Embed Size (px)
Citation preview

Rescue of the Mucocutaneous Manifestations byHuman Cord Blood Derived NonhematopoieticStem Cells in a Mouse Model of RecessiveDystrophic Epidermolysis Bullosa
YANLING LIAO,a LARISA IVANOVA,a HONGWEN ZHU,b ASHLIN YAHR,a JANET AYELLO,a
CARMELLA VAN DE VEN,a AHMED RASHAD,a JOUNI UITTO,c ANGELA M. CHRISTIANO,d
MITCHELL S. CAIROa,e,f,g,h
Key Words. Stem cell transplantation • Human cord blood stem cells • Cytotherapy • Inheriteddiseases • Skin blistering diseases
ABSTRACT
Recessive dystrophic epidermolysis bullosa (RDEB) is a severe skin blistering disease caused bymutations in COL7A1-encoding type VII collagen (C7). Currently, there is no curative therapy forpatients with RDEB. Our previous studies demonstrated that human umbilical cord blood(HUCB) derived unrestricted somatic stem cells (USSCs) express C7 and facilitate wound healingin a murine wounding model. The primary objective of this study is to investigate the therapeu-tic functions of USSCs in the C7 null (Col7a1
2/2) C57BL6/J mice, a murine model of RDEB. Wedemonstrated that intrahepatic administration of USSCs significantly improved the blisteringphenotype and enhanced the life span in the recipients. The injected USSCs trafficked to thesites of blistering and were incorporated in short-term in the recipients’ skin and gastrointesti-nal tract. Consistent with an overall histological improvement in the epidermal-dermal adher-ence following USSC treatment, the expression of C7 at the basement membrane zone wasdetected and the previously disorganized integrin a6 distribution was normalized. We also dem-onstrated that USSCs treatment induced an infiltration of macrophages with a regenerative“M2” phenotype. Our data suggest that HUCB-derived USSCs improved the RDEB phenotypethrough multiple mechanisms. This study has warranted future clinical investigation of USSCs asa novel and universal allogeneic stem cell donor source in selected patients with RDEB. STEM
CELLS 2015;33:1807–1817
INTRODUCTION
Recessive dystrophic epidermolysis bullosa(RDEB) is a severe inherited skin blistering dis-ease caused by mutations in the COL7A1 genethat encodes type VII collagen (C7) [1]. The C7protein is synthesized by both basal keratino-cytes and dermal fibroblasts and is a majorcomponent in the anchoring fibrils, an attach-ment structure that extends from the lowerportion of the basement membrane to thepapillary dermis [2, 3]. Clinically, patients withRDEB suffer from recurrent blisters in the skin,oral mucosa, gastrointestinal and genitourinarytracts [4, 5]. In time, blisters in the skin healwith scarring, resulting in severe deformities,including pseudosyndactyly [4, 5]. Moreover,patients with RDEB are at a significantly higherrisk of developing cutaneous squamous cellcarcinoma [6].
Currently, there is no curative treatmentfor patients with RDEB. Preclinical studies indi-
cated that injection of enriched congenic wild-type hematopoietic progenitor cells (HPC)significantly elongated the survival of some ofthe recipient Col7a1
2/2 (RDEB) mice [7].Furthermore, allogeneic HPC transplantations fol-lowing myeloablative chemotherapy in patientswith RDEB have increased C7 protein depositionand anchoring fibril formation at the dermal-epidermal junction [8]. However, the number andstructure of the anchoring fibrils after transplan-tation were still abnormal [9]. Interestingly, ascompared to the preclinical studies where onlyenriched hematopoietic progenitor cell therapyprolonged the survival of the Col7a12/2 mice,clinical studies demonstrated the engraftment ofboth CD451 and CD452CD312 donor cells in theskin of the patients following allogeneic HPCtransplantation [9, 10]. The identities of thesenonhematopoietic cells have not been estab-lished. However, it is likely that they contributeto the C7 expression and clinical improvementsin recipient RDEB skin.
aDepartment of Pediatrics,eDepartment of Medicine,fDepartment of Pathology,gDepartment of Immunology andMicrobiology, and hDepartmentof Cell Biology and Anatomy,New York Medical College,Valhalla, New York, USA;bDepartment of Surgery, TianjinHospital, Tianjin Academy ofIntegrative Medicine, Tianjin,People’s Republic of China;cDepartment of Dermatology andCutaneous Biology, JeffersonMedical College, Philadelphia,Pennsylvania, USA; dDepartmentof Dermatology, ColumbiaUniversity Medical Center, NewYork, New York, USA
Correspondence: Mitchell S. Cairo,M.D., Department of Pediatrics,Medicine, Pathology, Microbiology &Immunology and Cell Biology &Anatomy, Maria Fareri Children’sHospital at Westchester MedicalCenter, New York Medical College,40 Sunshine Cottage Road, Skyline-1N-D12, Valhalla, New York 10595,USA. Telephone: 914-594-2150;Fax: 914-594-2151;e-mail: [email protected] orYanling Liao, Ph.D., Department ofPediatrics, New York MedicalCollege, Vosburgh 318a, Valhalla,New York, New York 10595, USA.Telephone: 914-594-3777; Fax: 914-594-2151; E-mail: [email protected]
Received August 19, 2014;accepted for publication January6, 2015; first published online inSTEM CELLS EXPRESS March 18,2015.
VC AlphaMed Press1066-5099/2015/$30.00/0
http://dx.doi.org/10.1002/stem.1966
STEM CELLS 2015;33:1807–1817 www.StemCells.com VC AlphaMed Press 2015
REGENERATIVE MEDICINE

Recently, mesenchymal stem cells (MSCs) have been usedas third-party facilitator cell therapy following allogeneic HPCtransplantation in patients with RDEB [9]. The clinical benefitsof adding MSCs remain unclear, but they may reduce the inci-dence and/or severity of acute graft-versus-host disease [9].MSCs have been shown to express C7 protein, and intrader-mal injection of MSCs has alleviated the RDEB phenotypeboth in a RDEB animal model and in human subjects [11, 12].
We and other have demonstrated that human cord bloodis rich with stem cells and progenitors [13]. One of the cordblood-derived nonhematopoietic stem cells, unrestrictedsomatic stem cells (USSCs), appear to be an immature meso-derm progenitor cell critical in MSC development and can bedistinguished from MSCs by their higher expansion capacity,broader differentiation ability, and differential expression ofgenes including d-like 1/preadipocyte factor 1 and the HOXgene clusters [14]. USSCs constitutively express a series ofcytokines at a significantly higher level than MSCs do andhave strong hematopoietic stimulating activity [15].
Our previous studies demonstrated that HUCB-derivedUSSCs express C7 protein and are capable of differentiationinto keratinocyte-like cells in vitro. In a murine wound healingmodel, USSCs engineered to express green fluorescent protein(GFP)/luciferase reporter genes (USSC GFP/Luc) were demon-strated to migrate to the wound and promote wound healing[16]. In this study, we further identified significant therapeuticfunctions of USSCs in a murine model of RDEB.
MATERIALS AND METHODS
Mice
C57BL6/J Col7a11/2 mice were developed by targeted abla-
tion of the COL7A1 gene through out-of-frame deletion ofexons 14–18 [17]. The Col7a1
2/2 RDEB mice were generatedby breeding of the heterozygous mice, and genotype wasdetermined by polymerase chain reaction (PCR) [17]. All ani-mal studies were conducted using IACUC protocols approvedby New York Medical College Animal Use Committee.
Human Stem Cell or USSC-Conditioned MediumInjection in Col7a12/2 Mice
USSCs GFP/Luc or cord blood mononuclear cells (CB MNCs)(0.25 3 106) were resuspended in 20 ml phosphate bufferedsaline (PBS) and injected into the liver of newborn RDEB mice(D0) using 31G insulin syringes (BD, Franklin Lakes, NJ, http://www.bd.com). In the group with repeated USSC injections, asecond dose (0.25 3 106) was injected in D5–8 RDEB mice inthe liver. For intradermal injection, USSCs GFP/Luc (0.25 3 106)were injected into the dorsal skin of D0 RDEB mice. USSC con-ditioned medium (CM) (25 mg total protein in 20 mL serum freeDulbecco’s modified Eagle’s medium) was injected in the liverof the D0 RDEB mice. Kaplan-Meier analysis was used for sur-vival curves, and log-rank (Mantel-Cox) test was applied tocompare the survival between different experimental groups(GraphPad Prism 6). A p value< .05 was considered significant.
Histological and Immunocytochemical Analyses
The tissues including skin and gastrointestinal (GI) tract wereexcised from the selected mice, embedded in Tissue-Tec OCTCompound (Sakura Finetek, Torrance, CA, http://www.sakura.
eu/) and stored at 280�C freezer. Six mm serial sections werecut for each specimen. H&E staining was processed followingstandard procedures, at the Core Histology Lab of New YorkMedical College. Images were acquired using Nikon 90iEclipse microscope (Nikon Instrument, NY, http://www.niko-ninstruments.com/). To evaluate the histology of the treatedand untreated RDEB skin, the periphery and the length of theattached skin were, respectively, measured by tracing theoverall epidermis and the epidermis that was only attached tothe dermis per section (NIS-Elements Software, NikonInstrument). At least three sections that included the begin-ning, middle, and end of the serial sectioning per samplewere chosen for the calculation. Sections for histocytochemicalstaining, unless otherwise indicated, were fixed in either acetoneor 4% paraformaldehyde and blocked with M.O.M. blocking rea-gent (for antibodies raised in mouse) (Vector Laboratories,Burlingame, CA, http://www.vectorlabs.com) or 10% horse serum(Gibco, Grand Island, NY, http://www.invitrogen.com) with 0.1%Triton (Sigma, MO, http://www.sigmaaldrich.com/). The slideswere then incubated with primary and corresponding secondaryantibodies (Supporting Information Table S1) and mounted inVectashield mounting medium containing 40,6-diamidino-2-phe-nylindole (DAPI) (Vector Laboratories). The immunocytochemistrystaining with rabbit anti-COL7A1 (EMD Chemicals, NJ, http://www.emdmillipore.com/) was conducted following manufac-ture’s recommendation: the cryostat sections were incubatedwith 0.5% bovine serum albumin (BSA)/PBS for 10 minutes at42�C followed by hyaluronidase (2 mg/mL) digestion for 30minutes, before incubation with the primary and secondary anti-bodies. Additional methods can be found in the SupportingInformation data.
RESULTS
Human Cord Blood-Derived USSCs Enhanced theLifespan of the Col7a12/2 Mice and Ameliorated theBlistering Phenotype
To investigate the effects of cord blood-derived USSCs on RDEB,we injected USSCs GFP/Luc (0.25 3 106) into the liver of theCol7a12/2 mice within 24 hours of their births (D0), withoutpreconditioning or immunosuppression. Col7a1
2/2 mice exhibithemorrhagic blisters and erosions on the paws and mucousmembranes and normally die within the first few postnatal days,as described before [17]. After a single intrahepatic USSC injec-tion, the median life span of the RDEB mice was elongated to 8days (n 5 21), as compared to 2 days (n 5 18) in PBS-injectedanimals (p< .0001) (Fig. 1A). Importantly, about 30% (n 5 6) ofthe USSC-treated mice lived beyond 2 weeks of life span andone animal survived for 4 weeks. In comparison, a single intra-hepatic injection of CB-MNCs (0.25 3 106) only moderatelyenhanced the median life span of recipient mice (3.5 days).
Notably, the hemorrhagic blistering phenotype wasresolved within a few days following USSC intrahepatic injec-tion. After a week, a few red marks remained in the paws ofmost of the treated mice (Fig. 1B). Occasionally, scars wereobserved during the healing processes, but they were quicklyresolved in later days, leaving no apparent signs of scarring(Fig. 1C). Histological analysis on the footpad skin of theRDEB mice after a single intrahepatic USSC injection demon-strated that the percentage of the skin with intact epidermal-
1808 Cord Blood Stem Cell Therapy for RDEB
VC AlphaMed Press 2015 STEM CELLS
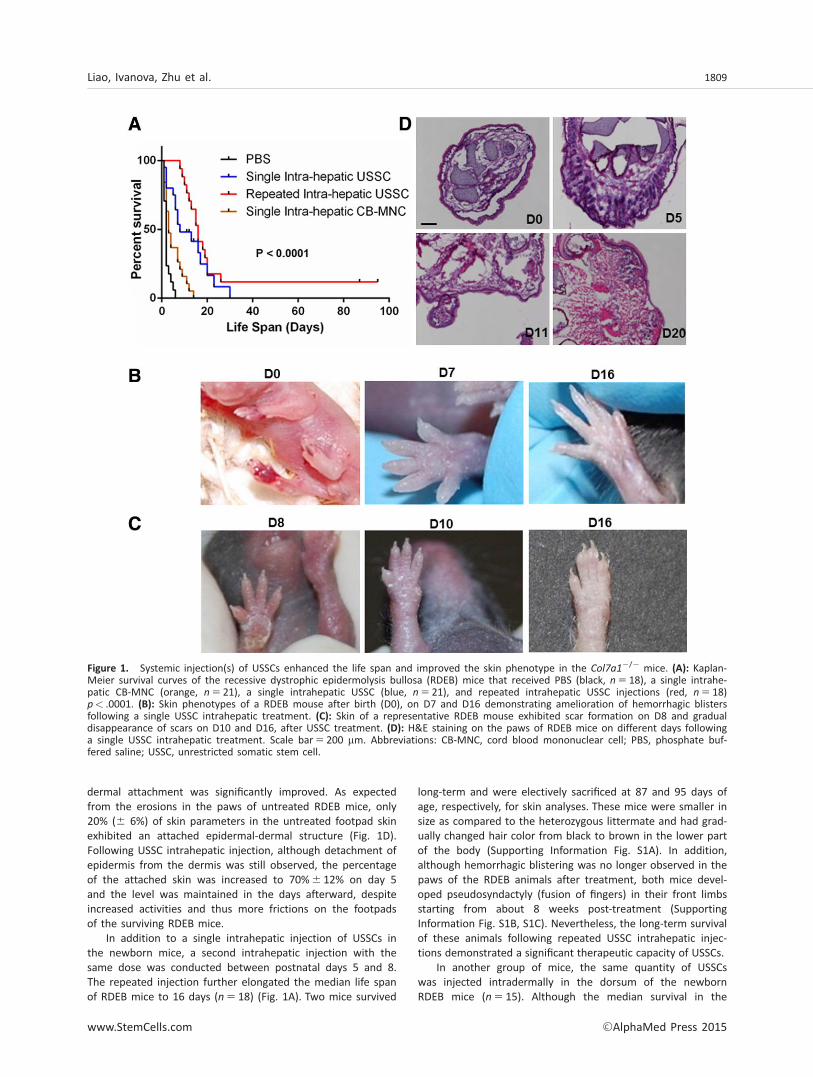
dermal attachment was significantly improved. As expectedfrom the erosions in the paws of untreated RDEB mice, only20% (6 6%) of skin parameters in the untreated footpad skinexhibited an attached epidermal-dermal structure (Fig. 1D).Following USSC intrahepatic injection, although detachment ofepidermis from the dermis was still observed, the percentageof the attached skin was increased to 70%6 12% on day 5and the level was maintained in the days afterward, despiteincreased activities and thus more frictions on the footpadsof the surviving RDEB mice.
In addition to a single intrahepatic injection of USSCs inthe newborn mice, a second intrahepatic injection with thesame dose was conducted between postnatal days 5 and 8.The repeated injection further elongated the median life spanof RDEB mice to 16 days (n 5 18) (Fig. 1A). Two mice survived
long-term and were electively sacrificed at 87 and 95 days ofage, respectively, for skin analyses. These mice were smaller insize as compared to the heterozygous littermate and had grad-ually changed hair color from black to brown in the lower partof the body (Supporting Information Fig. S1A). In addition,although hemorrhagic blistering was no longer observed in thepaws of the RDEB animals after treatment, both mice devel-oped pseudosyndactyly (fusion of fingers) in their front limbsstarting from about 8 weeks post-treatment (SupportingInformation Fig. S1B, S1C). Nevertheless, the long-term survivalof these animals following repeated USSC intrahepatic injec-tions demonstrated a significant therapeutic capacity of USSCs.
In another group of mice, the same quantity of USSCswas injected intradermally in the dorsum of the newbornRDEB mice (n 5 15). Although the median survival in the
Figure 1. Systemic injection(s) of USSCs enhanced the life span and improved the skin phenotype in the Col7a12/2 mice. (A): Kaplan-Meier survival curves of the recessive dystrophic epidermolysis bullosa (RDEB) mice that received PBS (black, n 5 18), a single intrahe-patic CB-MNC (orange, n 5 21), a single intrahepatic USSC (blue, n 5 21), and repeated intrahepatic USSC injections (red, n 5 18)p< .0001. (B): Skin phenotypes of a RDEB mouse after birth (D0), on D7 and D16 demonstrating amelioration of hemorrhagic blistersfollowing a single USSC intrahepatic treatment. (C): Skin of a representative RDEB mouse exhibited scar formation on D8 and gradualdisappearance of scars on D10 and D16, after USSC treatment. (D): H&E staining on the paws of RDEB mice on different days followinga single USSC intrahepatic treatment. Scale bar5 200 mm. Abbreviations: CB-MNC, cord blood mononuclear cell; PBS, phosphate buf-fered saline; USSC, unrestricted somatic stem cell.
Liao, Ivanova, Zhu et al. 1809
www.StemCells.com VC AlphaMed Press 2015
intradermal injection group was enhanced (4 days in intrader-mal injection group versus 2 days in PBS-injected group;p< .001), there was no substantial long term survival in therecipients (Supporting Information Fig. S2A). We further pre-pared CM from USSCs (USSC-CM) to determine whethersecreted factors from USSCs, for example C7, as confirmed byWestern blotting analysis (Supporting Information Fig. S2B)would alleviate the RDEB phenotype. However, the medianlife span of RDEB mice (n 5 18) was only elongated to 3 daysfollowing intrahepatic injection of USSC-CM (p 5 0.001)(Supporting Information Fig. S2A). Collectively, these data sug-gest that circulation of the USSC cell population was neces-sary for the extended survival of the RDEB mice.
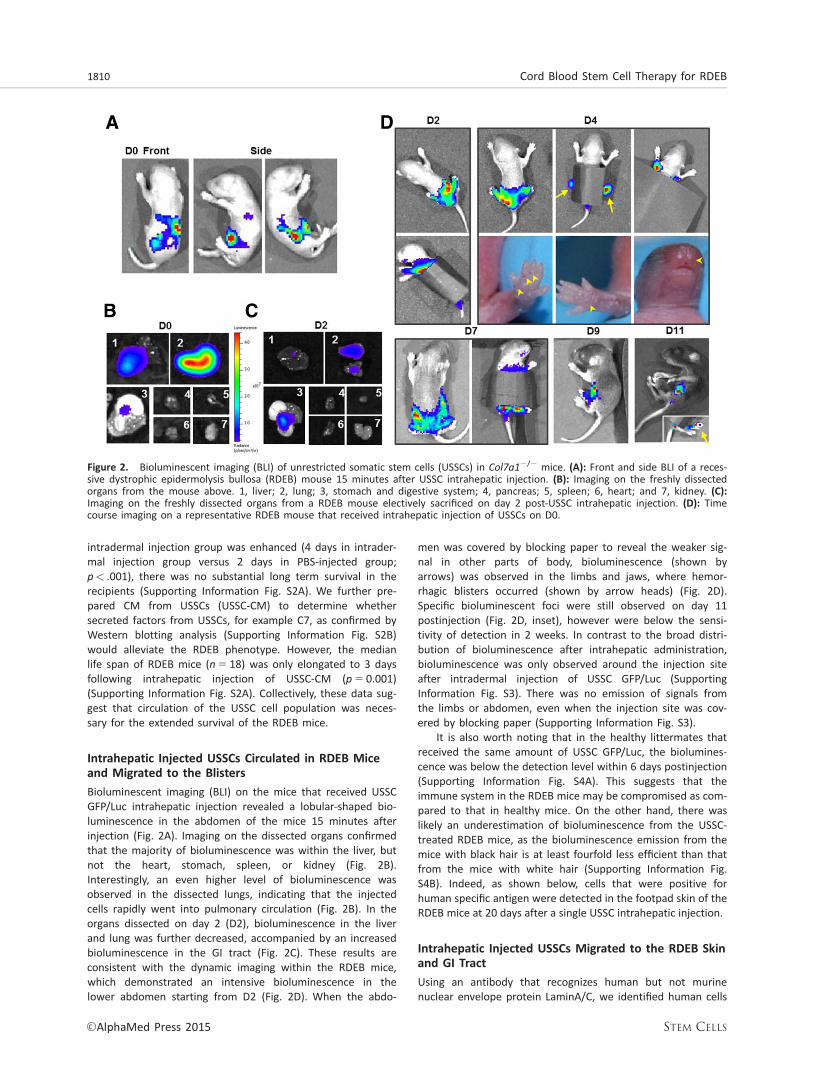
Intrahepatic Injected USSCs Circulated in RDEB Miceand Migrated to the Blisters
Bioluminescent imaging (BLI) on the mice that received USSCGFP/Luc intrahepatic injection revealed a lobular-shaped bio-luminescence in the abdomen of the mice 15 minutes afterinjection (Fig. 2A). Imaging on the dissected organs confirmedthat the majority of bioluminescence was within the liver, butnot the heart, stomach, spleen, or kidney (Fig. 2B).Interestingly, an even higher level of bioluminescence wasobserved in the dissected lungs, indicating that the injectedcells rapidly went into pulmonary circulation (Fig. 2B). In theorgans dissected on day 2 (D2), bioluminescence in the liverand lung was further decreased, accompanied by an increasedbioluminescence in the GI tract (Fig. 2C). These results areconsistent with the dynamic imaging within the RDEB mice,which demonstrated an intensive bioluminescence in thelower abdomen starting from D2 (Fig. 2D). When the abdo-
men was covered by blocking paper to reveal the weaker sig-nal in other parts of body, bioluminescence (shown byarrows) was observed in the limbs and jaws, where hemor-rhagic blisters occurred (shown by arrow heads) (Fig. 2D).Specific bioluminescent foci were still observed on day 11postinjection (Fig. 2D, inset), however were below the sensi-tivity of detection in 2 weeks. In contrast to the broad distri-bution of bioluminescence after intrahepatic administration,bioluminescence was only observed around the injection siteafter intradermal injection of USSC GFP/Luc (SupportingInformation Fig. S3). There was no emission of signals fromthe limbs or abdomen, even when the injection site was cov-ered by blocking paper (Supporting Information Fig. S3).
It is also worth noting that in the healthy littermates thatreceived the same amount of USSC GFP/Luc, the biolumines-cence was below the detection level within 6 days postinjection(Supporting Information Fig. S4A). This suggests that theimmune system in the RDEB mice may be compromised as com-pared to that in healthy mice. On the other hand, there waslikely an underestimation of bioluminescence from the USSC-treated RDEB mice, as the bioluminescence emission from themice with black hair is at least fourfold less efficient than thatfrom the mice with white hair (Supporting Information Fig.S4B). Indeed, as shown below, cells that were positive forhuman specific antigen were detected in the footpad skin of theRDEB mice at 20 days after a single USSC intrahepatic injection.
Intrahepatic Injected USSCs Migrated to the RDEB Skinand GI Tract
Using an antibody that recognizes human but not murinenuclear envelope protein LaminA/C, we identified human cells
Figure 2. Bioluminescent imaging (BLI) of unrestricted somatic stem cells (USSCs) in Col7a12/2 mice. (A): Front and side BLI of a reces-sive dystrophic epidermolysis bullosa (RDEB) mouse 15 minutes after USSC intrahepatic injection. (B): Imaging on the freshly dissectedorgans from the mouse above. 1, liver; 2, lung; 3, stomach and digestive system; 4, pancreas; 5, spleen; 6, heart; and 7, kidney. (C):Imaging on the freshly dissected organs from a RDEB mouse electively sacrificed on day 2 post-USSC intrahepatic injection. (D): Timecourse imaging on a representative RDEB mouse that received intrahepatic injection of USSCs on D0.
1810 Cord Blood Stem Cell Therapy for RDEB
VC AlphaMed Press 2015 STEM CELLS
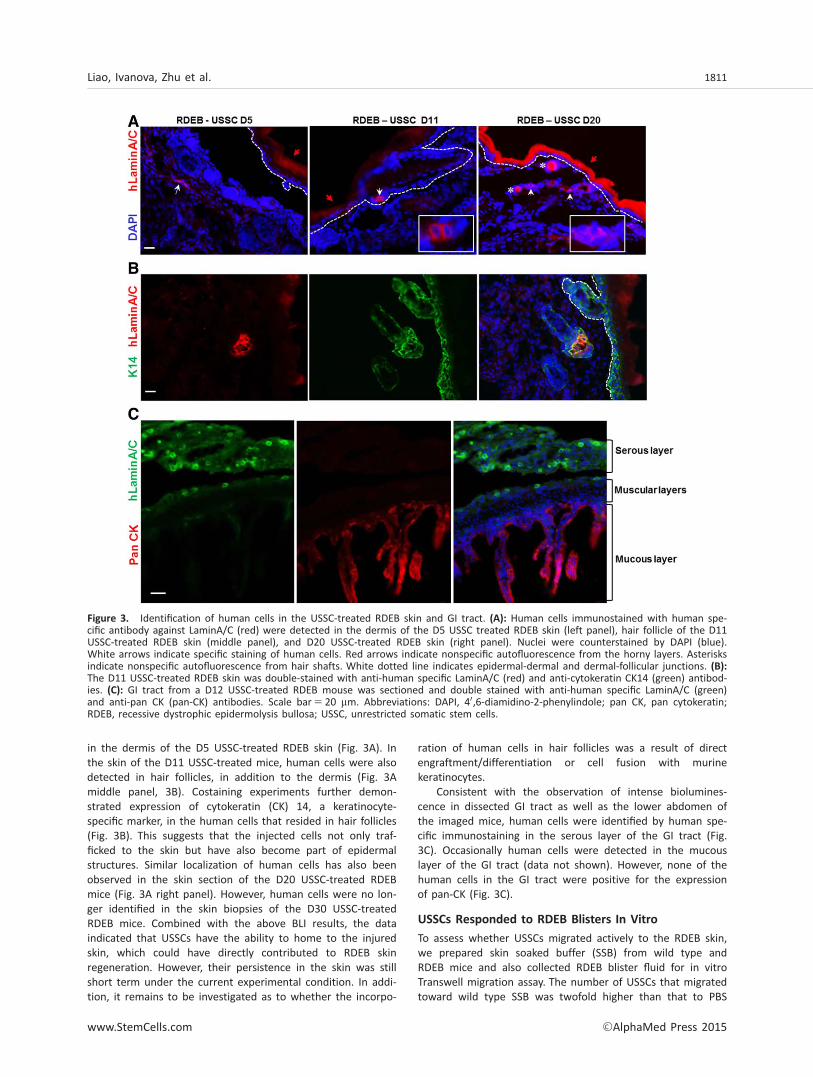
in the dermis of the D5 USSC-treated RDEB skin (Fig. 3A). Inthe skin of the D11 USSC-treated mice, human cells were alsodetected in hair follicles, in addition to the dermis (Fig. 3Amiddle panel, 3B). Costaining experiments further demon-strated expression of cytokeratin (CK) 14, a keratinocyte-specific marker, in the human cells that resided in hair follicles(Fig. 3B). This suggests that the injected cells not only traf-ficked to the skin but have also become part of epidermalstructures. Similar localization of human cells has also beenobserved in the skin section of the D20 USSC-treated RDEBmice (Fig. 3A right panel). However, human cells were no lon-ger identified in the skin biopsies of the D30 USSC-treatedRDEB mice. Combined with the above BLI results, the dataindicated that USSCs have the ability to home to the injuredskin, which could have directly contributed to RDEB skinregeneration. However, their persistence in the skin was stillshort term under the current experimental condition. In addi-tion, it remains to be investigated as to whether the incorpo-
ration of human cells in hair follicles was a result of directengraftment/differentiation or cell fusion with murinekeratinocytes.
Consistent with the observation of intense biolumines-cence in dissected GI tract as well as the lower abdomen ofthe imaged mice, human cells were identified by human spe-cific immunostaining in the serous layer of the GI tract (Fig.3C). Occasionally human cells were detected in the mucouslayer of the GI tract (data not shown). However, none of thehuman cells in the GI tract were positive for the expressionof pan-CK (Fig. 3C).
USSCs Responded to RDEB Blisters In Vitro
To assess whether USSCs migrated actively to the RDEB skin,we prepared skin soaked buffer (SSB) from wild type andRDEB mice and also collected RDEB blister fluid for in vitroTranswell migration assay. The number of USSCs that migratedtoward wild type SSB was twofold higher than that to PBS
Figure 3. Identification of human cells in the USSC-treated RDEB skin and GI tract. (A): Human cells immunostained with human spe-cific antibody against LaminA/C (red) were detected in the dermis of the D5 USSC treated RDEB skin (left panel), hair follicle of the D11USSC-treated RDEB skin (middle panel), and D20 USSC-treated RDEB skin (right panel). Nuclei were counterstained by DAPI (blue).White arrows indicate specific staining of human cells. Red arrows indicate nonspecific autofluorescence from the horny layers. Asterisksindicate nonspecific autofluorescence from hair shafts. White dotted line indicates epidermal-dermal and dermal-follicular junctions. (B):The D11 USSC-treated RDEB skin was double-stained with anti-human specific LaminA/C (red) and anti-cytokeratin CK14 (green) antibod-ies. (C): GI tract from a D12 USSC-treated RDEB mouse was sectioned and double stained with anti-human specific LaminA/C (green)and anti-pan CK (pan-CK) antibodies. Scale bar5 20 mm. Abbreviations: DAPI, 40,6-diamidino-2-phenylindole; pan CK, pan cytokeratin;RDEB, recessive dystrophic epidermolysis bullosa; USSC, unrestricted somatic stem cells.
Liao, Ivanova, Zhu et al. 1811
www.StemCells.com VC AlphaMed Press 2015

(Fig. 4). In significant contrast, the number of USSCs thatresponded to the RDEB SSB was more than 40-fold higherthan that to PBS, and there was a further threefold increasein the number of the cells that responded to the RDEB blisterfluid (Fig. 4). These data suggest that some factors specificallyin the RDEB skin and/or blister fluid are significant chemoat-tractants for USSCs. Our previous studies demonstrated thatUSSCs are positive for the expression of CXCR4, CCR7, andPDGFRa, receptors for chemoattractant proteins SDF1, CCL21,and HMGB1, respectively [16]. In a Transwell migration assay,USSCs significantly migrated to SDF1, marginally to CCL21 butrarely to HMGB1 (data not shown). Moreover, preblockingUSSCs with antibodies against CXCR4, CCR7, or PDGFRa hadno significant effects on the migratory ability of USSCs toRDEB SSB in the Transwell Assay (data not shown). It is possi-ble that the concentration of antibodies used in this assaywas not sufficient to block the specific interactions.Alternatively, combinations of these chemotactic interactionsor other novel interactions are responsible for the migrationof USSCs to the RDEB skin.
USSC-Treated RDEB Mice Exhibited C7 Expression andReorganization of Integrin a6 in the Skin
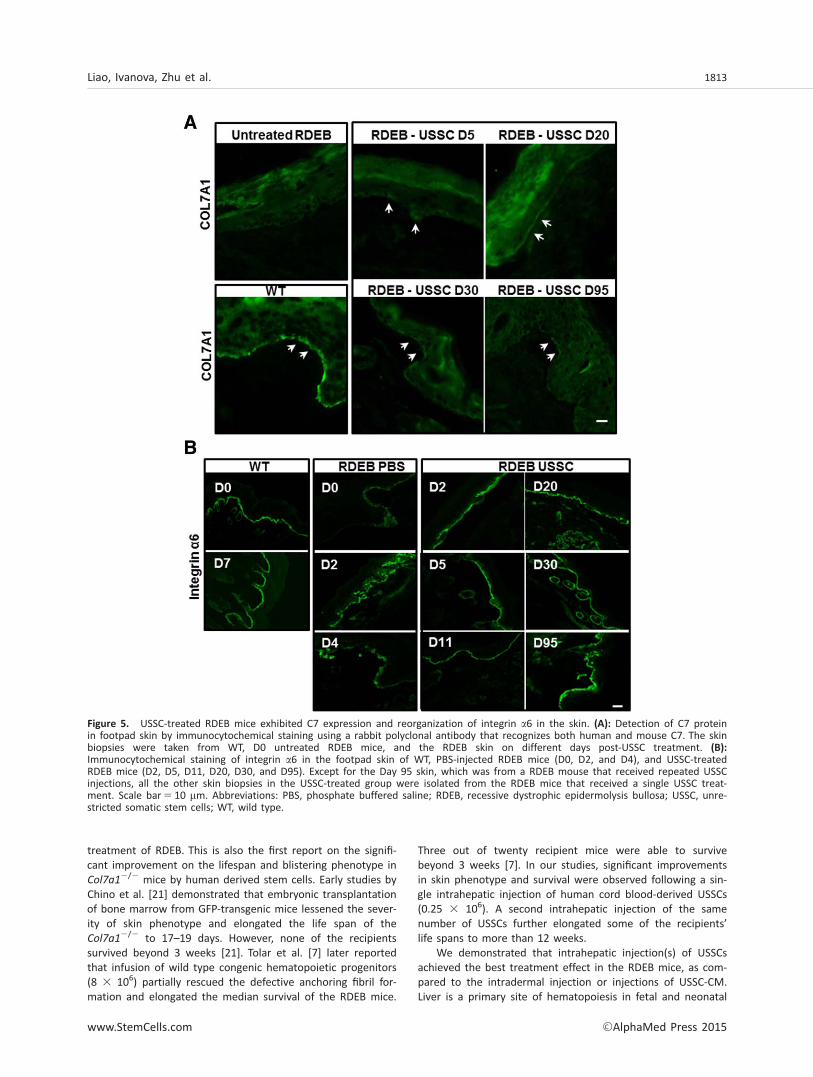
As demonstrated previously and also shown in SupportingInformation Figure S2B, USSCs express Col7A1 mRNA andsecrete C7 protein [16]. To investigate whether USSC intrahe-patic injection led to C7 expression in the affected RDEB skin,we conducted immunocytochemical staining on skin biopsiesusing rabbit polyclonal antibody that recognizes both humanand murine C7 protein. In contrast to a complete absence ofC7 in the untreated RDEB skin, C7 immunostaining started toappear in RDEB footpad skin 5 days post-USSC treatment (Fig.5A). The level of C7 staining was increased in the skin biopsies20 days post a single treatment, although still weaker than inthe wild type mice (Fig. 5A). A residual C7 was also detectedin skin biopsies 95 days post-USSC injection (Fig. 5A). The C7staining was not observed in all of the skin specimens, sug-gesting that USSC injections resulted in a partial rescue of C7protein in the recipient RDEB mice.
Nystrom et al. [18] recently reported that C7 was requiredfor re-epithelialization of wounds and its loss led to altereddistribution of laminin-332 and integrin a6b4 during wound
healing. Here, we assessed the distribution of integrin a6 inthe experimental RDEB mice. The wild-type mouse skin exhib-ited a sharp and linear expression of integrin a6 at theepidermal-dermal junction (Fig. 5B). In contrast, a broad,patchy, and suprabasal expression of integrin a6 was observedin majority of the skin from untreated newborn RDEB mice(Fig. 5B). The expression of integrin a6 appeared to beincreased, but also more disorganized, in skins from the D2and D4 PBS-injected RDEB mice. Importantly, an even and lin-ear distribution of integrin a6 resembling that in wild typewas observed in the skin of RDEB mice treated with USSCs,starting from D2 (Fig. 5B). These data suggest that USSC treat-ment increased the skin integrity of RDEB mice and normal-ized the aberrant distribution of integrin a6.
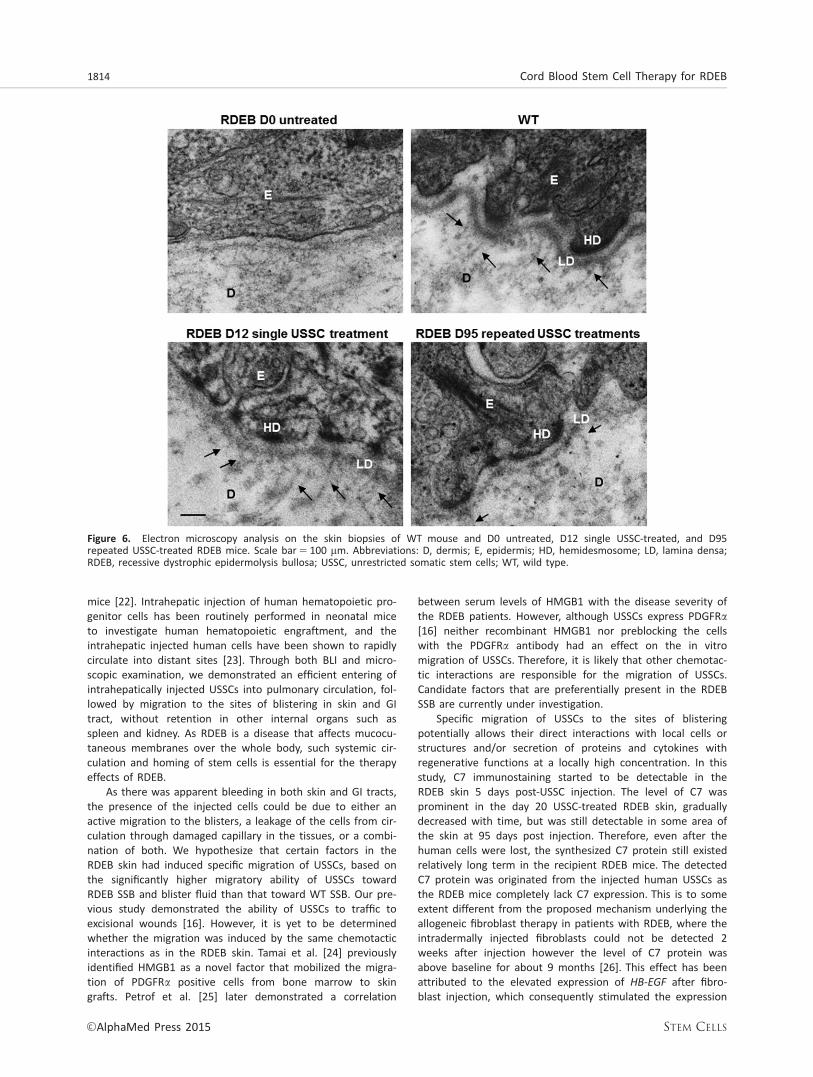
Electron microscopy of untreated RDEB mouse skin biop-sies showed a complete lack of anchoring fibrils beneath lam-ina densa, which was also poorly formed as compared to thewild-type structure (Fig. 6). In contrast, anchoring fibrils origi-nated from lamina densa were identified in the skin biopsiesfrom the D12 USSC-treated RDEB mice. The anchoring fibrils,although scanty and rudimentary, were still discernible in theskin of the D95 USSC-treated RDEB mouse.
Macrophages in the USSC-treated RDEB Skin Exhibitedan Anti-inflammatory Phenotype
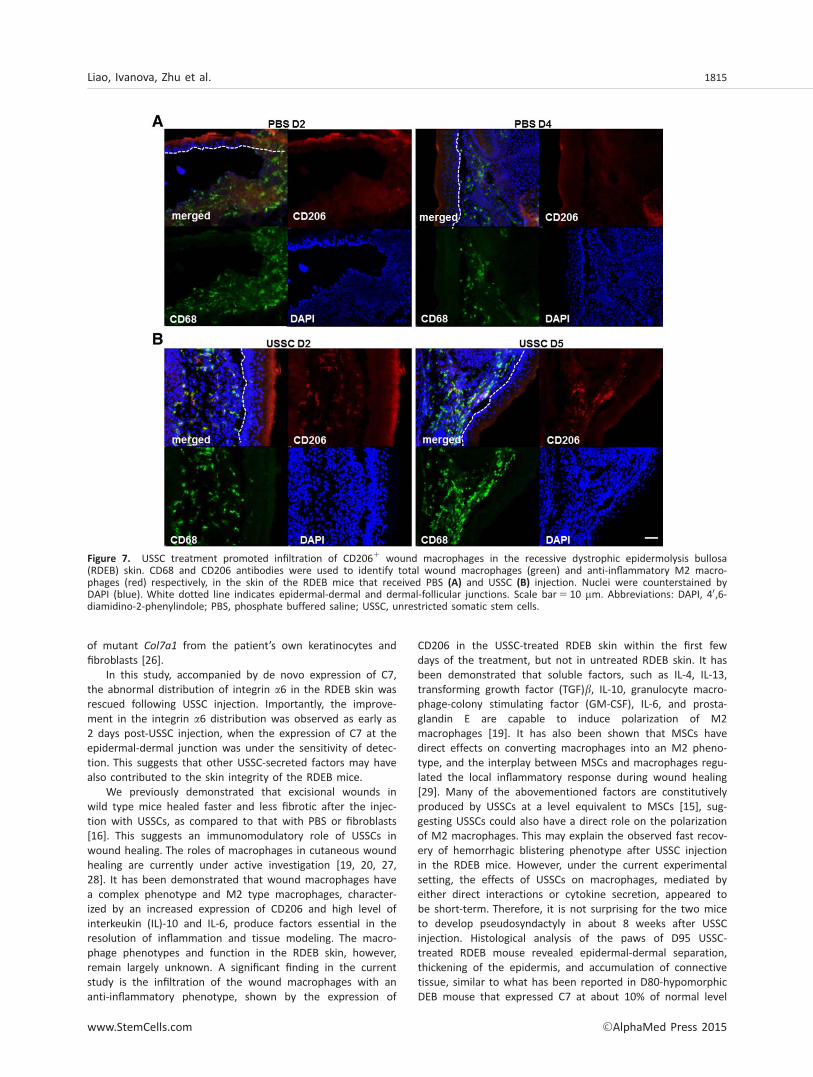
Macrophages have been known to play an important role inremoval of cellular debris from necrosis, which is usuallyloaded with endogenous danger signals including heat-shockproteins, nuclear proteins (such as HMGB1), histones, andcomponents of extracellular matrix [19]. Macrophages havealso been endowed with extraordinary phenotypic plasticity,varying their functional repertoire according to the physiologicor pathologic sites to which they are recruited. Two best char-acterized in vitro phenotypes are a proinflammatory “M1”phenotype and an anti-inflammatory “M2a” phenotype, fre-quently termed “wound healing” Mp [20]. Here, for the firsttime, we investigated the immune phenotypes of macro-phages in the RDEB mice with and without USSC intrahepaticinjection. In untreated RDEB skin, infiltration of macrophages,as demonstrated by positive CD68 staining, was observedfrom immediately after birth to the first few postnatal days(Fig. 7A). Rarely these CD681 total macrophages were stainedpositive for mannose receptor CD206, a typical cell surfacemarker for the alternatively activated M2a macrophages.Remarkably, on day 2 following USSC treatment, there was asignificant number of CD681 macrophages that were alsopositive for CD206 (Fig. 7B). Similarly, double positive stainingfor CD68 and CD206 was observed in the day 5 RDEB skintreated with USSCs. However, on day 11 and afterward, onlyCD68, but not CD206 staining, was observed in the skin.These data suggest that USSC treatment in the short termpromoted alternative activation of host macrophages in theRDEB skin, potentially contributing to the regulation of theinflammatory response and enhancing the healing of thedermis.
DISCUSSION
In this study, we reported the potential use of a novel stemcell source, that is, human cord blood-derived USSCs for the
Figure 4. In vitro cell migration assay. Actual (A) and relative (B)migration of unrestricted somatic stem cells to PBS, WT and RDEBSSB, and RDEB blister fluid. The RDEB blister fluid was diluted 1:6in PBS. Experiments were repeated three times with duplicatewells for each condition. Abbreviations: PBS, phosphate bufferedsaline; RDEB, recessive dystrophic epidermolysis bullosa; SSB, skinsoaked buffer; WT, wild type.
1812 Cord Blood Stem Cell Therapy for RDEB
VC AlphaMed Press 2015 STEM CELLS
treatment of RDEB. This is also the first report on the signifi-cant improvement on the lifespan and blistering phenotype inCol7a1
2/2 mice by human derived stem cells. Early studies byChino et al. [21] demonstrated that embryonic transplantationof bone marrow from GFP-transgenic mice lessened the sever-ity of skin phenotype and elongated the life span of theCol7a1
2/2 to 17–19 days. However, none of the recipientssurvived beyond 3 weeks [21]. Tolar et al. [7] later reportedthat infusion of wild type congenic hematopoietic progenitors(8 3 106) partially rescued the defective anchoring fibril for-mation and elongated the median survival of the RDEB mice.
Three out of twenty recipient mice were able to survivebeyond 3 weeks [7]. In our studies, significant improvementsin skin phenotype and survival were observed following a sin-gle intrahepatic injection of human cord blood-derived USSCs(0.25 3 106). A second intrahepatic injection of the samenumber of USSCs further elongated some of the recipients’life spans to more than 12 weeks.
We demonstrated that intrahepatic injection(s) of USSCsachieved the best treatment effect in the RDEB mice, as com-pared to the intradermal injection or injections of USSC-CM.Liver is a primary site of hematopoiesis in fetal and neonatal
Figure 5. USSC-treated RDEB mice exhibited C7 expression and reorganization of integrin a6 in the skin. (A): Detection of C7 proteinin footpad skin by immunocytochemical staining using a rabbit polyclonal antibody that recognizes both human and mouse C7. The skinbiopsies were taken from WT, D0 untreated RDEB mice, and the RDEB skin on different days post-USSC treatment. (B):Immunocytochemical staining of integrin a6 in the footpad skin of WT, PBS-injected RDEB mice (D0, D2, and D4), and USSC-treatedRDEB mice (D2, D5, D11, D20, D30, and D95). Except for the Day 95 skin, which was from a RDEB mouse that received repeated USSCinjections, all the other skin biopsies in the USSC-treated group were isolated from the RDEB mice that received a single USSC treat-ment. Scale bar5 10 mm. Abbreviations: PBS, phosphate buffered saline; RDEB, recessive dystrophic epidermolysis bullosa; USSC, unre-stricted somatic stem cells; WT, wild type.
Liao, Ivanova, Zhu et al. 1813
www.StemCells.com VC AlphaMed Press 2015
mice [22]. Intrahepatic injection of human hematopoietic pro-genitor cells has been routinely performed in neonatal miceto investigate human hematopoietic engraftment, and theintrahepatic injected human cells have been shown to rapidlycirculate into distant sites [23]. Through both BLI and micro-scopic examination, we demonstrated an efficient entering ofintrahepatically injected USSCs into pulmonary circulation, fol-lowed by migration to the sites of blistering in skin and GItract, without retention in other internal organs such asspleen and kidney. As RDEB is a disease that affects mucocu-taneous membranes over the whole body, such systemic cir-culation and homing of stem cells is essential for the therapyeffects of RDEB.
As there was apparent bleeding in both skin and GI tracts,the presence of the injected cells could be due to either anactive migration to the blisters, a leakage of the cells from cir-culation through damaged capillary in the tissues, or a combi-nation of both. We hypothesize that certain factors in theRDEB skin had induced specific migration of USSCs, based onthe significantly higher migratory ability of USSCs towardRDEB SSB and blister fluid than that toward WT SSB. Our pre-vious study demonstrated the ability of USSCs to traffic toexcisional wounds [16]. However, it is yet to be determinedwhether the migration was induced by the same chemotacticinteractions as in the RDEB skin. Tamai et al. [24] previouslyidentified HMGB1 as a novel factor that mobilized the migra-tion of PDGFRa positive cells from bone marrow to skingrafts. Petrof et al. [25] later demonstrated a correlation
between serum levels of HMGB1 with the disease severity ofthe RDEB patients. However, although USSCs express PDGFRa[16] neither recombinant HMGB1 nor preblocking the cellswith the PDGFRa antibody had an effect on the in vitromigration of USSCs. Therefore, it is likely that other chemotac-tic interactions are responsible for the migration of USSCs.Candidate factors that are preferentially present in the RDEBSSB are currently under investigation.
Specific migration of USSCs to the sites of blisteringpotentially allows their direct interactions with local cells orstructures and/or secretion of proteins and cytokines withregenerative functions at a locally high concentration. In thisstudy, C7 immunostaining started to be detectable in theRDEB skin 5 days post-USSC injection. The level of C7 wasprominent in the day 20 USSC-treated RDEB skin, graduallydecreased with time, but was still detectable in some area ofthe skin at 95 days post injection. Therefore, even after thehuman cells were lost, the synthesized C7 protein still existedrelatively long term in the recipient RDEB mice. The detectedC7 protein was originated from the injected human USSCs asthe RDEB mice completely lack C7 expression. This is to someextent different from the proposed mechanism underlying theallogeneic fibroblast therapy in patients with RDEB, where theintradermally injected fibroblasts could not be detected 2weeks after injection however the level of C7 protein wasabove baseline for about 9 months [26]. This effect has beenattributed to the elevated expression of HB-EGF after fibro-blast injection, which consequently stimulated the expression
Figure 6. Electron microscopy analysis on the skin biopsies of WT mouse and D0 untreated, D12 single USSC-treated, and D95repeated USSC-treated RDEB mice. Scale bar5 100 mm. Abbreviations: D, dermis; E, epidermis; HD, hemidesmosome; LD, lamina densa;RDEB, recessive dystrophic epidermolysis bullosa; USSC, unrestricted somatic stem cells; WT, wild type.
1814 Cord Blood Stem Cell Therapy for RDEB
VC AlphaMed Press 2015 STEM CELLS
of mutant Col7a1 from the patient’s own keratinocytes andfibroblasts [26].
In this study, accompanied by de novo expression of C7,the abnormal distribution of integrin a6 in the RDEB skin wasrescued following USSC injection. Importantly, the improve-ment in the integrin a6 distribution was observed as early as2 days post-USSC injection, when the expression of C7 at theepidermal-dermal junction was under the sensitivity of detec-tion. This suggests that other USSC-secreted factors may havealso contributed to the skin integrity of the RDEB mice.
We previously demonstrated that excisional wounds inwild type mice healed faster and less fibrotic after the injec-tion with USSCs, as compared to that with PBS or fibroblasts[16]. This suggests an immunomodulatory role of USSCs inwound healing. The roles of macrophages in cutaneous woundhealing are currently under active investigation [19, 20, 27,28]. It has been demonstrated that wound macrophages havea complex phenotype and M2 type macrophages, character-ized by an increased expression of CD206 and high level ofinterkeukin (IL)-10 and IL-6, produce factors essential in theresolution of inflammation and tissue modeling. The macro-phage phenotypes and function in the RDEB skin, however,remain largely unknown. A significant finding in the currentstudy is the infiltration of the wound macrophages with ananti-inflammatory phenotype, shown by the expression of
CD206 in the USSC-treated RDEB skin within the first fewdays of the treatment, but not in untreated RDEB skin. It hasbeen demonstrated that soluble factors, such as IL-4, IL-13,transforming growth factor (TGF)b, IL-10, granulocyte macro-phage-colony stimulating factor (GM-CSF), IL-6, and prosta-glandin E are capable to induce polarization of M2macrophages [19]. It has also been shown that MSCs havedirect effects on converting macrophages into an M2 pheno-type, and the interplay between MSCs and macrophages regu-lated the local inflammatory response during wound healing[29]. Many of the abovementioned factors are constitutivelyproduced by USSCs at a level equivalent to MSCs [15], sug-gesting USSCs could also have a direct role on the polarizationof M2 macrophages. This may explain the observed fast recov-ery of hemorrhagic blistering phenotype after USSC injectionin the RDEB mice. However, under the current experimentalsetting, the effects of USSCs on macrophages, mediated byeither direct interactions or cytokine secretion, appeared tobe short-term. Therefore, it is not surprising for the two miceto develop pseudosyndactyly in about 8 weeks after USSCinjection. Histological analysis of the paws of D95 USSC-treated RDEB mouse revealed epidermal-dermal separation,thickening of the epidermis, and accumulation of connectivetissue, similar to what has been reported in D80-hypomorphicDEB mouse that expressed C7 at about 10% of normal level
Figure 7. USSC treatment promoted infiltration of CD2061 wound macrophages in the recessive dystrophic epidermolysis bullosa(RDEB) skin. CD68 and CD206 antibodies were used to identify total wound macrophages (green) and anti-inflammatory M2 macro-phages (red) respectively, in the skin of the RDEB mice that received PBS (A) and USSC (B) injection. Nuclei were counterstained byDAPI (blue). White dotted line indicates epidermal-dermal and dermal-follicular junctions. Scale bar5 10 mm. Abbreviations: DAPI, 40,6-diamidino-2-phenylindole; PBS, phosphate buffered saline; USSC, unrestricted somatic stem cells.
Liao, Ivanova, Zhu et al. 1815
www.StemCells.com VC AlphaMed Press 2015

[30]. Pseudosyndactyly has been suggested to be triggered byproinflammatory and profibrotic phenotype of dermal fibro-blasts. We speculate that if the modulatory effects of USSCson macrophages could be prolonged to diminish proinflamma-tion, the development of pseudosyndactyly may be preventedor further postponed. With that, further studies are warrantedto investigate interactions between USSCs and macrophagesand identify cytokines that mediate the USSC-induced M2macrophage polarization. It would also be interesting todetermine whether intradermal injection of USSCs would simi-larly activate M2 macrophages, and whether injections ofUSSCs both systemically and locally would further potentiatethe immunomodulatory effects of USSCs.
Our previous studies demonstrated that USSCs were capa-ble of differentiation into keratinocyte-like cells in vitro [16].In the excisional wounding model, intradermal injection ofUSSCs promoted wound healing without the evidence of epi-dermal engraftment and differentiation [16]. In this study,intrahepatically injected USSCs were detected in not only thedermis of the RDEB skin but also hair follicles. Moreover, thehuman cells that resided in hair follicles were also stainedpositive for keratinocyte marker CK14. It is likely that vascularcirculation has delivered USSCs into an epidermal niche thatenables their efficient maturation and incorporation into theskin. A possibility that we cannot exclude for now is thefusion of the human cells with the mouse keratinocytes inhair follicles. Another question that needs to be addressed iswhether the human cells undergo division or proliferation invivo. In this study, the human cells were identified in theRDEB skin on and before 20 days postinjection but were nolonger detectable in the days afterward. Although our abilityto identify human cells and their niche in skin biopsies hasbeen limited by the number of tissue sections analyzed andthe positioning of the specimen (longitudinal for a betteranalysis on hair follicles), we concluded from our study thatthe incorporation of USSCs was short-term in the RDEB mice.
This study was conducted without the use of immunosup-pression in the recipient RDEB mice. It is interesting to notethat compared to the loss of bioluminescence in 2 weeks inthe USSC-treated RDEB mice, the bioluminescence disappearedas early as 6 days in the parallel USSC-injected wild type litter-mates. This is consistent with an earlier study that suggested adeficiency in cellular immunity in the patients with RDEB [31].It is possible that the compromised immune system in theRDEB mice may have provided a physiological/immunologicalwindow for USSCs to persist and function. Future studies willdetermine the effect of immunosuppression on the long-termpersistence of USSCs. Human immune reconstitution in RDEBmice that recapitulates a clinical transplantation setting couldalso be an interesting platform to assess the potential long-term effects of USSCs. Earlier animal studies demonstrated thatcotransplantation of USSCs enhances human HSC engraftment[32]. Therefore, we speculate that USSCs could exert both ther-
apeutic and adjuvant roles when combined with humanimmune reconstitution in recipient RDEB mice.
CONCLUSION
Significant progress has been made in the treatment ofpatients with RDEB; however, there clearly remains a vastunmet medical need for the development of more effectivetherapies. In this study, human cord blood-derived USSCshave demonstrated several key properties, including migrationto mucocutaneous epithelia, secretion of C7, and immunomo-dulation that support its future use as allogeneic stem cellsfor the treatment of RDEB. Currently, isolation of USSCs,clinical-scale expansion, cryopreservation, and thawing ofUSSCs can be performed in compliance with good manufac-turing practice standards [33]. Furthermore, all of thereported preclinical investigations with USSCs, including ours,have not found any tumorigenic activity from USSCs [14, 32,33]. All these studies warrant future clinical investigation ofUSSCs as a universal allogeneic stem cell source in selectedpatients with RDEB.
ACKNOWLEDGMENTS
We thank Dr. Olga Igoucheva at Jefferson Medical College forthe generation and support of Col7a11/2 mice; Dr. GlennMerlino at the National Cancer Institute for providing thepSico PolII-eGFP-Luc2 plasmid. We greatly acknowledge tech-nical support from Jen Yu and Anne Sollas for the EM and his-tology analyses, respectively, and the scientific discussionsfrom colleagues especially Dr. Christopher Leonard atDepartment of Physiology and Drs. Tiwari Aradhana andXianzheng Zhou at the Pediatric Cancer Research Laboratoryat NYMC. The authors thank Erin Morris, RN and MiguelMuniz, AAS for the assistance in the preparation of thismanuscript. This work was supported by grants from DebraInternational and the Pediatric Cancer Research Foundation.
AUTHOR CONTRIBUTIONS
Y.L.: conception and design, collection and assembly of data,data analysis and interpretation, and manuscript writing; L.I.and H.Z.: collection and assembly of data and data analysis andinterpretation; A.Y. and A.R.: collection and assembly of data;J.A. and C.v.d.V.: administrative support and data analysis andinterpretation; J.U.: provision of study material and manuscriptwriting; A.M.C.: conception and design and manuscript writing;M.S.C.: conception and design, data analysis and interpretation,manuscript writing, and final approval of manuscript.
DISCLOSURE OF POTENTIAL CONFLICTS OF INTEREST
The authors indicate no potential conflicts of interests.
REFERENCES
1 Hilal L, Rochat A, Duquesnoy P et al. Ahomozygous insertion-deletion in the typeVII collagen gene (COL7A1) in Hallopeau-Siemens dystrophic epidermolysis bullosa.Nat Genet 1993;5:287–293.
2 Bruckner-Tuderman L, McGrath JA,Robinson EC et al. Progress in epidermolysisbullosa research: Summary of DEBRAInternational Research Conference 2012.J Invest Dermatol 2013;133:2121–2126.
3 Kiuru M, Itoh M, Cairo MS et al. Bonemarrow stem cell therapy for recessive dys-
trophic epidermolysis bullosa. Dermatol Clin2010;28:371–382, xii-xiii.
4 Fine JD, Eady RA, Bauer EA et al. The classi-fication of inherited epidermolysis bullosa (EB):Report of the Third International ConsensusMeeting on Diagnosis and Classification of EB.J Am Acad Dermatol 2008;58:931–950.
1816 Cord Blood Stem Cell Therapy for RDEB
VC AlphaMed Press 2015 STEM CELLS

5 Freeman EB, Koglmeier J, Martinez AEet al. Gastrointestinal complications of epi-dermolysis bullosa in children. Br J Dermatol2008;158:1308–1314.
6 Fine JD, Johnson LB, Weiner M et al.Cause-specific risks of childhood death ininherited epidermolysis bullosa. J Pediatr2008;152:276–280.
7 Tolar J, Ishida-Yamamoto A, Riddle Met al. Amelioration of epidermolysis bullosaby transfer of wild-type bone marrow cells.Blood 2009;113:1167–1174.
8 Wagner JE, Ishida-Yamamoto A, McGrathJA et al. Bone marrow transplantation forrecessive dystrophic epidermolysis bullosa. NEngl J Med 2010;363:629–639.
9 Tolar J, Wagner JE. Allogeneic blood andbone marrow cells for the treatment ofsevere epidermolysis bullosa: Repair of theextracellular matrix. Lancet 2013;382:1214–1223.10 Tolar J, Blazar BR, Wagner JE. Concisereview: Transplantation of human hemato-poietic cells for extracellular matrix proteindeficiency in epidermolysis bullosa. StemCells 2011;29:900–906.11 Alexeev V, Uitto J, Igoucheva O. Geneexpression signatures of mouse bonemarrow-derived mesenchymal stem cells inthe cutaneous environment and therapeuticimplications for blistering skin disorder.Cytotherapy 2011;13:30–45.12 Conget P, Rodriguez F, Kramer S et al.Replenishment of type VII collagen and re-epithelialization of chronically ulcerated skinafter intradermal administration of allogeneicmesenchymal stromal cells in two patientswith recessive dystrophic epidermolysis bul-losa. Cytotherapy 2010;12:429–431.13 Liao Y, Geyer MB, Yang AJ et al. Cordblood transplantation and stem cell regenera-tive potential. Exp Hematol 2011;39:393–412.14 Kogler G, Sensken S, Airey JA et al. Anew human somatic stem cell from placentalcord blood with intrinsic pluripotent differen-tiation potential. J Exp Med 2004;200:123–135.
15 Kogler G, Radke TF, Lefort A et al.Cytokine production and hematopoiesis sup-porting activity of cord blood-derived unre-stricted somatic stem cells. Exp Hematol2005;33:573–583.16 Liao Y, Itoh M, Yang A et al. Human cordblood-derived unrestricted somatic stem cellspromote wound healing and have therapeu-tic potential for patients with recessive dys-trophic epidermolysis bullosa. Cell Transplant2014;23:303–317.17 Heinonen S, Mannikko M, Klement JFet al. Targeted inactivation of the type VIIcollagen gene (Col7a1) in mice results insevere blistering phenotype: A model forrecessive dystrophic epidermolysis bullosa.J Cell Sci 1999;112 (Pt 21):3641–3648.18 Nystrom A, Velati D, Mittapalli VR et al.Collagen VII plays a dual role in wound heal-ing. J Clin Invest 2013;123:3498–3509.19 Mosser DM, Edwards JP. Exploring thefull spectrum of macrophage activation. NatRev Immunol 2008;8:958–969.20 Novak ML, Koh TJ. Macrophage pheno-types during tissue repair. J Leukocyte Biol2013;93:875–881.21 Chino T, Tamai K, Yamazaki T et al. Bonemarrow cell transfer into fetal circulation canameliorate genetic skin diseases by providingfibroblasts to the skin and inducing immunetolerance. Am J Pathol 2008;173:803–814.22 Palis J, Robertson S, Kennedy M et al.Development of erythroid and myeloid pro-genitors in the yolk sac and embryo properof the mouse. Development 1999;126:5073–5084.23 Wulf-Goldenberg A, Keil M, Fichtner Iet al. Intrahepatic transplantation of CD341
cord blood stem cells into newborn andadult NOD/SCID mice induce differentialorgan engraftment. Tissue Cell 2012;44:80–86.24 Tamai K, Yamazaki T, Chino T et al.PDGFRalpha-positive cells in bone marroware mobilized by high mobility group box 1(HMGB1) to regenerate injured epithelia.Proc Natl Acad Sci USA 2011;108:6609–6614.
25 Petrof G, Abdul-Wahab A, Proudfoot Let al. Serum levels of high mobility groupbox 1 correlate with disease severity inrecessive dystrophic epidermolysis bullosa.Exp Dermatol 2013;22:433–435.26 Nagy N, Almaani N, Tanaka A et al. HB-EGF induces COL7A1 expression in keratino-cytes and fibroblasts: Possible mechanismunderlying allogeneic fibroblast therapy inrecessive dystrophic epidermolysis Bullosa.J Invest Dermatol 2011;131:1771–1774.27 Mirza R, DiPietro LA, Koh TJ. Selectiveand specific macrophage ablation is detri-mental to wound healing in mice. Am JPathol 2009;175:2454–2462.28 Daley JM, Brancato SK, Thomay AAet al. The phenotype of murine wound mac-rophages. J Leukocyte Biol 2010;87:59–67.29 Zhang QZ, Su WR, Shi SH et al. Humangingiva-derived mesenchymal stem cells elicitpolarization of m2 macrophages and enhancecutaneous wound healing. Stem Cells 2010;28:1856–1868.30 Fritsch A, Loeckermann S, Kern JS et al.A hypomorphic mouse model of dystrophicepidermolysis bullosa reveals mechanisms ofdisease and response to fibroblast therapy.J Clin Invest 2008;118:1669–1679.31 Chopra V, Tyring SK, Johnson L et al.Patients with severe forms of inherited epi-dermolysis bullosa exhibit decreased lympho-kine and monokine production. J ClinImmunol 1990;10:321–329.32 Jeltsch KS, Radke TF, Laufs S et al.Unrestricted somatic stem cells: Interactionwith CD341 cells in vitro and in vivo, expres-sion of homing genes and exclusion oftumorigenic potential. Cytotherapy 2011;13:357–365.33 Aktas M, Buchheiser A, Houben A et al.Good manufacturing practice-grade productionof unrestricted somatic stem cell from freshcord blood. Cytotherapy 2010;12:338–348.34 Day CP, Carter J, Bonomi C et al.Lentivirus-mediated bifunctional cell labelingfor in vivo melanoma study. Pigment CellMelanoma Res 2009;22:283–295.
See www.StemCells.com for supporting information available online.
Liao, Ivanova, Zhu et al. 1817
www.StemCells.com VC AlphaMed Press 2015





























