Embed Size (px)
Citation preview

Contents lists available at SciVerse ScienceDirect
Ann Allergy Asthma Immunol 111 (2013) 32e37
Characterization of Cannabis sativa allergensAjay P. Nayak, PhD *; Brett J. Green, PhD *; Gordon Sussman, MD y; Noam Berlin, BSc z; Hemant Lata, PhD x;Suman Chandra x; Mahmoud A. ElSohly, PhD x; Justin M. Hettick, PhD *; and Donald H. Beezhold, PhD *
*Allergy and Clinical Immunology Branch, Health Effects Laboratory Division, National Institute for Occupational Safety and Health, Centers for Disease Control and Prevention,Morgantown, West VirginiayDivision of Allergy and Clinical Immunology, Department of Medicine, University of Toronto, Toronto, Ontario, CanadazGordon Sussman Clinical Allergy Research Inc, Toronto, Ontario, CanadaxNational Center for Natural Product Research, Research Institute of Pharmaceutical Sciences, School of Pharmacy, University of Mississippi, Mississippi
A R T I C L E I N F O
Article history:Received for publication March 26, 2013.Received in revised form April 17, 2013.Accepted for publication April 27, 2013.
A
BmkO
Reprints: Donald H. Beezhold, PhD, Allergy and CWillowdale Rd, National Institute for OccupationaWV 26505; E-mail: [email protected]: Authors have nothing to disclose.Disclaimer: The findings and the conclusions in thand do not necessarily represent the views of thtional Safety and Health.Funding Sources: This study was supported in paNS12-10 with the National Institute of EnvironCenters for Disease Control and Prevention andDrug Abuse, National Institute of Health, DepServices, contract N01DA-10-7773.
1081-1206/13/$36.00 - see front matter � 2013 Ahttp://dx.doi.org/10.1016/j.anai.2013.04.018
B S T R A C T
ackground: Allergic sensitization to Cannabis sativa is rarely reported, but the increasing consumption ofarijuana has resulted in an increase in the number of individuals who become sensitized. To date, little isnown about the causal allergens associated with C sativa.bjective: To characterize marijuana allergens in different components of the C sativa plant using serum IgE
from marijuana sensitized patients.Methods: Serum samples from 23 patients with a positive skin prick test result to a crude C sativa extractwere evaluated. IgE reactivity was variable between patients and C sativa extracts. IgE reactivity to C sativaproteins in Western blots was heterogeneous and ranged from 10 to 70 kDa. Putative allergens derived from2-dimensional gels were identified.Results: Prominent IgE reactive bands included a 23-kDa oxygen-evolving enhancer protein 2 and a 50-kDaprotein identified to be the photosynthetic enzyme ribulose-1,5-bisphosphate carboxylase/oxygenase.Additional proteins were identified in the proteomic analysis, including those from adenosine triphosphatesynthase, glyceraldehyde-3-phosphate dehydrogenase, phosphoglycerate kinase, and luminal bindingprotein (heat shock protein 70), suggesting these proteins are potential allergens. Deglycosylation studieshelped refine protein allergen identification and demonstrated significant IgE antibodies against plantoligosaccharides that could help explain cross-reactivity.Conclusion: Identification and characterization of allergens from C sativa may be helpful in furtherunderstanding allergic sensitization to this plant species.� 2013 American College of Allergy, Asthma & Immunology. Published by Elsevier Inc. All rights reserved.
1
IntroductionCannabis sativa is an herbaceous angiosperm belonging to thefamily Cannabaceae. C sativa and its resinous derivative hashish havea variety of industrial and more recently medicinal applications. Theplant is most well known for its use as a recreational drug because itcontains the psychoactive compound, D-9-tetrahydrocannabinol. Anincrease in marijuana consumption has been observed among
linical Immunology Branch, 1095l Safety and Health, Morgantown,
is report are those of the authorse National Institute for Occupa-
rt by interagency agreement 12-mental Health Sciences and thein part by National Institute onartment of Health and Human
merican College of Allergy, Asthma &
teenagers in the United States. As a result of the increasing social andmedical use, reports of allergic sensitization to marijuana areincreasing in the peer-reviewed literature.2e10
Cases have been reported where hypersensitivity and evenanaphylactic responses have been associated with marijuana use,and clinical symptoms include sore throat, nasal congestion,rhinitis, pharyngitis, wheezing, dyspnea, angioedema, and lac-rimation.2e4,11,12 In long-term and high-dose users, more severemanifestations of bronchitis and asthma with reduced vitalcapacity have been reported.12
Allergic sensitization to C sativa has been reported in occupa-tional settings as well.13 Hemp workers involved in processinghemp fibers at a textile mill had significantly higher prevalence ofchronic respiratory symptoms attributed to byssinosis. Sensitiza-tion of laboratory workers that handle and test marijuana has alsobeen reported.14e16
The allergens of C sativa and its various derivatives are poorlycharacterized. Although D-9-tetrahydrocannabinol has been sug-gested to be an allergen,11 more recent studies show type 1
Immunology. Published by Elsevier Inc. All rights reserved.

A.P. Nayak et al. / Ann Allergy Asthma Immunol 111 (2013) 32e37 33
hypersensitivity to high-molecular-weight proteins derived from Csativa.4,6,9,10,17,18 In a recent study in Spain, IgE-binding proteinswere observed with molecular weights ranging from 10 to 69 kDa.4
Others have identified patients sensitized to 2 prominent IgE-binding bands located at 10 and 14 kDa,9 and a 9-kDa lipid trans-fer protein was identified to bind IgE from a patient sensitized to Csativa.17 However, the identity of most allergens from C sativaremains unknown, and no allergens are currently listed by theInternational Union of Immunological Societies allergen nomen-clature subcommittee.
Previously, we described 17 individuals who tested skin pricktest (SPT) positive to crude extracts of marijuana buds and flowers.3
In all patients, exposure was primarily through smoking and directcontact with the plant. However, one patient was additionallyexposed through consumption of marijuana tea. In this study, wecharacterize patient IgE reactivity to root, leaf, flower, and budextracts in an attempt to identify potential allergens for patientssensitized to C sativa.
Methods
Patient Population
Serum samples were obtained from 17 individuals with inha-lation and contact symptoms who were SPT positive to a crude Csativa extract from buds and flowers (macerated in water for 15minutes), as previously reported.3 The most common symptomsafter exposure to marijuana included rhinitis and conjunctivitis,periorbital angioedema, wheezing, and contact urticaria. Most SPT-positive patients had primary exposure to C sativa through smokingor direct contact to the plant. One patient also consumedmarijuanatea, which resulted in gastric cramping, vomiting, and anaphylaxis.Exposure was due to recreational use of marijuana in all cases. Sixadditional patients with similar clinical presentation and positiveSPT results to C sativa were also recruited during the study. Inaddition, serum samples from 7 individuals who consumed C sativabut were SPT negative to C sativa extracts and serum from a labo-ratory worker with no known exposure to C sativa were used asnegative controls. Deidentified serum samples were sent to theNational Institute for Occupational Safety and Health for serologicanalysis of IgE-binding protein and proteomic analysis. Writtenconsent was obtained from human subjects, and the ethicsapproval was obtained for this study from the Canadian ShieldEthics Review Board.
C Sativa Extracts
For protein analysis, extracts from various parts of C sativa LMexican variant (leaf, root, female flowers, and buds) wereprepared at the National Center for Natural Product Research,Research Institute of Pharmaceutical Sciences, School of Pharmacyat the University of Mississippi. Unpollinated female flowers wereused for generation of extracts and were devoid of seed compo-nents. Proteins were extracted from 100 mg of root, leaves, buds,and flowers using the total plant protein extraction kit (Sigma-Aldrich, St Louis, Missouri), according to the manufacturer’sinstructions.
SDS-PAGE, 2-Dimensional Gel Electrophoresis, and Western BlotAnalysis
Individual C sativa protein extracts (25 mg) were separated bysodium dodecyl sulphateepolyacrylamide gel electrophoresis(SDS-PAGE) using 12% polyacrylamide gels and stained withImperial Blue (Thermo Scientific, Rockford, Illinois) according to themanufacturer’s instructions.
For Western blot analyses, 25 mg of individual C sativa proteinextracts were separated as described and transferred overnight to
a nitrocellulose membrane (0.22 mm; BioRad, Hercules, California).The membrane was blocked using Tris-buffered saline (TBS) con-taining 3% bovine serum albumin (blocking buffer) for 1 hour. Themembrane was incubated with pooled sera from SPT-positivepatients, diluted 1:5 (vol/vol) in blocking buffer for analyzingpatient reactivity to the extracts from different parts of C sativaplants. For individual patient screening, the membrane was trans-ferred to a multiscreen apparatus (BioRad) for analysis and incu-bated with either individual SPT-positive or individual SPT-negative patient sera diluted 1:5 (vol/vol) in blocking buffer andincubated overnight at 4�C on a rocker. The following morning, themembrane was washed and incubated with mouse anti-human IgEmonoclonal antibody (clone GE-1; Sigma-Aldrich) diluted 1:5,000(vol/vol) in blocking buffer for 2 hours on a rocker at roomtemperature (RT). The membrane was washed 3 times with TBScontaining 0.05% Tween-20 (TBST) and incubated with alkalinephosphataseeconjugated goat anti-mouse IgG antibody (HþL;Promega, Madison,Wisconsin) diluted 1:5,000 (vol/vol) in blockingbuffer for 1 hour on a rocker at RT. The membrane was incubatedwith 1-Step NBT/BCIP (Promega) substrate solution for 15 to 20min. The reaction was stopped by washing the membrane withdistilled water.
To investigate the role of glycosylation in IgE reactivity to Csativa proteins, extracts from roots, leaves, buds, and flowers werepartially deglycosylated as previously described.19 The extractswere first separated by SDS-PAGE and transferred to nitrocellulosemembranes, each membrane containing samples from the 4different plant sources. One membrane was treated with 50 mMsodium periodate in 50 mM acetate buffer for 1 hour in dark at RT,whereas the other membrane served as a control and was treatedwith 50 mM acetate buffer alone for the same duration. Themembranes were washed 3 times for 10 minutes with phosphatebuffered saline (PBS) (pH 7.4) containing 0.05% Tween 20 (PBST)and blocked in PBST containing 3% nonfat dry milk (PBST-M). Themembranes were then incubated with pooled serum from SPT-positive patients, diluted 1:5 (vol/vol) in PBST-M, and processedfor determination of IgE reactivity as described above.
For 2-dimensional gel electrophoresis, C sativa protein extractsof leaves were processed as described earlier. Briefly, proteins weresolubilized in rehydration solution containing 8 M urea, 20 mMdithiothreitol, 2% CHAPS, 0.5% IPG buffer, and bromophenol blue.The solution was loaded onto a 7-cm IPG strip pH 3-10 (GEHealthcare, Uppsala, Sweden) and focused using an IPGphorapparatus (GE Healthcare). The strip was equilibrated in 50mM Trishydrochloride pH 8.8 (with 6 M urea, 65 mM dithiothreitol, 30%glycerol, 2% SDS, and bromophenol blue) before separation offocused proteins in the second dimension using SDS-PAGE. Stripswere processed in duplicate for each extract sample, and one set ofgels was stained with Imperial blue protein stain, whereas theother was processed for Western blot analysis.
All Western blots were analyzed with secondary and tertiaryantibody control by incubating samples with mouse anti-humanIgE antibody and goat anti-mouse IgG (HþL) and developing withNBT/BCIP as described earlier.
Proteomic Analysis and Homology Searching
Protein spots of interest were excised from 2-dimensional SDS-PAGE gels. Spots were destained, reduced, alkylated, and digestedwith porcine trypsin (Sigma-Aldrich) according to the methods ofShevchenko et al.20 Reduction was performed with 20 mM tribu-tylphosphine (Sigma-Aldrich), and trypsin digestion was per-formed overnight at 37�C with shaking (500 rpm). Digestedsupernatant was combined with extraction buffer and concen-trated in a centrifugal evaporator (Savant DNA120; Thermo Scien-tific, Asheville, North Carolina) before mass spectrometry analysis.

A.P. Nayak et al. / Ann Allergy Asthma Immunol 111 (2013) 32e3734
Aliquots of individual digested protein spots were subjectedto ultraperformance liquid chromatography on a nanoACQUITYsystem (Waters, Milford, Massachusetts). Samples were desalted ona 180 � 20-mm Symmetry C18 (5-mm particle) trap column with100% solvent A (0.1% formic acid in distilled, deionized water).Analytical separation was performed on a BEH130 C18 (1.7-mmparticle) using a gradient of 97/3 A/B (0.1% formic acid in acetoni-trile) to 50/50 A/B.
Eluent from the ultraperformance liquid chromatographysystem was directed to the positive nanoelectrospray source ofa Synapt (Waters) quadrupole time-of-flight mass spectrometer.Dry nitrogen was supplied as a desolvation gas. Ultrahigh purityargon was used as the collision gas for collision-induced dissocia-tion. Data were acquired in a data-dependent fashion according tothe following criteria: MS survey scans were performed from 100 to1,500 unified atomic mass units for 1 second. A mass-to-chargeratio of interest was selected for tandem mass spectrometry if itmet the intensity threshold of 20 counts per second and was eitherdoubly or triply charged, as determined by examination of themass-to-charge ratio gap between isotopes.
Data were analyzed by using ProteinLynx Global Server version2.4 to search the entire nonredundant SwissProt protein database(www.uniprot.org). The search was constrained by using 100-ppmmass accuracy, carbamidomethyl cysteine as a fixed modification,trypsin as the enzyme, and a minimum of 2 peptides to match. Theentire nonredundant database was searched for the full data set toeliminate any false-positive matches from common in gel digestcontaminants, such as keratin or trypsin autodigestion products.Unmatched de novo peptide sequences were blasted againstSwissProt using the PAM30MS scoring matrix.
The de novo peptide sequences were further analyzed to iden-tify putative allergens. The translated nucleotide database ortranscriptome shotgun assembly of C sativa (taxid 3483) was usedfor identification of putative messenger RNA sequences that tran-scribe the de novo peptide sequences. The transcriptome shotgunassembly database for C sativa was recently published in theNational Center for Biotechnology Information database collectionwith accession numbers between GenBank numbers JP449145 andJP482359.21 Putative messenger RNA sequences were translatedusing EMBOSS Transeq software, and appropriate in-frame
Figure 1. Sodium dodecyl sulphateepolyacrylamide gel electrophoresis (SDS-PAGE) showextracts were derived from leaves, buds, flowers, and roots of C sativa. A, Imperial proteinin daltons is indicated in the left margin as estimated using Precision plus protein all blu
translated protein sequences were identified in BLAST searches forhomologous proteins.
Results
IgE Reactivity to Various Parts of C sativa Plant
The protein profiles of root, leaf, bud, and flower extracts of Csativa are demonstrated in Figure 1A. Protein profiles of C sativaleaves, buds, and to some extent flowers were similar witha prominent approximately 50-kDa band common to theseextracts. The extracts from flowers demonstrated prominentprotein bands at approximately 18 and approximately 35 kDa thatwere absent in the other extracts. In contrast, the protein profile ofC sativa root extract was distinct but was lacking the predominantbands present in the other extracts.
For extracts from each of the C sativa plant sources, IgE frompooled serum reacted to multiple protein bands ranging from 20 to100 kDa. Immunoblotting showed some variability in IgE binding toeach plant component (Fig 1B); however, IgE reactivity was largelysimilar for extracts collected from leaves, buds, and flowers.Prominent staining was observed at approximately 50 kDa and 23kDa in these extracts. The IgE reactivity of pooled serum samples toroot extract showed a profile distinct to extracts from the othersources, with prominent staining at approximately 35 kDa. Inaddition to these prominent bands, immunostaining was alsoobserved for multiple other bands as well.
Individual patients had a wide spectrum of IgE reactivityranging from 10 to 100 kDa, with considerable variability. Eightof 23 SPT-positive patients (35%) demonstrated a similar patternof IgE reactivity (Fig 2A). In leaf extracts, 13 of 23 patients (56.5%)reacted to the approximately 50-kDa protein band. Eight patients(patients 3, 8, and 16-21) had immunoreactivity to a bandlocalized at approximately 23 kDa. Minor bands associated withIgE binding were also localized at approximately 15, 35, and75 kDa.
With the root extract, 9 patients (39%) did not have IgEbinding to root extract proteins (Fig 2B). Of the remaining 14patients, the reactivity was variable and mostly restricted to a 37-kDa band. Minor bands were identified at approximately 10, 13,15, 17, 25, 50, and 100 kDa. The 8 patients with the strongest
ing Cannabis sativa protein extracts profiles and pooled-serum IgE reactivity. Proteinstained SDS-PAGE gel. B, Pooled patient serum IgE reactivity. Molecular weight (MW)e standards.
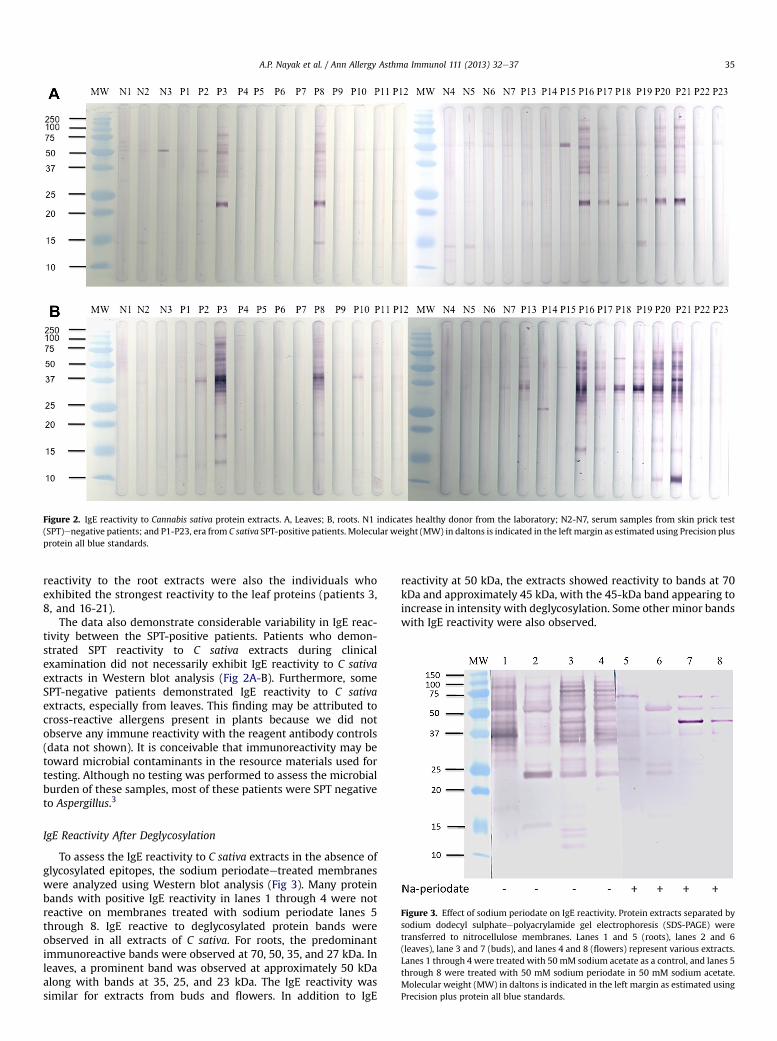
Figure 2. IgE reactivity to Cannabis sativa protein extracts. A, Leaves; B, roots. N1 indicates healthy donor from the laboratory; N2-N7, serum samples from skin prick test(SPT)enegative patients; and P1-P23, era from C sativa SPT-positive patients. Molecular weight (MW) in daltons is indicated in the left margin as estimated using Precision plusprotein all blue standards.
Figure 3. Effect of sodium periodate on IgE reactivity. Protein extracts separated bysodium dodecyl sulphateepolyacrylamide gel electrophoresis (SDS-PAGE) weretransferred to nitrocellulose membranes. Lanes 1 and 5 (roots), lanes 2 and 6(leaves), lane 3 and 7 (buds), and lanes 4 and 8 (flowers) represent various extracts.Lanes 1 through 4 were treated with 50 mM sodium acetate as a control, and lanes 5through 8 were treated with 50 mM sodium periodate in 50 mM sodium acetate.Molecular weight (MW) in daltons is indicated in the left margin as estimated usingPrecision plus protein all blue standards.
A.P. Nayak et al. / Ann Allergy Asthma Immunol 111 (2013) 32e37 35
reactivity to the root extracts were also the individuals whoexhibited the strongest reactivity to the leaf proteins (patients 3,8, and 16-21).
The data also demonstrate considerable variability in IgE reac-tivity between the SPT-positive patients. Patients who demon-strated SPT reactivity to C sativa extracts during clinicalexamination did not necessarily exhibit IgE reactivity to C sativaextracts in Western blot analysis (Fig 2A-B). Furthermore, someSPT-negative patients demonstrated IgE reactivity to C sativaextracts, especially from leaves. This finding may be attributed tocross-reactive allergens present in plants because we did notobserve any immune reactivity with the reagent antibody controls(data not shown). It is conceivable that immunoreactivity may betoward microbial contaminants in the resource materials used fortesting. Although no testing was performed to assess the microbialburden of these samples, most of these patients were SPT negativeto Aspergillus.3
IgE Reactivity After Deglycosylation
To assess the IgE reactivity to C sativa extracts in the absence ofglycosylated epitopes, the sodium periodateetreated membraneswere analyzed using Western blot analysis (Fig 3). Many proteinbands with positive IgE reactivity in lanes 1 through 4 were notreactive on membranes treated with sodium periodate lanes 5through 8. IgE reactive to deglycosylated protein bands wereobserved in all extracts of C sativa. For roots, the predominantimmunoreactive bands were observed at 70, 50, 35, and 27 kDa. Inleaves, a prominent band was observed at approximately 50 kDaalong with bands at 35, 25, and 23 kDa. The IgE reactivity wassimilar for extracts from buds and flowers. In addition to IgE
reactivity at 50 kDa, the extracts showed reactivity to bands at 70kDa and approximately 45 kDa, with the 45-kDa band appearing toincrease in intensity with deglycosylation. Some other minor bandswith IgE reactivity were also observed.

A.P. Nayak et al. / Ann Allergy Asthma Immunol 111 (2013) 32e3736
Two-Dimensional Gel Electrophoresis and Proteomic Analysis
Two-dimensional gel electrophoresis was combined withWestern blotting to select C sativa bands for allergen identification.Two-dimensional immunoblot demonstrated IgE reactivity atapproximately 50 kDa to a large area along the pH gradient (spots3-10) (Fig 4). A number of peptides could be identified by proteomicanalysis for spots 1 through 4, including the heavy chain subunit ofribulose-1,5-bisphosphate carboxylase/oxygenase (RuBisCO) andpeptides for adenosine triphosphate (ATP) synthase (spots 1 and 2)(eTable 1). IgE binding to the 2-dimensional blot appeared themostintense to a series of spots at approximately 23 kDa. Peptidesequences were obtained for these spots (spots 6-10) and identifiedas oxygen-evolving enhancer protein 2, a 23-kDa protein belongingto photosystem II. Spot 5 (approximately 45 kDa), which increasedin IgE-binding intensity with deglycosylation, contained peptidesfrom several enzymes, including RuBisCO activase 1 and 2, gluta-mine synthetase, phosphoglycerate kinase, glyceraldehyde-3-phosphate dehydrogenase, and camphor resistance protein. In theroot extract, we identified the ATP synthase b-subunit and luminalbinding protein (BiP) as potential allergens (data not shown).
Discussion
C sativa is widely used for various medicinal and industrialpurposes.22 Recently, an increase in the recreational use of C sativahas been reported.1,23 Studies have focused on the overall healtheffects of C sativa consumption with less attention to aspects suchas respiratory morbidity or allergic sensitization.22 Allergicresponses to C sativa are rarely reported; however, it is likely thatthe social and legal aspects surrounding the recreational use ofmarijuana may discourage patients from addressing their symp-toms with a physician. Exposure to C sativa has been reported tosensitize individuals and even results in serious anaphylacticreactions.3,11 Most cases described in the literature involve sensi-tization to C sativa via multiple exposures, presumably involvingrespiratory, cutaneous, and/or oral routes.6,24 Allergic sensitizationto C sativa has also been attributable to environmental C sativapollen exposures, particularly in the southwestern region of UnitedStates.25 To date, the allergens associated with C sativa exposureand sensitization have remained relatively uncharacterized.
Figure 4. Identification of IgE-binding antigens. Two-dimensional Western blotanalysis of IgE reactivity to Cannabis sativa leaf extracts using pooled serum IgE fromskin prick testepositive patients. Proteomic analysis was performed by de novosequencing of the peptides and matched with the existing UniProt database.Molecular weight (MW) in daltons is indicated in the left margin as estimated usingPrecision plus protein all blue standards.
In the present study, IgE binding to C sativa root, leaf, flower, andbud extracts was determined. The protein and IgE-binding profilesobserved in leaf, flower, or bud extracts were similar, indicatingsome common proteins in these parts of the plant. As expected, theprotein profile and the IgE reactivity of the root extract wassomewhat different. Analysis of individual SPT patient IgE reac-tivity demonstrated a variability in patterns of reactivity, consistentwith a previous observation.9 Some patients who were determinedto exhibit a positive SPT result did not show evidence of IgE bindingin Western blot analysis (Fig 2).
In the leaf extract, the predominant IgE reactivity was observedat 50 and 23 kDa, even after deglycosylation. Using proteomicanalysis, the 50-kDa protein was identified as the larger chainsubunit of RuBisCO, and the approximately 23-kDa protein wasidentified as oxygen-evolving enhancer protein 2 of the photo-system II, whereas multiple peptides and proteins were identifiedfor the 45-kDa protein. Previous studies have attempted to identifyallergens associatedwith C sativa sensitization,9,17 and sensitizationto a lipid transfer protein (LTP) was determined in one case report.17
A 10-kDa protein band, whichmay correspond to LTP, was proposedin European and Asian cohorts as a potential C sativa allergen.2,9,26
Although LTPs are common sensitizers in foods and plant-derivedaerosols,27,28 no LTP sequences were identified in our liquid chro-matographyemass spectrometry analysis; however, IgE reactivitywas observed at approximately 10 kDa for 2 patients with the Csativa root extract.
The present study follows previous reports of clinical determi-nation of hypersensitivity to C sativa.3,5 In this study, the prominentIgE reactive bands were identified as RuBisCO and oxygen-evolvingenhancer protein 2. Multiple peptides for the large (55 kDa) andsmall chains (14 kDa) of RuBisCO were identified in the liquidchromatographyemass spectrometry analysis. RuBisCO is the mostabundant protein in nature and catalyzes the conversion ofD-ribulose,1,5-bisphosphate to 2,3-phospho-D-glycerate in thepresence of carbon dioxide, a rate-limiting step for photo-synthesis.29e31 Although RuBisCO is generally susceptible to rapiddegradation by gastric fluid,32,33 IgE responses have been generatedon parenteral administration of this protein.34,35 Presumably, thepatients in the present study were sensitized by inhalation ordermal contact with dried marijuana, potentially bypassingdegradation by gastric fluid. Recently, several reports have sug-gested that RuBisCO can be an allergen in other plants.36e38
Another IgE-binding protein identified in this study was ATPsynthase. In one study, sequence homology was demonstratedbetween a bovine allergen and one of the subunits of ATPsynthase, which forms the basis for a potential cross-reactivity.39
Glyceraldehyde-3-phosphate dehydrogenase was also identifiedas one of the putative allergens of C sativa in our studies. Thisprotein has been identified as a major allergen of Triticum aestivum(wheat), several fungi, and a major allergen in rambutan(Nephelium lappaceum)einduced anaphylaxis.40e42 Several otherproteins identified in this analysis have also been reported as moldor pollen allergens in previous studies. Phosphoglycerate kinase isa characterized allergen of Candida albicans.43 BiP is a highlyconserved hazel pollen allergen (Cor a 10), which functions asa chaperone during protein synthesis and is reported to be a cross-reactive pollen allergen.44 BiP also shares a high degree of sequencehomology to heat shock protein 70, which has been reported as anallergen in many fungi.45,46 Oxygen-evolving enhancer protein wasa prominent allergen in our analysis; however, there are no studiesin the literature that currently report this protein to be an allergen.
Analysis of IgE reactivity on sodium periodateetreatedmembranes demonstrated an IgE response to carbohydrate deter-minants. It is possible that patients who tested positive during skinprick test analysis may have some IgE toward the glycosylatedmoieties on proteins as demonstrated by the Western blot analysis.

A.P. Nayak et al. / Ann Allergy Asthma Immunol 111 (2013) 32e37 37
To the best of our knowledge, no previous studies have examinedthis possibility in C sativa sensitization, and our studies suggest thatsome of the reactivity could be due to cross-reactivity carbohydratedeterminants. Further analysis is required to determine the identityand clinical relevance of these IgE-binding epitopes. It is not knownwhether these IgE reactive oligosaccharides are similar to othercross-reactive carbohydrate determinants that aremonovalent, andalthough they exhibit extensive cross-reactivity during analysis ofplant extracts in vitro, they have limited activity in SPT.47 Because oflimitations in the amount of samples, the cross-reactivity tocarbohydrates could not be further addressed in the current study.
In this study, sensitization to C sativa allergens has been char-acterized, and peptides from enzymes associated with theprimary metabolism of plants, such as RuBisCO, oxygen-evolvingenhancer protein, ATP synthase, phosphoglycerate kinase, andglyceraldehyde-3-phosphate dehydrogenase were identified asallergens. We also determined that IgE reactivity to carbohydratesalso exists. Because of the use of marijuana as a recreational drugand as an antiemetic in patients receiving chemotherapy, allergicsensitization to C sativa is expected to increase. Further researchis needed to understand the variability in underlying immuno-logic mechanisms and development of standardized reagents forclinical and diagnostic testing.
Supplementary Data
Supplementary data related to this article can be found at http://dx.doi.org/10.1016/j.anai.2013.04.018.
References
[1] National Institute on Drug Abuse. Marijuana abuse. In: Research Report Series.Bethesda, MD: National Institute on Drug Abuse; 2010.
[2] Perez JAJ. Allergic reaction associated with intravenous marijuana use. J EmergMed. 2000;18:260e261.
[3] Tessmer A, Berlin N, Sussman G, Chung EC, Beezhold D. Hypersensitivityreactions to marijuana. Ann Allergy Asthma Immunol. 2012;108:282e284.
[4] Perez-Bustamante S, Vazquez de la torre M, Villanueva A, Pelta R, Rubio M,Baeza M. Allergy to marijuana: case report. J Allergy Clin Immunol. 2007;119(suppl 1). S273-S273.
[5] Basharat P, Sussman G, Beezhold D, Leader N. Hypersensitivity reactions tomarijuana. J Allergy Clin Immunol. 2011;127(suppl 2). AB178-AB178.
[6] Stadtmauer G, Beyer K, Bardina L, Sicherer SH. Anaphylaxis to ingestion ofhempseed. J Allergy Clin Immunol. 2003;112:216e217.
[7] Stockli SS, Bircher AJ. Generalized pruritus in a patient sensitized to tobaccoand cannabis. J Dtsch Dermatol Ges. 2007;5:303e304.
[8] Stokes JR, Hartel R, Ford LB, Casale TB. Cannabis (hemp) positive skin tests andrespiratory symptoms. Ann Allergy Asthma Immunol. 2000;85:238e240.
[9] de Larramendi CH, Carnes J, Garcia-Abujeta JL, et al. Sensitization and allergyto Cannabis sativa leaves in a population of tomato (Lycopersicon esculentum)-sensitized patients. Int Arch Allergy Immunol. 2008;146:195e202.
[10] Mayoral M, Calderon H, Cano R, Lombardero M. Allergic rhinoconjunctivitiscaused by Cannabis sativa pollen. J Investig Allergol Clin Immunol. 2008;18:73e74.
[11] Liskow B, Liss JL, Parker CW. Allergy to marihuana. Ann Intern Med. 1971;75:571e573.
[12] Henderson RL, Tennant FS, Guerry R. Respiratory manifestations of hashishsmoking. Arch Otolaryngol. 1972;95:248e251.
[13] Zuskin E, Kanceljak B, Pokrajac D, Schachter EN, Witek TJ Jr. Respiratorysymptoms and lung function in hemp workers. Br J Ind Med. 1990;47:627e632.
[14] Williams C, Thompstone J, Wilkinson M. Work-related contact urticaria toCannabis sativa. Contact Dermatitis. 2008;58:62e63.
[15] Majmudar V, Azam NA, Finch T. Contact urticaria to Cannabis sativa. ContactDermatitis. 2006;54:127.
[16] Herzinger T, Schopf P, Przybilla B, Rueff F. IgE-mediated hypersensitivityreactions to cannabis in laboratory personnel. Int Arch Allergy Immunol. 2011;156:423e426.
[17] Gamboa P, Sanchez-Monge R, Luisa Sanz M, Palacin A, Salcedo G, Diaz-Perales A. Sensitization to Cannabis sativa caused by a novel allergenic lipidtransfer protein, Can s 3. J Allergy Clin Immunol. 2007;120:1459e1460.
[18] Armentia A, Castrodeza J, Ruiz-Munoz P, et al. Allergic hypersensitivity tocannabis in patients with allergy and illicit drug users. Allergol Immunopathol(Madr). 2011;39:271e279.
[19] Schmechel D, Simpson JP, Beezhold DH, Lewis DM. The development ofspecies-specific immunodiagnostics for Stachybotrys chartarum: the role ofcross-reactivity. J Immunol Methods. 2006;309:150e159.
[20] Shevchenko A, Tomas H, Havlis J, Olsen JV, Mann M. In-gel digestion for massspectrometric characterization of proteins and proteomes. Nat Protoc. 2006;1:2856e2860.
[21] van Bakel H, Stout JM, Cote AG, et al. The draft genome and transcriptome ofCannabis sativa. Genome Biol. 2011;12:R102.
[22] Hollister LE. Health aspects of cannabis. Pharmacol Rev. 1986;38:1e20.[23] Adlaf EM, Begin P, Sawka E. Canadian Addiction Survey (CAS): A National
Survey of Canadians’ Use of Alcohol and Other Drugs: Prevalence of Use andRelated Harms. Toronto, Ontario: Canadian Center on Substance Abuse; 2005.
[24] Tennant FS, Pendergast TJ. Medical manifestations associated with hashish.JAMA. 1971;216:1965e1969.
[25] Freeman GL. Allergic skin test reactivity to marijuana in the southwest. West JMed. 1983;138:829e831.
[26] Tanaka H, Degawa M, Kawata E, Hayashi J, Shoyama Y. Identification ofCannabis pollens using an allergic patient’s immunoglobulin E and purifica-tion and characterization of allergens in Cannabis pollens. Forensic Sci Int.1998;97:139e153.
[27] Salcedo G, Sanchez-Monge R, Diaz-Perales A, Garcia-Casado G, Barber D. Plantnon-sepcific lipid transfer proteins as food and pollen allergens. Clin ExpAllergy. 2004;34:1336e1341.
[28] Enrique E, Ahrazem O, Bartra J, et al. Lipid transfer protein is involved inrhinoconjunctivitis and asthma produced by rice inhalation. J Allergy ClinImmunol. 2005;116:926e928.
[29] Vrtala S, Ball T, Spitzauer S, et al. Immunization with purified natural andrecombinant allergens induces mouse IgG1 antibodies that recognize similarepitopes as human IgE and inhibit the human IgE-allergen interaction andallergen-induced basophil degranulation. J Immunol. 1998;160:6137e6144.
[30] Taylor SL. Protein allergenicity assessment of foods produced through agri-cultural biotechnology. Annu Rev Pharmacol Toxicol. 2002;42:99e112.
[31] Ellis RJ. The most abundant protein in the world. Trends Biochem Sci. 1979;4:241e244.
[32] Astwood JD, Leach JN, Fuchs RL. Stability of food allergens to digestionin vitro. Nat Biotechnol. 1996;14:1269e1273.
[33] Fu TJ, Abbott UR, Hatzos C. Digestibility of food allergens and nonallergenicproteins in simulated gastric fluid and stimulated intestinal fluid: a compar-ative study. J Agric Food Chem. 2002;50:7154e7160.
[34] Bowman CC, Selgrade MK. Differences in allergenic potential of food extractsfollowing oral exposure in mice reflect differences in digestibility: potentialapproaches to safety assessment. Toxicol Sci. 2008;102:100e109.
[35] Thomas K, Herouet C, Bannon G, et al. Evaluation of mouse models forassessing the allergenic potential of proteins. In: The Toxicologist. NewOrleans, LA: Society of Toxicology; 2005:1307.
[36] de Lacoste de Laval A, Ledent C, Mairesse M, Leduc V, Guerin L. Rubisco canact as a potent food and respiratory allergen. Paper presented at: EAACICongress; Vienna, Austria; June 10e14, 2006.
[37] Foti C, Damiani E, Zambonin CG, et al. Urticaria and angioedema to rubiscoallergen in spinach and tomato. Ann Allergy Asthma Immunol. 2012;108:60e61.
[38] Hoff M, Son DY, Gubesch M, et al. Serum testing of genetically modifiedsoybeans with special emphasis on potential allergenicity of the heterlogousprotein CP4 EPSPS. Mol Nutr Food Res. 2007;51:946e955.
[39] Parkkinen S, Rytkonen M, Pentikainen J, Virtanen T, Mantyjarvi R. Homologyof a bovine allergen and the oligomycin sensitivity-conferring protein of themitochondrial adenosine triphosphate synthase complex. J Allergy ClinImmunol. 1995;95:1255e1260.
[40] Sander I, Flagge A, Merget R, Halder TM, Meyer HE, Baur X. Identification ofwheat fluor allergens by means of 2-dimensional immunoblotting. J AllergyClin Immunol. 2001;107:907e913.
[41] Benndorf D, Muller A, Bock K, Manuwald O, Herbarth O, von Bergen M.Identification of spore allergens from the indoor mould Aspergillus versicolor.Allergy. 2008;63:454e460.
[42] Jirapongsananuruk O, Jirarattanasopa N, Pongpruska S, Vichyanond P,Piboonpocanun S. Glyceraldehyde-3-phosphate dehydrogenase as a majorallergen in rambutan-induced anaphylaxis. Ann Allergy Asthma Immunol.2011;106:545e546.
[43] Ishiguro A, Homma M, Torii S, Tanaka K. Identification of Candida albicansantigens reactive with immunoglobulin E antibody of human sera. InfectImmun. 1992;60:1550e1557.
[44] Gruehn S, Suphioglu C, O’Hehir RE, Volkmann D. Molecular cloning andcharacterization of hazel pollen protein (70 kD) as a luminal binding protein(BiP): a novel cross-reactive plant allergen. Int Arch Allergy Immunol. 2003;131:91e100.
[45] De Vouge MW, Thaker AJ, Zhang L, Muradia G, Rode H, Vijay HM. Molecularcloning of IgE-binding fragments of Alternaria alternata allergens. Int ArchAllergy Immunol. 1998;116:261e268.
[46] Andersson A, Rasool O, Schmidt M, et al. Cloning, expression and character-ization of two new IgE-binding proteins from the yeastMalassezia sympodialiswith sequence similarities to heat shock proteins and manganese superoxidedismutase. Eur J Biochem. 2004;271:1885e1894.
[47] Commins SP, Platts-Mills TAE. Allergenicity of carbohydrates and their role inanaphylactic events. Curr Allergy Asthma Rep. 2010;10:29e33.

eTable 1Proteomic analysis data for IgE-reactive spots to Cannabis sativa leaf extracta
Spot GenBankaccessionNo. (TSA)
Description MW Peptides Coverageamino acids, %)
Peptide(s) PeptideMW, Da
Signalpeptide
1 JP462165.1 ATP synthase subunit beta,chloroplastic Q95DR6.1(Agapanthus africanus)
53766 7 20.48 (R)INPTTSGSGVSTLEKK(N)(K)LSIFETGIK(V)(K)VVDLLAPYR(R)(K)TVLIMELINNIAK(A)(K)VALVYGQMNEPPGAR(M)(R)DVNEQDVLLFIDNIFR(F)(R)FVQAGSEVSALLGR(M)
1623.87771006.56991044.59681470.8481600.80321948.98951432.7675
No
JP454288.1 Ribulose-1,5- bisphosphatecarboxylase/oxygenaselarge subunit NP_084661.2(Oenothera elata subsp hookeri)
52783 9 27.78 (R)VTPQPGVPPEEAGAAVAAESSTGTWTTVWTDGLTSLDR(Y)
(R)IPTSYTK(T)(K)TFQGPPHGIQVER(D)(R)GGLDFTKDDENVNSQPFMR(W)(K)DDENVNSQPFMR(W)(R)DNGLLLHIHR(A)(R)DDFIEKDR(S)(R)VALEACVKAR(N)(K)WSPELAAACEVWK(E)
3853.8645808.4331464.74742168.97971450.61471186.65711036.48251115.61221545.7285
No
JP449419.1 Putative UDP-glucosyltransferaseXP_002513630.1 (Ricinus communis)
54238 1 5.49 (-)LMEGMKFLWRLHVAAAGLVSAAELLAR(-) 2952.6191 No
JP451360.1 Conserved hypothetical proteinXP_002521999.1 (Ricinus communis)
62831 1 1.60 (-)YTGPKEKDR(-) 1092.5564 No
JP473150.1 Predicted uncharacterized proteinXP_003518132.1 (Glycine max)
50615 1 2.66 (-)PELACYAAASPLWK(-) 1575.7755 No
NA Unmatched NA NA NA (-)SPPGDLHQKGDYGK(-)(-)NAPPHGLKLDR(-)(-)AHGRSVFYVHR(-)(-)AGQTPQEFRFPR(-)(-)RDRTDQKLNWFRLQQLLGNAAK(-)(-)LLRLEDAEQFEKEELRAAGLVSAAGRGRR(-)(-)ASVKVFKVK(-)(-)LSMLRAADKFLFKFDEAAAGLVKRHFSR(-)(-)LGSAMGLSAK(-)(-)GLALECSVKAR(-)(-)QGDGPKEFELGTR(-)(-)SSGSAAGTECLLSGSVKSGR(-)(-)SVNLHLDVLDGNEKFLGKAR(-)(-)SAGAEPVFLLTGGHEGPLVANTSLADYVPR(-)(-)TPSPELAAAPAGYWK(-)(-)LGLLHLHR(-)
1497.72131216.66761327.68981432.72112670.44683239.74851004.63833223.7437933.49541145.62271432.69471909.91632224.19653037.55051557.7828957.5872
NA
2 JP454288.1 Ribulose-1, 5-bisphosphatecarboxylase/oxygenaselarge subunit NP_084661.2(Oenothera elata subsp hookeri)
52783 5 12.63 (R)GGLDFTKDDENVNSQPFMR(W)(R)FIFCAEAIYK(A)(R)DNGLLLHIHR(A)(R)EITLGFVDLLR(D)(-)KGPPHGLKLDR(-)
2168.97971260.62131169.63041274.72351216.7040
No
NA Unmatched NA NA NA (-)FLEKQNKVLKTK(-)(-)ESTELMTLLSSRTAKK(-)(-)LLRAFAACDARDLLKVRAAAGLVCLLLLQK(-)(-)AVSTTFDRVNFVDSKGGRGR(-)(-)SRELNEKRARMK(-)(-)NWGPKEFKSKGR(-)(-)LTRALTFAERGPR(-)(-)RLWLEFLLLERGDAAKLVSAAAARRK(-)(-)FGRLWWFGAVFLMK(-)(-)KRQLLKLVRRLRRLQQLLVSAALEVLK(-)(-)SWPELAARGFLWK(-)
1474.88721793.95563223.87742168.10861516.82571432.75761486.83682952.71391756.92753240.08671559.8250
NA
3 JP454288.1 Ribulose-1,5- bisphosphatecarboxylase/oxygenaselarge subunit NP_084661.2(Oenothera elata subsp hookeri)
52783 3 15.36 (R)VTPQPGVPPEEAGAAVAAESSTGTWTTVWTDGLTSLDR(Y)
(R)GGLDFTKDDENVNSQPFMR(W)(R)VALEACVKAR(N)
3853.86452184.97441115.6122
No
NA Unmatched NA NA NA (-)FPHRRSNDPFTNSQKQAR(-)(-)SSGSAARMCLLSKHHLR(-)(-)LLARFAASVLGAAAAFLLRLFDKEDRFSR(-)(-)KERFEFKRRRHAAAAGLVSAALLLDK(-)(-)LFDDWDLLSSRGSKR(-)(-)LFQVSLNYLFGSVQVSGSVK(-)(-)SGGGGGGGLGSSKVR(-)(-)DGKGPKEFELSVK(-)(-)PASAFLKCLLK(-)(-)HVWKLDVLNWDKFLNRK(-)(-)PALPATSVKR(-)(-)KTLTLVSLPR(-)(-)TLLDLDKDK(-)
2185.08891909.97293223.79792952.68821793.90592171.16241231.62681432.75611189.68932210.21141038.61871126.70741059.5810
NA
A.P. Nayak et al. / Ann Allergy Asthma Immunol 111 (2013) 32e3737.e1

eTable 1 (continued )
Spot GenBankaccessionNo. (TSA)
Description MW Peptides Coverageamino acids, %)
Peptide(s) PeptideMW, Da
Signalpeptide
4 JP462165.1 ATP synthase b-subunit, chloroplasticQ95DR6.1 (Agapanthus africanus)
53766 4 11.44 (R)SAPAFIQLDTK(L)(K)VVDLLAPYR(R)(K)TVLIMELINNIAK(A)(R)DVNEQDVLLFIDNIFR(F)
1189.63431044.59681470.84801948.9895
No
JP454288.1 Ribulose-1,5-bisphosphatecarboxylase/oxygenase largesubunit NP_084661.2(Oenothera elata subsp hookeri)
52783 11 22.31 (R)LTYYTPEYQTK(D)(K)DTDILAAFR(V)(K)TFQGPPHGIQVER(D)(R)GGLDFTKDDENVNSQPFMR(W)(K)DDENVNSQPFMR(W)(R)EITLGFVDLLR(D)(R)EITLGFVDLLRDDFIEK(D)(R)DDFIEKDR(S)(R)VALEACVQAR(N)(K)WSPELAAACEIWK(E)(-)ANPPHGLQVER(-)
1405.67661020.5241464.74742168.97971450.61471274.72352022.06741036.48251115.57581559.74431216.6312
JP452260.1 Predicted probable aniontransporter 5 XP_002282945.1(Vitis vinifera)
48074 1 2.71 (-)NWGPQEFELGTR(-) 1432.6736 No
NA Unmatched NA NA NA (-)ASVKFRVAK(-)(-)PTLSAPLPTSY(-)(-)GAGNGPQEFELWK(-)(-)NENASCPVSTEDGYVWK(-)(-)FPQDNTLPAPKENSQPMVR(-)(-)QECEGTSLVSGWGSRK(-)(-)FLVLLQLDTK(-)(-)QLDPYKDHPVRPAFLQLDTK(-)(-)SLGEVDLQGEHR(-)(-)EGCEAASSLLSGWGSRK(-)(-)YWSCWNSFMSFSFDVCR(-)(-)LPSPRFAANSFVSAAPNKSR(-)(-)LMKVEAASVLGAAAAFLSAWSFESLEGLESR(-)(-)RDSLTSDYDRTLASLFAAAAGLVSAAPLESR(-)(-)GPEFLKASVLGAAAAFLSALTTEMSVESR(-)(-)PATMFLDSR(-)(-)VMQLAAPGVFFKPLCLLLLALGVSAAPELSR(-)
1004.61321145.59681431.67831954.83682168.06841779.8211188.71182380.25371338.65281793.83672220.88552116.11773239.6533223.6472952.52641036.50113223.8225
NA
5 JP451043.1 Predicted ribulose bisphosphatecarboxylase/oxygenase activase 2,chloroplastic isoform XP_002282236.1(Vitis vinifera)
51833 5 16.45 (K)GLVQDFSDDQQDITR(G)(K)VPLILGIWGGK(G)(R)VPIIVTGNDFSTLYAPLIR(D)(R)VYDDEVRK(W)(-)LLREYGNFLVQESNRVK(-)
1879.9011151.70672088.16191022.50322064.1116
No
JP450816.1 Putative plant glutamine synthetaseXP_002516801.1 (Ricinus communis)
47852 2 6.46 (K)IIAEYIWVGGSGIDLR(S)(K)AILNLSLR(H)
1760.9462898.5600
No
JP458176.1 Putative phosphoglycerate kinaseXP_002513353.1 (Ricinus communis)
50665 3 10.92 (K)LVASLPDGGVLLLENVR(F)(K)GVSLLLPSDVVIADK(F)(K)GVTTIIGGGDSVAAVEK(V)
1764.01441524.87631572.8359
No
JP481389.1 Predicted phosphoglycerate kinase,cytosolic-like XP_003546821.1(Glycine max)
42504 3 13.46 (K)LVASLPDGGVLLLENVR(F)(K)GVSLLLPSDVVIADK(F)(K)GVTTIIGGGDSVAAVEK(V)
1764.01441524.87631572.8359
No
JP452228.1 Glyceraldehyde-3-phosphatedehydrogenase B CBL43264.1(Solanum tuberosum)
48315 3 9.05 (K)YDSMLGTFK(A)(K)GISAEDVNAAFR(K)(K)VVAWYDNEWGYSQR(V)
1076.48471248.60991771.7954
No
JP473302.1 Camphor resistance CrcB-like proteinNP_565956.1 (Arabidopsis thaliana)
46943 1 2.33 (-)CGLGVLGLFK(-) 1005.5681 No
JP458088.1 Predicted ribulose bisphosphatecarboxylase/oxygenase activase 1,chloroplastic-like XP_002270571.2(Vitis vinifera)
48523 1 3.60 (-)LLREYGNFLVQESNRVK(-) 2064.1116 No
NA Unmatched NA NA NA (-)ANDPAPHLVLFDLFYEARK(-)(-)TPLSRGVETLGK(-)(-)KNDGLPWVASLPEAKGLDEQQDVK(-)(-)WTALYLGEAVALCQR(-)(-)YNFTLPNLK(-)(-)SLGTLLFPPSLPR(-)(-)APARTPDLDRRKESGNAWPAK(-)(-)HPQKRKDPSFPDVFAK(-)(-)YDDEKVR(-)(-)EWRCCLLSGLEFPK(-)(-)LNLFLLKYKRLGEARSDRR(-)(-)ERRRLDLLKTLSFELLAAGLVSAAAARQR(-)(-)YFCRLVVEELWK(-)(-)KLSFLELDRKLLLETLRRSVSAAARAQR(-)(-)TEYLFLNRQWER(-)
2215.14261256.7092636.34451692.86571108.59161396.80772335.21441896.0004923.43491793.85942347.363223.86281640.83853239.89431653.8263
NA
(continued on next page)
A.P. Nayak et al. / Ann Allergy Asthma Immunol 111 (2013) 32e37 37.e2

eTable 1 (continued )
Spot GenBankaccessionNo. (TSA)
Description MW Peptides Coverageamino acids, %)
Peptide(s) PeptideMW, Da
Signalpeptide
6 JP449733.1 Predicted calmodulin-like proteinXP_002279066.1 (Vitis vinifera)
17245 1 5.59 (-)SVSYYSWLK(-) 1131.5599 NA
NA Unmatched NA NA NA (-)QMTWASSLLSGVSTAKK(-)(-)SLKHGGGSTWRLARFLFLLGK(-)(-)QAYEDLAKK(-)(-)GNRLLQKVDLGFK(-)(-)RLWFRSFGAGAAYKK(-)(-)RFLALAASKNLLRLSRTTLGLVRGRKWK(-)(-)FQFSGALLMWK(-)(-)PANLQRYLLDLLMPGRPLLPRPLALENK(-)(K)GSCVILSTSASK(Y)
1793.93462343.33281064.55021486.8621756.95263223.96221326.6793210.84231208.607
NA
7 JP475070.1 23-kDa protein of oxygen evolvingsystem of photosystem II P16059(PSBP_PEA) (Pisum sativum)
28585 5 14.98 (-)ARLESSAPVLGGK(-)(-)EFPGQVLR(-)(-)QYYNISVLTR(-)(-)EREFPGQVLR(-)(-)TADGDEGGKHQLITATVK(-)
1283.7198944.44271255.62121229.49391853.7921
Yes
JP469123.1 Putative glycine-rich RNA-bindingprotein XP_002533374.1(Ricinus communis)
19941 1 6.59 (-)APQGFGFLSASK(-) 1208.6189 NA
NA Unmatched NA NA NA (-)LSSPAKSLSR(-)(-)VATVLALR(-)(-)YLLSVQLFQTYLLSVKEL(-)(-)NYLLLKLWNFTLAGK(-)(-)TALSSSFDRFRRNLLESRVVVKK(-)(-)LAATLTKK(-)(-)KDNFLNLVLETLKERRRLRSAALLLSR(-)(-)TQGKNLVPRTRRHLSASK(-)(-)PLGFNLNVRPTRRHLSASK(-)(-)LGKRLLFLRFTRWHASK(-)(-)PAGGFFQLSASK(-)
1044.5928841.53852156.21341793.0242707.5244844.53823223.8992162.21442162.21832215.28541208.6189
NA
8 JP474788.1 Conserved hypothetical proteinXP_002518775.1 (Ricinus communis)
21574 1 8.24 (-)SKTDFLPVPGTAGGSK(-) 1560.8147 NA
JP475070.1 23-kDa protein of oxygen evolvingsystem of photosystem II P16059(PSBP_PEA) (Pisum sativum)
28585 6 23.59 (K)QYYNISVLTR(T)(-)EREFPGQVLR(-)(-)ILESSAPVIGGK(-)(-)EFPGQVLR(-)(-)TADGDEGGKHQLITATVK(-)(-)TADGDEGGKHQLITATVKDGK(-)
1255.61061229.89151169.7180944.52491855.21182153.9761
Yes
NA Unmatched NA NA NA (-)LSSPATLKTK(-)(-)APANRRLVLDKLLTLHPAVK(-)(-)VATVALLR(-)(-)LSPKPLESLPR(-)(-)LWSELYLLSLLFLGK(-)(-)EYELKKKKK(-)(-)RRRDLRLLDESLQLESLLKVKK(-)(K)QFPYPKYEPVGVHDIMLLK(L)(-)AYWTALLLLFGK(-)(-)TVLDPAFLTVVK(-)(-)AYWTAFRKKPK(-)(-)SLLDPAFLTVVK(-)(-)PANEFCRSASK(-)(-)PANEKFMSASK(-)(-)LSFNLLLLLGFVVR(-)(-)DLRFRLLLGFVVR(-)
1044.61792224.3535841.53851235.72381794.03321192.7182707.61842273.19171394.79611301.75941394.69341301.71191208.57091208.57091602.83521602.8352
NA
9 NA Unmatched NA NA NA (-)LSSPATLKTK(-)(-)FQLSSQTLLSGLLLFK(-)(-)VATVLALR(-)(-)TSSFSKAFLLAR(-)(-)ALATLKTK(-)(-)RRLLRVLLSLALVFK(-)(-)SLVMPTTEEWFRSFLSASK(-)(-)NALKQHSVRFAPQGFLNRK(-)(-)PANEFCRSASK(-)(-)SLVMLWEEFWRSFLSASK(-)(-)SVSYYWLK(-)(-)NNDAQQMLLK(-)
1044.61791794.0292841.53851326.7296844.53811796.18762214.93652210.0861208.57092214.93651131.49571173.6466
NA
A.P. Nayak et al. / Ann Allergy Asthma Immunol 111 (2013) 32e3737.e3
eTable 1 (continued )
Spot GenBankaccessionNo. (TSA)
Description MW Peptides Coverageamino acids, %)
Peptide(s) PeptideMW, Da
Signalpeptide
10 NA Unmatched NA NA NA (-)LSSPAKSLSR(-)(-)AVRGFGSGGHLRLL(-)(-)KDTSDFTVALTK(-)(-)APQGRRLVLNFHGTLAAVYK(-)(-)APANRRLVLKELLTLHPAVK(-)(-)AAPLSLPR(-)(-)TFTLLSSGYKGLHRK(-)(-)KQSLSSVGVFKKK(-)(-)DRMLEFLLSVRLFR(-)
1044.59281438.81581324.68752210.24372238.3691823.49161706.94691434.8561793.9973
NA
Abbreviations: ATP, adenosine triphosphate; NA, not applicable; MW, molecular weight; TSA, transcriptome shotgun assembly.aPeptides identified from spots 1 through 10 in Figure 3A were analyzed against the Cannabis sativa database to determine sequence coverage. Information on homologousproteins in the database has been identified. No peptides were recovered for spots 11 and 12.
A.P. Nayak et al. / Ann Allergy Asthma Immunol 111 (2013) 32e37 37.e4





























