Embed Size (px)
Citation preview

(This is a sample cover image for this issue. The actual cover is not yet available at this time.)
This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Review
Regulation of VEGF signaling by membrane traffic
Arie Horowitz a,b,⁎, Himabindu Reddy Seerapu a
a Department of Molecular Cardiology, Lerner Research Institute, Cleveland Clinic Lerner College of Medicine, 9500 Euclid Avenue, Cleveland, OH 44195, USAb Department of Physiology and Biophysics, Case Western Reserve University School of Medicine, 10900 Euclid Avenue, Cleveland, OH 44106, USA
a b s t r a c ta r t i c l e i n f o
Article history:Received 23 April 2012Accepted 9 May 2012Available online 19 May 2012
Keywords:AngiogenesisEndocytosisMembrane trafficNeuropilin-1Rho GTPaseVEGFR2
Recent findings have drawn attention to the role of membrane traffic in the signaling of vascular endothelialgrowth factor (VEGF). The significance of this development stems from the pivotal function of VEGF invasculogenesis and angiogenesis. The outline of the regulation of VEGF receptor (VEGFR) signaling by mem-brane traffic is similar to that of the epidermal growth factor receptor (EGFR), a prototype of the intertwiningbetween membrane traffic and signaling. There are, however, unique features in VEGFR signaling that areconferred in part by the involvement of the co-receptor neuropilin (Nrp). Nrp1 and VEGFR2 are integratedinto membrane traffic through the adaptor protein synectin, which recruits myosin VI, a molecular motorthat drives inward trafficking [17,21,64]. The recent detection of only mild vascular defects in a knockinmouse model that expresses Nrp1 lacking a cytoplasmic domain [104], questions the co-receptor's role inVEGF signaling and membrane traffic. The regulation of endocytosis by ephrin-B2 is another feature uniqueto VEGR2/3 [18,19], but it awaits a mechanistic explanation. Current models do not fully explain how mem-brane traffic bridges between VEGFR and the downstream effectors that produce its functional outcome, suchas cell migration. VEGF-A appears to accomplish this task in part by recruiting endocytic vesicles carryingRhoA to internalized active VEGFR2 [58].
© 2012 Elsevier Inc. All rights reserved.
Contents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18111.1. Overview: regulation of receptor tyrosine kinase (RTK) signaling by membrane traffic . . . . . . . . . . . . . . . . . . . . . . . 18111.2. EGFR as a prototype for the regulation of RTK signaling by membrane traffic . . . . . . . . . . . . . . . . . . . . . . . . . . . 1811
2. Endocytosis and trafficking of VEGFR . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18122.1. Endocytic pathway of VEGFR . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18122.2. Regulation of VEGFR2 endocytosis and sorting by ubiquitination . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18142.3. Regulation of VEGFR endocytosis by ephrin . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1814
3. Endocytosis and trafficking of Nrp1 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18143.1. Association between Nrp1 and VEGFR2 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18143.2. Endocytic pathway of Nrp1 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18143.3. Role of Nrp1 in VEGF signaling . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1815
4. Endocytosis of adhesion receptors . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18154.1. Endocytosis of intercellular adhesion receptors . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18154.2. Endocytosis of extracellular matrix (ECM) receptors . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1816
Cellular Signalling 24 (2012) 1810–1820
Abbreviations: Actn4, actinin-4; Amot, angiomotin; APPL, adaptor protein phospho-tyrosine interaction, PH domain, and leucine zipper containing; Cbl, Casitas B-lineage lym-phoma; EC, endothelial cell; EGFR, epithelial growth factor receptor; ECM, extracellular matrix; EPS15, epidermal growth factor receptor substrate 15; FGFR, fibroblast growth factorreceptor; FAK, focal-adhesion kinase; GIPC, GAIP-interacting protein, C-terminus; GPI, glycosyl phosphatidylinositol; Grb2, growth factor receptor-bound protein 2; HECT, E3 ho-mologous to E6-AP C terminus; Hrs, hepatocyte growth factor-regulated tyrosine kinase substrate; JAM, junctional adhesion molecule; KI, knockin; LRP, lipoprotein receptor-relatedprotein; NIP, neuropilin-1 binding protein; Mical-l2, molecule interacting with CasL-like 2; MUPP1, multi PDZ domain protein 1; Nrp, neuropilin; PAK, p21-activated kinase; PDGFR,platelet derived growth factor receptor; PDZRN3, PDZ domain-containing RING finger protein 3; PEST, rich in proline [P], glutamic acid [E], serine [S], and threonine [T]; PAI,plasminogen activator inhibitor; PTP1b, protein tyrosine phosphatase 1b; PKD, protein kinase D; PKC, protein kinase C; RTK, receptor tyrosine kinase; SNARE, N-ethylmaleimide‐sensitive factor attachment protein receptor; SH, Src homology; Shc, Src homology and collagen homology; TRCP, β-transducin repeats-containing protein;uPAR, urokinase plasmin activator receptor; VEcad, vascular endothelial cadherin; VEGFR, vascular endothelial growth factor receptor; YFP, yellow fluorescent protein.⁎ Corresponding author at: Department of Molecular Cardiology, Lerner Research Institute, Cleveland Clinic Lerner College of Medicine, 9500 Euclid Avenue, Cleveland, OH 44195,
USA. Tel.: +1 216 636 9461; fax: +1 216 445 8204.E-mail address: [email protected] (A. Horowitz).
0898-6568/$ – see front matter © 2012 Elsevier Inc. All rights reserved.doi:10.1016/j.cellsig.2012.05.007
Contents lists available at SciVerse ScienceDirect
Cellular Signalling
j ourna l homepage: www.e lsev ie r .com/ locate /ce l l s ig

Author's personal copy
5. Endocytosis and trafficking of Rho GTPases . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18176. Concluding remarks . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1818References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1818
1. Introduction
1.1. Overview: regulation of receptor tyrosine kinase (RTK) signaling bymembrane traffic
Membrane traffic has been recognized for some time now as anessential component of RTK signaling in general [1,2], and particularlyin the vascular system, [3,4]. That recognition represents a conceptualdeparture from the previously prevailing view where the sole effectof endocytosis was thought to be the attenuation of cell surface recep-tors, including that of RTKs [5,6]. To date, endocytosis and membranetraffic have been shown to regulate the signaling pathways of multi-ple RTKs and their co-receptors. These include EGFR [6–8], the plate-let derived growth factor receptor (PDGFR) [9–11], the fibroblastgrowth factor receptor (FGFR) [12,13] and its co-receptor syndecan-4 [14], VEGFR [15–19] and its co-receptor Nrp [20–22], the insulinand insulin-like growth factor receptors [23–26], the hepatocytegrowth factor Met [27], the Eph receptors [28,29], and TrkA, thenerve growth factor receptor [30]. All the RTKs that are internalizedafter binding their soluble ligands undergo primarily clathrin-mediated endocytosis, and their trafficking is regulated by the sameadaptor proteins and small GTPases [3,4]. At the same time, the mo-lecular pathways that intertwine membrane traffic and their down-stream phosphorylation cascades appear to differ among RTK types,possibly reflecting the differences between their physiological func-tions. The mechanism and functional role of membrane traffic isbest understood in the signaling pathway of EGFR, and to a lesser de-gree in VEGFR signaling [31,32]. This review focuses solely on theendocytic pathways that are activated downstream of VEGF in endo-thelial cells (ECs), in order to discuss the state of knowledge and theimpact of this relatively recent aspect of VEGF signaling on the under-standing of angiogenesis.
1.2. EGFR as a prototype for the regulation of RTK signaling by membranetraffic
Though the reciprocal relation between EGFR signaling and mem-brane traffic has been described in a detailed and up-to-date mannerelsewhere [31,33], it is worth recapitulating its main components(Fig. 1) because it is known in more detail than that of other RTKs.EGFR undergoes mainly clathrin-dependent endocytosis that is trig-gered by the binding of its ligand [34]. EGFR dimerization and tyrosinephosphorylation were required for its recruitment to coated pits andclathrin-dependent endocytosis [35]. In the absence of tyrosine phos-phorylation, the rates of both coated-pit recruitment and endocytosisof EGFR were reduced [36]. The mechanism that regulates endocytosisof tyrosine-phosphorylated EGFR is complex and still not fully charac-terized. It appears that there are several redundant and possiblyinterdependent mechanisms, which involve: (a) EGFR binding to theclathrin endocytosis protein complex AP2, and to the phospho-tyrosine binding adaptor protein growth factor receptor-bound protein2 (Grb2); (b) ubiquitination of 15 lysines in the EGFR kinase domain bythe E3 ubiquitin ligase Casitas B-lineage lymphoma (Cbl), and of 6other lysines in the carboxy-terminus region; (c) acetylation of 3 ofthe latter lysines [37]. Knocking down Grb2, a phospho-tyrosine bind-ing protein that interacts with several RTKs, including VEGFR2 [38],had the largest inhibitory effect on EGFR endocytosis. The inhibitionresulted from loss of the recruitment of Cbl to EGFR [39], and loss ofbinding of the Src homology (SH)-3 domain of Grb2 to an unidentified
protein(s) containing poly-proline motif(s), possibly subunits of theAP2 complex [40]. The latter was required for the recruitment ofEGFR to clathrin-coated pits [41]. Ubiquitination is required for the re-cruitment of EGFR to clathrin via the adaptor protein epsin-1 [42]. Therole of lysine acetylation is not clear— it could have a positive effect, orit could inhibit EGFR endocytosis by competing with the ubiquitinationof these lysines.
Internalized EGFR follows the canonical clathrin-dependentendocytic route. EGFR trafficked to early endosomes located in thecell periphery, where it remained dimerized, bound to EGF [43], andphosphorylated [44]. From early endosomes, EGFR either recycledrapidly to the plasma membrane [45], or remained in the sameendocytic compartment during its maturation into multi-vesicularbodies (MVB). EGFR in MVBs was partitioned into two populations.The EGFR population on the limiting MVB membrane underwentslow, probably Rab11-dependent recycling that occurred within15–20 min after the initiation of endocytosis [46]. Another population
5 TyrUbEps1
AP2
Grb2
MVB/Lys
P
EE
ERK
RE
Akt
P
P
EGFR
Rab4
Rab11
Rab5
Fig. 1. Scheme of membrane traffic and signaling of EGFR. Recruitment to clathrin-coated pits requires autophosphorylation (on 5 Tyr residues), association with ofGrb2, and the binding of the AP2 complex to Tyr947, Lys1010, and Lys1011. The Cbl ligaseubiquinates (Ub) 21 lysines located in the cytoplasmic domain of the EGFR, which arerequired for internalization, in part by recruiting the clathrin adaptor protein epsin-1(Eps1). Phosphorylated EGFR traffics via Rab5 to early endosomes (EE), from whereit is recycled to the plasma membrane via Rab4, or to recycling endosomes (RE) fromwhere is recycled to plasma membrane, or to multi-vesicular bodies (MVB). EGFR lo-cated in the limiting membrane of the MVB is recycled to the plasma membrane,whereas the EGFR pool in internal vesicles is degraded when these vesicles fuse withthe lysosome. Akt activation requires endocytosis and proceeds from EGFR in earlyendosomes, whereas ERK activation is not endocytosis-dependent.
1811A. Horowitz, H.R. Seerapu / Cellular Signalling 24 (2012) 1810–1820
Author's personal copy
was located in internal MVB vesicles that fused with lysosomal vesi-cles, where EGFR was degraded [47]. EGFR regulates concomitantlytwo major signaling pathways, the phosphoinositide 3-kinase(PI3K)/Akt pathway, and the extracellular signal-regulated kinase(ERK)/mitogen-activated protein kinase (MAPK) pathway [39]. De-spite a long-standing convention that optimal EGF-triggered ERK/MAPK signaling requires EGFR endocytosis, whereas PI3K/Akt signal-ing does not (reviewed in Ref. [39]), recent studies reversed this view,showing that Akt phosphorylation was severely reduced in cells ex-pressing an internalization-deficient EGFR mutant, but ERK1/2 phos-phorylation level was increased [37]. The discrepancy has not beenexplained. It may arise from the difference between the approachesused to inhibit endocytosis. The studies in the first group used adominant-negative dynamin mutant that inhibited all clathrin-dependent endocytosis, whereas in the study of Goh et al. only theendocytosis of EGFR was inhibited by mutating ubiquitin and AP2binding motifs in its cytoplasmic domain [37].
2. Endocytosis and trafficking of VEGFR
The evolving understanding of VEGFRmembrane traffic appears toconform in some respects with that of EGFR, though the understand-ing of VEGFR2 membrane traffic has not reached the level of detail atwhich EGFR endocytosis is known. Some of the earliest indications forthe functional significance of VEGFR2 trafficking came from studieson guided cell migration in Drosophila [15]. The spatial localizationof PVR, an ortholog of vertebrate PDGFR and VEGFR, in border cellsof the Drosophila embryo, and the capability of the receptor toguide cell migration depended on proteins known to regulate endo-cytosis. Unlike EGFR, VEGFR signals in cooperation with the Nrpnon-catalytic co-receptor, and therefore the membrane traffic ofboth receptors is reviewed. Recently, the internalization of VEGFR2and VEGFR3 was found to depend on ephrin signaling, a pathwaythat was not known before to be linked to VEGFR signaling [18,19].The dependence on ephrin appears to be unique to VEGFRinternalization.
2.1. Endocytic pathway of VEGFR
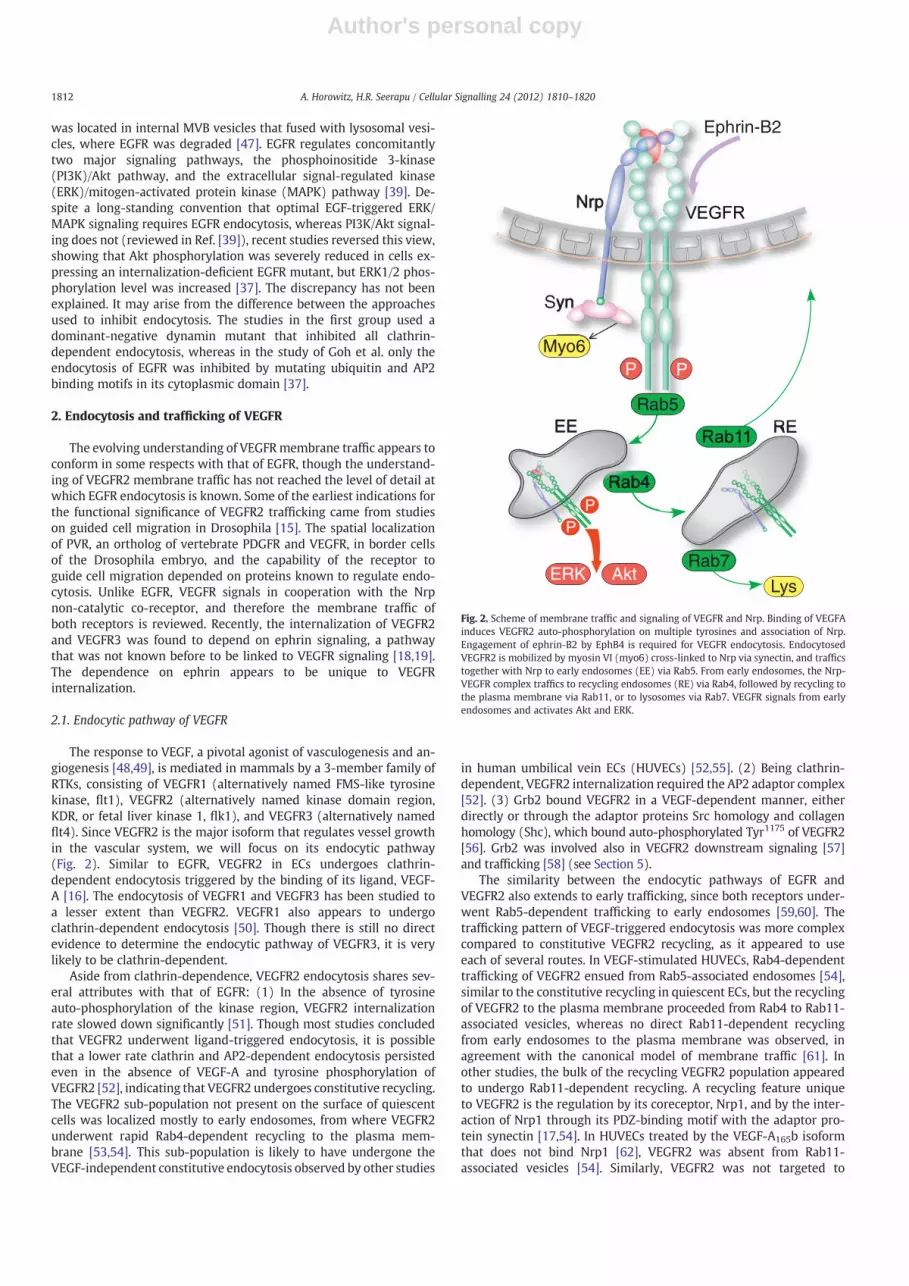
The response to VEGF, a pivotal agonist of vasculogenesis and an-giogenesis [48,49], is mediated in mammals by a 3-member family ofRTKs, consisting of VEGFR1 (alternatively named FMS-like tyrosinekinase, flt1), VEGFR2 (alternatively named kinase domain region,KDR, or fetal liver kinase 1, flk1), and VEGFR3 (alternatively namedflt4). Since VEGFR2 is the major isoform that regulates vessel growthin the vascular system, we will focus on its endocytic pathway(Fig. 2). Similar to EGFR, VEGFR2 in ECs undergoes clathrin-dependent endocytosis triggered by the binding of its ligand, VEGF-A [16]. The endocytosis of VEGFR1 and VEGFR3 has been studied toa lesser extent than VEGFR2. VEGFR1 also appears to undergoclathrin-dependent endocytosis [50]. Though there is still no directevidence to determine the endocytic pathway of VEGFR3, it is verylikely to be clathrin-dependent.
Aside from clathrin-dependence, VEGFR2 endocytosis shares sev-eral attributes with that of EGFR: (1) In the absence of tyrosineauto-phosphorylation of the kinase region, VEGFR2 internalizationrate slowed down significantly [51]. Though most studies concludedthat VEGFR2 underwent ligand-triggered endocytosis, it is possiblethat a lower rate clathrin and AP2-dependent endocytosis persistedeven in the absence of VEGF-A and tyrosine phosphorylation ofVEGFR2 [52], indicating that VEGFR2 undergoes constitutive recycling.The VEGFR2 sub-population not present on the surface of quiescentcells was localized mostly to early endosomes, from where VEGFR2underwent rapid Rab4-dependent recycling to the plasma mem-brane [53,54]. This sub-population is likely to have undergone theVEGF-independent constitutive endocytosis observed by other studies
in human umbilical vein ECs (HUVECs) [52,55]. (2) Being clathrin-dependent, VEGFR2 internalization required the AP2 adaptor complex[52]. (3) Grb2 bound VEGFR2 in a VEGF-dependent manner, eitherdirectly or through the adaptor proteins Src homology and collagenhomology (Shc), which bound auto-phosphorylated Tyr1175 of VEGFR2[56]. Grb2 was involved also in VEGFR2 downstream signaling [57]and trafficking [58] (see Section 5).
The similarity between the endocytic pathways of EGFR andVEGFR2 also extends to early trafficking, since both receptors under-went Rab5-dependent trafficking to early endosomes [59,60]. Thetrafficking pattern of VEGF-triggered endocytosis was more complexcompared to constitutive VEGFR2 recycling, as it appeared to useeach of several routes. In VEGF-stimulated HUVECs, Rab4-dependenttrafficking of VEGFR2 ensued from Rab5-associated endosomes [54],similar to the constitutive recycling in quiescent ECs, but the recyclingof VEGFR2 to the plasma membrane proceeded from Rab4 to Rab11-associated vesicles, whereas no direct Rab11-dependent recyclingfrom early endosomes to the plasma membrane was observed, inagreement with the canonical model of membrane traffic [61]. Inother studies, the bulk of the recycling VEGFR2 population appearedto undergo Rab11-dependent recycling. A recycling feature uniqueto VEGFR2 is the regulation by its coreceptor, Nrp1, and by the inter-action of Nrp1 through its PDZ-binding motif with the adaptor pro-tein synectin [17,54]. In HUVECs treated by the VEGF-A165b isoformthat does not bind Nrp1 [62], VEGFR2 was absent from Rab11-associated vesicles [54]. Similarly, VEGFR2 was not targeted to
Rab4P
EE RE
Nrp
Syn
Myo6
phrin-B2
LysERK Akt
Rab5
Rab7
P
E
Fig. 2. Scheme of membrane traffic and signaling of VEGFR and Nrp. Binding of VEGFAinduces VEGFR2 auto-phosphorylation on multiple tyrosines and association of Nrp.Engagement of ephrin-B2 by EphB4 is required for VEGFR endocytosis. EndocytosedVEGFR2 is mobilized by myosin VI (myo6) cross-linked to Nrp via synectin, and trafficstogether with Nrp to early endosomes (EE) via Rab5. From early endosomes, the Nrp-VEGFR complex traffics to recycling endosomes (RE) via Rab4, followed by recycling tothe plasma membrane via Rab11, or to lysosomes via Rab7. VEGFR signals from earlyendosomes and activates Akt and ERK.
1812 A. Horowitz, H.R. Seerapu / Cellular Signalling 24 (2012) 1810–1820

Author's personal copy
Rab11-associated vesicles in HUVECs expressing a Nrp1 mutant de-void of its carboxy-terminus PDZ binding motif [54]. These observa-tions suggest that Nrp1 and its interaction with PDZ domain-containing proteins are required for Rab11-dependent recycling ofVEGFR2. Synectin (also named neuropilin-1 binding protein, NIP,and GAIP-interacting protein, C-terminus, GIPC) is the only knownprotein that binds to the PDZ-binding motif of Nrp1 [63]. Synectinfunctions as a universal adaptor of the inward-traffic molecularmotor myosin VI [64]. This interpretation counters, however, previ-ous observations that narrowed down the role of both synectin andmyosin VI to the inward trafficking of uncoated vesicles up to their fu-sion with early endosomes [65], where trafficking is regulated byRab5 rather than by Rab11. Alternatively, other studies attributedthe dependence on synectin and myosin VI to VEGFR2 rather thanNrp1 [17]. Though VEGFR2 does not have a PDZ-binding motif, it wassuggested that by analogy with the nerve growth factor receptor TrkA,synectin could be recruited via the adaptor protein phospho-tyrosineinteraction, PH domain, and leucine zipper containing (APPL) [66].The study of Lanahan et al. [17] ruled out Nrp1-dependent traffickingof the VEGFR2-Nrp1 complex because VEGF-D, an isoform that doesnot bind Nrp1, produced a similarly lower activation of ERK1/2 as inECs isolated from synectin−/− mice. The reliance on VEGF-D counters,however, evidence for VEGF-D binding to Nrp1 [67]. Further, the puta-tive absence of Nrp1 in the VEGFR2–VEGF-D complex could not ruleout the dependence of VEGFR2 trafficking on Nrp1, because the exper-iment was carried out in synectin−/− ECs. The absence of synectinwould prevent VEGFR2 trafficking even if Nrp1 had been present inthe VEGFR2–VEGF-D complex.
Interestingly, the study of Ballmer-Hofer et al. [54] found that onceVEGFR2 entered the Rab11-associated trafficking compartment, itwas no longer phosphorylated at Tyr1175, a post-translational modifi-cation that is required for both MAPK and Akt activation [68]. Thisindicates that the endosomal compartment from which VEGFR2signaled preceded Rab11-dependent trafficking. The finding ofBallmer-Hofer et al. [54] concurs with the study of Lanahan et al.[17], where Rab11 knockdown increased MAPK and Akt activation,presumably because it slowed down the departure of VEGFR2 froma preceding compartment. Collectively, these findings suggest thatVEGFR2 signals from early or from recycling Rab4/Rab5-associatedendosomes. The two studies suggest different sites, however, for thedephosphorylation of VEGR2. Whereas the study of Ballmer-Hoferet al. [54] implies that VEGFR2 was dephosphorylated immediatelyafter exiting the signaling compartment and before Rab11-dependentrecycling, the study of Lanahan et al. suggests that VEGFR2 wasdephosphorylated by the protein tyrosine phosphatase 1b (PTP1b)near the plasma membrane. The localization of PTP1b activity nearthe plasma membrane is supported by other studies [69], whereasno PTP was identified in the study of Ballmer-Hofer et al. [54] as acandidate phosphatase of VEGFR2. There is consensus between severalstudies that part of the VEGFR2 population progressed from either earlyor recycling endosomes (Rab5 or Rab4-associated, respectively) toRab7-associated late endosomes and lysosomes, i.e. to degradation[17,54,60]. The lysosome-targeted VEGFR2 fractionwas small, however,as long as VEGFR2 co-trafficked with Nrp1. Once the VEGFR2-Nrp1 as-sociation was perturbed, a larger fraction of the VEGFR2 populationwas diverted to Rab7-associated endosomes [54]. While a mechanisticexplanation of the dependence of VEGFR2 routing on the associationwith Nrp1 is lacking, it is possible that the lateral binding of VEGFR2to Nrp1 interfered with the lysine ubiquitination required for RTKdegradation [37,70]. When VEGFR2 degradation was inhibited byknocking down Rab7, MAPK activation increased [17,60]. This obser-vation is in agreement with the premise that VEGFR2 signals fromearly and/or Rab4-associated recycling endosomes. A caveat thatmay be considered in regard to the studies on VEGFR2 traffickingdiscussed above is that their time resolution was relatively low, par-ticularly during the first 30 min after VEGF-A treatment. Since most
of the data was sampled at 30 min or later, it is possible that atleast part of the data corresponded to the steady state, and did notrepresent the transient phase of trafficking. In fact, 30 min after en-docytosis had been triggered, a large fraction of VEGFR2 in VEGF-A-treated mouse primary ECs had already recycled back to the plasmamembrane [21].
The colocalization of internalized VEGFR2 with Rab7, a GTPase re-quired for late endosome fusion with lysosomes [61], indicates thatVEGFR2 was targeted to that compartment rather than to theproteasome. However, recent evidence indicates that VEGFR2 wasprocessed by the proteasome prior to undergoing final degradationin the lysosome [32]. Proteolytic cleavage appears to have occurredimmediately after VEGFR2 passed through the early endosomes, pro-ducing a fragment that corresponds to the transmembrane andectoplasmic domains of the receptor. Expectedly, inhibiting theproteasome extended the duration of the activation of the down-stream effectors Akt and MAPK. The complementary fragmentcorresponding to the cytoplasmic domain of VEGFR2 could conceiv-ably still function as a tyrosine kinase and activate downstream effec-tors. Surprisingly, when the cytoplasmic domain was expressed as achimera fused at its amino-terminus to yellow fluorescent protein(YFP), Tyr1175 was phosphorylated in the absence of VEGF, and Aktand MAPK were activated. However, the putative endogenous cyto-plasmic domain was not detected directly, and its phosphorylationstate in either the presence or absence of VEGF was not determined.
Similar to EGFR, the main downstream signaling pathways ofVEGFR2 regulate either PLC /MAPK or PI3-Kinase/Akt [68]. A conflu-ence of evidence from numerous studies established unambiguouslythat internalization was required for the optimal tyrosine phosphory-lation of VEGFR2 and for the subsequent activation of both pathways[17–19,60,71–73]. Unlike EGFR signaling, the studies on VEGFR2 didnot find differential effects of the internalization on Akt versusMAPK activation, concluding, instead, that both signaling pathwayswere upregulated by endocytosis of VEGFR2. Interestingly, exposureof ECs to matrix-bound, rather than to soluble VEGF-A165 producedan increase in the activation of p38 [72], a MAPK that remodels theactin cytoskeleton and induces cell migration in response to VEGF[74]. The mechanism that accounts for the signaling specificity ofmatrix-bound VEGF-A165 is unknown. An exception to the similaritybetween the endocytic mechanisms of EGFR and VEGFR is the in-volvement of cerebral cavernous malformation 3 (Ccm3) [75], anadaptor/scaffolding protein that is mutated in patients sufferingfrom vascular malformation in the brain. Ccm3 bound to and col-ocalized with VEGFR2 on the plasma membrane in quiescent ECs.Ccm3 remained colocalized with VEGFR2 in cytoplasmic punctaethat seemed to be vesicles, upon VEGF treatment, but the vesicleswere not characterized by endocytic markers. Since knockdown ofCcm3 resulted in the internalization of VEGFR2, and reduced the acti-vation of MAPK and Akt, it was deduced that Ccm3 was required forstabilizing VEGFR2 at the cell surface and, therefore, prolonging itssignaling. This conclusion is at odds, however, with the recent evi-dence that endocytosis is required for VEGFR optimal signaling[17–19]. Since Ccm3 internalized and colocalized with VEGFR2, itcould hypothetically be involved in VEGFR2 sorting, similar toCcm1. The latter binds sorting nexin 17 [76], which diverts membranereceptors away from lysosomal degradation by preventing their entryinto the internal vesicles of the MVB [77]. This would explain the“destabilization” of VEGFR2 when Ccm3 was knocked down [75].
It was recently observed that as much as a 30% fraction of the cel-lular VEGFR2 population was localized to the trans-Golgi apparatus inquiescent HUVECs [78]. Upon treatment by VEGF-A165, VEGFR2 exitedalmost completely from the trans-Golgi to cytoplasmic vesicles. Thetrans-Golgi VEGFR2 pool was regulated by N-ethylmaleimide‐sensitive factor attachment protein receptor (SNARE) syntaxin-6[78]. Since syntaxin-6 regulates transport from the trans-Golgi tothe plasma membrane [79], it is likely that the study of Manickam
1813A. Horowitz, H.R. Seerapu / Cellular Signalling 24 (2012) 1810–1820

Author's personal copy
et al. [78] detected the exocytosis of the trans-Golgi VEGFR2 pool tothe plasma membrane in response to VEGF.
VEGF signaling is distinct from other RTK ligands in originatingfrom intracellular pools in an autocrine manner [80]. Despite the ab-sence of exogenous VEGF, hypoxia increased the tyrosine phosphory-lation of VEGFR2 [80]. Since only a cell-permeable inhibitor ofVEGFR2 had an effect on its tyrosine phosphorylation level, it appearsthat the receptor was activated by the pool of internalized VEGF. Thetrafficking pathway of the VEGFR2 population activated by intracellu-lar VEGF is unknown.
2.2. Regulation of VEGFR2 endocytosis and sorting by ubiquitination
Aside from the well-known role in marking proteins for degrada-tion, ubiquitination was required for the recruitment of adaptor pro-teins that sort endocytosed RTKs [81]. This has been demonstratedparticularly for EGFR [81]. Although it had been established thatVEGFR2 was ubiquitinated in a ligand-dependent manner in responseto VEGF [82], the role of ubiquitination in VEGF signaling is less un-derstood in comparison to EGFR. Early studies established that theE3 homologous to E6-AP C terminus (HECT) domain ubiquitin ligaseNedd4 mediated the proteasomal degradation of VEGFR2, and thatthis degradation could be inhibited by the sequestration of Nedd4by Grb10 [82], an adaptor protein that bound and was phosphorylat-ed by VEGFR2 [83]. However, Nedd4 was not shown to ubiquinateVEGFR2, or to regulate VEGFR2 sorting during internalization. TheE3 ligase Cbl bound phosphorylated Try1052/1057 of VEGFR2 in porcineECs, and ubiquitinated and inhibited PLC 1 bound to Tyr1173, but itwas not implicated in the endocytosis of VEGFR2 [84]. Recent studiesestablished that activated VEGFR2 bound the E3 ligase β-transducinrepeats-containing protein (TRCP) via Ser1188/1191 located in aVEGFR2 motif that appears to conform to the PEST (rich in proline[P], glutamic acid [E], serine [S], and threonine [T]) consensus se-quence [85]. While TRCP was shown to regulate the poly-ubiquitination of VEGFR2, it was not linked to VEGFR2 endocytosis[85]. RTK endocytosis is regulated by mono-ubiquitination [86],whereas poly-ubiquitination regulates proteolysis. Similar to EGFR[31], the endocytic sorting of VEGFR2 may be regulated by mono-ubiquitination of yet unidentified lysines that would form bindingsites to endocytic adaptor proteins such as epidermal growth factorreceptor substrate 15 (EPS15).
2.3. Regulation of VEGFR endocytosis by ephrin
Eph receptors are RTKs that guide neuronal patterning and facili-tate the segregation of developing vessels into arteries and veins[87]. Eph signaling is triggered by binding in trans to ephrin receptorson apposing cells. Eph-ephrin signaling is bi-directional, both for-ward, via a tyrosine kinase phosphorylation cascade (Eph), and in re-verse (ephrin), by binding other cell surface receptors in trans(ephrin A subfamily), or by binding cytoplasmic proteins (ephrin Bsubfamily). The expression patterns of EphB4 and ephrin-B2 in thevascular system are complementary, so that EphB4 is expressed pri-marily arteries, whereas ephrin-B2 is expressed in veins [88,89].Though the precise functions of Eph receptors and ephrin in the vas-culature have not been determined, their crucial role in vascular de-velopment was demonstrated in loss-of-function murine models.Disruption of the genes coding for either protein resulted in severeangiogenesis defects and embryonic lethality [88,89]. The angiogenicdefects in the EphB4−/− mouse were symmetric to those in theephrinB2−/− mouse, mirroring the complementary vein-artery distri-bution of EphB4 and ephrin-B2, respectively.
Two recent studies revealed an unexpected connection betweenEphB4-ephrin-B2 signaling, and between the internalization ofVGFR2 [18] and VEGFR3 [19]. Engagement of ephrin-B2 by its ligand,EphB4, was followed by internalization of VEGFR3 in lymphatic ECs
[19]. Other RTKs, e.g. PDGFR, were not internalized, indicating thatthe effect of ephrin-B2-EphB4 binding is specific to VEGFR. VEGFR3failed to undergo endocytosis in ECs from the ephrinB2−/− mouse.VEGF-C-mediated tyrosine phosphorylation of VEGFR3 and theactivation of Akt and of MAPK were markedly reduced in theephrinB2−/− ECs. Ephrin-B2-EphB4 binding caused endocytosis ofVEGFR3 in the absence of VEGF-C, but the internalized VEGFR3 wasnot auto-phosphorylated. VEGFR2 was observed to undergo a similarephrin-B2-dependent endocytosis in vascular ECs, in the presence ofVEGF-A [18].
It is unknown how ephrin-B2-EphB4 signaling induced the inter-nalization of VEGFR2/3, but the above studies provide some insightinto the underlying mechanism. VEGFR3 endocytosis appears tohave been triggered by transient association in cis between ephrin-B2 and VEGFR3 at the plasma membrane, but once internalized,ephrin-B2 and VEGFR2/3 no longer colocalized [19]. Notably, the in-teraction of ephrin-B2 with cytoplasmic proteins through its PDZ-binding motif was required for VEGFR2 endocytosis [18]. To date, atleast nine PDZ domain-containing proteins that bind ephrin-B2 havebeen identified (http://www.signaling-gateway.org). One of these,PDZ domain-containing RING finger protein 3 (PDZRN3), is an E3ubiquitin-protein ligase [90]. Thus, the lateral association ofVEGFR2/3 with ephrin-B2 may facilitate the ubiquitination ofVEGFR2/3 by ephrin-B2-bound PDZRN3. Though this has not beenas well established as for EGFR [37], it is conceivable that Cbl-mediated ubiquitination of VEGFR would be required for its endocy-tosis, thus accounting for the dependence of VEGFR2 endocytosis onephrin-B2.
3. Endocytosis and trafficking of Nrp1
3.1. Association between Nrp1 and VEGFR2
The uniqueness of the endocytic pathway of VEGFR is conferredmainly by its association with Nrp1. This is demonstrated by the con-sequences of perturbing the association between the two receptorson VEGFR2 signaling, as described above. Unlike the transientcolocalization between ephrin-B2 and VEGFR at the cell surface [19],Nrp1 and VEGFR were 50–60% colocalized even in quiescent mouseprimary ECs, and remained so up to 30 min after stimulation byVEGF-A165 [21]. Once VEGF-A was applied, Nrp1 and VEGFR2 becamephysically associated [21,91,92], and remained colocalized up to30 min after the application of VEGF-A165 [21]. It is unknownwhetherthe Nrp1-VEGFR2 association is direct or indirect, and which aminoacid sequences and structural motifs of each receptor are requiredfor VEGF-induced association. The dependence of the Nrp1-VEGFR2association on the binding of the PDZ adaptor protein synectin [93]suggests that the carboxy-terminus of Nrp1 is required for the associ-ation. Since VEGFR2 is not known to bind synectin, it is not clear whysynectin would be required for the association. If APPL binds VEGFR2,as suggested by Lanahan et al. [17], Nrp1 and APPL-bound synectindimers could cross link the two receptors [64,94,95], thus explainingthe dependence of the Nrp1-VEGFR2 association on synectin.
3.2. Endocytic pathway of Nrp1
Since Nrp1 colocalized extensively with VEGFR2 on the surface ofquiescent ECs [21], it could have conceivably accompanied VEGFR2when the latter underwent constitutive endocytosis [52]. There is di-rect evidence for VEGF-independent endocytosis and trafficking ofNrp1 together with integrin α5β1 from adhesion complexes, includ-ing focal-adhesions, to early endosomes [22]. The ligand-triggeredendocytosis of Nrp1 in response to VEGF-A is clearly clathrin depen-dent, in contrast to semaphorin 3C-induced endocytosis, which islipid raft-dependent [21]. In agreement with the finding that Nrp1and VEGFR2 remain largely colocalized throughout endocytosis and
1814 A. Horowitz, H.R. Seerapu / Cellular Signalling 24 (2012) 1810–1820

Author's personal copy
trafficking [21], Nrp1 traversed the same endocytic compartments asVEGFR2, i.e. early endosomes [21,54], followed by “slow” recycling viaRab11-dependent trafficking. Only a minor fraction of the VEGFR2–Nrp1 complexes was diverted via Rab7-associated endosomes to ly-sosomal degradation [54].
3.3. Role of Nrp1 in VEGF signaling
Several lines of evidence suggest that Nrp1 signals independentlyfrom VEGFR, or that it is required for transducing aspects of VEGF sig-naling that are not mediated by VEGFR alone. The putative contribu-tion of Nrp1 to VEGF signaling would obligately be mediated by thecytoplasmic domain. Since the carboxy-terminus of the same domainis used by synectin to cross-link Nrp1 to myosin VI, the molecularmotor that drives Nrp1 trafficking [17,21], it is likely that Nrp1-dependent signaling is tightly coupled to Nrp1 trafficking.
The first indications for Nrp1-triggered signaling came from stud-ies where the ectoplasmic domain of Nrp1 was replaced with that ofEGFR and expressed in HUVECs lacking endogenous EGFR [96]. EGFtreatment of these cells promoted their migration to a similar extentas VEGF-induced migration. Interestingly, when the EGFR-Nrp1 chi-mera consisted of a Nrp1 cytoplasmic domain lacking the carboxy-terminus PDZ-binding motif, VEGF failed to promote cell migration.These results imply that EGF-induced clustering of the cytoplasmicdomain of Nrp1 was sufficient to trigger downstream signaling, pre-sumably in a PI3K-dependent manner [96]. However, these conclu-sions rest on the finding that immunoblotting did not detect EGFRin the HUVECs used by Wang et al. [96]. It is possible that thoughEGFR was not detectable by immunoblotting, its expression levelwas sufficient to activate PI3K and elicit cell migration. Moreover,HUVECs are highly variable primary ECs, and are known to expressEGFR [97]. Thus, the results of Wang et al. [96] would be valid provid-ed the same EGFR-deficient batch of HUVECs had been used through-out the study.
The requirement for the binding of synectin to the carboxy-terminus of Nrp1 was suggested also by studies where nrp1 wasknocked down in the zebrafish, inhibiting the angiogenesis of theintersomitic vessels and the sub-intestinal vein [98]. Whereas thisvascular phenotype was rescued by the expression of human NRP1mRNA, expression of a mutant Nrp1 lacking the PDZ-binding was in-effective. The requirement for this motif in vascular development wasrecapitulated by the similarity between the nrp1 and the synectinmorphants.
While the above zebrafish studies suggest that Nrp1 can signal,they do not implicate it directly in transducing VEGF signaling. Evi-dence to that end was provided by studies in which antibodiesspecific to the ectoplasmic domain of Nrp1 were used to disrupt itsassociation with VEGFR2 [91]. Strikingly, while the tyrosine phos-phorylation level of VEGFR2 and the proliferative and permeabilityaspects of VEGF signaling were unchanged, the migration of HUVECstreated with Nrp1 antibodies was reduced drastically. The same anti-bodies strongly inhibited also the VEGF-driven outgrowth of HUVECsfrom beads encased in a fibrinogen matrix. These results suggest thatNrp1 was required for transducing the motility aspect of VEGF signal-ing. Similar conclusions could be drawn from a subsequent studyfrom the same group, showing that binding of the VEGF-A121 isoformto Nrp1 was similarly required for sustaining EC migration [99]. SinceVEGF-A121 binds Nrp1 but does cross-link it to VEGFR2, this result canbe construed to suggest that Nrp1 can signal independently fromVEGFR2. Were this scenario correct, how then would a short cytoplas-mic domain lacking catalytic activity transduce the binding of VEGF toits ectoplasmic domain? A conceivable mechanism is the recruitmentof a cytoplasmic protein complex that can signal once assembled.Aside from synectin, there is evidence for the binding of focal-adhesion kinase (FAK) [100] and the non-receptor tyrosine kinaseFer [101]. Both of these interactions do not appear to be involved in
VEGF signaling, but other studies implicated Nrp1 in VEGF-inducedtyrosine phosphorylation of FAK and, consequently, in the regulationof focal-adhesion assembly [102]. The potential involvement of Nrp1in the regulation of focal-adhesions was suggested also by its role inthe phosphorylation of p130Cas, an event that appeared to be medi-ated by PYK2, a tyrosine kinase that is closely related to FAK [103].Aside from the binding of synectin to the cytoplasmic domain ofNrp1, there is no evidence to implicate endocytosis and membranetrafficking of Nrp1 in any of the putative signaling mechanismsdescribed here.
Given the above convincing evidence for the migration/angiogen-esis-specific role of Nrp1 in VEGF signaling, it is surprising that amouse knockin (KI) model expressing a mutant nrp1 lacking the cyto-plasmic domain (nrp1cytoΔ/Δ) was not embryonic lethal, andappeared grossly normal [104]. Phenotyping of the embryonic vascu-lar system of the nrp1cytoΔ/Δ mouse detected no major defects, andanalysis of the retinal vasculature in the neonatal nrp1cytoΔ/Δmouse found a relatively minor defect consisting of excessiveartery-vein crossovers. This defect did not interfere with the forma-tion of an intact retinal vasculature. While the analysis of other vascu-lar beds in the nrp1cytoΔ/Δ was not reported, the presence of suchminor defects is perplexing given that antibody-mediated disruptionof the association between Nrp1 and VEGFR2 interfered with thedevelopment of the retinal vasculature [91]. The absence of vasculardefects cannot be attributed to compensation by Nrp2, since theretinal vasculature in mice expressing nrp1cytoΔ/Δ on a nrp2-nullbackground was indistinguishable from that in mice expressingnrp1cytoΔ/Δ on a normal genetic background [104]. The discrepancybetween the mild vascular phenotype of the nrp1cytoΔ/Δ KI mouseand the abundant evidence for the role of the Nrp1 cytoplasmic do-main in angiogenesis remains unreconciled. Further, the apparentfailure to rescue the zebrafish nrp1 vascular morphant by expressinghuman Nrp1 lacking the PDZ binding motif (Nrp1-1ΔSEA) suggeststhat at least in this model system the cytoplasmic domain, and in par-ticular its PDZ-binding motif, is required for angiogenesis [98]. How-ever, the implications of the nrp1cytoΔ/Δ genetic in-vivo model forthe function of Nrp1 in mammalian vascular development should ar-guably persevere over conclusions drawn from the numerous in-vitromodels, or from non-genetic in-vivo models [91]. It appears, there-fore, that at least during development, the primary signaling functionof the Nrp1 cytoplasmic domain is not the regulation of angiogenesis.Conceivable VEGF-dependent functions would be the regulation ofvessel permeability [105,106] or of endothelial cell adhesion via integrin[107]. There is no evidence to suggest that such functions entail Nrp1endocytosis and trafficking.
4. Endocytosis of adhesion receptors
VEGF signaling induces the endocytosis of twomajor classes of ad-hesive receptors: intercellular junction proteins, and ECM receptors.Both adherens and tight junction proteins, i.e. vascular endothelialcadherin (VEcad) and occludin, respectively, are endocytosed in re-sponse to VEGF [16,108,109]. Among ECM adhesion receptors, severaltypes of integrin α and β subunits are internalized in response toVEGF [110,111]. Though it is known primarily for its function in theplasminogen activation cascade, the urokinase plasmin activator re-ceptor (uPAR) is also reviewed here, since it interacts with the ECMprotein vitronectin and with several integrin β subunits [112].
4.1. Endocytosis of intercellular adhesion receptors
One of the major cell functions regulated by VEGF-A throughVEGFR2 is the dissociation of EC junctions and the subsequent in-crease in vessel permeability. At the cellular level, the loss of vesselpatency is caused by the disassembly of EC adherens and tight junc-tions in a process that involves the Src tyrosine kinase [113]. It
1815A. Horowitz, H.R. Seerapu / Cellular Signalling 24 (2012) 1810–1820

Author's personal copy
appears that in quiescent ECs, VEcad is associated with VEGFR2 viaβ-catenin [114], and functions to stabilize the receptor at the cellsurface [113,115] (Fig. 3). Once VEGFR2 was activated, VEcad disso-ciated from it and bound β-arrestin [108], an adaptor moleculeknown primarily for facilitating the endocytosis of G-protein coupledreceptors, which facilitates the formation of clathrin-coated vesicles[116]. The recruitment of β-arrestin to VEcad was initiated by Src-dependent phosphorylation and activation of the Rho GEF Vav2.The latter activated Rac, resulting in the activation of p21-activatedkinase (PAK), and the phosphorylation of VEcad on Ser665, whichserved as a β-arrestin-docking site [108]. VEcad underwent clathrin-dependent endocytosis [16,108] and trafficked to early endosomes[108], possibly employing myosin X, an actin-based molecular motor[117]. VEcad did not colocalize after endocytosis with VEGFR2 [16],in agreement with the reported dissociation of VEcad from activatedVEGFR2 [118].
Similar to adherens junctions, the disassembly of tight junctions inresponse to VEGF is mediated by the endocytosis of its components.Among the integral tight junction proteins, the endocytic mechanismof the 4-pass transmembrane protein occludin has been elucidated inprimary bovine retinal endothelial cells [109]. Occludin endocytosisfollowed the VEGF induced activation of protein kinase C (PKC) ζ,also a tight junction component [119], via an unknown pathway.PKCζ phosphorylated occludin at Ser490, resulting in its removalfrom the cell borders and its colocalization with clathrin, withclathrin-binding adaptor protein epsin-1, EPS15, and with hepatocyte
growth factor-regulated tyrosine kinase substrate (Hrs) (Fig. 4). Thelatter two proteins contain a ubiquitin-interacting motif and facilitatethe ubiquitination and targeting of their ligands to lysosomal degra-dation [120]. Following endocytosis, occludin reappeared in cytoplas-mic punctae that colocalized with early endosome and lysosomalmarkers [109]. After the Ser490 phosphorylation of occludin, the PDZdomain-containing protein ZO1 dissociated from occludin [121].Since ZO1 functions as a scaffold that cross-links tight junction trans-membrane proteins and the actin cytoskeleton [122], its dissociationfrom occludin is a key step in junction disassembly. Interestingly,Murakami et al. [109] observed that claudin-5, a 4-pass and ZO1-binding transmembrane protein like occludin, also underwent endo-cytosis in response to VEGF.
4.2. Endocytosis of extracellular matrix (ECM) receptors
As reviewed elsewhere [123], the endocytosis and trafficking ofintegrins is required for the regulation of Rho GTPases, and for theregulation of the signaling of integrin-associated growth factor recep-tors. VEGF-driven endocytosis and trafficking of integrin αvβ3, a fi-bronectin and vitronectin receptor [124], was mediated in HUVECsby protein kinase D1 (PKD1) [110]. PKD1 associated with integrinαvβ3 constitutively, regardless of VEGF-A signaling [110,125].VEGF-A activated PKD1 by inducing its phosphorylation of Ser744/748
in the activation loop, most likely by protein kinase Cα [126](Fig. 5). Additionally, VEGF-A targeted PKD1 to nascent focal-adhesions (focal complexes) on cell protrusions by inducing phos-phorylation of Ser916. Knockdown of PKD1 enhanced integrin αvβ3recycling, while inhibiting the formation of focal-adhesions and themigration of HUVECs [110]. It appears, therefore, that integrin αvβ3underwent constitutive Rab5 and Rab4-dependent recycling throughearly endosomes, and that the activation of PKD1 by VEGF-A inhibitedthe endocytosis of αvβ3 via an unknown mechanism. As a result,PKD1 knockdown tipped the balance maintained by constitutive
Src Rac
VEGFR VEcad
S665
PP β-arrβ-cat
EE EE
Vav2 P
PP
Pak
Fig. 3. Scheme of the endocytic mechanism of VEcad. Activated VEGFR induces Src to ac-tivate Vav2, which then activates Rac and PAK. The latter phosphorylates the VEcad cyto-plasmic domain on Ser665, which serves as a docking site for β-arrestin (β-arr). β-arrestinfacilitates the formation of clathrin-coated vesicles. During or after endocytosis, β-catenin(β-cat), which cross-links VEGFR2 and VEcad, dissociates from VEGFR2. VEcad andVEGFR2 proceed to traffic separately to early endosomes (EE).
S490
Occludin
Occludin
ZO1
PKCζ P
Fig. 4. Scheme of the endocytic mechanism of occludin. Activated VEGFR induces PKCζto phosphorylate occludin on Ser490. Following the phosphorylation, ZO1, which wasbound to occludin in tight junctions, dissociates from it. Occludin undergoes clathrin-dependent endocytosis.
1816 A. Horowitz, H.R. Seerapu / Cellular Signalling 24 (2012) 1810–1820

Author's personal copy
recycling between inward and outward trafficking towards the latter,resulting in a higher apparent presence of integrin αvβ3 on the cellsurface. Though VEGF treatment resulted in a relatively small increasein the recycling rate of integrin αvβ3 in comparison to constitutiverecycling, the VEGF-induced relative increase in EC migration ratewas far larger [110]. Integrin can undergo either clathrin orcaveolin-dependent endocytosis [127], but the study of di Blasio etal. [110] did not positively identify the endocytic pathway.
The VEGF-induced endocytosis of the β1 integrin subunit incorpo-rated uPAR, a glycosyl phosphatidylinositol (GPI)-anchored cellsurface receptor noted primarily for its role in the plasminogen acti-vation cascade. uPAR binds uPA, which cleaves plasminogen to gener-ate plasmin [128]. Aside from binding uPA, uPAR functions as areceptor for numerous ligands, including the ECM protein vitronectin,and binds in cis several integrin β subunits, including β1 [128,129].Whereas integrin β1 (in tandem with subunit α5) was localized inquiescent HUVECs mainly at the cell periphery, treatment withVEGF-A165 redistributed it to cytoplasmic vesicular structures,where it colocalized with uPAR [111]. Since previous studies hadshown that uPAR underwent clathrin-dependent endocytosis [130],it is likely that VEGF-A165 induced the endocytosis of integrin β1 inthe same manner. VEGF-A165 failed to induce clathrin-dependent en-docytosis of integrin β1 in ECs of uPAR−/− mice, or in ECs treated witha peptide that inhibited the interaction of uPAR with lipoproteinreceptor-related protein (LRP) [130]. These results suggest thatintegrin β1 endocytosis required coordination between LRP, uPAR,and integrin α5β1, as follows: LRP associated with uPAR that was en-gaged by uPA bound to PAI-1, followed by the association of uPAR to
integrin β1 [130] (Fig. 6). Blocking the integrin β1-uPAR associationinhibited EC migration towards a VEGF-A165 gradient [111], indicat-ing that VEGF-induced endocytosis of integrin α5β1 was requiredfor angiogenesis in-vivo. Though VEGF-A165 also induced endocytosisof integrin α3β1, this endocytosis did not require interaction ofintegrin β1 with uPAR [111].
It should be noted that VEGFR2 and integrin β1 [131] or β3[131–133] signaling were shown to have reciprocal functional rela-tions, but those were not reported to involve endocytosis or traffick-ing of either receptor.
5. Endocytosis and trafficking of Rho GTPases
Recent studies found that VEGF signaling can induce the traffick-ing not only of cell-surface receptors, but also of Rho GTPases [58].A previous study showed that hepatocyte growth factor, a ligand ofthe Met RTK, induced membrane trafficking of Rac1, and caused itsactivation on endosomes by the Tiam1 GEF in a Rab5-dependentmanner [134]. Similarly, VEGF-A165 induced the trafficking of RhoA,together with its GEF Syx [58]. Unlike Rac1, the trafficking of RhoAwas mediated by Rab13, a GTPase mainly known for regulating theendocytosis of occludin from tight junctions [135]. In primarymouse ECs migrating in response to a VEGF-A165 gradient, Rab13 aswell as RhoA and Syx were localized not only to endocytic vesicles,but also at the leading edge of the cells [58]. In agreement with previ-ous studies, it appears that Rab13 is involved not only in the regula-tion of tight junctions but also in the remodeling of the actin
Integrin
PKCα
P PPKD1
αv
S744/748
Rab5
Rab4
Fig. 5. Scheme of the endocytic mechanism of integrin αvβ3. In quiescent ECs, integrinαvβ3 undergoes constitutive recycling to early endosomes. VEGF activation of PKD1 viaphosphorylation of Ser744 and Ser748, most likely by PKCα, results in the binding ofPKD1 to the cytoplasmic domain of the β3 subunit, and inhibition of integrin αvβ3 en-docytosis. This produces an apparent increase in the integrin αvβ3 presence on the cellsurface.
Integrin
β1α5
uPAR
uPA
PAI-1
LRP
Fig. 6. Scheme of the endocytic mechanism of integrin α5β1 and uPAR. Downstream ofVEGF, uPA binds together with PAI-1 to uPAR on the EC surface. The binding is followedby lateral association with LRP, after which the uPAR-LRP complex associates laterallywith the integrin β1 subunit. Integrin α5β1 undergoes clathrin dependent endocytosistogether with uPAR, and possibly also with LRP.
1817A. Horowitz, H.R. Seerapu / Cellular Signalling 24 (2012) 1810–1820

Author's personal copy
cytoskeleton of migrating cells [136,137]. The hypothetical scenarioof VEGF-A-driven trafficking starts with the induction of clathrin-dependent endocytosis of tight junction proteins by VEGF (Fig. 7).Multi PDZ domain protein (MUPP1)-1, which binds the integraltight junction transmembrane proteins junctional adhesion molecule(JAM) and claudin [138,139], would then remain attached to their cy-toplasmic domains and would be removed from the tight junctions onvesicles engulfing JAM and claudin. Mupp1 is known to bind Syx[140] and angiomotin (Amot). Amot would facilitate the targeting ofMupp1 and Syx to the lipid bilayer of endocytic vesicles due to itshigh affinity for phosphatidylinositol 3-phosphate [141], which isenriched in the membranes of vesicles [142]. Rab13 would form acomplex with the actin-binding protein actinin-4 (Actn4) throughtheir common binding partner, molecule interacting with CasL-like 2(Mical-l2) [136]. The Rab13/Mical-l2/Actn4 complex would target theSyx-RhoA carrying vesicles to endocytosed tyrosine-phosphorylatedVEGFR2, since Actn4 binds Grb2, which binds VEGFR2 [38]. Thismechanism can account for the localization of RhoA, Syx, and Rab13to the leading edge of migrating ECs [58]. Knockdown of rab13 inthe zebrafish inhibited angiogenesis, thus supporting the in-vivo roleof VEGF-induced trafficking of RhoA [58].
6. Concluding remarks
The unique attributes of VEGFR signaling and its regulation bymembrane traffic are conferred largely by the association with theco-receptor Nrp. The phenotyping of the nrp1cytoΔ/Δ mouse ruledout, surprisingly, an obvious signaling role for the cytoplasmic do-main of Nrp1 in vasculogenesis and angiogenesis, aside from
arteriovenous patterning in the retina. It is likely that there is moreto the cytoplasmic domain of Nrp1 than the binding via its PDZmotif to a single protein, synectin. One potential function is the regu-lation of focal-adhesions in-vitro [102], but it is unclear what is the in-vivo equivalent of this function, since the in-vivo analog of two-dimensional focal adhesions in not known. Another aspect of VEGFRendocytosis that has not yet been determined is the mechanism bywhich it is triggered by ephrin-B2-EphB4 signaling.
VEGFR and other RTKs exploit the sorting mechanisms of theendosomal system to achieve signaling specificity. The early endo-somes appears to be the major sorting center for RTK signaling(reviewed in [81]), and the site wherefrom the receptors signal inthe cytoplasm. The sorting tools employed by VEGFR signaling in-clude Rab GTPases [60] and tyrosine phosphorylation [52]. Althoughthere is evidence for the involvement of ubiquitin ligases andubiquitin-adaptor proteins [82–84,143], this is probably only a partof the role of ubiquitination in regulation of VEGFR membrane trafficand signaling. Similar to EGFR, it is likely that VEGFR is regulated bymono-ubiquitination of lysines in its cytoplasmic domain, and byubiquitin-binding proteins such as epsin-1 and EPS15. Importantly,the number of animal models that addressed directly the role ofmembrane traffic in VEGF signaling is relatively small [17–19,58].Hence, the impact of membrane traffic on the full spectrum of VEGFsignaling remains unknown.
References
[1] J.E. Murphy, B.E. Padilla, B. Hasdemir, G.S. Cottrell, N.W. Bunnett, Proceedings ofthe National Academy of Sciences of the United States of America 106 (42)(2009) 17,615–17,622.
[2] Y. Zwang, Y. Yarden, Traffic 10 (4) (2009) 349–363.[3] S. Mukherjee, M. Tessema, A. Wandinger-Ness, Circulation Research 98 (6)
(2006) 743–756.[4] M. Santambrogio, D. Valdembri, G. Serini, Molecular Aspects of Medicine 32 (2)
(2011) 112–122.[5] H.T. Haigler, J.A. McKanna, S. Cohen, The Journal of Cell Biology 81 (2) (1979)
382–395.[6] A.V. Vieira, C. Lamaze, S.L. Schmid, Science 274 (5295) (1996) 2086–2089.[7] G.M. Di Guglielmo, P.C. Baass, W.J. Ou, B.I. Posner, J.J. Bergeron, The EMBO Jour-
nal 13 (18) (1994) 4269–4277.[8] W.H. Lai, P.H. Cameron, J.J. Doherty II, B.I. Posner, J.J. Bergeron, The Journal of Cell
Biology 109 (6 Pt 1) (1989) 2751–2760.[9] A. De Donatis, G. Comito, F. Buricchi, M.C. Vinci, A. Parenti, A. Caselli, G. Camici,
G. Manao, G. Ramponi, P. Cirri, The Journal of Biological Chemistry 283 (29)(2008) 19,948–19,956.
[10] Y. Wang, S.D. Pennock, X. Chen, A. Kazlauskas, Z. Wang, The Journal of BiologicalChemistry 279 (9) (2004) 8038–8046.
[11] K. Kawada, G. Upadhyay, S. Ferandon, S. Janarthanan, M. Hall, J.P. Vilardaga,V. Yajnik, Molecular and Cellular Biology 29 (16) (2009) 4508–4518.
[12] D.M. Bryant, F.G. Wylie, J.L. Stow, Molecular Biology of the Cell 16 (1) (2005)14–23.
[13] B. Hausott, A. Rietzler, N. Vallant, M. Auer, I. Haller, S. Perkhofer, L. Klimaschewski,Neuroscience 188 (2011) 13–22.
[14] E. Tkachenko, E. Lutgens, R.V. Stan, M. Simons, Journal of Cell Science 117 (Pt 15)(2004) 3189–3199.
[15] G. Jekely, H.H. Sung, C.M. Luque, P. Rorth, Developmental Cell 9 (2) (2005)197–207.
[16] M.G. Lampugnani, F. Orsenigo, M.C. Gagliani, C. Tacchetti, E. Dejana, The Journalof Cell Biology 174 (4) (2006) 593–604.
[17] A.A. Lanahan, K. Hermans, F. Claes, J.S. Kerley-Hamilton, Z.W. Zhuang, F.J. Giordano,P. Carmeliet, M. Simons, Developmental Cell 18 (5) (2010) 713–724.
[18] S. Sawamiphak, S. Seidel, C.L. Essmann, G.A. Wilkinson, M.E. Pitulescu, T. Acker,A. Acker-Palmer, Nature 465 (7297) (2010) 487–491.
[19] Y. Wang, M. Nakayama, M.E. Pitulescu, T.S. Schmidt, M.L. Bochenek, A. Sakakibara,S. Adams, A. Davy, U. Deutsch, U. Luthi, A. Barberis, L.E. Benjamin, T. Makinen,C.D. Nobes, R.H. Adams, Nature 465 (7297) (2010) 483–486.
[20] M. Narazaki, G. Tosato, Blood 107 (10) (2006) 3892–3901.[21] A. Salikhova, L. Wang, A.A. Lanahan, M. Liu, M. Simons, W.P. Leenders,
D. Mukhopadhyay, A. Horowitz, Circulation Research 103 (6) (2008) e71–e79.[22] D. Valdembri, P.T. Caswell, K.I. Anderson, J.P. Schwarz, I. Konig, E. Astanina,
F. Caccavari, J.C. Norman, M.J. Humphries, F. Bussolino, G. Serini, PLoS Biology7 (1) (2009) e25.
[23] J.M. Backer, C.R. Kahn, M.F. White, The Journal of Biological Chemistry 264 (3)(1989) 1694–1701.
[24] J.C. Chow, G. Condorelli, R.J. Smith, The Journal of Biological Chemistry 273 (8)(1998) 4672–4680.
[25] K.L. Kelly, N.B. Ruderman, The Journal of Biological Chemistry 268 (6) (1993)4391–4398.
PipTyr-VEGFR2
transmembraneprotein
Endocytic vesicle
Clathrinscaffold
Rab13
MICAL-L2Syx
Amot
RhoA•GDP
Grb2Mupp1
Endocytic vesicle
RhoA•GTP
Actinin-4
A
B
C
Fig. 7. Hypothetical mechanism of Syx and RhoA trafficking from cell junctions to theleading edge. Step A: VEGF-A binding to VEGFR2 triggers clathrin-dependent endocy-tosis of junctional transmembrane proteins. Step B: Amot targets the Syx-associatedcomplex, containing also Mupp1 and RhoA, to the cytoplasmic leaflet of uncoatedendocytic vesicles enclosing the endocytosed junctional transmembrane proteins.Step C: Rab13 associates with and mediates the trafficking of these vesicles. It recruitsGrb2, which targets the Rab13-associated vesicles to Tyr-phosphorylated VEGFR2. Syxactivates RhoA co-trafficking on the same vesicles, at the leading edge of the migratingcell.This figure was originally published in the Journal of Biological Chemistry by Wu et al.[58]. The American Society for Biochemistry and Molecular Biology©.
1818 A. Horowitz, H.R. Seerapu / Cellular Signalling 24 (2012) 1810–1820

Author's personal copy
[26] B. Kublaoui, J. Lee, P.F. Pilch, The Journal of Biological Chemistry 270 (1) (1995)59–65.
[27] D.E. Hammond, S. Carter, J. McCullough, S. Urbe, G. Vande Woude, M.J. Clague,Molecular Biology of the Cell 14 (4) (2003) 1346–1354.
[28] F. Mann, E. Miranda, C. Weinl, E. Harmer, C.E. Holt, Journal of Neurobiology 57(3) (2003) 323–336.
[29] M. Zimmer, A. Palmer, J. Kohler, R. Klein, Nature Cell Biology 5 (10) (2003)869–878.
[30] M.L. Grimes, J. Zhou, E.C. Beattie, E.C. Yuen, D.E. Hall, J.S. Valletta, K.S. Topp,J.H. LaVail, N.W. Bunnett, W.C. Mobley, The Journal of Neuroscience 16 (24)(1996) 7950–7964.
[31] R. Avraham, Y. Yarden, Nature Reviews. Molecular Cell Biology 12 (2) (2011)104–117.
[32] A.F. Bruns, L. Bao, J.H. Walker, S. Ponnambalam, Biochemical Society Transac-tions 37 (Pt 6) (2009) 1193–1197.
[33] A. Sorkin, L.K. Goh, Experimental Cell Research 314 (17) (2008) 3093–3106.[34] T. Yamazaki, K. Zaal, D. Hailey, J. Presley, J. Lippincott-Schwartz, L.E. Samelson,
Journal of Cell Science 115 (Pt 9) (2002) 1791–1802.[35] C. Lamaze, S.L. Schmid, The Journal of Cell Biology 129 (1) (1995) 47–54.[36] A.M. Honegger, T.J. Dull, S. Felder, E. Van Obberghen, F. Bellot, D. Szapary,
A. Schmidt, A. Ullrich, J. Schlessinger, Cell 51 (2) (1987) 199–209.[37] L.K. Goh, F. Huang, W. Kim, S. Gygi, A. Sorkin, The Journal of Cell Biology 189 (5)
(2010) 871–883.[38] G. D'Angelo, J.F. Martini, T. Iiri, W.J. Fantl, J. Martial, R.I. Weiner, Molecular Endo-
crinology 13 (5) (1999) 692–704.[39] A. Sorkin, M. von Zastrow, Nature Reviews. Molecular Cell Biology 10 (9) (2009)
609–622.[40] B. Blagoev, I. Kratchmarova, S.E. Ong, M. Nielsen, L.J. Foster, M. Mann, Nature
Biotechnology 21 (3) (2003) 315–318.[41] L.E. Johannessen, N.M. Pedersen, K.W. Pedersen, I.H. Madshus, E. Stang, Molecu-
lar and Cellular Biology 26 (2) (2006) 389–401.[42] M. Kazazic, V. Bertelsen, K.W. Pedersen, T.T. Vuong, M.V. Grandal, M.S. Rodland,
L.M. Traub, E. Stang, I.H. Madshus, Traffic 10 (2) (2009) 235–245.[43] A.D. Sorkin, L.V. Teslenko, N.N. Nikolsky, Experimental Cell Research 175 (1)
(1988) 192–205.[44] A. Sorkin, G. Carpenter, The Journal of Biological Chemistry 266 (34) (1991)
23,453–23,460.[45] A. Sorkin, E. Kornilova, L. Teslenko, A. Sorokin, N. Nikolsky, Biochimica et Bio-
physica Acta 1011 (1) (1989) 88–96.[46] A. Sorkin, S. Krolenko, N. Kudrjavtceva, J. Lazebnik, L. Teslenko, A.M. Soderquist,
N. Nikolsky, The Journal of Cell Biology 112 (1) (1991) 55–63.[47] C.M. Stoscheck, G. Carpenter, The Journal of Cell Biology 98 (3) (1984)
1048–1053.[48] N. Ferrara, H.P. Gerber, J. LeCouter, Nature Medicine 9 (6) (2003) 669–676.[49] C. Ruhrberg, BioEssays 25 (11) (2003) 1052–1060.[50] S. Kobayashi, A. Sawano, Y. Nojima, M. Shibuya, Y. Maru, The FASEB Journal 18
(7) (2004) 929–931.[51] M. Dougher, B.I. Terman, Oncogene 18 (8) (1999) 1619–1627.[52] L.C. Ewan, H.M. Jopling, H. Jia, S. Mittar, A. Bagherzadeh, G.J. Howell, J.H. Walker,
I.C. Zachary, S. Ponnambalam, Traffic 7 (9) (2006) 1270–1282.[53] A. Gampel, L. Moss, M.C. Jones, V. Brunton, J.C. Norman, H. Mellor, Blood 108 (8)
(2006) 2624–2631.[54] K. Ballmer-Hofer, A.E. Andersson, L.E. Ratcliffe, P. Berger, Blood 118 (3) (2011)
816–826.[55] H.M. Jopling, G.J. Howell, N. Gamper, S. Ponnambalam, Biochemical and Bio-
physical Research Communications 410 (2) (2011) 170–176.[56] J. Kroll, J. Waltenberger, The Journal of Biological Chemistry 272 (51) (1997)
32,521–32,527.[57] M. Laramee, C. Chabot, M. Cloutier, R. Stenne, M. Holgado-Madruga, A.J. Wong,
I. Royal, The Journal of Biological Chemistry 282 (11) (2007) 7758–7769.[58] C. Wu, S. Agrawal, A. Vasanji, J. Drazba, S. Sarkaria, J. Xie, C.M. Welsch, B. Anand-
Apte, A. Horowitz, The Journal of Biological Chemistry 286 (2011) 23511–23520.[59] M.A. Barbieri, R.L. Roberts, A. Gumusboga, H. Highfield, C. Alvarez-Dominguez,
A. Wells, P.D. Stahl, The Journal of Cell Biology 151 (3) (2000) 539–550.[60] H.M. Jopling, A.F. Odell, N.M. Hooper, I.C. Zachary, J.H. Walker, S. Ponnambalam,
Arteriosclerosis, Thrombosis, and Vascular Biology 29 (7) (2009) 1119–1124.[61] H. Stenmark, Nature Reviews. Molecular Cell Biology 10 (8) (2009) 513–525.[62] S. Cebe Suarez, M. Pieren, L. Cariolato, S. Arn, U. Hoffmann, A. Bogucki, C. Manlius,
J. Wood, K. Ballmer-Hofer, Cellular and Molecular Life Sciences 63 (17) (2006)2067–2077.
[63] H. Cai, R.R. Reed, The Journal of Neuroscience 19 (15) (1999) 6519–6527.[64] S.N. Naccache, T. Hasson, A. Horowitz, Proceedings of the National Academy of
Sciences of the United States of America 103 (34) (2006) 12,735–12,740.[65] L. Aschenbrenner, T. Lee, T. Hasson, Molecular Biology of the Cell 14 (7) (2003)
2728–2743.[66] T. Varsano, M.Q. Dong, I. Niesman, H. Gacula, X. Lou, T. Ma, J.R. Testa, J.R. Yates III,
M.G. Farquhar, Molecular and Cellular Biology 26 (23) (2006) 8942–8952.[67] T. Karpanen, C.A. Heckman, S. Keskitalo, M. Jeltsch, H. Ollila, G. Neufeld,
L. Tamagnone, K. Alitalo, The FASEB Journal 20 (9) (2006) 1462–1472.[68] S. Koch, S. Tugues, X. Li, L. Gualandi, L. Claesson-Welsh, The Biochemical Journal
437 (2) (2011) 169–183.[69] I. Anderie, I. Schulz, A. Schmid, Cellular Signalling 19 (3) (2007) 582–592.[70] E.M. Haugsten, J. Malecki, S.M. Bjorklund, S. Olsnes, J. Wesche, Molecular Biology
of the Cell 19 (8) (2008) 3390–3403.[71] A.F. Bruns, S.P. Herbert, A.F. Odell, H.M. Jopling, N.M. Hooper, I.C. Zachary,
J.H. Walker, S. Ponnambalam, Traffic 11 (1) (2010) 161–174.
[72] T.T. Chen, A. Luque, S. Lee, S.M. Anderson, T. Segura, M.L. Iruela-Arispe, The Jour-nal of Cell Biology 188 (4) (2010) 595–609.
[73] L.K. Hasseine, J. Murdaca, F. Suavet, S. Longnus, S. Giorgetti-Peraldi, E. VanObberghen, Experimental Cell Research 313 (9) (2007) 1927–1942.
[74] S. Rousseau, F. Houle, J. Landry, J. Huot, Oncogene 15 (18) (1997) 2169–2177.[75] Y. He, H. Zhang, L. Yu, M. Gunel, T.J. Boggon, H. Chen, W. Min, Science Signaling 3
(116) (2010) ra26.[76] M. Czubayko, P. Knauth, T. Schluter, V. Florian, R. Bohnensack, Biochemical and
Biophysical Research Communications 345 (3) (2006) 1264–1272.[77] P. van Kerkhof, J. Lee, L. McCormick, E. Tetrault, W. Lu, M. Schoenfish, V. Oorschot,
G.J. Strous, J. Klumperman, G. Bu, The EMBO Journal 24 (16) (2005) 2851–2861.[78] V. Manickam, A. Tiwari, J.J. Jung, R. Bhattacharya, A. Goel, D. Mukhopadhyay,
A. Choudhury, Blood 117 (4) (2011) 1425–1435.[79] A. Choudhury, D.L. Marks, K.M. Proctor, G.W. Gould, R.E. Pagano, Nature Cell Bi-
ology 8 (4) (2006) 317–328.[80] S. Lee, T.T. Chen, C.L. Barber, M.C. Jordan, J. Murdock, S. Desai, N. Ferrara, A. Nagy,
K.P. Roos, M.L. Iruela-Arispe, Cell 130 (4) (2007) 691–703.[81] S. Sigismund, S. Confalonieri, A. Ciliberto, S. Polo, G. Scita, P.P. Di Fiore, Physio-
logical Reviews 92 (1) (2012) 273–366.[82] J. Murdaca, C. Treins, M.N. Monthouel-Kartmann, R. Pontier-Bres, S. Kumar,
E. Van Obberghen, S. Giorgetti-Peraldi, The Journal of Biological Chemistry279 (25) (2004) 26,754–26,761.
[83] S. Giorgetti-Peraldi, J. Murdaca, J.C. Mas, E. Van Obberghen, Oncogene 20 (30)(2001) 3959–3968.
[84] A.J. Singh, R.D. Meyer, G. Navruzbekov, R. Shelke, L. Duan, H. Band, S.E. Leeman,N. Rahimi, Proceedings of the National Academy of Sciences of the United Statesof America 104 (13) (2007) 5413–5418.
[85] R.D. Meyer, S. Srinivasan, A.J. Singh, J.E. Mahoney, K.R. Gharahassanlou, N. Rahimi,Molecular and Cellular Biology 31 (10) (2011) 2010–2025.
[86] K. Haglund, S. Sigismund, S. Polo, I. Szymkiewicz, P.P. Di Fiore, I. Dikic, NatureCell Biology 5 (5) (2003) 461–466.
[87] M.E. Pitulescu, R.H. Adams, Genes & Development 24 (22) (2010) 2480–2492.[88] H.U. Wang, Z.F. Chen, D.J. Anderson, Cell 93 (5) (1998) 741–753.[89] S.S. Gerety,H.U.Wang, Z.F. Chen, D.J. Anderson,Molecular Cell 4 (3) (1999) 403–414.[90] G. Meyer, F. Varoqueaux, A. Neeb, M. Oschlies, N. Brose, Neuropharmacology 47
(5) (2004) 724–733.[91] Q. Pan, Y. Chanthery, W.C. Liang, S. Stawicki, J. Mak, N. Rathore, R.K. Tong,
J. Kowalski, S.F. Yee, G. Pacheco, S. Ross, Z. Cheng, J. Le Couter, G. Plowman,F. Peale, A.W. Koch, Y. Wu, A. Bagri, M. Tessier-Lavigne, R.J. Watts, Cancer Cell11 (1) (2007) 53–67.
[92] S. Soker, S. Takashima, H.Q. Miao, G. Neufeld, M. Klagsbrun, Cell 92 (6) (1998)735–745.
[93] C. Prahst, M. Heroult, A.A. Lanahan, N. Uziel, O. Kessler, N. Shraga-Heled,M. Simons,G. Neufeld, H.G. Augustin, The Journal of Biological Chemistry 283 (37) (2008)25,110–25,114.
[94] Y. Gao, M. Li, W. Chen, M. Simons, Journal of Cellular Physiology 184 (3) (2000)373–379.
[95] F. Jeanneteau, J. Diaz, P. Sokoloff, N. Griffon, Molecular Biology of the Cell 15 (2)(2004) 696–705.
[96] L. Wang, H. Zeng, P. Wang, S. Soker, D. Mukhopadhyay, The Journal of BiologicalChemistry 278 (49) (2003) 48,848–48,860.
[97] R. Soares, S. Guo, F. Gartner, F.C. Schmitt, J. Russo, Angiogenesis 6 (4) (2003)271–281.
[98] L. Wang, D. Mukhopadhyay, X. Xu, The FASEB Journal 20 (9) (2006) 1513–1515.[99] I. Bosanac, I.E. Wertz, B. Pan, C. Yu, S. Kusam, C. Lam, L. Phu, Q. Phung, B. Maurer,
D. Arnott, D.S. Kirkpatrick, V.M. Dixit, S.G. Hymowitz, Molecular Cell 40 (4)(2010) 548–557.
[100] A. Bechara, H. Nawabi, F. Moret, A. Yaron, E. Weaver, M. Bozon, K. Abouzid,J.L. Guan, M. Tessier-Lavigne, V. Lemmon, V. Castellani, The EMBO Journal 27(11) (2008) 1549–1562.
[101] S.X. Jiang, S. Whitehead, A. Aylsworth, J. Slinn, B. Zurakowski, K. Chan, J. Li,S.T. Hou, The Journal of Biological Chemistry 285 (13) (2010) 9908–9918.
[102] B. Herzog, C. Pellet-Many, G. Britton, B. Hartzoulakis, I.C. Zachary, Molecular Bi-ology of the Cell 22 (15) (2011) 2766–2776.
[103] H. Avraham, S.Y. Park, K. Schinkmann, S. Avraham, Cellular Signalling 12 (3)(2000) 123–133.
[104] A. Fantin, Q. Schwarz, K. Davidson, E.M. Normando, L. Denti, C. Ruhrberg, Devel-opment (Cambridge, England) 138 (19) (2011) 4185–4191.
[105] P.M. Becker, J. Waltenberger, R. Yachechko, T. Mirzapoiazova, J.S. Sham, C.G. Lee,J.A. Elias, A.D. Verin, Circulation Research 96 (12) (2005) 1257–1265.
[106] M. Murga, O. Fernandez-Capetillo, G. Tosato, Blood 105 (5) (2005) 1992–1999.[107] H.L. Goel, B. Pursell, C. Standley, K. Fogarty, A.M. Mercurio, Journal of Cell Science
125 (Pt 2) (2012) 497–506.[108] J. Gavard, J.S. Gutkind, Nature Cell Biology 8 (11) (2006) 1223–1234.[109] T. Murakami, E.A. Felinski, D.A. Antonetti, The Journal of Biological Chemistry
284 (31) (2009) 21,036–21,046.[110] L. di Blasio, S. Droetto, J. Norman, F. Bussolino, L. Primo, Traffic 11 (8) (2010)
1107–1118.[111] R.A. Alexander, G.W. Prager, J. Mihaly-Bison, P. Uhrin, S. Sunzenauer, B.R. Binder,
G.J. Schutz, M. Freissmuth, J.M. Breuss, Cardiovascular Research 94 (1) (2012)125–135.
[112] G. Eden, M. Archinti, F. Furlan, R. Murphy, B. Degryse, Current PharmaceuticalDesign 17 (19) (2011) 1874–1889.
[113] S. Weis, S. Shintani, A. Weber, R. Kirchmair, M. Wood, A. Cravens, H. McSharry,A. Iwakura, Y.S. Yoon, N. Himes, D. Burstein, J. Doukas, R. Soll, D. Losordo, D. Cheresh,The Journal of Clinical Investigation 113 (6) (2004) 885–894.
1819A. Horowitz, H.R. Seerapu / Cellular Signalling 24 (2012) 1810–1820

Author's personal copy
[114] P. Carmeliet, M.G. Lampugnani, L. Moons, F. Breviario, V. Compernolle, F. Bono,G. Balconi, R. Spagnuolo, B. Oosthuyse, M. Dewerchin, A. Zanetti, A. Angellilo,V. Mattot, D. Nuyens, E. Lutgens, F. Clotman, M.C. de Ruiter, A. Gittenberger-deGroot, R. Poelmann, F. Lupu, J.M. Herbert, D. Collen, E. Dejana, Cell 98 (2) (1999)147–157.
[115] M. Grazia Lampugnani, A. Zanetti, M. Corada, T. Takahashi, G. Balconi, F. Breviario,F. Orsenigo, A. Cattelino, R. Kemler, T.O. Daniel, E. Dejana, The Journal of CellBiology 161 (4) (2003) 793–804.
[116] A.K. Shukla, K. Xiao, R.J. Lefkowitz, Trends in Biochemical Sciences 36 (9) (2011)457–469.
[117] S. Almagro, C. Durmort, A. Chervin-Petinot, S. Heyraud, M. Dubois, O. Lambert,C. Maillefaud, E. Hewat, J.P. Schaal, P. Huber, D. Gulino-Debrac, Molecular andCellular Biology 30 (7) (2010) 1703–1717.
[118] Y. Wallez, F. Cand, F. Cruzalegui, C. Wernstedt, S. Souchelnytskyi, I. Vilgrain,P. Huber, Oncogene 26 (7) (2007) 1067–1077.
[119] V. Dodane, B. Kachar, The Journal of Membrane Biology 149 (3) (1996) 199–209.[120] C. Le Roy, J.L. Wrana, Nature Reviews. Molecular Cell Biology 6 (2) (2005)
112–126.[121] J.M. Sundstrom, B.R. Tash, T. Murakami, J.M. Flanagan, M.C. Bewley, B.A. Stanley,
K.B. Gonsar, D.A. Antonetti, Journal of Proteome Research 8 (2) (2009) 808–817.[122] M. Furuse, Cold Spring Harbor Perspectives in Biology 2 (1) (2010) a002907.[123] P.T. Caswell, S. Vadrevu, J.C. Norman, Nature Reviews. Molecular Cell Biology 10
(12) (2009) 843–853.[124] J.D. Humphries, A. Byron, M.J. Humphries, Journal of Cell Science 119 (Pt 19)
(2006) 3901–3903.[125] R. Soldi, S. Mitola, M. Strasly, P. Defilippi, G. Tarone, F. Bussolino, The EMBO Jour-
nal 18 (4) (1999) 882–892.[126] C. Wong, Z.G. Jin, The Journal of Biological Chemistry 280 (39) (2005)
33,262–33,269.[127] J. Ivaska, J. Heino, Annual Review of Cell and Developmental Biology 27 (2011)
291–320.
[128] F. Blasi, N. Sidenius, FEBS Letters 584 (9) (2010) 1923–1930.[129] M.C. Kugler, Y. Wei, H.A. Chapman, Current Pharmaceutical Design 9 (19)
(2003) 1565–1574.[130] R.P. Czekay, T.A. Kuemmel, R.A. Orlando, M.G. Farquhar, Molecular Biology of the
Cell 12 (5) (2001) 1467–1479.[131] T.V. Byzova, C.K. Goldman, N. Pampori, K.A. Thomas, A. Bett, S.J. Shattil, E.F. Plow,
Molecular Cell 6 (4) (2000) 851–860.[132] G.H. Mahabeleshwar, W. Feng, K. Reddy, E.F. Plow, T.V. Byzova, Circulation
Research 101 (6) (2007) 570–580.[133] X.Z. West, N. Meller, N.L. Malinin, L. Deshmukh, J. Meller, G.H. Mahabeleshwar,
M.E. Weber, B.A. Kerr, O. Vinogradova, T.V. Byzova, PloS One 7 (2) (2012)e31071.
[134] A. Palamidessi, E. Frittoli, M. Garre, M. Faretta, M. Mione, I. Testa, A. Diaspro,L. Lanzetti, G. Scita, P.P. Di Fiore, Cell 134 (1) (2008) 135–147.
[135] S. Morimoto, N. Nishimura, T. Terai, S. Manabe, Y. Yamamoto, W. Shinahara,H. Miyake, S. Tashiro, M. Shimada, T. Sasaki, The Journal of Biological Chemistry280 (3) (2005) 2220–2228.
[136] T. Terai, N. Nishimura, I. Kanda, N. Yasui, T. Sasaki, Molecular Biology of the Cell17 (5) (2006) 2465–2475.
[137] S. Mili, K. Moissoglu, I.G. Macara, Nature 453 (7191) (2008) 115–119.[138] B. Jeansonne, Q. Lu, D.A. Goodenough, Y.H. Chen, Cellular and Molecular Biology
(Noisy-le-Grand, France) 49 (1) (2003) 13–21.[139] Y. Hamazaki, M. Itoh, H. Sasaki, M. Furuse, S. Tsukita, The Journal of Biological
Chemistry 277 (1) (2002) 455–461.[140] M.A. Estevez, J.A. Henderson, D. Ahn, X.R. Zhu, G. Poschmann, H. Lubbert, R. Marx,
J.M. Baraban, Journal of Neurochemistry 106 (3) (2008) 1287–1297.[141] B. Heller, E. Adu-Gyamfi, W. Smith-Kinnaman, C. Babbey, M. Vora, Y. Xue,
R. Bittman, R.V. Stahelin, C.D. Wells, The Journal of Biological Chemistry 285(16) (2010) 12,308–12,320.
[142] K. Lindmo, H. Stenmark, Journal of Cell Science 119 (Pt 4) (2006) 605–614.[143] R.D. Meyer, D. Husain, N. Rahimi, Oncogene 30 (19) (2011) 2198–2206.
1820 A. Horowitz, H.R. Seerapu / Cellular Signalling 24 (2012) 1810–1820





























