the Kavirondo Gulf of Victoria Nyanza Geogr J100 22e31
Kordos L Begun DR 2001 A new cranium of Dryopithecus from RudabanyaHungary J Hum Evol 15 689e700
Le Gros Clark WE Leakey LSB 1950 Diagnoses of East African Miocene Homi-noidea Q J Geol Soc Lond 105 260e263
Le Gros Clark WE Leakey LSB 1951 The Miocene Hominoidea of East Africa InFossil Mammals of Africa No 1 British Museum (Natural History) London
Leakey LSB 1943 A Miocene anthropoid mandible from Rusinga Kenya Nature152 319e320
Leakey LSB 1963 East African fossil Hominoidea and the classification within thissuper-family In Washburn SL (Ed) Classification and Human EvolutionAldine Chicago pp 32e49
Leakey LSB 1967 Notes on the mammalian faunas from the Miocene and Pleis-tocene of East Africa In Bishop WW Clark JD (Eds) Background to Evolu-tion in Africa University of Chicago Press Chicago pp 7e29
Leakey RE Leakey MG 1986 A new Miocene hominoid from Kenya Nature 324143e146
Leakey RE Leakey MG Walker AC 1988 Morphology of Afropithecus turka-nensis from Kenya Am J Phys Anthropol 76 289e307
Leakey MG Ungar PS Walker A 1995 A new genus of large primate from theLate Oligocene of Lothidok Turkana District Kenya J Hum Evol 28519e531
Linnaeus C 1758 Systema naturae per regna tria naturae secundum classesordines genera species cum characteribus differentris synonymis locis tenthed Laurentii Salvii Stockholm
MacInnes DG 1943 Notes on the East African Miocene primates J E Afr UgandaNat Hist Soc 17 141e181
MacLatchy L 2004 The oldest ape Evol Anthropol 13 90e103MacLatchy L Bossert WH 1996 An analysis of the articular surface distribution of
the femoral head and acetabulum in anthropoids with implications for hipfunction in Miocene hominoids J Hum Evol 31 425e453
MacLatchy L Rossie JB 2005 The Napak hominoid still Proconsul major InLieberman DE Smith RJ Kelley J (Eds) Interpreting the Past Essays onHuman Primate and Mammal Evolution in Honor of David Pilbeam Brill Ac-ademic Publishers Boston pp 15e28
Martin L 1981 New specimens of Proconsul from Koru Kenya J Hum Evol 10139e150
McCollum MS Peppe DJ McNulty KP Dunsworth HM Harcourt-Smith WEH Andrews AL 2013 Magnetostratigraphy of the early MioceneHiwegi Formation (Rusinga Island Lake Victoria Kenya) Geological Society ofAmerica Abstracts with Programs 45 p 12
McNulty KP 2010 Apes and tricksters the evolution and diversification ofhumans closest relatives Evol Edu Outreach 3 322e332
McNulty KP Harcourt-Smith WEH Dunsworth HM 2007 New primate fossilsfrom Rusinga Island Kenya Am J Phys Anthropol 132 (S44) 170
McNulty KP MacLatchy L Peppe DJ Nengo I Manthi FK Miller ERStevens NJ Cote S Lehmann T Kingston J 2014a A regional approach toEast African early Miocene paleobiology J Vert Paleontol 185 SVP Programand Abstracts Book 2014
McNulty KP MacLatchy L Rossie JB Peppe DJ Deino AL Mbua ENManthi FK Nengo IO Stevens NJ Cote S Lehmann T Gutierrez M 2014b
Research on East African catarrhine and hominoid evolution results from thefirst year Am J Phys Anthropol 153 (S58) 182
Michel LA Peppe DJ Lutz JA Driese SG Dunsworth HM Harcourt-Smith WEH Horner W Lehmann T Nightengale S McNulty KP 2014Remnants of an ancient forest provide ecological context for Early Miocenefossil apes Nature Comm 5 3236
Myres JL 1932 Africa Man 32 208Nakatsukasa M Kunimatsu Y Nakano Y Takano T Ishida H 2007 Vertebral
morphology of Nacholapithecus kerioi based on KNM-BG 35250 J Hum Evol52 347e369
Nengo IO Rae TC 1992 New hominoid fossils from the early Miocene site ofSonghor Kenya J Hum Evol 23 423e429
Peppe DJ McNulty KP Cote SM Harcourt-Smith WEH Dunsworth HM VanCouvering JA 2009 Stratigraphic interpretation of the Kulu Formation (EarlyMiocene Rusinga Island Kenya) and its implications for primate evolutionJ Hum Evol 56 447e461
Pickford M 1981 Preliminary Miocene mammalian biostratigraphy for westernKenya J Hum Evol 10 73e97
Pickford M 1986 Sexual dimorphism in Proconsul Hum Evol 1 111e148Pickford M Andrews PJ 1981 The Tinderet Miocene sequence in Kenya J Hum
Evol 10 11e33Pickford M Kunimatsu Y 2005 Catarrhines from the Middle Miocene (ca 145
Ma) of Kipsaraman Tugen Hills Kenya Anthropol Sci 113 189e224Pickford M Senut B Gommery D Musiime E 2009 Distinctiveness of Ugan-
dapithecus from Proconsul Estudios Geol 65 183e241Pilbeam DR 1969 Tertiary Pongidae of East Africa evolutionary relationships and
taxonomy Bulletin of the Peabody Museum of Natural History 31 pp 1e185Rae TC 1993 Phylogenetic analysis of proconsulid facial morphology PhD
Dissertation State University of New York at Stony BrookRae TC 1999 Mosaic evolution in the origin of the Hominoidea Folia Primatol 70
125e135Rafferty KL Walker A Ruff C Rose MD Andrews PJ 1995 Postcranial esti-
mates of body weights in Proconsul with a note on a distal tibia of P major fromNapak Uganda Am J Phys Anthropol 97 391e402
Rossie JB MacLatchy L 2013 Dentognathic remains of an Afropithecus individualfrom Kalodirr Kenya J Hum Evol 65 199e208
Senut B Pickford M Gommery D Kunimatsu Y 2000 Un nouveau genredrsquohominoiumlde du Miocene inferieur drsquoAfrique orientale Ugandapithecus major(Le Gros Clark and Leakey 1950) CR Acad Sci Paris 331 227e233
Simons EL Pilbeam DR 1965 Preliminary revision of the Dryopithecinae (Pon-gidae Anthropoidea) Folia Primatol 3 81e152
Smith TM Martin LB Leakey MG 2003 Enamel thickness microstructure anddevelopment in Afropithecus turkanensis J Hum Evol 44 283e306
Stevens NJ Seiffert ER OConnor PM Roberts EM Schmitz MD Krause CGorscak E Ngasala S Hieronymus TL Temu J 2013 Palaeontological evi-dence for an Oligocene divergence between Old World monkeys and apesNature 497 611e614
Suwa G Kono RT Katoh S Asfaw B Beyene Y 2007 A new species of great apefrom the late Miocene epoch in Ethiopia Nature 448 921e924
Szalay FS Delson E 1979 Evolutionary History of the Primates Academic PressNew York
Teaford MF Beard KC Leakey RE Walker A 1988 New hominoid facial skel-eton from the Early Miocene of Rusinga Island Kenya and its bearing on therelationship between Proconsul nyanzae and Proconsul africanus J Hum Evol17 461e477
Van Couvering JAH Miller J 1969 Miocene stratigraphy and age determinationsRusinga Island Kenya Nature 221 628e632
Walker A 1997 Proconsul function and phylogeny In Begun DR Ward CVRose MD (Eds) Function Phylogeny and Fossils Miocene Hominoid Evolu-tion and Adaptation Plenum Press New York pp 209e224
Walker A Pickford M 1983 New postcranial fossils of Proconsul africanus andProconsul nyanzae In Ciochon RL Corruccini RS (Eds) New Interpretationsof Ape and Human Ancestry Plenum Press New York pp 325e351
Walker A Teaford M 1988 The Kaswanga primate site an Early Miocene hom-inoid site on Rusinga Island Kenya J Hum Evol 17 539e544
Walker A Teaford M 1989 The hunt for Proconsul Sci Am 260 76e82Walker A Teaford MF Martin L Andrews P 1993 A new species of Proconsul
from the early Miocene of RusingaMfangano Islands Kenya J Hum Evol 2543e56
Ward CV Ruff CB Walker A Rose MD Teaford MF Nengo IO 1995 Func-tional morphology of Proconsul patellas from Rusinga Island Kenya with im-plications for other Miocene-Pliocene catarrhines J Hum Evol 29 1e19
Ward S Brown B Hill A Kelley J Downs W 1999 Equatorius a new hominoidgenus from the Middle Miocene of Kenya Science 285 1382e1386
Werdelin L Sanders WJ 2010 Cenozoic Mammals of Africa University of Cali-fornia Press Berkeley

KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 43
Leakey 1943) the consensus view emerged of this genus as a basalhominoid (eg Andrews 1985) Around this time Harrison (19821987) raised the possibility that Proconsul and other East Africannon-cercopithecoid catarrhines from the early Miocene might bebetter placed among stem catarrhines phylogenetically precedingthe divergence of the hominoid and cercopithecoid lineagesWalkers (Walker and Pickford 1983 Walker and Teaford 1989Walker 1997 see also Rae 1993 1999) resurrection of the homi-nid (frac14 great apethorn human) status for Proconsulwas ultimately shortlived with most researchers maintaining Proconsul as a basalmember of the Hominoidea
Taxonomic history of Proconsul
Germane to the phylogenetic debate is the taxonomic frame-work within which these fossils are analyzed Proconsul (typespecies Proconsul africanus) was erected for fossil primate speci-mens discovered from 1926 to 1931 at Koru a Western Kenya lo-cality situated on the periphery of the extinct Tinderet volcano(Hopwood 1933 Le Gros Clark and Leakey 1951) In that firstpublication Hopwood (1933460) made oblique but favorablecomparison to recently discovered but unpublished specimensfrom Rusinga Island Yet LSB Leakeys initial presentation of theRusinga material characterized it as being quite different fromHopwoods Proconsul leading Sir Arthur Keith to liken it to theyounger hominoid Dryopithecus (see Myres 1932 Pickford 1986)Leakeys formal publication however included both Koru andRusinga specimens in a single genus which was thought torepresent the base of the human-ape clade (Leakey 1943)
At this time early Miocene primate localities in Western Kenyawere all geographically clustered around either the Kisingiri vol-cano (Rusinga Island Mfangano Island Karungu Uyoma) or theTinderet volcano (Koru Songhor Legetet Hill and others Fig 1)Continued collecting in both regions by the East African Archaeo-logical Expedition and British-Kenya Miocene Expedition graduallyrevealed substantial size variation in the type species Proconsulafricanus and both the Kisingiri and Tinderet localities were re-interpreted as having a smaller and a larger Proconsul species
Figure 1 Map depicting the major Proconsul localities Early Miocene localities clus-tering around the Tinderet volcano include Koru Legetet Hill Chamtwara MeswaBridge Songhor Kapurtay and Mteitei Valley Those associated with Kisingiri areRusinga Island Mfangano Island Karungu and the Uyoma peninsula The middleMiocene type locality of P gitongai is not shown here
This prompted Le Gros Clark and Leakey (1950) to erect two newspecies Proconsul major for the large specimens from Tinderet andProconsul nyanzae for the large Kisingiri specimens The smallerspecies P africanus continued to be recognized in both areas bymost workers Large-bodied catarrhines discovered in Ugandaexpanded the geographic range of P major (Bishop 1964) althoughsome of this sample was later reassigned to a new taxon Moroto-pithecus bishopi by Gebo et al (1997)
This basic taxonomic arrangement remained relatively stable forthree decades although individual specimen identifications did notalways adhere to these simple geographic distinctions (egAndrews 1978) Subsequent studies by Bosler (1981) Harrison(1982 1987) Kelley (1986) and others (Pickford 1986 Teafordet al 1988 Walker and Teaford 1988) further standardized thetaxonomic attributions of specimens and thereby reduced thenumber of Proconsul species recognized at each site P nyanzae and(for some) P africanus associated with Kisingiri localities P majorand P africanus associated with Tinderet localities Ugandan Pro-consul restricted to P major
Fieldwork on Rusinga in the 1980s generated sufficient newevidence to confirm that the smaller Kisingiri Proconsul specimensdiffer morphologically from P africanus (Kelley 1986) leadingWalker et al (1993) to assign this material to a new species Pro-consul heseloni The presence of two species of Proconsul on Rusingaand Mfangano was not universally accepted however Kelley forexample referred all Kisingiri specimens to P nyanzae voicingconcerns over the unlikely distribution of sexes (cf Kelley 1986)and the disregard of likely body size variation within the twospecies (Kelley 1993) Likewise Pickford (1986) recognized onlyP nyanzae from Rusinga and Mfangano notably highlighting thedistinct morphology of Kisingiri Proconsul when compared to Tin-deret and Ugandan specimens Despite these and other objectionsthe presence of P nyanzae and P heseloni at the Kisingiri localitieshas come to be universally accepted (Begun et al 1994 Leakeyet al 1995 Rafferty et al 1995 Ward et al 1995 MacLatchy andBossert 1996 Harrison 2002 2010 Smith et al 2003 Ishidaet al 2004 Nakatsukasa et al 2007 Deane 2009 Harrison andAndrews 2009 Peppe et al 2009 Pickford et al 2009 Michelet al 2014)
The naming of P heseloni after 30 years of relative taxonomicstability heralded a new period of systematic revision First Senutet al (2000) transferred P major into a new genus ldquoUgandapithe-cusrdquo a genus whose validity is debated within the paleoanthro-pological community (see Begun 2007 Harrison and Andrews2009 Harrison 2010) and which we consider to be a junior syn-onym of Proconsul (see below) Pickford and Kunimatsu (2005)subsequently allocated a few isolated teeth from Kipsaraman tothis genus under the species name ldquoUgandapithecusrdquo gitongai Inaddition the proconsulid material from Meswa Bridgedlongrecognized as a distinct species (Andrews et al 1981)dwas giventhe name Proconsul meswae by Harrison and Andrews (2009) Inthat same year however Pickford et al (2009) published acomprehensive taxonomic revision of Proconsul (including ldquoUgan-dapithecusrdquo) based on cranial mandibular and dental material Indoing so they identified a number of similarities between ldquoUgan-dapithecusrdquo major and the smaller Tinderet Proconsul specimensparticularly with regard to mandibular morphology In thatmanner specimens that have been historically referred toP africanus plus the smallest specimens referred to P major (cfPilbeam 1969 Bosler 1981) were transferred to a new speciesldquoUgandapithecusrdquo legetetensis (Pickford et al 2009) Likewise themandible KNM-SO 1112dregarded as P africanus by otherworkersdwas referred to P meswae and the latter species trans-ferred to ldquoUgandapithecusrdquo (Pickford et al 2009) As a result of thissystematic revision P africanus was left almost entirely without
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6144
mandibular representation whereas ldquoUrdquo legetetensis was repre-sented mostly by mandibles and lower teeth (Pickford et al 2009)
Table 1 summarizes the relevant taxonomic allocations used bydifferent authors according to the geographic distribution of lo-calities In its broadest conception (eg Harrison and Andrews2009) the genus Proconsul is quite speciose relative to otherMiocene hominoid genera including as many as six distinct spe-cies This number is substantially reduced under the taxonomicscheme proposed by Pickford et al (2009) wherein half of thesespecies are referred to ldquoUgandapithecusrdquo Those preferring tomaintain all of this material in Proconsul however have noted thatthe greatest distinction within that sample is between the Kisingiriassemblages on the one hand and those from Tinderet and Ugandaon the other (Walker et al 1993 Begun 2001 2007MacLatchy andRossie 2005 Harrison and Andrews 2009 Harrison 2010)daninterpretation presaged in some ways by the taxonomy proposedby Kelley (1986) and Pickford (1986) and ultimately echoing theoriginal presentation by LSB Leakey (Myres 1932)
In accordancewith other researchers (eg Harrison 2002 2010MacLatchy 2004 MacLatchy and Rossie 2005 Suwa et al 2007Harrison and Andrews 2009 McNulty 2010 Hill et al 2013) wedo not recognize the generic distinction of ldquoUgandapithecusrdquo fromProconsul This is due in part to our findings detailed below of adistinct clade uniting P africanus with species assigned to ldquoUgan-dapithecusrdquo In addition the most recent characterization of
Table 1Identifications of Proconsul (sensu lato) species by different authors listed byregiona
Authors KisingiriLocalities
TinderetLocalities
UgandanLocalities
Hopwood 1933 e P africanusb e
MacInnes 1943 P africanus P africanus e
Le Gros Clark andLeakey 1950
P africanus P africanus e
P nyanzaeb P majorb
Andrews 1978 P africanus P africanus P majorP nyanzae P majorP major
Bosler 1981 P africanus P africanus P majorP nyanzae P nyanzaeP major P major
Kelley 1986 P nyanzae P africanus P majorP major
Pickford 1986 P nyanzae P africanus P majorP major
Walker et al 1993 P heselonib P africanus P majorP nyanzae P major
Senut et al 2000 P nyanzae P africanus U majorU majorc
Harrison andAndrews 2009d
P heseloni P africanus P majorP nyanzae P major
P meswaeb
Pickford et al 2009 P heseloni P africanus U majorP nyanzae U major U legetetensis
U legetetensisb U gitongaiU meswaec
This paperd E heselonic P africanus P majorE nyanzaec P major
P meswae
a Very small samples from other regions (eg Buluk) that have been attributed toProconsul (sensu lato) are not included here as they are not critical to understandingthe placement of Kisingiri specimens
b New genus andor species named in publicationc Existing species transferred to a new genusd Proconsul gitongai from the middle Miocene Muruyur Formation (Kipsaraman)
is also recognized but not at the early Miocene localities assessed in this paper
ldquoUgandapithecusrdquo which removed most Tinderet specimens fromProconsul and distributed them among new and existing species ofldquoUgandapithecusrdquo was based largely on contrasts with Rusingaspecimens rather than with the type species of Proconsul (Pickfordet al 2009) This is particularly true for mandibles and the lowerdentitiondspecimens less securely tied to P africanus because thetype specimen M 14084 is a partial maxilla (eg Bosler 1981) Weargue here that if specimens are sorted without preconceived taxo-nomic assumptions the purported diagnostic features of ldquoUganda-pithecusrdquo (sensu Pickford et al 2009) serve instead to differentiateall specimens in the Tinderet and Uganda samples of Proconsul(Proconsul sensu stricto) from those found at Kisingiri localities (cfKelley 1986 Walker et al 1993 Begun 2001 2007 MacLatchy andRossie 2005 Harrison and Andrews 2009 Harrison 2010)
Geochronology of Proconsul localities
The chronological framework of Proconsul localities provides animportant context for understanding historical and current in-terpretations of species distributions and relationships Early re-searchers while acknowledging the limitations of theirstratigraphic and faunal correlations were nevertheless able tocorrectly position the Western Kenyan localities within the lowerMiocene associating some of the deposits with the Burdigalian astage defined by marine sediments in Europe and dated betweenabout 16 and 205 Ma (Kent 1942 MacInnes 1943) The applicationof radiometric dating techniques helped refine these estimateswith some of the earliest KeAr studies conducted on East Africanlocalities associated with Proconsul (Evernden et al 1964 Bishopet al 1969 Van Couvering and Miller 1969) These datesconfirmed an early Miocene age for most localities with estimatesranging between 23 and 16 Ma However the Kisingiri depositsyielded discrepant results due to post-depositional alteration of thedatable biotites (Drake et al 1988) Re-sampling and further ana-lyses yielded more satisfactory and repeatable age estimates of179e178 Ma but notably these were based on only five samplesconfined to a small segment of the Rusinga geological section(Drake et al 1988)
Radiometric dates in conjunction with studies of the associatedfaunal communities (eg Pickford 1981) created a geochronolog-ical framework for Proconsul wherein the Tinderet localities areoldest dating to perhaps 225 Ma at Meswa Bridge (see review inHarrison and Andrews 2009) with Napak in Uganda beingapproximately contemporaneous with Songhor and Koru (Bishopet al 1969 Pickford and Andrews 1981) and the Kisingiri local-ities preserving the youngest sediments (Pickford1981 Drake et al1988) However themost recent re-dating of Kisingiri sites presentsa more complex scenario Rather than being younger than all of theTinderet sites the Kisingiri stratigraphic sequence is substantiallylonger than previously thought with the oldest fossil stratacontemporaneous with those from Koru and Legetet Hill atapproximately 20e19 Ma (McCollum et al 2013) The youngestMiocene fauna from Rusinga is not associated with radiometricdates butmagnetostratigraphic analysis suggests an age near 17Maor younger (Peppe et al 2009) Hence differences between speciesof Proconsul and in particular between assemblages from TinderetandKisingiri cannot be simply explained bychronology Indeed thegreat length of the Kisingiri sequence suggests that taxonomicuniformity throughout this timeperiodmight require re-evaluation
Aims of this study
Motivated by new specimens analyses and interpretations ofProconsul thathave appeared in the last severalyearsweundertookathorough review of the craniodental and mandibular specimens
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 45
attributed to this genus and here provide an alternate taxonomicinterpretation to those in current use Our primary objective was toevaluate the morphological homogeneity of the genus Proconsul astraditionally constituted (Proconsul sensu lato) and explore thepossibility that theKisingiri specimens formadistinct group (Walkeret al1993 Begun 2001 2007MacLatchyandRossie 2005Harrisonand Andrews 2009) While we agree with Pickford et al (2009) thatthe material traditionally referred to Proconsul exceeds what can becomfortably accommodated within a single genus our resultsconform to the suggestion proposed by Begun (2001 2007) and laterby MacLatchy and Rossie (2005) and Harrison and Andrews (2009)that the Kisingiri specimens from Rusinga and Mfangano Islandsconstitute a distinct cladewith respect to Proconsul from theTinderetand Ugandan localities Our assessment of the taxonomic andphyletic distinction of the Kisingiri material necessitates genericdifferentiation of this sample from Proconsul
Systematics
Order Primates Linnaeus 1758Infraorder Catarrhini E Geoffroy Saint-Hilaire 1812Superfamily Hominoidea Gray 1825Family incertae sedisEkembo gen novurn lsidzoobankorgactB11CECF0-9DD1-4265-9C06-A60E4BEE974E
Synonymy
1950 Proconsul Hopwood Le Gros Clark and Leakey 2601965 Dryopithecus (Proconsul) (Hopwood) Simons and Pilbeam
1201978 Proconsul Hopwood Andrews 901986 Proconsul Hopwood Kelley 4791993 Proconsul Hopwood Walker et al 512010 Proconsul Hopwood Harrison 437
Type species
Ekembo nyanzae comb nov (Le Gros Clark and Leakey 1950)
Included species
Ekembo nyanzae (Le Gros Clark and Leakey 1950) Ekemboheseloni comb nov (Walker et al 1993)
Etymology
Ekembo means ldquoaperdquo or ldquomonkeyrdquo in the Suba language whichwas historically spoken by peoples in western Kenya includingthose who settled Rusinga and Mfangano Islands (Jack Wanyendepers comm)
Diagnosis
Medium- to large-bodied and sexually dimorphic hominoidcharacterized by the following combination of features Frontalbone moderately wide at the coronal suture narrowing anteri-orly toward gracile supraorbital rims which are not demarcatedby a post-toral sulcus Large frontal sinuses behind a prominentglabella Minor postorbital constriction Temporal lines promi-nent and situated high on frontal and parietal bones convergingposteriorly without forming a sagittal crest (in female
specimens) Nasal bones long and thin Broad interorbital regionflat rather than projecting Upper face more vertically orientedrelative to the mid- and lower face Nasolacrimal duct positionedwithin a broad orbit Rostrum broad with premaxilla extendingup to the nasal bones Canine jugum prominent in both sexes butcanine fossa developed only in males Nasal aperture broad withwidest portion inferior and sometimes near the base Inferioraperture margin concave or incised in the midline Nasoalveolarclivus intermediate in height between hylobatids and hominidsClivus does not overlap palatine process of the maxilla pre-senting typical mammalian incisive fenestration ante-roposteriorly positioned at or posterior to mid-canine Maxillarysinus extensive reaching anteriorly to P3P4 junction Zygomati-coalveolar crest arises low on the face near M2 Nearly verticalmalar region Broad anterior palate A single palatine foramenpositioned near the M2M3 junction Broad mandibular ramusangled slightly posteriorly from vertical leads to a graduallyrounded and gracile gonial region Mandibular corpora rangefrom short and gracile to taller and moderately robust Sym-physis characterized by moderately developed superior trans-verse torus and variably developed inferior transverse torus andgenioglossal pit Symphyseal planum and subplanum regionvertically or nearly vertically oriented A single mental foramenpositioned near the P3P4 junction approximately halfway up thecorpus or higher
Upper central and lateral incisors heteromorphic Centralincisor characterized by a narrow lingual tubercle often consist-ing of two or three narrow ridges and distinct but thin marginalridges that are continuous with the lingual cingulum with themesial marginal ridge joining the cingulum at a sharp angle incontrast to a more continuously curving transition from cingulumto distal marginal ridge resulting in an asymmetric ridge andcingulum morphology Lateral incisors have well-developedincisal edges and are asymmetric with a moderately developedlingual cingulum Narrow lower incisors with central and lateralincisors having similar morphology Upper and lower caninesstrongly sexually dimorphic with only slight bilateral compres-sion and moderately low-crowned in relation to basal crown di-mensions with apices terminating at a simple point Canine rootssmooth and conical Cheek tooth cusps low and broad and con-nected by inflated intercuspal crests Upper premolars morpho-logically similar with well-developed mesial foveae onlymoderate cusp heteromorphy and cusps positioned at theapproximate mesiodistal midpoint of the crown resulting innearly symmetric buccal profiles Molars with moderate bucco-lingual flare and reduced cingulum development Upper first andsecond molars approximately square with nearly straight distalmargins and buccally oriented posthypocone cristae Lower firstand second molars rectangular with M1 substantially narrowerthan M2 Upper and lower third molars vary in degree of distalcusp development
Forelimbs and hind limbs of similar length Long vertebral col-umn with six lumbar vertebrae characterized by transverse pro-cesses that arise from the dorsal margin of the vertebral centrumand prominent accessory processes for erector spinae musclesPossessing a coccyx rather than a tail Thorax long narrow anddorsoventrally deep Broad flat sternebrae Pelvis long and narrowwith sagittally oriented ilia an elongate ischium lacking osseousevidence for the presence of callosities and a narrow sacrumwith asmall sacro-iliac joint Scapula positioned laterally on thorax with anarrow ovoid and ventrally oriented glenoid fossa Humerus witha posteriorly directed head retroflexed shaft with little torsion anda distal articular surface lacking a well-differentiated trochlea andcapitulum and having a narrow zona conoidea and shallowtrochlear notch but lacking an entepicondylar foramen Radius
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6146
with a small ovoid head and flat distal articular surface that mainlycontacts the scaphoid Ulna displays a large olecranon processnarrow semilunar notch with only a slight keel anteriorly posi-tioned radial notch and a long distal styloid process that retainsarticulation with triquetrum and pisiform Carpals relatively nar-row and include an unfused os centrale a hamate with a smallhamulus and a flat medially oriented triquetral surface Handcharacterized by straight metacarpals with narrow bases andheads Pollex well developed Manual and pedal phalanges gener-ally similar to each other but less so than in Old World monkeysFemur slender with a small head long neck with a high neck-shaftangle centrally placed fovea capitis reduced greater trochanterand a broad distal end with the medial condyle slightly larger thanthe lateral Broad flat patella Tibia long and slender but fibularobust Tarsals and metatarsals elongate with a robust hallux
Differential diagnosis
Ekembo can be readily distinguished from all other medium-and large-bodied catarrhine genera from the early Miocene Cra-nial mandibular and dental differences between Ekembo andProconsul are described in detail below and summarized inTable 2 Ekembo also differs from Proconsul in having a rounderradial head and a femur with a shorter greater trochanter poste-riorly projecting lesser trochanter less extensive inter-trochantericcrest more robust phalanges with more strongly developedfibrous flexor sheath ridges and broader proximal and interme-diate phalangeal distal joint surfaces (condyles) with widertrochlear grooves and less sharply defined trochlear ridges(Harrison 1982 Andrews et al 1997 Senut et al 2000 Pickfordet al 2009)
Ekembo differs from Afropithecus in having a more inflatedglabellar region no frontal trigon less strongly developed anteriortemporal ridges less postorbital constriction a stronger mandib-ular superior transverse torus less posterior shallowing of themandibular corpus higher-crowned lower canines in relation tomesiodistal length buccolingually narrower upper premolarsrelative to molars upper premolars with cusps positioned closer tothe mesiodistal midline reduced P3 cusp heteromorphy a moresymmetric I2 and substantially less buccolingual flare in the cheekteeth (Leakey et al 1988 Rossie and MacLatchy 2013)
Ekembo differs from Heliopithecus in having buccolinguallynarrower premolars relative to molars premolar cusps positionedcloser to the mesiodistal midline and reduced P3 cusp hetero-morphy (Harrison 2010)
Ekembo differs from Equatorius in having a more gracilemandible a larger superior but smaller inferior mandibular trans-verse torus labiolingually narrower lower incisors with straightermesial and distal margins higher-crowned lower canines in rela-tion to mesiodistal length greater molar cingulum development aless posteriorly directed humeral medial epicondyle a more ante-riorly directed radioulnar joint and narrower metacarpal heads(Ward et al 1999)
Ekembo differs from Nacholapithecus in having a shorter sub-nasal clivus a more expansive incisive foramen a broader nasalaperture at the base a minimally developed mandibular inferiortransverse torus a narrower lingual tubercle on I1 higher-crownedlower canines in relation to mesiodistal length a larger humeralradial fossa a shallower coronoid fossa a less well-developed zonaconoidea a narrower and distally slanted ulnar coronoid process alonger and lower femoral neck a more posteriorly positioned lessertrochanter and a smaller posteromedial tubercle on the talus(Ishida et al 2004)
Ekembo differs from Nyanzapithecus in having more anteriorlyextended maxillary sinuses a relatively taller mandibular corpus
less buccolingually compressed upper and lower canines with lesscurved crowns lower and more rounded cusps with more inflatedocclusal crests on the cheek teeth P3 and P4 that more closelyresemble each other and have much less cingular development abroader P4 relative to M1 M1e2 occlusal outline more closelyapproximating a square and lower molars that are broader relativeto mesiodistal length with a more mesially placed entoconid rela-tive to the hypoconid (Harrison 1986 2010)
Ekembo differs from Rangwapithecus in having a broad andexpansive palatal fenestration a wider palate relative to length amore robust mandible with a well-defined superior transversetorus broader upper and lower incisors relative to height lessbuccolingually compressed canines without scimitar-shapedcrowns upper premolars with reduced cingulum and cusps posi-tioned near the mesiodistal midpoint resulting in expanded mesialfoveae and symmetric buccal profiles a broader P4 relative to M1upper and lower molars that are buccolingually broader relative tolength and have more rounded cusps and inflated occlusal crestsmore squared upper molars with substantially less cingulumdevelopment and lower molars with a more mesially placedentoconid (Andrews 1974 Harrison 2002 2010 Cote et al 2013Hill et al 2013)
Ekembo differs from Turkanapithecus in having an expandedglabellar region less postorbital constriction a narrower temporalfossa with less flaring zygomatic arches the lacrimal fossa withinthe orbital margin a relatively broader palate with non-convergingtooth rows narrower nasal bones and interorbital region a distinctmandibular superior transverse torus a more robust mandibularcorpus taller mandibular ramus upper canines more rounded incross-section a reduced cingulum on cheek teeth awell-developedhypocone on M1e2 a more squared M2 with a protocone subequalto paracone and broader lower molars relative to length (Leakeyand Leakey 1986 Harrison 2002 2010)
Ekembo differs from Mabokopithecus in having a relativelybroader M3 crown without distal recurvature of the crown andlacking a buccal concavity a hypoconulid that is in-line with thebuccal cusps more bunodont cusps a well-defined protocristidand a broad open talonid without an accessory cuspid (Harrison2002 2010)
Ekembo differs from Xenopithecus in having reduced uppermolar flare a substantially reduced cingulum no prehypoconecrista molar occlusal profiles more closely approximating a squareand less constricted trigons (Hopwood 1933 Le Gros Clark andLeakey 1951 Harrison 2002 2010)
Ekembo differs from Morotopithecus in having a broader inter-orbital region a better developed canine fossa in males premolarssmaller relative to molars greatly reduced or absent buccalcingula on upper molars lumbar vertebrae with transverse pro-cesses positioned more ventrally on the centrum and a narrowerdistal femur with a deeper patellar groove and similarly-sizedmedial and lateral femoral condyles (Gebo et al 1997MacLatchy 2004)
Recently Stevens et al (2013) likened the new genus Rukwapi-thecus represented by a mandible fragment with well-preservedP4-M3 to nyanzapithecine species Ekembo can be distinguishedfrom this genus in having a more gradually rounded gonial angle aless obliquely oriented P4 with subequal protoconid and metaconidand a broader mesial fovea less mesially positioned metaconidreduced buccal cingula and smaller mesial foveae on the molarssmaller hypoconulids broader M1e2 relative to length a singularrather than twinned metaconid no accessory cuspules in thelingual notch and a well-defined crest between entoconid andhypoconulid
Family Proconsulidae (Leakey 1963)
TabDen
Fe
G
U
U
U
U
U
M
Lo
Lo
Lo
a
b
c
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 47
Genus Proconsul Hopwood 1933
Type species
Proconsul africanus Hopwood 1933
Included species
Proconsul africanus (Hopwood 1933) Proconsul major (Le GrosClark and Leakey 1950) Proconsul gitongai comb nov (Pickfordand Kunimatsu 2005) Proconsul meswae (Harrison and Andrews2009)
Diagnosis (replaces the emended diagnosis of Harrison 2010)
Proconsul species are medium- to large-bodied sexually dimor-phic catarrhines that can be distinguished from other proconsulidgenera by the following combination of features Robust mandib-ular corpus and symphysis characterized by a long shallowlysloping planum alveolare terminating at a large superior transversetorus and a subplanum surface oriented posteroinferiorly Upper
le 2tognathic features that distinguish Ekembo from Proconsula
ature Ekembo
eneralCusp morphology More bunodont occupying more of theMolar crests Inflated incorporated into cuspsCingulum Reducedpper central incisorLingual pillar Narrow generally low reliefMarginal ridges and cingulum Asymmetric with sharply angled cingulu
ridge transitionpper canineTip PointRoot fluting AbsentRoot striations Rare and weakly developedpper third premolarb
Mesiodistal cusp position Crown midpointMesial fovea Well developedBuccal profile SymmetricMesial crown margin Straight or slightly curvedDistal crown margin Slight to moderately curvedCusp heteromorphy ReducedCrown shape Converges on P4 shapeMesiobuccal flange Nopper fourth premolarParacone height Taller relative to crown lengthpper second molarc
Occlusal crown shape SquarishDistal margin Straight to slightly curvedBuccal cingulum Absent or rudimentaryDistolingual cingulum Merges into hypoconePosthypocone crista Distobuccally to buccally orientedBuccolingual flare ReducedandiblePlanum alveolare Short vertically inclinedSuperior transverse torus Moderate to well-developedOrientation of subplanum area More vertically orientedCorpus robusticity More gracilewer incisorsCrown shape Narrow relative to heightwer canineTip Pointwer molarsBuccolingual flare ReducedM1 M2 breadth M1 narrower relative to M2
Although presented here as discrete characteristics for descriptive purposes some oNearly all of these features tend to distinguish P4s as well but in that tooth the diffeThese features tend to distinguish M1s as well but in that tooth the differences are
incisors heteromorphic with I1 exhibiting an inflated lingual tu-bercle and a near symmetrical configuration of lingual marginalridges and cingulum with a curving transition from cingulum tothe mesial marginal ridge Lower incisors broad relative to crownheight Upper and lower canines with distinct blade-like or burin-like tips in both deciduous and adult teeth Upper canine rootswith broad longitudinal grooves or fluting defining two or threedistinct columns plus numerous small longitudinal striationsextending cervically from the root apex Upper P3s morphologicallydissimilar from P4s P3 has substantial cusp heteromorphy (para-cone much taller than protocone) with mesially positioned cuspsand a notched mesial border that together limit or eliminate amesial fovea A long steeply angled preparacrista contributes to ahighly asymmetric buccal profile before being interrupted near thetooth cervix by a mesially projecting flange The distal margin isstrongly curved P4 has a tall paracone relative to crown lengthMolars characterized by extensive cingular development individ-uated conical cusps connected by sharp occlusal crests and sub-stantial buccolingual flare Upper molars rhomboid in shaperesulting from a broad trigon and a distally expanded hypocone M2
has a distinct buccal shelf between paracone and metacone a dis-tolingual cingulum that wraps fully around the hypocone rather
Proconsul
occlusal surface area More distinct individuated conesSharper more distinct from cuspsExtensive
Inflatedbulbousm-mesial More symmetric with curving cingulum-mesial
ridge transition
Blade-likePresentVery common and well developed
MesialAbsent or reducedAsymmetricNotchedStrongly curvedStrongDistinct from P4
Yes
Shorter relative to crown length
RhomboidStrongly curvedDistinct cingular shelfDistinct from hypoconeDistally to distobuccally orientedPronounced
Long more horizontally orientedStrongly developedMore posteriorly inclinedMore robust
Broad relative to height
Blade-like
PronouncedM1 more similar to M2
f these features are undoubtedly correlated aspects of single evolutionary changesrences are not as pronouncednot as pronounced
Figure 2 Upper central incisors of Proconsul and Ekembo Lingual views of Ekembo(KNM-RU 1685 cast) left and Proconsul (KNM-CA 1300) right Note the differences inthe transition between the lingual cingulum and the mesial marginal ridge which ismore rounded in Proconsul and more angled in Ekembo The central pillar is also morebulbous or inflated and without ridging in Proconsul Additional specimens illustratingthe Proconsul morphology are shown in Pickford et al (2009 Fig 17 p 197) butreferred in that manuscript to Ugandapithecus
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6148
thanmerging into the hypocone on its lingual surface and a distallyoriented posthypocone crista Lower first and second molars nearlyequal in buccolingual breadth
Comparative morphology of Ekembo and Proconsul
Samples and comparative methods
The above diagnoses are based on extensive survey and analysisof the original fossil material Specimens housed in the NationalMuseums of Kenya (NMK) and the Natural History Museum (Lon-don) were analyzed by the authors during several separate and twojoint (NMK) research visits Fossils in the Uganda National Museumwere assessed by one of us (KPM) during two visits All measure-ments were collected by KPM Dental terminology follows Szalayand Delson (1979)
The following results are organized with respect to themorphology of the Kisingiri sample (attributed to E nyanzae andE heseloni) versus that of Proconsul from Tinderet and Napak(traditionally referred to P africanus P major and more recentlyP meswae) Specimens from additional localities that have at onetime been attributed to Proconsul (eg Andrews1978 Pickford andKunimatsu 2005 [cf Harrison 2010]) are for the most part notconsidered here most are middle Miocene in age and have beentransferred to other genera (Harrison 2002 2010) Of those that arefrom early Miocene sites there is growing evidence that P majorcan also be found at Moroto and differentiated fromMorotopithecus(Pickford et al 2009) an issue of some interest but ultimately notcritical to the taxonomic questions being considered here Whereappropriate however we do provide observations on a few earlyMiocene specimens attributed to ldquoUrdquo gitongai (Pickford andKunimatsu 2005 Pickford et al 2009 see Harrison 2010) Yetwhile we consider the middle Miocene holotype likely to representa distinct species here referred to Proconsul we concur withHarrison (2010) that many specimens placed in this species byPickford et al (2009) cannot be distinguished from P major
Sample sizes particularly from Tinderet and Ugandan localitiestend to be small but where possible standard univariate (eg t-tests) and bivariate (regression ANCOVA) statistical analyses wereperformed A single eigendecomposition of the covariance matrixof symphyseal measurements was done to characterize the multi-variate shape of the mandibular symphysis after accounting for sizedifferences (sensu Jolicouer 1963) Quantitative results are illus-trated as box-and-whisker plots of shape indices or bivariate plotsof the measurements For samples of fewer than five specimensshape index values are plotted in lieu of box-and-whiskersummaries
Morphological comparisons
Upper incisors Two features of the Ekembo upper central incisorsdistinguish them from those of Proconsul including specimenswidely attributed to the type species P africanus The lingual tu-bercle in Ekembo is quite narrow generallywith low relief and oftenscored by two or more irregular ridges in contrast to the muchmore inflated or bulbous tubercle in Proconsul I1s (Fig 2) Ekembospecimens also have a characteristic morphology in the transitionfrom the lingual cingulum to the mesial marginal ridge (Fig 2)from the mesiodistal mid-point of the crown the cingulumextends either directly mesially or even angles slightly cervicallyto intersect the mesial marginal ridge at a fairly sharp angle Incontrast in I1s of Proconsul the cingulum angles incisally from themid-point and transitions into the mesial marginal ridge in agradual curve The expression of these traits varies somewhat inthe smaller Tinderet specimens attributed to P africanus
compared with those of P major (cf Pickford et al 2009)However these variations are relatively minor with respect to thedifferences between the two genera Of 25 I1s known fromKisingiri localities only one specimen KNM-MW 562 somewhatresembles the Tinderet sample in both of these featuresdhavinga somewhat less angled transition from the mesial marginal ridgeto the cingulum and a more bulbous lingual tubercle This isparticularly interesting as the specimen was discovered in theMakira Beds now dated to older than 190 Ma and contemporarywith the Tinderet localities (McCollum et al 2013) Neverthelessits overall morphology is more similar to other Ekembospecimens than to Proconsul incisors Two other Ekembospecimens KNM-RU 1677 and RU 1933 have somewhat moreinflated lingual tubercles but otherwise resemble other Ekembo I1s
No consistent differences were found in upper lateral incisors
Upper canines Upper canine crowns of Ekembo are typical of thoseof most catarrhines in terminating in an evenly tapering point Thisdiffers from the Proconsul specimens which have a blade-like(Kelley 1986 Senut et al 2000 Harrison 2010) or ldquoburin-likerdquo(Pickford et al 2009) canine tip (Fig 3) While blade-like apicalmorphology has long been recognized as a distinctivecharacteristic of P major we confirm the report by Pickford et al(2009) that this feature is also present in the P africanus typespecimen (Fig 3a) although somewhat understated as expectedin the more slender and higher-crowned P africanus caninesUnfortunately other canines attributed to P africanus (eg M44837 KNM SO 419 LG 921 CA 1910 SO 5353 Kelley 1986) havebroken apices and therefore cannot be evaluated for this featureA blade-like tip is also present in the deciduous upper canines ofP major (eg KNM-SO 371 MV 10 NAP IV UMP 62-20) andP meswae (KNM ME 11) Hence the blade-like upper canine tipunites all Tinderet and Napak Proconsul species but is neverexpressed in Ekembo specimens from Rusinga and Mfangano
Two features of the canine roots also differentiate these generalongitudinal grooving or fluting along the length of the root (cfKelley 1986) and the development of numerous finer longitudinal
Figure 3 Canines of Proconsul and Ekembo a) Upper canine of the P africanus holotype (M 14084) lingual view b) Upper canine of P major (KNM-CA 2127) lingual view c) Lowercanine of P major buccal view d) lower canine of Ekembo cf nyanzae (KNM-RU 1676) buccal view and e) Upper canine of P major (KNM-SO 584) distal view Note the blade-like orldquoburin-likerdquo tip of the unworn Proconsul canines including on the nearly unworn canine tip of the P africanus type specimen (a) Ekembo canines have more typical pointed tipsProconsul canines also have longitudinal grooves and fine striations on the root (e) Ekembo lacks these features Scale bars frac14 1 cm
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 49
striations near the root apex that extend to varying degrees towardthe cervix of the tooth Longitudinal ldquoflutingrdquo consists of broadgrooves that segment the root into distinct sections (cf Pickfordet al 2009 Fig 3e) Most often the root in cross-section has theappearance of a cylinder that has been sectioned longitudinallywith the two halves offset from each other but in some cases thereare three segments and the root cross-section is more triangular inappearance (Kelley 1986) This characteristic was noted by Kelley(1986) in the P africanus canines but not in those of P majorHere we affirm that fluting is present on all of the visible uppercanine roots attributed to both P africanus and P major rootmorphology could not be evaluated in the deciduous upper caninefrom Meswa Bridge since it is implanted in the maxillary alveolusNone of the Ekembo specimens display this morphology havinginstead a simpler round to ovoid cross-section along the length ofthe root
The fine longitudinal striations on Proconsul upper canine rootsvary in the degree to which they extend toward the cervix (Fig 3e)but they are visible in all specimens attributed to P africanus(KNM-SO 419 521 921 5353 KNM-X 502) and five of six uppercanines of P major (excepting KNM-SO 1614) This morphology isalso present in UMP 62-12 which was included as part of theMorotopithecus bishopi hypodigm (Gebo et al 1997) Fine striationsalong the upper canine root are almost entirely absent in EkemboTwenty of twenty-three specimens lack this feature three speci-mens (KNM-RU 1763 1845 1891) exhibit incipient to slight
striations but these are never developed to the extent seen inProconsul canines
Upper premolars Upper third premolars display some of thegreatest differences between Ekembo and Proconsul P4s tend todiffer in similar ways but the differences are not nearly as markedEkembo P3s have cusps positioned near the mesiodistal midpoint ofthe crown well-developed mesial foveae pre- and postparacristaeof similar length and slope resulting in symmetric or nearsymmetric buccal profiles a straight or slightly curved mesialmargin and only moderately curved distal margin (Fig 4)Proconsul P3s are more similar in appearance to those of small-bodied catarrhines from the early Miocene with cusps positionedmesially on the crown greatly reducing or eliminating the mesialfovea They also have a steep preparacrista and more shallowlysloped postparacrista resulting in a highly asymmetric buccalprofile and an inwardly angled or notched mesial margin but astrongly curved distal margin This morphology is bestcharacterized in the P africanus holotype (M 14084) but can alsobe seen in other specimens attributed to this species as well asthose referred to P major (Fig 4d e) Also noteworthy is adistinctive mesiobuccal flange that mesially extends the buccalportion of the crown but only near the cervical line of the toothPickford et al (2009) describe this feature in some detail forldquoUgandapithecusrdquo but in fact this morphology is clearly expressedin the P africanus holotype as well (Fig 4a d) A single adult P3
Figure 4 Upper premolars of Proconsul and Ekembo occlusal (aec) and lingual (def) views a) P3e4 of the P africanus holotype (M 14084) b) P3 of P major (M 14331) c) P3e4 of theE heseloni holotype (KNM-RU 2036) d) P3 of the P africanus holotype (M 14084) e) P3 of P major (M 14331) f) P3 of the E heseloni holotype (KNM-RU 2036) Note the mesiallypositioned cusps lack of mesial fovea and strongly asymmetric crown in Proconsul compared to Ekembo (see text for further comparisons) Image of M 14331 reversed for easiercomparison Photo 4a courtesy of Rutger Jansma Scale bars frac14 1 cm
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6150
germ and two dP3s attributed to P meswae look remarkably similarto those of other Proconsul species including the presence of thisflange In contrast the Ekembo P3 has a well-developed mesialfovea that terminates at a mesiobuccal shoulder positionedapproximately halfway between the paracone tip and the cervixFrom this shoulder the mesial margin angles slightly distallydown to the cervix (Fig 4f)
The degree of cusp height heteromorphy in P3 also differentiatesEkembo from Proconsul (contra Pickford et al 2009) Only four non-Kisingiri specimens were sufficiently unworn to take measure-ments but more worn specimens also appear to express this dif-ferential in cusp height Figure 5a shows the ratio of protoconeparacone height measured from the tooth cervix and with only oneexception Ekembo P3s have substantially less cusp heteromorphyThis exception KNM-RU 1677 does not in fact have greater heter-omorphy but instead has an abnormal extension of the enamel ontothe buccal (paracone) root which skews the measurement Hencecusp relief relative to the occlusal surface in RU 1677 is not nearly asheteromoprhic as in Proconsul and when this is taken into accountthe difference between genera is significant (t-test dffrac14 11 t frac14 283p frac14 00165) The adult P3 germ from Meswa Bridge is incompletelyformed and therefore could not be measured However the differ-ence between paracone and protocone heights with respect to theocclusal surface appears to be as great as that in the P africanusholotype It does not appear that sex is an important determinant ofthis feature since well-established male (eg M16647 KNM-RU16000) and female (KNM-RU 2036 7290) specimens do not segre-gate across the Kisingiri range of variation
Upper fourth premolars of Ekembo are broadly similar inmorphology to Ekembo P3s in fact the similarity between P3 and P4
is substantially greater in Ekembo than in Proconsul (Figs 4a c)Likewise most of the morphological differences that discriminatethe P3s of these genera also tend to differentiate the P4s but less
clearly Cusp heteromorphy does not distinguish P4s of thesegenera but the overall height of the P4 paracone in Ekembo issignificantly greater relative to crown length than in Proconsul P4s(t-test df frac14 14 t frac14 316 p frac14 00069 Fig 5b)
Upper molars Cusps of Ekembo upper molars are uniformly morebunodont than those of Proconsul occupying more of the occlusalsurface Intercuspal crests are more fully incorporated into thecusps in Ekembo molars giving them a more inflated appearanceThis is particularly evident in the crista obliqua which is very broadin Ekembo but thin and knife-like in Proconsul (Fig 6) Cusps ofProconsul molars stand more as individual cones Pickford et al(2009) have further differentiated the cusp morphology ofsmaller and larger Tinderet specimens arguing that the latterhave more bulbous cusps While accurate this difference is minorcompared with the morphological difference between Ekemboand Proconsul and we regard the within-Proconsul cuspalvariation as size-related In fact the intermediately sizedP meswae has cusp morphology that is appropriatelyintermediate between that of P africanus and the larger P major
Other differences between Ekembo and Proconsul upper molarsare most evident in M2 We concur with previous assessments thatthe Kisingiri upper molars have less developed cingula than non-Kisingiri molars (eg Senut et al 2000 Harrison 2002 2010Harrison and Andrews 2009) More specifically Ekembo M2s lackor have only rudimentary buccal cingula between the paracone andmetacone Likewise the lingual cingulum is discontinuous merg-ing into the hypocone lingually and then re-emerging as a distalmarginal ridge (Fig 6) Proconsul M2s display substantially morecingular development overall and in particular exhibit a prominentbuccal shelf between the paracone and metacone and a distinctdistolingual cingulum that is continuous around the hypocone towhere it merges with the distal marginal ridge
Figure 5 Box-and-whisker plots a) P3 cusp heteromorphy index (protocone heightparacone height) Results of a t-test show this difference to be significant (p frac14 00165) Theexcluded Ekembo outlier KNM-RU 1677 only appears to have extreme heteromorphy because of extensive expansion of the buccal enamel onto the anterior root (see text) b) P4
relative paracone height (paracone heightmesiodistal crown length) Results of a t-test show this difference to be significant (p frac14 00069)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 51
The occlusal profiles of Ekembo M2s are squarish in shape withrelatively straight and equal sides with a posthypocone crista thatis distobucally to buccally directed In contrast the occlusal profilesof Proconsul M2s have a more rhomboid shape with substantialbasal flare around the mesial cusps and a distally expanded hypo-cone with a distally directed posthypocone crista resulting in astrongly curved distal margin (Fig 6) Even in specimens with lesscurved distal margins the posthypocone crista is still more distallydirected compared to the condition in Ekembo These features arecharacteristic not only of P africanus P meswae and P major butare also expressed in the isolatedM2 fromMoroto Uganda (MOR IIb2098) attributed by Pickford et al (2009) to ldquoUrdquo gitongai butregarded here as belonging to P major
As noted by others (eg Harrison and Andrews 2009) the de-gree of buccolingual molar flare differs among these samplesQuantified as the distance between the tips of the mesial cuspsdivided by maximum crown breadth the degree of molar flare in
Ekembo M2s is significantly less than in Proconsul M2s (t-testdf frac14 29 t frac14 396 p frac14 00004) In a bivariate plot of these twomeasures the distributions of Ekembo and Proconsul teeth barelyoverlap with only KNM-RU 1677 encroaching into Proconsul ter-ritory (Fig 7)
These differences in M2 morphology also tend to distinguishEkembo from Proconsul M1s but not always as clearly Whereas M2smore consistently express distinct morphotypes the M1s in bothgenera show more variation in these features
The degree of upper third molar development (ie whether M3
has four distinct cusps or the distal cusps are reduced or absent) haslong been cited as taxonomically informative for these species orgroups of these species (eg Hopwood 1933 Le Gros Clark andLeakey 1951 Andrews 1978 Pickford et al 2009) However wefound this tooth to be unreliable for taxonomic purposes Withinprimates M3s are typically the most variable molars in both sizeand shape (Blankenship et al 2007 Kavanagh et al 2007) and this
Figure 6 Occlusal view of the M2s of Proconsul and Ekembo Left P africanus holotype(M 14084) Right E heseloni holotype (KNM-RU 2036) Note in Proconsul the overalldifference in crown shape as well as the more conical individuated cusps and nar-rower cristae strongly developed lingual cingulum that continues to the distal aspectof the crown broad buccal cingular shelf and more distally directed posthypoconecrista In Ekembo the cusps are more bunodont and expanded the cristae are broaderwith better developed accessory tubercles the lingual cingulum merges into thehypocone there is no buccal shelf and the posthypocone crista is more buccallydirected
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6152
is observable even in small samples of Proconsul species TheP africanus holotype has a substantially reduced M3 whereas inKNM-CA 2250dattributed by most authors to this species(Andrews 1978 Pickford et al 2009)dit is fully developed Simi-larly the hypodigm of P major includes both fully developed M3s(eg KNM-CA 389 LG 1815) and specimens with reduced ormissing hypocones (KNM-CA 397 1299) Ekembo displays compa-rable ranges of variation from highly reduced to fully developedM3s in both species Thus these species and genera cannot bedistinguished based on M3 development and we did not find anyother distinguishing characteristics between Ekembo and Proconsulin the upper third molars
Figure 7 Scatter plot depicting the de
Mandible Pickford et al (2009) enumerated several traits thatreadily distinguish Kisingiri from non-Kisingiri mandibles andthese distinctions are entirely consistent with the distinction ofEkembo from Proconsul Our analyses specifically underscoredifferences in the symphyseal region The planum alveolare inboth male and female specimens of Ekembo is short and morevertically oriented even the most elongate of these (KNM-RU47805 McNulty et al 2007) is substantially more verticalcompared to specimens of Proconsul In contrast the planumalveolare in Proconsul is conspicuously long and nearlyhorizontally oriented Well-preserved mandibles attributed toP major (eg KNM-SO 396 and 404) exhibit this morphology inextreme fashion (Fig 8) smaller specimens (KNM-SO 1112P africanus) show less extreme expression but are still readilydistinguishable from Ekembo mandibles For its size KNM-LG 452has a surprisingly short planum alveolare though not to thedegree seen in Rusinga mandibles Correlated to the shortplanum alveolare in Ekembo is a reduced superior transversetorus which while well-developed does not approach therobusticity expressed in Proconsul mandibles This difference isreflected in an index of symphyseal robusticity (thicknessperpendicular to the long axis of the symphysislength of thelong axis) which shows Ekembo and Proconsul to be significantlydifferent (t-test df frac14 14 t frac14 430 p frac14 00007)
Development of the inferior transverse torusdoften used todifferentiate among species of Proconsul (eg Pickford et al 2009Harrison 2010)dwas found to be highly variable and inconsis-tently expressed within these samples However the overallorientation of the subplanum region (as defined by a line con-necting the posterior-most midline points of the superior andinferior transverse tori or the posterior-most midline point of theinferior margin of the symphysis in the absence of the latter) doesdistinguish Ekembo from Proconsul mandibles The orientation ofthis line in relation to the alveolar margin in the vicinity of thepostcanine teeth was assessed qualitatively by placing a small rod
gree of buccolingual flare in M2
Figure 8 Mandibles of Proconsul and Ekembo Occlusal views of P major a) KNM-SO 396 b) KNM-SO 404 c) KNM-LG 452 Occlusal views of Ekembo d) KNM-RU 2087 e) KNM-RU1674 f) KNM-RU 7290 Occlusal views of P africanus g) KNM-SO 372 h) KNM-SO 1112 i) Lingual view of P africanus (KNM-SO 1112) Note the long more horizontally orientedplanum alveolare in Proconsul resulting in a pronounced superior transverse torus The planum alveolare is shorter and more vertical in Ekembo such that one can see the inferiormandibular border from occlusal view
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 53
(eg a dental pick) in themidline internally along the two referencepoints and recording the point in lingual perspective at which therod intersected the alveolar margin (Fig 9) In Ekembo the sub-planum region is more vertically oriented so that the rod intersectsthe alveolar margin most often in the premolar field Not surpris-ingly there is a difference between sexes with females having areduced superior transverse torus and therefore a more verticallyoriented subplanum region compared to males In contrast withEkembo Proconsul mandibles have much more inclined subplanumsurfaces with the rod intersecting the lingual alveolar margin inthe vicinity of the M1 (females) or M2 (males Fig 9) When sex istaken into account therefore the distributions of Ekembo andProconsul in this feature are discrete Note that male and femalemandibles attributed to both P africanus and P major are includedin this assessment and are surprisingly similar despite dramaticsize differences
We also quantified internal symphyseal morphology using threemeasures symphyseal height (infradentaleegnathion) planumalveolare length (posterior-most point on the superior transversetoruseinfradentale) and subplanum length (posterior-most pointon the superior transverse torusegnathion) a combination thatincorporates not only the lengths of these intervals but also theangular relationships among them To compare symphyseal shapeacross a range of sizes we chose a simple principal componentsapproach to partition size onto the first eigenvector (cf Jolicouer1963) While this is not the preferred method for partitioning sizein most datasets (Jungers et al 1995) for this particular sample andset of measurements Mosimann shape variables were still
significantly correlatedwith size (see discussion in Coleman 2008)Hence we assessed differences in symphyseal morphology be-tween Ekembo and Proconsul using the second and third principalcomponents of the three variables Distributions of the two generaare distinct in this shape space (Fig 10 Supplementary OnlineMaterial [SOM] Table 1)
Robusticity of the mandibular corpus also differs between thesesamples though it is more difficult to test due to differentialpreservation among specimens Corpus thickness and depth underP4 can be measured in the greatest number of specimens andcomparison of these measures suggests a difference betweengroups in the scaling of corpus robusticity (Fig 11) Among largepresumably male specimens Ekembo mandibles are substantiallyless robust than those of Proconsul However among smallermandibles only a single Proconsul specimen (KNM-SO 372) could beincluded and it is similar in robusticity to Ekembomandibles of thissize Considered together the impression is that corpus robusticitymay scale differently in Ekembo than it does in Proconsul and a testfor differences in slopes using an ANCOVA design of the loggedvariables was significant (df frac14 2 F frac14 2481 p lt 00001) Never-theless additional specimens are needed to properly evaluate theinfluence of taxonomy sex and size on corpus robusticity areasonable alternative interpretation of Figure 11 is that it dem-onstrates a significant difference between P major and all otherspecies of both Proconsul and Ekembo
Lower incisors Ekembo specimens are uniformly narrower inmesiodistal length than incisors attributed to P africanus P major
Figure 10 Scatter plot of principal component scores based on an eigenanalysis of threeposterior point on the superior transverse toruseinfradentale) and subplanum length (mprimarily captures size differences among specimens (Jolicouer 1963 see text and SOM Trespectively) in the symphyseal variables
Figure 9 Line drawing of a hemimandible depicting different orientations of the sub-planum area in Ekembo and Proconsul Females are denoted using shorter vectors maleswith longer vectors Vector is estimated in M 14086 which is broken at the midlineEkembo specimens have a more vertical subplanum compared to those from Songhorand Legetet but within both groups females have more vertically oriented regions thanmales Hence even the most ventrally rotated subplanum in the largest Ekembo malespecimen (KNM-RU 47805) groups with the Tinderet females in this feature
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6154
and P meswae both absolutely and in relation to crown height(Fig 12) This is true for both I1 and I2 A t-test adjusted for non-independent observations (eg I1 and I2 from the same mandible)confirmed the difference as significant (t-test df frac14 12 t frac14 258p frac14 00242) Moreover while the measurable Proconsul samplewas small this same pattern is observable in other specimenswhose tips were slightly to moderately worn and therefore notmeasured
Lower canines The only lower canine trait that discretely sepa-rates these genera is the pointed crown tip in Ekembo (Fig 3d)contrasted with a blade-like (or ldquoburinrdquo) tip in Proconsul(Fig 3c) This feature is well established for P major but is alsoexpressed in KNM-CA 1772 which was referred to P africanus byKelley (1986) and is too small to be either P major or P meswaeTwo other specimens commonly attributed to P africanus KNM-CA 2149 and KNM-SO 1112 both have broken canine tipsLikewise a new specimen (Kor 65004) attributed to P africanusby Pickford et al (2009) also has a broken tip and cannot beevaluated (see Pickford et al 2009 Fig 23) As with the uppercanines the blade-like tip is also present in the deciduous lowercanines of P major (eg KNM-CA 361 S0 5352) and P meswae(KNM-ME 1)
Lower premolars Like most previous authors (eg Andrews 1978Harrison 2002 2010 Pickford et al 2009) we found neitherqualitative nor quantitative characteristics of the lower premolarswith any taxonomic value
Lower molars Differences in lower molar morphology are few andrelatively subtle but in some cases mirror those seen in the uppermolars Cusps on the lower molars of Ekembo are more bunodontthan those in Proconsul and occupy more of the occlusal surfaceBroad inter-cuspal crests are incorporated into the Ekembo cuspsmaking them appear more inflated Molar cusps of Proconsul are
variables symphyseal height (infradentaleegnathion) planum alveolare length (mostost posterior point on the superior transverse torusegnathion) The first componentable 1) and thus PC 2 and PC 3 illustrate the residual shape variance (72 and 28
Figure 11 Scatter plot showing mandibular corpus robusticity under P4 Trend lines were computed using ordinary least squares (OLS) regression Similar relationships areobservable at other parts of the corpus but differential preservation results in smaller samples at those positions Slopes are significantly different (p lt 00001 tested on loggedvariables)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 55
more distinct from one another and are linked by narrower sharperinter-cuspal crests The lower molars of Ekembo also have asubstantially less developed buccal cingulum than those ofProconsul (Harrison 2002 2010 Harrison and Andrews 2009)
Figure 12 Scatter plot of lower incisor crown proportions Differenc
Lastly and again as seen in the upper molars the two genera canbe distinguished by the significantly greater amount of lowermolar flare in Proconsul (Fig 13 t-test df frac14 59 t frac14 713p lt 00001 see also Harrison and Andrews 2009)
es between Ekembo and Proconsul are significant (p frac14 00242)
Figure 13 Box-and-whisker plot of buccolingual flare in lower molars measured asthe average distance between the mesial buccal and lingual cusps divided by overallbuccolingual crown breadth Differences between Ekembo and Proconsul are highlysignificant (p lt 00001)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6156
Relative molar size has been used to distinguish species ofProconsul but we observed the primary difference to be in relativebuccolingual breadths of M1 and M2 In Ekembo M1 is substantiallynarrower than M2 than is the case in Proconsul (Fig 14 Harrison2002 2010) Although none of the smaller Proconsul mandiblespreserve complete crowns for both M1 and M2 KNM-SO 1112 hasan intact M2 and at least preserves the cervix for M1 However evenif the cervical measurement is used for its M1 breadthdthusunderestimating the true crown breadthdKNM-SO 1112 still has abroader first molar in relation to M2 than would be expected inEkembo If crown breadth is estimated from the ratio of cervixbreadth to maximum crown breadth in other Proconsul fossils therange of likely breadths for the SO 1112 M1 clearly distinguishes it
from the Ekembo sample (Fig 14) Testing this relationship onlogged variables using the mean estimate for the SO 1112 M1(Fig 14) reveals a significant difference between the two groups(ANCOVA df frac14 1 F frac14 2424 p frac14 00002)
As with upper third molars we find a great deal of variation inM3 morphology and mistrust the taxonomic significance some-times placed on characteristics of this toothdat least with regard tothese taxa Harrison (2002 2010) cited the mesiodistal position ofthe M3 entoconid as differentiating Kisingiri third molars fromthose attributed to P africanus We concur with his assessment butfound the entoconid position to vary in specimens attributed toP major Thus this character does not differentiate Ekembo from allProconsul species but may distinguish it from the type speciesP africanus
Discussion
The distinctiveness of the Kisingiri large catarrhine materialfrom that at Tinderet and Ugandan localities has been noted pre-viously by several authors (Pickford 1986 Walker et al 1993Begun 2001 2007 MacLatchy and Rossie 2005 Harrison andAndrews 2009 Harrison 2010) What had not been fully docu-mented was the nearly complete lack of overlap between thesesamples in a number of features despite very large assemblagesfrom Rusinga and Mfangano On descriptive criteria alone (Table 2)many isolated teeth (I1 C1 P3 M2 C1) and all of the mandibles canbe assigned to either Proconsul or Ekembowith very little chance ofmisattribution That some of the most diagnostic features occur inthe upper dentitiondthe only anatomy preserved in theP africanus type specimendadds weight to the taxonomicarrangement proposed here The morphology of the upper teeth inall Proconsul species compellingly unites them into a single cladedistinct from E heseloni and E nyanzae This is reinforced bynumerous features of the mandible and mandibular dentition thatfurther distinguish these samples The number and variety of thesefeatures are such that Proconsul and Ekembo are more differentfrom one another than are many closely-related modern catarrhinegenera
Derived morphology of Ekembo and Proconsul
It is premature to formally assign primitive and derived status tothe individual features of Ekembo and Proconsul before completinga formal phylogenetic analysis This is especially true given theabundance of evidence available for Ekembo and hence the poten-tial impact that our generic revision could have on phylogeneticinterpretations Nevertheless in comparison with both extanthominoids and the small-bodied catarrhines from the earlyMiocene it is informative to differentiate the evolutionary grades ofthese genera Many characteristics of Ekembo are more advanced inthe direction of modern apes compared to those of Proconsul(Table 2) General trends such as expanded and more bunodontcusps broader molar crests reduced cingulum and morphologicalsimilarity between P3 and P4 all represent morphologies thatpresage those of later fossil and extant hominoids The moreadvanced morphologies of P3 and M2 are particularly conspicuouscompared not only with those of Proconsul but to most earlyMiocene catarrhines In contrast the mesially positioned stronglyheteromorphic cusps on the P3 of Proconsul plus the correspondingasymmetric buccal profile notched mesial margin and missingmesial fovea are characteristic of other early Miocene catarrhinessuch as Dendropithecus Rangwapithecus and Limnopithecus andare likely primitive among East African catarrhines The moresymmetric P3 of Ekembo with midline positioned cusps andreduced cusp heteromorphy is reminiscent of later hominoids and
Figure 14 Relative buccolingual dimensions of M1 and M2 KNM-SO 1112 has a preserved M2 but only the cervix of M1 Its M1 breadth was estimated based on the ratio to cervixbreadth to maximum crown breadth in other Tinderet fossils Even the cervical breadth of SO 1112 is broader than would be predicted for the crown breadth of a Kisingiri M1 Trendlines computed using OLS regression and using the average estimate of SO 1112 M1 breadth Group differences were significant (p frac14 00002)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 57
even hominid taxa such as Rudapithecus (eg Kordos and Begun2001)
Proconsul is likewise united by a suite of features that are for themost part characteristic only of this lineage (Table 2) Themorphology of both upper and lower canines is unique to thisgenus and shared by all three well-known species plus the poorlyrepresented middle Miocene P gitongai (Pickford and Kunimatsu2005) The mandibular symphysis is also distinctive in Proconsulcompared with other catarrhines with its very long and shallowlysloping planum alveolare and steeply inclined subplanum surfaceIndeed even the small edentulous anterior mandibular fragmentKNM-SO 372 is readily placed within Proconsul and excluded fromother similarly sized catarrhine taxa such as Rangwapithecus andEkembo based on this unique symphyseal morphology (Figs 8 and9)
Other differences between Ekembo and Proconsul are of uncer-tain evolutionary polarity (eg orientation of the posthypoconecrista) or may be of taxonomic relevance without broader phylo-genetic significance (eg degree of molar flare Table 2) Werecognize that there are morphological differences within Procon-sul particularly between the better known species P africanus andP major However many of these differences are likely related tosize and represent variations on a single generic theme that aremarkedly different from the morphology of Ekembo The recentdescription (Harrison and Andrews 2009) of the intermediatelysized P meswae underscores the broad morphological unity withinProconsul and further substantiates a well-defined Proconsul cladecomprising Tinderet and Ugandan species
Based on the abundant morphological and morphometric evi-dence supporting genus-level distinction between Proconsul andEkembo we are currently conducting a more detailed phylogeneticanalysis of all early Miocene catarrhines in order to better under-stand the distribution and evolutionary significance of these andother morphological features We anticipate that the debates sur-rounding the phylogenetic relationships of an historically consti-tuted sample of Proconsul can be more satisfactorily resolved nowthat the Kisingiri material is appropriately recognized as a distinctgenus
The generic disposition of ldquoUgandapithecusrdquo species
A great many features said to characterize ldquoUgandapithecusrdquowere added to the original diagnosis of the genus by Pickford et al(2009) in their systematic revision of Proconsul (sensu lato) As partof that study they also erected a new species ldquoUrdquo legetetensis andtransferred P meswae to ldquoUgandapithecusrdquo Evaluating the same setof features across the entire available sample however we wereunable to find support for this more expansive conception ofldquoUgandapithecusrdquo for several reasons First some diagnostic fea-tures cited in Pickford et al (2009) were quite variable within theldquoUgandapithecusrdquo sample As an example marked reduction of theM3 entoconid to a bead on the lingual cingulum is not shared byseveral ldquoUrdquo major specimens such as KNM-CA 393 NAP I 49000and Kor 253004 (see Pickford et al 2009 Fig 33e) and was likelynot a feature of a few of the more worn specimens eg BUMP 600and BUMP 601 or even the holotype of P major M 16648 Second
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6158
some diagnostic features of ldquoUgandapithecusrdquo were found to bevariably present in Ekembo arguing against their utility for dis-tinguishing ldquoUgandapithecusrdquo from Pickford et als (2009) com-bined P africanus thorn Kisingiri conception of Proconsul For examplesignificantly reduced curvature of the I1 distal margin in ldquoUganda-pithecusrdquo is also found in some Kisingiri specimens including KNM-RU 1685 1712 1714 1831 1846 and 1951 several more Rusingaspecimens have moderately reduced curvature Third some fea-tures that purportedly distinguish ldquoUgandapithecusrdquo fromP africanus are likely related to size For example the somewhatmore central position of the P3 paracone in ldquoUrdquomajor is not seen inthe smaller ldquoUrdquo meswae whose paracone is positioned similarly tothat of the holotype of P africanus (see Harrison and Andrews2009 Fig 6d)
However the major part of the evidence for recognizingldquoUgandapithecusrdquo (sensu Pickford et al 2009) was assembled by re-assigning specimens traditionally attributed to P africanus into newspecies of ldquoUgandapithecusrdquo The P africanus holotype was left inassociation with the Rusinga sample despite having morphologyoverwhelmingly characteristic of other Tinderet specimens Theresulting specimen allocations left P africanus morphologicallyimpoverisheddstripped of much of its hypodigmdso that Pro-consul for Pickford et al (2009) came to be characterized primarilyby the Rusinga sample (see similar comments in Harrison andAndrews 2009) Within that conceptual framework any spec-imen that differs from those at Rusinga could not be Proconsul butmust instead belong to ldquoUgandapithecusrdquo Thus specimens such asthe KNM-SO 1112 mandible were erroneously assigned to ldquoUgan-dapithecusrdquo because they were (correctly) perceived as differingfrom their Rusinga counterparts (Pickford et al 2009)
Here we restore the P africanus hypodigm and associate it withthe other Tinderet fossils according to their shared morphologySpecimens assigned to ldquoUgandapithecusrdquo revert to Proconsul andmany of the features cited to support a speciose ldquoUgandapithecusrdquo(Pickford et al 2009) instead become diagnostic features of Pro-consul This correction serves to highlight the distinctiveness of theKisingiri remains and the need to recognize this distinctiveness atthe generic level
As demonstrated above specimens from Kisingiri do not displaythe morphology of Proconsul as represented by the holotype ofP africanus M 14084 Given that this specimen preserves only theupper canine and cheek teeth similarities with or departures fromthis morphology represent the most compelling arguments forassessing inclusivity in the genus However for the mandible andlower dentition if one simply sorts the specimens by elementwithout relying on taxonomic preconceptions the overwhelmingdistinction within the traditionally constituted Proconsul sample isbetween Kisingiri material on the one hand and that from Tinderetand Uganda on the other (cf Walker et al 1993 Begun 2001 2007MacLatchy and Rossie 2005 Harrison and Andrews 2009Harrison 2010) Additionally with the Kisingiri specimensremoved to Ekembo the postcranial differences used to partlyjustify a distinct ldquoUgandapithecusrdquo lose their relevance (seeHarrison and Andrews 2009)
We have differed among ourselves in our past treatment ofP major as generically distinct from the smaller species of Pro-consul (Begun 2007 McNulty 2010) Morphologically many ofthe unique features of P major can be interpreted as size-relatedvariants on the smaller P africanus morphology Even if so thequestion is whether these are sufficiently different and numerousto warrant generic distinction that is do they suggest thatP major belongs in a separate clade from P africanus andP meswae As we have demonstrated differences among thesethree species are certainly fewer and show less morphologicaldistinction than those between Ekembo and any of the three
Nevertheless while we ultimately agree with Harrison andAndrews (2009 see also Begun 2007) that the features dis-tinguishing ldquoUrdquo major from P africanus are consistent with thosethat distinguish congeneric species and are at least partly due todifferences in size we concede the possibility that the largestProconsul species could belong in a separate genus (Senut et al2000 Pickford and Kunimatsu 2005) However whether or notone recognizes a separate genus for P major the P africanus ma-terial does not sort morphologically with the Kisingiri materialTherefore the taxonomic alternatives are either two generaProconsul and Ekembo or three Proconsul Ugandapithecus andEkembo with Proconsul and Ugandapithecus united in a cladeexclusive of Ekembo Thus while we strongly favor the firstalternative recognizing Ugandapithecus does not change thefundamental argument concerning the need to refer the Kisingirispecies to a generically and cladistically distinct taxon from thespecies represented at Tinderet and the Ugandan sites
Specimens of interest
Despite the substantial dental and gnathic differences betweenProconsul and Ekembo there are a few specimens whose taxonomicaffinities require additional scrutiny
KNM-RU 1676-77 This set of associated upper and lower teeth is aconsistent outlier among E heseloni and E nyanzae specimens Insome ways this specimen more closely resembles Tinderet Pro-consul in the features that distinguish it from Ekembo Molar cuspmorphology and cingulum development I1 lingual tubercledevelopment and M2 shape and posthypocone crista orientationare more consistent with specimens from the Tinderet sites On theother hand it otherwise resembles Ekembo and lacks the strongestdiagnostic features that unite species of Proconsul such as thedistinctive canine and P3 morphology Overall this specimen doesnot fit unambiguously into either genus in fact the degree ofbuccolingual flare cusp morphology enamel wrinkling and espe-cially the long extension of enamel onto the anterior root in P3 arein some ways reminiscent of Afropithecus although it clearly differsfrom that genus as well (cf Rossie and MacLatchy 2013)
Were these characteristics individually distributed among thelarge Kisingiri sample one might attribute this to intra-genericvariation but when found together in a single specimen from alarge sample it suggests a unique individual One of us (KPM)thinks it likely that the specimen was not actually found at aKisingiri locality based on its unique preservation compared toother Rusinga fossils and on other examples of mis-proveniencedspecimens (Leakey 1967 Andrews and Molleson 1979 Pickford1986 Kelley 1986) However we recognize that relying toofrequently on this explanation allows almost any scenario to bejustified and suggest some alternative possibilities One is that thespecimen was found on Rusinga but in the much older WayandoFormation strata instead of the Hiwegi Formation The differencesin morphology in that case might reflect more than a million yearsof temporal change (McCollum et al 2013) with KNM-RU 1676-77preserving certain more primitive Proconsul-like features As analternative it is reasonable to assume that fossil assemblages asvast as those from Rusingawill include not only abundant taxa butalso rare or infrequent inhabitants of the local biome There aremany examples of this from Rusingas mammalian communityeg ldquoHemicyonrdquo Kulutherium Tadarida cf Hyainailouros cfArchaeobeledon Kelba (see relevant chapters in Werdelin andSanders 2010) As such KNM-RU 1676-77 may simply representa rare and well-preserved specimen of some other catarrhinetaxon that lived among or visited a primate community dominatedby Ekembo
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 59
KNM-SO 1112 This specimen was initially referred to Rangwapi-thecus but soon after was transferred to Proconsul afrianus (Bosler1981 Martin 1981)da designation that most subsequent authorshave followed (Pickford 1986 Nengo and Rae 1992 Kelley 19861995 Walker et al 1993 MacLatchy and Rossie 2005 Hill et al2013) Pickford et al (2009) more recently referred this specimento ldquoUrdquo meswae citing two primary reasons First they reiteratedBoslers (1981) concern that its P3 differs from others attributedto P africanus However at that time the majority of what wasconsidered to be P africanus would have been from RusingaIsland In fact while noting its contrasting morphology Boslersuggested that differences between KNM-SO 1112 and other P3smight reasonably be attributed to sexual dimorphism notingthat this specimen provides ldquoa not implausible match for the Pafricanus holotype maxilla M 14084rdquo (1981152) Thatassessment is borne out in this study Using the much largersample of Ekembo P3s as a model variation can be seen toencompass specimens with well-developed mesial beaks andthose with more bilaterally compressed crowns and parallelsides The latter morphology is predominantly found amongmale specimens (KNM-RU 1674 1924 and KPS I and III) thussupporting Boslers (1981) suggestion that this variation isrelated to sexual dimorphism and hence that KNM-SO 1112 is amale P africanus
The second feature cited by Pickford et al (2009) in referringthis specimen to ldquoUrdquo meswae was posterior shallowing of themandibular corpus However because they redefined ldquoUgandapi-thecusrdquo to include all of the well-preserved Tinderet mandibles(Pickford et al 2009)dleaving Proconsul to be represented only byRusinga specimensdtheir only alternative within this scheme for aspecimen the size of SO 1112 was to refer it to ldquoUrdquomeswae It seemsodd though to assign this specimen to a species from a (perhapsmuch) older site when the Proconsul africanus holotype is ofappropriate size and similar age and as there are no other adultmandibles from either species with which to compare it In anycase mandibular shallowing is variable in both Ekembo and Pro-consul with some Kisingiri specimens exhibiting considerableshallowing (eg KNM-RU 2087) and some ldquoUgandapithecusrdquo spec-imens having very little (eg M 14086)
We therefore see no morphological grounds to assign KNM-SO1112 to P meswae but do find some morphological evidence toassign it as a male P africanus which occurs at Songhor
KNM-LG 452 This very complete mandible was originally recog-nized as a female of P major based on the size of the dentition androbusticity of the mandible (Martin 1981) Bosler (1981) howeverleft this specimen unassigned noting similarities to P major in P3-M1 size but having distinctly smaller M2e3 In this she likenedKNM-LG 452 to a few other specimens (notably M 14086) thatare smaller in their distal molars in a manner more consistentwith taxonomic rather than sex differences (Bosler 1981 see alsoPilbeam 1969) Kelley (1986 1995) also assigned this specimen asa female of P major based on canine size and morphology butnoted that the canine lacked the shallowly-sloping mesial ridgetypical of most female anthropoid canines This is also truehowever of two other canines assigned as female P major KO 9and SO 373 (Kelley 1986) and so might simply be an unusualcharacteristic of P major female lower canines In the end Kelley(1995) concluded that considering all of the criteria fordistinguishing male and female canines the evidence that KNM-LG 452 is female remains questionable
Pickford et al (2009) further developed the idea that two spe-cies are represented within the P major sample and made KNM-LG452 the holotype of a new species ldquoUrdquo legetetensis We concur thatthis specimen is unique among the well-preserved Tinderet
mandibles in some important features Whereas male and femalemandibles of P major have long internal symphyseal planesexpansive extra- and retromolar sulci and rami that cross theocclusal plane at distal M3 KNM-LG 452 has a shorter symphysealplane (though not to the extent seen in Ekembo) a moderateextramolar sulcus but no retromolar sulcus and a ramus thatcrosses the occlusal plane more mesially If KNM-SO 396 and SO404 represent males and females respectively of P major then themorphology of KNM-LG 452 appears distinct
At present however we do not think there is sufficient evidenceto differentiate this specimen from either P major or P meswaePickford et al (2009) argue that KNM-LG 452 cannot belong to thelatter species because of its larger size The size differences how-ever are well within the range of variation seen in P majorE heseloni and E nyanzae Moreover whereas the Legetet mandibleis larger than most of the P meswae hypodigm an adult P4 referredto this species (KNM-ME 25) substantially exceeds its counterpartin KNM-LG 452 Should the Legetetmandible ultimately prove to bea male specimen size alone would not rule out its attribution toP meswae On the other hand if it is female then it becomes harderto justify its inclusion in that sample of mostly smaller specimens Iffemale therefore it could represent a separate species for whichthe binomial Proconsul legetetensis comb nov is available Recentlycollected fossils from the SonghorKapurtay area provide someadditional support for the possibility that three species of Proconsulexisted at these localities (McNulty 2014ab) However toconvincingly demonstrate this and determine whether a thirdspecies can be differentiated from P meswae will require a moredetailed analysis of variation among Proconsul species a corre-sponding study of variation in modern taxa anddalmost certain-lydadditional fossil specimens For now we regard ldquoUrdquo legetetensisas a subjective junior synonym of P major
Conclusions
The genus Proconsul has been central to studies of ape and hu-man evolution since its first description by Hopwood (1933) As oneof the best known fossil catarrhines represented by nearly everyskeletal element our knowledge of Proconsuls functional anatomypositional behavior and overall morphology has made it thefoundation for paleoanthropological research in the early Mioceneas well as studies on the origin and early diversification of homi-noids With respect to systematics however Proconsul is less wellunderstood the type species P africanus is known from a rela-tively small number of specimens from sites in the Tinderet areaand no postcranial bones can be unambiguously assigned to itRather it is the copious and well-preserved specimens from theKisingiri sites of Rusinga and Mfangano Islands that have formedthe basis for the common image of Proconsul
We argue here that the Kisingiri species traditionally referredto ProconsuldP heseloni and P nyanzaedare generically distinctfrom P africanus and hereby transfer them into a new genusEkembo based on a substantial number of morphological differ-ences (Table 2) Considering only those traits preserved in theP africanus holotype the entire sample from Rusinga and Mfan-gano differs markedly in the morphology of the upper canine P3P4 and M2 Additional differences in the I1 lower incisors andcanines molar cusp and occlusal crest morphology degree of flarein all molars and especially mandibular features clearly distin-guish the Kisingiri from non-Kisingiri samples in a way that isincompatible with congeneric species With the transfer ofE heseloni and E nyanzae Proconsul becomes a well-defined cladecomprising at least three speciesdP africanus P major andP meswaedunited by several features that are unique for thegenus (Table 2)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6160
We found no support in this study for the genus ldquoUgandapi-thecusrdquo as constructed by Pickford et al (2009) In that paper theytransferred the vast majority of Proconsul fossils from the Tinderetand Napak localities into ldquoUgandapithecusrdquo leaving P africanusrepresented only by its holotype and a small number of isolatedteeth and aligning it with the material from Rusinga and Mfan-ganoWhen the P africanus hypodigm is reconstituted and properlyaffiliated with the other Tinderet species according to itsmorphology much of the evidence marshaled by Pickford et al(2009) for ldquoUgandapithecusrdquo actually supports the distinction be-tween the Tinderet and Kisingiri samples ie the generic distinc-tion of Ekembo In addition some of the traits enumerated byPickford and colleagues in support of ldquoUgandapithecusrdquo wererevealed in our analysis to be variable within one or more speciesand therefore of questionable taxonomic utility We do not rejectout of hand the original conception of ldquoUgandapithecusrdquo (Senutet al 2000) as either a monotypic genus distinguishing ldquoUrdquo ma-jor or perhaps including a closely allied middle Miocene ldquoUrdquogitongai as well (Pickford and Kunimatsu 2005) but we have fornow retained the early Miocene specimens of both in Proconsulpending further study However even if Ugandapithecus is a taxo-nomically valid genus comprising just the latter two species theirmorphology indicates that theywould still be united in a cladewithProconsul and they lack several more advanced features that partlydefine Ekembo Better understanding of the role that body size playsinmediating differences within Proconsulwill help to further clarifythe relationships among these species
Despite the existence of one well-represented and several lesscomplete skeletons of what was long assumed to be Proconsul thephylogenetic status of this genus has defied consensus This islargely due in our view to Proconsul having been an admixture oftwo distinct genera one more advanced in the direction of extantapes than the other As we demonstrate here the material fromRusinga and Mfangano traditionally referred to Proconsul andwhich includes all of the more complete skeletal material isdistinct from the Tinderet and Napak material and belongs in aseparate genus Ekembo By disentangling these distinct clades weanticipate being able to achieve better phylogenetic resolution forboth genera thereby improving our knowledge of early hominoidevolution and diversification
Acknowledgments
We thank many people and agencies that assisted with thedevelopment and completion of this project This research wasinitiated as a result of funding from the Wenner-Gren Foundationand the Johns Hopkins School of Medicine to DR Begun in1987e1989 and was renewed following research and travelgenerously funded by multiple Leakey Foundation grants to KMcNulty National Science Foundation grants to K McNulty(0852609 1241807) the McKnight Foundation the University ofMinnesota the Lloyd A Wilford Endowment the LeverhulmeFoundation and the Turkana Basin Institute J Kelley gratefullyacknowledges support from the Institute of Human Origins Ari-zona State University DR Begun also wishes to acknowledge theAlexander Humboldt Stiftung and NSERC for funding support Weare grateful to the National Museums of Kenya the UgandaMuseum the Natural History Museum and to Rose Mwanja EzraMusiime Tom Mukhuyu Samuel Muteti James Yatich and MaryMuungu for access to and assistance with specimens in their careMany people provided assistance comments criticisms insightsand suggestions on various aspects of this work and we especiallythank Holly Dunsworth Will Harcourt-Smith Tom Lehmann DanPeppe Nicole Garrett Kirsten Jenkins Rutger Jansma Susy CoteIsaiah Nengo Andrew Hill and Brenda Benefit K McNulty
acknowledges the evolutionary anthropology faculty and studentsat Durham University for their helpful comments on an earlyversion of this manuscript and for their gracious hospitality Weare likewise grateful to the anonymous reviewers associate editorand editor whose comments strengthened the paper This manu-script is publication 1 supporting Research on East AfricanCatarrhine and Hominoid Evolution (REACHE)
Appendix A Supplementary data
Supplementary data related to this article can be found at httpdxdoiorg101016jjhevol201503009
References
Andrews P 1974 New species of Dryopithecus from Kenya Nature 249 188e190Andrews P 1978 A revision of the Miocene Hominoidea of East Africa Bull Brit
Mus (Nat Hist) Geol 30 85e225Andrews P 1985 Family group systematics and evolution among catarrhine pri-
mates In Delson E (Ed) Ancestors The Hard Evidence Liss New Yorkpp 14e22
Andrews P Harrison T Martin L Pickford M 1981 Hominoid primates from anew Miocene locality named Meswa Bridge in Kenya J Hum Evol 10123e128
Andrews P Begun D Zylstra M 1997 Interrelationships between functionalmorphology and paleoenvironments in Miocene hominoids In Begun DWard C Rose M (Eds) Function Phylogeny and Fossils Miocene HominoidEvolution and Adaptations Plenum Press New York pp 29e58
Andrews P Molleson T 1979 The provenance of Sivapithecus africanus Bull BritMus (Nat Hist) Geol 32 19e23
Begun DR 2001 African and Eurasian Miocene hominoids and the origins of theHominidae In de Bonis L Koufos GD Andrews P (Eds) Hominoid Evolutionand Climatic Change in Europe Phylogeny of the Neogene Hominoid Primatesof Eurasia Cambridge University Press London pp 231e253
Begun DR 2007 Fossil record of Miocene hominoids In Henke W Tattersall I(Eds) Handbook of Paleoanthropology Springer Berlin pp 921e977
Begun DR Teaford MF Walker AC 1994 Comparative and functional anatomyof Proconsul phalanges from the Kaswanga primate site Rusinga Island KenyaJ Hum Evol 26 89e165
Bishop WW 1964 More fossil primates and other Miocene mammals from North-East Uganda Nature 203 1327e1331
Bishop WW Miller JA Fitch FJ 1969 New potassium-argon age determinationsrelevant to the Miocene fossil mammal sequence in East Africa Am J Sci 267669e699
Blankenship JA Mincer HH Anderson KM Woods MA Burton EL 2007Third molar development in the estimation of chronologic age in Americanblacks as compared with whites J Forensic Sci 52 428e433
Bosler W 1981 Species groupings of early Miocene dryopithecine teeth from EastAfrica J Hum Evol 10 151e158
Coleman MN 2008 What does geometric mean mean geometrically Assessingthe utility of geometric mean and other size variables in studies of skullallometry Am J Phys Anthropol 135 404e415
Cote S Malit N Nengo I 2013 Additional mandibles of Rangwapithecus gordonian early Miocene catarrhine from the Tinderet localities of Western KenyaJ Hum Evol 153 341e352
Deane AS 2009 Early Miocene catarrhine dietary behavior the influence of theRed Queen Effect on incisor shape and curvature J Hum Evol 56 275e285
Drake R Van Couvering JA Pickford M Curtis G Harris JA 1988 New chro-nology for the Early Miocene mammalian faunas of Kisingiri Western KenyaJ Geol Soc Lond 145 479e491
Evernden JF Savage DE Curtis GH James GT 1964 Potassium-argon datesand the Cenozoic mammalian chronology of North America Am J Sci 262145e198
Gebo DL MacLatchy L Kityo R Deino A Kingston J Pilbeam D 1997A hominoid genus from the Early Miocene of Uganda Science 276 401e404
Geoffroy Saint-Hilaire E 1812 Tableau des quadrumanes 1 Ord QuadrumanesAnnls Mus Hist Nat Paris 19 85e122
Gray JE 1825 Outline of an attempt at the disposition of the Mammalia into tribesand families with a list of the genera apparently appertaining to each tribeAnnls Philos 10 337e344
Harrison T 1982 Small-bodied apes from the Miocene of East Africa PhDDissertation University of London
Harrison T 1986 New fossil anthropoids from the middle Miocene of East Africaand their bearing on the origin of the Oreopithecidae Am J Phys Anthropol 71265e284
Harrison T 1987 The phylogenetic relationships of the early catarrhine primates areview of the current evidence J Hum Evol 16 41e80
Harrison T 2002 Late Oligocene to middle Miocene catarrhines from Afro-ArabiaIn Hartwig WC (Ed) The Primate Fossil Record Cambridge University PressCambridge pp 311e338
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 61
Harrison T 2010 Dendropithecoidea Proconsuloidea and Hominoidea (CatarrhiniPrimates) In Werdelin L Sanders WJ (Eds) Cenozoic Mammals of AfricaUniversity of California Press Berkeley pp 429e469
Harrison T Andrews P 2009 The anatomy and systematic position of a newspecies of early Miocene proconsulid from Meswa Bridge Kenya J Hum Evol56 479e496
Hill A Nengo IO Rossie JB 2013 A Rangwapithecus gordoni mandible from theearly Miocene site of Songhor Kenya J Hum Evol 65 490e500
Hopwood AT 1933 Miocene primates from Kenya J Linn Soc Zool 38 437e464Ishida H Kunimatsu Y Takano T Nakano Y Nakatsukasa M 2004 Nacholapi-
thecus skeleton from the Middle Miocene of Kenya J Hum Evol 46 69e103Jolicouer P 1963 The multivariate generalization of the allometry equation Bio-
metrics 19 497e499Jungers WL Falsetti AB Wall CE 1995 Shape relative size and size-
adjustments in morphometrics Am J Phys Anthropol 38 (S2) 137e161Kavanagh KD Evans AR Jernvall J 2007 Predicting evolutionary patterns of
mammalian teeth from development Nature 449 427e433Kelley J 1986 Species recognition and sexual dimorphism in Proconsul and
Rangwapithecus J Hum Evol 15 461e495Kelley J 1995 Sex determination in Miocene catarrhine primates Am J Phys
Anthropol 96 390e417Kelley J 1993 Taxonomic implications of sexual dimorphism in Lufengpithecus In
Kimbel WH Martin LB (Eds) Species Species Concepts and Primate Evo-lution Springer New York pp 429e458
Kent PE 1942 The country round the Kavirondo Gulf of Victoria Nyanza Geogr J100 22e31
Kordos L Begun DR 2001 A new cranium of Dryopithecus from RudabanyaHungary J Hum Evol 15 689e700
Le Gros Clark WE Leakey LSB 1950 Diagnoses of East African Miocene Homi-noidea Q J Geol Soc Lond 105 260e263
Le Gros Clark WE Leakey LSB 1951 The Miocene Hominoidea of East Africa InFossil Mammals of Africa No 1 British Museum (Natural History) London
Leakey LSB 1943 A Miocene anthropoid mandible from Rusinga Kenya Nature152 319e320
Leakey LSB 1963 East African fossil Hominoidea and the classification within thissuper-family In Washburn SL (Ed) Classification and Human EvolutionAldine Chicago pp 32e49
Leakey LSB 1967 Notes on the mammalian faunas from the Miocene and Pleis-tocene of East Africa In Bishop WW Clark JD (Eds) Background to Evolu-tion in Africa University of Chicago Press Chicago pp 7e29
Leakey RE Leakey MG 1986 A new Miocene hominoid from Kenya Nature 324143e146
Leakey RE Leakey MG Walker AC 1988 Morphology of Afropithecus turka-nensis from Kenya Am J Phys Anthropol 76 289e307
Leakey MG Ungar PS Walker A 1995 A new genus of large primate from theLate Oligocene of Lothidok Turkana District Kenya J Hum Evol 28519e531
Linnaeus C 1758 Systema naturae per regna tria naturae secundum classesordines genera species cum characteribus differentris synonymis locis tenthed Laurentii Salvii Stockholm
MacInnes DG 1943 Notes on the East African Miocene primates J E Afr UgandaNat Hist Soc 17 141e181
MacLatchy L 2004 The oldest ape Evol Anthropol 13 90e103MacLatchy L Bossert WH 1996 An analysis of the articular surface distribution of
the femoral head and acetabulum in anthropoids with implications for hipfunction in Miocene hominoids J Hum Evol 31 425e453
MacLatchy L Rossie JB 2005 The Napak hominoid still Proconsul major InLieberman DE Smith RJ Kelley J (Eds) Interpreting the Past Essays onHuman Primate and Mammal Evolution in Honor of David Pilbeam Brill Ac-ademic Publishers Boston pp 15e28
Martin L 1981 New specimens of Proconsul from Koru Kenya J Hum Evol 10139e150
McCollum MS Peppe DJ McNulty KP Dunsworth HM Harcourt-Smith WEH Andrews AL 2013 Magnetostratigraphy of the early MioceneHiwegi Formation (Rusinga Island Lake Victoria Kenya) Geological Society ofAmerica Abstracts with Programs 45 p 12
McNulty KP 2010 Apes and tricksters the evolution and diversification ofhumans closest relatives Evol Edu Outreach 3 322e332
McNulty KP Harcourt-Smith WEH Dunsworth HM 2007 New primate fossilsfrom Rusinga Island Kenya Am J Phys Anthropol 132 (S44) 170
McNulty KP MacLatchy L Peppe DJ Nengo I Manthi FK Miller ERStevens NJ Cote S Lehmann T Kingston J 2014a A regional approach toEast African early Miocene paleobiology J Vert Paleontol 185 SVP Programand Abstracts Book 2014
McNulty KP MacLatchy L Rossie JB Peppe DJ Deino AL Mbua ENManthi FK Nengo IO Stevens NJ Cote S Lehmann T Gutierrez M 2014b
Research on East African catarrhine and hominoid evolution results from thefirst year Am J Phys Anthropol 153 (S58) 182
Michel LA Peppe DJ Lutz JA Driese SG Dunsworth HM Harcourt-Smith WEH Horner W Lehmann T Nightengale S McNulty KP 2014Remnants of an ancient forest provide ecological context for Early Miocenefossil apes Nature Comm 5 3236
Myres JL 1932 Africa Man 32 208Nakatsukasa M Kunimatsu Y Nakano Y Takano T Ishida H 2007 Vertebral
morphology of Nacholapithecus kerioi based on KNM-BG 35250 J Hum Evol52 347e369
Nengo IO Rae TC 1992 New hominoid fossils from the early Miocene site ofSonghor Kenya J Hum Evol 23 423e429
Peppe DJ McNulty KP Cote SM Harcourt-Smith WEH Dunsworth HM VanCouvering JA 2009 Stratigraphic interpretation of the Kulu Formation (EarlyMiocene Rusinga Island Kenya) and its implications for primate evolutionJ Hum Evol 56 447e461
Pickford M 1981 Preliminary Miocene mammalian biostratigraphy for westernKenya J Hum Evol 10 73e97
Pickford M 1986 Sexual dimorphism in Proconsul Hum Evol 1 111e148Pickford M Andrews PJ 1981 The Tinderet Miocene sequence in Kenya J Hum
Evol 10 11e33Pickford M Kunimatsu Y 2005 Catarrhines from the Middle Miocene (ca 145
Ma) of Kipsaraman Tugen Hills Kenya Anthropol Sci 113 189e224Pickford M Senut B Gommery D Musiime E 2009 Distinctiveness of Ugan-
dapithecus from Proconsul Estudios Geol 65 183e241Pilbeam DR 1969 Tertiary Pongidae of East Africa evolutionary relationships and
taxonomy Bulletin of the Peabody Museum of Natural History 31 pp 1e185Rae TC 1993 Phylogenetic analysis of proconsulid facial morphology PhD
Dissertation State University of New York at Stony BrookRae TC 1999 Mosaic evolution in the origin of the Hominoidea Folia Primatol 70
125e135Rafferty KL Walker A Ruff C Rose MD Andrews PJ 1995 Postcranial esti-
mates of body weights in Proconsul with a note on a distal tibia of P major fromNapak Uganda Am J Phys Anthropol 97 391e402
Rossie JB MacLatchy L 2013 Dentognathic remains of an Afropithecus individualfrom Kalodirr Kenya J Hum Evol 65 199e208
Senut B Pickford M Gommery D Kunimatsu Y 2000 Un nouveau genredrsquohominoiumlde du Miocene inferieur drsquoAfrique orientale Ugandapithecus major(Le Gros Clark and Leakey 1950) CR Acad Sci Paris 331 227e233
Simons EL Pilbeam DR 1965 Preliminary revision of the Dryopithecinae (Pon-gidae Anthropoidea) Folia Primatol 3 81e152
Smith TM Martin LB Leakey MG 2003 Enamel thickness microstructure anddevelopment in Afropithecus turkanensis J Hum Evol 44 283e306
Stevens NJ Seiffert ER OConnor PM Roberts EM Schmitz MD Krause CGorscak E Ngasala S Hieronymus TL Temu J 2013 Palaeontological evi-dence for an Oligocene divergence between Old World monkeys and apesNature 497 611e614
Suwa G Kono RT Katoh S Asfaw B Beyene Y 2007 A new species of great apefrom the late Miocene epoch in Ethiopia Nature 448 921e924
Szalay FS Delson E 1979 Evolutionary History of the Primates Academic PressNew York
Teaford MF Beard KC Leakey RE Walker A 1988 New hominoid facial skel-eton from the Early Miocene of Rusinga Island Kenya and its bearing on therelationship between Proconsul nyanzae and Proconsul africanus J Hum Evol17 461e477
Van Couvering JAH Miller J 1969 Miocene stratigraphy and age determinationsRusinga Island Kenya Nature 221 628e632
Walker A 1997 Proconsul function and phylogeny In Begun DR Ward CVRose MD (Eds) Function Phylogeny and Fossils Miocene Hominoid Evolu-tion and Adaptation Plenum Press New York pp 209e224
Walker A Pickford M 1983 New postcranial fossils of Proconsul africanus andProconsul nyanzae In Ciochon RL Corruccini RS (Eds) New Interpretationsof Ape and Human Ancestry Plenum Press New York pp 325e351
Walker A Teaford M 1988 The Kaswanga primate site an Early Miocene hom-inoid site on Rusinga Island Kenya J Hum Evol 17 539e544
Walker A Teaford M 1989 The hunt for Proconsul Sci Am 260 76e82Walker A Teaford MF Martin L Andrews P 1993 A new species of Proconsul
from the early Miocene of RusingaMfangano Islands Kenya J Hum Evol 2543e56
Ward CV Ruff CB Walker A Rose MD Teaford MF Nengo IO 1995 Func-tional morphology of Proconsul patellas from Rusinga Island Kenya with im-plications for other Miocene-Pliocene catarrhines J Hum Evol 29 1e19
Ward S Brown B Hill A Kelley J Downs W 1999 Equatorius a new hominoidgenus from the Middle Miocene of Kenya Science 285 1382e1386
Werdelin L Sanders WJ 2010 Cenozoic Mammals of Africa University of Cali-fornia Press Berkeley

KP McNulty et al Journal of Human Evolution 84 (2015) 42e6144
mandibular representation whereas ldquoUrdquo legetetensis was repre-sented mostly by mandibles and lower teeth (Pickford et al 2009)
Table 1 summarizes the relevant taxonomic allocations used bydifferent authors according to the geographic distribution of lo-calities In its broadest conception (eg Harrison and Andrews2009) the genus Proconsul is quite speciose relative to otherMiocene hominoid genera including as many as six distinct spe-cies This number is substantially reduced under the taxonomicscheme proposed by Pickford et al (2009) wherein half of thesespecies are referred to ldquoUgandapithecusrdquo Those preferring tomaintain all of this material in Proconsul however have noted thatthe greatest distinction within that sample is between the Kisingiriassemblages on the one hand and those from Tinderet and Ugandaon the other (Walker et al 1993 Begun 2001 2007MacLatchy andRossie 2005 Harrison and Andrews 2009 Harrison 2010)daninterpretation presaged in some ways by the taxonomy proposedby Kelley (1986) and Pickford (1986) and ultimately echoing theoriginal presentation by LSB Leakey (Myres 1932)
In accordancewith other researchers (eg Harrison 2002 2010MacLatchy 2004 MacLatchy and Rossie 2005 Suwa et al 2007Harrison and Andrews 2009 McNulty 2010 Hill et al 2013) wedo not recognize the generic distinction of ldquoUgandapithecusrdquo fromProconsul This is due in part to our findings detailed below of adistinct clade uniting P africanus with species assigned to ldquoUgan-dapithecusrdquo In addition the most recent characterization of
Table 1Identifications of Proconsul (sensu lato) species by different authors listed byregiona
Authors KisingiriLocalities
TinderetLocalities
UgandanLocalities
Hopwood 1933 e P africanusb e
MacInnes 1943 P africanus P africanus e
Le Gros Clark andLeakey 1950
P africanus P africanus e
P nyanzaeb P majorb
Andrews 1978 P africanus P africanus P majorP nyanzae P majorP major
Bosler 1981 P africanus P africanus P majorP nyanzae P nyanzaeP major P major
Kelley 1986 P nyanzae P africanus P majorP major
Pickford 1986 P nyanzae P africanus P majorP major
Walker et al 1993 P heselonib P africanus P majorP nyanzae P major
Senut et al 2000 P nyanzae P africanus U majorU majorc
Harrison andAndrews 2009d
P heseloni P africanus P majorP nyanzae P major
P meswaeb
Pickford et al 2009 P heseloni P africanus U majorP nyanzae U major U legetetensis
U legetetensisb U gitongaiU meswaec
This paperd E heselonic P africanus P majorE nyanzaec P major
P meswae
a Very small samples from other regions (eg Buluk) that have been attributed toProconsul (sensu lato) are not included here as they are not critical to understandingthe placement of Kisingiri specimens
b New genus andor species named in publicationc Existing species transferred to a new genusd Proconsul gitongai from the middle Miocene Muruyur Formation (Kipsaraman)
is also recognized but not at the early Miocene localities assessed in this paper
ldquoUgandapithecusrdquo which removed most Tinderet specimens fromProconsul and distributed them among new and existing species ofldquoUgandapithecusrdquo was based largely on contrasts with Rusingaspecimens rather than with the type species of Proconsul (Pickfordet al 2009) This is particularly true for mandibles and the lowerdentitiondspecimens less securely tied to P africanus because thetype specimen M 14084 is a partial maxilla (eg Bosler 1981) Weargue here that if specimens are sorted without preconceived taxo-nomic assumptions the purported diagnostic features of ldquoUganda-pithecusrdquo (sensu Pickford et al 2009) serve instead to differentiateall specimens in the Tinderet and Uganda samples of Proconsul(Proconsul sensu stricto) from those found at Kisingiri localities (cfKelley 1986 Walker et al 1993 Begun 2001 2007 MacLatchy andRossie 2005 Harrison and Andrews 2009 Harrison 2010)
Geochronology of Proconsul localities
The chronological framework of Proconsul localities provides animportant context for understanding historical and current in-terpretations of species distributions and relationships Early re-searchers while acknowledging the limitations of theirstratigraphic and faunal correlations were nevertheless able tocorrectly position the Western Kenyan localities within the lowerMiocene associating some of the deposits with the Burdigalian astage defined by marine sediments in Europe and dated betweenabout 16 and 205 Ma (Kent 1942 MacInnes 1943) The applicationof radiometric dating techniques helped refine these estimateswith some of the earliest KeAr studies conducted on East Africanlocalities associated with Proconsul (Evernden et al 1964 Bishopet al 1969 Van Couvering and Miller 1969) These datesconfirmed an early Miocene age for most localities with estimatesranging between 23 and 16 Ma However the Kisingiri depositsyielded discrepant results due to post-depositional alteration of thedatable biotites (Drake et al 1988) Re-sampling and further ana-lyses yielded more satisfactory and repeatable age estimates of179e178 Ma but notably these were based on only five samplesconfined to a small segment of the Rusinga geological section(Drake et al 1988)
Radiometric dates in conjunction with studies of the associatedfaunal communities (eg Pickford 1981) created a geochronolog-ical framework for Proconsul wherein the Tinderet localities areoldest dating to perhaps 225 Ma at Meswa Bridge (see review inHarrison and Andrews 2009) with Napak in Uganda beingapproximately contemporaneous with Songhor and Koru (Bishopet al 1969 Pickford and Andrews 1981) and the Kisingiri local-ities preserving the youngest sediments (Pickford1981 Drake et al1988) However themost recent re-dating of Kisingiri sites presentsa more complex scenario Rather than being younger than all of theTinderet sites the Kisingiri stratigraphic sequence is substantiallylonger than previously thought with the oldest fossil stratacontemporaneous with those from Koru and Legetet Hill atapproximately 20e19 Ma (McCollum et al 2013) The youngestMiocene fauna from Rusinga is not associated with radiometricdates butmagnetostratigraphic analysis suggests an age near 17Maor younger (Peppe et al 2009) Hence differences between speciesof Proconsul and in particular between assemblages from TinderetandKisingiri cannot be simply explained bychronology Indeed thegreat length of the Kisingiri sequence suggests that taxonomicuniformity throughout this timeperiodmight require re-evaluation
Aims of this study
Motivated by new specimens analyses and interpretations ofProconsul thathave appeared in the last severalyearsweundertookathorough review of the craniodental and mandibular specimens
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 45
attributed to this genus and here provide an alternate taxonomicinterpretation to those in current use Our primary objective was toevaluate the morphological homogeneity of the genus Proconsul astraditionally constituted (Proconsul sensu lato) and explore thepossibility that theKisingiri specimens formadistinct group (Walkeret al1993 Begun 2001 2007MacLatchyandRossie 2005Harrisonand Andrews 2009) While we agree with Pickford et al (2009) thatthe material traditionally referred to Proconsul exceeds what can becomfortably accommodated within a single genus our resultsconform to the suggestion proposed by Begun (2001 2007) and laterby MacLatchy and Rossie (2005) and Harrison and Andrews (2009)that the Kisingiri specimens from Rusinga and Mfangano Islandsconstitute a distinct cladewith respect to Proconsul from theTinderetand Ugandan localities Our assessment of the taxonomic andphyletic distinction of the Kisingiri material necessitates genericdifferentiation of this sample from Proconsul
Systematics
Order Primates Linnaeus 1758Infraorder Catarrhini E Geoffroy Saint-Hilaire 1812Superfamily Hominoidea Gray 1825Family incertae sedisEkembo gen novurn lsidzoobankorgactB11CECF0-9DD1-4265-9C06-A60E4BEE974E
Synonymy
1950 Proconsul Hopwood Le Gros Clark and Leakey 2601965 Dryopithecus (Proconsul) (Hopwood) Simons and Pilbeam
1201978 Proconsul Hopwood Andrews 901986 Proconsul Hopwood Kelley 4791993 Proconsul Hopwood Walker et al 512010 Proconsul Hopwood Harrison 437
Type species
Ekembo nyanzae comb nov (Le Gros Clark and Leakey 1950)
Included species
Ekembo nyanzae (Le Gros Clark and Leakey 1950) Ekemboheseloni comb nov (Walker et al 1993)
Etymology
Ekembo means ldquoaperdquo or ldquomonkeyrdquo in the Suba language whichwas historically spoken by peoples in western Kenya includingthose who settled Rusinga and Mfangano Islands (Jack Wanyendepers comm)
Diagnosis
Medium- to large-bodied and sexually dimorphic hominoidcharacterized by the following combination of features Frontalbone moderately wide at the coronal suture narrowing anteri-orly toward gracile supraorbital rims which are not demarcatedby a post-toral sulcus Large frontal sinuses behind a prominentglabella Minor postorbital constriction Temporal lines promi-nent and situated high on frontal and parietal bones convergingposteriorly without forming a sagittal crest (in female
specimens) Nasal bones long and thin Broad interorbital regionflat rather than projecting Upper face more vertically orientedrelative to the mid- and lower face Nasolacrimal duct positionedwithin a broad orbit Rostrum broad with premaxilla extendingup to the nasal bones Canine jugum prominent in both sexes butcanine fossa developed only in males Nasal aperture broad withwidest portion inferior and sometimes near the base Inferioraperture margin concave or incised in the midline Nasoalveolarclivus intermediate in height between hylobatids and hominidsClivus does not overlap palatine process of the maxilla pre-senting typical mammalian incisive fenestration ante-roposteriorly positioned at or posterior to mid-canine Maxillarysinus extensive reaching anteriorly to P3P4 junction Zygomati-coalveolar crest arises low on the face near M2 Nearly verticalmalar region Broad anterior palate A single palatine foramenpositioned near the M2M3 junction Broad mandibular ramusangled slightly posteriorly from vertical leads to a graduallyrounded and gracile gonial region Mandibular corpora rangefrom short and gracile to taller and moderately robust Sym-physis characterized by moderately developed superior trans-verse torus and variably developed inferior transverse torus andgenioglossal pit Symphyseal planum and subplanum regionvertically or nearly vertically oriented A single mental foramenpositioned near the P3P4 junction approximately halfway up thecorpus or higher
Upper central and lateral incisors heteromorphic Centralincisor characterized by a narrow lingual tubercle often consist-ing of two or three narrow ridges and distinct but thin marginalridges that are continuous with the lingual cingulum with themesial marginal ridge joining the cingulum at a sharp angle incontrast to a more continuously curving transition from cingulumto distal marginal ridge resulting in an asymmetric ridge andcingulum morphology Lateral incisors have well-developedincisal edges and are asymmetric with a moderately developedlingual cingulum Narrow lower incisors with central and lateralincisors having similar morphology Upper and lower caninesstrongly sexually dimorphic with only slight bilateral compres-sion and moderately low-crowned in relation to basal crown di-mensions with apices terminating at a simple point Canine rootssmooth and conical Cheek tooth cusps low and broad and con-nected by inflated intercuspal crests Upper premolars morpho-logically similar with well-developed mesial foveae onlymoderate cusp heteromorphy and cusps positioned at theapproximate mesiodistal midpoint of the crown resulting innearly symmetric buccal profiles Molars with moderate bucco-lingual flare and reduced cingulum development Upper first andsecond molars approximately square with nearly straight distalmargins and buccally oriented posthypocone cristae Lower firstand second molars rectangular with M1 substantially narrowerthan M2 Upper and lower third molars vary in degree of distalcusp development
Forelimbs and hind limbs of similar length Long vertebral col-umn with six lumbar vertebrae characterized by transverse pro-cesses that arise from the dorsal margin of the vertebral centrumand prominent accessory processes for erector spinae musclesPossessing a coccyx rather than a tail Thorax long narrow anddorsoventrally deep Broad flat sternebrae Pelvis long and narrowwith sagittally oriented ilia an elongate ischium lacking osseousevidence for the presence of callosities and a narrow sacrumwith asmall sacro-iliac joint Scapula positioned laterally on thorax with anarrow ovoid and ventrally oriented glenoid fossa Humerus witha posteriorly directed head retroflexed shaft with little torsion anda distal articular surface lacking a well-differentiated trochlea andcapitulum and having a narrow zona conoidea and shallowtrochlear notch but lacking an entepicondylar foramen Radius
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6146
with a small ovoid head and flat distal articular surface that mainlycontacts the scaphoid Ulna displays a large olecranon processnarrow semilunar notch with only a slight keel anteriorly posi-tioned radial notch and a long distal styloid process that retainsarticulation with triquetrum and pisiform Carpals relatively nar-row and include an unfused os centrale a hamate with a smallhamulus and a flat medially oriented triquetral surface Handcharacterized by straight metacarpals with narrow bases andheads Pollex well developed Manual and pedal phalanges gener-ally similar to each other but less so than in Old World monkeysFemur slender with a small head long neck with a high neck-shaftangle centrally placed fovea capitis reduced greater trochanterand a broad distal end with the medial condyle slightly larger thanthe lateral Broad flat patella Tibia long and slender but fibularobust Tarsals and metatarsals elongate with a robust hallux
Differential diagnosis
Ekembo can be readily distinguished from all other medium-and large-bodied catarrhine genera from the early Miocene Cra-nial mandibular and dental differences between Ekembo andProconsul are described in detail below and summarized inTable 2 Ekembo also differs from Proconsul in having a rounderradial head and a femur with a shorter greater trochanter poste-riorly projecting lesser trochanter less extensive inter-trochantericcrest more robust phalanges with more strongly developedfibrous flexor sheath ridges and broader proximal and interme-diate phalangeal distal joint surfaces (condyles) with widertrochlear grooves and less sharply defined trochlear ridges(Harrison 1982 Andrews et al 1997 Senut et al 2000 Pickfordet al 2009)
Ekembo differs from Afropithecus in having a more inflatedglabellar region no frontal trigon less strongly developed anteriortemporal ridges less postorbital constriction a stronger mandib-ular superior transverse torus less posterior shallowing of themandibular corpus higher-crowned lower canines in relation tomesiodistal length buccolingually narrower upper premolarsrelative to molars upper premolars with cusps positioned closer tothe mesiodistal midline reduced P3 cusp heteromorphy a moresymmetric I2 and substantially less buccolingual flare in the cheekteeth (Leakey et al 1988 Rossie and MacLatchy 2013)
Ekembo differs from Heliopithecus in having buccolinguallynarrower premolars relative to molars premolar cusps positionedcloser to the mesiodistal midline and reduced P3 cusp hetero-morphy (Harrison 2010)
Ekembo differs from Equatorius in having a more gracilemandible a larger superior but smaller inferior mandibular trans-verse torus labiolingually narrower lower incisors with straightermesial and distal margins higher-crowned lower canines in rela-tion to mesiodistal length greater molar cingulum development aless posteriorly directed humeral medial epicondyle a more ante-riorly directed radioulnar joint and narrower metacarpal heads(Ward et al 1999)
Ekembo differs from Nacholapithecus in having a shorter sub-nasal clivus a more expansive incisive foramen a broader nasalaperture at the base a minimally developed mandibular inferiortransverse torus a narrower lingual tubercle on I1 higher-crownedlower canines in relation to mesiodistal length a larger humeralradial fossa a shallower coronoid fossa a less well-developed zonaconoidea a narrower and distally slanted ulnar coronoid process alonger and lower femoral neck a more posteriorly positioned lessertrochanter and a smaller posteromedial tubercle on the talus(Ishida et al 2004)
Ekembo differs from Nyanzapithecus in having more anteriorlyextended maxillary sinuses a relatively taller mandibular corpus
less buccolingually compressed upper and lower canines with lesscurved crowns lower and more rounded cusps with more inflatedocclusal crests on the cheek teeth P3 and P4 that more closelyresemble each other and have much less cingular development abroader P4 relative to M1 M1e2 occlusal outline more closelyapproximating a square and lower molars that are broader relativeto mesiodistal length with a more mesially placed entoconid rela-tive to the hypoconid (Harrison 1986 2010)
Ekembo differs from Rangwapithecus in having a broad andexpansive palatal fenestration a wider palate relative to length amore robust mandible with a well-defined superior transversetorus broader upper and lower incisors relative to height lessbuccolingually compressed canines without scimitar-shapedcrowns upper premolars with reduced cingulum and cusps posi-tioned near the mesiodistal midpoint resulting in expanded mesialfoveae and symmetric buccal profiles a broader P4 relative to M1upper and lower molars that are buccolingually broader relative tolength and have more rounded cusps and inflated occlusal crestsmore squared upper molars with substantially less cingulumdevelopment and lower molars with a more mesially placedentoconid (Andrews 1974 Harrison 2002 2010 Cote et al 2013Hill et al 2013)
Ekembo differs from Turkanapithecus in having an expandedglabellar region less postorbital constriction a narrower temporalfossa with less flaring zygomatic arches the lacrimal fossa withinthe orbital margin a relatively broader palate with non-convergingtooth rows narrower nasal bones and interorbital region a distinctmandibular superior transverse torus a more robust mandibularcorpus taller mandibular ramus upper canines more rounded incross-section a reduced cingulum on cheek teeth awell-developedhypocone on M1e2 a more squared M2 with a protocone subequalto paracone and broader lower molars relative to length (Leakeyand Leakey 1986 Harrison 2002 2010)
Ekembo differs from Mabokopithecus in having a relativelybroader M3 crown without distal recurvature of the crown andlacking a buccal concavity a hypoconulid that is in-line with thebuccal cusps more bunodont cusps a well-defined protocristidand a broad open talonid without an accessory cuspid (Harrison2002 2010)
Ekembo differs from Xenopithecus in having reduced uppermolar flare a substantially reduced cingulum no prehypoconecrista molar occlusal profiles more closely approximating a squareand less constricted trigons (Hopwood 1933 Le Gros Clark andLeakey 1951 Harrison 2002 2010)
Ekembo differs from Morotopithecus in having a broader inter-orbital region a better developed canine fossa in males premolarssmaller relative to molars greatly reduced or absent buccalcingula on upper molars lumbar vertebrae with transverse pro-cesses positioned more ventrally on the centrum and a narrowerdistal femur with a deeper patellar groove and similarly-sizedmedial and lateral femoral condyles (Gebo et al 1997MacLatchy 2004)
Recently Stevens et al (2013) likened the new genus Rukwapi-thecus represented by a mandible fragment with well-preservedP4-M3 to nyanzapithecine species Ekembo can be distinguishedfrom this genus in having a more gradually rounded gonial angle aless obliquely oriented P4 with subequal protoconid and metaconidand a broader mesial fovea less mesially positioned metaconidreduced buccal cingula and smaller mesial foveae on the molarssmaller hypoconulids broader M1e2 relative to length a singularrather than twinned metaconid no accessory cuspules in thelingual notch and a well-defined crest between entoconid andhypoconulid
Family Proconsulidae (Leakey 1963)
TabDen
Fe
G
U
U
U
U
U
M
Lo
Lo
Lo
a
b
c
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 47
Genus Proconsul Hopwood 1933
Type species
Proconsul africanus Hopwood 1933
Included species
Proconsul africanus (Hopwood 1933) Proconsul major (Le GrosClark and Leakey 1950) Proconsul gitongai comb nov (Pickfordand Kunimatsu 2005) Proconsul meswae (Harrison and Andrews2009)
Diagnosis (replaces the emended diagnosis of Harrison 2010)
Proconsul species are medium- to large-bodied sexually dimor-phic catarrhines that can be distinguished from other proconsulidgenera by the following combination of features Robust mandib-ular corpus and symphysis characterized by a long shallowlysloping planum alveolare terminating at a large superior transversetorus and a subplanum surface oriented posteroinferiorly Upper
le 2tognathic features that distinguish Ekembo from Proconsula
ature Ekembo
eneralCusp morphology More bunodont occupying more of theMolar crests Inflated incorporated into cuspsCingulum Reducedpper central incisorLingual pillar Narrow generally low reliefMarginal ridges and cingulum Asymmetric with sharply angled cingulu
ridge transitionpper canineTip PointRoot fluting AbsentRoot striations Rare and weakly developedpper third premolarb
Mesiodistal cusp position Crown midpointMesial fovea Well developedBuccal profile SymmetricMesial crown margin Straight or slightly curvedDistal crown margin Slight to moderately curvedCusp heteromorphy ReducedCrown shape Converges on P4 shapeMesiobuccal flange Nopper fourth premolarParacone height Taller relative to crown lengthpper second molarc
Occlusal crown shape SquarishDistal margin Straight to slightly curvedBuccal cingulum Absent or rudimentaryDistolingual cingulum Merges into hypoconePosthypocone crista Distobuccally to buccally orientedBuccolingual flare ReducedandiblePlanum alveolare Short vertically inclinedSuperior transverse torus Moderate to well-developedOrientation of subplanum area More vertically orientedCorpus robusticity More gracilewer incisorsCrown shape Narrow relative to heightwer canineTip Pointwer molarsBuccolingual flare ReducedM1 M2 breadth M1 narrower relative to M2
Although presented here as discrete characteristics for descriptive purposes some oNearly all of these features tend to distinguish P4s as well but in that tooth the diffeThese features tend to distinguish M1s as well but in that tooth the differences are
incisors heteromorphic with I1 exhibiting an inflated lingual tu-bercle and a near symmetrical configuration of lingual marginalridges and cingulum with a curving transition from cingulum tothe mesial marginal ridge Lower incisors broad relative to crownheight Upper and lower canines with distinct blade-like or burin-like tips in both deciduous and adult teeth Upper canine rootswith broad longitudinal grooves or fluting defining two or threedistinct columns plus numerous small longitudinal striationsextending cervically from the root apex Upper P3s morphologicallydissimilar from P4s P3 has substantial cusp heteromorphy (para-cone much taller than protocone) with mesially positioned cuspsand a notched mesial border that together limit or eliminate amesial fovea A long steeply angled preparacrista contributes to ahighly asymmetric buccal profile before being interrupted near thetooth cervix by a mesially projecting flange The distal margin isstrongly curved P4 has a tall paracone relative to crown lengthMolars characterized by extensive cingular development individ-uated conical cusps connected by sharp occlusal crests and sub-stantial buccolingual flare Upper molars rhomboid in shaperesulting from a broad trigon and a distally expanded hypocone M2
has a distinct buccal shelf between paracone and metacone a dis-tolingual cingulum that wraps fully around the hypocone rather
Proconsul
occlusal surface area More distinct individuated conesSharper more distinct from cuspsExtensive
Inflatedbulbousm-mesial More symmetric with curving cingulum-mesial
ridge transition
Blade-likePresentVery common and well developed
MesialAbsent or reducedAsymmetricNotchedStrongly curvedStrongDistinct from P4
Yes
Shorter relative to crown length
RhomboidStrongly curvedDistinct cingular shelfDistinct from hypoconeDistally to distobuccally orientedPronounced
Long more horizontally orientedStrongly developedMore posteriorly inclinedMore robust
Broad relative to height
Blade-like
PronouncedM1 more similar to M2
f these features are undoubtedly correlated aspects of single evolutionary changesrences are not as pronouncednot as pronounced
Figure 2 Upper central incisors of Proconsul and Ekembo Lingual views of Ekembo(KNM-RU 1685 cast) left and Proconsul (KNM-CA 1300) right Note the differences inthe transition between the lingual cingulum and the mesial marginal ridge which ismore rounded in Proconsul and more angled in Ekembo The central pillar is also morebulbous or inflated and without ridging in Proconsul Additional specimens illustratingthe Proconsul morphology are shown in Pickford et al (2009 Fig 17 p 197) butreferred in that manuscript to Ugandapithecus
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6148
thanmerging into the hypocone on its lingual surface and a distallyoriented posthypocone crista Lower first and second molars nearlyequal in buccolingual breadth
Comparative morphology of Ekembo and Proconsul
Samples and comparative methods
The above diagnoses are based on extensive survey and analysisof the original fossil material Specimens housed in the NationalMuseums of Kenya (NMK) and the Natural History Museum (Lon-don) were analyzed by the authors during several separate and twojoint (NMK) research visits Fossils in the Uganda National Museumwere assessed by one of us (KPM) during two visits All measure-ments were collected by KPM Dental terminology follows Szalayand Delson (1979)
The following results are organized with respect to themorphology of the Kisingiri sample (attributed to E nyanzae andE heseloni) versus that of Proconsul from Tinderet and Napak(traditionally referred to P africanus P major and more recentlyP meswae) Specimens from additional localities that have at onetime been attributed to Proconsul (eg Andrews1978 Pickford andKunimatsu 2005 [cf Harrison 2010]) are for the most part notconsidered here most are middle Miocene in age and have beentransferred to other genera (Harrison 2002 2010) Of those that arefrom early Miocene sites there is growing evidence that P majorcan also be found at Moroto and differentiated fromMorotopithecus(Pickford et al 2009) an issue of some interest but ultimately notcritical to the taxonomic questions being considered here Whereappropriate however we do provide observations on a few earlyMiocene specimens attributed to ldquoUrdquo gitongai (Pickford andKunimatsu 2005 Pickford et al 2009 see Harrison 2010) Yetwhile we consider the middle Miocene holotype likely to representa distinct species here referred to Proconsul we concur withHarrison (2010) that many specimens placed in this species byPickford et al (2009) cannot be distinguished from P major
Sample sizes particularly from Tinderet and Ugandan localitiestend to be small but where possible standard univariate (eg t-tests) and bivariate (regression ANCOVA) statistical analyses wereperformed A single eigendecomposition of the covariance matrixof symphyseal measurements was done to characterize the multi-variate shape of the mandibular symphysis after accounting for sizedifferences (sensu Jolicouer 1963) Quantitative results are illus-trated as box-and-whisker plots of shape indices or bivariate plotsof the measurements For samples of fewer than five specimensshape index values are plotted in lieu of box-and-whiskersummaries
Morphological comparisons
Upper incisors Two features of the Ekembo upper central incisorsdistinguish them from those of Proconsul including specimenswidely attributed to the type species P africanus The lingual tu-bercle in Ekembo is quite narrow generallywith low relief and oftenscored by two or more irregular ridges in contrast to the muchmore inflated or bulbous tubercle in Proconsul I1s (Fig 2) Ekembospecimens also have a characteristic morphology in the transitionfrom the lingual cingulum to the mesial marginal ridge (Fig 2)from the mesiodistal mid-point of the crown the cingulumextends either directly mesially or even angles slightly cervicallyto intersect the mesial marginal ridge at a fairly sharp angle Incontrast in I1s of Proconsul the cingulum angles incisally from themid-point and transitions into the mesial marginal ridge in agradual curve The expression of these traits varies somewhat inthe smaller Tinderet specimens attributed to P africanus
compared with those of P major (cf Pickford et al 2009)However these variations are relatively minor with respect to thedifferences between the two genera Of 25 I1s known fromKisingiri localities only one specimen KNM-MW 562 somewhatresembles the Tinderet sample in both of these featuresdhavinga somewhat less angled transition from the mesial marginal ridgeto the cingulum and a more bulbous lingual tubercle This isparticularly interesting as the specimen was discovered in theMakira Beds now dated to older than 190 Ma and contemporarywith the Tinderet localities (McCollum et al 2013) Neverthelessits overall morphology is more similar to other Ekembospecimens than to Proconsul incisors Two other Ekembospecimens KNM-RU 1677 and RU 1933 have somewhat moreinflated lingual tubercles but otherwise resemble other Ekembo I1s
No consistent differences were found in upper lateral incisors
Upper canines Upper canine crowns of Ekembo are typical of thoseof most catarrhines in terminating in an evenly tapering point Thisdiffers from the Proconsul specimens which have a blade-like(Kelley 1986 Senut et al 2000 Harrison 2010) or ldquoburin-likerdquo(Pickford et al 2009) canine tip (Fig 3) While blade-like apicalmorphology has long been recognized as a distinctivecharacteristic of P major we confirm the report by Pickford et al(2009) that this feature is also present in the P africanus typespecimen (Fig 3a) although somewhat understated as expectedin the more slender and higher-crowned P africanus caninesUnfortunately other canines attributed to P africanus (eg M44837 KNM SO 419 LG 921 CA 1910 SO 5353 Kelley 1986) havebroken apices and therefore cannot be evaluated for this featureA blade-like tip is also present in the deciduous upper canines ofP major (eg KNM-SO 371 MV 10 NAP IV UMP 62-20) andP meswae (KNM ME 11) Hence the blade-like upper canine tipunites all Tinderet and Napak Proconsul species but is neverexpressed in Ekembo specimens from Rusinga and Mfangano
Two features of the canine roots also differentiate these generalongitudinal grooving or fluting along the length of the root (cfKelley 1986) and the development of numerous finer longitudinal
Figure 3 Canines of Proconsul and Ekembo a) Upper canine of the P africanus holotype (M 14084) lingual view b) Upper canine of P major (KNM-CA 2127) lingual view c) Lowercanine of P major buccal view d) lower canine of Ekembo cf nyanzae (KNM-RU 1676) buccal view and e) Upper canine of P major (KNM-SO 584) distal view Note the blade-like orldquoburin-likerdquo tip of the unworn Proconsul canines including on the nearly unworn canine tip of the P africanus type specimen (a) Ekembo canines have more typical pointed tipsProconsul canines also have longitudinal grooves and fine striations on the root (e) Ekembo lacks these features Scale bars frac14 1 cm
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 49
striations near the root apex that extend to varying degrees towardthe cervix of the tooth Longitudinal ldquoflutingrdquo consists of broadgrooves that segment the root into distinct sections (cf Pickfordet al 2009 Fig 3e) Most often the root in cross-section has theappearance of a cylinder that has been sectioned longitudinallywith the two halves offset from each other but in some cases thereare three segments and the root cross-section is more triangular inappearance (Kelley 1986) This characteristic was noted by Kelley(1986) in the P africanus canines but not in those of P majorHere we affirm that fluting is present on all of the visible uppercanine roots attributed to both P africanus and P major rootmorphology could not be evaluated in the deciduous upper caninefrom Meswa Bridge since it is implanted in the maxillary alveolusNone of the Ekembo specimens display this morphology havinginstead a simpler round to ovoid cross-section along the length ofthe root
The fine longitudinal striations on Proconsul upper canine rootsvary in the degree to which they extend toward the cervix (Fig 3e)but they are visible in all specimens attributed to P africanus(KNM-SO 419 521 921 5353 KNM-X 502) and five of six uppercanines of P major (excepting KNM-SO 1614) This morphology isalso present in UMP 62-12 which was included as part of theMorotopithecus bishopi hypodigm (Gebo et al 1997) Fine striationsalong the upper canine root are almost entirely absent in EkemboTwenty of twenty-three specimens lack this feature three speci-mens (KNM-RU 1763 1845 1891) exhibit incipient to slight
striations but these are never developed to the extent seen inProconsul canines
Upper premolars Upper third premolars display some of thegreatest differences between Ekembo and Proconsul P4s tend todiffer in similar ways but the differences are not nearly as markedEkembo P3s have cusps positioned near the mesiodistal midpoint ofthe crown well-developed mesial foveae pre- and postparacristaeof similar length and slope resulting in symmetric or nearsymmetric buccal profiles a straight or slightly curved mesialmargin and only moderately curved distal margin (Fig 4)Proconsul P3s are more similar in appearance to those of small-bodied catarrhines from the early Miocene with cusps positionedmesially on the crown greatly reducing or eliminating the mesialfovea They also have a steep preparacrista and more shallowlysloped postparacrista resulting in a highly asymmetric buccalprofile and an inwardly angled or notched mesial margin but astrongly curved distal margin This morphology is bestcharacterized in the P africanus holotype (M 14084) but can alsobe seen in other specimens attributed to this species as well asthose referred to P major (Fig 4d e) Also noteworthy is adistinctive mesiobuccal flange that mesially extends the buccalportion of the crown but only near the cervical line of the toothPickford et al (2009) describe this feature in some detail forldquoUgandapithecusrdquo but in fact this morphology is clearly expressedin the P africanus holotype as well (Fig 4a d) A single adult P3
Figure 4 Upper premolars of Proconsul and Ekembo occlusal (aec) and lingual (def) views a) P3e4 of the P africanus holotype (M 14084) b) P3 of P major (M 14331) c) P3e4 of theE heseloni holotype (KNM-RU 2036) d) P3 of the P africanus holotype (M 14084) e) P3 of P major (M 14331) f) P3 of the E heseloni holotype (KNM-RU 2036) Note the mesiallypositioned cusps lack of mesial fovea and strongly asymmetric crown in Proconsul compared to Ekembo (see text for further comparisons) Image of M 14331 reversed for easiercomparison Photo 4a courtesy of Rutger Jansma Scale bars frac14 1 cm
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6150
germ and two dP3s attributed to P meswae look remarkably similarto those of other Proconsul species including the presence of thisflange In contrast the Ekembo P3 has a well-developed mesialfovea that terminates at a mesiobuccal shoulder positionedapproximately halfway between the paracone tip and the cervixFrom this shoulder the mesial margin angles slightly distallydown to the cervix (Fig 4f)
The degree of cusp height heteromorphy in P3 also differentiatesEkembo from Proconsul (contra Pickford et al 2009) Only four non-Kisingiri specimens were sufficiently unworn to take measure-ments but more worn specimens also appear to express this dif-ferential in cusp height Figure 5a shows the ratio of protoconeparacone height measured from the tooth cervix and with only oneexception Ekembo P3s have substantially less cusp heteromorphyThis exception KNM-RU 1677 does not in fact have greater heter-omorphy but instead has an abnormal extension of the enamel ontothe buccal (paracone) root which skews the measurement Hencecusp relief relative to the occlusal surface in RU 1677 is not nearly asheteromoprhic as in Proconsul and when this is taken into accountthe difference between genera is significant (t-test dffrac14 11 t frac14 283p frac14 00165) The adult P3 germ from Meswa Bridge is incompletelyformed and therefore could not be measured However the differ-ence between paracone and protocone heights with respect to theocclusal surface appears to be as great as that in the P africanusholotype It does not appear that sex is an important determinant ofthis feature since well-established male (eg M16647 KNM-RU16000) and female (KNM-RU 2036 7290) specimens do not segre-gate across the Kisingiri range of variation
Upper fourth premolars of Ekembo are broadly similar inmorphology to Ekembo P3s in fact the similarity between P3 and P4
is substantially greater in Ekembo than in Proconsul (Figs 4a c)Likewise most of the morphological differences that discriminatethe P3s of these genera also tend to differentiate the P4s but less
clearly Cusp heteromorphy does not distinguish P4s of thesegenera but the overall height of the P4 paracone in Ekembo issignificantly greater relative to crown length than in Proconsul P4s(t-test df frac14 14 t frac14 316 p frac14 00069 Fig 5b)
Upper molars Cusps of Ekembo upper molars are uniformly morebunodont than those of Proconsul occupying more of the occlusalsurface Intercuspal crests are more fully incorporated into thecusps in Ekembo molars giving them a more inflated appearanceThis is particularly evident in the crista obliqua which is very broadin Ekembo but thin and knife-like in Proconsul (Fig 6) Cusps ofProconsul molars stand more as individual cones Pickford et al(2009) have further differentiated the cusp morphology ofsmaller and larger Tinderet specimens arguing that the latterhave more bulbous cusps While accurate this difference is minorcompared with the morphological difference between Ekemboand Proconsul and we regard the within-Proconsul cuspalvariation as size-related In fact the intermediately sizedP meswae has cusp morphology that is appropriatelyintermediate between that of P africanus and the larger P major
Other differences between Ekembo and Proconsul upper molarsare most evident in M2 We concur with previous assessments thatthe Kisingiri upper molars have less developed cingula than non-Kisingiri molars (eg Senut et al 2000 Harrison 2002 2010Harrison and Andrews 2009) More specifically Ekembo M2s lackor have only rudimentary buccal cingula between the paracone andmetacone Likewise the lingual cingulum is discontinuous merg-ing into the hypocone lingually and then re-emerging as a distalmarginal ridge (Fig 6) Proconsul M2s display substantially morecingular development overall and in particular exhibit a prominentbuccal shelf between the paracone and metacone and a distinctdistolingual cingulum that is continuous around the hypocone towhere it merges with the distal marginal ridge
Figure 5 Box-and-whisker plots a) P3 cusp heteromorphy index (protocone heightparacone height) Results of a t-test show this difference to be significant (p frac14 00165) Theexcluded Ekembo outlier KNM-RU 1677 only appears to have extreme heteromorphy because of extensive expansion of the buccal enamel onto the anterior root (see text) b) P4
relative paracone height (paracone heightmesiodistal crown length) Results of a t-test show this difference to be significant (p frac14 00069)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 51
The occlusal profiles of Ekembo M2s are squarish in shape withrelatively straight and equal sides with a posthypocone crista thatis distobucally to buccally directed In contrast the occlusal profilesof Proconsul M2s have a more rhomboid shape with substantialbasal flare around the mesial cusps and a distally expanded hypo-cone with a distally directed posthypocone crista resulting in astrongly curved distal margin (Fig 6) Even in specimens with lesscurved distal margins the posthypocone crista is still more distallydirected compared to the condition in Ekembo These features arecharacteristic not only of P africanus P meswae and P major butare also expressed in the isolatedM2 fromMoroto Uganda (MOR IIb2098) attributed by Pickford et al (2009) to ldquoUrdquo gitongai butregarded here as belonging to P major
As noted by others (eg Harrison and Andrews 2009) the de-gree of buccolingual molar flare differs among these samplesQuantified as the distance between the tips of the mesial cuspsdivided by maximum crown breadth the degree of molar flare in
Ekembo M2s is significantly less than in Proconsul M2s (t-testdf frac14 29 t frac14 396 p frac14 00004) In a bivariate plot of these twomeasures the distributions of Ekembo and Proconsul teeth barelyoverlap with only KNM-RU 1677 encroaching into Proconsul ter-ritory (Fig 7)
These differences in M2 morphology also tend to distinguishEkembo from Proconsul M1s but not always as clearly Whereas M2smore consistently express distinct morphotypes the M1s in bothgenera show more variation in these features
The degree of upper third molar development (ie whether M3
has four distinct cusps or the distal cusps are reduced or absent) haslong been cited as taxonomically informative for these species orgroups of these species (eg Hopwood 1933 Le Gros Clark andLeakey 1951 Andrews 1978 Pickford et al 2009) However wefound this tooth to be unreliable for taxonomic purposes Withinprimates M3s are typically the most variable molars in both sizeand shape (Blankenship et al 2007 Kavanagh et al 2007) and this
Figure 6 Occlusal view of the M2s of Proconsul and Ekembo Left P africanus holotype(M 14084) Right E heseloni holotype (KNM-RU 2036) Note in Proconsul the overalldifference in crown shape as well as the more conical individuated cusps and nar-rower cristae strongly developed lingual cingulum that continues to the distal aspectof the crown broad buccal cingular shelf and more distally directed posthypoconecrista In Ekembo the cusps are more bunodont and expanded the cristae are broaderwith better developed accessory tubercles the lingual cingulum merges into thehypocone there is no buccal shelf and the posthypocone crista is more buccallydirected
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6152
is observable even in small samples of Proconsul species TheP africanus holotype has a substantially reduced M3 whereas inKNM-CA 2250dattributed by most authors to this species(Andrews 1978 Pickford et al 2009)dit is fully developed Simi-larly the hypodigm of P major includes both fully developed M3s(eg KNM-CA 389 LG 1815) and specimens with reduced ormissing hypocones (KNM-CA 397 1299) Ekembo displays compa-rable ranges of variation from highly reduced to fully developedM3s in both species Thus these species and genera cannot bedistinguished based on M3 development and we did not find anyother distinguishing characteristics between Ekembo and Proconsulin the upper third molars
Figure 7 Scatter plot depicting the de
Mandible Pickford et al (2009) enumerated several traits thatreadily distinguish Kisingiri from non-Kisingiri mandibles andthese distinctions are entirely consistent with the distinction ofEkembo from Proconsul Our analyses specifically underscoredifferences in the symphyseal region The planum alveolare inboth male and female specimens of Ekembo is short and morevertically oriented even the most elongate of these (KNM-RU47805 McNulty et al 2007) is substantially more verticalcompared to specimens of Proconsul In contrast the planumalveolare in Proconsul is conspicuously long and nearlyhorizontally oriented Well-preserved mandibles attributed toP major (eg KNM-SO 396 and 404) exhibit this morphology inextreme fashion (Fig 8) smaller specimens (KNM-SO 1112P africanus) show less extreme expression but are still readilydistinguishable from Ekembo mandibles For its size KNM-LG 452has a surprisingly short planum alveolare though not to thedegree seen in Rusinga mandibles Correlated to the shortplanum alveolare in Ekembo is a reduced superior transversetorus which while well-developed does not approach therobusticity expressed in Proconsul mandibles This difference isreflected in an index of symphyseal robusticity (thicknessperpendicular to the long axis of the symphysislength of thelong axis) which shows Ekembo and Proconsul to be significantlydifferent (t-test df frac14 14 t frac14 430 p frac14 00007)
Development of the inferior transverse torusdoften used todifferentiate among species of Proconsul (eg Pickford et al 2009Harrison 2010)dwas found to be highly variable and inconsis-tently expressed within these samples However the overallorientation of the subplanum region (as defined by a line con-necting the posterior-most midline points of the superior andinferior transverse tori or the posterior-most midline point of theinferior margin of the symphysis in the absence of the latter) doesdistinguish Ekembo from Proconsul mandibles The orientation ofthis line in relation to the alveolar margin in the vicinity of thepostcanine teeth was assessed qualitatively by placing a small rod
gree of buccolingual flare in M2
Figure 8 Mandibles of Proconsul and Ekembo Occlusal views of P major a) KNM-SO 396 b) KNM-SO 404 c) KNM-LG 452 Occlusal views of Ekembo d) KNM-RU 2087 e) KNM-RU1674 f) KNM-RU 7290 Occlusal views of P africanus g) KNM-SO 372 h) KNM-SO 1112 i) Lingual view of P africanus (KNM-SO 1112) Note the long more horizontally orientedplanum alveolare in Proconsul resulting in a pronounced superior transverse torus The planum alveolare is shorter and more vertical in Ekembo such that one can see the inferiormandibular border from occlusal view
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 53
(eg a dental pick) in themidline internally along the two referencepoints and recording the point in lingual perspective at which therod intersected the alveolar margin (Fig 9) In Ekembo the sub-planum region is more vertically oriented so that the rod intersectsthe alveolar margin most often in the premolar field Not surpris-ingly there is a difference between sexes with females having areduced superior transverse torus and therefore a more verticallyoriented subplanum region compared to males In contrast withEkembo Proconsul mandibles have much more inclined subplanumsurfaces with the rod intersecting the lingual alveolar margin inthe vicinity of the M1 (females) or M2 (males Fig 9) When sex istaken into account therefore the distributions of Ekembo andProconsul in this feature are discrete Note that male and femalemandibles attributed to both P africanus and P major are includedin this assessment and are surprisingly similar despite dramaticsize differences
We also quantified internal symphyseal morphology using threemeasures symphyseal height (infradentaleegnathion) planumalveolare length (posterior-most point on the superior transversetoruseinfradentale) and subplanum length (posterior-most pointon the superior transverse torusegnathion) a combination thatincorporates not only the lengths of these intervals but also theangular relationships among them To compare symphyseal shapeacross a range of sizes we chose a simple principal componentsapproach to partition size onto the first eigenvector (cf Jolicouer1963) While this is not the preferred method for partitioning sizein most datasets (Jungers et al 1995) for this particular sample andset of measurements Mosimann shape variables were still
significantly correlatedwith size (see discussion in Coleman 2008)Hence we assessed differences in symphyseal morphology be-tween Ekembo and Proconsul using the second and third principalcomponents of the three variables Distributions of the two generaare distinct in this shape space (Fig 10 Supplementary OnlineMaterial [SOM] Table 1)
Robusticity of the mandibular corpus also differs between thesesamples though it is more difficult to test due to differentialpreservation among specimens Corpus thickness and depth underP4 can be measured in the greatest number of specimens andcomparison of these measures suggests a difference betweengroups in the scaling of corpus robusticity (Fig 11) Among largepresumably male specimens Ekembo mandibles are substantiallyless robust than those of Proconsul However among smallermandibles only a single Proconsul specimen (KNM-SO 372) could beincluded and it is similar in robusticity to Ekembomandibles of thissize Considered together the impression is that corpus robusticitymay scale differently in Ekembo than it does in Proconsul and a testfor differences in slopes using an ANCOVA design of the loggedvariables was significant (df frac14 2 F frac14 2481 p lt 00001) Never-theless additional specimens are needed to properly evaluate theinfluence of taxonomy sex and size on corpus robusticity areasonable alternative interpretation of Figure 11 is that it dem-onstrates a significant difference between P major and all otherspecies of both Proconsul and Ekembo
Lower incisors Ekembo specimens are uniformly narrower inmesiodistal length than incisors attributed to P africanus P major
Figure 10 Scatter plot of principal component scores based on an eigenanalysis of threeposterior point on the superior transverse toruseinfradentale) and subplanum length (mprimarily captures size differences among specimens (Jolicouer 1963 see text and SOM Trespectively) in the symphyseal variables
Figure 9 Line drawing of a hemimandible depicting different orientations of the sub-planum area in Ekembo and Proconsul Females are denoted using shorter vectors maleswith longer vectors Vector is estimated in M 14086 which is broken at the midlineEkembo specimens have a more vertical subplanum compared to those from Songhorand Legetet but within both groups females have more vertically oriented regions thanmales Hence even the most ventrally rotated subplanum in the largest Ekembo malespecimen (KNM-RU 47805) groups with the Tinderet females in this feature
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6154
and P meswae both absolutely and in relation to crown height(Fig 12) This is true for both I1 and I2 A t-test adjusted for non-independent observations (eg I1 and I2 from the same mandible)confirmed the difference as significant (t-test df frac14 12 t frac14 258p frac14 00242) Moreover while the measurable Proconsul samplewas small this same pattern is observable in other specimenswhose tips were slightly to moderately worn and therefore notmeasured
Lower canines The only lower canine trait that discretely sepa-rates these genera is the pointed crown tip in Ekembo (Fig 3d)contrasted with a blade-like (or ldquoburinrdquo) tip in Proconsul(Fig 3c) This feature is well established for P major but is alsoexpressed in KNM-CA 1772 which was referred to P africanus byKelley (1986) and is too small to be either P major or P meswaeTwo other specimens commonly attributed to P africanus KNM-CA 2149 and KNM-SO 1112 both have broken canine tipsLikewise a new specimen (Kor 65004) attributed to P africanusby Pickford et al (2009) also has a broken tip and cannot beevaluated (see Pickford et al 2009 Fig 23) As with the uppercanines the blade-like tip is also present in the deciduous lowercanines of P major (eg KNM-CA 361 S0 5352) and P meswae(KNM-ME 1)
Lower premolars Like most previous authors (eg Andrews 1978Harrison 2002 2010 Pickford et al 2009) we found neitherqualitative nor quantitative characteristics of the lower premolarswith any taxonomic value
Lower molars Differences in lower molar morphology are few andrelatively subtle but in some cases mirror those seen in the uppermolars Cusps on the lower molars of Ekembo are more bunodontthan those in Proconsul and occupy more of the occlusal surfaceBroad inter-cuspal crests are incorporated into the Ekembo cuspsmaking them appear more inflated Molar cusps of Proconsul are
variables symphyseal height (infradentaleegnathion) planum alveolare length (mostost posterior point on the superior transverse torusegnathion) The first componentable 1) and thus PC 2 and PC 3 illustrate the residual shape variance (72 and 28
Figure 11 Scatter plot showing mandibular corpus robusticity under P4 Trend lines were computed using ordinary least squares (OLS) regression Similar relationships areobservable at other parts of the corpus but differential preservation results in smaller samples at those positions Slopes are significantly different (p lt 00001 tested on loggedvariables)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 55
more distinct from one another and are linked by narrower sharperinter-cuspal crests The lower molars of Ekembo also have asubstantially less developed buccal cingulum than those ofProconsul (Harrison 2002 2010 Harrison and Andrews 2009)
Figure 12 Scatter plot of lower incisor crown proportions Differenc
Lastly and again as seen in the upper molars the two genera canbe distinguished by the significantly greater amount of lowermolar flare in Proconsul (Fig 13 t-test df frac14 59 t frac14 713p lt 00001 see also Harrison and Andrews 2009)
es between Ekembo and Proconsul are significant (p frac14 00242)
Figure 13 Box-and-whisker plot of buccolingual flare in lower molars measured asthe average distance between the mesial buccal and lingual cusps divided by overallbuccolingual crown breadth Differences between Ekembo and Proconsul are highlysignificant (p lt 00001)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6156
Relative molar size has been used to distinguish species ofProconsul but we observed the primary difference to be in relativebuccolingual breadths of M1 and M2 In Ekembo M1 is substantiallynarrower than M2 than is the case in Proconsul (Fig 14 Harrison2002 2010) Although none of the smaller Proconsul mandiblespreserve complete crowns for both M1 and M2 KNM-SO 1112 hasan intact M2 and at least preserves the cervix for M1 However evenif the cervical measurement is used for its M1 breadthdthusunderestimating the true crown breadthdKNM-SO 1112 still has abroader first molar in relation to M2 than would be expected inEkembo If crown breadth is estimated from the ratio of cervixbreadth to maximum crown breadth in other Proconsul fossils therange of likely breadths for the SO 1112 M1 clearly distinguishes it
from the Ekembo sample (Fig 14) Testing this relationship onlogged variables using the mean estimate for the SO 1112 M1(Fig 14) reveals a significant difference between the two groups(ANCOVA df frac14 1 F frac14 2424 p frac14 00002)
As with upper third molars we find a great deal of variation inM3 morphology and mistrust the taxonomic significance some-times placed on characteristics of this toothdat least with regard tothese taxa Harrison (2002 2010) cited the mesiodistal position ofthe M3 entoconid as differentiating Kisingiri third molars fromthose attributed to P africanus We concur with his assessment butfound the entoconid position to vary in specimens attributed toP major Thus this character does not differentiate Ekembo from allProconsul species but may distinguish it from the type speciesP africanus
Discussion
The distinctiveness of the Kisingiri large catarrhine materialfrom that at Tinderet and Ugandan localities has been noted pre-viously by several authors (Pickford 1986 Walker et al 1993Begun 2001 2007 MacLatchy and Rossie 2005 Harrison andAndrews 2009 Harrison 2010) What had not been fully docu-mented was the nearly complete lack of overlap between thesesamples in a number of features despite very large assemblagesfrom Rusinga and Mfangano On descriptive criteria alone (Table 2)many isolated teeth (I1 C1 P3 M2 C1) and all of the mandibles canbe assigned to either Proconsul or Ekembowith very little chance ofmisattribution That some of the most diagnostic features occur inthe upper dentitiondthe only anatomy preserved in theP africanus type specimendadds weight to the taxonomicarrangement proposed here The morphology of the upper teeth inall Proconsul species compellingly unites them into a single cladedistinct from E heseloni and E nyanzae This is reinforced bynumerous features of the mandible and mandibular dentition thatfurther distinguish these samples The number and variety of thesefeatures are such that Proconsul and Ekembo are more differentfrom one another than are many closely-related modern catarrhinegenera
Derived morphology of Ekembo and Proconsul
It is premature to formally assign primitive and derived status tothe individual features of Ekembo and Proconsul before completinga formal phylogenetic analysis This is especially true given theabundance of evidence available for Ekembo and hence the poten-tial impact that our generic revision could have on phylogeneticinterpretations Nevertheless in comparison with both extanthominoids and the small-bodied catarrhines from the earlyMiocene it is informative to differentiate the evolutionary grades ofthese genera Many characteristics of Ekembo are more advanced inthe direction of modern apes compared to those of Proconsul(Table 2) General trends such as expanded and more bunodontcusps broader molar crests reduced cingulum and morphologicalsimilarity between P3 and P4 all represent morphologies thatpresage those of later fossil and extant hominoids The moreadvanced morphologies of P3 and M2 are particularly conspicuouscompared not only with those of Proconsul but to most earlyMiocene catarrhines In contrast the mesially positioned stronglyheteromorphic cusps on the P3 of Proconsul plus the correspondingasymmetric buccal profile notched mesial margin and missingmesial fovea are characteristic of other early Miocene catarrhinessuch as Dendropithecus Rangwapithecus and Limnopithecus andare likely primitive among East African catarrhines The moresymmetric P3 of Ekembo with midline positioned cusps andreduced cusp heteromorphy is reminiscent of later hominoids and
Figure 14 Relative buccolingual dimensions of M1 and M2 KNM-SO 1112 has a preserved M2 but only the cervix of M1 Its M1 breadth was estimated based on the ratio to cervixbreadth to maximum crown breadth in other Tinderet fossils Even the cervical breadth of SO 1112 is broader than would be predicted for the crown breadth of a Kisingiri M1 Trendlines computed using OLS regression and using the average estimate of SO 1112 M1 breadth Group differences were significant (p frac14 00002)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 57
even hominid taxa such as Rudapithecus (eg Kordos and Begun2001)
Proconsul is likewise united by a suite of features that are for themost part characteristic only of this lineage (Table 2) Themorphology of both upper and lower canines is unique to thisgenus and shared by all three well-known species plus the poorlyrepresented middle Miocene P gitongai (Pickford and Kunimatsu2005) The mandibular symphysis is also distinctive in Proconsulcompared with other catarrhines with its very long and shallowlysloping planum alveolare and steeply inclined subplanum surfaceIndeed even the small edentulous anterior mandibular fragmentKNM-SO 372 is readily placed within Proconsul and excluded fromother similarly sized catarrhine taxa such as Rangwapithecus andEkembo based on this unique symphyseal morphology (Figs 8 and9)
Other differences between Ekembo and Proconsul are of uncer-tain evolutionary polarity (eg orientation of the posthypoconecrista) or may be of taxonomic relevance without broader phylo-genetic significance (eg degree of molar flare Table 2) Werecognize that there are morphological differences within Procon-sul particularly between the better known species P africanus andP major However many of these differences are likely related tosize and represent variations on a single generic theme that aremarkedly different from the morphology of Ekembo The recentdescription (Harrison and Andrews 2009) of the intermediatelysized P meswae underscores the broad morphological unity withinProconsul and further substantiates a well-defined Proconsul cladecomprising Tinderet and Ugandan species
Based on the abundant morphological and morphometric evi-dence supporting genus-level distinction between Proconsul andEkembo we are currently conducting a more detailed phylogeneticanalysis of all early Miocene catarrhines in order to better under-stand the distribution and evolutionary significance of these andother morphological features We anticipate that the debates sur-rounding the phylogenetic relationships of an historically consti-tuted sample of Proconsul can be more satisfactorily resolved nowthat the Kisingiri material is appropriately recognized as a distinctgenus
The generic disposition of ldquoUgandapithecusrdquo species
A great many features said to characterize ldquoUgandapithecusrdquowere added to the original diagnosis of the genus by Pickford et al(2009) in their systematic revision of Proconsul (sensu lato) As partof that study they also erected a new species ldquoUrdquo legetetensis andtransferred P meswae to ldquoUgandapithecusrdquo Evaluating the same setof features across the entire available sample however we wereunable to find support for this more expansive conception ofldquoUgandapithecusrdquo for several reasons First some diagnostic fea-tures cited in Pickford et al (2009) were quite variable within theldquoUgandapithecusrdquo sample As an example marked reduction of theM3 entoconid to a bead on the lingual cingulum is not shared byseveral ldquoUrdquo major specimens such as KNM-CA 393 NAP I 49000and Kor 253004 (see Pickford et al 2009 Fig 33e) and was likelynot a feature of a few of the more worn specimens eg BUMP 600and BUMP 601 or even the holotype of P major M 16648 Second
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6158
some diagnostic features of ldquoUgandapithecusrdquo were found to bevariably present in Ekembo arguing against their utility for dis-tinguishing ldquoUgandapithecusrdquo from Pickford et als (2009) com-bined P africanus thorn Kisingiri conception of Proconsul For examplesignificantly reduced curvature of the I1 distal margin in ldquoUganda-pithecusrdquo is also found in some Kisingiri specimens including KNM-RU 1685 1712 1714 1831 1846 and 1951 several more Rusingaspecimens have moderately reduced curvature Third some fea-tures that purportedly distinguish ldquoUgandapithecusrdquo fromP africanus are likely related to size For example the somewhatmore central position of the P3 paracone in ldquoUrdquomajor is not seen inthe smaller ldquoUrdquo meswae whose paracone is positioned similarly tothat of the holotype of P africanus (see Harrison and Andrews2009 Fig 6d)
However the major part of the evidence for recognizingldquoUgandapithecusrdquo (sensu Pickford et al 2009) was assembled by re-assigning specimens traditionally attributed to P africanus into newspecies of ldquoUgandapithecusrdquo The P africanus holotype was left inassociation with the Rusinga sample despite having morphologyoverwhelmingly characteristic of other Tinderet specimens Theresulting specimen allocations left P africanus morphologicallyimpoverisheddstripped of much of its hypodigmdso that Pro-consul for Pickford et al (2009) came to be characterized primarilyby the Rusinga sample (see similar comments in Harrison andAndrews 2009) Within that conceptual framework any spec-imen that differs from those at Rusinga could not be Proconsul butmust instead belong to ldquoUgandapithecusrdquo Thus specimens such asthe KNM-SO 1112 mandible were erroneously assigned to ldquoUgan-dapithecusrdquo because they were (correctly) perceived as differingfrom their Rusinga counterparts (Pickford et al 2009)
Here we restore the P africanus hypodigm and associate it withthe other Tinderet fossils according to their shared morphologySpecimens assigned to ldquoUgandapithecusrdquo revert to Proconsul andmany of the features cited to support a speciose ldquoUgandapithecusrdquo(Pickford et al 2009) instead become diagnostic features of Pro-consul This correction serves to highlight the distinctiveness of theKisingiri remains and the need to recognize this distinctiveness atthe generic level
As demonstrated above specimens from Kisingiri do not displaythe morphology of Proconsul as represented by the holotype ofP africanus M 14084 Given that this specimen preserves only theupper canine and cheek teeth similarities with or departures fromthis morphology represent the most compelling arguments forassessing inclusivity in the genus However for the mandible andlower dentition if one simply sorts the specimens by elementwithout relying on taxonomic preconceptions the overwhelmingdistinction within the traditionally constituted Proconsul sample isbetween Kisingiri material on the one hand and that from Tinderetand Uganda on the other (cf Walker et al 1993 Begun 2001 2007MacLatchy and Rossie 2005 Harrison and Andrews 2009Harrison 2010) Additionally with the Kisingiri specimensremoved to Ekembo the postcranial differences used to partlyjustify a distinct ldquoUgandapithecusrdquo lose their relevance (seeHarrison and Andrews 2009)
We have differed among ourselves in our past treatment ofP major as generically distinct from the smaller species of Pro-consul (Begun 2007 McNulty 2010) Morphologically many ofthe unique features of P major can be interpreted as size-relatedvariants on the smaller P africanus morphology Even if so thequestion is whether these are sufficiently different and numerousto warrant generic distinction that is do they suggest thatP major belongs in a separate clade from P africanus andP meswae As we have demonstrated differences among thesethree species are certainly fewer and show less morphologicaldistinction than those between Ekembo and any of the three
Nevertheless while we ultimately agree with Harrison andAndrews (2009 see also Begun 2007) that the features dis-tinguishing ldquoUrdquo major from P africanus are consistent with thosethat distinguish congeneric species and are at least partly due todifferences in size we concede the possibility that the largestProconsul species could belong in a separate genus (Senut et al2000 Pickford and Kunimatsu 2005) However whether or notone recognizes a separate genus for P major the P africanus ma-terial does not sort morphologically with the Kisingiri materialTherefore the taxonomic alternatives are either two generaProconsul and Ekembo or three Proconsul Ugandapithecus andEkembo with Proconsul and Ugandapithecus united in a cladeexclusive of Ekembo Thus while we strongly favor the firstalternative recognizing Ugandapithecus does not change thefundamental argument concerning the need to refer the Kisingirispecies to a generically and cladistically distinct taxon from thespecies represented at Tinderet and the Ugandan sites
Specimens of interest
Despite the substantial dental and gnathic differences betweenProconsul and Ekembo there are a few specimens whose taxonomicaffinities require additional scrutiny
KNM-RU 1676-77 This set of associated upper and lower teeth is aconsistent outlier among E heseloni and E nyanzae specimens Insome ways this specimen more closely resembles Tinderet Pro-consul in the features that distinguish it from Ekembo Molar cuspmorphology and cingulum development I1 lingual tubercledevelopment and M2 shape and posthypocone crista orientationare more consistent with specimens from the Tinderet sites On theother hand it otherwise resembles Ekembo and lacks the strongestdiagnostic features that unite species of Proconsul such as thedistinctive canine and P3 morphology Overall this specimen doesnot fit unambiguously into either genus in fact the degree ofbuccolingual flare cusp morphology enamel wrinkling and espe-cially the long extension of enamel onto the anterior root in P3 arein some ways reminiscent of Afropithecus although it clearly differsfrom that genus as well (cf Rossie and MacLatchy 2013)
Were these characteristics individually distributed among thelarge Kisingiri sample one might attribute this to intra-genericvariation but when found together in a single specimen from alarge sample it suggests a unique individual One of us (KPM)thinks it likely that the specimen was not actually found at aKisingiri locality based on its unique preservation compared toother Rusinga fossils and on other examples of mis-proveniencedspecimens (Leakey 1967 Andrews and Molleson 1979 Pickford1986 Kelley 1986) However we recognize that relying toofrequently on this explanation allows almost any scenario to bejustified and suggest some alternative possibilities One is that thespecimen was found on Rusinga but in the much older WayandoFormation strata instead of the Hiwegi Formation The differencesin morphology in that case might reflect more than a million yearsof temporal change (McCollum et al 2013) with KNM-RU 1676-77preserving certain more primitive Proconsul-like features As analternative it is reasonable to assume that fossil assemblages asvast as those from Rusingawill include not only abundant taxa butalso rare or infrequent inhabitants of the local biome There aremany examples of this from Rusingas mammalian communityeg ldquoHemicyonrdquo Kulutherium Tadarida cf Hyainailouros cfArchaeobeledon Kelba (see relevant chapters in Werdelin andSanders 2010) As such KNM-RU 1676-77 may simply representa rare and well-preserved specimen of some other catarrhinetaxon that lived among or visited a primate community dominatedby Ekembo
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 59
KNM-SO 1112 This specimen was initially referred to Rangwapi-thecus but soon after was transferred to Proconsul afrianus (Bosler1981 Martin 1981)da designation that most subsequent authorshave followed (Pickford 1986 Nengo and Rae 1992 Kelley 19861995 Walker et al 1993 MacLatchy and Rossie 2005 Hill et al2013) Pickford et al (2009) more recently referred this specimento ldquoUrdquo meswae citing two primary reasons First they reiteratedBoslers (1981) concern that its P3 differs from others attributedto P africanus However at that time the majority of what wasconsidered to be P africanus would have been from RusingaIsland In fact while noting its contrasting morphology Boslersuggested that differences between KNM-SO 1112 and other P3smight reasonably be attributed to sexual dimorphism notingthat this specimen provides ldquoa not implausible match for the Pafricanus holotype maxilla M 14084rdquo (1981152) Thatassessment is borne out in this study Using the much largersample of Ekembo P3s as a model variation can be seen toencompass specimens with well-developed mesial beaks andthose with more bilaterally compressed crowns and parallelsides The latter morphology is predominantly found amongmale specimens (KNM-RU 1674 1924 and KPS I and III) thussupporting Boslers (1981) suggestion that this variation isrelated to sexual dimorphism and hence that KNM-SO 1112 is amale P africanus
The second feature cited by Pickford et al (2009) in referringthis specimen to ldquoUrdquo meswae was posterior shallowing of themandibular corpus However because they redefined ldquoUgandapi-thecusrdquo to include all of the well-preserved Tinderet mandibles(Pickford et al 2009)dleaving Proconsul to be represented only byRusinga specimensdtheir only alternative within this scheme for aspecimen the size of SO 1112 was to refer it to ldquoUrdquomeswae It seemsodd though to assign this specimen to a species from a (perhapsmuch) older site when the Proconsul africanus holotype is ofappropriate size and similar age and as there are no other adultmandibles from either species with which to compare it In anycase mandibular shallowing is variable in both Ekembo and Pro-consul with some Kisingiri specimens exhibiting considerableshallowing (eg KNM-RU 2087) and some ldquoUgandapithecusrdquo spec-imens having very little (eg M 14086)
We therefore see no morphological grounds to assign KNM-SO1112 to P meswae but do find some morphological evidence toassign it as a male P africanus which occurs at Songhor
KNM-LG 452 This very complete mandible was originally recog-nized as a female of P major based on the size of the dentition androbusticity of the mandible (Martin 1981) Bosler (1981) howeverleft this specimen unassigned noting similarities to P major in P3-M1 size but having distinctly smaller M2e3 In this she likenedKNM-LG 452 to a few other specimens (notably M 14086) thatare smaller in their distal molars in a manner more consistentwith taxonomic rather than sex differences (Bosler 1981 see alsoPilbeam 1969) Kelley (1986 1995) also assigned this specimen asa female of P major based on canine size and morphology butnoted that the canine lacked the shallowly-sloping mesial ridgetypical of most female anthropoid canines This is also truehowever of two other canines assigned as female P major KO 9and SO 373 (Kelley 1986) and so might simply be an unusualcharacteristic of P major female lower canines In the end Kelley(1995) concluded that considering all of the criteria fordistinguishing male and female canines the evidence that KNM-LG 452 is female remains questionable
Pickford et al (2009) further developed the idea that two spe-cies are represented within the P major sample and made KNM-LG452 the holotype of a new species ldquoUrdquo legetetensis We concur thatthis specimen is unique among the well-preserved Tinderet
mandibles in some important features Whereas male and femalemandibles of P major have long internal symphyseal planesexpansive extra- and retromolar sulci and rami that cross theocclusal plane at distal M3 KNM-LG 452 has a shorter symphysealplane (though not to the extent seen in Ekembo) a moderateextramolar sulcus but no retromolar sulcus and a ramus thatcrosses the occlusal plane more mesially If KNM-SO 396 and SO404 represent males and females respectively of P major then themorphology of KNM-LG 452 appears distinct
At present however we do not think there is sufficient evidenceto differentiate this specimen from either P major or P meswaePickford et al (2009) argue that KNM-LG 452 cannot belong to thelatter species because of its larger size The size differences how-ever are well within the range of variation seen in P majorE heseloni and E nyanzae Moreover whereas the Legetet mandibleis larger than most of the P meswae hypodigm an adult P4 referredto this species (KNM-ME 25) substantially exceeds its counterpartin KNM-LG 452 Should the Legetetmandible ultimately prove to bea male specimen size alone would not rule out its attribution toP meswae On the other hand if it is female then it becomes harderto justify its inclusion in that sample of mostly smaller specimens Iffemale therefore it could represent a separate species for whichthe binomial Proconsul legetetensis comb nov is available Recentlycollected fossils from the SonghorKapurtay area provide someadditional support for the possibility that three species of Proconsulexisted at these localities (McNulty 2014ab) However toconvincingly demonstrate this and determine whether a thirdspecies can be differentiated from P meswae will require a moredetailed analysis of variation among Proconsul species a corre-sponding study of variation in modern taxa anddalmost certain-lydadditional fossil specimens For now we regard ldquoUrdquo legetetensisas a subjective junior synonym of P major
Conclusions
The genus Proconsul has been central to studies of ape and hu-man evolution since its first description by Hopwood (1933) As oneof the best known fossil catarrhines represented by nearly everyskeletal element our knowledge of Proconsuls functional anatomypositional behavior and overall morphology has made it thefoundation for paleoanthropological research in the early Mioceneas well as studies on the origin and early diversification of homi-noids With respect to systematics however Proconsul is less wellunderstood the type species P africanus is known from a rela-tively small number of specimens from sites in the Tinderet areaand no postcranial bones can be unambiguously assigned to itRather it is the copious and well-preserved specimens from theKisingiri sites of Rusinga and Mfangano Islands that have formedthe basis for the common image of Proconsul
We argue here that the Kisingiri species traditionally referredto ProconsuldP heseloni and P nyanzaedare generically distinctfrom P africanus and hereby transfer them into a new genusEkembo based on a substantial number of morphological differ-ences (Table 2) Considering only those traits preserved in theP africanus holotype the entire sample from Rusinga and Mfan-gano differs markedly in the morphology of the upper canine P3P4 and M2 Additional differences in the I1 lower incisors andcanines molar cusp and occlusal crest morphology degree of flarein all molars and especially mandibular features clearly distin-guish the Kisingiri from non-Kisingiri samples in a way that isincompatible with congeneric species With the transfer ofE heseloni and E nyanzae Proconsul becomes a well-defined cladecomprising at least three speciesdP africanus P major andP meswaedunited by several features that are unique for thegenus (Table 2)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6160
We found no support in this study for the genus ldquoUgandapi-thecusrdquo as constructed by Pickford et al (2009) In that paper theytransferred the vast majority of Proconsul fossils from the Tinderetand Napak localities into ldquoUgandapithecusrdquo leaving P africanusrepresented only by its holotype and a small number of isolatedteeth and aligning it with the material from Rusinga and Mfan-ganoWhen the P africanus hypodigm is reconstituted and properlyaffiliated with the other Tinderet species according to itsmorphology much of the evidence marshaled by Pickford et al(2009) for ldquoUgandapithecusrdquo actually supports the distinction be-tween the Tinderet and Kisingiri samples ie the generic distinc-tion of Ekembo In addition some of the traits enumerated byPickford and colleagues in support of ldquoUgandapithecusrdquo wererevealed in our analysis to be variable within one or more speciesand therefore of questionable taxonomic utility We do not rejectout of hand the original conception of ldquoUgandapithecusrdquo (Senutet al 2000) as either a monotypic genus distinguishing ldquoUrdquo ma-jor or perhaps including a closely allied middle Miocene ldquoUrdquogitongai as well (Pickford and Kunimatsu 2005) but we have fornow retained the early Miocene specimens of both in Proconsulpending further study However even if Ugandapithecus is a taxo-nomically valid genus comprising just the latter two species theirmorphology indicates that theywould still be united in a cladewithProconsul and they lack several more advanced features that partlydefine Ekembo Better understanding of the role that body size playsinmediating differences within Proconsulwill help to further clarifythe relationships among these species
Despite the existence of one well-represented and several lesscomplete skeletons of what was long assumed to be Proconsul thephylogenetic status of this genus has defied consensus This islargely due in our view to Proconsul having been an admixture oftwo distinct genera one more advanced in the direction of extantapes than the other As we demonstrate here the material fromRusinga and Mfangano traditionally referred to Proconsul andwhich includes all of the more complete skeletal material isdistinct from the Tinderet and Napak material and belongs in aseparate genus Ekembo By disentangling these distinct clades weanticipate being able to achieve better phylogenetic resolution forboth genera thereby improving our knowledge of early hominoidevolution and diversification
Acknowledgments
We thank many people and agencies that assisted with thedevelopment and completion of this project This research wasinitiated as a result of funding from the Wenner-Gren Foundationand the Johns Hopkins School of Medicine to DR Begun in1987e1989 and was renewed following research and travelgenerously funded by multiple Leakey Foundation grants to KMcNulty National Science Foundation grants to K McNulty(0852609 1241807) the McKnight Foundation the University ofMinnesota the Lloyd A Wilford Endowment the LeverhulmeFoundation and the Turkana Basin Institute J Kelley gratefullyacknowledges support from the Institute of Human Origins Ari-zona State University DR Begun also wishes to acknowledge theAlexander Humboldt Stiftung and NSERC for funding support Weare grateful to the National Museums of Kenya the UgandaMuseum the Natural History Museum and to Rose Mwanja EzraMusiime Tom Mukhuyu Samuel Muteti James Yatich and MaryMuungu for access to and assistance with specimens in their careMany people provided assistance comments criticisms insightsand suggestions on various aspects of this work and we especiallythank Holly Dunsworth Will Harcourt-Smith Tom Lehmann DanPeppe Nicole Garrett Kirsten Jenkins Rutger Jansma Susy CoteIsaiah Nengo Andrew Hill and Brenda Benefit K McNulty
acknowledges the evolutionary anthropology faculty and studentsat Durham University for their helpful comments on an earlyversion of this manuscript and for their gracious hospitality Weare likewise grateful to the anonymous reviewers associate editorand editor whose comments strengthened the paper This manu-script is publication 1 supporting Research on East AfricanCatarrhine and Hominoid Evolution (REACHE)
Appendix A Supplementary data
Supplementary data related to this article can be found at httpdxdoiorg101016jjhevol201503009
References
Andrews P 1974 New species of Dryopithecus from Kenya Nature 249 188e190Andrews P 1978 A revision of the Miocene Hominoidea of East Africa Bull Brit
Mus (Nat Hist) Geol 30 85e225Andrews P 1985 Family group systematics and evolution among catarrhine pri-
mates In Delson E (Ed) Ancestors The Hard Evidence Liss New Yorkpp 14e22
Andrews P Harrison T Martin L Pickford M 1981 Hominoid primates from anew Miocene locality named Meswa Bridge in Kenya J Hum Evol 10123e128
Andrews P Begun D Zylstra M 1997 Interrelationships between functionalmorphology and paleoenvironments in Miocene hominoids In Begun DWard C Rose M (Eds) Function Phylogeny and Fossils Miocene HominoidEvolution and Adaptations Plenum Press New York pp 29e58
Andrews P Molleson T 1979 The provenance of Sivapithecus africanus Bull BritMus (Nat Hist) Geol 32 19e23
Begun DR 2001 African and Eurasian Miocene hominoids and the origins of theHominidae In de Bonis L Koufos GD Andrews P (Eds) Hominoid Evolutionand Climatic Change in Europe Phylogeny of the Neogene Hominoid Primatesof Eurasia Cambridge University Press London pp 231e253
Begun DR 2007 Fossil record of Miocene hominoids In Henke W Tattersall I(Eds) Handbook of Paleoanthropology Springer Berlin pp 921e977
Begun DR Teaford MF Walker AC 1994 Comparative and functional anatomyof Proconsul phalanges from the Kaswanga primate site Rusinga Island KenyaJ Hum Evol 26 89e165
Bishop WW 1964 More fossil primates and other Miocene mammals from North-East Uganda Nature 203 1327e1331
Bishop WW Miller JA Fitch FJ 1969 New potassium-argon age determinationsrelevant to the Miocene fossil mammal sequence in East Africa Am J Sci 267669e699
Blankenship JA Mincer HH Anderson KM Woods MA Burton EL 2007Third molar development in the estimation of chronologic age in Americanblacks as compared with whites J Forensic Sci 52 428e433
Bosler W 1981 Species groupings of early Miocene dryopithecine teeth from EastAfrica J Hum Evol 10 151e158
Coleman MN 2008 What does geometric mean mean geometrically Assessingthe utility of geometric mean and other size variables in studies of skullallometry Am J Phys Anthropol 135 404e415
Cote S Malit N Nengo I 2013 Additional mandibles of Rangwapithecus gordonian early Miocene catarrhine from the Tinderet localities of Western KenyaJ Hum Evol 153 341e352
Deane AS 2009 Early Miocene catarrhine dietary behavior the influence of theRed Queen Effect on incisor shape and curvature J Hum Evol 56 275e285
Drake R Van Couvering JA Pickford M Curtis G Harris JA 1988 New chro-nology for the Early Miocene mammalian faunas of Kisingiri Western KenyaJ Geol Soc Lond 145 479e491
Evernden JF Savage DE Curtis GH James GT 1964 Potassium-argon datesand the Cenozoic mammalian chronology of North America Am J Sci 262145e198
Gebo DL MacLatchy L Kityo R Deino A Kingston J Pilbeam D 1997A hominoid genus from the Early Miocene of Uganda Science 276 401e404
Geoffroy Saint-Hilaire E 1812 Tableau des quadrumanes 1 Ord QuadrumanesAnnls Mus Hist Nat Paris 19 85e122
Gray JE 1825 Outline of an attempt at the disposition of the Mammalia into tribesand families with a list of the genera apparently appertaining to each tribeAnnls Philos 10 337e344
Harrison T 1982 Small-bodied apes from the Miocene of East Africa PhDDissertation University of London
Harrison T 1986 New fossil anthropoids from the middle Miocene of East Africaand their bearing on the origin of the Oreopithecidae Am J Phys Anthropol 71265e284
Harrison T 1987 The phylogenetic relationships of the early catarrhine primates areview of the current evidence J Hum Evol 16 41e80
Harrison T 2002 Late Oligocene to middle Miocene catarrhines from Afro-ArabiaIn Hartwig WC (Ed) The Primate Fossil Record Cambridge University PressCambridge pp 311e338
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 61
Harrison T 2010 Dendropithecoidea Proconsuloidea and Hominoidea (CatarrhiniPrimates) In Werdelin L Sanders WJ (Eds) Cenozoic Mammals of AfricaUniversity of California Press Berkeley pp 429e469
Harrison T Andrews P 2009 The anatomy and systematic position of a newspecies of early Miocene proconsulid from Meswa Bridge Kenya J Hum Evol56 479e496
Hill A Nengo IO Rossie JB 2013 A Rangwapithecus gordoni mandible from theearly Miocene site of Songhor Kenya J Hum Evol 65 490e500
Hopwood AT 1933 Miocene primates from Kenya J Linn Soc Zool 38 437e464Ishida H Kunimatsu Y Takano T Nakano Y Nakatsukasa M 2004 Nacholapi-
thecus skeleton from the Middle Miocene of Kenya J Hum Evol 46 69e103Jolicouer P 1963 The multivariate generalization of the allometry equation Bio-
metrics 19 497e499Jungers WL Falsetti AB Wall CE 1995 Shape relative size and size-
adjustments in morphometrics Am J Phys Anthropol 38 (S2) 137e161Kavanagh KD Evans AR Jernvall J 2007 Predicting evolutionary patterns of
mammalian teeth from development Nature 449 427e433Kelley J 1986 Species recognition and sexual dimorphism in Proconsul and
Rangwapithecus J Hum Evol 15 461e495Kelley J 1995 Sex determination in Miocene catarrhine primates Am J Phys
Anthropol 96 390e417Kelley J 1993 Taxonomic implications of sexual dimorphism in Lufengpithecus In
Kimbel WH Martin LB (Eds) Species Species Concepts and Primate Evo-lution Springer New York pp 429e458
Kent PE 1942 The country round the Kavirondo Gulf of Victoria Nyanza Geogr J100 22e31
Kordos L Begun DR 2001 A new cranium of Dryopithecus from RudabanyaHungary J Hum Evol 15 689e700
Le Gros Clark WE Leakey LSB 1950 Diagnoses of East African Miocene Homi-noidea Q J Geol Soc Lond 105 260e263
Le Gros Clark WE Leakey LSB 1951 The Miocene Hominoidea of East Africa InFossil Mammals of Africa No 1 British Museum (Natural History) London
Leakey LSB 1943 A Miocene anthropoid mandible from Rusinga Kenya Nature152 319e320
Leakey LSB 1963 East African fossil Hominoidea and the classification within thissuper-family In Washburn SL (Ed) Classification and Human EvolutionAldine Chicago pp 32e49
Leakey LSB 1967 Notes on the mammalian faunas from the Miocene and Pleis-tocene of East Africa In Bishop WW Clark JD (Eds) Background to Evolu-tion in Africa University of Chicago Press Chicago pp 7e29
Leakey RE Leakey MG 1986 A new Miocene hominoid from Kenya Nature 324143e146
Leakey RE Leakey MG Walker AC 1988 Morphology of Afropithecus turka-nensis from Kenya Am J Phys Anthropol 76 289e307
Leakey MG Ungar PS Walker A 1995 A new genus of large primate from theLate Oligocene of Lothidok Turkana District Kenya J Hum Evol 28519e531
Linnaeus C 1758 Systema naturae per regna tria naturae secundum classesordines genera species cum characteribus differentris synonymis locis tenthed Laurentii Salvii Stockholm
MacInnes DG 1943 Notes on the East African Miocene primates J E Afr UgandaNat Hist Soc 17 141e181
MacLatchy L 2004 The oldest ape Evol Anthropol 13 90e103MacLatchy L Bossert WH 1996 An analysis of the articular surface distribution of
the femoral head and acetabulum in anthropoids with implications for hipfunction in Miocene hominoids J Hum Evol 31 425e453
MacLatchy L Rossie JB 2005 The Napak hominoid still Proconsul major InLieberman DE Smith RJ Kelley J (Eds) Interpreting the Past Essays onHuman Primate and Mammal Evolution in Honor of David Pilbeam Brill Ac-ademic Publishers Boston pp 15e28
Martin L 1981 New specimens of Proconsul from Koru Kenya J Hum Evol 10139e150
McCollum MS Peppe DJ McNulty KP Dunsworth HM Harcourt-Smith WEH Andrews AL 2013 Magnetostratigraphy of the early MioceneHiwegi Formation (Rusinga Island Lake Victoria Kenya) Geological Society ofAmerica Abstracts with Programs 45 p 12
McNulty KP 2010 Apes and tricksters the evolution and diversification ofhumans closest relatives Evol Edu Outreach 3 322e332
McNulty KP Harcourt-Smith WEH Dunsworth HM 2007 New primate fossilsfrom Rusinga Island Kenya Am J Phys Anthropol 132 (S44) 170
McNulty KP MacLatchy L Peppe DJ Nengo I Manthi FK Miller ERStevens NJ Cote S Lehmann T Kingston J 2014a A regional approach toEast African early Miocene paleobiology J Vert Paleontol 185 SVP Programand Abstracts Book 2014
McNulty KP MacLatchy L Rossie JB Peppe DJ Deino AL Mbua ENManthi FK Nengo IO Stevens NJ Cote S Lehmann T Gutierrez M 2014b
Research on East African catarrhine and hominoid evolution results from thefirst year Am J Phys Anthropol 153 (S58) 182
Michel LA Peppe DJ Lutz JA Driese SG Dunsworth HM Harcourt-Smith WEH Horner W Lehmann T Nightengale S McNulty KP 2014Remnants of an ancient forest provide ecological context for Early Miocenefossil apes Nature Comm 5 3236
Myres JL 1932 Africa Man 32 208Nakatsukasa M Kunimatsu Y Nakano Y Takano T Ishida H 2007 Vertebral
morphology of Nacholapithecus kerioi based on KNM-BG 35250 J Hum Evol52 347e369
Nengo IO Rae TC 1992 New hominoid fossils from the early Miocene site ofSonghor Kenya J Hum Evol 23 423e429
Peppe DJ McNulty KP Cote SM Harcourt-Smith WEH Dunsworth HM VanCouvering JA 2009 Stratigraphic interpretation of the Kulu Formation (EarlyMiocene Rusinga Island Kenya) and its implications for primate evolutionJ Hum Evol 56 447e461
Pickford M 1981 Preliminary Miocene mammalian biostratigraphy for westernKenya J Hum Evol 10 73e97
Pickford M 1986 Sexual dimorphism in Proconsul Hum Evol 1 111e148Pickford M Andrews PJ 1981 The Tinderet Miocene sequence in Kenya J Hum
Evol 10 11e33Pickford M Kunimatsu Y 2005 Catarrhines from the Middle Miocene (ca 145
Ma) of Kipsaraman Tugen Hills Kenya Anthropol Sci 113 189e224Pickford M Senut B Gommery D Musiime E 2009 Distinctiveness of Ugan-
dapithecus from Proconsul Estudios Geol 65 183e241Pilbeam DR 1969 Tertiary Pongidae of East Africa evolutionary relationships and
taxonomy Bulletin of the Peabody Museum of Natural History 31 pp 1e185Rae TC 1993 Phylogenetic analysis of proconsulid facial morphology PhD
Dissertation State University of New York at Stony BrookRae TC 1999 Mosaic evolution in the origin of the Hominoidea Folia Primatol 70
125e135Rafferty KL Walker A Ruff C Rose MD Andrews PJ 1995 Postcranial esti-
mates of body weights in Proconsul with a note on a distal tibia of P major fromNapak Uganda Am J Phys Anthropol 97 391e402
Rossie JB MacLatchy L 2013 Dentognathic remains of an Afropithecus individualfrom Kalodirr Kenya J Hum Evol 65 199e208
Senut B Pickford M Gommery D Kunimatsu Y 2000 Un nouveau genredrsquohominoiumlde du Miocene inferieur drsquoAfrique orientale Ugandapithecus major(Le Gros Clark and Leakey 1950) CR Acad Sci Paris 331 227e233
Simons EL Pilbeam DR 1965 Preliminary revision of the Dryopithecinae (Pon-gidae Anthropoidea) Folia Primatol 3 81e152
Smith TM Martin LB Leakey MG 2003 Enamel thickness microstructure anddevelopment in Afropithecus turkanensis J Hum Evol 44 283e306
Stevens NJ Seiffert ER OConnor PM Roberts EM Schmitz MD Krause CGorscak E Ngasala S Hieronymus TL Temu J 2013 Palaeontological evi-dence for an Oligocene divergence between Old World monkeys and apesNature 497 611e614
Suwa G Kono RT Katoh S Asfaw B Beyene Y 2007 A new species of great apefrom the late Miocene epoch in Ethiopia Nature 448 921e924
Szalay FS Delson E 1979 Evolutionary History of the Primates Academic PressNew York
Teaford MF Beard KC Leakey RE Walker A 1988 New hominoid facial skel-eton from the Early Miocene of Rusinga Island Kenya and its bearing on therelationship between Proconsul nyanzae and Proconsul africanus J Hum Evol17 461e477
Van Couvering JAH Miller J 1969 Miocene stratigraphy and age determinationsRusinga Island Kenya Nature 221 628e632
Walker A 1997 Proconsul function and phylogeny In Begun DR Ward CVRose MD (Eds) Function Phylogeny and Fossils Miocene Hominoid Evolu-tion and Adaptation Plenum Press New York pp 209e224
Walker A Pickford M 1983 New postcranial fossils of Proconsul africanus andProconsul nyanzae In Ciochon RL Corruccini RS (Eds) New Interpretationsof Ape and Human Ancestry Plenum Press New York pp 325e351
Walker A Teaford M 1988 The Kaswanga primate site an Early Miocene hom-inoid site on Rusinga Island Kenya J Hum Evol 17 539e544
Walker A Teaford M 1989 The hunt for Proconsul Sci Am 260 76e82Walker A Teaford MF Martin L Andrews P 1993 A new species of Proconsul
from the early Miocene of RusingaMfangano Islands Kenya J Hum Evol 2543e56
Ward CV Ruff CB Walker A Rose MD Teaford MF Nengo IO 1995 Func-tional morphology of Proconsul patellas from Rusinga Island Kenya with im-plications for other Miocene-Pliocene catarrhines J Hum Evol 29 1e19
Ward S Brown B Hill A Kelley J Downs W 1999 Equatorius a new hominoidgenus from the Middle Miocene of Kenya Science 285 1382e1386
Werdelin L Sanders WJ 2010 Cenozoic Mammals of Africa University of Cali-fornia Press Berkeley

KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 45
attributed to this genus and here provide an alternate taxonomicinterpretation to those in current use Our primary objective was toevaluate the morphological homogeneity of the genus Proconsul astraditionally constituted (Proconsul sensu lato) and explore thepossibility that theKisingiri specimens formadistinct group (Walkeret al1993 Begun 2001 2007MacLatchyandRossie 2005Harrisonand Andrews 2009) While we agree with Pickford et al (2009) thatthe material traditionally referred to Proconsul exceeds what can becomfortably accommodated within a single genus our resultsconform to the suggestion proposed by Begun (2001 2007) and laterby MacLatchy and Rossie (2005) and Harrison and Andrews (2009)that the Kisingiri specimens from Rusinga and Mfangano Islandsconstitute a distinct cladewith respect to Proconsul from theTinderetand Ugandan localities Our assessment of the taxonomic andphyletic distinction of the Kisingiri material necessitates genericdifferentiation of this sample from Proconsul
Systematics
Order Primates Linnaeus 1758Infraorder Catarrhini E Geoffroy Saint-Hilaire 1812Superfamily Hominoidea Gray 1825Family incertae sedisEkembo gen novurn lsidzoobankorgactB11CECF0-9DD1-4265-9C06-A60E4BEE974E
Synonymy
1950 Proconsul Hopwood Le Gros Clark and Leakey 2601965 Dryopithecus (Proconsul) (Hopwood) Simons and Pilbeam
1201978 Proconsul Hopwood Andrews 901986 Proconsul Hopwood Kelley 4791993 Proconsul Hopwood Walker et al 512010 Proconsul Hopwood Harrison 437
Type species
Ekembo nyanzae comb nov (Le Gros Clark and Leakey 1950)
Included species
Ekembo nyanzae (Le Gros Clark and Leakey 1950) Ekemboheseloni comb nov (Walker et al 1993)
Etymology
Ekembo means ldquoaperdquo or ldquomonkeyrdquo in the Suba language whichwas historically spoken by peoples in western Kenya includingthose who settled Rusinga and Mfangano Islands (Jack Wanyendepers comm)
Diagnosis
Medium- to large-bodied and sexually dimorphic hominoidcharacterized by the following combination of features Frontalbone moderately wide at the coronal suture narrowing anteri-orly toward gracile supraorbital rims which are not demarcatedby a post-toral sulcus Large frontal sinuses behind a prominentglabella Minor postorbital constriction Temporal lines promi-nent and situated high on frontal and parietal bones convergingposteriorly without forming a sagittal crest (in female
specimens) Nasal bones long and thin Broad interorbital regionflat rather than projecting Upper face more vertically orientedrelative to the mid- and lower face Nasolacrimal duct positionedwithin a broad orbit Rostrum broad with premaxilla extendingup to the nasal bones Canine jugum prominent in both sexes butcanine fossa developed only in males Nasal aperture broad withwidest portion inferior and sometimes near the base Inferioraperture margin concave or incised in the midline Nasoalveolarclivus intermediate in height between hylobatids and hominidsClivus does not overlap palatine process of the maxilla pre-senting typical mammalian incisive fenestration ante-roposteriorly positioned at or posterior to mid-canine Maxillarysinus extensive reaching anteriorly to P3P4 junction Zygomati-coalveolar crest arises low on the face near M2 Nearly verticalmalar region Broad anterior palate A single palatine foramenpositioned near the M2M3 junction Broad mandibular ramusangled slightly posteriorly from vertical leads to a graduallyrounded and gracile gonial region Mandibular corpora rangefrom short and gracile to taller and moderately robust Sym-physis characterized by moderately developed superior trans-verse torus and variably developed inferior transverse torus andgenioglossal pit Symphyseal planum and subplanum regionvertically or nearly vertically oriented A single mental foramenpositioned near the P3P4 junction approximately halfway up thecorpus or higher
Upper central and lateral incisors heteromorphic Centralincisor characterized by a narrow lingual tubercle often consist-ing of two or three narrow ridges and distinct but thin marginalridges that are continuous with the lingual cingulum with themesial marginal ridge joining the cingulum at a sharp angle incontrast to a more continuously curving transition from cingulumto distal marginal ridge resulting in an asymmetric ridge andcingulum morphology Lateral incisors have well-developedincisal edges and are asymmetric with a moderately developedlingual cingulum Narrow lower incisors with central and lateralincisors having similar morphology Upper and lower caninesstrongly sexually dimorphic with only slight bilateral compres-sion and moderately low-crowned in relation to basal crown di-mensions with apices terminating at a simple point Canine rootssmooth and conical Cheek tooth cusps low and broad and con-nected by inflated intercuspal crests Upper premolars morpho-logically similar with well-developed mesial foveae onlymoderate cusp heteromorphy and cusps positioned at theapproximate mesiodistal midpoint of the crown resulting innearly symmetric buccal profiles Molars with moderate bucco-lingual flare and reduced cingulum development Upper first andsecond molars approximately square with nearly straight distalmargins and buccally oriented posthypocone cristae Lower firstand second molars rectangular with M1 substantially narrowerthan M2 Upper and lower third molars vary in degree of distalcusp development
Forelimbs and hind limbs of similar length Long vertebral col-umn with six lumbar vertebrae characterized by transverse pro-cesses that arise from the dorsal margin of the vertebral centrumand prominent accessory processes for erector spinae musclesPossessing a coccyx rather than a tail Thorax long narrow anddorsoventrally deep Broad flat sternebrae Pelvis long and narrowwith sagittally oriented ilia an elongate ischium lacking osseousevidence for the presence of callosities and a narrow sacrumwith asmall sacro-iliac joint Scapula positioned laterally on thorax with anarrow ovoid and ventrally oriented glenoid fossa Humerus witha posteriorly directed head retroflexed shaft with little torsion anda distal articular surface lacking a well-differentiated trochlea andcapitulum and having a narrow zona conoidea and shallowtrochlear notch but lacking an entepicondylar foramen Radius
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6146
with a small ovoid head and flat distal articular surface that mainlycontacts the scaphoid Ulna displays a large olecranon processnarrow semilunar notch with only a slight keel anteriorly posi-tioned radial notch and a long distal styloid process that retainsarticulation with triquetrum and pisiform Carpals relatively nar-row and include an unfused os centrale a hamate with a smallhamulus and a flat medially oriented triquetral surface Handcharacterized by straight metacarpals with narrow bases andheads Pollex well developed Manual and pedal phalanges gener-ally similar to each other but less so than in Old World monkeysFemur slender with a small head long neck with a high neck-shaftangle centrally placed fovea capitis reduced greater trochanterand a broad distal end with the medial condyle slightly larger thanthe lateral Broad flat patella Tibia long and slender but fibularobust Tarsals and metatarsals elongate with a robust hallux
Differential diagnosis
Ekembo can be readily distinguished from all other medium-and large-bodied catarrhine genera from the early Miocene Cra-nial mandibular and dental differences between Ekembo andProconsul are described in detail below and summarized inTable 2 Ekembo also differs from Proconsul in having a rounderradial head and a femur with a shorter greater trochanter poste-riorly projecting lesser trochanter less extensive inter-trochantericcrest more robust phalanges with more strongly developedfibrous flexor sheath ridges and broader proximal and interme-diate phalangeal distal joint surfaces (condyles) with widertrochlear grooves and less sharply defined trochlear ridges(Harrison 1982 Andrews et al 1997 Senut et al 2000 Pickfordet al 2009)
Ekembo differs from Afropithecus in having a more inflatedglabellar region no frontal trigon less strongly developed anteriortemporal ridges less postorbital constriction a stronger mandib-ular superior transverse torus less posterior shallowing of themandibular corpus higher-crowned lower canines in relation tomesiodistal length buccolingually narrower upper premolarsrelative to molars upper premolars with cusps positioned closer tothe mesiodistal midline reduced P3 cusp heteromorphy a moresymmetric I2 and substantially less buccolingual flare in the cheekteeth (Leakey et al 1988 Rossie and MacLatchy 2013)
Ekembo differs from Heliopithecus in having buccolinguallynarrower premolars relative to molars premolar cusps positionedcloser to the mesiodistal midline and reduced P3 cusp hetero-morphy (Harrison 2010)
Ekembo differs from Equatorius in having a more gracilemandible a larger superior but smaller inferior mandibular trans-verse torus labiolingually narrower lower incisors with straightermesial and distal margins higher-crowned lower canines in rela-tion to mesiodistal length greater molar cingulum development aless posteriorly directed humeral medial epicondyle a more ante-riorly directed radioulnar joint and narrower metacarpal heads(Ward et al 1999)
Ekembo differs from Nacholapithecus in having a shorter sub-nasal clivus a more expansive incisive foramen a broader nasalaperture at the base a minimally developed mandibular inferiortransverse torus a narrower lingual tubercle on I1 higher-crownedlower canines in relation to mesiodistal length a larger humeralradial fossa a shallower coronoid fossa a less well-developed zonaconoidea a narrower and distally slanted ulnar coronoid process alonger and lower femoral neck a more posteriorly positioned lessertrochanter and a smaller posteromedial tubercle on the talus(Ishida et al 2004)
Ekembo differs from Nyanzapithecus in having more anteriorlyextended maxillary sinuses a relatively taller mandibular corpus
less buccolingually compressed upper and lower canines with lesscurved crowns lower and more rounded cusps with more inflatedocclusal crests on the cheek teeth P3 and P4 that more closelyresemble each other and have much less cingular development abroader P4 relative to M1 M1e2 occlusal outline more closelyapproximating a square and lower molars that are broader relativeto mesiodistal length with a more mesially placed entoconid rela-tive to the hypoconid (Harrison 1986 2010)
Ekembo differs from Rangwapithecus in having a broad andexpansive palatal fenestration a wider palate relative to length amore robust mandible with a well-defined superior transversetorus broader upper and lower incisors relative to height lessbuccolingually compressed canines without scimitar-shapedcrowns upper premolars with reduced cingulum and cusps posi-tioned near the mesiodistal midpoint resulting in expanded mesialfoveae and symmetric buccal profiles a broader P4 relative to M1upper and lower molars that are buccolingually broader relative tolength and have more rounded cusps and inflated occlusal crestsmore squared upper molars with substantially less cingulumdevelopment and lower molars with a more mesially placedentoconid (Andrews 1974 Harrison 2002 2010 Cote et al 2013Hill et al 2013)
Ekembo differs from Turkanapithecus in having an expandedglabellar region less postorbital constriction a narrower temporalfossa with less flaring zygomatic arches the lacrimal fossa withinthe orbital margin a relatively broader palate with non-convergingtooth rows narrower nasal bones and interorbital region a distinctmandibular superior transverse torus a more robust mandibularcorpus taller mandibular ramus upper canines more rounded incross-section a reduced cingulum on cheek teeth awell-developedhypocone on M1e2 a more squared M2 with a protocone subequalto paracone and broader lower molars relative to length (Leakeyand Leakey 1986 Harrison 2002 2010)
Ekembo differs from Mabokopithecus in having a relativelybroader M3 crown without distal recurvature of the crown andlacking a buccal concavity a hypoconulid that is in-line with thebuccal cusps more bunodont cusps a well-defined protocristidand a broad open talonid without an accessory cuspid (Harrison2002 2010)
Ekembo differs from Xenopithecus in having reduced uppermolar flare a substantially reduced cingulum no prehypoconecrista molar occlusal profiles more closely approximating a squareand less constricted trigons (Hopwood 1933 Le Gros Clark andLeakey 1951 Harrison 2002 2010)
Ekembo differs from Morotopithecus in having a broader inter-orbital region a better developed canine fossa in males premolarssmaller relative to molars greatly reduced or absent buccalcingula on upper molars lumbar vertebrae with transverse pro-cesses positioned more ventrally on the centrum and a narrowerdistal femur with a deeper patellar groove and similarly-sizedmedial and lateral femoral condyles (Gebo et al 1997MacLatchy 2004)
Recently Stevens et al (2013) likened the new genus Rukwapi-thecus represented by a mandible fragment with well-preservedP4-M3 to nyanzapithecine species Ekembo can be distinguishedfrom this genus in having a more gradually rounded gonial angle aless obliquely oriented P4 with subequal protoconid and metaconidand a broader mesial fovea less mesially positioned metaconidreduced buccal cingula and smaller mesial foveae on the molarssmaller hypoconulids broader M1e2 relative to length a singularrather than twinned metaconid no accessory cuspules in thelingual notch and a well-defined crest between entoconid andhypoconulid
Family Proconsulidae (Leakey 1963)
TabDen
Fe
G
U
U
U
U
U
M
Lo
Lo
Lo
a
b
c
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 47
Genus Proconsul Hopwood 1933
Type species
Proconsul africanus Hopwood 1933
Included species
Proconsul africanus (Hopwood 1933) Proconsul major (Le GrosClark and Leakey 1950) Proconsul gitongai comb nov (Pickfordand Kunimatsu 2005) Proconsul meswae (Harrison and Andrews2009)
Diagnosis (replaces the emended diagnosis of Harrison 2010)
Proconsul species are medium- to large-bodied sexually dimor-phic catarrhines that can be distinguished from other proconsulidgenera by the following combination of features Robust mandib-ular corpus and symphysis characterized by a long shallowlysloping planum alveolare terminating at a large superior transversetorus and a subplanum surface oriented posteroinferiorly Upper
le 2tognathic features that distinguish Ekembo from Proconsula
ature Ekembo
eneralCusp morphology More bunodont occupying more of theMolar crests Inflated incorporated into cuspsCingulum Reducedpper central incisorLingual pillar Narrow generally low reliefMarginal ridges and cingulum Asymmetric with sharply angled cingulu
ridge transitionpper canineTip PointRoot fluting AbsentRoot striations Rare and weakly developedpper third premolarb
Mesiodistal cusp position Crown midpointMesial fovea Well developedBuccal profile SymmetricMesial crown margin Straight or slightly curvedDistal crown margin Slight to moderately curvedCusp heteromorphy ReducedCrown shape Converges on P4 shapeMesiobuccal flange Nopper fourth premolarParacone height Taller relative to crown lengthpper second molarc
Occlusal crown shape SquarishDistal margin Straight to slightly curvedBuccal cingulum Absent or rudimentaryDistolingual cingulum Merges into hypoconePosthypocone crista Distobuccally to buccally orientedBuccolingual flare ReducedandiblePlanum alveolare Short vertically inclinedSuperior transverse torus Moderate to well-developedOrientation of subplanum area More vertically orientedCorpus robusticity More gracilewer incisorsCrown shape Narrow relative to heightwer canineTip Pointwer molarsBuccolingual flare ReducedM1 M2 breadth M1 narrower relative to M2
Although presented here as discrete characteristics for descriptive purposes some oNearly all of these features tend to distinguish P4s as well but in that tooth the diffeThese features tend to distinguish M1s as well but in that tooth the differences are
incisors heteromorphic with I1 exhibiting an inflated lingual tu-bercle and a near symmetrical configuration of lingual marginalridges and cingulum with a curving transition from cingulum tothe mesial marginal ridge Lower incisors broad relative to crownheight Upper and lower canines with distinct blade-like or burin-like tips in both deciduous and adult teeth Upper canine rootswith broad longitudinal grooves or fluting defining two or threedistinct columns plus numerous small longitudinal striationsextending cervically from the root apex Upper P3s morphologicallydissimilar from P4s P3 has substantial cusp heteromorphy (para-cone much taller than protocone) with mesially positioned cuspsand a notched mesial border that together limit or eliminate amesial fovea A long steeply angled preparacrista contributes to ahighly asymmetric buccal profile before being interrupted near thetooth cervix by a mesially projecting flange The distal margin isstrongly curved P4 has a tall paracone relative to crown lengthMolars characterized by extensive cingular development individ-uated conical cusps connected by sharp occlusal crests and sub-stantial buccolingual flare Upper molars rhomboid in shaperesulting from a broad trigon and a distally expanded hypocone M2
has a distinct buccal shelf between paracone and metacone a dis-tolingual cingulum that wraps fully around the hypocone rather
Proconsul
occlusal surface area More distinct individuated conesSharper more distinct from cuspsExtensive
Inflatedbulbousm-mesial More symmetric with curving cingulum-mesial
ridge transition
Blade-likePresentVery common and well developed
MesialAbsent or reducedAsymmetricNotchedStrongly curvedStrongDistinct from P4
Yes
Shorter relative to crown length
RhomboidStrongly curvedDistinct cingular shelfDistinct from hypoconeDistally to distobuccally orientedPronounced
Long more horizontally orientedStrongly developedMore posteriorly inclinedMore robust
Broad relative to height
Blade-like
PronouncedM1 more similar to M2
f these features are undoubtedly correlated aspects of single evolutionary changesrences are not as pronouncednot as pronounced
Figure 2 Upper central incisors of Proconsul and Ekembo Lingual views of Ekembo(KNM-RU 1685 cast) left and Proconsul (KNM-CA 1300) right Note the differences inthe transition between the lingual cingulum and the mesial marginal ridge which ismore rounded in Proconsul and more angled in Ekembo The central pillar is also morebulbous or inflated and without ridging in Proconsul Additional specimens illustratingthe Proconsul morphology are shown in Pickford et al (2009 Fig 17 p 197) butreferred in that manuscript to Ugandapithecus
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6148
thanmerging into the hypocone on its lingual surface and a distallyoriented posthypocone crista Lower first and second molars nearlyequal in buccolingual breadth
Comparative morphology of Ekembo and Proconsul
Samples and comparative methods
The above diagnoses are based on extensive survey and analysisof the original fossil material Specimens housed in the NationalMuseums of Kenya (NMK) and the Natural History Museum (Lon-don) were analyzed by the authors during several separate and twojoint (NMK) research visits Fossils in the Uganda National Museumwere assessed by one of us (KPM) during two visits All measure-ments were collected by KPM Dental terminology follows Szalayand Delson (1979)
The following results are organized with respect to themorphology of the Kisingiri sample (attributed to E nyanzae andE heseloni) versus that of Proconsul from Tinderet and Napak(traditionally referred to P africanus P major and more recentlyP meswae) Specimens from additional localities that have at onetime been attributed to Proconsul (eg Andrews1978 Pickford andKunimatsu 2005 [cf Harrison 2010]) are for the most part notconsidered here most are middle Miocene in age and have beentransferred to other genera (Harrison 2002 2010) Of those that arefrom early Miocene sites there is growing evidence that P majorcan also be found at Moroto and differentiated fromMorotopithecus(Pickford et al 2009) an issue of some interest but ultimately notcritical to the taxonomic questions being considered here Whereappropriate however we do provide observations on a few earlyMiocene specimens attributed to ldquoUrdquo gitongai (Pickford andKunimatsu 2005 Pickford et al 2009 see Harrison 2010) Yetwhile we consider the middle Miocene holotype likely to representa distinct species here referred to Proconsul we concur withHarrison (2010) that many specimens placed in this species byPickford et al (2009) cannot be distinguished from P major
Sample sizes particularly from Tinderet and Ugandan localitiestend to be small but where possible standard univariate (eg t-tests) and bivariate (regression ANCOVA) statistical analyses wereperformed A single eigendecomposition of the covariance matrixof symphyseal measurements was done to characterize the multi-variate shape of the mandibular symphysis after accounting for sizedifferences (sensu Jolicouer 1963) Quantitative results are illus-trated as box-and-whisker plots of shape indices or bivariate plotsof the measurements For samples of fewer than five specimensshape index values are plotted in lieu of box-and-whiskersummaries
Morphological comparisons
Upper incisors Two features of the Ekembo upper central incisorsdistinguish them from those of Proconsul including specimenswidely attributed to the type species P africanus The lingual tu-bercle in Ekembo is quite narrow generallywith low relief and oftenscored by two or more irregular ridges in contrast to the muchmore inflated or bulbous tubercle in Proconsul I1s (Fig 2) Ekembospecimens also have a characteristic morphology in the transitionfrom the lingual cingulum to the mesial marginal ridge (Fig 2)from the mesiodistal mid-point of the crown the cingulumextends either directly mesially or even angles slightly cervicallyto intersect the mesial marginal ridge at a fairly sharp angle Incontrast in I1s of Proconsul the cingulum angles incisally from themid-point and transitions into the mesial marginal ridge in agradual curve The expression of these traits varies somewhat inthe smaller Tinderet specimens attributed to P africanus
compared with those of P major (cf Pickford et al 2009)However these variations are relatively minor with respect to thedifferences between the two genera Of 25 I1s known fromKisingiri localities only one specimen KNM-MW 562 somewhatresembles the Tinderet sample in both of these featuresdhavinga somewhat less angled transition from the mesial marginal ridgeto the cingulum and a more bulbous lingual tubercle This isparticularly interesting as the specimen was discovered in theMakira Beds now dated to older than 190 Ma and contemporarywith the Tinderet localities (McCollum et al 2013) Neverthelessits overall morphology is more similar to other Ekembospecimens than to Proconsul incisors Two other Ekembospecimens KNM-RU 1677 and RU 1933 have somewhat moreinflated lingual tubercles but otherwise resemble other Ekembo I1s
No consistent differences were found in upper lateral incisors
Upper canines Upper canine crowns of Ekembo are typical of thoseof most catarrhines in terminating in an evenly tapering point Thisdiffers from the Proconsul specimens which have a blade-like(Kelley 1986 Senut et al 2000 Harrison 2010) or ldquoburin-likerdquo(Pickford et al 2009) canine tip (Fig 3) While blade-like apicalmorphology has long been recognized as a distinctivecharacteristic of P major we confirm the report by Pickford et al(2009) that this feature is also present in the P africanus typespecimen (Fig 3a) although somewhat understated as expectedin the more slender and higher-crowned P africanus caninesUnfortunately other canines attributed to P africanus (eg M44837 KNM SO 419 LG 921 CA 1910 SO 5353 Kelley 1986) havebroken apices and therefore cannot be evaluated for this featureA blade-like tip is also present in the deciduous upper canines ofP major (eg KNM-SO 371 MV 10 NAP IV UMP 62-20) andP meswae (KNM ME 11) Hence the blade-like upper canine tipunites all Tinderet and Napak Proconsul species but is neverexpressed in Ekembo specimens from Rusinga and Mfangano
Two features of the canine roots also differentiate these generalongitudinal grooving or fluting along the length of the root (cfKelley 1986) and the development of numerous finer longitudinal
Figure 3 Canines of Proconsul and Ekembo a) Upper canine of the P africanus holotype (M 14084) lingual view b) Upper canine of P major (KNM-CA 2127) lingual view c) Lowercanine of P major buccal view d) lower canine of Ekembo cf nyanzae (KNM-RU 1676) buccal view and e) Upper canine of P major (KNM-SO 584) distal view Note the blade-like orldquoburin-likerdquo tip of the unworn Proconsul canines including on the nearly unworn canine tip of the P africanus type specimen (a) Ekembo canines have more typical pointed tipsProconsul canines also have longitudinal grooves and fine striations on the root (e) Ekembo lacks these features Scale bars frac14 1 cm
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 49
striations near the root apex that extend to varying degrees towardthe cervix of the tooth Longitudinal ldquoflutingrdquo consists of broadgrooves that segment the root into distinct sections (cf Pickfordet al 2009 Fig 3e) Most often the root in cross-section has theappearance of a cylinder that has been sectioned longitudinallywith the two halves offset from each other but in some cases thereare three segments and the root cross-section is more triangular inappearance (Kelley 1986) This characteristic was noted by Kelley(1986) in the P africanus canines but not in those of P majorHere we affirm that fluting is present on all of the visible uppercanine roots attributed to both P africanus and P major rootmorphology could not be evaluated in the deciduous upper caninefrom Meswa Bridge since it is implanted in the maxillary alveolusNone of the Ekembo specimens display this morphology havinginstead a simpler round to ovoid cross-section along the length ofthe root
The fine longitudinal striations on Proconsul upper canine rootsvary in the degree to which they extend toward the cervix (Fig 3e)but they are visible in all specimens attributed to P africanus(KNM-SO 419 521 921 5353 KNM-X 502) and five of six uppercanines of P major (excepting KNM-SO 1614) This morphology isalso present in UMP 62-12 which was included as part of theMorotopithecus bishopi hypodigm (Gebo et al 1997) Fine striationsalong the upper canine root are almost entirely absent in EkemboTwenty of twenty-three specimens lack this feature three speci-mens (KNM-RU 1763 1845 1891) exhibit incipient to slight
striations but these are never developed to the extent seen inProconsul canines
Upper premolars Upper third premolars display some of thegreatest differences between Ekembo and Proconsul P4s tend todiffer in similar ways but the differences are not nearly as markedEkembo P3s have cusps positioned near the mesiodistal midpoint ofthe crown well-developed mesial foveae pre- and postparacristaeof similar length and slope resulting in symmetric or nearsymmetric buccal profiles a straight or slightly curved mesialmargin and only moderately curved distal margin (Fig 4)Proconsul P3s are more similar in appearance to those of small-bodied catarrhines from the early Miocene with cusps positionedmesially on the crown greatly reducing or eliminating the mesialfovea They also have a steep preparacrista and more shallowlysloped postparacrista resulting in a highly asymmetric buccalprofile and an inwardly angled or notched mesial margin but astrongly curved distal margin This morphology is bestcharacterized in the P africanus holotype (M 14084) but can alsobe seen in other specimens attributed to this species as well asthose referred to P major (Fig 4d e) Also noteworthy is adistinctive mesiobuccal flange that mesially extends the buccalportion of the crown but only near the cervical line of the toothPickford et al (2009) describe this feature in some detail forldquoUgandapithecusrdquo but in fact this morphology is clearly expressedin the P africanus holotype as well (Fig 4a d) A single adult P3
Figure 4 Upper premolars of Proconsul and Ekembo occlusal (aec) and lingual (def) views a) P3e4 of the P africanus holotype (M 14084) b) P3 of P major (M 14331) c) P3e4 of theE heseloni holotype (KNM-RU 2036) d) P3 of the P africanus holotype (M 14084) e) P3 of P major (M 14331) f) P3 of the E heseloni holotype (KNM-RU 2036) Note the mesiallypositioned cusps lack of mesial fovea and strongly asymmetric crown in Proconsul compared to Ekembo (see text for further comparisons) Image of M 14331 reversed for easiercomparison Photo 4a courtesy of Rutger Jansma Scale bars frac14 1 cm
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6150
germ and two dP3s attributed to P meswae look remarkably similarto those of other Proconsul species including the presence of thisflange In contrast the Ekembo P3 has a well-developed mesialfovea that terminates at a mesiobuccal shoulder positionedapproximately halfway between the paracone tip and the cervixFrom this shoulder the mesial margin angles slightly distallydown to the cervix (Fig 4f)
The degree of cusp height heteromorphy in P3 also differentiatesEkembo from Proconsul (contra Pickford et al 2009) Only four non-Kisingiri specimens were sufficiently unworn to take measure-ments but more worn specimens also appear to express this dif-ferential in cusp height Figure 5a shows the ratio of protoconeparacone height measured from the tooth cervix and with only oneexception Ekembo P3s have substantially less cusp heteromorphyThis exception KNM-RU 1677 does not in fact have greater heter-omorphy but instead has an abnormal extension of the enamel ontothe buccal (paracone) root which skews the measurement Hencecusp relief relative to the occlusal surface in RU 1677 is not nearly asheteromoprhic as in Proconsul and when this is taken into accountthe difference between genera is significant (t-test dffrac14 11 t frac14 283p frac14 00165) The adult P3 germ from Meswa Bridge is incompletelyformed and therefore could not be measured However the differ-ence between paracone and protocone heights with respect to theocclusal surface appears to be as great as that in the P africanusholotype It does not appear that sex is an important determinant ofthis feature since well-established male (eg M16647 KNM-RU16000) and female (KNM-RU 2036 7290) specimens do not segre-gate across the Kisingiri range of variation
Upper fourth premolars of Ekembo are broadly similar inmorphology to Ekembo P3s in fact the similarity between P3 and P4
is substantially greater in Ekembo than in Proconsul (Figs 4a c)Likewise most of the morphological differences that discriminatethe P3s of these genera also tend to differentiate the P4s but less
clearly Cusp heteromorphy does not distinguish P4s of thesegenera but the overall height of the P4 paracone in Ekembo issignificantly greater relative to crown length than in Proconsul P4s(t-test df frac14 14 t frac14 316 p frac14 00069 Fig 5b)
Upper molars Cusps of Ekembo upper molars are uniformly morebunodont than those of Proconsul occupying more of the occlusalsurface Intercuspal crests are more fully incorporated into thecusps in Ekembo molars giving them a more inflated appearanceThis is particularly evident in the crista obliqua which is very broadin Ekembo but thin and knife-like in Proconsul (Fig 6) Cusps ofProconsul molars stand more as individual cones Pickford et al(2009) have further differentiated the cusp morphology ofsmaller and larger Tinderet specimens arguing that the latterhave more bulbous cusps While accurate this difference is minorcompared with the morphological difference between Ekemboand Proconsul and we regard the within-Proconsul cuspalvariation as size-related In fact the intermediately sizedP meswae has cusp morphology that is appropriatelyintermediate between that of P africanus and the larger P major
Other differences between Ekembo and Proconsul upper molarsare most evident in M2 We concur with previous assessments thatthe Kisingiri upper molars have less developed cingula than non-Kisingiri molars (eg Senut et al 2000 Harrison 2002 2010Harrison and Andrews 2009) More specifically Ekembo M2s lackor have only rudimentary buccal cingula between the paracone andmetacone Likewise the lingual cingulum is discontinuous merg-ing into the hypocone lingually and then re-emerging as a distalmarginal ridge (Fig 6) Proconsul M2s display substantially morecingular development overall and in particular exhibit a prominentbuccal shelf between the paracone and metacone and a distinctdistolingual cingulum that is continuous around the hypocone towhere it merges with the distal marginal ridge
Figure 5 Box-and-whisker plots a) P3 cusp heteromorphy index (protocone heightparacone height) Results of a t-test show this difference to be significant (p frac14 00165) Theexcluded Ekembo outlier KNM-RU 1677 only appears to have extreme heteromorphy because of extensive expansion of the buccal enamel onto the anterior root (see text) b) P4
relative paracone height (paracone heightmesiodistal crown length) Results of a t-test show this difference to be significant (p frac14 00069)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 51
The occlusal profiles of Ekembo M2s are squarish in shape withrelatively straight and equal sides with a posthypocone crista thatis distobucally to buccally directed In contrast the occlusal profilesof Proconsul M2s have a more rhomboid shape with substantialbasal flare around the mesial cusps and a distally expanded hypo-cone with a distally directed posthypocone crista resulting in astrongly curved distal margin (Fig 6) Even in specimens with lesscurved distal margins the posthypocone crista is still more distallydirected compared to the condition in Ekembo These features arecharacteristic not only of P africanus P meswae and P major butare also expressed in the isolatedM2 fromMoroto Uganda (MOR IIb2098) attributed by Pickford et al (2009) to ldquoUrdquo gitongai butregarded here as belonging to P major
As noted by others (eg Harrison and Andrews 2009) the de-gree of buccolingual molar flare differs among these samplesQuantified as the distance between the tips of the mesial cuspsdivided by maximum crown breadth the degree of molar flare in
Ekembo M2s is significantly less than in Proconsul M2s (t-testdf frac14 29 t frac14 396 p frac14 00004) In a bivariate plot of these twomeasures the distributions of Ekembo and Proconsul teeth barelyoverlap with only KNM-RU 1677 encroaching into Proconsul ter-ritory (Fig 7)
These differences in M2 morphology also tend to distinguishEkembo from Proconsul M1s but not always as clearly Whereas M2smore consistently express distinct morphotypes the M1s in bothgenera show more variation in these features
The degree of upper third molar development (ie whether M3
has four distinct cusps or the distal cusps are reduced or absent) haslong been cited as taxonomically informative for these species orgroups of these species (eg Hopwood 1933 Le Gros Clark andLeakey 1951 Andrews 1978 Pickford et al 2009) However wefound this tooth to be unreliable for taxonomic purposes Withinprimates M3s are typically the most variable molars in both sizeand shape (Blankenship et al 2007 Kavanagh et al 2007) and this
Figure 6 Occlusal view of the M2s of Proconsul and Ekembo Left P africanus holotype(M 14084) Right E heseloni holotype (KNM-RU 2036) Note in Proconsul the overalldifference in crown shape as well as the more conical individuated cusps and nar-rower cristae strongly developed lingual cingulum that continues to the distal aspectof the crown broad buccal cingular shelf and more distally directed posthypoconecrista In Ekembo the cusps are more bunodont and expanded the cristae are broaderwith better developed accessory tubercles the lingual cingulum merges into thehypocone there is no buccal shelf and the posthypocone crista is more buccallydirected
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6152
is observable even in small samples of Proconsul species TheP africanus holotype has a substantially reduced M3 whereas inKNM-CA 2250dattributed by most authors to this species(Andrews 1978 Pickford et al 2009)dit is fully developed Simi-larly the hypodigm of P major includes both fully developed M3s(eg KNM-CA 389 LG 1815) and specimens with reduced ormissing hypocones (KNM-CA 397 1299) Ekembo displays compa-rable ranges of variation from highly reduced to fully developedM3s in both species Thus these species and genera cannot bedistinguished based on M3 development and we did not find anyother distinguishing characteristics between Ekembo and Proconsulin the upper third molars
Figure 7 Scatter plot depicting the de
Mandible Pickford et al (2009) enumerated several traits thatreadily distinguish Kisingiri from non-Kisingiri mandibles andthese distinctions are entirely consistent with the distinction ofEkembo from Proconsul Our analyses specifically underscoredifferences in the symphyseal region The planum alveolare inboth male and female specimens of Ekembo is short and morevertically oriented even the most elongate of these (KNM-RU47805 McNulty et al 2007) is substantially more verticalcompared to specimens of Proconsul In contrast the planumalveolare in Proconsul is conspicuously long and nearlyhorizontally oriented Well-preserved mandibles attributed toP major (eg KNM-SO 396 and 404) exhibit this morphology inextreme fashion (Fig 8) smaller specimens (KNM-SO 1112P africanus) show less extreme expression but are still readilydistinguishable from Ekembo mandibles For its size KNM-LG 452has a surprisingly short planum alveolare though not to thedegree seen in Rusinga mandibles Correlated to the shortplanum alveolare in Ekembo is a reduced superior transversetorus which while well-developed does not approach therobusticity expressed in Proconsul mandibles This difference isreflected in an index of symphyseal robusticity (thicknessperpendicular to the long axis of the symphysislength of thelong axis) which shows Ekembo and Proconsul to be significantlydifferent (t-test df frac14 14 t frac14 430 p frac14 00007)
Development of the inferior transverse torusdoften used todifferentiate among species of Proconsul (eg Pickford et al 2009Harrison 2010)dwas found to be highly variable and inconsis-tently expressed within these samples However the overallorientation of the subplanum region (as defined by a line con-necting the posterior-most midline points of the superior andinferior transverse tori or the posterior-most midline point of theinferior margin of the symphysis in the absence of the latter) doesdistinguish Ekembo from Proconsul mandibles The orientation ofthis line in relation to the alveolar margin in the vicinity of thepostcanine teeth was assessed qualitatively by placing a small rod
gree of buccolingual flare in M2
Figure 8 Mandibles of Proconsul and Ekembo Occlusal views of P major a) KNM-SO 396 b) KNM-SO 404 c) KNM-LG 452 Occlusal views of Ekembo d) KNM-RU 2087 e) KNM-RU1674 f) KNM-RU 7290 Occlusal views of P africanus g) KNM-SO 372 h) KNM-SO 1112 i) Lingual view of P africanus (KNM-SO 1112) Note the long more horizontally orientedplanum alveolare in Proconsul resulting in a pronounced superior transverse torus The planum alveolare is shorter and more vertical in Ekembo such that one can see the inferiormandibular border from occlusal view
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 53
(eg a dental pick) in themidline internally along the two referencepoints and recording the point in lingual perspective at which therod intersected the alveolar margin (Fig 9) In Ekembo the sub-planum region is more vertically oriented so that the rod intersectsthe alveolar margin most often in the premolar field Not surpris-ingly there is a difference between sexes with females having areduced superior transverse torus and therefore a more verticallyoriented subplanum region compared to males In contrast withEkembo Proconsul mandibles have much more inclined subplanumsurfaces with the rod intersecting the lingual alveolar margin inthe vicinity of the M1 (females) or M2 (males Fig 9) When sex istaken into account therefore the distributions of Ekembo andProconsul in this feature are discrete Note that male and femalemandibles attributed to both P africanus and P major are includedin this assessment and are surprisingly similar despite dramaticsize differences
We also quantified internal symphyseal morphology using threemeasures symphyseal height (infradentaleegnathion) planumalveolare length (posterior-most point on the superior transversetoruseinfradentale) and subplanum length (posterior-most pointon the superior transverse torusegnathion) a combination thatincorporates not only the lengths of these intervals but also theangular relationships among them To compare symphyseal shapeacross a range of sizes we chose a simple principal componentsapproach to partition size onto the first eigenvector (cf Jolicouer1963) While this is not the preferred method for partitioning sizein most datasets (Jungers et al 1995) for this particular sample andset of measurements Mosimann shape variables were still
significantly correlatedwith size (see discussion in Coleman 2008)Hence we assessed differences in symphyseal morphology be-tween Ekembo and Proconsul using the second and third principalcomponents of the three variables Distributions of the two generaare distinct in this shape space (Fig 10 Supplementary OnlineMaterial [SOM] Table 1)
Robusticity of the mandibular corpus also differs between thesesamples though it is more difficult to test due to differentialpreservation among specimens Corpus thickness and depth underP4 can be measured in the greatest number of specimens andcomparison of these measures suggests a difference betweengroups in the scaling of corpus robusticity (Fig 11) Among largepresumably male specimens Ekembo mandibles are substantiallyless robust than those of Proconsul However among smallermandibles only a single Proconsul specimen (KNM-SO 372) could beincluded and it is similar in robusticity to Ekembomandibles of thissize Considered together the impression is that corpus robusticitymay scale differently in Ekembo than it does in Proconsul and a testfor differences in slopes using an ANCOVA design of the loggedvariables was significant (df frac14 2 F frac14 2481 p lt 00001) Never-theless additional specimens are needed to properly evaluate theinfluence of taxonomy sex and size on corpus robusticity areasonable alternative interpretation of Figure 11 is that it dem-onstrates a significant difference between P major and all otherspecies of both Proconsul and Ekembo
Lower incisors Ekembo specimens are uniformly narrower inmesiodistal length than incisors attributed to P africanus P major
Figure 10 Scatter plot of principal component scores based on an eigenanalysis of threeposterior point on the superior transverse toruseinfradentale) and subplanum length (mprimarily captures size differences among specimens (Jolicouer 1963 see text and SOM Trespectively) in the symphyseal variables
Figure 9 Line drawing of a hemimandible depicting different orientations of the sub-planum area in Ekembo and Proconsul Females are denoted using shorter vectors maleswith longer vectors Vector is estimated in M 14086 which is broken at the midlineEkembo specimens have a more vertical subplanum compared to those from Songhorand Legetet but within both groups females have more vertically oriented regions thanmales Hence even the most ventrally rotated subplanum in the largest Ekembo malespecimen (KNM-RU 47805) groups with the Tinderet females in this feature
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6154
and P meswae both absolutely and in relation to crown height(Fig 12) This is true for both I1 and I2 A t-test adjusted for non-independent observations (eg I1 and I2 from the same mandible)confirmed the difference as significant (t-test df frac14 12 t frac14 258p frac14 00242) Moreover while the measurable Proconsul samplewas small this same pattern is observable in other specimenswhose tips were slightly to moderately worn and therefore notmeasured
Lower canines The only lower canine trait that discretely sepa-rates these genera is the pointed crown tip in Ekembo (Fig 3d)contrasted with a blade-like (or ldquoburinrdquo) tip in Proconsul(Fig 3c) This feature is well established for P major but is alsoexpressed in KNM-CA 1772 which was referred to P africanus byKelley (1986) and is too small to be either P major or P meswaeTwo other specimens commonly attributed to P africanus KNM-CA 2149 and KNM-SO 1112 both have broken canine tipsLikewise a new specimen (Kor 65004) attributed to P africanusby Pickford et al (2009) also has a broken tip and cannot beevaluated (see Pickford et al 2009 Fig 23) As with the uppercanines the blade-like tip is also present in the deciduous lowercanines of P major (eg KNM-CA 361 S0 5352) and P meswae(KNM-ME 1)
Lower premolars Like most previous authors (eg Andrews 1978Harrison 2002 2010 Pickford et al 2009) we found neitherqualitative nor quantitative characteristics of the lower premolarswith any taxonomic value
Lower molars Differences in lower molar morphology are few andrelatively subtle but in some cases mirror those seen in the uppermolars Cusps on the lower molars of Ekembo are more bunodontthan those in Proconsul and occupy more of the occlusal surfaceBroad inter-cuspal crests are incorporated into the Ekembo cuspsmaking them appear more inflated Molar cusps of Proconsul are
variables symphyseal height (infradentaleegnathion) planum alveolare length (mostost posterior point on the superior transverse torusegnathion) The first componentable 1) and thus PC 2 and PC 3 illustrate the residual shape variance (72 and 28
Figure 11 Scatter plot showing mandibular corpus robusticity under P4 Trend lines were computed using ordinary least squares (OLS) regression Similar relationships areobservable at other parts of the corpus but differential preservation results in smaller samples at those positions Slopes are significantly different (p lt 00001 tested on loggedvariables)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 55
more distinct from one another and are linked by narrower sharperinter-cuspal crests The lower molars of Ekembo also have asubstantially less developed buccal cingulum than those ofProconsul (Harrison 2002 2010 Harrison and Andrews 2009)
Figure 12 Scatter plot of lower incisor crown proportions Differenc
Lastly and again as seen in the upper molars the two genera canbe distinguished by the significantly greater amount of lowermolar flare in Proconsul (Fig 13 t-test df frac14 59 t frac14 713p lt 00001 see also Harrison and Andrews 2009)
es between Ekembo and Proconsul are significant (p frac14 00242)
Figure 13 Box-and-whisker plot of buccolingual flare in lower molars measured asthe average distance between the mesial buccal and lingual cusps divided by overallbuccolingual crown breadth Differences between Ekembo and Proconsul are highlysignificant (p lt 00001)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6156
Relative molar size has been used to distinguish species ofProconsul but we observed the primary difference to be in relativebuccolingual breadths of M1 and M2 In Ekembo M1 is substantiallynarrower than M2 than is the case in Proconsul (Fig 14 Harrison2002 2010) Although none of the smaller Proconsul mandiblespreserve complete crowns for both M1 and M2 KNM-SO 1112 hasan intact M2 and at least preserves the cervix for M1 However evenif the cervical measurement is used for its M1 breadthdthusunderestimating the true crown breadthdKNM-SO 1112 still has abroader first molar in relation to M2 than would be expected inEkembo If crown breadth is estimated from the ratio of cervixbreadth to maximum crown breadth in other Proconsul fossils therange of likely breadths for the SO 1112 M1 clearly distinguishes it
from the Ekembo sample (Fig 14) Testing this relationship onlogged variables using the mean estimate for the SO 1112 M1(Fig 14) reveals a significant difference between the two groups(ANCOVA df frac14 1 F frac14 2424 p frac14 00002)
As with upper third molars we find a great deal of variation inM3 morphology and mistrust the taxonomic significance some-times placed on characteristics of this toothdat least with regard tothese taxa Harrison (2002 2010) cited the mesiodistal position ofthe M3 entoconid as differentiating Kisingiri third molars fromthose attributed to P africanus We concur with his assessment butfound the entoconid position to vary in specimens attributed toP major Thus this character does not differentiate Ekembo from allProconsul species but may distinguish it from the type speciesP africanus
Discussion
The distinctiveness of the Kisingiri large catarrhine materialfrom that at Tinderet and Ugandan localities has been noted pre-viously by several authors (Pickford 1986 Walker et al 1993Begun 2001 2007 MacLatchy and Rossie 2005 Harrison andAndrews 2009 Harrison 2010) What had not been fully docu-mented was the nearly complete lack of overlap between thesesamples in a number of features despite very large assemblagesfrom Rusinga and Mfangano On descriptive criteria alone (Table 2)many isolated teeth (I1 C1 P3 M2 C1) and all of the mandibles canbe assigned to either Proconsul or Ekembowith very little chance ofmisattribution That some of the most diagnostic features occur inthe upper dentitiondthe only anatomy preserved in theP africanus type specimendadds weight to the taxonomicarrangement proposed here The morphology of the upper teeth inall Proconsul species compellingly unites them into a single cladedistinct from E heseloni and E nyanzae This is reinforced bynumerous features of the mandible and mandibular dentition thatfurther distinguish these samples The number and variety of thesefeatures are such that Proconsul and Ekembo are more differentfrom one another than are many closely-related modern catarrhinegenera
Derived morphology of Ekembo and Proconsul
It is premature to formally assign primitive and derived status tothe individual features of Ekembo and Proconsul before completinga formal phylogenetic analysis This is especially true given theabundance of evidence available for Ekembo and hence the poten-tial impact that our generic revision could have on phylogeneticinterpretations Nevertheless in comparison with both extanthominoids and the small-bodied catarrhines from the earlyMiocene it is informative to differentiate the evolutionary grades ofthese genera Many characteristics of Ekembo are more advanced inthe direction of modern apes compared to those of Proconsul(Table 2) General trends such as expanded and more bunodontcusps broader molar crests reduced cingulum and morphologicalsimilarity between P3 and P4 all represent morphologies thatpresage those of later fossil and extant hominoids The moreadvanced morphologies of P3 and M2 are particularly conspicuouscompared not only with those of Proconsul but to most earlyMiocene catarrhines In contrast the mesially positioned stronglyheteromorphic cusps on the P3 of Proconsul plus the correspondingasymmetric buccal profile notched mesial margin and missingmesial fovea are characteristic of other early Miocene catarrhinessuch as Dendropithecus Rangwapithecus and Limnopithecus andare likely primitive among East African catarrhines The moresymmetric P3 of Ekembo with midline positioned cusps andreduced cusp heteromorphy is reminiscent of later hominoids and
Figure 14 Relative buccolingual dimensions of M1 and M2 KNM-SO 1112 has a preserved M2 but only the cervix of M1 Its M1 breadth was estimated based on the ratio to cervixbreadth to maximum crown breadth in other Tinderet fossils Even the cervical breadth of SO 1112 is broader than would be predicted for the crown breadth of a Kisingiri M1 Trendlines computed using OLS regression and using the average estimate of SO 1112 M1 breadth Group differences were significant (p frac14 00002)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 57
even hominid taxa such as Rudapithecus (eg Kordos and Begun2001)
Proconsul is likewise united by a suite of features that are for themost part characteristic only of this lineage (Table 2) Themorphology of both upper and lower canines is unique to thisgenus and shared by all three well-known species plus the poorlyrepresented middle Miocene P gitongai (Pickford and Kunimatsu2005) The mandibular symphysis is also distinctive in Proconsulcompared with other catarrhines with its very long and shallowlysloping planum alveolare and steeply inclined subplanum surfaceIndeed even the small edentulous anterior mandibular fragmentKNM-SO 372 is readily placed within Proconsul and excluded fromother similarly sized catarrhine taxa such as Rangwapithecus andEkembo based on this unique symphyseal morphology (Figs 8 and9)
Other differences between Ekembo and Proconsul are of uncer-tain evolutionary polarity (eg orientation of the posthypoconecrista) or may be of taxonomic relevance without broader phylo-genetic significance (eg degree of molar flare Table 2) Werecognize that there are morphological differences within Procon-sul particularly between the better known species P africanus andP major However many of these differences are likely related tosize and represent variations on a single generic theme that aremarkedly different from the morphology of Ekembo The recentdescription (Harrison and Andrews 2009) of the intermediatelysized P meswae underscores the broad morphological unity withinProconsul and further substantiates a well-defined Proconsul cladecomprising Tinderet and Ugandan species
Based on the abundant morphological and morphometric evi-dence supporting genus-level distinction between Proconsul andEkembo we are currently conducting a more detailed phylogeneticanalysis of all early Miocene catarrhines in order to better under-stand the distribution and evolutionary significance of these andother morphological features We anticipate that the debates sur-rounding the phylogenetic relationships of an historically consti-tuted sample of Proconsul can be more satisfactorily resolved nowthat the Kisingiri material is appropriately recognized as a distinctgenus
The generic disposition of ldquoUgandapithecusrdquo species
A great many features said to characterize ldquoUgandapithecusrdquowere added to the original diagnosis of the genus by Pickford et al(2009) in their systematic revision of Proconsul (sensu lato) As partof that study they also erected a new species ldquoUrdquo legetetensis andtransferred P meswae to ldquoUgandapithecusrdquo Evaluating the same setof features across the entire available sample however we wereunable to find support for this more expansive conception ofldquoUgandapithecusrdquo for several reasons First some diagnostic fea-tures cited in Pickford et al (2009) were quite variable within theldquoUgandapithecusrdquo sample As an example marked reduction of theM3 entoconid to a bead on the lingual cingulum is not shared byseveral ldquoUrdquo major specimens such as KNM-CA 393 NAP I 49000and Kor 253004 (see Pickford et al 2009 Fig 33e) and was likelynot a feature of a few of the more worn specimens eg BUMP 600and BUMP 601 or even the holotype of P major M 16648 Second
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6158
some diagnostic features of ldquoUgandapithecusrdquo were found to bevariably present in Ekembo arguing against their utility for dis-tinguishing ldquoUgandapithecusrdquo from Pickford et als (2009) com-bined P africanus thorn Kisingiri conception of Proconsul For examplesignificantly reduced curvature of the I1 distal margin in ldquoUganda-pithecusrdquo is also found in some Kisingiri specimens including KNM-RU 1685 1712 1714 1831 1846 and 1951 several more Rusingaspecimens have moderately reduced curvature Third some fea-tures that purportedly distinguish ldquoUgandapithecusrdquo fromP africanus are likely related to size For example the somewhatmore central position of the P3 paracone in ldquoUrdquomajor is not seen inthe smaller ldquoUrdquo meswae whose paracone is positioned similarly tothat of the holotype of P africanus (see Harrison and Andrews2009 Fig 6d)
However the major part of the evidence for recognizingldquoUgandapithecusrdquo (sensu Pickford et al 2009) was assembled by re-assigning specimens traditionally attributed to P africanus into newspecies of ldquoUgandapithecusrdquo The P africanus holotype was left inassociation with the Rusinga sample despite having morphologyoverwhelmingly characteristic of other Tinderet specimens Theresulting specimen allocations left P africanus morphologicallyimpoverisheddstripped of much of its hypodigmdso that Pro-consul for Pickford et al (2009) came to be characterized primarilyby the Rusinga sample (see similar comments in Harrison andAndrews 2009) Within that conceptual framework any spec-imen that differs from those at Rusinga could not be Proconsul butmust instead belong to ldquoUgandapithecusrdquo Thus specimens such asthe KNM-SO 1112 mandible were erroneously assigned to ldquoUgan-dapithecusrdquo because they were (correctly) perceived as differingfrom their Rusinga counterparts (Pickford et al 2009)
Here we restore the P africanus hypodigm and associate it withthe other Tinderet fossils according to their shared morphologySpecimens assigned to ldquoUgandapithecusrdquo revert to Proconsul andmany of the features cited to support a speciose ldquoUgandapithecusrdquo(Pickford et al 2009) instead become diagnostic features of Pro-consul This correction serves to highlight the distinctiveness of theKisingiri remains and the need to recognize this distinctiveness atthe generic level
As demonstrated above specimens from Kisingiri do not displaythe morphology of Proconsul as represented by the holotype ofP africanus M 14084 Given that this specimen preserves only theupper canine and cheek teeth similarities with or departures fromthis morphology represent the most compelling arguments forassessing inclusivity in the genus However for the mandible andlower dentition if one simply sorts the specimens by elementwithout relying on taxonomic preconceptions the overwhelmingdistinction within the traditionally constituted Proconsul sample isbetween Kisingiri material on the one hand and that from Tinderetand Uganda on the other (cf Walker et al 1993 Begun 2001 2007MacLatchy and Rossie 2005 Harrison and Andrews 2009Harrison 2010) Additionally with the Kisingiri specimensremoved to Ekembo the postcranial differences used to partlyjustify a distinct ldquoUgandapithecusrdquo lose their relevance (seeHarrison and Andrews 2009)
We have differed among ourselves in our past treatment ofP major as generically distinct from the smaller species of Pro-consul (Begun 2007 McNulty 2010) Morphologically many ofthe unique features of P major can be interpreted as size-relatedvariants on the smaller P africanus morphology Even if so thequestion is whether these are sufficiently different and numerousto warrant generic distinction that is do they suggest thatP major belongs in a separate clade from P africanus andP meswae As we have demonstrated differences among thesethree species are certainly fewer and show less morphologicaldistinction than those between Ekembo and any of the three
Nevertheless while we ultimately agree with Harrison andAndrews (2009 see also Begun 2007) that the features dis-tinguishing ldquoUrdquo major from P africanus are consistent with thosethat distinguish congeneric species and are at least partly due todifferences in size we concede the possibility that the largestProconsul species could belong in a separate genus (Senut et al2000 Pickford and Kunimatsu 2005) However whether or notone recognizes a separate genus for P major the P africanus ma-terial does not sort morphologically with the Kisingiri materialTherefore the taxonomic alternatives are either two generaProconsul and Ekembo or three Proconsul Ugandapithecus andEkembo with Proconsul and Ugandapithecus united in a cladeexclusive of Ekembo Thus while we strongly favor the firstalternative recognizing Ugandapithecus does not change thefundamental argument concerning the need to refer the Kisingirispecies to a generically and cladistically distinct taxon from thespecies represented at Tinderet and the Ugandan sites
Specimens of interest
Despite the substantial dental and gnathic differences betweenProconsul and Ekembo there are a few specimens whose taxonomicaffinities require additional scrutiny
KNM-RU 1676-77 This set of associated upper and lower teeth is aconsistent outlier among E heseloni and E nyanzae specimens Insome ways this specimen more closely resembles Tinderet Pro-consul in the features that distinguish it from Ekembo Molar cuspmorphology and cingulum development I1 lingual tubercledevelopment and M2 shape and posthypocone crista orientationare more consistent with specimens from the Tinderet sites On theother hand it otherwise resembles Ekembo and lacks the strongestdiagnostic features that unite species of Proconsul such as thedistinctive canine and P3 morphology Overall this specimen doesnot fit unambiguously into either genus in fact the degree ofbuccolingual flare cusp morphology enamel wrinkling and espe-cially the long extension of enamel onto the anterior root in P3 arein some ways reminiscent of Afropithecus although it clearly differsfrom that genus as well (cf Rossie and MacLatchy 2013)
Were these characteristics individually distributed among thelarge Kisingiri sample one might attribute this to intra-genericvariation but when found together in a single specimen from alarge sample it suggests a unique individual One of us (KPM)thinks it likely that the specimen was not actually found at aKisingiri locality based on its unique preservation compared toother Rusinga fossils and on other examples of mis-proveniencedspecimens (Leakey 1967 Andrews and Molleson 1979 Pickford1986 Kelley 1986) However we recognize that relying toofrequently on this explanation allows almost any scenario to bejustified and suggest some alternative possibilities One is that thespecimen was found on Rusinga but in the much older WayandoFormation strata instead of the Hiwegi Formation The differencesin morphology in that case might reflect more than a million yearsof temporal change (McCollum et al 2013) with KNM-RU 1676-77preserving certain more primitive Proconsul-like features As analternative it is reasonable to assume that fossil assemblages asvast as those from Rusingawill include not only abundant taxa butalso rare or infrequent inhabitants of the local biome There aremany examples of this from Rusingas mammalian communityeg ldquoHemicyonrdquo Kulutherium Tadarida cf Hyainailouros cfArchaeobeledon Kelba (see relevant chapters in Werdelin andSanders 2010) As such KNM-RU 1676-77 may simply representa rare and well-preserved specimen of some other catarrhinetaxon that lived among or visited a primate community dominatedby Ekembo
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 59
KNM-SO 1112 This specimen was initially referred to Rangwapi-thecus but soon after was transferred to Proconsul afrianus (Bosler1981 Martin 1981)da designation that most subsequent authorshave followed (Pickford 1986 Nengo and Rae 1992 Kelley 19861995 Walker et al 1993 MacLatchy and Rossie 2005 Hill et al2013) Pickford et al (2009) more recently referred this specimento ldquoUrdquo meswae citing two primary reasons First they reiteratedBoslers (1981) concern that its P3 differs from others attributedto P africanus However at that time the majority of what wasconsidered to be P africanus would have been from RusingaIsland In fact while noting its contrasting morphology Boslersuggested that differences between KNM-SO 1112 and other P3smight reasonably be attributed to sexual dimorphism notingthat this specimen provides ldquoa not implausible match for the Pafricanus holotype maxilla M 14084rdquo (1981152) Thatassessment is borne out in this study Using the much largersample of Ekembo P3s as a model variation can be seen toencompass specimens with well-developed mesial beaks andthose with more bilaterally compressed crowns and parallelsides The latter morphology is predominantly found amongmale specimens (KNM-RU 1674 1924 and KPS I and III) thussupporting Boslers (1981) suggestion that this variation isrelated to sexual dimorphism and hence that KNM-SO 1112 is amale P africanus
The second feature cited by Pickford et al (2009) in referringthis specimen to ldquoUrdquo meswae was posterior shallowing of themandibular corpus However because they redefined ldquoUgandapi-thecusrdquo to include all of the well-preserved Tinderet mandibles(Pickford et al 2009)dleaving Proconsul to be represented only byRusinga specimensdtheir only alternative within this scheme for aspecimen the size of SO 1112 was to refer it to ldquoUrdquomeswae It seemsodd though to assign this specimen to a species from a (perhapsmuch) older site when the Proconsul africanus holotype is ofappropriate size and similar age and as there are no other adultmandibles from either species with which to compare it In anycase mandibular shallowing is variable in both Ekembo and Pro-consul with some Kisingiri specimens exhibiting considerableshallowing (eg KNM-RU 2087) and some ldquoUgandapithecusrdquo spec-imens having very little (eg M 14086)
We therefore see no morphological grounds to assign KNM-SO1112 to P meswae but do find some morphological evidence toassign it as a male P africanus which occurs at Songhor
KNM-LG 452 This very complete mandible was originally recog-nized as a female of P major based on the size of the dentition androbusticity of the mandible (Martin 1981) Bosler (1981) howeverleft this specimen unassigned noting similarities to P major in P3-M1 size but having distinctly smaller M2e3 In this she likenedKNM-LG 452 to a few other specimens (notably M 14086) thatare smaller in their distal molars in a manner more consistentwith taxonomic rather than sex differences (Bosler 1981 see alsoPilbeam 1969) Kelley (1986 1995) also assigned this specimen asa female of P major based on canine size and morphology butnoted that the canine lacked the shallowly-sloping mesial ridgetypical of most female anthropoid canines This is also truehowever of two other canines assigned as female P major KO 9and SO 373 (Kelley 1986) and so might simply be an unusualcharacteristic of P major female lower canines In the end Kelley(1995) concluded that considering all of the criteria fordistinguishing male and female canines the evidence that KNM-LG 452 is female remains questionable
Pickford et al (2009) further developed the idea that two spe-cies are represented within the P major sample and made KNM-LG452 the holotype of a new species ldquoUrdquo legetetensis We concur thatthis specimen is unique among the well-preserved Tinderet
mandibles in some important features Whereas male and femalemandibles of P major have long internal symphyseal planesexpansive extra- and retromolar sulci and rami that cross theocclusal plane at distal M3 KNM-LG 452 has a shorter symphysealplane (though not to the extent seen in Ekembo) a moderateextramolar sulcus but no retromolar sulcus and a ramus thatcrosses the occlusal plane more mesially If KNM-SO 396 and SO404 represent males and females respectively of P major then themorphology of KNM-LG 452 appears distinct
At present however we do not think there is sufficient evidenceto differentiate this specimen from either P major or P meswaePickford et al (2009) argue that KNM-LG 452 cannot belong to thelatter species because of its larger size The size differences how-ever are well within the range of variation seen in P majorE heseloni and E nyanzae Moreover whereas the Legetet mandibleis larger than most of the P meswae hypodigm an adult P4 referredto this species (KNM-ME 25) substantially exceeds its counterpartin KNM-LG 452 Should the Legetetmandible ultimately prove to bea male specimen size alone would not rule out its attribution toP meswae On the other hand if it is female then it becomes harderto justify its inclusion in that sample of mostly smaller specimens Iffemale therefore it could represent a separate species for whichthe binomial Proconsul legetetensis comb nov is available Recentlycollected fossils from the SonghorKapurtay area provide someadditional support for the possibility that three species of Proconsulexisted at these localities (McNulty 2014ab) However toconvincingly demonstrate this and determine whether a thirdspecies can be differentiated from P meswae will require a moredetailed analysis of variation among Proconsul species a corre-sponding study of variation in modern taxa anddalmost certain-lydadditional fossil specimens For now we regard ldquoUrdquo legetetensisas a subjective junior synonym of P major
Conclusions
The genus Proconsul has been central to studies of ape and hu-man evolution since its first description by Hopwood (1933) As oneof the best known fossil catarrhines represented by nearly everyskeletal element our knowledge of Proconsuls functional anatomypositional behavior and overall morphology has made it thefoundation for paleoanthropological research in the early Mioceneas well as studies on the origin and early diversification of homi-noids With respect to systematics however Proconsul is less wellunderstood the type species P africanus is known from a rela-tively small number of specimens from sites in the Tinderet areaand no postcranial bones can be unambiguously assigned to itRather it is the copious and well-preserved specimens from theKisingiri sites of Rusinga and Mfangano Islands that have formedthe basis for the common image of Proconsul
We argue here that the Kisingiri species traditionally referredto ProconsuldP heseloni and P nyanzaedare generically distinctfrom P africanus and hereby transfer them into a new genusEkembo based on a substantial number of morphological differ-ences (Table 2) Considering only those traits preserved in theP africanus holotype the entire sample from Rusinga and Mfan-gano differs markedly in the morphology of the upper canine P3P4 and M2 Additional differences in the I1 lower incisors andcanines molar cusp and occlusal crest morphology degree of flarein all molars and especially mandibular features clearly distin-guish the Kisingiri from non-Kisingiri samples in a way that isincompatible with congeneric species With the transfer ofE heseloni and E nyanzae Proconsul becomes a well-defined cladecomprising at least three speciesdP africanus P major andP meswaedunited by several features that are unique for thegenus (Table 2)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6160
We found no support in this study for the genus ldquoUgandapi-thecusrdquo as constructed by Pickford et al (2009) In that paper theytransferred the vast majority of Proconsul fossils from the Tinderetand Napak localities into ldquoUgandapithecusrdquo leaving P africanusrepresented only by its holotype and a small number of isolatedteeth and aligning it with the material from Rusinga and Mfan-ganoWhen the P africanus hypodigm is reconstituted and properlyaffiliated with the other Tinderet species according to itsmorphology much of the evidence marshaled by Pickford et al(2009) for ldquoUgandapithecusrdquo actually supports the distinction be-tween the Tinderet and Kisingiri samples ie the generic distinc-tion of Ekembo In addition some of the traits enumerated byPickford and colleagues in support of ldquoUgandapithecusrdquo wererevealed in our analysis to be variable within one or more speciesand therefore of questionable taxonomic utility We do not rejectout of hand the original conception of ldquoUgandapithecusrdquo (Senutet al 2000) as either a monotypic genus distinguishing ldquoUrdquo ma-jor or perhaps including a closely allied middle Miocene ldquoUrdquogitongai as well (Pickford and Kunimatsu 2005) but we have fornow retained the early Miocene specimens of both in Proconsulpending further study However even if Ugandapithecus is a taxo-nomically valid genus comprising just the latter two species theirmorphology indicates that theywould still be united in a cladewithProconsul and they lack several more advanced features that partlydefine Ekembo Better understanding of the role that body size playsinmediating differences within Proconsulwill help to further clarifythe relationships among these species
Despite the existence of one well-represented and several lesscomplete skeletons of what was long assumed to be Proconsul thephylogenetic status of this genus has defied consensus This islargely due in our view to Proconsul having been an admixture oftwo distinct genera one more advanced in the direction of extantapes than the other As we demonstrate here the material fromRusinga and Mfangano traditionally referred to Proconsul andwhich includes all of the more complete skeletal material isdistinct from the Tinderet and Napak material and belongs in aseparate genus Ekembo By disentangling these distinct clades weanticipate being able to achieve better phylogenetic resolution forboth genera thereby improving our knowledge of early hominoidevolution and diversification
Acknowledgments
We thank many people and agencies that assisted with thedevelopment and completion of this project This research wasinitiated as a result of funding from the Wenner-Gren Foundationand the Johns Hopkins School of Medicine to DR Begun in1987e1989 and was renewed following research and travelgenerously funded by multiple Leakey Foundation grants to KMcNulty National Science Foundation grants to K McNulty(0852609 1241807) the McKnight Foundation the University ofMinnesota the Lloyd A Wilford Endowment the LeverhulmeFoundation and the Turkana Basin Institute J Kelley gratefullyacknowledges support from the Institute of Human Origins Ari-zona State University DR Begun also wishes to acknowledge theAlexander Humboldt Stiftung and NSERC for funding support Weare grateful to the National Museums of Kenya the UgandaMuseum the Natural History Museum and to Rose Mwanja EzraMusiime Tom Mukhuyu Samuel Muteti James Yatich and MaryMuungu for access to and assistance with specimens in their careMany people provided assistance comments criticisms insightsand suggestions on various aspects of this work and we especiallythank Holly Dunsworth Will Harcourt-Smith Tom Lehmann DanPeppe Nicole Garrett Kirsten Jenkins Rutger Jansma Susy CoteIsaiah Nengo Andrew Hill and Brenda Benefit K McNulty
acknowledges the evolutionary anthropology faculty and studentsat Durham University for their helpful comments on an earlyversion of this manuscript and for their gracious hospitality Weare likewise grateful to the anonymous reviewers associate editorand editor whose comments strengthened the paper This manu-script is publication 1 supporting Research on East AfricanCatarrhine and Hominoid Evolution (REACHE)
Appendix A Supplementary data
Supplementary data related to this article can be found at httpdxdoiorg101016jjhevol201503009
References
Andrews P 1974 New species of Dryopithecus from Kenya Nature 249 188e190Andrews P 1978 A revision of the Miocene Hominoidea of East Africa Bull Brit
Mus (Nat Hist) Geol 30 85e225Andrews P 1985 Family group systematics and evolution among catarrhine pri-
mates In Delson E (Ed) Ancestors The Hard Evidence Liss New Yorkpp 14e22
Andrews P Harrison T Martin L Pickford M 1981 Hominoid primates from anew Miocene locality named Meswa Bridge in Kenya J Hum Evol 10123e128
Andrews P Begun D Zylstra M 1997 Interrelationships between functionalmorphology and paleoenvironments in Miocene hominoids In Begun DWard C Rose M (Eds) Function Phylogeny and Fossils Miocene HominoidEvolution and Adaptations Plenum Press New York pp 29e58
Andrews P Molleson T 1979 The provenance of Sivapithecus africanus Bull BritMus (Nat Hist) Geol 32 19e23
Begun DR 2001 African and Eurasian Miocene hominoids and the origins of theHominidae In de Bonis L Koufos GD Andrews P (Eds) Hominoid Evolutionand Climatic Change in Europe Phylogeny of the Neogene Hominoid Primatesof Eurasia Cambridge University Press London pp 231e253
Begun DR 2007 Fossil record of Miocene hominoids In Henke W Tattersall I(Eds) Handbook of Paleoanthropology Springer Berlin pp 921e977
Begun DR Teaford MF Walker AC 1994 Comparative and functional anatomyof Proconsul phalanges from the Kaswanga primate site Rusinga Island KenyaJ Hum Evol 26 89e165
Bishop WW 1964 More fossil primates and other Miocene mammals from North-East Uganda Nature 203 1327e1331
Bishop WW Miller JA Fitch FJ 1969 New potassium-argon age determinationsrelevant to the Miocene fossil mammal sequence in East Africa Am J Sci 267669e699
Blankenship JA Mincer HH Anderson KM Woods MA Burton EL 2007Third molar development in the estimation of chronologic age in Americanblacks as compared with whites J Forensic Sci 52 428e433
Bosler W 1981 Species groupings of early Miocene dryopithecine teeth from EastAfrica J Hum Evol 10 151e158
Coleman MN 2008 What does geometric mean mean geometrically Assessingthe utility of geometric mean and other size variables in studies of skullallometry Am J Phys Anthropol 135 404e415
Cote S Malit N Nengo I 2013 Additional mandibles of Rangwapithecus gordonian early Miocene catarrhine from the Tinderet localities of Western KenyaJ Hum Evol 153 341e352
Deane AS 2009 Early Miocene catarrhine dietary behavior the influence of theRed Queen Effect on incisor shape and curvature J Hum Evol 56 275e285
Drake R Van Couvering JA Pickford M Curtis G Harris JA 1988 New chro-nology for the Early Miocene mammalian faunas of Kisingiri Western KenyaJ Geol Soc Lond 145 479e491
Evernden JF Savage DE Curtis GH James GT 1964 Potassium-argon datesand the Cenozoic mammalian chronology of North America Am J Sci 262145e198
Gebo DL MacLatchy L Kityo R Deino A Kingston J Pilbeam D 1997A hominoid genus from the Early Miocene of Uganda Science 276 401e404
Geoffroy Saint-Hilaire E 1812 Tableau des quadrumanes 1 Ord QuadrumanesAnnls Mus Hist Nat Paris 19 85e122
Gray JE 1825 Outline of an attempt at the disposition of the Mammalia into tribesand families with a list of the genera apparently appertaining to each tribeAnnls Philos 10 337e344
Harrison T 1982 Small-bodied apes from the Miocene of East Africa PhDDissertation University of London
Harrison T 1986 New fossil anthropoids from the middle Miocene of East Africaand their bearing on the origin of the Oreopithecidae Am J Phys Anthropol 71265e284
Harrison T 1987 The phylogenetic relationships of the early catarrhine primates areview of the current evidence J Hum Evol 16 41e80
Harrison T 2002 Late Oligocene to middle Miocene catarrhines from Afro-ArabiaIn Hartwig WC (Ed) The Primate Fossil Record Cambridge University PressCambridge pp 311e338
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 61
Harrison T 2010 Dendropithecoidea Proconsuloidea and Hominoidea (CatarrhiniPrimates) In Werdelin L Sanders WJ (Eds) Cenozoic Mammals of AfricaUniversity of California Press Berkeley pp 429e469
Harrison T Andrews P 2009 The anatomy and systematic position of a newspecies of early Miocene proconsulid from Meswa Bridge Kenya J Hum Evol56 479e496
Hill A Nengo IO Rossie JB 2013 A Rangwapithecus gordoni mandible from theearly Miocene site of Songhor Kenya J Hum Evol 65 490e500
Hopwood AT 1933 Miocene primates from Kenya J Linn Soc Zool 38 437e464Ishida H Kunimatsu Y Takano T Nakano Y Nakatsukasa M 2004 Nacholapi-
thecus skeleton from the Middle Miocene of Kenya J Hum Evol 46 69e103Jolicouer P 1963 The multivariate generalization of the allometry equation Bio-
metrics 19 497e499Jungers WL Falsetti AB Wall CE 1995 Shape relative size and size-
adjustments in morphometrics Am J Phys Anthropol 38 (S2) 137e161Kavanagh KD Evans AR Jernvall J 2007 Predicting evolutionary patterns of
mammalian teeth from development Nature 449 427e433Kelley J 1986 Species recognition and sexual dimorphism in Proconsul and
Rangwapithecus J Hum Evol 15 461e495Kelley J 1995 Sex determination in Miocene catarrhine primates Am J Phys
Anthropol 96 390e417Kelley J 1993 Taxonomic implications of sexual dimorphism in Lufengpithecus In
Kimbel WH Martin LB (Eds) Species Species Concepts and Primate Evo-lution Springer New York pp 429e458
Kent PE 1942 The country round the Kavirondo Gulf of Victoria Nyanza Geogr J100 22e31
Kordos L Begun DR 2001 A new cranium of Dryopithecus from RudabanyaHungary J Hum Evol 15 689e700
Le Gros Clark WE Leakey LSB 1950 Diagnoses of East African Miocene Homi-noidea Q J Geol Soc Lond 105 260e263
Le Gros Clark WE Leakey LSB 1951 The Miocene Hominoidea of East Africa InFossil Mammals of Africa No 1 British Museum (Natural History) London
Leakey LSB 1943 A Miocene anthropoid mandible from Rusinga Kenya Nature152 319e320
Leakey LSB 1963 East African fossil Hominoidea and the classification within thissuper-family In Washburn SL (Ed) Classification and Human EvolutionAldine Chicago pp 32e49
Leakey LSB 1967 Notes on the mammalian faunas from the Miocene and Pleis-tocene of East Africa In Bishop WW Clark JD (Eds) Background to Evolu-tion in Africa University of Chicago Press Chicago pp 7e29
Leakey RE Leakey MG 1986 A new Miocene hominoid from Kenya Nature 324143e146
Leakey RE Leakey MG Walker AC 1988 Morphology of Afropithecus turka-nensis from Kenya Am J Phys Anthropol 76 289e307
Leakey MG Ungar PS Walker A 1995 A new genus of large primate from theLate Oligocene of Lothidok Turkana District Kenya J Hum Evol 28519e531
Linnaeus C 1758 Systema naturae per regna tria naturae secundum classesordines genera species cum characteribus differentris synonymis locis tenthed Laurentii Salvii Stockholm
MacInnes DG 1943 Notes on the East African Miocene primates J E Afr UgandaNat Hist Soc 17 141e181
MacLatchy L 2004 The oldest ape Evol Anthropol 13 90e103MacLatchy L Bossert WH 1996 An analysis of the articular surface distribution of
the femoral head and acetabulum in anthropoids with implications for hipfunction in Miocene hominoids J Hum Evol 31 425e453
MacLatchy L Rossie JB 2005 The Napak hominoid still Proconsul major InLieberman DE Smith RJ Kelley J (Eds) Interpreting the Past Essays onHuman Primate and Mammal Evolution in Honor of David Pilbeam Brill Ac-ademic Publishers Boston pp 15e28
Martin L 1981 New specimens of Proconsul from Koru Kenya J Hum Evol 10139e150
McCollum MS Peppe DJ McNulty KP Dunsworth HM Harcourt-Smith WEH Andrews AL 2013 Magnetostratigraphy of the early MioceneHiwegi Formation (Rusinga Island Lake Victoria Kenya) Geological Society ofAmerica Abstracts with Programs 45 p 12
McNulty KP 2010 Apes and tricksters the evolution and diversification ofhumans closest relatives Evol Edu Outreach 3 322e332
McNulty KP Harcourt-Smith WEH Dunsworth HM 2007 New primate fossilsfrom Rusinga Island Kenya Am J Phys Anthropol 132 (S44) 170
McNulty KP MacLatchy L Peppe DJ Nengo I Manthi FK Miller ERStevens NJ Cote S Lehmann T Kingston J 2014a A regional approach toEast African early Miocene paleobiology J Vert Paleontol 185 SVP Programand Abstracts Book 2014
McNulty KP MacLatchy L Rossie JB Peppe DJ Deino AL Mbua ENManthi FK Nengo IO Stevens NJ Cote S Lehmann T Gutierrez M 2014b
Research on East African catarrhine and hominoid evolution results from thefirst year Am J Phys Anthropol 153 (S58) 182
Michel LA Peppe DJ Lutz JA Driese SG Dunsworth HM Harcourt-Smith WEH Horner W Lehmann T Nightengale S McNulty KP 2014Remnants of an ancient forest provide ecological context for Early Miocenefossil apes Nature Comm 5 3236
Myres JL 1932 Africa Man 32 208Nakatsukasa M Kunimatsu Y Nakano Y Takano T Ishida H 2007 Vertebral
morphology of Nacholapithecus kerioi based on KNM-BG 35250 J Hum Evol52 347e369
Nengo IO Rae TC 1992 New hominoid fossils from the early Miocene site ofSonghor Kenya J Hum Evol 23 423e429
Peppe DJ McNulty KP Cote SM Harcourt-Smith WEH Dunsworth HM VanCouvering JA 2009 Stratigraphic interpretation of the Kulu Formation (EarlyMiocene Rusinga Island Kenya) and its implications for primate evolutionJ Hum Evol 56 447e461
Pickford M 1981 Preliminary Miocene mammalian biostratigraphy for westernKenya J Hum Evol 10 73e97
Pickford M 1986 Sexual dimorphism in Proconsul Hum Evol 1 111e148Pickford M Andrews PJ 1981 The Tinderet Miocene sequence in Kenya J Hum
Evol 10 11e33Pickford M Kunimatsu Y 2005 Catarrhines from the Middle Miocene (ca 145
Ma) of Kipsaraman Tugen Hills Kenya Anthropol Sci 113 189e224Pickford M Senut B Gommery D Musiime E 2009 Distinctiveness of Ugan-
dapithecus from Proconsul Estudios Geol 65 183e241Pilbeam DR 1969 Tertiary Pongidae of East Africa evolutionary relationships and
taxonomy Bulletin of the Peabody Museum of Natural History 31 pp 1e185Rae TC 1993 Phylogenetic analysis of proconsulid facial morphology PhD
Dissertation State University of New York at Stony BrookRae TC 1999 Mosaic evolution in the origin of the Hominoidea Folia Primatol 70
125e135Rafferty KL Walker A Ruff C Rose MD Andrews PJ 1995 Postcranial esti-
mates of body weights in Proconsul with a note on a distal tibia of P major fromNapak Uganda Am J Phys Anthropol 97 391e402
Rossie JB MacLatchy L 2013 Dentognathic remains of an Afropithecus individualfrom Kalodirr Kenya J Hum Evol 65 199e208
Senut B Pickford M Gommery D Kunimatsu Y 2000 Un nouveau genredrsquohominoiumlde du Miocene inferieur drsquoAfrique orientale Ugandapithecus major(Le Gros Clark and Leakey 1950) CR Acad Sci Paris 331 227e233
Simons EL Pilbeam DR 1965 Preliminary revision of the Dryopithecinae (Pon-gidae Anthropoidea) Folia Primatol 3 81e152
Smith TM Martin LB Leakey MG 2003 Enamel thickness microstructure anddevelopment in Afropithecus turkanensis J Hum Evol 44 283e306
Stevens NJ Seiffert ER OConnor PM Roberts EM Schmitz MD Krause CGorscak E Ngasala S Hieronymus TL Temu J 2013 Palaeontological evi-dence for an Oligocene divergence between Old World monkeys and apesNature 497 611e614
Suwa G Kono RT Katoh S Asfaw B Beyene Y 2007 A new species of great apefrom the late Miocene epoch in Ethiopia Nature 448 921e924
Szalay FS Delson E 1979 Evolutionary History of the Primates Academic PressNew York
Teaford MF Beard KC Leakey RE Walker A 1988 New hominoid facial skel-eton from the Early Miocene of Rusinga Island Kenya and its bearing on therelationship between Proconsul nyanzae and Proconsul africanus J Hum Evol17 461e477
Van Couvering JAH Miller J 1969 Miocene stratigraphy and age determinationsRusinga Island Kenya Nature 221 628e632
Walker A 1997 Proconsul function and phylogeny In Begun DR Ward CVRose MD (Eds) Function Phylogeny and Fossils Miocene Hominoid Evolu-tion and Adaptation Plenum Press New York pp 209e224
Walker A Pickford M 1983 New postcranial fossils of Proconsul africanus andProconsul nyanzae In Ciochon RL Corruccini RS (Eds) New Interpretationsof Ape and Human Ancestry Plenum Press New York pp 325e351
Walker A Teaford M 1988 The Kaswanga primate site an Early Miocene hom-inoid site on Rusinga Island Kenya J Hum Evol 17 539e544
Walker A Teaford M 1989 The hunt for Proconsul Sci Am 260 76e82Walker A Teaford MF Martin L Andrews P 1993 A new species of Proconsul
from the early Miocene of RusingaMfangano Islands Kenya J Hum Evol 2543e56
Ward CV Ruff CB Walker A Rose MD Teaford MF Nengo IO 1995 Func-tional morphology of Proconsul patellas from Rusinga Island Kenya with im-plications for other Miocene-Pliocene catarrhines J Hum Evol 29 1e19
Ward S Brown B Hill A Kelley J Downs W 1999 Equatorius a new hominoidgenus from the Middle Miocene of Kenya Science 285 1382e1386
Werdelin L Sanders WJ 2010 Cenozoic Mammals of Africa University of Cali-fornia Press Berkeley

KP McNulty et al Journal of Human Evolution 84 (2015) 42e6146
with a small ovoid head and flat distal articular surface that mainlycontacts the scaphoid Ulna displays a large olecranon processnarrow semilunar notch with only a slight keel anteriorly posi-tioned radial notch and a long distal styloid process that retainsarticulation with triquetrum and pisiform Carpals relatively nar-row and include an unfused os centrale a hamate with a smallhamulus and a flat medially oriented triquetral surface Handcharacterized by straight metacarpals with narrow bases andheads Pollex well developed Manual and pedal phalanges gener-ally similar to each other but less so than in Old World monkeysFemur slender with a small head long neck with a high neck-shaftangle centrally placed fovea capitis reduced greater trochanterand a broad distal end with the medial condyle slightly larger thanthe lateral Broad flat patella Tibia long and slender but fibularobust Tarsals and metatarsals elongate with a robust hallux
Differential diagnosis
Ekembo can be readily distinguished from all other medium-and large-bodied catarrhine genera from the early Miocene Cra-nial mandibular and dental differences between Ekembo andProconsul are described in detail below and summarized inTable 2 Ekembo also differs from Proconsul in having a rounderradial head and a femur with a shorter greater trochanter poste-riorly projecting lesser trochanter less extensive inter-trochantericcrest more robust phalanges with more strongly developedfibrous flexor sheath ridges and broader proximal and interme-diate phalangeal distal joint surfaces (condyles) with widertrochlear grooves and less sharply defined trochlear ridges(Harrison 1982 Andrews et al 1997 Senut et al 2000 Pickfordet al 2009)
Ekembo differs from Afropithecus in having a more inflatedglabellar region no frontal trigon less strongly developed anteriortemporal ridges less postorbital constriction a stronger mandib-ular superior transverse torus less posterior shallowing of themandibular corpus higher-crowned lower canines in relation tomesiodistal length buccolingually narrower upper premolarsrelative to molars upper premolars with cusps positioned closer tothe mesiodistal midline reduced P3 cusp heteromorphy a moresymmetric I2 and substantially less buccolingual flare in the cheekteeth (Leakey et al 1988 Rossie and MacLatchy 2013)
Ekembo differs from Heliopithecus in having buccolinguallynarrower premolars relative to molars premolar cusps positionedcloser to the mesiodistal midline and reduced P3 cusp hetero-morphy (Harrison 2010)
Ekembo differs from Equatorius in having a more gracilemandible a larger superior but smaller inferior mandibular trans-verse torus labiolingually narrower lower incisors with straightermesial and distal margins higher-crowned lower canines in rela-tion to mesiodistal length greater molar cingulum development aless posteriorly directed humeral medial epicondyle a more ante-riorly directed radioulnar joint and narrower metacarpal heads(Ward et al 1999)
Ekembo differs from Nacholapithecus in having a shorter sub-nasal clivus a more expansive incisive foramen a broader nasalaperture at the base a minimally developed mandibular inferiortransverse torus a narrower lingual tubercle on I1 higher-crownedlower canines in relation to mesiodistal length a larger humeralradial fossa a shallower coronoid fossa a less well-developed zonaconoidea a narrower and distally slanted ulnar coronoid process alonger and lower femoral neck a more posteriorly positioned lessertrochanter and a smaller posteromedial tubercle on the talus(Ishida et al 2004)
Ekembo differs from Nyanzapithecus in having more anteriorlyextended maxillary sinuses a relatively taller mandibular corpus
less buccolingually compressed upper and lower canines with lesscurved crowns lower and more rounded cusps with more inflatedocclusal crests on the cheek teeth P3 and P4 that more closelyresemble each other and have much less cingular development abroader P4 relative to M1 M1e2 occlusal outline more closelyapproximating a square and lower molars that are broader relativeto mesiodistal length with a more mesially placed entoconid rela-tive to the hypoconid (Harrison 1986 2010)
Ekembo differs from Rangwapithecus in having a broad andexpansive palatal fenestration a wider palate relative to length amore robust mandible with a well-defined superior transversetorus broader upper and lower incisors relative to height lessbuccolingually compressed canines without scimitar-shapedcrowns upper premolars with reduced cingulum and cusps posi-tioned near the mesiodistal midpoint resulting in expanded mesialfoveae and symmetric buccal profiles a broader P4 relative to M1upper and lower molars that are buccolingually broader relative tolength and have more rounded cusps and inflated occlusal crestsmore squared upper molars with substantially less cingulumdevelopment and lower molars with a more mesially placedentoconid (Andrews 1974 Harrison 2002 2010 Cote et al 2013Hill et al 2013)
Ekembo differs from Turkanapithecus in having an expandedglabellar region less postorbital constriction a narrower temporalfossa with less flaring zygomatic arches the lacrimal fossa withinthe orbital margin a relatively broader palate with non-convergingtooth rows narrower nasal bones and interorbital region a distinctmandibular superior transverse torus a more robust mandibularcorpus taller mandibular ramus upper canines more rounded incross-section a reduced cingulum on cheek teeth awell-developedhypocone on M1e2 a more squared M2 with a protocone subequalto paracone and broader lower molars relative to length (Leakeyand Leakey 1986 Harrison 2002 2010)
Ekembo differs from Mabokopithecus in having a relativelybroader M3 crown without distal recurvature of the crown andlacking a buccal concavity a hypoconulid that is in-line with thebuccal cusps more bunodont cusps a well-defined protocristidand a broad open talonid without an accessory cuspid (Harrison2002 2010)
Ekembo differs from Xenopithecus in having reduced uppermolar flare a substantially reduced cingulum no prehypoconecrista molar occlusal profiles more closely approximating a squareand less constricted trigons (Hopwood 1933 Le Gros Clark andLeakey 1951 Harrison 2002 2010)
Ekembo differs from Morotopithecus in having a broader inter-orbital region a better developed canine fossa in males premolarssmaller relative to molars greatly reduced or absent buccalcingula on upper molars lumbar vertebrae with transverse pro-cesses positioned more ventrally on the centrum and a narrowerdistal femur with a deeper patellar groove and similarly-sizedmedial and lateral femoral condyles (Gebo et al 1997MacLatchy 2004)
Recently Stevens et al (2013) likened the new genus Rukwapi-thecus represented by a mandible fragment with well-preservedP4-M3 to nyanzapithecine species Ekembo can be distinguishedfrom this genus in having a more gradually rounded gonial angle aless obliquely oriented P4 with subequal protoconid and metaconidand a broader mesial fovea less mesially positioned metaconidreduced buccal cingula and smaller mesial foveae on the molarssmaller hypoconulids broader M1e2 relative to length a singularrather than twinned metaconid no accessory cuspules in thelingual notch and a well-defined crest between entoconid andhypoconulid
Family Proconsulidae (Leakey 1963)
TabDen
Fe
G
U
U
U
U
U
M
Lo
Lo
Lo
a
b
c
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 47
Genus Proconsul Hopwood 1933
Type species
Proconsul africanus Hopwood 1933
Included species
Proconsul africanus (Hopwood 1933) Proconsul major (Le GrosClark and Leakey 1950) Proconsul gitongai comb nov (Pickfordand Kunimatsu 2005) Proconsul meswae (Harrison and Andrews2009)
Diagnosis (replaces the emended diagnosis of Harrison 2010)
Proconsul species are medium- to large-bodied sexually dimor-phic catarrhines that can be distinguished from other proconsulidgenera by the following combination of features Robust mandib-ular corpus and symphysis characterized by a long shallowlysloping planum alveolare terminating at a large superior transversetorus and a subplanum surface oriented posteroinferiorly Upper
le 2tognathic features that distinguish Ekembo from Proconsula
ature Ekembo
eneralCusp morphology More bunodont occupying more of theMolar crests Inflated incorporated into cuspsCingulum Reducedpper central incisorLingual pillar Narrow generally low reliefMarginal ridges and cingulum Asymmetric with sharply angled cingulu
ridge transitionpper canineTip PointRoot fluting AbsentRoot striations Rare and weakly developedpper third premolarb
Mesiodistal cusp position Crown midpointMesial fovea Well developedBuccal profile SymmetricMesial crown margin Straight or slightly curvedDistal crown margin Slight to moderately curvedCusp heteromorphy ReducedCrown shape Converges on P4 shapeMesiobuccal flange Nopper fourth premolarParacone height Taller relative to crown lengthpper second molarc
Occlusal crown shape SquarishDistal margin Straight to slightly curvedBuccal cingulum Absent or rudimentaryDistolingual cingulum Merges into hypoconePosthypocone crista Distobuccally to buccally orientedBuccolingual flare ReducedandiblePlanum alveolare Short vertically inclinedSuperior transverse torus Moderate to well-developedOrientation of subplanum area More vertically orientedCorpus robusticity More gracilewer incisorsCrown shape Narrow relative to heightwer canineTip Pointwer molarsBuccolingual flare ReducedM1 M2 breadth M1 narrower relative to M2
Although presented here as discrete characteristics for descriptive purposes some oNearly all of these features tend to distinguish P4s as well but in that tooth the diffeThese features tend to distinguish M1s as well but in that tooth the differences are
incisors heteromorphic with I1 exhibiting an inflated lingual tu-bercle and a near symmetrical configuration of lingual marginalridges and cingulum with a curving transition from cingulum tothe mesial marginal ridge Lower incisors broad relative to crownheight Upper and lower canines with distinct blade-like or burin-like tips in both deciduous and adult teeth Upper canine rootswith broad longitudinal grooves or fluting defining two or threedistinct columns plus numerous small longitudinal striationsextending cervically from the root apex Upper P3s morphologicallydissimilar from P4s P3 has substantial cusp heteromorphy (para-cone much taller than protocone) with mesially positioned cuspsand a notched mesial border that together limit or eliminate amesial fovea A long steeply angled preparacrista contributes to ahighly asymmetric buccal profile before being interrupted near thetooth cervix by a mesially projecting flange The distal margin isstrongly curved P4 has a tall paracone relative to crown lengthMolars characterized by extensive cingular development individ-uated conical cusps connected by sharp occlusal crests and sub-stantial buccolingual flare Upper molars rhomboid in shaperesulting from a broad trigon and a distally expanded hypocone M2
has a distinct buccal shelf between paracone and metacone a dis-tolingual cingulum that wraps fully around the hypocone rather
Proconsul
occlusal surface area More distinct individuated conesSharper more distinct from cuspsExtensive
Inflatedbulbousm-mesial More symmetric with curving cingulum-mesial
ridge transition
Blade-likePresentVery common and well developed
MesialAbsent or reducedAsymmetricNotchedStrongly curvedStrongDistinct from P4
Yes
Shorter relative to crown length
RhomboidStrongly curvedDistinct cingular shelfDistinct from hypoconeDistally to distobuccally orientedPronounced
Long more horizontally orientedStrongly developedMore posteriorly inclinedMore robust
Broad relative to height
Blade-like
PronouncedM1 more similar to M2
f these features are undoubtedly correlated aspects of single evolutionary changesrences are not as pronouncednot as pronounced
Figure 2 Upper central incisors of Proconsul and Ekembo Lingual views of Ekembo(KNM-RU 1685 cast) left and Proconsul (KNM-CA 1300) right Note the differences inthe transition between the lingual cingulum and the mesial marginal ridge which ismore rounded in Proconsul and more angled in Ekembo The central pillar is also morebulbous or inflated and without ridging in Proconsul Additional specimens illustratingthe Proconsul morphology are shown in Pickford et al (2009 Fig 17 p 197) butreferred in that manuscript to Ugandapithecus
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6148
thanmerging into the hypocone on its lingual surface and a distallyoriented posthypocone crista Lower first and second molars nearlyequal in buccolingual breadth
Comparative morphology of Ekembo and Proconsul
Samples and comparative methods
The above diagnoses are based on extensive survey and analysisof the original fossil material Specimens housed in the NationalMuseums of Kenya (NMK) and the Natural History Museum (Lon-don) were analyzed by the authors during several separate and twojoint (NMK) research visits Fossils in the Uganda National Museumwere assessed by one of us (KPM) during two visits All measure-ments were collected by KPM Dental terminology follows Szalayand Delson (1979)
The following results are organized with respect to themorphology of the Kisingiri sample (attributed to E nyanzae andE heseloni) versus that of Proconsul from Tinderet and Napak(traditionally referred to P africanus P major and more recentlyP meswae) Specimens from additional localities that have at onetime been attributed to Proconsul (eg Andrews1978 Pickford andKunimatsu 2005 [cf Harrison 2010]) are for the most part notconsidered here most are middle Miocene in age and have beentransferred to other genera (Harrison 2002 2010) Of those that arefrom early Miocene sites there is growing evidence that P majorcan also be found at Moroto and differentiated fromMorotopithecus(Pickford et al 2009) an issue of some interest but ultimately notcritical to the taxonomic questions being considered here Whereappropriate however we do provide observations on a few earlyMiocene specimens attributed to ldquoUrdquo gitongai (Pickford andKunimatsu 2005 Pickford et al 2009 see Harrison 2010) Yetwhile we consider the middle Miocene holotype likely to representa distinct species here referred to Proconsul we concur withHarrison (2010) that many specimens placed in this species byPickford et al (2009) cannot be distinguished from P major
Sample sizes particularly from Tinderet and Ugandan localitiestend to be small but where possible standard univariate (eg t-tests) and bivariate (regression ANCOVA) statistical analyses wereperformed A single eigendecomposition of the covariance matrixof symphyseal measurements was done to characterize the multi-variate shape of the mandibular symphysis after accounting for sizedifferences (sensu Jolicouer 1963) Quantitative results are illus-trated as box-and-whisker plots of shape indices or bivariate plotsof the measurements For samples of fewer than five specimensshape index values are plotted in lieu of box-and-whiskersummaries
Morphological comparisons
Upper incisors Two features of the Ekembo upper central incisorsdistinguish them from those of Proconsul including specimenswidely attributed to the type species P africanus The lingual tu-bercle in Ekembo is quite narrow generallywith low relief and oftenscored by two or more irregular ridges in contrast to the muchmore inflated or bulbous tubercle in Proconsul I1s (Fig 2) Ekembospecimens also have a characteristic morphology in the transitionfrom the lingual cingulum to the mesial marginal ridge (Fig 2)from the mesiodistal mid-point of the crown the cingulumextends either directly mesially or even angles slightly cervicallyto intersect the mesial marginal ridge at a fairly sharp angle Incontrast in I1s of Proconsul the cingulum angles incisally from themid-point and transitions into the mesial marginal ridge in agradual curve The expression of these traits varies somewhat inthe smaller Tinderet specimens attributed to P africanus
compared with those of P major (cf Pickford et al 2009)However these variations are relatively minor with respect to thedifferences between the two genera Of 25 I1s known fromKisingiri localities only one specimen KNM-MW 562 somewhatresembles the Tinderet sample in both of these featuresdhavinga somewhat less angled transition from the mesial marginal ridgeto the cingulum and a more bulbous lingual tubercle This isparticularly interesting as the specimen was discovered in theMakira Beds now dated to older than 190 Ma and contemporarywith the Tinderet localities (McCollum et al 2013) Neverthelessits overall morphology is more similar to other Ekembospecimens than to Proconsul incisors Two other Ekembospecimens KNM-RU 1677 and RU 1933 have somewhat moreinflated lingual tubercles but otherwise resemble other Ekembo I1s
No consistent differences were found in upper lateral incisors
Upper canines Upper canine crowns of Ekembo are typical of thoseof most catarrhines in terminating in an evenly tapering point Thisdiffers from the Proconsul specimens which have a blade-like(Kelley 1986 Senut et al 2000 Harrison 2010) or ldquoburin-likerdquo(Pickford et al 2009) canine tip (Fig 3) While blade-like apicalmorphology has long been recognized as a distinctivecharacteristic of P major we confirm the report by Pickford et al(2009) that this feature is also present in the P africanus typespecimen (Fig 3a) although somewhat understated as expectedin the more slender and higher-crowned P africanus caninesUnfortunately other canines attributed to P africanus (eg M44837 KNM SO 419 LG 921 CA 1910 SO 5353 Kelley 1986) havebroken apices and therefore cannot be evaluated for this featureA blade-like tip is also present in the deciduous upper canines ofP major (eg KNM-SO 371 MV 10 NAP IV UMP 62-20) andP meswae (KNM ME 11) Hence the blade-like upper canine tipunites all Tinderet and Napak Proconsul species but is neverexpressed in Ekembo specimens from Rusinga and Mfangano
Two features of the canine roots also differentiate these generalongitudinal grooving or fluting along the length of the root (cfKelley 1986) and the development of numerous finer longitudinal
Figure 3 Canines of Proconsul and Ekembo a) Upper canine of the P africanus holotype (M 14084) lingual view b) Upper canine of P major (KNM-CA 2127) lingual view c) Lowercanine of P major buccal view d) lower canine of Ekembo cf nyanzae (KNM-RU 1676) buccal view and e) Upper canine of P major (KNM-SO 584) distal view Note the blade-like orldquoburin-likerdquo tip of the unworn Proconsul canines including on the nearly unworn canine tip of the P africanus type specimen (a) Ekembo canines have more typical pointed tipsProconsul canines also have longitudinal grooves and fine striations on the root (e) Ekembo lacks these features Scale bars frac14 1 cm
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 49
striations near the root apex that extend to varying degrees towardthe cervix of the tooth Longitudinal ldquoflutingrdquo consists of broadgrooves that segment the root into distinct sections (cf Pickfordet al 2009 Fig 3e) Most often the root in cross-section has theappearance of a cylinder that has been sectioned longitudinallywith the two halves offset from each other but in some cases thereare three segments and the root cross-section is more triangular inappearance (Kelley 1986) This characteristic was noted by Kelley(1986) in the P africanus canines but not in those of P majorHere we affirm that fluting is present on all of the visible uppercanine roots attributed to both P africanus and P major rootmorphology could not be evaluated in the deciduous upper caninefrom Meswa Bridge since it is implanted in the maxillary alveolusNone of the Ekembo specimens display this morphology havinginstead a simpler round to ovoid cross-section along the length ofthe root
The fine longitudinal striations on Proconsul upper canine rootsvary in the degree to which they extend toward the cervix (Fig 3e)but they are visible in all specimens attributed to P africanus(KNM-SO 419 521 921 5353 KNM-X 502) and five of six uppercanines of P major (excepting KNM-SO 1614) This morphology isalso present in UMP 62-12 which was included as part of theMorotopithecus bishopi hypodigm (Gebo et al 1997) Fine striationsalong the upper canine root are almost entirely absent in EkemboTwenty of twenty-three specimens lack this feature three speci-mens (KNM-RU 1763 1845 1891) exhibit incipient to slight
striations but these are never developed to the extent seen inProconsul canines
Upper premolars Upper third premolars display some of thegreatest differences between Ekembo and Proconsul P4s tend todiffer in similar ways but the differences are not nearly as markedEkembo P3s have cusps positioned near the mesiodistal midpoint ofthe crown well-developed mesial foveae pre- and postparacristaeof similar length and slope resulting in symmetric or nearsymmetric buccal profiles a straight or slightly curved mesialmargin and only moderately curved distal margin (Fig 4)Proconsul P3s are more similar in appearance to those of small-bodied catarrhines from the early Miocene with cusps positionedmesially on the crown greatly reducing or eliminating the mesialfovea They also have a steep preparacrista and more shallowlysloped postparacrista resulting in a highly asymmetric buccalprofile and an inwardly angled or notched mesial margin but astrongly curved distal margin This morphology is bestcharacterized in the P africanus holotype (M 14084) but can alsobe seen in other specimens attributed to this species as well asthose referred to P major (Fig 4d e) Also noteworthy is adistinctive mesiobuccal flange that mesially extends the buccalportion of the crown but only near the cervical line of the toothPickford et al (2009) describe this feature in some detail forldquoUgandapithecusrdquo but in fact this morphology is clearly expressedin the P africanus holotype as well (Fig 4a d) A single adult P3
Figure 4 Upper premolars of Proconsul and Ekembo occlusal (aec) and lingual (def) views a) P3e4 of the P africanus holotype (M 14084) b) P3 of P major (M 14331) c) P3e4 of theE heseloni holotype (KNM-RU 2036) d) P3 of the P africanus holotype (M 14084) e) P3 of P major (M 14331) f) P3 of the E heseloni holotype (KNM-RU 2036) Note the mesiallypositioned cusps lack of mesial fovea and strongly asymmetric crown in Proconsul compared to Ekembo (see text for further comparisons) Image of M 14331 reversed for easiercomparison Photo 4a courtesy of Rutger Jansma Scale bars frac14 1 cm
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6150
germ and two dP3s attributed to P meswae look remarkably similarto those of other Proconsul species including the presence of thisflange In contrast the Ekembo P3 has a well-developed mesialfovea that terminates at a mesiobuccal shoulder positionedapproximately halfway between the paracone tip and the cervixFrom this shoulder the mesial margin angles slightly distallydown to the cervix (Fig 4f)
The degree of cusp height heteromorphy in P3 also differentiatesEkembo from Proconsul (contra Pickford et al 2009) Only four non-Kisingiri specimens were sufficiently unworn to take measure-ments but more worn specimens also appear to express this dif-ferential in cusp height Figure 5a shows the ratio of protoconeparacone height measured from the tooth cervix and with only oneexception Ekembo P3s have substantially less cusp heteromorphyThis exception KNM-RU 1677 does not in fact have greater heter-omorphy but instead has an abnormal extension of the enamel ontothe buccal (paracone) root which skews the measurement Hencecusp relief relative to the occlusal surface in RU 1677 is not nearly asheteromoprhic as in Proconsul and when this is taken into accountthe difference between genera is significant (t-test dffrac14 11 t frac14 283p frac14 00165) The adult P3 germ from Meswa Bridge is incompletelyformed and therefore could not be measured However the differ-ence between paracone and protocone heights with respect to theocclusal surface appears to be as great as that in the P africanusholotype It does not appear that sex is an important determinant ofthis feature since well-established male (eg M16647 KNM-RU16000) and female (KNM-RU 2036 7290) specimens do not segre-gate across the Kisingiri range of variation
Upper fourth premolars of Ekembo are broadly similar inmorphology to Ekembo P3s in fact the similarity between P3 and P4
is substantially greater in Ekembo than in Proconsul (Figs 4a c)Likewise most of the morphological differences that discriminatethe P3s of these genera also tend to differentiate the P4s but less
clearly Cusp heteromorphy does not distinguish P4s of thesegenera but the overall height of the P4 paracone in Ekembo issignificantly greater relative to crown length than in Proconsul P4s(t-test df frac14 14 t frac14 316 p frac14 00069 Fig 5b)
Upper molars Cusps of Ekembo upper molars are uniformly morebunodont than those of Proconsul occupying more of the occlusalsurface Intercuspal crests are more fully incorporated into thecusps in Ekembo molars giving them a more inflated appearanceThis is particularly evident in the crista obliqua which is very broadin Ekembo but thin and knife-like in Proconsul (Fig 6) Cusps ofProconsul molars stand more as individual cones Pickford et al(2009) have further differentiated the cusp morphology ofsmaller and larger Tinderet specimens arguing that the latterhave more bulbous cusps While accurate this difference is minorcompared with the morphological difference between Ekemboand Proconsul and we regard the within-Proconsul cuspalvariation as size-related In fact the intermediately sizedP meswae has cusp morphology that is appropriatelyintermediate between that of P africanus and the larger P major
Other differences between Ekembo and Proconsul upper molarsare most evident in M2 We concur with previous assessments thatthe Kisingiri upper molars have less developed cingula than non-Kisingiri molars (eg Senut et al 2000 Harrison 2002 2010Harrison and Andrews 2009) More specifically Ekembo M2s lackor have only rudimentary buccal cingula between the paracone andmetacone Likewise the lingual cingulum is discontinuous merg-ing into the hypocone lingually and then re-emerging as a distalmarginal ridge (Fig 6) Proconsul M2s display substantially morecingular development overall and in particular exhibit a prominentbuccal shelf between the paracone and metacone and a distinctdistolingual cingulum that is continuous around the hypocone towhere it merges with the distal marginal ridge
Figure 5 Box-and-whisker plots a) P3 cusp heteromorphy index (protocone heightparacone height) Results of a t-test show this difference to be significant (p frac14 00165) Theexcluded Ekembo outlier KNM-RU 1677 only appears to have extreme heteromorphy because of extensive expansion of the buccal enamel onto the anterior root (see text) b) P4
relative paracone height (paracone heightmesiodistal crown length) Results of a t-test show this difference to be significant (p frac14 00069)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 51
The occlusal profiles of Ekembo M2s are squarish in shape withrelatively straight and equal sides with a posthypocone crista thatis distobucally to buccally directed In contrast the occlusal profilesof Proconsul M2s have a more rhomboid shape with substantialbasal flare around the mesial cusps and a distally expanded hypo-cone with a distally directed posthypocone crista resulting in astrongly curved distal margin (Fig 6) Even in specimens with lesscurved distal margins the posthypocone crista is still more distallydirected compared to the condition in Ekembo These features arecharacteristic not only of P africanus P meswae and P major butare also expressed in the isolatedM2 fromMoroto Uganda (MOR IIb2098) attributed by Pickford et al (2009) to ldquoUrdquo gitongai butregarded here as belonging to P major
As noted by others (eg Harrison and Andrews 2009) the de-gree of buccolingual molar flare differs among these samplesQuantified as the distance between the tips of the mesial cuspsdivided by maximum crown breadth the degree of molar flare in
Ekembo M2s is significantly less than in Proconsul M2s (t-testdf frac14 29 t frac14 396 p frac14 00004) In a bivariate plot of these twomeasures the distributions of Ekembo and Proconsul teeth barelyoverlap with only KNM-RU 1677 encroaching into Proconsul ter-ritory (Fig 7)
These differences in M2 morphology also tend to distinguishEkembo from Proconsul M1s but not always as clearly Whereas M2smore consistently express distinct morphotypes the M1s in bothgenera show more variation in these features
The degree of upper third molar development (ie whether M3
has four distinct cusps or the distal cusps are reduced or absent) haslong been cited as taxonomically informative for these species orgroups of these species (eg Hopwood 1933 Le Gros Clark andLeakey 1951 Andrews 1978 Pickford et al 2009) However wefound this tooth to be unreliable for taxonomic purposes Withinprimates M3s are typically the most variable molars in both sizeand shape (Blankenship et al 2007 Kavanagh et al 2007) and this
Figure 6 Occlusal view of the M2s of Proconsul and Ekembo Left P africanus holotype(M 14084) Right E heseloni holotype (KNM-RU 2036) Note in Proconsul the overalldifference in crown shape as well as the more conical individuated cusps and nar-rower cristae strongly developed lingual cingulum that continues to the distal aspectof the crown broad buccal cingular shelf and more distally directed posthypoconecrista In Ekembo the cusps are more bunodont and expanded the cristae are broaderwith better developed accessory tubercles the lingual cingulum merges into thehypocone there is no buccal shelf and the posthypocone crista is more buccallydirected
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6152
is observable even in small samples of Proconsul species TheP africanus holotype has a substantially reduced M3 whereas inKNM-CA 2250dattributed by most authors to this species(Andrews 1978 Pickford et al 2009)dit is fully developed Simi-larly the hypodigm of P major includes both fully developed M3s(eg KNM-CA 389 LG 1815) and specimens with reduced ormissing hypocones (KNM-CA 397 1299) Ekembo displays compa-rable ranges of variation from highly reduced to fully developedM3s in both species Thus these species and genera cannot bedistinguished based on M3 development and we did not find anyother distinguishing characteristics between Ekembo and Proconsulin the upper third molars
Figure 7 Scatter plot depicting the de
Mandible Pickford et al (2009) enumerated several traits thatreadily distinguish Kisingiri from non-Kisingiri mandibles andthese distinctions are entirely consistent with the distinction ofEkembo from Proconsul Our analyses specifically underscoredifferences in the symphyseal region The planum alveolare inboth male and female specimens of Ekembo is short and morevertically oriented even the most elongate of these (KNM-RU47805 McNulty et al 2007) is substantially more verticalcompared to specimens of Proconsul In contrast the planumalveolare in Proconsul is conspicuously long and nearlyhorizontally oriented Well-preserved mandibles attributed toP major (eg KNM-SO 396 and 404) exhibit this morphology inextreme fashion (Fig 8) smaller specimens (KNM-SO 1112P africanus) show less extreme expression but are still readilydistinguishable from Ekembo mandibles For its size KNM-LG 452has a surprisingly short planum alveolare though not to thedegree seen in Rusinga mandibles Correlated to the shortplanum alveolare in Ekembo is a reduced superior transversetorus which while well-developed does not approach therobusticity expressed in Proconsul mandibles This difference isreflected in an index of symphyseal robusticity (thicknessperpendicular to the long axis of the symphysislength of thelong axis) which shows Ekembo and Proconsul to be significantlydifferent (t-test df frac14 14 t frac14 430 p frac14 00007)
Development of the inferior transverse torusdoften used todifferentiate among species of Proconsul (eg Pickford et al 2009Harrison 2010)dwas found to be highly variable and inconsis-tently expressed within these samples However the overallorientation of the subplanum region (as defined by a line con-necting the posterior-most midline points of the superior andinferior transverse tori or the posterior-most midline point of theinferior margin of the symphysis in the absence of the latter) doesdistinguish Ekembo from Proconsul mandibles The orientation ofthis line in relation to the alveolar margin in the vicinity of thepostcanine teeth was assessed qualitatively by placing a small rod
gree of buccolingual flare in M2
Figure 8 Mandibles of Proconsul and Ekembo Occlusal views of P major a) KNM-SO 396 b) KNM-SO 404 c) KNM-LG 452 Occlusal views of Ekembo d) KNM-RU 2087 e) KNM-RU1674 f) KNM-RU 7290 Occlusal views of P africanus g) KNM-SO 372 h) KNM-SO 1112 i) Lingual view of P africanus (KNM-SO 1112) Note the long more horizontally orientedplanum alveolare in Proconsul resulting in a pronounced superior transverse torus The planum alveolare is shorter and more vertical in Ekembo such that one can see the inferiormandibular border from occlusal view
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 53
(eg a dental pick) in themidline internally along the two referencepoints and recording the point in lingual perspective at which therod intersected the alveolar margin (Fig 9) In Ekembo the sub-planum region is more vertically oriented so that the rod intersectsthe alveolar margin most often in the premolar field Not surpris-ingly there is a difference between sexes with females having areduced superior transverse torus and therefore a more verticallyoriented subplanum region compared to males In contrast withEkembo Proconsul mandibles have much more inclined subplanumsurfaces with the rod intersecting the lingual alveolar margin inthe vicinity of the M1 (females) or M2 (males Fig 9) When sex istaken into account therefore the distributions of Ekembo andProconsul in this feature are discrete Note that male and femalemandibles attributed to both P africanus and P major are includedin this assessment and are surprisingly similar despite dramaticsize differences
We also quantified internal symphyseal morphology using threemeasures symphyseal height (infradentaleegnathion) planumalveolare length (posterior-most point on the superior transversetoruseinfradentale) and subplanum length (posterior-most pointon the superior transverse torusegnathion) a combination thatincorporates not only the lengths of these intervals but also theangular relationships among them To compare symphyseal shapeacross a range of sizes we chose a simple principal componentsapproach to partition size onto the first eigenvector (cf Jolicouer1963) While this is not the preferred method for partitioning sizein most datasets (Jungers et al 1995) for this particular sample andset of measurements Mosimann shape variables were still
significantly correlatedwith size (see discussion in Coleman 2008)Hence we assessed differences in symphyseal morphology be-tween Ekembo and Proconsul using the second and third principalcomponents of the three variables Distributions of the two generaare distinct in this shape space (Fig 10 Supplementary OnlineMaterial [SOM] Table 1)
Robusticity of the mandibular corpus also differs between thesesamples though it is more difficult to test due to differentialpreservation among specimens Corpus thickness and depth underP4 can be measured in the greatest number of specimens andcomparison of these measures suggests a difference betweengroups in the scaling of corpus robusticity (Fig 11) Among largepresumably male specimens Ekembo mandibles are substantiallyless robust than those of Proconsul However among smallermandibles only a single Proconsul specimen (KNM-SO 372) could beincluded and it is similar in robusticity to Ekembomandibles of thissize Considered together the impression is that corpus robusticitymay scale differently in Ekembo than it does in Proconsul and a testfor differences in slopes using an ANCOVA design of the loggedvariables was significant (df frac14 2 F frac14 2481 p lt 00001) Never-theless additional specimens are needed to properly evaluate theinfluence of taxonomy sex and size on corpus robusticity areasonable alternative interpretation of Figure 11 is that it dem-onstrates a significant difference between P major and all otherspecies of both Proconsul and Ekembo
Lower incisors Ekembo specimens are uniformly narrower inmesiodistal length than incisors attributed to P africanus P major
Figure 10 Scatter plot of principal component scores based on an eigenanalysis of threeposterior point on the superior transverse toruseinfradentale) and subplanum length (mprimarily captures size differences among specimens (Jolicouer 1963 see text and SOM Trespectively) in the symphyseal variables
Figure 9 Line drawing of a hemimandible depicting different orientations of the sub-planum area in Ekembo and Proconsul Females are denoted using shorter vectors maleswith longer vectors Vector is estimated in M 14086 which is broken at the midlineEkembo specimens have a more vertical subplanum compared to those from Songhorand Legetet but within both groups females have more vertically oriented regions thanmales Hence even the most ventrally rotated subplanum in the largest Ekembo malespecimen (KNM-RU 47805) groups with the Tinderet females in this feature
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6154
and P meswae both absolutely and in relation to crown height(Fig 12) This is true for both I1 and I2 A t-test adjusted for non-independent observations (eg I1 and I2 from the same mandible)confirmed the difference as significant (t-test df frac14 12 t frac14 258p frac14 00242) Moreover while the measurable Proconsul samplewas small this same pattern is observable in other specimenswhose tips were slightly to moderately worn and therefore notmeasured
Lower canines The only lower canine trait that discretely sepa-rates these genera is the pointed crown tip in Ekembo (Fig 3d)contrasted with a blade-like (or ldquoburinrdquo) tip in Proconsul(Fig 3c) This feature is well established for P major but is alsoexpressed in KNM-CA 1772 which was referred to P africanus byKelley (1986) and is too small to be either P major or P meswaeTwo other specimens commonly attributed to P africanus KNM-CA 2149 and KNM-SO 1112 both have broken canine tipsLikewise a new specimen (Kor 65004) attributed to P africanusby Pickford et al (2009) also has a broken tip and cannot beevaluated (see Pickford et al 2009 Fig 23) As with the uppercanines the blade-like tip is also present in the deciduous lowercanines of P major (eg KNM-CA 361 S0 5352) and P meswae(KNM-ME 1)
Lower premolars Like most previous authors (eg Andrews 1978Harrison 2002 2010 Pickford et al 2009) we found neitherqualitative nor quantitative characteristics of the lower premolarswith any taxonomic value
Lower molars Differences in lower molar morphology are few andrelatively subtle but in some cases mirror those seen in the uppermolars Cusps on the lower molars of Ekembo are more bunodontthan those in Proconsul and occupy more of the occlusal surfaceBroad inter-cuspal crests are incorporated into the Ekembo cuspsmaking them appear more inflated Molar cusps of Proconsul are
variables symphyseal height (infradentaleegnathion) planum alveolare length (mostost posterior point on the superior transverse torusegnathion) The first componentable 1) and thus PC 2 and PC 3 illustrate the residual shape variance (72 and 28
Figure 11 Scatter plot showing mandibular corpus robusticity under P4 Trend lines were computed using ordinary least squares (OLS) regression Similar relationships areobservable at other parts of the corpus but differential preservation results in smaller samples at those positions Slopes are significantly different (p lt 00001 tested on loggedvariables)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 55
more distinct from one another and are linked by narrower sharperinter-cuspal crests The lower molars of Ekembo also have asubstantially less developed buccal cingulum than those ofProconsul (Harrison 2002 2010 Harrison and Andrews 2009)
Figure 12 Scatter plot of lower incisor crown proportions Differenc
Lastly and again as seen in the upper molars the two genera canbe distinguished by the significantly greater amount of lowermolar flare in Proconsul (Fig 13 t-test df frac14 59 t frac14 713p lt 00001 see also Harrison and Andrews 2009)
es between Ekembo and Proconsul are significant (p frac14 00242)
Figure 13 Box-and-whisker plot of buccolingual flare in lower molars measured asthe average distance between the mesial buccal and lingual cusps divided by overallbuccolingual crown breadth Differences between Ekembo and Proconsul are highlysignificant (p lt 00001)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6156
Relative molar size has been used to distinguish species ofProconsul but we observed the primary difference to be in relativebuccolingual breadths of M1 and M2 In Ekembo M1 is substantiallynarrower than M2 than is the case in Proconsul (Fig 14 Harrison2002 2010) Although none of the smaller Proconsul mandiblespreserve complete crowns for both M1 and M2 KNM-SO 1112 hasan intact M2 and at least preserves the cervix for M1 However evenif the cervical measurement is used for its M1 breadthdthusunderestimating the true crown breadthdKNM-SO 1112 still has abroader first molar in relation to M2 than would be expected inEkembo If crown breadth is estimated from the ratio of cervixbreadth to maximum crown breadth in other Proconsul fossils therange of likely breadths for the SO 1112 M1 clearly distinguishes it
from the Ekembo sample (Fig 14) Testing this relationship onlogged variables using the mean estimate for the SO 1112 M1(Fig 14) reveals a significant difference between the two groups(ANCOVA df frac14 1 F frac14 2424 p frac14 00002)
As with upper third molars we find a great deal of variation inM3 morphology and mistrust the taxonomic significance some-times placed on characteristics of this toothdat least with regard tothese taxa Harrison (2002 2010) cited the mesiodistal position ofthe M3 entoconid as differentiating Kisingiri third molars fromthose attributed to P africanus We concur with his assessment butfound the entoconid position to vary in specimens attributed toP major Thus this character does not differentiate Ekembo from allProconsul species but may distinguish it from the type speciesP africanus
Discussion
The distinctiveness of the Kisingiri large catarrhine materialfrom that at Tinderet and Ugandan localities has been noted pre-viously by several authors (Pickford 1986 Walker et al 1993Begun 2001 2007 MacLatchy and Rossie 2005 Harrison andAndrews 2009 Harrison 2010) What had not been fully docu-mented was the nearly complete lack of overlap between thesesamples in a number of features despite very large assemblagesfrom Rusinga and Mfangano On descriptive criteria alone (Table 2)many isolated teeth (I1 C1 P3 M2 C1) and all of the mandibles canbe assigned to either Proconsul or Ekembowith very little chance ofmisattribution That some of the most diagnostic features occur inthe upper dentitiondthe only anatomy preserved in theP africanus type specimendadds weight to the taxonomicarrangement proposed here The morphology of the upper teeth inall Proconsul species compellingly unites them into a single cladedistinct from E heseloni and E nyanzae This is reinforced bynumerous features of the mandible and mandibular dentition thatfurther distinguish these samples The number and variety of thesefeatures are such that Proconsul and Ekembo are more differentfrom one another than are many closely-related modern catarrhinegenera
Derived morphology of Ekembo and Proconsul
It is premature to formally assign primitive and derived status tothe individual features of Ekembo and Proconsul before completinga formal phylogenetic analysis This is especially true given theabundance of evidence available for Ekembo and hence the poten-tial impact that our generic revision could have on phylogeneticinterpretations Nevertheless in comparison with both extanthominoids and the small-bodied catarrhines from the earlyMiocene it is informative to differentiate the evolutionary grades ofthese genera Many characteristics of Ekembo are more advanced inthe direction of modern apes compared to those of Proconsul(Table 2) General trends such as expanded and more bunodontcusps broader molar crests reduced cingulum and morphologicalsimilarity between P3 and P4 all represent morphologies thatpresage those of later fossil and extant hominoids The moreadvanced morphologies of P3 and M2 are particularly conspicuouscompared not only with those of Proconsul but to most earlyMiocene catarrhines In contrast the mesially positioned stronglyheteromorphic cusps on the P3 of Proconsul plus the correspondingasymmetric buccal profile notched mesial margin and missingmesial fovea are characteristic of other early Miocene catarrhinessuch as Dendropithecus Rangwapithecus and Limnopithecus andare likely primitive among East African catarrhines The moresymmetric P3 of Ekembo with midline positioned cusps andreduced cusp heteromorphy is reminiscent of later hominoids and
Figure 14 Relative buccolingual dimensions of M1 and M2 KNM-SO 1112 has a preserved M2 but only the cervix of M1 Its M1 breadth was estimated based on the ratio to cervixbreadth to maximum crown breadth in other Tinderet fossils Even the cervical breadth of SO 1112 is broader than would be predicted for the crown breadth of a Kisingiri M1 Trendlines computed using OLS regression and using the average estimate of SO 1112 M1 breadth Group differences were significant (p frac14 00002)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 57
even hominid taxa such as Rudapithecus (eg Kordos and Begun2001)
Proconsul is likewise united by a suite of features that are for themost part characteristic only of this lineage (Table 2) Themorphology of both upper and lower canines is unique to thisgenus and shared by all three well-known species plus the poorlyrepresented middle Miocene P gitongai (Pickford and Kunimatsu2005) The mandibular symphysis is also distinctive in Proconsulcompared with other catarrhines with its very long and shallowlysloping planum alveolare and steeply inclined subplanum surfaceIndeed even the small edentulous anterior mandibular fragmentKNM-SO 372 is readily placed within Proconsul and excluded fromother similarly sized catarrhine taxa such as Rangwapithecus andEkembo based on this unique symphyseal morphology (Figs 8 and9)
Other differences between Ekembo and Proconsul are of uncer-tain evolutionary polarity (eg orientation of the posthypoconecrista) or may be of taxonomic relevance without broader phylo-genetic significance (eg degree of molar flare Table 2) Werecognize that there are morphological differences within Procon-sul particularly between the better known species P africanus andP major However many of these differences are likely related tosize and represent variations on a single generic theme that aremarkedly different from the morphology of Ekembo The recentdescription (Harrison and Andrews 2009) of the intermediatelysized P meswae underscores the broad morphological unity withinProconsul and further substantiates a well-defined Proconsul cladecomprising Tinderet and Ugandan species
Based on the abundant morphological and morphometric evi-dence supporting genus-level distinction between Proconsul andEkembo we are currently conducting a more detailed phylogeneticanalysis of all early Miocene catarrhines in order to better under-stand the distribution and evolutionary significance of these andother morphological features We anticipate that the debates sur-rounding the phylogenetic relationships of an historically consti-tuted sample of Proconsul can be more satisfactorily resolved nowthat the Kisingiri material is appropriately recognized as a distinctgenus
The generic disposition of ldquoUgandapithecusrdquo species
A great many features said to characterize ldquoUgandapithecusrdquowere added to the original diagnosis of the genus by Pickford et al(2009) in their systematic revision of Proconsul (sensu lato) As partof that study they also erected a new species ldquoUrdquo legetetensis andtransferred P meswae to ldquoUgandapithecusrdquo Evaluating the same setof features across the entire available sample however we wereunable to find support for this more expansive conception ofldquoUgandapithecusrdquo for several reasons First some diagnostic fea-tures cited in Pickford et al (2009) were quite variable within theldquoUgandapithecusrdquo sample As an example marked reduction of theM3 entoconid to a bead on the lingual cingulum is not shared byseveral ldquoUrdquo major specimens such as KNM-CA 393 NAP I 49000and Kor 253004 (see Pickford et al 2009 Fig 33e) and was likelynot a feature of a few of the more worn specimens eg BUMP 600and BUMP 601 or even the holotype of P major M 16648 Second
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6158
some diagnostic features of ldquoUgandapithecusrdquo were found to bevariably present in Ekembo arguing against their utility for dis-tinguishing ldquoUgandapithecusrdquo from Pickford et als (2009) com-bined P africanus thorn Kisingiri conception of Proconsul For examplesignificantly reduced curvature of the I1 distal margin in ldquoUganda-pithecusrdquo is also found in some Kisingiri specimens including KNM-RU 1685 1712 1714 1831 1846 and 1951 several more Rusingaspecimens have moderately reduced curvature Third some fea-tures that purportedly distinguish ldquoUgandapithecusrdquo fromP africanus are likely related to size For example the somewhatmore central position of the P3 paracone in ldquoUrdquomajor is not seen inthe smaller ldquoUrdquo meswae whose paracone is positioned similarly tothat of the holotype of P africanus (see Harrison and Andrews2009 Fig 6d)
However the major part of the evidence for recognizingldquoUgandapithecusrdquo (sensu Pickford et al 2009) was assembled by re-assigning specimens traditionally attributed to P africanus into newspecies of ldquoUgandapithecusrdquo The P africanus holotype was left inassociation with the Rusinga sample despite having morphologyoverwhelmingly characteristic of other Tinderet specimens Theresulting specimen allocations left P africanus morphologicallyimpoverisheddstripped of much of its hypodigmdso that Pro-consul for Pickford et al (2009) came to be characterized primarilyby the Rusinga sample (see similar comments in Harrison andAndrews 2009) Within that conceptual framework any spec-imen that differs from those at Rusinga could not be Proconsul butmust instead belong to ldquoUgandapithecusrdquo Thus specimens such asthe KNM-SO 1112 mandible were erroneously assigned to ldquoUgan-dapithecusrdquo because they were (correctly) perceived as differingfrom their Rusinga counterparts (Pickford et al 2009)
Here we restore the P africanus hypodigm and associate it withthe other Tinderet fossils according to their shared morphologySpecimens assigned to ldquoUgandapithecusrdquo revert to Proconsul andmany of the features cited to support a speciose ldquoUgandapithecusrdquo(Pickford et al 2009) instead become diagnostic features of Pro-consul This correction serves to highlight the distinctiveness of theKisingiri remains and the need to recognize this distinctiveness atthe generic level
As demonstrated above specimens from Kisingiri do not displaythe morphology of Proconsul as represented by the holotype ofP africanus M 14084 Given that this specimen preserves only theupper canine and cheek teeth similarities with or departures fromthis morphology represent the most compelling arguments forassessing inclusivity in the genus However for the mandible andlower dentition if one simply sorts the specimens by elementwithout relying on taxonomic preconceptions the overwhelmingdistinction within the traditionally constituted Proconsul sample isbetween Kisingiri material on the one hand and that from Tinderetand Uganda on the other (cf Walker et al 1993 Begun 2001 2007MacLatchy and Rossie 2005 Harrison and Andrews 2009Harrison 2010) Additionally with the Kisingiri specimensremoved to Ekembo the postcranial differences used to partlyjustify a distinct ldquoUgandapithecusrdquo lose their relevance (seeHarrison and Andrews 2009)
We have differed among ourselves in our past treatment ofP major as generically distinct from the smaller species of Pro-consul (Begun 2007 McNulty 2010) Morphologically many ofthe unique features of P major can be interpreted as size-relatedvariants on the smaller P africanus morphology Even if so thequestion is whether these are sufficiently different and numerousto warrant generic distinction that is do they suggest thatP major belongs in a separate clade from P africanus andP meswae As we have demonstrated differences among thesethree species are certainly fewer and show less morphologicaldistinction than those between Ekembo and any of the three
Nevertheless while we ultimately agree with Harrison andAndrews (2009 see also Begun 2007) that the features dis-tinguishing ldquoUrdquo major from P africanus are consistent with thosethat distinguish congeneric species and are at least partly due todifferences in size we concede the possibility that the largestProconsul species could belong in a separate genus (Senut et al2000 Pickford and Kunimatsu 2005) However whether or notone recognizes a separate genus for P major the P africanus ma-terial does not sort morphologically with the Kisingiri materialTherefore the taxonomic alternatives are either two generaProconsul and Ekembo or three Proconsul Ugandapithecus andEkembo with Proconsul and Ugandapithecus united in a cladeexclusive of Ekembo Thus while we strongly favor the firstalternative recognizing Ugandapithecus does not change thefundamental argument concerning the need to refer the Kisingirispecies to a generically and cladistically distinct taxon from thespecies represented at Tinderet and the Ugandan sites
Specimens of interest
Despite the substantial dental and gnathic differences betweenProconsul and Ekembo there are a few specimens whose taxonomicaffinities require additional scrutiny
KNM-RU 1676-77 This set of associated upper and lower teeth is aconsistent outlier among E heseloni and E nyanzae specimens Insome ways this specimen more closely resembles Tinderet Pro-consul in the features that distinguish it from Ekembo Molar cuspmorphology and cingulum development I1 lingual tubercledevelopment and M2 shape and posthypocone crista orientationare more consistent with specimens from the Tinderet sites On theother hand it otherwise resembles Ekembo and lacks the strongestdiagnostic features that unite species of Proconsul such as thedistinctive canine and P3 morphology Overall this specimen doesnot fit unambiguously into either genus in fact the degree ofbuccolingual flare cusp morphology enamel wrinkling and espe-cially the long extension of enamel onto the anterior root in P3 arein some ways reminiscent of Afropithecus although it clearly differsfrom that genus as well (cf Rossie and MacLatchy 2013)
Were these characteristics individually distributed among thelarge Kisingiri sample one might attribute this to intra-genericvariation but when found together in a single specimen from alarge sample it suggests a unique individual One of us (KPM)thinks it likely that the specimen was not actually found at aKisingiri locality based on its unique preservation compared toother Rusinga fossils and on other examples of mis-proveniencedspecimens (Leakey 1967 Andrews and Molleson 1979 Pickford1986 Kelley 1986) However we recognize that relying toofrequently on this explanation allows almost any scenario to bejustified and suggest some alternative possibilities One is that thespecimen was found on Rusinga but in the much older WayandoFormation strata instead of the Hiwegi Formation The differencesin morphology in that case might reflect more than a million yearsof temporal change (McCollum et al 2013) with KNM-RU 1676-77preserving certain more primitive Proconsul-like features As analternative it is reasonable to assume that fossil assemblages asvast as those from Rusingawill include not only abundant taxa butalso rare or infrequent inhabitants of the local biome There aremany examples of this from Rusingas mammalian communityeg ldquoHemicyonrdquo Kulutherium Tadarida cf Hyainailouros cfArchaeobeledon Kelba (see relevant chapters in Werdelin andSanders 2010) As such KNM-RU 1676-77 may simply representa rare and well-preserved specimen of some other catarrhinetaxon that lived among or visited a primate community dominatedby Ekembo
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 59
KNM-SO 1112 This specimen was initially referred to Rangwapi-thecus but soon after was transferred to Proconsul afrianus (Bosler1981 Martin 1981)da designation that most subsequent authorshave followed (Pickford 1986 Nengo and Rae 1992 Kelley 19861995 Walker et al 1993 MacLatchy and Rossie 2005 Hill et al2013) Pickford et al (2009) more recently referred this specimento ldquoUrdquo meswae citing two primary reasons First they reiteratedBoslers (1981) concern that its P3 differs from others attributedto P africanus However at that time the majority of what wasconsidered to be P africanus would have been from RusingaIsland In fact while noting its contrasting morphology Boslersuggested that differences between KNM-SO 1112 and other P3smight reasonably be attributed to sexual dimorphism notingthat this specimen provides ldquoa not implausible match for the Pafricanus holotype maxilla M 14084rdquo (1981152) Thatassessment is borne out in this study Using the much largersample of Ekembo P3s as a model variation can be seen toencompass specimens with well-developed mesial beaks andthose with more bilaterally compressed crowns and parallelsides The latter morphology is predominantly found amongmale specimens (KNM-RU 1674 1924 and KPS I and III) thussupporting Boslers (1981) suggestion that this variation isrelated to sexual dimorphism and hence that KNM-SO 1112 is amale P africanus
The second feature cited by Pickford et al (2009) in referringthis specimen to ldquoUrdquo meswae was posterior shallowing of themandibular corpus However because they redefined ldquoUgandapi-thecusrdquo to include all of the well-preserved Tinderet mandibles(Pickford et al 2009)dleaving Proconsul to be represented only byRusinga specimensdtheir only alternative within this scheme for aspecimen the size of SO 1112 was to refer it to ldquoUrdquomeswae It seemsodd though to assign this specimen to a species from a (perhapsmuch) older site when the Proconsul africanus holotype is ofappropriate size and similar age and as there are no other adultmandibles from either species with which to compare it In anycase mandibular shallowing is variable in both Ekembo and Pro-consul with some Kisingiri specimens exhibiting considerableshallowing (eg KNM-RU 2087) and some ldquoUgandapithecusrdquo spec-imens having very little (eg M 14086)
We therefore see no morphological grounds to assign KNM-SO1112 to P meswae but do find some morphological evidence toassign it as a male P africanus which occurs at Songhor
KNM-LG 452 This very complete mandible was originally recog-nized as a female of P major based on the size of the dentition androbusticity of the mandible (Martin 1981) Bosler (1981) howeverleft this specimen unassigned noting similarities to P major in P3-M1 size but having distinctly smaller M2e3 In this she likenedKNM-LG 452 to a few other specimens (notably M 14086) thatare smaller in their distal molars in a manner more consistentwith taxonomic rather than sex differences (Bosler 1981 see alsoPilbeam 1969) Kelley (1986 1995) also assigned this specimen asa female of P major based on canine size and morphology butnoted that the canine lacked the shallowly-sloping mesial ridgetypical of most female anthropoid canines This is also truehowever of two other canines assigned as female P major KO 9and SO 373 (Kelley 1986) and so might simply be an unusualcharacteristic of P major female lower canines In the end Kelley(1995) concluded that considering all of the criteria fordistinguishing male and female canines the evidence that KNM-LG 452 is female remains questionable
Pickford et al (2009) further developed the idea that two spe-cies are represented within the P major sample and made KNM-LG452 the holotype of a new species ldquoUrdquo legetetensis We concur thatthis specimen is unique among the well-preserved Tinderet
mandibles in some important features Whereas male and femalemandibles of P major have long internal symphyseal planesexpansive extra- and retromolar sulci and rami that cross theocclusal plane at distal M3 KNM-LG 452 has a shorter symphysealplane (though not to the extent seen in Ekembo) a moderateextramolar sulcus but no retromolar sulcus and a ramus thatcrosses the occlusal plane more mesially If KNM-SO 396 and SO404 represent males and females respectively of P major then themorphology of KNM-LG 452 appears distinct
At present however we do not think there is sufficient evidenceto differentiate this specimen from either P major or P meswaePickford et al (2009) argue that KNM-LG 452 cannot belong to thelatter species because of its larger size The size differences how-ever are well within the range of variation seen in P majorE heseloni and E nyanzae Moreover whereas the Legetet mandibleis larger than most of the P meswae hypodigm an adult P4 referredto this species (KNM-ME 25) substantially exceeds its counterpartin KNM-LG 452 Should the Legetetmandible ultimately prove to bea male specimen size alone would not rule out its attribution toP meswae On the other hand if it is female then it becomes harderto justify its inclusion in that sample of mostly smaller specimens Iffemale therefore it could represent a separate species for whichthe binomial Proconsul legetetensis comb nov is available Recentlycollected fossils from the SonghorKapurtay area provide someadditional support for the possibility that three species of Proconsulexisted at these localities (McNulty 2014ab) However toconvincingly demonstrate this and determine whether a thirdspecies can be differentiated from P meswae will require a moredetailed analysis of variation among Proconsul species a corre-sponding study of variation in modern taxa anddalmost certain-lydadditional fossil specimens For now we regard ldquoUrdquo legetetensisas a subjective junior synonym of P major
Conclusions
The genus Proconsul has been central to studies of ape and hu-man evolution since its first description by Hopwood (1933) As oneof the best known fossil catarrhines represented by nearly everyskeletal element our knowledge of Proconsuls functional anatomypositional behavior and overall morphology has made it thefoundation for paleoanthropological research in the early Mioceneas well as studies on the origin and early diversification of homi-noids With respect to systematics however Proconsul is less wellunderstood the type species P africanus is known from a rela-tively small number of specimens from sites in the Tinderet areaand no postcranial bones can be unambiguously assigned to itRather it is the copious and well-preserved specimens from theKisingiri sites of Rusinga and Mfangano Islands that have formedthe basis for the common image of Proconsul
We argue here that the Kisingiri species traditionally referredto ProconsuldP heseloni and P nyanzaedare generically distinctfrom P africanus and hereby transfer them into a new genusEkembo based on a substantial number of morphological differ-ences (Table 2) Considering only those traits preserved in theP africanus holotype the entire sample from Rusinga and Mfan-gano differs markedly in the morphology of the upper canine P3P4 and M2 Additional differences in the I1 lower incisors andcanines molar cusp and occlusal crest morphology degree of flarein all molars and especially mandibular features clearly distin-guish the Kisingiri from non-Kisingiri samples in a way that isincompatible with congeneric species With the transfer ofE heseloni and E nyanzae Proconsul becomes a well-defined cladecomprising at least three speciesdP africanus P major andP meswaedunited by several features that are unique for thegenus (Table 2)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6160
We found no support in this study for the genus ldquoUgandapi-thecusrdquo as constructed by Pickford et al (2009) In that paper theytransferred the vast majority of Proconsul fossils from the Tinderetand Napak localities into ldquoUgandapithecusrdquo leaving P africanusrepresented only by its holotype and a small number of isolatedteeth and aligning it with the material from Rusinga and Mfan-ganoWhen the P africanus hypodigm is reconstituted and properlyaffiliated with the other Tinderet species according to itsmorphology much of the evidence marshaled by Pickford et al(2009) for ldquoUgandapithecusrdquo actually supports the distinction be-tween the Tinderet and Kisingiri samples ie the generic distinc-tion of Ekembo In addition some of the traits enumerated byPickford and colleagues in support of ldquoUgandapithecusrdquo wererevealed in our analysis to be variable within one or more speciesand therefore of questionable taxonomic utility We do not rejectout of hand the original conception of ldquoUgandapithecusrdquo (Senutet al 2000) as either a monotypic genus distinguishing ldquoUrdquo ma-jor or perhaps including a closely allied middle Miocene ldquoUrdquogitongai as well (Pickford and Kunimatsu 2005) but we have fornow retained the early Miocene specimens of both in Proconsulpending further study However even if Ugandapithecus is a taxo-nomically valid genus comprising just the latter two species theirmorphology indicates that theywould still be united in a cladewithProconsul and they lack several more advanced features that partlydefine Ekembo Better understanding of the role that body size playsinmediating differences within Proconsulwill help to further clarifythe relationships among these species
Despite the existence of one well-represented and several lesscomplete skeletons of what was long assumed to be Proconsul thephylogenetic status of this genus has defied consensus This islargely due in our view to Proconsul having been an admixture oftwo distinct genera one more advanced in the direction of extantapes than the other As we demonstrate here the material fromRusinga and Mfangano traditionally referred to Proconsul andwhich includes all of the more complete skeletal material isdistinct from the Tinderet and Napak material and belongs in aseparate genus Ekembo By disentangling these distinct clades weanticipate being able to achieve better phylogenetic resolution forboth genera thereby improving our knowledge of early hominoidevolution and diversification
Acknowledgments
We thank many people and agencies that assisted with thedevelopment and completion of this project This research wasinitiated as a result of funding from the Wenner-Gren Foundationand the Johns Hopkins School of Medicine to DR Begun in1987e1989 and was renewed following research and travelgenerously funded by multiple Leakey Foundation grants to KMcNulty National Science Foundation grants to K McNulty(0852609 1241807) the McKnight Foundation the University ofMinnesota the Lloyd A Wilford Endowment the LeverhulmeFoundation and the Turkana Basin Institute J Kelley gratefullyacknowledges support from the Institute of Human Origins Ari-zona State University DR Begun also wishes to acknowledge theAlexander Humboldt Stiftung and NSERC for funding support Weare grateful to the National Museums of Kenya the UgandaMuseum the Natural History Museum and to Rose Mwanja EzraMusiime Tom Mukhuyu Samuel Muteti James Yatich and MaryMuungu for access to and assistance with specimens in their careMany people provided assistance comments criticisms insightsand suggestions on various aspects of this work and we especiallythank Holly Dunsworth Will Harcourt-Smith Tom Lehmann DanPeppe Nicole Garrett Kirsten Jenkins Rutger Jansma Susy CoteIsaiah Nengo Andrew Hill and Brenda Benefit K McNulty
acknowledges the evolutionary anthropology faculty and studentsat Durham University for their helpful comments on an earlyversion of this manuscript and for their gracious hospitality Weare likewise grateful to the anonymous reviewers associate editorand editor whose comments strengthened the paper This manu-script is publication 1 supporting Research on East AfricanCatarrhine and Hominoid Evolution (REACHE)
Appendix A Supplementary data
Supplementary data related to this article can be found at httpdxdoiorg101016jjhevol201503009
References
Andrews P 1974 New species of Dryopithecus from Kenya Nature 249 188e190Andrews P 1978 A revision of the Miocene Hominoidea of East Africa Bull Brit
Mus (Nat Hist) Geol 30 85e225Andrews P 1985 Family group systematics and evolution among catarrhine pri-
mates In Delson E (Ed) Ancestors The Hard Evidence Liss New Yorkpp 14e22
Andrews P Harrison T Martin L Pickford M 1981 Hominoid primates from anew Miocene locality named Meswa Bridge in Kenya J Hum Evol 10123e128
Andrews P Begun D Zylstra M 1997 Interrelationships between functionalmorphology and paleoenvironments in Miocene hominoids In Begun DWard C Rose M (Eds) Function Phylogeny and Fossils Miocene HominoidEvolution and Adaptations Plenum Press New York pp 29e58
Andrews P Molleson T 1979 The provenance of Sivapithecus africanus Bull BritMus (Nat Hist) Geol 32 19e23
Begun DR 2001 African and Eurasian Miocene hominoids and the origins of theHominidae In de Bonis L Koufos GD Andrews P (Eds) Hominoid Evolutionand Climatic Change in Europe Phylogeny of the Neogene Hominoid Primatesof Eurasia Cambridge University Press London pp 231e253
Begun DR 2007 Fossil record of Miocene hominoids In Henke W Tattersall I(Eds) Handbook of Paleoanthropology Springer Berlin pp 921e977
Begun DR Teaford MF Walker AC 1994 Comparative and functional anatomyof Proconsul phalanges from the Kaswanga primate site Rusinga Island KenyaJ Hum Evol 26 89e165
Bishop WW 1964 More fossil primates and other Miocene mammals from North-East Uganda Nature 203 1327e1331
Bishop WW Miller JA Fitch FJ 1969 New potassium-argon age determinationsrelevant to the Miocene fossil mammal sequence in East Africa Am J Sci 267669e699
Blankenship JA Mincer HH Anderson KM Woods MA Burton EL 2007Third molar development in the estimation of chronologic age in Americanblacks as compared with whites J Forensic Sci 52 428e433
Bosler W 1981 Species groupings of early Miocene dryopithecine teeth from EastAfrica J Hum Evol 10 151e158
Coleman MN 2008 What does geometric mean mean geometrically Assessingthe utility of geometric mean and other size variables in studies of skullallometry Am J Phys Anthropol 135 404e415
Cote S Malit N Nengo I 2013 Additional mandibles of Rangwapithecus gordonian early Miocene catarrhine from the Tinderet localities of Western KenyaJ Hum Evol 153 341e352
Deane AS 2009 Early Miocene catarrhine dietary behavior the influence of theRed Queen Effect on incisor shape and curvature J Hum Evol 56 275e285
Drake R Van Couvering JA Pickford M Curtis G Harris JA 1988 New chro-nology for the Early Miocene mammalian faunas of Kisingiri Western KenyaJ Geol Soc Lond 145 479e491
Evernden JF Savage DE Curtis GH James GT 1964 Potassium-argon datesand the Cenozoic mammalian chronology of North America Am J Sci 262145e198
Gebo DL MacLatchy L Kityo R Deino A Kingston J Pilbeam D 1997A hominoid genus from the Early Miocene of Uganda Science 276 401e404
Geoffroy Saint-Hilaire E 1812 Tableau des quadrumanes 1 Ord QuadrumanesAnnls Mus Hist Nat Paris 19 85e122
Gray JE 1825 Outline of an attempt at the disposition of the Mammalia into tribesand families with a list of the genera apparently appertaining to each tribeAnnls Philos 10 337e344
Harrison T 1982 Small-bodied apes from the Miocene of East Africa PhDDissertation University of London
Harrison T 1986 New fossil anthropoids from the middle Miocene of East Africaand their bearing on the origin of the Oreopithecidae Am J Phys Anthropol 71265e284
Harrison T 1987 The phylogenetic relationships of the early catarrhine primates areview of the current evidence J Hum Evol 16 41e80
Harrison T 2002 Late Oligocene to middle Miocene catarrhines from Afro-ArabiaIn Hartwig WC (Ed) The Primate Fossil Record Cambridge University PressCambridge pp 311e338
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 61
Harrison T 2010 Dendropithecoidea Proconsuloidea and Hominoidea (CatarrhiniPrimates) In Werdelin L Sanders WJ (Eds) Cenozoic Mammals of AfricaUniversity of California Press Berkeley pp 429e469
Harrison T Andrews P 2009 The anatomy and systematic position of a newspecies of early Miocene proconsulid from Meswa Bridge Kenya J Hum Evol56 479e496
Hill A Nengo IO Rossie JB 2013 A Rangwapithecus gordoni mandible from theearly Miocene site of Songhor Kenya J Hum Evol 65 490e500
Hopwood AT 1933 Miocene primates from Kenya J Linn Soc Zool 38 437e464Ishida H Kunimatsu Y Takano T Nakano Y Nakatsukasa M 2004 Nacholapi-
thecus skeleton from the Middle Miocene of Kenya J Hum Evol 46 69e103Jolicouer P 1963 The multivariate generalization of the allometry equation Bio-
metrics 19 497e499Jungers WL Falsetti AB Wall CE 1995 Shape relative size and size-
adjustments in morphometrics Am J Phys Anthropol 38 (S2) 137e161Kavanagh KD Evans AR Jernvall J 2007 Predicting evolutionary patterns of
mammalian teeth from development Nature 449 427e433Kelley J 1986 Species recognition and sexual dimorphism in Proconsul and
Rangwapithecus J Hum Evol 15 461e495Kelley J 1995 Sex determination in Miocene catarrhine primates Am J Phys
Anthropol 96 390e417Kelley J 1993 Taxonomic implications of sexual dimorphism in Lufengpithecus In
Kimbel WH Martin LB (Eds) Species Species Concepts and Primate Evo-lution Springer New York pp 429e458
Kent PE 1942 The country round the Kavirondo Gulf of Victoria Nyanza Geogr J100 22e31
Kordos L Begun DR 2001 A new cranium of Dryopithecus from RudabanyaHungary J Hum Evol 15 689e700
Le Gros Clark WE Leakey LSB 1950 Diagnoses of East African Miocene Homi-noidea Q J Geol Soc Lond 105 260e263
Le Gros Clark WE Leakey LSB 1951 The Miocene Hominoidea of East Africa InFossil Mammals of Africa No 1 British Museum (Natural History) London
Leakey LSB 1943 A Miocene anthropoid mandible from Rusinga Kenya Nature152 319e320
Leakey LSB 1963 East African fossil Hominoidea and the classification within thissuper-family In Washburn SL (Ed) Classification and Human EvolutionAldine Chicago pp 32e49
Leakey LSB 1967 Notes on the mammalian faunas from the Miocene and Pleis-tocene of East Africa In Bishop WW Clark JD (Eds) Background to Evolu-tion in Africa University of Chicago Press Chicago pp 7e29
Leakey RE Leakey MG 1986 A new Miocene hominoid from Kenya Nature 324143e146
Leakey RE Leakey MG Walker AC 1988 Morphology of Afropithecus turka-nensis from Kenya Am J Phys Anthropol 76 289e307
Leakey MG Ungar PS Walker A 1995 A new genus of large primate from theLate Oligocene of Lothidok Turkana District Kenya J Hum Evol 28519e531
Linnaeus C 1758 Systema naturae per regna tria naturae secundum classesordines genera species cum characteribus differentris synonymis locis tenthed Laurentii Salvii Stockholm
MacInnes DG 1943 Notes on the East African Miocene primates J E Afr UgandaNat Hist Soc 17 141e181
MacLatchy L 2004 The oldest ape Evol Anthropol 13 90e103MacLatchy L Bossert WH 1996 An analysis of the articular surface distribution of
the femoral head and acetabulum in anthropoids with implications for hipfunction in Miocene hominoids J Hum Evol 31 425e453
MacLatchy L Rossie JB 2005 The Napak hominoid still Proconsul major InLieberman DE Smith RJ Kelley J (Eds) Interpreting the Past Essays onHuman Primate and Mammal Evolution in Honor of David Pilbeam Brill Ac-ademic Publishers Boston pp 15e28
Martin L 1981 New specimens of Proconsul from Koru Kenya J Hum Evol 10139e150
McCollum MS Peppe DJ McNulty KP Dunsworth HM Harcourt-Smith WEH Andrews AL 2013 Magnetostratigraphy of the early MioceneHiwegi Formation (Rusinga Island Lake Victoria Kenya) Geological Society ofAmerica Abstracts with Programs 45 p 12
McNulty KP 2010 Apes and tricksters the evolution and diversification ofhumans closest relatives Evol Edu Outreach 3 322e332
McNulty KP Harcourt-Smith WEH Dunsworth HM 2007 New primate fossilsfrom Rusinga Island Kenya Am J Phys Anthropol 132 (S44) 170
McNulty KP MacLatchy L Peppe DJ Nengo I Manthi FK Miller ERStevens NJ Cote S Lehmann T Kingston J 2014a A regional approach toEast African early Miocene paleobiology J Vert Paleontol 185 SVP Programand Abstracts Book 2014
McNulty KP MacLatchy L Rossie JB Peppe DJ Deino AL Mbua ENManthi FK Nengo IO Stevens NJ Cote S Lehmann T Gutierrez M 2014b
Research on East African catarrhine and hominoid evolution results from thefirst year Am J Phys Anthropol 153 (S58) 182
Michel LA Peppe DJ Lutz JA Driese SG Dunsworth HM Harcourt-Smith WEH Horner W Lehmann T Nightengale S McNulty KP 2014Remnants of an ancient forest provide ecological context for Early Miocenefossil apes Nature Comm 5 3236
Myres JL 1932 Africa Man 32 208Nakatsukasa M Kunimatsu Y Nakano Y Takano T Ishida H 2007 Vertebral
morphology of Nacholapithecus kerioi based on KNM-BG 35250 J Hum Evol52 347e369
Nengo IO Rae TC 1992 New hominoid fossils from the early Miocene site ofSonghor Kenya J Hum Evol 23 423e429
Peppe DJ McNulty KP Cote SM Harcourt-Smith WEH Dunsworth HM VanCouvering JA 2009 Stratigraphic interpretation of the Kulu Formation (EarlyMiocene Rusinga Island Kenya) and its implications for primate evolutionJ Hum Evol 56 447e461
Pickford M 1981 Preliminary Miocene mammalian biostratigraphy for westernKenya J Hum Evol 10 73e97
Pickford M 1986 Sexual dimorphism in Proconsul Hum Evol 1 111e148Pickford M Andrews PJ 1981 The Tinderet Miocene sequence in Kenya J Hum
Evol 10 11e33Pickford M Kunimatsu Y 2005 Catarrhines from the Middle Miocene (ca 145
Ma) of Kipsaraman Tugen Hills Kenya Anthropol Sci 113 189e224Pickford M Senut B Gommery D Musiime E 2009 Distinctiveness of Ugan-
dapithecus from Proconsul Estudios Geol 65 183e241Pilbeam DR 1969 Tertiary Pongidae of East Africa evolutionary relationships and
taxonomy Bulletin of the Peabody Museum of Natural History 31 pp 1e185Rae TC 1993 Phylogenetic analysis of proconsulid facial morphology PhD
Dissertation State University of New York at Stony BrookRae TC 1999 Mosaic evolution in the origin of the Hominoidea Folia Primatol 70
125e135Rafferty KL Walker A Ruff C Rose MD Andrews PJ 1995 Postcranial esti-
mates of body weights in Proconsul with a note on a distal tibia of P major fromNapak Uganda Am J Phys Anthropol 97 391e402
Rossie JB MacLatchy L 2013 Dentognathic remains of an Afropithecus individualfrom Kalodirr Kenya J Hum Evol 65 199e208
Senut B Pickford M Gommery D Kunimatsu Y 2000 Un nouveau genredrsquohominoiumlde du Miocene inferieur drsquoAfrique orientale Ugandapithecus major(Le Gros Clark and Leakey 1950) CR Acad Sci Paris 331 227e233
Simons EL Pilbeam DR 1965 Preliminary revision of the Dryopithecinae (Pon-gidae Anthropoidea) Folia Primatol 3 81e152
Smith TM Martin LB Leakey MG 2003 Enamel thickness microstructure anddevelopment in Afropithecus turkanensis J Hum Evol 44 283e306
Stevens NJ Seiffert ER OConnor PM Roberts EM Schmitz MD Krause CGorscak E Ngasala S Hieronymus TL Temu J 2013 Palaeontological evi-dence for an Oligocene divergence between Old World monkeys and apesNature 497 611e614
Suwa G Kono RT Katoh S Asfaw B Beyene Y 2007 A new species of great apefrom the late Miocene epoch in Ethiopia Nature 448 921e924
Szalay FS Delson E 1979 Evolutionary History of the Primates Academic PressNew York
Teaford MF Beard KC Leakey RE Walker A 1988 New hominoid facial skel-eton from the Early Miocene of Rusinga Island Kenya and its bearing on therelationship between Proconsul nyanzae and Proconsul africanus J Hum Evol17 461e477
Van Couvering JAH Miller J 1969 Miocene stratigraphy and age determinationsRusinga Island Kenya Nature 221 628e632
Walker A 1997 Proconsul function and phylogeny In Begun DR Ward CVRose MD (Eds) Function Phylogeny and Fossils Miocene Hominoid Evolu-tion and Adaptation Plenum Press New York pp 209e224
Walker A Pickford M 1983 New postcranial fossils of Proconsul africanus andProconsul nyanzae In Ciochon RL Corruccini RS (Eds) New Interpretationsof Ape and Human Ancestry Plenum Press New York pp 325e351
Walker A Teaford M 1988 The Kaswanga primate site an Early Miocene hom-inoid site on Rusinga Island Kenya J Hum Evol 17 539e544
Walker A Teaford M 1989 The hunt for Proconsul Sci Am 260 76e82Walker A Teaford MF Martin L Andrews P 1993 A new species of Proconsul
from the early Miocene of RusingaMfangano Islands Kenya J Hum Evol 2543e56
Ward CV Ruff CB Walker A Rose MD Teaford MF Nengo IO 1995 Func-tional morphology of Proconsul patellas from Rusinga Island Kenya with im-plications for other Miocene-Pliocene catarrhines J Hum Evol 29 1e19
Ward S Brown B Hill A Kelley J Downs W 1999 Equatorius a new hominoidgenus from the Middle Miocene of Kenya Science 285 1382e1386
Werdelin L Sanders WJ 2010 Cenozoic Mammals of Africa University of Cali-fornia Press Berkeley

TabDen
Fe
G
U
U
U
U
U
M
Lo
Lo
Lo
a
b
c
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 47
Genus Proconsul Hopwood 1933
Type species
Proconsul africanus Hopwood 1933
Included species
Proconsul africanus (Hopwood 1933) Proconsul major (Le GrosClark and Leakey 1950) Proconsul gitongai comb nov (Pickfordand Kunimatsu 2005) Proconsul meswae (Harrison and Andrews2009)
Diagnosis (replaces the emended diagnosis of Harrison 2010)
Proconsul species are medium- to large-bodied sexually dimor-phic catarrhines that can be distinguished from other proconsulidgenera by the following combination of features Robust mandib-ular corpus and symphysis characterized by a long shallowlysloping planum alveolare terminating at a large superior transversetorus and a subplanum surface oriented posteroinferiorly Upper
le 2tognathic features that distinguish Ekembo from Proconsula
ature Ekembo
eneralCusp morphology More bunodont occupying more of theMolar crests Inflated incorporated into cuspsCingulum Reducedpper central incisorLingual pillar Narrow generally low reliefMarginal ridges and cingulum Asymmetric with sharply angled cingulu
ridge transitionpper canineTip PointRoot fluting AbsentRoot striations Rare and weakly developedpper third premolarb
Mesiodistal cusp position Crown midpointMesial fovea Well developedBuccal profile SymmetricMesial crown margin Straight or slightly curvedDistal crown margin Slight to moderately curvedCusp heteromorphy ReducedCrown shape Converges on P4 shapeMesiobuccal flange Nopper fourth premolarParacone height Taller relative to crown lengthpper second molarc
Occlusal crown shape SquarishDistal margin Straight to slightly curvedBuccal cingulum Absent or rudimentaryDistolingual cingulum Merges into hypoconePosthypocone crista Distobuccally to buccally orientedBuccolingual flare ReducedandiblePlanum alveolare Short vertically inclinedSuperior transverse torus Moderate to well-developedOrientation of subplanum area More vertically orientedCorpus robusticity More gracilewer incisorsCrown shape Narrow relative to heightwer canineTip Pointwer molarsBuccolingual flare ReducedM1 M2 breadth M1 narrower relative to M2
Although presented here as discrete characteristics for descriptive purposes some oNearly all of these features tend to distinguish P4s as well but in that tooth the diffeThese features tend to distinguish M1s as well but in that tooth the differences are
incisors heteromorphic with I1 exhibiting an inflated lingual tu-bercle and a near symmetrical configuration of lingual marginalridges and cingulum with a curving transition from cingulum tothe mesial marginal ridge Lower incisors broad relative to crownheight Upper and lower canines with distinct blade-like or burin-like tips in both deciduous and adult teeth Upper canine rootswith broad longitudinal grooves or fluting defining two or threedistinct columns plus numerous small longitudinal striationsextending cervically from the root apex Upper P3s morphologicallydissimilar from P4s P3 has substantial cusp heteromorphy (para-cone much taller than protocone) with mesially positioned cuspsand a notched mesial border that together limit or eliminate amesial fovea A long steeply angled preparacrista contributes to ahighly asymmetric buccal profile before being interrupted near thetooth cervix by a mesially projecting flange The distal margin isstrongly curved P4 has a tall paracone relative to crown lengthMolars characterized by extensive cingular development individ-uated conical cusps connected by sharp occlusal crests and sub-stantial buccolingual flare Upper molars rhomboid in shaperesulting from a broad trigon and a distally expanded hypocone M2
has a distinct buccal shelf between paracone and metacone a dis-tolingual cingulum that wraps fully around the hypocone rather
Proconsul
occlusal surface area More distinct individuated conesSharper more distinct from cuspsExtensive
Inflatedbulbousm-mesial More symmetric with curving cingulum-mesial
ridge transition
Blade-likePresentVery common and well developed
MesialAbsent or reducedAsymmetricNotchedStrongly curvedStrongDistinct from P4
Yes
Shorter relative to crown length
RhomboidStrongly curvedDistinct cingular shelfDistinct from hypoconeDistally to distobuccally orientedPronounced
Long more horizontally orientedStrongly developedMore posteriorly inclinedMore robust
Broad relative to height
Blade-like
PronouncedM1 more similar to M2
f these features are undoubtedly correlated aspects of single evolutionary changesrences are not as pronouncednot as pronounced
Figure 2 Upper central incisors of Proconsul and Ekembo Lingual views of Ekembo(KNM-RU 1685 cast) left and Proconsul (KNM-CA 1300) right Note the differences inthe transition between the lingual cingulum and the mesial marginal ridge which ismore rounded in Proconsul and more angled in Ekembo The central pillar is also morebulbous or inflated and without ridging in Proconsul Additional specimens illustratingthe Proconsul morphology are shown in Pickford et al (2009 Fig 17 p 197) butreferred in that manuscript to Ugandapithecus
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6148
thanmerging into the hypocone on its lingual surface and a distallyoriented posthypocone crista Lower first and second molars nearlyequal in buccolingual breadth
Comparative morphology of Ekembo and Proconsul
Samples and comparative methods
The above diagnoses are based on extensive survey and analysisof the original fossil material Specimens housed in the NationalMuseums of Kenya (NMK) and the Natural History Museum (Lon-don) were analyzed by the authors during several separate and twojoint (NMK) research visits Fossils in the Uganda National Museumwere assessed by one of us (KPM) during two visits All measure-ments were collected by KPM Dental terminology follows Szalayand Delson (1979)
The following results are organized with respect to themorphology of the Kisingiri sample (attributed to E nyanzae andE heseloni) versus that of Proconsul from Tinderet and Napak(traditionally referred to P africanus P major and more recentlyP meswae) Specimens from additional localities that have at onetime been attributed to Proconsul (eg Andrews1978 Pickford andKunimatsu 2005 [cf Harrison 2010]) are for the most part notconsidered here most are middle Miocene in age and have beentransferred to other genera (Harrison 2002 2010) Of those that arefrom early Miocene sites there is growing evidence that P majorcan also be found at Moroto and differentiated fromMorotopithecus(Pickford et al 2009) an issue of some interest but ultimately notcritical to the taxonomic questions being considered here Whereappropriate however we do provide observations on a few earlyMiocene specimens attributed to ldquoUrdquo gitongai (Pickford andKunimatsu 2005 Pickford et al 2009 see Harrison 2010) Yetwhile we consider the middle Miocene holotype likely to representa distinct species here referred to Proconsul we concur withHarrison (2010) that many specimens placed in this species byPickford et al (2009) cannot be distinguished from P major
Sample sizes particularly from Tinderet and Ugandan localitiestend to be small but where possible standard univariate (eg t-tests) and bivariate (regression ANCOVA) statistical analyses wereperformed A single eigendecomposition of the covariance matrixof symphyseal measurements was done to characterize the multi-variate shape of the mandibular symphysis after accounting for sizedifferences (sensu Jolicouer 1963) Quantitative results are illus-trated as box-and-whisker plots of shape indices or bivariate plotsof the measurements For samples of fewer than five specimensshape index values are plotted in lieu of box-and-whiskersummaries
Morphological comparisons
Upper incisors Two features of the Ekembo upper central incisorsdistinguish them from those of Proconsul including specimenswidely attributed to the type species P africanus The lingual tu-bercle in Ekembo is quite narrow generallywith low relief and oftenscored by two or more irregular ridges in contrast to the muchmore inflated or bulbous tubercle in Proconsul I1s (Fig 2) Ekembospecimens also have a characteristic morphology in the transitionfrom the lingual cingulum to the mesial marginal ridge (Fig 2)from the mesiodistal mid-point of the crown the cingulumextends either directly mesially or even angles slightly cervicallyto intersect the mesial marginal ridge at a fairly sharp angle Incontrast in I1s of Proconsul the cingulum angles incisally from themid-point and transitions into the mesial marginal ridge in agradual curve The expression of these traits varies somewhat inthe smaller Tinderet specimens attributed to P africanus
compared with those of P major (cf Pickford et al 2009)However these variations are relatively minor with respect to thedifferences between the two genera Of 25 I1s known fromKisingiri localities only one specimen KNM-MW 562 somewhatresembles the Tinderet sample in both of these featuresdhavinga somewhat less angled transition from the mesial marginal ridgeto the cingulum and a more bulbous lingual tubercle This isparticularly interesting as the specimen was discovered in theMakira Beds now dated to older than 190 Ma and contemporarywith the Tinderet localities (McCollum et al 2013) Neverthelessits overall morphology is more similar to other Ekembospecimens than to Proconsul incisors Two other Ekembospecimens KNM-RU 1677 and RU 1933 have somewhat moreinflated lingual tubercles but otherwise resemble other Ekembo I1s
No consistent differences were found in upper lateral incisors
Upper canines Upper canine crowns of Ekembo are typical of thoseof most catarrhines in terminating in an evenly tapering point Thisdiffers from the Proconsul specimens which have a blade-like(Kelley 1986 Senut et al 2000 Harrison 2010) or ldquoburin-likerdquo(Pickford et al 2009) canine tip (Fig 3) While blade-like apicalmorphology has long been recognized as a distinctivecharacteristic of P major we confirm the report by Pickford et al(2009) that this feature is also present in the P africanus typespecimen (Fig 3a) although somewhat understated as expectedin the more slender and higher-crowned P africanus caninesUnfortunately other canines attributed to P africanus (eg M44837 KNM SO 419 LG 921 CA 1910 SO 5353 Kelley 1986) havebroken apices and therefore cannot be evaluated for this featureA blade-like tip is also present in the deciduous upper canines ofP major (eg KNM-SO 371 MV 10 NAP IV UMP 62-20) andP meswae (KNM ME 11) Hence the blade-like upper canine tipunites all Tinderet and Napak Proconsul species but is neverexpressed in Ekembo specimens from Rusinga and Mfangano
Two features of the canine roots also differentiate these generalongitudinal grooving or fluting along the length of the root (cfKelley 1986) and the development of numerous finer longitudinal
Figure 3 Canines of Proconsul and Ekembo a) Upper canine of the P africanus holotype (M 14084) lingual view b) Upper canine of P major (KNM-CA 2127) lingual view c) Lowercanine of P major buccal view d) lower canine of Ekembo cf nyanzae (KNM-RU 1676) buccal view and e) Upper canine of P major (KNM-SO 584) distal view Note the blade-like orldquoburin-likerdquo tip of the unworn Proconsul canines including on the nearly unworn canine tip of the P africanus type specimen (a) Ekembo canines have more typical pointed tipsProconsul canines also have longitudinal grooves and fine striations on the root (e) Ekembo lacks these features Scale bars frac14 1 cm
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 49
striations near the root apex that extend to varying degrees towardthe cervix of the tooth Longitudinal ldquoflutingrdquo consists of broadgrooves that segment the root into distinct sections (cf Pickfordet al 2009 Fig 3e) Most often the root in cross-section has theappearance of a cylinder that has been sectioned longitudinallywith the two halves offset from each other but in some cases thereare three segments and the root cross-section is more triangular inappearance (Kelley 1986) This characteristic was noted by Kelley(1986) in the P africanus canines but not in those of P majorHere we affirm that fluting is present on all of the visible uppercanine roots attributed to both P africanus and P major rootmorphology could not be evaluated in the deciduous upper caninefrom Meswa Bridge since it is implanted in the maxillary alveolusNone of the Ekembo specimens display this morphology havinginstead a simpler round to ovoid cross-section along the length ofthe root
The fine longitudinal striations on Proconsul upper canine rootsvary in the degree to which they extend toward the cervix (Fig 3e)but they are visible in all specimens attributed to P africanus(KNM-SO 419 521 921 5353 KNM-X 502) and five of six uppercanines of P major (excepting KNM-SO 1614) This morphology isalso present in UMP 62-12 which was included as part of theMorotopithecus bishopi hypodigm (Gebo et al 1997) Fine striationsalong the upper canine root are almost entirely absent in EkemboTwenty of twenty-three specimens lack this feature three speci-mens (KNM-RU 1763 1845 1891) exhibit incipient to slight
striations but these are never developed to the extent seen inProconsul canines
Upper premolars Upper third premolars display some of thegreatest differences between Ekembo and Proconsul P4s tend todiffer in similar ways but the differences are not nearly as markedEkembo P3s have cusps positioned near the mesiodistal midpoint ofthe crown well-developed mesial foveae pre- and postparacristaeof similar length and slope resulting in symmetric or nearsymmetric buccal profiles a straight or slightly curved mesialmargin and only moderately curved distal margin (Fig 4)Proconsul P3s are more similar in appearance to those of small-bodied catarrhines from the early Miocene with cusps positionedmesially on the crown greatly reducing or eliminating the mesialfovea They also have a steep preparacrista and more shallowlysloped postparacrista resulting in a highly asymmetric buccalprofile and an inwardly angled or notched mesial margin but astrongly curved distal margin This morphology is bestcharacterized in the P africanus holotype (M 14084) but can alsobe seen in other specimens attributed to this species as well asthose referred to P major (Fig 4d e) Also noteworthy is adistinctive mesiobuccal flange that mesially extends the buccalportion of the crown but only near the cervical line of the toothPickford et al (2009) describe this feature in some detail forldquoUgandapithecusrdquo but in fact this morphology is clearly expressedin the P africanus holotype as well (Fig 4a d) A single adult P3
Figure 4 Upper premolars of Proconsul and Ekembo occlusal (aec) and lingual (def) views a) P3e4 of the P africanus holotype (M 14084) b) P3 of P major (M 14331) c) P3e4 of theE heseloni holotype (KNM-RU 2036) d) P3 of the P africanus holotype (M 14084) e) P3 of P major (M 14331) f) P3 of the E heseloni holotype (KNM-RU 2036) Note the mesiallypositioned cusps lack of mesial fovea and strongly asymmetric crown in Proconsul compared to Ekembo (see text for further comparisons) Image of M 14331 reversed for easiercomparison Photo 4a courtesy of Rutger Jansma Scale bars frac14 1 cm
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6150
germ and two dP3s attributed to P meswae look remarkably similarto those of other Proconsul species including the presence of thisflange In contrast the Ekembo P3 has a well-developed mesialfovea that terminates at a mesiobuccal shoulder positionedapproximately halfway between the paracone tip and the cervixFrom this shoulder the mesial margin angles slightly distallydown to the cervix (Fig 4f)
The degree of cusp height heteromorphy in P3 also differentiatesEkembo from Proconsul (contra Pickford et al 2009) Only four non-Kisingiri specimens were sufficiently unworn to take measure-ments but more worn specimens also appear to express this dif-ferential in cusp height Figure 5a shows the ratio of protoconeparacone height measured from the tooth cervix and with only oneexception Ekembo P3s have substantially less cusp heteromorphyThis exception KNM-RU 1677 does not in fact have greater heter-omorphy but instead has an abnormal extension of the enamel ontothe buccal (paracone) root which skews the measurement Hencecusp relief relative to the occlusal surface in RU 1677 is not nearly asheteromoprhic as in Proconsul and when this is taken into accountthe difference between genera is significant (t-test dffrac14 11 t frac14 283p frac14 00165) The adult P3 germ from Meswa Bridge is incompletelyformed and therefore could not be measured However the differ-ence between paracone and protocone heights with respect to theocclusal surface appears to be as great as that in the P africanusholotype It does not appear that sex is an important determinant ofthis feature since well-established male (eg M16647 KNM-RU16000) and female (KNM-RU 2036 7290) specimens do not segre-gate across the Kisingiri range of variation
Upper fourth premolars of Ekembo are broadly similar inmorphology to Ekembo P3s in fact the similarity between P3 and P4
is substantially greater in Ekembo than in Proconsul (Figs 4a c)Likewise most of the morphological differences that discriminatethe P3s of these genera also tend to differentiate the P4s but less
clearly Cusp heteromorphy does not distinguish P4s of thesegenera but the overall height of the P4 paracone in Ekembo issignificantly greater relative to crown length than in Proconsul P4s(t-test df frac14 14 t frac14 316 p frac14 00069 Fig 5b)
Upper molars Cusps of Ekembo upper molars are uniformly morebunodont than those of Proconsul occupying more of the occlusalsurface Intercuspal crests are more fully incorporated into thecusps in Ekembo molars giving them a more inflated appearanceThis is particularly evident in the crista obliqua which is very broadin Ekembo but thin and knife-like in Proconsul (Fig 6) Cusps ofProconsul molars stand more as individual cones Pickford et al(2009) have further differentiated the cusp morphology ofsmaller and larger Tinderet specimens arguing that the latterhave more bulbous cusps While accurate this difference is minorcompared with the morphological difference between Ekemboand Proconsul and we regard the within-Proconsul cuspalvariation as size-related In fact the intermediately sizedP meswae has cusp morphology that is appropriatelyintermediate between that of P africanus and the larger P major
Other differences between Ekembo and Proconsul upper molarsare most evident in M2 We concur with previous assessments thatthe Kisingiri upper molars have less developed cingula than non-Kisingiri molars (eg Senut et al 2000 Harrison 2002 2010Harrison and Andrews 2009) More specifically Ekembo M2s lackor have only rudimentary buccal cingula between the paracone andmetacone Likewise the lingual cingulum is discontinuous merg-ing into the hypocone lingually and then re-emerging as a distalmarginal ridge (Fig 6) Proconsul M2s display substantially morecingular development overall and in particular exhibit a prominentbuccal shelf between the paracone and metacone and a distinctdistolingual cingulum that is continuous around the hypocone towhere it merges with the distal marginal ridge
Figure 5 Box-and-whisker plots a) P3 cusp heteromorphy index (protocone heightparacone height) Results of a t-test show this difference to be significant (p frac14 00165) Theexcluded Ekembo outlier KNM-RU 1677 only appears to have extreme heteromorphy because of extensive expansion of the buccal enamel onto the anterior root (see text) b) P4
relative paracone height (paracone heightmesiodistal crown length) Results of a t-test show this difference to be significant (p frac14 00069)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 51
The occlusal profiles of Ekembo M2s are squarish in shape withrelatively straight and equal sides with a posthypocone crista thatis distobucally to buccally directed In contrast the occlusal profilesof Proconsul M2s have a more rhomboid shape with substantialbasal flare around the mesial cusps and a distally expanded hypo-cone with a distally directed posthypocone crista resulting in astrongly curved distal margin (Fig 6) Even in specimens with lesscurved distal margins the posthypocone crista is still more distallydirected compared to the condition in Ekembo These features arecharacteristic not only of P africanus P meswae and P major butare also expressed in the isolatedM2 fromMoroto Uganda (MOR IIb2098) attributed by Pickford et al (2009) to ldquoUrdquo gitongai butregarded here as belonging to P major
As noted by others (eg Harrison and Andrews 2009) the de-gree of buccolingual molar flare differs among these samplesQuantified as the distance between the tips of the mesial cuspsdivided by maximum crown breadth the degree of molar flare in
Ekembo M2s is significantly less than in Proconsul M2s (t-testdf frac14 29 t frac14 396 p frac14 00004) In a bivariate plot of these twomeasures the distributions of Ekembo and Proconsul teeth barelyoverlap with only KNM-RU 1677 encroaching into Proconsul ter-ritory (Fig 7)
These differences in M2 morphology also tend to distinguishEkembo from Proconsul M1s but not always as clearly Whereas M2smore consistently express distinct morphotypes the M1s in bothgenera show more variation in these features
The degree of upper third molar development (ie whether M3
has four distinct cusps or the distal cusps are reduced or absent) haslong been cited as taxonomically informative for these species orgroups of these species (eg Hopwood 1933 Le Gros Clark andLeakey 1951 Andrews 1978 Pickford et al 2009) However wefound this tooth to be unreliable for taxonomic purposes Withinprimates M3s are typically the most variable molars in both sizeand shape (Blankenship et al 2007 Kavanagh et al 2007) and this
Figure 6 Occlusal view of the M2s of Proconsul and Ekembo Left P africanus holotype(M 14084) Right E heseloni holotype (KNM-RU 2036) Note in Proconsul the overalldifference in crown shape as well as the more conical individuated cusps and nar-rower cristae strongly developed lingual cingulum that continues to the distal aspectof the crown broad buccal cingular shelf and more distally directed posthypoconecrista In Ekembo the cusps are more bunodont and expanded the cristae are broaderwith better developed accessory tubercles the lingual cingulum merges into thehypocone there is no buccal shelf and the posthypocone crista is more buccallydirected
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6152
is observable even in small samples of Proconsul species TheP africanus holotype has a substantially reduced M3 whereas inKNM-CA 2250dattributed by most authors to this species(Andrews 1978 Pickford et al 2009)dit is fully developed Simi-larly the hypodigm of P major includes both fully developed M3s(eg KNM-CA 389 LG 1815) and specimens with reduced ormissing hypocones (KNM-CA 397 1299) Ekembo displays compa-rable ranges of variation from highly reduced to fully developedM3s in both species Thus these species and genera cannot bedistinguished based on M3 development and we did not find anyother distinguishing characteristics between Ekembo and Proconsulin the upper third molars
Figure 7 Scatter plot depicting the de
Mandible Pickford et al (2009) enumerated several traits thatreadily distinguish Kisingiri from non-Kisingiri mandibles andthese distinctions are entirely consistent with the distinction ofEkembo from Proconsul Our analyses specifically underscoredifferences in the symphyseal region The planum alveolare inboth male and female specimens of Ekembo is short and morevertically oriented even the most elongate of these (KNM-RU47805 McNulty et al 2007) is substantially more verticalcompared to specimens of Proconsul In contrast the planumalveolare in Proconsul is conspicuously long and nearlyhorizontally oriented Well-preserved mandibles attributed toP major (eg KNM-SO 396 and 404) exhibit this morphology inextreme fashion (Fig 8) smaller specimens (KNM-SO 1112P africanus) show less extreme expression but are still readilydistinguishable from Ekembo mandibles For its size KNM-LG 452has a surprisingly short planum alveolare though not to thedegree seen in Rusinga mandibles Correlated to the shortplanum alveolare in Ekembo is a reduced superior transversetorus which while well-developed does not approach therobusticity expressed in Proconsul mandibles This difference isreflected in an index of symphyseal robusticity (thicknessperpendicular to the long axis of the symphysislength of thelong axis) which shows Ekembo and Proconsul to be significantlydifferent (t-test df frac14 14 t frac14 430 p frac14 00007)
Development of the inferior transverse torusdoften used todifferentiate among species of Proconsul (eg Pickford et al 2009Harrison 2010)dwas found to be highly variable and inconsis-tently expressed within these samples However the overallorientation of the subplanum region (as defined by a line con-necting the posterior-most midline points of the superior andinferior transverse tori or the posterior-most midline point of theinferior margin of the symphysis in the absence of the latter) doesdistinguish Ekembo from Proconsul mandibles The orientation ofthis line in relation to the alveolar margin in the vicinity of thepostcanine teeth was assessed qualitatively by placing a small rod
gree of buccolingual flare in M2
Figure 8 Mandibles of Proconsul and Ekembo Occlusal views of P major a) KNM-SO 396 b) KNM-SO 404 c) KNM-LG 452 Occlusal views of Ekembo d) KNM-RU 2087 e) KNM-RU1674 f) KNM-RU 7290 Occlusal views of P africanus g) KNM-SO 372 h) KNM-SO 1112 i) Lingual view of P africanus (KNM-SO 1112) Note the long more horizontally orientedplanum alveolare in Proconsul resulting in a pronounced superior transverse torus The planum alveolare is shorter and more vertical in Ekembo such that one can see the inferiormandibular border from occlusal view
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 53
(eg a dental pick) in themidline internally along the two referencepoints and recording the point in lingual perspective at which therod intersected the alveolar margin (Fig 9) In Ekembo the sub-planum region is more vertically oriented so that the rod intersectsthe alveolar margin most often in the premolar field Not surpris-ingly there is a difference between sexes with females having areduced superior transverse torus and therefore a more verticallyoriented subplanum region compared to males In contrast withEkembo Proconsul mandibles have much more inclined subplanumsurfaces with the rod intersecting the lingual alveolar margin inthe vicinity of the M1 (females) or M2 (males Fig 9) When sex istaken into account therefore the distributions of Ekembo andProconsul in this feature are discrete Note that male and femalemandibles attributed to both P africanus and P major are includedin this assessment and are surprisingly similar despite dramaticsize differences
We also quantified internal symphyseal morphology using threemeasures symphyseal height (infradentaleegnathion) planumalveolare length (posterior-most point on the superior transversetoruseinfradentale) and subplanum length (posterior-most pointon the superior transverse torusegnathion) a combination thatincorporates not only the lengths of these intervals but also theangular relationships among them To compare symphyseal shapeacross a range of sizes we chose a simple principal componentsapproach to partition size onto the first eigenvector (cf Jolicouer1963) While this is not the preferred method for partitioning sizein most datasets (Jungers et al 1995) for this particular sample andset of measurements Mosimann shape variables were still
significantly correlatedwith size (see discussion in Coleman 2008)Hence we assessed differences in symphyseal morphology be-tween Ekembo and Proconsul using the second and third principalcomponents of the three variables Distributions of the two generaare distinct in this shape space (Fig 10 Supplementary OnlineMaterial [SOM] Table 1)
Robusticity of the mandibular corpus also differs between thesesamples though it is more difficult to test due to differentialpreservation among specimens Corpus thickness and depth underP4 can be measured in the greatest number of specimens andcomparison of these measures suggests a difference betweengroups in the scaling of corpus robusticity (Fig 11) Among largepresumably male specimens Ekembo mandibles are substantiallyless robust than those of Proconsul However among smallermandibles only a single Proconsul specimen (KNM-SO 372) could beincluded and it is similar in robusticity to Ekembomandibles of thissize Considered together the impression is that corpus robusticitymay scale differently in Ekembo than it does in Proconsul and a testfor differences in slopes using an ANCOVA design of the loggedvariables was significant (df frac14 2 F frac14 2481 p lt 00001) Never-theless additional specimens are needed to properly evaluate theinfluence of taxonomy sex and size on corpus robusticity areasonable alternative interpretation of Figure 11 is that it dem-onstrates a significant difference between P major and all otherspecies of both Proconsul and Ekembo
Lower incisors Ekembo specimens are uniformly narrower inmesiodistal length than incisors attributed to P africanus P major
Figure 10 Scatter plot of principal component scores based on an eigenanalysis of threeposterior point on the superior transverse toruseinfradentale) and subplanum length (mprimarily captures size differences among specimens (Jolicouer 1963 see text and SOM Trespectively) in the symphyseal variables
Figure 9 Line drawing of a hemimandible depicting different orientations of the sub-planum area in Ekembo and Proconsul Females are denoted using shorter vectors maleswith longer vectors Vector is estimated in M 14086 which is broken at the midlineEkembo specimens have a more vertical subplanum compared to those from Songhorand Legetet but within both groups females have more vertically oriented regions thanmales Hence even the most ventrally rotated subplanum in the largest Ekembo malespecimen (KNM-RU 47805) groups with the Tinderet females in this feature
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6154
and P meswae both absolutely and in relation to crown height(Fig 12) This is true for both I1 and I2 A t-test adjusted for non-independent observations (eg I1 and I2 from the same mandible)confirmed the difference as significant (t-test df frac14 12 t frac14 258p frac14 00242) Moreover while the measurable Proconsul samplewas small this same pattern is observable in other specimenswhose tips were slightly to moderately worn and therefore notmeasured
Lower canines The only lower canine trait that discretely sepa-rates these genera is the pointed crown tip in Ekembo (Fig 3d)contrasted with a blade-like (or ldquoburinrdquo) tip in Proconsul(Fig 3c) This feature is well established for P major but is alsoexpressed in KNM-CA 1772 which was referred to P africanus byKelley (1986) and is too small to be either P major or P meswaeTwo other specimens commonly attributed to P africanus KNM-CA 2149 and KNM-SO 1112 both have broken canine tipsLikewise a new specimen (Kor 65004) attributed to P africanusby Pickford et al (2009) also has a broken tip and cannot beevaluated (see Pickford et al 2009 Fig 23) As with the uppercanines the blade-like tip is also present in the deciduous lowercanines of P major (eg KNM-CA 361 S0 5352) and P meswae(KNM-ME 1)
Lower premolars Like most previous authors (eg Andrews 1978Harrison 2002 2010 Pickford et al 2009) we found neitherqualitative nor quantitative characteristics of the lower premolarswith any taxonomic value
Lower molars Differences in lower molar morphology are few andrelatively subtle but in some cases mirror those seen in the uppermolars Cusps on the lower molars of Ekembo are more bunodontthan those in Proconsul and occupy more of the occlusal surfaceBroad inter-cuspal crests are incorporated into the Ekembo cuspsmaking them appear more inflated Molar cusps of Proconsul are
variables symphyseal height (infradentaleegnathion) planum alveolare length (mostost posterior point on the superior transverse torusegnathion) The first componentable 1) and thus PC 2 and PC 3 illustrate the residual shape variance (72 and 28
Figure 11 Scatter plot showing mandibular corpus robusticity under P4 Trend lines were computed using ordinary least squares (OLS) regression Similar relationships areobservable at other parts of the corpus but differential preservation results in smaller samples at those positions Slopes are significantly different (p lt 00001 tested on loggedvariables)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 55
more distinct from one another and are linked by narrower sharperinter-cuspal crests The lower molars of Ekembo also have asubstantially less developed buccal cingulum than those ofProconsul (Harrison 2002 2010 Harrison and Andrews 2009)
Figure 12 Scatter plot of lower incisor crown proportions Differenc
Lastly and again as seen in the upper molars the two genera canbe distinguished by the significantly greater amount of lowermolar flare in Proconsul (Fig 13 t-test df frac14 59 t frac14 713p lt 00001 see also Harrison and Andrews 2009)
es between Ekembo and Proconsul are significant (p frac14 00242)
Figure 13 Box-and-whisker plot of buccolingual flare in lower molars measured asthe average distance between the mesial buccal and lingual cusps divided by overallbuccolingual crown breadth Differences between Ekembo and Proconsul are highlysignificant (p lt 00001)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6156
Relative molar size has been used to distinguish species ofProconsul but we observed the primary difference to be in relativebuccolingual breadths of M1 and M2 In Ekembo M1 is substantiallynarrower than M2 than is the case in Proconsul (Fig 14 Harrison2002 2010) Although none of the smaller Proconsul mandiblespreserve complete crowns for both M1 and M2 KNM-SO 1112 hasan intact M2 and at least preserves the cervix for M1 However evenif the cervical measurement is used for its M1 breadthdthusunderestimating the true crown breadthdKNM-SO 1112 still has abroader first molar in relation to M2 than would be expected inEkembo If crown breadth is estimated from the ratio of cervixbreadth to maximum crown breadth in other Proconsul fossils therange of likely breadths for the SO 1112 M1 clearly distinguishes it
from the Ekembo sample (Fig 14) Testing this relationship onlogged variables using the mean estimate for the SO 1112 M1(Fig 14) reveals a significant difference between the two groups(ANCOVA df frac14 1 F frac14 2424 p frac14 00002)
As with upper third molars we find a great deal of variation inM3 morphology and mistrust the taxonomic significance some-times placed on characteristics of this toothdat least with regard tothese taxa Harrison (2002 2010) cited the mesiodistal position ofthe M3 entoconid as differentiating Kisingiri third molars fromthose attributed to P africanus We concur with his assessment butfound the entoconid position to vary in specimens attributed toP major Thus this character does not differentiate Ekembo from allProconsul species but may distinguish it from the type speciesP africanus
Discussion
The distinctiveness of the Kisingiri large catarrhine materialfrom that at Tinderet and Ugandan localities has been noted pre-viously by several authors (Pickford 1986 Walker et al 1993Begun 2001 2007 MacLatchy and Rossie 2005 Harrison andAndrews 2009 Harrison 2010) What had not been fully docu-mented was the nearly complete lack of overlap between thesesamples in a number of features despite very large assemblagesfrom Rusinga and Mfangano On descriptive criteria alone (Table 2)many isolated teeth (I1 C1 P3 M2 C1) and all of the mandibles canbe assigned to either Proconsul or Ekembowith very little chance ofmisattribution That some of the most diagnostic features occur inthe upper dentitiondthe only anatomy preserved in theP africanus type specimendadds weight to the taxonomicarrangement proposed here The morphology of the upper teeth inall Proconsul species compellingly unites them into a single cladedistinct from E heseloni and E nyanzae This is reinforced bynumerous features of the mandible and mandibular dentition thatfurther distinguish these samples The number and variety of thesefeatures are such that Proconsul and Ekembo are more differentfrom one another than are many closely-related modern catarrhinegenera
Derived morphology of Ekembo and Proconsul
It is premature to formally assign primitive and derived status tothe individual features of Ekembo and Proconsul before completinga formal phylogenetic analysis This is especially true given theabundance of evidence available for Ekembo and hence the poten-tial impact that our generic revision could have on phylogeneticinterpretations Nevertheless in comparison with both extanthominoids and the small-bodied catarrhines from the earlyMiocene it is informative to differentiate the evolutionary grades ofthese genera Many characteristics of Ekembo are more advanced inthe direction of modern apes compared to those of Proconsul(Table 2) General trends such as expanded and more bunodontcusps broader molar crests reduced cingulum and morphologicalsimilarity between P3 and P4 all represent morphologies thatpresage those of later fossil and extant hominoids The moreadvanced morphologies of P3 and M2 are particularly conspicuouscompared not only with those of Proconsul but to most earlyMiocene catarrhines In contrast the mesially positioned stronglyheteromorphic cusps on the P3 of Proconsul plus the correspondingasymmetric buccal profile notched mesial margin and missingmesial fovea are characteristic of other early Miocene catarrhinessuch as Dendropithecus Rangwapithecus and Limnopithecus andare likely primitive among East African catarrhines The moresymmetric P3 of Ekembo with midline positioned cusps andreduced cusp heteromorphy is reminiscent of later hominoids and
Figure 14 Relative buccolingual dimensions of M1 and M2 KNM-SO 1112 has a preserved M2 but only the cervix of M1 Its M1 breadth was estimated based on the ratio to cervixbreadth to maximum crown breadth in other Tinderet fossils Even the cervical breadth of SO 1112 is broader than would be predicted for the crown breadth of a Kisingiri M1 Trendlines computed using OLS regression and using the average estimate of SO 1112 M1 breadth Group differences were significant (p frac14 00002)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 57
even hominid taxa such as Rudapithecus (eg Kordos and Begun2001)
Proconsul is likewise united by a suite of features that are for themost part characteristic only of this lineage (Table 2) Themorphology of both upper and lower canines is unique to thisgenus and shared by all three well-known species plus the poorlyrepresented middle Miocene P gitongai (Pickford and Kunimatsu2005) The mandibular symphysis is also distinctive in Proconsulcompared with other catarrhines with its very long and shallowlysloping planum alveolare and steeply inclined subplanum surfaceIndeed even the small edentulous anterior mandibular fragmentKNM-SO 372 is readily placed within Proconsul and excluded fromother similarly sized catarrhine taxa such as Rangwapithecus andEkembo based on this unique symphyseal morphology (Figs 8 and9)
Other differences between Ekembo and Proconsul are of uncer-tain evolutionary polarity (eg orientation of the posthypoconecrista) or may be of taxonomic relevance without broader phylo-genetic significance (eg degree of molar flare Table 2) Werecognize that there are morphological differences within Procon-sul particularly between the better known species P africanus andP major However many of these differences are likely related tosize and represent variations on a single generic theme that aremarkedly different from the morphology of Ekembo The recentdescription (Harrison and Andrews 2009) of the intermediatelysized P meswae underscores the broad morphological unity withinProconsul and further substantiates a well-defined Proconsul cladecomprising Tinderet and Ugandan species
Based on the abundant morphological and morphometric evi-dence supporting genus-level distinction between Proconsul andEkembo we are currently conducting a more detailed phylogeneticanalysis of all early Miocene catarrhines in order to better under-stand the distribution and evolutionary significance of these andother morphological features We anticipate that the debates sur-rounding the phylogenetic relationships of an historically consti-tuted sample of Proconsul can be more satisfactorily resolved nowthat the Kisingiri material is appropriately recognized as a distinctgenus
The generic disposition of ldquoUgandapithecusrdquo species
A great many features said to characterize ldquoUgandapithecusrdquowere added to the original diagnosis of the genus by Pickford et al(2009) in their systematic revision of Proconsul (sensu lato) As partof that study they also erected a new species ldquoUrdquo legetetensis andtransferred P meswae to ldquoUgandapithecusrdquo Evaluating the same setof features across the entire available sample however we wereunable to find support for this more expansive conception ofldquoUgandapithecusrdquo for several reasons First some diagnostic fea-tures cited in Pickford et al (2009) were quite variable within theldquoUgandapithecusrdquo sample As an example marked reduction of theM3 entoconid to a bead on the lingual cingulum is not shared byseveral ldquoUrdquo major specimens such as KNM-CA 393 NAP I 49000and Kor 253004 (see Pickford et al 2009 Fig 33e) and was likelynot a feature of a few of the more worn specimens eg BUMP 600and BUMP 601 or even the holotype of P major M 16648 Second
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6158
some diagnostic features of ldquoUgandapithecusrdquo were found to bevariably present in Ekembo arguing against their utility for dis-tinguishing ldquoUgandapithecusrdquo from Pickford et als (2009) com-bined P africanus thorn Kisingiri conception of Proconsul For examplesignificantly reduced curvature of the I1 distal margin in ldquoUganda-pithecusrdquo is also found in some Kisingiri specimens including KNM-RU 1685 1712 1714 1831 1846 and 1951 several more Rusingaspecimens have moderately reduced curvature Third some fea-tures that purportedly distinguish ldquoUgandapithecusrdquo fromP africanus are likely related to size For example the somewhatmore central position of the P3 paracone in ldquoUrdquomajor is not seen inthe smaller ldquoUrdquo meswae whose paracone is positioned similarly tothat of the holotype of P africanus (see Harrison and Andrews2009 Fig 6d)
However the major part of the evidence for recognizingldquoUgandapithecusrdquo (sensu Pickford et al 2009) was assembled by re-assigning specimens traditionally attributed to P africanus into newspecies of ldquoUgandapithecusrdquo The P africanus holotype was left inassociation with the Rusinga sample despite having morphologyoverwhelmingly characteristic of other Tinderet specimens Theresulting specimen allocations left P africanus morphologicallyimpoverisheddstripped of much of its hypodigmdso that Pro-consul for Pickford et al (2009) came to be characterized primarilyby the Rusinga sample (see similar comments in Harrison andAndrews 2009) Within that conceptual framework any spec-imen that differs from those at Rusinga could not be Proconsul butmust instead belong to ldquoUgandapithecusrdquo Thus specimens such asthe KNM-SO 1112 mandible were erroneously assigned to ldquoUgan-dapithecusrdquo because they were (correctly) perceived as differingfrom their Rusinga counterparts (Pickford et al 2009)
Here we restore the P africanus hypodigm and associate it withthe other Tinderet fossils according to their shared morphologySpecimens assigned to ldquoUgandapithecusrdquo revert to Proconsul andmany of the features cited to support a speciose ldquoUgandapithecusrdquo(Pickford et al 2009) instead become diagnostic features of Pro-consul This correction serves to highlight the distinctiveness of theKisingiri remains and the need to recognize this distinctiveness atthe generic level
As demonstrated above specimens from Kisingiri do not displaythe morphology of Proconsul as represented by the holotype ofP africanus M 14084 Given that this specimen preserves only theupper canine and cheek teeth similarities with or departures fromthis morphology represent the most compelling arguments forassessing inclusivity in the genus However for the mandible andlower dentition if one simply sorts the specimens by elementwithout relying on taxonomic preconceptions the overwhelmingdistinction within the traditionally constituted Proconsul sample isbetween Kisingiri material on the one hand and that from Tinderetand Uganda on the other (cf Walker et al 1993 Begun 2001 2007MacLatchy and Rossie 2005 Harrison and Andrews 2009Harrison 2010) Additionally with the Kisingiri specimensremoved to Ekembo the postcranial differences used to partlyjustify a distinct ldquoUgandapithecusrdquo lose their relevance (seeHarrison and Andrews 2009)
We have differed among ourselves in our past treatment ofP major as generically distinct from the smaller species of Pro-consul (Begun 2007 McNulty 2010) Morphologically many ofthe unique features of P major can be interpreted as size-relatedvariants on the smaller P africanus morphology Even if so thequestion is whether these are sufficiently different and numerousto warrant generic distinction that is do they suggest thatP major belongs in a separate clade from P africanus andP meswae As we have demonstrated differences among thesethree species are certainly fewer and show less morphologicaldistinction than those between Ekembo and any of the three
Nevertheless while we ultimately agree with Harrison andAndrews (2009 see also Begun 2007) that the features dis-tinguishing ldquoUrdquo major from P africanus are consistent with thosethat distinguish congeneric species and are at least partly due todifferences in size we concede the possibility that the largestProconsul species could belong in a separate genus (Senut et al2000 Pickford and Kunimatsu 2005) However whether or notone recognizes a separate genus for P major the P africanus ma-terial does not sort morphologically with the Kisingiri materialTherefore the taxonomic alternatives are either two generaProconsul and Ekembo or three Proconsul Ugandapithecus andEkembo with Proconsul and Ugandapithecus united in a cladeexclusive of Ekembo Thus while we strongly favor the firstalternative recognizing Ugandapithecus does not change thefundamental argument concerning the need to refer the Kisingirispecies to a generically and cladistically distinct taxon from thespecies represented at Tinderet and the Ugandan sites
Specimens of interest
Despite the substantial dental and gnathic differences betweenProconsul and Ekembo there are a few specimens whose taxonomicaffinities require additional scrutiny
KNM-RU 1676-77 This set of associated upper and lower teeth is aconsistent outlier among E heseloni and E nyanzae specimens Insome ways this specimen more closely resembles Tinderet Pro-consul in the features that distinguish it from Ekembo Molar cuspmorphology and cingulum development I1 lingual tubercledevelopment and M2 shape and posthypocone crista orientationare more consistent with specimens from the Tinderet sites On theother hand it otherwise resembles Ekembo and lacks the strongestdiagnostic features that unite species of Proconsul such as thedistinctive canine and P3 morphology Overall this specimen doesnot fit unambiguously into either genus in fact the degree ofbuccolingual flare cusp morphology enamel wrinkling and espe-cially the long extension of enamel onto the anterior root in P3 arein some ways reminiscent of Afropithecus although it clearly differsfrom that genus as well (cf Rossie and MacLatchy 2013)
Were these characteristics individually distributed among thelarge Kisingiri sample one might attribute this to intra-genericvariation but when found together in a single specimen from alarge sample it suggests a unique individual One of us (KPM)thinks it likely that the specimen was not actually found at aKisingiri locality based on its unique preservation compared toother Rusinga fossils and on other examples of mis-proveniencedspecimens (Leakey 1967 Andrews and Molleson 1979 Pickford1986 Kelley 1986) However we recognize that relying toofrequently on this explanation allows almost any scenario to bejustified and suggest some alternative possibilities One is that thespecimen was found on Rusinga but in the much older WayandoFormation strata instead of the Hiwegi Formation The differencesin morphology in that case might reflect more than a million yearsof temporal change (McCollum et al 2013) with KNM-RU 1676-77preserving certain more primitive Proconsul-like features As analternative it is reasonable to assume that fossil assemblages asvast as those from Rusingawill include not only abundant taxa butalso rare or infrequent inhabitants of the local biome There aremany examples of this from Rusingas mammalian communityeg ldquoHemicyonrdquo Kulutherium Tadarida cf Hyainailouros cfArchaeobeledon Kelba (see relevant chapters in Werdelin andSanders 2010) As such KNM-RU 1676-77 may simply representa rare and well-preserved specimen of some other catarrhinetaxon that lived among or visited a primate community dominatedby Ekembo
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 59
KNM-SO 1112 This specimen was initially referred to Rangwapi-thecus but soon after was transferred to Proconsul afrianus (Bosler1981 Martin 1981)da designation that most subsequent authorshave followed (Pickford 1986 Nengo and Rae 1992 Kelley 19861995 Walker et al 1993 MacLatchy and Rossie 2005 Hill et al2013) Pickford et al (2009) more recently referred this specimento ldquoUrdquo meswae citing two primary reasons First they reiteratedBoslers (1981) concern that its P3 differs from others attributedto P africanus However at that time the majority of what wasconsidered to be P africanus would have been from RusingaIsland In fact while noting its contrasting morphology Boslersuggested that differences between KNM-SO 1112 and other P3smight reasonably be attributed to sexual dimorphism notingthat this specimen provides ldquoa not implausible match for the Pafricanus holotype maxilla M 14084rdquo (1981152) Thatassessment is borne out in this study Using the much largersample of Ekembo P3s as a model variation can be seen toencompass specimens with well-developed mesial beaks andthose with more bilaterally compressed crowns and parallelsides The latter morphology is predominantly found amongmale specimens (KNM-RU 1674 1924 and KPS I and III) thussupporting Boslers (1981) suggestion that this variation isrelated to sexual dimorphism and hence that KNM-SO 1112 is amale P africanus
The second feature cited by Pickford et al (2009) in referringthis specimen to ldquoUrdquo meswae was posterior shallowing of themandibular corpus However because they redefined ldquoUgandapi-thecusrdquo to include all of the well-preserved Tinderet mandibles(Pickford et al 2009)dleaving Proconsul to be represented only byRusinga specimensdtheir only alternative within this scheme for aspecimen the size of SO 1112 was to refer it to ldquoUrdquomeswae It seemsodd though to assign this specimen to a species from a (perhapsmuch) older site when the Proconsul africanus holotype is ofappropriate size and similar age and as there are no other adultmandibles from either species with which to compare it In anycase mandibular shallowing is variable in both Ekembo and Pro-consul with some Kisingiri specimens exhibiting considerableshallowing (eg KNM-RU 2087) and some ldquoUgandapithecusrdquo spec-imens having very little (eg M 14086)
We therefore see no morphological grounds to assign KNM-SO1112 to P meswae but do find some morphological evidence toassign it as a male P africanus which occurs at Songhor
KNM-LG 452 This very complete mandible was originally recog-nized as a female of P major based on the size of the dentition androbusticity of the mandible (Martin 1981) Bosler (1981) howeverleft this specimen unassigned noting similarities to P major in P3-M1 size but having distinctly smaller M2e3 In this she likenedKNM-LG 452 to a few other specimens (notably M 14086) thatare smaller in their distal molars in a manner more consistentwith taxonomic rather than sex differences (Bosler 1981 see alsoPilbeam 1969) Kelley (1986 1995) also assigned this specimen asa female of P major based on canine size and morphology butnoted that the canine lacked the shallowly-sloping mesial ridgetypical of most female anthropoid canines This is also truehowever of two other canines assigned as female P major KO 9and SO 373 (Kelley 1986) and so might simply be an unusualcharacteristic of P major female lower canines In the end Kelley(1995) concluded that considering all of the criteria fordistinguishing male and female canines the evidence that KNM-LG 452 is female remains questionable
Pickford et al (2009) further developed the idea that two spe-cies are represented within the P major sample and made KNM-LG452 the holotype of a new species ldquoUrdquo legetetensis We concur thatthis specimen is unique among the well-preserved Tinderet
mandibles in some important features Whereas male and femalemandibles of P major have long internal symphyseal planesexpansive extra- and retromolar sulci and rami that cross theocclusal plane at distal M3 KNM-LG 452 has a shorter symphysealplane (though not to the extent seen in Ekembo) a moderateextramolar sulcus but no retromolar sulcus and a ramus thatcrosses the occlusal plane more mesially If KNM-SO 396 and SO404 represent males and females respectively of P major then themorphology of KNM-LG 452 appears distinct
At present however we do not think there is sufficient evidenceto differentiate this specimen from either P major or P meswaePickford et al (2009) argue that KNM-LG 452 cannot belong to thelatter species because of its larger size The size differences how-ever are well within the range of variation seen in P majorE heseloni and E nyanzae Moreover whereas the Legetet mandibleis larger than most of the P meswae hypodigm an adult P4 referredto this species (KNM-ME 25) substantially exceeds its counterpartin KNM-LG 452 Should the Legetetmandible ultimately prove to bea male specimen size alone would not rule out its attribution toP meswae On the other hand if it is female then it becomes harderto justify its inclusion in that sample of mostly smaller specimens Iffemale therefore it could represent a separate species for whichthe binomial Proconsul legetetensis comb nov is available Recentlycollected fossils from the SonghorKapurtay area provide someadditional support for the possibility that three species of Proconsulexisted at these localities (McNulty 2014ab) However toconvincingly demonstrate this and determine whether a thirdspecies can be differentiated from P meswae will require a moredetailed analysis of variation among Proconsul species a corre-sponding study of variation in modern taxa anddalmost certain-lydadditional fossil specimens For now we regard ldquoUrdquo legetetensisas a subjective junior synonym of P major
Conclusions
The genus Proconsul has been central to studies of ape and hu-man evolution since its first description by Hopwood (1933) As oneof the best known fossil catarrhines represented by nearly everyskeletal element our knowledge of Proconsuls functional anatomypositional behavior and overall morphology has made it thefoundation for paleoanthropological research in the early Mioceneas well as studies on the origin and early diversification of homi-noids With respect to systematics however Proconsul is less wellunderstood the type species P africanus is known from a rela-tively small number of specimens from sites in the Tinderet areaand no postcranial bones can be unambiguously assigned to itRather it is the copious and well-preserved specimens from theKisingiri sites of Rusinga and Mfangano Islands that have formedthe basis for the common image of Proconsul
We argue here that the Kisingiri species traditionally referredto ProconsuldP heseloni and P nyanzaedare generically distinctfrom P africanus and hereby transfer them into a new genusEkembo based on a substantial number of morphological differ-ences (Table 2) Considering only those traits preserved in theP africanus holotype the entire sample from Rusinga and Mfan-gano differs markedly in the morphology of the upper canine P3P4 and M2 Additional differences in the I1 lower incisors andcanines molar cusp and occlusal crest morphology degree of flarein all molars and especially mandibular features clearly distin-guish the Kisingiri from non-Kisingiri samples in a way that isincompatible with congeneric species With the transfer ofE heseloni and E nyanzae Proconsul becomes a well-defined cladecomprising at least three speciesdP africanus P major andP meswaedunited by several features that are unique for thegenus (Table 2)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6160
We found no support in this study for the genus ldquoUgandapi-thecusrdquo as constructed by Pickford et al (2009) In that paper theytransferred the vast majority of Proconsul fossils from the Tinderetand Napak localities into ldquoUgandapithecusrdquo leaving P africanusrepresented only by its holotype and a small number of isolatedteeth and aligning it with the material from Rusinga and Mfan-ganoWhen the P africanus hypodigm is reconstituted and properlyaffiliated with the other Tinderet species according to itsmorphology much of the evidence marshaled by Pickford et al(2009) for ldquoUgandapithecusrdquo actually supports the distinction be-tween the Tinderet and Kisingiri samples ie the generic distinc-tion of Ekembo In addition some of the traits enumerated byPickford and colleagues in support of ldquoUgandapithecusrdquo wererevealed in our analysis to be variable within one or more speciesand therefore of questionable taxonomic utility We do not rejectout of hand the original conception of ldquoUgandapithecusrdquo (Senutet al 2000) as either a monotypic genus distinguishing ldquoUrdquo ma-jor or perhaps including a closely allied middle Miocene ldquoUrdquogitongai as well (Pickford and Kunimatsu 2005) but we have fornow retained the early Miocene specimens of both in Proconsulpending further study However even if Ugandapithecus is a taxo-nomically valid genus comprising just the latter two species theirmorphology indicates that theywould still be united in a cladewithProconsul and they lack several more advanced features that partlydefine Ekembo Better understanding of the role that body size playsinmediating differences within Proconsulwill help to further clarifythe relationships among these species
Despite the existence of one well-represented and several lesscomplete skeletons of what was long assumed to be Proconsul thephylogenetic status of this genus has defied consensus This islargely due in our view to Proconsul having been an admixture oftwo distinct genera one more advanced in the direction of extantapes than the other As we demonstrate here the material fromRusinga and Mfangano traditionally referred to Proconsul andwhich includes all of the more complete skeletal material isdistinct from the Tinderet and Napak material and belongs in aseparate genus Ekembo By disentangling these distinct clades weanticipate being able to achieve better phylogenetic resolution forboth genera thereby improving our knowledge of early hominoidevolution and diversification
Acknowledgments
We thank many people and agencies that assisted with thedevelopment and completion of this project This research wasinitiated as a result of funding from the Wenner-Gren Foundationand the Johns Hopkins School of Medicine to DR Begun in1987e1989 and was renewed following research and travelgenerously funded by multiple Leakey Foundation grants to KMcNulty National Science Foundation grants to K McNulty(0852609 1241807) the McKnight Foundation the University ofMinnesota the Lloyd A Wilford Endowment the LeverhulmeFoundation and the Turkana Basin Institute J Kelley gratefullyacknowledges support from the Institute of Human Origins Ari-zona State University DR Begun also wishes to acknowledge theAlexander Humboldt Stiftung and NSERC for funding support Weare grateful to the National Museums of Kenya the UgandaMuseum the Natural History Museum and to Rose Mwanja EzraMusiime Tom Mukhuyu Samuel Muteti James Yatich and MaryMuungu for access to and assistance with specimens in their careMany people provided assistance comments criticisms insightsand suggestions on various aspects of this work and we especiallythank Holly Dunsworth Will Harcourt-Smith Tom Lehmann DanPeppe Nicole Garrett Kirsten Jenkins Rutger Jansma Susy CoteIsaiah Nengo Andrew Hill and Brenda Benefit K McNulty
acknowledges the evolutionary anthropology faculty and studentsat Durham University for their helpful comments on an earlyversion of this manuscript and for their gracious hospitality Weare likewise grateful to the anonymous reviewers associate editorand editor whose comments strengthened the paper This manu-script is publication 1 supporting Research on East AfricanCatarrhine and Hominoid Evolution (REACHE)
Appendix A Supplementary data
Supplementary data related to this article can be found at httpdxdoiorg101016jjhevol201503009
References
Andrews P 1974 New species of Dryopithecus from Kenya Nature 249 188e190Andrews P 1978 A revision of the Miocene Hominoidea of East Africa Bull Brit
Mus (Nat Hist) Geol 30 85e225Andrews P 1985 Family group systematics and evolution among catarrhine pri-
mates In Delson E (Ed) Ancestors The Hard Evidence Liss New Yorkpp 14e22
Andrews P Harrison T Martin L Pickford M 1981 Hominoid primates from anew Miocene locality named Meswa Bridge in Kenya J Hum Evol 10123e128
Andrews P Begun D Zylstra M 1997 Interrelationships between functionalmorphology and paleoenvironments in Miocene hominoids In Begun DWard C Rose M (Eds) Function Phylogeny and Fossils Miocene HominoidEvolution and Adaptations Plenum Press New York pp 29e58
Andrews P Molleson T 1979 The provenance of Sivapithecus africanus Bull BritMus (Nat Hist) Geol 32 19e23
Begun DR 2001 African and Eurasian Miocene hominoids and the origins of theHominidae In de Bonis L Koufos GD Andrews P (Eds) Hominoid Evolutionand Climatic Change in Europe Phylogeny of the Neogene Hominoid Primatesof Eurasia Cambridge University Press London pp 231e253
Begun DR 2007 Fossil record of Miocene hominoids In Henke W Tattersall I(Eds) Handbook of Paleoanthropology Springer Berlin pp 921e977
Begun DR Teaford MF Walker AC 1994 Comparative and functional anatomyof Proconsul phalanges from the Kaswanga primate site Rusinga Island KenyaJ Hum Evol 26 89e165
Bishop WW 1964 More fossil primates and other Miocene mammals from North-East Uganda Nature 203 1327e1331
Bishop WW Miller JA Fitch FJ 1969 New potassium-argon age determinationsrelevant to the Miocene fossil mammal sequence in East Africa Am J Sci 267669e699
Blankenship JA Mincer HH Anderson KM Woods MA Burton EL 2007Third molar development in the estimation of chronologic age in Americanblacks as compared with whites J Forensic Sci 52 428e433
Bosler W 1981 Species groupings of early Miocene dryopithecine teeth from EastAfrica J Hum Evol 10 151e158
Coleman MN 2008 What does geometric mean mean geometrically Assessingthe utility of geometric mean and other size variables in studies of skullallometry Am J Phys Anthropol 135 404e415
Cote S Malit N Nengo I 2013 Additional mandibles of Rangwapithecus gordonian early Miocene catarrhine from the Tinderet localities of Western KenyaJ Hum Evol 153 341e352
Deane AS 2009 Early Miocene catarrhine dietary behavior the influence of theRed Queen Effect on incisor shape and curvature J Hum Evol 56 275e285
Drake R Van Couvering JA Pickford M Curtis G Harris JA 1988 New chro-nology for the Early Miocene mammalian faunas of Kisingiri Western KenyaJ Geol Soc Lond 145 479e491
Evernden JF Savage DE Curtis GH James GT 1964 Potassium-argon datesand the Cenozoic mammalian chronology of North America Am J Sci 262145e198
Gebo DL MacLatchy L Kityo R Deino A Kingston J Pilbeam D 1997A hominoid genus from the Early Miocene of Uganda Science 276 401e404
Geoffroy Saint-Hilaire E 1812 Tableau des quadrumanes 1 Ord QuadrumanesAnnls Mus Hist Nat Paris 19 85e122
Gray JE 1825 Outline of an attempt at the disposition of the Mammalia into tribesand families with a list of the genera apparently appertaining to each tribeAnnls Philos 10 337e344
Harrison T 1982 Small-bodied apes from the Miocene of East Africa PhDDissertation University of London
Harrison T 1986 New fossil anthropoids from the middle Miocene of East Africaand their bearing on the origin of the Oreopithecidae Am J Phys Anthropol 71265e284
Harrison T 1987 The phylogenetic relationships of the early catarrhine primates areview of the current evidence J Hum Evol 16 41e80
Harrison T 2002 Late Oligocene to middle Miocene catarrhines from Afro-ArabiaIn Hartwig WC (Ed) The Primate Fossil Record Cambridge University PressCambridge pp 311e338
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 61
Harrison T 2010 Dendropithecoidea Proconsuloidea and Hominoidea (CatarrhiniPrimates) In Werdelin L Sanders WJ (Eds) Cenozoic Mammals of AfricaUniversity of California Press Berkeley pp 429e469
Harrison T Andrews P 2009 The anatomy and systematic position of a newspecies of early Miocene proconsulid from Meswa Bridge Kenya J Hum Evol56 479e496
Hill A Nengo IO Rossie JB 2013 A Rangwapithecus gordoni mandible from theearly Miocene site of Songhor Kenya J Hum Evol 65 490e500
Hopwood AT 1933 Miocene primates from Kenya J Linn Soc Zool 38 437e464Ishida H Kunimatsu Y Takano T Nakano Y Nakatsukasa M 2004 Nacholapi-
thecus skeleton from the Middle Miocene of Kenya J Hum Evol 46 69e103Jolicouer P 1963 The multivariate generalization of the allometry equation Bio-
metrics 19 497e499Jungers WL Falsetti AB Wall CE 1995 Shape relative size and size-
adjustments in morphometrics Am J Phys Anthropol 38 (S2) 137e161Kavanagh KD Evans AR Jernvall J 2007 Predicting evolutionary patterns of
mammalian teeth from development Nature 449 427e433Kelley J 1986 Species recognition and sexual dimorphism in Proconsul and
Rangwapithecus J Hum Evol 15 461e495Kelley J 1995 Sex determination in Miocene catarrhine primates Am J Phys
Anthropol 96 390e417Kelley J 1993 Taxonomic implications of sexual dimorphism in Lufengpithecus In
Kimbel WH Martin LB (Eds) Species Species Concepts and Primate Evo-lution Springer New York pp 429e458
Kent PE 1942 The country round the Kavirondo Gulf of Victoria Nyanza Geogr J100 22e31
Kordos L Begun DR 2001 A new cranium of Dryopithecus from RudabanyaHungary J Hum Evol 15 689e700
Le Gros Clark WE Leakey LSB 1950 Diagnoses of East African Miocene Homi-noidea Q J Geol Soc Lond 105 260e263
Le Gros Clark WE Leakey LSB 1951 The Miocene Hominoidea of East Africa InFossil Mammals of Africa No 1 British Museum (Natural History) London
Leakey LSB 1943 A Miocene anthropoid mandible from Rusinga Kenya Nature152 319e320
Leakey LSB 1963 East African fossil Hominoidea and the classification within thissuper-family In Washburn SL (Ed) Classification and Human EvolutionAldine Chicago pp 32e49
Leakey LSB 1967 Notes on the mammalian faunas from the Miocene and Pleis-tocene of East Africa In Bishop WW Clark JD (Eds) Background to Evolu-tion in Africa University of Chicago Press Chicago pp 7e29
Leakey RE Leakey MG 1986 A new Miocene hominoid from Kenya Nature 324143e146
Leakey RE Leakey MG Walker AC 1988 Morphology of Afropithecus turka-nensis from Kenya Am J Phys Anthropol 76 289e307
Leakey MG Ungar PS Walker A 1995 A new genus of large primate from theLate Oligocene of Lothidok Turkana District Kenya J Hum Evol 28519e531
Linnaeus C 1758 Systema naturae per regna tria naturae secundum classesordines genera species cum characteribus differentris synonymis locis tenthed Laurentii Salvii Stockholm
MacInnes DG 1943 Notes on the East African Miocene primates J E Afr UgandaNat Hist Soc 17 141e181
MacLatchy L 2004 The oldest ape Evol Anthropol 13 90e103MacLatchy L Bossert WH 1996 An analysis of the articular surface distribution of
the femoral head and acetabulum in anthropoids with implications for hipfunction in Miocene hominoids J Hum Evol 31 425e453
MacLatchy L Rossie JB 2005 The Napak hominoid still Proconsul major InLieberman DE Smith RJ Kelley J (Eds) Interpreting the Past Essays onHuman Primate and Mammal Evolution in Honor of David Pilbeam Brill Ac-ademic Publishers Boston pp 15e28
Martin L 1981 New specimens of Proconsul from Koru Kenya J Hum Evol 10139e150
McCollum MS Peppe DJ McNulty KP Dunsworth HM Harcourt-Smith WEH Andrews AL 2013 Magnetostratigraphy of the early MioceneHiwegi Formation (Rusinga Island Lake Victoria Kenya) Geological Society ofAmerica Abstracts with Programs 45 p 12
McNulty KP 2010 Apes and tricksters the evolution and diversification ofhumans closest relatives Evol Edu Outreach 3 322e332
McNulty KP Harcourt-Smith WEH Dunsworth HM 2007 New primate fossilsfrom Rusinga Island Kenya Am J Phys Anthropol 132 (S44) 170
McNulty KP MacLatchy L Peppe DJ Nengo I Manthi FK Miller ERStevens NJ Cote S Lehmann T Kingston J 2014a A regional approach toEast African early Miocene paleobiology J Vert Paleontol 185 SVP Programand Abstracts Book 2014
McNulty KP MacLatchy L Rossie JB Peppe DJ Deino AL Mbua ENManthi FK Nengo IO Stevens NJ Cote S Lehmann T Gutierrez M 2014b
Research on East African catarrhine and hominoid evolution results from thefirst year Am J Phys Anthropol 153 (S58) 182
Michel LA Peppe DJ Lutz JA Driese SG Dunsworth HM Harcourt-Smith WEH Horner W Lehmann T Nightengale S McNulty KP 2014Remnants of an ancient forest provide ecological context for Early Miocenefossil apes Nature Comm 5 3236
Myres JL 1932 Africa Man 32 208Nakatsukasa M Kunimatsu Y Nakano Y Takano T Ishida H 2007 Vertebral
morphology of Nacholapithecus kerioi based on KNM-BG 35250 J Hum Evol52 347e369
Nengo IO Rae TC 1992 New hominoid fossils from the early Miocene site ofSonghor Kenya J Hum Evol 23 423e429
Peppe DJ McNulty KP Cote SM Harcourt-Smith WEH Dunsworth HM VanCouvering JA 2009 Stratigraphic interpretation of the Kulu Formation (EarlyMiocene Rusinga Island Kenya) and its implications for primate evolutionJ Hum Evol 56 447e461
Pickford M 1981 Preliminary Miocene mammalian biostratigraphy for westernKenya J Hum Evol 10 73e97
Pickford M 1986 Sexual dimorphism in Proconsul Hum Evol 1 111e148Pickford M Andrews PJ 1981 The Tinderet Miocene sequence in Kenya J Hum
Evol 10 11e33Pickford M Kunimatsu Y 2005 Catarrhines from the Middle Miocene (ca 145
Ma) of Kipsaraman Tugen Hills Kenya Anthropol Sci 113 189e224Pickford M Senut B Gommery D Musiime E 2009 Distinctiveness of Ugan-
dapithecus from Proconsul Estudios Geol 65 183e241Pilbeam DR 1969 Tertiary Pongidae of East Africa evolutionary relationships and
taxonomy Bulletin of the Peabody Museum of Natural History 31 pp 1e185Rae TC 1993 Phylogenetic analysis of proconsulid facial morphology PhD
Dissertation State University of New York at Stony BrookRae TC 1999 Mosaic evolution in the origin of the Hominoidea Folia Primatol 70
125e135Rafferty KL Walker A Ruff C Rose MD Andrews PJ 1995 Postcranial esti-
mates of body weights in Proconsul with a note on a distal tibia of P major fromNapak Uganda Am J Phys Anthropol 97 391e402
Rossie JB MacLatchy L 2013 Dentognathic remains of an Afropithecus individualfrom Kalodirr Kenya J Hum Evol 65 199e208
Senut B Pickford M Gommery D Kunimatsu Y 2000 Un nouveau genredrsquohominoiumlde du Miocene inferieur drsquoAfrique orientale Ugandapithecus major(Le Gros Clark and Leakey 1950) CR Acad Sci Paris 331 227e233
Simons EL Pilbeam DR 1965 Preliminary revision of the Dryopithecinae (Pon-gidae Anthropoidea) Folia Primatol 3 81e152
Smith TM Martin LB Leakey MG 2003 Enamel thickness microstructure anddevelopment in Afropithecus turkanensis J Hum Evol 44 283e306
Stevens NJ Seiffert ER OConnor PM Roberts EM Schmitz MD Krause CGorscak E Ngasala S Hieronymus TL Temu J 2013 Palaeontological evi-dence for an Oligocene divergence between Old World monkeys and apesNature 497 611e614
Suwa G Kono RT Katoh S Asfaw B Beyene Y 2007 A new species of great apefrom the late Miocene epoch in Ethiopia Nature 448 921e924
Szalay FS Delson E 1979 Evolutionary History of the Primates Academic PressNew York
Teaford MF Beard KC Leakey RE Walker A 1988 New hominoid facial skel-eton from the Early Miocene of Rusinga Island Kenya and its bearing on therelationship between Proconsul nyanzae and Proconsul africanus J Hum Evol17 461e477
Van Couvering JAH Miller J 1969 Miocene stratigraphy and age determinationsRusinga Island Kenya Nature 221 628e632
Walker A 1997 Proconsul function and phylogeny In Begun DR Ward CVRose MD (Eds) Function Phylogeny and Fossils Miocene Hominoid Evolu-tion and Adaptation Plenum Press New York pp 209e224
Walker A Pickford M 1983 New postcranial fossils of Proconsul africanus andProconsul nyanzae In Ciochon RL Corruccini RS (Eds) New Interpretationsof Ape and Human Ancestry Plenum Press New York pp 325e351
Walker A Teaford M 1988 The Kaswanga primate site an Early Miocene hom-inoid site on Rusinga Island Kenya J Hum Evol 17 539e544
Walker A Teaford M 1989 The hunt for Proconsul Sci Am 260 76e82Walker A Teaford MF Martin L Andrews P 1993 A new species of Proconsul
from the early Miocene of RusingaMfangano Islands Kenya J Hum Evol 2543e56
Ward CV Ruff CB Walker A Rose MD Teaford MF Nengo IO 1995 Func-tional morphology of Proconsul patellas from Rusinga Island Kenya with im-plications for other Miocene-Pliocene catarrhines J Hum Evol 29 1e19
Ward S Brown B Hill A Kelley J Downs W 1999 Equatorius a new hominoidgenus from the Middle Miocene of Kenya Science 285 1382e1386
Werdelin L Sanders WJ 2010 Cenozoic Mammals of Africa University of Cali-fornia Press Berkeley

Figure 2 Upper central incisors of Proconsul and Ekembo Lingual views of Ekembo(KNM-RU 1685 cast) left and Proconsul (KNM-CA 1300) right Note the differences inthe transition between the lingual cingulum and the mesial marginal ridge which ismore rounded in Proconsul and more angled in Ekembo The central pillar is also morebulbous or inflated and without ridging in Proconsul Additional specimens illustratingthe Proconsul morphology are shown in Pickford et al (2009 Fig 17 p 197) butreferred in that manuscript to Ugandapithecus
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6148
thanmerging into the hypocone on its lingual surface and a distallyoriented posthypocone crista Lower first and second molars nearlyequal in buccolingual breadth
Comparative morphology of Ekembo and Proconsul
Samples and comparative methods
The above diagnoses are based on extensive survey and analysisof the original fossil material Specimens housed in the NationalMuseums of Kenya (NMK) and the Natural History Museum (Lon-don) were analyzed by the authors during several separate and twojoint (NMK) research visits Fossils in the Uganda National Museumwere assessed by one of us (KPM) during two visits All measure-ments were collected by KPM Dental terminology follows Szalayand Delson (1979)
The following results are organized with respect to themorphology of the Kisingiri sample (attributed to E nyanzae andE heseloni) versus that of Proconsul from Tinderet and Napak(traditionally referred to P africanus P major and more recentlyP meswae) Specimens from additional localities that have at onetime been attributed to Proconsul (eg Andrews1978 Pickford andKunimatsu 2005 [cf Harrison 2010]) are for the most part notconsidered here most are middle Miocene in age and have beentransferred to other genera (Harrison 2002 2010) Of those that arefrom early Miocene sites there is growing evidence that P majorcan also be found at Moroto and differentiated fromMorotopithecus(Pickford et al 2009) an issue of some interest but ultimately notcritical to the taxonomic questions being considered here Whereappropriate however we do provide observations on a few earlyMiocene specimens attributed to ldquoUrdquo gitongai (Pickford andKunimatsu 2005 Pickford et al 2009 see Harrison 2010) Yetwhile we consider the middle Miocene holotype likely to representa distinct species here referred to Proconsul we concur withHarrison (2010) that many specimens placed in this species byPickford et al (2009) cannot be distinguished from P major
Sample sizes particularly from Tinderet and Ugandan localitiestend to be small but where possible standard univariate (eg t-tests) and bivariate (regression ANCOVA) statistical analyses wereperformed A single eigendecomposition of the covariance matrixof symphyseal measurements was done to characterize the multi-variate shape of the mandibular symphysis after accounting for sizedifferences (sensu Jolicouer 1963) Quantitative results are illus-trated as box-and-whisker plots of shape indices or bivariate plotsof the measurements For samples of fewer than five specimensshape index values are plotted in lieu of box-and-whiskersummaries
Morphological comparisons
Upper incisors Two features of the Ekembo upper central incisorsdistinguish them from those of Proconsul including specimenswidely attributed to the type species P africanus The lingual tu-bercle in Ekembo is quite narrow generallywith low relief and oftenscored by two or more irregular ridges in contrast to the muchmore inflated or bulbous tubercle in Proconsul I1s (Fig 2) Ekembospecimens also have a characteristic morphology in the transitionfrom the lingual cingulum to the mesial marginal ridge (Fig 2)from the mesiodistal mid-point of the crown the cingulumextends either directly mesially or even angles slightly cervicallyto intersect the mesial marginal ridge at a fairly sharp angle Incontrast in I1s of Proconsul the cingulum angles incisally from themid-point and transitions into the mesial marginal ridge in agradual curve The expression of these traits varies somewhat inthe smaller Tinderet specimens attributed to P africanus
compared with those of P major (cf Pickford et al 2009)However these variations are relatively minor with respect to thedifferences between the two genera Of 25 I1s known fromKisingiri localities only one specimen KNM-MW 562 somewhatresembles the Tinderet sample in both of these featuresdhavinga somewhat less angled transition from the mesial marginal ridgeto the cingulum and a more bulbous lingual tubercle This isparticularly interesting as the specimen was discovered in theMakira Beds now dated to older than 190 Ma and contemporarywith the Tinderet localities (McCollum et al 2013) Neverthelessits overall morphology is more similar to other Ekembospecimens than to Proconsul incisors Two other Ekembospecimens KNM-RU 1677 and RU 1933 have somewhat moreinflated lingual tubercles but otherwise resemble other Ekembo I1s
No consistent differences were found in upper lateral incisors
Upper canines Upper canine crowns of Ekembo are typical of thoseof most catarrhines in terminating in an evenly tapering point Thisdiffers from the Proconsul specimens which have a blade-like(Kelley 1986 Senut et al 2000 Harrison 2010) or ldquoburin-likerdquo(Pickford et al 2009) canine tip (Fig 3) While blade-like apicalmorphology has long been recognized as a distinctivecharacteristic of P major we confirm the report by Pickford et al(2009) that this feature is also present in the P africanus typespecimen (Fig 3a) although somewhat understated as expectedin the more slender and higher-crowned P africanus caninesUnfortunately other canines attributed to P africanus (eg M44837 KNM SO 419 LG 921 CA 1910 SO 5353 Kelley 1986) havebroken apices and therefore cannot be evaluated for this featureA blade-like tip is also present in the deciduous upper canines ofP major (eg KNM-SO 371 MV 10 NAP IV UMP 62-20) andP meswae (KNM ME 11) Hence the blade-like upper canine tipunites all Tinderet and Napak Proconsul species but is neverexpressed in Ekembo specimens from Rusinga and Mfangano
Two features of the canine roots also differentiate these generalongitudinal grooving or fluting along the length of the root (cfKelley 1986) and the development of numerous finer longitudinal
Figure 3 Canines of Proconsul and Ekembo a) Upper canine of the P africanus holotype (M 14084) lingual view b) Upper canine of P major (KNM-CA 2127) lingual view c) Lowercanine of P major buccal view d) lower canine of Ekembo cf nyanzae (KNM-RU 1676) buccal view and e) Upper canine of P major (KNM-SO 584) distal view Note the blade-like orldquoburin-likerdquo tip of the unworn Proconsul canines including on the nearly unworn canine tip of the P africanus type specimen (a) Ekembo canines have more typical pointed tipsProconsul canines also have longitudinal grooves and fine striations on the root (e) Ekembo lacks these features Scale bars frac14 1 cm
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 49
striations near the root apex that extend to varying degrees towardthe cervix of the tooth Longitudinal ldquoflutingrdquo consists of broadgrooves that segment the root into distinct sections (cf Pickfordet al 2009 Fig 3e) Most often the root in cross-section has theappearance of a cylinder that has been sectioned longitudinallywith the two halves offset from each other but in some cases thereare three segments and the root cross-section is more triangular inappearance (Kelley 1986) This characteristic was noted by Kelley(1986) in the P africanus canines but not in those of P majorHere we affirm that fluting is present on all of the visible uppercanine roots attributed to both P africanus and P major rootmorphology could not be evaluated in the deciduous upper caninefrom Meswa Bridge since it is implanted in the maxillary alveolusNone of the Ekembo specimens display this morphology havinginstead a simpler round to ovoid cross-section along the length ofthe root
The fine longitudinal striations on Proconsul upper canine rootsvary in the degree to which they extend toward the cervix (Fig 3e)but they are visible in all specimens attributed to P africanus(KNM-SO 419 521 921 5353 KNM-X 502) and five of six uppercanines of P major (excepting KNM-SO 1614) This morphology isalso present in UMP 62-12 which was included as part of theMorotopithecus bishopi hypodigm (Gebo et al 1997) Fine striationsalong the upper canine root are almost entirely absent in EkemboTwenty of twenty-three specimens lack this feature three speci-mens (KNM-RU 1763 1845 1891) exhibit incipient to slight
striations but these are never developed to the extent seen inProconsul canines
Upper premolars Upper third premolars display some of thegreatest differences between Ekembo and Proconsul P4s tend todiffer in similar ways but the differences are not nearly as markedEkembo P3s have cusps positioned near the mesiodistal midpoint ofthe crown well-developed mesial foveae pre- and postparacristaeof similar length and slope resulting in symmetric or nearsymmetric buccal profiles a straight or slightly curved mesialmargin and only moderately curved distal margin (Fig 4)Proconsul P3s are more similar in appearance to those of small-bodied catarrhines from the early Miocene with cusps positionedmesially on the crown greatly reducing or eliminating the mesialfovea They also have a steep preparacrista and more shallowlysloped postparacrista resulting in a highly asymmetric buccalprofile and an inwardly angled or notched mesial margin but astrongly curved distal margin This morphology is bestcharacterized in the P africanus holotype (M 14084) but can alsobe seen in other specimens attributed to this species as well asthose referred to P major (Fig 4d e) Also noteworthy is adistinctive mesiobuccal flange that mesially extends the buccalportion of the crown but only near the cervical line of the toothPickford et al (2009) describe this feature in some detail forldquoUgandapithecusrdquo but in fact this morphology is clearly expressedin the P africanus holotype as well (Fig 4a d) A single adult P3
Figure 4 Upper premolars of Proconsul and Ekembo occlusal (aec) and lingual (def) views a) P3e4 of the P africanus holotype (M 14084) b) P3 of P major (M 14331) c) P3e4 of theE heseloni holotype (KNM-RU 2036) d) P3 of the P africanus holotype (M 14084) e) P3 of P major (M 14331) f) P3 of the E heseloni holotype (KNM-RU 2036) Note the mesiallypositioned cusps lack of mesial fovea and strongly asymmetric crown in Proconsul compared to Ekembo (see text for further comparisons) Image of M 14331 reversed for easiercomparison Photo 4a courtesy of Rutger Jansma Scale bars frac14 1 cm
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6150
germ and two dP3s attributed to P meswae look remarkably similarto those of other Proconsul species including the presence of thisflange In contrast the Ekembo P3 has a well-developed mesialfovea that terminates at a mesiobuccal shoulder positionedapproximately halfway between the paracone tip and the cervixFrom this shoulder the mesial margin angles slightly distallydown to the cervix (Fig 4f)
The degree of cusp height heteromorphy in P3 also differentiatesEkembo from Proconsul (contra Pickford et al 2009) Only four non-Kisingiri specimens were sufficiently unworn to take measure-ments but more worn specimens also appear to express this dif-ferential in cusp height Figure 5a shows the ratio of protoconeparacone height measured from the tooth cervix and with only oneexception Ekembo P3s have substantially less cusp heteromorphyThis exception KNM-RU 1677 does not in fact have greater heter-omorphy but instead has an abnormal extension of the enamel ontothe buccal (paracone) root which skews the measurement Hencecusp relief relative to the occlusal surface in RU 1677 is not nearly asheteromoprhic as in Proconsul and when this is taken into accountthe difference between genera is significant (t-test dffrac14 11 t frac14 283p frac14 00165) The adult P3 germ from Meswa Bridge is incompletelyformed and therefore could not be measured However the differ-ence between paracone and protocone heights with respect to theocclusal surface appears to be as great as that in the P africanusholotype It does not appear that sex is an important determinant ofthis feature since well-established male (eg M16647 KNM-RU16000) and female (KNM-RU 2036 7290) specimens do not segre-gate across the Kisingiri range of variation
Upper fourth premolars of Ekembo are broadly similar inmorphology to Ekembo P3s in fact the similarity between P3 and P4
is substantially greater in Ekembo than in Proconsul (Figs 4a c)Likewise most of the morphological differences that discriminatethe P3s of these genera also tend to differentiate the P4s but less
clearly Cusp heteromorphy does not distinguish P4s of thesegenera but the overall height of the P4 paracone in Ekembo issignificantly greater relative to crown length than in Proconsul P4s(t-test df frac14 14 t frac14 316 p frac14 00069 Fig 5b)
Upper molars Cusps of Ekembo upper molars are uniformly morebunodont than those of Proconsul occupying more of the occlusalsurface Intercuspal crests are more fully incorporated into thecusps in Ekembo molars giving them a more inflated appearanceThis is particularly evident in the crista obliqua which is very broadin Ekembo but thin and knife-like in Proconsul (Fig 6) Cusps ofProconsul molars stand more as individual cones Pickford et al(2009) have further differentiated the cusp morphology ofsmaller and larger Tinderet specimens arguing that the latterhave more bulbous cusps While accurate this difference is minorcompared with the morphological difference between Ekemboand Proconsul and we regard the within-Proconsul cuspalvariation as size-related In fact the intermediately sizedP meswae has cusp morphology that is appropriatelyintermediate between that of P africanus and the larger P major
Other differences between Ekembo and Proconsul upper molarsare most evident in M2 We concur with previous assessments thatthe Kisingiri upper molars have less developed cingula than non-Kisingiri molars (eg Senut et al 2000 Harrison 2002 2010Harrison and Andrews 2009) More specifically Ekembo M2s lackor have only rudimentary buccal cingula between the paracone andmetacone Likewise the lingual cingulum is discontinuous merg-ing into the hypocone lingually and then re-emerging as a distalmarginal ridge (Fig 6) Proconsul M2s display substantially morecingular development overall and in particular exhibit a prominentbuccal shelf between the paracone and metacone and a distinctdistolingual cingulum that is continuous around the hypocone towhere it merges with the distal marginal ridge
Figure 5 Box-and-whisker plots a) P3 cusp heteromorphy index (protocone heightparacone height) Results of a t-test show this difference to be significant (p frac14 00165) Theexcluded Ekembo outlier KNM-RU 1677 only appears to have extreme heteromorphy because of extensive expansion of the buccal enamel onto the anterior root (see text) b) P4
relative paracone height (paracone heightmesiodistal crown length) Results of a t-test show this difference to be significant (p frac14 00069)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 51
The occlusal profiles of Ekembo M2s are squarish in shape withrelatively straight and equal sides with a posthypocone crista thatis distobucally to buccally directed In contrast the occlusal profilesof Proconsul M2s have a more rhomboid shape with substantialbasal flare around the mesial cusps and a distally expanded hypo-cone with a distally directed posthypocone crista resulting in astrongly curved distal margin (Fig 6) Even in specimens with lesscurved distal margins the posthypocone crista is still more distallydirected compared to the condition in Ekembo These features arecharacteristic not only of P africanus P meswae and P major butare also expressed in the isolatedM2 fromMoroto Uganda (MOR IIb2098) attributed by Pickford et al (2009) to ldquoUrdquo gitongai butregarded here as belonging to P major
As noted by others (eg Harrison and Andrews 2009) the de-gree of buccolingual molar flare differs among these samplesQuantified as the distance between the tips of the mesial cuspsdivided by maximum crown breadth the degree of molar flare in
Ekembo M2s is significantly less than in Proconsul M2s (t-testdf frac14 29 t frac14 396 p frac14 00004) In a bivariate plot of these twomeasures the distributions of Ekembo and Proconsul teeth barelyoverlap with only KNM-RU 1677 encroaching into Proconsul ter-ritory (Fig 7)
These differences in M2 morphology also tend to distinguishEkembo from Proconsul M1s but not always as clearly Whereas M2smore consistently express distinct morphotypes the M1s in bothgenera show more variation in these features
The degree of upper third molar development (ie whether M3
has four distinct cusps or the distal cusps are reduced or absent) haslong been cited as taxonomically informative for these species orgroups of these species (eg Hopwood 1933 Le Gros Clark andLeakey 1951 Andrews 1978 Pickford et al 2009) However wefound this tooth to be unreliable for taxonomic purposes Withinprimates M3s are typically the most variable molars in both sizeand shape (Blankenship et al 2007 Kavanagh et al 2007) and this
Figure 6 Occlusal view of the M2s of Proconsul and Ekembo Left P africanus holotype(M 14084) Right E heseloni holotype (KNM-RU 2036) Note in Proconsul the overalldifference in crown shape as well as the more conical individuated cusps and nar-rower cristae strongly developed lingual cingulum that continues to the distal aspectof the crown broad buccal cingular shelf and more distally directed posthypoconecrista In Ekembo the cusps are more bunodont and expanded the cristae are broaderwith better developed accessory tubercles the lingual cingulum merges into thehypocone there is no buccal shelf and the posthypocone crista is more buccallydirected
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6152
is observable even in small samples of Proconsul species TheP africanus holotype has a substantially reduced M3 whereas inKNM-CA 2250dattributed by most authors to this species(Andrews 1978 Pickford et al 2009)dit is fully developed Simi-larly the hypodigm of P major includes both fully developed M3s(eg KNM-CA 389 LG 1815) and specimens with reduced ormissing hypocones (KNM-CA 397 1299) Ekembo displays compa-rable ranges of variation from highly reduced to fully developedM3s in both species Thus these species and genera cannot bedistinguished based on M3 development and we did not find anyother distinguishing characteristics between Ekembo and Proconsulin the upper third molars
Figure 7 Scatter plot depicting the de
Mandible Pickford et al (2009) enumerated several traits thatreadily distinguish Kisingiri from non-Kisingiri mandibles andthese distinctions are entirely consistent with the distinction ofEkembo from Proconsul Our analyses specifically underscoredifferences in the symphyseal region The planum alveolare inboth male and female specimens of Ekembo is short and morevertically oriented even the most elongate of these (KNM-RU47805 McNulty et al 2007) is substantially more verticalcompared to specimens of Proconsul In contrast the planumalveolare in Proconsul is conspicuously long and nearlyhorizontally oriented Well-preserved mandibles attributed toP major (eg KNM-SO 396 and 404) exhibit this morphology inextreme fashion (Fig 8) smaller specimens (KNM-SO 1112P africanus) show less extreme expression but are still readilydistinguishable from Ekembo mandibles For its size KNM-LG 452has a surprisingly short planum alveolare though not to thedegree seen in Rusinga mandibles Correlated to the shortplanum alveolare in Ekembo is a reduced superior transversetorus which while well-developed does not approach therobusticity expressed in Proconsul mandibles This difference isreflected in an index of symphyseal robusticity (thicknessperpendicular to the long axis of the symphysislength of thelong axis) which shows Ekembo and Proconsul to be significantlydifferent (t-test df frac14 14 t frac14 430 p frac14 00007)
Development of the inferior transverse torusdoften used todifferentiate among species of Proconsul (eg Pickford et al 2009Harrison 2010)dwas found to be highly variable and inconsis-tently expressed within these samples However the overallorientation of the subplanum region (as defined by a line con-necting the posterior-most midline points of the superior andinferior transverse tori or the posterior-most midline point of theinferior margin of the symphysis in the absence of the latter) doesdistinguish Ekembo from Proconsul mandibles The orientation ofthis line in relation to the alveolar margin in the vicinity of thepostcanine teeth was assessed qualitatively by placing a small rod
gree of buccolingual flare in M2
Figure 8 Mandibles of Proconsul and Ekembo Occlusal views of P major a) KNM-SO 396 b) KNM-SO 404 c) KNM-LG 452 Occlusal views of Ekembo d) KNM-RU 2087 e) KNM-RU1674 f) KNM-RU 7290 Occlusal views of P africanus g) KNM-SO 372 h) KNM-SO 1112 i) Lingual view of P africanus (KNM-SO 1112) Note the long more horizontally orientedplanum alveolare in Proconsul resulting in a pronounced superior transverse torus The planum alveolare is shorter and more vertical in Ekembo such that one can see the inferiormandibular border from occlusal view
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 53
(eg a dental pick) in themidline internally along the two referencepoints and recording the point in lingual perspective at which therod intersected the alveolar margin (Fig 9) In Ekembo the sub-planum region is more vertically oriented so that the rod intersectsthe alveolar margin most often in the premolar field Not surpris-ingly there is a difference between sexes with females having areduced superior transverse torus and therefore a more verticallyoriented subplanum region compared to males In contrast withEkembo Proconsul mandibles have much more inclined subplanumsurfaces with the rod intersecting the lingual alveolar margin inthe vicinity of the M1 (females) or M2 (males Fig 9) When sex istaken into account therefore the distributions of Ekembo andProconsul in this feature are discrete Note that male and femalemandibles attributed to both P africanus and P major are includedin this assessment and are surprisingly similar despite dramaticsize differences
We also quantified internal symphyseal morphology using threemeasures symphyseal height (infradentaleegnathion) planumalveolare length (posterior-most point on the superior transversetoruseinfradentale) and subplanum length (posterior-most pointon the superior transverse torusegnathion) a combination thatincorporates not only the lengths of these intervals but also theangular relationships among them To compare symphyseal shapeacross a range of sizes we chose a simple principal componentsapproach to partition size onto the first eigenvector (cf Jolicouer1963) While this is not the preferred method for partitioning sizein most datasets (Jungers et al 1995) for this particular sample andset of measurements Mosimann shape variables were still
significantly correlatedwith size (see discussion in Coleman 2008)Hence we assessed differences in symphyseal morphology be-tween Ekembo and Proconsul using the second and third principalcomponents of the three variables Distributions of the two generaare distinct in this shape space (Fig 10 Supplementary OnlineMaterial [SOM] Table 1)
Robusticity of the mandibular corpus also differs between thesesamples though it is more difficult to test due to differentialpreservation among specimens Corpus thickness and depth underP4 can be measured in the greatest number of specimens andcomparison of these measures suggests a difference betweengroups in the scaling of corpus robusticity (Fig 11) Among largepresumably male specimens Ekembo mandibles are substantiallyless robust than those of Proconsul However among smallermandibles only a single Proconsul specimen (KNM-SO 372) could beincluded and it is similar in robusticity to Ekembomandibles of thissize Considered together the impression is that corpus robusticitymay scale differently in Ekembo than it does in Proconsul and a testfor differences in slopes using an ANCOVA design of the loggedvariables was significant (df frac14 2 F frac14 2481 p lt 00001) Never-theless additional specimens are needed to properly evaluate theinfluence of taxonomy sex and size on corpus robusticity areasonable alternative interpretation of Figure 11 is that it dem-onstrates a significant difference between P major and all otherspecies of both Proconsul and Ekembo
Lower incisors Ekembo specimens are uniformly narrower inmesiodistal length than incisors attributed to P africanus P major
Figure 10 Scatter plot of principal component scores based on an eigenanalysis of threeposterior point on the superior transverse toruseinfradentale) and subplanum length (mprimarily captures size differences among specimens (Jolicouer 1963 see text and SOM Trespectively) in the symphyseal variables
Figure 9 Line drawing of a hemimandible depicting different orientations of the sub-planum area in Ekembo and Proconsul Females are denoted using shorter vectors maleswith longer vectors Vector is estimated in M 14086 which is broken at the midlineEkembo specimens have a more vertical subplanum compared to those from Songhorand Legetet but within both groups females have more vertically oriented regions thanmales Hence even the most ventrally rotated subplanum in the largest Ekembo malespecimen (KNM-RU 47805) groups with the Tinderet females in this feature
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6154
and P meswae both absolutely and in relation to crown height(Fig 12) This is true for both I1 and I2 A t-test adjusted for non-independent observations (eg I1 and I2 from the same mandible)confirmed the difference as significant (t-test df frac14 12 t frac14 258p frac14 00242) Moreover while the measurable Proconsul samplewas small this same pattern is observable in other specimenswhose tips were slightly to moderately worn and therefore notmeasured
Lower canines The only lower canine trait that discretely sepa-rates these genera is the pointed crown tip in Ekembo (Fig 3d)contrasted with a blade-like (or ldquoburinrdquo) tip in Proconsul(Fig 3c) This feature is well established for P major but is alsoexpressed in KNM-CA 1772 which was referred to P africanus byKelley (1986) and is too small to be either P major or P meswaeTwo other specimens commonly attributed to P africanus KNM-CA 2149 and KNM-SO 1112 both have broken canine tipsLikewise a new specimen (Kor 65004) attributed to P africanusby Pickford et al (2009) also has a broken tip and cannot beevaluated (see Pickford et al 2009 Fig 23) As with the uppercanines the blade-like tip is also present in the deciduous lowercanines of P major (eg KNM-CA 361 S0 5352) and P meswae(KNM-ME 1)
Lower premolars Like most previous authors (eg Andrews 1978Harrison 2002 2010 Pickford et al 2009) we found neitherqualitative nor quantitative characteristics of the lower premolarswith any taxonomic value
Lower molars Differences in lower molar morphology are few andrelatively subtle but in some cases mirror those seen in the uppermolars Cusps on the lower molars of Ekembo are more bunodontthan those in Proconsul and occupy more of the occlusal surfaceBroad inter-cuspal crests are incorporated into the Ekembo cuspsmaking them appear more inflated Molar cusps of Proconsul are
variables symphyseal height (infradentaleegnathion) planum alveolare length (mostost posterior point on the superior transverse torusegnathion) The first componentable 1) and thus PC 2 and PC 3 illustrate the residual shape variance (72 and 28
Figure 11 Scatter plot showing mandibular corpus robusticity under P4 Trend lines were computed using ordinary least squares (OLS) regression Similar relationships areobservable at other parts of the corpus but differential preservation results in smaller samples at those positions Slopes are significantly different (p lt 00001 tested on loggedvariables)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 55
more distinct from one another and are linked by narrower sharperinter-cuspal crests The lower molars of Ekembo also have asubstantially less developed buccal cingulum than those ofProconsul (Harrison 2002 2010 Harrison and Andrews 2009)
Figure 12 Scatter plot of lower incisor crown proportions Differenc
Lastly and again as seen in the upper molars the two genera canbe distinguished by the significantly greater amount of lowermolar flare in Proconsul (Fig 13 t-test df frac14 59 t frac14 713p lt 00001 see also Harrison and Andrews 2009)
es between Ekembo and Proconsul are significant (p frac14 00242)
Figure 13 Box-and-whisker plot of buccolingual flare in lower molars measured asthe average distance between the mesial buccal and lingual cusps divided by overallbuccolingual crown breadth Differences between Ekembo and Proconsul are highlysignificant (p lt 00001)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6156
Relative molar size has been used to distinguish species ofProconsul but we observed the primary difference to be in relativebuccolingual breadths of M1 and M2 In Ekembo M1 is substantiallynarrower than M2 than is the case in Proconsul (Fig 14 Harrison2002 2010) Although none of the smaller Proconsul mandiblespreserve complete crowns for both M1 and M2 KNM-SO 1112 hasan intact M2 and at least preserves the cervix for M1 However evenif the cervical measurement is used for its M1 breadthdthusunderestimating the true crown breadthdKNM-SO 1112 still has abroader first molar in relation to M2 than would be expected inEkembo If crown breadth is estimated from the ratio of cervixbreadth to maximum crown breadth in other Proconsul fossils therange of likely breadths for the SO 1112 M1 clearly distinguishes it
from the Ekembo sample (Fig 14) Testing this relationship onlogged variables using the mean estimate for the SO 1112 M1(Fig 14) reveals a significant difference between the two groups(ANCOVA df frac14 1 F frac14 2424 p frac14 00002)
As with upper third molars we find a great deal of variation inM3 morphology and mistrust the taxonomic significance some-times placed on characteristics of this toothdat least with regard tothese taxa Harrison (2002 2010) cited the mesiodistal position ofthe M3 entoconid as differentiating Kisingiri third molars fromthose attributed to P africanus We concur with his assessment butfound the entoconid position to vary in specimens attributed toP major Thus this character does not differentiate Ekembo from allProconsul species but may distinguish it from the type speciesP africanus
Discussion
The distinctiveness of the Kisingiri large catarrhine materialfrom that at Tinderet and Ugandan localities has been noted pre-viously by several authors (Pickford 1986 Walker et al 1993Begun 2001 2007 MacLatchy and Rossie 2005 Harrison andAndrews 2009 Harrison 2010) What had not been fully docu-mented was the nearly complete lack of overlap between thesesamples in a number of features despite very large assemblagesfrom Rusinga and Mfangano On descriptive criteria alone (Table 2)many isolated teeth (I1 C1 P3 M2 C1) and all of the mandibles canbe assigned to either Proconsul or Ekembowith very little chance ofmisattribution That some of the most diagnostic features occur inthe upper dentitiondthe only anatomy preserved in theP africanus type specimendadds weight to the taxonomicarrangement proposed here The morphology of the upper teeth inall Proconsul species compellingly unites them into a single cladedistinct from E heseloni and E nyanzae This is reinforced bynumerous features of the mandible and mandibular dentition thatfurther distinguish these samples The number and variety of thesefeatures are such that Proconsul and Ekembo are more differentfrom one another than are many closely-related modern catarrhinegenera
Derived morphology of Ekembo and Proconsul
It is premature to formally assign primitive and derived status tothe individual features of Ekembo and Proconsul before completinga formal phylogenetic analysis This is especially true given theabundance of evidence available for Ekembo and hence the poten-tial impact that our generic revision could have on phylogeneticinterpretations Nevertheless in comparison with both extanthominoids and the small-bodied catarrhines from the earlyMiocene it is informative to differentiate the evolutionary grades ofthese genera Many characteristics of Ekembo are more advanced inthe direction of modern apes compared to those of Proconsul(Table 2) General trends such as expanded and more bunodontcusps broader molar crests reduced cingulum and morphologicalsimilarity between P3 and P4 all represent morphologies thatpresage those of later fossil and extant hominoids The moreadvanced morphologies of P3 and M2 are particularly conspicuouscompared not only with those of Proconsul but to most earlyMiocene catarrhines In contrast the mesially positioned stronglyheteromorphic cusps on the P3 of Proconsul plus the correspondingasymmetric buccal profile notched mesial margin and missingmesial fovea are characteristic of other early Miocene catarrhinessuch as Dendropithecus Rangwapithecus and Limnopithecus andare likely primitive among East African catarrhines The moresymmetric P3 of Ekembo with midline positioned cusps andreduced cusp heteromorphy is reminiscent of later hominoids and
Figure 14 Relative buccolingual dimensions of M1 and M2 KNM-SO 1112 has a preserved M2 but only the cervix of M1 Its M1 breadth was estimated based on the ratio to cervixbreadth to maximum crown breadth in other Tinderet fossils Even the cervical breadth of SO 1112 is broader than would be predicted for the crown breadth of a Kisingiri M1 Trendlines computed using OLS regression and using the average estimate of SO 1112 M1 breadth Group differences were significant (p frac14 00002)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 57
even hominid taxa such as Rudapithecus (eg Kordos and Begun2001)
Proconsul is likewise united by a suite of features that are for themost part characteristic only of this lineage (Table 2) Themorphology of both upper and lower canines is unique to thisgenus and shared by all three well-known species plus the poorlyrepresented middle Miocene P gitongai (Pickford and Kunimatsu2005) The mandibular symphysis is also distinctive in Proconsulcompared with other catarrhines with its very long and shallowlysloping planum alveolare and steeply inclined subplanum surfaceIndeed even the small edentulous anterior mandibular fragmentKNM-SO 372 is readily placed within Proconsul and excluded fromother similarly sized catarrhine taxa such as Rangwapithecus andEkembo based on this unique symphyseal morphology (Figs 8 and9)
Other differences between Ekembo and Proconsul are of uncer-tain evolutionary polarity (eg orientation of the posthypoconecrista) or may be of taxonomic relevance without broader phylo-genetic significance (eg degree of molar flare Table 2) Werecognize that there are morphological differences within Procon-sul particularly between the better known species P africanus andP major However many of these differences are likely related tosize and represent variations on a single generic theme that aremarkedly different from the morphology of Ekembo The recentdescription (Harrison and Andrews 2009) of the intermediatelysized P meswae underscores the broad morphological unity withinProconsul and further substantiates a well-defined Proconsul cladecomprising Tinderet and Ugandan species
Based on the abundant morphological and morphometric evi-dence supporting genus-level distinction between Proconsul andEkembo we are currently conducting a more detailed phylogeneticanalysis of all early Miocene catarrhines in order to better under-stand the distribution and evolutionary significance of these andother morphological features We anticipate that the debates sur-rounding the phylogenetic relationships of an historically consti-tuted sample of Proconsul can be more satisfactorily resolved nowthat the Kisingiri material is appropriately recognized as a distinctgenus
The generic disposition of ldquoUgandapithecusrdquo species
A great many features said to characterize ldquoUgandapithecusrdquowere added to the original diagnosis of the genus by Pickford et al(2009) in their systematic revision of Proconsul (sensu lato) As partof that study they also erected a new species ldquoUrdquo legetetensis andtransferred P meswae to ldquoUgandapithecusrdquo Evaluating the same setof features across the entire available sample however we wereunable to find support for this more expansive conception ofldquoUgandapithecusrdquo for several reasons First some diagnostic fea-tures cited in Pickford et al (2009) were quite variable within theldquoUgandapithecusrdquo sample As an example marked reduction of theM3 entoconid to a bead on the lingual cingulum is not shared byseveral ldquoUrdquo major specimens such as KNM-CA 393 NAP I 49000and Kor 253004 (see Pickford et al 2009 Fig 33e) and was likelynot a feature of a few of the more worn specimens eg BUMP 600and BUMP 601 or even the holotype of P major M 16648 Second
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6158
some diagnostic features of ldquoUgandapithecusrdquo were found to bevariably present in Ekembo arguing against their utility for dis-tinguishing ldquoUgandapithecusrdquo from Pickford et als (2009) com-bined P africanus thorn Kisingiri conception of Proconsul For examplesignificantly reduced curvature of the I1 distal margin in ldquoUganda-pithecusrdquo is also found in some Kisingiri specimens including KNM-RU 1685 1712 1714 1831 1846 and 1951 several more Rusingaspecimens have moderately reduced curvature Third some fea-tures that purportedly distinguish ldquoUgandapithecusrdquo fromP africanus are likely related to size For example the somewhatmore central position of the P3 paracone in ldquoUrdquomajor is not seen inthe smaller ldquoUrdquo meswae whose paracone is positioned similarly tothat of the holotype of P africanus (see Harrison and Andrews2009 Fig 6d)
However the major part of the evidence for recognizingldquoUgandapithecusrdquo (sensu Pickford et al 2009) was assembled by re-assigning specimens traditionally attributed to P africanus into newspecies of ldquoUgandapithecusrdquo The P africanus holotype was left inassociation with the Rusinga sample despite having morphologyoverwhelmingly characteristic of other Tinderet specimens Theresulting specimen allocations left P africanus morphologicallyimpoverisheddstripped of much of its hypodigmdso that Pro-consul for Pickford et al (2009) came to be characterized primarilyby the Rusinga sample (see similar comments in Harrison andAndrews 2009) Within that conceptual framework any spec-imen that differs from those at Rusinga could not be Proconsul butmust instead belong to ldquoUgandapithecusrdquo Thus specimens such asthe KNM-SO 1112 mandible were erroneously assigned to ldquoUgan-dapithecusrdquo because they were (correctly) perceived as differingfrom their Rusinga counterparts (Pickford et al 2009)
Here we restore the P africanus hypodigm and associate it withthe other Tinderet fossils according to their shared morphologySpecimens assigned to ldquoUgandapithecusrdquo revert to Proconsul andmany of the features cited to support a speciose ldquoUgandapithecusrdquo(Pickford et al 2009) instead become diagnostic features of Pro-consul This correction serves to highlight the distinctiveness of theKisingiri remains and the need to recognize this distinctiveness atthe generic level
As demonstrated above specimens from Kisingiri do not displaythe morphology of Proconsul as represented by the holotype ofP africanus M 14084 Given that this specimen preserves only theupper canine and cheek teeth similarities with or departures fromthis morphology represent the most compelling arguments forassessing inclusivity in the genus However for the mandible andlower dentition if one simply sorts the specimens by elementwithout relying on taxonomic preconceptions the overwhelmingdistinction within the traditionally constituted Proconsul sample isbetween Kisingiri material on the one hand and that from Tinderetand Uganda on the other (cf Walker et al 1993 Begun 2001 2007MacLatchy and Rossie 2005 Harrison and Andrews 2009Harrison 2010) Additionally with the Kisingiri specimensremoved to Ekembo the postcranial differences used to partlyjustify a distinct ldquoUgandapithecusrdquo lose their relevance (seeHarrison and Andrews 2009)
We have differed among ourselves in our past treatment ofP major as generically distinct from the smaller species of Pro-consul (Begun 2007 McNulty 2010) Morphologically many ofthe unique features of P major can be interpreted as size-relatedvariants on the smaller P africanus morphology Even if so thequestion is whether these are sufficiently different and numerousto warrant generic distinction that is do they suggest thatP major belongs in a separate clade from P africanus andP meswae As we have demonstrated differences among thesethree species are certainly fewer and show less morphologicaldistinction than those between Ekembo and any of the three
Nevertheless while we ultimately agree with Harrison andAndrews (2009 see also Begun 2007) that the features dis-tinguishing ldquoUrdquo major from P africanus are consistent with thosethat distinguish congeneric species and are at least partly due todifferences in size we concede the possibility that the largestProconsul species could belong in a separate genus (Senut et al2000 Pickford and Kunimatsu 2005) However whether or notone recognizes a separate genus for P major the P africanus ma-terial does not sort morphologically with the Kisingiri materialTherefore the taxonomic alternatives are either two generaProconsul and Ekembo or three Proconsul Ugandapithecus andEkembo with Proconsul and Ugandapithecus united in a cladeexclusive of Ekembo Thus while we strongly favor the firstalternative recognizing Ugandapithecus does not change thefundamental argument concerning the need to refer the Kisingirispecies to a generically and cladistically distinct taxon from thespecies represented at Tinderet and the Ugandan sites
Specimens of interest
Despite the substantial dental and gnathic differences betweenProconsul and Ekembo there are a few specimens whose taxonomicaffinities require additional scrutiny
KNM-RU 1676-77 This set of associated upper and lower teeth is aconsistent outlier among E heseloni and E nyanzae specimens Insome ways this specimen more closely resembles Tinderet Pro-consul in the features that distinguish it from Ekembo Molar cuspmorphology and cingulum development I1 lingual tubercledevelopment and M2 shape and posthypocone crista orientationare more consistent with specimens from the Tinderet sites On theother hand it otherwise resembles Ekembo and lacks the strongestdiagnostic features that unite species of Proconsul such as thedistinctive canine and P3 morphology Overall this specimen doesnot fit unambiguously into either genus in fact the degree ofbuccolingual flare cusp morphology enamel wrinkling and espe-cially the long extension of enamel onto the anterior root in P3 arein some ways reminiscent of Afropithecus although it clearly differsfrom that genus as well (cf Rossie and MacLatchy 2013)
Were these characteristics individually distributed among thelarge Kisingiri sample one might attribute this to intra-genericvariation but when found together in a single specimen from alarge sample it suggests a unique individual One of us (KPM)thinks it likely that the specimen was not actually found at aKisingiri locality based on its unique preservation compared toother Rusinga fossils and on other examples of mis-proveniencedspecimens (Leakey 1967 Andrews and Molleson 1979 Pickford1986 Kelley 1986) However we recognize that relying toofrequently on this explanation allows almost any scenario to bejustified and suggest some alternative possibilities One is that thespecimen was found on Rusinga but in the much older WayandoFormation strata instead of the Hiwegi Formation The differencesin morphology in that case might reflect more than a million yearsof temporal change (McCollum et al 2013) with KNM-RU 1676-77preserving certain more primitive Proconsul-like features As analternative it is reasonable to assume that fossil assemblages asvast as those from Rusingawill include not only abundant taxa butalso rare or infrequent inhabitants of the local biome There aremany examples of this from Rusingas mammalian communityeg ldquoHemicyonrdquo Kulutherium Tadarida cf Hyainailouros cfArchaeobeledon Kelba (see relevant chapters in Werdelin andSanders 2010) As such KNM-RU 1676-77 may simply representa rare and well-preserved specimen of some other catarrhinetaxon that lived among or visited a primate community dominatedby Ekembo
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 59
KNM-SO 1112 This specimen was initially referred to Rangwapi-thecus but soon after was transferred to Proconsul afrianus (Bosler1981 Martin 1981)da designation that most subsequent authorshave followed (Pickford 1986 Nengo and Rae 1992 Kelley 19861995 Walker et al 1993 MacLatchy and Rossie 2005 Hill et al2013) Pickford et al (2009) more recently referred this specimento ldquoUrdquo meswae citing two primary reasons First they reiteratedBoslers (1981) concern that its P3 differs from others attributedto P africanus However at that time the majority of what wasconsidered to be P africanus would have been from RusingaIsland In fact while noting its contrasting morphology Boslersuggested that differences between KNM-SO 1112 and other P3smight reasonably be attributed to sexual dimorphism notingthat this specimen provides ldquoa not implausible match for the Pafricanus holotype maxilla M 14084rdquo (1981152) Thatassessment is borne out in this study Using the much largersample of Ekembo P3s as a model variation can be seen toencompass specimens with well-developed mesial beaks andthose with more bilaterally compressed crowns and parallelsides The latter morphology is predominantly found amongmale specimens (KNM-RU 1674 1924 and KPS I and III) thussupporting Boslers (1981) suggestion that this variation isrelated to sexual dimorphism and hence that KNM-SO 1112 is amale P africanus
The second feature cited by Pickford et al (2009) in referringthis specimen to ldquoUrdquo meswae was posterior shallowing of themandibular corpus However because they redefined ldquoUgandapi-thecusrdquo to include all of the well-preserved Tinderet mandibles(Pickford et al 2009)dleaving Proconsul to be represented only byRusinga specimensdtheir only alternative within this scheme for aspecimen the size of SO 1112 was to refer it to ldquoUrdquomeswae It seemsodd though to assign this specimen to a species from a (perhapsmuch) older site when the Proconsul africanus holotype is ofappropriate size and similar age and as there are no other adultmandibles from either species with which to compare it In anycase mandibular shallowing is variable in both Ekembo and Pro-consul with some Kisingiri specimens exhibiting considerableshallowing (eg KNM-RU 2087) and some ldquoUgandapithecusrdquo spec-imens having very little (eg M 14086)
We therefore see no morphological grounds to assign KNM-SO1112 to P meswae but do find some morphological evidence toassign it as a male P africanus which occurs at Songhor
KNM-LG 452 This very complete mandible was originally recog-nized as a female of P major based on the size of the dentition androbusticity of the mandible (Martin 1981) Bosler (1981) howeverleft this specimen unassigned noting similarities to P major in P3-M1 size but having distinctly smaller M2e3 In this she likenedKNM-LG 452 to a few other specimens (notably M 14086) thatare smaller in their distal molars in a manner more consistentwith taxonomic rather than sex differences (Bosler 1981 see alsoPilbeam 1969) Kelley (1986 1995) also assigned this specimen asa female of P major based on canine size and morphology butnoted that the canine lacked the shallowly-sloping mesial ridgetypical of most female anthropoid canines This is also truehowever of two other canines assigned as female P major KO 9and SO 373 (Kelley 1986) and so might simply be an unusualcharacteristic of P major female lower canines In the end Kelley(1995) concluded that considering all of the criteria fordistinguishing male and female canines the evidence that KNM-LG 452 is female remains questionable
Pickford et al (2009) further developed the idea that two spe-cies are represented within the P major sample and made KNM-LG452 the holotype of a new species ldquoUrdquo legetetensis We concur thatthis specimen is unique among the well-preserved Tinderet
mandibles in some important features Whereas male and femalemandibles of P major have long internal symphyseal planesexpansive extra- and retromolar sulci and rami that cross theocclusal plane at distal M3 KNM-LG 452 has a shorter symphysealplane (though not to the extent seen in Ekembo) a moderateextramolar sulcus but no retromolar sulcus and a ramus thatcrosses the occlusal plane more mesially If KNM-SO 396 and SO404 represent males and females respectively of P major then themorphology of KNM-LG 452 appears distinct
At present however we do not think there is sufficient evidenceto differentiate this specimen from either P major or P meswaePickford et al (2009) argue that KNM-LG 452 cannot belong to thelatter species because of its larger size The size differences how-ever are well within the range of variation seen in P majorE heseloni and E nyanzae Moreover whereas the Legetet mandibleis larger than most of the P meswae hypodigm an adult P4 referredto this species (KNM-ME 25) substantially exceeds its counterpartin KNM-LG 452 Should the Legetetmandible ultimately prove to bea male specimen size alone would not rule out its attribution toP meswae On the other hand if it is female then it becomes harderto justify its inclusion in that sample of mostly smaller specimens Iffemale therefore it could represent a separate species for whichthe binomial Proconsul legetetensis comb nov is available Recentlycollected fossils from the SonghorKapurtay area provide someadditional support for the possibility that three species of Proconsulexisted at these localities (McNulty 2014ab) However toconvincingly demonstrate this and determine whether a thirdspecies can be differentiated from P meswae will require a moredetailed analysis of variation among Proconsul species a corre-sponding study of variation in modern taxa anddalmost certain-lydadditional fossil specimens For now we regard ldquoUrdquo legetetensisas a subjective junior synonym of P major
Conclusions
The genus Proconsul has been central to studies of ape and hu-man evolution since its first description by Hopwood (1933) As oneof the best known fossil catarrhines represented by nearly everyskeletal element our knowledge of Proconsuls functional anatomypositional behavior and overall morphology has made it thefoundation for paleoanthropological research in the early Mioceneas well as studies on the origin and early diversification of homi-noids With respect to systematics however Proconsul is less wellunderstood the type species P africanus is known from a rela-tively small number of specimens from sites in the Tinderet areaand no postcranial bones can be unambiguously assigned to itRather it is the copious and well-preserved specimens from theKisingiri sites of Rusinga and Mfangano Islands that have formedthe basis for the common image of Proconsul
We argue here that the Kisingiri species traditionally referredto ProconsuldP heseloni and P nyanzaedare generically distinctfrom P africanus and hereby transfer them into a new genusEkembo based on a substantial number of morphological differ-ences (Table 2) Considering only those traits preserved in theP africanus holotype the entire sample from Rusinga and Mfan-gano differs markedly in the morphology of the upper canine P3P4 and M2 Additional differences in the I1 lower incisors andcanines molar cusp and occlusal crest morphology degree of flarein all molars and especially mandibular features clearly distin-guish the Kisingiri from non-Kisingiri samples in a way that isincompatible with congeneric species With the transfer ofE heseloni and E nyanzae Proconsul becomes a well-defined cladecomprising at least three speciesdP africanus P major andP meswaedunited by several features that are unique for thegenus (Table 2)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6160
We found no support in this study for the genus ldquoUgandapi-thecusrdquo as constructed by Pickford et al (2009) In that paper theytransferred the vast majority of Proconsul fossils from the Tinderetand Napak localities into ldquoUgandapithecusrdquo leaving P africanusrepresented only by its holotype and a small number of isolatedteeth and aligning it with the material from Rusinga and Mfan-ganoWhen the P africanus hypodigm is reconstituted and properlyaffiliated with the other Tinderet species according to itsmorphology much of the evidence marshaled by Pickford et al(2009) for ldquoUgandapithecusrdquo actually supports the distinction be-tween the Tinderet and Kisingiri samples ie the generic distinc-tion of Ekembo In addition some of the traits enumerated byPickford and colleagues in support of ldquoUgandapithecusrdquo wererevealed in our analysis to be variable within one or more speciesand therefore of questionable taxonomic utility We do not rejectout of hand the original conception of ldquoUgandapithecusrdquo (Senutet al 2000) as either a monotypic genus distinguishing ldquoUrdquo ma-jor or perhaps including a closely allied middle Miocene ldquoUrdquogitongai as well (Pickford and Kunimatsu 2005) but we have fornow retained the early Miocene specimens of both in Proconsulpending further study However even if Ugandapithecus is a taxo-nomically valid genus comprising just the latter two species theirmorphology indicates that theywould still be united in a cladewithProconsul and they lack several more advanced features that partlydefine Ekembo Better understanding of the role that body size playsinmediating differences within Proconsulwill help to further clarifythe relationships among these species
Despite the existence of one well-represented and several lesscomplete skeletons of what was long assumed to be Proconsul thephylogenetic status of this genus has defied consensus This islargely due in our view to Proconsul having been an admixture oftwo distinct genera one more advanced in the direction of extantapes than the other As we demonstrate here the material fromRusinga and Mfangano traditionally referred to Proconsul andwhich includes all of the more complete skeletal material isdistinct from the Tinderet and Napak material and belongs in aseparate genus Ekembo By disentangling these distinct clades weanticipate being able to achieve better phylogenetic resolution forboth genera thereby improving our knowledge of early hominoidevolution and diversification
Acknowledgments
We thank many people and agencies that assisted with thedevelopment and completion of this project This research wasinitiated as a result of funding from the Wenner-Gren Foundationand the Johns Hopkins School of Medicine to DR Begun in1987e1989 and was renewed following research and travelgenerously funded by multiple Leakey Foundation grants to KMcNulty National Science Foundation grants to K McNulty(0852609 1241807) the McKnight Foundation the University ofMinnesota the Lloyd A Wilford Endowment the LeverhulmeFoundation and the Turkana Basin Institute J Kelley gratefullyacknowledges support from the Institute of Human Origins Ari-zona State University DR Begun also wishes to acknowledge theAlexander Humboldt Stiftung and NSERC for funding support Weare grateful to the National Museums of Kenya the UgandaMuseum the Natural History Museum and to Rose Mwanja EzraMusiime Tom Mukhuyu Samuel Muteti James Yatich and MaryMuungu for access to and assistance with specimens in their careMany people provided assistance comments criticisms insightsand suggestions on various aspects of this work and we especiallythank Holly Dunsworth Will Harcourt-Smith Tom Lehmann DanPeppe Nicole Garrett Kirsten Jenkins Rutger Jansma Susy CoteIsaiah Nengo Andrew Hill and Brenda Benefit K McNulty
acknowledges the evolutionary anthropology faculty and studentsat Durham University for their helpful comments on an earlyversion of this manuscript and for their gracious hospitality Weare likewise grateful to the anonymous reviewers associate editorand editor whose comments strengthened the paper This manu-script is publication 1 supporting Research on East AfricanCatarrhine and Hominoid Evolution (REACHE)
Appendix A Supplementary data
Supplementary data related to this article can be found at httpdxdoiorg101016jjhevol201503009
References
Andrews P 1974 New species of Dryopithecus from Kenya Nature 249 188e190Andrews P 1978 A revision of the Miocene Hominoidea of East Africa Bull Brit
Mus (Nat Hist) Geol 30 85e225Andrews P 1985 Family group systematics and evolution among catarrhine pri-
mates In Delson E (Ed) Ancestors The Hard Evidence Liss New Yorkpp 14e22
Andrews P Harrison T Martin L Pickford M 1981 Hominoid primates from anew Miocene locality named Meswa Bridge in Kenya J Hum Evol 10123e128
Andrews P Begun D Zylstra M 1997 Interrelationships between functionalmorphology and paleoenvironments in Miocene hominoids In Begun DWard C Rose M (Eds) Function Phylogeny and Fossils Miocene HominoidEvolution and Adaptations Plenum Press New York pp 29e58
Andrews P Molleson T 1979 The provenance of Sivapithecus africanus Bull BritMus (Nat Hist) Geol 32 19e23
Begun DR 2001 African and Eurasian Miocene hominoids and the origins of theHominidae In de Bonis L Koufos GD Andrews P (Eds) Hominoid Evolutionand Climatic Change in Europe Phylogeny of the Neogene Hominoid Primatesof Eurasia Cambridge University Press London pp 231e253
Begun DR 2007 Fossil record of Miocene hominoids In Henke W Tattersall I(Eds) Handbook of Paleoanthropology Springer Berlin pp 921e977
Begun DR Teaford MF Walker AC 1994 Comparative and functional anatomyof Proconsul phalanges from the Kaswanga primate site Rusinga Island KenyaJ Hum Evol 26 89e165
Bishop WW 1964 More fossil primates and other Miocene mammals from North-East Uganda Nature 203 1327e1331
Bishop WW Miller JA Fitch FJ 1969 New potassium-argon age determinationsrelevant to the Miocene fossil mammal sequence in East Africa Am J Sci 267669e699
Blankenship JA Mincer HH Anderson KM Woods MA Burton EL 2007Third molar development in the estimation of chronologic age in Americanblacks as compared with whites J Forensic Sci 52 428e433
Bosler W 1981 Species groupings of early Miocene dryopithecine teeth from EastAfrica J Hum Evol 10 151e158
Coleman MN 2008 What does geometric mean mean geometrically Assessingthe utility of geometric mean and other size variables in studies of skullallometry Am J Phys Anthropol 135 404e415
Cote S Malit N Nengo I 2013 Additional mandibles of Rangwapithecus gordonian early Miocene catarrhine from the Tinderet localities of Western KenyaJ Hum Evol 153 341e352
Deane AS 2009 Early Miocene catarrhine dietary behavior the influence of theRed Queen Effect on incisor shape and curvature J Hum Evol 56 275e285
Drake R Van Couvering JA Pickford M Curtis G Harris JA 1988 New chro-nology for the Early Miocene mammalian faunas of Kisingiri Western KenyaJ Geol Soc Lond 145 479e491
Evernden JF Savage DE Curtis GH James GT 1964 Potassium-argon datesand the Cenozoic mammalian chronology of North America Am J Sci 262145e198
Gebo DL MacLatchy L Kityo R Deino A Kingston J Pilbeam D 1997A hominoid genus from the Early Miocene of Uganda Science 276 401e404
Geoffroy Saint-Hilaire E 1812 Tableau des quadrumanes 1 Ord QuadrumanesAnnls Mus Hist Nat Paris 19 85e122
Gray JE 1825 Outline of an attempt at the disposition of the Mammalia into tribesand families with a list of the genera apparently appertaining to each tribeAnnls Philos 10 337e344
Harrison T 1982 Small-bodied apes from the Miocene of East Africa PhDDissertation University of London
Harrison T 1986 New fossil anthropoids from the middle Miocene of East Africaand their bearing on the origin of the Oreopithecidae Am J Phys Anthropol 71265e284
Harrison T 1987 The phylogenetic relationships of the early catarrhine primates areview of the current evidence J Hum Evol 16 41e80
Harrison T 2002 Late Oligocene to middle Miocene catarrhines from Afro-ArabiaIn Hartwig WC (Ed) The Primate Fossil Record Cambridge University PressCambridge pp 311e338
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 61
Harrison T 2010 Dendropithecoidea Proconsuloidea and Hominoidea (CatarrhiniPrimates) In Werdelin L Sanders WJ (Eds) Cenozoic Mammals of AfricaUniversity of California Press Berkeley pp 429e469
Harrison T Andrews P 2009 The anatomy and systematic position of a newspecies of early Miocene proconsulid from Meswa Bridge Kenya J Hum Evol56 479e496
Hill A Nengo IO Rossie JB 2013 A Rangwapithecus gordoni mandible from theearly Miocene site of Songhor Kenya J Hum Evol 65 490e500
Hopwood AT 1933 Miocene primates from Kenya J Linn Soc Zool 38 437e464Ishida H Kunimatsu Y Takano T Nakano Y Nakatsukasa M 2004 Nacholapi-
thecus skeleton from the Middle Miocene of Kenya J Hum Evol 46 69e103Jolicouer P 1963 The multivariate generalization of the allometry equation Bio-
metrics 19 497e499Jungers WL Falsetti AB Wall CE 1995 Shape relative size and size-
adjustments in morphometrics Am J Phys Anthropol 38 (S2) 137e161Kavanagh KD Evans AR Jernvall J 2007 Predicting evolutionary patterns of
mammalian teeth from development Nature 449 427e433Kelley J 1986 Species recognition and sexual dimorphism in Proconsul and
Rangwapithecus J Hum Evol 15 461e495Kelley J 1995 Sex determination in Miocene catarrhine primates Am J Phys
Anthropol 96 390e417Kelley J 1993 Taxonomic implications of sexual dimorphism in Lufengpithecus In
Kimbel WH Martin LB (Eds) Species Species Concepts and Primate Evo-lution Springer New York pp 429e458
Kent PE 1942 The country round the Kavirondo Gulf of Victoria Nyanza Geogr J100 22e31
Kordos L Begun DR 2001 A new cranium of Dryopithecus from RudabanyaHungary J Hum Evol 15 689e700
Le Gros Clark WE Leakey LSB 1950 Diagnoses of East African Miocene Homi-noidea Q J Geol Soc Lond 105 260e263
Le Gros Clark WE Leakey LSB 1951 The Miocene Hominoidea of East Africa InFossil Mammals of Africa No 1 British Museum (Natural History) London
Leakey LSB 1943 A Miocene anthropoid mandible from Rusinga Kenya Nature152 319e320
Leakey LSB 1963 East African fossil Hominoidea and the classification within thissuper-family In Washburn SL (Ed) Classification and Human EvolutionAldine Chicago pp 32e49
Leakey LSB 1967 Notes on the mammalian faunas from the Miocene and Pleis-tocene of East Africa In Bishop WW Clark JD (Eds) Background to Evolu-tion in Africa University of Chicago Press Chicago pp 7e29
Leakey RE Leakey MG 1986 A new Miocene hominoid from Kenya Nature 324143e146
Leakey RE Leakey MG Walker AC 1988 Morphology of Afropithecus turka-nensis from Kenya Am J Phys Anthropol 76 289e307
Leakey MG Ungar PS Walker A 1995 A new genus of large primate from theLate Oligocene of Lothidok Turkana District Kenya J Hum Evol 28519e531
Linnaeus C 1758 Systema naturae per regna tria naturae secundum classesordines genera species cum characteribus differentris synonymis locis tenthed Laurentii Salvii Stockholm
MacInnes DG 1943 Notes on the East African Miocene primates J E Afr UgandaNat Hist Soc 17 141e181
MacLatchy L 2004 The oldest ape Evol Anthropol 13 90e103MacLatchy L Bossert WH 1996 An analysis of the articular surface distribution of
the femoral head and acetabulum in anthropoids with implications for hipfunction in Miocene hominoids J Hum Evol 31 425e453
MacLatchy L Rossie JB 2005 The Napak hominoid still Proconsul major InLieberman DE Smith RJ Kelley J (Eds) Interpreting the Past Essays onHuman Primate and Mammal Evolution in Honor of David Pilbeam Brill Ac-ademic Publishers Boston pp 15e28
Martin L 1981 New specimens of Proconsul from Koru Kenya J Hum Evol 10139e150
McCollum MS Peppe DJ McNulty KP Dunsworth HM Harcourt-Smith WEH Andrews AL 2013 Magnetostratigraphy of the early MioceneHiwegi Formation (Rusinga Island Lake Victoria Kenya) Geological Society ofAmerica Abstracts with Programs 45 p 12
McNulty KP 2010 Apes and tricksters the evolution and diversification ofhumans closest relatives Evol Edu Outreach 3 322e332
McNulty KP Harcourt-Smith WEH Dunsworth HM 2007 New primate fossilsfrom Rusinga Island Kenya Am J Phys Anthropol 132 (S44) 170
McNulty KP MacLatchy L Peppe DJ Nengo I Manthi FK Miller ERStevens NJ Cote S Lehmann T Kingston J 2014a A regional approach toEast African early Miocene paleobiology J Vert Paleontol 185 SVP Programand Abstracts Book 2014
McNulty KP MacLatchy L Rossie JB Peppe DJ Deino AL Mbua ENManthi FK Nengo IO Stevens NJ Cote S Lehmann T Gutierrez M 2014b
Research on East African catarrhine and hominoid evolution results from thefirst year Am J Phys Anthropol 153 (S58) 182
Michel LA Peppe DJ Lutz JA Driese SG Dunsworth HM Harcourt-Smith WEH Horner W Lehmann T Nightengale S McNulty KP 2014Remnants of an ancient forest provide ecological context for Early Miocenefossil apes Nature Comm 5 3236
Myres JL 1932 Africa Man 32 208Nakatsukasa M Kunimatsu Y Nakano Y Takano T Ishida H 2007 Vertebral
morphology of Nacholapithecus kerioi based on KNM-BG 35250 J Hum Evol52 347e369
Nengo IO Rae TC 1992 New hominoid fossils from the early Miocene site ofSonghor Kenya J Hum Evol 23 423e429
Peppe DJ McNulty KP Cote SM Harcourt-Smith WEH Dunsworth HM VanCouvering JA 2009 Stratigraphic interpretation of the Kulu Formation (EarlyMiocene Rusinga Island Kenya) and its implications for primate evolutionJ Hum Evol 56 447e461
Pickford M 1981 Preliminary Miocene mammalian biostratigraphy for westernKenya J Hum Evol 10 73e97
Pickford M 1986 Sexual dimorphism in Proconsul Hum Evol 1 111e148Pickford M Andrews PJ 1981 The Tinderet Miocene sequence in Kenya J Hum
Evol 10 11e33Pickford M Kunimatsu Y 2005 Catarrhines from the Middle Miocene (ca 145
Ma) of Kipsaraman Tugen Hills Kenya Anthropol Sci 113 189e224Pickford M Senut B Gommery D Musiime E 2009 Distinctiveness of Ugan-
dapithecus from Proconsul Estudios Geol 65 183e241Pilbeam DR 1969 Tertiary Pongidae of East Africa evolutionary relationships and
taxonomy Bulletin of the Peabody Museum of Natural History 31 pp 1e185Rae TC 1993 Phylogenetic analysis of proconsulid facial morphology PhD
Dissertation State University of New York at Stony BrookRae TC 1999 Mosaic evolution in the origin of the Hominoidea Folia Primatol 70
125e135Rafferty KL Walker A Ruff C Rose MD Andrews PJ 1995 Postcranial esti-
mates of body weights in Proconsul with a note on a distal tibia of P major fromNapak Uganda Am J Phys Anthropol 97 391e402
Rossie JB MacLatchy L 2013 Dentognathic remains of an Afropithecus individualfrom Kalodirr Kenya J Hum Evol 65 199e208
Senut B Pickford M Gommery D Kunimatsu Y 2000 Un nouveau genredrsquohominoiumlde du Miocene inferieur drsquoAfrique orientale Ugandapithecus major(Le Gros Clark and Leakey 1950) CR Acad Sci Paris 331 227e233
Simons EL Pilbeam DR 1965 Preliminary revision of the Dryopithecinae (Pon-gidae Anthropoidea) Folia Primatol 3 81e152
Smith TM Martin LB Leakey MG 2003 Enamel thickness microstructure anddevelopment in Afropithecus turkanensis J Hum Evol 44 283e306
Stevens NJ Seiffert ER OConnor PM Roberts EM Schmitz MD Krause CGorscak E Ngasala S Hieronymus TL Temu J 2013 Palaeontological evi-dence for an Oligocene divergence between Old World monkeys and apesNature 497 611e614
Suwa G Kono RT Katoh S Asfaw B Beyene Y 2007 A new species of great apefrom the late Miocene epoch in Ethiopia Nature 448 921e924
Szalay FS Delson E 1979 Evolutionary History of the Primates Academic PressNew York
Teaford MF Beard KC Leakey RE Walker A 1988 New hominoid facial skel-eton from the Early Miocene of Rusinga Island Kenya and its bearing on therelationship between Proconsul nyanzae and Proconsul africanus J Hum Evol17 461e477
Van Couvering JAH Miller J 1969 Miocene stratigraphy and age determinationsRusinga Island Kenya Nature 221 628e632
Walker A 1997 Proconsul function and phylogeny In Begun DR Ward CVRose MD (Eds) Function Phylogeny and Fossils Miocene Hominoid Evolu-tion and Adaptation Plenum Press New York pp 209e224
Walker A Pickford M 1983 New postcranial fossils of Proconsul africanus andProconsul nyanzae In Ciochon RL Corruccini RS (Eds) New Interpretationsof Ape and Human Ancestry Plenum Press New York pp 325e351
Walker A Teaford M 1988 The Kaswanga primate site an Early Miocene hom-inoid site on Rusinga Island Kenya J Hum Evol 17 539e544
Walker A Teaford M 1989 The hunt for Proconsul Sci Am 260 76e82Walker A Teaford MF Martin L Andrews P 1993 A new species of Proconsul
from the early Miocene of RusingaMfangano Islands Kenya J Hum Evol 2543e56
Ward CV Ruff CB Walker A Rose MD Teaford MF Nengo IO 1995 Func-tional morphology of Proconsul patellas from Rusinga Island Kenya with im-plications for other Miocene-Pliocene catarrhines J Hum Evol 29 1e19
Ward S Brown B Hill A Kelley J Downs W 1999 Equatorius a new hominoidgenus from the Middle Miocene of Kenya Science 285 1382e1386
Werdelin L Sanders WJ 2010 Cenozoic Mammals of Africa University of Cali-fornia Press Berkeley

Figure 3 Canines of Proconsul and Ekembo a) Upper canine of the P africanus holotype (M 14084) lingual view b) Upper canine of P major (KNM-CA 2127) lingual view c) Lowercanine of P major buccal view d) lower canine of Ekembo cf nyanzae (KNM-RU 1676) buccal view and e) Upper canine of P major (KNM-SO 584) distal view Note the blade-like orldquoburin-likerdquo tip of the unworn Proconsul canines including on the nearly unworn canine tip of the P africanus type specimen (a) Ekembo canines have more typical pointed tipsProconsul canines also have longitudinal grooves and fine striations on the root (e) Ekembo lacks these features Scale bars frac14 1 cm
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 49
striations near the root apex that extend to varying degrees towardthe cervix of the tooth Longitudinal ldquoflutingrdquo consists of broadgrooves that segment the root into distinct sections (cf Pickfordet al 2009 Fig 3e) Most often the root in cross-section has theappearance of a cylinder that has been sectioned longitudinallywith the two halves offset from each other but in some cases thereare three segments and the root cross-section is more triangular inappearance (Kelley 1986) This characteristic was noted by Kelley(1986) in the P africanus canines but not in those of P majorHere we affirm that fluting is present on all of the visible uppercanine roots attributed to both P africanus and P major rootmorphology could not be evaluated in the deciduous upper caninefrom Meswa Bridge since it is implanted in the maxillary alveolusNone of the Ekembo specimens display this morphology havinginstead a simpler round to ovoid cross-section along the length ofthe root
The fine longitudinal striations on Proconsul upper canine rootsvary in the degree to which they extend toward the cervix (Fig 3e)but they are visible in all specimens attributed to P africanus(KNM-SO 419 521 921 5353 KNM-X 502) and five of six uppercanines of P major (excepting KNM-SO 1614) This morphology isalso present in UMP 62-12 which was included as part of theMorotopithecus bishopi hypodigm (Gebo et al 1997) Fine striationsalong the upper canine root are almost entirely absent in EkemboTwenty of twenty-three specimens lack this feature three speci-mens (KNM-RU 1763 1845 1891) exhibit incipient to slight
striations but these are never developed to the extent seen inProconsul canines
Upper premolars Upper third premolars display some of thegreatest differences between Ekembo and Proconsul P4s tend todiffer in similar ways but the differences are not nearly as markedEkembo P3s have cusps positioned near the mesiodistal midpoint ofthe crown well-developed mesial foveae pre- and postparacristaeof similar length and slope resulting in symmetric or nearsymmetric buccal profiles a straight or slightly curved mesialmargin and only moderately curved distal margin (Fig 4)Proconsul P3s are more similar in appearance to those of small-bodied catarrhines from the early Miocene with cusps positionedmesially on the crown greatly reducing or eliminating the mesialfovea They also have a steep preparacrista and more shallowlysloped postparacrista resulting in a highly asymmetric buccalprofile and an inwardly angled or notched mesial margin but astrongly curved distal margin This morphology is bestcharacterized in the P africanus holotype (M 14084) but can alsobe seen in other specimens attributed to this species as well asthose referred to P major (Fig 4d e) Also noteworthy is adistinctive mesiobuccal flange that mesially extends the buccalportion of the crown but only near the cervical line of the toothPickford et al (2009) describe this feature in some detail forldquoUgandapithecusrdquo but in fact this morphology is clearly expressedin the P africanus holotype as well (Fig 4a d) A single adult P3
Figure 4 Upper premolars of Proconsul and Ekembo occlusal (aec) and lingual (def) views a) P3e4 of the P africanus holotype (M 14084) b) P3 of P major (M 14331) c) P3e4 of theE heseloni holotype (KNM-RU 2036) d) P3 of the P africanus holotype (M 14084) e) P3 of P major (M 14331) f) P3 of the E heseloni holotype (KNM-RU 2036) Note the mesiallypositioned cusps lack of mesial fovea and strongly asymmetric crown in Proconsul compared to Ekembo (see text for further comparisons) Image of M 14331 reversed for easiercomparison Photo 4a courtesy of Rutger Jansma Scale bars frac14 1 cm
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6150
germ and two dP3s attributed to P meswae look remarkably similarto those of other Proconsul species including the presence of thisflange In contrast the Ekembo P3 has a well-developed mesialfovea that terminates at a mesiobuccal shoulder positionedapproximately halfway between the paracone tip and the cervixFrom this shoulder the mesial margin angles slightly distallydown to the cervix (Fig 4f)
The degree of cusp height heteromorphy in P3 also differentiatesEkembo from Proconsul (contra Pickford et al 2009) Only four non-Kisingiri specimens were sufficiently unworn to take measure-ments but more worn specimens also appear to express this dif-ferential in cusp height Figure 5a shows the ratio of protoconeparacone height measured from the tooth cervix and with only oneexception Ekembo P3s have substantially less cusp heteromorphyThis exception KNM-RU 1677 does not in fact have greater heter-omorphy but instead has an abnormal extension of the enamel ontothe buccal (paracone) root which skews the measurement Hencecusp relief relative to the occlusal surface in RU 1677 is not nearly asheteromoprhic as in Proconsul and when this is taken into accountthe difference between genera is significant (t-test dffrac14 11 t frac14 283p frac14 00165) The adult P3 germ from Meswa Bridge is incompletelyformed and therefore could not be measured However the differ-ence between paracone and protocone heights with respect to theocclusal surface appears to be as great as that in the P africanusholotype It does not appear that sex is an important determinant ofthis feature since well-established male (eg M16647 KNM-RU16000) and female (KNM-RU 2036 7290) specimens do not segre-gate across the Kisingiri range of variation
Upper fourth premolars of Ekembo are broadly similar inmorphology to Ekembo P3s in fact the similarity between P3 and P4
is substantially greater in Ekembo than in Proconsul (Figs 4a c)Likewise most of the morphological differences that discriminatethe P3s of these genera also tend to differentiate the P4s but less
clearly Cusp heteromorphy does not distinguish P4s of thesegenera but the overall height of the P4 paracone in Ekembo issignificantly greater relative to crown length than in Proconsul P4s(t-test df frac14 14 t frac14 316 p frac14 00069 Fig 5b)
Upper molars Cusps of Ekembo upper molars are uniformly morebunodont than those of Proconsul occupying more of the occlusalsurface Intercuspal crests are more fully incorporated into thecusps in Ekembo molars giving them a more inflated appearanceThis is particularly evident in the crista obliqua which is very broadin Ekembo but thin and knife-like in Proconsul (Fig 6) Cusps ofProconsul molars stand more as individual cones Pickford et al(2009) have further differentiated the cusp morphology ofsmaller and larger Tinderet specimens arguing that the latterhave more bulbous cusps While accurate this difference is minorcompared with the morphological difference between Ekemboand Proconsul and we regard the within-Proconsul cuspalvariation as size-related In fact the intermediately sizedP meswae has cusp morphology that is appropriatelyintermediate between that of P africanus and the larger P major
Other differences between Ekembo and Proconsul upper molarsare most evident in M2 We concur with previous assessments thatthe Kisingiri upper molars have less developed cingula than non-Kisingiri molars (eg Senut et al 2000 Harrison 2002 2010Harrison and Andrews 2009) More specifically Ekembo M2s lackor have only rudimentary buccal cingula between the paracone andmetacone Likewise the lingual cingulum is discontinuous merg-ing into the hypocone lingually and then re-emerging as a distalmarginal ridge (Fig 6) Proconsul M2s display substantially morecingular development overall and in particular exhibit a prominentbuccal shelf between the paracone and metacone and a distinctdistolingual cingulum that is continuous around the hypocone towhere it merges with the distal marginal ridge
Figure 5 Box-and-whisker plots a) P3 cusp heteromorphy index (protocone heightparacone height) Results of a t-test show this difference to be significant (p frac14 00165) Theexcluded Ekembo outlier KNM-RU 1677 only appears to have extreme heteromorphy because of extensive expansion of the buccal enamel onto the anterior root (see text) b) P4
relative paracone height (paracone heightmesiodistal crown length) Results of a t-test show this difference to be significant (p frac14 00069)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 51
The occlusal profiles of Ekembo M2s are squarish in shape withrelatively straight and equal sides with a posthypocone crista thatis distobucally to buccally directed In contrast the occlusal profilesof Proconsul M2s have a more rhomboid shape with substantialbasal flare around the mesial cusps and a distally expanded hypo-cone with a distally directed posthypocone crista resulting in astrongly curved distal margin (Fig 6) Even in specimens with lesscurved distal margins the posthypocone crista is still more distallydirected compared to the condition in Ekembo These features arecharacteristic not only of P africanus P meswae and P major butare also expressed in the isolatedM2 fromMoroto Uganda (MOR IIb2098) attributed by Pickford et al (2009) to ldquoUrdquo gitongai butregarded here as belonging to P major
As noted by others (eg Harrison and Andrews 2009) the de-gree of buccolingual molar flare differs among these samplesQuantified as the distance between the tips of the mesial cuspsdivided by maximum crown breadth the degree of molar flare in
Ekembo M2s is significantly less than in Proconsul M2s (t-testdf frac14 29 t frac14 396 p frac14 00004) In a bivariate plot of these twomeasures the distributions of Ekembo and Proconsul teeth barelyoverlap with only KNM-RU 1677 encroaching into Proconsul ter-ritory (Fig 7)
These differences in M2 morphology also tend to distinguishEkembo from Proconsul M1s but not always as clearly Whereas M2smore consistently express distinct morphotypes the M1s in bothgenera show more variation in these features
The degree of upper third molar development (ie whether M3
has four distinct cusps or the distal cusps are reduced or absent) haslong been cited as taxonomically informative for these species orgroups of these species (eg Hopwood 1933 Le Gros Clark andLeakey 1951 Andrews 1978 Pickford et al 2009) However wefound this tooth to be unreliable for taxonomic purposes Withinprimates M3s are typically the most variable molars in both sizeand shape (Blankenship et al 2007 Kavanagh et al 2007) and this
Figure 6 Occlusal view of the M2s of Proconsul and Ekembo Left P africanus holotype(M 14084) Right E heseloni holotype (KNM-RU 2036) Note in Proconsul the overalldifference in crown shape as well as the more conical individuated cusps and nar-rower cristae strongly developed lingual cingulum that continues to the distal aspectof the crown broad buccal cingular shelf and more distally directed posthypoconecrista In Ekembo the cusps are more bunodont and expanded the cristae are broaderwith better developed accessory tubercles the lingual cingulum merges into thehypocone there is no buccal shelf and the posthypocone crista is more buccallydirected
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6152
is observable even in small samples of Proconsul species TheP africanus holotype has a substantially reduced M3 whereas inKNM-CA 2250dattributed by most authors to this species(Andrews 1978 Pickford et al 2009)dit is fully developed Simi-larly the hypodigm of P major includes both fully developed M3s(eg KNM-CA 389 LG 1815) and specimens with reduced ormissing hypocones (KNM-CA 397 1299) Ekembo displays compa-rable ranges of variation from highly reduced to fully developedM3s in both species Thus these species and genera cannot bedistinguished based on M3 development and we did not find anyother distinguishing characteristics between Ekembo and Proconsulin the upper third molars
Figure 7 Scatter plot depicting the de
Mandible Pickford et al (2009) enumerated several traits thatreadily distinguish Kisingiri from non-Kisingiri mandibles andthese distinctions are entirely consistent with the distinction ofEkembo from Proconsul Our analyses specifically underscoredifferences in the symphyseal region The planum alveolare inboth male and female specimens of Ekembo is short and morevertically oriented even the most elongate of these (KNM-RU47805 McNulty et al 2007) is substantially more verticalcompared to specimens of Proconsul In contrast the planumalveolare in Proconsul is conspicuously long and nearlyhorizontally oriented Well-preserved mandibles attributed toP major (eg KNM-SO 396 and 404) exhibit this morphology inextreme fashion (Fig 8) smaller specimens (KNM-SO 1112P africanus) show less extreme expression but are still readilydistinguishable from Ekembo mandibles For its size KNM-LG 452has a surprisingly short planum alveolare though not to thedegree seen in Rusinga mandibles Correlated to the shortplanum alveolare in Ekembo is a reduced superior transversetorus which while well-developed does not approach therobusticity expressed in Proconsul mandibles This difference isreflected in an index of symphyseal robusticity (thicknessperpendicular to the long axis of the symphysislength of thelong axis) which shows Ekembo and Proconsul to be significantlydifferent (t-test df frac14 14 t frac14 430 p frac14 00007)
Development of the inferior transverse torusdoften used todifferentiate among species of Proconsul (eg Pickford et al 2009Harrison 2010)dwas found to be highly variable and inconsis-tently expressed within these samples However the overallorientation of the subplanum region (as defined by a line con-necting the posterior-most midline points of the superior andinferior transverse tori or the posterior-most midline point of theinferior margin of the symphysis in the absence of the latter) doesdistinguish Ekembo from Proconsul mandibles The orientation ofthis line in relation to the alveolar margin in the vicinity of thepostcanine teeth was assessed qualitatively by placing a small rod
gree of buccolingual flare in M2
Figure 8 Mandibles of Proconsul and Ekembo Occlusal views of P major a) KNM-SO 396 b) KNM-SO 404 c) KNM-LG 452 Occlusal views of Ekembo d) KNM-RU 2087 e) KNM-RU1674 f) KNM-RU 7290 Occlusal views of P africanus g) KNM-SO 372 h) KNM-SO 1112 i) Lingual view of P africanus (KNM-SO 1112) Note the long more horizontally orientedplanum alveolare in Proconsul resulting in a pronounced superior transverse torus The planum alveolare is shorter and more vertical in Ekembo such that one can see the inferiormandibular border from occlusal view
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 53
(eg a dental pick) in themidline internally along the two referencepoints and recording the point in lingual perspective at which therod intersected the alveolar margin (Fig 9) In Ekembo the sub-planum region is more vertically oriented so that the rod intersectsthe alveolar margin most often in the premolar field Not surpris-ingly there is a difference between sexes with females having areduced superior transverse torus and therefore a more verticallyoriented subplanum region compared to males In contrast withEkembo Proconsul mandibles have much more inclined subplanumsurfaces with the rod intersecting the lingual alveolar margin inthe vicinity of the M1 (females) or M2 (males Fig 9) When sex istaken into account therefore the distributions of Ekembo andProconsul in this feature are discrete Note that male and femalemandibles attributed to both P africanus and P major are includedin this assessment and are surprisingly similar despite dramaticsize differences
We also quantified internal symphyseal morphology using threemeasures symphyseal height (infradentaleegnathion) planumalveolare length (posterior-most point on the superior transversetoruseinfradentale) and subplanum length (posterior-most pointon the superior transverse torusegnathion) a combination thatincorporates not only the lengths of these intervals but also theangular relationships among them To compare symphyseal shapeacross a range of sizes we chose a simple principal componentsapproach to partition size onto the first eigenvector (cf Jolicouer1963) While this is not the preferred method for partitioning sizein most datasets (Jungers et al 1995) for this particular sample andset of measurements Mosimann shape variables were still
significantly correlatedwith size (see discussion in Coleman 2008)Hence we assessed differences in symphyseal morphology be-tween Ekembo and Proconsul using the second and third principalcomponents of the three variables Distributions of the two generaare distinct in this shape space (Fig 10 Supplementary OnlineMaterial [SOM] Table 1)
Robusticity of the mandibular corpus also differs between thesesamples though it is more difficult to test due to differentialpreservation among specimens Corpus thickness and depth underP4 can be measured in the greatest number of specimens andcomparison of these measures suggests a difference betweengroups in the scaling of corpus robusticity (Fig 11) Among largepresumably male specimens Ekembo mandibles are substantiallyless robust than those of Proconsul However among smallermandibles only a single Proconsul specimen (KNM-SO 372) could beincluded and it is similar in robusticity to Ekembomandibles of thissize Considered together the impression is that corpus robusticitymay scale differently in Ekembo than it does in Proconsul and a testfor differences in slopes using an ANCOVA design of the loggedvariables was significant (df frac14 2 F frac14 2481 p lt 00001) Never-theless additional specimens are needed to properly evaluate theinfluence of taxonomy sex and size on corpus robusticity areasonable alternative interpretation of Figure 11 is that it dem-onstrates a significant difference between P major and all otherspecies of both Proconsul and Ekembo
Lower incisors Ekembo specimens are uniformly narrower inmesiodistal length than incisors attributed to P africanus P major
Figure 10 Scatter plot of principal component scores based on an eigenanalysis of threeposterior point on the superior transverse toruseinfradentale) and subplanum length (mprimarily captures size differences among specimens (Jolicouer 1963 see text and SOM Trespectively) in the symphyseal variables
Figure 9 Line drawing of a hemimandible depicting different orientations of the sub-planum area in Ekembo and Proconsul Females are denoted using shorter vectors maleswith longer vectors Vector is estimated in M 14086 which is broken at the midlineEkembo specimens have a more vertical subplanum compared to those from Songhorand Legetet but within both groups females have more vertically oriented regions thanmales Hence even the most ventrally rotated subplanum in the largest Ekembo malespecimen (KNM-RU 47805) groups with the Tinderet females in this feature
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6154
and P meswae both absolutely and in relation to crown height(Fig 12) This is true for both I1 and I2 A t-test adjusted for non-independent observations (eg I1 and I2 from the same mandible)confirmed the difference as significant (t-test df frac14 12 t frac14 258p frac14 00242) Moreover while the measurable Proconsul samplewas small this same pattern is observable in other specimenswhose tips were slightly to moderately worn and therefore notmeasured
Lower canines The only lower canine trait that discretely sepa-rates these genera is the pointed crown tip in Ekembo (Fig 3d)contrasted with a blade-like (or ldquoburinrdquo) tip in Proconsul(Fig 3c) This feature is well established for P major but is alsoexpressed in KNM-CA 1772 which was referred to P africanus byKelley (1986) and is too small to be either P major or P meswaeTwo other specimens commonly attributed to P africanus KNM-CA 2149 and KNM-SO 1112 both have broken canine tipsLikewise a new specimen (Kor 65004) attributed to P africanusby Pickford et al (2009) also has a broken tip and cannot beevaluated (see Pickford et al 2009 Fig 23) As with the uppercanines the blade-like tip is also present in the deciduous lowercanines of P major (eg KNM-CA 361 S0 5352) and P meswae(KNM-ME 1)
Lower premolars Like most previous authors (eg Andrews 1978Harrison 2002 2010 Pickford et al 2009) we found neitherqualitative nor quantitative characteristics of the lower premolarswith any taxonomic value
Lower molars Differences in lower molar morphology are few andrelatively subtle but in some cases mirror those seen in the uppermolars Cusps on the lower molars of Ekembo are more bunodontthan those in Proconsul and occupy more of the occlusal surfaceBroad inter-cuspal crests are incorporated into the Ekembo cuspsmaking them appear more inflated Molar cusps of Proconsul are
variables symphyseal height (infradentaleegnathion) planum alveolare length (mostost posterior point on the superior transverse torusegnathion) The first componentable 1) and thus PC 2 and PC 3 illustrate the residual shape variance (72 and 28
Figure 11 Scatter plot showing mandibular corpus robusticity under P4 Trend lines were computed using ordinary least squares (OLS) regression Similar relationships areobservable at other parts of the corpus but differential preservation results in smaller samples at those positions Slopes are significantly different (p lt 00001 tested on loggedvariables)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 55
more distinct from one another and are linked by narrower sharperinter-cuspal crests The lower molars of Ekembo also have asubstantially less developed buccal cingulum than those ofProconsul (Harrison 2002 2010 Harrison and Andrews 2009)
Figure 12 Scatter plot of lower incisor crown proportions Differenc
Lastly and again as seen in the upper molars the two genera canbe distinguished by the significantly greater amount of lowermolar flare in Proconsul (Fig 13 t-test df frac14 59 t frac14 713p lt 00001 see also Harrison and Andrews 2009)
es between Ekembo and Proconsul are significant (p frac14 00242)
Figure 13 Box-and-whisker plot of buccolingual flare in lower molars measured asthe average distance between the mesial buccal and lingual cusps divided by overallbuccolingual crown breadth Differences between Ekembo and Proconsul are highlysignificant (p lt 00001)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6156
Relative molar size has been used to distinguish species ofProconsul but we observed the primary difference to be in relativebuccolingual breadths of M1 and M2 In Ekembo M1 is substantiallynarrower than M2 than is the case in Proconsul (Fig 14 Harrison2002 2010) Although none of the smaller Proconsul mandiblespreserve complete crowns for both M1 and M2 KNM-SO 1112 hasan intact M2 and at least preserves the cervix for M1 However evenif the cervical measurement is used for its M1 breadthdthusunderestimating the true crown breadthdKNM-SO 1112 still has abroader first molar in relation to M2 than would be expected inEkembo If crown breadth is estimated from the ratio of cervixbreadth to maximum crown breadth in other Proconsul fossils therange of likely breadths for the SO 1112 M1 clearly distinguishes it
from the Ekembo sample (Fig 14) Testing this relationship onlogged variables using the mean estimate for the SO 1112 M1(Fig 14) reveals a significant difference between the two groups(ANCOVA df frac14 1 F frac14 2424 p frac14 00002)
As with upper third molars we find a great deal of variation inM3 morphology and mistrust the taxonomic significance some-times placed on characteristics of this toothdat least with regard tothese taxa Harrison (2002 2010) cited the mesiodistal position ofthe M3 entoconid as differentiating Kisingiri third molars fromthose attributed to P africanus We concur with his assessment butfound the entoconid position to vary in specimens attributed toP major Thus this character does not differentiate Ekembo from allProconsul species but may distinguish it from the type speciesP africanus
Discussion
The distinctiveness of the Kisingiri large catarrhine materialfrom that at Tinderet and Ugandan localities has been noted pre-viously by several authors (Pickford 1986 Walker et al 1993Begun 2001 2007 MacLatchy and Rossie 2005 Harrison andAndrews 2009 Harrison 2010) What had not been fully docu-mented was the nearly complete lack of overlap between thesesamples in a number of features despite very large assemblagesfrom Rusinga and Mfangano On descriptive criteria alone (Table 2)many isolated teeth (I1 C1 P3 M2 C1) and all of the mandibles canbe assigned to either Proconsul or Ekembowith very little chance ofmisattribution That some of the most diagnostic features occur inthe upper dentitiondthe only anatomy preserved in theP africanus type specimendadds weight to the taxonomicarrangement proposed here The morphology of the upper teeth inall Proconsul species compellingly unites them into a single cladedistinct from E heseloni and E nyanzae This is reinforced bynumerous features of the mandible and mandibular dentition thatfurther distinguish these samples The number and variety of thesefeatures are such that Proconsul and Ekembo are more differentfrom one another than are many closely-related modern catarrhinegenera
Derived morphology of Ekembo and Proconsul
It is premature to formally assign primitive and derived status tothe individual features of Ekembo and Proconsul before completinga formal phylogenetic analysis This is especially true given theabundance of evidence available for Ekembo and hence the poten-tial impact that our generic revision could have on phylogeneticinterpretations Nevertheless in comparison with both extanthominoids and the small-bodied catarrhines from the earlyMiocene it is informative to differentiate the evolutionary grades ofthese genera Many characteristics of Ekembo are more advanced inthe direction of modern apes compared to those of Proconsul(Table 2) General trends such as expanded and more bunodontcusps broader molar crests reduced cingulum and morphologicalsimilarity between P3 and P4 all represent morphologies thatpresage those of later fossil and extant hominoids The moreadvanced morphologies of P3 and M2 are particularly conspicuouscompared not only with those of Proconsul but to most earlyMiocene catarrhines In contrast the mesially positioned stronglyheteromorphic cusps on the P3 of Proconsul plus the correspondingasymmetric buccal profile notched mesial margin and missingmesial fovea are characteristic of other early Miocene catarrhinessuch as Dendropithecus Rangwapithecus and Limnopithecus andare likely primitive among East African catarrhines The moresymmetric P3 of Ekembo with midline positioned cusps andreduced cusp heteromorphy is reminiscent of later hominoids and
Figure 14 Relative buccolingual dimensions of M1 and M2 KNM-SO 1112 has a preserved M2 but only the cervix of M1 Its M1 breadth was estimated based on the ratio to cervixbreadth to maximum crown breadth in other Tinderet fossils Even the cervical breadth of SO 1112 is broader than would be predicted for the crown breadth of a Kisingiri M1 Trendlines computed using OLS regression and using the average estimate of SO 1112 M1 breadth Group differences were significant (p frac14 00002)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 57
even hominid taxa such as Rudapithecus (eg Kordos and Begun2001)
Proconsul is likewise united by a suite of features that are for themost part characteristic only of this lineage (Table 2) Themorphology of both upper and lower canines is unique to thisgenus and shared by all three well-known species plus the poorlyrepresented middle Miocene P gitongai (Pickford and Kunimatsu2005) The mandibular symphysis is also distinctive in Proconsulcompared with other catarrhines with its very long and shallowlysloping planum alveolare and steeply inclined subplanum surfaceIndeed even the small edentulous anterior mandibular fragmentKNM-SO 372 is readily placed within Proconsul and excluded fromother similarly sized catarrhine taxa such as Rangwapithecus andEkembo based on this unique symphyseal morphology (Figs 8 and9)
Other differences between Ekembo and Proconsul are of uncer-tain evolutionary polarity (eg orientation of the posthypoconecrista) or may be of taxonomic relevance without broader phylo-genetic significance (eg degree of molar flare Table 2) Werecognize that there are morphological differences within Procon-sul particularly between the better known species P africanus andP major However many of these differences are likely related tosize and represent variations on a single generic theme that aremarkedly different from the morphology of Ekembo The recentdescription (Harrison and Andrews 2009) of the intermediatelysized P meswae underscores the broad morphological unity withinProconsul and further substantiates a well-defined Proconsul cladecomprising Tinderet and Ugandan species
Based on the abundant morphological and morphometric evi-dence supporting genus-level distinction between Proconsul andEkembo we are currently conducting a more detailed phylogeneticanalysis of all early Miocene catarrhines in order to better under-stand the distribution and evolutionary significance of these andother morphological features We anticipate that the debates sur-rounding the phylogenetic relationships of an historically consti-tuted sample of Proconsul can be more satisfactorily resolved nowthat the Kisingiri material is appropriately recognized as a distinctgenus
The generic disposition of ldquoUgandapithecusrdquo species
A great many features said to characterize ldquoUgandapithecusrdquowere added to the original diagnosis of the genus by Pickford et al(2009) in their systematic revision of Proconsul (sensu lato) As partof that study they also erected a new species ldquoUrdquo legetetensis andtransferred P meswae to ldquoUgandapithecusrdquo Evaluating the same setof features across the entire available sample however we wereunable to find support for this more expansive conception ofldquoUgandapithecusrdquo for several reasons First some diagnostic fea-tures cited in Pickford et al (2009) were quite variable within theldquoUgandapithecusrdquo sample As an example marked reduction of theM3 entoconid to a bead on the lingual cingulum is not shared byseveral ldquoUrdquo major specimens such as KNM-CA 393 NAP I 49000and Kor 253004 (see Pickford et al 2009 Fig 33e) and was likelynot a feature of a few of the more worn specimens eg BUMP 600and BUMP 601 or even the holotype of P major M 16648 Second
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6158
some diagnostic features of ldquoUgandapithecusrdquo were found to bevariably present in Ekembo arguing against their utility for dis-tinguishing ldquoUgandapithecusrdquo from Pickford et als (2009) com-bined P africanus thorn Kisingiri conception of Proconsul For examplesignificantly reduced curvature of the I1 distal margin in ldquoUganda-pithecusrdquo is also found in some Kisingiri specimens including KNM-RU 1685 1712 1714 1831 1846 and 1951 several more Rusingaspecimens have moderately reduced curvature Third some fea-tures that purportedly distinguish ldquoUgandapithecusrdquo fromP africanus are likely related to size For example the somewhatmore central position of the P3 paracone in ldquoUrdquomajor is not seen inthe smaller ldquoUrdquo meswae whose paracone is positioned similarly tothat of the holotype of P africanus (see Harrison and Andrews2009 Fig 6d)
However the major part of the evidence for recognizingldquoUgandapithecusrdquo (sensu Pickford et al 2009) was assembled by re-assigning specimens traditionally attributed to P africanus into newspecies of ldquoUgandapithecusrdquo The P africanus holotype was left inassociation with the Rusinga sample despite having morphologyoverwhelmingly characteristic of other Tinderet specimens Theresulting specimen allocations left P africanus morphologicallyimpoverisheddstripped of much of its hypodigmdso that Pro-consul for Pickford et al (2009) came to be characterized primarilyby the Rusinga sample (see similar comments in Harrison andAndrews 2009) Within that conceptual framework any spec-imen that differs from those at Rusinga could not be Proconsul butmust instead belong to ldquoUgandapithecusrdquo Thus specimens such asthe KNM-SO 1112 mandible were erroneously assigned to ldquoUgan-dapithecusrdquo because they were (correctly) perceived as differingfrom their Rusinga counterparts (Pickford et al 2009)
Here we restore the P africanus hypodigm and associate it withthe other Tinderet fossils according to their shared morphologySpecimens assigned to ldquoUgandapithecusrdquo revert to Proconsul andmany of the features cited to support a speciose ldquoUgandapithecusrdquo(Pickford et al 2009) instead become diagnostic features of Pro-consul This correction serves to highlight the distinctiveness of theKisingiri remains and the need to recognize this distinctiveness atthe generic level
As demonstrated above specimens from Kisingiri do not displaythe morphology of Proconsul as represented by the holotype ofP africanus M 14084 Given that this specimen preserves only theupper canine and cheek teeth similarities with or departures fromthis morphology represent the most compelling arguments forassessing inclusivity in the genus However for the mandible andlower dentition if one simply sorts the specimens by elementwithout relying on taxonomic preconceptions the overwhelmingdistinction within the traditionally constituted Proconsul sample isbetween Kisingiri material on the one hand and that from Tinderetand Uganda on the other (cf Walker et al 1993 Begun 2001 2007MacLatchy and Rossie 2005 Harrison and Andrews 2009Harrison 2010) Additionally with the Kisingiri specimensremoved to Ekembo the postcranial differences used to partlyjustify a distinct ldquoUgandapithecusrdquo lose their relevance (seeHarrison and Andrews 2009)
We have differed among ourselves in our past treatment ofP major as generically distinct from the smaller species of Pro-consul (Begun 2007 McNulty 2010) Morphologically many ofthe unique features of P major can be interpreted as size-relatedvariants on the smaller P africanus morphology Even if so thequestion is whether these are sufficiently different and numerousto warrant generic distinction that is do they suggest thatP major belongs in a separate clade from P africanus andP meswae As we have demonstrated differences among thesethree species are certainly fewer and show less morphologicaldistinction than those between Ekembo and any of the three
Nevertheless while we ultimately agree with Harrison andAndrews (2009 see also Begun 2007) that the features dis-tinguishing ldquoUrdquo major from P africanus are consistent with thosethat distinguish congeneric species and are at least partly due todifferences in size we concede the possibility that the largestProconsul species could belong in a separate genus (Senut et al2000 Pickford and Kunimatsu 2005) However whether or notone recognizes a separate genus for P major the P africanus ma-terial does not sort morphologically with the Kisingiri materialTherefore the taxonomic alternatives are either two generaProconsul and Ekembo or three Proconsul Ugandapithecus andEkembo with Proconsul and Ugandapithecus united in a cladeexclusive of Ekembo Thus while we strongly favor the firstalternative recognizing Ugandapithecus does not change thefundamental argument concerning the need to refer the Kisingirispecies to a generically and cladistically distinct taxon from thespecies represented at Tinderet and the Ugandan sites
Specimens of interest
Despite the substantial dental and gnathic differences betweenProconsul and Ekembo there are a few specimens whose taxonomicaffinities require additional scrutiny
KNM-RU 1676-77 This set of associated upper and lower teeth is aconsistent outlier among E heseloni and E nyanzae specimens Insome ways this specimen more closely resembles Tinderet Pro-consul in the features that distinguish it from Ekembo Molar cuspmorphology and cingulum development I1 lingual tubercledevelopment and M2 shape and posthypocone crista orientationare more consistent with specimens from the Tinderet sites On theother hand it otherwise resembles Ekembo and lacks the strongestdiagnostic features that unite species of Proconsul such as thedistinctive canine and P3 morphology Overall this specimen doesnot fit unambiguously into either genus in fact the degree ofbuccolingual flare cusp morphology enamel wrinkling and espe-cially the long extension of enamel onto the anterior root in P3 arein some ways reminiscent of Afropithecus although it clearly differsfrom that genus as well (cf Rossie and MacLatchy 2013)
Were these characteristics individually distributed among thelarge Kisingiri sample one might attribute this to intra-genericvariation but when found together in a single specimen from alarge sample it suggests a unique individual One of us (KPM)thinks it likely that the specimen was not actually found at aKisingiri locality based on its unique preservation compared toother Rusinga fossils and on other examples of mis-proveniencedspecimens (Leakey 1967 Andrews and Molleson 1979 Pickford1986 Kelley 1986) However we recognize that relying toofrequently on this explanation allows almost any scenario to bejustified and suggest some alternative possibilities One is that thespecimen was found on Rusinga but in the much older WayandoFormation strata instead of the Hiwegi Formation The differencesin morphology in that case might reflect more than a million yearsof temporal change (McCollum et al 2013) with KNM-RU 1676-77preserving certain more primitive Proconsul-like features As analternative it is reasonable to assume that fossil assemblages asvast as those from Rusingawill include not only abundant taxa butalso rare or infrequent inhabitants of the local biome There aremany examples of this from Rusingas mammalian communityeg ldquoHemicyonrdquo Kulutherium Tadarida cf Hyainailouros cfArchaeobeledon Kelba (see relevant chapters in Werdelin andSanders 2010) As such KNM-RU 1676-77 may simply representa rare and well-preserved specimen of some other catarrhinetaxon that lived among or visited a primate community dominatedby Ekembo
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 59
KNM-SO 1112 This specimen was initially referred to Rangwapi-thecus but soon after was transferred to Proconsul afrianus (Bosler1981 Martin 1981)da designation that most subsequent authorshave followed (Pickford 1986 Nengo and Rae 1992 Kelley 19861995 Walker et al 1993 MacLatchy and Rossie 2005 Hill et al2013) Pickford et al (2009) more recently referred this specimento ldquoUrdquo meswae citing two primary reasons First they reiteratedBoslers (1981) concern that its P3 differs from others attributedto P africanus However at that time the majority of what wasconsidered to be P africanus would have been from RusingaIsland In fact while noting its contrasting morphology Boslersuggested that differences between KNM-SO 1112 and other P3smight reasonably be attributed to sexual dimorphism notingthat this specimen provides ldquoa not implausible match for the Pafricanus holotype maxilla M 14084rdquo (1981152) Thatassessment is borne out in this study Using the much largersample of Ekembo P3s as a model variation can be seen toencompass specimens with well-developed mesial beaks andthose with more bilaterally compressed crowns and parallelsides The latter morphology is predominantly found amongmale specimens (KNM-RU 1674 1924 and KPS I and III) thussupporting Boslers (1981) suggestion that this variation isrelated to sexual dimorphism and hence that KNM-SO 1112 is amale P africanus
The second feature cited by Pickford et al (2009) in referringthis specimen to ldquoUrdquo meswae was posterior shallowing of themandibular corpus However because they redefined ldquoUgandapi-thecusrdquo to include all of the well-preserved Tinderet mandibles(Pickford et al 2009)dleaving Proconsul to be represented only byRusinga specimensdtheir only alternative within this scheme for aspecimen the size of SO 1112 was to refer it to ldquoUrdquomeswae It seemsodd though to assign this specimen to a species from a (perhapsmuch) older site when the Proconsul africanus holotype is ofappropriate size and similar age and as there are no other adultmandibles from either species with which to compare it In anycase mandibular shallowing is variable in both Ekembo and Pro-consul with some Kisingiri specimens exhibiting considerableshallowing (eg KNM-RU 2087) and some ldquoUgandapithecusrdquo spec-imens having very little (eg M 14086)
We therefore see no morphological grounds to assign KNM-SO1112 to P meswae but do find some morphological evidence toassign it as a male P africanus which occurs at Songhor
KNM-LG 452 This very complete mandible was originally recog-nized as a female of P major based on the size of the dentition androbusticity of the mandible (Martin 1981) Bosler (1981) howeverleft this specimen unassigned noting similarities to P major in P3-M1 size but having distinctly smaller M2e3 In this she likenedKNM-LG 452 to a few other specimens (notably M 14086) thatare smaller in their distal molars in a manner more consistentwith taxonomic rather than sex differences (Bosler 1981 see alsoPilbeam 1969) Kelley (1986 1995) also assigned this specimen asa female of P major based on canine size and morphology butnoted that the canine lacked the shallowly-sloping mesial ridgetypical of most female anthropoid canines This is also truehowever of two other canines assigned as female P major KO 9and SO 373 (Kelley 1986) and so might simply be an unusualcharacteristic of P major female lower canines In the end Kelley(1995) concluded that considering all of the criteria fordistinguishing male and female canines the evidence that KNM-LG 452 is female remains questionable
Pickford et al (2009) further developed the idea that two spe-cies are represented within the P major sample and made KNM-LG452 the holotype of a new species ldquoUrdquo legetetensis We concur thatthis specimen is unique among the well-preserved Tinderet
mandibles in some important features Whereas male and femalemandibles of P major have long internal symphyseal planesexpansive extra- and retromolar sulci and rami that cross theocclusal plane at distal M3 KNM-LG 452 has a shorter symphysealplane (though not to the extent seen in Ekembo) a moderateextramolar sulcus but no retromolar sulcus and a ramus thatcrosses the occlusal plane more mesially If KNM-SO 396 and SO404 represent males and females respectively of P major then themorphology of KNM-LG 452 appears distinct
At present however we do not think there is sufficient evidenceto differentiate this specimen from either P major or P meswaePickford et al (2009) argue that KNM-LG 452 cannot belong to thelatter species because of its larger size The size differences how-ever are well within the range of variation seen in P majorE heseloni and E nyanzae Moreover whereas the Legetet mandibleis larger than most of the P meswae hypodigm an adult P4 referredto this species (KNM-ME 25) substantially exceeds its counterpartin KNM-LG 452 Should the Legetetmandible ultimately prove to bea male specimen size alone would not rule out its attribution toP meswae On the other hand if it is female then it becomes harderto justify its inclusion in that sample of mostly smaller specimens Iffemale therefore it could represent a separate species for whichthe binomial Proconsul legetetensis comb nov is available Recentlycollected fossils from the SonghorKapurtay area provide someadditional support for the possibility that three species of Proconsulexisted at these localities (McNulty 2014ab) However toconvincingly demonstrate this and determine whether a thirdspecies can be differentiated from P meswae will require a moredetailed analysis of variation among Proconsul species a corre-sponding study of variation in modern taxa anddalmost certain-lydadditional fossil specimens For now we regard ldquoUrdquo legetetensisas a subjective junior synonym of P major
Conclusions
The genus Proconsul has been central to studies of ape and hu-man evolution since its first description by Hopwood (1933) As oneof the best known fossil catarrhines represented by nearly everyskeletal element our knowledge of Proconsuls functional anatomypositional behavior and overall morphology has made it thefoundation for paleoanthropological research in the early Mioceneas well as studies on the origin and early diversification of homi-noids With respect to systematics however Proconsul is less wellunderstood the type species P africanus is known from a rela-tively small number of specimens from sites in the Tinderet areaand no postcranial bones can be unambiguously assigned to itRather it is the copious and well-preserved specimens from theKisingiri sites of Rusinga and Mfangano Islands that have formedthe basis for the common image of Proconsul
We argue here that the Kisingiri species traditionally referredto ProconsuldP heseloni and P nyanzaedare generically distinctfrom P africanus and hereby transfer them into a new genusEkembo based on a substantial number of morphological differ-ences (Table 2) Considering only those traits preserved in theP africanus holotype the entire sample from Rusinga and Mfan-gano differs markedly in the morphology of the upper canine P3P4 and M2 Additional differences in the I1 lower incisors andcanines molar cusp and occlusal crest morphology degree of flarein all molars and especially mandibular features clearly distin-guish the Kisingiri from non-Kisingiri samples in a way that isincompatible with congeneric species With the transfer ofE heseloni and E nyanzae Proconsul becomes a well-defined cladecomprising at least three speciesdP africanus P major andP meswaedunited by several features that are unique for thegenus (Table 2)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6160
We found no support in this study for the genus ldquoUgandapi-thecusrdquo as constructed by Pickford et al (2009) In that paper theytransferred the vast majority of Proconsul fossils from the Tinderetand Napak localities into ldquoUgandapithecusrdquo leaving P africanusrepresented only by its holotype and a small number of isolatedteeth and aligning it with the material from Rusinga and Mfan-ganoWhen the P africanus hypodigm is reconstituted and properlyaffiliated with the other Tinderet species according to itsmorphology much of the evidence marshaled by Pickford et al(2009) for ldquoUgandapithecusrdquo actually supports the distinction be-tween the Tinderet and Kisingiri samples ie the generic distinc-tion of Ekembo In addition some of the traits enumerated byPickford and colleagues in support of ldquoUgandapithecusrdquo wererevealed in our analysis to be variable within one or more speciesand therefore of questionable taxonomic utility We do not rejectout of hand the original conception of ldquoUgandapithecusrdquo (Senutet al 2000) as either a monotypic genus distinguishing ldquoUrdquo ma-jor or perhaps including a closely allied middle Miocene ldquoUrdquogitongai as well (Pickford and Kunimatsu 2005) but we have fornow retained the early Miocene specimens of both in Proconsulpending further study However even if Ugandapithecus is a taxo-nomically valid genus comprising just the latter two species theirmorphology indicates that theywould still be united in a cladewithProconsul and they lack several more advanced features that partlydefine Ekembo Better understanding of the role that body size playsinmediating differences within Proconsulwill help to further clarifythe relationships among these species
Despite the existence of one well-represented and several lesscomplete skeletons of what was long assumed to be Proconsul thephylogenetic status of this genus has defied consensus This islargely due in our view to Proconsul having been an admixture oftwo distinct genera one more advanced in the direction of extantapes than the other As we demonstrate here the material fromRusinga and Mfangano traditionally referred to Proconsul andwhich includes all of the more complete skeletal material isdistinct from the Tinderet and Napak material and belongs in aseparate genus Ekembo By disentangling these distinct clades weanticipate being able to achieve better phylogenetic resolution forboth genera thereby improving our knowledge of early hominoidevolution and diversification
Acknowledgments
We thank many people and agencies that assisted with thedevelopment and completion of this project This research wasinitiated as a result of funding from the Wenner-Gren Foundationand the Johns Hopkins School of Medicine to DR Begun in1987e1989 and was renewed following research and travelgenerously funded by multiple Leakey Foundation grants to KMcNulty National Science Foundation grants to K McNulty(0852609 1241807) the McKnight Foundation the University ofMinnesota the Lloyd A Wilford Endowment the LeverhulmeFoundation and the Turkana Basin Institute J Kelley gratefullyacknowledges support from the Institute of Human Origins Ari-zona State University DR Begun also wishes to acknowledge theAlexander Humboldt Stiftung and NSERC for funding support Weare grateful to the National Museums of Kenya the UgandaMuseum the Natural History Museum and to Rose Mwanja EzraMusiime Tom Mukhuyu Samuel Muteti James Yatich and MaryMuungu for access to and assistance with specimens in their careMany people provided assistance comments criticisms insightsand suggestions on various aspects of this work and we especiallythank Holly Dunsworth Will Harcourt-Smith Tom Lehmann DanPeppe Nicole Garrett Kirsten Jenkins Rutger Jansma Susy CoteIsaiah Nengo Andrew Hill and Brenda Benefit K McNulty
acknowledges the evolutionary anthropology faculty and studentsat Durham University for their helpful comments on an earlyversion of this manuscript and for their gracious hospitality Weare likewise grateful to the anonymous reviewers associate editorand editor whose comments strengthened the paper This manu-script is publication 1 supporting Research on East AfricanCatarrhine and Hominoid Evolution (REACHE)
Appendix A Supplementary data
Supplementary data related to this article can be found at httpdxdoiorg101016jjhevol201503009
References
Andrews P 1974 New species of Dryopithecus from Kenya Nature 249 188e190Andrews P 1978 A revision of the Miocene Hominoidea of East Africa Bull Brit
Mus (Nat Hist) Geol 30 85e225Andrews P 1985 Family group systematics and evolution among catarrhine pri-
mates In Delson E (Ed) Ancestors The Hard Evidence Liss New Yorkpp 14e22
Andrews P Harrison T Martin L Pickford M 1981 Hominoid primates from anew Miocene locality named Meswa Bridge in Kenya J Hum Evol 10123e128
Andrews P Begun D Zylstra M 1997 Interrelationships between functionalmorphology and paleoenvironments in Miocene hominoids In Begun DWard C Rose M (Eds) Function Phylogeny and Fossils Miocene HominoidEvolution and Adaptations Plenum Press New York pp 29e58
Andrews P Molleson T 1979 The provenance of Sivapithecus africanus Bull BritMus (Nat Hist) Geol 32 19e23
Begun DR 2001 African and Eurasian Miocene hominoids and the origins of theHominidae In de Bonis L Koufos GD Andrews P (Eds) Hominoid Evolutionand Climatic Change in Europe Phylogeny of the Neogene Hominoid Primatesof Eurasia Cambridge University Press London pp 231e253
Begun DR 2007 Fossil record of Miocene hominoids In Henke W Tattersall I(Eds) Handbook of Paleoanthropology Springer Berlin pp 921e977
Begun DR Teaford MF Walker AC 1994 Comparative and functional anatomyof Proconsul phalanges from the Kaswanga primate site Rusinga Island KenyaJ Hum Evol 26 89e165
Bishop WW 1964 More fossil primates and other Miocene mammals from North-East Uganda Nature 203 1327e1331
Bishop WW Miller JA Fitch FJ 1969 New potassium-argon age determinationsrelevant to the Miocene fossil mammal sequence in East Africa Am J Sci 267669e699
Blankenship JA Mincer HH Anderson KM Woods MA Burton EL 2007Third molar development in the estimation of chronologic age in Americanblacks as compared with whites J Forensic Sci 52 428e433
Bosler W 1981 Species groupings of early Miocene dryopithecine teeth from EastAfrica J Hum Evol 10 151e158
Coleman MN 2008 What does geometric mean mean geometrically Assessingthe utility of geometric mean and other size variables in studies of skullallometry Am J Phys Anthropol 135 404e415
Cote S Malit N Nengo I 2013 Additional mandibles of Rangwapithecus gordonian early Miocene catarrhine from the Tinderet localities of Western KenyaJ Hum Evol 153 341e352
Deane AS 2009 Early Miocene catarrhine dietary behavior the influence of theRed Queen Effect on incisor shape and curvature J Hum Evol 56 275e285
Drake R Van Couvering JA Pickford M Curtis G Harris JA 1988 New chro-nology for the Early Miocene mammalian faunas of Kisingiri Western KenyaJ Geol Soc Lond 145 479e491
Evernden JF Savage DE Curtis GH James GT 1964 Potassium-argon datesand the Cenozoic mammalian chronology of North America Am J Sci 262145e198
Gebo DL MacLatchy L Kityo R Deino A Kingston J Pilbeam D 1997A hominoid genus from the Early Miocene of Uganda Science 276 401e404
Geoffroy Saint-Hilaire E 1812 Tableau des quadrumanes 1 Ord QuadrumanesAnnls Mus Hist Nat Paris 19 85e122
Gray JE 1825 Outline of an attempt at the disposition of the Mammalia into tribesand families with a list of the genera apparently appertaining to each tribeAnnls Philos 10 337e344
Harrison T 1982 Small-bodied apes from the Miocene of East Africa PhDDissertation University of London
Harrison T 1986 New fossil anthropoids from the middle Miocene of East Africaand their bearing on the origin of the Oreopithecidae Am J Phys Anthropol 71265e284
Harrison T 1987 The phylogenetic relationships of the early catarrhine primates areview of the current evidence J Hum Evol 16 41e80
Harrison T 2002 Late Oligocene to middle Miocene catarrhines from Afro-ArabiaIn Hartwig WC (Ed) The Primate Fossil Record Cambridge University PressCambridge pp 311e338
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 61
Harrison T 2010 Dendropithecoidea Proconsuloidea and Hominoidea (CatarrhiniPrimates) In Werdelin L Sanders WJ (Eds) Cenozoic Mammals of AfricaUniversity of California Press Berkeley pp 429e469
Harrison T Andrews P 2009 The anatomy and systematic position of a newspecies of early Miocene proconsulid from Meswa Bridge Kenya J Hum Evol56 479e496
Hill A Nengo IO Rossie JB 2013 A Rangwapithecus gordoni mandible from theearly Miocene site of Songhor Kenya J Hum Evol 65 490e500
Hopwood AT 1933 Miocene primates from Kenya J Linn Soc Zool 38 437e464Ishida H Kunimatsu Y Takano T Nakano Y Nakatsukasa M 2004 Nacholapi-
thecus skeleton from the Middle Miocene of Kenya J Hum Evol 46 69e103Jolicouer P 1963 The multivariate generalization of the allometry equation Bio-
metrics 19 497e499Jungers WL Falsetti AB Wall CE 1995 Shape relative size and size-
adjustments in morphometrics Am J Phys Anthropol 38 (S2) 137e161Kavanagh KD Evans AR Jernvall J 2007 Predicting evolutionary patterns of
mammalian teeth from development Nature 449 427e433Kelley J 1986 Species recognition and sexual dimorphism in Proconsul and
Rangwapithecus J Hum Evol 15 461e495Kelley J 1995 Sex determination in Miocene catarrhine primates Am J Phys
Anthropol 96 390e417Kelley J 1993 Taxonomic implications of sexual dimorphism in Lufengpithecus In
Kimbel WH Martin LB (Eds) Species Species Concepts and Primate Evo-lution Springer New York pp 429e458
Kent PE 1942 The country round the Kavirondo Gulf of Victoria Nyanza Geogr J100 22e31
Kordos L Begun DR 2001 A new cranium of Dryopithecus from RudabanyaHungary J Hum Evol 15 689e700
Le Gros Clark WE Leakey LSB 1950 Diagnoses of East African Miocene Homi-noidea Q J Geol Soc Lond 105 260e263
Le Gros Clark WE Leakey LSB 1951 The Miocene Hominoidea of East Africa InFossil Mammals of Africa No 1 British Museum (Natural History) London
Leakey LSB 1943 A Miocene anthropoid mandible from Rusinga Kenya Nature152 319e320
Leakey LSB 1963 East African fossil Hominoidea and the classification within thissuper-family In Washburn SL (Ed) Classification and Human EvolutionAldine Chicago pp 32e49
Leakey LSB 1967 Notes on the mammalian faunas from the Miocene and Pleis-tocene of East Africa In Bishop WW Clark JD (Eds) Background to Evolu-tion in Africa University of Chicago Press Chicago pp 7e29
Leakey RE Leakey MG 1986 A new Miocene hominoid from Kenya Nature 324143e146
Leakey RE Leakey MG Walker AC 1988 Morphology of Afropithecus turka-nensis from Kenya Am J Phys Anthropol 76 289e307
Leakey MG Ungar PS Walker A 1995 A new genus of large primate from theLate Oligocene of Lothidok Turkana District Kenya J Hum Evol 28519e531
Linnaeus C 1758 Systema naturae per regna tria naturae secundum classesordines genera species cum characteribus differentris synonymis locis tenthed Laurentii Salvii Stockholm
MacInnes DG 1943 Notes on the East African Miocene primates J E Afr UgandaNat Hist Soc 17 141e181
MacLatchy L 2004 The oldest ape Evol Anthropol 13 90e103MacLatchy L Bossert WH 1996 An analysis of the articular surface distribution of
the femoral head and acetabulum in anthropoids with implications for hipfunction in Miocene hominoids J Hum Evol 31 425e453
MacLatchy L Rossie JB 2005 The Napak hominoid still Proconsul major InLieberman DE Smith RJ Kelley J (Eds) Interpreting the Past Essays onHuman Primate and Mammal Evolution in Honor of David Pilbeam Brill Ac-ademic Publishers Boston pp 15e28
Martin L 1981 New specimens of Proconsul from Koru Kenya J Hum Evol 10139e150
McCollum MS Peppe DJ McNulty KP Dunsworth HM Harcourt-Smith WEH Andrews AL 2013 Magnetostratigraphy of the early MioceneHiwegi Formation (Rusinga Island Lake Victoria Kenya) Geological Society ofAmerica Abstracts with Programs 45 p 12
McNulty KP 2010 Apes and tricksters the evolution and diversification ofhumans closest relatives Evol Edu Outreach 3 322e332
McNulty KP Harcourt-Smith WEH Dunsworth HM 2007 New primate fossilsfrom Rusinga Island Kenya Am J Phys Anthropol 132 (S44) 170
McNulty KP MacLatchy L Peppe DJ Nengo I Manthi FK Miller ERStevens NJ Cote S Lehmann T Kingston J 2014a A regional approach toEast African early Miocene paleobiology J Vert Paleontol 185 SVP Programand Abstracts Book 2014
McNulty KP MacLatchy L Rossie JB Peppe DJ Deino AL Mbua ENManthi FK Nengo IO Stevens NJ Cote S Lehmann T Gutierrez M 2014b
Research on East African catarrhine and hominoid evolution results from thefirst year Am J Phys Anthropol 153 (S58) 182
Michel LA Peppe DJ Lutz JA Driese SG Dunsworth HM Harcourt-Smith WEH Horner W Lehmann T Nightengale S McNulty KP 2014Remnants of an ancient forest provide ecological context for Early Miocenefossil apes Nature Comm 5 3236
Myres JL 1932 Africa Man 32 208Nakatsukasa M Kunimatsu Y Nakano Y Takano T Ishida H 2007 Vertebral
morphology of Nacholapithecus kerioi based on KNM-BG 35250 J Hum Evol52 347e369
Nengo IO Rae TC 1992 New hominoid fossils from the early Miocene site ofSonghor Kenya J Hum Evol 23 423e429
Peppe DJ McNulty KP Cote SM Harcourt-Smith WEH Dunsworth HM VanCouvering JA 2009 Stratigraphic interpretation of the Kulu Formation (EarlyMiocene Rusinga Island Kenya) and its implications for primate evolutionJ Hum Evol 56 447e461
Pickford M 1981 Preliminary Miocene mammalian biostratigraphy for westernKenya J Hum Evol 10 73e97
Pickford M 1986 Sexual dimorphism in Proconsul Hum Evol 1 111e148Pickford M Andrews PJ 1981 The Tinderet Miocene sequence in Kenya J Hum
Evol 10 11e33Pickford M Kunimatsu Y 2005 Catarrhines from the Middle Miocene (ca 145
Ma) of Kipsaraman Tugen Hills Kenya Anthropol Sci 113 189e224Pickford M Senut B Gommery D Musiime E 2009 Distinctiveness of Ugan-
dapithecus from Proconsul Estudios Geol 65 183e241Pilbeam DR 1969 Tertiary Pongidae of East Africa evolutionary relationships and
taxonomy Bulletin of the Peabody Museum of Natural History 31 pp 1e185Rae TC 1993 Phylogenetic analysis of proconsulid facial morphology PhD
Dissertation State University of New York at Stony BrookRae TC 1999 Mosaic evolution in the origin of the Hominoidea Folia Primatol 70
125e135Rafferty KL Walker A Ruff C Rose MD Andrews PJ 1995 Postcranial esti-
mates of body weights in Proconsul with a note on a distal tibia of P major fromNapak Uganda Am J Phys Anthropol 97 391e402
Rossie JB MacLatchy L 2013 Dentognathic remains of an Afropithecus individualfrom Kalodirr Kenya J Hum Evol 65 199e208
Senut B Pickford M Gommery D Kunimatsu Y 2000 Un nouveau genredrsquohominoiumlde du Miocene inferieur drsquoAfrique orientale Ugandapithecus major(Le Gros Clark and Leakey 1950) CR Acad Sci Paris 331 227e233
Simons EL Pilbeam DR 1965 Preliminary revision of the Dryopithecinae (Pon-gidae Anthropoidea) Folia Primatol 3 81e152
Smith TM Martin LB Leakey MG 2003 Enamel thickness microstructure anddevelopment in Afropithecus turkanensis J Hum Evol 44 283e306
Stevens NJ Seiffert ER OConnor PM Roberts EM Schmitz MD Krause CGorscak E Ngasala S Hieronymus TL Temu J 2013 Palaeontological evi-dence for an Oligocene divergence between Old World monkeys and apesNature 497 611e614
Suwa G Kono RT Katoh S Asfaw B Beyene Y 2007 A new species of great apefrom the late Miocene epoch in Ethiopia Nature 448 921e924
Szalay FS Delson E 1979 Evolutionary History of the Primates Academic PressNew York
Teaford MF Beard KC Leakey RE Walker A 1988 New hominoid facial skel-eton from the Early Miocene of Rusinga Island Kenya and its bearing on therelationship between Proconsul nyanzae and Proconsul africanus J Hum Evol17 461e477
Van Couvering JAH Miller J 1969 Miocene stratigraphy and age determinationsRusinga Island Kenya Nature 221 628e632
Walker A 1997 Proconsul function and phylogeny In Begun DR Ward CVRose MD (Eds) Function Phylogeny and Fossils Miocene Hominoid Evolu-tion and Adaptation Plenum Press New York pp 209e224
Walker A Pickford M 1983 New postcranial fossils of Proconsul africanus andProconsul nyanzae In Ciochon RL Corruccini RS (Eds) New Interpretationsof Ape and Human Ancestry Plenum Press New York pp 325e351
Walker A Teaford M 1988 The Kaswanga primate site an Early Miocene hom-inoid site on Rusinga Island Kenya J Hum Evol 17 539e544
Walker A Teaford M 1989 The hunt for Proconsul Sci Am 260 76e82Walker A Teaford MF Martin L Andrews P 1993 A new species of Proconsul
from the early Miocene of RusingaMfangano Islands Kenya J Hum Evol 2543e56
Ward CV Ruff CB Walker A Rose MD Teaford MF Nengo IO 1995 Func-tional morphology of Proconsul patellas from Rusinga Island Kenya with im-plications for other Miocene-Pliocene catarrhines J Hum Evol 29 1e19
Ward S Brown B Hill A Kelley J Downs W 1999 Equatorius a new hominoidgenus from the Middle Miocene of Kenya Science 285 1382e1386
Werdelin L Sanders WJ 2010 Cenozoic Mammals of Africa University of Cali-fornia Press Berkeley

Figure 4 Upper premolars of Proconsul and Ekembo occlusal (aec) and lingual (def) views a) P3e4 of the P africanus holotype (M 14084) b) P3 of P major (M 14331) c) P3e4 of theE heseloni holotype (KNM-RU 2036) d) P3 of the P africanus holotype (M 14084) e) P3 of P major (M 14331) f) P3 of the E heseloni holotype (KNM-RU 2036) Note the mesiallypositioned cusps lack of mesial fovea and strongly asymmetric crown in Proconsul compared to Ekembo (see text for further comparisons) Image of M 14331 reversed for easiercomparison Photo 4a courtesy of Rutger Jansma Scale bars frac14 1 cm
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6150
germ and two dP3s attributed to P meswae look remarkably similarto those of other Proconsul species including the presence of thisflange In contrast the Ekembo P3 has a well-developed mesialfovea that terminates at a mesiobuccal shoulder positionedapproximately halfway between the paracone tip and the cervixFrom this shoulder the mesial margin angles slightly distallydown to the cervix (Fig 4f)
The degree of cusp height heteromorphy in P3 also differentiatesEkembo from Proconsul (contra Pickford et al 2009) Only four non-Kisingiri specimens were sufficiently unworn to take measure-ments but more worn specimens also appear to express this dif-ferential in cusp height Figure 5a shows the ratio of protoconeparacone height measured from the tooth cervix and with only oneexception Ekembo P3s have substantially less cusp heteromorphyThis exception KNM-RU 1677 does not in fact have greater heter-omorphy but instead has an abnormal extension of the enamel ontothe buccal (paracone) root which skews the measurement Hencecusp relief relative to the occlusal surface in RU 1677 is not nearly asheteromoprhic as in Proconsul and when this is taken into accountthe difference between genera is significant (t-test dffrac14 11 t frac14 283p frac14 00165) The adult P3 germ from Meswa Bridge is incompletelyformed and therefore could not be measured However the differ-ence between paracone and protocone heights with respect to theocclusal surface appears to be as great as that in the P africanusholotype It does not appear that sex is an important determinant ofthis feature since well-established male (eg M16647 KNM-RU16000) and female (KNM-RU 2036 7290) specimens do not segre-gate across the Kisingiri range of variation
Upper fourth premolars of Ekembo are broadly similar inmorphology to Ekembo P3s in fact the similarity between P3 and P4
is substantially greater in Ekembo than in Proconsul (Figs 4a c)Likewise most of the morphological differences that discriminatethe P3s of these genera also tend to differentiate the P4s but less
clearly Cusp heteromorphy does not distinguish P4s of thesegenera but the overall height of the P4 paracone in Ekembo issignificantly greater relative to crown length than in Proconsul P4s(t-test df frac14 14 t frac14 316 p frac14 00069 Fig 5b)
Upper molars Cusps of Ekembo upper molars are uniformly morebunodont than those of Proconsul occupying more of the occlusalsurface Intercuspal crests are more fully incorporated into thecusps in Ekembo molars giving them a more inflated appearanceThis is particularly evident in the crista obliqua which is very broadin Ekembo but thin and knife-like in Proconsul (Fig 6) Cusps ofProconsul molars stand more as individual cones Pickford et al(2009) have further differentiated the cusp morphology ofsmaller and larger Tinderet specimens arguing that the latterhave more bulbous cusps While accurate this difference is minorcompared with the morphological difference between Ekemboand Proconsul and we regard the within-Proconsul cuspalvariation as size-related In fact the intermediately sizedP meswae has cusp morphology that is appropriatelyintermediate between that of P africanus and the larger P major
Other differences between Ekembo and Proconsul upper molarsare most evident in M2 We concur with previous assessments thatthe Kisingiri upper molars have less developed cingula than non-Kisingiri molars (eg Senut et al 2000 Harrison 2002 2010Harrison and Andrews 2009) More specifically Ekembo M2s lackor have only rudimentary buccal cingula between the paracone andmetacone Likewise the lingual cingulum is discontinuous merg-ing into the hypocone lingually and then re-emerging as a distalmarginal ridge (Fig 6) Proconsul M2s display substantially morecingular development overall and in particular exhibit a prominentbuccal shelf between the paracone and metacone and a distinctdistolingual cingulum that is continuous around the hypocone towhere it merges with the distal marginal ridge
Figure 5 Box-and-whisker plots a) P3 cusp heteromorphy index (protocone heightparacone height) Results of a t-test show this difference to be significant (p frac14 00165) Theexcluded Ekembo outlier KNM-RU 1677 only appears to have extreme heteromorphy because of extensive expansion of the buccal enamel onto the anterior root (see text) b) P4
relative paracone height (paracone heightmesiodistal crown length) Results of a t-test show this difference to be significant (p frac14 00069)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 51
The occlusal profiles of Ekembo M2s are squarish in shape withrelatively straight and equal sides with a posthypocone crista thatis distobucally to buccally directed In contrast the occlusal profilesof Proconsul M2s have a more rhomboid shape with substantialbasal flare around the mesial cusps and a distally expanded hypo-cone with a distally directed posthypocone crista resulting in astrongly curved distal margin (Fig 6) Even in specimens with lesscurved distal margins the posthypocone crista is still more distallydirected compared to the condition in Ekembo These features arecharacteristic not only of P africanus P meswae and P major butare also expressed in the isolatedM2 fromMoroto Uganda (MOR IIb2098) attributed by Pickford et al (2009) to ldquoUrdquo gitongai butregarded here as belonging to P major
As noted by others (eg Harrison and Andrews 2009) the de-gree of buccolingual molar flare differs among these samplesQuantified as the distance between the tips of the mesial cuspsdivided by maximum crown breadth the degree of molar flare in
Ekembo M2s is significantly less than in Proconsul M2s (t-testdf frac14 29 t frac14 396 p frac14 00004) In a bivariate plot of these twomeasures the distributions of Ekembo and Proconsul teeth barelyoverlap with only KNM-RU 1677 encroaching into Proconsul ter-ritory (Fig 7)
These differences in M2 morphology also tend to distinguishEkembo from Proconsul M1s but not always as clearly Whereas M2smore consistently express distinct morphotypes the M1s in bothgenera show more variation in these features
The degree of upper third molar development (ie whether M3
has four distinct cusps or the distal cusps are reduced or absent) haslong been cited as taxonomically informative for these species orgroups of these species (eg Hopwood 1933 Le Gros Clark andLeakey 1951 Andrews 1978 Pickford et al 2009) However wefound this tooth to be unreliable for taxonomic purposes Withinprimates M3s are typically the most variable molars in both sizeand shape (Blankenship et al 2007 Kavanagh et al 2007) and this
Figure 6 Occlusal view of the M2s of Proconsul and Ekembo Left P africanus holotype(M 14084) Right E heseloni holotype (KNM-RU 2036) Note in Proconsul the overalldifference in crown shape as well as the more conical individuated cusps and nar-rower cristae strongly developed lingual cingulum that continues to the distal aspectof the crown broad buccal cingular shelf and more distally directed posthypoconecrista In Ekembo the cusps are more bunodont and expanded the cristae are broaderwith better developed accessory tubercles the lingual cingulum merges into thehypocone there is no buccal shelf and the posthypocone crista is more buccallydirected
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6152
is observable even in small samples of Proconsul species TheP africanus holotype has a substantially reduced M3 whereas inKNM-CA 2250dattributed by most authors to this species(Andrews 1978 Pickford et al 2009)dit is fully developed Simi-larly the hypodigm of P major includes both fully developed M3s(eg KNM-CA 389 LG 1815) and specimens with reduced ormissing hypocones (KNM-CA 397 1299) Ekembo displays compa-rable ranges of variation from highly reduced to fully developedM3s in both species Thus these species and genera cannot bedistinguished based on M3 development and we did not find anyother distinguishing characteristics between Ekembo and Proconsulin the upper third molars
Figure 7 Scatter plot depicting the de
Mandible Pickford et al (2009) enumerated several traits thatreadily distinguish Kisingiri from non-Kisingiri mandibles andthese distinctions are entirely consistent with the distinction ofEkembo from Proconsul Our analyses specifically underscoredifferences in the symphyseal region The planum alveolare inboth male and female specimens of Ekembo is short and morevertically oriented even the most elongate of these (KNM-RU47805 McNulty et al 2007) is substantially more verticalcompared to specimens of Proconsul In contrast the planumalveolare in Proconsul is conspicuously long and nearlyhorizontally oriented Well-preserved mandibles attributed toP major (eg KNM-SO 396 and 404) exhibit this morphology inextreme fashion (Fig 8) smaller specimens (KNM-SO 1112P africanus) show less extreme expression but are still readilydistinguishable from Ekembo mandibles For its size KNM-LG 452has a surprisingly short planum alveolare though not to thedegree seen in Rusinga mandibles Correlated to the shortplanum alveolare in Ekembo is a reduced superior transversetorus which while well-developed does not approach therobusticity expressed in Proconsul mandibles This difference isreflected in an index of symphyseal robusticity (thicknessperpendicular to the long axis of the symphysislength of thelong axis) which shows Ekembo and Proconsul to be significantlydifferent (t-test df frac14 14 t frac14 430 p frac14 00007)
Development of the inferior transverse torusdoften used todifferentiate among species of Proconsul (eg Pickford et al 2009Harrison 2010)dwas found to be highly variable and inconsis-tently expressed within these samples However the overallorientation of the subplanum region (as defined by a line con-necting the posterior-most midline points of the superior andinferior transverse tori or the posterior-most midline point of theinferior margin of the symphysis in the absence of the latter) doesdistinguish Ekembo from Proconsul mandibles The orientation ofthis line in relation to the alveolar margin in the vicinity of thepostcanine teeth was assessed qualitatively by placing a small rod
gree of buccolingual flare in M2
Figure 8 Mandibles of Proconsul and Ekembo Occlusal views of P major a) KNM-SO 396 b) KNM-SO 404 c) KNM-LG 452 Occlusal views of Ekembo d) KNM-RU 2087 e) KNM-RU1674 f) KNM-RU 7290 Occlusal views of P africanus g) KNM-SO 372 h) KNM-SO 1112 i) Lingual view of P africanus (KNM-SO 1112) Note the long more horizontally orientedplanum alveolare in Proconsul resulting in a pronounced superior transverse torus The planum alveolare is shorter and more vertical in Ekembo such that one can see the inferiormandibular border from occlusal view
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 53
(eg a dental pick) in themidline internally along the two referencepoints and recording the point in lingual perspective at which therod intersected the alveolar margin (Fig 9) In Ekembo the sub-planum region is more vertically oriented so that the rod intersectsthe alveolar margin most often in the premolar field Not surpris-ingly there is a difference between sexes with females having areduced superior transverse torus and therefore a more verticallyoriented subplanum region compared to males In contrast withEkembo Proconsul mandibles have much more inclined subplanumsurfaces with the rod intersecting the lingual alveolar margin inthe vicinity of the M1 (females) or M2 (males Fig 9) When sex istaken into account therefore the distributions of Ekembo andProconsul in this feature are discrete Note that male and femalemandibles attributed to both P africanus and P major are includedin this assessment and are surprisingly similar despite dramaticsize differences
We also quantified internal symphyseal morphology using threemeasures symphyseal height (infradentaleegnathion) planumalveolare length (posterior-most point on the superior transversetoruseinfradentale) and subplanum length (posterior-most pointon the superior transverse torusegnathion) a combination thatincorporates not only the lengths of these intervals but also theangular relationships among them To compare symphyseal shapeacross a range of sizes we chose a simple principal componentsapproach to partition size onto the first eigenvector (cf Jolicouer1963) While this is not the preferred method for partitioning sizein most datasets (Jungers et al 1995) for this particular sample andset of measurements Mosimann shape variables were still
significantly correlatedwith size (see discussion in Coleman 2008)Hence we assessed differences in symphyseal morphology be-tween Ekembo and Proconsul using the second and third principalcomponents of the three variables Distributions of the two generaare distinct in this shape space (Fig 10 Supplementary OnlineMaterial [SOM] Table 1)
Robusticity of the mandibular corpus also differs between thesesamples though it is more difficult to test due to differentialpreservation among specimens Corpus thickness and depth underP4 can be measured in the greatest number of specimens andcomparison of these measures suggests a difference betweengroups in the scaling of corpus robusticity (Fig 11) Among largepresumably male specimens Ekembo mandibles are substantiallyless robust than those of Proconsul However among smallermandibles only a single Proconsul specimen (KNM-SO 372) could beincluded and it is similar in robusticity to Ekembomandibles of thissize Considered together the impression is that corpus robusticitymay scale differently in Ekembo than it does in Proconsul and a testfor differences in slopes using an ANCOVA design of the loggedvariables was significant (df frac14 2 F frac14 2481 p lt 00001) Never-theless additional specimens are needed to properly evaluate theinfluence of taxonomy sex and size on corpus robusticity areasonable alternative interpretation of Figure 11 is that it dem-onstrates a significant difference between P major and all otherspecies of both Proconsul and Ekembo
Lower incisors Ekembo specimens are uniformly narrower inmesiodistal length than incisors attributed to P africanus P major
Figure 10 Scatter plot of principal component scores based on an eigenanalysis of threeposterior point on the superior transverse toruseinfradentale) and subplanum length (mprimarily captures size differences among specimens (Jolicouer 1963 see text and SOM Trespectively) in the symphyseal variables
Figure 9 Line drawing of a hemimandible depicting different orientations of the sub-planum area in Ekembo and Proconsul Females are denoted using shorter vectors maleswith longer vectors Vector is estimated in M 14086 which is broken at the midlineEkembo specimens have a more vertical subplanum compared to those from Songhorand Legetet but within both groups females have more vertically oriented regions thanmales Hence even the most ventrally rotated subplanum in the largest Ekembo malespecimen (KNM-RU 47805) groups with the Tinderet females in this feature
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6154
and P meswae both absolutely and in relation to crown height(Fig 12) This is true for both I1 and I2 A t-test adjusted for non-independent observations (eg I1 and I2 from the same mandible)confirmed the difference as significant (t-test df frac14 12 t frac14 258p frac14 00242) Moreover while the measurable Proconsul samplewas small this same pattern is observable in other specimenswhose tips were slightly to moderately worn and therefore notmeasured
Lower canines The only lower canine trait that discretely sepa-rates these genera is the pointed crown tip in Ekembo (Fig 3d)contrasted with a blade-like (or ldquoburinrdquo) tip in Proconsul(Fig 3c) This feature is well established for P major but is alsoexpressed in KNM-CA 1772 which was referred to P africanus byKelley (1986) and is too small to be either P major or P meswaeTwo other specimens commonly attributed to P africanus KNM-CA 2149 and KNM-SO 1112 both have broken canine tipsLikewise a new specimen (Kor 65004) attributed to P africanusby Pickford et al (2009) also has a broken tip and cannot beevaluated (see Pickford et al 2009 Fig 23) As with the uppercanines the blade-like tip is also present in the deciduous lowercanines of P major (eg KNM-CA 361 S0 5352) and P meswae(KNM-ME 1)
Lower premolars Like most previous authors (eg Andrews 1978Harrison 2002 2010 Pickford et al 2009) we found neitherqualitative nor quantitative characteristics of the lower premolarswith any taxonomic value
Lower molars Differences in lower molar morphology are few andrelatively subtle but in some cases mirror those seen in the uppermolars Cusps on the lower molars of Ekembo are more bunodontthan those in Proconsul and occupy more of the occlusal surfaceBroad inter-cuspal crests are incorporated into the Ekembo cuspsmaking them appear more inflated Molar cusps of Proconsul are
variables symphyseal height (infradentaleegnathion) planum alveolare length (mostost posterior point on the superior transverse torusegnathion) The first componentable 1) and thus PC 2 and PC 3 illustrate the residual shape variance (72 and 28
Figure 11 Scatter plot showing mandibular corpus robusticity under P4 Trend lines were computed using ordinary least squares (OLS) regression Similar relationships areobservable at other parts of the corpus but differential preservation results in smaller samples at those positions Slopes are significantly different (p lt 00001 tested on loggedvariables)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 55
more distinct from one another and are linked by narrower sharperinter-cuspal crests The lower molars of Ekembo also have asubstantially less developed buccal cingulum than those ofProconsul (Harrison 2002 2010 Harrison and Andrews 2009)
Figure 12 Scatter plot of lower incisor crown proportions Differenc
Lastly and again as seen in the upper molars the two genera canbe distinguished by the significantly greater amount of lowermolar flare in Proconsul (Fig 13 t-test df frac14 59 t frac14 713p lt 00001 see also Harrison and Andrews 2009)
es between Ekembo and Proconsul are significant (p frac14 00242)
Figure 13 Box-and-whisker plot of buccolingual flare in lower molars measured asthe average distance between the mesial buccal and lingual cusps divided by overallbuccolingual crown breadth Differences between Ekembo and Proconsul are highlysignificant (p lt 00001)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6156
Relative molar size has been used to distinguish species ofProconsul but we observed the primary difference to be in relativebuccolingual breadths of M1 and M2 In Ekembo M1 is substantiallynarrower than M2 than is the case in Proconsul (Fig 14 Harrison2002 2010) Although none of the smaller Proconsul mandiblespreserve complete crowns for both M1 and M2 KNM-SO 1112 hasan intact M2 and at least preserves the cervix for M1 However evenif the cervical measurement is used for its M1 breadthdthusunderestimating the true crown breadthdKNM-SO 1112 still has abroader first molar in relation to M2 than would be expected inEkembo If crown breadth is estimated from the ratio of cervixbreadth to maximum crown breadth in other Proconsul fossils therange of likely breadths for the SO 1112 M1 clearly distinguishes it
from the Ekembo sample (Fig 14) Testing this relationship onlogged variables using the mean estimate for the SO 1112 M1(Fig 14) reveals a significant difference between the two groups(ANCOVA df frac14 1 F frac14 2424 p frac14 00002)
As with upper third molars we find a great deal of variation inM3 morphology and mistrust the taxonomic significance some-times placed on characteristics of this toothdat least with regard tothese taxa Harrison (2002 2010) cited the mesiodistal position ofthe M3 entoconid as differentiating Kisingiri third molars fromthose attributed to P africanus We concur with his assessment butfound the entoconid position to vary in specimens attributed toP major Thus this character does not differentiate Ekembo from allProconsul species but may distinguish it from the type speciesP africanus
Discussion
The distinctiveness of the Kisingiri large catarrhine materialfrom that at Tinderet and Ugandan localities has been noted pre-viously by several authors (Pickford 1986 Walker et al 1993Begun 2001 2007 MacLatchy and Rossie 2005 Harrison andAndrews 2009 Harrison 2010) What had not been fully docu-mented was the nearly complete lack of overlap between thesesamples in a number of features despite very large assemblagesfrom Rusinga and Mfangano On descriptive criteria alone (Table 2)many isolated teeth (I1 C1 P3 M2 C1) and all of the mandibles canbe assigned to either Proconsul or Ekembowith very little chance ofmisattribution That some of the most diagnostic features occur inthe upper dentitiondthe only anatomy preserved in theP africanus type specimendadds weight to the taxonomicarrangement proposed here The morphology of the upper teeth inall Proconsul species compellingly unites them into a single cladedistinct from E heseloni and E nyanzae This is reinforced bynumerous features of the mandible and mandibular dentition thatfurther distinguish these samples The number and variety of thesefeatures are such that Proconsul and Ekembo are more differentfrom one another than are many closely-related modern catarrhinegenera
Derived morphology of Ekembo and Proconsul
It is premature to formally assign primitive and derived status tothe individual features of Ekembo and Proconsul before completinga formal phylogenetic analysis This is especially true given theabundance of evidence available for Ekembo and hence the poten-tial impact that our generic revision could have on phylogeneticinterpretations Nevertheless in comparison with both extanthominoids and the small-bodied catarrhines from the earlyMiocene it is informative to differentiate the evolutionary grades ofthese genera Many characteristics of Ekembo are more advanced inthe direction of modern apes compared to those of Proconsul(Table 2) General trends such as expanded and more bunodontcusps broader molar crests reduced cingulum and morphologicalsimilarity between P3 and P4 all represent morphologies thatpresage those of later fossil and extant hominoids The moreadvanced morphologies of P3 and M2 are particularly conspicuouscompared not only with those of Proconsul but to most earlyMiocene catarrhines In contrast the mesially positioned stronglyheteromorphic cusps on the P3 of Proconsul plus the correspondingasymmetric buccal profile notched mesial margin and missingmesial fovea are characteristic of other early Miocene catarrhinessuch as Dendropithecus Rangwapithecus and Limnopithecus andare likely primitive among East African catarrhines The moresymmetric P3 of Ekembo with midline positioned cusps andreduced cusp heteromorphy is reminiscent of later hominoids and
Figure 14 Relative buccolingual dimensions of M1 and M2 KNM-SO 1112 has a preserved M2 but only the cervix of M1 Its M1 breadth was estimated based on the ratio to cervixbreadth to maximum crown breadth in other Tinderet fossils Even the cervical breadth of SO 1112 is broader than would be predicted for the crown breadth of a Kisingiri M1 Trendlines computed using OLS regression and using the average estimate of SO 1112 M1 breadth Group differences were significant (p frac14 00002)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 57
even hominid taxa such as Rudapithecus (eg Kordos and Begun2001)
Proconsul is likewise united by a suite of features that are for themost part characteristic only of this lineage (Table 2) Themorphology of both upper and lower canines is unique to thisgenus and shared by all three well-known species plus the poorlyrepresented middle Miocene P gitongai (Pickford and Kunimatsu2005) The mandibular symphysis is also distinctive in Proconsulcompared with other catarrhines with its very long and shallowlysloping planum alveolare and steeply inclined subplanum surfaceIndeed even the small edentulous anterior mandibular fragmentKNM-SO 372 is readily placed within Proconsul and excluded fromother similarly sized catarrhine taxa such as Rangwapithecus andEkembo based on this unique symphyseal morphology (Figs 8 and9)
Other differences between Ekembo and Proconsul are of uncer-tain evolutionary polarity (eg orientation of the posthypoconecrista) or may be of taxonomic relevance without broader phylo-genetic significance (eg degree of molar flare Table 2) Werecognize that there are morphological differences within Procon-sul particularly between the better known species P africanus andP major However many of these differences are likely related tosize and represent variations on a single generic theme that aremarkedly different from the morphology of Ekembo The recentdescription (Harrison and Andrews 2009) of the intermediatelysized P meswae underscores the broad morphological unity withinProconsul and further substantiates a well-defined Proconsul cladecomprising Tinderet and Ugandan species
Based on the abundant morphological and morphometric evi-dence supporting genus-level distinction between Proconsul andEkembo we are currently conducting a more detailed phylogeneticanalysis of all early Miocene catarrhines in order to better under-stand the distribution and evolutionary significance of these andother morphological features We anticipate that the debates sur-rounding the phylogenetic relationships of an historically consti-tuted sample of Proconsul can be more satisfactorily resolved nowthat the Kisingiri material is appropriately recognized as a distinctgenus
The generic disposition of ldquoUgandapithecusrdquo species
A great many features said to characterize ldquoUgandapithecusrdquowere added to the original diagnosis of the genus by Pickford et al(2009) in their systematic revision of Proconsul (sensu lato) As partof that study they also erected a new species ldquoUrdquo legetetensis andtransferred P meswae to ldquoUgandapithecusrdquo Evaluating the same setof features across the entire available sample however we wereunable to find support for this more expansive conception ofldquoUgandapithecusrdquo for several reasons First some diagnostic fea-tures cited in Pickford et al (2009) were quite variable within theldquoUgandapithecusrdquo sample As an example marked reduction of theM3 entoconid to a bead on the lingual cingulum is not shared byseveral ldquoUrdquo major specimens such as KNM-CA 393 NAP I 49000and Kor 253004 (see Pickford et al 2009 Fig 33e) and was likelynot a feature of a few of the more worn specimens eg BUMP 600and BUMP 601 or even the holotype of P major M 16648 Second
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6158
some diagnostic features of ldquoUgandapithecusrdquo were found to bevariably present in Ekembo arguing against their utility for dis-tinguishing ldquoUgandapithecusrdquo from Pickford et als (2009) com-bined P africanus thorn Kisingiri conception of Proconsul For examplesignificantly reduced curvature of the I1 distal margin in ldquoUganda-pithecusrdquo is also found in some Kisingiri specimens including KNM-RU 1685 1712 1714 1831 1846 and 1951 several more Rusingaspecimens have moderately reduced curvature Third some fea-tures that purportedly distinguish ldquoUgandapithecusrdquo fromP africanus are likely related to size For example the somewhatmore central position of the P3 paracone in ldquoUrdquomajor is not seen inthe smaller ldquoUrdquo meswae whose paracone is positioned similarly tothat of the holotype of P africanus (see Harrison and Andrews2009 Fig 6d)
However the major part of the evidence for recognizingldquoUgandapithecusrdquo (sensu Pickford et al 2009) was assembled by re-assigning specimens traditionally attributed to P africanus into newspecies of ldquoUgandapithecusrdquo The P africanus holotype was left inassociation with the Rusinga sample despite having morphologyoverwhelmingly characteristic of other Tinderet specimens Theresulting specimen allocations left P africanus morphologicallyimpoverisheddstripped of much of its hypodigmdso that Pro-consul for Pickford et al (2009) came to be characterized primarilyby the Rusinga sample (see similar comments in Harrison andAndrews 2009) Within that conceptual framework any spec-imen that differs from those at Rusinga could not be Proconsul butmust instead belong to ldquoUgandapithecusrdquo Thus specimens such asthe KNM-SO 1112 mandible were erroneously assigned to ldquoUgan-dapithecusrdquo because they were (correctly) perceived as differingfrom their Rusinga counterparts (Pickford et al 2009)
Here we restore the P africanus hypodigm and associate it withthe other Tinderet fossils according to their shared morphologySpecimens assigned to ldquoUgandapithecusrdquo revert to Proconsul andmany of the features cited to support a speciose ldquoUgandapithecusrdquo(Pickford et al 2009) instead become diagnostic features of Pro-consul This correction serves to highlight the distinctiveness of theKisingiri remains and the need to recognize this distinctiveness atthe generic level
As demonstrated above specimens from Kisingiri do not displaythe morphology of Proconsul as represented by the holotype ofP africanus M 14084 Given that this specimen preserves only theupper canine and cheek teeth similarities with or departures fromthis morphology represent the most compelling arguments forassessing inclusivity in the genus However for the mandible andlower dentition if one simply sorts the specimens by elementwithout relying on taxonomic preconceptions the overwhelmingdistinction within the traditionally constituted Proconsul sample isbetween Kisingiri material on the one hand and that from Tinderetand Uganda on the other (cf Walker et al 1993 Begun 2001 2007MacLatchy and Rossie 2005 Harrison and Andrews 2009Harrison 2010) Additionally with the Kisingiri specimensremoved to Ekembo the postcranial differences used to partlyjustify a distinct ldquoUgandapithecusrdquo lose their relevance (seeHarrison and Andrews 2009)
We have differed among ourselves in our past treatment ofP major as generically distinct from the smaller species of Pro-consul (Begun 2007 McNulty 2010) Morphologically many ofthe unique features of P major can be interpreted as size-relatedvariants on the smaller P africanus morphology Even if so thequestion is whether these are sufficiently different and numerousto warrant generic distinction that is do they suggest thatP major belongs in a separate clade from P africanus andP meswae As we have demonstrated differences among thesethree species are certainly fewer and show less morphologicaldistinction than those between Ekembo and any of the three
Nevertheless while we ultimately agree with Harrison andAndrews (2009 see also Begun 2007) that the features dis-tinguishing ldquoUrdquo major from P africanus are consistent with thosethat distinguish congeneric species and are at least partly due todifferences in size we concede the possibility that the largestProconsul species could belong in a separate genus (Senut et al2000 Pickford and Kunimatsu 2005) However whether or notone recognizes a separate genus for P major the P africanus ma-terial does not sort morphologically with the Kisingiri materialTherefore the taxonomic alternatives are either two generaProconsul and Ekembo or three Proconsul Ugandapithecus andEkembo with Proconsul and Ugandapithecus united in a cladeexclusive of Ekembo Thus while we strongly favor the firstalternative recognizing Ugandapithecus does not change thefundamental argument concerning the need to refer the Kisingirispecies to a generically and cladistically distinct taxon from thespecies represented at Tinderet and the Ugandan sites
Specimens of interest
Despite the substantial dental and gnathic differences betweenProconsul and Ekembo there are a few specimens whose taxonomicaffinities require additional scrutiny
KNM-RU 1676-77 This set of associated upper and lower teeth is aconsistent outlier among E heseloni and E nyanzae specimens Insome ways this specimen more closely resembles Tinderet Pro-consul in the features that distinguish it from Ekembo Molar cuspmorphology and cingulum development I1 lingual tubercledevelopment and M2 shape and posthypocone crista orientationare more consistent with specimens from the Tinderet sites On theother hand it otherwise resembles Ekembo and lacks the strongestdiagnostic features that unite species of Proconsul such as thedistinctive canine and P3 morphology Overall this specimen doesnot fit unambiguously into either genus in fact the degree ofbuccolingual flare cusp morphology enamel wrinkling and espe-cially the long extension of enamel onto the anterior root in P3 arein some ways reminiscent of Afropithecus although it clearly differsfrom that genus as well (cf Rossie and MacLatchy 2013)
Were these characteristics individually distributed among thelarge Kisingiri sample one might attribute this to intra-genericvariation but when found together in a single specimen from alarge sample it suggests a unique individual One of us (KPM)thinks it likely that the specimen was not actually found at aKisingiri locality based on its unique preservation compared toother Rusinga fossils and on other examples of mis-proveniencedspecimens (Leakey 1967 Andrews and Molleson 1979 Pickford1986 Kelley 1986) However we recognize that relying toofrequently on this explanation allows almost any scenario to bejustified and suggest some alternative possibilities One is that thespecimen was found on Rusinga but in the much older WayandoFormation strata instead of the Hiwegi Formation The differencesin morphology in that case might reflect more than a million yearsof temporal change (McCollum et al 2013) with KNM-RU 1676-77preserving certain more primitive Proconsul-like features As analternative it is reasonable to assume that fossil assemblages asvast as those from Rusingawill include not only abundant taxa butalso rare or infrequent inhabitants of the local biome There aremany examples of this from Rusingas mammalian communityeg ldquoHemicyonrdquo Kulutherium Tadarida cf Hyainailouros cfArchaeobeledon Kelba (see relevant chapters in Werdelin andSanders 2010) As such KNM-RU 1676-77 may simply representa rare and well-preserved specimen of some other catarrhinetaxon that lived among or visited a primate community dominatedby Ekembo
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 59
KNM-SO 1112 This specimen was initially referred to Rangwapi-thecus but soon after was transferred to Proconsul afrianus (Bosler1981 Martin 1981)da designation that most subsequent authorshave followed (Pickford 1986 Nengo and Rae 1992 Kelley 19861995 Walker et al 1993 MacLatchy and Rossie 2005 Hill et al2013) Pickford et al (2009) more recently referred this specimento ldquoUrdquo meswae citing two primary reasons First they reiteratedBoslers (1981) concern that its P3 differs from others attributedto P africanus However at that time the majority of what wasconsidered to be P africanus would have been from RusingaIsland In fact while noting its contrasting morphology Boslersuggested that differences between KNM-SO 1112 and other P3smight reasonably be attributed to sexual dimorphism notingthat this specimen provides ldquoa not implausible match for the Pafricanus holotype maxilla M 14084rdquo (1981152) Thatassessment is borne out in this study Using the much largersample of Ekembo P3s as a model variation can be seen toencompass specimens with well-developed mesial beaks andthose with more bilaterally compressed crowns and parallelsides The latter morphology is predominantly found amongmale specimens (KNM-RU 1674 1924 and KPS I and III) thussupporting Boslers (1981) suggestion that this variation isrelated to sexual dimorphism and hence that KNM-SO 1112 is amale P africanus
The second feature cited by Pickford et al (2009) in referringthis specimen to ldquoUrdquo meswae was posterior shallowing of themandibular corpus However because they redefined ldquoUgandapi-thecusrdquo to include all of the well-preserved Tinderet mandibles(Pickford et al 2009)dleaving Proconsul to be represented only byRusinga specimensdtheir only alternative within this scheme for aspecimen the size of SO 1112 was to refer it to ldquoUrdquomeswae It seemsodd though to assign this specimen to a species from a (perhapsmuch) older site when the Proconsul africanus holotype is ofappropriate size and similar age and as there are no other adultmandibles from either species with which to compare it In anycase mandibular shallowing is variable in both Ekembo and Pro-consul with some Kisingiri specimens exhibiting considerableshallowing (eg KNM-RU 2087) and some ldquoUgandapithecusrdquo spec-imens having very little (eg M 14086)
We therefore see no morphological grounds to assign KNM-SO1112 to P meswae but do find some morphological evidence toassign it as a male P africanus which occurs at Songhor
KNM-LG 452 This very complete mandible was originally recog-nized as a female of P major based on the size of the dentition androbusticity of the mandible (Martin 1981) Bosler (1981) howeverleft this specimen unassigned noting similarities to P major in P3-M1 size but having distinctly smaller M2e3 In this she likenedKNM-LG 452 to a few other specimens (notably M 14086) thatare smaller in their distal molars in a manner more consistentwith taxonomic rather than sex differences (Bosler 1981 see alsoPilbeam 1969) Kelley (1986 1995) also assigned this specimen asa female of P major based on canine size and morphology butnoted that the canine lacked the shallowly-sloping mesial ridgetypical of most female anthropoid canines This is also truehowever of two other canines assigned as female P major KO 9and SO 373 (Kelley 1986) and so might simply be an unusualcharacteristic of P major female lower canines In the end Kelley(1995) concluded that considering all of the criteria fordistinguishing male and female canines the evidence that KNM-LG 452 is female remains questionable
Pickford et al (2009) further developed the idea that two spe-cies are represented within the P major sample and made KNM-LG452 the holotype of a new species ldquoUrdquo legetetensis We concur thatthis specimen is unique among the well-preserved Tinderet
mandibles in some important features Whereas male and femalemandibles of P major have long internal symphyseal planesexpansive extra- and retromolar sulci and rami that cross theocclusal plane at distal M3 KNM-LG 452 has a shorter symphysealplane (though not to the extent seen in Ekembo) a moderateextramolar sulcus but no retromolar sulcus and a ramus thatcrosses the occlusal plane more mesially If KNM-SO 396 and SO404 represent males and females respectively of P major then themorphology of KNM-LG 452 appears distinct
At present however we do not think there is sufficient evidenceto differentiate this specimen from either P major or P meswaePickford et al (2009) argue that KNM-LG 452 cannot belong to thelatter species because of its larger size The size differences how-ever are well within the range of variation seen in P majorE heseloni and E nyanzae Moreover whereas the Legetet mandibleis larger than most of the P meswae hypodigm an adult P4 referredto this species (KNM-ME 25) substantially exceeds its counterpartin KNM-LG 452 Should the Legetetmandible ultimately prove to bea male specimen size alone would not rule out its attribution toP meswae On the other hand if it is female then it becomes harderto justify its inclusion in that sample of mostly smaller specimens Iffemale therefore it could represent a separate species for whichthe binomial Proconsul legetetensis comb nov is available Recentlycollected fossils from the SonghorKapurtay area provide someadditional support for the possibility that three species of Proconsulexisted at these localities (McNulty 2014ab) However toconvincingly demonstrate this and determine whether a thirdspecies can be differentiated from P meswae will require a moredetailed analysis of variation among Proconsul species a corre-sponding study of variation in modern taxa anddalmost certain-lydadditional fossil specimens For now we regard ldquoUrdquo legetetensisas a subjective junior synonym of P major
Conclusions
The genus Proconsul has been central to studies of ape and hu-man evolution since its first description by Hopwood (1933) As oneof the best known fossil catarrhines represented by nearly everyskeletal element our knowledge of Proconsuls functional anatomypositional behavior and overall morphology has made it thefoundation for paleoanthropological research in the early Mioceneas well as studies on the origin and early diversification of homi-noids With respect to systematics however Proconsul is less wellunderstood the type species P africanus is known from a rela-tively small number of specimens from sites in the Tinderet areaand no postcranial bones can be unambiguously assigned to itRather it is the copious and well-preserved specimens from theKisingiri sites of Rusinga and Mfangano Islands that have formedthe basis for the common image of Proconsul
We argue here that the Kisingiri species traditionally referredto ProconsuldP heseloni and P nyanzaedare generically distinctfrom P africanus and hereby transfer them into a new genusEkembo based on a substantial number of morphological differ-ences (Table 2) Considering only those traits preserved in theP africanus holotype the entire sample from Rusinga and Mfan-gano differs markedly in the morphology of the upper canine P3P4 and M2 Additional differences in the I1 lower incisors andcanines molar cusp and occlusal crest morphology degree of flarein all molars and especially mandibular features clearly distin-guish the Kisingiri from non-Kisingiri samples in a way that isincompatible with congeneric species With the transfer ofE heseloni and E nyanzae Proconsul becomes a well-defined cladecomprising at least three speciesdP africanus P major andP meswaedunited by several features that are unique for thegenus (Table 2)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6160
We found no support in this study for the genus ldquoUgandapi-thecusrdquo as constructed by Pickford et al (2009) In that paper theytransferred the vast majority of Proconsul fossils from the Tinderetand Napak localities into ldquoUgandapithecusrdquo leaving P africanusrepresented only by its holotype and a small number of isolatedteeth and aligning it with the material from Rusinga and Mfan-ganoWhen the P africanus hypodigm is reconstituted and properlyaffiliated with the other Tinderet species according to itsmorphology much of the evidence marshaled by Pickford et al(2009) for ldquoUgandapithecusrdquo actually supports the distinction be-tween the Tinderet and Kisingiri samples ie the generic distinc-tion of Ekembo In addition some of the traits enumerated byPickford and colleagues in support of ldquoUgandapithecusrdquo wererevealed in our analysis to be variable within one or more speciesand therefore of questionable taxonomic utility We do not rejectout of hand the original conception of ldquoUgandapithecusrdquo (Senutet al 2000) as either a monotypic genus distinguishing ldquoUrdquo ma-jor or perhaps including a closely allied middle Miocene ldquoUrdquogitongai as well (Pickford and Kunimatsu 2005) but we have fornow retained the early Miocene specimens of both in Proconsulpending further study However even if Ugandapithecus is a taxo-nomically valid genus comprising just the latter two species theirmorphology indicates that theywould still be united in a cladewithProconsul and they lack several more advanced features that partlydefine Ekembo Better understanding of the role that body size playsinmediating differences within Proconsulwill help to further clarifythe relationships among these species
Despite the existence of one well-represented and several lesscomplete skeletons of what was long assumed to be Proconsul thephylogenetic status of this genus has defied consensus This islargely due in our view to Proconsul having been an admixture oftwo distinct genera one more advanced in the direction of extantapes than the other As we demonstrate here the material fromRusinga and Mfangano traditionally referred to Proconsul andwhich includes all of the more complete skeletal material isdistinct from the Tinderet and Napak material and belongs in aseparate genus Ekembo By disentangling these distinct clades weanticipate being able to achieve better phylogenetic resolution forboth genera thereby improving our knowledge of early hominoidevolution and diversification
Acknowledgments
We thank many people and agencies that assisted with thedevelopment and completion of this project This research wasinitiated as a result of funding from the Wenner-Gren Foundationand the Johns Hopkins School of Medicine to DR Begun in1987e1989 and was renewed following research and travelgenerously funded by multiple Leakey Foundation grants to KMcNulty National Science Foundation grants to K McNulty(0852609 1241807) the McKnight Foundation the University ofMinnesota the Lloyd A Wilford Endowment the LeverhulmeFoundation and the Turkana Basin Institute J Kelley gratefullyacknowledges support from the Institute of Human Origins Ari-zona State University DR Begun also wishes to acknowledge theAlexander Humboldt Stiftung and NSERC for funding support Weare grateful to the National Museums of Kenya the UgandaMuseum the Natural History Museum and to Rose Mwanja EzraMusiime Tom Mukhuyu Samuel Muteti James Yatich and MaryMuungu for access to and assistance with specimens in their careMany people provided assistance comments criticisms insightsand suggestions on various aspects of this work and we especiallythank Holly Dunsworth Will Harcourt-Smith Tom Lehmann DanPeppe Nicole Garrett Kirsten Jenkins Rutger Jansma Susy CoteIsaiah Nengo Andrew Hill and Brenda Benefit K McNulty
acknowledges the evolutionary anthropology faculty and studentsat Durham University for their helpful comments on an earlyversion of this manuscript and for their gracious hospitality Weare likewise grateful to the anonymous reviewers associate editorand editor whose comments strengthened the paper This manu-script is publication 1 supporting Research on East AfricanCatarrhine and Hominoid Evolution (REACHE)
Appendix A Supplementary data
Supplementary data related to this article can be found at httpdxdoiorg101016jjhevol201503009
References
Andrews P 1974 New species of Dryopithecus from Kenya Nature 249 188e190Andrews P 1978 A revision of the Miocene Hominoidea of East Africa Bull Brit
Mus (Nat Hist) Geol 30 85e225Andrews P 1985 Family group systematics and evolution among catarrhine pri-
mates In Delson E (Ed) Ancestors The Hard Evidence Liss New Yorkpp 14e22
Andrews P Harrison T Martin L Pickford M 1981 Hominoid primates from anew Miocene locality named Meswa Bridge in Kenya J Hum Evol 10123e128
Andrews P Begun D Zylstra M 1997 Interrelationships between functionalmorphology and paleoenvironments in Miocene hominoids In Begun DWard C Rose M (Eds) Function Phylogeny and Fossils Miocene HominoidEvolution and Adaptations Plenum Press New York pp 29e58
Andrews P Molleson T 1979 The provenance of Sivapithecus africanus Bull BritMus (Nat Hist) Geol 32 19e23
Begun DR 2001 African and Eurasian Miocene hominoids and the origins of theHominidae In de Bonis L Koufos GD Andrews P (Eds) Hominoid Evolutionand Climatic Change in Europe Phylogeny of the Neogene Hominoid Primatesof Eurasia Cambridge University Press London pp 231e253
Begun DR 2007 Fossil record of Miocene hominoids In Henke W Tattersall I(Eds) Handbook of Paleoanthropology Springer Berlin pp 921e977
Begun DR Teaford MF Walker AC 1994 Comparative and functional anatomyof Proconsul phalanges from the Kaswanga primate site Rusinga Island KenyaJ Hum Evol 26 89e165
Bishop WW 1964 More fossil primates and other Miocene mammals from North-East Uganda Nature 203 1327e1331
Bishop WW Miller JA Fitch FJ 1969 New potassium-argon age determinationsrelevant to the Miocene fossil mammal sequence in East Africa Am J Sci 267669e699
Blankenship JA Mincer HH Anderson KM Woods MA Burton EL 2007Third molar development in the estimation of chronologic age in Americanblacks as compared with whites J Forensic Sci 52 428e433
Bosler W 1981 Species groupings of early Miocene dryopithecine teeth from EastAfrica J Hum Evol 10 151e158
Coleman MN 2008 What does geometric mean mean geometrically Assessingthe utility of geometric mean and other size variables in studies of skullallometry Am J Phys Anthropol 135 404e415
Cote S Malit N Nengo I 2013 Additional mandibles of Rangwapithecus gordonian early Miocene catarrhine from the Tinderet localities of Western KenyaJ Hum Evol 153 341e352
Deane AS 2009 Early Miocene catarrhine dietary behavior the influence of theRed Queen Effect on incisor shape and curvature J Hum Evol 56 275e285
Drake R Van Couvering JA Pickford M Curtis G Harris JA 1988 New chro-nology for the Early Miocene mammalian faunas of Kisingiri Western KenyaJ Geol Soc Lond 145 479e491
Evernden JF Savage DE Curtis GH James GT 1964 Potassium-argon datesand the Cenozoic mammalian chronology of North America Am J Sci 262145e198
Gebo DL MacLatchy L Kityo R Deino A Kingston J Pilbeam D 1997A hominoid genus from the Early Miocene of Uganda Science 276 401e404
Geoffroy Saint-Hilaire E 1812 Tableau des quadrumanes 1 Ord QuadrumanesAnnls Mus Hist Nat Paris 19 85e122
Gray JE 1825 Outline of an attempt at the disposition of the Mammalia into tribesand families with a list of the genera apparently appertaining to each tribeAnnls Philos 10 337e344
Harrison T 1982 Small-bodied apes from the Miocene of East Africa PhDDissertation University of London
Harrison T 1986 New fossil anthropoids from the middle Miocene of East Africaand their bearing on the origin of the Oreopithecidae Am J Phys Anthropol 71265e284
Harrison T 1987 The phylogenetic relationships of the early catarrhine primates areview of the current evidence J Hum Evol 16 41e80
Harrison T 2002 Late Oligocene to middle Miocene catarrhines from Afro-ArabiaIn Hartwig WC (Ed) The Primate Fossil Record Cambridge University PressCambridge pp 311e338
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 61
Harrison T 2010 Dendropithecoidea Proconsuloidea and Hominoidea (CatarrhiniPrimates) In Werdelin L Sanders WJ (Eds) Cenozoic Mammals of AfricaUniversity of California Press Berkeley pp 429e469
Harrison T Andrews P 2009 The anatomy and systematic position of a newspecies of early Miocene proconsulid from Meswa Bridge Kenya J Hum Evol56 479e496
Hill A Nengo IO Rossie JB 2013 A Rangwapithecus gordoni mandible from theearly Miocene site of Songhor Kenya J Hum Evol 65 490e500
Hopwood AT 1933 Miocene primates from Kenya J Linn Soc Zool 38 437e464Ishida H Kunimatsu Y Takano T Nakano Y Nakatsukasa M 2004 Nacholapi-
thecus skeleton from the Middle Miocene of Kenya J Hum Evol 46 69e103Jolicouer P 1963 The multivariate generalization of the allometry equation Bio-
metrics 19 497e499Jungers WL Falsetti AB Wall CE 1995 Shape relative size and size-
adjustments in morphometrics Am J Phys Anthropol 38 (S2) 137e161Kavanagh KD Evans AR Jernvall J 2007 Predicting evolutionary patterns of
mammalian teeth from development Nature 449 427e433Kelley J 1986 Species recognition and sexual dimorphism in Proconsul and
Rangwapithecus J Hum Evol 15 461e495Kelley J 1995 Sex determination in Miocene catarrhine primates Am J Phys
Anthropol 96 390e417Kelley J 1993 Taxonomic implications of sexual dimorphism in Lufengpithecus In
Kimbel WH Martin LB (Eds) Species Species Concepts and Primate Evo-lution Springer New York pp 429e458
Kent PE 1942 The country round the Kavirondo Gulf of Victoria Nyanza Geogr J100 22e31
Kordos L Begun DR 2001 A new cranium of Dryopithecus from RudabanyaHungary J Hum Evol 15 689e700
Le Gros Clark WE Leakey LSB 1950 Diagnoses of East African Miocene Homi-noidea Q J Geol Soc Lond 105 260e263
Le Gros Clark WE Leakey LSB 1951 The Miocene Hominoidea of East Africa InFossil Mammals of Africa No 1 British Museum (Natural History) London
Leakey LSB 1943 A Miocene anthropoid mandible from Rusinga Kenya Nature152 319e320
Leakey LSB 1963 East African fossil Hominoidea and the classification within thissuper-family In Washburn SL (Ed) Classification and Human EvolutionAldine Chicago pp 32e49
Leakey LSB 1967 Notes on the mammalian faunas from the Miocene and Pleis-tocene of East Africa In Bishop WW Clark JD (Eds) Background to Evolu-tion in Africa University of Chicago Press Chicago pp 7e29
Leakey RE Leakey MG 1986 A new Miocene hominoid from Kenya Nature 324143e146
Leakey RE Leakey MG Walker AC 1988 Morphology of Afropithecus turka-nensis from Kenya Am J Phys Anthropol 76 289e307
Leakey MG Ungar PS Walker A 1995 A new genus of large primate from theLate Oligocene of Lothidok Turkana District Kenya J Hum Evol 28519e531
Linnaeus C 1758 Systema naturae per regna tria naturae secundum classesordines genera species cum characteribus differentris synonymis locis tenthed Laurentii Salvii Stockholm
MacInnes DG 1943 Notes on the East African Miocene primates J E Afr UgandaNat Hist Soc 17 141e181
MacLatchy L 2004 The oldest ape Evol Anthropol 13 90e103MacLatchy L Bossert WH 1996 An analysis of the articular surface distribution of
the femoral head and acetabulum in anthropoids with implications for hipfunction in Miocene hominoids J Hum Evol 31 425e453
MacLatchy L Rossie JB 2005 The Napak hominoid still Proconsul major InLieberman DE Smith RJ Kelley J (Eds) Interpreting the Past Essays onHuman Primate and Mammal Evolution in Honor of David Pilbeam Brill Ac-ademic Publishers Boston pp 15e28
Martin L 1981 New specimens of Proconsul from Koru Kenya J Hum Evol 10139e150
McCollum MS Peppe DJ McNulty KP Dunsworth HM Harcourt-Smith WEH Andrews AL 2013 Magnetostratigraphy of the early MioceneHiwegi Formation (Rusinga Island Lake Victoria Kenya) Geological Society ofAmerica Abstracts with Programs 45 p 12
McNulty KP 2010 Apes and tricksters the evolution and diversification ofhumans closest relatives Evol Edu Outreach 3 322e332
McNulty KP Harcourt-Smith WEH Dunsworth HM 2007 New primate fossilsfrom Rusinga Island Kenya Am J Phys Anthropol 132 (S44) 170
McNulty KP MacLatchy L Peppe DJ Nengo I Manthi FK Miller ERStevens NJ Cote S Lehmann T Kingston J 2014a A regional approach toEast African early Miocene paleobiology J Vert Paleontol 185 SVP Programand Abstracts Book 2014
McNulty KP MacLatchy L Rossie JB Peppe DJ Deino AL Mbua ENManthi FK Nengo IO Stevens NJ Cote S Lehmann T Gutierrez M 2014b
Research on East African catarrhine and hominoid evolution results from thefirst year Am J Phys Anthropol 153 (S58) 182
Michel LA Peppe DJ Lutz JA Driese SG Dunsworth HM Harcourt-Smith WEH Horner W Lehmann T Nightengale S McNulty KP 2014Remnants of an ancient forest provide ecological context for Early Miocenefossil apes Nature Comm 5 3236
Myres JL 1932 Africa Man 32 208Nakatsukasa M Kunimatsu Y Nakano Y Takano T Ishida H 2007 Vertebral
morphology of Nacholapithecus kerioi based on KNM-BG 35250 J Hum Evol52 347e369
Nengo IO Rae TC 1992 New hominoid fossils from the early Miocene site ofSonghor Kenya J Hum Evol 23 423e429
Peppe DJ McNulty KP Cote SM Harcourt-Smith WEH Dunsworth HM VanCouvering JA 2009 Stratigraphic interpretation of the Kulu Formation (EarlyMiocene Rusinga Island Kenya) and its implications for primate evolutionJ Hum Evol 56 447e461
Pickford M 1981 Preliminary Miocene mammalian biostratigraphy for westernKenya J Hum Evol 10 73e97
Pickford M 1986 Sexual dimorphism in Proconsul Hum Evol 1 111e148Pickford M Andrews PJ 1981 The Tinderet Miocene sequence in Kenya J Hum
Evol 10 11e33Pickford M Kunimatsu Y 2005 Catarrhines from the Middle Miocene (ca 145
Ma) of Kipsaraman Tugen Hills Kenya Anthropol Sci 113 189e224Pickford M Senut B Gommery D Musiime E 2009 Distinctiveness of Ugan-
dapithecus from Proconsul Estudios Geol 65 183e241Pilbeam DR 1969 Tertiary Pongidae of East Africa evolutionary relationships and
taxonomy Bulletin of the Peabody Museum of Natural History 31 pp 1e185Rae TC 1993 Phylogenetic analysis of proconsulid facial morphology PhD
Dissertation State University of New York at Stony BrookRae TC 1999 Mosaic evolution in the origin of the Hominoidea Folia Primatol 70
125e135Rafferty KL Walker A Ruff C Rose MD Andrews PJ 1995 Postcranial esti-
mates of body weights in Proconsul with a note on a distal tibia of P major fromNapak Uganda Am J Phys Anthropol 97 391e402
Rossie JB MacLatchy L 2013 Dentognathic remains of an Afropithecus individualfrom Kalodirr Kenya J Hum Evol 65 199e208
Senut B Pickford M Gommery D Kunimatsu Y 2000 Un nouveau genredrsquohominoiumlde du Miocene inferieur drsquoAfrique orientale Ugandapithecus major(Le Gros Clark and Leakey 1950) CR Acad Sci Paris 331 227e233
Simons EL Pilbeam DR 1965 Preliminary revision of the Dryopithecinae (Pon-gidae Anthropoidea) Folia Primatol 3 81e152
Smith TM Martin LB Leakey MG 2003 Enamel thickness microstructure anddevelopment in Afropithecus turkanensis J Hum Evol 44 283e306
Stevens NJ Seiffert ER OConnor PM Roberts EM Schmitz MD Krause CGorscak E Ngasala S Hieronymus TL Temu J 2013 Palaeontological evi-dence for an Oligocene divergence between Old World monkeys and apesNature 497 611e614
Suwa G Kono RT Katoh S Asfaw B Beyene Y 2007 A new species of great apefrom the late Miocene epoch in Ethiopia Nature 448 921e924
Szalay FS Delson E 1979 Evolutionary History of the Primates Academic PressNew York
Teaford MF Beard KC Leakey RE Walker A 1988 New hominoid facial skel-eton from the Early Miocene of Rusinga Island Kenya and its bearing on therelationship between Proconsul nyanzae and Proconsul africanus J Hum Evol17 461e477
Van Couvering JAH Miller J 1969 Miocene stratigraphy and age determinationsRusinga Island Kenya Nature 221 628e632
Walker A 1997 Proconsul function and phylogeny In Begun DR Ward CVRose MD (Eds) Function Phylogeny and Fossils Miocene Hominoid Evolu-tion and Adaptation Plenum Press New York pp 209e224
Walker A Pickford M 1983 New postcranial fossils of Proconsul africanus andProconsul nyanzae In Ciochon RL Corruccini RS (Eds) New Interpretationsof Ape and Human Ancestry Plenum Press New York pp 325e351
Walker A Teaford M 1988 The Kaswanga primate site an Early Miocene hom-inoid site on Rusinga Island Kenya J Hum Evol 17 539e544
Walker A Teaford M 1989 The hunt for Proconsul Sci Am 260 76e82Walker A Teaford MF Martin L Andrews P 1993 A new species of Proconsul
from the early Miocene of RusingaMfangano Islands Kenya J Hum Evol 2543e56
Ward CV Ruff CB Walker A Rose MD Teaford MF Nengo IO 1995 Func-tional morphology of Proconsul patellas from Rusinga Island Kenya with im-plications for other Miocene-Pliocene catarrhines J Hum Evol 29 1e19
Ward S Brown B Hill A Kelley J Downs W 1999 Equatorius a new hominoidgenus from the Middle Miocene of Kenya Science 285 1382e1386
Werdelin L Sanders WJ 2010 Cenozoic Mammals of Africa University of Cali-fornia Press Berkeley

Figure 5 Box-and-whisker plots a) P3 cusp heteromorphy index (protocone heightparacone height) Results of a t-test show this difference to be significant (p frac14 00165) Theexcluded Ekembo outlier KNM-RU 1677 only appears to have extreme heteromorphy because of extensive expansion of the buccal enamel onto the anterior root (see text) b) P4
relative paracone height (paracone heightmesiodistal crown length) Results of a t-test show this difference to be significant (p frac14 00069)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 51
The occlusal profiles of Ekembo M2s are squarish in shape withrelatively straight and equal sides with a posthypocone crista thatis distobucally to buccally directed In contrast the occlusal profilesof Proconsul M2s have a more rhomboid shape with substantialbasal flare around the mesial cusps and a distally expanded hypo-cone with a distally directed posthypocone crista resulting in astrongly curved distal margin (Fig 6) Even in specimens with lesscurved distal margins the posthypocone crista is still more distallydirected compared to the condition in Ekembo These features arecharacteristic not only of P africanus P meswae and P major butare also expressed in the isolatedM2 fromMoroto Uganda (MOR IIb2098) attributed by Pickford et al (2009) to ldquoUrdquo gitongai butregarded here as belonging to P major
As noted by others (eg Harrison and Andrews 2009) the de-gree of buccolingual molar flare differs among these samplesQuantified as the distance between the tips of the mesial cuspsdivided by maximum crown breadth the degree of molar flare in
Ekembo M2s is significantly less than in Proconsul M2s (t-testdf frac14 29 t frac14 396 p frac14 00004) In a bivariate plot of these twomeasures the distributions of Ekembo and Proconsul teeth barelyoverlap with only KNM-RU 1677 encroaching into Proconsul ter-ritory (Fig 7)
These differences in M2 morphology also tend to distinguishEkembo from Proconsul M1s but not always as clearly Whereas M2smore consistently express distinct morphotypes the M1s in bothgenera show more variation in these features
The degree of upper third molar development (ie whether M3
has four distinct cusps or the distal cusps are reduced or absent) haslong been cited as taxonomically informative for these species orgroups of these species (eg Hopwood 1933 Le Gros Clark andLeakey 1951 Andrews 1978 Pickford et al 2009) However wefound this tooth to be unreliable for taxonomic purposes Withinprimates M3s are typically the most variable molars in both sizeand shape (Blankenship et al 2007 Kavanagh et al 2007) and this
Figure 6 Occlusal view of the M2s of Proconsul and Ekembo Left P africanus holotype(M 14084) Right E heseloni holotype (KNM-RU 2036) Note in Proconsul the overalldifference in crown shape as well as the more conical individuated cusps and nar-rower cristae strongly developed lingual cingulum that continues to the distal aspectof the crown broad buccal cingular shelf and more distally directed posthypoconecrista In Ekembo the cusps are more bunodont and expanded the cristae are broaderwith better developed accessory tubercles the lingual cingulum merges into thehypocone there is no buccal shelf and the posthypocone crista is more buccallydirected
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6152
is observable even in small samples of Proconsul species TheP africanus holotype has a substantially reduced M3 whereas inKNM-CA 2250dattributed by most authors to this species(Andrews 1978 Pickford et al 2009)dit is fully developed Simi-larly the hypodigm of P major includes both fully developed M3s(eg KNM-CA 389 LG 1815) and specimens with reduced ormissing hypocones (KNM-CA 397 1299) Ekembo displays compa-rable ranges of variation from highly reduced to fully developedM3s in both species Thus these species and genera cannot bedistinguished based on M3 development and we did not find anyother distinguishing characteristics between Ekembo and Proconsulin the upper third molars
Figure 7 Scatter plot depicting the de
Mandible Pickford et al (2009) enumerated several traits thatreadily distinguish Kisingiri from non-Kisingiri mandibles andthese distinctions are entirely consistent with the distinction ofEkembo from Proconsul Our analyses specifically underscoredifferences in the symphyseal region The planum alveolare inboth male and female specimens of Ekembo is short and morevertically oriented even the most elongate of these (KNM-RU47805 McNulty et al 2007) is substantially more verticalcompared to specimens of Proconsul In contrast the planumalveolare in Proconsul is conspicuously long and nearlyhorizontally oriented Well-preserved mandibles attributed toP major (eg KNM-SO 396 and 404) exhibit this morphology inextreme fashion (Fig 8) smaller specimens (KNM-SO 1112P africanus) show less extreme expression but are still readilydistinguishable from Ekembo mandibles For its size KNM-LG 452has a surprisingly short planum alveolare though not to thedegree seen in Rusinga mandibles Correlated to the shortplanum alveolare in Ekembo is a reduced superior transversetorus which while well-developed does not approach therobusticity expressed in Proconsul mandibles This difference isreflected in an index of symphyseal robusticity (thicknessperpendicular to the long axis of the symphysislength of thelong axis) which shows Ekembo and Proconsul to be significantlydifferent (t-test df frac14 14 t frac14 430 p frac14 00007)
Development of the inferior transverse torusdoften used todifferentiate among species of Proconsul (eg Pickford et al 2009Harrison 2010)dwas found to be highly variable and inconsis-tently expressed within these samples However the overallorientation of the subplanum region (as defined by a line con-necting the posterior-most midline points of the superior andinferior transverse tori or the posterior-most midline point of theinferior margin of the symphysis in the absence of the latter) doesdistinguish Ekembo from Proconsul mandibles The orientation ofthis line in relation to the alveolar margin in the vicinity of thepostcanine teeth was assessed qualitatively by placing a small rod
gree of buccolingual flare in M2
Figure 8 Mandibles of Proconsul and Ekembo Occlusal views of P major a) KNM-SO 396 b) KNM-SO 404 c) KNM-LG 452 Occlusal views of Ekembo d) KNM-RU 2087 e) KNM-RU1674 f) KNM-RU 7290 Occlusal views of P africanus g) KNM-SO 372 h) KNM-SO 1112 i) Lingual view of P africanus (KNM-SO 1112) Note the long more horizontally orientedplanum alveolare in Proconsul resulting in a pronounced superior transverse torus The planum alveolare is shorter and more vertical in Ekembo such that one can see the inferiormandibular border from occlusal view
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 53
(eg a dental pick) in themidline internally along the two referencepoints and recording the point in lingual perspective at which therod intersected the alveolar margin (Fig 9) In Ekembo the sub-planum region is more vertically oriented so that the rod intersectsthe alveolar margin most often in the premolar field Not surpris-ingly there is a difference between sexes with females having areduced superior transverse torus and therefore a more verticallyoriented subplanum region compared to males In contrast withEkembo Proconsul mandibles have much more inclined subplanumsurfaces with the rod intersecting the lingual alveolar margin inthe vicinity of the M1 (females) or M2 (males Fig 9) When sex istaken into account therefore the distributions of Ekembo andProconsul in this feature are discrete Note that male and femalemandibles attributed to both P africanus and P major are includedin this assessment and are surprisingly similar despite dramaticsize differences
We also quantified internal symphyseal morphology using threemeasures symphyseal height (infradentaleegnathion) planumalveolare length (posterior-most point on the superior transversetoruseinfradentale) and subplanum length (posterior-most pointon the superior transverse torusegnathion) a combination thatincorporates not only the lengths of these intervals but also theangular relationships among them To compare symphyseal shapeacross a range of sizes we chose a simple principal componentsapproach to partition size onto the first eigenvector (cf Jolicouer1963) While this is not the preferred method for partitioning sizein most datasets (Jungers et al 1995) for this particular sample andset of measurements Mosimann shape variables were still
significantly correlatedwith size (see discussion in Coleman 2008)Hence we assessed differences in symphyseal morphology be-tween Ekembo and Proconsul using the second and third principalcomponents of the three variables Distributions of the two generaare distinct in this shape space (Fig 10 Supplementary OnlineMaterial [SOM] Table 1)
Robusticity of the mandibular corpus also differs between thesesamples though it is more difficult to test due to differentialpreservation among specimens Corpus thickness and depth underP4 can be measured in the greatest number of specimens andcomparison of these measures suggests a difference betweengroups in the scaling of corpus robusticity (Fig 11) Among largepresumably male specimens Ekembo mandibles are substantiallyless robust than those of Proconsul However among smallermandibles only a single Proconsul specimen (KNM-SO 372) could beincluded and it is similar in robusticity to Ekembomandibles of thissize Considered together the impression is that corpus robusticitymay scale differently in Ekembo than it does in Proconsul and a testfor differences in slopes using an ANCOVA design of the loggedvariables was significant (df frac14 2 F frac14 2481 p lt 00001) Never-theless additional specimens are needed to properly evaluate theinfluence of taxonomy sex and size on corpus robusticity areasonable alternative interpretation of Figure 11 is that it dem-onstrates a significant difference between P major and all otherspecies of both Proconsul and Ekembo
Lower incisors Ekembo specimens are uniformly narrower inmesiodistal length than incisors attributed to P africanus P major
Figure 10 Scatter plot of principal component scores based on an eigenanalysis of threeposterior point on the superior transverse toruseinfradentale) and subplanum length (mprimarily captures size differences among specimens (Jolicouer 1963 see text and SOM Trespectively) in the symphyseal variables
Figure 9 Line drawing of a hemimandible depicting different orientations of the sub-planum area in Ekembo and Proconsul Females are denoted using shorter vectors maleswith longer vectors Vector is estimated in M 14086 which is broken at the midlineEkembo specimens have a more vertical subplanum compared to those from Songhorand Legetet but within both groups females have more vertically oriented regions thanmales Hence even the most ventrally rotated subplanum in the largest Ekembo malespecimen (KNM-RU 47805) groups with the Tinderet females in this feature
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6154
and P meswae both absolutely and in relation to crown height(Fig 12) This is true for both I1 and I2 A t-test adjusted for non-independent observations (eg I1 and I2 from the same mandible)confirmed the difference as significant (t-test df frac14 12 t frac14 258p frac14 00242) Moreover while the measurable Proconsul samplewas small this same pattern is observable in other specimenswhose tips were slightly to moderately worn and therefore notmeasured
Lower canines The only lower canine trait that discretely sepa-rates these genera is the pointed crown tip in Ekembo (Fig 3d)contrasted with a blade-like (or ldquoburinrdquo) tip in Proconsul(Fig 3c) This feature is well established for P major but is alsoexpressed in KNM-CA 1772 which was referred to P africanus byKelley (1986) and is too small to be either P major or P meswaeTwo other specimens commonly attributed to P africanus KNM-CA 2149 and KNM-SO 1112 both have broken canine tipsLikewise a new specimen (Kor 65004) attributed to P africanusby Pickford et al (2009) also has a broken tip and cannot beevaluated (see Pickford et al 2009 Fig 23) As with the uppercanines the blade-like tip is also present in the deciduous lowercanines of P major (eg KNM-CA 361 S0 5352) and P meswae(KNM-ME 1)
Lower premolars Like most previous authors (eg Andrews 1978Harrison 2002 2010 Pickford et al 2009) we found neitherqualitative nor quantitative characteristics of the lower premolarswith any taxonomic value
Lower molars Differences in lower molar morphology are few andrelatively subtle but in some cases mirror those seen in the uppermolars Cusps on the lower molars of Ekembo are more bunodontthan those in Proconsul and occupy more of the occlusal surfaceBroad inter-cuspal crests are incorporated into the Ekembo cuspsmaking them appear more inflated Molar cusps of Proconsul are
variables symphyseal height (infradentaleegnathion) planum alveolare length (mostost posterior point on the superior transverse torusegnathion) The first componentable 1) and thus PC 2 and PC 3 illustrate the residual shape variance (72 and 28
Figure 11 Scatter plot showing mandibular corpus robusticity under P4 Trend lines were computed using ordinary least squares (OLS) regression Similar relationships areobservable at other parts of the corpus but differential preservation results in smaller samples at those positions Slopes are significantly different (p lt 00001 tested on loggedvariables)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 55
more distinct from one another and are linked by narrower sharperinter-cuspal crests The lower molars of Ekembo also have asubstantially less developed buccal cingulum than those ofProconsul (Harrison 2002 2010 Harrison and Andrews 2009)
Figure 12 Scatter plot of lower incisor crown proportions Differenc
Lastly and again as seen in the upper molars the two genera canbe distinguished by the significantly greater amount of lowermolar flare in Proconsul (Fig 13 t-test df frac14 59 t frac14 713p lt 00001 see also Harrison and Andrews 2009)
es between Ekembo and Proconsul are significant (p frac14 00242)
Figure 13 Box-and-whisker plot of buccolingual flare in lower molars measured asthe average distance between the mesial buccal and lingual cusps divided by overallbuccolingual crown breadth Differences between Ekembo and Proconsul are highlysignificant (p lt 00001)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6156
Relative molar size has been used to distinguish species ofProconsul but we observed the primary difference to be in relativebuccolingual breadths of M1 and M2 In Ekembo M1 is substantiallynarrower than M2 than is the case in Proconsul (Fig 14 Harrison2002 2010) Although none of the smaller Proconsul mandiblespreserve complete crowns for both M1 and M2 KNM-SO 1112 hasan intact M2 and at least preserves the cervix for M1 However evenif the cervical measurement is used for its M1 breadthdthusunderestimating the true crown breadthdKNM-SO 1112 still has abroader first molar in relation to M2 than would be expected inEkembo If crown breadth is estimated from the ratio of cervixbreadth to maximum crown breadth in other Proconsul fossils therange of likely breadths for the SO 1112 M1 clearly distinguishes it
from the Ekembo sample (Fig 14) Testing this relationship onlogged variables using the mean estimate for the SO 1112 M1(Fig 14) reveals a significant difference between the two groups(ANCOVA df frac14 1 F frac14 2424 p frac14 00002)
As with upper third molars we find a great deal of variation inM3 morphology and mistrust the taxonomic significance some-times placed on characteristics of this toothdat least with regard tothese taxa Harrison (2002 2010) cited the mesiodistal position ofthe M3 entoconid as differentiating Kisingiri third molars fromthose attributed to P africanus We concur with his assessment butfound the entoconid position to vary in specimens attributed toP major Thus this character does not differentiate Ekembo from allProconsul species but may distinguish it from the type speciesP africanus
Discussion
The distinctiveness of the Kisingiri large catarrhine materialfrom that at Tinderet and Ugandan localities has been noted pre-viously by several authors (Pickford 1986 Walker et al 1993Begun 2001 2007 MacLatchy and Rossie 2005 Harrison andAndrews 2009 Harrison 2010) What had not been fully docu-mented was the nearly complete lack of overlap between thesesamples in a number of features despite very large assemblagesfrom Rusinga and Mfangano On descriptive criteria alone (Table 2)many isolated teeth (I1 C1 P3 M2 C1) and all of the mandibles canbe assigned to either Proconsul or Ekembowith very little chance ofmisattribution That some of the most diagnostic features occur inthe upper dentitiondthe only anatomy preserved in theP africanus type specimendadds weight to the taxonomicarrangement proposed here The morphology of the upper teeth inall Proconsul species compellingly unites them into a single cladedistinct from E heseloni and E nyanzae This is reinforced bynumerous features of the mandible and mandibular dentition thatfurther distinguish these samples The number and variety of thesefeatures are such that Proconsul and Ekembo are more differentfrom one another than are many closely-related modern catarrhinegenera
Derived morphology of Ekembo and Proconsul
It is premature to formally assign primitive and derived status tothe individual features of Ekembo and Proconsul before completinga formal phylogenetic analysis This is especially true given theabundance of evidence available for Ekembo and hence the poten-tial impact that our generic revision could have on phylogeneticinterpretations Nevertheless in comparison with both extanthominoids and the small-bodied catarrhines from the earlyMiocene it is informative to differentiate the evolutionary grades ofthese genera Many characteristics of Ekembo are more advanced inthe direction of modern apes compared to those of Proconsul(Table 2) General trends such as expanded and more bunodontcusps broader molar crests reduced cingulum and morphologicalsimilarity between P3 and P4 all represent morphologies thatpresage those of later fossil and extant hominoids The moreadvanced morphologies of P3 and M2 are particularly conspicuouscompared not only with those of Proconsul but to most earlyMiocene catarrhines In contrast the mesially positioned stronglyheteromorphic cusps on the P3 of Proconsul plus the correspondingasymmetric buccal profile notched mesial margin and missingmesial fovea are characteristic of other early Miocene catarrhinessuch as Dendropithecus Rangwapithecus and Limnopithecus andare likely primitive among East African catarrhines The moresymmetric P3 of Ekembo with midline positioned cusps andreduced cusp heteromorphy is reminiscent of later hominoids and
Figure 14 Relative buccolingual dimensions of M1 and M2 KNM-SO 1112 has a preserved M2 but only the cervix of M1 Its M1 breadth was estimated based on the ratio to cervixbreadth to maximum crown breadth in other Tinderet fossils Even the cervical breadth of SO 1112 is broader than would be predicted for the crown breadth of a Kisingiri M1 Trendlines computed using OLS regression and using the average estimate of SO 1112 M1 breadth Group differences were significant (p frac14 00002)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 57
even hominid taxa such as Rudapithecus (eg Kordos and Begun2001)
Proconsul is likewise united by a suite of features that are for themost part characteristic only of this lineage (Table 2) Themorphology of both upper and lower canines is unique to thisgenus and shared by all three well-known species plus the poorlyrepresented middle Miocene P gitongai (Pickford and Kunimatsu2005) The mandibular symphysis is also distinctive in Proconsulcompared with other catarrhines with its very long and shallowlysloping planum alveolare and steeply inclined subplanum surfaceIndeed even the small edentulous anterior mandibular fragmentKNM-SO 372 is readily placed within Proconsul and excluded fromother similarly sized catarrhine taxa such as Rangwapithecus andEkembo based on this unique symphyseal morphology (Figs 8 and9)
Other differences between Ekembo and Proconsul are of uncer-tain evolutionary polarity (eg orientation of the posthypoconecrista) or may be of taxonomic relevance without broader phylo-genetic significance (eg degree of molar flare Table 2) Werecognize that there are morphological differences within Procon-sul particularly between the better known species P africanus andP major However many of these differences are likely related tosize and represent variations on a single generic theme that aremarkedly different from the morphology of Ekembo The recentdescription (Harrison and Andrews 2009) of the intermediatelysized P meswae underscores the broad morphological unity withinProconsul and further substantiates a well-defined Proconsul cladecomprising Tinderet and Ugandan species
Based on the abundant morphological and morphometric evi-dence supporting genus-level distinction between Proconsul andEkembo we are currently conducting a more detailed phylogeneticanalysis of all early Miocene catarrhines in order to better under-stand the distribution and evolutionary significance of these andother morphological features We anticipate that the debates sur-rounding the phylogenetic relationships of an historically consti-tuted sample of Proconsul can be more satisfactorily resolved nowthat the Kisingiri material is appropriately recognized as a distinctgenus
The generic disposition of ldquoUgandapithecusrdquo species
A great many features said to characterize ldquoUgandapithecusrdquowere added to the original diagnosis of the genus by Pickford et al(2009) in their systematic revision of Proconsul (sensu lato) As partof that study they also erected a new species ldquoUrdquo legetetensis andtransferred P meswae to ldquoUgandapithecusrdquo Evaluating the same setof features across the entire available sample however we wereunable to find support for this more expansive conception ofldquoUgandapithecusrdquo for several reasons First some diagnostic fea-tures cited in Pickford et al (2009) were quite variable within theldquoUgandapithecusrdquo sample As an example marked reduction of theM3 entoconid to a bead on the lingual cingulum is not shared byseveral ldquoUrdquo major specimens such as KNM-CA 393 NAP I 49000and Kor 253004 (see Pickford et al 2009 Fig 33e) and was likelynot a feature of a few of the more worn specimens eg BUMP 600and BUMP 601 or even the holotype of P major M 16648 Second
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6158
some diagnostic features of ldquoUgandapithecusrdquo were found to bevariably present in Ekembo arguing against their utility for dis-tinguishing ldquoUgandapithecusrdquo from Pickford et als (2009) com-bined P africanus thorn Kisingiri conception of Proconsul For examplesignificantly reduced curvature of the I1 distal margin in ldquoUganda-pithecusrdquo is also found in some Kisingiri specimens including KNM-RU 1685 1712 1714 1831 1846 and 1951 several more Rusingaspecimens have moderately reduced curvature Third some fea-tures that purportedly distinguish ldquoUgandapithecusrdquo fromP africanus are likely related to size For example the somewhatmore central position of the P3 paracone in ldquoUrdquomajor is not seen inthe smaller ldquoUrdquo meswae whose paracone is positioned similarly tothat of the holotype of P africanus (see Harrison and Andrews2009 Fig 6d)
However the major part of the evidence for recognizingldquoUgandapithecusrdquo (sensu Pickford et al 2009) was assembled by re-assigning specimens traditionally attributed to P africanus into newspecies of ldquoUgandapithecusrdquo The P africanus holotype was left inassociation with the Rusinga sample despite having morphologyoverwhelmingly characteristic of other Tinderet specimens Theresulting specimen allocations left P africanus morphologicallyimpoverisheddstripped of much of its hypodigmdso that Pro-consul for Pickford et al (2009) came to be characterized primarilyby the Rusinga sample (see similar comments in Harrison andAndrews 2009) Within that conceptual framework any spec-imen that differs from those at Rusinga could not be Proconsul butmust instead belong to ldquoUgandapithecusrdquo Thus specimens such asthe KNM-SO 1112 mandible were erroneously assigned to ldquoUgan-dapithecusrdquo because they were (correctly) perceived as differingfrom their Rusinga counterparts (Pickford et al 2009)
Here we restore the P africanus hypodigm and associate it withthe other Tinderet fossils according to their shared morphologySpecimens assigned to ldquoUgandapithecusrdquo revert to Proconsul andmany of the features cited to support a speciose ldquoUgandapithecusrdquo(Pickford et al 2009) instead become diagnostic features of Pro-consul This correction serves to highlight the distinctiveness of theKisingiri remains and the need to recognize this distinctiveness atthe generic level
As demonstrated above specimens from Kisingiri do not displaythe morphology of Proconsul as represented by the holotype ofP africanus M 14084 Given that this specimen preserves only theupper canine and cheek teeth similarities with or departures fromthis morphology represent the most compelling arguments forassessing inclusivity in the genus However for the mandible andlower dentition if one simply sorts the specimens by elementwithout relying on taxonomic preconceptions the overwhelmingdistinction within the traditionally constituted Proconsul sample isbetween Kisingiri material on the one hand and that from Tinderetand Uganda on the other (cf Walker et al 1993 Begun 2001 2007MacLatchy and Rossie 2005 Harrison and Andrews 2009Harrison 2010) Additionally with the Kisingiri specimensremoved to Ekembo the postcranial differences used to partlyjustify a distinct ldquoUgandapithecusrdquo lose their relevance (seeHarrison and Andrews 2009)
We have differed among ourselves in our past treatment ofP major as generically distinct from the smaller species of Pro-consul (Begun 2007 McNulty 2010) Morphologically many ofthe unique features of P major can be interpreted as size-relatedvariants on the smaller P africanus morphology Even if so thequestion is whether these are sufficiently different and numerousto warrant generic distinction that is do they suggest thatP major belongs in a separate clade from P africanus andP meswae As we have demonstrated differences among thesethree species are certainly fewer and show less morphologicaldistinction than those between Ekembo and any of the three
Nevertheless while we ultimately agree with Harrison andAndrews (2009 see also Begun 2007) that the features dis-tinguishing ldquoUrdquo major from P africanus are consistent with thosethat distinguish congeneric species and are at least partly due todifferences in size we concede the possibility that the largestProconsul species could belong in a separate genus (Senut et al2000 Pickford and Kunimatsu 2005) However whether or notone recognizes a separate genus for P major the P africanus ma-terial does not sort morphologically with the Kisingiri materialTherefore the taxonomic alternatives are either two generaProconsul and Ekembo or three Proconsul Ugandapithecus andEkembo with Proconsul and Ugandapithecus united in a cladeexclusive of Ekembo Thus while we strongly favor the firstalternative recognizing Ugandapithecus does not change thefundamental argument concerning the need to refer the Kisingirispecies to a generically and cladistically distinct taxon from thespecies represented at Tinderet and the Ugandan sites
Specimens of interest
Despite the substantial dental and gnathic differences betweenProconsul and Ekembo there are a few specimens whose taxonomicaffinities require additional scrutiny
KNM-RU 1676-77 This set of associated upper and lower teeth is aconsistent outlier among E heseloni and E nyanzae specimens Insome ways this specimen more closely resembles Tinderet Pro-consul in the features that distinguish it from Ekembo Molar cuspmorphology and cingulum development I1 lingual tubercledevelopment and M2 shape and posthypocone crista orientationare more consistent with specimens from the Tinderet sites On theother hand it otherwise resembles Ekembo and lacks the strongestdiagnostic features that unite species of Proconsul such as thedistinctive canine and P3 morphology Overall this specimen doesnot fit unambiguously into either genus in fact the degree ofbuccolingual flare cusp morphology enamel wrinkling and espe-cially the long extension of enamel onto the anterior root in P3 arein some ways reminiscent of Afropithecus although it clearly differsfrom that genus as well (cf Rossie and MacLatchy 2013)
Were these characteristics individually distributed among thelarge Kisingiri sample one might attribute this to intra-genericvariation but when found together in a single specimen from alarge sample it suggests a unique individual One of us (KPM)thinks it likely that the specimen was not actually found at aKisingiri locality based on its unique preservation compared toother Rusinga fossils and on other examples of mis-proveniencedspecimens (Leakey 1967 Andrews and Molleson 1979 Pickford1986 Kelley 1986) However we recognize that relying toofrequently on this explanation allows almost any scenario to bejustified and suggest some alternative possibilities One is that thespecimen was found on Rusinga but in the much older WayandoFormation strata instead of the Hiwegi Formation The differencesin morphology in that case might reflect more than a million yearsof temporal change (McCollum et al 2013) with KNM-RU 1676-77preserving certain more primitive Proconsul-like features As analternative it is reasonable to assume that fossil assemblages asvast as those from Rusingawill include not only abundant taxa butalso rare or infrequent inhabitants of the local biome There aremany examples of this from Rusingas mammalian communityeg ldquoHemicyonrdquo Kulutherium Tadarida cf Hyainailouros cfArchaeobeledon Kelba (see relevant chapters in Werdelin andSanders 2010) As such KNM-RU 1676-77 may simply representa rare and well-preserved specimen of some other catarrhinetaxon that lived among or visited a primate community dominatedby Ekembo
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 59
KNM-SO 1112 This specimen was initially referred to Rangwapi-thecus but soon after was transferred to Proconsul afrianus (Bosler1981 Martin 1981)da designation that most subsequent authorshave followed (Pickford 1986 Nengo and Rae 1992 Kelley 19861995 Walker et al 1993 MacLatchy and Rossie 2005 Hill et al2013) Pickford et al (2009) more recently referred this specimento ldquoUrdquo meswae citing two primary reasons First they reiteratedBoslers (1981) concern that its P3 differs from others attributedto P africanus However at that time the majority of what wasconsidered to be P africanus would have been from RusingaIsland In fact while noting its contrasting morphology Boslersuggested that differences between KNM-SO 1112 and other P3smight reasonably be attributed to sexual dimorphism notingthat this specimen provides ldquoa not implausible match for the Pafricanus holotype maxilla M 14084rdquo (1981152) Thatassessment is borne out in this study Using the much largersample of Ekembo P3s as a model variation can be seen toencompass specimens with well-developed mesial beaks andthose with more bilaterally compressed crowns and parallelsides The latter morphology is predominantly found amongmale specimens (KNM-RU 1674 1924 and KPS I and III) thussupporting Boslers (1981) suggestion that this variation isrelated to sexual dimorphism and hence that KNM-SO 1112 is amale P africanus
The second feature cited by Pickford et al (2009) in referringthis specimen to ldquoUrdquo meswae was posterior shallowing of themandibular corpus However because they redefined ldquoUgandapi-thecusrdquo to include all of the well-preserved Tinderet mandibles(Pickford et al 2009)dleaving Proconsul to be represented only byRusinga specimensdtheir only alternative within this scheme for aspecimen the size of SO 1112 was to refer it to ldquoUrdquomeswae It seemsodd though to assign this specimen to a species from a (perhapsmuch) older site when the Proconsul africanus holotype is ofappropriate size and similar age and as there are no other adultmandibles from either species with which to compare it In anycase mandibular shallowing is variable in both Ekembo and Pro-consul with some Kisingiri specimens exhibiting considerableshallowing (eg KNM-RU 2087) and some ldquoUgandapithecusrdquo spec-imens having very little (eg M 14086)
We therefore see no morphological grounds to assign KNM-SO1112 to P meswae but do find some morphological evidence toassign it as a male P africanus which occurs at Songhor
KNM-LG 452 This very complete mandible was originally recog-nized as a female of P major based on the size of the dentition androbusticity of the mandible (Martin 1981) Bosler (1981) howeverleft this specimen unassigned noting similarities to P major in P3-M1 size but having distinctly smaller M2e3 In this she likenedKNM-LG 452 to a few other specimens (notably M 14086) thatare smaller in their distal molars in a manner more consistentwith taxonomic rather than sex differences (Bosler 1981 see alsoPilbeam 1969) Kelley (1986 1995) also assigned this specimen asa female of P major based on canine size and morphology butnoted that the canine lacked the shallowly-sloping mesial ridgetypical of most female anthropoid canines This is also truehowever of two other canines assigned as female P major KO 9and SO 373 (Kelley 1986) and so might simply be an unusualcharacteristic of P major female lower canines In the end Kelley(1995) concluded that considering all of the criteria fordistinguishing male and female canines the evidence that KNM-LG 452 is female remains questionable
Pickford et al (2009) further developed the idea that two spe-cies are represented within the P major sample and made KNM-LG452 the holotype of a new species ldquoUrdquo legetetensis We concur thatthis specimen is unique among the well-preserved Tinderet
mandibles in some important features Whereas male and femalemandibles of P major have long internal symphyseal planesexpansive extra- and retromolar sulci and rami that cross theocclusal plane at distal M3 KNM-LG 452 has a shorter symphysealplane (though not to the extent seen in Ekembo) a moderateextramolar sulcus but no retromolar sulcus and a ramus thatcrosses the occlusal plane more mesially If KNM-SO 396 and SO404 represent males and females respectively of P major then themorphology of KNM-LG 452 appears distinct
At present however we do not think there is sufficient evidenceto differentiate this specimen from either P major or P meswaePickford et al (2009) argue that KNM-LG 452 cannot belong to thelatter species because of its larger size The size differences how-ever are well within the range of variation seen in P majorE heseloni and E nyanzae Moreover whereas the Legetet mandibleis larger than most of the P meswae hypodigm an adult P4 referredto this species (KNM-ME 25) substantially exceeds its counterpartin KNM-LG 452 Should the Legetetmandible ultimately prove to bea male specimen size alone would not rule out its attribution toP meswae On the other hand if it is female then it becomes harderto justify its inclusion in that sample of mostly smaller specimens Iffemale therefore it could represent a separate species for whichthe binomial Proconsul legetetensis comb nov is available Recentlycollected fossils from the SonghorKapurtay area provide someadditional support for the possibility that three species of Proconsulexisted at these localities (McNulty 2014ab) However toconvincingly demonstrate this and determine whether a thirdspecies can be differentiated from P meswae will require a moredetailed analysis of variation among Proconsul species a corre-sponding study of variation in modern taxa anddalmost certain-lydadditional fossil specimens For now we regard ldquoUrdquo legetetensisas a subjective junior synonym of P major
Conclusions
The genus Proconsul has been central to studies of ape and hu-man evolution since its first description by Hopwood (1933) As oneof the best known fossil catarrhines represented by nearly everyskeletal element our knowledge of Proconsuls functional anatomypositional behavior and overall morphology has made it thefoundation for paleoanthropological research in the early Mioceneas well as studies on the origin and early diversification of homi-noids With respect to systematics however Proconsul is less wellunderstood the type species P africanus is known from a rela-tively small number of specimens from sites in the Tinderet areaand no postcranial bones can be unambiguously assigned to itRather it is the copious and well-preserved specimens from theKisingiri sites of Rusinga and Mfangano Islands that have formedthe basis for the common image of Proconsul
We argue here that the Kisingiri species traditionally referredto ProconsuldP heseloni and P nyanzaedare generically distinctfrom P africanus and hereby transfer them into a new genusEkembo based on a substantial number of morphological differ-ences (Table 2) Considering only those traits preserved in theP africanus holotype the entire sample from Rusinga and Mfan-gano differs markedly in the morphology of the upper canine P3P4 and M2 Additional differences in the I1 lower incisors andcanines molar cusp and occlusal crest morphology degree of flarein all molars and especially mandibular features clearly distin-guish the Kisingiri from non-Kisingiri samples in a way that isincompatible with congeneric species With the transfer ofE heseloni and E nyanzae Proconsul becomes a well-defined cladecomprising at least three speciesdP africanus P major andP meswaedunited by several features that are unique for thegenus (Table 2)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6160
We found no support in this study for the genus ldquoUgandapi-thecusrdquo as constructed by Pickford et al (2009) In that paper theytransferred the vast majority of Proconsul fossils from the Tinderetand Napak localities into ldquoUgandapithecusrdquo leaving P africanusrepresented only by its holotype and a small number of isolatedteeth and aligning it with the material from Rusinga and Mfan-ganoWhen the P africanus hypodigm is reconstituted and properlyaffiliated with the other Tinderet species according to itsmorphology much of the evidence marshaled by Pickford et al(2009) for ldquoUgandapithecusrdquo actually supports the distinction be-tween the Tinderet and Kisingiri samples ie the generic distinc-tion of Ekembo In addition some of the traits enumerated byPickford and colleagues in support of ldquoUgandapithecusrdquo wererevealed in our analysis to be variable within one or more speciesand therefore of questionable taxonomic utility We do not rejectout of hand the original conception of ldquoUgandapithecusrdquo (Senutet al 2000) as either a monotypic genus distinguishing ldquoUrdquo ma-jor or perhaps including a closely allied middle Miocene ldquoUrdquogitongai as well (Pickford and Kunimatsu 2005) but we have fornow retained the early Miocene specimens of both in Proconsulpending further study However even if Ugandapithecus is a taxo-nomically valid genus comprising just the latter two species theirmorphology indicates that theywould still be united in a cladewithProconsul and they lack several more advanced features that partlydefine Ekembo Better understanding of the role that body size playsinmediating differences within Proconsulwill help to further clarifythe relationships among these species
Despite the existence of one well-represented and several lesscomplete skeletons of what was long assumed to be Proconsul thephylogenetic status of this genus has defied consensus This islargely due in our view to Proconsul having been an admixture oftwo distinct genera one more advanced in the direction of extantapes than the other As we demonstrate here the material fromRusinga and Mfangano traditionally referred to Proconsul andwhich includes all of the more complete skeletal material isdistinct from the Tinderet and Napak material and belongs in aseparate genus Ekembo By disentangling these distinct clades weanticipate being able to achieve better phylogenetic resolution forboth genera thereby improving our knowledge of early hominoidevolution and diversification
Acknowledgments
We thank many people and agencies that assisted with thedevelopment and completion of this project This research wasinitiated as a result of funding from the Wenner-Gren Foundationand the Johns Hopkins School of Medicine to DR Begun in1987e1989 and was renewed following research and travelgenerously funded by multiple Leakey Foundation grants to KMcNulty National Science Foundation grants to K McNulty(0852609 1241807) the McKnight Foundation the University ofMinnesota the Lloyd A Wilford Endowment the LeverhulmeFoundation and the Turkana Basin Institute J Kelley gratefullyacknowledges support from the Institute of Human Origins Ari-zona State University DR Begun also wishes to acknowledge theAlexander Humboldt Stiftung and NSERC for funding support Weare grateful to the National Museums of Kenya the UgandaMuseum the Natural History Museum and to Rose Mwanja EzraMusiime Tom Mukhuyu Samuel Muteti James Yatich and MaryMuungu for access to and assistance with specimens in their careMany people provided assistance comments criticisms insightsand suggestions on various aspects of this work and we especiallythank Holly Dunsworth Will Harcourt-Smith Tom Lehmann DanPeppe Nicole Garrett Kirsten Jenkins Rutger Jansma Susy CoteIsaiah Nengo Andrew Hill and Brenda Benefit K McNulty
acknowledges the evolutionary anthropology faculty and studentsat Durham University for their helpful comments on an earlyversion of this manuscript and for their gracious hospitality Weare likewise grateful to the anonymous reviewers associate editorand editor whose comments strengthened the paper This manu-script is publication 1 supporting Research on East AfricanCatarrhine and Hominoid Evolution (REACHE)
Appendix A Supplementary data
Supplementary data related to this article can be found at httpdxdoiorg101016jjhevol201503009
References
Andrews P 1974 New species of Dryopithecus from Kenya Nature 249 188e190Andrews P 1978 A revision of the Miocene Hominoidea of East Africa Bull Brit
Mus (Nat Hist) Geol 30 85e225Andrews P 1985 Family group systematics and evolution among catarrhine pri-
mates In Delson E (Ed) Ancestors The Hard Evidence Liss New Yorkpp 14e22
Andrews P Harrison T Martin L Pickford M 1981 Hominoid primates from anew Miocene locality named Meswa Bridge in Kenya J Hum Evol 10123e128
Andrews P Begun D Zylstra M 1997 Interrelationships between functionalmorphology and paleoenvironments in Miocene hominoids In Begun DWard C Rose M (Eds) Function Phylogeny and Fossils Miocene HominoidEvolution and Adaptations Plenum Press New York pp 29e58
Andrews P Molleson T 1979 The provenance of Sivapithecus africanus Bull BritMus (Nat Hist) Geol 32 19e23
Begun DR 2001 African and Eurasian Miocene hominoids and the origins of theHominidae In de Bonis L Koufos GD Andrews P (Eds) Hominoid Evolutionand Climatic Change in Europe Phylogeny of the Neogene Hominoid Primatesof Eurasia Cambridge University Press London pp 231e253
Begun DR 2007 Fossil record of Miocene hominoids In Henke W Tattersall I(Eds) Handbook of Paleoanthropology Springer Berlin pp 921e977
Begun DR Teaford MF Walker AC 1994 Comparative and functional anatomyof Proconsul phalanges from the Kaswanga primate site Rusinga Island KenyaJ Hum Evol 26 89e165
Bishop WW 1964 More fossil primates and other Miocene mammals from North-East Uganda Nature 203 1327e1331
Bishop WW Miller JA Fitch FJ 1969 New potassium-argon age determinationsrelevant to the Miocene fossil mammal sequence in East Africa Am J Sci 267669e699
Blankenship JA Mincer HH Anderson KM Woods MA Burton EL 2007Third molar development in the estimation of chronologic age in Americanblacks as compared with whites J Forensic Sci 52 428e433
Bosler W 1981 Species groupings of early Miocene dryopithecine teeth from EastAfrica J Hum Evol 10 151e158
Coleman MN 2008 What does geometric mean mean geometrically Assessingthe utility of geometric mean and other size variables in studies of skullallometry Am J Phys Anthropol 135 404e415
Cote S Malit N Nengo I 2013 Additional mandibles of Rangwapithecus gordonian early Miocene catarrhine from the Tinderet localities of Western KenyaJ Hum Evol 153 341e352
Deane AS 2009 Early Miocene catarrhine dietary behavior the influence of theRed Queen Effect on incisor shape and curvature J Hum Evol 56 275e285
Drake R Van Couvering JA Pickford M Curtis G Harris JA 1988 New chro-nology for the Early Miocene mammalian faunas of Kisingiri Western KenyaJ Geol Soc Lond 145 479e491
Evernden JF Savage DE Curtis GH James GT 1964 Potassium-argon datesand the Cenozoic mammalian chronology of North America Am J Sci 262145e198
Gebo DL MacLatchy L Kityo R Deino A Kingston J Pilbeam D 1997A hominoid genus from the Early Miocene of Uganda Science 276 401e404
Geoffroy Saint-Hilaire E 1812 Tableau des quadrumanes 1 Ord QuadrumanesAnnls Mus Hist Nat Paris 19 85e122
Gray JE 1825 Outline of an attempt at the disposition of the Mammalia into tribesand families with a list of the genera apparently appertaining to each tribeAnnls Philos 10 337e344
Harrison T 1982 Small-bodied apes from the Miocene of East Africa PhDDissertation University of London
Harrison T 1986 New fossil anthropoids from the middle Miocene of East Africaand their bearing on the origin of the Oreopithecidae Am J Phys Anthropol 71265e284
Harrison T 1987 The phylogenetic relationships of the early catarrhine primates areview of the current evidence J Hum Evol 16 41e80
Harrison T 2002 Late Oligocene to middle Miocene catarrhines from Afro-ArabiaIn Hartwig WC (Ed) The Primate Fossil Record Cambridge University PressCambridge pp 311e338
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 61
Harrison T 2010 Dendropithecoidea Proconsuloidea and Hominoidea (CatarrhiniPrimates) In Werdelin L Sanders WJ (Eds) Cenozoic Mammals of AfricaUniversity of California Press Berkeley pp 429e469
Harrison T Andrews P 2009 The anatomy and systematic position of a newspecies of early Miocene proconsulid from Meswa Bridge Kenya J Hum Evol56 479e496
Hill A Nengo IO Rossie JB 2013 A Rangwapithecus gordoni mandible from theearly Miocene site of Songhor Kenya J Hum Evol 65 490e500
Hopwood AT 1933 Miocene primates from Kenya J Linn Soc Zool 38 437e464Ishida H Kunimatsu Y Takano T Nakano Y Nakatsukasa M 2004 Nacholapi-
thecus skeleton from the Middle Miocene of Kenya J Hum Evol 46 69e103Jolicouer P 1963 The multivariate generalization of the allometry equation Bio-
metrics 19 497e499Jungers WL Falsetti AB Wall CE 1995 Shape relative size and size-
adjustments in morphometrics Am J Phys Anthropol 38 (S2) 137e161Kavanagh KD Evans AR Jernvall J 2007 Predicting evolutionary patterns of
mammalian teeth from development Nature 449 427e433Kelley J 1986 Species recognition and sexual dimorphism in Proconsul and
Rangwapithecus J Hum Evol 15 461e495Kelley J 1995 Sex determination in Miocene catarrhine primates Am J Phys
Anthropol 96 390e417Kelley J 1993 Taxonomic implications of sexual dimorphism in Lufengpithecus In
Kimbel WH Martin LB (Eds) Species Species Concepts and Primate Evo-lution Springer New York pp 429e458
Kent PE 1942 The country round the Kavirondo Gulf of Victoria Nyanza Geogr J100 22e31
Kordos L Begun DR 2001 A new cranium of Dryopithecus from RudabanyaHungary J Hum Evol 15 689e700
Le Gros Clark WE Leakey LSB 1950 Diagnoses of East African Miocene Homi-noidea Q J Geol Soc Lond 105 260e263
Le Gros Clark WE Leakey LSB 1951 The Miocene Hominoidea of East Africa InFossil Mammals of Africa No 1 British Museum (Natural History) London
Leakey LSB 1943 A Miocene anthropoid mandible from Rusinga Kenya Nature152 319e320
Leakey LSB 1963 East African fossil Hominoidea and the classification within thissuper-family In Washburn SL (Ed) Classification and Human EvolutionAldine Chicago pp 32e49
Leakey LSB 1967 Notes on the mammalian faunas from the Miocene and Pleis-tocene of East Africa In Bishop WW Clark JD (Eds) Background to Evolu-tion in Africa University of Chicago Press Chicago pp 7e29
Leakey RE Leakey MG 1986 A new Miocene hominoid from Kenya Nature 324143e146
Leakey RE Leakey MG Walker AC 1988 Morphology of Afropithecus turka-nensis from Kenya Am J Phys Anthropol 76 289e307
Leakey MG Ungar PS Walker A 1995 A new genus of large primate from theLate Oligocene of Lothidok Turkana District Kenya J Hum Evol 28519e531
Linnaeus C 1758 Systema naturae per regna tria naturae secundum classesordines genera species cum characteribus differentris synonymis locis tenthed Laurentii Salvii Stockholm
MacInnes DG 1943 Notes on the East African Miocene primates J E Afr UgandaNat Hist Soc 17 141e181
MacLatchy L 2004 The oldest ape Evol Anthropol 13 90e103MacLatchy L Bossert WH 1996 An analysis of the articular surface distribution of
the femoral head and acetabulum in anthropoids with implications for hipfunction in Miocene hominoids J Hum Evol 31 425e453
MacLatchy L Rossie JB 2005 The Napak hominoid still Proconsul major InLieberman DE Smith RJ Kelley J (Eds) Interpreting the Past Essays onHuman Primate and Mammal Evolution in Honor of David Pilbeam Brill Ac-ademic Publishers Boston pp 15e28
Martin L 1981 New specimens of Proconsul from Koru Kenya J Hum Evol 10139e150
McCollum MS Peppe DJ McNulty KP Dunsworth HM Harcourt-Smith WEH Andrews AL 2013 Magnetostratigraphy of the early MioceneHiwegi Formation (Rusinga Island Lake Victoria Kenya) Geological Society ofAmerica Abstracts with Programs 45 p 12
McNulty KP 2010 Apes and tricksters the evolution and diversification ofhumans closest relatives Evol Edu Outreach 3 322e332
McNulty KP Harcourt-Smith WEH Dunsworth HM 2007 New primate fossilsfrom Rusinga Island Kenya Am J Phys Anthropol 132 (S44) 170
McNulty KP MacLatchy L Peppe DJ Nengo I Manthi FK Miller ERStevens NJ Cote S Lehmann T Kingston J 2014a A regional approach toEast African early Miocene paleobiology J Vert Paleontol 185 SVP Programand Abstracts Book 2014
McNulty KP MacLatchy L Rossie JB Peppe DJ Deino AL Mbua ENManthi FK Nengo IO Stevens NJ Cote S Lehmann T Gutierrez M 2014b
Research on East African catarrhine and hominoid evolution results from thefirst year Am J Phys Anthropol 153 (S58) 182
Michel LA Peppe DJ Lutz JA Driese SG Dunsworth HM Harcourt-Smith WEH Horner W Lehmann T Nightengale S McNulty KP 2014Remnants of an ancient forest provide ecological context for Early Miocenefossil apes Nature Comm 5 3236
Myres JL 1932 Africa Man 32 208Nakatsukasa M Kunimatsu Y Nakano Y Takano T Ishida H 2007 Vertebral
morphology of Nacholapithecus kerioi based on KNM-BG 35250 J Hum Evol52 347e369
Nengo IO Rae TC 1992 New hominoid fossils from the early Miocene site ofSonghor Kenya J Hum Evol 23 423e429
Peppe DJ McNulty KP Cote SM Harcourt-Smith WEH Dunsworth HM VanCouvering JA 2009 Stratigraphic interpretation of the Kulu Formation (EarlyMiocene Rusinga Island Kenya) and its implications for primate evolutionJ Hum Evol 56 447e461
Pickford M 1981 Preliminary Miocene mammalian biostratigraphy for westernKenya J Hum Evol 10 73e97
Pickford M 1986 Sexual dimorphism in Proconsul Hum Evol 1 111e148Pickford M Andrews PJ 1981 The Tinderet Miocene sequence in Kenya J Hum
Evol 10 11e33Pickford M Kunimatsu Y 2005 Catarrhines from the Middle Miocene (ca 145
Ma) of Kipsaraman Tugen Hills Kenya Anthropol Sci 113 189e224Pickford M Senut B Gommery D Musiime E 2009 Distinctiveness of Ugan-
dapithecus from Proconsul Estudios Geol 65 183e241Pilbeam DR 1969 Tertiary Pongidae of East Africa evolutionary relationships and
taxonomy Bulletin of the Peabody Museum of Natural History 31 pp 1e185Rae TC 1993 Phylogenetic analysis of proconsulid facial morphology PhD
Dissertation State University of New York at Stony BrookRae TC 1999 Mosaic evolution in the origin of the Hominoidea Folia Primatol 70
125e135Rafferty KL Walker A Ruff C Rose MD Andrews PJ 1995 Postcranial esti-
mates of body weights in Proconsul with a note on a distal tibia of P major fromNapak Uganda Am J Phys Anthropol 97 391e402
Rossie JB MacLatchy L 2013 Dentognathic remains of an Afropithecus individualfrom Kalodirr Kenya J Hum Evol 65 199e208
Senut B Pickford M Gommery D Kunimatsu Y 2000 Un nouveau genredrsquohominoiumlde du Miocene inferieur drsquoAfrique orientale Ugandapithecus major(Le Gros Clark and Leakey 1950) CR Acad Sci Paris 331 227e233
Simons EL Pilbeam DR 1965 Preliminary revision of the Dryopithecinae (Pon-gidae Anthropoidea) Folia Primatol 3 81e152
Smith TM Martin LB Leakey MG 2003 Enamel thickness microstructure anddevelopment in Afropithecus turkanensis J Hum Evol 44 283e306
Stevens NJ Seiffert ER OConnor PM Roberts EM Schmitz MD Krause CGorscak E Ngasala S Hieronymus TL Temu J 2013 Palaeontological evi-dence for an Oligocene divergence between Old World monkeys and apesNature 497 611e614
Suwa G Kono RT Katoh S Asfaw B Beyene Y 2007 A new species of great apefrom the late Miocene epoch in Ethiopia Nature 448 921e924
Szalay FS Delson E 1979 Evolutionary History of the Primates Academic PressNew York
Teaford MF Beard KC Leakey RE Walker A 1988 New hominoid facial skel-eton from the Early Miocene of Rusinga Island Kenya and its bearing on therelationship between Proconsul nyanzae and Proconsul africanus J Hum Evol17 461e477
Van Couvering JAH Miller J 1969 Miocene stratigraphy and age determinationsRusinga Island Kenya Nature 221 628e632
Walker A 1997 Proconsul function and phylogeny In Begun DR Ward CVRose MD (Eds) Function Phylogeny and Fossils Miocene Hominoid Evolu-tion and Adaptation Plenum Press New York pp 209e224
Walker A Pickford M 1983 New postcranial fossils of Proconsul africanus andProconsul nyanzae In Ciochon RL Corruccini RS (Eds) New Interpretationsof Ape and Human Ancestry Plenum Press New York pp 325e351
Walker A Teaford M 1988 The Kaswanga primate site an Early Miocene hom-inoid site on Rusinga Island Kenya J Hum Evol 17 539e544
Walker A Teaford M 1989 The hunt for Proconsul Sci Am 260 76e82Walker A Teaford MF Martin L Andrews P 1993 A new species of Proconsul
from the early Miocene of RusingaMfangano Islands Kenya J Hum Evol 2543e56
Ward CV Ruff CB Walker A Rose MD Teaford MF Nengo IO 1995 Func-tional morphology of Proconsul patellas from Rusinga Island Kenya with im-plications for other Miocene-Pliocene catarrhines J Hum Evol 29 1e19
Ward S Brown B Hill A Kelley J Downs W 1999 Equatorius a new hominoidgenus from the Middle Miocene of Kenya Science 285 1382e1386
Werdelin L Sanders WJ 2010 Cenozoic Mammals of Africa University of Cali-fornia Press Berkeley

Figure 6 Occlusal view of the M2s of Proconsul and Ekembo Left P africanus holotype(M 14084) Right E heseloni holotype (KNM-RU 2036) Note in Proconsul the overalldifference in crown shape as well as the more conical individuated cusps and nar-rower cristae strongly developed lingual cingulum that continues to the distal aspectof the crown broad buccal cingular shelf and more distally directed posthypoconecrista In Ekembo the cusps are more bunodont and expanded the cristae are broaderwith better developed accessory tubercles the lingual cingulum merges into thehypocone there is no buccal shelf and the posthypocone crista is more buccallydirected
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6152
is observable even in small samples of Proconsul species TheP africanus holotype has a substantially reduced M3 whereas inKNM-CA 2250dattributed by most authors to this species(Andrews 1978 Pickford et al 2009)dit is fully developed Simi-larly the hypodigm of P major includes both fully developed M3s(eg KNM-CA 389 LG 1815) and specimens with reduced ormissing hypocones (KNM-CA 397 1299) Ekembo displays compa-rable ranges of variation from highly reduced to fully developedM3s in both species Thus these species and genera cannot bedistinguished based on M3 development and we did not find anyother distinguishing characteristics between Ekembo and Proconsulin the upper third molars
Figure 7 Scatter plot depicting the de
Mandible Pickford et al (2009) enumerated several traits thatreadily distinguish Kisingiri from non-Kisingiri mandibles andthese distinctions are entirely consistent with the distinction ofEkembo from Proconsul Our analyses specifically underscoredifferences in the symphyseal region The planum alveolare inboth male and female specimens of Ekembo is short and morevertically oriented even the most elongate of these (KNM-RU47805 McNulty et al 2007) is substantially more verticalcompared to specimens of Proconsul In contrast the planumalveolare in Proconsul is conspicuously long and nearlyhorizontally oriented Well-preserved mandibles attributed toP major (eg KNM-SO 396 and 404) exhibit this morphology inextreme fashion (Fig 8) smaller specimens (KNM-SO 1112P africanus) show less extreme expression but are still readilydistinguishable from Ekembo mandibles For its size KNM-LG 452has a surprisingly short planum alveolare though not to thedegree seen in Rusinga mandibles Correlated to the shortplanum alveolare in Ekembo is a reduced superior transversetorus which while well-developed does not approach therobusticity expressed in Proconsul mandibles This difference isreflected in an index of symphyseal robusticity (thicknessperpendicular to the long axis of the symphysislength of thelong axis) which shows Ekembo and Proconsul to be significantlydifferent (t-test df frac14 14 t frac14 430 p frac14 00007)
Development of the inferior transverse torusdoften used todifferentiate among species of Proconsul (eg Pickford et al 2009Harrison 2010)dwas found to be highly variable and inconsis-tently expressed within these samples However the overallorientation of the subplanum region (as defined by a line con-necting the posterior-most midline points of the superior andinferior transverse tori or the posterior-most midline point of theinferior margin of the symphysis in the absence of the latter) doesdistinguish Ekembo from Proconsul mandibles The orientation ofthis line in relation to the alveolar margin in the vicinity of thepostcanine teeth was assessed qualitatively by placing a small rod
gree of buccolingual flare in M2
Figure 8 Mandibles of Proconsul and Ekembo Occlusal views of P major a) KNM-SO 396 b) KNM-SO 404 c) KNM-LG 452 Occlusal views of Ekembo d) KNM-RU 2087 e) KNM-RU1674 f) KNM-RU 7290 Occlusal views of P africanus g) KNM-SO 372 h) KNM-SO 1112 i) Lingual view of P africanus (KNM-SO 1112) Note the long more horizontally orientedplanum alveolare in Proconsul resulting in a pronounced superior transverse torus The planum alveolare is shorter and more vertical in Ekembo such that one can see the inferiormandibular border from occlusal view
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 53
(eg a dental pick) in themidline internally along the two referencepoints and recording the point in lingual perspective at which therod intersected the alveolar margin (Fig 9) In Ekembo the sub-planum region is more vertically oriented so that the rod intersectsthe alveolar margin most often in the premolar field Not surpris-ingly there is a difference between sexes with females having areduced superior transverse torus and therefore a more verticallyoriented subplanum region compared to males In contrast withEkembo Proconsul mandibles have much more inclined subplanumsurfaces with the rod intersecting the lingual alveolar margin inthe vicinity of the M1 (females) or M2 (males Fig 9) When sex istaken into account therefore the distributions of Ekembo andProconsul in this feature are discrete Note that male and femalemandibles attributed to both P africanus and P major are includedin this assessment and are surprisingly similar despite dramaticsize differences
We also quantified internal symphyseal morphology using threemeasures symphyseal height (infradentaleegnathion) planumalveolare length (posterior-most point on the superior transversetoruseinfradentale) and subplanum length (posterior-most pointon the superior transverse torusegnathion) a combination thatincorporates not only the lengths of these intervals but also theangular relationships among them To compare symphyseal shapeacross a range of sizes we chose a simple principal componentsapproach to partition size onto the first eigenvector (cf Jolicouer1963) While this is not the preferred method for partitioning sizein most datasets (Jungers et al 1995) for this particular sample andset of measurements Mosimann shape variables were still
significantly correlatedwith size (see discussion in Coleman 2008)Hence we assessed differences in symphyseal morphology be-tween Ekembo and Proconsul using the second and third principalcomponents of the three variables Distributions of the two generaare distinct in this shape space (Fig 10 Supplementary OnlineMaterial [SOM] Table 1)
Robusticity of the mandibular corpus also differs between thesesamples though it is more difficult to test due to differentialpreservation among specimens Corpus thickness and depth underP4 can be measured in the greatest number of specimens andcomparison of these measures suggests a difference betweengroups in the scaling of corpus robusticity (Fig 11) Among largepresumably male specimens Ekembo mandibles are substantiallyless robust than those of Proconsul However among smallermandibles only a single Proconsul specimen (KNM-SO 372) could beincluded and it is similar in robusticity to Ekembomandibles of thissize Considered together the impression is that corpus robusticitymay scale differently in Ekembo than it does in Proconsul and a testfor differences in slopes using an ANCOVA design of the loggedvariables was significant (df frac14 2 F frac14 2481 p lt 00001) Never-theless additional specimens are needed to properly evaluate theinfluence of taxonomy sex and size on corpus robusticity areasonable alternative interpretation of Figure 11 is that it dem-onstrates a significant difference between P major and all otherspecies of both Proconsul and Ekembo
Lower incisors Ekembo specimens are uniformly narrower inmesiodistal length than incisors attributed to P africanus P major
Figure 10 Scatter plot of principal component scores based on an eigenanalysis of threeposterior point on the superior transverse toruseinfradentale) and subplanum length (mprimarily captures size differences among specimens (Jolicouer 1963 see text and SOM Trespectively) in the symphyseal variables
Figure 9 Line drawing of a hemimandible depicting different orientations of the sub-planum area in Ekembo and Proconsul Females are denoted using shorter vectors maleswith longer vectors Vector is estimated in M 14086 which is broken at the midlineEkembo specimens have a more vertical subplanum compared to those from Songhorand Legetet but within both groups females have more vertically oriented regions thanmales Hence even the most ventrally rotated subplanum in the largest Ekembo malespecimen (KNM-RU 47805) groups with the Tinderet females in this feature
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6154
and P meswae both absolutely and in relation to crown height(Fig 12) This is true for both I1 and I2 A t-test adjusted for non-independent observations (eg I1 and I2 from the same mandible)confirmed the difference as significant (t-test df frac14 12 t frac14 258p frac14 00242) Moreover while the measurable Proconsul samplewas small this same pattern is observable in other specimenswhose tips were slightly to moderately worn and therefore notmeasured
Lower canines The only lower canine trait that discretely sepa-rates these genera is the pointed crown tip in Ekembo (Fig 3d)contrasted with a blade-like (or ldquoburinrdquo) tip in Proconsul(Fig 3c) This feature is well established for P major but is alsoexpressed in KNM-CA 1772 which was referred to P africanus byKelley (1986) and is too small to be either P major or P meswaeTwo other specimens commonly attributed to P africanus KNM-CA 2149 and KNM-SO 1112 both have broken canine tipsLikewise a new specimen (Kor 65004) attributed to P africanusby Pickford et al (2009) also has a broken tip and cannot beevaluated (see Pickford et al 2009 Fig 23) As with the uppercanines the blade-like tip is also present in the deciduous lowercanines of P major (eg KNM-CA 361 S0 5352) and P meswae(KNM-ME 1)
Lower premolars Like most previous authors (eg Andrews 1978Harrison 2002 2010 Pickford et al 2009) we found neitherqualitative nor quantitative characteristics of the lower premolarswith any taxonomic value
Lower molars Differences in lower molar morphology are few andrelatively subtle but in some cases mirror those seen in the uppermolars Cusps on the lower molars of Ekembo are more bunodontthan those in Proconsul and occupy more of the occlusal surfaceBroad inter-cuspal crests are incorporated into the Ekembo cuspsmaking them appear more inflated Molar cusps of Proconsul are
variables symphyseal height (infradentaleegnathion) planum alveolare length (mostost posterior point on the superior transverse torusegnathion) The first componentable 1) and thus PC 2 and PC 3 illustrate the residual shape variance (72 and 28
Figure 11 Scatter plot showing mandibular corpus robusticity under P4 Trend lines were computed using ordinary least squares (OLS) regression Similar relationships areobservable at other parts of the corpus but differential preservation results in smaller samples at those positions Slopes are significantly different (p lt 00001 tested on loggedvariables)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 55
more distinct from one another and are linked by narrower sharperinter-cuspal crests The lower molars of Ekembo also have asubstantially less developed buccal cingulum than those ofProconsul (Harrison 2002 2010 Harrison and Andrews 2009)
Figure 12 Scatter plot of lower incisor crown proportions Differenc
Lastly and again as seen in the upper molars the two genera canbe distinguished by the significantly greater amount of lowermolar flare in Proconsul (Fig 13 t-test df frac14 59 t frac14 713p lt 00001 see also Harrison and Andrews 2009)
es between Ekembo and Proconsul are significant (p frac14 00242)
Figure 13 Box-and-whisker plot of buccolingual flare in lower molars measured asthe average distance between the mesial buccal and lingual cusps divided by overallbuccolingual crown breadth Differences between Ekembo and Proconsul are highlysignificant (p lt 00001)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6156
Relative molar size has been used to distinguish species ofProconsul but we observed the primary difference to be in relativebuccolingual breadths of M1 and M2 In Ekembo M1 is substantiallynarrower than M2 than is the case in Proconsul (Fig 14 Harrison2002 2010) Although none of the smaller Proconsul mandiblespreserve complete crowns for both M1 and M2 KNM-SO 1112 hasan intact M2 and at least preserves the cervix for M1 However evenif the cervical measurement is used for its M1 breadthdthusunderestimating the true crown breadthdKNM-SO 1112 still has abroader first molar in relation to M2 than would be expected inEkembo If crown breadth is estimated from the ratio of cervixbreadth to maximum crown breadth in other Proconsul fossils therange of likely breadths for the SO 1112 M1 clearly distinguishes it
from the Ekembo sample (Fig 14) Testing this relationship onlogged variables using the mean estimate for the SO 1112 M1(Fig 14) reveals a significant difference between the two groups(ANCOVA df frac14 1 F frac14 2424 p frac14 00002)
As with upper third molars we find a great deal of variation inM3 morphology and mistrust the taxonomic significance some-times placed on characteristics of this toothdat least with regard tothese taxa Harrison (2002 2010) cited the mesiodistal position ofthe M3 entoconid as differentiating Kisingiri third molars fromthose attributed to P africanus We concur with his assessment butfound the entoconid position to vary in specimens attributed toP major Thus this character does not differentiate Ekembo from allProconsul species but may distinguish it from the type speciesP africanus
Discussion
The distinctiveness of the Kisingiri large catarrhine materialfrom that at Tinderet and Ugandan localities has been noted pre-viously by several authors (Pickford 1986 Walker et al 1993Begun 2001 2007 MacLatchy and Rossie 2005 Harrison andAndrews 2009 Harrison 2010) What had not been fully docu-mented was the nearly complete lack of overlap between thesesamples in a number of features despite very large assemblagesfrom Rusinga and Mfangano On descriptive criteria alone (Table 2)many isolated teeth (I1 C1 P3 M2 C1) and all of the mandibles canbe assigned to either Proconsul or Ekembowith very little chance ofmisattribution That some of the most diagnostic features occur inthe upper dentitiondthe only anatomy preserved in theP africanus type specimendadds weight to the taxonomicarrangement proposed here The morphology of the upper teeth inall Proconsul species compellingly unites them into a single cladedistinct from E heseloni and E nyanzae This is reinforced bynumerous features of the mandible and mandibular dentition thatfurther distinguish these samples The number and variety of thesefeatures are such that Proconsul and Ekembo are more differentfrom one another than are many closely-related modern catarrhinegenera
Derived morphology of Ekembo and Proconsul
It is premature to formally assign primitive and derived status tothe individual features of Ekembo and Proconsul before completinga formal phylogenetic analysis This is especially true given theabundance of evidence available for Ekembo and hence the poten-tial impact that our generic revision could have on phylogeneticinterpretations Nevertheless in comparison with both extanthominoids and the small-bodied catarrhines from the earlyMiocene it is informative to differentiate the evolutionary grades ofthese genera Many characteristics of Ekembo are more advanced inthe direction of modern apes compared to those of Proconsul(Table 2) General trends such as expanded and more bunodontcusps broader molar crests reduced cingulum and morphologicalsimilarity between P3 and P4 all represent morphologies thatpresage those of later fossil and extant hominoids The moreadvanced morphologies of P3 and M2 are particularly conspicuouscompared not only with those of Proconsul but to most earlyMiocene catarrhines In contrast the mesially positioned stronglyheteromorphic cusps on the P3 of Proconsul plus the correspondingasymmetric buccal profile notched mesial margin and missingmesial fovea are characteristic of other early Miocene catarrhinessuch as Dendropithecus Rangwapithecus and Limnopithecus andare likely primitive among East African catarrhines The moresymmetric P3 of Ekembo with midline positioned cusps andreduced cusp heteromorphy is reminiscent of later hominoids and
Figure 14 Relative buccolingual dimensions of M1 and M2 KNM-SO 1112 has a preserved M2 but only the cervix of M1 Its M1 breadth was estimated based on the ratio to cervixbreadth to maximum crown breadth in other Tinderet fossils Even the cervical breadth of SO 1112 is broader than would be predicted for the crown breadth of a Kisingiri M1 Trendlines computed using OLS regression and using the average estimate of SO 1112 M1 breadth Group differences were significant (p frac14 00002)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 57
even hominid taxa such as Rudapithecus (eg Kordos and Begun2001)
Proconsul is likewise united by a suite of features that are for themost part characteristic only of this lineage (Table 2) Themorphology of both upper and lower canines is unique to thisgenus and shared by all three well-known species plus the poorlyrepresented middle Miocene P gitongai (Pickford and Kunimatsu2005) The mandibular symphysis is also distinctive in Proconsulcompared with other catarrhines with its very long and shallowlysloping planum alveolare and steeply inclined subplanum surfaceIndeed even the small edentulous anterior mandibular fragmentKNM-SO 372 is readily placed within Proconsul and excluded fromother similarly sized catarrhine taxa such as Rangwapithecus andEkembo based on this unique symphyseal morphology (Figs 8 and9)
Other differences between Ekembo and Proconsul are of uncer-tain evolutionary polarity (eg orientation of the posthypoconecrista) or may be of taxonomic relevance without broader phylo-genetic significance (eg degree of molar flare Table 2) Werecognize that there are morphological differences within Procon-sul particularly between the better known species P africanus andP major However many of these differences are likely related tosize and represent variations on a single generic theme that aremarkedly different from the morphology of Ekembo The recentdescription (Harrison and Andrews 2009) of the intermediatelysized P meswae underscores the broad morphological unity withinProconsul and further substantiates a well-defined Proconsul cladecomprising Tinderet and Ugandan species
Based on the abundant morphological and morphometric evi-dence supporting genus-level distinction between Proconsul andEkembo we are currently conducting a more detailed phylogeneticanalysis of all early Miocene catarrhines in order to better under-stand the distribution and evolutionary significance of these andother morphological features We anticipate that the debates sur-rounding the phylogenetic relationships of an historically consti-tuted sample of Proconsul can be more satisfactorily resolved nowthat the Kisingiri material is appropriately recognized as a distinctgenus
The generic disposition of ldquoUgandapithecusrdquo species
A great many features said to characterize ldquoUgandapithecusrdquowere added to the original diagnosis of the genus by Pickford et al(2009) in their systematic revision of Proconsul (sensu lato) As partof that study they also erected a new species ldquoUrdquo legetetensis andtransferred P meswae to ldquoUgandapithecusrdquo Evaluating the same setof features across the entire available sample however we wereunable to find support for this more expansive conception ofldquoUgandapithecusrdquo for several reasons First some diagnostic fea-tures cited in Pickford et al (2009) were quite variable within theldquoUgandapithecusrdquo sample As an example marked reduction of theM3 entoconid to a bead on the lingual cingulum is not shared byseveral ldquoUrdquo major specimens such as KNM-CA 393 NAP I 49000and Kor 253004 (see Pickford et al 2009 Fig 33e) and was likelynot a feature of a few of the more worn specimens eg BUMP 600and BUMP 601 or even the holotype of P major M 16648 Second
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6158
some diagnostic features of ldquoUgandapithecusrdquo were found to bevariably present in Ekembo arguing against their utility for dis-tinguishing ldquoUgandapithecusrdquo from Pickford et als (2009) com-bined P africanus thorn Kisingiri conception of Proconsul For examplesignificantly reduced curvature of the I1 distal margin in ldquoUganda-pithecusrdquo is also found in some Kisingiri specimens including KNM-RU 1685 1712 1714 1831 1846 and 1951 several more Rusingaspecimens have moderately reduced curvature Third some fea-tures that purportedly distinguish ldquoUgandapithecusrdquo fromP africanus are likely related to size For example the somewhatmore central position of the P3 paracone in ldquoUrdquomajor is not seen inthe smaller ldquoUrdquo meswae whose paracone is positioned similarly tothat of the holotype of P africanus (see Harrison and Andrews2009 Fig 6d)
However the major part of the evidence for recognizingldquoUgandapithecusrdquo (sensu Pickford et al 2009) was assembled by re-assigning specimens traditionally attributed to P africanus into newspecies of ldquoUgandapithecusrdquo The P africanus holotype was left inassociation with the Rusinga sample despite having morphologyoverwhelmingly characteristic of other Tinderet specimens Theresulting specimen allocations left P africanus morphologicallyimpoverisheddstripped of much of its hypodigmdso that Pro-consul for Pickford et al (2009) came to be characterized primarilyby the Rusinga sample (see similar comments in Harrison andAndrews 2009) Within that conceptual framework any spec-imen that differs from those at Rusinga could not be Proconsul butmust instead belong to ldquoUgandapithecusrdquo Thus specimens such asthe KNM-SO 1112 mandible were erroneously assigned to ldquoUgan-dapithecusrdquo because they were (correctly) perceived as differingfrom their Rusinga counterparts (Pickford et al 2009)
Here we restore the P africanus hypodigm and associate it withthe other Tinderet fossils according to their shared morphologySpecimens assigned to ldquoUgandapithecusrdquo revert to Proconsul andmany of the features cited to support a speciose ldquoUgandapithecusrdquo(Pickford et al 2009) instead become diagnostic features of Pro-consul This correction serves to highlight the distinctiveness of theKisingiri remains and the need to recognize this distinctiveness atthe generic level
As demonstrated above specimens from Kisingiri do not displaythe morphology of Proconsul as represented by the holotype ofP africanus M 14084 Given that this specimen preserves only theupper canine and cheek teeth similarities with or departures fromthis morphology represent the most compelling arguments forassessing inclusivity in the genus However for the mandible andlower dentition if one simply sorts the specimens by elementwithout relying on taxonomic preconceptions the overwhelmingdistinction within the traditionally constituted Proconsul sample isbetween Kisingiri material on the one hand and that from Tinderetand Uganda on the other (cf Walker et al 1993 Begun 2001 2007MacLatchy and Rossie 2005 Harrison and Andrews 2009Harrison 2010) Additionally with the Kisingiri specimensremoved to Ekembo the postcranial differences used to partlyjustify a distinct ldquoUgandapithecusrdquo lose their relevance (seeHarrison and Andrews 2009)
We have differed among ourselves in our past treatment ofP major as generically distinct from the smaller species of Pro-consul (Begun 2007 McNulty 2010) Morphologically many ofthe unique features of P major can be interpreted as size-relatedvariants on the smaller P africanus morphology Even if so thequestion is whether these are sufficiently different and numerousto warrant generic distinction that is do they suggest thatP major belongs in a separate clade from P africanus andP meswae As we have demonstrated differences among thesethree species are certainly fewer and show less morphologicaldistinction than those between Ekembo and any of the three
Nevertheless while we ultimately agree with Harrison andAndrews (2009 see also Begun 2007) that the features dis-tinguishing ldquoUrdquo major from P africanus are consistent with thosethat distinguish congeneric species and are at least partly due todifferences in size we concede the possibility that the largestProconsul species could belong in a separate genus (Senut et al2000 Pickford and Kunimatsu 2005) However whether or notone recognizes a separate genus for P major the P africanus ma-terial does not sort morphologically with the Kisingiri materialTherefore the taxonomic alternatives are either two generaProconsul and Ekembo or three Proconsul Ugandapithecus andEkembo with Proconsul and Ugandapithecus united in a cladeexclusive of Ekembo Thus while we strongly favor the firstalternative recognizing Ugandapithecus does not change thefundamental argument concerning the need to refer the Kisingirispecies to a generically and cladistically distinct taxon from thespecies represented at Tinderet and the Ugandan sites
Specimens of interest
Despite the substantial dental and gnathic differences betweenProconsul and Ekembo there are a few specimens whose taxonomicaffinities require additional scrutiny
KNM-RU 1676-77 This set of associated upper and lower teeth is aconsistent outlier among E heseloni and E nyanzae specimens Insome ways this specimen more closely resembles Tinderet Pro-consul in the features that distinguish it from Ekembo Molar cuspmorphology and cingulum development I1 lingual tubercledevelopment and M2 shape and posthypocone crista orientationare more consistent with specimens from the Tinderet sites On theother hand it otherwise resembles Ekembo and lacks the strongestdiagnostic features that unite species of Proconsul such as thedistinctive canine and P3 morphology Overall this specimen doesnot fit unambiguously into either genus in fact the degree ofbuccolingual flare cusp morphology enamel wrinkling and espe-cially the long extension of enamel onto the anterior root in P3 arein some ways reminiscent of Afropithecus although it clearly differsfrom that genus as well (cf Rossie and MacLatchy 2013)
Were these characteristics individually distributed among thelarge Kisingiri sample one might attribute this to intra-genericvariation but when found together in a single specimen from alarge sample it suggests a unique individual One of us (KPM)thinks it likely that the specimen was not actually found at aKisingiri locality based on its unique preservation compared toother Rusinga fossils and on other examples of mis-proveniencedspecimens (Leakey 1967 Andrews and Molleson 1979 Pickford1986 Kelley 1986) However we recognize that relying toofrequently on this explanation allows almost any scenario to bejustified and suggest some alternative possibilities One is that thespecimen was found on Rusinga but in the much older WayandoFormation strata instead of the Hiwegi Formation The differencesin morphology in that case might reflect more than a million yearsof temporal change (McCollum et al 2013) with KNM-RU 1676-77preserving certain more primitive Proconsul-like features As analternative it is reasonable to assume that fossil assemblages asvast as those from Rusingawill include not only abundant taxa butalso rare or infrequent inhabitants of the local biome There aremany examples of this from Rusingas mammalian communityeg ldquoHemicyonrdquo Kulutherium Tadarida cf Hyainailouros cfArchaeobeledon Kelba (see relevant chapters in Werdelin andSanders 2010) As such KNM-RU 1676-77 may simply representa rare and well-preserved specimen of some other catarrhinetaxon that lived among or visited a primate community dominatedby Ekembo
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 59
KNM-SO 1112 This specimen was initially referred to Rangwapi-thecus but soon after was transferred to Proconsul afrianus (Bosler1981 Martin 1981)da designation that most subsequent authorshave followed (Pickford 1986 Nengo and Rae 1992 Kelley 19861995 Walker et al 1993 MacLatchy and Rossie 2005 Hill et al2013) Pickford et al (2009) more recently referred this specimento ldquoUrdquo meswae citing two primary reasons First they reiteratedBoslers (1981) concern that its P3 differs from others attributedto P africanus However at that time the majority of what wasconsidered to be P africanus would have been from RusingaIsland In fact while noting its contrasting morphology Boslersuggested that differences between KNM-SO 1112 and other P3smight reasonably be attributed to sexual dimorphism notingthat this specimen provides ldquoa not implausible match for the Pafricanus holotype maxilla M 14084rdquo (1981152) Thatassessment is borne out in this study Using the much largersample of Ekembo P3s as a model variation can be seen toencompass specimens with well-developed mesial beaks andthose with more bilaterally compressed crowns and parallelsides The latter morphology is predominantly found amongmale specimens (KNM-RU 1674 1924 and KPS I and III) thussupporting Boslers (1981) suggestion that this variation isrelated to sexual dimorphism and hence that KNM-SO 1112 is amale P africanus
The second feature cited by Pickford et al (2009) in referringthis specimen to ldquoUrdquo meswae was posterior shallowing of themandibular corpus However because they redefined ldquoUgandapi-thecusrdquo to include all of the well-preserved Tinderet mandibles(Pickford et al 2009)dleaving Proconsul to be represented only byRusinga specimensdtheir only alternative within this scheme for aspecimen the size of SO 1112 was to refer it to ldquoUrdquomeswae It seemsodd though to assign this specimen to a species from a (perhapsmuch) older site when the Proconsul africanus holotype is ofappropriate size and similar age and as there are no other adultmandibles from either species with which to compare it In anycase mandibular shallowing is variable in both Ekembo and Pro-consul with some Kisingiri specimens exhibiting considerableshallowing (eg KNM-RU 2087) and some ldquoUgandapithecusrdquo spec-imens having very little (eg M 14086)
We therefore see no morphological grounds to assign KNM-SO1112 to P meswae but do find some morphological evidence toassign it as a male P africanus which occurs at Songhor
KNM-LG 452 This very complete mandible was originally recog-nized as a female of P major based on the size of the dentition androbusticity of the mandible (Martin 1981) Bosler (1981) howeverleft this specimen unassigned noting similarities to P major in P3-M1 size but having distinctly smaller M2e3 In this she likenedKNM-LG 452 to a few other specimens (notably M 14086) thatare smaller in their distal molars in a manner more consistentwith taxonomic rather than sex differences (Bosler 1981 see alsoPilbeam 1969) Kelley (1986 1995) also assigned this specimen asa female of P major based on canine size and morphology butnoted that the canine lacked the shallowly-sloping mesial ridgetypical of most female anthropoid canines This is also truehowever of two other canines assigned as female P major KO 9and SO 373 (Kelley 1986) and so might simply be an unusualcharacteristic of P major female lower canines In the end Kelley(1995) concluded that considering all of the criteria fordistinguishing male and female canines the evidence that KNM-LG 452 is female remains questionable
Pickford et al (2009) further developed the idea that two spe-cies are represented within the P major sample and made KNM-LG452 the holotype of a new species ldquoUrdquo legetetensis We concur thatthis specimen is unique among the well-preserved Tinderet
mandibles in some important features Whereas male and femalemandibles of P major have long internal symphyseal planesexpansive extra- and retromolar sulci and rami that cross theocclusal plane at distal M3 KNM-LG 452 has a shorter symphysealplane (though not to the extent seen in Ekembo) a moderateextramolar sulcus but no retromolar sulcus and a ramus thatcrosses the occlusal plane more mesially If KNM-SO 396 and SO404 represent males and females respectively of P major then themorphology of KNM-LG 452 appears distinct
At present however we do not think there is sufficient evidenceto differentiate this specimen from either P major or P meswaePickford et al (2009) argue that KNM-LG 452 cannot belong to thelatter species because of its larger size The size differences how-ever are well within the range of variation seen in P majorE heseloni and E nyanzae Moreover whereas the Legetet mandibleis larger than most of the P meswae hypodigm an adult P4 referredto this species (KNM-ME 25) substantially exceeds its counterpartin KNM-LG 452 Should the Legetetmandible ultimately prove to bea male specimen size alone would not rule out its attribution toP meswae On the other hand if it is female then it becomes harderto justify its inclusion in that sample of mostly smaller specimens Iffemale therefore it could represent a separate species for whichthe binomial Proconsul legetetensis comb nov is available Recentlycollected fossils from the SonghorKapurtay area provide someadditional support for the possibility that three species of Proconsulexisted at these localities (McNulty 2014ab) However toconvincingly demonstrate this and determine whether a thirdspecies can be differentiated from P meswae will require a moredetailed analysis of variation among Proconsul species a corre-sponding study of variation in modern taxa anddalmost certain-lydadditional fossil specimens For now we regard ldquoUrdquo legetetensisas a subjective junior synonym of P major
Conclusions
The genus Proconsul has been central to studies of ape and hu-man evolution since its first description by Hopwood (1933) As oneof the best known fossil catarrhines represented by nearly everyskeletal element our knowledge of Proconsuls functional anatomypositional behavior and overall morphology has made it thefoundation for paleoanthropological research in the early Mioceneas well as studies on the origin and early diversification of homi-noids With respect to systematics however Proconsul is less wellunderstood the type species P africanus is known from a rela-tively small number of specimens from sites in the Tinderet areaand no postcranial bones can be unambiguously assigned to itRather it is the copious and well-preserved specimens from theKisingiri sites of Rusinga and Mfangano Islands that have formedthe basis for the common image of Proconsul
We argue here that the Kisingiri species traditionally referredto ProconsuldP heseloni and P nyanzaedare generically distinctfrom P africanus and hereby transfer them into a new genusEkembo based on a substantial number of morphological differ-ences (Table 2) Considering only those traits preserved in theP africanus holotype the entire sample from Rusinga and Mfan-gano differs markedly in the morphology of the upper canine P3P4 and M2 Additional differences in the I1 lower incisors andcanines molar cusp and occlusal crest morphology degree of flarein all molars and especially mandibular features clearly distin-guish the Kisingiri from non-Kisingiri samples in a way that isincompatible with congeneric species With the transfer ofE heseloni and E nyanzae Proconsul becomes a well-defined cladecomprising at least three speciesdP africanus P major andP meswaedunited by several features that are unique for thegenus (Table 2)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6160
We found no support in this study for the genus ldquoUgandapi-thecusrdquo as constructed by Pickford et al (2009) In that paper theytransferred the vast majority of Proconsul fossils from the Tinderetand Napak localities into ldquoUgandapithecusrdquo leaving P africanusrepresented only by its holotype and a small number of isolatedteeth and aligning it with the material from Rusinga and Mfan-ganoWhen the P africanus hypodigm is reconstituted and properlyaffiliated with the other Tinderet species according to itsmorphology much of the evidence marshaled by Pickford et al(2009) for ldquoUgandapithecusrdquo actually supports the distinction be-tween the Tinderet and Kisingiri samples ie the generic distinc-tion of Ekembo In addition some of the traits enumerated byPickford and colleagues in support of ldquoUgandapithecusrdquo wererevealed in our analysis to be variable within one or more speciesand therefore of questionable taxonomic utility We do not rejectout of hand the original conception of ldquoUgandapithecusrdquo (Senutet al 2000) as either a monotypic genus distinguishing ldquoUrdquo ma-jor or perhaps including a closely allied middle Miocene ldquoUrdquogitongai as well (Pickford and Kunimatsu 2005) but we have fornow retained the early Miocene specimens of both in Proconsulpending further study However even if Ugandapithecus is a taxo-nomically valid genus comprising just the latter two species theirmorphology indicates that theywould still be united in a cladewithProconsul and they lack several more advanced features that partlydefine Ekembo Better understanding of the role that body size playsinmediating differences within Proconsulwill help to further clarifythe relationships among these species
Despite the existence of one well-represented and several lesscomplete skeletons of what was long assumed to be Proconsul thephylogenetic status of this genus has defied consensus This islargely due in our view to Proconsul having been an admixture oftwo distinct genera one more advanced in the direction of extantapes than the other As we demonstrate here the material fromRusinga and Mfangano traditionally referred to Proconsul andwhich includes all of the more complete skeletal material isdistinct from the Tinderet and Napak material and belongs in aseparate genus Ekembo By disentangling these distinct clades weanticipate being able to achieve better phylogenetic resolution forboth genera thereby improving our knowledge of early hominoidevolution and diversification
Acknowledgments
We thank many people and agencies that assisted with thedevelopment and completion of this project This research wasinitiated as a result of funding from the Wenner-Gren Foundationand the Johns Hopkins School of Medicine to DR Begun in1987e1989 and was renewed following research and travelgenerously funded by multiple Leakey Foundation grants to KMcNulty National Science Foundation grants to K McNulty(0852609 1241807) the McKnight Foundation the University ofMinnesota the Lloyd A Wilford Endowment the LeverhulmeFoundation and the Turkana Basin Institute J Kelley gratefullyacknowledges support from the Institute of Human Origins Ari-zona State University DR Begun also wishes to acknowledge theAlexander Humboldt Stiftung and NSERC for funding support Weare grateful to the National Museums of Kenya the UgandaMuseum the Natural History Museum and to Rose Mwanja EzraMusiime Tom Mukhuyu Samuel Muteti James Yatich and MaryMuungu for access to and assistance with specimens in their careMany people provided assistance comments criticisms insightsand suggestions on various aspects of this work and we especiallythank Holly Dunsworth Will Harcourt-Smith Tom Lehmann DanPeppe Nicole Garrett Kirsten Jenkins Rutger Jansma Susy CoteIsaiah Nengo Andrew Hill and Brenda Benefit K McNulty
acknowledges the evolutionary anthropology faculty and studentsat Durham University for their helpful comments on an earlyversion of this manuscript and for their gracious hospitality Weare likewise grateful to the anonymous reviewers associate editorand editor whose comments strengthened the paper This manu-script is publication 1 supporting Research on East AfricanCatarrhine and Hominoid Evolution (REACHE)
Appendix A Supplementary data
Supplementary data related to this article can be found at httpdxdoiorg101016jjhevol201503009
References
Andrews P 1974 New species of Dryopithecus from Kenya Nature 249 188e190Andrews P 1978 A revision of the Miocene Hominoidea of East Africa Bull Brit
Mus (Nat Hist) Geol 30 85e225Andrews P 1985 Family group systematics and evolution among catarrhine pri-
mates In Delson E (Ed) Ancestors The Hard Evidence Liss New Yorkpp 14e22
Andrews P Harrison T Martin L Pickford M 1981 Hominoid primates from anew Miocene locality named Meswa Bridge in Kenya J Hum Evol 10123e128
Andrews P Begun D Zylstra M 1997 Interrelationships between functionalmorphology and paleoenvironments in Miocene hominoids In Begun DWard C Rose M (Eds) Function Phylogeny and Fossils Miocene HominoidEvolution and Adaptations Plenum Press New York pp 29e58
Andrews P Molleson T 1979 The provenance of Sivapithecus africanus Bull BritMus (Nat Hist) Geol 32 19e23
Begun DR 2001 African and Eurasian Miocene hominoids and the origins of theHominidae In de Bonis L Koufos GD Andrews P (Eds) Hominoid Evolutionand Climatic Change in Europe Phylogeny of the Neogene Hominoid Primatesof Eurasia Cambridge University Press London pp 231e253
Begun DR 2007 Fossil record of Miocene hominoids In Henke W Tattersall I(Eds) Handbook of Paleoanthropology Springer Berlin pp 921e977
Begun DR Teaford MF Walker AC 1994 Comparative and functional anatomyof Proconsul phalanges from the Kaswanga primate site Rusinga Island KenyaJ Hum Evol 26 89e165
Bishop WW 1964 More fossil primates and other Miocene mammals from North-East Uganda Nature 203 1327e1331
Bishop WW Miller JA Fitch FJ 1969 New potassium-argon age determinationsrelevant to the Miocene fossil mammal sequence in East Africa Am J Sci 267669e699
Blankenship JA Mincer HH Anderson KM Woods MA Burton EL 2007Third molar development in the estimation of chronologic age in Americanblacks as compared with whites J Forensic Sci 52 428e433
Bosler W 1981 Species groupings of early Miocene dryopithecine teeth from EastAfrica J Hum Evol 10 151e158
Coleman MN 2008 What does geometric mean mean geometrically Assessingthe utility of geometric mean and other size variables in studies of skullallometry Am J Phys Anthropol 135 404e415
Cote S Malit N Nengo I 2013 Additional mandibles of Rangwapithecus gordonian early Miocene catarrhine from the Tinderet localities of Western KenyaJ Hum Evol 153 341e352
Deane AS 2009 Early Miocene catarrhine dietary behavior the influence of theRed Queen Effect on incisor shape and curvature J Hum Evol 56 275e285
Drake R Van Couvering JA Pickford M Curtis G Harris JA 1988 New chro-nology for the Early Miocene mammalian faunas of Kisingiri Western KenyaJ Geol Soc Lond 145 479e491
Evernden JF Savage DE Curtis GH James GT 1964 Potassium-argon datesand the Cenozoic mammalian chronology of North America Am J Sci 262145e198
Gebo DL MacLatchy L Kityo R Deino A Kingston J Pilbeam D 1997A hominoid genus from the Early Miocene of Uganda Science 276 401e404
Geoffroy Saint-Hilaire E 1812 Tableau des quadrumanes 1 Ord QuadrumanesAnnls Mus Hist Nat Paris 19 85e122
Gray JE 1825 Outline of an attempt at the disposition of the Mammalia into tribesand families with a list of the genera apparently appertaining to each tribeAnnls Philos 10 337e344
Harrison T 1982 Small-bodied apes from the Miocene of East Africa PhDDissertation University of London
Harrison T 1986 New fossil anthropoids from the middle Miocene of East Africaand their bearing on the origin of the Oreopithecidae Am J Phys Anthropol 71265e284
Harrison T 1987 The phylogenetic relationships of the early catarrhine primates areview of the current evidence J Hum Evol 16 41e80
Harrison T 2002 Late Oligocene to middle Miocene catarrhines from Afro-ArabiaIn Hartwig WC (Ed) The Primate Fossil Record Cambridge University PressCambridge pp 311e338
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 61
Harrison T 2010 Dendropithecoidea Proconsuloidea and Hominoidea (CatarrhiniPrimates) In Werdelin L Sanders WJ (Eds) Cenozoic Mammals of AfricaUniversity of California Press Berkeley pp 429e469
Harrison T Andrews P 2009 The anatomy and systematic position of a newspecies of early Miocene proconsulid from Meswa Bridge Kenya J Hum Evol56 479e496
Hill A Nengo IO Rossie JB 2013 A Rangwapithecus gordoni mandible from theearly Miocene site of Songhor Kenya J Hum Evol 65 490e500
Hopwood AT 1933 Miocene primates from Kenya J Linn Soc Zool 38 437e464Ishida H Kunimatsu Y Takano T Nakano Y Nakatsukasa M 2004 Nacholapi-
thecus skeleton from the Middle Miocene of Kenya J Hum Evol 46 69e103Jolicouer P 1963 The multivariate generalization of the allometry equation Bio-
metrics 19 497e499Jungers WL Falsetti AB Wall CE 1995 Shape relative size and size-
adjustments in morphometrics Am J Phys Anthropol 38 (S2) 137e161Kavanagh KD Evans AR Jernvall J 2007 Predicting evolutionary patterns of
mammalian teeth from development Nature 449 427e433Kelley J 1986 Species recognition and sexual dimorphism in Proconsul and
Rangwapithecus J Hum Evol 15 461e495Kelley J 1995 Sex determination in Miocene catarrhine primates Am J Phys
Anthropol 96 390e417Kelley J 1993 Taxonomic implications of sexual dimorphism in Lufengpithecus In
Kimbel WH Martin LB (Eds) Species Species Concepts and Primate Evo-lution Springer New York pp 429e458
Kent PE 1942 The country round the Kavirondo Gulf of Victoria Nyanza Geogr J100 22e31
Kordos L Begun DR 2001 A new cranium of Dryopithecus from RudabanyaHungary J Hum Evol 15 689e700
Le Gros Clark WE Leakey LSB 1950 Diagnoses of East African Miocene Homi-noidea Q J Geol Soc Lond 105 260e263
Le Gros Clark WE Leakey LSB 1951 The Miocene Hominoidea of East Africa InFossil Mammals of Africa No 1 British Museum (Natural History) London
Leakey LSB 1943 A Miocene anthropoid mandible from Rusinga Kenya Nature152 319e320
Leakey LSB 1963 East African fossil Hominoidea and the classification within thissuper-family In Washburn SL (Ed) Classification and Human EvolutionAldine Chicago pp 32e49
Leakey LSB 1967 Notes on the mammalian faunas from the Miocene and Pleis-tocene of East Africa In Bishop WW Clark JD (Eds) Background to Evolu-tion in Africa University of Chicago Press Chicago pp 7e29
Leakey RE Leakey MG 1986 A new Miocene hominoid from Kenya Nature 324143e146
Leakey RE Leakey MG Walker AC 1988 Morphology of Afropithecus turka-nensis from Kenya Am J Phys Anthropol 76 289e307
Leakey MG Ungar PS Walker A 1995 A new genus of large primate from theLate Oligocene of Lothidok Turkana District Kenya J Hum Evol 28519e531
Linnaeus C 1758 Systema naturae per regna tria naturae secundum classesordines genera species cum characteribus differentris synonymis locis tenthed Laurentii Salvii Stockholm
MacInnes DG 1943 Notes on the East African Miocene primates J E Afr UgandaNat Hist Soc 17 141e181
MacLatchy L 2004 The oldest ape Evol Anthropol 13 90e103MacLatchy L Bossert WH 1996 An analysis of the articular surface distribution of
the femoral head and acetabulum in anthropoids with implications for hipfunction in Miocene hominoids J Hum Evol 31 425e453
MacLatchy L Rossie JB 2005 The Napak hominoid still Proconsul major InLieberman DE Smith RJ Kelley J (Eds) Interpreting the Past Essays onHuman Primate and Mammal Evolution in Honor of David Pilbeam Brill Ac-ademic Publishers Boston pp 15e28
Martin L 1981 New specimens of Proconsul from Koru Kenya J Hum Evol 10139e150
McCollum MS Peppe DJ McNulty KP Dunsworth HM Harcourt-Smith WEH Andrews AL 2013 Magnetostratigraphy of the early MioceneHiwegi Formation (Rusinga Island Lake Victoria Kenya) Geological Society ofAmerica Abstracts with Programs 45 p 12
McNulty KP 2010 Apes and tricksters the evolution and diversification ofhumans closest relatives Evol Edu Outreach 3 322e332
McNulty KP Harcourt-Smith WEH Dunsworth HM 2007 New primate fossilsfrom Rusinga Island Kenya Am J Phys Anthropol 132 (S44) 170
McNulty KP MacLatchy L Peppe DJ Nengo I Manthi FK Miller ERStevens NJ Cote S Lehmann T Kingston J 2014a A regional approach toEast African early Miocene paleobiology J Vert Paleontol 185 SVP Programand Abstracts Book 2014
McNulty KP MacLatchy L Rossie JB Peppe DJ Deino AL Mbua ENManthi FK Nengo IO Stevens NJ Cote S Lehmann T Gutierrez M 2014b
Research on East African catarrhine and hominoid evolution results from thefirst year Am J Phys Anthropol 153 (S58) 182
Michel LA Peppe DJ Lutz JA Driese SG Dunsworth HM Harcourt-Smith WEH Horner W Lehmann T Nightengale S McNulty KP 2014Remnants of an ancient forest provide ecological context for Early Miocenefossil apes Nature Comm 5 3236
Myres JL 1932 Africa Man 32 208Nakatsukasa M Kunimatsu Y Nakano Y Takano T Ishida H 2007 Vertebral
morphology of Nacholapithecus kerioi based on KNM-BG 35250 J Hum Evol52 347e369
Nengo IO Rae TC 1992 New hominoid fossils from the early Miocene site ofSonghor Kenya J Hum Evol 23 423e429
Peppe DJ McNulty KP Cote SM Harcourt-Smith WEH Dunsworth HM VanCouvering JA 2009 Stratigraphic interpretation of the Kulu Formation (EarlyMiocene Rusinga Island Kenya) and its implications for primate evolutionJ Hum Evol 56 447e461
Pickford M 1981 Preliminary Miocene mammalian biostratigraphy for westernKenya J Hum Evol 10 73e97
Pickford M 1986 Sexual dimorphism in Proconsul Hum Evol 1 111e148Pickford M Andrews PJ 1981 The Tinderet Miocene sequence in Kenya J Hum
Evol 10 11e33Pickford M Kunimatsu Y 2005 Catarrhines from the Middle Miocene (ca 145
Ma) of Kipsaraman Tugen Hills Kenya Anthropol Sci 113 189e224Pickford M Senut B Gommery D Musiime E 2009 Distinctiveness of Ugan-
dapithecus from Proconsul Estudios Geol 65 183e241Pilbeam DR 1969 Tertiary Pongidae of East Africa evolutionary relationships and
taxonomy Bulletin of the Peabody Museum of Natural History 31 pp 1e185Rae TC 1993 Phylogenetic analysis of proconsulid facial morphology PhD
Dissertation State University of New York at Stony BrookRae TC 1999 Mosaic evolution in the origin of the Hominoidea Folia Primatol 70
125e135Rafferty KL Walker A Ruff C Rose MD Andrews PJ 1995 Postcranial esti-
mates of body weights in Proconsul with a note on a distal tibia of P major fromNapak Uganda Am J Phys Anthropol 97 391e402
Rossie JB MacLatchy L 2013 Dentognathic remains of an Afropithecus individualfrom Kalodirr Kenya J Hum Evol 65 199e208
Senut B Pickford M Gommery D Kunimatsu Y 2000 Un nouveau genredrsquohominoiumlde du Miocene inferieur drsquoAfrique orientale Ugandapithecus major(Le Gros Clark and Leakey 1950) CR Acad Sci Paris 331 227e233
Simons EL Pilbeam DR 1965 Preliminary revision of the Dryopithecinae (Pon-gidae Anthropoidea) Folia Primatol 3 81e152
Smith TM Martin LB Leakey MG 2003 Enamel thickness microstructure anddevelopment in Afropithecus turkanensis J Hum Evol 44 283e306
Stevens NJ Seiffert ER OConnor PM Roberts EM Schmitz MD Krause CGorscak E Ngasala S Hieronymus TL Temu J 2013 Palaeontological evi-dence for an Oligocene divergence between Old World monkeys and apesNature 497 611e614
Suwa G Kono RT Katoh S Asfaw B Beyene Y 2007 A new species of great apefrom the late Miocene epoch in Ethiopia Nature 448 921e924
Szalay FS Delson E 1979 Evolutionary History of the Primates Academic PressNew York
Teaford MF Beard KC Leakey RE Walker A 1988 New hominoid facial skel-eton from the Early Miocene of Rusinga Island Kenya and its bearing on therelationship between Proconsul nyanzae and Proconsul africanus J Hum Evol17 461e477
Van Couvering JAH Miller J 1969 Miocene stratigraphy and age determinationsRusinga Island Kenya Nature 221 628e632
Walker A 1997 Proconsul function and phylogeny In Begun DR Ward CVRose MD (Eds) Function Phylogeny and Fossils Miocene Hominoid Evolu-tion and Adaptation Plenum Press New York pp 209e224
Walker A Pickford M 1983 New postcranial fossils of Proconsul africanus andProconsul nyanzae In Ciochon RL Corruccini RS (Eds) New Interpretationsof Ape and Human Ancestry Plenum Press New York pp 325e351
Walker A Teaford M 1988 The Kaswanga primate site an Early Miocene hom-inoid site on Rusinga Island Kenya J Hum Evol 17 539e544
Walker A Teaford M 1989 The hunt for Proconsul Sci Am 260 76e82Walker A Teaford MF Martin L Andrews P 1993 A new species of Proconsul
from the early Miocene of RusingaMfangano Islands Kenya J Hum Evol 2543e56
Ward CV Ruff CB Walker A Rose MD Teaford MF Nengo IO 1995 Func-tional morphology of Proconsul patellas from Rusinga Island Kenya with im-plications for other Miocene-Pliocene catarrhines J Hum Evol 29 1e19
Ward S Brown B Hill A Kelley J Downs W 1999 Equatorius a new hominoidgenus from the Middle Miocene of Kenya Science 285 1382e1386
Werdelin L Sanders WJ 2010 Cenozoic Mammals of Africa University of Cali-fornia Press Berkeley
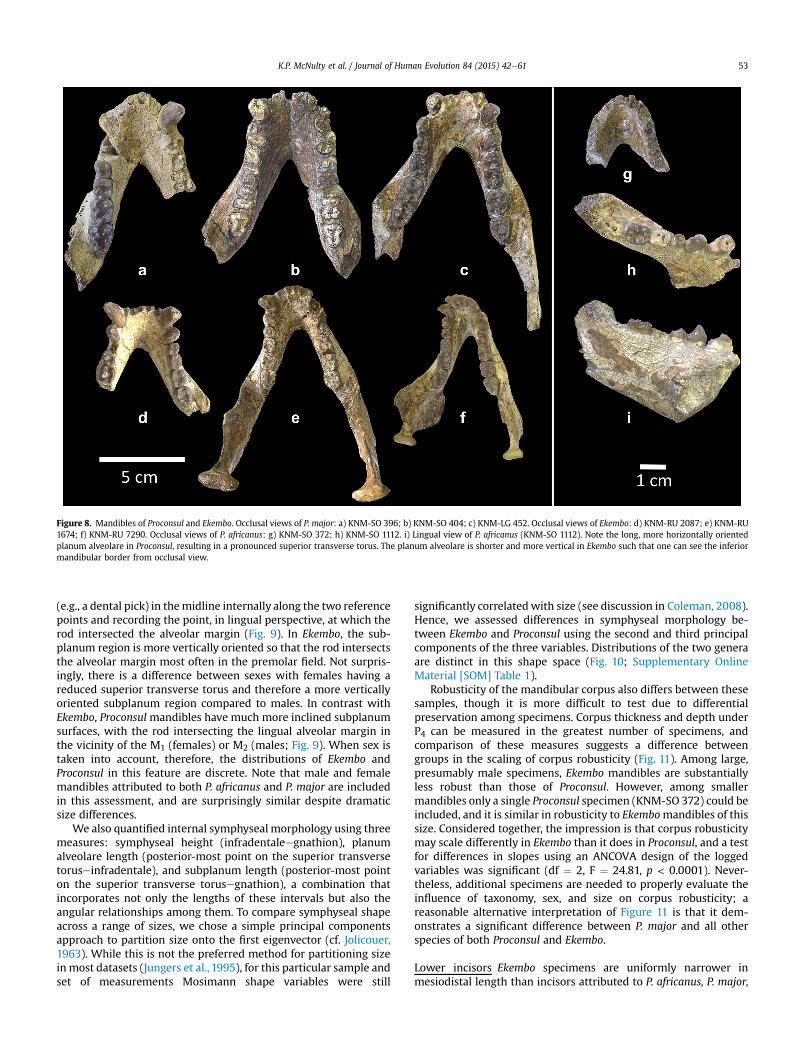
Figure 8 Mandibles of Proconsul and Ekembo Occlusal views of P major a) KNM-SO 396 b) KNM-SO 404 c) KNM-LG 452 Occlusal views of Ekembo d) KNM-RU 2087 e) KNM-RU1674 f) KNM-RU 7290 Occlusal views of P africanus g) KNM-SO 372 h) KNM-SO 1112 i) Lingual view of P africanus (KNM-SO 1112) Note the long more horizontally orientedplanum alveolare in Proconsul resulting in a pronounced superior transverse torus The planum alveolare is shorter and more vertical in Ekembo such that one can see the inferiormandibular border from occlusal view
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 53
(eg a dental pick) in themidline internally along the two referencepoints and recording the point in lingual perspective at which therod intersected the alveolar margin (Fig 9) In Ekembo the sub-planum region is more vertically oriented so that the rod intersectsthe alveolar margin most often in the premolar field Not surpris-ingly there is a difference between sexes with females having areduced superior transverse torus and therefore a more verticallyoriented subplanum region compared to males In contrast withEkembo Proconsul mandibles have much more inclined subplanumsurfaces with the rod intersecting the lingual alveolar margin inthe vicinity of the M1 (females) or M2 (males Fig 9) When sex istaken into account therefore the distributions of Ekembo andProconsul in this feature are discrete Note that male and femalemandibles attributed to both P africanus and P major are includedin this assessment and are surprisingly similar despite dramaticsize differences
We also quantified internal symphyseal morphology using threemeasures symphyseal height (infradentaleegnathion) planumalveolare length (posterior-most point on the superior transversetoruseinfradentale) and subplanum length (posterior-most pointon the superior transverse torusegnathion) a combination thatincorporates not only the lengths of these intervals but also theangular relationships among them To compare symphyseal shapeacross a range of sizes we chose a simple principal componentsapproach to partition size onto the first eigenvector (cf Jolicouer1963) While this is not the preferred method for partitioning sizein most datasets (Jungers et al 1995) for this particular sample andset of measurements Mosimann shape variables were still
significantly correlatedwith size (see discussion in Coleman 2008)Hence we assessed differences in symphyseal morphology be-tween Ekembo and Proconsul using the second and third principalcomponents of the three variables Distributions of the two generaare distinct in this shape space (Fig 10 Supplementary OnlineMaterial [SOM] Table 1)
Robusticity of the mandibular corpus also differs between thesesamples though it is more difficult to test due to differentialpreservation among specimens Corpus thickness and depth underP4 can be measured in the greatest number of specimens andcomparison of these measures suggests a difference betweengroups in the scaling of corpus robusticity (Fig 11) Among largepresumably male specimens Ekembo mandibles are substantiallyless robust than those of Proconsul However among smallermandibles only a single Proconsul specimen (KNM-SO 372) could beincluded and it is similar in robusticity to Ekembomandibles of thissize Considered together the impression is that corpus robusticitymay scale differently in Ekembo than it does in Proconsul and a testfor differences in slopes using an ANCOVA design of the loggedvariables was significant (df frac14 2 F frac14 2481 p lt 00001) Never-theless additional specimens are needed to properly evaluate theinfluence of taxonomy sex and size on corpus robusticity areasonable alternative interpretation of Figure 11 is that it dem-onstrates a significant difference between P major and all otherspecies of both Proconsul and Ekembo
Lower incisors Ekembo specimens are uniformly narrower inmesiodistal length than incisors attributed to P africanus P major
Figure 10 Scatter plot of principal component scores based on an eigenanalysis of threeposterior point on the superior transverse toruseinfradentale) and subplanum length (mprimarily captures size differences among specimens (Jolicouer 1963 see text and SOM Trespectively) in the symphyseal variables
Figure 9 Line drawing of a hemimandible depicting different orientations of the sub-planum area in Ekembo and Proconsul Females are denoted using shorter vectors maleswith longer vectors Vector is estimated in M 14086 which is broken at the midlineEkembo specimens have a more vertical subplanum compared to those from Songhorand Legetet but within both groups females have more vertically oriented regions thanmales Hence even the most ventrally rotated subplanum in the largest Ekembo malespecimen (KNM-RU 47805) groups with the Tinderet females in this feature
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6154
and P meswae both absolutely and in relation to crown height(Fig 12) This is true for both I1 and I2 A t-test adjusted for non-independent observations (eg I1 and I2 from the same mandible)confirmed the difference as significant (t-test df frac14 12 t frac14 258p frac14 00242) Moreover while the measurable Proconsul samplewas small this same pattern is observable in other specimenswhose tips were slightly to moderately worn and therefore notmeasured
Lower canines The only lower canine trait that discretely sepa-rates these genera is the pointed crown tip in Ekembo (Fig 3d)contrasted with a blade-like (or ldquoburinrdquo) tip in Proconsul(Fig 3c) This feature is well established for P major but is alsoexpressed in KNM-CA 1772 which was referred to P africanus byKelley (1986) and is too small to be either P major or P meswaeTwo other specimens commonly attributed to P africanus KNM-CA 2149 and KNM-SO 1112 both have broken canine tipsLikewise a new specimen (Kor 65004) attributed to P africanusby Pickford et al (2009) also has a broken tip and cannot beevaluated (see Pickford et al 2009 Fig 23) As with the uppercanines the blade-like tip is also present in the deciduous lowercanines of P major (eg KNM-CA 361 S0 5352) and P meswae(KNM-ME 1)
Lower premolars Like most previous authors (eg Andrews 1978Harrison 2002 2010 Pickford et al 2009) we found neitherqualitative nor quantitative characteristics of the lower premolarswith any taxonomic value
Lower molars Differences in lower molar morphology are few andrelatively subtle but in some cases mirror those seen in the uppermolars Cusps on the lower molars of Ekembo are more bunodontthan those in Proconsul and occupy more of the occlusal surfaceBroad inter-cuspal crests are incorporated into the Ekembo cuspsmaking them appear more inflated Molar cusps of Proconsul are
variables symphyseal height (infradentaleegnathion) planum alveolare length (mostost posterior point on the superior transverse torusegnathion) The first componentable 1) and thus PC 2 and PC 3 illustrate the residual shape variance (72 and 28
Figure 11 Scatter plot showing mandibular corpus robusticity under P4 Trend lines were computed using ordinary least squares (OLS) regression Similar relationships areobservable at other parts of the corpus but differential preservation results in smaller samples at those positions Slopes are significantly different (p lt 00001 tested on loggedvariables)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 55
more distinct from one another and are linked by narrower sharperinter-cuspal crests The lower molars of Ekembo also have asubstantially less developed buccal cingulum than those ofProconsul (Harrison 2002 2010 Harrison and Andrews 2009)
Figure 12 Scatter plot of lower incisor crown proportions Differenc
Lastly and again as seen in the upper molars the two genera canbe distinguished by the significantly greater amount of lowermolar flare in Proconsul (Fig 13 t-test df frac14 59 t frac14 713p lt 00001 see also Harrison and Andrews 2009)
es between Ekembo and Proconsul are significant (p frac14 00242)
Figure 13 Box-and-whisker plot of buccolingual flare in lower molars measured asthe average distance between the mesial buccal and lingual cusps divided by overallbuccolingual crown breadth Differences between Ekembo and Proconsul are highlysignificant (p lt 00001)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6156
Relative molar size has been used to distinguish species ofProconsul but we observed the primary difference to be in relativebuccolingual breadths of M1 and M2 In Ekembo M1 is substantiallynarrower than M2 than is the case in Proconsul (Fig 14 Harrison2002 2010) Although none of the smaller Proconsul mandiblespreserve complete crowns for both M1 and M2 KNM-SO 1112 hasan intact M2 and at least preserves the cervix for M1 However evenif the cervical measurement is used for its M1 breadthdthusunderestimating the true crown breadthdKNM-SO 1112 still has abroader first molar in relation to M2 than would be expected inEkembo If crown breadth is estimated from the ratio of cervixbreadth to maximum crown breadth in other Proconsul fossils therange of likely breadths for the SO 1112 M1 clearly distinguishes it
from the Ekembo sample (Fig 14) Testing this relationship onlogged variables using the mean estimate for the SO 1112 M1(Fig 14) reveals a significant difference between the two groups(ANCOVA df frac14 1 F frac14 2424 p frac14 00002)
As with upper third molars we find a great deal of variation inM3 morphology and mistrust the taxonomic significance some-times placed on characteristics of this toothdat least with regard tothese taxa Harrison (2002 2010) cited the mesiodistal position ofthe M3 entoconid as differentiating Kisingiri third molars fromthose attributed to P africanus We concur with his assessment butfound the entoconid position to vary in specimens attributed toP major Thus this character does not differentiate Ekembo from allProconsul species but may distinguish it from the type speciesP africanus
Discussion
The distinctiveness of the Kisingiri large catarrhine materialfrom that at Tinderet and Ugandan localities has been noted pre-viously by several authors (Pickford 1986 Walker et al 1993Begun 2001 2007 MacLatchy and Rossie 2005 Harrison andAndrews 2009 Harrison 2010) What had not been fully docu-mented was the nearly complete lack of overlap between thesesamples in a number of features despite very large assemblagesfrom Rusinga and Mfangano On descriptive criteria alone (Table 2)many isolated teeth (I1 C1 P3 M2 C1) and all of the mandibles canbe assigned to either Proconsul or Ekembowith very little chance ofmisattribution That some of the most diagnostic features occur inthe upper dentitiondthe only anatomy preserved in theP africanus type specimendadds weight to the taxonomicarrangement proposed here The morphology of the upper teeth inall Proconsul species compellingly unites them into a single cladedistinct from E heseloni and E nyanzae This is reinforced bynumerous features of the mandible and mandibular dentition thatfurther distinguish these samples The number and variety of thesefeatures are such that Proconsul and Ekembo are more differentfrom one another than are many closely-related modern catarrhinegenera
Derived morphology of Ekembo and Proconsul
It is premature to formally assign primitive and derived status tothe individual features of Ekembo and Proconsul before completinga formal phylogenetic analysis This is especially true given theabundance of evidence available for Ekembo and hence the poten-tial impact that our generic revision could have on phylogeneticinterpretations Nevertheless in comparison with both extanthominoids and the small-bodied catarrhines from the earlyMiocene it is informative to differentiate the evolutionary grades ofthese genera Many characteristics of Ekembo are more advanced inthe direction of modern apes compared to those of Proconsul(Table 2) General trends such as expanded and more bunodontcusps broader molar crests reduced cingulum and morphologicalsimilarity between P3 and P4 all represent morphologies thatpresage those of later fossil and extant hominoids The moreadvanced morphologies of P3 and M2 are particularly conspicuouscompared not only with those of Proconsul but to most earlyMiocene catarrhines In contrast the mesially positioned stronglyheteromorphic cusps on the P3 of Proconsul plus the correspondingasymmetric buccal profile notched mesial margin and missingmesial fovea are characteristic of other early Miocene catarrhinessuch as Dendropithecus Rangwapithecus and Limnopithecus andare likely primitive among East African catarrhines The moresymmetric P3 of Ekembo with midline positioned cusps andreduced cusp heteromorphy is reminiscent of later hominoids and
Figure 14 Relative buccolingual dimensions of M1 and M2 KNM-SO 1112 has a preserved M2 but only the cervix of M1 Its M1 breadth was estimated based on the ratio to cervixbreadth to maximum crown breadth in other Tinderet fossils Even the cervical breadth of SO 1112 is broader than would be predicted for the crown breadth of a Kisingiri M1 Trendlines computed using OLS regression and using the average estimate of SO 1112 M1 breadth Group differences were significant (p frac14 00002)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 57
even hominid taxa such as Rudapithecus (eg Kordos and Begun2001)
Proconsul is likewise united by a suite of features that are for themost part characteristic only of this lineage (Table 2) Themorphology of both upper and lower canines is unique to thisgenus and shared by all three well-known species plus the poorlyrepresented middle Miocene P gitongai (Pickford and Kunimatsu2005) The mandibular symphysis is also distinctive in Proconsulcompared with other catarrhines with its very long and shallowlysloping planum alveolare and steeply inclined subplanum surfaceIndeed even the small edentulous anterior mandibular fragmentKNM-SO 372 is readily placed within Proconsul and excluded fromother similarly sized catarrhine taxa such as Rangwapithecus andEkembo based on this unique symphyseal morphology (Figs 8 and9)
Other differences between Ekembo and Proconsul are of uncer-tain evolutionary polarity (eg orientation of the posthypoconecrista) or may be of taxonomic relevance without broader phylo-genetic significance (eg degree of molar flare Table 2) Werecognize that there are morphological differences within Procon-sul particularly between the better known species P africanus andP major However many of these differences are likely related tosize and represent variations on a single generic theme that aremarkedly different from the morphology of Ekembo The recentdescription (Harrison and Andrews 2009) of the intermediatelysized P meswae underscores the broad morphological unity withinProconsul and further substantiates a well-defined Proconsul cladecomprising Tinderet and Ugandan species
Based on the abundant morphological and morphometric evi-dence supporting genus-level distinction between Proconsul andEkembo we are currently conducting a more detailed phylogeneticanalysis of all early Miocene catarrhines in order to better under-stand the distribution and evolutionary significance of these andother morphological features We anticipate that the debates sur-rounding the phylogenetic relationships of an historically consti-tuted sample of Proconsul can be more satisfactorily resolved nowthat the Kisingiri material is appropriately recognized as a distinctgenus
The generic disposition of ldquoUgandapithecusrdquo species
A great many features said to characterize ldquoUgandapithecusrdquowere added to the original diagnosis of the genus by Pickford et al(2009) in their systematic revision of Proconsul (sensu lato) As partof that study they also erected a new species ldquoUrdquo legetetensis andtransferred P meswae to ldquoUgandapithecusrdquo Evaluating the same setof features across the entire available sample however we wereunable to find support for this more expansive conception ofldquoUgandapithecusrdquo for several reasons First some diagnostic fea-tures cited in Pickford et al (2009) were quite variable within theldquoUgandapithecusrdquo sample As an example marked reduction of theM3 entoconid to a bead on the lingual cingulum is not shared byseveral ldquoUrdquo major specimens such as KNM-CA 393 NAP I 49000and Kor 253004 (see Pickford et al 2009 Fig 33e) and was likelynot a feature of a few of the more worn specimens eg BUMP 600and BUMP 601 or even the holotype of P major M 16648 Second
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6158
some diagnostic features of ldquoUgandapithecusrdquo were found to bevariably present in Ekembo arguing against their utility for dis-tinguishing ldquoUgandapithecusrdquo from Pickford et als (2009) com-bined P africanus thorn Kisingiri conception of Proconsul For examplesignificantly reduced curvature of the I1 distal margin in ldquoUganda-pithecusrdquo is also found in some Kisingiri specimens including KNM-RU 1685 1712 1714 1831 1846 and 1951 several more Rusingaspecimens have moderately reduced curvature Third some fea-tures that purportedly distinguish ldquoUgandapithecusrdquo fromP africanus are likely related to size For example the somewhatmore central position of the P3 paracone in ldquoUrdquomajor is not seen inthe smaller ldquoUrdquo meswae whose paracone is positioned similarly tothat of the holotype of P africanus (see Harrison and Andrews2009 Fig 6d)
However the major part of the evidence for recognizingldquoUgandapithecusrdquo (sensu Pickford et al 2009) was assembled by re-assigning specimens traditionally attributed to P africanus into newspecies of ldquoUgandapithecusrdquo The P africanus holotype was left inassociation with the Rusinga sample despite having morphologyoverwhelmingly characteristic of other Tinderet specimens Theresulting specimen allocations left P africanus morphologicallyimpoverisheddstripped of much of its hypodigmdso that Pro-consul for Pickford et al (2009) came to be characterized primarilyby the Rusinga sample (see similar comments in Harrison andAndrews 2009) Within that conceptual framework any spec-imen that differs from those at Rusinga could not be Proconsul butmust instead belong to ldquoUgandapithecusrdquo Thus specimens such asthe KNM-SO 1112 mandible were erroneously assigned to ldquoUgan-dapithecusrdquo because they were (correctly) perceived as differingfrom their Rusinga counterparts (Pickford et al 2009)
Here we restore the P africanus hypodigm and associate it withthe other Tinderet fossils according to their shared morphologySpecimens assigned to ldquoUgandapithecusrdquo revert to Proconsul andmany of the features cited to support a speciose ldquoUgandapithecusrdquo(Pickford et al 2009) instead become diagnostic features of Pro-consul This correction serves to highlight the distinctiveness of theKisingiri remains and the need to recognize this distinctiveness atthe generic level
As demonstrated above specimens from Kisingiri do not displaythe morphology of Proconsul as represented by the holotype ofP africanus M 14084 Given that this specimen preserves only theupper canine and cheek teeth similarities with or departures fromthis morphology represent the most compelling arguments forassessing inclusivity in the genus However for the mandible andlower dentition if one simply sorts the specimens by elementwithout relying on taxonomic preconceptions the overwhelmingdistinction within the traditionally constituted Proconsul sample isbetween Kisingiri material on the one hand and that from Tinderetand Uganda on the other (cf Walker et al 1993 Begun 2001 2007MacLatchy and Rossie 2005 Harrison and Andrews 2009Harrison 2010) Additionally with the Kisingiri specimensremoved to Ekembo the postcranial differences used to partlyjustify a distinct ldquoUgandapithecusrdquo lose their relevance (seeHarrison and Andrews 2009)
We have differed among ourselves in our past treatment ofP major as generically distinct from the smaller species of Pro-consul (Begun 2007 McNulty 2010) Morphologically many ofthe unique features of P major can be interpreted as size-relatedvariants on the smaller P africanus morphology Even if so thequestion is whether these are sufficiently different and numerousto warrant generic distinction that is do they suggest thatP major belongs in a separate clade from P africanus andP meswae As we have demonstrated differences among thesethree species are certainly fewer and show less morphologicaldistinction than those between Ekembo and any of the three
Nevertheless while we ultimately agree with Harrison andAndrews (2009 see also Begun 2007) that the features dis-tinguishing ldquoUrdquo major from P africanus are consistent with thosethat distinguish congeneric species and are at least partly due todifferences in size we concede the possibility that the largestProconsul species could belong in a separate genus (Senut et al2000 Pickford and Kunimatsu 2005) However whether or notone recognizes a separate genus for P major the P africanus ma-terial does not sort morphologically with the Kisingiri materialTherefore the taxonomic alternatives are either two generaProconsul and Ekembo or three Proconsul Ugandapithecus andEkembo with Proconsul and Ugandapithecus united in a cladeexclusive of Ekembo Thus while we strongly favor the firstalternative recognizing Ugandapithecus does not change thefundamental argument concerning the need to refer the Kisingirispecies to a generically and cladistically distinct taxon from thespecies represented at Tinderet and the Ugandan sites
Specimens of interest
Despite the substantial dental and gnathic differences betweenProconsul and Ekembo there are a few specimens whose taxonomicaffinities require additional scrutiny
KNM-RU 1676-77 This set of associated upper and lower teeth is aconsistent outlier among E heseloni and E nyanzae specimens Insome ways this specimen more closely resembles Tinderet Pro-consul in the features that distinguish it from Ekembo Molar cuspmorphology and cingulum development I1 lingual tubercledevelopment and M2 shape and posthypocone crista orientationare more consistent with specimens from the Tinderet sites On theother hand it otherwise resembles Ekembo and lacks the strongestdiagnostic features that unite species of Proconsul such as thedistinctive canine and P3 morphology Overall this specimen doesnot fit unambiguously into either genus in fact the degree ofbuccolingual flare cusp morphology enamel wrinkling and espe-cially the long extension of enamel onto the anterior root in P3 arein some ways reminiscent of Afropithecus although it clearly differsfrom that genus as well (cf Rossie and MacLatchy 2013)
Were these characteristics individually distributed among thelarge Kisingiri sample one might attribute this to intra-genericvariation but when found together in a single specimen from alarge sample it suggests a unique individual One of us (KPM)thinks it likely that the specimen was not actually found at aKisingiri locality based on its unique preservation compared toother Rusinga fossils and on other examples of mis-proveniencedspecimens (Leakey 1967 Andrews and Molleson 1979 Pickford1986 Kelley 1986) However we recognize that relying toofrequently on this explanation allows almost any scenario to bejustified and suggest some alternative possibilities One is that thespecimen was found on Rusinga but in the much older WayandoFormation strata instead of the Hiwegi Formation The differencesin morphology in that case might reflect more than a million yearsof temporal change (McCollum et al 2013) with KNM-RU 1676-77preserving certain more primitive Proconsul-like features As analternative it is reasonable to assume that fossil assemblages asvast as those from Rusingawill include not only abundant taxa butalso rare or infrequent inhabitants of the local biome There aremany examples of this from Rusingas mammalian communityeg ldquoHemicyonrdquo Kulutherium Tadarida cf Hyainailouros cfArchaeobeledon Kelba (see relevant chapters in Werdelin andSanders 2010) As such KNM-RU 1676-77 may simply representa rare and well-preserved specimen of some other catarrhinetaxon that lived among or visited a primate community dominatedby Ekembo
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 59
KNM-SO 1112 This specimen was initially referred to Rangwapi-thecus but soon after was transferred to Proconsul afrianus (Bosler1981 Martin 1981)da designation that most subsequent authorshave followed (Pickford 1986 Nengo and Rae 1992 Kelley 19861995 Walker et al 1993 MacLatchy and Rossie 2005 Hill et al2013) Pickford et al (2009) more recently referred this specimento ldquoUrdquo meswae citing two primary reasons First they reiteratedBoslers (1981) concern that its P3 differs from others attributedto P africanus However at that time the majority of what wasconsidered to be P africanus would have been from RusingaIsland In fact while noting its contrasting morphology Boslersuggested that differences between KNM-SO 1112 and other P3smight reasonably be attributed to sexual dimorphism notingthat this specimen provides ldquoa not implausible match for the Pafricanus holotype maxilla M 14084rdquo (1981152) Thatassessment is borne out in this study Using the much largersample of Ekembo P3s as a model variation can be seen toencompass specimens with well-developed mesial beaks andthose with more bilaterally compressed crowns and parallelsides The latter morphology is predominantly found amongmale specimens (KNM-RU 1674 1924 and KPS I and III) thussupporting Boslers (1981) suggestion that this variation isrelated to sexual dimorphism and hence that KNM-SO 1112 is amale P africanus
The second feature cited by Pickford et al (2009) in referringthis specimen to ldquoUrdquo meswae was posterior shallowing of themandibular corpus However because they redefined ldquoUgandapi-thecusrdquo to include all of the well-preserved Tinderet mandibles(Pickford et al 2009)dleaving Proconsul to be represented only byRusinga specimensdtheir only alternative within this scheme for aspecimen the size of SO 1112 was to refer it to ldquoUrdquomeswae It seemsodd though to assign this specimen to a species from a (perhapsmuch) older site when the Proconsul africanus holotype is ofappropriate size and similar age and as there are no other adultmandibles from either species with which to compare it In anycase mandibular shallowing is variable in both Ekembo and Pro-consul with some Kisingiri specimens exhibiting considerableshallowing (eg KNM-RU 2087) and some ldquoUgandapithecusrdquo spec-imens having very little (eg M 14086)
We therefore see no morphological grounds to assign KNM-SO1112 to P meswae but do find some morphological evidence toassign it as a male P africanus which occurs at Songhor
KNM-LG 452 This very complete mandible was originally recog-nized as a female of P major based on the size of the dentition androbusticity of the mandible (Martin 1981) Bosler (1981) howeverleft this specimen unassigned noting similarities to P major in P3-M1 size but having distinctly smaller M2e3 In this she likenedKNM-LG 452 to a few other specimens (notably M 14086) thatare smaller in their distal molars in a manner more consistentwith taxonomic rather than sex differences (Bosler 1981 see alsoPilbeam 1969) Kelley (1986 1995) also assigned this specimen asa female of P major based on canine size and morphology butnoted that the canine lacked the shallowly-sloping mesial ridgetypical of most female anthropoid canines This is also truehowever of two other canines assigned as female P major KO 9and SO 373 (Kelley 1986) and so might simply be an unusualcharacteristic of P major female lower canines In the end Kelley(1995) concluded that considering all of the criteria fordistinguishing male and female canines the evidence that KNM-LG 452 is female remains questionable
Pickford et al (2009) further developed the idea that two spe-cies are represented within the P major sample and made KNM-LG452 the holotype of a new species ldquoUrdquo legetetensis We concur thatthis specimen is unique among the well-preserved Tinderet
mandibles in some important features Whereas male and femalemandibles of P major have long internal symphyseal planesexpansive extra- and retromolar sulci and rami that cross theocclusal plane at distal M3 KNM-LG 452 has a shorter symphysealplane (though not to the extent seen in Ekembo) a moderateextramolar sulcus but no retromolar sulcus and a ramus thatcrosses the occlusal plane more mesially If KNM-SO 396 and SO404 represent males and females respectively of P major then themorphology of KNM-LG 452 appears distinct
At present however we do not think there is sufficient evidenceto differentiate this specimen from either P major or P meswaePickford et al (2009) argue that KNM-LG 452 cannot belong to thelatter species because of its larger size The size differences how-ever are well within the range of variation seen in P majorE heseloni and E nyanzae Moreover whereas the Legetet mandibleis larger than most of the P meswae hypodigm an adult P4 referredto this species (KNM-ME 25) substantially exceeds its counterpartin KNM-LG 452 Should the Legetetmandible ultimately prove to bea male specimen size alone would not rule out its attribution toP meswae On the other hand if it is female then it becomes harderto justify its inclusion in that sample of mostly smaller specimens Iffemale therefore it could represent a separate species for whichthe binomial Proconsul legetetensis comb nov is available Recentlycollected fossils from the SonghorKapurtay area provide someadditional support for the possibility that three species of Proconsulexisted at these localities (McNulty 2014ab) However toconvincingly demonstrate this and determine whether a thirdspecies can be differentiated from P meswae will require a moredetailed analysis of variation among Proconsul species a corre-sponding study of variation in modern taxa anddalmost certain-lydadditional fossil specimens For now we regard ldquoUrdquo legetetensisas a subjective junior synonym of P major
Conclusions
The genus Proconsul has been central to studies of ape and hu-man evolution since its first description by Hopwood (1933) As oneof the best known fossil catarrhines represented by nearly everyskeletal element our knowledge of Proconsuls functional anatomypositional behavior and overall morphology has made it thefoundation for paleoanthropological research in the early Mioceneas well as studies on the origin and early diversification of homi-noids With respect to systematics however Proconsul is less wellunderstood the type species P africanus is known from a rela-tively small number of specimens from sites in the Tinderet areaand no postcranial bones can be unambiguously assigned to itRather it is the copious and well-preserved specimens from theKisingiri sites of Rusinga and Mfangano Islands that have formedthe basis for the common image of Proconsul
We argue here that the Kisingiri species traditionally referredto ProconsuldP heseloni and P nyanzaedare generically distinctfrom P africanus and hereby transfer them into a new genusEkembo based on a substantial number of morphological differ-ences (Table 2) Considering only those traits preserved in theP africanus holotype the entire sample from Rusinga and Mfan-gano differs markedly in the morphology of the upper canine P3P4 and M2 Additional differences in the I1 lower incisors andcanines molar cusp and occlusal crest morphology degree of flarein all molars and especially mandibular features clearly distin-guish the Kisingiri from non-Kisingiri samples in a way that isincompatible with congeneric species With the transfer ofE heseloni and E nyanzae Proconsul becomes a well-defined cladecomprising at least three speciesdP africanus P major andP meswaedunited by several features that are unique for thegenus (Table 2)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6160
We found no support in this study for the genus ldquoUgandapi-thecusrdquo as constructed by Pickford et al (2009) In that paper theytransferred the vast majority of Proconsul fossils from the Tinderetand Napak localities into ldquoUgandapithecusrdquo leaving P africanusrepresented only by its holotype and a small number of isolatedteeth and aligning it with the material from Rusinga and Mfan-ganoWhen the P africanus hypodigm is reconstituted and properlyaffiliated with the other Tinderet species according to itsmorphology much of the evidence marshaled by Pickford et al(2009) for ldquoUgandapithecusrdquo actually supports the distinction be-tween the Tinderet and Kisingiri samples ie the generic distinc-tion of Ekembo In addition some of the traits enumerated byPickford and colleagues in support of ldquoUgandapithecusrdquo wererevealed in our analysis to be variable within one or more speciesand therefore of questionable taxonomic utility We do not rejectout of hand the original conception of ldquoUgandapithecusrdquo (Senutet al 2000) as either a monotypic genus distinguishing ldquoUrdquo ma-jor or perhaps including a closely allied middle Miocene ldquoUrdquogitongai as well (Pickford and Kunimatsu 2005) but we have fornow retained the early Miocene specimens of both in Proconsulpending further study However even if Ugandapithecus is a taxo-nomically valid genus comprising just the latter two species theirmorphology indicates that theywould still be united in a cladewithProconsul and they lack several more advanced features that partlydefine Ekembo Better understanding of the role that body size playsinmediating differences within Proconsulwill help to further clarifythe relationships among these species
Despite the existence of one well-represented and several lesscomplete skeletons of what was long assumed to be Proconsul thephylogenetic status of this genus has defied consensus This islargely due in our view to Proconsul having been an admixture oftwo distinct genera one more advanced in the direction of extantapes than the other As we demonstrate here the material fromRusinga and Mfangano traditionally referred to Proconsul andwhich includes all of the more complete skeletal material isdistinct from the Tinderet and Napak material and belongs in aseparate genus Ekembo By disentangling these distinct clades weanticipate being able to achieve better phylogenetic resolution forboth genera thereby improving our knowledge of early hominoidevolution and diversification
Acknowledgments
We thank many people and agencies that assisted with thedevelopment and completion of this project This research wasinitiated as a result of funding from the Wenner-Gren Foundationand the Johns Hopkins School of Medicine to DR Begun in1987e1989 and was renewed following research and travelgenerously funded by multiple Leakey Foundation grants to KMcNulty National Science Foundation grants to K McNulty(0852609 1241807) the McKnight Foundation the University ofMinnesota the Lloyd A Wilford Endowment the LeverhulmeFoundation and the Turkana Basin Institute J Kelley gratefullyacknowledges support from the Institute of Human Origins Ari-zona State University DR Begun also wishes to acknowledge theAlexander Humboldt Stiftung and NSERC for funding support Weare grateful to the National Museums of Kenya the UgandaMuseum the Natural History Museum and to Rose Mwanja EzraMusiime Tom Mukhuyu Samuel Muteti James Yatich and MaryMuungu for access to and assistance with specimens in their careMany people provided assistance comments criticisms insightsand suggestions on various aspects of this work and we especiallythank Holly Dunsworth Will Harcourt-Smith Tom Lehmann DanPeppe Nicole Garrett Kirsten Jenkins Rutger Jansma Susy CoteIsaiah Nengo Andrew Hill and Brenda Benefit K McNulty
acknowledges the evolutionary anthropology faculty and studentsat Durham University for their helpful comments on an earlyversion of this manuscript and for their gracious hospitality Weare likewise grateful to the anonymous reviewers associate editorand editor whose comments strengthened the paper This manu-script is publication 1 supporting Research on East AfricanCatarrhine and Hominoid Evolution (REACHE)
Appendix A Supplementary data
Supplementary data related to this article can be found at httpdxdoiorg101016jjhevol201503009
References
Andrews P 1974 New species of Dryopithecus from Kenya Nature 249 188e190Andrews P 1978 A revision of the Miocene Hominoidea of East Africa Bull Brit
Mus (Nat Hist) Geol 30 85e225Andrews P 1985 Family group systematics and evolution among catarrhine pri-
mates In Delson E (Ed) Ancestors The Hard Evidence Liss New Yorkpp 14e22
Andrews P Harrison T Martin L Pickford M 1981 Hominoid primates from anew Miocene locality named Meswa Bridge in Kenya J Hum Evol 10123e128
Andrews P Begun D Zylstra M 1997 Interrelationships between functionalmorphology and paleoenvironments in Miocene hominoids In Begun DWard C Rose M (Eds) Function Phylogeny and Fossils Miocene HominoidEvolution and Adaptations Plenum Press New York pp 29e58
Andrews P Molleson T 1979 The provenance of Sivapithecus africanus Bull BritMus (Nat Hist) Geol 32 19e23
Begun DR 2001 African and Eurasian Miocene hominoids and the origins of theHominidae In de Bonis L Koufos GD Andrews P (Eds) Hominoid Evolutionand Climatic Change in Europe Phylogeny of the Neogene Hominoid Primatesof Eurasia Cambridge University Press London pp 231e253
Begun DR 2007 Fossil record of Miocene hominoids In Henke W Tattersall I(Eds) Handbook of Paleoanthropology Springer Berlin pp 921e977
Begun DR Teaford MF Walker AC 1994 Comparative and functional anatomyof Proconsul phalanges from the Kaswanga primate site Rusinga Island KenyaJ Hum Evol 26 89e165
Bishop WW 1964 More fossil primates and other Miocene mammals from North-East Uganda Nature 203 1327e1331
Bishop WW Miller JA Fitch FJ 1969 New potassium-argon age determinationsrelevant to the Miocene fossil mammal sequence in East Africa Am J Sci 267669e699
Blankenship JA Mincer HH Anderson KM Woods MA Burton EL 2007Third molar development in the estimation of chronologic age in Americanblacks as compared with whites J Forensic Sci 52 428e433
Bosler W 1981 Species groupings of early Miocene dryopithecine teeth from EastAfrica J Hum Evol 10 151e158
Coleman MN 2008 What does geometric mean mean geometrically Assessingthe utility of geometric mean and other size variables in studies of skullallometry Am J Phys Anthropol 135 404e415
Cote S Malit N Nengo I 2013 Additional mandibles of Rangwapithecus gordonian early Miocene catarrhine from the Tinderet localities of Western KenyaJ Hum Evol 153 341e352
Deane AS 2009 Early Miocene catarrhine dietary behavior the influence of theRed Queen Effect on incisor shape and curvature J Hum Evol 56 275e285
Drake R Van Couvering JA Pickford M Curtis G Harris JA 1988 New chro-nology for the Early Miocene mammalian faunas of Kisingiri Western KenyaJ Geol Soc Lond 145 479e491
Evernden JF Savage DE Curtis GH James GT 1964 Potassium-argon datesand the Cenozoic mammalian chronology of North America Am J Sci 262145e198
Gebo DL MacLatchy L Kityo R Deino A Kingston J Pilbeam D 1997A hominoid genus from the Early Miocene of Uganda Science 276 401e404
Geoffroy Saint-Hilaire E 1812 Tableau des quadrumanes 1 Ord QuadrumanesAnnls Mus Hist Nat Paris 19 85e122
Gray JE 1825 Outline of an attempt at the disposition of the Mammalia into tribesand families with a list of the genera apparently appertaining to each tribeAnnls Philos 10 337e344
Harrison T 1982 Small-bodied apes from the Miocene of East Africa PhDDissertation University of London
Harrison T 1986 New fossil anthropoids from the middle Miocene of East Africaand their bearing on the origin of the Oreopithecidae Am J Phys Anthropol 71265e284
Harrison T 1987 The phylogenetic relationships of the early catarrhine primates areview of the current evidence J Hum Evol 16 41e80
Harrison T 2002 Late Oligocene to middle Miocene catarrhines from Afro-ArabiaIn Hartwig WC (Ed) The Primate Fossil Record Cambridge University PressCambridge pp 311e338
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 61
Harrison T 2010 Dendropithecoidea Proconsuloidea and Hominoidea (CatarrhiniPrimates) In Werdelin L Sanders WJ (Eds) Cenozoic Mammals of AfricaUniversity of California Press Berkeley pp 429e469
Harrison T Andrews P 2009 The anatomy and systematic position of a newspecies of early Miocene proconsulid from Meswa Bridge Kenya J Hum Evol56 479e496
Hill A Nengo IO Rossie JB 2013 A Rangwapithecus gordoni mandible from theearly Miocene site of Songhor Kenya J Hum Evol 65 490e500
Hopwood AT 1933 Miocene primates from Kenya J Linn Soc Zool 38 437e464Ishida H Kunimatsu Y Takano T Nakano Y Nakatsukasa M 2004 Nacholapi-
thecus skeleton from the Middle Miocene of Kenya J Hum Evol 46 69e103Jolicouer P 1963 The multivariate generalization of the allometry equation Bio-
metrics 19 497e499Jungers WL Falsetti AB Wall CE 1995 Shape relative size and size-
adjustments in morphometrics Am J Phys Anthropol 38 (S2) 137e161Kavanagh KD Evans AR Jernvall J 2007 Predicting evolutionary patterns of
mammalian teeth from development Nature 449 427e433Kelley J 1986 Species recognition and sexual dimorphism in Proconsul and
Rangwapithecus J Hum Evol 15 461e495Kelley J 1995 Sex determination in Miocene catarrhine primates Am J Phys
Anthropol 96 390e417Kelley J 1993 Taxonomic implications of sexual dimorphism in Lufengpithecus In
Kimbel WH Martin LB (Eds) Species Species Concepts and Primate Evo-lution Springer New York pp 429e458
Kent PE 1942 The country round the Kavirondo Gulf of Victoria Nyanza Geogr J100 22e31
Kordos L Begun DR 2001 A new cranium of Dryopithecus from RudabanyaHungary J Hum Evol 15 689e700
Le Gros Clark WE Leakey LSB 1950 Diagnoses of East African Miocene Homi-noidea Q J Geol Soc Lond 105 260e263
Le Gros Clark WE Leakey LSB 1951 The Miocene Hominoidea of East Africa InFossil Mammals of Africa No 1 British Museum (Natural History) London
Leakey LSB 1943 A Miocene anthropoid mandible from Rusinga Kenya Nature152 319e320
Leakey LSB 1963 East African fossil Hominoidea and the classification within thissuper-family In Washburn SL (Ed) Classification and Human EvolutionAldine Chicago pp 32e49
Leakey LSB 1967 Notes on the mammalian faunas from the Miocene and Pleis-tocene of East Africa In Bishop WW Clark JD (Eds) Background to Evolu-tion in Africa University of Chicago Press Chicago pp 7e29
Leakey RE Leakey MG 1986 A new Miocene hominoid from Kenya Nature 324143e146
Leakey RE Leakey MG Walker AC 1988 Morphology of Afropithecus turka-nensis from Kenya Am J Phys Anthropol 76 289e307
Leakey MG Ungar PS Walker A 1995 A new genus of large primate from theLate Oligocene of Lothidok Turkana District Kenya J Hum Evol 28519e531
Linnaeus C 1758 Systema naturae per regna tria naturae secundum classesordines genera species cum characteribus differentris synonymis locis tenthed Laurentii Salvii Stockholm
MacInnes DG 1943 Notes on the East African Miocene primates J E Afr UgandaNat Hist Soc 17 141e181
MacLatchy L 2004 The oldest ape Evol Anthropol 13 90e103MacLatchy L Bossert WH 1996 An analysis of the articular surface distribution of
the femoral head and acetabulum in anthropoids with implications for hipfunction in Miocene hominoids J Hum Evol 31 425e453
MacLatchy L Rossie JB 2005 The Napak hominoid still Proconsul major InLieberman DE Smith RJ Kelley J (Eds) Interpreting the Past Essays onHuman Primate and Mammal Evolution in Honor of David Pilbeam Brill Ac-ademic Publishers Boston pp 15e28
Martin L 1981 New specimens of Proconsul from Koru Kenya J Hum Evol 10139e150
McCollum MS Peppe DJ McNulty KP Dunsworth HM Harcourt-Smith WEH Andrews AL 2013 Magnetostratigraphy of the early MioceneHiwegi Formation (Rusinga Island Lake Victoria Kenya) Geological Society ofAmerica Abstracts with Programs 45 p 12
McNulty KP 2010 Apes and tricksters the evolution and diversification ofhumans closest relatives Evol Edu Outreach 3 322e332
McNulty KP Harcourt-Smith WEH Dunsworth HM 2007 New primate fossilsfrom Rusinga Island Kenya Am J Phys Anthropol 132 (S44) 170
McNulty KP MacLatchy L Peppe DJ Nengo I Manthi FK Miller ERStevens NJ Cote S Lehmann T Kingston J 2014a A regional approach toEast African early Miocene paleobiology J Vert Paleontol 185 SVP Programand Abstracts Book 2014
McNulty KP MacLatchy L Rossie JB Peppe DJ Deino AL Mbua ENManthi FK Nengo IO Stevens NJ Cote S Lehmann T Gutierrez M 2014b
Research on East African catarrhine and hominoid evolution results from thefirst year Am J Phys Anthropol 153 (S58) 182
Michel LA Peppe DJ Lutz JA Driese SG Dunsworth HM Harcourt-Smith WEH Horner W Lehmann T Nightengale S McNulty KP 2014Remnants of an ancient forest provide ecological context for Early Miocenefossil apes Nature Comm 5 3236
Myres JL 1932 Africa Man 32 208Nakatsukasa M Kunimatsu Y Nakano Y Takano T Ishida H 2007 Vertebral
morphology of Nacholapithecus kerioi based on KNM-BG 35250 J Hum Evol52 347e369
Nengo IO Rae TC 1992 New hominoid fossils from the early Miocene site ofSonghor Kenya J Hum Evol 23 423e429
Peppe DJ McNulty KP Cote SM Harcourt-Smith WEH Dunsworth HM VanCouvering JA 2009 Stratigraphic interpretation of the Kulu Formation (EarlyMiocene Rusinga Island Kenya) and its implications for primate evolutionJ Hum Evol 56 447e461
Pickford M 1981 Preliminary Miocene mammalian biostratigraphy for westernKenya J Hum Evol 10 73e97
Pickford M 1986 Sexual dimorphism in Proconsul Hum Evol 1 111e148Pickford M Andrews PJ 1981 The Tinderet Miocene sequence in Kenya J Hum
Evol 10 11e33Pickford M Kunimatsu Y 2005 Catarrhines from the Middle Miocene (ca 145
Ma) of Kipsaraman Tugen Hills Kenya Anthropol Sci 113 189e224Pickford M Senut B Gommery D Musiime E 2009 Distinctiveness of Ugan-
dapithecus from Proconsul Estudios Geol 65 183e241Pilbeam DR 1969 Tertiary Pongidae of East Africa evolutionary relationships and
taxonomy Bulletin of the Peabody Museum of Natural History 31 pp 1e185Rae TC 1993 Phylogenetic analysis of proconsulid facial morphology PhD
Dissertation State University of New York at Stony BrookRae TC 1999 Mosaic evolution in the origin of the Hominoidea Folia Primatol 70
125e135Rafferty KL Walker A Ruff C Rose MD Andrews PJ 1995 Postcranial esti-
mates of body weights in Proconsul with a note on a distal tibia of P major fromNapak Uganda Am J Phys Anthropol 97 391e402
Rossie JB MacLatchy L 2013 Dentognathic remains of an Afropithecus individualfrom Kalodirr Kenya J Hum Evol 65 199e208
Senut B Pickford M Gommery D Kunimatsu Y 2000 Un nouveau genredrsquohominoiumlde du Miocene inferieur drsquoAfrique orientale Ugandapithecus major(Le Gros Clark and Leakey 1950) CR Acad Sci Paris 331 227e233
Simons EL Pilbeam DR 1965 Preliminary revision of the Dryopithecinae (Pon-gidae Anthropoidea) Folia Primatol 3 81e152
Smith TM Martin LB Leakey MG 2003 Enamel thickness microstructure anddevelopment in Afropithecus turkanensis J Hum Evol 44 283e306
Stevens NJ Seiffert ER OConnor PM Roberts EM Schmitz MD Krause CGorscak E Ngasala S Hieronymus TL Temu J 2013 Palaeontological evi-dence for an Oligocene divergence between Old World monkeys and apesNature 497 611e614
Suwa G Kono RT Katoh S Asfaw B Beyene Y 2007 A new species of great apefrom the late Miocene epoch in Ethiopia Nature 448 921e924
Szalay FS Delson E 1979 Evolutionary History of the Primates Academic PressNew York
Teaford MF Beard KC Leakey RE Walker A 1988 New hominoid facial skel-eton from the Early Miocene of Rusinga Island Kenya and its bearing on therelationship between Proconsul nyanzae and Proconsul africanus J Hum Evol17 461e477
Van Couvering JAH Miller J 1969 Miocene stratigraphy and age determinationsRusinga Island Kenya Nature 221 628e632
Walker A 1997 Proconsul function and phylogeny In Begun DR Ward CVRose MD (Eds) Function Phylogeny and Fossils Miocene Hominoid Evolu-tion and Adaptation Plenum Press New York pp 209e224
Walker A Pickford M 1983 New postcranial fossils of Proconsul africanus andProconsul nyanzae In Ciochon RL Corruccini RS (Eds) New Interpretationsof Ape and Human Ancestry Plenum Press New York pp 325e351
Walker A Teaford M 1988 The Kaswanga primate site an Early Miocene hom-inoid site on Rusinga Island Kenya J Hum Evol 17 539e544
Walker A Teaford M 1989 The hunt for Proconsul Sci Am 260 76e82Walker A Teaford MF Martin L Andrews P 1993 A new species of Proconsul
from the early Miocene of RusingaMfangano Islands Kenya J Hum Evol 2543e56
Ward CV Ruff CB Walker A Rose MD Teaford MF Nengo IO 1995 Func-tional morphology of Proconsul patellas from Rusinga Island Kenya with im-plications for other Miocene-Pliocene catarrhines J Hum Evol 29 1e19
Ward S Brown B Hill A Kelley J Downs W 1999 Equatorius a new hominoidgenus from the Middle Miocene of Kenya Science 285 1382e1386
Werdelin L Sanders WJ 2010 Cenozoic Mammals of Africa University of Cali-fornia Press Berkeley

Figure 10 Scatter plot of principal component scores based on an eigenanalysis of threeposterior point on the superior transverse toruseinfradentale) and subplanum length (mprimarily captures size differences among specimens (Jolicouer 1963 see text and SOM Trespectively) in the symphyseal variables
Figure 9 Line drawing of a hemimandible depicting different orientations of the sub-planum area in Ekembo and Proconsul Females are denoted using shorter vectors maleswith longer vectors Vector is estimated in M 14086 which is broken at the midlineEkembo specimens have a more vertical subplanum compared to those from Songhorand Legetet but within both groups females have more vertically oriented regions thanmales Hence even the most ventrally rotated subplanum in the largest Ekembo malespecimen (KNM-RU 47805) groups with the Tinderet females in this feature
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6154
and P meswae both absolutely and in relation to crown height(Fig 12) This is true for both I1 and I2 A t-test adjusted for non-independent observations (eg I1 and I2 from the same mandible)confirmed the difference as significant (t-test df frac14 12 t frac14 258p frac14 00242) Moreover while the measurable Proconsul samplewas small this same pattern is observable in other specimenswhose tips were slightly to moderately worn and therefore notmeasured
Lower canines The only lower canine trait that discretely sepa-rates these genera is the pointed crown tip in Ekembo (Fig 3d)contrasted with a blade-like (or ldquoburinrdquo) tip in Proconsul(Fig 3c) This feature is well established for P major but is alsoexpressed in KNM-CA 1772 which was referred to P africanus byKelley (1986) and is too small to be either P major or P meswaeTwo other specimens commonly attributed to P africanus KNM-CA 2149 and KNM-SO 1112 both have broken canine tipsLikewise a new specimen (Kor 65004) attributed to P africanusby Pickford et al (2009) also has a broken tip and cannot beevaluated (see Pickford et al 2009 Fig 23) As with the uppercanines the blade-like tip is also present in the deciduous lowercanines of P major (eg KNM-CA 361 S0 5352) and P meswae(KNM-ME 1)
Lower premolars Like most previous authors (eg Andrews 1978Harrison 2002 2010 Pickford et al 2009) we found neitherqualitative nor quantitative characteristics of the lower premolarswith any taxonomic value
Lower molars Differences in lower molar morphology are few andrelatively subtle but in some cases mirror those seen in the uppermolars Cusps on the lower molars of Ekembo are more bunodontthan those in Proconsul and occupy more of the occlusal surfaceBroad inter-cuspal crests are incorporated into the Ekembo cuspsmaking them appear more inflated Molar cusps of Proconsul are
variables symphyseal height (infradentaleegnathion) planum alveolare length (mostost posterior point on the superior transverse torusegnathion) The first componentable 1) and thus PC 2 and PC 3 illustrate the residual shape variance (72 and 28
Figure 11 Scatter plot showing mandibular corpus robusticity under P4 Trend lines were computed using ordinary least squares (OLS) regression Similar relationships areobservable at other parts of the corpus but differential preservation results in smaller samples at those positions Slopes are significantly different (p lt 00001 tested on loggedvariables)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 55
more distinct from one another and are linked by narrower sharperinter-cuspal crests The lower molars of Ekembo also have asubstantially less developed buccal cingulum than those ofProconsul (Harrison 2002 2010 Harrison and Andrews 2009)
Figure 12 Scatter plot of lower incisor crown proportions Differenc
Lastly and again as seen in the upper molars the two genera canbe distinguished by the significantly greater amount of lowermolar flare in Proconsul (Fig 13 t-test df frac14 59 t frac14 713p lt 00001 see also Harrison and Andrews 2009)
es between Ekembo and Proconsul are significant (p frac14 00242)
Figure 13 Box-and-whisker plot of buccolingual flare in lower molars measured asthe average distance between the mesial buccal and lingual cusps divided by overallbuccolingual crown breadth Differences between Ekembo and Proconsul are highlysignificant (p lt 00001)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6156
Relative molar size has been used to distinguish species ofProconsul but we observed the primary difference to be in relativebuccolingual breadths of M1 and M2 In Ekembo M1 is substantiallynarrower than M2 than is the case in Proconsul (Fig 14 Harrison2002 2010) Although none of the smaller Proconsul mandiblespreserve complete crowns for both M1 and M2 KNM-SO 1112 hasan intact M2 and at least preserves the cervix for M1 However evenif the cervical measurement is used for its M1 breadthdthusunderestimating the true crown breadthdKNM-SO 1112 still has abroader first molar in relation to M2 than would be expected inEkembo If crown breadth is estimated from the ratio of cervixbreadth to maximum crown breadth in other Proconsul fossils therange of likely breadths for the SO 1112 M1 clearly distinguishes it
from the Ekembo sample (Fig 14) Testing this relationship onlogged variables using the mean estimate for the SO 1112 M1(Fig 14) reveals a significant difference between the two groups(ANCOVA df frac14 1 F frac14 2424 p frac14 00002)
As with upper third molars we find a great deal of variation inM3 morphology and mistrust the taxonomic significance some-times placed on characteristics of this toothdat least with regard tothese taxa Harrison (2002 2010) cited the mesiodistal position ofthe M3 entoconid as differentiating Kisingiri third molars fromthose attributed to P africanus We concur with his assessment butfound the entoconid position to vary in specimens attributed toP major Thus this character does not differentiate Ekembo from allProconsul species but may distinguish it from the type speciesP africanus
Discussion
The distinctiveness of the Kisingiri large catarrhine materialfrom that at Tinderet and Ugandan localities has been noted pre-viously by several authors (Pickford 1986 Walker et al 1993Begun 2001 2007 MacLatchy and Rossie 2005 Harrison andAndrews 2009 Harrison 2010) What had not been fully docu-mented was the nearly complete lack of overlap between thesesamples in a number of features despite very large assemblagesfrom Rusinga and Mfangano On descriptive criteria alone (Table 2)many isolated teeth (I1 C1 P3 M2 C1) and all of the mandibles canbe assigned to either Proconsul or Ekembowith very little chance ofmisattribution That some of the most diagnostic features occur inthe upper dentitiondthe only anatomy preserved in theP africanus type specimendadds weight to the taxonomicarrangement proposed here The morphology of the upper teeth inall Proconsul species compellingly unites them into a single cladedistinct from E heseloni and E nyanzae This is reinforced bynumerous features of the mandible and mandibular dentition thatfurther distinguish these samples The number and variety of thesefeatures are such that Proconsul and Ekembo are more differentfrom one another than are many closely-related modern catarrhinegenera
Derived morphology of Ekembo and Proconsul
It is premature to formally assign primitive and derived status tothe individual features of Ekembo and Proconsul before completinga formal phylogenetic analysis This is especially true given theabundance of evidence available for Ekembo and hence the poten-tial impact that our generic revision could have on phylogeneticinterpretations Nevertheless in comparison with both extanthominoids and the small-bodied catarrhines from the earlyMiocene it is informative to differentiate the evolutionary grades ofthese genera Many characteristics of Ekembo are more advanced inthe direction of modern apes compared to those of Proconsul(Table 2) General trends such as expanded and more bunodontcusps broader molar crests reduced cingulum and morphologicalsimilarity between P3 and P4 all represent morphologies thatpresage those of later fossil and extant hominoids The moreadvanced morphologies of P3 and M2 are particularly conspicuouscompared not only with those of Proconsul but to most earlyMiocene catarrhines In contrast the mesially positioned stronglyheteromorphic cusps on the P3 of Proconsul plus the correspondingasymmetric buccal profile notched mesial margin and missingmesial fovea are characteristic of other early Miocene catarrhinessuch as Dendropithecus Rangwapithecus and Limnopithecus andare likely primitive among East African catarrhines The moresymmetric P3 of Ekembo with midline positioned cusps andreduced cusp heteromorphy is reminiscent of later hominoids and
Figure 14 Relative buccolingual dimensions of M1 and M2 KNM-SO 1112 has a preserved M2 but only the cervix of M1 Its M1 breadth was estimated based on the ratio to cervixbreadth to maximum crown breadth in other Tinderet fossils Even the cervical breadth of SO 1112 is broader than would be predicted for the crown breadth of a Kisingiri M1 Trendlines computed using OLS regression and using the average estimate of SO 1112 M1 breadth Group differences were significant (p frac14 00002)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 57
even hominid taxa such as Rudapithecus (eg Kordos and Begun2001)
Proconsul is likewise united by a suite of features that are for themost part characteristic only of this lineage (Table 2) Themorphology of both upper and lower canines is unique to thisgenus and shared by all three well-known species plus the poorlyrepresented middle Miocene P gitongai (Pickford and Kunimatsu2005) The mandibular symphysis is also distinctive in Proconsulcompared with other catarrhines with its very long and shallowlysloping planum alveolare and steeply inclined subplanum surfaceIndeed even the small edentulous anterior mandibular fragmentKNM-SO 372 is readily placed within Proconsul and excluded fromother similarly sized catarrhine taxa such as Rangwapithecus andEkembo based on this unique symphyseal morphology (Figs 8 and9)
Other differences between Ekembo and Proconsul are of uncer-tain evolutionary polarity (eg orientation of the posthypoconecrista) or may be of taxonomic relevance without broader phylo-genetic significance (eg degree of molar flare Table 2) Werecognize that there are morphological differences within Procon-sul particularly between the better known species P africanus andP major However many of these differences are likely related tosize and represent variations on a single generic theme that aremarkedly different from the morphology of Ekembo The recentdescription (Harrison and Andrews 2009) of the intermediatelysized P meswae underscores the broad morphological unity withinProconsul and further substantiates a well-defined Proconsul cladecomprising Tinderet and Ugandan species
Based on the abundant morphological and morphometric evi-dence supporting genus-level distinction between Proconsul andEkembo we are currently conducting a more detailed phylogeneticanalysis of all early Miocene catarrhines in order to better under-stand the distribution and evolutionary significance of these andother morphological features We anticipate that the debates sur-rounding the phylogenetic relationships of an historically consti-tuted sample of Proconsul can be more satisfactorily resolved nowthat the Kisingiri material is appropriately recognized as a distinctgenus
The generic disposition of ldquoUgandapithecusrdquo species
A great many features said to characterize ldquoUgandapithecusrdquowere added to the original diagnosis of the genus by Pickford et al(2009) in their systematic revision of Proconsul (sensu lato) As partof that study they also erected a new species ldquoUrdquo legetetensis andtransferred P meswae to ldquoUgandapithecusrdquo Evaluating the same setof features across the entire available sample however we wereunable to find support for this more expansive conception ofldquoUgandapithecusrdquo for several reasons First some diagnostic fea-tures cited in Pickford et al (2009) were quite variable within theldquoUgandapithecusrdquo sample As an example marked reduction of theM3 entoconid to a bead on the lingual cingulum is not shared byseveral ldquoUrdquo major specimens such as KNM-CA 393 NAP I 49000and Kor 253004 (see Pickford et al 2009 Fig 33e) and was likelynot a feature of a few of the more worn specimens eg BUMP 600and BUMP 601 or even the holotype of P major M 16648 Second
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6158
some diagnostic features of ldquoUgandapithecusrdquo were found to bevariably present in Ekembo arguing against their utility for dis-tinguishing ldquoUgandapithecusrdquo from Pickford et als (2009) com-bined P africanus thorn Kisingiri conception of Proconsul For examplesignificantly reduced curvature of the I1 distal margin in ldquoUganda-pithecusrdquo is also found in some Kisingiri specimens including KNM-RU 1685 1712 1714 1831 1846 and 1951 several more Rusingaspecimens have moderately reduced curvature Third some fea-tures that purportedly distinguish ldquoUgandapithecusrdquo fromP africanus are likely related to size For example the somewhatmore central position of the P3 paracone in ldquoUrdquomajor is not seen inthe smaller ldquoUrdquo meswae whose paracone is positioned similarly tothat of the holotype of P africanus (see Harrison and Andrews2009 Fig 6d)
However the major part of the evidence for recognizingldquoUgandapithecusrdquo (sensu Pickford et al 2009) was assembled by re-assigning specimens traditionally attributed to P africanus into newspecies of ldquoUgandapithecusrdquo The P africanus holotype was left inassociation with the Rusinga sample despite having morphologyoverwhelmingly characteristic of other Tinderet specimens Theresulting specimen allocations left P africanus morphologicallyimpoverisheddstripped of much of its hypodigmdso that Pro-consul for Pickford et al (2009) came to be characterized primarilyby the Rusinga sample (see similar comments in Harrison andAndrews 2009) Within that conceptual framework any spec-imen that differs from those at Rusinga could not be Proconsul butmust instead belong to ldquoUgandapithecusrdquo Thus specimens such asthe KNM-SO 1112 mandible were erroneously assigned to ldquoUgan-dapithecusrdquo because they were (correctly) perceived as differingfrom their Rusinga counterparts (Pickford et al 2009)
Here we restore the P africanus hypodigm and associate it withthe other Tinderet fossils according to their shared morphologySpecimens assigned to ldquoUgandapithecusrdquo revert to Proconsul andmany of the features cited to support a speciose ldquoUgandapithecusrdquo(Pickford et al 2009) instead become diagnostic features of Pro-consul This correction serves to highlight the distinctiveness of theKisingiri remains and the need to recognize this distinctiveness atthe generic level
As demonstrated above specimens from Kisingiri do not displaythe morphology of Proconsul as represented by the holotype ofP africanus M 14084 Given that this specimen preserves only theupper canine and cheek teeth similarities with or departures fromthis morphology represent the most compelling arguments forassessing inclusivity in the genus However for the mandible andlower dentition if one simply sorts the specimens by elementwithout relying on taxonomic preconceptions the overwhelmingdistinction within the traditionally constituted Proconsul sample isbetween Kisingiri material on the one hand and that from Tinderetand Uganda on the other (cf Walker et al 1993 Begun 2001 2007MacLatchy and Rossie 2005 Harrison and Andrews 2009Harrison 2010) Additionally with the Kisingiri specimensremoved to Ekembo the postcranial differences used to partlyjustify a distinct ldquoUgandapithecusrdquo lose their relevance (seeHarrison and Andrews 2009)
We have differed among ourselves in our past treatment ofP major as generically distinct from the smaller species of Pro-consul (Begun 2007 McNulty 2010) Morphologically many ofthe unique features of P major can be interpreted as size-relatedvariants on the smaller P africanus morphology Even if so thequestion is whether these are sufficiently different and numerousto warrant generic distinction that is do they suggest thatP major belongs in a separate clade from P africanus andP meswae As we have demonstrated differences among thesethree species are certainly fewer and show less morphologicaldistinction than those between Ekembo and any of the three
Nevertheless while we ultimately agree with Harrison andAndrews (2009 see also Begun 2007) that the features dis-tinguishing ldquoUrdquo major from P africanus are consistent with thosethat distinguish congeneric species and are at least partly due todifferences in size we concede the possibility that the largestProconsul species could belong in a separate genus (Senut et al2000 Pickford and Kunimatsu 2005) However whether or notone recognizes a separate genus for P major the P africanus ma-terial does not sort morphologically with the Kisingiri materialTherefore the taxonomic alternatives are either two generaProconsul and Ekembo or three Proconsul Ugandapithecus andEkembo with Proconsul and Ugandapithecus united in a cladeexclusive of Ekembo Thus while we strongly favor the firstalternative recognizing Ugandapithecus does not change thefundamental argument concerning the need to refer the Kisingirispecies to a generically and cladistically distinct taxon from thespecies represented at Tinderet and the Ugandan sites
Specimens of interest
Despite the substantial dental and gnathic differences betweenProconsul and Ekembo there are a few specimens whose taxonomicaffinities require additional scrutiny
KNM-RU 1676-77 This set of associated upper and lower teeth is aconsistent outlier among E heseloni and E nyanzae specimens Insome ways this specimen more closely resembles Tinderet Pro-consul in the features that distinguish it from Ekembo Molar cuspmorphology and cingulum development I1 lingual tubercledevelopment and M2 shape and posthypocone crista orientationare more consistent with specimens from the Tinderet sites On theother hand it otherwise resembles Ekembo and lacks the strongestdiagnostic features that unite species of Proconsul such as thedistinctive canine and P3 morphology Overall this specimen doesnot fit unambiguously into either genus in fact the degree ofbuccolingual flare cusp morphology enamel wrinkling and espe-cially the long extension of enamel onto the anterior root in P3 arein some ways reminiscent of Afropithecus although it clearly differsfrom that genus as well (cf Rossie and MacLatchy 2013)
Were these characteristics individually distributed among thelarge Kisingiri sample one might attribute this to intra-genericvariation but when found together in a single specimen from alarge sample it suggests a unique individual One of us (KPM)thinks it likely that the specimen was not actually found at aKisingiri locality based on its unique preservation compared toother Rusinga fossils and on other examples of mis-proveniencedspecimens (Leakey 1967 Andrews and Molleson 1979 Pickford1986 Kelley 1986) However we recognize that relying toofrequently on this explanation allows almost any scenario to bejustified and suggest some alternative possibilities One is that thespecimen was found on Rusinga but in the much older WayandoFormation strata instead of the Hiwegi Formation The differencesin morphology in that case might reflect more than a million yearsof temporal change (McCollum et al 2013) with KNM-RU 1676-77preserving certain more primitive Proconsul-like features As analternative it is reasonable to assume that fossil assemblages asvast as those from Rusingawill include not only abundant taxa butalso rare or infrequent inhabitants of the local biome There aremany examples of this from Rusingas mammalian communityeg ldquoHemicyonrdquo Kulutherium Tadarida cf Hyainailouros cfArchaeobeledon Kelba (see relevant chapters in Werdelin andSanders 2010) As such KNM-RU 1676-77 may simply representa rare and well-preserved specimen of some other catarrhinetaxon that lived among or visited a primate community dominatedby Ekembo
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 59
KNM-SO 1112 This specimen was initially referred to Rangwapi-thecus but soon after was transferred to Proconsul afrianus (Bosler1981 Martin 1981)da designation that most subsequent authorshave followed (Pickford 1986 Nengo and Rae 1992 Kelley 19861995 Walker et al 1993 MacLatchy and Rossie 2005 Hill et al2013) Pickford et al (2009) more recently referred this specimento ldquoUrdquo meswae citing two primary reasons First they reiteratedBoslers (1981) concern that its P3 differs from others attributedto P africanus However at that time the majority of what wasconsidered to be P africanus would have been from RusingaIsland In fact while noting its contrasting morphology Boslersuggested that differences between KNM-SO 1112 and other P3smight reasonably be attributed to sexual dimorphism notingthat this specimen provides ldquoa not implausible match for the Pafricanus holotype maxilla M 14084rdquo (1981152) Thatassessment is borne out in this study Using the much largersample of Ekembo P3s as a model variation can be seen toencompass specimens with well-developed mesial beaks andthose with more bilaterally compressed crowns and parallelsides The latter morphology is predominantly found amongmale specimens (KNM-RU 1674 1924 and KPS I and III) thussupporting Boslers (1981) suggestion that this variation isrelated to sexual dimorphism and hence that KNM-SO 1112 is amale P africanus
The second feature cited by Pickford et al (2009) in referringthis specimen to ldquoUrdquo meswae was posterior shallowing of themandibular corpus However because they redefined ldquoUgandapi-thecusrdquo to include all of the well-preserved Tinderet mandibles(Pickford et al 2009)dleaving Proconsul to be represented only byRusinga specimensdtheir only alternative within this scheme for aspecimen the size of SO 1112 was to refer it to ldquoUrdquomeswae It seemsodd though to assign this specimen to a species from a (perhapsmuch) older site when the Proconsul africanus holotype is ofappropriate size and similar age and as there are no other adultmandibles from either species with which to compare it In anycase mandibular shallowing is variable in both Ekembo and Pro-consul with some Kisingiri specimens exhibiting considerableshallowing (eg KNM-RU 2087) and some ldquoUgandapithecusrdquo spec-imens having very little (eg M 14086)
We therefore see no morphological grounds to assign KNM-SO1112 to P meswae but do find some morphological evidence toassign it as a male P africanus which occurs at Songhor
KNM-LG 452 This very complete mandible was originally recog-nized as a female of P major based on the size of the dentition androbusticity of the mandible (Martin 1981) Bosler (1981) howeverleft this specimen unassigned noting similarities to P major in P3-M1 size but having distinctly smaller M2e3 In this she likenedKNM-LG 452 to a few other specimens (notably M 14086) thatare smaller in their distal molars in a manner more consistentwith taxonomic rather than sex differences (Bosler 1981 see alsoPilbeam 1969) Kelley (1986 1995) also assigned this specimen asa female of P major based on canine size and morphology butnoted that the canine lacked the shallowly-sloping mesial ridgetypical of most female anthropoid canines This is also truehowever of two other canines assigned as female P major KO 9and SO 373 (Kelley 1986) and so might simply be an unusualcharacteristic of P major female lower canines In the end Kelley(1995) concluded that considering all of the criteria fordistinguishing male and female canines the evidence that KNM-LG 452 is female remains questionable
Pickford et al (2009) further developed the idea that two spe-cies are represented within the P major sample and made KNM-LG452 the holotype of a new species ldquoUrdquo legetetensis We concur thatthis specimen is unique among the well-preserved Tinderet
mandibles in some important features Whereas male and femalemandibles of P major have long internal symphyseal planesexpansive extra- and retromolar sulci and rami that cross theocclusal plane at distal M3 KNM-LG 452 has a shorter symphysealplane (though not to the extent seen in Ekembo) a moderateextramolar sulcus but no retromolar sulcus and a ramus thatcrosses the occlusal plane more mesially If KNM-SO 396 and SO404 represent males and females respectively of P major then themorphology of KNM-LG 452 appears distinct
At present however we do not think there is sufficient evidenceto differentiate this specimen from either P major or P meswaePickford et al (2009) argue that KNM-LG 452 cannot belong to thelatter species because of its larger size The size differences how-ever are well within the range of variation seen in P majorE heseloni and E nyanzae Moreover whereas the Legetet mandibleis larger than most of the P meswae hypodigm an adult P4 referredto this species (KNM-ME 25) substantially exceeds its counterpartin KNM-LG 452 Should the Legetetmandible ultimately prove to bea male specimen size alone would not rule out its attribution toP meswae On the other hand if it is female then it becomes harderto justify its inclusion in that sample of mostly smaller specimens Iffemale therefore it could represent a separate species for whichthe binomial Proconsul legetetensis comb nov is available Recentlycollected fossils from the SonghorKapurtay area provide someadditional support for the possibility that three species of Proconsulexisted at these localities (McNulty 2014ab) However toconvincingly demonstrate this and determine whether a thirdspecies can be differentiated from P meswae will require a moredetailed analysis of variation among Proconsul species a corre-sponding study of variation in modern taxa anddalmost certain-lydadditional fossil specimens For now we regard ldquoUrdquo legetetensisas a subjective junior synonym of P major
Conclusions
The genus Proconsul has been central to studies of ape and hu-man evolution since its first description by Hopwood (1933) As oneof the best known fossil catarrhines represented by nearly everyskeletal element our knowledge of Proconsuls functional anatomypositional behavior and overall morphology has made it thefoundation for paleoanthropological research in the early Mioceneas well as studies on the origin and early diversification of homi-noids With respect to systematics however Proconsul is less wellunderstood the type species P africanus is known from a rela-tively small number of specimens from sites in the Tinderet areaand no postcranial bones can be unambiguously assigned to itRather it is the copious and well-preserved specimens from theKisingiri sites of Rusinga and Mfangano Islands that have formedthe basis for the common image of Proconsul
We argue here that the Kisingiri species traditionally referredto ProconsuldP heseloni and P nyanzaedare generically distinctfrom P africanus and hereby transfer them into a new genusEkembo based on a substantial number of morphological differ-ences (Table 2) Considering only those traits preserved in theP africanus holotype the entire sample from Rusinga and Mfan-gano differs markedly in the morphology of the upper canine P3P4 and M2 Additional differences in the I1 lower incisors andcanines molar cusp and occlusal crest morphology degree of flarein all molars and especially mandibular features clearly distin-guish the Kisingiri from non-Kisingiri samples in a way that isincompatible with congeneric species With the transfer ofE heseloni and E nyanzae Proconsul becomes a well-defined cladecomprising at least three speciesdP africanus P major andP meswaedunited by several features that are unique for thegenus (Table 2)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6160
We found no support in this study for the genus ldquoUgandapi-thecusrdquo as constructed by Pickford et al (2009) In that paper theytransferred the vast majority of Proconsul fossils from the Tinderetand Napak localities into ldquoUgandapithecusrdquo leaving P africanusrepresented only by its holotype and a small number of isolatedteeth and aligning it with the material from Rusinga and Mfan-ganoWhen the P africanus hypodigm is reconstituted and properlyaffiliated with the other Tinderet species according to itsmorphology much of the evidence marshaled by Pickford et al(2009) for ldquoUgandapithecusrdquo actually supports the distinction be-tween the Tinderet and Kisingiri samples ie the generic distinc-tion of Ekembo In addition some of the traits enumerated byPickford and colleagues in support of ldquoUgandapithecusrdquo wererevealed in our analysis to be variable within one or more speciesand therefore of questionable taxonomic utility We do not rejectout of hand the original conception of ldquoUgandapithecusrdquo (Senutet al 2000) as either a monotypic genus distinguishing ldquoUrdquo ma-jor or perhaps including a closely allied middle Miocene ldquoUrdquogitongai as well (Pickford and Kunimatsu 2005) but we have fornow retained the early Miocene specimens of both in Proconsulpending further study However even if Ugandapithecus is a taxo-nomically valid genus comprising just the latter two species theirmorphology indicates that theywould still be united in a cladewithProconsul and they lack several more advanced features that partlydefine Ekembo Better understanding of the role that body size playsinmediating differences within Proconsulwill help to further clarifythe relationships among these species
Despite the existence of one well-represented and several lesscomplete skeletons of what was long assumed to be Proconsul thephylogenetic status of this genus has defied consensus This islargely due in our view to Proconsul having been an admixture oftwo distinct genera one more advanced in the direction of extantapes than the other As we demonstrate here the material fromRusinga and Mfangano traditionally referred to Proconsul andwhich includes all of the more complete skeletal material isdistinct from the Tinderet and Napak material and belongs in aseparate genus Ekembo By disentangling these distinct clades weanticipate being able to achieve better phylogenetic resolution forboth genera thereby improving our knowledge of early hominoidevolution and diversification
Acknowledgments
We thank many people and agencies that assisted with thedevelopment and completion of this project This research wasinitiated as a result of funding from the Wenner-Gren Foundationand the Johns Hopkins School of Medicine to DR Begun in1987e1989 and was renewed following research and travelgenerously funded by multiple Leakey Foundation grants to KMcNulty National Science Foundation grants to K McNulty(0852609 1241807) the McKnight Foundation the University ofMinnesota the Lloyd A Wilford Endowment the LeverhulmeFoundation and the Turkana Basin Institute J Kelley gratefullyacknowledges support from the Institute of Human Origins Ari-zona State University DR Begun also wishes to acknowledge theAlexander Humboldt Stiftung and NSERC for funding support Weare grateful to the National Museums of Kenya the UgandaMuseum the Natural History Museum and to Rose Mwanja EzraMusiime Tom Mukhuyu Samuel Muteti James Yatich and MaryMuungu for access to and assistance with specimens in their careMany people provided assistance comments criticisms insightsand suggestions on various aspects of this work and we especiallythank Holly Dunsworth Will Harcourt-Smith Tom Lehmann DanPeppe Nicole Garrett Kirsten Jenkins Rutger Jansma Susy CoteIsaiah Nengo Andrew Hill and Brenda Benefit K McNulty
acknowledges the evolutionary anthropology faculty and studentsat Durham University for their helpful comments on an earlyversion of this manuscript and for their gracious hospitality Weare likewise grateful to the anonymous reviewers associate editorand editor whose comments strengthened the paper This manu-script is publication 1 supporting Research on East AfricanCatarrhine and Hominoid Evolution (REACHE)
Appendix A Supplementary data
Supplementary data related to this article can be found at httpdxdoiorg101016jjhevol201503009
References
Andrews P 1974 New species of Dryopithecus from Kenya Nature 249 188e190Andrews P 1978 A revision of the Miocene Hominoidea of East Africa Bull Brit
Mus (Nat Hist) Geol 30 85e225Andrews P 1985 Family group systematics and evolution among catarrhine pri-
mates In Delson E (Ed) Ancestors The Hard Evidence Liss New Yorkpp 14e22
Andrews P Harrison T Martin L Pickford M 1981 Hominoid primates from anew Miocene locality named Meswa Bridge in Kenya J Hum Evol 10123e128
Andrews P Begun D Zylstra M 1997 Interrelationships between functionalmorphology and paleoenvironments in Miocene hominoids In Begun DWard C Rose M (Eds) Function Phylogeny and Fossils Miocene HominoidEvolution and Adaptations Plenum Press New York pp 29e58
Andrews P Molleson T 1979 The provenance of Sivapithecus africanus Bull BritMus (Nat Hist) Geol 32 19e23
Begun DR 2001 African and Eurasian Miocene hominoids and the origins of theHominidae In de Bonis L Koufos GD Andrews P (Eds) Hominoid Evolutionand Climatic Change in Europe Phylogeny of the Neogene Hominoid Primatesof Eurasia Cambridge University Press London pp 231e253
Begun DR 2007 Fossil record of Miocene hominoids In Henke W Tattersall I(Eds) Handbook of Paleoanthropology Springer Berlin pp 921e977
Begun DR Teaford MF Walker AC 1994 Comparative and functional anatomyof Proconsul phalanges from the Kaswanga primate site Rusinga Island KenyaJ Hum Evol 26 89e165
Bishop WW 1964 More fossil primates and other Miocene mammals from North-East Uganda Nature 203 1327e1331
Bishop WW Miller JA Fitch FJ 1969 New potassium-argon age determinationsrelevant to the Miocene fossil mammal sequence in East Africa Am J Sci 267669e699
Blankenship JA Mincer HH Anderson KM Woods MA Burton EL 2007Third molar development in the estimation of chronologic age in Americanblacks as compared with whites J Forensic Sci 52 428e433
Bosler W 1981 Species groupings of early Miocene dryopithecine teeth from EastAfrica J Hum Evol 10 151e158
Coleman MN 2008 What does geometric mean mean geometrically Assessingthe utility of geometric mean and other size variables in studies of skullallometry Am J Phys Anthropol 135 404e415
Cote S Malit N Nengo I 2013 Additional mandibles of Rangwapithecus gordonian early Miocene catarrhine from the Tinderet localities of Western KenyaJ Hum Evol 153 341e352
Deane AS 2009 Early Miocene catarrhine dietary behavior the influence of theRed Queen Effect on incisor shape and curvature J Hum Evol 56 275e285
Drake R Van Couvering JA Pickford M Curtis G Harris JA 1988 New chro-nology for the Early Miocene mammalian faunas of Kisingiri Western KenyaJ Geol Soc Lond 145 479e491
Evernden JF Savage DE Curtis GH James GT 1964 Potassium-argon datesand the Cenozoic mammalian chronology of North America Am J Sci 262145e198
Gebo DL MacLatchy L Kityo R Deino A Kingston J Pilbeam D 1997A hominoid genus from the Early Miocene of Uganda Science 276 401e404
Geoffroy Saint-Hilaire E 1812 Tableau des quadrumanes 1 Ord QuadrumanesAnnls Mus Hist Nat Paris 19 85e122
Gray JE 1825 Outline of an attempt at the disposition of the Mammalia into tribesand families with a list of the genera apparently appertaining to each tribeAnnls Philos 10 337e344
Harrison T 1982 Small-bodied apes from the Miocene of East Africa PhDDissertation University of London
Harrison T 1986 New fossil anthropoids from the middle Miocene of East Africaand their bearing on the origin of the Oreopithecidae Am J Phys Anthropol 71265e284
Harrison T 1987 The phylogenetic relationships of the early catarrhine primates areview of the current evidence J Hum Evol 16 41e80
Harrison T 2002 Late Oligocene to middle Miocene catarrhines from Afro-ArabiaIn Hartwig WC (Ed) The Primate Fossil Record Cambridge University PressCambridge pp 311e338
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 61
Harrison T 2010 Dendropithecoidea Proconsuloidea and Hominoidea (CatarrhiniPrimates) In Werdelin L Sanders WJ (Eds) Cenozoic Mammals of AfricaUniversity of California Press Berkeley pp 429e469
Harrison T Andrews P 2009 The anatomy and systematic position of a newspecies of early Miocene proconsulid from Meswa Bridge Kenya J Hum Evol56 479e496
Hill A Nengo IO Rossie JB 2013 A Rangwapithecus gordoni mandible from theearly Miocene site of Songhor Kenya J Hum Evol 65 490e500
Hopwood AT 1933 Miocene primates from Kenya J Linn Soc Zool 38 437e464Ishida H Kunimatsu Y Takano T Nakano Y Nakatsukasa M 2004 Nacholapi-
thecus skeleton from the Middle Miocene of Kenya J Hum Evol 46 69e103Jolicouer P 1963 The multivariate generalization of the allometry equation Bio-
metrics 19 497e499Jungers WL Falsetti AB Wall CE 1995 Shape relative size and size-
adjustments in morphometrics Am J Phys Anthropol 38 (S2) 137e161Kavanagh KD Evans AR Jernvall J 2007 Predicting evolutionary patterns of
mammalian teeth from development Nature 449 427e433Kelley J 1986 Species recognition and sexual dimorphism in Proconsul and
Rangwapithecus J Hum Evol 15 461e495Kelley J 1995 Sex determination in Miocene catarrhine primates Am J Phys
Anthropol 96 390e417Kelley J 1993 Taxonomic implications of sexual dimorphism in Lufengpithecus In
Kimbel WH Martin LB (Eds) Species Species Concepts and Primate Evo-lution Springer New York pp 429e458
Kent PE 1942 The country round the Kavirondo Gulf of Victoria Nyanza Geogr J100 22e31
Kordos L Begun DR 2001 A new cranium of Dryopithecus from RudabanyaHungary J Hum Evol 15 689e700
Le Gros Clark WE Leakey LSB 1950 Diagnoses of East African Miocene Homi-noidea Q J Geol Soc Lond 105 260e263
Le Gros Clark WE Leakey LSB 1951 The Miocene Hominoidea of East Africa InFossil Mammals of Africa No 1 British Museum (Natural History) London
Leakey LSB 1943 A Miocene anthropoid mandible from Rusinga Kenya Nature152 319e320
Leakey LSB 1963 East African fossil Hominoidea and the classification within thissuper-family In Washburn SL (Ed) Classification and Human EvolutionAldine Chicago pp 32e49
Leakey LSB 1967 Notes on the mammalian faunas from the Miocene and Pleis-tocene of East Africa In Bishop WW Clark JD (Eds) Background to Evolu-tion in Africa University of Chicago Press Chicago pp 7e29
Leakey RE Leakey MG 1986 A new Miocene hominoid from Kenya Nature 324143e146
Leakey RE Leakey MG Walker AC 1988 Morphology of Afropithecus turka-nensis from Kenya Am J Phys Anthropol 76 289e307
Leakey MG Ungar PS Walker A 1995 A new genus of large primate from theLate Oligocene of Lothidok Turkana District Kenya J Hum Evol 28519e531
Linnaeus C 1758 Systema naturae per regna tria naturae secundum classesordines genera species cum characteribus differentris synonymis locis tenthed Laurentii Salvii Stockholm
MacInnes DG 1943 Notes on the East African Miocene primates J E Afr UgandaNat Hist Soc 17 141e181
MacLatchy L 2004 The oldest ape Evol Anthropol 13 90e103MacLatchy L Bossert WH 1996 An analysis of the articular surface distribution of
the femoral head and acetabulum in anthropoids with implications for hipfunction in Miocene hominoids J Hum Evol 31 425e453
MacLatchy L Rossie JB 2005 The Napak hominoid still Proconsul major InLieberman DE Smith RJ Kelley J (Eds) Interpreting the Past Essays onHuman Primate and Mammal Evolution in Honor of David Pilbeam Brill Ac-ademic Publishers Boston pp 15e28
Martin L 1981 New specimens of Proconsul from Koru Kenya J Hum Evol 10139e150
McCollum MS Peppe DJ McNulty KP Dunsworth HM Harcourt-Smith WEH Andrews AL 2013 Magnetostratigraphy of the early MioceneHiwegi Formation (Rusinga Island Lake Victoria Kenya) Geological Society ofAmerica Abstracts with Programs 45 p 12
McNulty KP 2010 Apes and tricksters the evolution and diversification ofhumans closest relatives Evol Edu Outreach 3 322e332
McNulty KP Harcourt-Smith WEH Dunsworth HM 2007 New primate fossilsfrom Rusinga Island Kenya Am J Phys Anthropol 132 (S44) 170
McNulty KP MacLatchy L Peppe DJ Nengo I Manthi FK Miller ERStevens NJ Cote S Lehmann T Kingston J 2014a A regional approach toEast African early Miocene paleobiology J Vert Paleontol 185 SVP Programand Abstracts Book 2014
McNulty KP MacLatchy L Rossie JB Peppe DJ Deino AL Mbua ENManthi FK Nengo IO Stevens NJ Cote S Lehmann T Gutierrez M 2014b
Research on East African catarrhine and hominoid evolution results from thefirst year Am J Phys Anthropol 153 (S58) 182
Michel LA Peppe DJ Lutz JA Driese SG Dunsworth HM Harcourt-Smith WEH Horner W Lehmann T Nightengale S McNulty KP 2014Remnants of an ancient forest provide ecological context for Early Miocenefossil apes Nature Comm 5 3236
Myres JL 1932 Africa Man 32 208Nakatsukasa M Kunimatsu Y Nakano Y Takano T Ishida H 2007 Vertebral
morphology of Nacholapithecus kerioi based on KNM-BG 35250 J Hum Evol52 347e369
Nengo IO Rae TC 1992 New hominoid fossils from the early Miocene site ofSonghor Kenya J Hum Evol 23 423e429
Peppe DJ McNulty KP Cote SM Harcourt-Smith WEH Dunsworth HM VanCouvering JA 2009 Stratigraphic interpretation of the Kulu Formation (EarlyMiocene Rusinga Island Kenya) and its implications for primate evolutionJ Hum Evol 56 447e461
Pickford M 1981 Preliminary Miocene mammalian biostratigraphy for westernKenya J Hum Evol 10 73e97
Pickford M 1986 Sexual dimorphism in Proconsul Hum Evol 1 111e148Pickford M Andrews PJ 1981 The Tinderet Miocene sequence in Kenya J Hum
Evol 10 11e33Pickford M Kunimatsu Y 2005 Catarrhines from the Middle Miocene (ca 145
Ma) of Kipsaraman Tugen Hills Kenya Anthropol Sci 113 189e224Pickford M Senut B Gommery D Musiime E 2009 Distinctiveness of Ugan-
dapithecus from Proconsul Estudios Geol 65 183e241Pilbeam DR 1969 Tertiary Pongidae of East Africa evolutionary relationships and
taxonomy Bulletin of the Peabody Museum of Natural History 31 pp 1e185Rae TC 1993 Phylogenetic analysis of proconsulid facial morphology PhD
Dissertation State University of New York at Stony BrookRae TC 1999 Mosaic evolution in the origin of the Hominoidea Folia Primatol 70
125e135Rafferty KL Walker A Ruff C Rose MD Andrews PJ 1995 Postcranial esti-
mates of body weights in Proconsul with a note on a distal tibia of P major fromNapak Uganda Am J Phys Anthropol 97 391e402
Rossie JB MacLatchy L 2013 Dentognathic remains of an Afropithecus individualfrom Kalodirr Kenya J Hum Evol 65 199e208
Senut B Pickford M Gommery D Kunimatsu Y 2000 Un nouveau genredrsquohominoiumlde du Miocene inferieur drsquoAfrique orientale Ugandapithecus major(Le Gros Clark and Leakey 1950) CR Acad Sci Paris 331 227e233
Simons EL Pilbeam DR 1965 Preliminary revision of the Dryopithecinae (Pon-gidae Anthropoidea) Folia Primatol 3 81e152
Smith TM Martin LB Leakey MG 2003 Enamel thickness microstructure anddevelopment in Afropithecus turkanensis J Hum Evol 44 283e306
Stevens NJ Seiffert ER OConnor PM Roberts EM Schmitz MD Krause CGorscak E Ngasala S Hieronymus TL Temu J 2013 Palaeontological evi-dence for an Oligocene divergence between Old World monkeys and apesNature 497 611e614
Suwa G Kono RT Katoh S Asfaw B Beyene Y 2007 A new species of great apefrom the late Miocene epoch in Ethiopia Nature 448 921e924
Szalay FS Delson E 1979 Evolutionary History of the Primates Academic PressNew York
Teaford MF Beard KC Leakey RE Walker A 1988 New hominoid facial skel-eton from the Early Miocene of Rusinga Island Kenya and its bearing on therelationship between Proconsul nyanzae and Proconsul africanus J Hum Evol17 461e477
Van Couvering JAH Miller J 1969 Miocene stratigraphy and age determinationsRusinga Island Kenya Nature 221 628e632
Walker A 1997 Proconsul function and phylogeny In Begun DR Ward CVRose MD (Eds) Function Phylogeny and Fossils Miocene Hominoid Evolu-tion and Adaptation Plenum Press New York pp 209e224
Walker A Pickford M 1983 New postcranial fossils of Proconsul africanus andProconsul nyanzae In Ciochon RL Corruccini RS (Eds) New Interpretationsof Ape and Human Ancestry Plenum Press New York pp 325e351
Walker A Teaford M 1988 The Kaswanga primate site an Early Miocene hom-inoid site on Rusinga Island Kenya J Hum Evol 17 539e544
Walker A Teaford M 1989 The hunt for Proconsul Sci Am 260 76e82Walker A Teaford MF Martin L Andrews P 1993 A new species of Proconsul
from the early Miocene of RusingaMfangano Islands Kenya J Hum Evol 2543e56
Ward CV Ruff CB Walker A Rose MD Teaford MF Nengo IO 1995 Func-tional morphology of Proconsul patellas from Rusinga Island Kenya with im-plications for other Miocene-Pliocene catarrhines J Hum Evol 29 1e19
Ward S Brown B Hill A Kelley J Downs W 1999 Equatorius a new hominoidgenus from the Middle Miocene of Kenya Science 285 1382e1386
Werdelin L Sanders WJ 2010 Cenozoic Mammals of Africa University of Cali-fornia Press Berkeley

Figure 11 Scatter plot showing mandibular corpus robusticity under P4 Trend lines were computed using ordinary least squares (OLS) regression Similar relationships areobservable at other parts of the corpus but differential preservation results in smaller samples at those positions Slopes are significantly different (p lt 00001 tested on loggedvariables)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 55
more distinct from one another and are linked by narrower sharperinter-cuspal crests The lower molars of Ekembo also have asubstantially less developed buccal cingulum than those ofProconsul (Harrison 2002 2010 Harrison and Andrews 2009)
Figure 12 Scatter plot of lower incisor crown proportions Differenc
Lastly and again as seen in the upper molars the two genera canbe distinguished by the significantly greater amount of lowermolar flare in Proconsul (Fig 13 t-test df frac14 59 t frac14 713p lt 00001 see also Harrison and Andrews 2009)
es between Ekembo and Proconsul are significant (p frac14 00242)
Figure 13 Box-and-whisker plot of buccolingual flare in lower molars measured asthe average distance between the mesial buccal and lingual cusps divided by overallbuccolingual crown breadth Differences between Ekembo and Proconsul are highlysignificant (p lt 00001)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6156
Relative molar size has been used to distinguish species ofProconsul but we observed the primary difference to be in relativebuccolingual breadths of M1 and M2 In Ekembo M1 is substantiallynarrower than M2 than is the case in Proconsul (Fig 14 Harrison2002 2010) Although none of the smaller Proconsul mandiblespreserve complete crowns for both M1 and M2 KNM-SO 1112 hasan intact M2 and at least preserves the cervix for M1 However evenif the cervical measurement is used for its M1 breadthdthusunderestimating the true crown breadthdKNM-SO 1112 still has abroader first molar in relation to M2 than would be expected inEkembo If crown breadth is estimated from the ratio of cervixbreadth to maximum crown breadth in other Proconsul fossils therange of likely breadths for the SO 1112 M1 clearly distinguishes it
from the Ekembo sample (Fig 14) Testing this relationship onlogged variables using the mean estimate for the SO 1112 M1(Fig 14) reveals a significant difference between the two groups(ANCOVA df frac14 1 F frac14 2424 p frac14 00002)
As with upper third molars we find a great deal of variation inM3 morphology and mistrust the taxonomic significance some-times placed on characteristics of this toothdat least with regard tothese taxa Harrison (2002 2010) cited the mesiodistal position ofthe M3 entoconid as differentiating Kisingiri third molars fromthose attributed to P africanus We concur with his assessment butfound the entoconid position to vary in specimens attributed toP major Thus this character does not differentiate Ekembo from allProconsul species but may distinguish it from the type speciesP africanus
Discussion
The distinctiveness of the Kisingiri large catarrhine materialfrom that at Tinderet and Ugandan localities has been noted pre-viously by several authors (Pickford 1986 Walker et al 1993Begun 2001 2007 MacLatchy and Rossie 2005 Harrison andAndrews 2009 Harrison 2010) What had not been fully docu-mented was the nearly complete lack of overlap between thesesamples in a number of features despite very large assemblagesfrom Rusinga and Mfangano On descriptive criteria alone (Table 2)many isolated teeth (I1 C1 P3 M2 C1) and all of the mandibles canbe assigned to either Proconsul or Ekembowith very little chance ofmisattribution That some of the most diagnostic features occur inthe upper dentitiondthe only anatomy preserved in theP africanus type specimendadds weight to the taxonomicarrangement proposed here The morphology of the upper teeth inall Proconsul species compellingly unites them into a single cladedistinct from E heseloni and E nyanzae This is reinforced bynumerous features of the mandible and mandibular dentition thatfurther distinguish these samples The number and variety of thesefeatures are such that Proconsul and Ekembo are more differentfrom one another than are many closely-related modern catarrhinegenera
Derived morphology of Ekembo and Proconsul
It is premature to formally assign primitive and derived status tothe individual features of Ekembo and Proconsul before completinga formal phylogenetic analysis This is especially true given theabundance of evidence available for Ekembo and hence the poten-tial impact that our generic revision could have on phylogeneticinterpretations Nevertheless in comparison with both extanthominoids and the small-bodied catarrhines from the earlyMiocene it is informative to differentiate the evolutionary grades ofthese genera Many characteristics of Ekembo are more advanced inthe direction of modern apes compared to those of Proconsul(Table 2) General trends such as expanded and more bunodontcusps broader molar crests reduced cingulum and morphologicalsimilarity between P3 and P4 all represent morphologies thatpresage those of later fossil and extant hominoids The moreadvanced morphologies of P3 and M2 are particularly conspicuouscompared not only with those of Proconsul but to most earlyMiocene catarrhines In contrast the mesially positioned stronglyheteromorphic cusps on the P3 of Proconsul plus the correspondingasymmetric buccal profile notched mesial margin and missingmesial fovea are characteristic of other early Miocene catarrhinessuch as Dendropithecus Rangwapithecus and Limnopithecus andare likely primitive among East African catarrhines The moresymmetric P3 of Ekembo with midline positioned cusps andreduced cusp heteromorphy is reminiscent of later hominoids and
Figure 14 Relative buccolingual dimensions of M1 and M2 KNM-SO 1112 has a preserved M2 but only the cervix of M1 Its M1 breadth was estimated based on the ratio to cervixbreadth to maximum crown breadth in other Tinderet fossils Even the cervical breadth of SO 1112 is broader than would be predicted for the crown breadth of a Kisingiri M1 Trendlines computed using OLS regression and using the average estimate of SO 1112 M1 breadth Group differences were significant (p frac14 00002)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 57
even hominid taxa such as Rudapithecus (eg Kordos and Begun2001)
Proconsul is likewise united by a suite of features that are for themost part characteristic only of this lineage (Table 2) Themorphology of both upper and lower canines is unique to thisgenus and shared by all three well-known species plus the poorlyrepresented middle Miocene P gitongai (Pickford and Kunimatsu2005) The mandibular symphysis is also distinctive in Proconsulcompared with other catarrhines with its very long and shallowlysloping planum alveolare and steeply inclined subplanum surfaceIndeed even the small edentulous anterior mandibular fragmentKNM-SO 372 is readily placed within Proconsul and excluded fromother similarly sized catarrhine taxa such as Rangwapithecus andEkembo based on this unique symphyseal morphology (Figs 8 and9)
Other differences between Ekembo and Proconsul are of uncer-tain evolutionary polarity (eg orientation of the posthypoconecrista) or may be of taxonomic relevance without broader phylo-genetic significance (eg degree of molar flare Table 2) Werecognize that there are morphological differences within Procon-sul particularly between the better known species P africanus andP major However many of these differences are likely related tosize and represent variations on a single generic theme that aremarkedly different from the morphology of Ekembo The recentdescription (Harrison and Andrews 2009) of the intermediatelysized P meswae underscores the broad morphological unity withinProconsul and further substantiates a well-defined Proconsul cladecomprising Tinderet and Ugandan species
Based on the abundant morphological and morphometric evi-dence supporting genus-level distinction between Proconsul andEkembo we are currently conducting a more detailed phylogeneticanalysis of all early Miocene catarrhines in order to better under-stand the distribution and evolutionary significance of these andother morphological features We anticipate that the debates sur-rounding the phylogenetic relationships of an historically consti-tuted sample of Proconsul can be more satisfactorily resolved nowthat the Kisingiri material is appropriately recognized as a distinctgenus
The generic disposition of ldquoUgandapithecusrdquo species
A great many features said to characterize ldquoUgandapithecusrdquowere added to the original diagnosis of the genus by Pickford et al(2009) in their systematic revision of Proconsul (sensu lato) As partof that study they also erected a new species ldquoUrdquo legetetensis andtransferred P meswae to ldquoUgandapithecusrdquo Evaluating the same setof features across the entire available sample however we wereunable to find support for this more expansive conception ofldquoUgandapithecusrdquo for several reasons First some diagnostic fea-tures cited in Pickford et al (2009) were quite variable within theldquoUgandapithecusrdquo sample As an example marked reduction of theM3 entoconid to a bead on the lingual cingulum is not shared byseveral ldquoUrdquo major specimens such as KNM-CA 393 NAP I 49000and Kor 253004 (see Pickford et al 2009 Fig 33e) and was likelynot a feature of a few of the more worn specimens eg BUMP 600and BUMP 601 or even the holotype of P major M 16648 Second
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6158
some diagnostic features of ldquoUgandapithecusrdquo were found to bevariably present in Ekembo arguing against their utility for dis-tinguishing ldquoUgandapithecusrdquo from Pickford et als (2009) com-bined P africanus thorn Kisingiri conception of Proconsul For examplesignificantly reduced curvature of the I1 distal margin in ldquoUganda-pithecusrdquo is also found in some Kisingiri specimens including KNM-RU 1685 1712 1714 1831 1846 and 1951 several more Rusingaspecimens have moderately reduced curvature Third some fea-tures that purportedly distinguish ldquoUgandapithecusrdquo fromP africanus are likely related to size For example the somewhatmore central position of the P3 paracone in ldquoUrdquomajor is not seen inthe smaller ldquoUrdquo meswae whose paracone is positioned similarly tothat of the holotype of P africanus (see Harrison and Andrews2009 Fig 6d)
However the major part of the evidence for recognizingldquoUgandapithecusrdquo (sensu Pickford et al 2009) was assembled by re-assigning specimens traditionally attributed to P africanus into newspecies of ldquoUgandapithecusrdquo The P africanus holotype was left inassociation with the Rusinga sample despite having morphologyoverwhelmingly characteristic of other Tinderet specimens Theresulting specimen allocations left P africanus morphologicallyimpoverisheddstripped of much of its hypodigmdso that Pro-consul for Pickford et al (2009) came to be characterized primarilyby the Rusinga sample (see similar comments in Harrison andAndrews 2009) Within that conceptual framework any spec-imen that differs from those at Rusinga could not be Proconsul butmust instead belong to ldquoUgandapithecusrdquo Thus specimens such asthe KNM-SO 1112 mandible were erroneously assigned to ldquoUgan-dapithecusrdquo because they were (correctly) perceived as differingfrom their Rusinga counterparts (Pickford et al 2009)
Here we restore the P africanus hypodigm and associate it withthe other Tinderet fossils according to their shared morphologySpecimens assigned to ldquoUgandapithecusrdquo revert to Proconsul andmany of the features cited to support a speciose ldquoUgandapithecusrdquo(Pickford et al 2009) instead become diagnostic features of Pro-consul This correction serves to highlight the distinctiveness of theKisingiri remains and the need to recognize this distinctiveness atthe generic level
As demonstrated above specimens from Kisingiri do not displaythe morphology of Proconsul as represented by the holotype ofP africanus M 14084 Given that this specimen preserves only theupper canine and cheek teeth similarities with or departures fromthis morphology represent the most compelling arguments forassessing inclusivity in the genus However for the mandible andlower dentition if one simply sorts the specimens by elementwithout relying on taxonomic preconceptions the overwhelmingdistinction within the traditionally constituted Proconsul sample isbetween Kisingiri material on the one hand and that from Tinderetand Uganda on the other (cf Walker et al 1993 Begun 2001 2007MacLatchy and Rossie 2005 Harrison and Andrews 2009Harrison 2010) Additionally with the Kisingiri specimensremoved to Ekembo the postcranial differences used to partlyjustify a distinct ldquoUgandapithecusrdquo lose their relevance (seeHarrison and Andrews 2009)
We have differed among ourselves in our past treatment ofP major as generically distinct from the smaller species of Pro-consul (Begun 2007 McNulty 2010) Morphologically many ofthe unique features of P major can be interpreted as size-relatedvariants on the smaller P africanus morphology Even if so thequestion is whether these are sufficiently different and numerousto warrant generic distinction that is do they suggest thatP major belongs in a separate clade from P africanus andP meswae As we have demonstrated differences among thesethree species are certainly fewer and show less morphologicaldistinction than those between Ekembo and any of the three
Nevertheless while we ultimately agree with Harrison andAndrews (2009 see also Begun 2007) that the features dis-tinguishing ldquoUrdquo major from P africanus are consistent with thosethat distinguish congeneric species and are at least partly due todifferences in size we concede the possibility that the largestProconsul species could belong in a separate genus (Senut et al2000 Pickford and Kunimatsu 2005) However whether or notone recognizes a separate genus for P major the P africanus ma-terial does not sort morphologically with the Kisingiri materialTherefore the taxonomic alternatives are either two generaProconsul and Ekembo or three Proconsul Ugandapithecus andEkembo with Proconsul and Ugandapithecus united in a cladeexclusive of Ekembo Thus while we strongly favor the firstalternative recognizing Ugandapithecus does not change thefundamental argument concerning the need to refer the Kisingirispecies to a generically and cladistically distinct taxon from thespecies represented at Tinderet and the Ugandan sites
Specimens of interest
Despite the substantial dental and gnathic differences betweenProconsul and Ekembo there are a few specimens whose taxonomicaffinities require additional scrutiny
KNM-RU 1676-77 This set of associated upper and lower teeth is aconsistent outlier among E heseloni and E nyanzae specimens Insome ways this specimen more closely resembles Tinderet Pro-consul in the features that distinguish it from Ekembo Molar cuspmorphology and cingulum development I1 lingual tubercledevelopment and M2 shape and posthypocone crista orientationare more consistent with specimens from the Tinderet sites On theother hand it otherwise resembles Ekembo and lacks the strongestdiagnostic features that unite species of Proconsul such as thedistinctive canine and P3 morphology Overall this specimen doesnot fit unambiguously into either genus in fact the degree ofbuccolingual flare cusp morphology enamel wrinkling and espe-cially the long extension of enamel onto the anterior root in P3 arein some ways reminiscent of Afropithecus although it clearly differsfrom that genus as well (cf Rossie and MacLatchy 2013)
Were these characteristics individually distributed among thelarge Kisingiri sample one might attribute this to intra-genericvariation but when found together in a single specimen from alarge sample it suggests a unique individual One of us (KPM)thinks it likely that the specimen was not actually found at aKisingiri locality based on its unique preservation compared toother Rusinga fossils and on other examples of mis-proveniencedspecimens (Leakey 1967 Andrews and Molleson 1979 Pickford1986 Kelley 1986) However we recognize that relying toofrequently on this explanation allows almost any scenario to bejustified and suggest some alternative possibilities One is that thespecimen was found on Rusinga but in the much older WayandoFormation strata instead of the Hiwegi Formation The differencesin morphology in that case might reflect more than a million yearsof temporal change (McCollum et al 2013) with KNM-RU 1676-77preserving certain more primitive Proconsul-like features As analternative it is reasonable to assume that fossil assemblages asvast as those from Rusingawill include not only abundant taxa butalso rare or infrequent inhabitants of the local biome There aremany examples of this from Rusingas mammalian communityeg ldquoHemicyonrdquo Kulutherium Tadarida cf Hyainailouros cfArchaeobeledon Kelba (see relevant chapters in Werdelin andSanders 2010) As such KNM-RU 1676-77 may simply representa rare and well-preserved specimen of some other catarrhinetaxon that lived among or visited a primate community dominatedby Ekembo
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 59
KNM-SO 1112 This specimen was initially referred to Rangwapi-thecus but soon after was transferred to Proconsul afrianus (Bosler1981 Martin 1981)da designation that most subsequent authorshave followed (Pickford 1986 Nengo and Rae 1992 Kelley 19861995 Walker et al 1993 MacLatchy and Rossie 2005 Hill et al2013) Pickford et al (2009) more recently referred this specimento ldquoUrdquo meswae citing two primary reasons First they reiteratedBoslers (1981) concern that its P3 differs from others attributedto P africanus However at that time the majority of what wasconsidered to be P africanus would have been from RusingaIsland In fact while noting its contrasting morphology Boslersuggested that differences between KNM-SO 1112 and other P3smight reasonably be attributed to sexual dimorphism notingthat this specimen provides ldquoa not implausible match for the Pafricanus holotype maxilla M 14084rdquo (1981152) Thatassessment is borne out in this study Using the much largersample of Ekembo P3s as a model variation can be seen toencompass specimens with well-developed mesial beaks andthose with more bilaterally compressed crowns and parallelsides The latter morphology is predominantly found amongmale specimens (KNM-RU 1674 1924 and KPS I and III) thussupporting Boslers (1981) suggestion that this variation isrelated to sexual dimorphism and hence that KNM-SO 1112 is amale P africanus
The second feature cited by Pickford et al (2009) in referringthis specimen to ldquoUrdquo meswae was posterior shallowing of themandibular corpus However because they redefined ldquoUgandapi-thecusrdquo to include all of the well-preserved Tinderet mandibles(Pickford et al 2009)dleaving Proconsul to be represented only byRusinga specimensdtheir only alternative within this scheme for aspecimen the size of SO 1112 was to refer it to ldquoUrdquomeswae It seemsodd though to assign this specimen to a species from a (perhapsmuch) older site when the Proconsul africanus holotype is ofappropriate size and similar age and as there are no other adultmandibles from either species with which to compare it In anycase mandibular shallowing is variable in both Ekembo and Pro-consul with some Kisingiri specimens exhibiting considerableshallowing (eg KNM-RU 2087) and some ldquoUgandapithecusrdquo spec-imens having very little (eg M 14086)
We therefore see no morphological grounds to assign KNM-SO1112 to P meswae but do find some morphological evidence toassign it as a male P africanus which occurs at Songhor
KNM-LG 452 This very complete mandible was originally recog-nized as a female of P major based on the size of the dentition androbusticity of the mandible (Martin 1981) Bosler (1981) howeverleft this specimen unassigned noting similarities to P major in P3-M1 size but having distinctly smaller M2e3 In this she likenedKNM-LG 452 to a few other specimens (notably M 14086) thatare smaller in their distal molars in a manner more consistentwith taxonomic rather than sex differences (Bosler 1981 see alsoPilbeam 1969) Kelley (1986 1995) also assigned this specimen asa female of P major based on canine size and morphology butnoted that the canine lacked the shallowly-sloping mesial ridgetypical of most female anthropoid canines This is also truehowever of two other canines assigned as female P major KO 9and SO 373 (Kelley 1986) and so might simply be an unusualcharacteristic of P major female lower canines In the end Kelley(1995) concluded that considering all of the criteria fordistinguishing male and female canines the evidence that KNM-LG 452 is female remains questionable
Pickford et al (2009) further developed the idea that two spe-cies are represented within the P major sample and made KNM-LG452 the holotype of a new species ldquoUrdquo legetetensis We concur thatthis specimen is unique among the well-preserved Tinderet
mandibles in some important features Whereas male and femalemandibles of P major have long internal symphyseal planesexpansive extra- and retromolar sulci and rami that cross theocclusal plane at distal M3 KNM-LG 452 has a shorter symphysealplane (though not to the extent seen in Ekembo) a moderateextramolar sulcus but no retromolar sulcus and a ramus thatcrosses the occlusal plane more mesially If KNM-SO 396 and SO404 represent males and females respectively of P major then themorphology of KNM-LG 452 appears distinct
At present however we do not think there is sufficient evidenceto differentiate this specimen from either P major or P meswaePickford et al (2009) argue that KNM-LG 452 cannot belong to thelatter species because of its larger size The size differences how-ever are well within the range of variation seen in P majorE heseloni and E nyanzae Moreover whereas the Legetet mandibleis larger than most of the P meswae hypodigm an adult P4 referredto this species (KNM-ME 25) substantially exceeds its counterpartin KNM-LG 452 Should the Legetetmandible ultimately prove to bea male specimen size alone would not rule out its attribution toP meswae On the other hand if it is female then it becomes harderto justify its inclusion in that sample of mostly smaller specimens Iffemale therefore it could represent a separate species for whichthe binomial Proconsul legetetensis comb nov is available Recentlycollected fossils from the SonghorKapurtay area provide someadditional support for the possibility that three species of Proconsulexisted at these localities (McNulty 2014ab) However toconvincingly demonstrate this and determine whether a thirdspecies can be differentiated from P meswae will require a moredetailed analysis of variation among Proconsul species a corre-sponding study of variation in modern taxa anddalmost certain-lydadditional fossil specimens For now we regard ldquoUrdquo legetetensisas a subjective junior synonym of P major
Conclusions
The genus Proconsul has been central to studies of ape and hu-man evolution since its first description by Hopwood (1933) As oneof the best known fossil catarrhines represented by nearly everyskeletal element our knowledge of Proconsuls functional anatomypositional behavior and overall morphology has made it thefoundation for paleoanthropological research in the early Mioceneas well as studies on the origin and early diversification of homi-noids With respect to systematics however Proconsul is less wellunderstood the type species P africanus is known from a rela-tively small number of specimens from sites in the Tinderet areaand no postcranial bones can be unambiguously assigned to itRather it is the copious and well-preserved specimens from theKisingiri sites of Rusinga and Mfangano Islands that have formedthe basis for the common image of Proconsul
We argue here that the Kisingiri species traditionally referredto ProconsuldP heseloni and P nyanzaedare generically distinctfrom P africanus and hereby transfer them into a new genusEkembo based on a substantial number of morphological differ-ences (Table 2) Considering only those traits preserved in theP africanus holotype the entire sample from Rusinga and Mfan-gano differs markedly in the morphology of the upper canine P3P4 and M2 Additional differences in the I1 lower incisors andcanines molar cusp and occlusal crest morphology degree of flarein all molars and especially mandibular features clearly distin-guish the Kisingiri from non-Kisingiri samples in a way that isincompatible with congeneric species With the transfer ofE heseloni and E nyanzae Proconsul becomes a well-defined cladecomprising at least three speciesdP africanus P major andP meswaedunited by several features that are unique for thegenus (Table 2)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6160
We found no support in this study for the genus ldquoUgandapi-thecusrdquo as constructed by Pickford et al (2009) In that paper theytransferred the vast majority of Proconsul fossils from the Tinderetand Napak localities into ldquoUgandapithecusrdquo leaving P africanusrepresented only by its holotype and a small number of isolatedteeth and aligning it with the material from Rusinga and Mfan-ganoWhen the P africanus hypodigm is reconstituted and properlyaffiliated with the other Tinderet species according to itsmorphology much of the evidence marshaled by Pickford et al(2009) for ldquoUgandapithecusrdquo actually supports the distinction be-tween the Tinderet and Kisingiri samples ie the generic distinc-tion of Ekembo In addition some of the traits enumerated byPickford and colleagues in support of ldquoUgandapithecusrdquo wererevealed in our analysis to be variable within one or more speciesand therefore of questionable taxonomic utility We do not rejectout of hand the original conception of ldquoUgandapithecusrdquo (Senutet al 2000) as either a monotypic genus distinguishing ldquoUrdquo ma-jor or perhaps including a closely allied middle Miocene ldquoUrdquogitongai as well (Pickford and Kunimatsu 2005) but we have fornow retained the early Miocene specimens of both in Proconsulpending further study However even if Ugandapithecus is a taxo-nomically valid genus comprising just the latter two species theirmorphology indicates that theywould still be united in a cladewithProconsul and they lack several more advanced features that partlydefine Ekembo Better understanding of the role that body size playsinmediating differences within Proconsulwill help to further clarifythe relationships among these species
Despite the existence of one well-represented and several lesscomplete skeletons of what was long assumed to be Proconsul thephylogenetic status of this genus has defied consensus This islargely due in our view to Proconsul having been an admixture oftwo distinct genera one more advanced in the direction of extantapes than the other As we demonstrate here the material fromRusinga and Mfangano traditionally referred to Proconsul andwhich includes all of the more complete skeletal material isdistinct from the Tinderet and Napak material and belongs in aseparate genus Ekembo By disentangling these distinct clades weanticipate being able to achieve better phylogenetic resolution forboth genera thereby improving our knowledge of early hominoidevolution and diversification
Acknowledgments
We thank many people and agencies that assisted with thedevelopment and completion of this project This research wasinitiated as a result of funding from the Wenner-Gren Foundationand the Johns Hopkins School of Medicine to DR Begun in1987e1989 and was renewed following research and travelgenerously funded by multiple Leakey Foundation grants to KMcNulty National Science Foundation grants to K McNulty(0852609 1241807) the McKnight Foundation the University ofMinnesota the Lloyd A Wilford Endowment the LeverhulmeFoundation and the Turkana Basin Institute J Kelley gratefullyacknowledges support from the Institute of Human Origins Ari-zona State University DR Begun also wishes to acknowledge theAlexander Humboldt Stiftung and NSERC for funding support Weare grateful to the National Museums of Kenya the UgandaMuseum the Natural History Museum and to Rose Mwanja EzraMusiime Tom Mukhuyu Samuel Muteti James Yatich and MaryMuungu for access to and assistance with specimens in their careMany people provided assistance comments criticisms insightsand suggestions on various aspects of this work and we especiallythank Holly Dunsworth Will Harcourt-Smith Tom Lehmann DanPeppe Nicole Garrett Kirsten Jenkins Rutger Jansma Susy CoteIsaiah Nengo Andrew Hill and Brenda Benefit K McNulty
acknowledges the evolutionary anthropology faculty and studentsat Durham University for their helpful comments on an earlyversion of this manuscript and for their gracious hospitality Weare likewise grateful to the anonymous reviewers associate editorand editor whose comments strengthened the paper This manu-script is publication 1 supporting Research on East AfricanCatarrhine and Hominoid Evolution (REACHE)
Appendix A Supplementary data
Supplementary data related to this article can be found at httpdxdoiorg101016jjhevol201503009
References
Andrews P 1974 New species of Dryopithecus from Kenya Nature 249 188e190Andrews P 1978 A revision of the Miocene Hominoidea of East Africa Bull Brit
Mus (Nat Hist) Geol 30 85e225Andrews P 1985 Family group systematics and evolution among catarrhine pri-
mates In Delson E (Ed) Ancestors The Hard Evidence Liss New Yorkpp 14e22
Andrews P Harrison T Martin L Pickford M 1981 Hominoid primates from anew Miocene locality named Meswa Bridge in Kenya J Hum Evol 10123e128
Andrews P Begun D Zylstra M 1997 Interrelationships between functionalmorphology and paleoenvironments in Miocene hominoids In Begun DWard C Rose M (Eds) Function Phylogeny and Fossils Miocene HominoidEvolution and Adaptations Plenum Press New York pp 29e58
Andrews P Molleson T 1979 The provenance of Sivapithecus africanus Bull BritMus (Nat Hist) Geol 32 19e23
Begun DR 2001 African and Eurasian Miocene hominoids and the origins of theHominidae In de Bonis L Koufos GD Andrews P (Eds) Hominoid Evolutionand Climatic Change in Europe Phylogeny of the Neogene Hominoid Primatesof Eurasia Cambridge University Press London pp 231e253
Begun DR 2007 Fossil record of Miocene hominoids In Henke W Tattersall I(Eds) Handbook of Paleoanthropology Springer Berlin pp 921e977
Begun DR Teaford MF Walker AC 1994 Comparative and functional anatomyof Proconsul phalanges from the Kaswanga primate site Rusinga Island KenyaJ Hum Evol 26 89e165
Bishop WW 1964 More fossil primates and other Miocene mammals from North-East Uganda Nature 203 1327e1331
Bishop WW Miller JA Fitch FJ 1969 New potassium-argon age determinationsrelevant to the Miocene fossil mammal sequence in East Africa Am J Sci 267669e699
Blankenship JA Mincer HH Anderson KM Woods MA Burton EL 2007Third molar development in the estimation of chronologic age in Americanblacks as compared with whites J Forensic Sci 52 428e433
Bosler W 1981 Species groupings of early Miocene dryopithecine teeth from EastAfrica J Hum Evol 10 151e158
Coleman MN 2008 What does geometric mean mean geometrically Assessingthe utility of geometric mean and other size variables in studies of skullallometry Am J Phys Anthropol 135 404e415
Cote S Malit N Nengo I 2013 Additional mandibles of Rangwapithecus gordonian early Miocene catarrhine from the Tinderet localities of Western KenyaJ Hum Evol 153 341e352
Deane AS 2009 Early Miocene catarrhine dietary behavior the influence of theRed Queen Effect on incisor shape and curvature J Hum Evol 56 275e285
Drake R Van Couvering JA Pickford M Curtis G Harris JA 1988 New chro-nology for the Early Miocene mammalian faunas of Kisingiri Western KenyaJ Geol Soc Lond 145 479e491
Evernden JF Savage DE Curtis GH James GT 1964 Potassium-argon datesand the Cenozoic mammalian chronology of North America Am J Sci 262145e198
Gebo DL MacLatchy L Kityo R Deino A Kingston J Pilbeam D 1997A hominoid genus from the Early Miocene of Uganda Science 276 401e404
Geoffroy Saint-Hilaire E 1812 Tableau des quadrumanes 1 Ord QuadrumanesAnnls Mus Hist Nat Paris 19 85e122
Gray JE 1825 Outline of an attempt at the disposition of the Mammalia into tribesand families with a list of the genera apparently appertaining to each tribeAnnls Philos 10 337e344
Harrison T 1982 Small-bodied apes from the Miocene of East Africa PhDDissertation University of London
Harrison T 1986 New fossil anthropoids from the middle Miocene of East Africaand their bearing on the origin of the Oreopithecidae Am J Phys Anthropol 71265e284
Harrison T 1987 The phylogenetic relationships of the early catarrhine primates areview of the current evidence J Hum Evol 16 41e80
Harrison T 2002 Late Oligocene to middle Miocene catarrhines from Afro-ArabiaIn Hartwig WC (Ed) The Primate Fossil Record Cambridge University PressCambridge pp 311e338
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 61
Harrison T 2010 Dendropithecoidea Proconsuloidea and Hominoidea (CatarrhiniPrimates) In Werdelin L Sanders WJ (Eds) Cenozoic Mammals of AfricaUniversity of California Press Berkeley pp 429e469
Harrison T Andrews P 2009 The anatomy and systematic position of a newspecies of early Miocene proconsulid from Meswa Bridge Kenya J Hum Evol56 479e496
Hill A Nengo IO Rossie JB 2013 A Rangwapithecus gordoni mandible from theearly Miocene site of Songhor Kenya J Hum Evol 65 490e500
Hopwood AT 1933 Miocene primates from Kenya J Linn Soc Zool 38 437e464Ishida H Kunimatsu Y Takano T Nakano Y Nakatsukasa M 2004 Nacholapi-
thecus skeleton from the Middle Miocene of Kenya J Hum Evol 46 69e103Jolicouer P 1963 The multivariate generalization of the allometry equation Bio-
metrics 19 497e499Jungers WL Falsetti AB Wall CE 1995 Shape relative size and size-
adjustments in morphometrics Am J Phys Anthropol 38 (S2) 137e161Kavanagh KD Evans AR Jernvall J 2007 Predicting evolutionary patterns of
mammalian teeth from development Nature 449 427e433Kelley J 1986 Species recognition and sexual dimorphism in Proconsul and
Rangwapithecus J Hum Evol 15 461e495Kelley J 1995 Sex determination in Miocene catarrhine primates Am J Phys
Anthropol 96 390e417Kelley J 1993 Taxonomic implications of sexual dimorphism in Lufengpithecus In
Kimbel WH Martin LB (Eds) Species Species Concepts and Primate Evo-lution Springer New York pp 429e458
Kent PE 1942 The country round the Kavirondo Gulf of Victoria Nyanza Geogr J100 22e31
Kordos L Begun DR 2001 A new cranium of Dryopithecus from RudabanyaHungary J Hum Evol 15 689e700
Le Gros Clark WE Leakey LSB 1950 Diagnoses of East African Miocene Homi-noidea Q J Geol Soc Lond 105 260e263
Le Gros Clark WE Leakey LSB 1951 The Miocene Hominoidea of East Africa InFossil Mammals of Africa No 1 British Museum (Natural History) London
Leakey LSB 1943 A Miocene anthropoid mandible from Rusinga Kenya Nature152 319e320
Leakey LSB 1963 East African fossil Hominoidea and the classification within thissuper-family In Washburn SL (Ed) Classification and Human EvolutionAldine Chicago pp 32e49
Leakey LSB 1967 Notes on the mammalian faunas from the Miocene and Pleis-tocene of East Africa In Bishop WW Clark JD (Eds) Background to Evolu-tion in Africa University of Chicago Press Chicago pp 7e29
Leakey RE Leakey MG 1986 A new Miocene hominoid from Kenya Nature 324143e146
Leakey RE Leakey MG Walker AC 1988 Morphology of Afropithecus turka-nensis from Kenya Am J Phys Anthropol 76 289e307
Leakey MG Ungar PS Walker A 1995 A new genus of large primate from theLate Oligocene of Lothidok Turkana District Kenya J Hum Evol 28519e531
Linnaeus C 1758 Systema naturae per regna tria naturae secundum classesordines genera species cum characteribus differentris synonymis locis tenthed Laurentii Salvii Stockholm
MacInnes DG 1943 Notes on the East African Miocene primates J E Afr UgandaNat Hist Soc 17 141e181
MacLatchy L 2004 The oldest ape Evol Anthropol 13 90e103MacLatchy L Bossert WH 1996 An analysis of the articular surface distribution of
the femoral head and acetabulum in anthropoids with implications for hipfunction in Miocene hominoids J Hum Evol 31 425e453
MacLatchy L Rossie JB 2005 The Napak hominoid still Proconsul major InLieberman DE Smith RJ Kelley J (Eds) Interpreting the Past Essays onHuman Primate and Mammal Evolution in Honor of David Pilbeam Brill Ac-ademic Publishers Boston pp 15e28
Martin L 1981 New specimens of Proconsul from Koru Kenya J Hum Evol 10139e150
McCollum MS Peppe DJ McNulty KP Dunsworth HM Harcourt-Smith WEH Andrews AL 2013 Magnetostratigraphy of the early MioceneHiwegi Formation (Rusinga Island Lake Victoria Kenya) Geological Society ofAmerica Abstracts with Programs 45 p 12
McNulty KP 2010 Apes and tricksters the evolution and diversification ofhumans closest relatives Evol Edu Outreach 3 322e332
McNulty KP Harcourt-Smith WEH Dunsworth HM 2007 New primate fossilsfrom Rusinga Island Kenya Am J Phys Anthropol 132 (S44) 170
McNulty KP MacLatchy L Peppe DJ Nengo I Manthi FK Miller ERStevens NJ Cote S Lehmann T Kingston J 2014a A regional approach toEast African early Miocene paleobiology J Vert Paleontol 185 SVP Programand Abstracts Book 2014
McNulty KP MacLatchy L Rossie JB Peppe DJ Deino AL Mbua ENManthi FK Nengo IO Stevens NJ Cote S Lehmann T Gutierrez M 2014b
Research on East African catarrhine and hominoid evolution results from thefirst year Am J Phys Anthropol 153 (S58) 182
Michel LA Peppe DJ Lutz JA Driese SG Dunsworth HM Harcourt-Smith WEH Horner W Lehmann T Nightengale S McNulty KP 2014Remnants of an ancient forest provide ecological context for Early Miocenefossil apes Nature Comm 5 3236
Myres JL 1932 Africa Man 32 208Nakatsukasa M Kunimatsu Y Nakano Y Takano T Ishida H 2007 Vertebral
morphology of Nacholapithecus kerioi based on KNM-BG 35250 J Hum Evol52 347e369
Nengo IO Rae TC 1992 New hominoid fossils from the early Miocene site ofSonghor Kenya J Hum Evol 23 423e429
Peppe DJ McNulty KP Cote SM Harcourt-Smith WEH Dunsworth HM VanCouvering JA 2009 Stratigraphic interpretation of the Kulu Formation (EarlyMiocene Rusinga Island Kenya) and its implications for primate evolutionJ Hum Evol 56 447e461
Pickford M 1981 Preliminary Miocene mammalian biostratigraphy for westernKenya J Hum Evol 10 73e97
Pickford M 1986 Sexual dimorphism in Proconsul Hum Evol 1 111e148Pickford M Andrews PJ 1981 The Tinderet Miocene sequence in Kenya J Hum
Evol 10 11e33Pickford M Kunimatsu Y 2005 Catarrhines from the Middle Miocene (ca 145
Ma) of Kipsaraman Tugen Hills Kenya Anthropol Sci 113 189e224Pickford M Senut B Gommery D Musiime E 2009 Distinctiveness of Ugan-
dapithecus from Proconsul Estudios Geol 65 183e241Pilbeam DR 1969 Tertiary Pongidae of East Africa evolutionary relationships and
taxonomy Bulletin of the Peabody Museum of Natural History 31 pp 1e185Rae TC 1993 Phylogenetic analysis of proconsulid facial morphology PhD
Dissertation State University of New York at Stony BrookRae TC 1999 Mosaic evolution in the origin of the Hominoidea Folia Primatol 70
125e135Rafferty KL Walker A Ruff C Rose MD Andrews PJ 1995 Postcranial esti-
mates of body weights in Proconsul with a note on a distal tibia of P major fromNapak Uganda Am J Phys Anthropol 97 391e402
Rossie JB MacLatchy L 2013 Dentognathic remains of an Afropithecus individualfrom Kalodirr Kenya J Hum Evol 65 199e208
Senut B Pickford M Gommery D Kunimatsu Y 2000 Un nouveau genredrsquohominoiumlde du Miocene inferieur drsquoAfrique orientale Ugandapithecus major(Le Gros Clark and Leakey 1950) CR Acad Sci Paris 331 227e233
Simons EL Pilbeam DR 1965 Preliminary revision of the Dryopithecinae (Pon-gidae Anthropoidea) Folia Primatol 3 81e152
Smith TM Martin LB Leakey MG 2003 Enamel thickness microstructure anddevelopment in Afropithecus turkanensis J Hum Evol 44 283e306
Stevens NJ Seiffert ER OConnor PM Roberts EM Schmitz MD Krause CGorscak E Ngasala S Hieronymus TL Temu J 2013 Palaeontological evi-dence for an Oligocene divergence between Old World monkeys and apesNature 497 611e614
Suwa G Kono RT Katoh S Asfaw B Beyene Y 2007 A new species of great apefrom the late Miocene epoch in Ethiopia Nature 448 921e924
Szalay FS Delson E 1979 Evolutionary History of the Primates Academic PressNew York
Teaford MF Beard KC Leakey RE Walker A 1988 New hominoid facial skel-eton from the Early Miocene of Rusinga Island Kenya and its bearing on therelationship between Proconsul nyanzae and Proconsul africanus J Hum Evol17 461e477
Van Couvering JAH Miller J 1969 Miocene stratigraphy and age determinationsRusinga Island Kenya Nature 221 628e632
Walker A 1997 Proconsul function and phylogeny In Begun DR Ward CVRose MD (Eds) Function Phylogeny and Fossils Miocene Hominoid Evolu-tion and Adaptation Plenum Press New York pp 209e224
Walker A Pickford M 1983 New postcranial fossils of Proconsul africanus andProconsul nyanzae In Ciochon RL Corruccini RS (Eds) New Interpretationsof Ape and Human Ancestry Plenum Press New York pp 325e351
Walker A Teaford M 1988 The Kaswanga primate site an Early Miocene hom-inoid site on Rusinga Island Kenya J Hum Evol 17 539e544
Walker A Teaford M 1989 The hunt for Proconsul Sci Am 260 76e82Walker A Teaford MF Martin L Andrews P 1993 A new species of Proconsul
from the early Miocene of RusingaMfangano Islands Kenya J Hum Evol 2543e56
Ward CV Ruff CB Walker A Rose MD Teaford MF Nengo IO 1995 Func-tional morphology of Proconsul patellas from Rusinga Island Kenya with im-plications for other Miocene-Pliocene catarrhines J Hum Evol 29 1e19
Ward S Brown B Hill A Kelley J Downs W 1999 Equatorius a new hominoidgenus from the Middle Miocene of Kenya Science 285 1382e1386
Werdelin L Sanders WJ 2010 Cenozoic Mammals of Africa University of Cali-fornia Press Berkeley

Figure 13 Box-and-whisker plot of buccolingual flare in lower molars measured asthe average distance between the mesial buccal and lingual cusps divided by overallbuccolingual crown breadth Differences between Ekembo and Proconsul are highlysignificant (p lt 00001)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6156
Relative molar size has been used to distinguish species ofProconsul but we observed the primary difference to be in relativebuccolingual breadths of M1 and M2 In Ekembo M1 is substantiallynarrower than M2 than is the case in Proconsul (Fig 14 Harrison2002 2010) Although none of the smaller Proconsul mandiblespreserve complete crowns for both M1 and M2 KNM-SO 1112 hasan intact M2 and at least preserves the cervix for M1 However evenif the cervical measurement is used for its M1 breadthdthusunderestimating the true crown breadthdKNM-SO 1112 still has abroader first molar in relation to M2 than would be expected inEkembo If crown breadth is estimated from the ratio of cervixbreadth to maximum crown breadth in other Proconsul fossils therange of likely breadths for the SO 1112 M1 clearly distinguishes it
from the Ekembo sample (Fig 14) Testing this relationship onlogged variables using the mean estimate for the SO 1112 M1(Fig 14) reveals a significant difference between the two groups(ANCOVA df frac14 1 F frac14 2424 p frac14 00002)
As with upper third molars we find a great deal of variation inM3 morphology and mistrust the taxonomic significance some-times placed on characteristics of this toothdat least with regard tothese taxa Harrison (2002 2010) cited the mesiodistal position ofthe M3 entoconid as differentiating Kisingiri third molars fromthose attributed to P africanus We concur with his assessment butfound the entoconid position to vary in specimens attributed toP major Thus this character does not differentiate Ekembo from allProconsul species but may distinguish it from the type speciesP africanus
Discussion
The distinctiveness of the Kisingiri large catarrhine materialfrom that at Tinderet and Ugandan localities has been noted pre-viously by several authors (Pickford 1986 Walker et al 1993Begun 2001 2007 MacLatchy and Rossie 2005 Harrison andAndrews 2009 Harrison 2010) What had not been fully docu-mented was the nearly complete lack of overlap between thesesamples in a number of features despite very large assemblagesfrom Rusinga and Mfangano On descriptive criteria alone (Table 2)many isolated teeth (I1 C1 P3 M2 C1) and all of the mandibles canbe assigned to either Proconsul or Ekembowith very little chance ofmisattribution That some of the most diagnostic features occur inthe upper dentitiondthe only anatomy preserved in theP africanus type specimendadds weight to the taxonomicarrangement proposed here The morphology of the upper teeth inall Proconsul species compellingly unites them into a single cladedistinct from E heseloni and E nyanzae This is reinforced bynumerous features of the mandible and mandibular dentition thatfurther distinguish these samples The number and variety of thesefeatures are such that Proconsul and Ekembo are more differentfrom one another than are many closely-related modern catarrhinegenera
Derived morphology of Ekembo and Proconsul
It is premature to formally assign primitive and derived status tothe individual features of Ekembo and Proconsul before completinga formal phylogenetic analysis This is especially true given theabundance of evidence available for Ekembo and hence the poten-tial impact that our generic revision could have on phylogeneticinterpretations Nevertheless in comparison with both extanthominoids and the small-bodied catarrhines from the earlyMiocene it is informative to differentiate the evolutionary grades ofthese genera Many characteristics of Ekembo are more advanced inthe direction of modern apes compared to those of Proconsul(Table 2) General trends such as expanded and more bunodontcusps broader molar crests reduced cingulum and morphologicalsimilarity between P3 and P4 all represent morphologies thatpresage those of later fossil and extant hominoids The moreadvanced morphologies of P3 and M2 are particularly conspicuouscompared not only with those of Proconsul but to most earlyMiocene catarrhines In contrast the mesially positioned stronglyheteromorphic cusps on the P3 of Proconsul plus the correspondingasymmetric buccal profile notched mesial margin and missingmesial fovea are characteristic of other early Miocene catarrhinessuch as Dendropithecus Rangwapithecus and Limnopithecus andare likely primitive among East African catarrhines The moresymmetric P3 of Ekembo with midline positioned cusps andreduced cusp heteromorphy is reminiscent of later hominoids and
Figure 14 Relative buccolingual dimensions of M1 and M2 KNM-SO 1112 has a preserved M2 but only the cervix of M1 Its M1 breadth was estimated based on the ratio to cervixbreadth to maximum crown breadth in other Tinderet fossils Even the cervical breadth of SO 1112 is broader than would be predicted for the crown breadth of a Kisingiri M1 Trendlines computed using OLS regression and using the average estimate of SO 1112 M1 breadth Group differences were significant (p frac14 00002)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 57
even hominid taxa such as Rudapithecus (eg Kordos and Begun2001)
Proconsul is likewise united by a suite of features that are for themost part characteristic only of this lineage (Table 2) Themorphology of both upper and lower canines is unique to thisgenus and shared by all three well-known species plus the poorlyrepresented middle Miocene P gitongai (Pickford and Kunimatsu2005) The mandibular symphysis is also distinctive in Proconsulcompared with other catarrhines with its very long and shallowlysloping planum alveolare and steeply inclined subplanum surfaceIndeed even the small edentulous anterior mandibular fragmentKNM-SO 372 is readily placed within Proconsul and excluded fromother similarly sized catarrhine taxa such as Rangwapithecus andEkembo based on this unique symphyseal morphology (Figs 8 and9)
Other differences between Ekembo and Proconsul are of uncer-tain evolutionary polarity (eg orientation of the posthypoconecrista) or may be of taxonomic relevance without broader phylo-genetic significance (eg degree of molar flare Table 2) Werecognize that there are morphological differences within Procon-sul particularly between the better known species P africanus andP major However many of these differences are likely related tosize and represent variations on a single generic theme that aremarkedly different from the morphology of Ekembo The recentdescription (Harrison and Andrews 2009) of the intermediatelysized P meswae underscores the broad morphological unity withinProconsul and further substantiates a well-defined Proconsul cladecomprising Tinderet and Ugandan species
Based on the abundant morphological and morphometric evi-dence supporting genus-level distinction between Proconsul andEkembo we are currently conducting a more detailed phylogeneticanalysis of all early Miocene catarrhines in order to better under-stand the distribution and evolutionary significance of these andother morphological features We anticipate that the debates sur-rounding the phylogenetic relationships of an historically consti-tuted sample of Proconsul can be more satisfactorily resolved nowthat the Kisingiri material is appropriately recognized as a distinctgenus
The generic disposition of ldquoUgandapithecusrdquo species
A great many features said to characterize ldquoUgandapithecusrdquowere added to the original diagnosis of the genus by Pickford et al(2009) in their systematic revision of Proconsul (sensu lato) As partof that study they also erected a new species ldquoUrdquo legetetensis andtransferred P meswae to ldquoUgandapithecusrdquo Evaluating the same setof features across the entire available sample however we wereunable to find support for this more expansive conception ofldquoUgandapithecusrdquo for several reasons First some diagnostic fea-tures cited in Pickford et al (2009) were quite variable within theldquoUgandapithecusrdquo sample As an example marked reduction of theM3 entoconid to a bead on the lingual cingulum is not shared byseveral ldquoUrdquo major specimens such as KNM-CA 393 NAP I 49000and Kor 253004 (see Pickford et al 2009 Fig 33e) and was likelynot a feature of a few of the more worn specimens eg BUMP 600and BUMP 601 or even the holotype of P major M 16648 Second
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6158
some diagnostic features of ldquoUgandapithecusrdquo were found to bevariably present in Ekembo arguing against their utility for dis-tinguishing ldquoUgandapithecusrdquo from Pickford et als (2009) com-bined P africanus thorn Kisingiri conception of Proconsul For examplesignificantly reduced curvature of the I1 distal margin in ldquoUganda-pithecusrdquo is also found in some Kisingiri specimens including KNM-RU 1685 1712 1714 1831 1846 and 1951 several more Rusingaspecimens have moderately reduced curvature Third some fea-tures that purportedly distinguish ldquoUgandapithecusrdquo fromP africanus are likely related to size For example the somewhatmore central position of the P3 paracone in ldquoUrdquomajor is not seen inthe smaller ldquoUrdquo meswae whose paracone is positioned similarly tothat of the holotype of P africanus (see Harrison and Andrews2009 Fig 6d)
However the major part of the evidence for recognizingldquoUgandapithecusrdquo (sensu Pickford et al 2009) was assembled by re-assigning specimens traditionally attributed to P africanus into newspecies of ldquoUgandapithecusrdquo The P africanus holotype was left inassociation with the Rusinga sample despite having morphologyoverwhelmingly characteristic of other Tinderet specimens Theresulting specimen allocations left P africanus morphologicallyimpoverisheddstripped of much of its hypodigmdso that Pro-consul for Pickford et al (2009) came to be characterized primarilyby the Rusinga sample (see similar comments in Harrison andAndrews 2009) Within that conceptual framework any spec-imen that differs from those at Rusinga could not be Proconsul butmust instead belong to ldquoUgandapithecusrdquo Thus specimens such asthe KNM-SO 1112 mandible were erroneously assigned to ldquoUgan-dapithecusrdquo because they were (correctly) perceived as differingfrom their Rusinga counterparts (Pickford et al 2009)
Here we restore the P africanus hypodigm and associate it withthe other Tinderet fossils according to their shared morphologySpecimens assigned to ldquoUgandapithecusrdquo revert to Proconsul andmany of the features cited to support a speciose ldquoUgandapithecusrdquo(Pickford et al 2009) instead become diagnostic features of Pro-consul This correction serves to highlight the distinctiveness of theKisingiri remains and the need to recognize this distinctiveness atthe generic level
As demonstrated above specimens from Kisingiri do not displaythe morphology of Proconsul as represented by the holotype ofP africanus M 14084 Given that this specimen preserves only theupper canine and cheek teeth similarities with or departures fromthis morphology represent the most compelling arguments forassessing inclusivity in the genus However for the mandible andlower dentition if one simply sorts the specimens by elementwithout relying on taxonomic preconceptions the overwhelmingdistinction within the traditionally constituted Proconsul sample isbetween Kisingiri material on the one hand and that from Tinderetand Uganda on the other (cf Walker et al 1993 Begun 2001 2007MacLatchy and Rossie 2005 Harrison and Andrews 2009Harrison 2010) Additionally with the Kisingiri specimensremoved to Ekembo the postcranial differences used to partlyjustify a distinct ldquoUgandapithecusrdquo lose their relevance (seeHarrison and Andrews 2009)
We have differed among ourselves in our past treatment ofP major as generically distinct from the smaller species of Pro-consul (Begun 2007 McNulty 2010) Morphologically many ofthe unique features of P major can be interpreted as size-relatedvariants on the smaller P africanus morphology Even if so thequestion is whether these are sufficiently different and numerousto warrant generic distinction that is do they suggest thatP major belongs in a separate clade from P africanus andP meswae As we have demonstrated differences among thesethree species are certainly fewer and show less morphologicaldistinction than those between Ekembo and any of the three
Nevertheless while we ultimately agree with Harrison andAndrews (2009 see also Begun 2007) that the features dis-tinguishing ldquoUrdquo major from P africanus are consistent with thosethat distinguish congeneric species and are at least partly due todifferences in size we concede the possibility that the largestProconsul species could belong in a separate genus (Senut et al2000 Pickford and Kunimatsu 2005) However whether or notone recognizes a separate genus for P major the P africanus ma-terial does not sort morphologically with the Kisingiri materialTherefore the taxonomic alternatives are either two generaProconsul and Ekembo or three Proconsul Ugandapithecus andEkembo with Proconsul and Ugandapithecus united in a cladeexclusive of Ekembo Thus while we strongly favor the firstalternative recognizing Ugandapithecus does not change thefundamental argument concerning the need to refer the Kisingirispecies to a generically and cladistically distinct taxon from thespecies represented at Tinderet and the Ugandan sites
Specimens of interest
Despite the substantial dental and gnathic differences betweenProconsul and Ekembo there are a few specimens whose taxonomicaffinities require additional scrutiny
KNM-RU 1676-77 This set of associated upper and lower teeth is aconsistent outlier among E heseloni and E nyanzae specimens Insome ways this specimen more closely resembles Tinderet Pro-consul in the features that distinguish it from Ekembo Molar cuspmorphology and cingulum development I1 lingual tubercledevelopment and M2 shape and posthypocone crista orientationare more consistent with specimens from the Tinderet sites On theother hand it otherwise resembles Ekembo and lacks the strongestdiagnostic features that unite species of Proconsul such as thedistinctive canine and P3 morphology Overall this specimen doesnot fit unambiguously into either genus in fact the degree ofbuccolingual flare cusp morphology enamel wrinkling and espe-cially the long extension of enamel onto the anterior root in P3 arein some ways reminiscent of Afropithecus although it clearly differsfrom that genus as well (cf Rossie and MacLatchy 2013)
Were these characteristics individually distributed among thelarge Kisingiri sample one might attribute this to intra-genericvariation but when found together in a single specimen from alarge sample it suggests a unique individual One of us (KPM)thinks it likely that the specimen was not actually found at aKisingiri locality based on its unique preservation compared toother Rusinga fossils and on other examples of mis-proveniencedspecimens (Leakey 1967 Andrews and Molleson 1979 Pickford1986 Kelley 1986) However we recognize that relying toofrequently on this explanation allows almost any scenario to bejustified and suggest some alternative possibilities One is that thespecimen was found on Rusinga but in the much older WayandoFormation strata instead of the Hiwegi Formation The differencesin morphology in that case might reflect more than a million yearsof temporal change (McCollum et al 2013) with KNM-RU 1676-77preserving certain more primitive Proconsul-like features As analternative it is reasonable to assume that fossil assemblages asvast as those from Rusingawill include not only abundant taxa butalso rare or infrequent inhabitants of the local biome There aremany examples of this from Rusingas mammalian communityeg ldquoHemicyonrdquo Kulutherium Tadarida cf Hyainailouros cfArchaeobeledon Kelba (see relevant chapters in Werdelin andSanders 2010) As such KNM-RU 1676-77 may simply representa rare and well-preserved specimen of some other catarrhinetaxon that lived among or visited a primate community dominatedby Ekembo
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 59
KNM-SO 1112 This specimen was initially referred to Rangwapi-thecus but soon after was transferred to Proconsul afrianus (Bosler1981 Martin 1981)da designation that most subsequent authorshave followed (Pickford 1986 Nengo and Rae 1992 Kelley 19861995 Walker et al 1993 MacLatchy and Rossie 2005 Hill et al2013) Pickford et al (2009) more recently referred this specimento ldquoUrdquo meswae citing two primary reasons First they reiteratedBoslers (1981) concern that its P3 differs from others attributedto P africanus However at that time the majority of what wasconsidered to be P africanus would have been from RusingaIsland In fact while noting its contrasting morphology Boslersuggested that differences between KNM-SO 1112 and other P3smight reasonably be attributed to sexual dimorphism notingthat this specimen provides ldquoa not implausible match for the Pafricanus holotype maxilla M 14084rdquo (1981152) Thatassessment is borne out in this study Using the much largersample of Ekembo P3s as a model variation can be seen toencompass specimens with well-developed mesial beaks andthose with more bilaterally compressed crowns and parallelsides The latter morphology is predominantly found amongmale specimens (KNM-RU 1674 1924 and KPS I and III) thussupporting Boslers (1981) suggestion that this variation isrelated to sexual dimorphism and hence that KNM-SO 1112 is amale P africanus
The second feature cited by Pickford et al (2009) in referringthis specimen to ldquoUrdquo meswae was posterior shallowing of themandibular corpus However because they redefined ldquoUgandapi-thecusrdquo to include all of the well-preserved Tinderet mandibles(Pickford et al 2009)dleaving Proconsul to be represented only byRusinga specimensdtheir only alternative within this scheme for aspecimen the size of SO 1112 was to refer it to ldquoUrdquomeswae It seemsodd though to assign this specimen to a species from a (perhapsmuch) older site when the Proconsul africanus holotype is ofappropriate size and similar age and as there are no other adultmandibles from either species with which to compare it In anycase mandibular shallowing is variable in both Ekembo and Pro-consul with some Kisingiri specimens exhibiting considerableshallowing (eg KNM-RU 2087) and some ldquoUgandapithecusrdquo spec-imens having very little (eg M 14086)
We therefore see no morphological grounds to assign KNM-SO1112 to P meswae but do find some morphological evidence toassign it as a male P africanus which occurs at Songhor
KNM-LG 452 This very complete mandible was originally recog-nized as a female of P major based on the size of the dentition androbusticity of the mandible (Martin 1981) Bosler (1981) howeverleft this specimen unassigned noting similarities to P major in P3-M1 size but having distinctly smaller M2e3 In this she likenedKNM-LG 452 to a few other specimens (notably M 14086) thatare smaller in their distal molars in a manner more consistentwith taxonomic rather than sex differences (Bosler 1981 see alsoPilbeam 1969) Kelley (1986 1995) also assigned this specimen asa female of P major based on canine size and morphology butnoted that the canine lacked the shallowly-sloping mesial ridgetypical of most female anthropoid canines This is also truehowever of two other canines assigned as female P major KO 9and SO 373 (Kelley 1986) and so might simply be an unusualcharacteristic of P major female lower canines In the end Kelley(1995) concluded that considering all of the criteria fordistinguishing male and female canines the evidence that KNM-LG 452 is female remains questionable
Pickford et al (2009) further developed the idea that two spe-cies are represented within the P major sample and made KNM-LG452 the holotype of a new species ldquoUrdquo legetetensis We concur thatthis specimen is unique among the well-preserved Tinderet
mandibles in some important features Whereas male and femalemandibles of P major have long internal symphyseal planesexpansive extra- and retromolar sulci and rami that cross theocclusal plane at distal M3 KNM-LG 452 has a shorter symphysealplane (though not to the extent seen in Ekembo) a moderateextramolar sulcus but no retromolar sulcus and a ramus thatcrosses the occlusal plane more mesially If KNM-SO 396 and SO404 represent males and females respectively of P major then themorphology of KNM-LG 452 appears distinct
At present however we do not think there is sufficient evidenceto differentiate this specimen from either P major or P meswaePickford et al (2009) argue that KNM-LG 452 cannot belong to thelatter species because of its larger size The size differences how-ever are well within the range of variation seen in P majorE heseloni and E nyanzae Moreover whereas the Legetet mandibleis larger than most of the P meswae hypodigm an adult P4 referredto this species (KNM-ME 25) substantially exceeds its counterpartin KNM-LG 452 Should the Legetetmandible ultimately prove to bea male specimen size alone would not rule out its attribution toP meswae On the other hand if it is female then it becomes harderto justify its inclusion in that sample of mostly smaller specimens Iffemale therefore it could represent a separate species for whichthe binomial Proconsul legetetensis comb nov is available Recentlycollected fossils from the SonghorKapurtay area provide someadditional support for the possibility that three species of Proconsulexisted at these localities (McNulty 2014ab) However toconvincingly demonstrate this and determine whether a thirdspecies can be differentiated from P meswae will require a moredetailed analysis of variation among Proconsul species a corre-sponding study of variation in modern taxa anddalmost certain-lydadditional fossil specimens For now we regard ldquoUrdquo legetetensisas a subjective junior synonym of P major
Conclusions
The genus Proconsul has been central to studies of ape and hu-man evolution since its first description by Hopwood (1933) As oneof the best known fossil catarrhines represented by nearly everyskeletal element our knowledge of Proconsuls functional anatomypositional behavior and overall morphology has made it thefoundation for paleoanthropological research in the early Mioceneas well as studies on the origin and early diversification of homi-noids With respect to systematics however Proconsul is less wellunderstood the type species P africanus is known from a rela-tively small number of specimens from sites in the Tinderet areaand no postcranial bones can be unambiguously assigned to itRather it is the copious and well-preserved specimens from theKisingiri sites of Rusinga and Mfangano Islands that have formedthe basis for the common image of Proconsul
We argue here that the Kisingiri species traditionally referredto ProconsuldP heseloni and P nyanzaedare generically distinctfrom P africanus and hereby transfer them into a new genusEkembo based on a substantial number of morphological differ-ences (Table 2) Considering only those traits preserved in theP africanus holotype the entire sample from Rusinga and Mfan-gano differs markedly in the morphology of the upper canine P3P4 and M2 Additional differences in the I1 lower incisors andcanines molar cusp and occlusal crest morphology degree of flarein all molars and especially mandibular features clearly distin-guish the Kisingiri from non-Kisingiri samples in a way that isincompatible with congeneric species With the transfer ofE heseloni and E nyanzae Proconsul becomes a well-defined cladecomprising at least three speciesdP africanus P major andP meswaedunited by several features that are unique for thegenus (Table 2)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6160
We found no support in this study for the genus ldquoUgandapi-thecusrdquo as constructed by Pickford et al (2009) In that paper theytransferred the vast majority of Proconsul fossils from the Tinderetand Napak localities into ldquoUgandapithecusrdquo leaving P africanusrepresented only by its holotype and a small number of isolatedteeth and aligning it with the material from Rusinga and Mfan-ganoWhen the P africanus hypodigm is reconstituted and properlyaffiliated with the other Tinderet species according to itsmorphology much of the evidence marshaled by Pickford et al(2009) for ldquoUgandapithecusrdquo actually supports the distinction be-tween the Tinderet and Kisingiri samples ie the generic distinc-tion of Ekembo In addition some of the traits enumerated byPickford and colleagues in support of ldquoUgandapithecusrdquo wererevealed in our analysis to be variable within one or more speciesand therefore of questionable taxonomic utility We do not rejectout of hand the original conception of ldquoUgandapithecusrdquo (Senutet al 2000) as either a monotypic genus distinguishing ldquoUrdquo ma-jor or perhaps including a closely allied middle Miocene ldquoUrdquogitongai as well (Pickford and Kunimatsu 2005) but we have fornow retained the early Miocene specimens of both in Proconsulpending further study However even if Ugandapithecus is a taxo-nomically valid genus comprising just the latter two species theirmorphology indicates that theywould still be united in a cladewithProconsul and they lack several more advanced features that partlydefine Ekembo Better understanding of the role that body size playsinmediating differences within Proconsulwill help to further clarifythe relationships among these species
Despite the existence of one well-represented and several lesscomplete skeletons of what was long assumed to be Proconsul thephylogenetic status of this genus has defied consensus This islargely due in our view to Proconsul having been an admixture oftwo distinct genera one more advanced in the direction of extantapes than the other As we demonstrate here the material fromRusinga and Mfangano traditionally referred to Proconsul andwhich includes all of the more complete skeletal material isdistinct from the Tinderet and Napak material and belongs in aseparate genus Ekembo By disentangling these distinct clades weanticipate being able to achieve better phylogenetic resolution forboth genera thereby improving our knowledge of early hominoidevolution and diversification
Acknowledgments
We thank many people and agencies that assisted with thedevelopment and completion of this project This research wasinitiated as a result of funding from the Wenner-Gren Foundationand the Johns Hopkins School of Medicine to DR Begun in1987e1989 and was renewed following research and travelgenerously funded by multiple Leakey Foundation grants to KMcNulty National Science Foundation grants to K McNulty(0852609 1241807) the McKnight Foundation the University ofMinnesota the Lloyd A Wilford Endowment the LeverhulmeFoundation and the Turkana Basin Institute J Kelley gratefullyacknowledges support from the Institute of Human Origins Ari-zona State University DR Begun also wishes to acknowledge theAlexander Humboldt Stiftung and NSERC for funding support Weare grateful to the National Museums of Kenya the UgandaMuseum the Natural History Museum and to Rose Mwanja EzraMusiime Tom Mukhuyu Samuel Muteti James Yatich and MaryMuungu for access to and assistance with specimens in their careMany people provided assistance comments criticisms insightsand suggestions on various aspects of this work and we especiallythank Holly Dunsworth Will Harcourt-Smith Tom Lehmann DanPeppe Nicole Garrett Kirsten Jenkins Rutger Jansma Susy CoteIsaiah Nengo Andrew Hill and Brenda Benefit K McNulty
acknowledges the evolutionary anthropology faculty and studentsat Durham University for their helpful comments on an earlyversion of this manuscript and for their gracious hospitality Weare likewise grateful to the anonymous reviewers associate editorand editor whose comments strengthened the paper This manu-script is publication 1 supporting Research on East AfricanCatarrhine and Hominoid Evolution (REACHE)
Appendix A Supplementary data
Supplementary data related to this article can be found at httpdxdoiorg101016jjhevol201503009
References
Andrews P 1974 New species of Dryopithecus from Kenya Nature 249 188e190Andrews P 1978 A revision of the Miocene Hominoidea of East Africa Bull Brit
Mus (Nat Hist) Geol 30 85e225Andrews P 1985 Family group systematics and evolution among catarrhine pri-
mates In Delson E (Ed) Ancestors The Hard Evidence Liss New Yorkpp 14e22
Andrews P Harrison T Martin L Pickford M 1981 Hominoid primates from anew Miocene locality named Meswa Bridge in Kenya J Hum Evol 10123e128
Andrews P Begun D Zylstra M 1997 Interrelationships between functionalmorphology and paleoenvironments in Miocene hominoids In Begun DWard C Rose M (Eds) Function Phylogeny and Fossils Miocene HominoidEvolution and Adaptations Plenum Press New York pp 29e58
Andrews P Molleson T 1979 The provenance of Sivapithecus africanus Bull BritMus (Nat Hist) Geol 32 19e23
Begun DR 2001 African and Eurasian Miocene hominoids and the origins of theHominidae In de Bonis L Koufos GD Andrews P (Eds) Hominoid Evolutionand Climatic Change in Europe Phylogeny of the Neogene Hominoid Primatesof Eurasia Cambridge University Press London pp 231e253
Begun DR 2007 Fossil record of Miocene hominoids In Henke W Tattersall I(Eds) Handbook of Paleoanthropology Springer Berlin pp 921e977
Begun DR Teaford MF Walker AC 1994 Comparative and functional anatomyof Proconsul phalanges from the Kaswanga primate site Rusinga Island KenyaJ Hum Evol 26 89e165
Bishop WW 1964 More fossil primates and other Miocene mammals from North-East Uganda Nature 203 1327e1331
Bishop WW Miller JA Fitch FJ 1969 New potassium-argon age determinationsrelevant to the Miocene fossil mammal sequence in East Africa Am J Sci 267669e699
Blankenship JA Mincer HH Anderson KM Woods MA Burton EL 2007Third molar development in the estimation of chronologic age in Americanblacks as compared with whites J Forensic Sci 52 428e433
Bosler W 1981 Species groupings of early Miocene dryopithecine teeth from EastAfrica J Hum Evol 10 151e158
Coleman MN 2008 What does geometric mean mean geometrically Assessingthe utility of geometric mean and other size variables in studies of skullallometry Am J Phys Anthropol 135 404e415
Cote S Malit N Nengo I 2013 Additional mandibles of Rangwapithecus gordonian early Miocene catarrhine from the Tinderet localities of Western KenyaJ Hum Evol 153 341e352
Deane AS 2009 Early Miocene catarrhine dietary behavior the influence of theRed Queen Effect on incisor shape and curvature J Hum Evol 56 275e285
Drake R Van Couvering JA Pickford M Curtis G Harris JA 1988 New chro-nology for the Early Miocene mammalian faunas of Kisingiri Western KenyaJ Geol Soc Lond 145 479e491
Evernden JF Savage DE Curtis GH James GT 1964 Potassium-argon datesand the Cenozoic mammalian chronology of North America Am J Sci 262145e198
Gebo DL MacLatchy L Kityo R Deino A Kingston J Pilbeam D 1997A hominoid genus from the Early Miocene of Uganda Science 276 401e404
Geoffroy Saint-Hilaire E 1812 Tableau des quadrumanes 1 Ord QuadrumanesAnnls Mus Hist Nat Paris 19 85e122
Gray JE 1825 Outline of an attempt at the disposition of the Mammalia into tribesand families with a list of the genera apparently appertaining to each tribeAnnls Philos 10 337e344
Harrison T 1982 Small-bodied apes from the Miocene of East Africa PhDDissertation University of London
Harrison T 1986 New fossil anthropoids from the middle Miocene of East Africaand their bearing on the origin of the Oreopithecidae Am J Phys Anthropol 71265e284
Harrison T 1987 The phylogenetic relationships of the early catarrhine primates areview of the current evidence J Hum Evol 16 41e80
Harrison T 2002 Late Oligocene to middle Miocene catarrhines from Afro-ArabiaIn Hartwig WC (Ed) The Primate Fossil Record Cambridge University PressCambridge pp 311e338
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 61
Harrison T 2010 Dendropithecoidea Proconsuloidea and Hominoidea (CatarrhiniPrimates) In Werdelin L Sanders WJ (Eds) Cenozoic Mammals of AfricaUniversity of California Press Berkeley pp 429e469
Harrison T Andrews P 2009 The anatomy and systematic position of a newspecies of early Miocene proconsulid from Meswa Bridge Kenya J Hum Evol56 479e496
Hill A Nengo IO Rossie JB 2013 A Rangwapithecus gordoni mandible from theearly Miocene site of Songhor Kenya J Hum Evol 65 490e500
Hopwood AT 1933 Miocene primates from Kenya J Linn Soc Zool 38 437e464Ishida H Kunimatsu Y Takano T Nakano Y Nakatsukasa M 2004 Nacholapi-
thecus skeleton from the Middle Miocene of Kenya J Hum Evol 46 69e103Jolicouer P 1963 The multivariate generalization of the allometry equation Bio-
metrics 19 497e499Jungers WL Falsetti AB Wall CE 1995 Shape relative size and size-
adjustments in morphometrics Am J Phys Anthropol 38 (S2) 137e161Kavanagh KD Evans AR Jernvall J 2007 Predicting evolutionary patterns of
mammalian teeth from development Nature 449 427e433Kelley J 1986 Species recognition and sexual dimorphism in Proconsul and
Rangwapithecus J Hum Evol 15 461e495Kelley J 1995 Sex determination in Miocene catarrhine primates Am J Phys
Anthropol 96 390e417Kelley J 1993 Taxonomic implications of sexual dimorphism in Lufengpithecus In
Kimbel WH Martin LB (Eds) Species Species Concepts and Primate Evo-lution Springer New York pp 429e458
Kent PE 1942 The country round the Kavirondo Gulf of Victoria Nyanza Geogr J100 22e31
Kordos L Begun DR 2001 A new cranium of Dryopithecus from RudabanyaHungary J Hum Evol 15 689e700
Le Gros Clark WE Leakey LSB 1950 Diagnoses of East African Miocene Homi-noidea Q J Geol Soc Lond 105 260e263
Le Gros Clark WE Leakey LSB 1951 The Miocene Hominoidea of East Africa InFossil Mammals of Africa No 1 British Museum (Natural History) London
Leakey LSB 1943 A Miocene anthropoid mandible from Rusinga Kenya Nature152 319e320
Leakey LSB 1963 East African fossil Hominoidea and the classification within thissuper-family In Washburn SL (Ed) Classification and Human EvolutionAldine Chicago pp 32e49
Leakey LSB 1967 Notes on the mammalian faunas from the Miocene and Pleis-tocene of East Africa In Bishop WW Clark JD (Eds) Background to Evolu-tion in Africa University of Chicago Press Chicago pp 7e29
Leakey RE Leakey MG 1986 A new Miocene hominoid from Kenya Nature 324143e146
Leakey RE Leakey MG Walker AC 1988 Morphology of Afropithecus turka-nensis from Kenya Am J Phys Anthropol 76 289e307
Leakey MG Ungar PS Walker A 1995 A new genus of large primate from theLate Oligocene of Lothidok Turkana District Kenya J Hum Evol 28519e531
Linnaeus C 1758 Systema naturae per regna tria naturae secundum classesordines genera species cum characteribus differentris synonymis locis tenthed Laurentii Salvii Stockholm
MacInnes DG 1943 Notes on the East African Miocene primates J E Afr UgandaNat Hist Soc 17 141e181
MacLatchy L 2004 The oldest ape Evol Anthropol 13 90e103MacLatchy L Bossert WH 1996 An analysis of the articular surface distribution of
the femoral head and acetabulum in anthropoids with implications for hipfunction in Miocene hominoids J Hum Evol 31 425e453
MacLatchy L Rossie JB 2005 The Napak hominoid still Proconsul major InLieberman DE Smith RJ Kelley J (Eds) Interpreting the Past Essays onHuman Primate and Mammal Evolution in Honor of David Pilbeam Brill Ac-ademic Publishers Boston pp 15e28
Martin L 1981 New specimens of Proconsul from Koru Kenya J Hum Evol 10139e150
McCollum MS Peppe DJ McNulty KP Dunsworth HM Harcourt-Smith WEH Andrews AL 2013 Magnetostratigraphy of the early MioceneHiwegi Formation (Rusinga Island Lake Victoria Kenya) Geological Society ofAmerica Abstracts with Programs 45 p 12
McNulty KP 2010 Apes and tricksters the evolution and diversification ofhumans closest relatives Evol Edu Outreach 3 322e332
McNulty KP Harcourt-Smith WEH Dunsworth HM 2007 New primate fossilsfrom Rusinga Island Kenya Am J Phys Anthropol 132 (S44) 170
McNulty KP MacLatchy L Peppe DJ Nengo I Manthi FK Miller ERStevens NJ Cote S Lehmann T Kingston J 2014a A regional approach toEast African early Miocene paleobiology J Vert Paleontol 185 SVP Programand Abstracts Book 2014
McNulty KP MacLatchy L Rossie JB Peppe DJ Deino AL Mbua ENManthi FK Nengo IO Stevens NJ Cote S Lehmann T Gutierrez M 2014b
Research on East African catarrhine and hominoid evolution results from thefirst year Am J Phys Anthropol 153 (S58) 182
Michel LA Peppe DJ Lutz JA Driese SG Dunsworth HM Harcourt-Smith WEH Horner W Lehmann T Nightengale S McNulty KP 2014Remnants of an ancient forest provide ecological context for Early Miocenefossil apes Nature Comm 5 3236
Myres JL 1932 Africa Man 32 208Nakatsukasa M Kunimatsu Y Nakano Y Takano T Ishida H 2007 Vertebral
morphology of Nacholapithecus kerioi based on KNM-BG 35250 J Hum Evol52 347e369
Nengo IO Rae TC 1992 New hominoid fossils from the early Miocene site ofSonghor Kenya J Hum Evol 23 423e429
Peppe DJ McNulty KP Cote SM Harcourt-Smith WEH Dunsworth HM VanCouvering JA 2009 Stratigraphic interpretation of the Kulu Formation (EarlyMiocene Rusinga Island Kenya) and its implications for primate evolutionJ Hum Evol 56 447e461
Pickford M 1981 Preliminary Miocene mammalian biostratigraphy for westernKenya J Hum Evol 10 73e97
Pickford M 1986 Sexual dimorphism in Proconsul Hum Evol 1 111e148Pickford M Andrews PJ 1981 The Tinderet Miocene sequence in Kenya J Hum
Evol 10 11e33Pickford M Kunimatsu Y 2005 Catarrhines from the Middle Miocene (ca 145
Ma) of Kipsaraman Tugen Hills Kenya Anthropol Sci 113 189e224Pickford M Senut B Gommery D Musiime E 2009 Distinctiveness of Ugan-
dapithecus from Proconsul Estudios Geol 65 183e241Pilbeam DR 1969 Tertiary Pongidae of East Africa evolutionary relationships and
taxonomy Bulletin of the Peabody Museum of Natural History 31 pp 1e185Rae TC 1993 Phylogenetic analysis of proconsulid facial morphology PhD
Dissertation State University of New York at Stony BrookRae TC 1999 Mosaic evolution in the origin of the Hominoidea Folia Primatol 70
125e135Rafferty KL Walker A Ruff C Rose MD Andrews PJ 1995 Postcranial esti-
mates of body weights in Proconsul with a note on a distal tibia of P major fromNapak Uganda Am J Phys Anthropol 97 391e402
Rossie JB MacLatchy L 2013 Dentognathic remains of an Afropithecus individualfrom Kalodirr Kenya J Hum Evol 65 199e208
Senut B Pickford M Gommery D Kunimatsu Y 2000 Un nouveau genredrsquohominoiumlde du Miocene inferieur drsquoAfrique orientale Ugandapithecus major(Le Gros Clark and Leakey 1950) CR Acad Sci Paris 331 227e233
Simons EL Pilbeam DR 1965 Preliminary revision of the Dryopithecinae (Pon-gidae Anthropoidea) Folia Primatol 3 81e152
Smith TM Martin LB Leakey MG 2003 Enamel thickness microstructure anddevelopment in Afropithecus turkanensis J Hum Evol 44 283e306
Stevens NJ Seiffert ER OConnor PM Roberts EM Schmitz MD Krause CGorscak E Ngasala S Hieronymus TL Temu J 2013 Palaeontological evi-dence for an Oligocene divergence between Old World monkeys and apesNature 497 611e614
Suwa G Kono RT Katoh S Asfaw B Beyene Y 2007 A new species of great apefrom the late Miocene epoch in Ethiopia Nature 448 921e924
Szalay FS Delson E 1979 Evolutionary History of the Primates Academic PressNew York
Teaford MF Beard KC Leakey RE Walker A 1988 New hominoid facial skel-eton from the Early Miocene of Rusinga Island Kenya and its bearing on therelationship between Proconsul nyanzae and Proconsul africanus J Hum Evol17 461e477
Van Couvering JAH Miller J 1969 Miocene stratigraphy and age determinationsRusinga Island Kenya Nature 221 628e632
Walker A 1997 Proconsul function and phylogeny In Begun DR Ward CVRose MD (Eds) Function Phylogeny and Fossils Miocene Hominoid Evolu-tion and Adaptation Plenum Press New York pp 209e224
Walker A Pickford M 1983 New postcranial fossils of Proconsul africanus andProconsul nyanzae In Ciochon RL Corruccini RS (Eds) New Interpretationsof Ape and Human Ancestry Plenum Press New York pp 325e351
Walker A Teaford M 1988 The Kaswanga primate site an Early Miocene hom-inoid site on Rusinga Island Kenya J Hum Evol 17 539e544
Walker A Teaford M 1989 The hunt for Proconsul Sci Am 260 76e82Walker A Teaford MF Martin L Andrews P 1993 A new species of Proconsul
from the early Miocene of RusingaMfangano Islands Kenya J Hum Evol 2543e56
Ward CV Ruff CB Walker A Rose MD Teaford MF Nengo IO 1995 Func-tional morphology of Proconsul patellas from Rusinga Island Kenya with im-plications for other Miocene-Pliocene catarrhines J Hum Evol 29 1e19
Ward S Brown B Hill A Kelley J Downs W 1999 Equatorius a new hominoidgenus from the Middle Miocene of Kenya Science 285 1382e1386
Werdelin L Sanders WJ 2010 Cenozoic Mammals of Africa University of Cali-fornia Press Berkeley

Figure 14 Relative buccolingual dimensions of M1 and M2 KNM-SO 1112 has a preserved M2 but only the cervix of M1 Its M1 breadth was estimated based on the ratio to cervixbreadth to maximum crown breadth in other Tinderet fossils Even the cervical breadth of SO 1112 is broader than would be predicted for the crown breadth of a Kisingiri M1 Trendlines computed using OLS regression and using the average estimate of SO 1112 M1 breadth Group differences were significant (p frac14 00002)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 57
even hominid taxa such as Rudapithecus (eg Kordos and Begun2001)
Proconsul is likewise united by a suite of features that are for themost part characteristic only of this lineage (Table 2) Themorphology of both upper and lower canines is unique to thisgenus and shared by all three well-known species plus the poorlyrepresented middle Miocene P gitongai (Pickford and Kunimatsu2005) The mandibular symphysis is also distinctive in Proconsulcompared with other catarrhines with its very long and shallowlysloping planum alveolare and steeply inclined subplanum surfaceIndeed even the small edentulous anterior mandibular fragmentKNM-SO 372 is readily placed within Proconsul and excluded fromother similarly sized catarrhine taxa such as Rangwapithecus andEkembo based on this unique symphyseal morphology (Figs 8 and9)
Other differences between Ekembo and Proconsul are of uncer-tain evolutionary polarity (eg orientation of the posthypoconecrista) or may be of taxonomic relevance without broader phylo-genetic significance (eg degree of molar flare Table 2) Werecognize that there are morphological differences within Procon-sul particularly between the better known species P africanus andP major However many of these differences are likely related tosize and represent variations on a single generic theme that aremarkedly different from the morphology of Ekembo The recentdescription (Harrison and Andrews 2009) of the intermediatelysized P meswae underscores the broad morphological unity withinProconsul and further substantiates a well-defined Proconsul cladecomprising Tinderet and Ugandan species
Based on the abundant morphological and morphometric evi-dence supporting genus-level distinction between Proconsul andEkembo we are currently conducting a more detailed phylogeneticanalysis of all early Miocene catarrhines in order to better under-stand the distribution and evolutionary significance of these andother morphological features We anticipate that the debates sur-rounding the phylogenetic relationships of an historically consti-tuted sample of Proconsul can be more satisfactorily resolved nowthat the Kisingiri material is appropriately recognized as a distinctgenus
The generic disposition of ldquoUgandapithecusrdquo species
A great many features said to characterize ldquoUgandapithecusrdquowere added to the original diagnosis of the genus by Pickford et al(2009) in their systematic revision of Proconsul (sensu lato) As partof that study they also erected a new species ldquoUrdquo legetetensis andtransferred P meswae to ldquoUgandapithecusrdquo Evaluating the same setof features across the entire available sample however we wereunable to find support for this more expansive conception ofldquoUgandapithecusrdquo for several reasons First some diagnostic fea-tures cited in Pickford et al (2009) were quite variable within theldquoUgandapithecusrdquo sample As an example marked reduction of theM3 entoconid to a bead on the lingual cingulum is not shared byseveral ldquoUrdquo major specimens such as KNM-CA 393 NAP I 49000and Kor 253004 (see Pickford et al 2009 Fig 33e) and was likelynot a feature of a few of the more worn specimens eg BUMP 600and BUMP 601 or even the holotype of P major M 16648 Second
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6158
some diagnostic features of ldquoUgandapithecusrdquo were found to bevariably present in Ekembo arguing against their utility for dis-tinguishing ldquoUgandapithecusrdquo from Pickford et als (2009) com-bined P africanus thorn Kisingiri conception of Proconsul For examplesignificantly reduced curvature of the I1 distal margin in ldquoUganda-pithecusrdquo is also found in some Kisingiri specimens including KNM-RU 1685 1712 1714 1831 1846 and 1951 several more Rusingaspecimens have moderately reduced curvature Third some fea-tures that purportedly distinguish ldquoUgandapithecusrdquo fromP africanus are likely related to size For example the somewhatmore central position of the P3 paracone in ldquoUrdquomajor is not seen inthe smaller ldquoUrdquo meswae whose paracone is positioned similarly tothat of the holotype of P africanus (see Harrison and Andrews2009 Fig 6d)
However the major part of the evidence for recognizingldquoUgandapithecusrdquo (sensu Pickford et al 2009) was assembled by re-assigning specimens traditionally attributed to P africanus into newspecies of ldquoUgandapithecusrdquo The P africanus holotype was left inassociation with the Rusinga sample despite having morphologyoverwhelmingly characteristic of other Tinderet specimens Theresulting specimen allocations left P africanus morphologicallyimpoverisheddstripped of much of its hypodigmdso that Pro-consul for Pickford et al (2009) came to be characterized primarilyby the Rusinga sample (see similar comments in Harrison andAndrews 2009) Within that conceptual framework any spec-imen that differs from those at Rusinga could not be Proconsul butmust instead belong to ldquoUgandapithecusrdquo Thus specimens such asthe KNM-SO 1112 mandible were erroneously assigned to ldquoUgan-dapithecusrdquo because they were (correctly) perceived as differingfrom their Rusinga counterparts (Pickford et al 2009)
Here we restore the P africanus hypodigm and associate it withthe other Tinderet fossils according to their shared morphologySpecimens assigned to ldquoUgandapithecusrdquo revert to Proconsul andmany of the features cited to support a speciose ldquoUgandapithecusrdquo(Pickford et al 2009) instead become diagnostic features of Pro-consul This correction serves to highlight the distinctiveness of theKisingiri remains and the need to recognize this distinctiveness atthe generic level
As demonstrated above specimens from Kisingiri do not displaythe morphology of Proconsul as represented by the holotype ofP africanus M 14084 Given that this specimen preserves only theupper canine and cheek teeth similarities with or departures fromthis morphology represent the most compelling arguments forassessing inclusivity in the genus However for the mandible andlower dentition if one simply sorts the specimens by elementwithout relying on taxonomic preconceptions the overwhelmingdistinction within the traditionally constituted Proconsul sample isbetween Kisingiri material on the one hand and that from Tinderetand Uganda on the other (cf Walker et al 1993 Begun 2001 2007MacLatchy and Rossie 2005 Harrison and Andrews 2009Harrison 2010) Additionally with the Kisingiri specimensremoved to Ekembo the postcranial differences used to partlyjustify a distinct ldquoUgandapithecusrdquo lose their relevance (seeHarrison and Andrews 2009)
We have differed among ourselves in our past treatment ofP major as generically distinct from the smaller species of Pro-consul (Begun 2007 McNulty 2010) Morphologically many ofthe unique features of P major can be interpreted as size-relatedvariants on the smaller P africanus morphology Even if so thequestion is whether these are sufficiently different and numerousto warrant generic distinction that is do they suggest thatP major belongs in a separate clade from P africanus andP meswae As we have demonstrated differences among thesethree species are certainly fewer and show less morphologicaldistinction than those between Ekembo and any of the three
Nevertheless while we ultimately agree with Harrison andAndrews (2009 see also Begun 2007) that the features dis-tinguishing ldquoUrdquo major from P africanus are consistent with thosethat distinguish congeneric species and are at least partly due todifferences in size we concede the possibility that the largestProconsul species could belong in a separate genus (Senut et al2000 Pickford and Kunimatsu 2005) However whether or notone recognizes a separate genus for P major the P africanus ma-terial does not sort morphologically with the Kisingiri materialTherefore the taxonomic alternatives are either two generaProconsul and Ekembo or three Proconsul Ugandapithecus andEkembo with Proconsul and Ugandapithecus united in a cladeexclusive of Ekembo Thus while we strongly favor the firstalternative recognizing Ugandapithecus does not change thefundamental argument concerning the need to refer the Kisingirispecies to a generically and cladistically distinct taxon from thespecies represented at Tinderet and the Ugandan sites
Specimens of interest
Despite the substantial dental and gnathic differences betweenProconsul and Ekembo there are a few specimens whose taxonomicaffinities require additional scrutiny
KNM-RU 1676-77 This set of associated upper and lower teeth is aconsistent outlier among E heseloni and E nyanzae specimens Insome ways this specimen more closely resembles Tinderet Pro-consul in the features that distinguish it from Ekembo Molar cuspmorphology and cingulum development I1 lingual tubercledevelopment and M2 shape and posthypocone crista orientationare more consistent with specimens from the Tinderet sites On theother hand it otherwise resembles Ekembo and lacks the strongestdiagnostic features that unite species of Proconsul such as thedistinctive canine and P3 morphology Overall this specimen doesnot fit unambiguously into either genus in fact the degree ofbuccolingual flare cusp morphology enamel wrinkling and espe-cially the long extension of enamel onto the anterior root in P3 arein some ways reminiscent of Afropithecus although it clearly differsfrom that genus as well (cf Rossie and MacLatchy 2013)
Were these characteristics individually distributed among thelarge Kisingiri sample one might attribute this to intra-genericvariation but when found together in a single specimen from alarge sample it suggests a unique individual One of us (KPM)thinks it likely that the specimen was not actually found at aKisingiri locality based on its unique preservation compared toother Rusinga fossils and on other examples of mis-proveniencedspecimens (Leakey 1967 Andrews and Molleson 1979 Pickford1986 Kelley 1986) However we recognize that relying toofrequently on this explanation allows almost any scenario to bejustified and suggest some alternative possibilities One is that thespecimen was found on Rusinga but in the much older WayandoFormation strata instead of the Hiwegi Formation The differencesin morphology in that case might reflect more than a million yearsof temporal change (McCollum et al 2013) with KNM-RU 1676-77preserving certain more primitive Proconsul-like features As analternative it is reasonable to assume that fossil assemblages asvast as those from Rusingawill include not only abundant taxa butalso rare or infrequent inhabitants of the local biome There aremany examples of this from Rusingas mammalian communityeg ldquoHemicyonrdquo Kulutherium Tadarida cf Hyainailouros cfArchaeobeledon Kelba (see relevant chapters in Werdelin andSanders 2010) As such KNM-RU 1676-77 may simply representa rare and well-preserved specimen of some other catarrhinetaxon that lived among or visited a primate community dominatedby Ekembo
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 59
KNM-SO 1112 This specimen was initially referred to Rangwapi-thecus but soon after was transferred to Proconsul afrianus (Bosler1981 Martin 1981)da designation that most subsequent authorshave followed (Pickford 1986 Nengo and Rae 1992 Kelley 19861995 Walker et al 1993 MacLatchy and Rossie 2005 Hill et al2013) Pickford et al (2009) more recently referred this specimento ldquoUrdquo meswae citing two primary reasons First they reiteratedBoslers (1981) concern that its P3 differs from others attributedto P africanus However at that time the majority of what wasconsidered to be P africanus would have been from RusingaIsland In fact while noting its contrasting morphology Boslersuggested that differences between KNM-SO 1112 and other P3smight reasonably be attributed to sexual dimorphism notingthat this specimen provides ldquoa not implausible match for the Pafricanus holotype maxilla M 14084rdquo (1981152) Thatassessment is borne out in this study Using the much largersample of Ekembo P3s as a model variation can be seen toencompass specimens with well-developed mesial beaks andthose with more bilaterally compressed crowns and parallelsides The latter morphology is predominantly found amongmale specimens (KNM-RU 1674 1924 and KPS I and III) thussupporting Boslers (1981) suggestion that this variation isrelated to sexual dimorphism and hence that KNM-SO 1112 is amale P africanus
The second feature cited by Pickford et al (2009) in referringthis specimen to ldquoUrdquo meswae was posterior shallowing of themandibular corpus However because they redefined ldquoUgandapi-thecusrdquo to include all of the well-preserved Tinderet mandibles(Pickford et al 2009)dleaving Proconsul to be represented only byRusinga specimensdtheir only alternative within this scheme for aspecimen the size of SO 1112 was to refer it to ldquoUrdquomeswae It seemsodd though to assign this specimen to a species from a (perhapsmuch) older site when the Proconsul africanus holotype is ofappropriate size and similar age and as there are no other adultmandibles from either species with which to compare it In anycase mandibular shallowing is variable in both Ekembo and Pro-consul with some Kisingiri specimens exhibiting considerableshallowing (eg KNM-RU 2087) and some ldquoUgandapithecusrdquo spec-imens having very little (eg M 14086)
We therefore see no morphological grounds to assign KNM-SO1112 to P meswae but do find some morphological evidence toassign it as a male P africanus which occurs at Songhor
KNM-LG 452 This very complete mandible was originally recog-nized as a female of P major based on the size of the dentition androbusticity of the mandible (Martin 1981) Bosler (1981) howeverleft this specimen unassigned noting similarities to P major in P3-M1 size but having distinctly smaller M2e3 In this she likenedKNM-LG 452 to a few other specimens (notably M 14086) thatare smaller in their distal molars in a manner more consistentwith taxonomic rather than sex differences (Bosler 1981 see alsoPilbeam 1969) Kelley (1986 1995) also assigned this specimen asa female of P major based on canine size and morphology butnoted that the canine lacked the shallowly-sloping mesial ridgetypical of most female anthropoid canines This is also truehowever of two other canines assigned as female P major KO 9and SO 373 (Kelley 1986) and so might simply be an unusualcharacteristic of P major female lower canines In the end Kelley(1995) concluded that considering all of the criteria fordistinguishing male and female canines the evidence that KNM-LG 452 is female remains questionable
Pickford et al (2009) further developed the idea that two spe-cies are represented within the P major sample and made KNM-LG452 the holotype of a new species ldquoUrdquo legetetensis We concur thatthis specimen is unique among the well-preserved Tinderet
mandibles in some important features Whereas male and femalemandibles of P major have long internal symphyseal planesexpansive extra- and retromolar sulci and rami that cross theocclusal plane at distal M3 KNM-LG 452 has a shorter symphysealplane (though not to the extent seen in Ekembo) a moderateextramolar sulcus but no retromolar sulcus and a ramus thatcrosses the occlusal plane more mesially If KNM-SO 396 and SO404 represent males and females respectively of P major then themorphology of KNM-LG 452 appears distinct
At present however we do not think there is sufficient evidenceto differentiate this specimen from either P major or P meswaePickford et al (2009) argue that KNM-LG 452 cannot belong to thelatter species because of its larger size The size differences how-ever are well within the range of variation seen in P majorE heseloni and E nyanzae Moreover whereas the Legetet mandibleis larger than most of the P meswae hypodigm an adult P4 referredto this species (KNM-ME 25) substantially exceeds its counterpartin KNM-LG 452 Should the Legetetmandible ultimately prove to bea male specimen size alone would not rule out its attribution toP meswae On the other hand if it is female then it becomes harderto justify its inclusion in that sample of mostly smaller specimens Iffemale therefore it could represent a separate species for whichthe binomial Proconsul legetetensis comb nov is available Recentlycollected fossils from the SonghorKapurtay area provide someadditional support for the possibility that three species of Proconsulexisted at these localities (McNulty 2014ab) However toconvincingly demonstrate this and determine whether a thirdspecies can be differentiated from P meswae will require a moredetailed analysis of variation among Proconsul species a corre-sponding study of variation in modern taxa anddalmost certain-lydadditional fossil specimens For now we regard ldquoUrdquo legetetensisas a subjective junior synonym of P major
Conclusions
The genus Proconsul has been central to studies of ape and hu-man evolution since its first description by Hopwood (1933) As oneof the best known fossil catarrhines represented by nearly everyskeletal element our knowledge of Proconsuls functional anatomypositional behavior and overall morphology has made it thefoundation for paleoanthropological research in the early Mioceneas well as studies on the origin and early diversification of homi-noids With respect to systematics however Proconsul is less wellunderstood the type species P africanus is known from a rela-tively small number of specimens from sites in the Tinderet areaand no postcranial bones can be unambiguously assigned to itRather it is the copious and well-preserved specimens from theKisingiri sites of Rusinga and Mfangano Islands that have formedthe basis for the common image of Proconsul
We argue here that the Kisingiri species traditionally referredto ProconsuldP heseloni and P nyanzaedare generically distinctfrom P africanus and hereby transfer them into a new genusEkembo based on a substantial number of morphological differ-ences (Table 2) Considering only those traits preserved in theP africanus holotype the entire sample from Rusinga and Mfan-gano differs markedly in the morphology of the upper canine P3P4 and M2 Additional differences in the I1 lower incisors andcanines molar cusp and occlusal crest morphology degree of flarein all molars and especially mandibular features clearly distin-guish the Kisingiri from non-Kisingiri samples in a way that isincompatible with congeneric species With the transfer ofE heseloni and E nyanzae Proconsul becomes a well-defined cladecomprising at least three speciesdP africanus P major andP meswaedunited by several features that are unique for thegenus (Table 2)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6160
We found no support in this study for the genus ldquoUgandapi-thecusrdquo as constructed by Pickford et al (2009) In that paper theytransferred the vast majority of Proconsul fossils from the Tinderetand Napak localities into ldquoUgandapithecusrdquo leaving P africanusrepresented only by its holotype and a small number of isolatedteeth and aligning it with the material from Rusinga and Mfan-ganoWhen the P africanus hypodigm is reconstituted and properlyaffiliated with the other Tinderet species according to itsmorphology much of the evidence marshaled by Pickford et al(2009) for ldquoUgandapithecusrdquo actually supports the distinction be-tween the Tinderet and Kisingiri samples ie the generic distinc-tion of Ekembo In addition some of the traits enumerated byPickford and colleagues in support of ldquoUgandapithecusrdquo wererevealed in our analysis to be variable within one or more speciesand therefore of questionable taxonomic utility We do not rejectout of hand the original conception of ldquoUgandapithecusrdquo (Senutet al 2000) as either a monotypic genus distinguishing ldquoUrdquo ma-jor or perhaps including a closely allied middle Miocene ldquoUrdquogitongai as well (Pickford and Kunimatsu 2005) but we have fornow retained the early Miocene specimens of both in Proconsulpending further study However even if Ugandapithecus is a taxo-nomically valid genus comprising just the latter two species theirmorphology indicates that theywould still be united in a cladewithProconsul and they lack several more advanced features that partlydefine Ekembo Better understanding of the role that body size playsinmediating differences within Proconsulwill help to further clarifythe relationships among these species
Despite the existence of one well-represented and several lesscomplete skeletons of what was long assumed to be Proconsul thephylogenetic status of this genus has defied consensus This islargely due in our view to Proconsul having been an admixture oftwo distinct genera one more advanced in the direction of extantapes than the other As we demonstrate here the material fromRusinga and Mfangano traditionally referred to Proconsul andwhich includes all of the more complete skeletal material isdistinct from the Tinderet and Napak material and belongs in aseparate genus Ekembo By disentangling these distinct clades weanticipate being able to achieve better phylogenetic resolution forboth genera thereby improving our knowledge of early hominoidevolution and diversification
Acknowledgments
We thank many people and agencies that assisted with thedevelopment and completion of this project This research wasinitiated as a result of funding from the Wenner-Gren Foundationand the Johns Hopkins School of Medicine to DR Begun in1987e1989 and was renewed following research and travelgenerously funded by multiple Leakey Foundation grants to KMcNulty National Science Foundation grants to K McNulty(0852609 1241807) the McKnight Foundation the University ofMinnesota the Lloyd A Wilford Endowment the LeverhulmeFoundation and the Turkana Basin Institute J Kelley gratefullyacknowledges support from the Institute of Human Origins Ari-zona State University DR Begun also wishes to acknowledge theAlexander Humboldt Stiftung and NSERC for funding support Weare grateful to the National Museums of Kenya the UgandaMuseum the Natural History Museum and to Rose Mwanja EzraMusiime Tom Mukhuyu Samuel Muteti James Yatich and MaryMuungu for access to and assistance with specimens in their careMany people provided assistance comments criticisms insightsand suggestions on various aspects of this work and we especiallythank Holly Dunsworth Will Harcourt-Smith Tom Lehmann DanPeppe Nicole Garrett Kirsten Jenkins Rutger Jansma Susy CoteIsaiah Nengo Andrew Hill and Brenda Benefit K McNulty
acknowledges the evolutionary anthropology faculty and studentsat Durham University for their helpful comments on an earlyversion of this manuscript and for their gracious hospitality Weare likewise grateful to the anonymous reviewers associate editorand editor whose comments strengthened the paper This manu-script is publication 1 supporting Research on East AfricanCatarrhine and Hominoid Evolution (REACHE)
Appendix A Supplementary data
Supplementary data related to this article can be found at httpdxdoiorg101016jjhevol201503009
References
Andrews P 1974 New species of Dryopithecus from Kenya Nature 249 188e190Andrews P 1978 A revision of the Miocene Hominoidea of East Africa Bull Brit
Mus (Nat Hist) Geol 30 85e225Andrews P 1985 Family group systematics and evolution among catarrhine pri-
mates In Delson E (Ed) Ancestors The Hard Evidence Liss New Yorkpp 14e22
Andrews P Harrison T Martin L Pickford M 1981 Hominoid primates from anew Miocene locality named Meswa Bridge in Kenya J Hum Evol 10123e128
Andrews P Begun D Zylstra M 1997 Interrelationships between functionalmorphology and paleoenvironments in Miocene hominoids In Begun DWard C Rose M (Eds) Function Phylogeny and Fossils Miocene HominoidEvolution and Adaptations Plenum Press New York pp 29e58
Andrews P Molleson T 1979 The provenance of Sivapithecus africanus Bull BritMus (Nat Hist) Geol 32 19e23
Begun DR 2001 African and Eurasian Miocene hominoids and the origins of theHominidae In de Bonis L Koufos GD Andrews P (Eds) Hominoid Evolutionand Climatic Change in Europe Phylogeny of the Neogene Hominoid Primatesof Eurasia Cambridge University Press London pp 231e253
Begun DR 2007 Fossil record of Miocene hominoids In Henke W Tattersall I(Eds) Handbook of Paleoanthropology Springer Berlin pp 921e977
Begun DR Teaford MF Walker AC 1994 Comparative and functional anatomyof Proconsul phalanges from the Kaswanga primate site Rusinga Island KenyaJ Hum Evol 26 89e165
Bishop WW 1964 More fossil primates and other Miocene mammals from North-East Uganda Nature 203 1327e1331
Bishop WW Miller JA Fitch FJ 1969 New potassium-argon age determinationsrelevant to the Miocene fossil mammal sequence in East Africa Am J Sci 267669e699
Blankenship JA Mincer HH Anderson KM Woods MA Burton EL 2007Third molar development in the estimation of chronologic age in Americanblacks as compared with whites J Forensic Sci 52 428e433
Bosler W 1981 Species groupings of early Miocene dryopithecine teeth from EastAfrica J Hum Evol 10 151e158
Coleman MN 2008 What does geometric mean mean geometrically Assessingthe utility of geometric mean and other size variables in studies of skullallometry Am J Phys Anthropol 135 404e415
Cote S Malit N Nengo I 2013 Additional mandibles of Rangwapithecus gordonian early Miocene catarrhine from the Tinderet localities of Western KenyaJ Hum Evol 153 341e352
Deane AS 2009 Early Miocene catarrhine dietary behavior the influence of theRed Queen Effect on incisor shape and curvature J Hum Evol 56 275e285
Drake R Van Couvering JA Pickford M Curtis G Harris JA 1988 New chro-nology for the Early Miocene mammalian faunas of Kisingiri Western KenyaJ Geol Soc Lond 145 479e491
Evernden JF Savage DE Curtis GH James GT 1964 Potassium-argon datesand the Cenozoic mammalian chronology of North America Am J Sci 262145e198
Gebo DL MacLatchy L Kityo R Deino A Kingston J Pilbeam D 1997A hominoid genus from the Early Miocene of Uganda Science 276 401e404
Geoffroy Saint-Hilaire E 1812 Tableau des quadrumanes 1 Ord QuadrumanesAnnls Mus Hist Nat Paris 19 85e122
Gray JE 1825 Outline of an attempt at the disposition of the Mammalia into tribesand families with a list of the genera apparently appertaining to each tribeAnnls Philos 10 337e344
Harrison T 1982 Small-bodied apes from the Miocene of East Africa PhDDissertation University of London
Harrison T 1986 New fossil anthropoids from the middle Miocene of East Africaand their bearing on the origin of the Oreopithecidae Am J Phys Anthropol 71265e284
Harrison T 1987 The phylogenetic relationships of the early catarrhine primates areview of the current evidence J Hum Evol 16 41e80
Harrison T 2002 Late Oligocene to middle Miocene catarrhines from Afro-ArabiaIn Hartwig WC (Ed) The Primate Fossil Record Cambridge University PressCambridge pp 311e338
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 61
Harrison T 2010 Dendropithecoidea Proconsuloidea and Hominoidea (CatarrhiniPrimates) In Werdelin L Sanders WJ (Eds) Cenozoic Mammals of AfricaUniversity of California Press Berkeley pp 429e469
Harrison T Andrews P 2009 The anatomy and systematic position of a newspecies of early Miocene proconsulid from Meswa Bridge Kenya J Hum Evol56 479e496
Hill A Nengo IO Rossie JB 2013 A Rangwapithecus gordoni mandible from theearly Miocene site of Songhor Kenya J Hum Evol 65 490e500
Hopwood AT 1933 Miocene primates from Kenya J Linn Soc Zool 38 437e464Ishida H Kunimatsu Y Takano T Nakano Y Nakatsukasa M 2004 Nacholapi-
thecus skeleton from the Middle Miocene of Kenya J Hum Evol 46 69e103Jolicouer P 1963 The multivariate generalization of the allometry equation Bio-
metrics 19 497e499Jungers WL Falsetti AB Wall CE 1995 Shape relative size and size-
adjustments in morphometrics Am J Phys Anthropol 38 (S2) 137e161Kavanagh KD Evans AR Jernvall J 2007 Predicting evolutionary patterns of
mammalian teeth from development Nature 449 427e433Kelley J 1986 Species recognition and sexual dimorphism in Proconsul and
Rangwapithecus J Hum Evol 15 461e495Kelley J 1995 Sex determination in Miocene catarrhine primates Am J Phys
Anthropol 96 390e417Kelley J 1993 Taxonomic implications of sexual dimorphism in Lufengpithecus In
Kimbel WH Martin LB (Eds) Species Species Concepts and Primate Evo-lution Springer New York pp 429e458
Kent PE 1942 The country round the Kavirondo Gulf of Victoria Nyanza Geogr J100 22e31
Kordos L Begun DR 2001 A new cranium of Dryopithecus from RudabanyaHungary J Hum Evol 15 689e700
Le Gros Clark WE Leakey LSB 1950 Diagnoses of East African Miocene Homi-noidea Q J Geol Soc Lond 105 260e263
Le Gros Clark WE Leakey LSB 1951 The Miocene Hominoidea of East Africa InFossil Mammals of Africa No 1 British Museum (Natural History) London
Leakey LSB 1943 A Miocene anthropoid mandible from Rusinga Kenya Nature152 319e320
Leakey LSB 1963 East African fossil Hominoidea and the classification within thissuper-family In Washburn SL (Ed) Classification and Human EvolutionAldine Chicago pp 32e49
Leakey LSB 1967 Notes on the mammalian faunas from the Miocene and Pleis-tocene of East Africa In Bishop WW Clark JD (Eds) Background to Evolu-tion in Africa University of Chicago Press Chicago pp 7e29
Leakey RE Leakey MG 1986 A new Miocene hominoid from Kenya Nature 324143e146
Leakey RE Leakey MG Walker AC 1988 Morphology of Afropithecus turka-nensis from Kenya Am J Phys Anthropol 76 289e307
Leakey MG Ungar PS Walker A 1995 A new genus of large primate from theLate Oligocene of Lothidok Turkana District Kenya J Hum Evol 28519e531
Linnaeus C 1758 Systema naturae per regna tria naturae secundum classesordines genera species cum characteribus differentris synonymis locis tenthed Laurentii Salvii Stockholm
MacInnes DG 1943 Notes on the East African Miocene primates J E Afr UgandaNat Hist Soc 17 141e181
MacLatchy L 2004 The oldest ape Evol Anthropol 13 90e103MacLatchy L Bossert WH 1996 An analysis of the articular surface distribution of
the femoral head and acetabulum in anthropoids with implications for hipfunction in Miocene hominoids J Hum Evol 31 425e453
MacLatchy L Rossie JB 2005 The Napak hominoid still Proconsul major InLieberman DE Smith RJ Kelley J (Eds) Interpreting the Past Essays onHuman Primate and Mammal Evolution in Honor of David Pilbeam Brill Ac-ademic Publishers Boston pp 15e28
Martin L 1981 New specimens of Proconsul from Koru Kenya J Hum Evol 10139e150
McCollum MS Peppe DJ McNulty KP Dunsworth HM Harcourt-Smith WEH Andrews AL 2013 Magnetostratigraphy of the early MioceneHiwegi Formation (Rusinga Island Lake Victoria Kenya) Geological Society ofAmerica Abstracts with Programs 45 p 12
McNulty KP 2010 Apes and tricksters the evolution and diversification ofhumans closest relatives Evol Edu Outreach 3 322e332
McNulty KP Harcourt-Smith WEH Dunsworth HM 2007 New primate fossilsfrom Rusinga Island Kenya Am J Phys Anthropol 132 (S44) 170
McNulty KP MacLatchy L Peppe DJ Nengo I Manthi FK Miller ERStevens NJ Cote S Lehmann T Kingston J 2014a A regional approach toEast African early Miocene paleobiology J Vert Paleontol 185 SVP Programand Abstracts Book 2014
McNulty KP MacLatchy L Rossie JB Peppe DJ Deino AL Mbua ENManthi FK Nengo IO Stevens NJ Cote S Lehmann T Gutierrez M 2014b
Research on East African catarrhine and hominoid evolution results from thefirst year Am J Phys Anthropol 153 (S58) 182
Michel LA Peppe DJ Lutz JA Driese SG Dunsworth HM Harcourt-Smith WEH Horner W Lehmann T Nightengale S McNulty KP 2014Remnants of an ancient forest provide ecological context for Early Miocenefossil apes Nature Comm 5 3236
Myres JL 1932 Africa Man 32 208Nakatsukasa M Kunimatsu Y Nakano Y Takano T Ishida H 2007 Vertebral
morphology of Nacholapithecus kerioi based on KNM-BG 35250 J Hum Evol52 347e369
Nengo IO Rae TC 1992 New hominoid fossils from the early Miocene site ofSonghor Kenya J Hum Evol 23 423e429
Peppe DJ McNulty KP Cote SM Harcourt-Smith WEH Dunsworth HM VanCouvering JA 2009 Stratigraphic interpretation of the Kulu Formation (EarlyMiocene Rusinga Island Kenya) and its implications for primate evolutionJ Hum Evol 56 447e461
Pickford M 1981 Preliminary Miocene mammalian biostratigraphy for westernKenya J Hum Evol 10 73e97
Pickford M 1986 Sexual dimorphism in Proconsul Hum Evol 1 111e148Pickford M Andrews PJ 1981 The Tinderet Miocene sequence in Kenya J Hum
Evol 10 11e33Pickford M Kunimatsu Y 2005 Catarrhines from the Middle Miocene (ca 145
Ma) of Kipsaraman Tugen Hills Kenya Anthropol Sci 113 189e224Pickford M Senut B Gommery D Musiime E 2009 Distinctiveness of Ugan-
dapithecus from Proconsul Estudios Geol 65 183e241Pilbeam DR 1969 Tertiary Pongidae of East Africa evolutionary relationships and
taxonomy Bulletin of the Peabody Museum of Natural History 31 pp 1e185Rae TC 1993 Phylogenetic analysis of proconsulid facial morphology PhD
Dissertation State University of New York at Stony BrookRae TC 1999 Mosaic evolution in the origin of the Hominoidea Folia Primatol 70
125e135Rafferty KL Walker A Ruff C Rose MD Andrews PJ 1995 Postcranial esti-
mates of body weights in Proconsul with a note on a distal tibia of P major fromNapak Uganda Am J Phys Anthropol 97 391e402
Rossie JB MacLatchy L 2013 Dentognathic remains of an Afropithecus individualfrom Kalodirr Kenya J Hum Evol 65 199e208
Senut B Pickford M Gommery D Kunimatsu Y 2000 Un nouveau genredrsquohominoiumlde du Miocene inferieur drsquoAfrique orientale Ugandapithecus major(Le Gros Clark and Leakey 1950) CR Acad Sci Paris 331 227e233
Simons EL Pilbeam DR 1965 Preliminary revision of the Dryopithecinae (Pon-gidae Anthropoidea) Folia Primatol 3 81e152
Smith TM Martin LB Leakey MG 2003 Enamel thickness microstructure anddevelopment in Afropithecus turkanensis J Hum Evol 44 283e306
Stevens NJ Seiffert ER OConnor PM Roberts EM Schmitz MD Krause CGorscak E Ngasala S Hieronymus TL Temu J 2013 Palaeontological evi-dence for an Oligocene divergence between Old World monkeys and apesNature 497 611e614
Suwa G Kono RT Katoh S Asfaw B Beyene Y 2007 A new species of great apefrom the late Miocene epoch in Ethiopia Nature 448 921e924
Szalay FS Delson E 1979 Evolutionary History of the Primates Academic PressNew York
Teaford MF Beard KC Leakey RE Walker A 1988 New hominoid facial skel-eton from the Early Miocene of Rusinga Island Kenya and its bearing on therelationship between Proconsul nyanzae and Proconsul africanus J Hum Evol17 461e477
Van Couvering JAH Miller J 1969 Miocene stratigraphy and age determinationsRusinga Island Kenya Nature 221 628e632
Walker A 1997 Proconsul function and phylogeny In Begun DR Ward CVRose MD (Eds) Function Phylogeny and Fossils Miocene Hominoid Evolu-tion and Adaptation Plenum Press New York pp 209e224
Walker A Pickford M 1983 New postcranial fossils of Proconsul africanus andProconsul nyanzae In Ciochon RL Corruccini RS (Eds) New Interpretationsof Ape and Human Ancestry Plenum Press New York pp 325e351
Walker A Teaford M 1988 The Kaswanga primate site an Early Miocene hom-inoid site on Rusinga Island Kenya J Hum Evol 17 539e544
Walker A Teaford M 1989 The hunt for Proconsul Sci Am 260 76e82Walker A Teaford MF Martin L Andrews P 1993 A new species of Proconsul
from the early Miocene of RusingaMfangano Islands Kenya J Hum Evol 2543e56
Ward CV Ruff CB Walker A Rose MD Teaford MF Nengo IO 1995 Func-tional morphology of Proconsul patellas from Rusinga Island Kenya with im-plications for other Miocene-Pliocene catarrhines J Hum Evol 29 1e19
Ward S Brown B Hill A Kelley J Downs W 1999 Equatorius a new hominoidgenus from the Middle Miocene of Kenya Science 285 1382e1386
Werdelin L Sanders WJ 2010 Cenozoic Mammals of Africa University of Cali-fornia Press Berkeley

KP McNulty et al Journal of Human Evolution 84 (2015) 42e6158
some diagnostic features of ldquoUgandapithecusrdquo were found to bevariably present in Ekembo arguing against their utility for dis-tinguishing ldquoUgandapithecusrdquo from Pickford et als (2009) com-bined P africanus thorn Kisingiri conception of Proconsul For examplesignificantly reduced curvature of the I1 distal margin in ldquoUganda-pithecusrdquo is also found in some Kisingiri specimens including KNM-RU 1685 1712 1714 1831 1846 and 1951 several more Rusingaspecimens have moderately reduced curvature Third some fea-tures that purportedly distinguish ldquoUgandapithecusrdquo fromP africanus are likely related to size For example the somewhatmore central position of the P3 paracone in ldquoUrdquomajor is not seen inthe smaller ldquoUrdquo meswae whose paracone is positioned similarly tothat of the holotype of P africanus (see Harrison and Andrews2009 Fig 6d)
However the major part of the evidence for recognizingldquoUgandapithecusrdquo (sensu Pickford et al 2009) was assembled by re-assigning specimens traditionally attributed to P africanus into newspecies of ldquoUgandapithecusrdquo The P africanus holotype was left inassociation with the Rusinga sample despite having morphologyoverwhelmingly characteristic of other Tinderet specimens Theresulting specimen allocations left P africanus morphologicallyimpoverisheddstripped of much of its hypodigmdso that Pro-consul for Pickford et al (2009) came to be characterized primarilyby the Rusinga sample (see similar comments in Harrison andAndrews 2009) Within that conceptual framework any spec-imen that differs from those at Rusinga could not be Proconsul butmust instead belong to ldquoUgandapithecusrdquo Thus specimens such asthe KNM-SO 1112 mandible were erroneously assigned to ldquoUgan-dapithecusrdquo because they were (correctly) perceived as differingfrom their Rusinga counterparts (Pickford et al 2009)
Here we restore the P africanus hypodigm and associate it withthe other Tinderet fossils according to their shared morphologySpecimens assigned to ldquoUgandapithecusrdquo revert to Proconsul andmany of the features cited to support a speciose ldquoUgandapithecusrdquo(Pickford et al 2009) instead become diagnostic features of Pro-consul This correction serves to highlight the distinctiveness of theKisingiri remains and the need to recognize this distinctiveness atthe generic level
As demonstrated above specimens from Kisingiri do not displaythe morphology of Proconsul as represented by the holotype ofP africanus M 14084 Given that this specimen preserves only theupper canine and cheek teeth similarities with or departures fromthis morphology represent the most compelling arguments forassessing inclusivity in the genus However for the mandible andlower dentition if one simply sorts the specimens by elementwithout relying on taxonomic preconceptions the overwhelmingdistinction within the traditionally constituted Proconsul sample isbetween Kisingiri material on the one hand and that from Tinderetand Uganda on the other (cf Walker et al 1993 Begun 2001 2007MacLatchy and Rossie 2005 Harrison and Andrews 2009Harrison 2010) Additionally with the Kisingiri specimensremoved to Ekembo the postcranial differences used to partlyjustify a distinct ldquoUgandapithecusrdquo lose their relevance (seeHarrison and Andrews 2009)
We have differed among ourselves in our past treatment ofP major as generically distinct from the smaller species of Pro-consul (Begun 2007 McNulty 2010) Morphologically many ofthe unique features of P major can be interpreted as size-relatedvariants on the smaller P africanus morphology Even if so thequestion is whether these are sufficiently different and numerousto warrant generic distinction that is do they suggest thatP major belongs in a separate clade from P africanus andP meswae As we have demonstrated differences among thesethree species are certainly fewer and show less morphologicaldistinction than those between Ekembo and any of the three
Nevertheless while we ultimately agree with Harrison andAndrews (2009 see also Begun 2007) that the features dis-tinguishing ldquoUrdquo major from P africanus are consistent with thosethat distinguish congeneric species and are at least partly due todifferences in size we concede the possibility that the largestProconsul species could belong in a separate genus (Senut et al2000 Pickford and Kunimatsu 2005) However whether or notone recognizes a separate genus for P major the P africanus ma-terial does not sort morphologically with the Kisingiri materialTherefore the taxonomic alternatives are either two generaProconsul and Ekembo or three Proconsul Ugandapithecus andEkembo with Proconsul and Ugandapithecus united in a cladeexclusive of Ekembo Thus while we strongly favor the firstalternative recognizing Ugandapithecus does not change thefundamental argument concerning the need to refer the Kisingirispecies to a generically and cladistically distinct taxon from thespecies represented at Tinderet and the Ugandan sites
Specimens of interest
Despite the substantial dental and gnathic differences betweenProconsul and Ekembo there are a few specimens whose taxonomicaffinities require additional scrutiny
KNM-RU 1676-77 This set of associated upper and lower teeth is aconsistent outlier among E heseloni and E nyanzae specimens Insome ways this specimen more closely resembles Tinderet Pro-consul in the features that distinguish it from Ekembo Molar cuspmorphology and cingulum development I1 lingual tubercledevelopment and M2 shape and posthypocone crista orientationare more consistent with specimens from the Tinderet sites On theother hand it otherwise resembles Ekembo and lacks the strongestdiagnostic features that unite species of Proconsul such as thedistinctive canine and P3 morphology Overall this specimen doesnot fit unambiguously into either genus in fact the degree ofbuccolingual flare cusp morphology enamel wrinkling and espe-cially the long extension of enamel onto the anterior root in P3 arein some ways reminiscent of Afropithecus although it clearly differsfrom that genus as well (cf Rossie and MacLatchy 2013)
Were these characteristics individually distributed among thelarge Kisingiri sample one might attribute this to intra-genericvariation but when found together in a single specimen from alarge sample it suggests a unique individual One of us (KPM)thinks it likely that the specimen was not actually found at aKisingiri locality based on its unique preservation compared toother Rusinga fossils and on other examples of mis-proveniencedspecimens (Leakey 1967 Andrews and Molleson 1979 Pickford1986 Kelley 1986) However we recognize that relying toofrequently on this explanation allows almost any scenario to bejustified and suggest some alternative possibilities One is that thespecimen was found on Rusinga but in the much older WayandoFormation strata instead of the Hiwegi Formation The differencesin morphology in that case might reflect more than a million yearsof temporal change (McCollum et al 2013) with KNM-RU 1676-77preserving certain more primitive Proconsul-like features As analternative it is reasonable to assume that fossil assemblages asvast as those from Rusingawill include not only abundant taxa butalso rare or infrequent inhabitants of the local biome There aremany examples of this from Rusingas mammalian communityeg ldquoHemicyonrdquo Kulutherium Tadarida cf Hyainailouros cfArchaeobeledon Kelba (see relevant chapters in Werdelin andSanders 2010) As such KNM-RU 1676-77 may simply representa rare and well-preserved specimen of some other catarrhinetaxon that lived among or visited a primate community dominatedby Ekembo
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 59
KNM-SO 1112 This specimen was initially referred to Rangwapi-thecus but soon after was transferred to Proconsul afrianus (Bosler1981 Martin 1981)da designation that most subsequent authorshave followed (Pickford 1986 Nengo and Rae 1992 Kelley 19861995 Walker et al 1993 MacLatchy and Rossie 2005 Hill et al2013) Pickford et al (2009) more recently referred this specimento ldquoUrdquo meswae citing two primary reasons First they reiteratedBoslers (1981) concern that its P3 differs from others attributedto P africanus However at that time the majority of what wasconsidered to be P africanus would have been from RusingaIsland In fact while noting its contrasting morphology Boslersuggested that differences between KNM-SO 1112 and other P3smight reasonably be attributed to sexual dimorphism notingthat this specimen provides ldquoa not implausible match for the Pafricanus holotype maxilla M 14084rdquo (1981152) Thatassessment is borne out in this study Using the much largersample of Ekembo P3s as a model variation can be seen toencompass specimens with well-developed mesial beaks andthose with more bilaterally compressed crowns and parallelsides The latter morphology is predominantly found amongmale specimens (KNM-RU 1674 1924 and KPS I and III) thussupporting Boslers (1981) suggestion that this variation isrelated to sexual dimorphism and hence that KNM-SO 1112 is amale P africanus
The second feature cited by Pickford et al (2009) in referringthis specimen to ldquoUrdquo meswae was posterior shallowing of themandibular corpus However because they redefined ldquoUgandapi-thecusrdquo to include all of the well-preserved Tinderet mandibles(Pickford et al 2009)dleaving Proconsul to be represented only byRusinga specimensdtheir only alternative within this scheme for aspecimen the size of SO 1112 was to refer it to ldquoUrdquomeswae It seemsodd though to assign this specimen to a species from a (perhapsmuch) older site when the Proconsul africanus holotype is ofappropriate size and similar age and as there are no other adultmandibles from either species with which to compare it In anycase mandibular shallowing is variable in both Ekembo and Pro-consul with some Kisingiri specimens exhibiting considerableshallowing (eg KNM-RU 2087) and some ldquoUgandapithecusrdquo spec-imens having very little (eg M 14086)
We therefore see no morphological grounds to assign KNM-SO1112 to P meswae but do find some morphological evidence toassign it as a male P africanus which occurs at Songhor
KNM-LG 452 This very complete mandible was originally recog-nized as a female of P major based on the size of the dentition androbusticity of the mandible (Martin 1981) Bosler (1981) howeverleft this specimen unassigned noting similarities to P major in P3-M1 size but having distinctly smaller M2e3 In this she likenedKNM-LG 452 to a few other specimens (notably M 14086) thatare smaller in their distal molars in a manner more consistentwith taxonomic rather than sex differences (Bosler 1981 see alsoPilbeam 1969) Kelley (1986 1995) also assigned this specimen asa female of P major based on canine size and morphology butnoted that the canine lacked the shallowly-sloping mesial ridgetypical of most female anthropoid canines This is also truehowever of two other canines assigned as female P major KO 9and SO 373 (Kelley 1986) and so might simply be an unusualcharacteristic of P major female lower canines In the end Kelley(1995) concluded that considering all of the criteria fordistinguishing male and female canines the evidence that KNM-LG 452 is female remains questionable
Pickford et al (2009) further developed the idea that two spe-cies are represented within the P major sample and made KNM-LG452 the holotype of a new species ldquoUrdquo legetetensis We concur thatthis specimen is unique among the well-preserved Tinderet
mandibles in some important features Whereas male and femalemandibles of P major have long internal symphyseal planesexpansive extra- and retromolar sulci and rami that cross theocclusal plane at distal M3 KNM-LG 452 has a shorter symphysealplane (though not to the extent seen in Ekembo) a moderateextramolar sulcus but no retromolar sulcus and a ramus thatcrosses the occlusal plane more mesially If KNM-SO 396 and SO404 represent males and females respectively of P major then themorphology of KNM-LG 452 appears distinct
At present however we do not think there is sufficient evidenceto differentiate this specimen from either P major or P meswaePickford et al (2009) argue that KNM-LG 452 cannot belong to thelatter species because of its larger size The size differences how-ever are well within the range of variation seen in P majorE heseloni and E nyanzae Moreover whereas the Legetet mandibleis larger than most of the P meswae hypodigm an adult P4 referredto this species (KNM-ME 25) substantially exceeds its counterpartin KNM-LG 452 Should the Legetetmandible ultimately prove to bea male specimen size alone would not rule out its attribution toP meswae On the other hand if it is female then it becomes harderto justify its inclusion in that sample of mostly smaller specimens Iffemale therefore it could represent a separate species for whichthe binomial Proconsul legetetensis comb nov is available Recentlycollected fossils from the SonghorKapurtay area provide someadditional support for the possibility that three species of Proconsulexisted at these localities (McNulty 2014ab) However toconvincingly demonstrate this and determine whether a thirdspecies can be differentiated from P meswae will require a moredetailed analysis of variation among Proconsul species a corre-sponding study of variation in modern taxa anddalmost certain-lydadditional fossil specimens For now we regard ldquoUrdquo legetetensisas a subjective junior synonym of P major
Conclusions
The genus Proconsul has been central to studies of ape and hu-man evolution since its first description by Hopwood (1933) As oneof the best known fossil catarrhines represented by nearly everyskeletal element our knowledge of Proconsuls functional anatomypositional behavior and overall morphology has made it thefoundation for paleoanthropological research in the early Mioceneas well as studies on the origin and early diversification of homi-noids With respect to systematics however Proconsul is less wellunderstood the type species P africanus is known from a rela-tively small number of specimens from sites in the Tinderet areaand no postcranial bones can be unambiguously assigned to itRather it is the copious and well-preserved specimens from theKisingiri sites of Rusinga and Mfangano Islands that have formedthe basis for the common image of Proconsul
We argue here that the Kisingiri species traditionally referredto ProconsuldP heseloni and P nyanzaedare generically distinctfrom P africanus and hereby transfer them into a new genusEkembo based on a substantial number of morphological differ-ences (Table 2) Considering only those traits preserved in theP africanus holotype the entire sample from Rusinga and Mfan-gano differs markedly in the morphology of the upper canine P3P4 and M2 Additional differences in the I1 lower incisors andcanines molar cusp and occlusal crest morphology degree of flarein all molars and especially mandibular features clearly distin-guish the Kisingiri from non-Kisingiri samples in a way that isincompatible with congeneric species With the transfer ofE heseloni and E nyanzae Proconsul becomes a well-defined cladecomprising at least three speciesdP africanus P major andP meswaedunited by several features that are unique for thegenus (Table 2)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6160
We found no support in this study for the genus ldquoUgandapi-thecusrdquo as constructed by Pickford et al (2009) In that paper theytransferred the vast majority of Proconsul fossils from the Tinderetand Napak localities into ldquoUgandapithecusrdquo leaving P africanusrepresented only by its holotype and a small number of isolatedteeth and aligning it with the material from Rusinga and Mfan-ganoWhen the P africanus hypodigm is reconstituted and properlyaffiliated with the other Tinderet species according to itsmorphology much of the evidence marshaled by Pickford et al(2009) for ldquoUgandapithecusrdquo actually supports the distinction be-tween the Tinderet and Kisingiri samples ie the generic distinc-tion of Ekembo In addition some of the traits enumerated byPickford and colleagues in support of ldquoUgandapithecusrdquo wererevealed in our analysis to be variable within one or more speciesand therefore of questionable taxonomic utility We do not rejectout of hand the original conception of ldquoUgandapithecusrdquo (Senutet al 2000) as either a monotypic genus distinguishing ldquoUrdquo ma-jor or perhaps including a closely allied middle Miocene ldquoUrdquogitongai as well (Pickford and Kunimatsu 2005) but we have fornow retained the early Miocene specimens of both in Proconsulpending further study However even if Ugandapithecus is a taxo-nomically valid genus comprising just the latter two species theirmorphology indicates that theywould still be united in a cladewithProconsul and they lack several more advanced features that partlydefine Ekembo Better understanding of the role that body size playsinmediating differences within Proconsulwill help to further clarifythe relationships among these species
Despite the existence of one well-represented and several lesscomplete skeletons of what was long assumed to be Proconsul thephylogenetic status of this genus has defied consensus This islargely due in our view to Proconsul having been an admixture oftwo distinct genera one more advanced in the direction of extantapes than the other As we demonstrate here the material fromRusinga and Mfangano traditionally referred to Proconsul andwhich includes all of the more complete skeletal material isdistinct from the Tinderet and Napak material and belongs in aseparate genus Ekembo By disentangling these distinct clades weanticipate being able to achieve better phylogenetic resolution forboth genera thereby improving our knowledge of early hominoidevolution and diversification
Acknowledgments
We thank many people and agencies that assisted with thedevelopment and completion of this project This research wasinitiated as a result of funding from the Wenner-Gren Foundationand the Johns Hopkins School of Medicine to DR Begun in1987e1989 and was renewed following research and travelgenerously funded by multiple Leakey Foundation grants to KMcNulty National Science Foundation grants to K McNulty(0852609 1241807) the McKnight Foundation the University ofMinnesota the Lloyd A Wilford Endowment the LeverhulmeFoundation and the Turkana Basin Institute J Kelley gratefullyacknowledges support from the Institute of Human Origins Ari-zona State University DR Begun also wishes to acknowledge theAlexander Humboldt Stiftung and NSERC for funding support Weare grateful to the National Museums of Kenya the UgandaMuseum the Natural History Museum and to Rose Mwanja EzraMusiime Tom Mukhuyu Samuel Muteti James Yatich and MaryMuungu for access to and assistance with specimens in their careMany people provided assistance comments criticisms insightsand suggestions on various aspects of this work and we especiallythank Holly Dunsworth Will Harcourt-Smith Tom Lehmann DanPeppe Nicole Garrett Kirsten Jenkins Rutger Jansma Susy CoteIsaiah Nengo Andrew Hill and Brenda Benefit K McNulty
acknowledges the evolutionary anthropology faculty and studentsat Durham University for their helpful comments on an earlyversion of this manuscript and for their gracious hospitality Weare likewise grateful to the anonymous reviewers associate editorand editor whose comments strengthened the paper This manu-script is publication 1 supporting Research on East AfricanCatarrhine and Hominoid Evolution (REACHE)
Appendix A Supplementary data
Supplementary data related to this article can be found at httpdxdoiorg101016jjhevol201503009
References
Andrews P 1974 New species of Dryopithecus from Kenya Nature 249 188e190Andrews P 1978 A revision of the Miocene Hominoidea of East Africa Bull Brit
Mus (Nat Hist) Geol 30 85e225Andrews P 1985 Family group systematics and evolution among catarrhine pri-
mates In Delson E (Ed) Ancestors The Hard Evidence Liss New Yorkpp 14e22
Andrews P Harrison T Martin L Pickford M 1981 Hominoid primates from anew Miocene locality named Meswa Bridge in Kenya J Hum Evol 10123e128
Andrews P Begun D Zylstra M 1997 Interrelationships between functionalmorphology and paleoenvironments in Miocene hominoids In Begun DWard C Rose M (Eds) Function Phylogeny and Fossils Miocene HominoidEvolution and Adaptations Plenum Press New York pp 29e58
Andrews P Molleson T 1979 The provenance of Sivapithecus africanus Bull BritMus (Nat Hist) Geol 32 19e23
Begun DR 2001 African and Eurasian Miocene hominoids and the origins of theHominidae In de Bonis L Koufos GD Andrews P (Eds) Hominoid Evolutionand Climatic Change in Europe Phylogeny of the Neogene Hominoid Primatesof Eurasia Cambridge University Press London pp 231e253
Begun DR 2007 Fossil record of Miocene hominoids In Henke W Tattersall I(Eds) Handbook of Paleoanthropology Springer Berlin pp 921e977
Begun DR Teaford MF Walker AC 1994 Comparative and functional anatomyof Proconsul phalanges from the Kaswanga primate site Rusinga Island KenyaJ Hum Evol 26 89e165
Bishop WW 1964 More fossil primates and other Miocene mammals from North-East Uganda Nature 203 1327e1331
Bishop WW Miller JA Fitch FJ 1969 New potassium-argon age determinationsrelevant to the Miocene fossil mammal sequence in East Africa Am J Sci 267669e699
Blankenship JA Mincer HH Anderson KM Woods MA Burton EL 2007Third molar development in the estimation of chronologic age in Americanblacks as compared with whites J Forensic Sci 52 428e433
Bosler W 1981 Species groupings of early Miocene dryopithecine teeth from EastAfrica J Hum Evol 10 151e158
Coleman MN 2008 What does geometric mean mean geometrically Assessingthe utility of geometric mean and other size variables in studies of skullallometry Am J Phys Anthropol 135 404e415
Cote S Malit N Nengo I 2013 Additional mandibles of Rangwapithecus gordonian early Miocene catarrhine from the Tinderet localities of Western KenyaJ Hum Evol 153 341e352
Deane AS 2009 Early Miocene catarrhine dietary behavior the influence of theRed Queen Effect on incisor shape and curvature J Hum Evol 56 275e285
Drake R Van Couvering JA Pickford M Curtis G Harris JA 1988 New chro-nology for the Early Miocene mammalian faunas of Kisingiri Western KenyaJ Geol Soc Lond 145 479e491
Evernden JF Savage DE Curtis GH James GT 1964 Potassium-argon datesand the Cenozoic mammalian chronology of North America Am J Sci 262145e198
Gebo DL MacLatchy L Kityo R Deino A Kingston J Pilbeam D 1997A hominoid genus from the Early Miocene of Uganda Science 276 401e404
Geoffroy Saint-Hilaire E 1812 Tableau des quadrumanes 1 Ord QuadrumanesAnnls Mus Hist Nat Paris 19 85e122
Gray JE 1825 Outline of an attempt at the disposition of the Mammalia into tribesand families with a list of the genera apparently appertaining to each tribeAnnls Philos 10 337e344
Harrison T 1982 Small-bodied apes from the Miocene of East Africa PhDDissertation University of London
Harrison T 1986 New fossil anthropoids from the middle Miocene of East Africaand their bearing on the origin of the Oreopithecidae Am J Phys Anthropol 71265e284
Harrison T 1987 The phylogenetic relationships of the early catarrhine primates areview of the current evidence J Hum Evol 16 41e80
Harrison T 2002 Late Oligocene to middle Miocene catarrhines from Afro-ArabiaIn Hartwig WC (Ed) The Primate Fossil Record Cambridge University PressCambridge pp 311e338
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 61
Harrison T 2010 Dendropithecoidea Proconsuloidea and Hominoidea (CatarrhiniPrimates) In Werdelin L Sanders WJ (Eds) Cenozoic Mammals of AfricaUniversity of California Press Berkeley pp 429e469
Harrison T Andrews P 2009 The anatomy and systematic position of a newspecies of early Miocene proconsulid from Meswa Bridge Kenya J Hum Evol56 479e496
Hill A Nengo IO Rossie JB 2013 A Rangwapithecus gordoni mandible from theearly Miocene site of Songhor Kenya J Hum Evol 65 490e500
Hopwood AT 1933 Miocene primates from Kenya J Linn Soc Zool 38 437e464Ishida H Kunimatsu Y Takano T Nakano Y Nakatsukasa M 2004 Nacholapi-
thecus skeleton from the Middle Miocene of Kenya J Hum Evol 46 69e103Jolicouer P 1963 The multivariate generalization of the allometry equation Bio-
metrics 19 497e499Jungers WL Falsetti AB Wall CE 1995 Shape relative size and size-
adjustments in morphometrics Am J Phys Anthropol 38 (S2) 137e161Kavanagh KD Evans AR Jernvall J 2007 Predicting evolutionary patterns of
mammalian teeth from development Nature 449 427e433Kelley J 1986 Species recognition and sexual dimorphism in Proconsul and
Rangwapithecus J Hum Evol 15 461e495Kelley J 1995 Sex determination in Miocene catarrhine primates Am J Phys
Anthropol 96 390e417Kelley J 1993 Taxonomic implications of sexual dimorphism in Lufengpithecus In
Kimbel WH Martin LB (Eds) Species Species Concepts and Primate Evo-lution Springer New York pp 429e458
Kent PE 1942 The country round the Kavirondo Gulf of Victoria Nyanza Geogr J100 22e31
Kordos L Begun DR 2001 A new cranium of Dryopithecus from RudabanyaHungary J Hum Evol 15 689e700
Le Gros Clark WE Leakey LSB 1950 Diagnoses of East African Miocene Homi-noidea Q J Geol Soc Lond 105 260e263
Le Gros Clark WE Leakey LSB 1951 The Miocene Hominoidea of East Africa InFossil Mammals of Africa No 1 British Museum (Natural History) London
Leakey LSB 1943 A Miocene anthropoid mandible from Rusinga Kenya Nature152 319e320
Leakey LSB 1963 East African fossil Hominoidea and the classification within thissuper-family In Washburn SL (Ed) Classification and Human EvolutionAldine Chicago pp 32e49
Leakey LSB 1967 Notes on the mammalian faunas from the Miocene and Pleis-tocene of East Africa In Bishop WW Clark JD (Eds) Background to Evolu-tion in Africa University of Chicago Press Chicago pp 7e29
Leakey RE Leakey MG 1986 A new Miocene hominoid from Kenya Nature 324143e146
Leakey RE Leakey MG Walker AC 1988 Morphology of Afropithecus turka-nensis from Kenya Am J Phys Anthropol 76 289e307
Leakey MG Ungar PS Walker A 1995 A new genus of large primate from theLate Oligocene of Lothidok Turkana District Kenya J Hum Evol 28519e531
Linnaeus C 1758 Systema naturae per regna tria naturae secundum classesordines genera species cum characteribus differentris synonymis locis tenthed Laurentii Salvii Stockholm
MacInnes DG 1943 Notes on the East African Miocene primates J E Afr UgandaNat Hist Soc 17 141e181
MacLatchy L 2004 The oldest ape Evol Anthropol 13 90e103MacLatchy L Bossert WH 1996 An analysis of the articular surface distribution of
the femoral head and acetabulum in anthropoids with implications for hipfunction in Miocene hominoids J Hum Evol 31 425e453
MacLatchy L Rossie JB 2005 The Napak hominoid still Proconsul major InLieberman DE Smith RJ Kelley J (Eds) Interpreting the Past Essays onHuman Primate and Mammal Evolution in Honor of David Pilbeam Brill Ac-ademic Publishers Boston pp 15e28
Martin L 1981 New specimens of Proconsul from Koru Kenya J Hum Evol 10139e150
McCollum MS Peppe DJ McNulty KP Dunsworth HM Harcourt-Smith WEH Andrews AL 2013 Magnetostratigraphy of the early MioceneHiwegi Formation (Rusinga Island Lake Victoria Kenya) Geological Society ofAmerica Abstracts with Programs 45 p 12
McNulty KP 2010 Apes and tricksters the evolution and diversification ofhumans closest relatives Evol Edu Outreach 3 322e332
McNulty KP Harcourt-Smith WEH Dunsworth HM 2007 New primate fossilsfrom Rusinga Island Kenya Am J Phys Anthropol 132 (S44) 170
McNulty KP MacLatchy L Peppe DJ Nengo I Manthi FK Miller ERStevens NJ Cote S Lehmann T Kingston J 2014a A regional approach toEast African early Miocene paleobiology J Vert Paleontol 185 SVP Programand Abstracts Book 2014
McNulty KP MacLatchy L Rossie JB Peppe DJ Deino AL Mbua ENManthi FK Nengo IO Stevens NJ Cote S Lehmann T Gutierrez M 2014b
Research on East African catarrhine and hominoid evolution results from thefirst year Am J Phys Anthropol 153 (S58) 182
Michel LA Peppe DJ Lutz JA Driese SG Dunsworth HM Harcourt-Smith WEH Horner W Lehmann T Nightengale S McNulty KP 2014Remnants of an ancient forest provide ecological context for Early Miocenefossil apes Nature Comm 5 3236
Myres JL 1932 Africa Man 32 208Nakatsukasa M Kunimatsu Y Nakano Y Takano T Ishida H 2007 Vertebral
morphology of Nacholapithecus kerioi based on KNM-BG 35250 J Hum Evol52 347e369
Nengo IO Rae TC 1992 New hominoid fossils from the early Miocene site ofSonghor Kenya J Hum Evol 23 423e429
Peppe DJ McNulty KP Cote SM Harcourt-Smith WEH Dunsworth HM VanCouvering JA 2009 Stratigraphic interpretation of the Kulu Formation (EarlyMiocene Rusinga Island Kenya) and its implications for primate evolutionJ Hum Evol 56 447e461
Pickford M 1981 Preliminary Miocene mammalian biostratigraphy for westernKenya J Hum Evol 10 73e97
Pickford M 1986 Sexual dimorphism in Proconsul Hum Evol 1 111e148Pickford M Andrews PJ 1981 The Tinderet Miocene sequence in Kenya J Hum
Evol 10 11e33Pickford M Kunimatsu Y 2005 Catarrhines from the Middle Miocene (ca 145
Ma) of Kipsaraman Tugen Hills Kenya Anthropol Sci 113 189e224Pickford M Senut B Gommery D Musiime E 2009 Distinctiveness of Ugan-
dapithecus from Proconsul Estudios Geol 65 183e241Pilbeam DR 1969 Tertiary Pongidae of East Africa evolutionary relationships and
taxonomy Bulletin of the Peabody Museum of Natural History 31 pp 1e185Rae TC 1993 Phylogenetic analysis of proconsulid facial morphology PhD
Dissertation State University of New York at Stony BrookRae TC 1999 Mosaic evolution in the origin of the Hominoidea Folia Primatol 70
125e135Rafferty KL Walker A Ruff C Rose MD Andrews PJ 1995 Postcranial esti-
mates of body weights in Proconsul with a note on a distal tibia of P major fromNapak Uganda Am J Phys Anthropol 97 391e402
Rossie JB MacLatchy L 2013 Dentognathic remains of an Afropithecus individualfrom Kalodirr Kenya J Hum Evol 65 199e208
Senut B Pickford M Gommery D Kunimatsu Y 2000 Un nouveau genredrsquohominoiumlde du Miocene inferieur drsquoAfrique orientale Ugandapithecus major(Le Gros Clark and Leakey 1950) CR Acad Sci Paris 331 227e233
Simons EL Pilbeam DR 1965 Preliminary revision of the Dryopithecinae (Pon-gidae Anthropoidea) Folia Primatol 3 81e152
Smith TM Martin LB Leakey MG 2003 Enamel thickness microstructure anddevelopment in Afropithecus turkanensis J Hum Evol 44 283e306
Stevens NJ Seiffert ER OConnor PM Roberts EM Schmitz MD Krause CGorscak E Ngasala S Hieronymus TL Temu J 2013 Palaeontological evi-dence for an Oligocene divergence between Old World monkeys and apesNature 497 611e614
Suwa G Kono RT Katoh S Asfaw B Beyene Y 2007 A new species of great apefrom the late Miocene epoch in Ethiopia Nature 448 921e924
Szalay FS Delson E 1979 Evolutionary History of the Primates Academic PressNew York
Teaford MF Beard KC Leakey RE Walker A 1988 New hominoid facial skel-eton from the Early Miocene of Rusinga Island Kenya and its bearing on therelationship between Proconsul nyanzae and Proconsul africanus J Hum Evol17 461e477
Van Couvering JAH Miller J 1969 Miocene stratigraphy and age determinationsRusinga Island Kenya Nature 221 628e632
Walker A 1997 Proconsul function and phylogeny In Begun DR Ward CVRose MD (Eds) Function Phylogeny and Fossils Miocene Hominoid Evolu-tion and Adaptation Plenum Press New York pp 209e224
Walker A Pickford M 1983 New postcranial fossils of Proconsul africanus andProconsul nyanzae In Ciochon RL Corruccini RS (Eds) New Interpretationsof Ape and Human Ancestry Plenum Press New York pp 325e351
Walker A Teaford M 1988 The Kaswanga primate site an Early Miocene hom-inoid site on Rusinga Island Kenya J Hum Evol 17 539e544
Walker A Teaford M 1989 The hunt for Proconsul Sci Am 260 76e82Walker A Teaford MF Martin L Andrews P 1993 A new species of Proconsul
from the early Miocene of RusingaMfangano Islands Kenya J Hum Evol 2543e56
Ward CV Ruff CB Walker A Rose MD Teaford MF Nengo IO 1995 Func-tional morphology of Proconsul patellas from Rusinga Island Kenya with im-plications for other Miocene-Pliocene catarrhines J Hum Evol 29 1e19
Ward S Brown B Hill A Kelley J Downs W 1999 Equatorius a new hominoidgenus from the Middle Miocene of Kenya Science 285 1382e1386
Werdelin L Sanders WJ 2010 Cenozoic Mammals of Africa University of Cali-fornia Press Berkeley

KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 59
KNM-SO 1112 This specimen was initially referred to Rangwapi-thecus but soon after was transferred to Proconsul afrianus (Bosler1981 Martin 1981)da designation that most subsequent authorshave followed (Pickford 1986 Nengo and Rae 1992 Kelley 19861995 Walker et al 1993 MacLatchy and Rossie 2005 Hill et al2013) Pickford et al (2009) more recently referred this specimento ldquoUrdquo meswae citing two primary reasons First they reiteratedBoslers (1981) concern that its P3 differs from others attributedto P africanus However at that time the majority of what wasconsidered to be P africanus would have been from RusingaIsland In fact while noting its contrasting morphology Boslersuggested that differences between KNM-SO 1112 and other P3smight reasonably be attributed to sexual dimorphism notingthat this specimen provides ldquoa not implausible match for the Pafricanus holotype maxilla M 14084rdquo (1981152) Thatassessment is borne out in this study Using the much largersample of Ekembo P3s as a model variation can be seen toencompass specimens with well-developed mesial beaks andthose with more bilaterally compressed crowns and parallelsides The latter morphology is predominantly found amongmale specimens (KNM-RU 1674 1924 and KPS I and III) thussupporting Boslers (1981) suggestion that this variation isrelated to sexual dimorphism and hence that KNM-SO 1112 is amale P africanus
The second feature cited by Pickford et al (2009) in referringthis specimen to ldquoUrdquo meswae was posterior shallowing of themandibular corpus However because they redefined ldquoUgandapi-thecusrdquo to include all of the well-preserved Tinderet mandibles(Pickford et al 2009)dleaving Proconsul to be represented only byRusinga specimensdtheir only alternative within this scheme for aspecimen the size of SO 1112 was to refer it to ldquoUrdquomeswae It seemsodd though to assign this specimen to a species from a (perhapsmuch) older site when the Proconsul africanus holotype is ofappropriate size and similar age and as there are no other adultmandibles from either species with which to compare it In anycase mandibular shallowing is variable in both Ekembo and Pro-consul with some Kisingiri specimens exhibiting considerableshallowing (eg KNM-RU 2087) and some ldquoUgandapithecusrdquo spec-imens having very little (eg M 14086)
We therefore see no morphological grounds to assign KNM-SO1112 to P meswae but do find some morphological evidence toassign it as a male P africanus which occurs at Songhor
KNM-LG 452 This very complete mandible was originally recog-nized as a female of P major based on the size of the dentition androbusticity of the mandible (Martin 1981) Bosler (1981) howeverleft this specimen unassigned noting similarities to P major in P3-M1 size but having distinctly smaller M2e3 In this she likenedKNM-LG 452 to a few other specimens (notably M 14086) thatare smaller in their distal molars in a manner more consistentwith taxonomic rather than sex differences (Bosler 1981 see alsoPilbeam 1969) Kelley (1986 1995) also assigned this specimen asa female of P major based on canine size and morphology butnoted that the canine lacked the shallowly-sloping mesial ridgetypical of most female anthropoid canines This is also truehowever of two other canines assigned as female P major KO 9and SO 373 (Kelley 1986) and so might simply be an unusualcharacteristic of P major female lower canines In the end Kelley(1995) concluded that considering all of the criteria fordistinguishing male and female canines the evidence that KNM-LG 452 is female remains questionable
Pickford et al (2009) further developed the idea that two spe-cies are represented within the P major sample and made KNM-LG452 the holotype of a new species ldquoUrdquo legetetensis We concur thatthis specimen is unique among the well-preserved Tinderet
mandibles in some important features Whereas male and femalemandibles of P major have long internal symphyseal planesexpansive extra- and retromolar sulci and rami that cross theocclusal plane at distal M3 KNM-LG 452 has a shorter symphysealplane (though not to the extent seen in Ekembo) a moderateextramolar sulcus but no retromolar sulcus and a ramus thatcrosses the occlusal plane more mesially If KNM-SO 396 and SO404 represent males and females respectively of P major then themorphology of KNM-LG 452 appears distinct
At present however we do not think there is sufficient evidenceto differentiate this specimen from either P major or P meswaePickford et al (2009) argue that KNM-LG 452 cannot belong to thelatter species because of its larger size The size differences how-ever are well within the range of variation seen in P majorE heseloni and E nyanzae Moreover whereas the Legetet mandibleis larger than most of the P meswae hypodigm an adult P4 referredto this species (KNM-ME 25) substantially exceeds its counterpartin KNM-LG 452 Should the Legetetmandible ultimately prove to bea male specimen size alone would not rule out its attribution toP meswae On the other hand if it is female then it becomes harderto justify its inclusion in that sample of mostly smaller specimens Iffemale therefore it could represent a separate species for whichthe binomial Proconsul legetetensis comb nov is available Recentlycollected fossils from the SonghorKapurtay area provide someadditional support for the possibility that three species of Proconsulexisted at these localities (McNulty 2014ab) However toconvincingly demonstrate this and determine whether a thirdspecies can be differentiated from P meswae will require a moredetailed analysis of variation among Proconsul species a corre-sponding study of variation in modern taxa anddalmost certain-lydadditional fossil specimens For now we regard ldquoUrdquo legetetensisas a subjective junior synonym of P major
Conclusions
The genus Proconsul has been central to studies of ape and hu-man evolution since its first description by Hopwood (1933) As oneof the best known fossil catarrhines represented by nearly everyskeletal element our knowledge of Proconsuls functional anatomypositional behavior and overall morphology has made it thefoundation for paleoanthropological research in the early Mioceneas well as studies on the origin and early diversification of homi-noids With respect to systematics however Proconsul is less wellunderstood the type species P africanus is known from a rela-tively small number of specimens from sites in the Tinderet areaand no postcranial bones can be unambiguously assigned to itRather it is the copious and well-preserved specimens from theKisingiri sites of Rusinga and Mfangano Islands that have formedthe basis for the common image of Proconsul
We argue here that the Kisingiri species traditionally referredto ProconsuldP heseloni and P nyanzaedare generically distinctfrom P africanus and hereby transfer them into a new genusEkembo based on a substantial number of morphological differ-ences (Table 2) Considering only those traits preserved in theP africanus holotype the entire sample from Rusinga and Mfan-gano differs markedly in the morphology of the upper canine P3P4 and M2 Additional differences in the I1 lower incisors andcanines molar cusp and occlusal crest morphology degree of flarein all molars and especially mandibular features clearly distin-guish the Kisingiri from non-Kisingiri samples in a way that isincompatible with congeneric species With the transfer ofE heseloni and E nyanzae Proconsul becomes a well-defined cladecomprising at least three speciesdP africanus P major andP meswaedunited by several features that are unique for thegenus (Table 2)
KP McNulty et al Journal of Human Evolution 84 (2015) 42e6160
We found no support in this study for the genus ldquoUgandapi-thecusrdquo as constructed by Pickford et al (2009) In that paper theytransferred the vast majority of Proconsul fossils from the Tinderetand Napak localities into ldquoUgandapithecusrdquo leaving P africanusrepresented only by its holotype and a small number of isolatedteeth and aligning it with the material from Rusinga and Mfan-ganoWhen the P africanus hypodigm is reconstituted and properlyaffiliated with the other Tinderet species according to itsmorphology much of the evidence marshaled by Pickford et al(2009) for ldquoUgandapithecusrdquo actually supports the distinction be-tween the Tinderet and Kisingiri samples ie the generic distinc-tion of Ekembo In addition some of the traits enumerated byPickford and colleagues in support of ldquoUgandapithecusrdquo wererevealed in our analysis to be variable within one or more speciesand therefore of questionable taxonomic utility We do not rejectout of hand the original conception of ldquoUgandapithecusrdquo (Senutet al 2000) as either a monotypic genus distinguishing ldquoUrdquo ma-jor or perhaps including a closely allied middle Miocene ldquoUrdquogitongai as well (Pickford and Kunimatsu 2005) but we have fornow retained the early Miocene specimens of both in Proconsulpending further study However even if Ugandapithecus is a taxo-nomically valid genus comprising just the latter two species theirmorphology indicates that theywould still be united in a cladewithProconsul and they lack several more advanced features that partlydefine Ekembo Better understanding of the role that body size playsinmediating differences within Proconsulwill help to further clarifythe relationships among these species
Despite the existence of one well-represented and several lesscomplete skeletons of what was long assumed to be Proconsul thephylogenetic status of this genus has defied consensus This islargely due in our view to Proconsul having been an admixture oftwo distinct genera one more advanced in the direction of extantapes than the other As we demonstrate here the material fromRusinga and Mfangano traditionally referred to Proconsul andwhich includes all of the more complete skeletal material isdistinct from the Tinderet and Napak material and belongs in aseparate genus Ekembo By disentangling these distinct clades weanticipate being able to achieve better phylogenetic resolution forboth genera thereby improving our knowledge of early hominoidevolution and diversification
Acknowledgments
We thank many people and agencies that assisted with thedevelopment and completion of this project This research wasinitiated as a result of funding from the Wenner-Gren Foundationand the Johns Hopkins School of Medicine to DR Begun in1987e1989 and was renewed following research and travelgenerously funded by multiple Leakey Foundation grants to KMcNulty National Science Foundation grants to K McNulty(0852609 1241807) the McKnight Foundation the University ofMinnesota the Lloyd A Wilford Endowment the LeverhulmeFoundation and the Turkana Basin Institute J Kelley gratefullyacknowledges support from the Institute of Human Origins Ari-zona State University DR Begun also wishes to acknowledge theAlexander Humboldt Stiftung and NSERC for funding support Weare grateful to the National Museums of Kenya the UgandaMuseum the Natural History Museum and to Rose Mwanja EzraMusiime Tom Mukhuyu Samuel Muteti James Yatich and MaryMuungu for access to and assistance with specimens in their careMany people provided assistance comments criticisms insightsand suggestions on various aspects of this work and we especiallythank Holly Dunsworth Will Harcourt-Smith Tom Lehmann DanPeppe Nicole Garrett Kirsten Jenkins Rutger Jansma Susy CoteIsaiah Nengo Andrew Hill and Brenda Benefit K McNulty
acknowledges the evolutionary anthropology faculty and studentsat Durham University for their helpful comments on an earlyversion of this manuscript and for their gracious hospitality Weare likewise grateful to the anonymous reviewers associate editorand editor whose comments strengthened the paper This manu-script is publication 1 supporting Research on East AfricanCatarrhine and Hominoid Evolution (REACHE)
Appendix A Supplementary data
Supplementary data related to this article can be found at httpdxdoiorg101016jjhevol201503009
References
Andrews P 1974 New species of Dryopithecus from Kenya Nature 249 188e190Andrews P 1978 A revision of the Miocene Hominoidea of East Africa Bull Brit
Mus (Nat Hist) Geol 30 85e225Andrews P 1985 Family group systematics and evolution among catarrhine pri-
mates In Delson E (Ed) Ancestors The Hard Evidence Liss New Yorkpp 14e22
Andrews P Harrison T Martin L Pickford M 1981 Hominoid primates from anew Miocene locality named Meswa Bridge in Kenya J Hum Evol 10123e128
Andrews P Begun D Zylstra M 1997 Interrelationships between functionalmorphology and paleoenvironments in Miocene hominoids In Begun DWard C Rose M (Eds) Function Phylogeny and Fossils Miocene HominoidEvolution and Adaptations Plenum Press New York pp 29e58
Andrews P Molleson T 1979 The provenance of Sivapithecus africanus Bull BritMus (Nat Hist) Geol 32 19e23
Begun DR 2001 African and Eurasian Miocene hominoids and the origins of theHominidae In de Bonis L Koufos GD Andrews P (Eds) Hominoid Evolutionand Climatic Change in Europe Phylogeny of the Neogene Hominoid Primatesof Eurasia Cambridge University Press London pp 231e253
Begun DR 2007 Fossil record of Miocene hominoids In Henke W Tattersall I(Eds) Handbook of Paleoanthropology Springer Berlin pp 921e977
Begun DR Teaford MF Walker AC 1994 Comparative and functional anatomyof Proconsul phalanges from the Kaswanga primate site Rusinga Island KenyaJ Hum Evol 26 89e165
Bishop WW 1964 More fossil primates and other Miocene mammals from North-East Uganda Nature 203 1327e1331
Bishop WW Miller JA Fitch FJ 1969 New potassium-argon age determinationsrelevant to the Miocene fossil mammal sequence in East Africa Am J Sci 267669e699
Blankenship JA Mincer HH Anderson KM Woods MA Burton EL 2007Third molar development in the estimation of chronologic age in Americanblacks as compared with whites J Forensic Sci 52 428e433
Bosler W 1981 Species groupings of early Miocene dryopithecine teeth from EastAfrica J Hum Evol 10 151e158
Coleman MN 2008 What does geometric mean mean geometrically Assessingthe utility of geometric mean and other size variables in studies of skullallometry Am J Phys Anthropol 135 404e415
Cote S Malit N Nengo I 2013 Additional mandibles of Rangwapithecus gordonian early Miocene catarrhine from the Tinderet localities of Western KenyaJ Hum Evol 153 341e352
Deane AS 2009 Early Miocene catarrhine dietary behavior the influence of theRed Queen Effect on incisor shape and curvature J Hum Evol 56 275e285
Drake R Van Couvering JA Pickford M Curtis G Harris JA 1988 New chro-nology for the Early Miocene mammalian faunas of Kisingiri Western KenyaJ Geol Soc Lond 145 479e491
Evernden JF Savage DE Curtis GH James GT 1964 Potassium-argon datesand the Cenozoic mammalian chronology of North America Am J Sci 262145e198
Gebo DL MacLatchy L Kityo R Deino A Kingston J Pilbeam D 1997A hominoid genus from the Early Miocene of Uganda Science 276 401e404
Geoffroy Saint-Hilaire E 1812 Tableau des quadrumanes 1 Ord QuadrumanesAnnls Mus Hist Nat Paris 19 85e122
Gray JE 1825 Outline of an attempt at the disposition of the Mammalia into tribesand families with a list of the genera apparently appertaining to each tribeAnnls Philos 10 337e344
Harrison T 1982 Small-bodied apes from the Miocene of East Africa PhDDissertation University of London
Harrison T 1986 New fossil anthropoids from the middle Miocene of East Africaand their bearing on the origin of the Oreopithecidae Am J Phys Anthropol 71265e284
Harrison T 1987 The phylogenetic relationships of the early catarrhine primates areview of the current evidence J Hum Evol 16 41e80
Harrison T 2002 Late Oligocene to middle Miocene catarrhines from Afro-ArabiaIn Hartwig WC (Ed) The Primate Fossil Record Cambridge University PressCambridge pp 311e338
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 61
Harrison T 2010 Dendropithecoidea Proconsuloidea and Hominoidea (CatarrhiniPrimates) In Werdelin L Sanders WJ (Eds) Cenozoic Mammals of AfricaUniversity of California Press Berkeley pp 429e469
Harrison T Andrews P 2009 The anatomy and systematic position of a newspecies of early Miocene proconsulid from Meswa Bridge Kenya J Hum Evol56 479e496
Hill A Nengo IO Rossie JB 2013 A Rangwapithecus gordoni mandible from theearly Miocene site of Songhor Kenya J Hum Evol 65 490e500
Hopwood AT 1933 Miocene primates from Kenya J Linn Soc Zool 38 437e464Ishida H Kunimatsu Y Takano T Nakano Y Nakatsukasa M 2004 Nacholapi-
thecus skeleton from the Middle Miocene of Kenya J Hum Evol 46 69e103Jolicouer P 1963 The multivariate generalization of the allometry equation Bio-
metrics 19 497e499Jungers WL Falsetti AB Wall CE 1995 Shape relative size and size-
adjustments in morphometrics Am J Phys Anthropol 38 (S2) 137e161Kavanagh KD Evans AR Jernvall J 2007 Predicting evolutionary patterns of
mammalian teeth from development Nature 449 427e433Kelley J 1986 Species recognition and sexual dimorphism in Proconsul and
Rangwapithecus J Hum Evol 15 461e495Kelley J 1995 Sex determination in Miocene catarrhine primates Am J Phys
Anthropol 96 390e417Kelley J 1993 Taxonomic implications of sexual dimorphism in Lufengpithecus In
Kimbel WH Martin LB (Eds) Species Species Concepts and Primate Evo-lution Springer New York pp 429e458
Kent PE 1942 The country round the Kavirondo Gulf of Victoria Nyanza Geogr J100 22e31
Kordos L Begun DR 2001 A new cranium of Dryopithecus from RudabanyaHungary J Hum Evol 15 689e700
Le Gros Clark WE Leakey LSB 1950 Diagnoses of East African Miocene Homi-noidea Q J Geol Soc Lond 105 260e263
Le Gros Clark WE Leakey LSB 1951 The Miocene Hominoidea of East Africa InFossil Mammals of Africa No 1 British Museum (Natural History) London
Leakey LSB 1943 A Miocene anthropoid mandible from Rusinga Kenya Nature152 319e320
Leakey LSB 1963 East African fossil Hominoidea and the classification within thissuper-family In Washburn SL (Ed) Classification and Human EvolutionAldine Chicago pp 32e49
Leakey LSB 1967 Notes on the mammalian faunas from the Miocene and Pleis-tocene of East Africa In Bishop WW Clark JD (Eds) Background to Evolu-tion in Africa University of Chicago Press Chicago pp 7e29
Leakey RE Leakey MG 1986 A new Miocene hominoid from Kenya Nature 324143e146
Leakey RE Leakey MG Walker AC 1988 Morphology of Afropithecus turka-nensis from Kenya Am J Phys Anthropol 76 289e307
Leakey MG Ungar PS Walker A 1995 A new genus of large primate from theLate Oligocene of Lothidok Turkana District Kenya J Hum Evol 28519e531
Linnaeus C 1758 Systema naturae per regna tria naturae secundum classesordines genera species cum characteribus differentris synonymis locis tenthed Laurentii Salvii Stockholm
MacInnes DG 1943 Notes on the East African Miocene primates J E Afr UgandaNat Hist Soc 17 141e181
MacLatchy L 2004 The oldest ape Evol Anthropol 13 90e103MacLatchy L Bossert WH 1996 An analysis of the articular surface distribution of
the femoral head and acetabulum in anthropoids with implications for hipfunction in Miocene hominoids J Hum Evol 31 425e453
MacLatchy L Rossie JB 2005 The Napak hominoid still Proconsul major InLieberman DE Smith RJ Kelley J (Eds) Interpreting the Past Essays onHuman Primate and Mammal Evolution in Honor of David Pilbeam Brill Ac-ademic Publishers Boston pp 15e28
Martin L 1981 New specimens of Proconsul from Koru Kenya J Hum Evol 10139e150
McCollum MS Peppe DJ McNulty KP Dunsworth HM Harcourt-Smith WEH Andrews AL 2013 Magnetostratigraphy of the early MioceneHiwegi Formation (Rusinga Island Lake Victoria Kenya) Geological Society ofAmerica Abstracts with Programs 45 p 12
McNulty KP 2010 Apes and tricksters the evolution and diversification ofhumans closest relatives Evol Edu Outreach 3 322e332
McNulty KP Harcourt-Smith WEH Dunsworth HM 2007 New primate fossilsfrom Rusinga Island Kenya Am J Phys Anthropol 132 (S44) 170
McNulty KP MacLatchy L Peppe DJ Nengo I Manthi FK Miller ERStevens NJ Cote S Lehmann T Kingston J 2014a A regional approach toEast African early Miocene paleobiology J Vert Paleontol 185 SVP Programand Abstracts Book 2014
McNulty KP MacLatchy L Rossie JB Peppe DJ Deino AL Mbua ENManthi FK Nengo IO Stevens NJ Cote S Lehmann T Gutierrez M 2014b
Research on East African catarrhine and hominoid evolution results from thefirst year Am J Phys Anthropol 153 (S58) 182
Michel LA Peppe DJ Lutz JA Driese SG Dunsworth HM Harcourt-Smith WEH Horner W Lehmann T Nightengale S McNulty KP 2014Remnants of an ancient forest provide ecological context for Early Miocenefossil apes Nature Comm 5 3236
Myres JL 1932 Africa Man 32 208Nakatsukasa M Kunimatsu Y Nakano Y Takano T Ishida H 2007 Vertebral
morphology of Nacholapithecus kerioi based on KNM-BG 35250 J Hum Evol52 347e369
Nengo IO Rae TC 1992 New hominoid fossils from the early Miocene site ofSonghor Kenya J Hum Evol 23 423e429
Peppe DJ McNulty KP Cote SM Harcourt-Smith WEH Dunsworth HM VanCouvering JA 2009 Stratigraphic interpretation of the Kulu Formation (EarlyMiocene Rusinga Island Kenya) and its implications for primate evolutionJ Hum Evol 56 447e461
Pickford M 1981 Preliminary Miocene mammalian biostratigraphy for westernKenya J Hum Evol 10 73e97
Pickford M 1986 Sexual dimorphism in Proconsul Hum Evol 1 111e148Pickford M Andrews PJ 1981 The Tinderet Miocene sequence in Kenya J Hum
Evol 10 11e33Pickford M Kunimatsu Y 2005 Catarrhines from the Middle Miocene (ca 145
Ma) of Kipsaraman Tugen Hills Kenya Anthropol Sci 113 189e224Pickford M Senut B Gommery D Musiime E 2009 Distinctiveness of Ugan-
dapithecus from Proconsul Estudios Geol 65 183e241Pilbeam DR 1969 Tertiary Pongidae of East Africa evolutionary relationships and
taxonomy Bulletin of the Peabody Museum of Natural History 31 pp 1e185Rae TC 1993 Phylogenetic analysis of proconsulid facial morphology PhD
Dissertation State University of New York at Stony BrookRae TC 1999 Mosaic evolution in the origin of the Hominoidea Folia Primatol 70
125e135Rafferty KL Walker A Ruff C Rose MD Andrews PJ 1995 Postcranial esti-
mates of body weights in Proconsul with a note on a distal tibia of P major fromNapak Uganda Am J Phys Anthropol 97 391e402
Rossie JB MacLatchy L 2013 Dentognathic remains of an Afropithecus individualfrom Kalodirr Kenya J Hum Evol 65 199e208
Senut B Pickford M Gommery D Kunimatsu Y 2000 Un nouveau genredrsquohominoiumlde du Miocene inferieur drsquoAfrique orientale Ugandapithecus major(Le Gros Clark and Leakey 1950) CR Acad Sci Paris 331 227e233
Simons EL Pilbeam DR 1965 Preliminary revision of the Dryopithecinae (Pon-gidae Anthropoidea) Folia Primatol 3 81e152
Smith TM Martin LB Leakey MG 2003 Enamel thickness microstructure anddevelopment in Afropithecus turkanensis J Hum Evol 44 283e306
Stevens NJ Seiffert ER OConnor PM Roberts EM Schmitz MD Krause CGorscak E Ngasala S Hieronymus TL Temu J 2013 Palaeontological evi-dence for an Oligocene divergence between Old World monkeys and apesNature 497 611e614
Suwa G Kono RT Katoh S Asfaw B Beyene Y 2007 A new species of great apefrom the late Miocene epoch in Ethiopia Nature 448 921e924
Szalay FS Delson E 1979 Evolutionary History of the Primates Academic PressNew York
Teaford MF Beard KC Leakey RE Walker A 1988 New hominoid facial skel-eton from the Early Miocene of Rusinga Island Kenya and its bearing on therelationship between Proconsul nyanzae and Proconsul africanus J Hum Evol17 461e477
Van Couvering JAH Miller J 1969 Miocene stratigraphy and age determinationsRusinga Island Kenya Nature 221 628e632
Walker A 1997 Proconsul function and phylogeny In Begun DR Ward CVRose MD (Eds) Function Phylogeny and Fossils Miocene Hominoid Evolu-tion and Adaptation Plenum Press New York pp 209e224
Walker A Pickford M 1983 New postcranial fossils of Proconsul africanus andProconsul nyanzae In Ciochon RL Corruccini RS (Eds) New Interpretationsof Ape and Human Ancestry Plenum Press New York pp 325e351
Walker A Teaford M 1988 The Kaswanga primate site an Early Miocene hom-inoid site on Rusinga Island Kenya J Hum Evol 17 539e544
Walker A Teaford M 1989 The hunt for Proconsul Sci Am 260 76e82Walker A Teaford MF Martin L Andrews P 1993 A new species of Proconsul
from the early Miocene of RusingaMfangano Islands Kenya J Hum Evol 2543e56
Ward CV Ruff CB Walker A Rose MD Teaford MF Nengo IO 1995 Func-tional morphology of Proconsul patellas from Rusinga Island Kenya with im-plications for other Miocene-Pliocene catarrhines J Hum Evol 29 1e19
Ward S Brown B Hill A Kelley J Downs W 1999 Equatorius a new hominoidgenus from the Middle Miocene of Kenya Science 285 1382e1386
Werdelin L Sanders WJ 2010 Cenozoic Mammals of Africa University of Cali-fornia Press Berkeley

KP McNulty et al Journal of Human Evolution 84 (2015) 42e6160
We found no support in this study for the genus ldquoUgandapi-thecusrdquo as constructed by Pickford et al (2009) In that paper theytransferred the vast majority of Proconsul fossils from the Tinderetand Napak localities into ldquoUgandapithecusrdquo leaving P africanusrepresented only by its holotype and a small number of isolatedteeth and aligning it with the material from Rusinga and Mfan-ganoWhen the P africanus hypodigm is reconstituted and properlyaffiliated with the other Tinderet species according to itsmorphology much of the evidence marshaled by Pickford et al(2009) for ldquoUgandapithecusrdquo actually supports the distinction be-tween the Tinderet and Kisingiri samples ie the generic distinc-tion of Ekembo In addition some of the traits enumerated byPickford and colleagues in support of ldquoUgandapithecusrdquo wererevealed in our analysis to be variable within one or more speciesand therefore of questionable taxonomic utility We do not rejectout of hand the original conception of ldquoUgandapithecusrdquo (Senutet al 2000) as either a monotypic genus distinguishing ldquoUrdquo ma-jor or perhaps including a closely allied middle Miocene ldquoUrdquogitongai as well (Pickford and Kunimatsu 2005) but we have fornow retained the early Miocene specimens of both in Proconsulpending further study However even if Ugandapithecus is a taxo-nomically valid genus comprising just the latter two species theirmorphology indicates that theywould still be united in a cladewithProconsul and they lack several more advanced features that partlydefine Ekembo Better understanding of the role that body size playsinmediating differences within Proconsulwill help to further clarifythe relationships among these species
Despite the existence of one well-represented and several lesscomplete skeletons of what was long assumed to be Proconsul thephylogenetic status of this genus has defied consensus This islargely due in our view to Proconsul having been an admixture oftwo distinct genera one more advanced in the direction of extantapes than the other As we demonstrate here the material fromRusinga and Mfangano traditionally referred to Proconsul andwhich includes all of the more complete skeletal material isdistinct from the Tinderet and Napak material and belongs in aseparate genus Ekembo By disentangling these distinct clades weanticipate being able to achieve better phylogenetic resolution forboth genera thereby improving our knowledge of early hominoidevolution and diversification
Acknowledgments
We thank many people and agencies that assisted with thedevelopment and completion of this project This research wasinitiated as a result of funding from the Wenner-Gren Foundationand the Johns Hopkins School of Medicine to DR Begun in1987e1989 and was renewed following research and travelgenerously funded by multiple Leakey Foundation grants to KMcNulty National Science Foundation grants to K McNulty(0852609 1241807) the McKnight Foundation the University ofMinnesota the Lloyd A Wilford Endowment the LeverhulmeFoundation and the Turkana Basin Institute J Kelley gratefullyacknowledges support from the Institute of Human Origins Ari-zona State University DR Begun also wishes to acknowledge theAlexander Humboldt Stiftung and NSERC for funding support Weare grateful to the National Museums of Kenya the UgandaMuseum the Natural History Museum and to Rose Mwanja EzraMusiime Tom Mukhuyu Samuel Muteti James Yatich and MaryMuungu for access to and assistance with specimens in their careMany people provided assistance comments criticisms insightsand suggestions on various aspects of this work and we especiallythank Holly Dunsworth Will Harcourt-Smith Tom Lehmann DanPeppe Nicole Garrett Kirsten Jenkins Rutger Jansma Susy CoteIsaiah Nengo Andrew Hill and Brenda Benefit K McNulty
acknowledges the evolutionary anthropology faculty and studentsat Durham University for their helpful comments on an earlyversion of this manuscript and for their gracious hospitality Weare likewise grateful to the anonymous reviewers associate editorand editor whose comments strengthened the paper This manu-script is publication 1 supporting Research on East AfricanCatarrhine and Hominoid Evolution (REACHE)
Appendix A Supplementary data
Supplementary data related to this article can be found at httpdxdoiorg101016jjhevol201503009
References
Andrews P 1974 New species of Dryopithecus from Kenya Nature 249 188e190Andrews P 1978 A revision of the Miocene Hominoidea of East Africa Bull Brit
Mus (Nat Hist) Geol 30 85e225Andrews P 1985 Family group systematics and evolution among catarrhine pri-
mates In Delson E (Ed) Ancestors The Hard Evidence Liss New Yorkpp 14e22
Andrews P Harrison T Martin L Pickford M 1981 Hominoid primates from anew Miocene locality named Meswa Bridge in Kenya J Hum Evol 10123e128
Andrews P Begun D Zylstra M 1997 Interrelationships between functionalmorphology and paleoenvironments in Miocene hominoids In Begun DWard C Rose M (Eds) Function Phylogeny and Fossils Miocene HominoidEvolution and Adaptations Plenum Press New York pp 29e58
Andrews P Molleson T 1979 The provenance of Sivapithecus africanus Bull BritMus (Nat Hist) Geol 32 19e23
Begun DR 2001 African and Eurasian Miocene hominoids and the origins of theHominidae In de Bonis L Koufos GD Andrews P (Eds) Hominoid Evolutionand Climatic Change in Europe Phylogeny of the Neogene Hominoid Primatesof Eurasia Cambridge University Press London pp 231e253
Begun DR 2007 Fossil record of Miocene hominoids In Henke W Tattersall I(Eds) Handbook of Paleoanthropology Springer Berlin pp 921e977
Begun DR Teaford MF Walker AC 1994 Comparative and functional anatomyof Proconsul phalanges from the Kaswanga primate site Rusinga Island KenyaJ Hum Evol 26 89e165
Bishop WW 1964 More fossil primates and other Miocene mammals from North-East Uganda Nature 203 1327e1331
Bishop WW Miller JA Fitch FJ 1969 New potassium-argon age determinationsrelevant to the Miocene fossil mammal sequence in East Africa Am J Sci 267669e699
Blankenship JA Mincer HH Anderson KM Woods MA Burton EL 2007Third molar development in the estimation of chronologic age in Americanblacks as compared with whites J Forensic Sci 52 428e433
Bosler W 1981 Species groupings of early Miocene dryopithecine teeth from EastAfrica J Hum Evol 10 151e158
Coleman MN 2008 What does geometric mean mean geometrically Assessingthe utility of geometric mean and other size variables in studies of skullallometry Am J Phys Anthropol 135 404e415
Cote S Malit N Nengo I 2013 Additional mandibles of Rangwapithecus gordonian early Miocene catarrhine from the Tinderet localities of Western KenyaJ Hum Evol 153 341e352
Deane AS 2009 Early Miocene catarrhine dietary behavior the influence of theRed Queen Effect on incisor shape and curvature J Hum Evol 56 275e285
Drake R Van Couvering JA Pickford M Curtis G Harris JA 1988 New chro-nology for the Early Miocene mammalian faunas of Kisingiri Western KenyaJ Geol Soc Lond 145 479e491
Evernden JF Savage DE Curtis GH James GT 1964 Potassium-argon datesand the Cenozoic mammalian chronology of North America Am J Sci 262145e198
Gebo DL MacLatchy L Kityo R Deino A Kingston J Pilbeam D 1997A hominoid genus from the Early Miocene of Uganda Science 276 401e404
Geoffroy Saint-Hilaire E 1812 Tableau des quadrumanes 1 Ord QuadrumanesAnnls Mus Hist Nat Paris 19 85e122
Gray JE 1825 Outline of an attempt at the disposition of the Mammalia into tribesand families with a list of the genera apparently appertaining to each tribeAnnls Philos 10 337e344
Harrison T 1982 Small-bodied apes from the Miocene of East Africa PhDDissertation University of London
Harrison T 1986 New fossil anthropoids from the middle Miocene of East Africaand their bearing on the origin of the Oreopithecidae Am J Phys Anthropol 71265e284
Harrison T 1987 The phylogenetic relationships of the early catarrhine primates areview of the current evidence J Hum Evol 16 41e80
Harrison T 2002 Late Oligocene to middle Miocene catarrhines from Afro-ArabiaIn Hartwig WC (Ed) The Primate Fossil Record Cambridge University PressCambridge pp 311e338
KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 61
Harrison T 2010 Dendropithecoidea Proconsuloidea and Hominoidea (CatarrhiniPrimates) In Werdelin L Sanders WJ (Eds) Cenozoic Mammals of AfricaUniversity of California Press Berkeley pp 429e469
Harrison T Andrews P 2009 The anatomy and systematic position of a newspecies of early Miocene proconsulid from Meswa Bridge Kenya J Hum Evol56 479e496
Hill A Nengo IO Rossie JB 2013 A Rangwapithecus gordoni mandible from theearly Miocene site of Songhor Kenya J Hum Evol 65 490e500
Hopwood AT 1933 Miocene primates from Kenya J Linn Soc Zool 38 437e464Ishida H Kunimatsu Y Takano T Nakano Y Nakatsukasa M 2004 Nacholapi-
thecus skeleton from the Middle Miocene of Kenya J Hum Evol 46 69e103Jolicouer P 1963 The multivariate generalization of the allometry equation Bio-
metrics 19 497e499Jungers WL Falsetti AB Wall CE 1995 Shape relative size and size-
adjustments in morphometrics Am J Phys Anthropol 38 (S2) 137e161Kavanagh KD Evans AR Jernvall J 2007 Predicting evolutionary patterns of
mammalian teeth from development Nature 449 427e433Kelley J 1986 Species recognition and sexual dimorphism in Proconsul and
Rangwapithecus J Hum Evol 15 461e495Kelley J 1995 Sex determination in Miocene catarrhine primates Am J Phys
Anthropol 96 390e417Kelley J 1993 Taxonomic implications of sexual dimorphism in Lufengpithecus In
Kimbel WH Martin LB (Eds) Species Species Concepts and Primate Evo-lution Springer New York pp 429e458
Kent PE 1942 The country round the Kavirondo Gulf of Victoria Nyanza Geogr J100 22e31
Kordos L Begun DR 2001 A new cranium of Dryopithecus from RudabanyaHungary J Hum Evol 15 689e700
Le Gros Clark WE Leakey LSB 1950 Diagnoses of East African Miocene Homi-noidea Q J Geol Soc Lond 105 260e263
Le Gros Clark WE Leakey LSB 1951 The Miocene Hominoidea of East Africa InFossil Mammals of Africa No 1 British Museum (Natural History) London
Leakey LSB 1943 A Miocene anthropoid mandible from Rusinga Kenya Nature152 319e320
Leakey LSB 1963 East African fossil Hominoidea and the classification within thissuper-family In Washburn SL (Ed) Classification and Human EvolutionAldine Chicago pp 32e49
Leakey LSB 1967 Notes on the mammalian faunas from the Miocene and Pleis-tocene of East Africa In Bishop WW Clark JD (Eds) Background to Evolu-tion in Africa University of Chicago Press Chicago pp 7e29
Leakey RE Leakey MG 1986 A new Miocene hominoid from Kenya Nature 324143e146
Leakey RE Leakey MG Walker AC 1988 Morphology of Afropithecus turka-nensis from Kenya Am J Phys Anthropol 76 289e307
Leakey MG Ungar PS Walker A 1995 A new genus of large primate from theLate Oligocene of Lothidok Turkana District Kenya J Hum Evol 28519e531
Linnaeus C 1758 Systema naturae per regna tria naturae secundum classesordines genera species cum characteribus differentris synonymis locis tenthed Laurentii Salvii Stockholm
MacInnes DG 1943 Notes on the East African Miocene primates J E Afr UgandaNat Hist Soc 17 141e181
MacLatchy L 2004 The oldest ape Evol Anthropol 13 90e103MacLatchy L Bossert WH 1996 An analysis of the articular surface distribution of
the femoral head and acetabulum in anthropoids with implications for hipfunction in Miocene hominoids J Hum Evol 31 425e453
MacLatchy L Rossie JB 2005 The Napak hominoid still Proconsul major InLieberman DE Smith RJ Kelley J (Eds) Interpreting the Past Essays onHuman Primate and Mammal Evolution in Honor of David Pilbeam Brill Ac-ademic Publishers Boston pp 15e28
Martin L 1981 New specimens of Proconsul from Koru Kenya J Hum Evol 10139e150
McCollum MS Peppe DJ McNulty KP Dunsworth HM Harcourt-Smith WEH Andrews AL 2013 Magnetostratigraphy of the early MioceneHiwegi Formation (Rusinga Island Lake Victoria Kenya) Geological Society ofAmerica Abstracts with Programs 45 p 12
McNulty KP 2010 Apes and tricksters the evolution and diversification ofhumans closest relatives Evol Edu Outreach 3 322e332
McNulty KP Harcourt-Smith WEH Dunsworth HM 2007 New primate fossilsfrom Rusinga Island Kenya Am J Phys Anthropol 132 (S44) 170
McNulty KP MacLatchy L Peppe DJ Nengo I Manthi FK Miller ERStevens NJ Cote S Lehmann T Kingston J 2014a A regional approach toEast African early Miocene paleobiology J Vert Paleontol 185 SVP Programand Abstracts Book 2014
McNulty KP MacLatchy L Rossie JB Peppe DJ Deino AL Mbua ENManthi FK Nengo IO Stevens NJ Cote S Lehmann T Gutierrez M 2014b
Research on East African catarrhine and hominoid evolution results from thefirst year Am J Phys Anthropol 153 (S58) 182
Michel LA Peppe DJ Lutz JA Driese SG Dunsworth HM Harcourt-Smith WEH Horner W Lehmann T Nightengale S McNulty KP 2014Remnants of an ancient forest provide ecological context for Early Miocenefossil apes Nature Comm 5 3236
Myres JL 1932 Africa Man 32 208Nakatsukasa M Kunimatsu Y Nakano Y Takano T Ishida H 2007 Vertebral
morphology of Nacholapithecus kerioi based on KNM-BG 35250 J Hum Evol52 347e369
Nengo IO Rae TC 1992 New hominoid fossils from the early Miocene site ofSonghor Kenya J Hum Evol 23 423e429
Peppe DJ McNulty KP Cote SM Harcourt-Smith WEH Dunsworth HM VanCouvering JA 2009 Stratigraphic interpretation of the Kulu Formation (EarlyMiocene Rusinga Island Kenya) and its implications for primate evolutionJ Hum Evol 56 447e461
Pickford M 1981 Preliminary Miocene mammalian biostratigraphy for westernKenya J Hum Evol 10 73e97
Pickford M 1986 Sexual dimorphism in Proconsul Hum Evol 1 111e148Pickford M Andrews PJ 1981 The Tinderet Miocene sequence in Kenya J Hum
Evol 10 11e33Pickford M Kunimatsu Y 2005 Catarrhines from the Middle Miocene (ca 145
Ma) of Kipsaraman Tugen Hills Kenya Anthropol Sci 113 189e224Pickford M Senut B Gommery D Musiime E 2009 Distinctiveness of Ugan-
dapithecus from Proconsul Estudios Geol 65 183e241Pilbeam DR 1969 Tertiary Pongidae of East Africa evolutionary relationships and
taxonomy Bulletin of the Peabody Museum of Natural History 31 pp 1e185Rae TC 1993 Phylogenetic analysis of proconsulid facial morphology PhD
Dissertation State University of New York at Stony BrookRae TC 1999 Mosaic evolution in the origin of the Hominoidea Folia Primatol 70
125e135Rafferty KL Walker A Ruff C Rose MD Andrews PJ 1995 Postcranial esti-
mates of body weights in Proconsul with a note on a distal tibia of P major fromNapak Uganda Am J Phys Anthropol 97 391e402
Rossie JB MacLatchy L 2013 Dentognathic remains of an Afropithecus individualfrom Kalodirr Kenya J Hum Evol 65 199e208
Senut B Pickford M Gommery D Kunimatsu Y 2000 Un nouveau genredrsquohominoiumlde du Miocene inferieur drsquoAfrique orientale Ugandapithecus major(Le Gros Clark and Leakey 1950) CR Acad Sci Paris 331 227e233
Simons EL Pilbeam DR 1965 Preliminary revision of the Dryopithecinae (Pon-gidae Anthropoidea) Folia Primatol 3 81e152
Smith TM Martin LB Leakey MG 2003 Enamel thickness microstructure anddevelopment in Afropithecus turkanensis J Hum Evol 44 283e306
Stevens NJ Seiffert ER OConnor PM Roberts EM Schmitz MD Krause CGorscak E Ngasala S Hieronymus TL Temu J 2013 Palaeontological evi-dence for an Oligocene divergence between Old World monkeys and apesNature 497 611e614
Suwa G Kono RT Katoh S Asfaw B Beyene Y 2007 A new species of great apefrom the late Miocene epoch in Ethiopia Nature 448 921e924
Szalay FS Delson E 1979 Evolutionary History of the Primates Academic PressNew York
Teaford MF Beard KC Leakey RE Walker A 1988 New hominoid facial skel-eton from the Early Miocene of Rusinga Island Kenya and its bearing on therelationship between Proconsul nyanzae and Proconsul africanus J Hum Evol17 461e477
Van Couvering JAH Miller J 1969 Miocene stratigraphy and age determinationsRusinga Island Kenya Nature 221 628e632
Walker A 1997 Proconsul function and phylogeny In Begun DR Ward CVRose MD (Eds) Function Phylogeny and Fossils Miocene Hominoid Evolu-tion and Adaptation Plenum Press New York pp 209e224
Walker A Pickford M 1983 New postcranial fossils of Proconsul africanus andProconsul nyanzae In Ciochon RL Corruccini RS (Eds) New Interpretationsof Ape and Human Ancestry Plenum Press New York pp 325e351
Walker A Teaford M 1988 The Kaswanga primate site an Early Miocene hom-inoid site on Rusinga Island Kenya J Hum Evol 17 539e544
Walker A Teaford M 1989 The hunt for Proconsul Sci Am 260 76e82Walker A Teaford MF Martin L Andrews P 1993 A new species of Proconsul
from the early Miocene of RusingaMfangano Islands Kenya J Hum Evol 2543e56
Ward CV Ruff CB Walker A Rose MD Teaford MF Nengo IO 1995 Func-tional morphology of Proconsul patellas from Rusinga Island Kenya with im-plications for other Miocene-Pliocene catarrhines J Hum Evol 29 1e19
Ward S Brown B Hill A Kelley J Downs W 1999 Equatorius a new hominoidgenus from the Middle Miocene of Kenya Science 285 1382e1386
Werdelin L Sanders WJ 2010 Cenozoic Mammals of Africa University of Cali-fornia Press Berkeley

KP McNulty et al Journal of Human Evolution 84 (2015) 42e61 61
Harrison T 2010 Dendropithecoidea Proconsuloidea and Hominoidea (CatarrhiniPrimates) In Werdelin L Sanders WJ (Eds) Cenozoic Mammals of AfricaUniversity of California Press Berkeley pp 429e469
Harrison T Andrews P 2009 The anatomy and systematic position of a newspecies of early Miocene proconsulid from Meswa Bridge Kenya J Hum Evol56 479e496
Hill A Nengo IO Rossie JB 2013 A Rangwapithecus gordoni mandible from theearly Miocene site of Songhor Kenya J Hum Evol 65 490e500
Hopwood AT 1933 Miocene primates from Kenya J Linn Soc Zool 38 437e464Ishida H Kunimatsu Y Takano T Nakano Y Nakatsukasa M 2004 Nacholapi-
thecus skeleton from the Middle Miocene of Kenya J Hum Evol 46 69e103Jolicouer P 1963 The multivariate generalization of the allometry equation Bio-
metrics 19 497e499Jungers WL Falsetti AB Wall CE 1995 Shape relative size and size-
adjustments in morphometrics Am J Phys Anthropol 38 (S2) 137e161Kavanagh KD Evans AR Jernvall J 2007 Predicting evolutionary patterns of
mammalian teeth from development Nature 449 427e433Kelley J 1986 Species recognition and sexual dimorphism in Proconsul and
Rangwapithecus J Hum Evol 15 461e495Kelley J 1995 Sex determination in Miocene catarrhine primates Am J Phys
Anthropol 96 390e417Kelley J 1993 Taxonomic implications of sexual dimorphism in Lufengpithecus In
Kimbel WH Martin LB (Eds) Species Species Concepts and Primate Evo-lution Springer New York pp 429e458
Kent PE 1942 The country round the Kavirondo Gulf of Victoria Nyanza Geogr J100 22e31
Kordos L Begun DR 2001 A new cranium of Dryopithecus from RudabanyaHungary J Hum Evol 15 689e700
Le Gros Clark WE Leakey LSB 1950 Diagnoses of East African Miocene Homi-noidea Q J Geol Soc Lond 105 260e263
Le Gros Clark WE Leakey LSB 1951 The Miocene Hominoidea of East Africa InFossil Mammals of Africa No 1 British Museum (Natural History) London
Leakey LSB 1943 A Miocene anthropoid mandible from Rusinga Kenya Nature152 319e320
Leakey LSB 1963 East African fossil Hominoidea and the classification within thissuper-family In Washburn SL (Ed) Classification and Human EvolutionAldine Chicago pp 32e49
Leakey LSB 1967 Notes on the mammalian faunas from the Miocene and Pleis-tocene of East Africa In Bishop WW Clark JD (Eds) Background to Evolu-tion in Africa University of Chicago Press Chicago pp 7e29
Leakey RE Leakey MG 1986 A new Miocene hominoid from Kenya Nature 324143e146
Leakey RE Leakey MG Walker AC 1988 Morphology of Afropithecus turka-nensis from Kenya Am J Phys Anthropol 76 289e307
Leakey MG Ungar PS Walker A 1995 A new genus of large primate from theLate Oligocene of Lothidok Turkana District Kenya J Hum Evol 28519e531
Linnaeus C 1758 Systema naturae per regna tria naturae secundum classesordines genera species cum characteribus differentris synonymis locis tenthed Laurentii Salvii Stockholm
MacInnes DG 1943 Notes on the East African Miocene primates J E Afr UgandaNat Hist Soc 17 141e181
MacLatchy L 2004 The oldest ape Evol Anthropol 13 90e103MacLatchy L Bossert WH 1996 An analysis of the articular surface distribution of
the femoral head and acetabulum in anthropoids with implications for hipfunction in Miocene hominoids J Hum Evol 31 425e453
MacLatchy L Rossie JB 2005 The Napak hominoid still Proconsul major InLieberman DE Smith RJ Kelley J (Eds) Interpreting the Past Essays onHuman Primate and Mammal Evolution in Honor of David Pilbeam Brill Ac-ademic Publishers Boston pp 15e28
Martin L 1981 New specimens of Proconsul from Koru Kenya J Hum Evol 10139e150
McCollum MS Peppe DJ McNulty KP Dunsworth HM Harcourt-Smith WEH Andrews AL 2013 Magnetostratigraphy of the early MioceneHiwegi Formation (Rusinga Island Lake Victoria Kenya) Geological Society ofAmerica Abstracts with Programs 45 p 12
McNulty KP 2010 Apes and tricksters the evolution and diversification ofhumans closest relatives Evol Edu Outreach 3 322e332
McNulty KP Harcourt-Smith WEH Dunsworth HM 2007 New primate fossilsfrom Rusinga Island Kenya Am J Phys Anthropol 132 (S44) 170
McNulty KP MacLatchy L Peppe DJ Nengo I Manthi FK Miller ERStevens NJ Cote S Lehmann T Kingston J 2014a A regional approach toEast African early Miocene paleobiology J Vert Paleontol 185 SVP Programand Abstracts Book 2014
McNulty KP MacLatchy L Rossie JB Peppe DJ Deino AL Mbua ENManthi FK Nengo IO Stevens NJ Cote S Lehmann T Gutierrez M 2014b
Research on East African catarrhine and hominoid evolution results from thefirst year Am J Phys Anthropol 153 (S58) 182
Michel LA Peppe DJ Lutz JA Driese SG Dunsworth HM Harcourt-Smith WEH Horner W Lehmann T Nightengale S McNulty KP 2014Remnants of an ancient forest provide ecological context for Early Miocenefossil apes Nature Comm 5 3236
Myres JL 1932 Africa Man 32 208Nakatsukasa M Kunimatsu Y Nakano Y Takano T Ishida H 2007 Vertebral
morphology of Nacholapithecus kerioi based on KNM-BG 35250 J Hum Evol52 347e369
Nengo IO Rae TC 1992 New hominoid fossils from the early Miocene site ofSonghor Kenya J Hum Evol 23 423e429
Peppe DJ McNulty KP Cote SM Harcourt-Smith WEH Dunsworth HM VanCouvering JA 2009 Stratigraphic interpretation of the Kulu Formation (EarlyMiocene Rusinga Island Kenya) and its implications for primate evolutionJ Hum Evol 56 447e461
Pickford M 1981 Preliminary Miocene mammalian biostratigraphy for westernKenya J Hum Evol 10 73e97
Pickford M 1986 Sexual dimorphism in Proconsul Hum Evol 1 111e148Pickford M Andrews PJ 1981 The Tinderet Miocene sequence in Kenya J Hum
Evol 10 11e33Pickford M Kunimatsu Y 2005 Catarrhines from the Middle Miocene (ca 145
Ma) of Kipsaraman Tugen Hills Kenya Anthropol Sci 113 189e224Pickford M Senut B Gommery D Musiime E 2009 Distinctiveness of Ugan-
dapithecus from Proconsul Estudios Geol 65 183e241Pilbeam DR 1969 Tertiary Pongidae of East Africa evolutionary relationships and
taxonomy Bulletin of the Peabody Museum of Natural History 31 pp 1e185Rae TC 1993 Phylogenetic analysis of proconsulid facial morphology PhD
Dissertation State University of New York at Stony BrookRae TC 1999 Mosaic evolution in the origin of the Hominoidea Folia Primatol 70
125e135Rafferty KL Walker A Ruff C Rose MD Andrews PJ 1995 Postcranial esti-
mates of body weights in Proconsul with a note on a distal tibia of P major fromNapak Uganda Am J Phys Anthropol 97 391e402
Rossie JB MacLatchy L 2013 Dentognathic remains of an Afropithecus individualfrom Kalodirr Kenya J Hum Evol 65 199e208
Senut B Pickford M Gommery D Kunimatsu Y 2000 Un nouveau genredrsquohominoiumlde du Miocene inferieur drsquoAfrique orientale Ugandapithecus major(Le Gros Clark and Leakey 1950) CR Acad Sci Paris 331 227e233
Simons EL Pilbeam DR 1965 Preliminary revision of the Dryopithecinae (Pon-gidae Anthropoidea) Folia Primatol 3 81e152
Smith TM Martin LB Leakey MG 2003 Enamel thickness microstructure anddevelopment in Afropithecus turkanensis J Hum Evol 44 283e306
Stevens NJ Seiffert ER OConnor PM Roberts EM Schmitz MD Krause CGorscak E Ngasala S Hieronymus TL Temu J 2013 Palaeontological evi-dence for an Oligocene divergence between Old World monkeys and apesNature 497 611e614
Suwa G Kono RT Katoh S Asfaw B Beyene Y 2007 A new species of great apefrom the late Miocene epoch in Ethiopia Nature 448 921e924
Szalay FS Delson E 1979 Evolutionary History of the Primates Academic PressNew York
Teaford MF Beard KC Leakey RE Walker A 1988 New hominoid facial skel-eton from the Early Miocene of Rusinga Island Kenya and its bearing on therelationship between Proconsul nyanzae and Proconsul africanus J Hum Evol17 461e477
Van Couvering JAH Miller J 1969 Miocene stratigraphy and age determinationsRusinga Island Kenya Nature 221 628e632
Walker A 1997 Proconsul function and phylogeny In Begun DR Ward CVRose MD (Eds) Function Phylogeny and Fossils Miocene Hominoid Evolu-tion and Adaptation Plenum Press New York pp 209e224
Walker A Pickford M 1983 New postcranial fossils of Proconsul africanus andProconsul nyanzae In Ciochon RL Corruccini RS (Eds) New Interpretationsof Ape and Human Ancestry Plenum Press New York pp 325e351
Walker A Teaford M 1988 The Kaswanga primate site an Early Miocene hom-inoid site on Rusinga Island Kenya J Hum Evol 17 539e544
Walker A Teaford M 1989 The hunt for Proconsul Sci Am 260 76e82Walker A Teaford MF Martin L Andrews P 1993 A new species of Proconsul
from the early Miocene of RusingaMfangano Islands Kenya J Hum Evol 2543e56
Ward CV Ruff CB Walker A Rose MD Teaford MF Nengo IO 1995 Func-tional morphology of Proconsul patellas from Rusinga Island Kenya with im-plications for other Miocene-Pliocene catarrhines J Hum Evol 29 1e19
Ward S Brown B Hill A Kelley J Downs W 1999 Equatorius a new hominoidgenus from the Middle Miocene of Kenya Science 285 1382e1386
Werdelin L Sanders WJ 2010 Cenozoic Mammals of Africa University of Cali-fornia Press Berkeley





























