Embed Size (px)
Citation preview

Excitatory amino acids and neurodegeneration: a hypotheticalrole of calcium precipitation
M.J. Rodrõ guez, F. Bernal, N. Andre s, Y. Malpesa, N. Mahy*
Unitat de BioquõÂmica, IDIBAPS (Institut d'Investigacions BiomeÁdiques August Pi i Sunyer), Facultat de Medicina, Universitat de Barcelona,
c/Casanova 143, E-08036, Barcelona, Spain
Received 11 March 1999; accepted 31 August 1999
Abstract
Activation of excitatory amino acid (EAA) receptors can induce neurodegeneration by two major mechanisms ofexcitotoxicity, one related to the in¯ux of Na+, Clÿ and water, and the other to the increase in intracellular calcium
concentration ([Ca2+]i). Thus, acute microinjection of EAAs in several areas of the central nervous system (CNS) has been usedto produce neurodegenerative models. We studied the excitotoxic pattern associated with acute microinjection of AMPA in rathippocampus, medial septum-diagonal band of Broca (MS-DBB), prefrontal cortex and retina. In all cases progressive neuronalloss, glial reaction and development of intra- and extracellular calcium concretions were observed. However, a CNS-area
di�erential vulnerability was revealed, as shown by the speci®c atrophy of MS-DBB and its limited calci®cation. Whethercalcium deposits are a defensive mechanism against the massive increment of free cytoplasmatic calcium is discussed on the basisof ultrastructural data and previous results. 7 2000 ISDN. Published by Elsevier Science Ltd. All rights reserved.
Keywords: Calci®cation; Calcium homeostasis; Excitatory amino acids; Glial lactate; Neurodegeneration; Compensatory mechanisms
Excitatory amino acids (EAAs) account for most ofthe excitatory synaptic activity in the central nervoussystem (CNS), and they are released by an estimated40% of all synapses [8]. Moreover, they have beenimplicated in processes of learning, memory, synapticplasticity and neurotrophic activity [27]. EAA recep-tors have been classi®ed into three groups: two iono-tropic groups ÐN-methyl-D-aspartate (NMDA) and a-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid(AMPA)-kainate receptors-; and one group of metabo-tropic receptors, which are coupled to G proteins.
Although initially non-NMDA receptors do not alter
the permeability of the membrane to Ca2+, EAA
release in the synaptic cleft increases post-synaptic and
glial membrane permeability. This leads to a transient
increase in the intracellular calcium concentration
([Ca2+]i) in neurons and glial cells [34].
Excessive activation of EAA receptors can induce
the death of receptive neurons through a process
characterized by a chronic glutamate release and the
consequent massive increase of [Ca2+]i [4]. This pro-
cess, de®ned as excitotoxicity, may also involve in¯u-
xes of Na+ and Clÿ and e�ux of K+ which produces
cell swelling [3,13]. Intracellular precipitation of cal-
cium has been observed in several human pathological
situations such as birth anoxia, Down's syndrome,
Fahr's disease, Alzheimer's disease, presenile dementia,
malakoplakia, glioma or some in¯ammatory and infec-
tious diseases (for reviews see Harrington et al. [10],
and Ellie et al. [6]). In our laboratory, stereotaxic
microinjection of glutamate analogues, such as quis-
Int. J. Devl Neuroscience 18 (2000) 299±307
0736-5748/00/$20.00 7 2000 ISDN. Published by Elsevier Science Ltd. All rights reserved.
PII: S0736-5748(99 )00098 -2
www.elsevier.com/locate/ijdevneu
* Corresponding author. Tel.: +34-93-402-45-25; fax: +34-93-403-
58-82.
E-mail address: [email protected] (N. Mahy).
Abbreviations: AMPA, a-amino-3-hydroxy-5-methyl-4-isoxazole
propionic acid; [Ca2+]i, intracellular calcium concentration; CNS,
central nervous system; EAA, excitatory amino acid; GP, globus pal-
lidus; IBO, ibotenic acid; MS-DBB, medial septum-diagonal band of
Broca; NMDA, N-methyl-D-aspartate; QUIS, quisqualic acid; TEM,
transmission electron microscopy.

qualic acid (QUIS) and ibotenic acid (IBO) in rat glo-bus pallidus (GP) or medial septum-diagonal band ofBroca (MS-DBB), has been used to produce exper-imental models of neurodegeneration [23,24,26,39]. Inall cases, neurodegeneration in GP was associated withintracellular calcium precipitates, which were notobserved in the lesioned medial septum.
In order to better understand the mechanisms thatunderlie CNS di�erences in vulnerability to excitotoxi-city, this study was focused in four areas: prefrontalcortex, hippocampus, and retina, which have shown todevelop calci®cation in several human pathologies[5,16,25,30,33,44,46], and medial septum as negativecontrol of calcium precipitation. The aim of this paperwas to investigate if [Ca2+]i increase associated withlong-term AMPA-induced neurodegeneration leads tothe formation of intracellular calcium precipitates simi-lar to those observed in human CNS. Whether theseprecipitates are part of the neurodegenerative processor a neuroprotective compensatory mechanism of the[Ca2+]i increase is discussed in view of the data pre-sented in this paper and other studies.
1. Experimental procedures
1.1. Animals
Thirty-six adult male Sprague±Dawley rats (bodyweight 250±300 g at the beginning of the study) wereobtained from the animal housing facilities of theSchool of Medicine (Universitat de Barcelona). Theywere kept on a 12 h/12 h day/night cycle, and housedwith free access to food and water. Principles of lab-oratory animal care were followed since all animalswere manipulated according to the European legis-lation (86/609/EEC). Moreover, procedures wereapproved by the Ethic Committee of the Universitatde Barcelona, under supervision of the Generalitat deCatalunya.
1.2. Surgery
Under equithesin anesthesia, rats were placed in astereotaxic instrument with the incisor bar set atÿ3.3 mm. A 5.0 ml Hamilton syringe held by themicromanipulator of the stereotaxic instrument wasused for the injection in the four areas of the CNS:medial septum, prefrontal cortex, hippocampus, andeye vitreous cavity. AMPA (obtained from SigmaChemical Co., St. Louis, MO, USA) was dissolved inphosphate-bu�ered saline (PBS; 50 mM, pH 7.4), and0.5 ml was infused at di�erent concentrations in eachbrain area; 1 ml was infused in retina at a ¯ow rate of0.2 ml/min. In each group, three rats received a 50 mMPBS injection and were used as control (Table 1). Atthe indicated time post-lesion animals were anesthe-tized and decapitated for histological studies, exceptthree animals with hippocampal lesion, which weretranscardially perfused for ultrastructural studies.
1.3. Histological procedures
After rat decapitation, brains were quickly removedand frozen. Coronal 12 mm crystat sections from theinjection site were obtained, mounted on untreatedslices and stored at ÿ308C for histological study. Ret-inas were isolated from the eye cup, removed andmounted on concave glass slices. Calcium depositswere visualized by means of Alizarin red staining [28](1% in 50 mM Tris, pH 7.6) counterstained with FastGreen (0.25% in 50 mM Tris). Adjacent sections werestained with Cresyl violet for Nissl standard staining.
1.4. Ultrastructural procedures
Three of the rats injected in hippocampus wereanesthetized and transcardially perfused with a paraf-ormaldehyde±glutaraldehyde solution (2% and 2.5%,respectively, in 0.1 M PB, pH 7.4). The brains werequickly removed, cut into 50-mm-thick sections with aMcIlwain Mechanical Tissue Chopper (The MLE Co.Ltd, England, UK) and a micropunch from the calci-®ed area was obtained. In order to determine the
Table 1
Conditions of AMPA-treatmenta
Area Number of animals Coordinatesb AMPA dose Needle inclination Time post-lesion
Medial septum 4 0.7 r/1.1 l/5.0 v 10.6 nmol 108 60 days
Prefrontal cortex 3 3.2 r/3.8 l/4.2 v 2.7 nmol 08 60 days
Hippocampus 6 3.3 c/2.2 l/2.9 v 2.7 nmol 08 15 days
Vitreous liquid 4 ± 21 nmol 08 30 days
Vitreous liquid 4 ± 10.6+21 nmolc 08 3 days
a In each group, three rats were injected with 50 mM PBS and sacri®ced at the indicated time post-lesion of the group.b All coordinates were taken according to the Atlas of Paxinos and Watson [35]. Abbreviations: c=caudal, l=lateral, r=rostral, v=ventral.c The second dose was performed ten days after the ®rst one.
M.J. RodrõÂguez et al. / Int. J. Devl Neuroscience 18 (2000) 299±307300
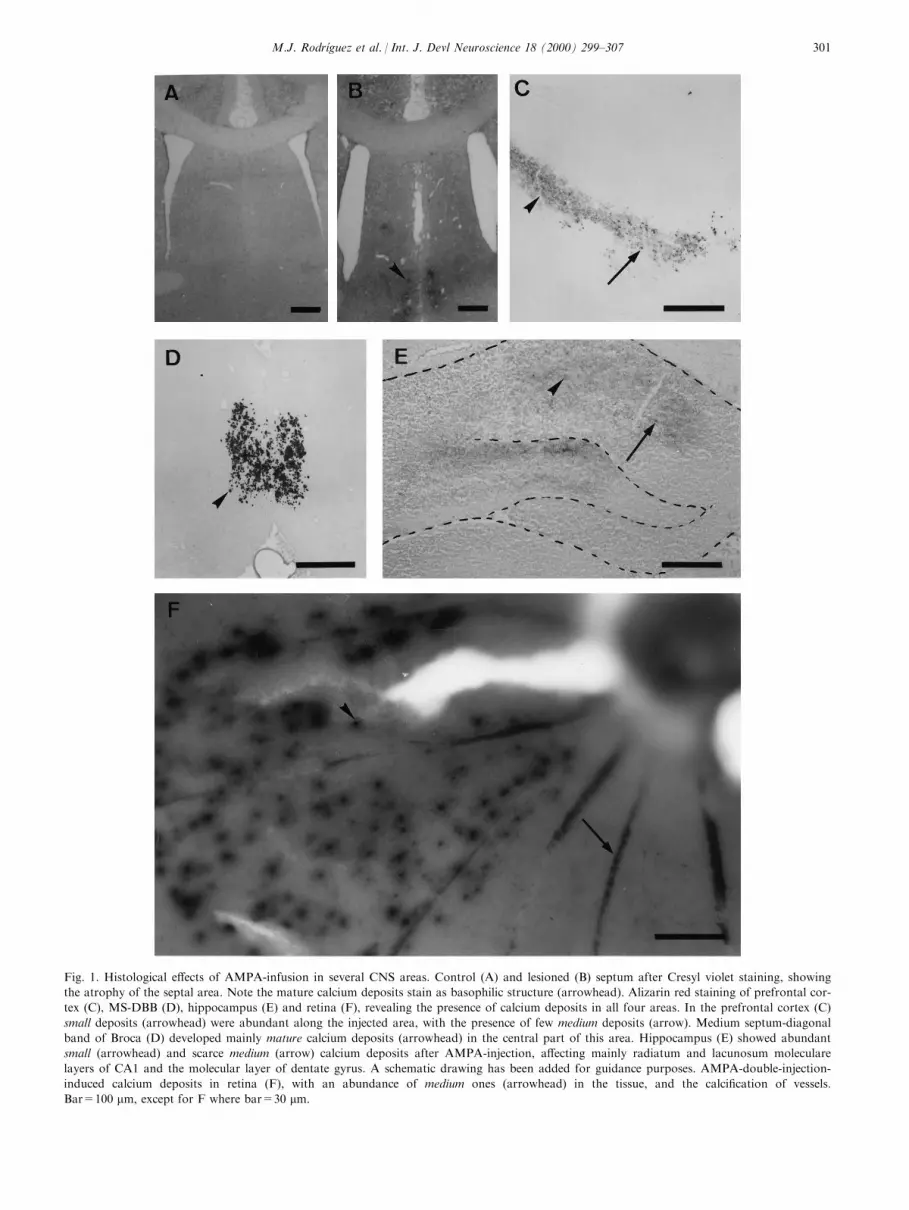
Fig. 1. Histological e�ects of AMPA-infusion in several CNS areas. Control (A) and lesioned (B) septum after Cresyl violet staining, showing
the atrophy of the septal area. Note the mature calcium deposits stain as basophilic structure (arrowhead). Alizarin red staining of prefrontal cor-
tex (C), MS-DBB (D), hippocampus (E) and retina (F), revealing the presence of calcium deposits in all four areas. In the prefrontal cortex (C)
small deposits (arrowhead) were abundant along the injected area, with the presence of few medium deposits (arrow). Medium septum-diagonal
band of Broca (D) developed mainly mature calcium deposits (arrowhead) in the central part of this area. Hippocampus (E) showed abundant
small (arrowhead) and scarce medium (arrow) calcium deposits after AMPA-injection, a�ecting mainly radiatum and lacunosum moleculare
layers of CA1 and the molecular layer of dentate gyrus. A schematic drawing has been added for guidance purposes. AMPA-double-injection-
induced calcium deposits in retina (F), with an abundance of medium ones (arrowhead) in the tissue, and the calci®cation of vessels.
Bar=100 mm, except for F where bar=30 mm.
M.J. RodrõÂguez et al. / Int. J. Devl Neuroscience 18 (2000) 299±307 301

deposit composition, half of the micropunches wereprocessed without post-®xation in 1% osmium tetrox-ide due to the overlap between osmium and phos-phorus X-ray spectra. The other half were processedby standard procedures for transmission electron mi-croscopy (TEM). In both situations, ultrathin sectionswere observed under a Phillips 301 TEM. At the sametime, X-ray spectra were acquired using an energy-dis-persive spectrometer (Princeton Gamma Tech, USA)attached to the electron microscope. The electron dif-fraction pattern was obtained from non-osmi®catedsamples using a Hitachi TEM 800MT at 75 and 100kV.
2. Results
2.1. Histological studies
In all the CNS areas examined, acute microinjectionof AMPA induced neurodegenerative damage, charac-terized by a neuronal loss, glial reaction and formationof calcium deposits. However, neurodegenerationappears to develop with some speci®city in these CNSareas, such as slight atrophy in the prefrontal cortexand signi®cant atropy (40%, P<0.05, Kruskal±Wallistest) in MS-DBB (Fig. 1A and 1B).
Alizarin red staining revealed calcium concretions inall AMPA-injected rats. Based on their size, rangingfrom 4 to 24 mm, these deposits were classi®ed in threegroups: large deposits (bigger than 10 mm), already vis-ible under the light microscope without staining, oftenshowed a dark core and were called `mature'. Smallerdeposits (5±10 mm) appeared in some areas; they werealso visible without any staining, but did not have adark core, and were called `medium' deposits. Finally,very small deposits (smaller than 5 mm) were detectedonly after Alizarin red staining. These deposits did nothave a dark core and were called `small'. In none ofthese brain areas calcium concretions were associatedto vessels; in contrast, abundant small deposits weredetected in the retina vessels.
After excitotoxic infusion in the prefrontal cortex,both medium and small calcium deposits were seenclose to the injection site (Fig. 1C). In the center of themedial septum, well delimited Alizarin red positivestaining showed mature calcium deposits (Fig. 1D),mainly localized where GABAergic cells were moreabundant. On the other hand, both small and mediumcalcium deposits were seen in all AMPA-lesioned hip-pocampi (Fig. 1E). Strong Alizarin red stainingextended from the injection site, covering more than ahalf of the hippocampus, and was especially abundantthroughout the radiatum and the lacunosum molecu-lare layers. Other sub®elds, such as the molecular layerof dentate gyrus, were also a�ected. Finally, in the ret-
ina no calcium precipitates were detected after a singleinjection of the highest dose of AMPA (data notshown). However, abundant medium concretions onthe retina surface and vessels were observed in all trea-ted rats 3 days after the second injection (Fig. 1F).
2.2. Ultrastructural studies
Fifteen days after AMPA injection, TEM analysisrevealed marked astroglial proliferation and extensivehypertrophy of the cell body and cytoplasmic pro-cesses, with dense bundles of intermediate ®lamentsand frequent intracytoplasmic deposits. These depositsranged from less than 0.5 to 10 mm in diameter, asmeasured directly on TEM photomicrographs. Theywere formed by numerous, small, needle-shaped crys-tals associated with a ®brilar matrix, and in close con-tact with cellular organelles such as microtubules,cisternae, vesicles or mitochondria, which presented nosign of ultrastructural degeneration (Fig. 2).
X-ray microanalysis of non-osmi®cated tissueshowed that intracytoplasmic deposits were formedessentially of calcium and phosphorus (Fig. 3), with amean Ca/P ratio of 1.320.2 (mean2SEM, n = 10).The four rings of di�erent intensity obtained by elec-tron di�raction of the particles (Fig. 4) indicated theircrystalline organization.
3. Discussion
The relationship between alteration of calciumhomeostasis and AMPA-induced neurodegeneration ofmedial septum, prefrontal cortex, hippocampus, andretina is evidenced here by the formation of calciumprecipitates in each of the lesioned areas.
The electron-di�raction ring pattern of the calciumdeposits was characteristic of a crystalline structuresimilar to apatites [11,15]. Moreover, the Ca/P ratio of1.32 0.2 of cytoplasmic deposits from TEM analysis,lower than the theoretical apatite value of 1.67, is con-sistent with experimental data obtained from otherbiological crystals which do not have an ideal organiz-ation [2]. Taken together, these data strongly indicatethat these deposits correspond to biological hydroxya-patites similar to those observed in rat GP [24] and inseveral human tissues [12,14,17].
Calcium precipitation is part of the on-going degen-erative process of the rat GP induced by an acutemicroinjection of AMPA, IBO or QUIS [23,36,39] andmay be similar to that associated with human basalganglia calci®cation [24]. As these deposits are alsoobserved in other rat brain areas after kainate, ibotenicacid or NMDA microinjection [18,31,37], their for-mation does not depend on the glutamate receptorsubtype initially stimulated. However, their size, num-
M.J. RodrõÂguez et al. / Int. J. Devl Neuroscience 18 (2000) 299±307302

ber and distribution vary not only with the excitotoxinbut also with the CNS area concerned. Thus, in MS-DBB, the on-going degenerative process associatedwith IBO and QUIS microinjection was characterizedby signi®cant atrophy and no calci®cation, whereas insimilar conditions, AMPA resulted in a similar atro-phy and calcium deposits localized at the site of injec-tion. Sensibility to AMPA-induced calci®cation wasdi�erent amongst the four studied CNS areas: in pre-frontal cortex and hippocampus, calcium deposits wereobserved after a dose of 2.7 nmol whereas in MS-DBB10.6 nmol were necessary; in retina only an injectionof 10.6 nmol followed by another one of 21 nmol ledto calcium concretions. In this last structure a singledose of 21 nmol AMPA resulted in a lesion devoid ofcalci®cation and, as demonstrated in chick retina [3],Clÿ movements might be responsible for this degenera-tive process. The calci®cation observed at the samedose following AMPA pretreatment reveals the acti-vation of a new Ca2+-dependent mechanism. Thus,the rat, in spite of being a dark-adapted animal, mightbe very helpful to understand the molecular basis ofhuman retinal calci®cation.
The permeability of AMPA receptors to Ca2+ isdetermined by the receptor subunit composition. The
GluR1 and GluR3 subunits can form channels per-meable to calcium, whereas the GluR2 subunit in thereceptor assembly prevents calcium permeability [9]. Inaddition, activation of metabotropic glutamate recep-tor II and III has shown to be neuroprotective againstexcitotoxicity [1]. Thus, together with variations incytoplasmic Ca2+ bu�ers and/or Ca2+ clearance sys-tems, which regulate altogether the [Ca2+]i increase[19,29], the glutamate-receptor-subtype compositionand distribution in each CNS area [34] may explainthis di�ering vulnerability.
On the other hand, glutamate CNS activation heigh-tened the brain energy needs. Many studies suggestthat glutamate stimulates glial glycolytic ¯ux, releasinglactate as aerobic energy substrate for neurons[20,21,22]. Similar results have been obtained usingglutamate-stimulated hippocampal slices, showing thatthe higher the level of lactate produced by glia, thehigher the concentration of glutamate that slices wouldtolerate, even at concentrations considered excitotoxicfor the cell [41]. Glial lactate also increases duringhuman hypoxia and is used as an obligatory aerobicneuronal energy substrate, even after hypoxia, toensure the preservation of neuronal viability [42]. Inview of these data, in all the four AMPA-lesioned
Fig. 2. Hippocampal intracellular calcium deposit by TEM. A reactive astrocyte showing an incipient calcium concretion into its cytoplasm.
Note the normal morphology of mitochondria (asterisk), which are in close contact with the deposit via ®laments (arrowhead). Bar=0.5 mm.
M.J. RodrõÂguez et al. / Int. J. Devl Neuroscience 18 (2000) 299±307 303

areas of this study, gliosis must be associated with anincrease in lactate, rapidly transported into neuronsfor its complete oxidation. In these cells, pH reduction,associated with the increased lactate concentration, fa-cilitates the solubility of Ca2+ and the formation ofH2PO
ÿ4 , HPO2ÿ
4 and PO3ÿ4 ions from inorganic phos-
phate [7]. Because of the very high Ca2+/H2POÿ4 ,
HPO2ÿ4 , PO3ÿ
4 a�nity, apatite nucleation may occurwith the subsequent growth of the crystalline for-mation along with the neurodegenerative process.After cell death, these deposits will be extracellular.
Thus, calci®cation of each lesioned area would dependnot only on the density and subtype of glutamatereceptors and calcium movements, but also on thedi�erential capacity of glial cells to release lactateduring degeneration. The lack of astrogliosis observedin MS-DBB after any excitotoxic lesion may explain,at least in part, the reduced calci®cation observed inthis area together with the important neuronal loss[23,29].
Human CNS calci®cation has frequently beenobserved in a wide range of diseases without any clear
Fig. 3. X-ray spectrum microanalysis of an hippocampal intracellular calcium deposit, showing an abundance of Ca and P, standing out against
the other elements. Copper and chlorine components are from the copper grid and the composition of the embedding resin, respectively. Note
the Ca/P ratio near to 1.6, characteristic of hydroxyapatites.
Fig. 4. Crystalline organization of calcium precipitates. (A) TEM image of a non-osmi®cated deposit, showing the needle-shaped crystals.
Bar=0.2 mm. (B) Electron-di�raction image, with a clear four-ring pattern (arrowheads), similar to that of hydroxyapatite.
M.J. RodrõÂguez et al. / Int. J. Devl Neuroscience 18 (2000) 299±307304

relationship between them [6,10]. However, in agree-ment with previous studies [18,23,24,32,39], our dataindicate that it may be related to glutamate receptoroverstimulation.
Although the signi®cance of cellular calci®cation isunknown, a number of points suggest that it may bepart of the compensatory mechanisms for excitotoxicneurodegeneration. In this paper, the fact that mito-chondria close to calcium concretions appear normalat the EM level argues in favor of the above hypoth-esis, despite the fact that mitochondrial dysfunctionconstitute a primary event in NMDA-induced degener-ation in cultured hippocampal neurons [40]. It has alsobeen described that very high levels of cytoplasmic cal-cium are not necessarily neurotoxic, and that an e�ec-tive uptake of calcium into mitochondria is required totrigger NMDA-receptor-stimulated neuronal death[43]. These data would indicate that, besides calcium,other causes such as Na+ and Clÿ in¯ux and K+
e�ux may induce neuronal damage.Furthermore, this hypothesis is consistent with the
fact that neurons undergoing prolonged stimulation
of NMDA receptors can survive in the presence of
[Ca2+]i chelators [45]. Thus, cytoplasmic precipitation
of calcium would mimick the decrease in [Ca2+]iinduced by these chelators: in rat GP, the AMPA
dose-response study between 0.54 and 21.6 mM has
evidenced that an increase in calci®cation is not ac-
companied by an increase in astrogliosis [36]; in similar
experiments, rat MS-DBB led to septal atrophy at the
low AMPA concentrations (0.54 mM, 2.7 mM and
5.4 mM) whereas concentrations higher than 5.4 mM
resulted in increasing calci®cation without any modi®-
cation of the atrophy [38]; ®nally, in hippocampus, the
selective adenosine-A2a-receptor antagonist 8-(3-chlor-
ostyryl)-ca�eine increased the NMDA-induced neur-
onal loss whilst calci®cation was decreased [37].
And, even if all these data support the neuroprotec-
tive hypothesis, further experiments are necessary to
demonstrate whether, as proposed in Fig. 5, calci®ca-
tion constitutes an optional step of the calcium homeo-
stasis. This new possibility would help compensate the
massive increment of free cytoplasmic ions associated
Fig. 5. Schematic drawing of the excitotoxic process induced by glutamate with the intracellular formation of precipitates as part of the calcium
homeostasis. The metabolic pathway of lactate with the communication between the glial and neuronal compartment is included in the diagram.
ADP; adenosine diphosphate; AMPA, AMPA receptor; ATP; adenosine trisphosphate; CBPs, calcium binding proteins; Glu, glutamate; Gln,
glutamine; IP3, inositol trisphosphate; NMDA, NMDA receptor; Pi, inorganic phosphate.
M.J. RodrõÂguez et al. / Int. J. Devl Neuroscience 18 (2000) 299±307 305

with the on-going degenerative process and the lactate-induced acidosis.
Acknowledgements
The authors wish to thank Dr Alberto Prats fromthe Department d'Anatomia and Mrs Mo nica Marti-nez for their scienti®c collaboration, and Mrs Nu riaCortadellas and Joaquim Portillo from the ServeisCienti®co-Te cnics of the Universitat de Barcelona fortheir technical help in the ultrastructural studies. Thiswork was supported by QuõÂmica Farmace utica BayerS.A. (Spain) and CIRIT PIC 96-98.
References
[1] Battaglia, G., Bruno, V., Ngomba, R. T., Di Grecia, R.,
Copani, A. and Nicoletti, F., Selective activation of group-II
metabotropic glutamate receptors is protective against excito-
toxic neuronal death. Eur. J. Pharmacol., 1998, 356, 271±274.
[2] Bloss, F. D., Crystallography and Crystal Chemistry. An
Introduction. Hold, Reinhart and Winston Inc, New York,
1971.
[3] Chen, Q., Olney, J., Lukasiewicz, P., Almli, T. and Romano,
C., Ca2+-independent excitotoxic neurodegeneration in isolated
retina, an intact neural net: a role for Clÿ and inhibitory trans-
mitters. Mol. Pharmacol., 1998, 53, 564±572.
[4] Choi, D., Calcium-mediated neurotoxicity: relationship to
speci®c channels types and roles in ischemic damage. Trends
Neurosci., 1988, 11, 465±469.
[5] Dowe, D. A., Heitzman, E. R. and Larkin, J. J., Human immu-
node®ciency virus infection in children. Clin. Imaging, 1992, 16,
145±151.
[6] Ellie, E., Julien, J. and Ferrer, X., Familial idiopathic striopalli-
dodentate calci®cations. Neurology, 1989, 39, 381±385.
[7] Farooqui, A. A., Haun, S. E. and Horrocks, L. A., Ischemia
and hypoxia. In Basic Neurochemistry, eds G. Siegel, B. W.
Agrano�, R. W. Alberts and P. B. Molino�. Raven Press, New
York, 1994, pp. 867±883.
[8] Fonnum, F., Glutamate: a neurotransmitter in mammalian
brain. J. Neurochem., 1984, 42, 1±11.
[9] Greenamyre, J. T. and Porter, R. H., Anatomy and physiology
of glutamate in the CNS. Neurology, 1994, 44, 7±13.
[10] Harrington, M. G., MacPherson, P., McIntosh, W. B., Allam,
B. F. and Bone, I., The signi®cance of the incidental ®nding of
basal ganglia calci®cation on computed tomography. J. Neurol.
Neurosurg. Psychiatry, 1981, 44, 1168±1170.
[11] Herrmann, G., StuÈ nitz, H. and Nitsch, C., Composition of ibo-
tenic acid-induced calci®cations in rat substantia nigra. Brain
Res., 1998, 786, 205±214.
[12] Honda E., Aoki M., Brunno M. and Ito A., Light and electron
microscopic study on surface and internal structure of human
brain stones with reference to some natural minerals. Bulletin de
l'Institut Oceanographique (Monaco), Spec. Iss. 14, 1994, 115±
120.
[13] Katayama, Y., Maeda, T., Koshinaga, M., Kawamata, T. and
Tsubokawa, T., Role of excitatory amino acid-mediated ionic
¯uxes in traumatic brain injury. Brain Pathology, 1995, 5, 427±
435.
[14] Khan, S. R. and Hackett, R. L., Identi®cation of urinary stone
and sediment crystals by scanning electron microscopy and X-
ray microanalysis. J. Urol., 1986, 135, 818±825.
[15] Kim, K. M., Apoptosis and calci®cation. Scanning Micosc.,
1995, 9, 1137±1178.
[16] Kobari, M., Nagawa, S., Sugimoto, Y. and Fukuuchi, Y.,
Familial idiopathic brain calci®cation with autosomal dominant
inheritance. Neurology, 1997, 48, 645±649.
[17] Kodaka, T., Mori, R., Debari, K. and Yamada, M., Scanning
electron microscopy and electron probe microanalysis studies of
human pineal concretions. J. Electron Microsc., 1994, 43, 307±
317.
[18] Korf, J. and Postema, F., Regional calcium accumulation and
cation shifts in rat brain by kainate. J. Neurochem., 1984, 43,
1052±1060.
[19] Kostyuk, P. and Verkhratsky, A., Calcium stores in neurons
and glia. Neuroscience, 1994, 63, 381±404.
[20] Larrabee, M. G., Lactate metabolism and its e�ects on glucose
metabolism in an excised neural tissue. J. Neurochem., 1995, 64,
1734±1741.
[21] Larrabee, M. G., Partitioning of CO2 production between glu-
cose and lactate in excised sympathetic ganglia, with impli-
cations for brain. J. Neurochem., 1996, 67, 1726±1734.
[22] Magistretti, P. J., Pellerin, L. and Martin, J. L., Brain energy
metabolism: an integrated cellular perspective. In
Psychopharmacology: the Fourth Generation of Progress, eds F.
E. Boom and D. J. Kupfer. Raven, New York, 1995, pp. 657±
670.
[23] Mahy, N., Bendahan, G., Boatell, M. L., Bjelke, B., Tinner, B.,
Olson, L. and Fuxe, K., Di�erential brain area vulnerability to
long-term subcortical excitotoxic lesions. Neuroscience, 1995, 65,
15±25.
[24] Mahy N., Prats A., Riveros A., Andre s N. and Bernal F., Basal
ganglia calci®cation induced by excitotoxicity: an experimental
model characterised by electron microscopy and X-ray microa-
nalysis. Acta Neuropathol., 1999, 98, 217±225.
[25] Mann, D., The pathological association between Down syn-
drome and Alzheimer disease. Mech. Ageing Dev., 1988, 43, 99±
136.
[26] McAlonan, G. M., Dawson, G. R., Wilkinson, L. O., Robbins,
T. W. and Everitt, B. J., The e�ects of AMPA-induced lesions
of the medial septum and vertical limb nucleus of the diagonal
band of Broca on spatial delayed non-matching to sample and
spatial learning in the water maze. Eur. J. Neurosci., 1995, 7,
1034±1049.
[27] McDonald, J. W. and Johnston, M. V., Physiological and
pathophysiological roles of excitatory amino acids during cen-
tral nervous system development. Brain Res. Rev., 1990, 15, 41±
70.
[28] McGee-Russel, S. M., Histological methods for calcium. J.
Histochem. Cytochem., 1958, 6, 22.
[29] Meyer, F. B., Calcium, neuronal hyperexcitability and ischemic
injury. Brain Res. Rev., 1989, 14, 227±243.
[30] Nagaratnam, N. and Plew, J. D., Extensive intracranial calci®-
cation secondary to hypoxia, presenting with dyspraxic gait.
Australas. Radiol., 1998, 42, 232±233.
[31] Nitsch, C. and Schaefer, F., Calcium deposits develop in rat
substantia nigra but not striatum several weeks after local ibote-
nic acid injection. Brain Res., 1990, 25, 769±773.
[32] Nitsch, C. and Scotti, A., Ibotenic acid-induced calcium depos-
its in rat substantia nigra. Ultrastructure of their time-depen-
dent formation. Acta Neuropathol., 1992, 85, 55±70.
[33] Ossoinig, K. C., Cennamo, G., Green, R. L. and Weyer, N. L.,
Echographic results in the diagnosis of retinoblastoma. Doc.
Ophtalmol. Proc. Series, 1981, 29, 103±107.
[34] Ozawa, S., Kamiya, H. and Tsuzuki, K., Glutamate receptors
in the mammalian central nervous system. Prog. Neurobiol.,
1998, 54, 581±618.
M.J. RodrõÂguez et al. / Int. J. Devl Neuroscience 18 (2000) 299±307306

[35] Paxinos, G. and Watson, C., The Rat Brain in Stereotaxic
Coordinates, 2nd ed. Academic Press, Sydney, 1986.
[36] Petegnief, V., Saura, J., Dewar, D., Cummins, D. J., Dragunow,
M., Mahy, N., Long-term e�ects of a-amino-3-hydroxy-5-
methyl-4-isoxazole propionate and 6-nitro-7-sulphamoylben-
zo(f)quinoxaline-2,3-dione in the rat basal ganglia: calci®cation,
changes in glutamate receptors and glial reactions. Neuroscience,
1999, 94, 105±115.
[37] Robledo, P., Ursu, G., Mahy, N., E�ects of adenosine and
gamma-aminobutiric acid A receptor antagonists on N-methyl-
D-aspartate induced neurotoxicity in the rat hippocampus.
Hippocampus, 1999, 9, 527±533.
[38] Rodrõ guez, M. J., Andre s, N., Malpesa, Y. and Mahy, N.,
Long-term excitotoxic lesion induced by AMPA injection in the
rat medial septum: a dose response study. Eur. J. Neurosci.,
1998, 10, 442.
[39] Saura, J., Boatell, M. L., Bendahan, G. and Mahy, N., Calcium
deposits formation and glial reaction in rat brain after ibotenic
acid-induced basal forebrain lesions. Eur. J. Neurosci., 1995, 7,
1569±1578.
[40] Schinder, A. F., Olson, E. C., Spitzer, N. C. and Montal, M.,
Mitochondrial dysfunction is a primary event in glutamate neu-
rotoxicity. J. Neurosci., 1996, 16, 6125±6133.
[41] Schurr, A., Miller, J. J., Payne, R. S. and Rigor, B. M., An
increase in lactate output by brain tissue serves to meet the
energy needs of glutamate-activated neurons. J. Neurosci., 1999,
19, 34±39.
[42] Sibson, N. R., Dhankhar, A., Mason, G. F., Rothman, D. L.,
Behar, K. L. and Schulman, R. G., Stoichiometric coupling of
brain glucose metabolism and glutamatergic neuronal activity.
Proc. Natl. Acad. Sci. USA, 1998, 95, 316±321.
[43] Stout, A. K., Raphael, H. M., Kanterewicz, B. I., Klann, E.
and Reynolds, J. J., Glutamate-induced neuron death requires
mitochondrial calcium uptake. Nat. Neurosci., 1998, 1, 366±373.
[44] Toti, P., Balestri, P., Cano, M., Galluzzi, P., Megha, T.,
Farnetani, M. A., Palmeri, M. L., Vascotto, M., Venturi, C.
and Fois, A., Celica disease with cerebral calcium and silica
deposits: X-ray spectroscopic ®ndings, an autopsy study.
Neurology, 1996, 46, 1088±1092.
[45] Tymianski, M., Wallace, M. C., Spigelman, I., Uno, M.,
Carlen, P. L., Tator, C. H. and Charlton, M. P., Cell-permeant
Ca2+ chelators reduce early excitotoxic and ischemic neuronal
injury in vitro and in vivo. Neuron, 1993, 11, 221±235.
[46] Wodarz, N., Becker, T. and Deckert, J., Musical hallucinations
associated with post-thyroidectomy hypoparathyroidism and
symmetric basal ganglia calci®cations. J. Neurol. Neurosurg.
Psychiatry, 1995, 58, 763±764.
M.J. RodrõÂguez et al. / Int. J. Devl Neuroscience 18 (2000) 299±307 307





























