Embed Size (px)
Citation preview

J O U R N A L O F P R O T E O M I C S 9 6 ( 2 0 1 4 ) 2 2 3 – 2 3 9
Ava i l ab l e on l i ne a t www.sc i enced i r ec t . com
ScienceDirectwww.e l sev i e r . com/ loca te / j p ro t
Activity of aaptamine and two derivatives,
demethyloxyaaptamine and isoaaptamine, incisplatin-resistant germ cell cancerSergey A. Dyshlovoya,b, Simone Venzc,d, Larisa K. Shubinab, Sergey N. Fedorovb,Reinhard Waltherc, Christine Jacobsena, Valentin A. Stonikb, Carsten Bokemeyera,Stefan Balabanova,e,1, Friedemann Honeckera,1,⁎aDepartment of Oncology, Haematology and Bone Marrow Transplantation with Section Pneumology, Hubertus Wald-Tumorzentrum,University Medical Center Hamburg-Eppendorf, Hamburg, GermanybLaboratory of Marine Natural Products Chemistry, G.B. Elyakov Pacific Institute of Bioorganic Chemistry, Far-East Branch, Russian Academyof Sciences, Vladivostok, Russian FederationcDepartment of Medical Biochemistry and Molecular Biology, University of Greifswald, Greifswald, GermanydInterfacultary Institute of Genetics and Functional Genomics, Department of Functional Genomics, University of Greifswald, Greifswald, GermanyeDivision of Hematology, University Hospital Zurich, Zurich, Switzerland
A R T I C L E I N F O
Abbreviations: 2-D WB, Mini-2D Westernfactor 5A-1; Hsp70, Heat shock protein 70 kD⁎ Corresponding author at: Tumor and Breast
2430043; fax: +41 71 2430044.E-mail address: friedemann.honecker@ze
1 These authors contributed equally to the
1874-3919/$ – see front matter © 2013 Elseviehttp://dx.doi.org/10.1016/j.jprot.2013.11.009
A B S T R A C T
Article history:Received 7 August 2013Accepted 12 November 2013
We analyzed the effects of all three marine alkaloids aaptamine, demethyloxyaaptamineand isoaaptamine in NT2-R, a cisplatin-resistant subline of the human embryonalcarcinoma cell line NT2. All aaptamines were found to be equally effective in both celllines, excluding cross-resistance between aaptamines and cisplatin in vitro. At theinhibitory concentration (IC50), aaptamine exerted an antiproliferative effect, whereasdemethyloxyaaptamine and isoaaptamine were strong inducers of apoptosis. Weanalyzed the changes in the proteome of NT2-R cells treated with these compounds. 16–22 proteins were found to be significantly altered, of which several were validated byWestern blotting and two-dimensional Western blotting analysis. Changes in theproteome pattern frequently resulted from post-transcriptional protein modifications,i.e. phosphorylation or hypusination in the case of eIF5A. Although the lists of alteredproteins were heterogeneous and compound-specific, gene ontology analyses identifiedrather similar profiles regarding the affected molecular functions. Ingenuity pathwayanalysis by IPA put the following factors in a central position of the hypothetical networks:myc and p53 for aaptamine; tumor necrosis factor (TNF) for demethyloxyaaptamine; andall three, myc, p53, and TNF for isoaaptamine. Our results represent an important steptowards a better understanding of the molecular basis underlying the observed bioactivityof these promising marine compounds.
Keywords:AaptamineMarine alkaloidsGerm cell cancerProteome analysisCisplatin resistance
blotting analysis; Cisplatin, Cis-diamminedichloroplatinum (II); eIF5A, Eukaryotic initiationa; MTT, 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide.Center ZeTuP St. Gallen, Rorschacher Str. 150, CH-9006 St., Gallen, Switzerland. Tel.: +41 71
tup.ch (F. Honecker).manuscript.
r B.V. All rights reserved.

224 J O U R N A L O F P R O T E O M I C S 9 6 ( 2 0 1 4 ) 2 2 3 – 2 3 9
Biological significanceWe characterized the mode of action of three aaptamines, marine natural compound withanti-tumor activity, using a functional proteomics approach and the cisplatin-resistantpluripotent human embryonal carcinoma cell line NT2-R. The manuscript is of particularscientific interest, as we could reveal the similarities and differences of the modes of action.Furthermore, wewere able to identify several new targets of these promising compounds.Wefound hypusination of eIF5A to be a prominent feature exclusively of aaptamine treatment, asthis was not observed upon treatment with demethyloxyaaptamine or isoaaptamine. Ourresults are a step towards unraveling the mode of action of these interesting compounds.
© 2013 Elsevier B.V. All rights reserved.
1. Introduction
Thirty years ago, Nakamura et al. reported the isolation ofaaptamine, a new benzo[de] [1,6]-naphthyridine alkaloid withα-adrenoreceptor blocking activity, from the marine spongeAaptos aaptos [1]. Later, other substances belonging to this classof heteroaromatic substances were isolated from marinesponges belonging to different families and orders. The mostcommonly occurring and therefore best characterized naturalaaptamine alkaloids are aaptamine (1), demethyloxyaaptamine(2), and isoaaptamine (3) [2] (Fig. 1A).
Aaptamines have previously been shown to possess antimi-crobial, antifungal, antiretroviral, cytotoxic, antifouling, enzymeinhibiting, antioxidant (for review, see [2]), as well as cancer-preventive activity [3]. At the same time, themolecular modes ofaction of aaptamine and its derivatives are largely unknown. Inrecent studies, it was shown that aaptamine intercalates intoDNAwith relativelyweak binding affinity [4], inhibits proteasomeactivity in a manner unrelated to cytotoxicity [5], and possessesantiproliferative properties (stipulated by p21 up-regulationand induction of a G2/M cell cycle arrest) at low [6–8], andpro-apoptotic properties at high concentrations [7,9]. Moleculartargets of aaptamine derivatives such as demethyloxyaaptamineand isoaaptamine are much less studied. However, forisoaaptamine, the ability to disturb cytoplasmic membranepermeability and thus induce potassium ion loss [10] as well asproteasome inhibitory activity [5] was recently shown. Fordemethyloxyaaptamine (2), to our knowledge, data onmolecularmechanisms of biological activity are completely elusive so far.
Building onour previous research onbioactivity of aaptaminealkaloids [3,7,9,11], this work presents a proteomic-basedscreening approach to identify molecular targets of aaptamineand the two natural derivatives in the cisplatin-resistant germcell cancer cell line NT2-R. The presented functional analysisexplores the similarities and differences in themode of action ofthese marine compounds and aims at unraveling relationshipsbetween structure and bioactivity.
2. Experimental section
2.1. Reagents and antibodies
Aaptamine (8,9-dimethoxy-1H-benzo[de][1,6]naphtyridine),demethyloxyaaptamine (9-methoxy-8-hydro-8-oxo-1H-benzo[de][1,6]naphtyridine), and isoaaptamine (1N-methyl-9-hydroxy-8-methyl-1H-benzo[de][1,6]naphtyridine) were isolated from the
marine spongeAaptos sp. asdescribedbefore [3]. Anisomycin andcisplatin (cis-diamminedichloroplatinum (II), 1 mg/ml in dH2O)were purchased fromNeoCorp (Weilheim, Germany). CoomassieBrilliant Blue G 250 was purchased from Carl Roth (Karlsruhe,Germany). Primary and secondary antibodies used are listed inthe Supplementary information.
2.2. Cell lines and culture conditions
NT2 cells (sometimes also referred to as NTERA2) [12] wereobtained from DSMZ (Braunschweig, Germany) The cisplatin-resistant subcells NT2-R have been generated as reported before[13,14]. Both NT2 and NT2-R cells were cultured in DMEMmedium supplemented with Glutamax™-I (Invitrogen, Paisley,UK) containing 10% fetal bovine serum (FBS) (Invitrogen) and 1%penicillin/streptomycin (Invitrogen). Cells were incubated at37 °C in a humidified atmosphere with 5% (v/v) CO2.
2.3. In vitro MTT-based drug sensitivity assay
The in vitro cytotoxic activity of aaptamine, demethyl-oxyaaptamine, and isoaaptamine was evaluated using theMTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bro-mide) assay, which was performed as previously described[7,15] with slight modifications. In brief, 6 × 103 cells/well wereseeded in 96-well plates and incubated overnight. The mediumwas replaced with medium containing the substances atindicated concentrations in a total volume of 0.1 ml/well, andcells were incubated for 24 h, 48 h, or 72 h. After addition of0.5 mg/ml MTT (Sigma, Taufkirchen, Germany), the cells werefurther incubated for 2 h. The medium was removed, and0.05 ml/well of DMSO (Sigma) was added. The plates wereagitated for 10 min and optical density was read with aphotometer (Victor2 Wallac 1420, PerkinElmer, Wellesley, MA,USA) at 570 nm. The background was read at 620 nm. Theexperiments were performed in triplicates, and the resultswere expressed as the drug concentrations that inhibit viablecells by 50% (inhibitory concentration, IC50), compared tocells treated with the equivalent volume of solvent (ethanol,0.5% [v/v]) alone.
2.4. In vitro trypan blue-based viability assay
The in vitro activity of aaptamine, demethyloxyaaptamine, andisoaaptamine on cell viability was furthermore evaluated withthe trypan blue exclusion assay using semi-automated cell countwith a Beckman Coulter Vi-CELL (Beckman Coulter, Krefeld,

225J O U R N A L O F P R O T E O M I C S 9 6 ( 2 0 1 4 ) 2 2 3 – 2 3 9
Germany) as described before [16]. In brief, 8 × 104 cells/well wereseeded in 12-well plates and incubated overnight. The mediumwas replaced with fresh medium containing the substances atindicated concentrations in a total volume of 2 ml/well, and cellswere incubated for 48 h. Drug-containing medium was removedand cells were washed with 0.5 ml of PBS and trypsinized with0.5 ml of trypsin-EDTA solution (Invitrogen). The number ofviable trypan blue excluding cells in both the medium andafter trypsination was evaluated with the Beckman CoulterVi-CELL counting trypan blue positive and negative cells. Assayswere performed in triplicates, and Student's t-test was used todetermine statistical significance.
2.5. Protein preparation
Preparation of protein extracts for two-dimensional electro-phoresis (2-DE) and Western blotting was performed asdescribed previously [17,18]. In brief, 2 × 106 cells/well wereseeded in culture flasks, incubated overnight, and treated withthe substances for 48 h. Cells were harvested with mediumflow, pelleted by centrifugation for 5 min at 453 ×g, andwashed3 times with PBS, followed by pelleting using the sameconditions. For 2-DE, cells were lysed with 450 μl of 2-DE lysisbuffer (9 M urea, 4% [w/v] CHAPS, 1% [v/v] pharmalyte, 1% [w/v]DTT, 0.01% [w/v] bromophenol blue) at room temperature for20 min. For Western blotting, cells were lysed with 100 μl ofWestern blotting lysis buffer (0.88% [w/v] NaCl, 50 mMTris–HCl(pH 7.6), 1% [v/v] NP-40, 0.25% [w/v] NaClO2, 1 mM PMSF, 1 mMNa3VO4) on ice for 20 min. Lysates were frozen overnight at−20 °C and centrifuged at 11,170 ×g for 10 min. Protein concen-tration in the supernatant was determined by Bradford assayusing BSA standards [19].
2.6. Western blotting
Western blotting was performed as previously described[20]. In brief, protein extracts were diluted with Westernblotting lysis buffer and loading dye up to a total proteinconcentration of 1–1.5 μg/μl, boiled for 5 min at 99 °C, andsubjected to electrophoresis in 12.5–15% SDS-polyacrylamidegels at 120 V. Proteins were transferred from gel to 0.2 μmpore PVDFmembrane (Millipore, Bedford, MA, USA) at 20 V for1 h. The membrane was blocked with 5% [w/v] non-fat drymilk in 0.05% [v/v] Tween-20/TBS and incubated with theprimary antibody according to the manufacturer's protocol(for antibodies used, see Supplementary information). Afterwashing, the membranes were incubated with an appropriatesecondary antibody for 1 h at RT. The signals were detectedusing the ECL chemiluminescence system (Thermo Scientific,Rockford, IL, USA) according to the manufacturer's protocol.
2.7. Two-dimensional electrophoresis (2D-PAGE)
2D-PAGE was performed as previously described [7,17]. In brief,aliquots of 450 μl solution containing 1 mg of protein extractwere loaded on a linear gradient Immobiline Dry Strip (IPG StrippH 4–7, 24 cm, Amersham Biosciences, Uppsala, Sweden),followed by rehydration of strips overnight and isoelectricfocusing (IEF) using the Protean IEF cell (Bio-Rad, Hercules, CA,USA). The first dimensionwas performed at 10,000 V for 80 kVh.
After IEF, IPG strips were equilibrated with 1% [w/v] DTT in ureabuffer (6 M urea, 4% [w/v] SDS and 50 mM of 1.5 M Tris–HCl(pH 8.8)) for 15 min at RT and then alkylated with 4.8% [w/v]iodoacetamide in urea buffer for 15 min at RT. The equilibratedstrips were directly loaded onto 15% SDS-polyacrylamide gels(27 cm × 21 cm × 1.5 mm) and overlaid with 0.6% [w/v] agarosein dH2O. Second dimension electrophoresis was conducted inSDS-running buffer (1.44% [w/v] glycine, 0.3% [w/v] Tris base,0.1% [w/v] SDS at 20 °C) for 14 h at 20 °C. Gels were fixed andstained with colloidal Coomassie Blue (400 ml/gel, containing0.13% [w/v] Coomassie Brilliant Blue G 250, 3.6% [v/v] H2SO4,1.44 M NaOH, 20.3% [w/v] CCl3COOH) overnight and destainedfor 48 h with dH2O. All experiments were performed intriplicates.
2.8. 2D gel image analysis
The 2D gels were scanned using the GS-800 CalibratedDensitometer (Bio-Rad). Spot matching, normalization of thedigital images (based on total optical density), and gel imageanalysis was performed using Delta 2D 4.0 software (Decodon,Greifswald, Germany). Quantitative analysis was performedusing fold change value and Student's t-test comparingtreated and non-treated cells. Assumed difference in spotintensity of relevant spots was ≥2 times, assumed confidencelevel for p-values was ≤0.05.
2.9. Protein identification by mass spectrometry
Protein spots of interest were excised from the gelmanually andtransferred to 96-well-plates. Digestion with trypsin and subse-quent spotting of peptide solutions onto theMALDI-targets wereperformed automatically in the Ettan Spot Handling Worksta-tion (Amersham-Biosciences) using a modified standard proto-col. In-gel protein digestion was performed as previouslydescribed [21]. The MALDI-MS measurement of spotted peptidesolutions was carried out on a 4800 MALDI-ToF/ToF™ Analyzer(Applied Biosystems, Foster City, CA, USA). The spectra wererecorded in reflector mode in a mass range from 800 to 4000 Dawith a focus mass of 2000 Da. The instrument was calibratedwith the autolytical fragment of trypsin, an internal calibrationwas automatically performed using this peak for one-point-calibration. Additionally, MALDI-MS/MS analysis was performedfor the five strongest peaks of theMS-spectrumafter subtractionof peaks corresponding to background or trypsin fragments.After calibration, a combined database search of MS and MS/MSmeasurements was performed using the GPS Explorer software(Ver. 3.6, Applied Biosystems, Foster City, CA, USA). Peak listswere compared with the SwissProt database v56.1 humantaxonomy. Peptide mixtures that yielded at least twice aMOWSE score of 56 (p < 0.05) for database results were regardedas positive identifications. The default significance thresholdwas p < 0.05.
2.10. 2D Western blotting analysis (2D-WB)
2D-WB analysis was used to confirm the results of 2D-PAGE.Protein samples were prepared as described for 2D-PAGE.Aliquots of 125 μl containing 30 μg of protein were loaded on alinear gradient Immobiline Dry Strip (IPG Strip pH 4–7, 7 cm

226 J O U R N A L O F P R O T E O M I C S 9 6 ( 2 0 1 4 ) 2 2 3 – 2 3 9
Amersham Biosciences), followed by rehydration of stripsovernight and isoelectric focusing (IEF) using the Protean IEFcell. The first dimension was carried out at 5000 V for 5 kVh.Consecutive equilibration and alkylation of the stripswere carried out as described for the 2D-PAGE experiment.The equilibrated strips were directly loaded onto 15% SDS-polyacrylamide gels, overlayedwith 0.6% [w/v] agarose in dH2O,and run 3 h at 65 V. Consecutive transfer, blocking, incubationwith antibodies, and detection steps were carried out asdescribed for the Western blotting experiments; see above.
2.11. Annexin-V-FLUOS/PI double staining
To examine the induction of apoptosis by aaptamine, demethyl-oxyaaptamine, and isoaaptamine, FACS-based analysis withAnnexin-V-FLUOS (Roche, Mannheim, Germany) and propidiumiodide (PI) (Sigma) double staining was performed as previ-ously described [22]. NT2-R cells were pre-incubated over-night in 6-well plates (2 × 105 cells/well). The medium waschangedwithmedium containing different concentrations ofthe substances. After 48 h of treatment, cells were harvestedwith trypsin-EDTA solution, washed with PBS, and incubatedwith 0.1 ml of Annexin-V-FLUOS and PI containing labelingbuffer for 30 min in the dark at RT. Cells were analyzed using
Fig. 1 – Effect of aaptamine, demethyloxyaaptamine, and isoaaptwith aaptamine, demethyloxyaaptamine, and isoaaptamine f. (Aisoaaptamine (3). (B) IC50 values of the substances 1, 2, and 3, (trypcells were detected; n.s.: not significant. (C) Flow cytometry analversus PI (double staining, upper row), and PI staining of DNA (lolower quadrants (upper row) or as a sub-G1 peak in the cell cycle(1 μM for 48 h) represent the positive control (D) Western blotting3, analyzing cleavage of PARP and caspase-3 as hallmarks of aporepresent the positive control.
the FACS Calibur (BD Bioscience, Bedford, MA, USA). The resultswere analyzed using the BD Bioscience Cell Quest Pro software.
2.12. Cell cycle analysis
The amount of dead (apoptotic) cells was measured byFACS-based cell cycle analysis, where apoptotic cells wereidentified as a sub-G1 peak [23]. NT2-R cells were treated asdescribed above (see Annexin-V-FLUOS/PI double staining), theneach samplewaswashedwith ice-cold PBSand fixedwith 1 ml of100% ethanol at −20 °C overnight. After addition of 0.5 ml of PBSand pelleting (5 min at 220 ×g), cells were resuspended in 0.2 mlof buffer containing RNase (0.2 mg/ml, Carl Roth) and PI (0.02 mg/ml), and incubated for 30 min in the dark on ice. Then 0.2 ml ofPBS was added, and cells were analyzed using the FACS Calibur.The results were quantitatively analyzed using the BD BioscienceCell Quest Pro software.
2.13. IPA system analysis
Anetwork analysiswas performed using the Ingenuity PathwaysAnalysis (IPA) (Ingenuity Systems, www.ingenuity.com) algo-rithm. The lists of differentially expressed proteins were enteredinto the IPA software to explore relevant biological networks
amine on cell proliferation and viability in NT2-R cells treated) Structure of aaptamine (1), demethyloxyaaptamine (2), andan blue assay). No difference in the values for NT2 and NT2-Rysis of NT2-R cells treated with 1, 2, or 3: Annexin-V-FLUOSwer row). Apoptotic cells appear in the right upper and rightanalysis (lower row). NT2-R cells treated with anisomycinanalysis of protein extracts of NT2-R cells treated with 1, 2, orptosis. NT2-R cells treated with cisplatin (5 μM for 48 h)

227J O U R N A L O F P R O T E O M I C S 9 6 ( 2 0 1 4 ) 2 2 3 – 2 3 9
and to assess interactions with other proteins. A hypotheticalglobal protein interaction network was constructed, showing themost relevant direct and indirect connections of proteins foundto be regulated under aaptamine, demethyloxyaaptamine, orisoaaptamine treatment.
2.14. PANTHER Classification System analysis
Gene ontology classification with respect to molecular func-tions and biological processes was performed with PANTHER7.2 software (Protein ANalysis THrough Evolutionary Relation-ships) Classification System analysis (Paul Thomas, http://www.pantherdb.org) [24].
3. Results
3.1. Effects of aaptamine, demethyloxyaaptamine, andisoaaptamine on NT2-R cell viability and proliferation
The cisplatin-resistant cell line NT2-R, generated in our labora-tory, has been shown to have an approximately 10-fold higherresistance to cisplatin when compared to the parental cisplatin-sensitive cell line NT2 [13]. This cell line was therefore used as amodel system to investigate the ability of aaptamine and itsderivatives to overcome cisplatin-resistance, a relevant clinicalproblem in the treatment of germ cell cancers. First, the trypanblue-based semi-automated cell counting assay was applied toevaluate the effect of aaptamines 1, 2, and 3 on cell proliferationand cell viability of bothNT2 andNT2-R cells. As shown in Fig. 1B,all three substances possess equal levels of activity in thecisplatin-sensitive and -resistant cell line after 48 h of treatment.IC50 values determined by the MTT assay after 48 h of treatmentwere slightly different from those determined by the trypan blueassay, showing lower values than the trypan blue method,especially for aaptamines 2 and 3 (Table 1). This observationhas been reported before for other drugs [25]. Nevertheless, thevalues obtained confirmed that there is no difference betweenactivities of the compounds in NT2 and NT2-R cells for any ofthree substances. Thisproves that theagents are equally effectivein cisplatin-sensitive and -resistant germ cell tumor cells andsuggests that the compounds could be promising agents in thetherapy of cisplatin-resistant tumors (Table 1). For furtherexperiments, we chose the cisplatin-resistant NT2-R cell line inorder to investigate themode of action of these alkaloids inmoredetail.
3.2. Effects of aaptamine, demethyloxyaaptamine, andisoaaptamine on induction of apoptosis
To understand whether the observed reduction of viable cellcount at the IC50 was due to inhibition of cell proliferation orresulted from induction of cell death, we performed flowcytometry analysis and Western blotting (Fig. 1C, D). Flowcytometry (FACS) analysis using Annexin-V-FLUOS/PI doublestaining or PI staining of DNA revealed a small amount ofapoptotic cells after treatment of NT2-R cells with 50 μM (IC50,determined by trypan blue method) of aaptamine after 48 h(Fig. 1C). In line with this finding, there was a lack of PARP orcaspase-3 cleavage in aaptamine-treated NT2-R cells as
determined by Western blotting (Fig. 1D). A similar effect wasdescribed by us before for cisplatin-sensitive NT2 cells undertreatment with aaptamine at the same concentration [7].Interestingly, the two derivatives demethyloxyaaptamine (2)and isoaaptamine (3) revealeda significant inductionof apoptosisin NT2-R cells at the respective IC50 (Fig. 1C, D). Anisomycinand cisplatin were used as positive controls for induction ofapoptosis.
3.3. Identification of differentially expressed proteins followingaaptamine, demethyloxyaaptamine, or isoaaptamine treatmentby 2D-PAGE
Based on the result of the trypan blue assay, we applied atreatment regimen with aaptamine (1), demethyloxyaaptamine(2), or isoaaptamine (3) at concentrations of 50 μM, 4.5 μM, or12.5 μM, respectively, for 48 h as the setting for a screen of theglobal proteomeby 2D-PAGE. Protein expressionpattern ofNT2-Rcells treated with substance 1, 2 or 3, compared to controlcells treated with the solvent alone, was analyzed by 2D-PAGE(Figs. 2–4). Digitalized gel images were analyzed with Delta 2Dsoftware to identify differentially expressed spots.
3.3.1. AaptamineFor NT2-R cells treated with aaptamine (1), 1086 spots on eachgel were analyzed and 22 of them were differently regulatedwith a fold change ≥2 (p < 0.05). When comparing aaptaminetreated and untreated cells, 15 spots showed down-regulation(Fig. 2A), and 7 spots up-regulation (Fig. 2B). Representativeexamples of regulated spots are depicted in Fig. 2C. All 22spots were excised from preparative Coomassie stained 2Dgels and were subjected to tandem mass spectrometry(Fig. 2A–C, Table 2).
3.3.2. DemethyloxyaaptamineFor NT2-R cells treated with demethyloxyaaptamine (2), 966spots on each gel were analyzed and 16 of themwere differentlyregulated with a fold change ≥2 (p < 0.05). When comparingdemethyloxyaaptamine treated and untreated cells, 10 spotsshowed down-regulation (Fig. 3A), and 6 spots up-regulation(Fig. 3B). Representative examples of regulated spots are depictedin Fig. 3C. All 16 spots were excised from preparative Coomassiestained2Dgels andwere subjected to tandemmass spectrometry(Fig. 3A–C, Table 3).
3.3.3. IsoaaptamineFor NT2-R cells treatedwith isoaaptamine (3), 1010 spots on eachgel were analyzed and 17 of themwere differently regulatedwitha fold change ≥2 (p < 0.05). When comparing isoaaptaminetreated and untreated cells, 4 spots showed down-regulation(Fig. 4A), and 13 spots up-regulation (Fig. 4B). Representativeexamples of regulated spots are depicted in Fig. 4C. All 17 spotswere excised from preparative Coomassie stained 2D gels andwere subjected to tandemmass spectrometry (Fig. 4A–C, Table 4).
3.4. Protein network analysis and gene ontology(GO) classification
Network analysis was computed using the Ingenuity Path-ways Analysis tool (IPA) and is presented in Fig. 5A. This

Table 1 – IC50 determined with MTT or trypan blue assay.
Substance IC50, μM
NT2 cells NT2-R cells
24 h 48 h 72 h 24 h 48 h 72 h
MTT assay 1 100.5 ± 10.3 54.5 ± 5.3 31.9 ± 7.6 106.2 ± 8.8 53.7 ± 4.5 18.6 ± 7.12 4.4 ± 1.5 1.7 ± 0.3 1.3 ± 0.3 4.5 ± 1.6 1.9 ± 0.4 1.1 ± 0.23 13.2 ± 2.4 6.3 ± 2.1 6.2 ± 2.4 12.5 ± 3.6 3.9 ± 0.9 4.1 ± 1.1
Trypan blue assay 1 – 54.3 ± 8.5 – – 50.0 ± 6.1 –2 – 4.9 ± 0.7 – – 4.5 ± 0.5 –3 – 11.8 ± 2.2 – – 12.5 ± 2.5 –
228 J O U R N A L O F P R O T E O M I C S 9 6 ( 2 0 1 4 ) 2 2 3 – 2 3 9
pathway analysis of proteins regulated under aaptamine,demethyloxyaaptamine, or isoaaptamine treatment in NT2-Rcells placed the following factors in a central position of thenetwork: proto-oncoprotein myc and tumor suppressorprotein p53 for aaptamine; tumor necrosis factor (TNF)for demethyloxyaaptamine; and myc, p53 and TNF forisoaaptamine. This suggests that these factors and pathwaysare potential targets or effectors of the observed bioactivity ofaaptamine and its derivatives (Fig. 5A). Finally, PANTHERClassification System analysis revealed that most discoveredproteins regulated under treatment of each substance areinvolved in metabolic and cellular processes. Regarding themolecular functions of regulated proteins, the majority isinvolved in catalytic and binding activities (Tables 2–4 andFig. 5B). In order to predict the possible side effects of usingaaptamine compounds as drugs we applied IPA systemtoxicology analysis. The major predicted target organs werekidney, liver and heart for all three compounds, indicating arather similar pattern of possible side effects (Fig. 6). Thisfurther supports the hypothesis that all three compoundsaffect cellular integrity through an overlapping set of cellularpathways.
3.5. Validation of proteomics data byWestern blotting and2D-WB
To validate the results of protein alterations identified by2D-PAGE, Western blotting and 2D-Western blotting analyseswere performed. In total, we analyzed six proteins, which wereselected from the lists shown in Tables 2–4. For aaptamine weselected eIF5A, cofilin-1 and vimentin for further validation.Interestingly, conventionalWestern blotting for all three proteinsrevealed no change in the overall protein expression of thesethree proteins, indicating that the observed alterations occurredas a result of post-translational protein modification rather thanchanges of the total protein amount. In order to assess the effectsof aaptamine on post-translational modification, we performed2D-Western blotting analyses to visualize drug treatment in-duced changes in theprotein isoformshavingdifferent isoelectricpoint (IP). For eIF5A [26] and cofilin-1 [27,28] regulation bypost-translational modification has been described before. Asdepicted in Fig. 7A, aaptamine treatment clearly affected the pIof eIF5A and colilin-1, indicated by a pH shift of a substantialpart of the proteins in the 2D-Western blotting. Interestingly,identical changes have been previously observed by us inaaptamine-treated cisplatin-sensitive NT2 cells [7]. Furthermore,alterations of vimentin isoforms, most probably caused by
post-translational modification (e.g. phosphorylation) ratherthan changes in the expression level, were detected in NT2-Rcells upon aaptamine treatment (Fig. 7A, regulated spots aremarked with arrows). Similar alterations in vimentin phosphor-ylation have been observed duringmitosis, and are thought to beassociated with vimentin filament assembly and reorganization[29,30].
In demethyloxyaaptamine-treated NT2-R cells, an up-regulation of the total protein amount of chaperon Hsp70 wasobserved (Fig. 7B). Examination of α-enolase alterations revealedseveral protein spots with different Mw and pI showingsimultaneous regulation (Fig. 7B, regulated spots are markedwith arrows). At the same time, conventional Western blottingdid not reveal any considerable change in the total amount ofα-enolase.
Treatment of NT2-R cells with isoaaptamine resulted inthe alteration of nucleolin isoforms. Up-regulation of anisoform at Mw = 100 kDa was observed both in conventionalWestern blotting as well as in 2D-Western blotting (Fig. 7C).We hypothesize that this could be the result of a partialdissociation of the basic complex of nucleolin from histone H1[31–33], which is detected by 2D-Western blotting as a majornucleolin spot with Mw = 110 kDa and pI ≥ 7.0.
Interestingly, the previously described increase in eIF5Ahypusination (and thereby activation) in aaptamine-treated NT2cells [7] was also detectable in aaptamine-treated NT2-R cells.However, neither demethyloxyaaptamine-, nor isoaaptamine-treated NT2-R cells showed a significant increase in eIF5Ahypusination (Fig. 7A–C).
4. Discussion
4.1. Aaptamine alkaloids inhibit cell proliferation andinduce apoptosis
Aaptamine alkaloids are marine natural compounds with awide range of biological activity and a so far largely undefinedmechanism of action. Herein, we describe and comparethe effects of aaptamine (1), demethyloxyaaptamine (2), andisoaaptamine (3) on the embryonal carcinoma cell linesNT-2 andNT2-R. MTT- and trypan blue-based methods showed com-pounds 1, 2, and 3 to be equally effective both in NT2 and thecisplatin-resistant subline NT2-R. We therefore conclude thatalthough both compounds, aaptamine and cisplatin, act via DNAintercalation, there is no cross-resistance between these agentsin vitro. This could make aaptamine alkaloids interesting agents
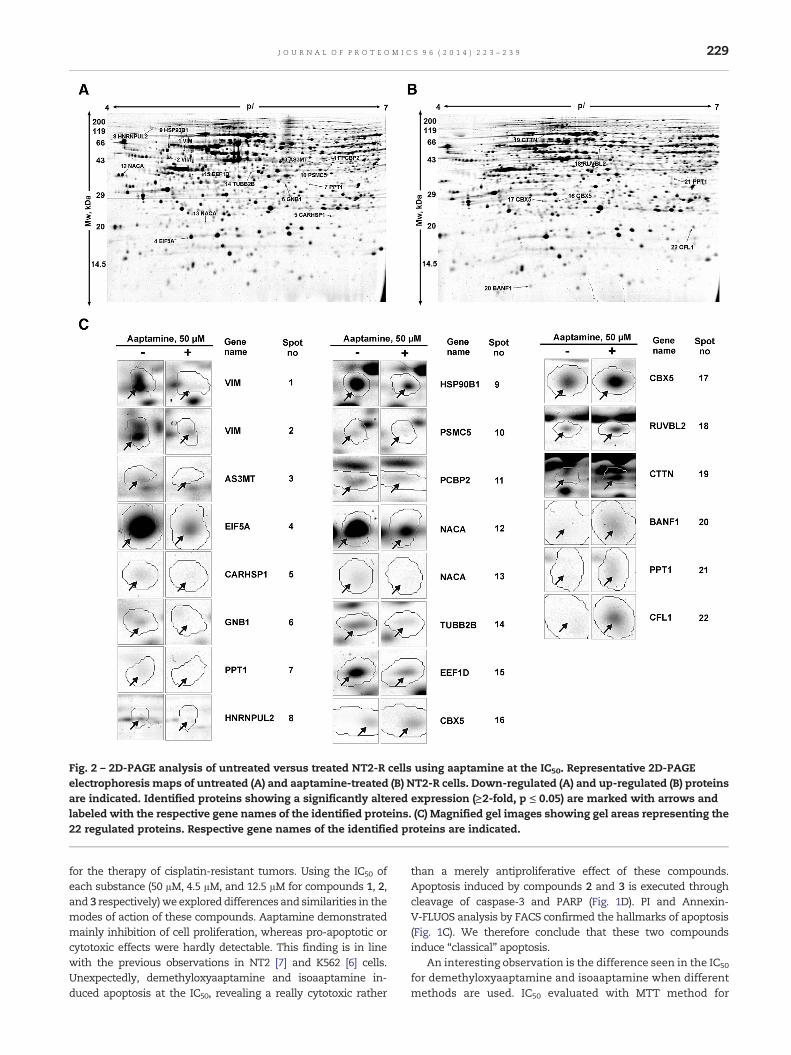
Fig. 2 – 2D-PAGE analysis of untreated versus treated NT2-R cells using aaptamine at the IC50. Representative 2D-PAGEelectrophoresis maps of untreated (A) and aaptamine-treated (B) NT2-R cells. Down-regulated (A) and up-regulated (B) proteinsare indicated. Identified proteins showing a significantly altered expression (≥2-fold, p ≤ 0.05) are marked with arrows andlabeled with the respective gene names of the identified proteins. (C) Magnified gel images showing gel areas representing the22 regulated proteins. Respective gene names of the identified proteins are indicated.
229J O U R N A L O F P R O T E O M I C S 9 6 ( 2 0 1 4 ) 2 2 3 – 2 3 9
for the therapy of cisplatin-resistant tumors. Using the IC50 ofeach substance (50 μM, 4.5 μM, and 12.5 μM for compounds 1, 2,and 3 respectively)we exploreddifferences and similarities in themodes of action of these compounds. Aaptamine demonstratedmainly inhibition of cell proliferation, whereas pro-apoptotic orcytotoxic effects were hardly detectable. This finding is in linewith the previous observations in NT2 [7] and K562 [6] cells.Unexpectedly, demethyloxyaaptamine and isoaaptamine in-duced apoptosis at the IC50, revealing a really cytotoxic rather
than a merely antiproliferative effect of these compounds.Apoptosis induced by compounds 2 and 3 is executed throughcleavage of caspase-3 and PARP (Fig. 1D). PI and Annexin-V-FLUOS analysis by FACS confirmed the hallmarks of apoptosis(Fig. 1C). We therefore conclude that these two compoundsinduce “classical” apoptosis.
An interesting observation is the difference seen in the IC50
for demethyloxyaaptamine and isoaaptamine when differentmethods are used. IC50 evaluated with MTT method for

Fig. 3 – 2D-PAGE analysis of untreated versus treated NT2-R cells using demethyloxyaaptamine at the IC50. Representative2D-PAGE electrophoresis maps of untreated (A) and demethyloxyaaptamine-treated (B) NT2-R cells. Down-regulated (A) andup-regulated (B) proteins are indicated. Identified proteins showing a significantly altered expression (≥2-fold, p ≤ 0.05) aremarked with arrows and labeled with the respective gene names of the identified proteins. (C) Magnified gel images showinggel areas representing the 16 regulated proteins. Respective gene names of the identified proteins are indicated.
230 J O U R N A L O F P R O T E O M I C S 9 6 ( 2 0 1 4 ) 2 2 3 – 2 3 9
compounds 2 and 3 (but not for compound 1) was 2-fold lowerthan those determined with the trypan blue-based method(Table 1). The MTT method can be used for the evaluation ofviability, proliferation or activation of metabolically activecells [15,34]. The ability of alive cells to cleave MTT intoformazan has been associated with the activity of cellularmitochondrial succinate dehydrogenase or mitochondrialdehydrogenase [35], although there are some doubts aboutthe exclusive role of mitochondria in this process [36]. Thetrypan blue based method is suitable for the determination ofthe number of viable cells. This method is based on the abilityof viable cells with an intact membrane to exclude trypan bluedye [37]. Based on the results of this experiment as wellas Western blotting and FACS assays we speculate that
compounds 2 and 3 exert an inhibitory effect on the cellularmetabolism at concentrations ~2-fold lower than thosecausing cell membrane permeability and cell death. Incontrast, compound 1 seems to act via inhibition of cellgrowth at concentrations when cell membrane permeabilityand metabolism still remain intact.
4.2. Protein network analysis and gene ontology classification
Proteome analysis revealed that 16–21 proteins were differentlyregulated under treatment with each substance at the IC50.Hypothetical pathway analysis performed by IPA system put thefollowing factors in a central position of the network: myc andp53 for aaptamine; TNF for demethyloxyaaptamine; and all three

Fig. 4 – 2D-PAGE analysis of untreated versus treated NT2-R cells using isoaaptamine at the IC50. Representative 2D-PAGEelectrophoresis maps of untreated (A) and isoaaptamine-treated (B) NT2-R cells. Down-regulated (A) and up-regulated(B) proteins are indicated. Identified proteins showing significantly altered expression (≥2-fold, p ≤ 0.05) are marked witharrows and labeled with the respective gene names of the identified proteins. (C) Magnified gel images showing gel areasrepresenting the 17 regulated proteins. Respective gene names of the identified proteins are indicated.
231J O U R N A L O F P R O T E O M I C S 9 6 ( 2 0 1 4 ) 2 2 3 – 2 3 9
proteins for isoaaptamine, thus identifying potential targets oreffectors of the substances (Fig. 5A). myc proteins play animportant role as a transcription factor regulating the expressionof a large number of genes. Themyc gene is frequentlymutated invarious types of human tumors, and is involved in key cellularprocesses such as cell cycle progression and induction ofapoptosis (for review, see [38]). p53 plays a central role in manycellular processes such as cell cycle arrest, senescence, anddifferentiation. One of the most prominent roles of p53 is itspro-apoptotic function in stressed or abnormal cells, therebyprotecting organisms from malignant transformation (reviewedin [39]). TNF proteins are cytokines involved in processes ofmaintenance and homeostasis of the immune system, inflam-mation, and host defense.While TNF possesses well determinedanti-cancer properties, its role in cancer biology ismore complex,
and there are several reports about TNF-promoted tumordevelopment and progression (for review, see [40]). More detailedanalysis of the activity of this transcription factor will benecessary to elucidate the putative role of these proteins in themode of action of the compounds under investigation. Surpris-ingly, despite the structural similarity of the compounds tested,there was not a single protein that showed universal regulationupon treatment with all three substances, and there were onlytwo proteins simultaneously regulated upon treatment withdemethyloxyaaptamine and isoaaptamine, namely Rab GDPdissociation inhibitor beta and polyglutamine-binding protein 1(corresponding gene names: GDI2 and PQBP1) (Tables 2, 3).However, despite the high degree of differences in proteomealterations, the distribution profiles of molecular functions andbiological processes of the altered proteins as determined by

Table 2 – List of proteins showing differential expression in untreated versus treated NT2-R cells using aaptamine (1), identified by MALDI-ToF-MSa. The proteins aregrouped based on their molecular functions as indicated by PANTHER 7.2 software.
N Gene name Protein name Accession no. ProteinMw
ProteinPI
Proteinescore
Peptidecount
Number ofsequenced peptides
Sequence coverage(%)
Fold change
Binding5 CARHSP1 Calcium-regulated heat stable protein 1 Q9Y2V2 15,882 8.4 97 5 1 49 0.348 HNRNPUL2 Heterogeneous nuclear ribonucleoprotein
U-like protein 2Q1KMD3 85,052 4.9 332 16 4 26 0.41
11 PCBP2 Poly(rC)-binding protein 2 Q15366 38,556 6.3 328 17 3 61 0.48
Catalytic activity3 AS3MT Arsenite methyltransferase Q9HBK9 41,720 5.8 167 20 1 61 0.326 GNB1 Guanine nucleotide-binding protein
G(I)/G(S)/G(T) subunit beta-1P62873 37,353 5.6 317 7 3 30 0.4
7 PPT1 Palmitoyl-protein thioesterase 1 P50897 34,171 6.1 509 10 5 38 0.410 PSMC5 26S protease regulatory subunit 8 P62195 45,597 7.1 228 23 2 65 0.4621 PPT1 Palmitoyl-protein thioesterase 1 P50897 34,171 6.1 379 9 4 35 3.1
Structural molecule activity1 VIM Vimentin P08670 53,619 5.1 726 29 4 74 0.292 VIM Vimentin P08670 53,619 5.1 610 23 4 63 0.314 TUBB2B Tubulin beta-2B chain Q9BVA1 49,921 4.8 302 18 3 40 0.4919 CTTN Src substrate cortactin Q14247 61,599 5.2 209 12 3 22 2.2922 CFL1 Cofilin-1 P23528 18,491 8.2 295 13 2 65 4.1
Transcription regulator activity12 NACA Nascent polypeptide-associated
complex subunit alphaQ13765 23,370 4.5 379 8 3 35 0.48
13 NACA Nascent polypeptide-associatedcomplex subunit alpha
Q13765 23,370 4.5 115 8 1 36 0.49
16 CBX5 Chromobox protein homolog 5 P45973 22,211 5.7 337 9 3 45 2.0217 CBX5 Chromobox protein homolog 5 P45973 22,211 5.7 593 19 5 74 2.1418 RUVBL2 RuvB-like 2 Q9Y230 51,125 5.5 532 27 4 57 2.23
Translation regulator activity4 EIF5A Eukaryotic translation initiation factor 5A-1 P63241 16,821 5.1 402 13 2 71 0.3415 EEF1D Elongation factor 1-delta P29692 31,103 4.9 384 15 3 55 0.5
Unmapped9 HSP90B1 Endoplasmin P14625 92,411 4.8 423 26 4 33 0.4220 BANF1 Barrier-to-autointegration factor O75531 10,052 5.8 173 8 1 60 2.36
a Spot numbers are equal to spot numbers in Fig. 2.
232JO
UR
NA
LO
FPR
OT
EO
MIC
S96
(2014)
223–239

Table 3 – List of proteins showing differential expression in untreated versus treated NT2-R cells using demethyloxyaaptamine (2), identified by MALDI-ToF-MSa. Theproteins are grouped based on their molecular functions as indicated by PANTHER 7.2 software.
N Gene name Protein name Accessionno.
ProteinMw
ProteinPI
Proteinescore
Peptidecount
Number ofsequencedpeptides
Sequencecoverage
(%)
Foldchange
Binding2 TFF2 STAM-binding protein Q03403 48,047 5.9 299 14 2 41 0.2596 RBM4 RNA-binding protein 4 Q9BWF3 40,289 6.6 659 25 3 66 0.419
Catalytic activity3 GDI2 Rab GDP dissociation inhibitor beta P50395 50,631 6.1 522 28 4 69 0.3754 AHCY Adenosylhomocysteinase P23526 47,685 5.9 374 16 3 38 0.3918 PHGDH D-3-phosphoglycerate dehydrogenase O43175 56,614 6.3 136 19 1 42 0.44211 PSMA5 Proteasome subunit alpha type-5 P28066 26,394 4.7 388 12 3 63 2.268
Structural molecule activity5 GRSF1 G-rich sequence factor 1 Q12849 50,138 5.7 353 21 4 55 0.40610 SEPT2 Septin-2 Q15019 41,461 6.2 772 24 5 66 0.49512 KRT18 Keratin. type I cytoskeletal 18 P05783 48,029 5.3 285 13 3 38 2.598
Transcription regulator activity9 CTBP2 C-terminal-binding protein 2 P56545 48,914 6.5 350 19 3 41 0.46514 PQBP1 Polyglutamine-binding protein 1 O60828 30,454 5.9 334 6 3 35 3.799
Translation regulator activity7 EIF6 Eukaryotic translation initiation factor 6 P56537 26,582 4.6 956 13 5 64 0.426
Unmapped1 WDR55 WD repeat-containing protein 55 Q9H6Y2 42,097 4.8 408 14 4 48 0.25313 HSPA1A Heat shock 70 kDa protein 1 P08107 70,009 5.5 637 31 4 44 2.79515 ENO1 Alpha-enolase P06733 47,139 7.0 379 23 2 61 4.14616 ENO1 Alpha-enolase P06733 47,139 7.0 97 12 1 38 12.125
a Spot numbers are equal to spot numbers in Fig. 3.
233JO
UR
NA
LO
FPR
OT
EO
MIC
S96
(2014)
223–239

Table 4 – List of proteins showing differential expression in untreated versus treated NT2-R cells using isoaaptamine (3), identified by MALDI-ToF-MSa. The proteins aregrouped based on their molecular functions as indicated by PANTHER 7.2 software.
N Gene name Protein name Accession No ProteinMw
ProteinPI
Proteinescore
Peptidecount
Number ofsequenced peptides
Sequencecoverage
(%)
Foldchange
Binding10 PSMD4 26S proteasome non-ATPase regulatory subunit 4 P55036 40,711 4.7 531 15 3 42 2.123
Catalytic activity2 GDI2 Rab GDP dissociation inhibitor beta P50395 50,631 6.1 430 26 4 65 0.4463 GART Trifunctional purine biosynthetic protein adenosine-3 P22102 107,699 6.3 418 27 3 36 0.4614 GSTP1 Glutathione S-transferase P P09211 23,341 5.4 211 10 2 55 0.4615 CTSD Cathepsin D P07339 44,524 6.1 490 20 5 54 2.0016 GANAB Neutral alpha-glucosidase AB Q14697 106,807 5.7 352 24 4 29 2.0089 UCHL1 Ubiquitin carboxyl-terminal hydrolase
isozyme L1P09936 24,808 5.3 339 13 2 69 2.092
16 NCL Nucleolin P19338 76,568 4.6 290 11 3 22 3.001
Structural molecule activity11 KRT8 Keratin. type II cytoskeletal 8 P05787 53,671 5.5 770 22 4 58 2.159
Transcription regulator activity13 PQBP1 Polyglutamine-binding protein 1 O60828 30,454 5.9 311 6 2 35 2.541
Translation regulator activity15 EIF3I Eukaryotic translation initiation factor 3 subunit I Q13347 36,479 5.4 545 17 5 56 2.878
Unmapped1 GRPEL1 GrpE protein homolog 1, mitochondrial Q9HAV7 24,264 8.2 388 10 4 49 0.3187 MAD1L1 Mitotic spindle assembly checkpoint protein MAD1 Q9Y6D9 83,016 5.7 421 24 4 38 2.0798 PDCD6IP Programmed cell death 6-interacting protein Q8WUM4 95,963 6.1 191 17 4 29 2.08112 SCRN1 Secernin-1 Q12765 46,353 4.7 226 15 2 44 2.17514 IMMT Mitochondrial inner membrane protein Q16891 83,626 6.1 287 17 3 26 2.62517 GRIPAP1 GRIP1-associated protein 1 Q4V328 95,931 5.1 149 14 2 25 3.131
a Spot numbers are equal to spot numbers in Fig. 4.
234JO
UR
NA
LO
FPR
OT
EO
MIC
S96
(2014)
223–239

Fig. 5 – Protein interaction network and functional analysis. (A) Protein interaction networks were constructed using theIngenuity Pathway Analysis tool (IPA). Relationships between proteins regulated under aaptamine, demethyloxyaaptamine, orisoaaptamine treatment (50 μM, 4.5 μM, and 12.5 μM, respectively, for 48 h), proteins predicted to be involved in interactions,as well as relevant signal pathways are illustrated. Gray nodes represent regulated proteins detected by proteomics aftertreatment with the substance. Dashed lines represent indirect relationships, solid lines represent direct relationships recordedin the IPA database. (B) PANTHER Classification System analysis of proteins regulated in NT2-R cells under aaptamine,demethyloxyaaptamine, or isoaaptamine treatment with respect to biological processes and molecular function ontology,revealing the illustrated distribution of involved processes.
235J O U R N A L O F P R O T E O M I C S 9 6 ( 2 0 1 4 ) 2 2 3 – 2 3 9
PANTHER were rather similar, especially for compounds 2 and 3(Fig. 5B). Based on the results of this analysis, we concludethat demethyloxyaaptamine and isoaaptamine significantlyinfluence the cellular metabolism, which correlates with theMTT and trypan blue assay data. IPA based hypotheticaltoxicological analyses predicted kidney, liver and heart as majortarget organs of the investigated compounds. The comparison oftoxicologic profiles of the compounds also revealed a significantdegree of similarity between compounds 1, 2, and 3, and again inparticular between demethyloxyaaptamine and isoaaptamine(Fig. 6).
4.3. Validation of differentially expressed proteins
Toconfirm theproteomics data,weanalyzed changes in severalselected proteins that showed up-regulation upon treatmentwith the three compounds. The alterations of vimentinand cofilin-1, the most significantly regulated proteins inaaptamine-treated NT2-R cells, as well as eIF5A were found toresult from post-translational modifications (Fig. 7A). Thesefindings are in line with our previous observations, wheresimilar changes, suggesting inactivation of cofilin-1 and activa-tion of eIF5A, were observed in aaptamine-treated NT2 cells [7].

Fig. 6 – Hypothetical toxicological analysis. The expected toxicity for different organs and systems as possible side effects ofusing aaptamine or its derivatives in humans, was evaluated using IPA analysis. p-Value is indicated. Overlapping of thepredicted drug toxicity profiles is presented by a Venn diagram.
236 J O U R N A L O F P R O T E O M I C S 9 6 ( 2 0 1 4 ) 2 2 3 – 2 3 9
Cofilin-1 is involved in actin remodeling processes during cellmotility, division, and axon development [41,42]. The observedphosphorylation, a post-translationalmodification of vimentin,regulates filament assembly and disassembly (reviewed in [43]).This process results in regulation of intermediate filamentstructure and alteration of several signaling pathways [29].
eIF5A is the only protein known to undergo hypusination [44].The exact role of this protein remains unclear, although severalstudies have shown an essential role of the hypusinated isoformof eIF5A in cell growth and differentiation [45–48]. At the sametime, we could show that forced hypusination of eIF5A did notalter the cell growth [7] in germ tumor cells.
We furthermore found that treatment with demethyl-oxyaaptamine resulted in up-regulation of Hsp70 (heat-shockprotein 70 kDa), as well as alterations of different isoforms ofalpha-enolase in NT2-R cells (Fig. 7B). The essential cellularfunctions of Hsp70 are protein folding and protection from celldeath. The role of this protein in cancer is double-edged, as it canprotect cancer cells from apoptosis, and on the other hand, itexerts an anti-cancer effect by playing a role in the presentationof cancer cells to the immune system, thus stimulating tumorimmunogenicity (reviewed in [49]). Alpha-enolase is a glycolyticenzyme that generatesATPduring glycolysis. It is associatedwithtumor progression, as it plays a role in growth control andhypoxia tolerance [50,51]. The changes observed in alpha-enolase
isoform regulation in this study are similar to those described forthe retinoic acid-induceddifferentiationofmurine germline stemcells and embryonic stem cells [46].
An up-regulation of the spot corresponding to the minornucleolin isoform was found in isoaaptamine-treated cells(Fig. 7C). Being one of the major nuclear proteins, nucleolin hasbeen reported to have various functions in activities such asself-cleaving, DNA helicase, RNA helicase, and DNA-dependentATPase (reviewed in [52]). Furthermore, it binds histone H1 andthus induces chromatin decondensation [33]. The observed effectcould be due to the dissociation of its complex with histone H1[31,33], whichwas also detected as amajor spot with pI ≥ 7.0 andbigger Mw.
4.4. Hypusination of eIF5A
Eukaryotic initiation factor 5A-1 is highly conserved in alleukaryotic species. This factor is known to be activated by ahypusination step — an addition of the unique amino acidhypusine at the Lys50 position. eIF5A is the only proteinknown to contain hypusine [44,53–56]. For the first time, werecently described the promotion of eIF5A hypusination inNT2 cells under aaptamine treatment [7]. In the present study,the same process was observed in aaptamine-treated NT2-Rcells (Fig. 7A). At the same time, in NT2-R cells treated with

Fig. 7 – Western blotting and 2D-WB analyses upon treatment with aaptamine, demethyloxyaaptamine, or isoaaptamine.Beta-actin or alpha-tubulin was used as loading control. (A) Analysis of changes in selected proteins in aaptamine treatedNT2-R cells. 2D-WB with anti-eIF5A antibody revealed down-regulation of the non-hypusinated (inactive) form of eIF5A(indicated with an arrow). 2D-WB analysis with anti-cofilin-1 antibody detected two specific protein spots with different pI.Up-regulation of the more acidic (phosphorylated) protein form is indicated with an arrow. 2D-WB analysis with anti-vimentinantibody detected three specific protein spots with different Mw and pI. Down-regulation of the more acidic (phosphorylated)protein form, initially detected with 2D-PAGE, is indicated with an arrow. The total expression (analyzed by Western blotting)of all three analyzed proteins is unchanged. (B) Analysis of changes in selected proteins in demethyloxyaaptamine treatedNT2-R cells. Western blotting analysis with anti-Hsp70 antibody revealed up-regulation of the total protein expression ofHsp70. 2D-WB with anti-α-enolase antibody revealed a number of α-enolase isoforms with different Mw and pI. The spotsidentified by 2D-PAGE are indicated with arrows. An increase in hypusination of eIF5A is not detected. (C) Analysis of changesin several proteins in isoaaptamine treated NT2-R cells. 2D-WB with anti-nucleolin antibody revealed up-regulation of a spotwith Mw = 100 kDa and pI = 5.4 (indicated with an arrow) together with a non-regulated major spot with Mw = 110 kDa andpI ≥ 7.0. An increase in hypusination of eIF5A is not observed.
237J O U R N A L O F P R O T E O M I C S 9 6 ( 2 0 1 4 ) 2 2 3 – 2 3 9

238 J O U R N A L O F P R O T E O M I C S 9 6 ( 2 0 1 4 ) 2 2 3 – 2 3 9
demethyloxyaaptamine and isoaaptamine, which are struc-turally closely related analogs of aaptamine, promotion ofeIF5A hypusination was not observed (Fig. 7B, C). Therefore,promotion of hypusination in NT2-R germ cell tumor cells is aunique and specific characteristic of aaptamine, and thiseffect is abolished in the two derivatives by the minorstructural alterations (see Fig. 1A) that are seen in the twocompounds in comparison to the lead compound.
5. Conclusion
Aaptamine and its derivatives demethyloxyaaptamine andisoaaptamine are equally effective as anti-cancer agents incisplatin-sensitive and -resistant germ cell tumor cells. Whileaaptamine mainly exerts an antiproliferative effect at the IC50,demethyloxyaaptamine and isoaaptamine show strong induc-tion of apoptosis. A global proteome screening approach by2D-PAGE revealed specific protein alterations upon treatmentwith aaptamine, demethyloxyaaptamine, and isoaaptamine.Interestingly, despite the heterogeneous list of altered proteins,gene ontology analyses identified rather similar profiles ofaffected molecular functions in treated cells. Ingenuity pathwayanalysis (IPA) of altered proteins suggestedmyc, p53, and TNF aspossible direct or indirect targets or effectors of the alkaloids.Hypusination of eIF5A, which is a prominent feature ofaaptamine treatment, was not observed upon treatment withdemethyloxyaaptamine or isoaaptamine. The identification ofchanges in protein expression andproteinmodifications inducedby these aaptamine alkaloids is an important step towards abetter understanding of the biological activity of these promisingagents on a molecular level.
Supplementary data to this article can be found online athttp://dx.doi.org/10.1016/j.jprot.2013.11.009.
Acknowledgment
This work was supported by an individual grant fromDeutscher Akademischer Austauschdienst (DAAD) (S.A.D.),by grant 546.2012.4, supporting the leading Russian scientificschools, by the Program of the Presidium of RAS “Molecularand Cell Biology”, and by grant 13-03-00986 from the RussianFoundation for Basic Research (S.A.D., S.N.F., L.K.S., V.A.S.), bythe Werner Otto Stiftung (F.H. and S.B.), and by FEB RAS grant13-III-В-05-009 (S.A.D.).
R E F E R E N C E S
[1] Nakamura H, Kobayashi J, Ohizumi Y. Isolation and structureof aaptamine a novel heteroaromatic substance possessingalpha-blocking activity from the sea sponge Aaptos aaptos.Tetrahedron Lett 1982;23:5555–8.
[2] Larghi EL, Bohn ML, Kaufman TS. Aaptamine and relatedproducts. Their isolation, chemical syntheses, and biologicalactivity. Tetrahedron 2009;65:4257–82.
[3] Shubina LK, Kalinovsky AI, Fedorov SN, Radchenko OS,Denisenko VA, Dmitrenok PS, et al. Aaptamine alkaloids fromthe Vietnamese sponge Aaptos sp. Nat Prod Commun2009;4:1085–8.
[4] Bowling JJ, Pennaka HK, Ivey K, Wahyuono S, Kelly M,Schinazi RF, et al. Antiviral and anticancer optimizationstudies of the DNA-binding marine natural productaaptamine. Chem Biol Drug Des 2008;71:205–15.
[5] Tsukamoto S, Yamanokuchi R, Yoshitomi M, Sato K, Ikeda T,Rotinsulu H, et al. Aaptamine, an alkaloid from the spongeAaptos suberitoides, functions as a proteasome inhibitor.Bioorg Med Chem Lett 2010;20:3341–3.
[6] Jin MH, Zhao WN, Zhang YW, Kobayashi M, Duan HQ, KongDX. Antiproliferative effect of aaptamine on human chronicmyeloid leukemia K562 cells. Int J Mol Sci 2011;12:7352–9.
[7] Dyshlovoy SA,Naeth I, Venz S, PreukschasM, Sievert H, JacobsenC, et al. Proteomic profiling of germ cell cancer cells treated withaaptamine, a marine alkaloid with antiproliferative activity. JProteome Res 2012;11:2316–30.
[8] Aoki S, Kong DX, Suna H, Sowa Y, Sakai T, Setiawan A, et al.Aaptamine, a spongean alkaloid, activates p21 promoter in ap53-independent manner. Biochem Biophys Res Commun2006;342:101–6.
[9] Shubina LK, Makarieva TN, Dyshlovoy SA, Fedorov SN,Dmitrenok PS, Stonik VA. Three new aaptamines from themarine sponge Aaptos sp. and their proapoptotic properties.Nat Prod Commun 2010;5:1881–4.
[10] Fedoreev SA, Prokof'eva NG, Denisenko VA, Rebachuk NM.Cytotoxic activity of aaptamines from suberitid marinesponges. Pharm Chem J 1988;22:615–8.
[11] Dyshlovoy S, Fedorov S, Shubina L, Honecker F, Stonik V.Anticancer activity of aaptamine and its derivatives isolatedfrom marine Vietnamese sponge Aaptos sp. Ann Oncol2011;22:33.
[12] Andrews PW. Retinoic acid induces neuronal differentiationof a cloned human embryonal carcinoma cell-line in vitro.Dev Biol 1984;103:285–93.
[13] Port M, Glaesener S, Ruf C, Riecke A, Bokemeyer C, Meineke V,et al. Micro-RNA expression in cisplatin resistant germ celltumor cell lines. Mol Cancer 2011;10.
[14] Oechsle K, Honecker F, Cheng T, Mayer F, Czaykowski P,Winquist E, et al. Preclinical and clinical activity of sunitinibin patients with cisplatin-refractory or multiply relapsedgerm cell tumors: a Canadian Urologic Oncology Group/German Testicular Cancer Study Group cooperative study.Ann Oncol 2011;22:2654–60.
[15] Sieuwerts AM, Klijn JGM, Peters HA, Foekens JA. The MTTtetrazolium salt assay scrutinized — how to use this assayreliably to measure metabolic-activity of cell-cultures in vitrofor the assessment of growth characteristics, IC50-values andcell survival. Eur J Clin Chem Clin 1995;33:813–23.
[16] Preukschas M, Hagel C, Schulte A, Weber K, Lamszus K,Sievert H, et al. Expression of eukaryotic initiation factor 5Aand hypusine forming enzymes in glioblastoma patientsamples: implications for new targeted therapies. Plos One2012;7.
[17] Glaesener S, Honecker F, Veltman IM, Gillis AJM, Rohlfing T,Streichert T, et al. Comparative proteome, transcriptome, andgenome analysis of a gonadal and an extragonadal germ celltumor cell. J Proteome Res 2008;7:3890–9.
[18] Ummanni R, Mundt F, Pospisil H, Venz S, Scharf C, Barett C,et al. Identification of clinically relevant protein targets inprostate cancer with 2D-DIGE coupled mass spectrometryand systems biology network platform. Plos One 2011;6.
[19] Bradford MM. A rapid and sensitive method for the quantitationof microgram quantities of protein utilizing the principle ofprotein-dye binding. Anal Biochem 1976;72:248–54.
[20] Dyshlovoy SA, Fedorov SN, Kalinovsky AI, Shubina LK,Bokemeyer C, Stonik VA, et al. Mycalamide A shows cytotoxicproperties and prevents EGF-induced neoplastic transformationthrough inhibition of nuclear factors. Mar Drugs 2012;10:1212–24.
[21] Junker H, Venz S, Zimmermann U, Thiele A, Scharf C,Walther R. Stage-related alterations in renal cell

239J O U R N A L O F P R O T E O M I C S 9 6 ( 2 0 1 4 ) 2 2 3 – 2 3 9
carcinoma — comprehensive quantitative analysis by2D-DIGE and protein network analysis. Plos One 2011;6:e21867.
[22] Koch S, Mayer F, Honecker F, Schittenhelm M, Bokemeyer C.Efficacy of cytotoxic agents used in the treatment oftesticular germ cell tumours under normoxic andhypoxic conditions in vitro. Brit J Cancer2003;89:2133–9.
[23] Nicoletti I, Migliorati G, Pagliacci MC, Grignani F, Riccardi C. Arapid and simple method for measuring thymocyte apoptosisby propidium iodide staining and flow cytometry. J ImmunolMethods 1991;139:271–9.
[24] Thomas PD, Kejariwal A, Campbell MJ, Mi HY, Diemer K, GuoN, et al. PANTHER: a browsable database of gene productsorganized by biological function, using curated protein familyand subfamily classification. Nucleic Acids Res2003;31:334–41.
[25] Paulsen MT, Ljungman M. The natural toxin juglone causesdegradation of p53 and induces rapid H2AX phosphorylationand cell death in human fibroblasts. Toxicol Appl Pharmacol2005;209:1–9.
[26] Park MH. The post-translational synthesis of apolyamine-derived amino acid, hypusine, in the eukaryotictranslation initiation factor 5A (eIF5A). J Biochem2006;139:161–9.
[27] Heyworth PG, Robinson JM, Ding JB, Ellis BA, Badwey JA.Cofilin undergoes rapid dephosphorylation in stimulatedneutrophils and translocates to ruffled membranes enrichedin products of the NADPH oxidase complex. Evidence for anovel cycle of phosphorylation and dephosphorylation.Histochem Cell Biol 1997;108:221–33.
[28] Kobayashi M, Nishita M, Mishima T, Ohashi K, Mizuno K.MAPKAPK-2-mediated LIM-kinase activation is critical forVEGF-induced actin remodeling and cell migration. EMBO J2006;25:713–26.
[29] Eriksson JE, He T, Trejo-Skalli AV, Harmala-Brasken AS,Hellman J, Chou YH, et al. Specific in vivo phosphorylationsites determine the assembly dynamics of vimentinintermediate filaments. J Cell Sci 2004;117:919–32.
[30] Inagaki M, Matsuoka Y, Tsujimura K, Ando S, Tokui T,Takahashi T, et al. Dynamic property of intermediatefilaments: regulation by phosphorylation. Bioessays1996;18:481–7.
[31] Gotzmann J, Eger A,MeissnerM, GrimmR, Gerner C, SauermannG, et al. Two-dimensional electrophoresis reveals a nuclearmatrix-associated nucleolin complex of basic isoelectric point.Electrophoresis 1997;18:2645–53.
[32] Jordan G. At the heart of the nucleolus. Nature1987;329:489–90.
[33] Erard MS, Belenguer P, Caizerguesferrer M, Pantaloni A,Amalric F. A major nucleolar protein, nucleolin, induceschromatin decondensation by binding to histone H1. Eur JBiochem 1988;175:525–30.
[34] Mosmann T. Rapid colorimetric assay for cellular growth andsurvival: application to proliferation and cytotoxicity assays. JImmunol Methods 1983;65:55–63.
[35] Slater TF. Studies on a succinate-neotetrazolium reductasesystem of rat liver: II. Points of coupling with the respiratorychain II. Points of coupling with the respiratory chain.Biochim Biophys Acta 1963;77:365–82.
[36] Liu YB, Peterson DA, Kimura H, Schubert D. Mechanism ofcellular 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazoliumbromide (MTT) reduction. J Neurochem 1997;69:581–93.
[37] Strober W. Trypan blue exclusion test of cell viability.Current Protocols in Immunology. John Wiley & Sons, Inc.;2001.
[38] Schmidt EV. The role of c-myc in regulation of translationinitiation. Oncogene 2004;23:3217–21.
[39] Vousden KH. p53: death star. Cell 2000;103:691–4.
[40] Bertazza L, Mocellin S. The dual role of tumor necrosisfactor (TNF) in cancer biology. Curr Med Chem2010;17:3337–52.
[41] Arber S, Barbayannis FA, Hanser H, Schneider C, Stanyon CA,Bernard O, et al. Regulation of actin dynamics throughphosphorylation of cofilin by LIM-kinase. Nature1998;393:805–9.
[42] Keezer SM, Ivie SE, Krutzsch HC, Tandle A, Libutti SK, RobertsDD. Angiogenesis inhibitors target the endothelial cellcytoskeleton through altered regulation of heat shock protein27 and cofilin. Cancer Res 2003;63:6405–12.
[43] Satelli A, Li SL. Vimentin in cancer and its potential as amolecular target for cancer therapy. Cell Mol Life Sci2011;68:3033–46.
[44] Park MH, Wolff EC, Folk JE. Hypusine: its post-translationalformation in eukaryotic initiation factor 5A and its potentialrole in cellular regulation. Biofactors 1993;4:95–104.
[45] Balabanov S, Ziegler P, Hartmann U, Kammer W, Copland M,Priemer M, et al. Inhibition of hypusination of eukaryoticinitiation factor 5a (eIF5A) as a novel a synergistic treatmentstrategy in imatinib-treated BCR-ABL positive leukemiasidentified by a global proteomics approach. Blood2005;106:565a–6a.
[46] Dihazi H, Dihazi GH, Jahn O, Meyer S, Nolte J, Asif AR, et al.Multipotent adult germline stem cells and embryonic stemcells functional proteomics revealed an important role ofeukaryotic initiation factor 5A (Eif5a) in stem celldifferentiation. J Proteome Res 2011;10:1962–73.
[47] Balabanov S, Gontarewicz A, Ziegler P, Hartmann U, KammerW, Copland M, et al. Hypusination of eukaryotic initiationfactor 5A (eIF5A): a novel therapeutic target inBCR-ABL-positive leukemias identified by a proteomicsapproach. Blood 2007;109:1701–11.
[48] Luchessi AD, Cambiaghi TD, Hirabara SM, Lambertucci RH,Silveira LR, Baptista IL, et al. Involvement of eukaryotictranslation initiation factor 5A (eIF5A) in skeletal musclestem cell differentiation. J Cell Physiol 2009;218:480–9.
[49] Gong J, Theriault J, Calderwood S. Hsp70-based anticancervaccines: chaperoning the immune response. In: CalderwoodS, Sherman M, Ciocca D, editors. Heat Shock Proteins inCancer. The Netherlands: Springer; 2007. p. 367–82.
[50] Subramanian A, Miller DM. Structural analysis of alpha-enolase—mapping the functional domains involved in down-regulationof the c-myc protooncogene. J Biol Chem 2000;275:5958–65.
[51] Trojanowicz B,Winkler A, Hammje K, ChenZX, Sekulla C, GlanzD, et al. Retinoic acid-mediated down-regulation of ENO1/MBP-1gene products caused decreased invasiveness of the follicularthyroid carcinoma cell lines. J Mol Endocrinol 2009;42:249–60.
[52] Tuteja R, Tuteja N. Nucleolin: a multifunctional majornucleolar phosphoprotein. Crit Rev Biochem Mol Biol1998;33:407–36.
[53] Jakus J, Wolff EC, Park MH, Folk JE. Features of thespermidine-binding site of deoxyhypusine synthase asderived from inhibition studies — effective inhibition bybis-guanylated and mono-guanylated diamines andpolyamines. J Biol Chem 1993;268:13151–9.
[54] Tome ME, Fiser SM, Payne CM, Gerner EW. Excess putrescineaccumulation inhibits the formation of modified eukaryoticinitiation factor 5A (eIF-5A) and induces apoptosis. Biochem J1997;328(Pt 3):847–54.
[55] Zhang JT, Dong ZZ, Arnold RJ, Yang YY, Park MH, Hrncirova P,et al. Modulation of differentiation-related gene 1 expressionby cell cycle blocker mimosine, revealed by proteomicanalysis. Mol Cell Proteomics 2005;4:993–1001.
[56] Thompson JE, Taylor CA, Sun Z, Cliche DO, Ming H, EshaqueB, et al. Eukaryotic translation initiation factor 5A inducesapoptosis in colon cancer cells and associates with thenucleus in response to tumour necrosis factor alphasignalling. Exp Cell Res 2007;313:437–49.





























