Embed Size (px)
Citation preview

DNA AND CELL BIOLOGYVolume 22 Number 10 2003copy Mary Ann Liebert IncPp 665ndash677
Specificity in Stress Response Epidermal KeratinocytesExhibit Specialized UV-Responsive
Signal Transduction Pathways
MAKOTO ADACHI15 ALIX GAZEL1 GIUSEPPE PINTUCCI1 ALYSSA SHUCK14 SHIVA SHIFTEH1
DOV GINSBURG1 LAXMI S RAO1 TAKEHIKO KANEKO15 IRWIN M FREEDBERG12
KUNIHIKO TAMAKI5 and MIROSLAV BLUMENBERG134
ABSTRACT
UV light a paradigmatic initiator of cell stress invokes responses that include signal transduction activationof transcription factors and changes in gene expression Consequently in epidermal keratinocytes its prin-cipal and frequent natural target UV regulates transcription of a distinctive set of genes Hypothesizing thatUV activates distinctive epidermal signal transduction pathways we compared the UV-responsive activationof the JNK and NFkB pathways in keratinocytes with the activation of the same pathways by other agentsand in other cell types Using of inhibitors and antisense oligonucleotides we found that in keratinocytes onlyUVBUVC activate JNK while in other cell types UVA heat shock and oxidative stress do as well Ker-atinocytes express JNK-1 and JNK-3 which is unexpected because JNK-3 expression is considered brain-spe-cific In keratinocytes ERK1 ERK2 and p38 are activated by growth factors but not by UV UVBUVC inkeratinocytes activates Elk1 and AP1 exclusively through the JNK pathway JNKK1 is essential for UVBUVCactivation of JNK in keratinocytes in vitro and in human skin in vivo In contrast in HeLa cells used as acontrol crosstalk among signal transduction pathways allows considerable laxity In parallel UVBUVC andTNFa activate the NFkB pathway via distinct mechanisms as shown using antisense oligonucleotides targetedagainst IKKb the active subunit of IKK This implies a specific UVBUVC responsive signal transductionpathway independent from other pathways Our results suggest that in epidermal keratinocytes specific sig-nal transduction pathways respond to UV light Based on these findings we propose that the UV light is nota genetic stress response inducer in these cells but a specific agent to which epidermis developed highly spe-cialized responses
665
INTRODUCTION
MULTIPLE EXTRACELLULAR DAMAGING AGENTS cause simi-lar intracellular responses jointly termed ldquothe stress re-
sponserdquo These agents include UV light osmotic shock arsenicoxidizing chemicals protein denaturing agents etc (Feige etal 1996) The stress response includes activation of particularsignal transducing pathways and transcription factors DNA re-pair enzymes as well as cytoskeletal changes (Leppa andBohmann 1999 Mercurio and Manning 1999) The common
responses to diverse agents imply that common pivotal sensorsof environmental stress activate a common predesignated setof signaling pathways (Williams 1999) However different celltypes are ordinarily exposed to different damaging agents andtherefore we hypothesized that in different cell types specificagents activate specific responses
A case in point is the epidermis our first line of defenseagainst the environment and the only tissue exposed to solar ul-traviolet radiation We hypothesize that unlike other cell typesin our body epidermal keratinocytes have developed specific
1Departments of Dermatology 2Cell Biology and 3Biochemistry and 4The NYU Cancer Institute New York University School of MedicineNew York New York
5Departments of Dermatology Faculty of Medicine University of Tokyo Bunkyo-ku Tokyo Japan
remedies not only to protect themselves from UV but also toprotect the underlying organism If this hypothesis is correctthen UV is not just another generic cause of stress to ker-atinocytes but a specific injury with a specific set of responsesTherefore in epidermal keratinocytes UV should elicit not onlythe general stress responses characterized in other cell typesbut also additional skin-specific protective mechanisms In-deed using DNA microarrays we found keratinocyte-specificregulation of gene expression by UV light (D Li et al 2001)Therefore we decided to examine the UV responding signaltransduction pathways in human epidermal keratinocytes a rel-evant target for UV light
The major effects of photodamage have been reviewed com-prehensively (Gilchrest 1995) The cellular effects of UV con-sistently include DNA damage apoptosis and transcriptionalchanges All three wavelengths UVC (100ndash290 nm) UVB(290ndash320 nm) and UVA (320ndash380 nm) cause DNA damageUVC is not present in terrestrial light UVB and UVC have sim-ilar molecular and biological effects in human skin whereas themolecular effects of UVA are different (Grether-Beck et al1997) The inherent responses of keratinocytes to UV by anal-ogy with responses to other extracellular signals can be sepa-rated into two phases the immediate which does not require denovo protein synthesis and delayed which depends on newly in-duced proteins The immediate phase comprises the cytoskeletalchanges and UV-specific signal transduction cascades which de-pends on protein phosphorylation and results in activation of tran-scription factors whereas the delayed phase consist of tran-scriptional and cytoskeletal changes (Hibi et al 1993 Rosetteand Karin 1996 Iordanov et al 1997 Bender et al 1998)
A major impetus for studies of the molecular response to UVcame with the identification of c-Jun amino-terminal kinases(JNK) the protein kinase that in response to UV binds to andactivates transcription factor c-Jun (Hibi et al 1993) Soon itwas realized that JNK responds to several stress signals in ad-dition to UV such as osmotic shock arsenate and proinflam-matory cytokines (Deacuterijard et al 1994 Kyriakis et al 1994)Three isoforms JNK1 JNK2 and JNK3 are often expressedin two splice variants designated ldquoardquo and ldquobrdquo (Karin 1995)JNK1 is a 46 kDa protein that when activated enters the nu-cleus and phosphorylates Ser63 and Ser73 of c-Jun The tran-scription factors activated by JNK are Elk1 SAP1 ATF2PEA3 and the eponymous Jun (Gupta et al 1996) Whilebound to c-Jun JNK1 can phosphorylate the Jun dimerizationpartners JunB JunD and ATF2 (Kallunki et al 1996) Thesignal-transducing cascade that activates JNK is analogous tothe Ste (for sterile) pathway in yeast the mating pheromone-responsive signal transduction cascade (Herskowitz 1995)JNK can be activated by one of two kinases designated JNKK1(aka SEK1 SKK1 and MKK4) and JNKK2 (MKK7) (Wu etal 1997) JNKKs in turn are activated by multiple JNKKKskinases that respond to a variety of stimuli (Fanger et al 1997)The UV-responsive JNKKK has not been identified yet
Another clear molecular effect of UV is the activation of theNFkB transcription factor (Barnes and Karin 1997 N Li andKarin 1998) The NFkBRel family of transcription factorsplays a particularly important role in the inflammatory and an-tiapoptotic processes (Baeuerle and Henkel 1994 Miyamotoand Verma 1995 Liu et al 1996 Tomic-Canic et al 1998)The activation of NFkB by UV is not associated with DNA
damage and occurs even in cytoplasts devoid of nuclear DNA(Devary et al 1993 Simon et al 1994) Inactive NFkB re-sides in the cytoplasm in complex with IkB protein Upon ac-tivation by diverse extracellular stimuli a large multiproteincomplex containing IKKa and IKKb phosphorylates IkB (Beget al 1993 Alkalay et al 1995 DiDonato et al 1997) Re-cently knockout mice of both IKKa and IKKb have been de-scribed (Delhase et al 1999 Hu et al 1999) It appears thatIKKb is responsible for cytokine signaling in most tissueswhile specifically in the epidermis IKKa is essential for nor-mal keratinocyte differentiation The mechanisms by which inflammatory cytokines such as TNFa or IL-1 as well as ion-izing radiation activate NFkB are known to involve phospho-rylation of two specific serine residues (Ser-32 and Ser-36)within the N-terminal regulatory domain of IkBa which is fol-lowed by polyubiquitination and degradation of IkB by a 26Sproteasome (Beg et al 1993 Alkalay et al 1995 Brown etal 1995 Bender et al 1998) The proteolysis of IkB resultsin the release of NFkB which is then free to enter the nucleusand activate gene transcription
Although UV can cause a release of TNFa and so indirectlyactivate the above pathway in direct response to UV an uniden-tified kinase activates NFkB (Bender et al 1998 N Li andKarin 1998) The direct UV-induced activation of NFkB maynot involve IKKb and is not dependent on phosphorylation atSer-32 and Ser-36 of IkBa (Bender et al 1998 Li and Karin1998) These experiments were performed with HeLa cellswhich are not a natural target of UV light
Several specific inhibitors of the NFkB signaling pathwayhave been described BAY11-7085 inhibits the cascade up-stream from IKK salicylates widely used anti-inflammatorydrugs have been recently identified as specific inhibitors ofIKKb while parthenolide directly inhibits the degradation ofIkBa (Kopp and Ghosh 1994 Grilli et al 1996 Pierce et al1996 Hehner et al 1998 Yin et al 1998)
Hypothesizing that skin has specific mechanisms to protectitself and the organism from UV damage we examined theMAPK and NFkB signaling pathways in human epidermal ker-atinocytes We found that in keratinocytes the activation of theJNK pathway by UV critically depends on JNKK1 and thatkeratinocytes express an unexpected assortment of JNK iso-forms Most important in keratinocytes unlike in other celltypes we examined the activation is very stringent UVB andUVC activate JNK while UVA does not Furthermore UVBand UVC do not activate parallel pathways and there is nocrosstalk between the UVB and UVC-dependent pathway andthe growth factor-dependent pathways Using specific inhibi-tors as well as antisense approaches we also compared the path-ways that activate NFkB in response to UV or TNFa in ker-atinocytes and HeLa cells We found that UVC and UVBactivate NFkB in both cell types but that UVA has no effectin either In addition we have shown that UVC and UVB usethe same signal transduction pathway to induce NFkB and thatthis pathway does not involve IKKb while TNFa uses a dif-ferent pathway to activate NFkB through IKKb Finally usingantisense oligonucleotides to target NFkB and IKKb we haveconfirmed that IKKb is essential for activation of NFkB byTNFa but plays no role in activation of NFkB by UVBUVCWe conclude that there is an independent UV responsive sig-nal transduction pathway activating NFkB in epidermis
ADACHI ET AL666
MATERIALS AND METHODS
Plasmids their growth and purification
The plasmid containing (NFkB)3-CAT was a gift from JVilcek NYU School of Medicine plasmid expressing Gal4Elkand GAL4CAT were a gift from J Blau Rockefeller Univer-sity and the control plasmids pRSVZ has been described pre-viously (Jiang et al 1993 1994 Klampfer et al 1994) Plas-mids were grown in JM101 Escherichia coli host to saturationdensity in LB medium DNAs were extracted and purified us-ing the Mega Prep kit from Promega (Madison WI)
Cell growth maintenance and transfection
Normal human foreskin epidermal keratinocytes were a gen-erous gift from Dr M Simon The cultures were initiated us-ing 3T3 feeder layers as described (Simon and Green 1984)and then frozen in liquid N2 until used Once thawed the ker-atinocytes were grown without feeder cells in defined serum-free keratinocyte medium supplemented with epidermal growthfactor and bovine pituitary extract (keratinocyte-SFM GIBCOCarlsbad CA) Cells were expanded through two 14 passagesbefore transfection and transfected at 90 confluency HeLacells were maintained in Dulbecco-modified Eaglersquos mediumsupplemented with 10 calf serum At the beginning of theseexperiments we optimized the doses of UV and concentrationsof EGF and TNFa to maximize the activation of the reportersin both HeLa cells and epidermal keratinocytes without killingthe cells (data not shown) All subsequent experiments wereperformed with optimal doses and concentrations
Transfections of keratinocytes with polybrene and DMSOshock were used to insert the DNA into the subconfluent ker-atinocytes as previously described (Jiang et al 1991) On theday of transfection cells were washed and incubated in the ba-sal medium without EGF and pituitary extract Each transfec-tion contained 10 mgdish of (NFkB)3-CAT and 3 mgdish ofpRSVZ The cells were incubated at 37degC in a 5 CO2 atmo-sphere and 24 h after transfection were illuminated with UVAUVB UVC or stimulated with TNFa EGF (100 ngml) in thepresence or absence of various inhibitors Treatment with as-pirin (0ndash100 mM from Sigma St Louis MO) BAY (0ndash5 mM)parthenolide (0ndash15 mM) and U0126 (0ndash1 mM all from BiomolPlymoth Meeting PA) lasted for another 24 h before harvest-ing We used two StratalinkerTM units (Stratagene La JollaCA) one equipped with FG15T8 bulbs as our UVB source andstandard GL-15 G15T8 bulbs as a source of UVC Before illu-mination the medium was removed from the cells and imme-diately after the illumination the same medium was restored tothe cells The whole illumination procedure takes less than aminute which we have shown does not cause drying or a heatshock The control cultures were treated identically except theactual illumination was omitted Cells were washed twice withphosphate-buffered saline 48 h after transfection and harvestedby scraping The cell disruption by repeated freezendashthaw cy-cles has also been described (Jiang et al 1991) All transfec-tions were performed in duplicate plates and each transfectionexperiment was repeated three to five times
HeLa cells were transfected using a modified calcium phos-phate precipitation procedure (Jiang et al 1991) Two days be-fore transfection cells were split by trypsinization and trans-
fected at 50 confluency Ten micrograms of the reporter plas-mid and 3 mg of the b-gal control plasmid were dissolved in atotal volume of 450 ml H2O To this mixture 50 ml of 25 MCaCl2 was added and mixed carefully Five hundred microlitersof BES-buffer pH 695 (50 mM N-Bis-2-hydroxyethyl-2-aminoethanesulfonic acid 250 mM NaCl 15 mM Na2HPO4)was added and mixed by inverting the tube The reaction mix-tures were incubated at room temperature for 10ndash15 min Onemilliliter of the precipitate suspension was added per platewhich was followed by incubation at 37degC in a 5 CO2 atmo-sphere Twenty-four hours after transfection cells were stimu-lated by TNFa or illuminated with UVA UVB or UVC withor without various inhibitors as indicated Cells were washedtwice with phosphate-buffered saline 36 to 48 h after transfec-tion and harvested by scraping
Enzyme assays
For b-galactosidase assays the substrate solution contained6 mg of O-nitrophenyl-D-galactoside (Sigma St Louis MO)freshly dissolved in PM buffer (66 mM Na2HPO4 33 mMNa2HPO4 40 mM mercaptoethanol 2 mM MgSO4 and 01 mMMnCl2) The reaction mixture contained 100 ml of substrate so-lution 300 ml of PM2 buffer and 50 ml of keratinocyte cell ex-tract or 20ndash30 ml of HeLa cell extract It was incubated at 37degCuntil development of a yellow color was obvious usually at 30min to 1 h The time of the reaction was recorded and the re-action stopped by addition of 04 ml of 1 M Na2CO3 OD420
was measured on a spectrophotometer (Labrepco HorshamPA)
The CAT reaction mixture contained 69 ml of 1 M Tris HClpH 78 1 ml of 14[C]-chloramphenicol (Sigma) 20 ml of 4 mMacetyl-CoA solution 30ndash60 ml of cell extract and enough wa-ter to bring the total reaction volume to 150 ml After incuba-tion at 37degC for 1 h the mixture was extracted into 1 ml ofethyl acetate phases were separated by brief centrifugation theorganic layer was transferred to a new tube and the solventevaporated The residue was dissolved in 25 ml of ethyl acetateand separated by thin layer chromatography on a silica gel platein chloroformmethanol 5 955 The plates were exposed to X-ray film for 12 to 24 h and the intensity of radioactive spotsdetermined using the Ambis Radioanalytic System (AmbisRochester NY) The conversion of chloramphenicol to itsmonoacetylated derivative was kept below 50 by varying theamount of extract or the duration of the reaction All CAT val-ues were normalized for transfection efficiency by calculatingthe ratio of CAT activity to b-galactosidase in each transfectedplate Each transfection experiment was separately performedthree or more times with each data point resulting from dupli-cate transfections
Use of antisense and sense oligonucleotides
Keratinocytes readily take up offered DNA Specificallyshort oligonucleotides can be introduced into these cells evenin the absence of cationic lipid (Nestle et al 1994 Reuning etal 1995) The oligonucleotides were synthesized in the phos-phorothiolate form to prolong their half-lives in the cells Wetargeted the antisense oligonucleotides to sequences includingand immediately upstream from the initiation codon for themost efficient antisense blocking At the same time we pre-
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 667
pared and used sense oligonucleotides as negative controls Ourapproach was to add the antisense oligonucleotides to the DNAtransfected into cells including the oligonucleotides into thetransfection solution which has the advantage of ensuring thatthe cells that received the transfected DNA also received theoligonucleotides The oligonucleotides were stored at 270degCin water until use and were added to the transfected DNA mix-ture In most cases we used 4 mg of the oligonucleotide pertransfection Their sequences are given in Table 1
Western blotting
The activated JNK antibody was purchased from Calbiochem(San Diego CA) and used at 1100 to 11000 dilution whilethe antirabbit immunoglobulin horseradish peroxidase-linkedantibodies and the ECL reagent were used as suggested by themanufacturer (Amersham Piscataway NJ) The anti-JNK3SAPK 1b antibody was purchased from Upstate Biotech-nology (Lake Placid NY) We used the antibody that recog-nizes JNK3 a1 (46KDa) and JNK3a2 (54KDa) at 11000dilution and the A431 cell lysate (Upstate Biotechnology) aspositive control according to manufacturerrsquos recommendationKeratinocytes were grown as described above At their thirdpassage at 80 confluence we incubated keratinocytes inKBM for 16 h to starve them of growth factors and then treatedas described above At various time intervals cells were har-vested by scraping washed twice with PBS and the cell pelletwas resuspended in 100 ml of the lysis buffer (2 SDS 10glycerol 10 mM Tris HCl pH 68) The cells were lysed by ul-trasonication for 30 sec using an ultrasonic homogenizer soni-cator (Cole-Parmer Vernon Hills IL) and boiling for 10 minAliquots of the supernatant obtained after centrifugation(2000 3 g 10 min) were stored at 270degC Protein content wasdetermined using the Bradford reagent (Perkin-Elmer BostonMA) Equal amounts of total protein were resolved by sodiumdodecyl sulphate-polyacrylamide gel electrophoresis (10 gel)
and electrophoretically transferred to nitrocellulose at 8 V for1 h After blocking the membranes with 5 nonfat dry milk inTris-buffered saline with 01 Tween (TBS-T) for 1 h at roomtemperature on an orbital shaker the blots were washed thrice(10 min each time) with TBS-T and incubated at 4degC with11000 dilution of the antibody Blots were washed 16 h laterwith TBS-T (3 3 10 min) and incubated at room temperaturewith 11000 dilution of antirabbit immunoglobin horseradishperoxidase-linked antibody for 2 h The chemiluminescenceECL reagent was used to develop the blots after washing themembranes three times 10 min with TBS-T and twice withTBS alone
JNK activity blots
Untreated control cells and cells treated with UV or EGFwere harvested 30 min after treatment The JNK activity wasmeasured by an in vitro kinase assay that detects phosphoryla-tion of a GST-Jun fusion protein The GST-Jun expressing con-struct was a generous gift from Dr Susan Logan New YorkUniversity School of Medicine This construct generates a fu-sion protein containing the 79 amino-terminal residues of c-junThe fusion protein GST-jun was used to purify JNK from thecell lysates and subsequently as a substrate for the same ki-nase The kinase assay was performed in kinase assay buffer(20 mM HEPES pH 75 20 mM beta-glycerol-phosphate 10mM MgCl2 10 mM DTT 50 mM sodium-orthovanadate 20mM ATP and 05 mlreaction of 32P-ATP (New England Nu-clear Amherst MA) The product of the kinase reaction wasthen separated by SDS-PAGE and the 32P-labeled GST-jun de-tected using autoradiography
RT-PCR
Cultured cells were harvested and total RNAs isolated uti-lizing RNeasy RNA extraction kit from Qiagen Between 1 and
ADACHI ET AL668
TABLE 1 OLIGONUCLEOTIDES USED
RT-PCR oligonucleotides used in detecting JNK ERK and p38 mRNAsJNK1 forward TCGCTACTACAGAGCA (570 bp expected product)JNK1 reverse GGAGAGGGCTGCCCCCJNK2 forward CTGGCCCGGACAGCGT (360 bp)JNK2 reverse GTTTTTATTTTGTCTCJNK3 forward TTATTACAGAGCCCCT (537 bp)JNK3 reverse TTTAGTCTTTTCTTCTGERK1 forward TTGCAGATCCAGACCA (295 bp)ERK1 reverse ATTTTTGTGTGGAAGAp38 forward GACGGATCGCAGAAGA (397 bp)p38 reverse TGCCAGGCACCCCGGT
Antisense oligonucleotides used in transfection of keratinocyte synthesized as phosphorotiolatesas-JNK1 CATGATGGCAAGCAATTAATGs-JNK1 GTACTACCGTTCGTTAATTACas-JNKK1 CATTGTTGGGAGTGAAGAGCCCAs-JNKK1 GTAACAACCCTCACTTCTCGGGTas-NFkB TGGATCATCTTCTGCCATTCTs-NKkB ACCTAGTAGAAGACGGTAAGAas-IkB CGCGTCGGCCGCCTGGAACATGCas-IKKa CATGGGGCGGGAGGGCAAGCGGCCas-IKKb ACTGATGTCGTGCTAACTCTGTCGs-IKKb TGACTACAGCACGATTGAGACAGC
15 mg of RNA were subjected to RT-PCR with Access RT-PCR kit system from Promega (Madison WI) for 32 cycleswith buffers and chemicals added as suggested by the manu-facturer In the supernumerary PCR experiments we used 42cycles The PCR products were subjected to either agarose orpolyacrylamide gel electrophoresis visualized with ethidiumbromide (Sigma) with a UV transilluminator from UVP andphotographed (Polaroid Waltham MA) The densities of bandswere quantified utilizing image scanner (GT-9000 Epson LongBeach CA)
Organ culture explants of normal human skin
Pieces of normal human skin were obtained immediately af-ter surgery They were cut into pieces approximately 3ndash5 mm3and incubated in keratinocyte basal medium KBM (Keratino-cyte-SFM Gibco-BRL Carlsbad CA) with or without TNFa
(50 ngml Intergen Norcross GA) in a humidified incubatorat 37degC for 24 h Generally we used 24-well culture disheswith up to five pieces in the same well and just enough mediumto cover the explants The antisense oligonucleotides wereadded at 300 ngml and when necessary replaced every 2 daysIllumination was performed epidermal side up with 100mJcm2 of UVB The explants were mounted in tissue Tec-OCTcompound (Sakura Finetek Japan) and frozen Sections 4 to 6mm thick were obtained with a cryostat (Miles LaboratoriesResearch Triangle Park NC) fixed with methanolacetone for10 min incubated with anti-JNK1 antibody (Calbiochem SanDiego CA) at 4degC overnight treated with peroxidase-conju-
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 669
FIG 1 UV activates JNK in keratinocytes The two left pan-els show the UV dose curve with each sample loaded in du-plicate The right panels show the time course of activation af-ter the illumination The top panels show the kinase activity ofJNK The activity was shown using GSTndashJun fusion protein asa substrate for immunoprecipitated JNK The bottom panelsshow the corresponding Western blots using antibody specificfor the activated phosphorylated form of JNKs The trianglepoints to the phosphorylated JNK substrate the doublet of openarrows to the JNK proteins The diffuse band in the top panelsis an uncharacterized artifact In the left panels we harvestedthe cells 10 min after irradiation and in the right panels weused 10 mJcm2 In each well we loaded 10 mg of protein
FIG 2 RT-PCR identification of JNK isotypes in keratinocytes HeLa cells and fibroblasts (a) Lanes K H and F representRNA from untreated keratinocytes HeLa cells and fibroblasts respectively Isotype-specific oligonucleotides were used in RT-PCR reactions The arrows point to the specific bands (b) Exaggerated PCR reactions for detecting low abundance mRNAs Du-plicate lanes show results of PCR reaction without (2) and with (1) input RNA preformed with supernumerary cycles (c) West-ern blots showing the presence of JNK3 lane 1 positive control (A431 extract) lane 2 HeLa cells and lane 3 keratinocytes
gated antimouse IgG secondary antibody (Vecstatin ABC-mouse IgG kit from Vector Laboratories Burlingame CA) atroom temperature for 1 h incubated with ABC complex (Vec-tor Laboratories) at room temperature for 1 h and treated with339-Diaminobenzidine-tetrahydrochloride (Dojindo Gaithers-burg MD) and 001 H2O2 in Tris pH 76 for 2 min The sam-ples were observed and photographed under the light micro-scope (Microphot-FXA Nikon Melville NY) Additionalantibodies used were from SantaCruz Biotechnology (SantaCruz CA)
RESULTS
UVB signal transduction pathway part I activation ofthe JNK pathway
To determine which JNK isoforms exist in keratinocytes andcan be activated by UVB we have grown primary cultures ofhuman epidermal keratinocytes and illuminated them with UVBat various doses As controls we used HeLa cells and normalhuman dermal fibroblasts The JNK activity was examined inthree ways First illuminated cells were stained with pan-JNKor active JNK-specific antibody to demonstrate the transloca-tion of JNK from cytoplasm to nucleus (not shown) Secondprotein extracts were prepared and analyzed using Western blotswith active JNK-specific antibody Third the kinase activitywas determined directly in affinity-precipitated JNK using aGST-tagged Jun protein fragment as the substrate The resultsof all three approaches were perfectly consistent UVB stronglyand specifically activates JNK in human epidermal ker-atinocytes (Fig 1)
Three isotypes of JNK have been described each with sev-eral splice variants It should be noted that there is confusionregarding which isotype is JNK2 and which is JNK3 in the lit-erature and even more egregiously in the blurbs of manufac-turers who provide antibodies We have adopted the unam-biguous designation of the NCBI databank To determine whichisotypes are expressed in keratinocytes we developed a set ofspecific RT-PCR oligonucleotides and used them to amplify theJNK mRNAs Surprisingly we found JNK1 and JNK3 in ker-atinocytes while JNK2 was absent (Fig 2a) This is an unex-pected finding because JNK1 and JNK2 are commonly almostubiquitously expressed while JNK3 was thought to be brain-specific (Yang et al 1997) Repeat of the PCR assay using alarge number of cycles confirmed the absence of JNK2 in ker-atinocytes (Fig 2b) Furthermore because of the importanceand unexpectedness of this result we decided to confirm theexpression of JNK3 protein in keratinocytes using Western blot-ting Indeed we find that JNK3 is expressed in keratinocytesbut not in HeLa cells (Fig 2c) Therefore keratinocytes expressboth the mRNA and the protein of JNK3 a kinase previouslythought to be brain-specific
To demonstrate that the activation of JNK is functional inthat it activates the Elk1 transcription factor in keratinocyteswe transfected into these cells a set of plasmids containing aGal4-CAT reporter and Gal4-DBDElk1-AF (Yankulov et al1994) The first plasmid contains a set of Gal4 binding sites up-stream from the CAT reporter gene the second expresses a fu-sion protein containing the Gal4 DNA binding domains fused
to the activating domain of transcription factor Elk1 When ac-tivated by UV via JNK the fusion protein activates transcrip-tion but if not activated it is silent Thus the system serves asan UV-responsive transcription reporter The transfected cellswere illuminated with UV and harvested 16 h later We foundthat both UVC and UVB activate the reporter but UVA doesnot (Fig 3) Note that at the 50 mJcm2 dose of UVC a signif-icant number of cells are killed These results mean that theJNK system in keratinocytes is present functional and specif-ically activated by short wavelength UV light UVBUVC butnot the long wavelength UVA We conclude that UVA doesnot activate the same system of signal transduction thatUVBUVC do
To identify the kinases responsible for UV signal we usedantisense approaches The antisense oligonucleotides were tar-geted against JNK1 and JNKK1 After 16 h to allow the anti-sense oligonucleotides to inhibit the synthesis of their targetsthe cells were irradiated with UV and after another 24 h har-vested for CAT determination We found that both antisensereagents attenuated the signals (Fig 4a and b) The antisenseeffects are specific because the sense oligonucleotides used ascontrols had no effect These results indicate that JNK1 andJNKK1 are essential for the transmission of the UVBUVC sig-
ADACHI ET AL670
FIG 3 Activation of transfected Elk-CAT reporter by UVBand UVC in keratinocytes GAL4-Elk1 fusion protein express-ing plasmid was cotransfected into keratinocytes with aGAL4CAT reporter The top panel shows the two cotransfectedplasmids and the effect of UV The transfected cells were illu-minated with indicated doses of UVA UVB or UVC harvested16 h later and the CAT levels determined Note that at the 50mJcm2 of UVC a significant number of cells is killed whichresults in significantly reduced levels of CAT activity (aster-isk) At the doses used UVA did not affect cell viability Thebars represent averages of duplicate transfections and the repli-cates differed by 25 or less
nal However they do not formally exclude the possibility thatJNK3 and JNKK2 also play a role
To determine whether the Elk1 activation is specific for theUV responsive signals in keratinocytes we tested the effects ofEGF In other cell types EGF can act through pathways thatactivate both JNK and Elk1 However when we attempted toactivate Elk1 using EGF in keratinocytes we were unsuccess-ful (Fig 4b) This means that in keratinocytes the EGF and UVresponsive pathways do not overlap on the contrary UV specif-ically activates the JNK pathway This suggests that the epi-dermal keratinocytes exhibit distinctive specialized UV-responsive signal transduction pathways
Indeed when we compared the responses to UV in severalcell types we found that the response in keratinocytes is sig-nificantly more stringently controlled than in other cell typesFor example both UV and EGF activated the JNK pathway inHeLa cells (Fig 4c and d) demonstrating a lack of specificityMoreover targeting JNK1 and JNKK1 with antisense oligonu-cleotides only partially inhibited the UV effects in HeLa cells
implying that in these cells parallel pathways can respond toUV to activate Elk1 (Fig 4c) Suspecting that EGF activatesElk1 through both the JNK and the ERKMAPK pathways inHeLa we inhibited the latter with U0126 a specific MEK-12inhibitor We found that U0126 significantly blocks the acti-vation by EGF but not by UV (Fig 4e) again demonstratingrelaxed stringency in HeLa cells
To approach these phenomena from another angle we usedkinase assays and Western blots Consistent with the above re-sults we found that while in keratinocytes only UVB (andUVC) could activate the JNK pathway in HeLa cells (and fi-broblasts not shown) heat shock UVA and osmotic shockcould activate the pathway as well (Fig 5a) In HeLa cells andfibroblasts UVB could activate the ERK pathway (not shown)but in keratinocytes it could not (Fig 5b) We conclude fromthese results that keratinocytes being often exposed to ambi-ent UV have developed more stringent and specific responsesto UV than other cell types which are not commonly exposedto UV These data are summarized in Figure 5c
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 671
FIG 4 UV specifically activates JNK in keratinocytes but not in HeLa cells GAL4-Elk1 and GAL4CAT plasmids were co-transfected into keratinocytes (a and b) or HeLa cells (c d and e) The transfected cells were treated with either UVC (5 mJcm2)or EGF (100 ngml) and harvested 24 h later for determination of CAT activity U0126a specific inhibitor of the MAPK path-way blocks the effects of EGF but not of UV Open bars represent the untreated control cells closed bars the UV-treated onesand the shaded bars the EGF treated ones
While there are only two JNKKs kinases that directly phos-phorylate JNK there is a large family of JNKKKs kinases thatphosphorylate JNKKs In a manuscript in preparation we de-scribe our analysis of the kinases expressed by keratinocytes
To demonstrate that the JNK system is responsive in vivowe used a novel ex vivo approach Biopsies of human skin weretreated with as-JNKK1 oligonucleotide or left untreated as con-trol Then both the treated and untreated biopsies were illumi-nated with UVC and shortly thereafter processed for immuno-histochemical detection of activated JNK (Fig 6) We found inthese biopsies that UV caused activation and nuclear translo-cation of JNK Furthermore this activation was abrogated bypretreatment with as-JNKK1 the antisense oligonucleotide tar-geting JNKK1 again demonstrating that JNKK1 is essential foractivation of JNK by UV in epidermis These results mean that
the specific signal transduction pathways we found operationalin cultured keratinocytes are also functional and working thesame way in human skin in vivo
UVB signal transduction pathway part II activationof the NFkB pathway
Having established that keratinocytes possess a specific UV-responding pathway to activate JNK we wanted to examine thepossibility that the UV-specific signal transduction pathwaysactivating NFkB are also specific in these cells The canonicalactivator of NFkB is TNFa however recent evidence suggeststhat UV and TNFa may activate NFkB through different mech-anisms (N Li and Karin 1998) Demonstrating a UV-specificpathway would strengthen even more our hypothesis that thereis specificity in UV responsive signal transduction in humanepidermal keratinocytes Therefore we transfected into ker-atinocytes and into HeLa cells a reporter plasmid containingthree tandem NFkB sites linked to the TK promoter-drivenCAT We found that the exposure of transfected cells to TNFaUVB or UVC resulted in the induction of the NFkB responseelement (Fig 7) Generally at the optimal conditions the in-duction of NFkB by UV was weaker than the induction byTNFa On the other hand NFkB was not activated by UVA atall even at high doses (Fig 7)
To compare the mechanisms of NFkB activation by UV andTNFa we have used a battery of specific inhibitors of the NFkBpathway which inhibit the pathway at different points (Fig 8a)Namely Bay11-7085 inhibits upstream from IKK (Pierce et al1997) aspirin has recently been found to be a competitive in-hibitor of ATP binding to IKKb (Yin et al 1998) whereasparthenolide directly inhibits the degradation of IkBa (Pierceet al 1997) When we treated transfected keratinocytes or HeLacells with these three inhibitors we found that Bay11-7085 in-hibited the induction of NFkB by TNFa in a dose-dependentmanner while it has almost no effect on the induction of NFkBby UVC (Fig 8b) The same results were obtained by using as-pirin (Fig 8c) In contrast parthenolide inhibited both the UVCand the TNFa effects in parallel (Fig 8d) These results con-firm that TNFa and UVC use different pathways to activateNFkB in HeLa cells Specifically while both UVC and TNFaactivate NFkB the TNFa signal goes via IKKs but the UVCsignal does not From these data we conclude that the conver-gence of the UVB and TNFa signals occurs downstream fromIKKs that is at the modification of IkB
One caveat in the use of inhibitors of specific kinases is thepossibility that such inhibitors may affect additional pathwaysnot currently recognized as affected by the given inhibitor Toaddress such specificity concerns and to examine the involve-ment of IKK in the induction of NFkB by TNFa UVC andUVB we used antisense oligonucleotides specifically targetingNFkB and IKKb as-NFkB as-IKKb (Table 1) (Reuning etal 1995 Komine et al 2000) In parallel we added the senseoligonucleotides s-NFkB s-IKKb as controls The antisenseexperiments confirmed the fact that UVBUVC use a clearlydifferent pathway to activate NFkB than TNFa does The sup-pression of NFkB synthesis in cultured keratinocytes by addingas-NFkB resulted in inhibition of activation of NFkB by allthree agents UVC UVB and TNFa as expected (Fig 8b) Onthe other hand the suppression of IKKb synthesis by adding
ADACHI ET AL672
FIG 5 Specific activation of JNK in keratinocytes andpromiscuous activation of JNK in HeLa cells (a) JNK activityin keratinocytes and HeLa cells treated with UVC UVA heatshock or EGF Duplicate experiments were performed with allagents except EGF Each lane contains 10 mg of protein InHeLa cells (bottom panel) UVC and EGF strongly while UVAand heat detectably activate JNK Cells were harvested 30 minafter treatment (b) Lack of activation of ERK by UVB in ker-atinocytes The top panel shows a Western blot with the anti-body specific for the activated ERKs the bottom for totalERK1 as a control for loading and transfer (c) Specificity insignal transduction In keratinocytes UVB specifically acti-vates JNK and EGF specifically activates ERKs whereas inHeLa cells all three extracellular signals UVB EGF and heatactivate both JNK and ERK Fibroblasts respond similar toHeLa cells (data not shown)
as-IKKb only inhibited the activation by TNFa but had no ef-fect on the activation by either UVC or UVB (Fig 8b) In theseexperiments we used s-NFkB and s-IKKa as controls and con-firmed that they had no effect in this pathway (Fig 8b) Fur-thermore as-NIK targeting a kinase functioning upstream fromIKK also partially inhibited the effects of TNFa Therefore
the antisense oligonucleotide experiments confirm our conclu-sions that the activation of NFkB by TNFa depends on the IKKpathway whereas the response to UVC or UVB occurs througha mechanism independent of IKKs
Furthermore as an additional control we used oligonu-cleotides targeting the JNK pathway as-JNKK1 and s-JNKK1Although they affected the JNK pathway (see Figs 4 and 6)they did not affect the NFkB pathway (Fig 8b) which indi-cates that the activation of the JNK pathway and the NFkBpathway are independent in keratinocytes This means that epi-dermal keratinocytes contain multiple specific UV responsivemechanisms
DISCUSSION
Our data suggest that UVBUVC illumination in epidermalkeratinocytes is not a generic stress response inducer but a spe-cific extracellular agent that activates specific signal transduc-tion pathways In particular we show that in keratinocytes UVexclusively activates the JNK pathway while in HeLa cellsUV also activates the parallel ERK and p38 pathways Con-versely in keratinocytes exclusively UV activates the JNKpathway while in HeLa cells growth factors can do so as wellIn parallel in keratinocytes UV uses a distinct pathway to ac-tivate the NFkB transcription factor a pathway separate fromthe more common TNFa activated one (Fig 9)
The specific activation of JNK by UVBUVC is particularlyrelevant because the JNK pathway is in different cell types atarget of multiple stress agents The balance between apoptosisand proliferation is at times regulated by JNK and the tran-scription factors activated by JNK regulate the expression ofmany genes (Karin and Hunter 1995) It is uniquely importantin epidermis that JNK exclusively responds to UVBUVC be-cause epidermis is the natural target of UVBUVC and the JNK-
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 673
FIG 6 Activation of JNK in vivo and the effect of as-JNKK1 In the control sample (left) JNK is not activated as shown us-ing an antibody specific for the activated phosphorylated form of JNK Treatment with 100 mJcm2 causes strong activation ofJNK (middle) Pretreatment for 3 h with the antisense oligonucleotide targeting JNKK1 abolishes this activation (right)
FIG 7 TNFa UVB and UVC activate NFkB while UVAdoes not Reporter plasmid NFkB3-CAT (top) which containsthree tandem NFkB sites upstream from the TK promoter wastransfected into keratinocytes The transfected cells were eithertreated with 50 ngml of TNFa or illuminated with indicateddoses of UV harvested 16 h later and the CAT levels deter-mined Untreated cells served as a control The replicates dif-fered by 25 or less
ADACHI ET AL674
FIG 8 Specific inhibitors can differentiate between the TNFa-and UVB-dependent signal transduction pathways (a) The effectsof specific inhibitors on NFkB activation Cultured keratinocyteswere treated with either UVB (8 mJcm2) or TNFa (50 ngml) af-ter transfection of the NFkB response element Inhibitor Bay11-7085 specifically inhibits upstream kinases of IKK aspirin inhibitsIKKb parthenolide blocks IkBa degradation (b) the effects of an-tisense oligonucleotides targeting signaling cascades on NFkB ac-tivation in keratinocytes Oligonucleotide as-NFkB targeting thetranscription factor blocks the activation by UVB and UVC whileoligonucleotides targeting IKKb JNK1 and JNKK1 are withouteffect (top two panels) In contrast oligonucleotides targetingIKKb and NIK partially or completely block the effects of TNFa(bottom panel) Note that the sense oligonucleotides (s-NFkB s-IKKb) used as controls had no effect on NFkB activation (c)Schematics of NFkB pathways activated by UV light and TNFaIn keratinocytes the two extracellular stimuli work through inde-pendent pathways that merge at the NFkBIkB complex
regulated transcription factors mediate both photodamage andits alleviation (Fisher et al 1997)
In keratinocytes the EGF and UV responsive pathways donot overlap but UV specifically activates the JNK pathwayThis suggests that the epidermal keratinocytes exhibit distinc-tive specialized UV-responsive signal transduction pathwaysIndeed when we compared the responses to UV in several celltypes we found that the response in keratinocytes is signifi-cantly more stringently controlled than in other cell types Forexample HeLa cells demonstrated a lack of specificity Usinga specific MEK-12 inhibitor we again demonstrated relaxedstringency in HeLa cells We conclude from these results thatkeratinocytes being often exposed to ambient UV have de-veloped more stringent and specific responses to UV than othercell types which are not commonly exposed to UV
At present we do not understand the relevance of JNK3 ex-pression in keratinocytes This isoform it was reported isspecifically expressed in the brain (Yoshida et al 2001) Wespeculate that perhaps the sequestration of JNK1 by the speci-ficity of the UV response creates a need for JNK3 as the moregeneral less restrictive stress signal transducer
TNFa one of proinflammatory cytokines important in cuta-neous inflammation activates NFkB via IKKs kinases that canbe activated by MEKK or NIK (Hirano et al 1996 Lee et al1997) In HeLa cells the phosphorylation of IkBa at Ser-32and Ser-36 residues is essential for the IKK mediated IkBa
degradation caused by TNFa IL-1 or ionizing radiation Us-ing specific inhibitors and antisense oligonucleotides we foundthat the NFkB activation by UVBUVC does not involve IKKs
The Ser-32 and Ser-36 residues are not involved in the UV-induced IkBa degradation (Bender et al 1997 N Li and Karin1998) Because IKKb specifically phosphorylates IkB at bothSer-32 and Ser-36 of IkBa these facts support our results Re-cently it was suggested that UV activation of NFkB in HeLacells may involve the RasndashRaf pathway although not in ker-atinocytes (Tobin et al 1996) This leads to the question howUV activates NFkB in keratinocytes Two new kinases IKKlaquo
and pp90rsk have been found to phosphorylate IkBa (Ghodaet al 1997 Schouten et al 1997 Shimada et al 1999 Israel2000 Peters et al 2000) However IKKlaquo phosphorylates theSer-36 residue while pp90rsk phosphorylates the Ser-32residue only Therefore these kinases are probably not involvedin the UV activated pathway Amino acids between IkBa po-sition 277 and 297 (the PEST region) are apparently requiredfor the UV induced degradation of IkB (Ito et al 1995 Ben-der et al 1998) but the signaling events that effect this degra-dation in keratinocytes are still unknown The data suggest thatTNFa and UVC use different pathways to activate NFkB inHeLa cells Specifically while both UVC and TNFa activateNFkB the TNFa signal goes via IKKs but the UVC signaldoes not From these data it appears that the convergence ofthe UVB and TNFa signals occurs downstream from IKKs thatis at the modification of IkB Overall these results imply thatthe epidermis a natural target of UV light has an exceptionalmolecular UV defense system distinct from UV responsive sys-tems in other tissues
The responses to the three different wavelengths of UV areespecially strictly controlled in keratinocytes Although allwavelengths cause DNA damage and a release of cytokines wefound that the inherent molecular responses to UVA are com-pletely different from the responses to UVBUVC For exam-ple UVBUVC but not UVA activate JNK and NFkB in ker-atinocytes Furthermore UVA activates the JNK pathway inHeLa cells but not in keratinocytes while UVBUVC activatethis pathway in both cells (Rosette and Karin 1996) Similarlyin both HeLa cells and cultured keratinocytes UVBUVC ac-tivate NFkB via a kinase different from IKKs while UVA doesnot activate NFkB at all We conclude that UVBUVC activatea completely different system of signal transduction from UVA
The results suggesting specific signal transduction pathwaysfor UVBUVC in keratinocytes presented here agree well withour studies of UVB-regulated genes using DNA micro arrays(Li et al 2001) We found that in response to UVB ker-atinocytes not only induce the immediate early genes and stressresponse genes but also elaborate responses designed to pro-tect the organism underneath For example chemokines of theIL-8 family are induced which alert the surrounding tissue tothe UV damage synthesis of the cornified envelope compo-nents is enhanced which thickens the stratum corneum etc Weconclude that human epidermal keratinocytes being naturallyexposed to UV light have elaborated highly specific molecu-lar responses to UVBUVC at the signal transduction level aswell as the gene expression level
ACKNOWLEDGMENTS
Our research was supported by the National Institutes ofHealth Grants AR30682 AR39176 AR40522 and AR41850
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 675
FIG 9 Specificity in stress response Epidermal ker-atinocytes exhibit specialized UV-responsive signal transduc-tion pathways which apparently do not network and intersectwith other pathways such as those responsive to EGF or TNFaIn contrast in other cell types such as HeLa multiple path-ways crossover interact and overlap
and by the Anaderm Research Corporation Special thinks goto Dr M Simon for the generous gift of keratinocytes We alsothank members of our laboratory for advice reagents and en-couragement
REFERENCES
ALKALAY I YARON A HATZUBAI A ORIAN ACIECHANOVER A and BEN-NERIAH Y (1995) Stimulation-dependent I kappa B alpha phosphorylation marks the NF-kappa Binhibitor for degradation via the ubiquitin-proteasome pathway ProcNatl Acad Sci USA 92 10599ndash10603
BAEUERLE PA and HENKEL T (1994) Function and activationof NF-kappa B in the immune system Annu Rev Immunol 12141ndash179
BARNES PJ and KARIN M (1997) Nuclear factor-kappaB A piv-otal transcription factor in chronic inflammatory diseases N EnglJ Med 336 1066ndash1071
BEG AA FINCO TS NANTERMET PV and BALDWIN AJR (1993) Tumor necrosis factor and interleukin-1 lead to phos-phorylation and loss of I kappa B alpha A mechanism for NF-kappaB activation Mol Cell Biol 13 3301ndash3310
BENDER K BLATTNER C KNEBEL A IORDANOV M HER-RLICH P and RAHMSDORF HJ (1997) UV-induced signaltransduction J Photochem Photobiol B 37 1ndash17
BENDER K GOTTLICHER M WHITESIDE S RAHMSDORFHJ and HERRLICH P (1998) Sequential DNA damage-indepen-dent and -dependent activation of NF-kappaB by UV EMBO J 175170ndash5181
BROWN K GERSTBERGER S CARLSON L FRANZOSO Gand SIEBENLIST U (1995) Control of I kappa B-alpha proteoly-sis by site-specific signal-induced phosphorylation Science 2671485ndash1488
DELHASE M HAYAKAWA M CHEN Y and KARIN M(1999) Positive and negative regulation of IkappaB kinase activitythrough IKKbeta subunit phosphorylation Science 284 309ndash313
DEacuteRIJARD B et al (1994) JNK1 A protein kinase stimulated byUV light and Ha-Ras that binds and phosphorylates the c-Jun acti-vation domain Cell 76 1025ndash1037
DEVARY Y ROSETTE C DIDONATO JA and KARIN M(1993) NF-kappa B activation by ultraviolet light not dependent ona nuclear signal Science 261 1442ndash1445
DIDONATO JA HAYAKAWA M ROTHWARF DM ZANDIE and KARIN M (1997) A cytokine-responsive IkappaB kinasethat activates the transcription factor NF-kappaB Nature 388548ndash554
FANGER GR GERWINS P WIDMANN C JARPE MB andJOHNSON GL (1997) MEKKs GCKs MLKs PAKs TAKs andtpls Upstream regulators of the c-Jun amino-terminal kinases CurrOpin Genet Dev 7 67ndash74
FEIGE U MORIMOTO RI YAHARA I and POLLA BS eds(1996) Stress-Inducible Cellular Responses (Birkhauser VerlagBasel Switzerland)
FISHER GJ WANG ZQ DATTA SC VARANI J KANG Sand VOORHEES JJ (1997) Pathophysiology of premature skin ag-ing induced by ultraviolet light N Engl J Med 337 1419ndash1428
GHODA L LIN X and GREENE WC (1997) The 90-kDa ribo-somal S6 kinase (pp90rsk) phosphorylates the N-terminal regulatorydomain of IkappaBalpha and stimulates its degradation in vitro JBiol Chem 272 21281ndash21288
GILCHREST B (1995) Photodamage (Blackwell Scientific NewYork)
GRETHER-BECK S BUETTNER R and KRUTMANN J (1997)Ultraviolet A radiation-induced expression of human genes Molec-ular and photobiological mechanisms Biol Chem 378 1231ndash1236
GRILLI M PIZZI M MEMO M and SPANO P (1996) Neuro-protection by aspirin and sodium salicylate through blockade of NF-kappaB activation Science 274 1383ndash1385
GUPTA S et al (1996) Selective interaction of JNK protein kinaseisoforms with transcription factors EMBO J 15 2760ndash2770
HEHNER SP et al (1998) Sesquiterpene lactones specifically in-hibit activation of NF-kappa B by preventing the degradation of Ikappa B-alpha and I kappa B-beta J Biol Chem 273 1288ndash1297
HERSKOWITZ I (1995) MAP kinase pathways in yeast For matingand more Cell 80 187ndash197
HIBI M LIN A SMEAL T MINDEN A and KARIN M (1993)Identification of an oncoprotein- and UV-responsive protein kinasethat binds and potentiates the c-Jun activation domain Genes Dev7 2135ndash2148
HIRANO M et al (1996) MEK kinase is involved in tumor necro-sis factor alpha-induced NF-kappaB activation and degradation ofIkappaB-alpha J Biol Chem 271 13234ndash13238
HU Y et al (1999) Abnormal morphogenesis but intact IKK activa-tion in mice lacking the IKKalpha subunit of IkappaB kinase Sci-ence 284 316ndash320
IORDANOV M et al (1997) CREB is activated by UVC through ap38HOG-1-dependent protein kinase EMBO J 16 1009ndash1022
ISREAL A (2000) The IKK complex An integrator of all signals thatactivate NF-kappaB Trends Cell Biol 10 129ndash133
ITO CY ADEY N BAUTCH VL and BALDWIN AS (1995)Structure and evolution of the human IKBA gene Genomics 29490ndash495
JIANG CK CONNOLLY D and BLUMENBERG M (1991)Comparison of methods for transfection of human epidermal ker-atinocytes J Invest Dermatol 97 969ndash973
JIANG CK FLANAGAN S OHTSUKI M SHUAI K FREED-BERG IM and BLUMENBERG M (1994) Disease-activatedtranscription factor Allergic reactions in human skin cause nucleartranscription of STAT-91 and induce synthesis of keratin K17 MolCell Biol 14 4759ndash4769
JIANG CK MAGNALDO T OHTSUKI M FREEDBERG IMBERNERD F and BLUMENBERG M (1993) Epidermal growthfactor and transforming growth factor alpha specifically induce theactivation- and hyperproliferation-associated keratins 6 and 16 ProcNatl Acad Sci USA 90 6786ndash6790
KALLUNKI T DENG T HIBI M and KARIN M (1996) c-Juncan recruit JNK to phosphorylate dimerization partners via specificdocking interactions Cell 87 929ndash939
KARIN M (1995) The regulation of AP-1 activity by mitogen-acti-vated protein kinases J Biol Chem 270 16483ndash16486
KARIN M and HUNTER T (1995) Transcriptional control by pro-tein phosphorylation Signal transmission from the cell surface to thenucleus Curr Biol 5 747ndash757
KLAMPFER L LEE TH HSU W VILCEK J and CHEN-KIANG S (1994) NF-IL6 and AP-1 cooperatively modulate the ac-tivation of the TSG-6 gene by tumor necrosis factor alpha and in-terleukin-1 Mol Cell Biol 14 6561ndash6569
KOMINE M et al (2000) Inflammatory versus proliferative pro-cesses in epidermis Tumor necrosis factor alpha induces K6b ker-atin synthesis through a transcriptional complex containing NFkappaB and CEBPbeta J Biol Chem 275 32077ndash32088
KOPP E and GHOSH S (1994) Inhibition of NF-kappa B by so-dium salicylate and aspirin Science 265 956ndash959
KYRIAKIS JM et al (1994) The stress-activated protein kinase sub-family of c-Jun kinases Nature 369 156ndash160
LEE FS HAGLER J CHEN ZJ and MANIATIS T (1997) Ac-tivation of the IkappaB alpha kinase complex by MEKK1 a kinaseof the JNK pathway Cell 88 213ndash222
LEPPA S and BOHMANN D (1999) Diverse functions of JNK sig-naling and c-Jun in stress response and apoptosis Oncogene 186158ndash6162
ADACHI ET AL676
LI D TURI TG SCHUCK A FREEDBERG IM KHITROVG and BLUMENBERG M (2001) Rays and arrays The tran-scriptional program in the response of human epidermal keratinocytesto UVB illumination FASEB J 15 2533ndash2535
LI N and KARIN M (1998) Ionizing radiation and short wavelengthUV activate NF-kappaB through two distinct mechanisms ProcNatl Acad Sci USA 95 13012ndash13017
LIU ZG HSU H GOEDDEL DV and KARIN M (1996) Dis-section of TNF receptor 1 effector functions JNK activation is notlinked to apoptosis while NF-kappaB activation prevents cell deathCell 87 565ndash576
MERCURIO F and MANNING AM (1999) NF-kappaB as a pri-mary regulator of the stress response Oncogene 18 6163ndash6171
MIYAMOTO S and VERMA IM (1995) RelNF-kappa BI kappaB story Adv Cancer Res 66 255ndash292
NESTLE FO MITRA RS BENNETT CF CHAN H andNICKOLOFF BJ (1994) Cationic lipid is not required for uptakeand selective inhibitory activity of ICAM-1 phosphorothioate anti-sense oligonucleotides in keratinocytes J Invest Dermatol 103569ndash575
PETERS RT LIAO SM and MANIATIS T (2000) IKKepsilonis part of a novel PMA-inducible IkappaB kinase complex Mol Cell5 513ndash522
PIERCE JW READ MA DING H LUSCINSKAS FW andCOLLINS T (1996) Salicylates inhibit I kappa B-alpha phos-phorylation endothelial-leukocyte adhesion molecule expressionand neutrophil transmigration J Immunol 156 3961ndash3969
PIERCE JW et al (1997) Novel inhibitors of cytokine-induced Ikap-paBalpha phosphorylation and endothelial cell adhesion molecule ex-pression show anti-inflammatory effects in vivo J Biol Chem 27221096ndash21103
REUNING U et al (1995) Inhibition of NF-kappa B-Rel A expres-sion by antisense oligodeoxynucleotides suppresses synthesis ofurokinase-type plasminogen activator (uPA) but not its inhibitor PAI-1 Nucleic Acids Res 23 3887ndash3893
ROSETTE C and KARIN M (1996) Ultraviolet light and osmoticstress activation of the JNK cascade through multiple growth factorand cytokine receptors Science 274 1194ndash1197
SCHOUTEN GJ et al (1997) IkappaB alpha is a target for the mitogen-activated 90 kDa ribosomal S6 kinase EMBO J 163133ndash3144
SHIMADA T et al (1999) IKK-i a novel lipopolysaccharide-inducible kinase that is related to IkappaB kinases Int Immunol 111357ndash1362
SIMON M and GREEN H (1984) Participation of membrane-asso-ciated proteins in the formation of the cross-linked envelope of thekeratinocyte Cell 36 827ndash834
SIMON MM ARAGANE Y SCHWARZ A LUGER TA andSCHWARZ T (1994) UVB light induces nuclear factor kappa B(NF kappa B) activity independently from chromosomal DNA dam-age in cell-free cytosolic extracts J Invest Dermatol 102 422ndash427
TOBIN D NILSSON M and TOFTGARD R (1996) Ras-inde-pendent activation of Rel-family transcription factors by UVB andTPA in cultured keratinocytes Oncogene 12 785ndash793
TOMIC-CANIC M KOMINE M FREEDBERG IM and BLU-MENBERG M (1998) Epidermal signal transduction and tran-scription factor activation in activated keratinocytes J Dermatol Sci17 167ndash181
WILLIAMS BR (1999) PKR A sentinel kinase for cellular stressOncogene 18 6112ndash6120
WU Z WU J JACINTO E and KARIN M (1997) Molecularcloning and characterization of human JNKK2 a novel Jun NH2-terminal kinase-specific kinase Mol Cell Biol 17 7407ndash7416
YANG DD et al (1997) Absence of excitotoxicity-induced apopto-sis in the hippocampus of mice lacking the Jnk3 gene Nature 389865ndash870
YANKULOV K BLAU J PURTON T ROBERTS S and BENT-LEY DL (1994) Transcriptional elongation by RNA polymeraseII is stimulated by transactivators Cell 77 749ndash759
YIN MJ YAMAMOTO Y and GAYNOR RB (1998) The anti-inflammatory agents aspirin and salicylate inhibit the activity ofI(kappa)B kinase-beta Nature 396 77ndash80
YOSHIDA S et al (2001) The c-Jun NH2-terminal kinase3 (JNK3)gene Genomic structure chromosomal assignment and loss of ex-pression in brain tumors J Hum Genet 46 182ndash187
Address reprint requests toM Blumenberg PhD
The NYU Cancer InstituteNew York University School of Medicine
550 First AveNew York NY 10016
E-mail blumem01mednyuedu
Received for publication June 5 2003 received in revised formJuly 15 2003 accepted August 14 2003
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 677

remedies not only to protect themselves from UV but also toprotect the underlying organism If this hypothesis is correctthen UV is not just another generic cause of stress to ker-atinocytes but a specific injury with a specific set of responsesTherefore in epidermal keratinocytes UV should elicit not onlythe general stress responses characterized in other cell typesbut also additional skin-specific protective mechanisms In-deed using DNA microarrays we found keratinocyte-specificregulation of gene expression by UV light (D Li et al 2001)Therefore we decided to examine the UV responding signaltransduction pathways in human epidermal keratinocytes a rel-evant target for UV light
The major effects of photodamage have been reviewed com-prehensively (Gilchrest 1995) The cellular effects of UV con-sistently include DNA damage apoptosis and transcriptionalchanges All three wavelengths UVC (100ndash290 nm) UVB(290ndash320 nm) and UVA (320ndash380 nm) cause DNA damageUVC is not present in terrestrial light UVB and UVC have sim-ilar molecular and biological effects in human skin whereas themolecular effects of UVA are different (Grether-Beck et al1997) The inherent responses of keratinocytes to UV by anal-ogy with responses to other extracellular signals can be sepa-rated into two phases the immediate which does not require denovo protein synthesis and delayed which depends on newly in-duced proteins The immediate phase comprises the cytoskeletalchanges and UV-specific signal transduction cascades which de-pends on protein phosphorylation and results in activation of tran-scription factors whereas the delayed phase consist of tran-scriptional and cytoskeletal changes (Hibi et al 1993 Rosetteand Karin 1996 Iordanov et al 1997 Bender et al 1998)
A major impetus for studies of the molecular response to UVcame with the identification of c-Jun amino-terminal kinases(JNK) the protein kinase that in response to UV binds to andactivates transcription factor c-Jun (Hibi et al 1993) Soon itwas realized that JNK responds to several stress signals in ad-dition to UV such as osmotic shock arsenate and proinflam-matory cytokines (Deacuterijard et al 1994 Kyriakis et al 1994)Three isoforms JNK1 JNK2 and JNK3 are often expressedin two splice variants designated ldquoardquo and ldquobrdquo (Karin 1995)JNK1 is a 46 kDa protein that when activated enters the nu-cleus and phosphorylates Ser63 and Ser73 of c-Jun The tran-scription factors activated by JNK are Elk1 SAP1 ATF2PEA3 and the eponymous Jun (Gupta et al 1996) Whilebound to c-Jun JNK1 can phosphorylate the Jun dimerizationpartners JunB JunD and ATF2 (Kallunki et al 1996) Thesignal-transducing cascade that activates JNK is analogous tothe Ste (for sterile) pathway in yeast the mating pheromone-responsive signal transduction cascade (Herskowitz 1995)JNK can be activated by one of two kinases designated JNKK1(aka SEK1 SKK1 and MKK4) and JNKK2 (MKK7) (Wu etal 1997) JNKKs in turn are activated by multiple JNKKKskinases that respond to a variety of stimuli (Fanger et al 1997)The UV-responsive JNKKK has not been identified yet
Another clear molecular effect of UV is the activation of theNFkB transcription factor (Barnes and Karin 1997 N Li andKarin 1998) The NFkBRel family of transcription factorsplays a particularly important role in the inflammatory and an-tiapoptotic processes (Baeuerle and Henkel 1994 Miyamotoand Verma 1995 Liu et al 1996 Tomic-Canic et al 1998)The activation of NFkB by UV is not associated with DNA
damage and occurs even in cytoplasts devoid of nuclear DNA(Devary et al 1993 Simon et al 1994) Inactive NFkB re-sides in the cytoplasm in complex with IkB protein Upon ac-tivation by diverse extracellular stimuli a large multiproteincomplex containing IKKa and IKKb phosphorylates IkB (Beget al 1993 Alkalay et al 1995 DiDonato et al 1997) Re-cently knockout mice of both IKKa and IKKb have been de-scribed (Delhase et al 1999 Hu et al 1999) It appears thatIKKb is responsible for cytokine signaling in most tissueswhile specifically in the epidermis IKKa is essential for nor-mal keratinocyte differentiation The mechanisms by which inflammatory cytokines such as TNFa or IL-1 as well as ion-izing radiation activate NFkB are known to involve phospho-rylation of two specific serine residues (Ser-32 and Ser-36)within the N-terminal regulatory domain of IkBa which is fol-lowed by polyubiquitination and degradation of IkB by a 26Sproteasome (Beg et al 1993 Alkalay et al 1995 Brown etal 1995 Bender et al 1998) The proteolysis of IkB resultsin the release of NFkB which is then free to enter the nucleusand activate gene transcription
Although UV can cause a release of TNFa and so indirectlyactivate the above pathway in direct response to UV an uniden-tified kinase activates NFkB (Bender et al 1998 N Li andKarin 1998) The direct UV-induced activation of NFkB maynot involve IKKb and is not dependent on phosphorylation atSer-32 and Ser-36 of IkBa (Bender et al 1998 Li and Karin1998) These experiments were performed with HeLa cellswhich are not a natural target of UV light
Several specific inhibitors of the NFkB signaling pathwayhave been described BAY11-7085 inhibits the cascade up-stream from IKK salicylates widely used anti-inflammatorydrugs have been recently identified as specific inhibitors ofIKKb while parthenolide directly inhibits the degradation ofIkBa (Kopp and Ghosh 1994 Grilli et al 1996 Pierce et al1996 Hehner et al 1998 Yin et al 1998)
Hypothesizing that skin has specific mechanisms to protectitself and the organism from UV damage we examined theMAPK and NFkB signaling pathways in human epidermal ker-atinocytes We found that in keratinocytes the activation of theJNK pathway by UV critically depends on JNKK1 and thatkeratinocytes express an unexpected assortment of JNK iso-forms Most important in keratinocytes unlike in other celltypes we examined the activation is very stringent UVB andUVC activate JNK while UVA does not Furthermore UVBand UVC do not activate parallel pathways and there is nocrosstalk between the UVB and UVC-dependent pathway andthe growth factor-dependent pathways Using specific inhibi-tors as well as antisense approaches we also compared the path-ways that activate NFkB in response to UV or TNFa in ker-atinocytes and HeLa cells We found that UVC and UVBactivate NFkB in both cell types but that UVA has no effectin either In addition we have shown that UVC and UVB usethe same signal transduction pathway to induce NFkB and thatthis pathway does not involve IKKb while TNFa uses a dif-ferent pathway to activate NFkB through IKKb Finally usingantisense oligonucleotides to target NFkB and IKKb we haveconfirmed that IKKb is essential for activation of NFkB byTNFa but plays no role in activation of NFkB by UVBUVCWe conclude that there is an independent UV responsive sig-nal transduction pathway activating NFkB in epidermis
ADACHI ET AL666
MATERIALS AND METHODS
Plasmids their growth and purification
The plasmid containing (NFkB)3-CAT was a gift from JVilcek NYU School of Medicine plasmid expressing Gal4Elkand GAL4CAT were a gift from J Blau Rockefeller Univer-sity and the control plasmids pRSVZ has been described pre-viously (Jiang et al 1993 1994 Klampfer et al 1994) Plas-mids were grown in JM101 Escherichia coli host to saturationdensity in LB medium DNAs were extracted and purified us-ing the Mega Prep kit from Promega (Madison WI)
Cell growth maintenance and transfection
Normal human foreskin epidermal keratinocytes were a gen-erous gift from Dr M Simon The cultures were initiated us-ing 3T3 feeder layers as described (Simon and Green 1984)and then frozen in liquid N2 until used Once thawed the ker-atinocytes were grown without feeder cells in defined serum-free keratinocyte medium supplemented with epidermal growthfactor and bovine pituitary extract (keratinocyte-SFM GIBCOCarlsbad CA) Cells were expanded through two 14 passagesbefore transfection and transfected at 90 confluency HeLacells were maintained in Dulbecco-modified Eaglersquos mediumsupplemented with 10 calf serum At the beginning of theseexperiments we optimized the doses of UV and concentrationsof EGF and TNFa to maximize the activation of the reportersin both HeLa cells and epidermal keratinocytes without killingthe cells (data not shown) All subsequent experiments wereperformed with optimal doses and concentrations
Transfections of keratinocytes with polybrene and DMSOshock were used to insert the DNA into the subconfluent ker-atinocytes as previously described (Jiang et al 1991) On theday of transfection cells were washed and incubated in the ba-sal medium without EGF and pituitary extract Each transfec-tion contained 10 mgdish of (NFkB)3-CAT and 3 mgdish ofpRSVZ The cells were incubated at 37degC in a 5 CO2 atmo-sphere and 24 h after transfection were illuminated with UVAUVB UVC or stimulated with TNFa EGF (100 ngml) in thepresence or absence of various inhibitors Treatment with as-pirin (0ndash100 mM from Sigma St Louis MO) BAY (0ndash5 mM)parthenolide (0ndash15 mM) and U0126 (0ndash1 mM all from BiomolPlymoth Meeting PA) lasted for another 24 h before harvest-ing We used two StratalinkerTM units (Stratagene La JollaCA) one equipped with FG15T8 bulbs as our UVB source andstandard GL-15 G15T8 bulbs as a source of UVC Before illu-mination the medium was removed from the cells and imme-diately after the illumination the same medium was restored tothe cells The whole illumination procedure takes less than aminute which we have shown does not cause drying or a heatshock The control cultures were treated identically except theactual illumination was omitted Cells were washed twice withphosphate-buffered saline 48 h after transfection and harvestedby scraping The cell disruption by repeated freezendashthaw cy-cles has also been described (Jiang et al 1991) All transfec-tions were performed in duplicate plates and each transfectionexperiment was repeated three to five times
HeLa cells were transfected using a modified calcium phos-phate precipitation procedure (Jiang et al 1991) Two days be-fore transfection cells were split by trypsinization and trans-
fected at 50 confluency Ten micrograms of the reporter plas-mid and 3 mg of the b-gal control plasmid were dissolved in atotal volume of 450 ml H2O To this mixture 50 ml of 25 MCaCl2 was added and mixed carefully Five hundred microlitersof BES-buffer pH 695 (50 mM N-Bis-2-hydroxyethyl-2-aminoethanesulfonic acid 250 mM NaCl 15 mM Na2HPO4)was added and mixed by inverting the tube The reaction mix-tures were incubated at room temperature for 10ndash15 min Onemilliliter of the precipitate suspension was added per platewhich was followed by incubation at 37degC in a 5 CO2 atmo-sphere Twenty-four hours after transfection cells were stimu-lated by TNFa or illuminated with UVA UVB or UVC withor without various inhibitors as indicated Cells were washedtwice with phosphate-buffered saline 36 to 48 h after transfec-tion and harvested by scraping
Enzyme assays
For b-galactosidase assays the substrate solution contained6 mg of O-nitrophenyl-D-galactoside (Sigma St Louis MO)freshly dissolved in PM buffer (66 mM Na2HPO4 33 mMNa2HPO4 40 mM mercaptoethanol 2 mM MgSO4 and 01 mMMnCl2) The reaction mixture contained 100 ml of substrate so-lution 300 ml of PM2 buffer and 50 ml of keratinocyte cell ex-tract or 20ndash30 ml of HeLa cell extract It was incubated at 37degCuntil development of a yellow color was obvious usually at 30min to 1 h The time of the reaction was recorded and the re-action stopped by addition of 04 ml of 1 M Na2CO3 OD420
was measured on a spectrophotometer (Labrepco HorshamPA)
The CAT reaction mixture contained 69 ml of 1 M Tris HClpH 78 1 ml of 14[C]-chloramphenicol (Sigma) 20 ml of 4 mMacetyl-CoA solution 30ndash60 ml of cell extract and enough wa-ter to bring the total reaction volume to 150 ml After incuba-tion at 37degC for 1 h the mixture was extracted into 1 ml ofethyl acetate phases were separated by brief centrifugation theorganic layer was transferred to a new tube and the solventevaporated The residue was dissolved in 25 ml of ethyl acetateand separated by thin layer chromatography on a silica gel platein chloroformmethanol 5 955 The plates were exposed to X-ray film for 12 to 24 h and the intensity of radioactive spotsdetermined using the Ambis Radioanalytic System (AmbisRochester NY) The conversion of chloramphenicol to itsmonoacetylated derivative was kept below 50 by varying theamount of extract or the duration of the reaction All CAT val-ues were normalized for transfection efficiency by calculatingthe ratio of CAT activity to b-galactosidase in each transfectedplate Each transfection experiment was separately performedthree or more times with each data point resulting from dupli-cate transfections
Use of antisense and sense oligonucleotides
Keratinocytes readily take up offered DNA Specificallyshort oligonucleotides can be introduced into these cells evenin the absence of cationic lipid (Nestle et al 1994 Reuning etal 1995) The oligonucleotides were synthesized in the phos-phorothiolate form to prolong their half-lives in the cells Wetargeted the antisense oligonucleotides to sequences includingand immediately upstream from the initiation codon for themost efficient antisense blocking At the same time we pre-
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 667
pared and used sense oligonucleotides as negative controls Ourapproach was to add the antisense oligonucleotides to the DNAtransfected into cells including the oligonucleotides into thetransfection solution which has the advantage of ensuring thatthe cells that received the transfected DNA also received theoligonucleotides The oligonucleotides were stored at 270degCin water until use and were added to the transfected DNA mix-ture In most cases we used 4 mg of the oligonucleotide pertransfection Their sequences are given in Table 1
Western blotting
The activated JNK antibody was purchased from Calbiochem(San Diego CA) and used at 1100 to 11000 dilution whilethe antirabbit immunoglobulin horseradish peroxidase-linkedantibodies and the ECL reagent were used as suggested by themanufacturer (Amersham Piscataway NJ) The anti-JNK3SAPK 1b antibody was purchased from Upstate Biotech-nology (Lake Placid NY) We used the antibody that recog-nizes JNK3 a1 (46KDa) and JNK3a2 (54KDa) at 11000dilution and the A431 cell lysate (Upstate Biotechnology) aspositive control according to manufacturerrsquos recommendationKeratinocytes were grown as described above At their thirdpassage at 80 confluence we incubated keratinocytes inKBM for 16 h to starve them of growth factors and then treatedas described above At various time intervals cells were har-vested by scraping washed twice with PBS and the cell pelletwas resuspended in 100 ml of the lysis buffer (2 SDS 10glycerol 10 mM Tris HCl pH 68) The cells were lysed by ul-trasonication for 30 sec using an ultrasonic homogenizer soni-cator (Cole-Parmer Vernon Hills IL) and boiling for 10 minAliquots of the supernatant obtained after centrifugation(2000 3 g 10 min) were stored at 270degC Protein content wasdetermined using the Bradford reagent (Perkin-Elmer BostonMA) Equal amounts of total protein were resolved by sodiumdodecyl sulphate-polyacrylamide gel electrophoresis (10 gel)
and electrophoretically transferred to nitrocellulose at 8 V for1 h After blocking the membranes with 5 nonfat dry milk inTris-buffered saline with 01 Tween (TBS-T) for 1 h at roomtemperature on an orbital shaker the blots were washed thrice(10 min each time) with TBS-T and incubated at 4degC with11000 dilution of the antibody Blots were washed 16 h laterwith TBS-T (3 3 10 min) and incubated at room temperaturewith 11000 dilution of antirabbit immunoglobin horseradishperoxidase-linked antibody for 2 h The chemiluminescenceECL reagent was used to develop the blots after washing themembranes three times 10 min with TBS-T and twice withTBS alone
JNK activity blots
Untreated control cells and cells treated with UV or EGFwere harvested 30 min after treatment The JNK activity wasmeasured by an in vitro kinase assay that detects phosphoryla-tion of a GST-Jun fusion protein The GST-Jun expressing con-struct was a generous gift from Dr Susan Logan New YorkUniversity School of Medicine This construct generates a fu-sion protein containing the 79 amino-terminal residues of c-junThe fusion protein GST-jun was used to purify JNK from thecell lysates and subsequently as a substrate for the same ki-nase The kinase assay was performed in kinase assay buffer(20 mM HEPES pH 75 20 mM beta-glycerol-phosphate 10mM MgCl2 10 mM DTT 50 mM sodium-orthovanadate 20mM ATP and 05 mlreaction of 32P-ATP (New England Nu-clear Amherst MA) The product of the kinase reaction wasthen separated by SDS-PAGE and the 32P-labeled GST-jun de-tected using autoradiography
RT-PCR
Cultured cells were harvested and total RNAs isolated uti-lizing RNeasy RNA extraction kit from Qiagen Between 1 and
ADACHI ET AL668
TABLE 1 OLIGONUCLEOTIDES USED
RT-PCR oligonucleotides used in detecting JNK ERK and p38 mRNAsJNK1 forward TCGCTACTACAGAGCA (570 bp expected product)JNK1 reverse GGAGAGGGCTGCCCCCJNK2 forward CTGGCCCGGACAGCGT (360 bp)JNK2 reverse GTTTTTATTTTGTCTCJNK3 forward TTATTACAGAGCCCCT (537 bp)JNK3 reverse TTTAGTCTTTTCTTCTGERK1 forward TTGCAGATCCAGACCA (295 bp)ERK1 reverse ATTTTTGTGTGGAAGAp38 forward GACGGATCGCAGAAGA (397 bp)p38 reverse TGCCAGGCACCCCGGT
Antisense oligonucleotides used in transfection of keratinocyte synthesized as phosphorotiolatesas-JNK1 CATGATGGCAAGCAATTAATGs-JNK1 GTACTACCGTTCGTTAATTACas-JNKK1 CATTGTTGGGAGTGAAGAGCCCAs-JNKK1 GTAACAACCCTCACTTCTCGGGTas-NFkB TGGATCATCTTCTGCCATTCTs-NKkB ACCTAGTAGAAGACGGTAAGAas-IkB CGCGTCGGCCGCCTGGAACATGCas-IKKa CATGGGGCGGGAGGGCAAGCGGCCas-IKKb ACTGATGTCGTGCTAACTCTGTCGs-IKKb TGACTACAGCACGATTGAGACAGC
15 mg of RNA were subjected to RT-PCR with Access RT-PCR kit system from Promega (Madison WI) for 32 cycleswith buffers and chemicals added as suggested by the manu-facturer In the supernumerary PCR experiments we used 42cycles The PCR products were subjected to either agarose orpolyacrylamide gel electrophoresis visualized with ethidiumbromide (Sigma) with a UV transilluminator from UVP andphotographed (Polaroid Waltham MA) The densities of bandswere quantified utilizing image scanner (GT-9000 Epson LongBeach CA)
Organ culture explants of normal human skin
Pieces of normal human skin were obtained immediately af-ter surgery They were cut into pieces approximately 3ndash5 mm3and incubated in keratinocyte basal medium KBM (Keratino-cyte-SFM Gibco-BRL Carlsbad CA) with or without TNFa
(50 ngml Intergen Norcross GA) in a humidified incubatorat 37degC for 24 h Generally we used 24-well culture disheswith up to five pieces in the same well and just enough mediumto cover the explants The antisense oligonucleotides wereadded at 300 ngml and when necessary replaced every 2 daysIllumination was performed epidermal side up with 100mJcm2 of UVB The explants were mounted in tissue Tec-OCTcompound (Sakura Finetek Japan) and frozen Sections 4 to 6mm thick were obtained with a cryostat (Miles LaboratoriesResearch Triangle Park NC) fixed with methanolacetone for10 min incubated with anti-JNK1 antibody (Calbiochem SanDiego CA) at 4degC overnight treated with peroxidase-conju-
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 669
FIG 1 UV activates JNK in keratinocytes The two left pan-els show the UV dose curve with each sample loaded in du-plicate The right panels show the time course of activation af-ter the illumination The top panels show the kinase activity ofJNK The activity was shown using GSTndashJun fusion protein asa substrate for immunoprecipitated JNK The bottom panelsshow the corresponding Western blots using antibody specificfor the activated phosphorylated form of JNKs The trianglepoints to the phosphorylated JNK substrate the doublet of openarrows to the JNK proteins The diffuse band in the top panelsis an uncharacterized artifact In the left panels we harvestedthe cells 10 min after irradiation and in the right panels weused 10 mJcm2 In each well we loaded 10 mg of protein
FIG 2 RT-PCR identification of JNK isotypes in keratinocytes HeLa cells and fibroblasts (a) Lanes K H and F representRNA from untreated keratinocytes HeLa cells and fibroblasts respectively Isotype-specific oligonucleotides were used in RT-PCR reactions The arrows point to the specific bands (b) Exaggerated PCR reactions for detecting low abundance mRNAs Du-plicate lanes show results of PCR reaction without (2) and with (1) input RNA preformed with supernumerary cycles (c) West-ern blots showing the presence of JNK3 lane 1 positive control (A431 extract) lane 2 HeLa cells and lane 3 keratinocytes
gated antimouse IgG secondary antibody (Vecstatin ABC-mouse IgG kit from Vector Laboratories Burlingame CA) atroom temperature for 1 h incubated with ABC complex (Vec-tor Laboratories) at room temperature for 1 h and treated with339-Diaminobenzidine-tetrahydrochloride (Dojindo Gaithers-burg MD) and 001 H2O2 in Tris pH 76 for 2 min The sam-ples were observed and photographed under the light micro-scope (Microphot-FXA Nikon Melville NY) Additionalantibodies used were from SantaCruz Biotechnology (SantaCruz CA)
RESULTS
UVB signal transduction pathway part I activation ofthe JNK pathway
To determine which JNK isoforms exist in keratinocytes andcan be activated by UVB we have grown primary cultures ofhuman epidermal keratinocytes and illuminated them with UVBat various doses As controls we used HeLa cells and normalhuman dermal fibroblasts The JNK activity was examined inthree ways First illuminated cells were stained with pan-JNKor active JNK-specific antibody to demonstrate the transloca-tion of JNK from cytoplasm to nucleus (not shown) Secondprotein extracts were prepared and analyzed using Western blotswith active JNK-specific antibody Third the kinase activitywas determined directly in affinity-precipitated JNK using aGST-tagged Jun protein fragment as the substrate The resultsof all three approaches were perfectly consistent UVB stronglyand specifically activates JNK in human epidermal ker-atinocytes (Fig 1)
Three isotypes of JNK have been described each with sev-eral splice variants It should be noted that there is confusionregarding which isotype is JNK2 and which is JNK3 in the lit-erature and even more egregiously in the blurbs of manufac-turers who provide antibodies We have adopted the unam-biguous designation of the NCBI databank To determine whichisotypes are expressed in keratinocytes we developed a set ofspecific RT-PCR oligonucleotides and used them to amplify theJNK mRNAs Surprisingly we found JNK1 and JNK3 in ker-atinocytes while JNK2 was absent (Fig 2a) This is an unex-pected finding because JNK1 and JNK2 are commonly almostubiquitously expressed while JNK3 was thought to be brain-specific (Yang et al 1997) Repeat of the PCR assay using alarge number of cycles confirmed the absence of JNK2 in ker-atinocytes (Fig 2b) Furthermore because of the importanceand unexpectedness of this result we decided to confirm theexpression of JNK3 protein in keratinocytes using Western blot-ting Indeed we find that JNK3 is expressed in keratinocytesbut not in HeLa cells (Fig 2c) Therefore keratinocytes expressboth the mRNA and the protein of JNK3 a kinase previouslythought to be brain-specific
To demonstrate that the activation of JNK is functional inthat it activates the Elk1 transcription factor in keratinocyteswe transfected into these cells a set of plasmids containing aGal4-CAT reporter and Gal4-DBDElk1-AF (Yankulov et al1994) The first plasmid contains a set of Gal4 binding sites up-stream from the CAT reporter gene the second expresses a fu-sion protein containing the Gal4 DNA binding domains fused
to the activating domain of transcription factor Elk1 When ac-tivated by UV via JNK the fusion protein activates transcrip-tion but if not activated it is silent Thus the system serves asan UV-responsive transcription reporter The transfected cellswere illuminated with UV and harvested 16 h later We foundthat both UVC and UVB activate the reporter but UVA doesnot (Fig 3) Note that at the 50 mJcm2 dose of UVC a signif-icant number of cells are killed These results mean that theJNK system in keratinocytes is present functional and specif-ically activated by short wavelength UV light UVBUVC butnot the long wavelength UVA We conclude that UVA doesnot activate the same system of signal transduction thatUVBUVC do
To identify the kinases responsible for UV signal we usedantisense approaches The antisense oligonucleotides were tar-geted against JNK1 and JNKK1 After 16 h to allow the anti-sense oligonucleotides to inhibit the synthesis of their targetsthe cells were irradiated with UV and after another 24 h har-vested for CAT determination We found that both antisensereagents attenuated the signals (Fig 4a and b) The antisenseeffects are specific because the sense oligonucleotides used ascontrols had no effect These results indicate that JNK1 andJNKK1 are essential for the transmission of the UVBUVC sig-
ADACHI ET AL670
FIG 3 Activation of transfected Elk-CAT reporter by UVBand UVC in keratinocytes GAL4-Elk1 fusion protein express-ing plasmid was cotransfected into keratinocytes with aGAL4CAT reporter The top panel shows the two cotransfectedplasmids and the effect of UV The transfected cells were illu-minated with indicated doses of UVA UVB or UVC harvested16 h later and the CAT levels determined Note that at the 50mJcm2 of UVC a significant number of cells is killed whichresults in significantly reduced levels of CAT activity (aster-isk) At the doses used UVA did not affect cell viability Thebars represent averages of duplicate transfections and the repli-cates differed by 25 or less
nal However they do not formally exclude the possibility thatJNK3 and JNKK2 also play a role
To determine whether the Elk1 activation is specific for theUV responsive signals in keratinocytes we tested the effects ofEGF In other cell types EGF can act through pathways thatactivate both JNK and Elk1 However when we attempted toactivate Elk1 using EGF in keratinocytes we were unsuccess-ful (Fig 4b) This means that in keratinocytes the EGF and UVresponsive pathways do not overlap on the contrary UV specif-ically activates the JNK pathway This suggests that the epi-dermal keratinocytes exhibit distinctive specialized UV-responsive signal transduction pathways
Indeed when we compared the responses to UV in severalcell types we found that the response in keratinocytes is sig-nificantly more stringently controlled than in other cell typesFor example both UV and EGF activated the JNK pathway inHeLa cells (Fig 4c and d) demonstrating a lack of specificityMoreover targeting JNK1 and JNKK1 with antisense oligonu-cleotides only partially inhibited the UV effects in HeLa cells
implying that in these cells parallel pathways can respond toUV to activate Elk1 (Fig 4c) Suspecting that EGF activatesElk1 through both the JNK and the ERKMAPK pathways inHeLa we inhibited the latter with U0126 a specific MEK-12inhibitor We found that U0126 significantly blocks the acti-vation by EGF but not by UV (Fig 4e) again demonstratingrelaxed stringency in HeLa cells
To approach these phenomena from another angle we usedkinase assays and Western blots Consistent with the above re-sults we found that while in keratinocytes only UVB (andUVC) could activate the JNK pathway in HeLa cells (and fi-broblasts not shown) heat shock UVA and osmotic shockcould activate the pathway as well (Fig 5a) In HeLa cells andfibroblasts UVB could activate the ERK pathway (not shown)but in keratinocytes it could not (Fig 5b) We conclude fromthese results that keratinocytes being often exposed to ambi-ent UV have developed more stringent and specific responsesto UV than other cell types which are not commonly exposedto UV These data are summarized in Figure 5c
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 671
FIG 4 UV specifically activates JNK in keratinocytes but not in HeLa cells GAL4-Elk1 and GAL4CAT plasmids were co-transfected into keratinocytes (a and b) or HeLa cells (c d and e) The transfected cells were treated with either UVC (5 mJcm2)or EGF (100 ngml) and harvested 24 h later for determination of CAT activity U0126a specific inhibitor of the MAPK path-way blocks the effects of EGF but not of UV Open bars represent the untreated control cells closed bars the UV-treated onesand the shaded bars the EGF treated ones
While there are only two JNKKs kinases that directly phos-phorylate JNK there is a large family of JNKKKs kinases thatphosphorylate JNKKs In a manuscript in preparation we de-scribe our analysis of the kinases expressed by keratinocytes
To demonstrate that the JNK system is responsive in vivowe used a novel ex vivo approach Biopsies of human skin weretreated with as-JNKK1 oligonucleotide or left untreated as con-trol Then both the treated and untreated biopsies were illumi-nated with UVC and shortly thereafter processed for immuno-histochemical detection of activated JNK (Fig 6) We found inthese biopsies that UV caused activation and nuclear translo-cation of JNK Furthermore this activation was abrogated bypretreatment with as-JNKK1 the antisense oligonucleotide tar-geting JNKK1 again demonstrating that JNKK1 is essential foractivation of JNK by UV in epidermis These results mean that
the specific signal transduction pathways we found operationalin cultured keratinocytes are also functional and working thesame way in human skin in vivo
UVB signal transduction pathway part II activationof the NFkB pathway
Having established that keratinocytes possess a specific UV-responding pathway to activate JNK we wanted to examine thepossibility that the UV-specific signal transduction pathwaysactivating NFkB are also specific in these cells The canonicalactivator of NFkB is TNFa however recent evidence suggeststhat UV and TNFa may activate NFkB through different mech-anisms (N Li and Karin 1998) Demonstrating a UV-specificpathway would strengthen even more our hypothesis that thereis specificity in UV responsive signal transduction in humanepidermal keratinocytes Therefore we transfected into ker-atinocytes and into HeLa cells a reporter plasmid containingthree tandem NFkB sites linked to the TK promoter-drivenCAT We found that the exposure of transfected cells to TNFaUVB or UVC resulted in the induction of the NFkB responseelement (Fig 7) Generally at the optimal conditions the in-duction of NFkB by UV was weaker than the induction byTNFa On the other hand NFkB was not activated by UVA atall even at high doses (Fig 7)
To compare the mechanisms of NFkB activation by UV andTNFa we have used a battery of specific inhibitors of the NFkBpathway which inhibit the pathway at different points (Fig 8a)Namely Bay11-7085 inhibits upstream from IKK (Pierce et al1997) aspirin has recently been found to be a competitive in-hibitor of ATP binding to IKKb (Yin et al 1998) whereasparthenolide directly inhibits the degradation of IkBa (Pierceet al 1997) When we treated transfected keratinocytes or HeLacells with these three inhibitors we found that Bay11-7085 in-hibited the induction of NFkB by TNFa in a dose-dependentmanner while it has almost no effect on the induction of NFkBby UVC (Fig 8b) The same results were obtained by using as-pirin (Fig 8c) In contrast parthenolide inhibited both the UVCand the TNFa effects in parallel (Fig 8d) These results con-firm that TNFa and UVC use different pathways to activateNFkB in HeLa cells Specifically while both UVC and TNFaactivate NFkB the TNFa signal goes via IKKs but the UVCsignal does not From these data we conclude that the conver-gence of the UVB and TNFa signals occurs downstream fromIKKs that is at the modification of IkB
One caveat in the use of inhibitors of specific kinases is thepossibility that such inhibitors may affect additional pathwaysnot currently recognized as affected by the given inhibitor Toaddress such specificity concerns and to examine the involve-ment of IKK in the induction of NFkB by TNFa UVC andUVB we used antisense oligonucleotides specifically targetingNFkB and IKKb as-NFkB as-IKKb (Table 1) (Reuning etal 1995 Komine et al 2000) In parallel we added the senseoligonucleotides s-NFkB s-IKKb as controls The antisenseexperiments confirmed the fact that UVBUVC use a clearlydifferent pathway to activate NFkB than TNFa does The sup-pression of NFkB synthesis in cultured keratinocytes by addingas-NFkB resulted in inhibition of activation of NFkB by allthree agents UVC UVB and TNFa as expected (Fig 8b) Onthe other hand the suppression of IKKb synthesis by adding
ADACHI ET AL672
FIG 5 Specific activation of JNK in keratinocytes andpromiscuous activation of JNK in HeLa cells (a) JNK activityin keratinocytes and HeLa cells treated with UVC UVA heatshock or EGF Duplicate experiments were performed with allagents except EGF Each lane contains 10 mg of protein InHeLa cells (bottom panel) UVC and EGF strongly while UVAand heat detectably activate JNK Cells were harvested 30 minafter treatment (b) Lack of activation of ERK by UVB in ker-atinocytes The top panel shows a Western blot with the anti-body specific for the activated ERKs the bottom for totalERK1 as a control for loading and transfer (c) Specificity insignal transduction In keratinocytes UVB specifically acti-vates JNK and EGF specifically activates ERKs whereas inHeLa cells all three extracellular signals UVB EGF and heatactivate both JNK and ERK Fibroblasts respond similar toHeLa cells (data not shown)
as-IKKb only inhibited the activation by TNFa but had no ef-fect on the activation by either UVC or UVB (Fig 8b) In theseexperiments we used s-NFkB and s-IKKa as controls and con-firmed that they had no effect in this pathway (Fig 8b) Fur-thermore as-NIK targeting a kinase functioning upstream fromIKK also partially inhibited the effects of TNFa Therefore
the antisense oligonucleotide experiments confirm our conclu-sions that the activation of NFkB by TNFa depends on the IKKpathway whereas the response to UVC or UVB occurs througha mechanism independent of IKKs
Furthermore as an additional control we used oligonu-cleotides targeting the JNK pathway as-JNKK1 and s-JNKK1Although they affected the JNK pathway (see Figs 4 and 6)they did not affect the NFkB pathway (Fig 8b) which indi-cates that the activation of the JNK pathway and the NFkBpathway are independent in keratinocytes This means that epi-dermal keratinocytes contain multiple specific UV responsivemechanisms
DISCUSSION
Our data suggest that UVBUVC illumination in epidermalkeratinocytes is not a generic stress response inducer but a spe-cific extracellular agent that activates specific signal transduc-tion pathways In particular we show that in keratinocytes UVexclusively activates the JNK pathway while in HeLa cellsUV also activates the parallel ERK and p38 pathways Con-versely in keratinocytes exclusively UV activates the JNKpathway while in HeLa cells growth factors can do so as wellIn parallel in keratinocytes UV uses a distinct pathway to ac-tivate the NFkB transcription factor a pathway separate fromthe more common TNFa activated one (Fig 9)
The specific activation of JNK by UVBUVC is particularlyrelevant because the JNK pathway is in different cell types atarget of multiple stress agents The balance between apoptosisand proliferation is at times regulated by JNK and the tran-scription factors activated by JNK regulate the expression ofmany genes (Karin and Hunter 1995) It is uniquely importantin epidermis that JNK exclusively responds to UVBUVC be-cause epidermis is the natural target of UVBUVC and the JNK-
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 673
FIG 6 Activation of JNK in vivo and the effect of as-JNKK1 In the control sample (left) JNK is not activated as shown us-ing an antibody specific for the activated phosphorylated form of JNK Treatment with 100 mJcm2 causes strong activation ofJNK (middle) Pretreatment for 3 h with the antisense oligonucleotide targeting JNKK1 abolishes this activation (right)
FIG 7 TNFa UVB and UVC activate NFkB while UVAdoes not Reporter plasmid NFkB3-CAT (top) which containsthree tandem NFkB sites upstream from the TK promoter wastransfected into keratinocytes The transfected cells were eithertreated with 50 ngml of TNFa or illuminated with indicateddoses of UV harvested 16 h later and the CAT levels deter-mined Untreated cells served as a control The replicates dif-fered by 25 or less
ADACHI ET AL674
FIG 8 Specific inhibitors can differentiate between the TNFa-and UVB-dependent signal transduction pathways (a) The effectsof specific inhibitors on NFkB activation Cultured keratinocyteswere treated with either UVB (8 mJcm2) or TNFa (50 ngml) af-ter transfection of the NFkB response element Inhibitor Bay11-7085 specifically inhibits upstream kinases of IKK aspirin inhibitsIKKb parthenolide blocks IkBa degradation (b) the effects of an-tisense oligonucleotides targeting signaling cascades on NFkB ac-tivation in keratinocytes Oligonucleotide as-NFkB targeting thetranscription factor blocks the activation by UVB and UVC whileoligonucleotides targeting IKKb JNK1 and JNKK1 are withouteffect (top two panels) In contrast oligonucleotides targetingIKKb and NIK partially or completely block the effects of TNFa(bottom panel) Note that the sense oligonucleotides (s-NFkB s-IKKb) used as controls had no effect on NFkB activation (c)Schematics of NFkB pathways activated by UV light and TNFaIn keratinocytes the two extracellular stimuli work through inde-pendent pathways that merge at the NFkBIkB complex
regulated transcription factors mediate both photodamage andits alleviation (Fisher et al 1997)
In keratinocytes the EGF and UV responsive pathways donot overlap but UV specifically activates the JNK pathwayThis suggests that the epidermal keratinocytes exhibit distinc-tive specialized UV-responsive signal transduction pathwaysIndeed when we compared the responses to UV in several celltypes we found that the response in keratinocytes is signifi-cantly more stringently controlled than in other cell types Forexample HeLa cells demonstrated a lack of specificity Usinga specific MEK-12 inhibitor we again demonstrated relaxedstringency in HeLa cells We conclude from these results thatkeratinocytes being often exposed to ambient UV have de-veloped more stringent and specific responses to UV than othercell types which are not commonly exposed to UV
At present we do not understand the relevance of JNK3 ex-pression in keratinocytes This isoform it was reported isspecifically expressed in the brain (Yoshida et al 2001) Wespeculate that perhaps the sequestration of JNK1 by the speci-ficity of the UV response creates a need for JNK3 as the moregeneral less restrictive stress signal transducer
TNFa one of proinflammatory cytokines important in cuta-neous inflammation activates NFkB via IKKs kinases that canbe activated by MEKK or NIK (Hirano et al 1996 Lee et al1997) In HeLa cells the phosphorylation of IkBa at Ser-32and Ser-36 residues is essential for the IKK mediated IkBa
degradation caused by TNFa IL-1 or ionizing radiation Us-ing specific inhibitors and antisense oligonucleotides we foundthat the NFkB activation by UVBUVC does not involve IKKs
The Ser-32 and Ser-36 residues are not involved in the UV-induced IkBa degradation (Bender et al 1997 N Li and Karin1998) Because IKKb specifically phosphorylates IkB at bothSer-32 and Ser-36 of IkBa these facts support our results Re-cently it was suggested that UV activation of NFkB in HeLacells may involve the RasndashRaf pathway although not in ker-atinocytes (Tobin et al 1996) This leads to the question howUV activates NFkB in keratinocytes Two new kinases IKKlaquo
and pp90rsk have been found to phosphorylate IkBa (Ghodaet al 1997 Schouten et al 1997 Shimada et al 1999 Israel2000 Peters et al 2000) However IKKlaquo phosphorylates theSer-36 residue while pp90rsk phosphorylates the Ser-32residue only Therefore these kinases are probably not involvedin the UV activated pathway Amino acids between IkBa po-sition 277 and 297 (the PEST region) are apparently requiredfor the UV induced degradation of IkB (Ito et al 1995 Ben-der et al 1998) but the signaling events that effect this degra-dation in keratinocytes are still unknown The data suggest thatTNFa and UVC use different pathways to activate NFkB inHeLa cells Specifically while both UVC and TNFa activateNFkB the TNFa signal goes via IKKs but the UVC signaldoes not From these data it appears that the convergence ofthe UVB and TNFa signals occurs downstream from IKKs thatis at the modification of IkB Overall these results imply thatthe epidermis a natural target of UV light has an exceptionalmolecular UV defense system distinct from UV responsive sys-tems in other tissues
The responses to the three different wavelengths of UV areespecially strictly controlled in keratinocytes Although allwavelengths cause DNA damage and a release of cytokines wefound that the inherent molecular responses to UVA are com-pletely different from the responses to UVBUVC For exam-ple UVBUVC but not UVA activate JNK and NFkB in ker-atinocytes Furthermore UVA activates the JNK pathway inHeLa cells but not in keratinocytes while UVBUVC activatethis pathway in both cells (Rosette and Karin 1996) Similarlyin both HeLa cells and cultured keratinocytes UVBUVC ac-tivate NFkB via a kinase different from IKKs while UVA doesnot activate NFkB at all We conclude that UVBUVC activatea completely different system of signal transduction from UVA
The results suggesting specific signal transduction pathwaysfor UVBUVC in keratinocytes presented here agree well withour studies of UVB-regulated genes using DNA micro arrays(Li et al 2001) We found that in response to UVB ker-atinocytes not only induce the immediate early genes and stressresponse genes but also elaborate responses designed to pro-tect the organism underneath For example chemokines of theIL-8 family are induced which alert the surrounding tissue tothe UV damage synthesis of the cornified envelope compo-nents is enhanced which thickens the stratum corneum etc Weconclude that human epidermal keratinocytes being naturallyexposed to UV light have elaborated highly specific molecu-lar responses to UVBUVC at the signal transduction level aswell as the gene expression level
ACKNOWLEDGMENTS
Our research was supported by the National Institutes ofHealth Grants AR30682 AR39176 AR40522 and AR41850
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 675
FIG 9 Specificity in stress response Epidermal ker-atinocytes exhibit specialized UV-responsive signal transduc-tion pathways which apparently do not network and intersectwith other pathways such as those responsive to EGF or TNFaIn contrast in other cell types such as HeLa multiple path-ways crossover interact and overlap
and by the Anaderm Research Corporation Special thinks goto Dr M Simon for the generous gift of keratinocytes We alsothank members of our laboratory for advice reagents and en-couragement
REFERENCES
ALKALAY I YARON A HATZUBAI A ORIAN ACIECHANOVER A and BEN-NERIAH Y (1995) Stimulation-dependent I kappa B alpha phosphorylation marks the NF-kappa Binhibitor for degradation via the ubiquitin-proteasome pathway ProcNatl Acad Sci USA 92 10599ndash10603
BAEUERLE PA and HENKEL T (1994) Function and activationof NF-kappa B in the immune system Annu Rev Immunol 12141ndash179
BARNES PJ and KARIN M (1997) Nuclear factor-kappaB A piv-otal transcription factor in chronic inflammatory diseases N EnglJ Med 336 1066ndash1071
BEG AA FINCO TS NANTERMET PV and BALDWIN AJR (1993) Tumor necrosis factor and interleukin-1 lead to phos-phorylation and loss of I kappa B alpha A mechanism for NF-kappaB activation Mol Cell Biol 13 3301ndash3310
BENDER K BLATTNER C KNEBEL A IORDANOV M HER-RLICH P and RAHMSDORF HJ (1997) UV-induced signaltransduction J Photochem Photobiol B 37 1ndash17
BENDER K GOTTLICHER M WHITESIDE S RAHMSDORFHJ and HERRLICH P (1998) Sequential DNA damage-indepen-dent and -dependent activation of NF-kappaB by UV EMBO J 175170ndash5181
BROWN K GERSTBERGER S CARLSON L FRANZOSO Gand SIEBENLIST U (1995) Control of I kappa B-alpha proteoly-sis by site-specific signal-induced phosphorylation Science 2671485ndash1488
DELHASE M HAYAKAWA M CHEN Y and KARIN M(1999) Positive and negative regulation of IkappaB kinase activitythrough IKKbeta subunit phosphorylation Science 284 309ndash313
DEacuteRIJARD B et al (1994) JNK1 A protein kinase stimulated byUV light and Ha-Ras that binds and phosphorylates the c-Jun acti-vation domain Cell 76 1025ndash1037
DEVARY Y ROSETTE C DIDONATO JA and KARIN M(1993) NF-kappa B activation by ultraviolet light not dependent ona nuclear signal Science 261 1442ndash1445
DIDONATO JA HAYAKAWA M ROTHWARF DM ZANDIE and KARIN M (1997) A cytokine-responsive IkappaB kinasethat activates the transcription factor NF-kappaB Nature 388548ndash554
FANGER GR GERWINS P WIDMANN C JARPE MB andJOHNSON GL (1997) MEKKs GCKs MLKs PAKs TAKs andtpls Upstream regulators of the c-Jun amino-terminal kinases CurrOpin Genet Dev 7 67ndash74
FEIGE U MORIMOTO RI YAHARA I and POLLA BS eds(1996) Stress-Inducible Cellular Responses (Birkhauser VerlagBasel Switzerland)
FISHER GJ WANG ZQ DATTA SC VARANI J KANG Sand VOORHEES JJ (1997) Pathophysiology of premature skin ag-ing induced by ultraviolet light N Engl J Med 337 1419ndash1428
GHODA L LIN X and GREENE WC (1997) The 90-kDa ribo-somal S6 kinase (pp90rsk) phosphorylates the N-terminal regulatorydomain of IkappaBalpha and stimulates its degradation in vitro JBiol Chem 272 21281ndash21288
GILCHREST B (1995) Photodamage (Blackwell Scientific NewYork)
GRETHER-BECK S BUETTNER R and KRUTMANN J (1997)Ultraviolet A radiation-induced expression of human genes Molec-ular and photobiological mechanisms Biol Chem 378 1231ndash1236
GRILLI M PIZZI M MEMO M and SPANO P (1996) Neuro-protection by aspirin and sodium salicylate through blockade of NF-kappaB activation Science 274 1383ndash1385
GUPTA S et al (1996) Selective interaction of JNK protein kinaseisoforms with transcription factors EMBO J 15 2760ndash2770
HEHNER SP et al (1998) Sesquiterpene lactones specifically in-hibit activation of NF-kappa B by preventing the degradation of Ikappa B-alpha and I kappa B-beta J Biol Chem 273 1288ndash1297
HERSKOWITZ I (1995) MAP kinase pathways in yeast For matingand more Cell 80 187ndash197
HIBI M LIN A SMEAL T MINDEN A and KARIN M (1993)Identification of an oncoprotein- and UV-responsive protein kinasethat binds and potentiates the c-Jun activation domain Genes Dev7 2135ndash2148
HIRANO M et al (1996) MEK kinase is involved in tumor necro-sis factor alpha-induced NF-kappaB activation and degradation ofIkappaB-alpha J Biol Chem 271 13234ndash13238
HU Y et al (1999) Abnormal morphogenesis but intact IKK activa-tion in mice lacking the IKKalpha subunit of IkappaB kinase Sci-ence 284 316ndash320
IORDANOV M et al (1997) CREB is activated by UVC through ap38HOG-1-dependent protein kinase EMBO J 16 1009ndash1022
ISREAL A (2000) The IKK complex An integrator of all signals thatactivate NF-kappaB Trends Cell Biol 10 129ndash133
ITO CY ADEY N BAUTCH VL and BALDWIN AS (1995)Structure and evolution of the human IKBA gene Genomics 29490ndash495
JIANG CK CONNOLLY D and BLUMENBERG M (1991)Comparison of methods for transfection of human epidermal ker-atinocytes J Invest Dermatol 97 969ndash973
JIANG CK FLANAGAN S OHTSUKI M SHUAI K FREED-BERG IM and BLUMENBERG M (1994) Disease-activatedtranscription factor Allergic reactions in human skin cause nucleartranscription of STAT-91 and induce synthesis of keratin K17 MolCell Biol 14 4759ndash4769
JIANG CK MAGNALDO T OHTSUKI M FREEDBERG IMBERNERD F and BLUMENBERG M (1993) Epidermal growthfactor and transforming growth factor alpha specifically induce theactivation- and hyperproliferation-associated keratins 6 and 16 ProcNatl Acad Sci USA 90 6786ndash6790
KALLUNKI T DENG T HIBI M and KARIN M (1996) c-Juncan recruit JNK to phosphorylate dimerization partners via specificdocking interactions Cell 87 929ndash939
KARIN M (1995) The regulation of AP-1 activity by mitogen-acti-vated protein kinases J Biol Chem 270 16483ndash16486
KARIN M and HUNTER T (1995) Transcriptional control by pro-tein phosphorylation Signal transmission from the cell surface to thenucleus Curr Biol 5 747ndash757
KLAMPFER L LEE TH HSU W VILCEK J and CHEN-KIANG S (1994) NF-IL6 and AP-1 cooperatively modulate the ac-tivation of the TSG-6 gene by tumor necrosis factor alpha and in-terleukin-1 Mol Cell Biol 14 6561ndash6569
KOMINE M et al (2000) Inflammatory versus proliferative pro-cesses in epidermis Tumor necrosis factor alpha induces K6b ker-atin synthesis through a transcriptional complex containing NFkappaB and CEBPbeta J Biol Chem 275 32077ndash32088
KOPP E and GHOSH S (1994) Inhibition of NF-kappa B by so-dium salicylate and aspirin Science 265 956ndash959
KYRIAKIS JM et al (1994) The stress-activated protein kinase sub-family of c-Jun kinases Nature 369 156ndash160
LEE FS HAGLER J CHEN ZJ and MANIATIS T (1997) Ac-tivation of the IkappaB alpha kinase complex by MEKK1 a kinaseof the JNK pathway Cell 88 213ndash222
LEPPA S and BOHMANN D (1999) Diverse functions of JNK sig-naling and c-Jun in stress response and apoptosis Oncogene 186158ndash6162
ADACHI ET AL676
LI D TURI TG SCHUCK A FREEDBERG IM KHITROVG and BLUMENBERG M (2001) Rays and arrays The tran-scriptional program in the response of human epidermal keratinocytesto UVB illumination FASEB J 15 2533ndash2535
LI N and KARIN M (1998) Ionizing radiation and short wavelengthUV activate NF-kappaB through two distinct mechanisms ProcNatl Acad Sci USA 95 13012ndash13017
LIU ZG HSU H GOEDDEL DV and KARIN M (1996) Dis-section of TNF receptor 1 effector functions JNK activation is notlinked to apoptosis while NF-kappaB activation prevents cell deathCell 87 565ndash576
MERCURIO F and MANNING AM (1999) NF-kappaB as a pri-mary regulator of the stress response Oncogene 18 6163ndash6171
MIYAMOTO S and VERMA IM (1995) RelNF-kappa BI kappaB story Adv Cancer Res 66 255ndash292
NESTLE FO MITRA RS BENNETT CF CHAN H andNICKOLOFF BJ (1994) Cationic lipid is not required for uptakeand selective inhibitory activity of ICAM-1 phosphorothioate anti-sense oligonucleotides in keratinocytes J Invest Dermatol 103569ndash575
PETERS RT LIAO SM and MANIATIS T (2000) IKKepsilonis part of a novel PMA-inducible IkappaB kinase complex Mol Cell5 513ndash522
PIERCE JW READ MA DING H LUSCINSKAS FW andCOLLINS T (1996) Salicylates inhibit I kappa B-alpha phos-phorylation endothelial-leukocyte adhesion molecule expressionand neutrophil transmigration J Immunol 156 3961ndash3969
PIERCE JW et al (1997) Novel inhibitors of cytokine-induced Ikap-paBalpha phosphorylation and endothelial cell adhesion molecule ex-pression show anti-inflammatory effects in vivo J Biol Chem 27221096ndash21103
REUNING U et al (1995) Inhibition of NF-kappa B-Rel A expres-sion by antisense oligodeoxynucleotides suppresses synthesis ofurokinase-type plasminogen activator (uPA) but not its inhibitor PAI-1 Nucleic Acids Res 23 3887ndash3893
ROSETTE C and KARIN M (1996) Ultraviolet light and osmoticstress activation of the JNK cascade through multiple growth factorand cytokine receptors Science 274 1194ndash1197
SCHOUTEN GJ et al (1997) IkappaB alpha is a target for the mitogen-activated 90 kDa ribosomal S6 kinase EMBO J 163133ndash3144
SHIMADA T et al (1999) IKK-i a novel lipopolysaccharide-inducible kinase that is related to IkappaB kinases Int Immunol 111357ndash1362
SIMON M and GREEN H (1984) Participation of membrane-asso-ciated proteins in the formation of the cross-linked envelope of thekeratinocyte Cell 36 827ndash834
SIMON MM ARAGANE Y SCHWARZ A LUGER TA andSCHWARZ T (1994) UVB light induces nuclear factor kappa B(NF kappa B) activity independently from chromosomal DNA dam-age in cell-free cytosolic extracts J Invest Dermatol 102 422ndash427
TOBIN D NILSSON M and TOFTGARD R (1996) Ras-inde-pendent activation of Rel-family transcription factors by UVB andTPA in cultured keratinocytes Oncogene 12 785ndash793
TOMIC-CANIC M KOMINE M FREEDBERG IM and BLU-MENBERG M (1998) Epidermal signal transduction and tran-scription factor activation in activated keratinocytes J Dermatol Sci17 167ndash181
WILLIAMS BR (1999) PKR A sentinel kinase for cellular stressOncogene 18 6112ndash6120
WU Z WU J JACINTO E and KARIN M (1997) Molecularcloning and characterization of human JNKK2 a novel Jun NH2-terminal kinase-specific kinase Mol Cell Biol 17 7407ndash7416
YANG DD et al (1997) Absence of excitotoxicity-induced apopto-sis in the hippocampus of mice lacking the Jnk3 gene Nature 389865ndash870
YANKULOV K BLAU J PURTON T ROBERTS S and BENT-LEY DL (1994) Transcriptional elongation by RNA polymeraseII is stimulated by transactivators Cell 77 749ndash759
YIN MJ YAMAMOTO Y and GAYNOR RB (1998) The anti-inflammatory agents aspirin and salicylate inhibit the activity ofI(kappa)B kinase-beta Nature 396 77ndash80
YOSHIDA S et al (2001) The c-Jun NH2-terminal kinase3 (JNK3)gene Genomic structure chromosomal assignment and loss of ex-pression in brain tumors J Hum Genet 46 182ndash187
Address reprint requests toM Blumenberg PhD
The NYU Cancer InstituteNew York University School of Medicine
550 First AveNew York NY 10016
E-mail blumem01mednyuedu
Received for publication June 5 2003 received in revised formJuly 15 2003 accepted August 14 2003
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 677

MATERIALS AND METHODS
Plasmids their growth and purification
The plasmid containing (NFkB)3-CAT was a gift from JVilcek NYU School of Medicine plasmid expressing Gal4Elkand GAL4CAT were a gift from J Blau Rockefeller Univer-sity and the control plasmids pRSVZ has been described pre-viously (Jiang et al 1993 1994 Klampfer et al 1994) Plas-mids were grown in JM101 Escherichia coli host to saturationdensity in LB medium DNAs were extracted and purified us-ing the Mega Prep kit from Promega (Madison WI)
Cell growth maintenance and transfection
Normal human foreskin epidermal keratinocytes were a gen-erous gift from Dr M Simon The cultures were initiated us-ing 3T3 feeder layers as described (Simon and Green 1984)and then frozen in liquid N2 until used Once thawed the ker-atinocytes were grown without feeder cells in defined serum-free keratinocyte medium supplemented with epidermal growthfactor and bovine pituitary extract (keratinocyte-SFM GIBCOCarlsbad CA) Cells were expanded through two 14 passagesbefore transfection and transfected at 90 confluency HeLacells were maintained in Dulbecco-modified Eaglersquos mediumsupplemented with 10 calf serum At the beginning of theseexperiments we optimized the doses of UV and concentrationsof EGF and TNFa to maximize the activation of the reportersin both HeLa cells and epidermal keratinocytes without killingthe cells (data not shown) All subsequent experiments wereperformed with optimal doses and concentrations
Transfections of keratinocytes with polybrene and DMSOshock were used to insert the DNA into the subconfluent ker-atinocytes as previously described (Jiang et al 1991) On theday of transfection cells were washed and incubated in the ba-sal medium without EGF and pituitary extract Each transfec-tion contained 10 mgdish of (NFkB)3-CAT and 3 mgdish ofpRSVZ The cells were incubated at 37degC in a 5 CO2 atmo-sphere and 24 h after transfection were illuminated with UVAUVB UVC or stimulated with TNFa EGF (100 ngml) in thepresence or absence of various inhibitors Treatment with as-pirin (0ndash100 mM from Sigma St Louis MO) BAY (0ndash5 mM)parthenolide (0ndash15 mM) and U0126 (0ndash1 mM all from BiomolPlymoth Meeting PA) lasted for another 24 h before harvest-ing We used two StratalinkerTM units (Stratagene La JollaCA) one equipped with FG15T8 bulbs as our UVB source andstandard GL-15 G15T8 bulbs as a source of UVC Before illu-mination the medium was removed from the cells and imme-diately after the illumination the same medium was restored tothe cells The whole illumination procedure takes less than aminute which we have shown does not cause drying or a heatshock The control cultures were treated identically except theactual illumination was omitted Cells were washed twice withphosphate-buffered saline 48 h after transfection and harvestedby scraping The cell disruption by repeated freezendashthaw cy-cles has also been described (Jiang et al 1991) All transfec-tions were performed in duplicate plates and each transfectionexperiment was repeated three to five times
HeLa cells were transfected using a modified calcium phos-phate precipitation procedure (Jiang et al 1991) Two days be-fore transfection cells were split by trypsinization and trans-
fected at 50 confluency Ten micrograms of the reporter plas-mid and 3 mg of the b-gal control plasmid were dissolved in atotal volume of 450 ml H2O To this mixture 50 ml of 25 MCaCl2 was added and mixed carefully Five hundred microlitersof BES-buffer pH 695 (50 mM N-Bis-2-hydroxyethyl-2-aminoethanesulfonic acid 250 mM NaCl 15 mM Na2HPO4)was added and mixed by inverting the tube The reaction mix-tures were incubated at room temperature for 10ndash15 min Onemilliliter of the precipitate suspension was added per platewhich was followed by incubation at 37degC in a 5 CO2 atmo-sphere Twenty-four hours after transfection cells were stimu-lated by TNFa or illuminated with UVA UVB or UVC withor without various inhibitors as indicated Cells were washedtwice with phosphate-buffered saline 36 to 48 h after transfec-tion and harvested by scraping
Enzyme assays
For b-galactosidase assays the substrate solution contained6 mg of O-nitrophenyl-D-galactoside (Sigma St Louis MO)freshly dissolved in PM buffer (66 mM Na2HPO4 33 mMNa2HPO4 40 mM mercaptoethanol 2 mM MgSO4 and 01 mMMnCl2) The reaction mixture contained 100 ml of substrate so-lution 300 ml of PM2 buffer and 50 ml of keratinocyte cell ex-tract or 20ndash30 ml of HeLa cell extract It was incubated at 37degCuntil development of a yellow color was obvious usually at 30min to 1 h The time of the reaction was recorded and the re-action stopped by addition of 04 ml of 1 M Na2CO3 OD420
was measured on a spectrophotometer (Labrepco HorshamPA)
The CAT reaction mixture contained 69 ml of 1 M Tris HClpH 78 1 ml of 14[C]-chloramphenicol (Sigma) 20 ml of 4 mMacetyl-CoA solution 30ndash60 ml of cell extract and enough wa-ter to bring the total reaction volume to 150 ml After incuba-tion at 37degC for 1 h the mixture was extracted into 1 ml ofethyl acetate phases were separated by brief centrifugation theorganic layer was transferred to a new tube and the solventevaporated The residue was dissolved in 25 ml of ethyl acetateand separated by thin layer chromatography on a silica gel platein chloroformmethanol 5 955 The plates were exposed to X-ray film for 12 to 24 h and the intensity of radioactive spotsdetermined using the Ambis Radioanalytic System (AmbisRochester NY) The conversion of chloramphenicol to itsmonoacetylated derivative was kept below 50 by varying theamount of extract or the duration of the reaction All CAT val-ues were normalized for transfection efficiency by calculatingthe ratio of CAT activity to b-galactosidase in each transfectedplate Each transfection experiment was separately performedthree or more times with each data point resulting from dupli-cate transfections
Use of antisense and sense oligonucleotides
Keratinocytes readily take up offered DNA Specificallyshort oligonucleotides can be introduced into these cells evenin the absence of cationic lipid (Nestle et al 1994 Reuning etal 1995) The oligonucleotides were synthesized in the phos-phorothiolate form to prolong their half-lives in the cells Wetargeted the antisense oligonucleotides to sequences includingand immediately upstream from the initiation codon for themost efficient antisense blocking At the same time we pre-
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 667
pared and used sense oligonucleotides as negative controls Ourapproach was to add the antisense oligonucleotides to the DNAtransfected into cells including the oligonucleotides into thetransfection solution which has the advantage of ensuring thatthe cells that received the transfected DNA also received theoligonucleotides The oligonucleotides were stored at 270degCin water until use and were added to the transfected DNA mix-ture In most cases we used 4 mg of the oligonucleotide pertransfection Their sequences are given in Table 1
Western blotting
The activated JNK antibody was purchased from Calbiochem(San Diego CA) and used at 1100 to 11000 dilution whilethe antirabbit immunoglobulin horseradish peroxidase-linkedantibodies and the ECL reagent were used as suggested by themanufacturer (Amersham Piscataway NJ) The anti-JNK3SAPK 1b antibody was purchased from Upstate Biotech-nology (Lake Placid NY) We used the antibody that recog-nizes JNK3 a1 (46KDa) and JNK3a2 (54KDa) at 11000dilution and the A431 cell lysate (Upstate Biotechnology) aspositive control according to manufacturerrsquos recommendationKeratinocytes were grown as described above At their thirdpassage at 80 confluence we incubated keratinocytes inKBM for 16 h to starve them of growth factors and then treatedas described above At various time intervals cells were har-vested by scraping washed twice with PBS and the cell pelletwas resuspended in 100 ml of the lysis buffer (2 SDS 10glycerol 10 mM Tris HCl pH 68) The cells were lysed by ul-trasonication for 30 sec using an ultrasonic homogenizer soni-cator (Cole-Parmer Vernon Hills IL) and boiling for 10 minAliquots of the supernatant obtained after centrifugation(2000 3 g 10 min) were stored at 270degC Protein content wasdetermined using the Bradford reagent (Perkin-Elmer BostonMA) Equal amounts of total protein were resolved by sodiumdodecyl sulphate-polyacrylamide gel electrophoresis (10 gel)
and electrophoretically transferred to nitrocellulose at 8 V for1 h After blocking the membranes with 5 nonfat dry milk inTris-buffered saline with 01 Tween (TBS-T) for 1 h at roomtemperature on an orbital shaker the blots were washed thrice(10 min each time) with TBS-T and incubated at 4degC with11000 dilution of the antibody Blots were washed 16 h laterwith TBS-T (3 3 10 min) and incubated at room temperaturewith 11000 dilution of antirabbit immunoglobin horseradishperoxidase-linked antibody for 2 h The chemiluminescenceECL reagent was used to develop the blots after washing themembranes three times 10 min with TBS-T and twice withTBS alone
JNK activity blots
Untreated control cells and cells treated with UV or EGFwere harvested 30 min after treatment The JNK activity wasmeasured by an in vitro kinase assay that detects phosphoryla-tion of a GST-Jun fusion protein The GST-Jun expressing con-struct was a generous gift from Dr Susan Logan New YorkUniversity School of Medicine This construct generates a fu-sion protein containing the 79 amino-terminal residues of c-junThe fusion protein GST-jun was used to purify JNK from thecell lysates and subsequently as a substrate for the same ki-nase The kinase assay was performed in kinase assay buffer(20 mM HEPES pH 75 20 mM beta-glycerol-phosphate 10mM MgCl2 10 mM DTT 50 mM sodium-orthovanadate 20mM ATP and 05 mlreaction of 32P-ATP (New England Nu-clear Amherst MA) The product of the kinase reaction wasthen separated by SDS-PAGE and the 32P-labeled GST-jun de-tected using autoradiography
RT-PCR
Cultured cells were harvested and total RNAs isolated uti-lizing RNeasy RNA extraction kit from Qiagen Between 1 and
ADACHI ET AL668
TABLE 1 OLIGONUCLEOTIDES USED
RT-PCR oligonucleotides used in detecting JNK ERK and p38 mRNAsJNK1 forward TCGCTACTACAGAGCA (570 bp expected product)JNK1 reverse GGAGAGGGCTGCCCCCJNK2 forward CTGGCCCGGACAGCGT (360 bp)JNK2 reverse GTTTTTATTTTGTCTCJNK3 forward TTATTACAGAGCCCCT (537 bp)JNK3 reverse TTTAGTCTTTTCTTCTGERK1 forward TTGCAGATCCAGACCA (295 bp)ERK1 reverse ATTTTTGTGTGGAAGAp38 forward GACGGATCGCAGAAGA (397 bp)p38 reverse TGCCAGGCACCCCGGT
Antisense oligonucleotides used in transfection of keratinocyte synthesized as phosphorotiolatesas-JNK1 CATGATGGCAAGCAATTAATGs-JNK1 GTACTACCGTTCGTTAATTACas-JNKK1 CATTGTTGGGAGTGAAGAGCCCAs-JNKK1 GTAACAACCCTCACTTCTCGGGTas-NFkB TGGATCATCTTCTGCCATTCTs-NKkB ACCTAGTAGAAGACGGTAAGAas-IkB CGCGTCGGCCGCCTGGAACATGCas-IKKa CATGGGGCGGGAGGGCAAGCGGCCas-IKKb ACTGATGTCGTGCTAACTCTGTCGs-IKKb TGACTACAGCACGATTGAGACAGC
15 mg of RNA were subjected to RT-PCR with Access RT-PCR kit system from Promega (Madison WI) for 32 cycleswith buffers and chemicals added as suggested by the manu-facturer In the supernumerary PCR experiments we used 42cycles The PCR products were subjected to either agarose orpolyacrylamide gel electrophoresis visualized with ethidiumbromide (Sigma) with a UV transilluminator from UVP andphotographed (Polaroid Waltham MA) The densities of bandswere quantified utilizing image scanner (GT-9000 Epson LongBeach CA)
Organ culture explants of normal human skin
Pieces of normal human skin were obtained immediately af-ter surgery They were cut into pieces approximately 3ndash5 mm3and incubated in keratinocyte basal medium KBM (Keratino-cyte-SFM Gibco-BRL Carlsbad CA) with or without TNFa
(50 ngml Intergen Norcross GA) in a humidified incubatorat 37degC for 24 h Generally we used 24-well culture disheswith up to five pieces in the same well and just enough mediumto cover the explants The antisense oligonucleotides wereadded at 300 ngml and when necessary replaced every 2 daysIllumination was performed epidermal side up with 100mJcm2 of UVB The explants were mounted in tissue Tec-OCTcompound (Sakura Finetek Japan) and frozen Sections 4 to 6mm thick were obtained with a cryostat (Miles LaboratoriesResearch Triangle Park NC) fixed with methanolacetone for10 min incubated with anti-JNK1 antibody (Calbiochem SanDiego CA) at 4degC overnight treated with peroxidase-conju-
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 669
FIG 1 UV activates JNK in keratinocytes The two left pan-els show the UV dose curve with each sample loaded in du-plicate The right panels show the time course of activation af-ter the illumination The top panels show the kinase activity ofJNK The activity was shown using GSTndashJun fusion protein asa substrate for immunoprecipitated JNK The bottom panelsshow the corresponding Western blots using antibody specificfor the activated phosphorylated form of JNKs The trianglepoints to the phosphorylated JNK substrate the doublet of openarrows to the JNK proteins The diffuse band in the top panelsis an uncharacterized artifact In the left panels we harvestedthe cells 10 min after irradiation and in the right panels weused 10 mJcm2 In each well we loaded 10 mg of protein
FIG 2 RT-PCR identification of JNK isotypes in keratinocytes HeLa cells and fibroblasts (a) Lanes K H and F representRNA from untreated keratinocytes HeLa cells and fibroblasts respectively Isotype-specific oligonucleotides were used in RT-PCR reactions The arrows point to the specific bands (b) Exaggerated PCR reactions for detecting low abundance mRNAs Du-plicate lanes show results of PCR reaction without (2) and with (1) input RNA preformed with supernumerary cycles (c) West-ern blots showing the presence of JNK3 lane 1 positive control (A431 extract) lane 2 HeLa cells and lane 3 keratinocytes
gated antimouse IgG secondary antibody (Vecstatin ABC-mouse IgG kit from Vector Laboratories Burlingame CA) atroom temperature for 1 h incubated with ABC complex (Vec-tor Laboratories) at room temperature for 1 h and treated with339-Diaminobenzidine-tetrahydrochloride (Dojindo Gaithers-burg MD) and 001 H2O2 in Tris pH 76 for 2 min The sam-ples were observed and photographed under the light micro-scope (Microphot-FXA Nikon Melville NY) Additionalantibodies used were from SantaCruz Biotechnology (SantaCruz CA)
RESULTS
UVB signal transduction pathway part I activation ofthe JNK pathway
To determine which JNK isoforms exist in keratinocytes andcan be activated by UVB we have grown primary cultures ofhuman epidermal keratinocytes and illuminated them with UVBat various doses As controls we used HeLa cells and normalhuman dermal fibroblasts The JNK activity was examined inthree ways First illuminated cells were stained with pan-JNKor active JNK-specific antibody to demonstrate the transloca-tion of JNK from cytoplasm to nucleus (not shown) Secondprotein extracts were prepared and analyzed using Western blotswith active JNK-specific antibody Third the kinase activitywas determined directly in affinity-precipitated JNK using aGST-tagged Jun protein fragment as the substrate The resultsof all three approaches were perfectly consistent UVB stronglyand specifically activates JNK in human epidermal ker-atinocytes (Fig 1)
Three isotypes of JNK have been described each with sev-eral splice variants It should be noted that there is confusionregarding which isotype is JNK2 and which is JNK3 in the lit-erature and even more egregiously in the blurbs of manufac-turers who provide antibodies We have adopted the unam-biguous designation of the NCBI databank To determine whichisotypes are expressed in keratinocytes we developed a set ofspecific RT-PCR oligonucleotides and used them to amplify theJNK mRNAs Surprisingly we found JNK1 and JNK3 in ker-atinocytes while JNK2 was absent (Fig 2a) This is an unex-pected finding because JNK1 and JNK2 are commonly almostubiquitously expressed while JNK3 was thought to be brain-specific (Yang et al 1997) Repeat of the PCR assay using alarge number of cycles confirmed the absence of JNK2 in ker-atinocytes (Fig 2b) Furthermore because of the importanceand unexpectedness of this result we decided to confirm theexpression of JNK3 protein in keratinocytes using Western blot-ting Indeed we find that JNK3 is expressed in keratinocytesbut not in HeLa cells (Fig 2c) Therefore keratinocytes expressboth the mRNA and the protein of JNK3 a kinase previouslythought to be brain-specific
To demonstrate that the activation of JNK is functional inthat it activates the Elk1 transcription factor in keratinocyteswe transfected into these cells a set of plasmids containing aGal4-CAT reporter and Gal4-DBDElk1-AF (Yankulov et al1994) The first plasmid contains a set of Gal4 binding sites up-stream from the CAT reporter gene the second expresses a fu-sion protein containing the Gal4 DNA binding domains fused
to the activating domain of transcription factor Elk1 When ac-tivated by UV via JNK the fusion protein activates transcrip-tion but if not activated it is silent Thus the system serves asan UV-responsive transcription reporter The transfected cellswere illuminated with UV and harvested 16 h later We foundthat both UVC and UVB activate the reporter but UVA doesnot (Fig 3) Note that at the 50 mJcm2 dose of UVC a signif-icant number of cells are killed These results mean that theJNK system in keratinocytes is present functional and specif-ically activated by short wavelength UV light UVBUVC butnot the long wavelength UVA We conclude that UVA doesnot activate the same system of signal transduction thatUVBUVC do
To identify the kinases responsible for UV signal we usedantisense approaches The antisense oligonucleotides were tar-geted against JNK1 and JNKK1 After 16 h to allow the anti-sense oligonucleotides to inhibit the synthesis of their targetsthe cells were irradiated with UV and after another 24 h har-vested for CAT determination We found that both antisensereagents attenuated the signals (Fig 4a and b) The antisenseeffects are specific because the sense oligonucleotides used ascontrols had no effect These results indicate that JNK1 andJNKK1 are essential for the transmission of the UVBUVC sig-
ADACHI ET AL670
FIG 3 Activation of transfected Elk-CAT reporter by UVBand UVC in keratinocytes GAL4-Elk1 fusion protein express-ing plasmid was cotransfected into keratinocytes with aGAL4CAT reporter The top panel shows the two cotransfectedplasmids and the effect of UV The transfected cells were illu-minated with indicated doses of UVA UVB or UVC harvested16 h later and the CAT levels determined Note that at the 50mJcm2 of UVC a significant number of cells is killed whichresults in significantly reduced levels of CAT activity (aster-isk) At the doses used UVA did not affect cell viability Thebars represent averages of duplicate transfections and the repli-cates differed by 25 or less
nal However they do not formally exclude the possibility thatJNK3 and JNKK2 also play a role
To determine whether the Elk1 activation is specific for theUV responsive signals in keratinocytes we tested the effects ofEGF In other cell types EGF can act through pathways thatactivate both JNK and Elk1 However when we attempted toactivate Elk1 using EGF in keratinocytes we were unsuccess-ful (Fig 4b) This means that in keratinocytes the EGF and UVresponsive pathways do not overlap on the contrary UV specif-ically activates the JNK pathway This suggests that the epi-dermal keratinocytes exhibit distinctive specialized UV-responsive signal transduction pathways
Indeed when we compared the responses to UV in severalcell types we found that the response in keratinocytes is sig-nificantly more stringently controlled than in other cell typesFor example both UV and EGF activated the JNK pathway inHeLa cells (Fig 4c and d) demonstrating a lack of specificityMoreover targeting JNK1 and JNKK1 with antisense oligonu-cleotides only partially inhibited the UV effects in HeLa cells
implying that in these cells parallel pathways can respond toUV to activate Elk1 (Fig 4c) Suspecting that EGF activatesElk1 through both the JNK and the ERKMAPK pathways inHeLa we inhibited the latter with U0126 a specific MEK-12inhibitor We found that U0126 significantly blocks the acti-vation by EGF but not by UV (Fig 4e) again demonstratingrelaxed stringency in HeLa cells
To approach these phenomena from another angle we usedkinase assays and Western blots Consistent with the above re-sults we found that while in keratinocytes only UVB (andUVC) could activate the JNK pathway in HeLa cells (and fi-broblasts not shown) heat shock UVA and osmotic shockcould activate the pathway as well (Fig 5a) In HeLa cells andfibroblasts UVB could activate the ERK pathway (not shown)but in keratinocytes it could not (Fig 5b) We conclude fromthese results that keratinocytes being often exposed to ambi-ent UV have developed more stringent and specific responsesto UV than other cell types which are not commonly exposedto UV These data are summarized in Figure 5c
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 671
FIG 4 UV specifically activates JNK in keratinocytes but not in HeLa cells GAL4-Elk1 and GAL4CAT plasmids were co-transfected into keratinocytes (a and b) or HeLa cells (c d and e) The transfected cells were treated with either UVC (5 mJcm2)or EGF (100 ngml) and harvested 24 h later for determination of CAT activity U0126a specific inhibitor of the MAPK path-way blocks the effects of EGF but not of UV Open bars represent the untreated control cells closed bars the UV-treated onesand the shaded bars the EGF treated ones
While there are only two JNKKs kinases that directly phos-phorylate JNK there is a large family of JNKKKs kinases thatphosphorylate JNKKs In a manuscript in preparation we de-scribe our analysis of the kinases expressed by keratinocytes
To demonstrate that the JNK system is responsive in vivowe used a novel ex vivo approach Biopsies of human skin weretreated with as-JNKK1 oligonucleotide or left untreated as con-trol Then both the treated and untreated biopsies were illumi-nated with UVC and shortly thereafter processed for immuno-histochemical detection of activated JNK (Fig 6) We found inthese biopsies that UV caused activation and nuclear translo-cation of JNK Furthermore this activation was abrogated bypretreatment with as-JNKK1 the antisense oligonucleotide tar-geting JNKK1 again demonstrating that JNKK1 is essential foractivation of JNK by UV in epidermis These results mean that
the specific signal transduction pathways we found operationalin cultured keratinocytes are also functional and working thesame way in human skin in vivo
UVB signal transduction pathway part II activationof the NFkB pathway
Having established that keratinocytes possess a specific UV-responding pathway to activate JNK we wanted to examine thepossibility that the UV-specific signal transduction pathwaysactivating NFkB are also specific in these cells The canonicalactivator of NFkB is TNFa however recent evidence suggeststhat UV and TNFa may activate NFkB through different mech-anisms (N Li and Karin 1998) Demonstrating a UV-specificpathway would strengthen even more our hypothesis that thereis specificity in UV responsive signal transduction in humanepidermal keratinocytes Therefore we transfected into ker-atinocytes and into HeLa cells a reporter plasmid containingthree tandem NFkB sites linked to the TK promoter-drivenCAT We found that the exposure of transfected cells to TNFaUVB or UVC resulted in the induction of the NFkB responseelement (Fig 7) Generally at the optimal conditions the in-duction of NFkB by UV was weaker than the induction byTNFa On the other hand NFkB was not activated by UVA atall even at high doses (Fig 7)
To compare the mechanisms of NFkB activation by UV andTNFa we have used a battery of specific inhibitors of the NFkBpathway which inhibit the pathway at different points (Fig 8a)Namely Bay11-7085 inhibits upstream from IKK (Pierce et al1997) aspirin has recently been found to be a competitive in-hibitor of ATP binding to IKKb (Yin et al 1998) whereasparthenolide directly inhibits the degradation of IkBa (Pierceet al 1997) When we treated transfected keratinocytes or HeLacells with these three inhibitors we found that Bay11-7085 in-hibited the induction of NFkB by TNFa in a dose-dependentmanner while it has almost no effect on the induction of NFkBby UVC (Fig 8b) The same results were obtained by using as-pirin (Fig 8c) In contrast parthenolide inhibited both the UVCand the TNFa effects in parallel (Fig 8d) These results con-firm that TNFa and UVC use different pathways to activateNFkB in HeLa cells Specifically while both UVC and TNFaactivate NFkB the TNFa signal goes via IKKs but the UVCsignal does not From these data we conclude that the conver-gence of the UVB and TNFa signals occurs downstream fromIKKs that is at the modification of IkB
One caveat in the use of inhibitors of specific kinases is thepossibility that such inhibitors may affect additional pathwaysnot currently recognized as affected by the given inhibitor Toaddress such specificity concerns and to examine the involve-ment of IKK in the induction of NFkB by TNFa UVC andUVB we used antisense oligonucleotides specifically targetingNFkB and IKKb as-NFkB as-IKKb (Table 1) (Reuning etal 1995 Komine et al 2000) In parallel we added the senseoligonucleotides s-NFkB s-IKKb as controls The antisenseexperiments confirmed the fact that UVBUVC use a clearlydifferent pathway to activate NFkB than TNFa does The sup-pression of NFkB synthesis in cultured keratinocytes by addingas-NFkB resulted in inhibition of activation of NFkB by allthree agents UVC UVB and TNFa as expected (Fig 8b) Onthe other hand the suppression of IKKb synthesis by adding
ADACHI ET AL672
FIG 5 Specific activation of JNK in keratinocytes andpromiscuous activation of JNK in HeLa cells (a) JNK activityin keratinocytes and HeLa cells treated with UVC UVA heatshock or EGF Duplicate experiments were performed with allagents except EGF Each lane contains 10 mg of protein InHeLa cells (bottom panel) UVC and EGF strongly while UVAand heat detectably activate JNK Cells were harvested 30 minafter treatment (b) Lack of activation of ERK by UVB in ker-atinocytes The top panel shows a Western blot with the anti-body specific for the activated ERKs the bottom for totalERK1 as a control for loading and transfer (c) Specificity insignal transduction In keratinocytes UVB specifically acti-vates JNK and EGF specifically activates ERKs whereas inHeLa cells all three extracellular signals UVB EGF and heatactivate both JNK and ERK Fibroblasts respond similar toHeLa cells (data not shown)
as-IKKb only inhibited the activation by TNFa but had no ef-fect on the activation by either UVC or UVB (Fig 8b) In theseexperiments we used s-NFkB and s-IKKa as controls and con-firmed that they had no effect in this pathway (Fig 8b) Fur-thermore as-NIK targeting a kinase functioning upstream fromIKK also partially inhibited the effects of TNFa Therefore
the antisense oligonucleotide experiments confirm our conclu-sions that the activation of NFkB by TNFa depends on the IKKpathway whereas the response to UVC or UVB occurs througha mechanism independent of IKKs
Furthermore as an additional control we used oligonu-cleotides targeting the JNK pathway as-JNKK1 and s-JNKK1Although they affected the JNK pathway (see Figs 4 and 6)they did not affect the NFkB pathway (Fig 8b) which indi-cates that the activation of the JNK pathway and the NFkBpathway are independent in keratinocytes This means that epi-dermal keratinocytes contain multiple specific UV responsivemechanisms
DISCUSSION
Our data suggest that UVBUVC illumination in epidermalkeratinocytes is not a generic stress response inducer but a spe-cific extracellular agent that activates specific signal transduc-tion pathways In particular we show that in keratinocytes UVexclusively activates the JNK pathway while in HeLa cellsUV also activates the parallel ERK and p38 pathways Con-versely in keratinocytes exclusively UV activates the JNKpathway while in HeLa cells growth factors can do so as wellIn parallel in keratinocytes UV uses a distinct pathway to ac-tivate the NFkB transcription factor a pathway separate fromthe more common TNFa activated one (Fig 9)
The specific activation of JNK by UVBUVC is particularlyrelevant because the JNK pathway is in different cell types atarget of multiple stress agents The balance between apoptosisand proliferation is at times regulated by JNK and the tran-scription factors activated by JNK regulate the expression ofmany genes (Karin and Hunter 1995) It is uniquely importantin epidermis that JNK exclusively responds to UVBUVC be-cause epidermis is the natural target of UVBUVC and the JNK-
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 673
FIG 6 Activation of JNK in vivo and the effect of as-JNKK1 In the control sample (left) JNK is not activated as shown us-ing an antibody specific for the activated phosphorylated form of JNK Treatment with 100 mJcm2 causes strong activation ofJNK (middle) Pretreatment for 3 h with the antisense oligonucleotide targeting JNKK1 abolishes this activation (right)
FIG 7 TNFa UVB and UVC activate NFkB while UVAdoes not Reporter plasmid NFkB3-CAT (top) which containsthree tandem NFkB sites upstream from the TK promoter wastransfected into keratinocytes The transfected cells were eithertreated with 50 ngml of TNFa or illuminated with indicateddoses of UV harvested 16 h later and the CAT levels deter-mined Untreated cells served as a control The replicates dif-fered by 25 or less
ADACHI ET AL674
FIG 8 Specific inhibitors can differentiate between the TNFa-and UVB-dependent signal transduction pathways (a) The effectsof specific inhibitors on NFkB activation Cultured keratinocyteswere treated with either UVB (8 mJcm2) or TNFa (50 ngml) af-ter transfection of the NFkB response element Inhibitor Bay11-7085 specifically inhibits upstream kinases of IKK aspirin inhibitsIKKb parthenolide blocks IkBa degradation (b) the effects of an-tisense oligonucleotides targeting signaling cascades on NFkB ac-tivation in keratinocytes Oligonucleotide as-NFkB targeting thetranscription factor blocks the activation by UVB and UVC whileoligonucleotides targeting IKKb JNK1 and JNKK1 are withouteffect (top two panels) In contrast oligonucleotides targetingIKKb and NIK partially or completely block the effects of TNFa(bottom panel) Note that the sense oligonucleotides (s-NFkB s-IKKb) used as controls had no effect on NFkB activation (c)Schematics of NFkB pathways activated by UV light and TNFaIn keratinocytes the two extracellular stimuli work through inde-pendent pathways that merge at the NFkBIkB complex
regulated transcription factors mediate both photodamage andits alleviation (Fisher et al 1997)
In keratinocytes the EGF and UV responsive pathways donot overlap but UV specifically activates the JNK pathwayThis suggests that the epidermal keratinocytes exhibit distinc-tive specialized UV-responsive signal transduction pathwaysIndeed when we compared the responses to UV in several celltypes we found that the response in keratinocytes is signifi-cantly more stringently controlled than in other cell types Forexample HeLa cells demonstrated a lack of specificity Usinga specific MEK-12 inhibitor we again demonstrated relaxedstringency in HeLa cells We conclude from these results thatkeratinocytes being often exposed to ambient UV have de-veloped more stringent and specific responses to UV than othercell types which are not commonly exposed to UV
At present we do not understand the relevance of JNK3 ex-pression in keratinocytes This isoform it was reported isspecifically expressed in the brain (Yoshida et al 2001) Wespeculate that perhaps the sequestration of JNK1 by the speci-ficity of the UV response creates a need for JNK3 as the moregeneral less restrictive stress signal transducer
TNFa one of proinflammatory cytokines important in cuta-neous inflammation activates NFkB via IKKs kinases that canbe activated by MEKK or NIK (Hirano et al 1996 Lee et al1997) In HeLa cells the phosphorylation of IkBa at Ser-32and Ser-36 residues is essential for the IKK mediated IkBa
degradation caused by TNFa IL-1 or ionizing radiation Us-ing specific inhibitors and antisense oligonucleotides we foundthat the NFkB activation by UVBUVC does not involve IKKs
The Ser-32 and Ser-36 residues are not involved in the UV-induced IkBa degradation (Bender et al 1997 N Li and Karin1998) Because IKKb specifically phosphorylates IkB at bothSer-32 and Ser-36 of IkBa these facts support our results Re-cently it was suggested that UV activation of NFkB in HeLacells may involve the RasndashRaf pathway although not in ker-atinocytes (Tobin et al 1996) This leads to the question howUV activates NFkB in keratinocytes Two new kinases IKKlaquo
and pp90rsk have been found to phosphorylate IkBa (Ghodaet al 1997 Schouten et al 1997 Shimada et al 1999 Israel2000 Peters et al 2000) However IKKlaquo phosphorylates theSer-36 residue while pp90rsk phosphorylates the Ser-32residue only Therefore these kinases are probably not involvedin the UV activated pathway Amino acids between IkBa po-sition 277 and 297 (the PEST region) are apparently requiredfor the UV induced degradation of IkB (Ito et al 1995 Ben-der et al 1998) but the signaling events that effect this degra-dation in keratinocytes are still unknown The data suggest thatTNFa and UVC use different pathways to activate NFkB inHeLa cells Specifically while both UVC and TNFa activateNFkB the TNFa signal goes via IKKs but the UVC signaldoes not From these data it appears that the convergence ofthe UVB and TNFa signals occurs downstream from IKKs thatis at the modification of IkB Overall these results imply thatthe epidermis a natural target of UV light has an exceptionalmolecular UV defense system distinct from UV responsive sys-tems in other tissues
The responses to the three different wavelengths of UV areespecially strictly controlled in keratinocytes Although allwavelengths cause DNA damage and a release of cytokines wefound that the inherent molecular responses to UVA are com-pletely different from the responses to UVBUVC For exam-ple UVBUVC but not UVA activate JNK and NFkB in ker-atinocytes Furthermore UVA activates the JNK pathway inHeLa cells but not in keratinocytes while UVBUVC activatethis pathway in both cells (Rosette and Karin 1996) Similarlyin both HeLa cells and cultured keratinocytes UVBUVC ac-tivate NFkB via a kinase different from IKKs while UVA doesnot activate NFkB at all We conclude that UVBUVC activatea completely different system of signal transduction from UVA
The results suggesting specific signal transduction pathwaysfor UVBUVC in keratinocytes presented here agree well withour studies of UVB-regulated genes using DNA micro arrays(Li et al 2001) We found that in response to UVB ker-atinocytes not only induce the immediate early genes and stressresponse genes but also elaborate responses designed to pro-tect the organism underneath For example chemokines of theIL-8 family are induced which alert the surrounding tissue tothe UV damage synthesis of the cornified envelope compo-nents is enhanced which thickens the stratum corneum etc Weconclude that human epidermal keratinocytes being naturallyexposed to UV light have elaborated highly specific molecu-lar responses to UVBUVC at the signal transduction level aswell as the gene expression level
ACKNOWLEDGMENTS
Our research was supported by the National Institutes ofHealth Grants AR30682 AR39176 AR40522 and AR41850
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 675
FIG 9 Specificity in stress response Epidermal ker-atinocytes exhibit specialized UV-responsive signal transduc-tion pathways which apparently do not network and intersectwith other pathways such as those responsive to EGF or TNFaIn contrast in other cell types such as HeLa multiple path-ways crossover interact and overlap
and by the Anaderm Research Corporation Special thinks goto Dr M Simon for the generous gift of keratinocytes We alsothank members of our laboratory for advice reagents and en-couragement
REFERENCES
ALKALAY I YARON A HATZUBAI A ORIAN ACIECHANOVER A and BEN-NERIAH Y (1995) Stimulation-dependent I kappa B alpha phosphorylation marks the NF-kappa Binhibitor for degradation via the ubiquitin-proteasome pathway ProcNatl Acad Sci USA 92 10599ndash10603
BAEUERLE PA and HENKEL T (1994) Function and activationof NF-kappa B in the immune system Annu Rev Immunol 12141ndash179
BARNES PJ and KARIN M (1997) Nuclear factor-kappaB A piv-otal transcription factor in chronic inflammatory diseases N EnglJ Med 336 1066ndash1071
BEG AA FINCO TS NANTERMET PV and BALDWIN AJR (1993) Tumor necrosis factor and interleukin-1 lead to phos-phorylation and loss of I kappa B alpha A mechanism for NF-kappaB activation Mol Cell Biol 13 3301ndash3310
BENDER K BLATTNER C KNEBEL A IORDANOV M HER-RLICH P and RAHMSDORF HJ (1997) UV-induced signaltransduction J Photochem Photobiol B 37 1ndash17
BENDER K GOTTLICHER M WHITESIDE S RAHMSDORFHJ and HERRLICH P (1998) Sequential DNA damage-indepen-dent and -dependent activation of NF-kappaB by UV EMBO J 175170ndash5181
BROWN K GERSTBERGER S CARLSON L FRANZOSO Gand SIEBENLIST U (1995) Control of I kappa B-alpha proteoly-sis by site-specific signal-induced phosphorylation Science 2671485ndash1488
DELHASE M HAYAKAWA M CHEN Y and KARIN M(1999) Positive and negative regulation of IkappaB kinase activitythrough IKKbeta subunit phosphorylation Science 284 309ndash313
DEacuteRIJARD B et al (1994) JNK1 A protein kinase stimulated byUV light and Ha-Ras that binds and phosphorylates the c-Jun acti-vation domain Cell 76 1025ndash1037
DEVARY Y ROSETTE C DIDONATO JA and KARIN M(1993) NF-kappa B activation by ultraviolet light not dependent ona nuclear signal Science 261 1442ndash1445
DIDONATO JA HAYAKAWA M ROTHWARF DM ZANDIE and KARIN M (1997) A cytokine-responsive IkappaB kinasethat activates the transcription factor NF-kappaB Nature 388548ndash554
FANGER GR GERWINS P WIDMANN C JARPE MB andJOHNSON GL (1997) MEKKs GCKs MLKs PAKs TAKs andtpls Upstream regulators of the c-Jun amino-terminal kinases CurrOpin Genet Dev 7 67ndash74
FEIGE U MORIMOTO RI YAHARA I and POLLA BS eds(1996) Stress-Inducible Cellular Responses (Birkhauser VerlagBasel Switzerland)
FISHER GJ WANG ZQ DATTA SC VARANI J KANG Sand VOORHEES JJ (1997) Pathophysiology of premature skin ag-ing induced by ultraviolet light N Engl J Med 337 1419ndash1428
GHODA L LIN X and GREENE WC (1997) The 90-kDa ribo-somal S6 kinase (pp90rsk) phosphorylates the N-terminal regulatorydomain of IkappaBalpha and stimulates its degradation in vitro JBiol Chem 272 21281ndash21288
GILCHREST B (1995) Photodamage (Blackwell Scientific NewYork)
GRETHER-BECK S BUETTNER R and KRUTMANN J (1997)Ultraviolet A radiation-induced expression of human genes Molec-ular and photobiological mechanisms Biol Chem 378 1231ndash1236
GRILLI M PIZZI M MEMO M and SPANO P (1996) Neuro-protection by aspirin and sodium salicylate through blockade of NF-kappaB activation Science 274 1383ndash1385
GUPTA S et al (1996) Selective interaction of JNK protein kinaseisoforms with transcription factors EMBO J 15 2760ndash2770
HEHNER SP et al (1998) Sesquiterpene lactones specifically in-hibit activation of NF-kappa B by preventing the degradation of Ikappa B-alpha and I kappa B-beta J Biol Chem 273 1288ndash1297
HERSKOWITZ I (1995) MAP kinase pathways in yeast For matingand more Cell 80 187ndash197
HIBI M LIN A SMEAL T MINDEN A and KARIN M (1993)Identification of an oncoprotein- and UV-responsive protein kinasethat binds and potentiates the c-Jun activation domain Genes Dev7 2135ndash2148
HIRANO M et al (1996) MEK kinase is involved in tumor necro-sis factor alpha-induced NF-kappaB activation and degradation ofIkappaB-alpha J Biol Chem 271 13234ndash13238
HU Y et al (1999) Abnormal morphogenesis but intact IKK activa-tion in mice lacking the IKKalpha subunit of IkappaB kinase Sci-ence 284 316ndash320
IORDANOV M et al (1997) CREB is activated by UVC through ap38HOG-1-dependent protein kinase EMBO J 16 1009ndash1022
ISREAL A (2000) The IKK complex An integrator of all signals thatactivate NF-kappaB Trends Cell Biol 10 129ndash133
ITO CY ADEY N BAUTCH VL and BALDWIN AS (1995)Structure and evolution of the human IKBA gene Genomics 29490ndash495
JIANG CK CONNOLLY D and BLUMENBERG M (1991)Comparison of methods for transfection of human epidermal ker-atinocytes J Invest Dermatol 97 969ndash973
JIANG CK FLANAGAN S OHTSUKI M SHUAI K FREED-BERG IM and BLUMENBERG M (1994) Disease-activatedtranscription factor Allergic reactions in human skin cause nucleartranscription of STAT-91 and induce synthesis of keratin K17 MolCell Biol 14 4759ndash4769
JIANG CK MAGNALDO T OHTSUKI M FREEDBERG IMBERNERD F and BLUMENBERG M (1993) Epidermal growthfactor and transforming growth factor alpha specifically induce theactivation- and hyperproliferation-associated keratins 6 and 16 ProcNatl Acad Sci USA 90 6786ndash6790
KALLUNKI T DENG T HIBI M and KARIN M (1996) c-Juncan recruit JNK to phosphorylate dimerization partners via specificdocking interactions Cell 87 929ndash939
KARIN M (1995) The regulation of AP-1 activity by mitogen-acti-vated protein kinases J Biol Chem 270 16483ndash16486
KARIN M and HUNTER T (1995) Transcriptional control by pro-tein phosphorylation Signal transmission from the cell surface to thenucleus Curr Biol 5 747ndash757
KLAMPFER L LEE TH HSU W VILCEK J and CHEN-KIANG S (1994) NF-IL6 and AP-1 cooperatively modulate the ac-tivation of the TSG-6 gene by tumor necrosis factor alpha and in-terleukin-1 Mol Cell Biol 14 6561ndash6569
KOMINE M et al (2000) Inflammatory versus proliferative pro-cesses in epidermis Tumor necrosis factor alpha induces K6b ker-atin synthesis through a transcriptional complex containing NFkappaB and CEBPbeta J Biol Chem 275 32077ndash32088
KOPP E and GHOSH S (1994) Inhibition of NF-kappa B by so-dium salicylate and aspirin Science 265 956ndash959
KYRIAKIS JM et al (1994) The stress-activated protein kinase sub-family of c-Jun kinases Nature 369 156ndash160
LEE FS HAGLER J CHEN ZJ and MANIATIS T (1997) Ac-tivation of the IkappaB alpha kinase complex by MEKK1 a kinaseof the JNK pathway Cell 88 213ndash222
LEPPA S and BOHMANN D (1999) Diverse functions of JNK sig-naling and c-Jun in stress response and apoptosis Oncogene 186158ndash6162
ADACHI ET AL676
LI D TURI TG SCHUCK A FREEDBERG IM KHITROVG and BLUMENBERG M (2001) Rays and arrays The tran-scriptional program in the response of human epidermal keratinocytesto UVB illumination FASEB J 15 2533ndash2535
LI N and KARIN M (1998) Ionizing radiation and short wavelengthUV activate NF-kappaB through two distinct mechanisms ProcNatl Acad Sci USA 95 13012ndash13017
LIU ZG HSU H GOEDDEL DV and KARIN M (1996) Dis-section of TNF receptor 1 effector functions JNK activation is notlinked to apoptosis while NF-kappaB activation prevents cell deathCell 87 565ndash576
MERCURIO F and MANNING AM (1999) NF-kappaB as a pri-mary regulator of the stress response Oncogene 18 6163ndash6171
MIYAMOTO S and VERMA IM (1995) RelNF-kappa BI kappaB story Adv Cancer Res 66 255ndash292
NESTLE FO MITRA RS BENNETT CF CHAN H andNICKOLOFF BJ (1994) Cationic lipid is not required for uptakeand selective inhibitory activity of ICAM-1 phosphorothioate anti-sense oligonucleotides in keratinocytes J Invest Dermatol 103569ndash575
PETERS RT LIAO SM and MANIATIS T (2000) IKKepsilonis part of a novel PMA-inducible IkappaB kinase complex Mol Cell5 513ndash522
PIERCE JW READ MA DING H LUSCINSKAS FW andCOLLINS T (1996) Salicylates inhibit I kappa B-alpha phos-phorylation endothelial-leukocyte adhesion molecule expressionand neutrophil transmigration J Immunol 156 3961ndash3969
PIERCE JW et al (1997) Novel inhibitors of cytokine-induced Ikap-paBalpha phosphorylation and endothelial cell adhesion molecule ex-pression show anti-inflammatory effects in vivo J Biol Chem 27221096ndash21103
REUNING U et al (1995) Inhibition of NF-kappa B-Rel A expres-sion by antisense oligodeoxynucleotides suppresses synthesis ofurokinase-type plasminogen activator (uPA) but not its inhibitor PAI-1 Nucleic Acids Res 23 3887ndash3893
ROSETTE C and KARIN M (1996) Ultraviolet light and osmoticstress activation of the JNK cascade through multiple growth factorand cytokine receptors Science 274 1194ndash1197
SCHOUTEN GJ et al (1997) IkappaB alpha is a target for the mitogen-activated 90 kDa ribosomal S6 kinase EMBO J 163133ndash3144
SHIMADA T et al (1999) IKK-i a novel lipopolysaccharide-inducible kinase that is related to IkappaB kinases Int Immunol 111357ndash1362
SIMON M and GREEN H (1984) Participation of membrane-asso-ciated proteins in the formation of the cross-linked envelope of thekeratinocyte Cell 36 827ndash834
SIMON MM ARAGANE Y SCHWARZ A LUGER TA andSCHWARZ T (1994) UVB light induces nuclear factor kappa B(NF kappa B) activity independently from chromosomal DNA dam-age in cell-free cytosolic extracts J Invest Dermatol 102 422ndash427
TOBIN D NILSSON M and TOFTGARD R (1996) Ras-inde-pendent activation of Rel-family transcription factors by UVB andTPA in cultured keratinocytes Oncogene 12 785ndash793
TOMIC-CANIC M KOMINE M FREEDBERG IM and BLU-MENBERG M (1998) Epidermal signal transduction and tran-scription factor activation in activated keratinocytes J Dermatol Sci17 167ndash181
WILLIAMS BR (1999) PKR A sentinel kinase for cellular stressOncogene 18 6112ndash6120
WU Z WU J JACINTO E and KARIN M (1997) Molecularcloning and characterization of human JNKK2 a novel Jun NH2-terminal kinase-specific kinase Mol Cell Biol 17 7407ndash7416
YANG DD et al (1997) Absence of excitotoxicity-induced apopto-sis in the hippocampus of mice lacking the Jnk3 gene Nature 389865ndash870
YANKULOV K BLAU J PURTON T ROBERTS S and BENT-LEY DL (1994) Transcriptional elongation by RNA polymeraseII is stimulated by transactivators Cell 77 749ndash759
YIN MJ YAMAMOTO Y and GAYNOR RB (1998) The anti-inflammatory agents aspirin and salicylate inhibit the activity ofI(kappa)B kinase-beta Nature 396 77ndash80
YOSHIDA S et al (2001) The c-Jun NH2-terminal kinase3 (JNK3)gene Genomic structure chromosomal assignment and loss of ex-pression in brain tumors J Hum Genet 46 182ndash187
Address reprint requests toM Blumenberg PhD
The NYU Cancer InstituteNew York University School of Medicine
550 First AveNew York NY 10016
E-mail blumem01mednyuedu
Received for publication June 5 2003 received in revised formJuly 15 2003 accepted August 14 2003
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 677

pared and used sense oligonucleotides as negative controls Ourapproach was to add the antisense oligonucleotides to the DNAtransfected into cells including the oligonucleotides into thetransfection solution which has the advantage of ensuring thatthe cells that received the transfected DNA also received theoligonucleotides The oligonucleotides were stored at 270degCin water until use and were added to the transfected DNA mix-ture In most cases we used 4 mg of the oligonucleotide pertransfection Their sequences are given in Table 1
Western blotting
The activated JNK antibody was purchased from Calbiochem(San Diego CA) and used at 1100 to 11000 dilution whilethe antirabbit immunoglobulin horseradish peroxidase-linkedantibodies and the ECL reagent were used as suggested by themanufacturer (Amersham Piscataway NJ) The anti-JNK3SAPK 1b antibody was purchased from Upstate Biotech-nology (Lake Placid NY) We used the antibody that recog-nizes JNK3 a1 (46KDa) and JNK3a2 (54KDa) at 11000dilution and the A431 cell lysate (Upstate Biotechnology) aspositive control according to manufacturerrsquos recommendationKeratinocytes were grown as described above At their thirdpassage at 80 confluence we incubated keratinocytes inKBM for 16 h to starve them of growth factors and then treatedas described above At various time intervals cells were har-vested by scraping washed twice with PBS and the cell pelletwas resuspended in 100 ml of the lysis buffer (2 SDS 10glycerol 10 mM Tris HCl pH 68) The cells were lysed by ul-trasonication for 30 sec using an ultrasonic homogenizer soni-cator (Cole-Parmer Vernon Hills IL) and boiling for 10 minAliquots of the supernatant obtained after centrifugation(2000 3 g 10 min) were stored at 270degC Protein content wasdetermined using the Bradford reagent (Perkin-Elmer BostonMA) Equal amounts of total protein were resolved by sodiumdodecyl sulphate-polyacrylamide gel electrophoresis (10 gel)
and electrophoretically transferred to nitrocellulose at 8 V for1 h After blocking the membranes with 5 nonfat dry milk inTris-buffered saline with 01 Tween (TBS-T) for 1 h at roomtemperature on an orbital shaker the blots were washed thrice(10 min each time) with TBS-T and incubated at 4degC with11000 dilution of the antibody Blots were washed 16 h laterwith TBS-T (3 3 10 min) and incubated at room temperaturewith 11000 dilution of antirabbit immunoglobin horseradishperoxidase-linked antibody for 2 h The chemiluminescenceECL reagent was used to develop the blots after washing themembranes three times 10 min with TBS-T and twice withTBS alone
JNK activity blots
Untreated control cells and cells treated with UV or EGFwere harvested 30 min after treatment The JNK activity wasmeasured by an in vitro kinase assay that detects phosphoryla-tion of a GST-Jun fusion protein The GST-Jun expressing con-struct was a generous gift from Dr Susan Logan New YorkUniversity School of Medicine This construct generates a fu-sion protein containing the 79 amino-terminal residues of c-junThe fusion protein GST-jun was used to purify JNK from thecell lysates and subsequently as a substrate for the same ki-nase The kinase assay was performed in kinase assay buffer(20 mM HEPES pH 75 20 mM beta-glycerol-phosphate 10mM MgCl2 10 mM DTT 50 mM sodium-orthovanadate 20mM ATP and 05 mlreaction of 32P-ATP (New England Nu-clear Amherst MA) The product of the kinase reaction wasthen separated by SDS-PAGE and the 32P-labeled GST-jun de-tected using autoradiography
RT-PCR
Cultured cells were harvested and total RNAs isolated uti-lizing RNeasy RNA extraction kit from Qiagen Between 1 and
ADACHI ET AL668
TABLE 1 OLIGONUCLEOTIDES USED
RT-PCR oligonucleotides used in detecting JNK ERK and p38 mRNAsJNK1 forward TCGCTACTACAGAGCA (570 bp expected product)JNK1 reverse GGAGAGGGCTGCCCCCJNK2 forward CTGGCCCGGACAGCGT (360 bp)JNK2 reverse GTTTTTATTTTGTCTCJNK3 forward TTATTACAGAGCCCCT (537 bp)JNK3 reverse TTTAGTCTTTTCTTCTGERK1 forward TTGCAGATCCAGACCA (295 bp)ERK1 reverse ATTTTTGTGTGGAAGAp38 forward GACGGATCGCAGAAGA (397 bp)p38 reverse TGCCAGGCACCCCGGT
Antisense oligonucleotides used in transfection of keratinocyte synthesized as phosphorotiolatesas-JNK1 CATGATGGCAAGCAATTAATGs-JNK1 GTACTACCGTTCGTTAATTACas-JNKK1 CATTGTTGGGAGTGAAGAGCCCAs-JNKK1 GTAACAACCCTCACTTCTCGGGTas-NFkB TGGATCATCTTCTGCCATTCTs-NKkB ACCTAGTAGAAGACGGTAAGAas-IkB CGCGTCGGCCGCCTGGAACATGCas-IKKa CATGGGGCGGGAGGGCAAGCGGCCas-IKKb ACTGATGTCGTGCTAACTCTGTCGs-IKKb TGACTACAGCACGATTGAGACAGC
15 mg of RNA were subjected to RT-PCR with Access RT-PCR kit system from Promega (Madison WI) for 32 cycleswith buffers and chemicals added as suggested by the manu-facturer In the supernumerary PCR experiments we used 42cycles The PCR products were subjected to either agarose orpolyacrylamide gel electrophoresis visualized with ethidiumbromide (Sigma) with a UV transilluminator from UVP andphotographed (Polaroid Waltham MA) The densities of bandswere quantified utilizing image scanner (GT-9000 Epson LongBeach CA)
Organ culture explants of normal human skin
Pieces of normal human skin were obtained immediately af-ter surgery They were cut into pieces approximately 3ndash5 mm3and incubated in keratinocyte basal medium KBM (Keratino-cyte-SFM Gibco-BRL Carlsbad CA) with or without TNFa
(50 ngml Intergen Norcross GA) in a humidified incubatorat 37degC for 24 h Generally we used 24-well culture disheswith up to five pieces in the same well and just enough mediumto cover the explants The antisense oligonucleotides wereadded at 300 ngml and when necessary replaced every 2 daysIllumination was performed epidermal side up with 100mJcm2 of UVB The explants were mounted in tissue Tec-OCTcompound (Sakura Finetek Japan) and frozen Sections 4 to 6mm thick were obtained with a cryostat (Miles LaboratoriesResearch Triangle Park NC) fixed with methanolacetone for10 min incubated with anti-JNK1 antibody (Calbiochem SanDiego CA) at 4degC overnight treated with peroxidase-conju-
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 669
FIG 1 UV activates JNK in keratinocytes The two left pan-els show the UV dose curve with each sample loaded in du-plicate The right panels show the time course of activation af-ter the illumination The top panels show the kinase activity ofJNK The activity was shown using GSTndashJun fusion protein asa substrate for immunoprecipitated JNK The bottom panelsshow the corresponding Western blots using antibody specificfor the activated phosphorylated form of JNKs The trianglepoints to the phosphorylated JNK substrate the doublet of openarrows to the JNK proteins The diffuse band in the top panelsis an uncharacterized artifact In the left panels we harvestedthe cells 10 min after irradiation and in the right panels weused 10 mJcm2 In each well we loaded 10 mg of protein
FIG 2 RT-PCR identification of JNK isotypes in keratinocytes HeLa cells and fibroblasts (a) Lanes K H and F representRNA from untreated keratinocytes HeLa cells and fibroblasts respectively Isotype-specific oligonucleotides were used in RT-PCR reactions The arrows point to the specific bands (b) Exaggerated PCR reactions for detecting low abundance mRNAs Du-plicate lanes show results of PCR reaction without (2) and with (1) input RNA preformed with supernumerary cycles (c) West-ern blots showing the presence of JNK3 lane 1 positive control (A431 extract) lane 2 HeLa cells and lane 3 keratinocytes
gated antimouse IgG secondary antibody (Vecstatin ABC-mouse IgG kit from Vector Laboratories Burlingame CA) atroom temperature for 1 h incubated with ABC complex (Vec-tor Laboratories) at room temperature for 1 h and treated with339-Diaminobenzidine-tetrahydrochloride (Dojindo Gaithers-burg MD) and 001 H2O2 in Tris pH 76 for 2 min The sam-ples were observed and photographed under the light micro-scope (Microphot-FXA Nikon Melville NY) Additionalantibodies used were from SantaCruz Biotechnology (SantaCruz CA)
RESULTS
UVB signal transduction pathway part I activation ofthe JNK pathway
To determine which JNK isoforms exist in keratinocytes andcan be activated by UVB we have grown primary cultures ofhuman epidermal keratinocytes and illuminated them with UVBat various doses As controls we used HeLa cells and normalhuman dermal fibroblasts The JNK activity was examined inthree ways First illuminated cells were stained with pan-JNKor active JNK-specific antibody to demonstrate the transloca-tion of JNK from cytoplasm to nucleus (not shown) Secondprotein extracts were prepared and analyzed using Western blotswith active JNK-specific antibody Third the kinase activitywas determined directly in affinity-precipitated JNK using aGST-tagged Jun protein fragment as the substrate The resultsof all three approaches were perfectly consistent UVB stronglyand specifically activates JNK in human epidermal ker-atinocytes (Fig 1)
Three isotypes of JNK have been described each with sev-eral splice variants It should be noted that there is confusionregarding which isotype is JNK2 and which is JNK3 in the lit-erature and even more egregiously in the blurbs of manufac-turers who provide antibodies We have adopted the unam-biguous designation of the NCBI databank To determine whichisotypes are expressed in keratinocytes we developed a set ofspecific RT-PCR oligonucleotides and used them to amplify theJNK mRNAs Surprisingly we found JNK1 and JNK3 in ker-atinocytes while JNK2 was absent (Fig 2a) This is an unex-pected finding because JNK1 and JNK2 are commonly almostubiquitously expressed while JNK3 was thought to be brain-specific (Yang et al 1997) Repeat of the PCR assay using alarge number of cycles confirmed the absence of JNK2 in ker-atinocytes (Fig 2b) Furthermore because of the importanceand unexpectedness of this result we decided to confirm theexpression of JNK3 protein in keratinocytes using Western blot-ting Indeed we find that JNK3 is expressed in keratinocytesbut not in HeLa cells (Fig 2c) Therefore keratinocytes expressboth the mRNA and the protein of JNK3 a kinase previouslythought to be brain-specific
To demonstrate that the activation of JNK is functional inthat it activates the Elk1 transcription factor in keratinocyteswe transfected into these cells a set of plasmids containing aGal4-CAT reporter and Gal4-DBDElk1-AF (Yankulov et al1994) The first plasmid contains a set of Gal4 binding sites up-stream from the CAT reporter gene the second expresses a fu-sion protein containing the Gal4 DNA binding domains fused
to the activating domain of transcription factor Elk1 When ac-tivated by UV via JNK the fusion protein activates transcrip-tion but if not activated it is silent Thus the system serves asan UV-responsive transcription reporter The transfected cellswere illuminated with UV and harvested 16 h later We foundthat both UVC and UVB activate the reporter but UVA doesnot (Fig 3) Note that at the 50 mJcm2 dose of UVC a signif-icant number of cells are killed These results mean that theJNK system in keratinocytes is present functional and specif-ically activated by short wavelength UV light UVBUVC butnot the long wavelength UVA We conclude that UVA doesnot activate the same system of signal transduction thatUVBUVC do
To identify the kinases responsible for UV signal we usedantisense approaches The antisense oligonucleotides were tar-geted against JNK1 and JNKK1 After 16 h to allow the anti-sense oligonucleotides to inhibit the synthesis of their targetsthe cells were irradiated with UV and after another 24 h har-vested for CAT determination We found that both antisensereagents attenuated the signals (Fig 4a and b) The antisenseeffects are specific because the sense oligonucleotides used ascontrols had no effect These results indicate that JNK1 andJNKK1 are essential for the transmission of the UVBUVC sig-
ADACHI ET AL670
FIG 3 Activation of transfected Elk-CAT reporter by UVBand UVC in keratinocytes GAL4-Elk1 fusion protein express-ing plasmid was cotransfected into keratinocytes with aGAL4CAT reporter The top panel shows the two cotransfectedplasmids and the effect of UV The transfected cells were illu-minated with indicated doses of UVA UVB or UVC harvested16 h later and the CAT levels determined Note that at the 50mJcm2 of UVC a significant number of cells is killed whichresults in significantly reduced levels of CAT activity (aster-isk) At the doses used UVA did not affect cell viability Thebars represent averages of duplicate transfections and the repli-cates differed by 25 or less
nal However they do not formally exclude the possibility thatJNK3 and JNKK2 also play a role
To determine whether the Elk1 activation is specific for theUV responsive signals in keratinocytes we tested the effects ofEGF In other cell types EGF can act through pathways thatactivate both JNK and Elk1 However when we attempted toactivate Elk1 using EGF in keratinocytes we were unsuccess-ful (Fig 4b) This means that in keratinocytes the EGF and UVresponsive pathways do not overlap on the contrary UV specif-ically activates the JNK pathway This suggests that the epi-dermal keratinocytes exhibit distinctive specialized UV-responsive signal transduction pathways
Indeed when we compared the responses to UV in severalcell types we found that the response in keratinocytes is sig-nificantly more stringently controlled than in other cell typesFor example both UV and EGF activated the JNK pathway inHeLa cells (Fig 4c and d) demonstrating a lack of specificityMoreover targeting JNK1 and JNKK1 with antisense oligonu-cleotides only partially inhibited the UV effects in HeLa cells
implying that in these cells parallel pathways can respond toUV to activate Elk1 (Fig 4c) Suspecting that EGF activatesElk1 through both the JNK and the ERKMAPK pathways inHeLa we inhibited the latter with U0126 a specific MEK-12inhibitor We found that U0126 significantly blocks the acti-vation by EGF but not by UV (Fig 4e) again demonstratingrelaxed stringency in HeLa cells
To approach these phenomena from another angle we usedkinase assays and Western blots Consistent with the above re-sults we found that while in keratinocytes only UVB (andUVC) could activate the JNK pathway in HeLa cells (and fi-broblasts not shown) heat shock UVA and osmotic shockcould activate the pathway as well (Fig 5a) In HeLa cells andfibroblasts UVB could activate the ERK pathway (not shown)but in keratinocytes it could not (Fig 5b) We conclude fromthese results that keratinocytes being often exposed to ambi-ent UV have developed more stringent and specific responsesto UV than other cell types which are not commonly exposedto UV These data are summarized in Figure 5c
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 671
FIG 4 UV specifically activates JNK in keratinocytes but not in HeLa cells GAL4-Elk1 and GAL4CAT plasmids were co-transfected into keratinocytes (a and b) or HeLa cells (c d and e) The transfected cells were treated with either UVC (5 mJcm2)or EGF (100 ngml) and harvested 24 h later for determination of CAT activity U0126a specific inhibitor of the MAPK path-way blocks the effects of EGF but not of UV Open bars represent the untreated control cells closed bars the UV-treated onesand the shaded bars the EGF treated ones
While there are only two JNKKs kinases that directly phos-phorylate JNK there is a large family of JNKKKs kinases thatphosphorylate JNKKs In a manuscript in preparation we de-scribe our analysis of the kinases expressed by keratinocytes
To demonstrate that the JNK system is responsive in vivowe used a novel ex vivo approach Biopsies of human skin weretreated with as-JNKK1 oligonucleotide or left untreated as con-trol Then both the treated and untreated biopsies were illumi-nated with UVC and shortly thereafter processed for immuno-histochemical detection of activated JNK (Fig 6) We found inthese biopsies that UV caused activation and nuclear translo-cation of JNK Furthermore this activation was abrogated bypretreatment with as-JNKK1 the antisense oligonucleotide tar-geting JNKK1 again demonstrating that JNKK1 is essential foractivation of JNK by UV in epidermis These results mean that
the specific signal transduction pathways we found operationalin cultured keratinocytes are also functional and working thesame way in human skin in vivo
UVB signal transduction pathway part II activationof the NFkB pathway
Having established that keratinocytes possess a specific UV-responding pathway to activate JNK we wanted to examine thepossibility that the UV-specific signal transduction pathwaysactivating NFkB are also specific in these cells The canonicalactivator of NFkB is TNFa however recent evidence suggeststhat UV and TNFa may activate NFkB through different mech-anisms (N Li and Karin 1998) Demonstrating a UV-specificpathway would strengthen even more our hypothesis that thereis specificity in UV responsive signal transduction in humanepidermal keratinocytes Therefore we transfected into ker-atinocytes and into HeLa cells a reporter plasmid containingthree tandem NFkB sites linked to the TK promoter-drivenCAT We found that the exposure of transfected cells to TNFaUVB or UVC resulted in the induction of the NFkB responseelement (Fig 7) Generally at the optimal conditions the in-duction of NFkB by UV was weaker than the induction byTNFa On the other hand NFkB was not activated by UVA atall even at high doses (Fig 7)
To compare the mechanisms of NFkB activation by UV andTNFa we have used a battery of specific inhibitors of the NFkBpathway which inhibit the pathway at different points (Fig 8a)Namely Bay11-7085 inhibits upstream from IKK (Pierce et al1997) aspirin has recently been found to be a competitive in-hibitor of ATP binding to IKKb (Yin et al 1998) whereasparthenolide directly inhibits the degradation of IkBa (Pierceet al 1997) When we treated transfected keratinocytes or HeLacells with these three inhibitors we found that Bay11-7085 in-hibited the induction of NFkB by TNFa in a dose-dependentmanner while it has almost no effect on the induction of NFkBby UVC (Fig 8b) The same results were obtained by using as-pirin (Fig 8c) In contrast parthenolide inhibited both the UVCand the TNFa effects in parallel (Fig 8d) These results con-firm that TNFa and UVC use different pathways to activateNFkB in HeLa cells Specifically while both UVC and TNFaactivate NFkB the TNFa signal goes via IKKs but the UVCsignal does not From these data we conclude that the conver-gence of the UVB and TNFa signals occurs downstream fromIKKs that is at the modification of IkB
One caveat in the use of inhibitors of specific kinases is thepossibility that such inhibitors may affect additional pathwaysnot currently recognized as affected by the given inhibitor Toaddress such specificity concerns and to examine the involve-ment of IKK in the induction of NFkB by TNFa UVC andUVB we used antisense oligonucleotides specifically targetingNFkB and IKKb as-NFkB as-IKKb (Table 1) (Reuning etal 1995 Komine et al 2000) In parallel we added the senseoligonucleotides s-NFkB s-IKKb as controls The antisenseexperiments confirmed the fact that UVBUVC use a clearlydifferent pathway to activate NFkB than TNFa does The sup-pression of NFkB synthesis in cultured keratinocytes by addingas-NFkB resulted in inhibition of activation of NFkB by allthree agents UVC UVB and TNFa as expected (Fig 8b) Onthe other hand the suppression of IKKb synthesis by adding
ADACHI ET AL672
FIG 5 Specific activation of JNK in keratinocytes andpromiscuous activation of JNK in HeLa cells (a) JNK activityin keratinocytes and HeLa cells treated with UVC UVA heatshock or EGF Duplicate experiments were performed with allagents except EGF Each lane contains 10 mg of protein InHeLa cells (bottom panel) UVC and EGF strongly while UVAand heat detectably activate JNK Cells were harvested 30 minafter treatment (b) Lack of activation of ERK by UVB in ker-atinocytes The top panel shows a Western blot with the anti-body specific for the activated ERKs the bottom for totalERK1 as a control for loading and transfer (c) Specificity insignal transduction In keratinocytes UVB specifically acti-vates JNK and EGF specifically activates ERKs whereas inHeLa cells all three extracellular signals UVB EGF and heatactivate both JNK and ERK Fibroblasts respond similar toHeLa cells (data not shown)
as-IKKb only inhibited the activation by TNFa but had no ef-fect on the activation by either UVC or UVB (Fig 8b) In theseexperiments we used s-NFkB and s-IKKa as controls and con-firmed that they had no effect in this pathway (Fig 8b) Fur-thermore as-NIK targeting a kinase functioning upstream fromIKK also partially inhibited the effects of TNFa Therefore
the antisense oligonucleotide experiments confirm our conclu-sions that the activation of NFkB by TNFa depends on the IKKpathway whereas the response to UVC or UVB occurs througha mechanism independent of IKKs
Furthermore as an additional control we used oligonu-cleotides targeting the JNK pathway as-JNKK1 and s-JNKK1Although they affected the JNK pathway (see Figs 4 and 6)they did not affect the NFkB pathway (Fig 8b) which indi-cates that the activation of the JNK pathway and the NFkBpathway are independent in keratinocytes This means that epi-dermal keratinocytes contain multiple specific UV responsivemechanisms
DISCUSSION
Our data suggest that UVBUVC illumination in epidermalkeratinocytes is not a generic stress response inducer but a spe-cific extracellular agent that activates specific signal transduc-tion pathways In particular we show that in keratinocytes UVexclusively activates the JNK pathway while in HeLa cellsUV also activates the parallel ERK and p38 pathways Con-versely in keratinocytes exclusively UV activates the JNKpathway while in HeLa cells growth factors can do so as wellIn parallel in keratinocytes UV uses a distinct pathway to ac-tivate the NFkB transcription factor a pathway separate fromthe more common TNFa activated one (Fig 9)
The specific activation of JNK by UVBUVC is particularlyrelevant because the JNK pathway is in different cell types atarget of multiple stress agents The balance between apoptosisand proliferation is at times regulated by JNK and the tran-scription factors activated by JNK regulate the expression ofmany genes (Karin and Hunter 1995) It is uniquely importantin epidermis that JNK exclusively responds to UVBUVC be-cause epidermis is the natural target of UVBUVC and the JNK-
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 673
FIG 6 Activation of JNK in vivo and the effect of as-JNKK1 In the control sample (left) JNK is not activated as shown us-ing an antibody specific for the activated phosphorylated form of JNK Treatment with 100 mJcm2 causes strong activation ofJNK (middle) Pretreatment for 3 h with the antisense oligonucleotide targeting JNKK1 abolishes this activation (right)
FIG 7 TNFa UVB and UVC activate NFkB while UVAdoes not Reporter plasmid NFkB3-CAT (top) which containsthree tandem NFkB sites upstream from the TK promoter wastransfected into keratinocytes The transfected cells were eithertreated with 50 ngml of TNFa or illuminated with indicateddoses of UV harvested 16 h later and the CAT levels deter-mined Untreated cells served as a control The replicates dif-fered by 25 or less
ADACHI ET AL674
FIG 8 Specific inhibitors can differentiate between the TNFa-and UVB-dependent signal transduction pathways (a) The effectsof specific inhibitors on NFkB activation Cultured keratinocyteswere treated with either UVB (8 mJcm2) or TNFa (50 ngml) af-ter transfection of the NFkB response element Inhibitor Bay11-7085 specifically inhibits upstream kinases of IKK aspirin inhibitsIKKb parthenolide blocks IkBa degradation (b) the effects of an-tisense oligonucleotides targeting signaling cascades on NFkB ac-tivation in keratinocytes Oligonucleotide as-NFkB targeting thetranscription factor blocks the activation by UVB and UVC whileoligonucleotides targeting IKKb JNK1 and JNKK1 are withouteffect (top two panels) In contrast oligonucleotides targetingIKKb and NIK partially or completely block the effects of TNFa(bottom panel) Note that the sense oligonucleotides (s-NFkB s-IKKb) used as controls had no effect on NFkB activation (c)Schematics of NFkB pathways activated by UV light and TNFaIn keratinocytes the two extracellular stimuli work through inde-pendent pathways that merge at the NFkBIkB complex
regulated transcription factors mediate both photodamage andits alleviation (Fisher et al 1997)
In keratinocytes the EGF and UV responsive pathways donot overlap but UV specifically activates the JNK pathwayThis suggests that the epidermal keratinocytes exhibit distinc-tive specialized UV-responsive signal transduction pathwaysIndeed when we compared the responses to UV in several celltypes we found that the response in keratinocytes is signifi-cantly more stringently controlled than in other cell types Forexample HeLa cells demonstrated a lack of specificity Usinga specific MEK-12 inhibitor we again demonstrated relaxedstringency in HeLa cells We conclude from these results thatkeratinocytes being often exposed to ambient UV have de-veloped more stringent and specific responses to UV than othercell types which are not commonly exposed to UV
At present we do not understand the relevance of JNK3 ex-pression in keratinocytes This isoform it was reported isspecifically expressed in the brain (Yoshida et al 2001) Wespeculate that perhaps the sequestration of JNK1 by the speci-ficity of the UV response creates a need for JNK3 as the moregeneral less restrictive stress signal transducer
TNFa one of proinflammatory cytokines important in cuta-neous inflammation activates NFkB via IKKs kinases that canbe activated by MEKK or NIK (Hirano et al 1996 Lee et al1997) In HeLa cells the phosphorylation of IkBa at Ser-32and Ser-36 residues is essential for the IKK mediated IkBa
degradation caused by TNFa IL-1 or ionizing radiation Us-ing specific inhibitors and antisense oligonucleotides we foundthat the NFkB activation by UVBUVC does not involve IKKs
The Ser-32 and Ser-36 residues are not involved in the UV-induced IkBa degradation (Bender et al 1997 N Li and Karin1998) Because IKKb specifically phosphorylates IkB at bothSer-32 and Ser-36 of IkBa these facts support our results Re-cently it was suggested that UV activation of NFkB in HeLacells may involve the RasndashRaf pathway although not in ker-atinocytes (Tobin et al 1996) This leads to the question howUV activates NFkB in keratinocytes Two new kinases IKKlaquo
and pp90rsk have been found to phosphorylate IkBa (Ghodaet al 1997 Schouten et al 1997 Shimada et al 1999 Israel2000 Peters et al 2000) However IKKlaquo phosphorylates theSer-36 residue while pp90rsk phosphorylates the Ser-32residue only Therefore these kinases are probably not involvedin the UV activated pathway Amino acids between IkBa po-sition 277 and 297 (the PEST region) are apparently requiredfor the UV induced degradation of IkB (Ito et al 1995 Ben-der et al 1998) but the signaling events that effect this degra-dation in keratinocytes are still unknown The data suggest thatTNFa and UVC use different pathways to activate NFkB inHeLa cells Specifically while both UVC and TNFa activateNFkB the TNFa signal goes via IKKs but the UVC signaldoes not From these data it appears that the convergence ofthe UVB and TNFa signals occurs downstream from IKKs thatis at the modification of IkB Overall these results imply thatthe epidermis a natural target of UV light has an exceptionalmolecular UV defense system distinct from UV responsive sys-tems in other tissues
The responses to the three different wavelengths of UV areespecially strictly controlled in keratinocytes Although allwavelengths cause DNA damage and a release of cytokines wefound that the inherent molecular responses to UVA are com-pletely different from the responses to UVBUVC For exam-ple UVBUVC but not UVA activate JNK and NFkB in ker-atinocytes Furthermore UVA activates the JNK pathway inHeLa cells but not in keratinocytes while UVBUVC activatethis pathway in both cells (Rosette and Karin 1996) Similarlyin both HeLa cells and cultured keratinocytes UVBUVC ac-tivate NFkB via a kinase different from IKKs while UVA doesnot activate NFkB at all We conclude that UVBUVC activatea completely different system of signal transduction from UVA
The results suggesting specific signal transduction pathwaysfor UVBUVC in keratinocytes presented here agree well withour studies of UVB-regulated genes using DNA micro arrays(Li et al 2001) We found that in response to UVB ker-atinocytes not only induce the immediate early genes and stressresponse genes but also elaborate responses designed to pro-tect the organism underneath For example chemokines of theIL-8 family are induced which alert the surrounding tissue tothe UV damage synthesis of the cornified envelope compo-nents is enhanced which thickens the stratum corneum etc Weconclude that human epidermal keratinocytes being naturallyexposed to UV light have elaborated highly specific molecu-lar responses to UVBUVC at the signal transduction level aswell as the gene expression level
ACKNOWLEDGMENTS
Our research was supported by the National Institutes ofHealth Grants AR30682 AR39176 AR40522 and AR41850
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 675
FIG 9 Specificity in stress response Epidermal ker-atinocytes exhibit specialized UV-responsive signal transduc-tion pathways which apparently do not network and intersectwith other pathways such as those responsive to EGF or TNFaIn contrast in other cell types such as HeLa multiple path-ways crossover interact and overlap
and by the Anaderm Research Corporation Special thinks goto Dr M Simon for the generous gift of keratinocytes We alsothank members of our laboratory for advice reagents and en-couragement
REFERENCES
ALKALAY I YARON A HATZUBAI A ORIAN ACIECHANOVER A and BEN-NERIAH Y (1995) Stimulation-dependent I kappa B alpha phosphorylation marks the NF-kappa Binhibitor for degradation via the ubiquitin-proteasome pathway ProcNatl Acad Sci USA 92 10599ndash10603
BAEUERLE PA and HENKEL T (1994) Function and activationof NF-kappa B in the immune system Annu Rev Immunol 12141ndash179
BARNES PJ and KARIN M (1997) Nuclear factor-kappaB A piv-otal transcription factor in chronic inflammatory diseases N EnglJ Med 336 1066ndash1071
BEG AA FINCO TS NANTERMET PV and BALDWIN AJR (1993) Tumor necrosis factor and interleukin-1 lead to phos-phorylation and loss of I kappa B alpha A mechanism for NF-kappaB activation Mol Cell Biol 13 3301ndash3310
BENDER K BLATTNER C KNEBEL A IORDANOV M HER-RLICH P and RAHMSDORF HJ (1997) UV-induced signaltransduction J Photochem Photobiol B 37 1ndash17
BENDER K GOTTLICHER M WHITESIDE S RAHMSDORFHJ and HERRLICH P (1998) Sequential DNA damage-indepen-dent and -dependent activation of NF-kappaB by UV EMBO J 175170ndash5181
BROWN K GERSTBERGER S CARLSON L FRANZOSO Gand SIEBENLIST U (1995) Control of I kappa B-alpha proteoly-sis by site-specific signal-induced phosphorylation Science 2671485ndash1488
DELHASE M HAYAKAWA M CHEN Y and KARIN M(1999) Positive and negative regulation of IkappaB kinase activitythrough IKKbeta subunit phosphorylation Science 284 309ndash313
DEacuteRIJARD B et al (1994) JNK1 A protein kinase stimulated byUV light and Ha-Ras that binds and phosphorylates the c-Jun acti-vation domain Cell 76 1025ndash1037
DEVARY Y ROSETTE C DIDONATO JA and KARIN M(1993) NF-kappa B activation by ultraviolet light not dependent ona nuclear signal Science 261 1442ndash1445
DIDONATO JA HAYAKAWA M ROTHWARF DM ZANDIE and KARIN M (1997) A cytokine-responsive IkappaB kinasethat activates the transcription factor NF-kappaB Nature 388548ndash554
FANGER GR GERWINS P WIDMANN C JARPE MB andJOHNSON GL (1997) MEKKs GCKs MLKs PAKs TAKs andtpls Upstream regulators of the c-Jun amino-terminal kinases CurrOpin Genet Dev 7 67ndash74
FEIGE U MORIMOTO RI YAHARA I and POLLA BS eds(1996) Stress-Inducible Cellular Responses (Birkhauser VerlagBasel Switzerland)
FISHER GJ WANG ZQ DATTA SC VARANI J KANG Sand VOORHEES JJ (1997) Pathophysiology of premature skin ag-ing induced by ultraviolet light N Engl J Med 337 1419ndash1428
GHODA L LIN X and GREENE WC (1997) The 90-kDa ribo-somal S6 kinase (pp90rsk) phosphorylates the N-terminal regulatorydomain of IkappaBalpha and stimulates its degradation in vitro JBiol Chem 272 21281ndash21288
GILCHREST B (1995) Photodamage (Blackwell Scientific NewYork)
GRETHER-BECK S BUETTNER R and KRUTMANN J (1997)Ultraviolet A radiation-induced expression of human genes Molec-ular and photobiological mechanisms Biol Chem 378 1231ndash1236
GRILLI M PIZZI M MEMO M and SPANO P (1996) Neuro-protection by aspirin and sodium salicylate through blockade of NF-kappaB activation Science 274 1383ndash1385
GUPTA S et al (1996) Selective interaction of JNK protein kinaseisoforms with transcription factors EMBO J 15 2760ndash2770
HEHNER SP et al (1998) Sesquiterpene lactones specifically in-hibit activation of NF-kappa B by preventing the degradation of Ikappa B-alpha and I kappa B-beta J Biol Chem 273 1288ndash1297
HERSKOWITZ I (1995) MAP kinase pathways in yeast For matingand more Cell 80 187ndash197
HIBI M LIN A SMEAL T MINDEN A and KARIN M (1993)Identification of an oncoprotein- and UV-responsive protein kinasethat binds and potentiates the c-Jun activation domain Genes Dev7 2135ndash2148
HIRANO M et al (1996) MEK kinase is involved in tumor necro-sis factor alpha-induced NF-kappaB activation and degradation ofIkappaB-alpha J Biol Chem 271 13234ndash13238
HU Y et al (1999) Abnormal morphogenesis but intact IKK activa-tion in mice lacking the IKKalpha subunit of IkappaB kinase Sci-ence 284 316ndash320
IORDANOV M et al (1997) CREB is activated by UVC through ap38HOG-1-dependent protein kinase EMBO J 16 1009ndash1022
ISREAL A (2000) The IKK complex An integrator of all signals thatactivate NF-kappaB Trends Cell Biol 10 129ndash133
ITO CY ADEY N BAUTCH VL and BALDWIN AS (1995)Structure and evolution of the human IKBA gene Genomics 29490ndash495
JIANG CK CONNOLLY D and BLUMENBERG M (1991)Comparison of methods for transfection of human epidermal ker-atinocytes J Invest Dermatol 97 969ndash973
JIANG CK FLANAGAN S OHTSUKI M SHUAI K FREED-BERG IM and BLUMENBERG M (1994) Disease-activatedtranscription factor Allergic reactions in human skin cause nucleartranscription of STAT-91 and induce synthesis of keratin K17 MolCell Biol 14 4759ndash4769
JIANG CK MAGNALDO T OHTSUKI M FREEDBERG IMBERNERD F and BLUMENBERG M (1993) Epidermal growthfactor and transforming growth factor alpha specifically induce theactivation- and hyperproliferation-associated keratins 6 and 16 ProcNatl Acad Sci USA 90 6786ndash6790
KALLUNKI T DENG T HIBI M and KARIN M (1996) c-Juncan recruit JNK to phosphorylate dimerization partners via specificdocking interactions Cell 87 929ndash939
KARIN M (1995) The regulation of AP-1 activity by mitogen-acti-vated protein kinases J Biol Chem 270 16483ndash16486
KARIN M and HUNTER T (1995) Transcriptional control by pro-tein phosphorylation Signal transmission from the cell surface to thenucleus Curr Biol 5 747ndash757
KLAMPFER L LEE TH HSU W VILCEK J and CHEN-KIANG S (1994) NF-IL6 and AP-1 cooperatively modulate the ac-tivation of the TSG-6 gene by tumor necrosis factor alpha and in-terleukin-1 Mol Cell Biol 14 6561ndash6569
KOMINE M et al (2000) Inflammatory versus proliferative pro-cesses in epidermis Tumor necrosis factor alpha induces K6b ker-atin synthesis through a transcriptional complex containing NFkappaB and CEBPbeta J Biol Chem 275 32077ndash32088
KOPP E and GHOSH S (1994) Inhibition of NF-kappa B by so-dium salicylate and aspirin Science 265 956ndash959
KYRIAKIS JM et al (1994) The stress-activated protein kinase sub-family of c-Jun kinases Nature 369 156ndash160
LEE FS HAGLER J CHEN ZJ and MANIATIS T (1997) Ac-tivation of the IkappaB alpha kinase complex by MEKK1 a kinaseof the JNK pathway Cell 88 213ndash222
LEPPA S and BOHMANN D (1999) Diverse functions of JNK sig-naling and c-Jun in stress response and apoptosis Oncogene 186158ndash6162
ADACHI ET AL676
LI D TURI TG SCHUCK A FREEDBERG IM KHITROVG and BLUMENBERG M (2001) Rays and arrays The tran-scriptional program in the response of human epidermal keratinocytesto UVB illumination FASEB J 15 2533ndash2535
LI N and KARIN M (1998) Ionizing radiation and short wavelengthUV activate NF-kappaB through two distinct mechanisms ProcNatl Acad Sci USA 95 13012ndash13017
LIU ZG HSU H GOEDDEL DV and KARIN M (1996) Dis-section of TNF receptor 1 effector functions JNK activation is notlinked to apoptosis while NF-kappaB activation prevents cell deathCell 87 565ndash576
MERCURIO F and MANNING AM (1999) NF-kappaB as a pri-mary regulator of the stress response Oncogene 18 6163ndash6171
MIYAMOTO S and VERMA IM (1995) RelNF-kappa BI kappaB story Adv Cancer Res 66 255ndash292
NESTLE FO MITRA RS BENNETT CF CHAN H andNICKOLOFF BJ (1994) Cationic lipid is not required for uptakeand selective inhibitory activity of ICAM-1 phosphorothioate anti-sense oligonucleotides in keratinocytes J Invest Dermatol 103569ndash575
PETERS RT LIAO SM and MANIATIS T (2000) IKKepsilonis part of a novel PMA-inducible IkappaB kinase complex Mol Cell5 513ndash522
PIERCE JW READ MA DING H LUSCINSKAS FW andCOLLINS T (1996) Salicylates inhibit I kappa B-alpha phos-phorylation endothelial-leukocyte adhesion molecule expressionand neutrophil transmigration J Immunol 156 3961ndash3969
PIERCE JW et al (1997) Novel inhibitors of cytokine-induced Ikap-paBalpha phosphorylation and endothelial cell adhesion molecule ex-pression show anti-inflammatory effects in vivo J Biol Chem 27221096ndash21103
REUNING U et al (1995) Inhibition of NF-kappa B-Rel A expres-sion by antisense oligodeoxynucleotides suppresses synthesis ofurokinase-type plasminogen activator (uPA) but not its inhibitor PAI-1 Nucleic Acids Res 23 3887ndash3893
ROSETTE C and KARIN M (1996) Ultraviolet light and osmoticstress activation of the JNK cascade through multiple growth factorand cytokine receptors Science 274 1194ndash1197
SCHOUTEN GJ et al (1997) IkappaB alpha is a target for the mitogen-activated 90 kDa ribosomal S6 kinase EMBO J 163133ndash3144
SHIMADA T et al (1999) IKK-i a novel lipopolysaccharide-inducible kinase that is related to IkappaB kinases Int Immunol 111357ndash1362
SIMON M and GREEN H (1984) Participation of membrane-asso-ciated proteins in the formation of the cross-linked envelope of thekeratinocyte Cell 36 827ndash834
SIMON MM ARAGANE Y SCHWARZ A LUGER TA andSCHWARZ T (1994) UVB light induces nuclear factor kappa B(NF kappa B) activity independently from chromosomal DNA dam-age in cell-free cytosolic extracts J Invest Dermatol 102 422ndash427
TOBIN D NILSSON M and TOFTGARD R (1996) Ras-inde-pendent activation of Rel-family transcription factors by UVB andTPA in cultured keratinocytes Oncogene 12 785ndash793
TOMIC-CANIC M KOMINE M FREEDBERG IM and BLU-MENBERG M (1998) Epidermal signal transduction and tran-scription factor activation in activated keratinocytes J Dermatol Sci17 167ndash181
WILLIAMS BR (1999) PKR A sentinel kinase for cellular stressOncogene 18 6112ndash6120
WU Z WU J JACINTO E and KARIN M (1997) Molecularcloning and characterization of human JNKK2 a novel Jun NH2-terminal kinase-specific kinase Mol Cell Biol 17 7407ndash7416
YANG DD et al (1997) Absence of excitotoxicity-induced apopto-sis in the hippocampus of mice lacking the Jnk3 gene Nature 389865ndash870
YANKULOV K BLAU J PURTON T ROBERTS S and BENT-LEY DL (1994) Transcriptional elongation by RNA polymeraseII is stimulated by transactivators Cell 77 749ndash759
YIN MJ YAMAMOTO Y and GAYNOR RB (1998) The anti-inflammatory agents aspirin and salicylate inhibit the activity ofI(kappa)B kinase-beta Nature 396 77ndash80
YOSHIDA S et al (2001) The c-Jun NH2-terminal kinase3 (JNK3)gene Genomic structure chromosomal assignment and loss of ex-pression in brain tumors J Hum Genet 46 182ndash187
Address reprint requests toM Blumenberg PhD
The NYU Cancer InstituteNew York University School of Medicine
550 First AveNew York NY 10016
E-mail blumem01mednyuedu
Received for publication June 5 2003 received in revised formJuly 15 2003 accepted August 14 2003
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 677
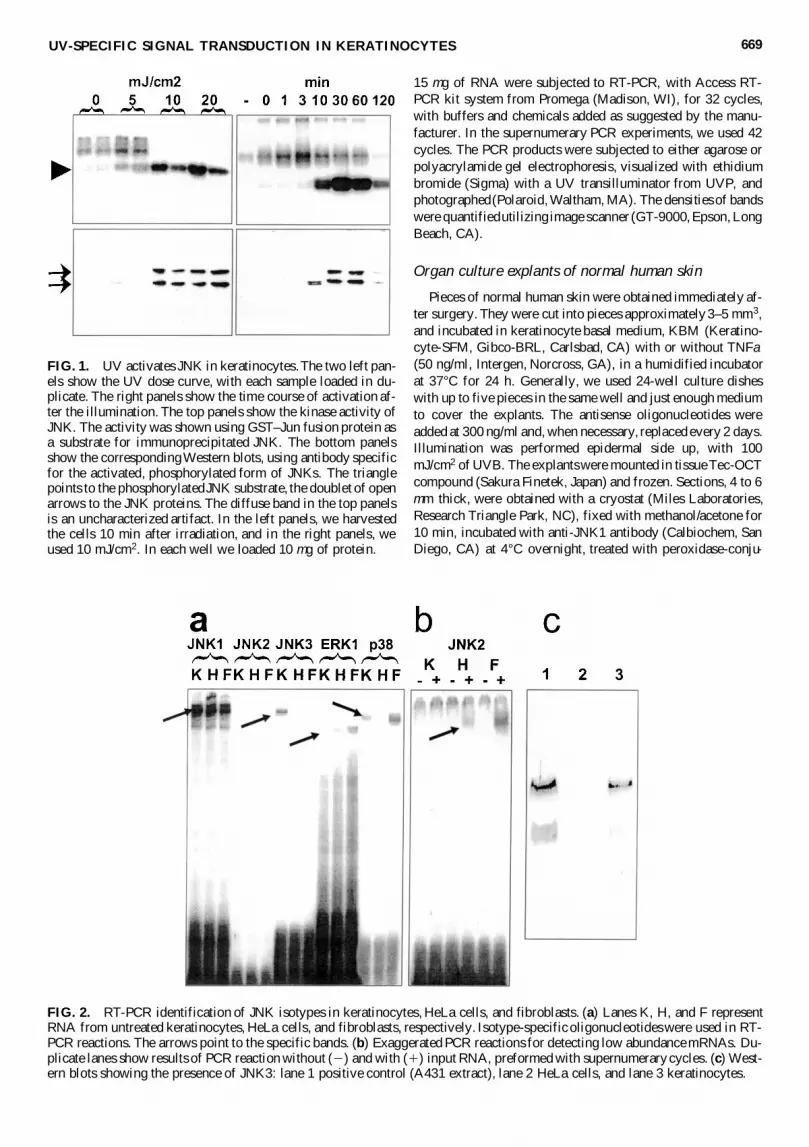
15 mg of RNA were subjected to RT-PCR with Access RT-PCR kit system from Promega (Madison WI) for 32 cycleswith buffers and chemicals added as suggested by the manu-facturer In the supernumerary PCR experiments we used 42cycles The PCR products were subjected to either agarose orpolyacrylamide gel electrophoresis visualized with ethidiumbromide (Sigma) with a UV transilluminator from UVP andphotographed (Polaroid Waltham MA) The densities of bandswere quantified utilizing image scanner (GT-9000 Epson LongBeach CA)
Organ culture explants of normal human skin
Pieces of normal human skin were obtained immediately af-ter surgery They were cut into pieces approximately 3ndash5 mm3and incubated in keratinocyte basal medium KBM (Keratino-cyte-SFM Gibco-BRL Carlsbad CA) with or without TNFa
(50 ngml Intergen Norcross GA) in a humidified incubatorat 37degC for 24 h Generally we used 24-well culture disheswith up to five pieces in the same well and just enough mediumto cover the explants The antisense oligonucleotides wereadded at 300 ngml and when necessary replaced every 2 daysIllumination was performed epidermal side up with 100mJcm2 of UVB The explants were mounted in tissue Tec-OCTcompound (Sakura Finetek Japan) and frozen Sections 4 to 6mm thick were obtained with a cryostat (Miles LaboratoriesResearch Triangle Park NC) fixed with methanolacetone for10 min incubated with anti-JNK1 antibody (Calbiochem SanDiego CA) at 4degC overnight treated with peroxidase-conju-
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 669
FIG 1 UV activates JNK in keratinocytes The two left pan-els show the UV dose curve with each sample loaded in du-plicate The right panels show the time course of activation af-ter the illumination The top panels show the kinase activity ofJNK The activity was shown using GSTndashJun fusion protein asa substrate for immunoprecipitated JNK The bottom panelsshow the corresponding Western blots using antibody specificfor the activated phosphorylated form of JNKs The trianglepoints to the phosphorylated JNK substrate the doublet of openarrows to the JNK proteins The diffuse band in the top panelsis an uncharacterized artifact In the left panels we harvestedthe cells 10 min after irradiation and in the right panels weused 10 mJcm2 In each well we loaded 10 mg of protein
FIG 2 RT-PCR identification of JNK isotypes in keratinocytes HeLa cells and fibroblasts (a) Lanes K H and F representRNA from untreated keratinocytes HeLa cells and fibroblasts respectively Isotype-specific oligonucleotides were used in RT-PCR reactions The arrows point to the specific bands (b) Exaggerated PCR reactions for detecting low abundance mRNAs Du-plicate lanes show results of PCR reaction without (2) and with (1) input RNA preformed with supernumerary cycles (c) West-ern blots showing the presence of JNK3 lane 1 positive control (A431 extract) lane 2 HeLa cells and lane 3 keratinocytes
gated antimouse IgG secondary antibody (Vecstatin ABC-mouse IgG kit from Vector Laboratories Burlingame CA) atroom temperature for 1 h incubated with ABC complex (Vec-tor Laboratories) at room temperature for 1 h and treated with339-Diaminobenzidine-tetrahydrochloride (Dojindo Gaithers-burg MD) and 001 H2O2 in Tris pH 76 for 2 min The sam-ples were observed and photographed under the light micro-scope (Microphot-FXA Nikon Melville NY) Additionalantibodies used were from SantaCruz Biotechnology (SantaCruz CA)
RESULTS
UVB signal transduction pathway part I activation ofthe JNK pathway
To determine which JNK isoforms exist in keratinocytes andcan be activated by UVB we have grown primary cultures ofhuman epidermal keratinocytes and illuminated them with UVBat various doses As controls we used HeLa cells and normalhuman dermal fibroblasts The JNK activity was examined inthree ways First illuminated cells were stained with pan-JNKor active JNK-specific antibody to demonstrate the transloca-tion of JNK from cytoplasm to nucleus (not shown) Secondprotein extracts were prepared and analyzed using Western blotswith active JNK-specific antibody Third the kinase activitywas determined directly in affinity-precipitated JNK using aGST-tagged Jun protein fragment as the substrate The resultsof all three approaches were perfectly consistent UVB stronglyand specifically activates JNK in human epidermal ker-atinocytes (Fig 1)
Three isotypes of JNK have been described each with sev-eral splice variants It should be noted that there is confusionregarding which isotype is JNK2 and which is JNK3 in the lit-erature and even more egregiously in the blurbs of manufac-turers who provide antibodies We have adopted the unam-biguous designation of the NCBI databank To determine whichisotypes are expressed in keratinocytes we developed a set ofspecific RT-PCR oligonucleotides and used them to amplify theJNK mRNAs Surprisingly we found JNK1 and JNK3 in ker-atinocytes while JNK2 was absent (Fig 2a) This is an unex-pected finding because JNK1 and JNK2 are commonly almostubiquitously expressed while JNK3 was thought to be brain-specific (Yang et al 1997) Repeat of the PCR assay using alarge number of cycles confirmed the absence of JNK2 in ker-atinocytes (Fig 2b) Furthermore because of the importanceand unexpectedness of this result we decided to confirm theexpression of JNK3 protein in keratinocytes using Western blot-ting Indeed we find that JNK3 is expressed in keratinocytesbut not in HeLa cells (Fig 2c) Therefore keratinocytes expressboth the mRNA and the protein of JNK3 a kinase previouslythought to be brain-specific
To demonstrate that the activation of JNK is functional inthat it activates the Elk1 transcription factor in keratinocyteswe transfected into these cells a set of plasmids containing aGal4-CAT reporter and Gal4-DBDElk1-AF (Yankulov et al1994) The first plasmid contains a set of Gal4 binding sites up-stream from the CAT reporter gene the second expresses a fu-sion protein containing the Gal4 DNA binding domains fused
to the activating domain of transcription factor Elk1 When ac-tivated by UV via JNK the fusion protein activates transcrip-tion but if not activated it is silent Thus the system serves asan UV-responsive transcription reporter The transfected cellswere illuminated with UV and harvested 16 h later We foundthat both UVC and UVB activate the reporter but UVA doesnot (Fig 3) Note that at the 50 mJcm2 dose of UVC a signif-icant number of cells are killed These results mean that theJNK system in keratinocytes is present functional and specif-ically activated by short wavelength UV light UVBUVC butnot the long wavelength UVA We conclude that UVA doesnot activate the same system of signal transduction thatUVBUVC do
To identify the kinases responsible for UV signal we usedantisense approaches The antisense oligonucleotides were tar-geted against JNK1 and JNKK1 After 16 h to allow the anti-sense oligonucleotides to inhibit the synthesis of their targetsthe cells were irradiated with UV and after another 24 h har-vested for CAT determination We found that both antisensereagents attenuated the signals (Fig 4a and b) The antisenseeffects are specific because the sense oligonucleotides used ascontrols had no effect These results indicate that JNK1 andJNKK1 are essential for the transmission of the UVBUVC sig-
ADACHI ET AL670
FIG 3 Activation of transfected Elk-CAT reporter by UVBand UVC in keratinocytes GAL4-Elk1 fusion protein express-ing plasmid was cotransfected into keratinocytes with aGAL4CAT reporter The top panel shows the two cotransfectedplasmids and the effect of UV The transfected cells were illu-minated with indicated doses of UVA UVB or UVC harvested16 h later and the CAT levels determined Note that at the 50mJcm2 of UVC a significant number of cells is killed whichresults in significantly reduced levels of CAT activity (aster-isk) At the doses used UVA did not affect cell viability Thebars represent averages of duplicate transfections and the repli-cates differed by 25 or less
nal However they do not formally exclude the possibility thatJNK3 and JNKK2 also play a role
To determine whether the Elk1 activation is specific for theUV responsive signals in keratinocytes we tested the effects ofEGF In other cell types EGF can act through pathways thatactivate both JNK and Elk1 However when we attempted toactivate Elk1 using EGF in keratinocytes we were unsuccess-ful (Fig 4b) This means that in keratinocytes the EGF and UVresponsive pathways do not overlap on the contrary UV specif-ically activates the JNK pathway This suggests that the epi-dermal keratinocytes exhibit distinctive specialized UV-responsive signal transduction pathways
Indeed when we compared the responses to UV in severalcell types we found that the response in keratinocytes is sig-nificantly more stringently controlled than in other cell typesFor example both UV and EGF activated the JNK pathway inHeLa cells (Fig 4c and d) demonstrating a lack of specificityMoreover targeting JNK1 and JNKK1 with antisense oligonu-cleotides only partially inhibited the UV effects in HeLa cells
implying that in these cells parallel pathways can respond toUV to activate Elk1 (Fig 4c) Suspecting that EGF activatesElk1 through both the JNK and the ERKMAPK pathways inHeLa we inhibited the latter with U0126 a specific MEK-12inhibitor We found that U0126 significantly blocks the acti-vation by EGF but not by UV (Fig 4e) again demonstratingrelaxed stringency in HeLa cells
To approach these phenomena from another angle we usedkinase assays and Western blots Consistent with the above re-sults we found that while in keratinocytes only UVB (andUVC) could activate the JNK pathway in HeLa cells (and fi-broblasts not shown) heat shock UVA and osmotic shockcould activate the pathway as well (Fig 5a) In HeLa cells andfibroblasts UVB could activate the ERK pathway (not shown)but in keratinocytes it could not (Fig 5b) We conclude fromthese results that keratinocytes being often exposed to ambi-ent UV have developed more stringent and specific responsesto UV than other cell types which are not commonly exposedto UV These data are summarized in Figure 5c
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 671
FIG 4 UV specifically activates JNK in keratinocytes but not in HeLa cells GAL4-Elk1 and GAL4CAT plasmids were co-transfected into keratinocytes (a and b) or HeLa cells (c d and e) The transfected cells were treated with either UVC (5 mJcm2)or EGF (100 ngml) and harvested 24 h later for determination of CAT activity U0126a specific inhibitor of the MAPK path-way blocks the effects of EGF but not of UV Open bars represent the untreated control cells closed bars the UV-treated onesand the shaded bars the EGF treated ones
While there are only two JNKKs kinases that directly phos-phorylate JNK there is a large family of JNKKKs kinases thatphosphorylate JNKKs In a manuscript in preparation we de-scribe our analysis of the kinases expressed by keratinocytes
To demonstrate that the JNK system is responsive in vivowe used a novel ex vivo approach Biopsies of human skin weretreated with as-JNKK1 oligonucleotide or left untreated as con-trol Then both the treated and untreated biopsies were illumi-nated with UVC and shortly thereafter processed for immuno-histochemical detection of activated JNK (Fig 6) We found inthese biopsies that UV caused activation and nuclear translo-cation of JNK Furthermore this activation was abrogated bypretreatment with as-JNKK1 the antisense oligonucleotide tar-geting JNKK1 again demonstrating that JNKK1 is essential foractivation of JNK by UV in epidermis These results mean that
the specific signal transduction pathways we found operationalin cultured keratinocytes are also functional and working thesame way in human skin in vivo
UVB signal transduction pathway part II activationof the NFkB pathway
Having established that keratinocytes possess a specific UV-responding pathway to activate JNK we wanted to examine thepossibility that the UV-specific signal transduction pathwaysactivating NFkB are also specific in these cells The canonicalactivator of NFkB is TNFa however recent evidence suggeststhat UV and TNFa may activate NFkB through different mech-anisms (N Li and Karin 1998) Demonstrating a UV-specificpathway would strengthen even more our hypothesis that thereis specificity in UV responsive signal transduction in humanepidermal keratinocytes Therefore we transfected into ker-atinocytes and into HeLa cells a reporter plasmid containingthree tandem NFkB sites linked to the TK promoter-drivenCAT We found that the exposure of transfected cells to TNFaUVB or UVC resulted in the induction of the NFkB responseelement (Fig 7) Generally at the optimal conditions the in-duction of NFkB by UV was weaker than the induction byTNFa On the other hand NFkB was not activated by UVA atall even at high doses (Fig 7)
To compare the mechanisms of NFkB activation by UV andTNFa we have used a battery of specific inhibitors of the NFkBpathway which inhibit the pathway at different points (Fig 8a)Namely Bay11-7085 inhibits upstream from IKK (Pierce et al1997) aspirin has recently been found to be a competitive in-hibitor of ATP binding to IKKb (Yin et al 1998) whereasparthenolide directly inhibits the degradation of IkBa (Pierceet al 1997) When we treated transfected keratinocytes or HeLacells with these three inhibitors we found that Bay11-7085 in-hibited the induction of NFkB by TNFa in a dose-dependentmanner while it has almost no effect on the induction of NFkBby UVC (Fig 8b) The same results were obtained by using as-pirin (Fig 8c) In contrast parthenolide inhibited both the UVCand the TNFa effects in parallel (Fig 8d) These results con-firm that TNFa and UVC use different pathways to activateNFkB in HeLa cells Specifically while both UVC and TNFaactivate NFkB the TNFa signal goes via IKKs but the UVCsignal does not From these data we conclude that the conver-gence of the UVB and TNFa signals occurs downstream fromIKKs that is at the modification of IkB
One caveat in the use of inhibitors of specific kinases is thepossibility that such inhibitors may affect additional pathwaysnot currently recognized as affected by the given inhibitor Toaddress such specificity concerns and to examine the involve-ment of IKK in the induction of NFkB by TNFa UVC andUVB we used antisense oligonucleotides specifically targetingNFkB and IKKb as-NFkB as-IKKb (Table 1) (Reuning etal 1995 Komine et al 2000) In parallel we added the senseoligonucleotides s-NFkB s-IKKb as controls The antisenseexperiments confirmed the fact that UVBUVC use a clearlydifferent pathway to activate NFkB than TNFa does The sup-pression of NFkB synthesis in cultured keratinocytes by addingas-NFkB resulted in inhibition of activation of NFkB by allthree agents UVC UVB and TNFa as expected (Fig 8b) Onthe other hand the suppression of IKKb synthesis by adding
ADACHI ET AL672
FIG 5 Specific activation of JNK in keratinocytes andpromiscuous activation of JNK in HeLa cells (a) JNK activityin keratinocytes and HeLa cells treated with UVC UVA heatshock or EGF Duplicate experiments were performed with allagents except EGF Each lane contains 10 mg of protein InHeLa cells (bottom panel) UVC and EGF strongly while UVAand heat detectably activate JNK Cells were harvested 30 minafter treatment (b) Lack of activation of ERK by UVB in ker-atinocytes The top panel shows a Western blot with the anti-body specific for the activated ERKs the bottom for totalERK1 as a control for loading and transfer (c) Specificity insignal transduction In keratinocytes UVB specifically acti-vates JNK and EGF specifically activates ERKs whereas inHeLa cells all three extracellular signals UVB EGF and heatactivate both JNK and ERK Fibroblasts respond similar toHeLa cells (data not shown)
as-IKKb only inhibited the activation by TNFa but had no ef-fect on the activation by either UVC or UVB (Fig 8b) In theseexperiments we used s-NFkB and s-IKKa as controls and con-firmed that they had no effect in this pathway (Fig 8b) Fur-thermore as-NIK targeting a kinase functioning upstream fromIKK also partially inhibited the effects of TNFa Therefore
the antisense oligonucleotide experiments confirm our conclu-sions that the activation of NFkB by TNFa depends on the IKKpathway whereas the response to UVC or UVB occurs througha mechanism independent of IKKs
Furthermore as an additional control we used oligonu-cleotides targeting the JNK pathway as-JNKK1 and s-JNKK1Although they affected the JNK pathway (see Figs 4 and 6)they did not affect the NFkB pathway (Fig 8b) which indi-cates that the activation of the JNK pathway and the NFkBpathway are independent in keratinocytes This means that epi-dermal keratinocytes contain multiple specific UV responsivemechanisms
DISCUSSION
Our data suggest that UVBUVC illumination in epidermalkeratinocytes is not a generic stress response inducer but a spe-cific extracellular agent that activates specific signal transduc-tion pathways In particular we show that in keratinocytes UVexclusively activates the JNK pathway while in HeLa cellsUV also activates the parallel ERK and p38 pathways Con-versely in keratinocytes exclusively UV activates the JNKpathway while in HeLa cells growth factors can do so as wellIn parallel in keratinocytes UV uses a distinct pathway to ac-tivate the NFkB transcription factor a pathway separate fromthe more common TNFa activated one (Fig 9)
The specific activation of JNK by UVBUVC is particularlyrelevant because the JNK pathway is in different cell types atarget of multiple stress agents The balance between apoptosisand proliferation is at times regulated by JNK and the tran-scription factors activated by JNK regulate the expression ofmany genes (Karin and Hunter 1995) It is uniquely importantin epidermis that JNK exclusively responds to UVBUVC be-cause epidermis is the natural target of UVBUVC and the JNK-
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 673
FIG 6 Activation of JNK in vivo and the effect of as-JNKK1 In the control sample (left) JNK is not activated as shown us-ing an antibody specific for the activated phosphorylated form of JNK Treatment with 100 mJcm2 causes strong activation ofJNK (middle) Pretreatment for 3 h with the antisense oligonucleotide targeting JNKK1 abolishes this activation (right)
FIG 7 TNFa UVB and UVC activate NFkB while UVAdoes not Reporter plasmid NFkB3-CAT (top) which containsthree tandem NFkB sites upstream from the TK promoter wastransfected into keratinocytes The transfected cells were eithertreated with 50 ngml of TNFa or illuminated with indicateddoses of UV harvested 16 h later and the CAT levels deter-mined Untreated cells served as a control The replicates dif-fered by 25 or less
ADACHI ET AL674
FIG 8 Specific inhibitors can differentiate between the TNFa-and UVB-dependent signal transduction pathways (a) The effectsof specific inhibitors on NFkB activation Cultured keratinocyteswere treated with either UVB (8 mJcm2) or TNFa (50 ngml) af-ter transfection of the NFkB response element Inhibitor Bay11-7085 specifically inhibits upstream kinases of IKK aspirin inhibitsIKKb parthenolide blocks IkBa degradation (b) the effects of an-tisense oligonucleotides targeting signaling cascades on NFkB ac-tivation in keratinocytes Oligonucleotide as-NFkB targeting thetranscription factor blocks the activation by UVB and UVC whileoligonucleotides targeting IKKb JNK1 and JNKK1 are withouteffect (top two panels) In contrast oligonucleotides targetingIKKb and NIK partially or completely block the effects of TNFa(bottom panel) Note that the sense oligonucleotides (s-NFkB s-IKKb) used as controls had no effect on NFkB activation (c)Schematics of NFkB pathways activated by UV light and TNFaIn keratinocytes the two extracellular stimuli work through inde-pendent pathways that merge at the NFkBIkB complex
regulated transcription factors mediate both photodamage andits alleviation (Fisher et al 1997)
In keratinocytes the EGF and UV responsive pathways donot overlap but UV specifically activates the JNK pathwayThis suggests that the epidermal keratinocytes exhibit distinc-tive specialized UV-responsive signal transduction pathwaysIndeed when we compared the responses to UV in several celltypes we found that the response in keratinocytes is signifi-cantly more stringently controlled than in other cell types Forexample HeLa cells demonstrated a lack of specificity Usinga specific MEK-12 inhibitor we again demonstrated relaxedstringency in HeLa cells We conclude from these results thatkeratinocytes being often exposed to ambient UV have de-veloped more stringent and specific responses to UV than othercell types which are not commonly exposed to UV
At present we do not understand the relevance of JNK3 ex-pression in keratinocytes This isoform it was reported isspecifically expressed in the brain (Yoshida et al 2001) Wespeculate that perhaps the sequestration of JNK1 by the speci-ficity of the UV response creates a need for JNK3 as the moregeneral less restrictive stress signal transducer
TNFa one of proinflammatory cytokines important in cuta-neous inflammation activates NFkB via IKKs kinases that canbe activated by MEKK or NIK (Hirano et al 1996 Lee et al1997) In HeLa cells the phosphorylation of IkBa at Ser-32and Ser-36 residues is essential for the IKK mediated IkBa
degradation caused by TNFa IL-1 or ionizing radiation Us-ing specific inhibitors and antisense oligonucleotides we foundthat the NFkB activation by UVBUVC does not involve IKKs
The Ser-32 and Ser-36 residues are not involved in the UV-induced IkBa degradation (Bender et al 1997 N Li and Karin1998) Because IKKb specifically phosphorylates IkB at bothSer-32 and Ser-36 of IkBa these facts support our results Re-cently it was suggested that UV activation of NFkB in HeLacells may involve the RasndashRaf pathway although not in ker-atinocytes (Tobin et al 1996) This leads to the question howUV activates NFkB in keratinocytes Two new kinases IKKlaquo
and pp90rsk have been found to phosphorylate IkBa (Ghodaet al 1997 Schouten et al 1997 Shimada et al 1999 Israel2000 Peters et al 2000) However IKKlaquo phosphorylates theSer-36 residue while pp90rsk phosphorylates the Ser-32residue only Therefore these kinases are probably not involvedin the UV activated pathway Amino acids between IkBa po-sition 277 and 297 (the PEST region) are apparently requiredfor the UV induced degradation of IkB (Ito et al 1995 Ben-der et al 1998) but the signaling events that effect this degra-dation in keratinocytes are still unknown The data suggest thatTNFa and UVC use different pathways to activate NFkB inHeLa cells Specifically while both UVC and TNFa activateNFkB the TNFa signal goes via IKKs but the UVC signaldoes not From these data it appears that the convergence ofthe UVB and TNFa signals occurs downstream from IKKs thatis at the modification of IkB Overall these results imply thatthe epidermis a natural target of UV light has an exceptionalmolecular UV defense system distinct from UV responsive sys-tems in other tissues
The responses to the three different wavelengths of UV areespecially strictly controlled in keratinocytes Although allwavelengths cause DNA damage and a release of cytokines wefound that the inherent molecular responses to UVA are com-pletely different from the responses to UVBUVC For exam-ple UVBUVC but not UVA activate JNK and NFkB in ker-atinocytes Furthermore UVA activates the JNK pathway inHeLa cells but not in keratinocytes while UVBUVC activatethis pathway in both cells (Rosette and Karin 1996) Similarlyin both HeLa cells and cultured keratinocytes UVBUVC ac-tivate NFkB via a kinase different from IKKs while UVA doesnot activate NFkB at all We conclude that UVBUVC activatea completely different system of signal transduction from UVA
The results suggesting specific signal transduction pathwaysfor UVBUVC in keratinocytes presented here agree well withour studies of UVB-regulated genes using DNA micro arrays(Li et al 2001) We found that in response to UVB ker-atinocytes not only induce the immediate early genes and stressresponse genes but also elaborate responses designed to pro-tect the organism underneath For example chemokines of theIL-8 family are induced which alert the surrounding tissue tothe UV damage synthesis of the cornified envelope compo-nents is enhanced which thickens the stratum corneum etc Weconclude that human epidermal keratinocytes being naturallyexposed to UV light have elaborated highly specific molecu-lar responses to UVBUVC at the signal transduction level aswell as the gene expression level
ACKNOWLEDGMENTS
Our research was supported by the National Institutes ofHealth Grants AR30682 AR39176 AR40522 and AR41850
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 675
FIG 9 Specificity in stress response Epidermal ker-atinocytes exhibit specialized UV-responsive signal transduc-tion pathways which apparently do not network and intersectwith other pathways such as those responsive to EGF or TNFaIn contrast in other cell types such as HeLa multiple path-ways crossover interact and overlap
and by the Anaderm Research Corporation Special thinks goto Dr M Simon for the generous gift of keratinocytes We alsothank members of our laboratory for advice reagents and en-couragement
REFERENCES
ALKALAY I YARON A HATZUBAI A ORIAN ACIECHANOVER A and BEN-NERIAH Y (1995) Stimulation-dependent I kappa B alpha phosphorylation marks the NF-kappa Binhibitor for degradation via the ubiquitin-proteasome pathway ProcNatl Acad Sci USA 92 10599ndash10603
BAEUERLE PA and HENKEL T (1994) Function and activationof NF-kappa B in the immune system Annu Rev Immunol 12141ndash179
BARNES PJ and KARIN M (1997) Nuclear factor-kappaB A piv-otal transcription factor in chronic inflammatory diseases N EnglJ Med 336 1066ndash1071
BEG AA FINCO TS NANTERMET PV and BALDWIN AJR (1993) Tumor necrosis factor and interleukin-1 lead to phos-phorylation and loss of I kappa B alpha A mechanism for NF-kappaB activation Mol Cell Biol 13 3301ndash3310
BENDER K BLATTNER C KNEBEL A IORDANOV M HER-RLICH P and RAHMSDORF HJ (1997) UV-induced signaltransduction J Photochem Photobiol B 37 1ndash17
BENDER K GOTTLICHER M WHITESIDE S RAHMSDORFHJ and HERRLICH P (1998) Sequential DNA damage-indepen-dent and -dependent activation of NF-kappaB by UV EMBO J 175170ndash5181
BROWN K GERSTBERGER S CARLSON L FRANZOSO Gand SIEBENLIST U (1995) Control of I kappa B-alpha proteoly-sis by site-specific signal-induced phosphorylation Science 2671485ndash1488
DELHASE M HAYAKAWA M CHEN Y and KARIN M(1999) Positive and negative regulation of IkappaB kinase activitythrough IKKbeta subunit phosphorylation Science 284 309ndash313
DEacuteRIJARD B et al (1994) JNK1 A protein kinase stimulated byUV light and Ha-Ras that binds and phosphorylates the c-Jun acti-vation domain Cell 76 1025ndash1037
DEVARY Y ROSETTE C DIDONATO JA and KARIN M(1993) NF-kappa B activation by ultraviolet light not dependent ona nuclear signal Science 261 1442ndash1445
DIDONATO JA HAYAKAWA M ROTHWARF DM ZANDIE and KARIN M (1997) A cytokine-responsive IkappaB kinasethat activates the transcription factor NF-kappaB Nature 388548ndash554
FANGER GR GERWINS P WIDMANN C JARPE MB andJOHNSON GL (1997) MEKKs GCKs MLKs PAKs TAKs andtpls Upstream regulators of the c-Jun amino-terminal kinases CurrOpin Genet Dev 7 67ndash74
FEIGE U MORIMOTO RI YAHARA I and POLLA BS eds(1996) Stress-Inducible Cellular Responses (Birkhauser VerlagBasel Switzerland)
FISHER GJ WANG ZQ DATTA SC VARANI J KANG Sand VOORHEES JJ (1997) Pathophysiology of premature skin ag-ing induced by ultraviolet light N Engl J Med 337 1419ndash1428
GHODA L LIN X and GREENE WC (1997) The 90-kDa ribo-somal S6 kinase (pp90rsk) phosphorylates the N-terminal regulatorydomain of IkappaBalpha and stimulates its degradation in vitro JBiol Chem 272 21281ndash21288
GILCHREST B (1995) Photodamage (Blackwell Scientific NewYork)
GRETHER-BECK S BUETTNER R and KRUTMANN J (1997)Ultraviolet A radiation-induced expression of human genes Molec-ular and photobiological mechanisms Biol Chem 378 1231ndash1236
GRILLI M PIZZI M MEMO M and SPANO P (1996) Neuro-protection by aspirin and sodium salicylate through blockade of NF-kappaB activation Science 274 1383ndash1385
GUPTA S et al (1996) Selective interaction of JNK protein kinaseisoforms with transcription factors EMBO J 15 2760ndash2770
HEHNER SP et al (1998) Sesquiterpene lactones specifically in-hibit activation of NF-kappa B by preventing the degradation of Ikappa B-alpha and I kappa B-beta J Biol Chem 273 1288ndash1297
HERSKOWITZ I (1995) MAP kinase pathways in yeast For matingand more Cell 80 187ndash197
HIBI M LIN A SMEAL T MINDEN A and KARIN M (1993)Identification of an oncoprotein- and UV-responsive protein kinasethat binds and potentiates the c-Jun activation domain Genes Dev7 2135ndash2148
HIRANO M et al (1996) MEK kinase is involved in tumor necro-sis factor alpha-induced NF-kappaB activation and degradation ofIkappaB-alpha J Biol Chem 271 13234ndash13238
HU Y et al (1999) Abnormal morphogenesis but intact IKK activa-tion in mice lacking the IKKalpha subunit of IkappaB kinase Sci-ence 284 316ndash320
IORDANOV M et al (1997) CREB is activated by UVC through ap38HOG-1-dependent protein kinase EMBO J 16 1009ndash1022
ISREAL A (2000) The IKK complex An integrator of all signals thatactivate NF-kappaB Trends Cell Biol 10 129ndash133
ITO CY ADEY N BAUTCH VL and BALDWIN AS (1995)Structure and evolution of the human IKBA gene Genomics 29490ndash495
JIANG CK CONNOLLY D and BLUMENBERG M (1991)Comparison of methods for transfection of human epidermal ker-atinocytes J Invest Dermatol 97 969ndash973
JIANG CK FLANAGAN S OHTSUKI M SHUAI K FREED-BERG IM and BLUMENBERG M (1994) Disease-activatedtranscription factor Allergic reactions in human skin cause nucleartranscription of STAT-91 and induce synthesis of keratin K17 MolCell Biol 14 4759ndash4769
JIANG CK MAGNALDO T OHTSUKI M FREEDBERG IMBERNERD F and BLUMENBERG M (1993) Epidermal growthfactor and transforming growth factor alpha specifically induce theactivation- and hyperproliferation-associated keratins 6 and 16 ProcNatl Acad Sci USA 90 6786ndash6790
KALLUNKI T DENG T HIBI M and KARIN M (1996) c-Juncan recruit JNK to phosphorylate dimerization partners via specificdocking interactions Cell 87 929ndash939
KARIN M (1995) The regulation of AP-1 activity by mitogen-acti-vated protein kinases J Biol Chem 270 16483ndash16486
KARIN M and HUNTER T (1995) Transcriptional control by pro-tein phosphorylation Signal transmission from the cell surface to thenucleus Curr Biol 5 747ndash757
KLAMPFER L LEE TH HSU W VILCEK J and CHEN-KIANG S (1994) NF-IL6 and AP-1 cooperatively modulate the ac-tivation of the TSG-6 gene by tumor necrosis factor alpha and in-terleukin-1 Mol Cell Biol 14 6561ndash6569
KOMINE M et al (2000) Inflammatory versus proliferative pro-cesses in epidermis Tumor necrosis factor alpha induces K6b ker-atin synthesis through a transcriptional complex containing NFkappaB and CEBPbeta J Biol Chem 275 32077ndash32088
KOPP E and GHOSH S (1994) Inhibition of NF-kappa B by so-dium salicylate and aspirin Science 265 956ndash959
KYRIAKIS JM et al (1994) The stress-activated protein kinase sub-family of c-Jun kinases Nature 369 156ndash160
LEE FS HAGLER J CHEN ZJ and MANIATIS T (1997) Ac-tivation of the IkappaB alpha kinase complex by MEKK1 a kinaseof the JNK pathway Cell 88 213ndash222
LEPPA S and BOHMANN D (1999) Diverse functions of JNK sig-naling and c-Jun in stress response and apoptosis Oncogene 186158ndash6162
ADACHI ET AL676
LI D TURI TG SCHUCK A FREEDBERG IM KHITROVG and BLUMENBERG M (2001) Rays and arrays The tran-scriptional program in the response of human epidermal keratinocytesto UVB illumination FASEB J 15 2533ndash2535
LI N and KARIN M (1998) Ionizing radiation and short wavelengthUV activate NF-kappaB through two distinct mechanisms ProcNatl Acad Sci USA 95 13012ndash13017
LIU ZG HSU H GOEDDEL DV and KARIN M (1996) Dis-section of TNF receptor 1 effector functions JNK activation is notlinked to apoptosis while NF-kappaB activation prevents cell deathCell 87 565ndash576
MERCURIO F and MANNING AM (1999) NF-kappaB as a pri-mary regulator of the stress response Oncogene 18 6163ndash6171
MIYAMOTO S and VERMA IM (1995) RelNF-kappa BI kappaB story Adv Cancer Res 66 255ndash292
NESTLE FO MITRA RS BENNETT CF CHAN H andNICKOLOFF BJ (1994) Cationic lipid is not required for uptakeand selective inhibitory activity of ICAM-1 phosphorothioate anti-sense oligonucleotides in keratinocytes J Invest Dermatol 103569ndash575
PETERS RT LIAO SM and MANIATIS T (2000) IKKepsilonis part of a novel PMA-inducible IkappaB kinase complex Mol Cell5 513ndash522
PIERCE JW READ MA DING H LUSCINSKAS FW andCOLLINS T (1996) Salicylates inhibit I kappa B-alpha phos-phorylation endothelial-leukocyte adhesion molecule expressionand neutrophil transmigration J Immunol 156 3961ndash3969
PIERCE JW et al (1997) Novel inhibitors of cytokine-induced Ikap-paBalpha phosphorylation and endothelial cell adhesion molecule ex-pression show anti-inflammatory effects in vivo J Biol Chem 27221096ndash21103
REUNING U et al (1995) Inhibition of NF-kappa B-Rel A expres-sion by antisense oligodeoxynucleotides suppresses synthesis ofurokinase-type plasminogen activator (uPA) but not its inhibitor PAI-1 Nucleic Acids Res 23 3887ndash3893
ROSETTE C and KARIN M (1996) Ultraviolet light and osmoticstress activation of the JNK cascade through multiple growth factorand cytokine receptors Science 274 1194ndash1197
SCHOUTEN GJ et al (1997) IkappaB alpha is a target for the mitogen-activated 90 kDa ribosomal S6 kinase EMBO J 163133ndash3144
SHIMADA T et al (1999) IKK-i a novel lipopolysaccharide-inducible kinase that is related to IkappaB kinases Int Immunol 111357ndash1362
SIMON M and GREEN H (1984) Participation of membrane-asso-ciated proteins in the formation of the cross-linked envelope of thekeratinocyte Cell 36 827ndash834
SIMON MM ARAGANE Y SCHWARZ A LUGER TA andSCHWARZ T (1994) UVB light induces nuclear factor kappa B(NF kappa B) activity independently from chromosomal DNA dam-age in cell-free cytosolic extracts J Invest Dermatol 102 422ndash427
TOBIN D NILSSON M and TOFTGARD R (1996) Ras-inde-pendent activation of Rel-family transcription factors by UVB andTPA in cultured keratinocytes Oncogene 12 785ndash793
TOMIC-CANIC M KOMINE M FREEDBERG IM and BLU-MENBERG M (1998) Epidermal signal transduction and tran-scription factor activation in activated keratinocytes J Dermatol Sci17 167ndash181
WILLIAMS BR (1999) PKR A sentinel kinase for cellular stressOncogene 18 6112ndash6120
WU Z WU J JACINTO E and KARIN M (1997) Molecularcloning and characterization of human JNKK2 a novel Jun NH2-terminal kinase-specific kinase Mol Cell Biol 17 7407ndash7416
YANG DD et al (1997) Absence of excitotoxicity-induced apopto-sis in the hippocampus of mice lacking the Jnk3 gene Nature 389865ndash870
YANKULOV K BLAU J PURTON T ROBERTS S and BENT-LEY DL (1994) Transcriptional elongation by RNA polymeraseII is stimulated by transactivators Cell 77 749ndash759
YIN MJ YAMAMOTO Y and GAYNOR RB (1998) The anti-inflammatory agents aspirin and salicylate inhibit the activity ofI(kappa)B kinase-beta Nature 396 77ndash80
YOSHIDA S et al (2001) The c-Jun NH2-terminal kinase3 (JNK3)gene Genomic structure chromosomal assignment and loss of ex-pression in brain tumors J Hum Genet 46 182ndash187
Address reprint requests toM Blumenberg PhD
The NYU Cancer InstituteNew York University School of Medicine
550 First AveNew York NY 10016
E-mail blumem01mednyuedu
Received for publication June 5 2003 received in revised formJuly 15 2003 accepted August 14 2003
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 677

gated antimouse IgG secondary antibody (Vecstatin ABC-mouse IgG kit from Vector Laboratories Burlingame CA) atroom temperature for 1 h incubated with ABC complex (Vec-tor Laboratories) at room temperature for 1 h and treated with339-Diaminobenzidine-tetrahydrochloride (Dojindo Gaithers-burg MD) and 001 H2O2 in Tris pH 76 for 2 min The sam-ples were observed and photographed under the light micro-scope (Microphot-FXA Nikon Melville NY) Additionalantibodies used were from SantaCruz Biotechnology (SantaCruz CA)
RESULTS
UVB signal transduction pathway part I activation ofthe JNK pathway
To determine which JNK isoforms exist in keratinocytes andcan be activated by UVB we have grown primary cultures ofhuman epidermal keratinocytes and illuminated them with UVBat various doses As controls we used HeLa cells and normalhuman dermal fibroblasts The JNK activity was examined inthree ways First illuminated cells were stained with pan-JNKor active JNK-specific antibody to demonstrate the transloca-tion of JNK from cytoplasm to nucleus (not shown) Secondprotein extracts were prepared and analyzed using Western blotswith active JNK-specific antibody Third the kinase activitywas determined directly in affinity-precipitated JNK using aGST-tagged Jun protein fragment as the substrate The resultsof all three approaches were perfectly consistent UVB stronglyand specifically activates JNK in human epidermal ker-atinocytes (Fig 1)
Three isotypes of JNK have been described each with sev-eral splice variants It should be noted that there is confusionregarding which isotype is JNK2 and which is JNK3 in the lit-erature and even more egregiously in the blurbs of manufac-turers who provide antibodies We have adopted the unam-biguous designation of the NCBI databank To determine whichisotypes are expressed in keratinocytes we developed a set ofspecific RT-PCR oligonucleotides and used them to amplify theJNK mRNAs Surprisingly we found JNK1 and JNK3 in ker-atinocytes while JNK2 was absent (Fig 2a) This is an unex-pected finding because JNK1 and JNK2 are commonly almostubiquitously expressed while JNK3 was thought to be brain-specific (Yang et al 1997) Repeat of the PCR assay using alarge number of cycles confirmed the absence of JNK2 in ker-atinocytes (Fig 2b) Furthermore because of the importanceand unexpectedness of this result we decided to confirm theexpression of JNK3 protein in keratinocytes using Western blot-ting Indeed we find that JNK3 is expressed in keratinocytesbut not in HeLa cells (Fig 2c) Therefore keratinocytes expressboth the mRNA and the protein of JNK3 a kinase previouslythought to be brain-specific
To demonstrate that the activation of JNK is functional inthat it activates the Elk1 transcription factor in keratinocyteswe transfected into these cells a set of plasmids containing aGal4-CAT reporter and Gal4-DBDElk1-AF (Yankulov et al1994) The first plasmid contains a set of Gal4 binding sites up-stream from the CAT reporter gene the second expresses a fu-sion protein containing the Gal4 DNA binding domains fused
to the activating domain of transcription factor Elk1 When ac-tivated by UV via JNK the fusion protein activates transcrip-tion but if not activated it is silent Thus the system serves asan UV-responsive transcription reporter The transfected cellswere illuminated with UV and harvested 16 h later We foundthat both UVC and UVB activate the reporter but UVA doesnot (Fig 3) Note that at the 50 mJcm2 dose of UVC a signif-icant number of cells are killed These results mean that theJNK system in keratinocytes is present functional and specif-ically activated by short wavelength UV light UVBUVC butnot the long wavelength UVA We conclude that UVA doesnot activate the same system of signal transduction thatUVBUVC do
To identify the kinases responsible for UV signal we usedantisense approaches The antisense oligonucleotides were tar-geted against JNK1 and JNKK1 After 16 h to allow the anti-sense oligonucleotides to inhibit the synthesis of their targetsthe cells were irradiated with UV and after another 24 h har-vested for CAT determination We found that both antisensereagents attenuated the signals (Fig 4a and b) The antisenseeffects are specific because the sense oligonucleotides used ascontrols had no effect These results indicate that JNK1 andJNKK1 are essential for the transmission of the UVBUVC sig-
ADACHI ET AL670
FIG 3 Activation of transfected Elk-CAT reporter by UVBand UVC in keratinocytes GAL4-Elk1 fusion protein express-ing plasmid was cotransfected into keratinocytes with aGAL4CAT reporter The top panel shows the two cotransfectedplasmids and the effect of UV The transfected cells were illu-minated with indicated doses of UVA UVB or UVC harvested16 h later and the CAT levels determined Note that at the 50mJcm2 of UVC a significant number of cells is killed whichresults in significantly reduced levels of CAT activity (aster-isk) At the doses used UVA did not affect cell viability Thebars represent averages of duplicate transfections and the repli-cates differed by 25 or less
nal However they do not formally exclude the possibility thatJNK3 and JNKK2 also play a role
To determine whether the Elk1 activation is specific for theUV responsive signals in keratinocytes we tested the effects ofEGF In other cell types EGF can act through pathways thatactivate both JNK and Elk1 However when we attempted toactivate Elk1 using EGF in keratinocytes we were unsuccess-ful (Fig 4b) This means that in keratinocytes the EGF and UVresponsive pathways do not overlap on the contrary UV specif-ically activates the JNK pathway This suggests that the epi-dermal keratinocytes exhibit distinctive specialized UV-responsive signal transduction pathways
Indeed when we compared the responses to UV in severalcell types we found that the response in keratinocytes is sig-nificantly more stringently controlled than in other cell typesFor example both UV and EGF activated the JNK pathway inHeLa cells (Fig 4c and d) demonstrating a lack of specificityMoreover targeting JNK1 and JNKK1 with antisense oligonu-cleotides only partially inhibited the UV effects in HeLa cells
implying that in these cells parallel pathways can respond toUV to activate Elk1 (Fig 4c) Suspecting that EGF activatesElk1 through both the JNK and the ERKMAPK pathways inHeLa we inhibited the latter with U0126 a specific MEK-12inhibitor We found that U0126 significantly blocks the acti-vation by EGF but not by UV (Fig 4e) again demonstratingrelaxed stringency in HeLa cells
To approach these phenomena from another angle we usedkinase assays and Western blots Consistent with the above re-sults we found that while in keratinocytes only UVB (andUVC) could activate the JNK pathway in HeLa cells (and fi-broblasts not shown) heat shock UVA and osmotic shockcould activate the pathway as well (Fig 5a) In HeLa cells andfibroblasts UVB could activate the ERK pathway (not shown)but in keratinocytes it could not (Fig 5b) We conclude fromthese results that keratinocytes being often exposed to ambi-ent UV have developed more stringent and specific responsesto UV than other cell types which are not commonly exposedto UV These data are summarized in Figure 5c
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 671
FIG 4 UV specifically activates JNK in keratinocytes but not in HeLa cells GAL4-Elk1 and GAL4CAT plasmids were co-transfected into keratinocytes (a and b) or HeLa cells (c d and e) The transfected cells were treated with either UVC (5 mJcm2)or EGF (100 ngml) and harvested 24 h later for determination of CAT activity U0126a specific inhibitor of the MAPK path-way blocks the effects of EGF but not of UV Open bars represent the untreated control cells closed bars the UV-treated onesand the shaded bars the EGF treated ones
While there are only two JNKKs kinases that directly phos-phorylate JNK there is a large family of JNKKKs kinases thatphosphorylate JNKKs In a manuscript in preparation we de-scribe our analysis of the kinases expressed by keratinocytes
To demonstrate that the JNK system is responsive in vivowe used a novel ex vivo approach Biopsies of human skin weretreated with as-JNKK1 oligonucleotide or left untreated as con-trol Then both the treated and untreated biopsies were illumi-nated with UVC and shortly thereafter processed for immuno-histochemical detection of activated JNK (Fig 6) We found inthese biopsies that UV caused activation and nuclear translo-cation of JNK Furthermore this activation was abrogated bypretreatment with as-JNKK1 the antisense oligonucleotide tar-geting JNKK1 again demonstrating that JNKK1 is essential foractivation of JNK by UV in epidermis These results mean that
the specific signal transduction pathways we found operationalin cultured keratinocytes are also functional and working thesame way in human skin in vivo
UVB signal transduction pathway part II activationof the NFkB pathway
Having established that keratinocytes possess a specific UV-responding pathway to activate JNK we wanted to examine thepossibility that the UV-specific signal transduction pathwaysactivating NFkB are also specific in these cells The canonicalactivator of NFkB is TNFa however recent evidence suggeststhat UV and TNFa may activate NFkB through different mech-anisms (N Li and Karin 1998) Demonstrating a UV-specificpathway would strengthen even more our hypothesis that thereis specificity in UV responsive signal transduction in humanepidermal keratinocytes Therefore we transfected into ker-atinocytes and into HeLa cells a reporter plasmid containingthree tandem NFkB sites linked to the TK promoter-drivenCAT We found that the exposure of transfected cells to TNFaUVB or UVC resulted in the induction of the NFkB responseelement (Fig 7) Generally at the optimal conditions the in-duction of NFkB by UV was weaker than the induction byTNFa On the other hand NFkB was not activated by UVA atall even at high doses (Fig 7)
To compare the mechanisms of NFkB activation by UV andTNFa we have used a battery of specific inhibitors of the NFkBpathway which inhibit the pathway at different points (Fig 8a)Namely Bay11-7085 inhibits upstream from IKK (Pierce et al1997) aspirin has recently been found to be a competitive in-hibitor of ATP binding to IKKb (Yin et al 1998) whereasparthenolide directly inhibits the degradation of IkBa (Pierceet al 1997) When we treated transfected keratinocytes or HeLacells with these three inhibitors we found that Bay11-7085 in-hibited the induction of NFkB by TNFa in a dose-dependentmanner while it has almost no effect on the induction of NFkBby UVC (Fig 8b) The same results were obtained by using as-pirin (Fig 8c) In contrast parthenolide inhibited both the UVCand the TNFa effects in parallel (Fig 8d) These results con-firm that TNFa and UVC use different pathways to activateNFkB in HeLa cells Specifically while both UVC and TNFaactivate NFkB the TNFa signal goes via IKKs but the UVCsignal does not From these data we conclude that the conver-gence of the UVB and TNFa signals occurs downstream fromIKKs that is at the modification of IkB
One caveat in the use of inhibitors of specific kinases is thepossibility that such inhibitors may affect additional pathwaysnot currently recognized as affected by the given inhibitor Toaddress such specificity concerns and to examine the involve-ment of IKK in the induction of NFkB by TNFa UVC andUVB we used antisense oligonucleotides specifically targetingNFkB and IKKb as-NFkB as-IKKb (Table 1) (Reuning etal 1995 Komine et al 2000) In parallel we added the senseoligonucleotides s-NFkB s-IKKb as controls The antisenseexperiments confirmed the fact that UVBUVC use a clearlydifferent pathway to activate NFkB than TNFa does The sup-pression of NFkB synthesis in cultured keratinocytes by addingas-NFkB resulted in inhibition of activation of NFkB by allthree agents UVC UVB and TNFa as expected (Fig 8b) Onthe other hand the suppression of IKKb synthesis by adding
ADACHI ET AL672
FIG 5 Specific activation of JNK in keratinocytes andpromiscuous activation of JNK in HeLa cells (a) JNK activityin keratinocytes and HeLa cells treated with UVC UVA heatshock or EGF Duplicate experiments were performed with allagents except EGF Each lane contains 10 mg of protein InHeLa cells (bottom panel) UVC and EGF strongly while UVAand heat detectably activate JNK Cells were harvested 30 minafter treatment (b) Lack of activation of ERK by UVB in ker-atinocytes The top panel shows a Western blot with the anti-body specific for the activated ERKs the bottom for totalERK1 as a control for loading and transfer (c) Specificity insignal transduction In keratinocytes UVB specifically acti-vates JNK and EGF specifically activates ERKs whereas inHeLa cells all three extracellular signals UVB EGF and heatactivate both JNK and ERK Fibroblasts respond similar toHeLa cells (data not shown)
as-IKKb only inhibited the activation by TNFa but had no ef-fect on the activation by either UVC or UVB (Fig 8b) In theseexperiments we used s-NFkB and s-IKKa as controls and con-firmed that they had no effect in this pathway (Fig 8b) Fur-thermore as-NIK targeting a kinase functioning upstream fromIKK also partially inhibited the effects of TNFa Therefore
the antisense oligonucleotide experiments confirm our conclu-sions that the activation of NFkB by TNFa depends on the IKKpathway whereas the response to UVC or UVB occurs througha mechanism independent of IKKs
Furthermore as an additional control we used oligonu-cleotides targeting the JNK pathway as-JNKK1 and s-JNKK1Although they affected the JNK pathway (see Figs 4 and 6)they did not affect the NFkB pathway (Fig 8b) which indi-cates that the activation of the JNK pathway and the NFkBpathway are independent in keratinocytes This means that epi-dermal keratinocytes contain multiple specific UV responsivemechanisms
DISCUSSION
Our data suggest that UVBUVC illumination in epidermalkeratinocytes is not a generic stress response inducer but a spe-cific extracellular agent that activates specific signal transduc-tion pathways In particular we show that in keratinocytes UVexclusively activates the JNK pathway while in HeLa cellsUV also activates the parallel ERK and p38 pathways Con-versely in keratinocytes exclusively UV activates the JNKpathway while in HeLa cells growth factors can do so as wellIn parallel in keratinocytes UV uses a distinct pathway to ac-tivate the NFkB transcription factor a pathway separate fromthe more common TNFa activated one (Fig 9)
The specific activation of JNK by UVBUVC is particularlyrelevant because the JNK pathway is in different cell types atarget of multiple stress agents The balance between apoptosisand proliferation is at times regulated by JNK and the tran-scription factors activated by JNK regulate the expression ofmany genes (Karin and Hunter 1995) It is uniquely importantin epidermis that JNK exclusively responds to UVBUVC be-cause epidermis is the natural target of UVBUVC and the JNK-
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 673
FIG 6 Activation of JNK in vivo and the effect of as-JNKK1 In the control sample (left) JNK is not activated as shown us-ing an antibody specific for the activated phosphorylated form of JNK Treatment with 100 mJcm2 causes strong activation ofJNK (middle) Pretreatment for 3 h with the antisense oligonucleotide targeting JNKK1 abolishes this activation (right)
FIG 7 TNFa UVB and UVC activate NFkB while UVAdoes not Reporter plasmid NFkB3-CAT (top) which containsthree tandem NFkB sites upstream from the TK promoter wastransfected into keratinocytes The transfected cells were eithertreated with 50 ngml of TNFa or illuminated with indicateddoses of UV harvested 16 h later and the CAT levels deter-mined Untreated cells served as a control The replicates dif-fered by 25 or less
ADACHI ET AL674
FIG 8 Specific inhibitors can differentiate between the TNFa-and UVB-dependent signal transduction pathways (a) The effectsof specific inhibitors on NFkB activation Cultured keratinocyteswere treated with either UVB (8 mJcm2) or TNFa (50 ngml) af-ter transfection of the NFkB response element Inhibitor Bay11-7085 specifically inhibits upstream kinases of IKK aspirin inhibitsIKKb parthenolide blocks IkBa degradation (b) the effects of an-tisense oligonucleotides targeting signaling cascades on NFkB ac-tivation in keratinocytes Oligonucleotide as-NFkB targeting thetranscription factor blocks the activation by UVB and UVC whileoligonucleotides targeting IKKb JNK1 and JNKK1 are withouteffect (top two panels) In contrast oligonucleotides targetingIKKb and NIK partially or completely block the effects of TNFa(bottom panel) Note that the sense oligonucleotides (s-NFkB s-IKKb) used as controls had no effect on NFkB activation (c)Schematics of NFkB pathways activated by UV light and TNFaIn keratinocytes the two extracellular stimuli work through inde-pendent pathways that merge at the NFkBIkB complex
regulated transcription factors mediate both photodamage andits alleviation (Fisher et al 1997)
In keratinocytes the EGF and UV responsive pathways donot overlap but UV specifically activates the JNK pathwayThis suggests that the epidermal keratinocytes exhibit distinc-tive specialized UV-responsive signal transduction pathwaysIndeed when we compared the responses to UV in several celltypes we found that the response in keratinocytes is signifi-cantly more stringently controlled than in other cell types Forexample HeLa cells demonstrated a lack of specificity Usinga specific MEK-12 inhibitor we again demonstrated relaxedstringency in HeLa cells We conclude from these results thatkeratinocytes being often exposed to ambient UV have de-veloped more stringent and specific responses to UV than othercell types which are not commonly exposed to UV
At present we do not understand the relevance of JNK3 ex-pression in keratinocytes This isoform it was reported isspecifically expressed in the brain (Yoshida et al 2001) Wespeculate that perhaps the sequestration of JNK1 by the speci-ficity of the UV response creates a need for JNK3 as the moregeneral less restrictive stress signal transducer
TNFa one of proinflammatory cytokines important in cuta-neous inflammation activates NFkB via IKKs kinases that canbe activated by MEKK or NIK (Hirano et al 1996 Lee et al1997) In HeLa cells the phosphorylation of IkBa at Ser-32and Ser-36 residues is essential for the IKK mediated IkBa
degradation caused by TNFa IL-1 or ionizing radiation Us-ing specific inhibitors and antisense oligonucleotides we foundthat the NFkB activation by UVBUVC does not involve IKKs
The Ser-32 and Ser-36 residues are not involved in the UV-induced IkBa degradation (Bender et al 1997 N Li and Karin1998) Because IKKb specifically phosphorylates IkB at bothSer-32 and Ser-36 of IkBa these facts support our results Re-cently it was suggested that UV activation of NFkB in HeLacells may involve the RasndashRaf pathway although not in ker-atinocytes (Tobin et al 1996) This leads to the question howUV activates NFkB in keratinocytes Two new kinases IKKlaquo
and pp90rsk have been found to phosphorylate IkBa (Ghodaet al 1997 Schouten et al 1997 Shimada et al 1999 Israel2000 Peters et al 2000) However IKKlaquo phosphorylates theSer-36 residue while pp90rsk phosphorylates the Ser-32residue only Therefore these kinases are probably not involvedin the UV activated pathway Amino acids between IkBa po-sition 277 and 297 (the PEST region) are apparently requiredfor the UV induced degradation of IkB (Ito et al 1995 Ben-der et al 1998) but the signaling events that effect this degra-dation in keratinocytes are still unknown The data suggest thatTNFa and UVC use different pathways to activate NFkB inHeLa cells Specifically while both UVC and TNFa activateNFkB the TNFa signal goes via IKKs but the UVC signaldoes not From these data it appears that the convergence ofthe UVB and TNFa signals occurs downstream from IKKs thatis at the modification of IkB Overall these results imply thatthe epidermis a natural target of UV light has an exceptionalmolecular UV defense system distinct from UV responsive sys-tems in other tissues
The responses to the three different wavelengths of UV areespecially strictly controlled in keratinocytes Although allwavelengths cause DNA damage and a release of cytokines wefound that the inherent molecular responses to UVA are com-pletely different from the responses to UVBUVC For exam-ple UVBUVC but not UVA activate JNK and NFkB in ker-atinocytes Furthermore UVA activates the JNK pathway inHeLa cells but not in keratinocytes while UVBUVC activatethis pathway in both cells (Rosette and Karin 1996) Similarlyin both HeLa cells and cultured keratinocytes UVBUVC ac-tivate NFkB via a kinase different from IKKs while UVA doesnot activate NFkB at all We conclude that UVBUVC activatea completely different system of signal transduction from UVA
The results suggesting specific signal transduction pathwaysfor UVBUVC in keratinocytes presented here agree well withour studies of UVB-regulated genes using DNA micro arrays(Li et al 2001) We found that in response to UVB ker-atinocytes not only induce the immediate early genes and stressresponse genes but also elaborate responses designed to pro-tect the organism underneath For example chemokines of theIL-8 family are induced which alert the surrounding tissue tothe UV damage synthesis of the cornified envelope compo-nents is enhanced which thickens the stratum corneum etc Weconclude that human epidermal keratinocytes being naturallyexposed to UV light have elaborated highly specific molecu-lar responses to UVBUVC at the signal transduction level aswell as the gene expression level
ACKNOWLEDGMENTS
Our research was supported by the National Institutes ofHealth Grants AR30682 AR39176 AR40522 and AR41850
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 675
FIG 9 Specificity in stress response Epidermal ker-atinocytes exhibit specialized UV-responsive signal transduc-tion pathways which apparently do not network and intersectwith other pathways such as those responsive to EGF or TNFaIn contrast in other cell types such as HeLa multiple path-ways crossover interact and overlap
and by the Anaderm Research Corporation Special thinks goto Dr M Simon for the generous gift of keratinocytes We alsothank members of our laboratory for advice reagents and en-couragement
REFERENCES
ALKALAY I YARON A HATZUBAI A ORIAN ACIECHANOVER A and BEN-NERIAH Y (1995) Stimulation-dependent I kappa B alpha phosphorylation marks the NF-kappa Binhibitor for degradation via the ubiquitin-proteasome pathway ProcNatl Acad Sci USA 92 10599ndash10603
BAEUERLE PA and HENKEL T (1994) Function and activationof NF-kappa B in the immune system Annu Rev Immunol 12141ndash179
BARNES PJ and KARIN M (1997) Nuclear factor-kappaB A piv-otal transcription factor in chronic inflammatory diseases N EnglJ Med 336 1066ndash1071
BEG AA FINCO TS NANTERMET PV and BALDWIN AJR (1993) Tumor necrosis factor and interleukin-1 lead to phos-phorylation and loss of I kappa B alpha A mechanism for NF-kappaB activation Mol Cell Biol 13 3301ndash3310
BENDER K BLATTNER C KNEBEL A IORDANOV M HER-RLICH P and RAHMSDORF HJ (1997) UV-induced signaltransduction J Photochem Photobiol B 37 1ndash17
BENDER K GOTTLICHER M WHITESIDE S RAHMSDORFHJ and HERRLICH P (1998) Sequential DNA damage-indepen-dent and -dependent activation of NF-kappaB by UV EMBO J 175170ndash5181
BROWN K GERSTBERGER S CARLSON L FRANZOSO Gand SIEBENLIST U (1995) Control of I kappa B-alpha proteoly-sis by site-specific signal-induced phosphorylation Science 2671485ndash1488
DELHASE M HAYAKAWA M CHEN Y and KARIN M(1999) Positive and negative regulation of IkappaB kinase activitythrough IKKbeta subunit phosphorylation Science 284 309ndash313
DEacuteRIJARD B et al (1994) JNK1 A protein kinase stimulated byUV light and Ha-Ras that binds and phosphorylates the c-Jun acti-vation domain Cell 76 1025ndash1037
DEVARY Y ROSETTE C DIDONATO JA and KARIN M(1993) NF-kappa B activation by ultraviolet light not dependent ona nuclear signal Science 261 1442ndash1445
DIDONATO JA HAYAKAWA M ROTHWARF DM ZANDIE and KARIN M (1997) A cytokine-responsive IkappaB kinasethat activates the transcription factor NF-kappaB Nature 388548ndash554
FANGER GR GERWINS P WIDMANN C JARPE MB andJOHNSON GL (1997) MEKKs GCKs MLKs PAKs TAKs andtpls Upstream regulators of the c-Jun amino-terminal kinases CurrOpin Genet Dev 7 67ndash74
FEIGE U MORIMOTO RI YAHARA I and POLLA BS eds(1996) Stress-Inducible Cellular Responses (Birkhauser VerlagBasel Switzerland)
FISHER GJ WANG ZQ DATTA SC VARANI J KANG Sand VOORHEES JJ (1997) Pathophysiology of premature skin ag-ing induced by ultraviolet light N Engl J Med 337 1419ndash1428
GHODA L LIN X and GREENE WC (1997) The 90-kDa ribo-somal S6 kinase (pp90rsk) phosphorylates the N-terminal regulatorydomain of IkappaBalpha and stimulates its degradation in vitro JBiol Chem 272 21281ndash21288
GILCHREST B (1995) Photodamage (Blackwell Scientific NewYork)
GRETHER-BECK S BUETTNER R and KRUTMANN J (1997)Ultraviolet A radiation-induced expression of human genes Molec-ular and photobiological mechanisms Biol Chem 378 1231ndash1236
GRILLI M PIZZI M MEMO M and SPANO P (1996) Neuro-protection by aspirin and sodium salicylate through blockade of NF-kappaB activation Science 274 1383ndash1385
GUPTA S et al (1996) Selective interaction of JNK protein kinaseisoforms with transcription factors EMBO J 15 2760ndash2770
HEHNER SP et al (1998) Sesquiterpene lactones specifically in-hibit activation of NF-kappa B by preventing the degradation of Ikappa B-alpha and I kappa B-beta J Biol Chem 273 1288ndash1297
HERSKOWITZ I (1995) MAP kinase pathways in yeast For matingand more Cell 80 187ndash197
HIBI M LIN A SMEAL T MINDEN A and KARIN M (1993)Identification of an oncoprotein- and UV-responsive protein kinasethat binds and potentiates the c-Jun activation domain Genes Dev7 2135ndash2148
HIRANO M et al (1996) MEK kinase is involved in tumor necro-sis factor alpha-induced NF-kappaB activation and degradation ofIkappaB-alpha J Biol Chem 271 13234ndash13238
HU Y et al (1999) Abnormal morphogenesis but intact IKK activa-tion in mice lacking the IKKalpha subunit of IkappaB kinase Sci-ence 284 316ndash320
IORDANOV M et al (1997) CREB is activated by UVC through ap38HOG-1-dependent protein kinase EMBO J 16 1009ndash1022
ISREAL A (2000) The IKK complex An integrator of all signals thatactivate NF-kappaB Trends Cell Biol 10 129ndash133
ITO CY ADEY N BAUTCH VL and BALDWIN AS (1995)Structure and evolution of the human IKBA gene Genomics 29490ndash495
JIANG CK CONNOLLY D and BLUMENBERG M (1991)Comparison of methods for transfection of human epidermal ker-atinocytes J Invest Dermatol 97 969ndash973
JIANG CK FLANAGAN S OHTSUKI M SHUAI K FREED-BERG IM and BLUMENBERG M (1994) Disease-activatedtranscription factor Allergic reactions in human skin cause nucleartranscription of STAT-91 and induce synthesis of keratin K17 MolCell Biol 14 4759ndash4769
JIANG CK MAGNALDO T OHTSUKI M FREEDBERG IMBERNERD F and BLUMENBERG M (1993) Epidermal growthfactor and transforming growth factor alpha specifically induce theactivation- and hyperproliferation-associated keratins 6 and 16 ProcNatl Acad Sci USA 90 6786ndash6790
KALLUNKI T DENG T HIBI M and KARIN M (1996) c-Juncan recruit JNK to phosphorylate dimerization partners via specificdocking interactions Cell 87 929ndash939
KARIN M (1995) The regulation of AP-1 activity by mitogen-acti-vated protein kinases J Biol Chem 270 16483ndash16486
KARIN M and HUNTER T (1995) Transcriptional control by pro-tein phosphorylation Signal transmission from the cell surface to thenucleus Curr Biol 5 747ndash757
KLAMPFER L LEE TH HSU W VILCEK J and CHEN-KIANG S (1994) NF-IL6 and AP-1 cooperatively modulate the ac-tivation of the TSG-6 gene by tumor necrosis factor alpha and in-terleukin-1 Mol Cell Biol 14 6561ndash6569
KOMINE M et al (2000) Inflammatory versus proliferative pro-cesses in epidermis Tumor necrosis factor alpha induces K6b ker-atin synthesis through a transcriptional complex containing NFkappaB and CEBPbeta J Biol Chem 275 32077ndash32088
KOPP E and GHOSH S (1994) Inhibition of NF-kappa B by so-dium salicylate and aspirin Science 265 956ndash959
KYRIAKIS JM et al (1994) The stress-activated protein kinase sub-family of c-Jun kinases Nature 369 156ndash160
LEE FS HAGLER J CHEN ZJ and MANIATIS T (1997) Ac-tivation of the IkappaB alpha kinase complex by MEKK1 a kinaseof the JNK pathway Cell 88 213ndash222
LEPPA S and BOHMANN D (1999) Diverse functions of JNK sig-naling and c-Jun in stress response and apoptosis Oncogene 186158ndash6162
ADACHI ET AL676
LI D TURI TG SCHUCK A FREEDBERG IM KHITROVG and BLUMENBERG M (2001) Rays and arrays The tran-scriptional program in the response of human epidermal keratinocytesto UVB illumination FASEB J 15 2533ndash2535
LI N and KARIN M (1998) Ionizing radiation and short wavelengthUV activate NF-kappaB through two distinct mechanisms ProcNatl Acad Sci USA 95 13012ndash13017
LIU ZG HSU H GOEDDEL DV and KARIN M (1996) Dis-section of TNF receptor 1 effector functions JNK activation is notlinked to apoptosis while NF-kappaB activation prevents cell deathCell 87 565ndash576
MERCURIO F and MANNING AM (1999) NF-kappaB as a pri-mary regulator of the stress response Oncogene 18 6163ndash6171
MIYAMOTO S and VERMA IM (1995) RelNF-kappa BI kappaB story Adv Cancer Res 66 255ndash292
NESTLE FO MITRA RS BENNETT CF CHAN H andNICKOLOFF BJ (1994) Cationic lipid is not required for uptakeand selective inhibitory activity of ICAM-1 phosphorothioate anti-sense oligonucleotides in keratinocytes J Invest Dermatol 103569ndash575
PETERS RT LIAO SM and MANIATIS T (2000) IKKepsilonis part of a novel PMA-inducible IkappaB kinase complex Mol Cell5 513ndash522
PIERCE JW READ MA DING H LUSCINSKAS FW andCOLLINS T (1996) Salicylates inhibit I kappa B-alpha phos-phorylation endothelial-leukocyte adhesion molecule expressionand neutrophil transmigration J Immunol 156 3961ndash3969
PIERCE JW et al (1997) Novel inhibitors of cytokine-induced Ikap-paBalpha phosphorylation and endothelial cell adhesion molecule ex-pression show anti-inflammatory effects in vivo J Biol Chem 27221096ndash21103
REUNING U et al (1995) Inhibition of NF-kappa B-Rel A expres-sion by antisense oligodeoxynucleotides suppresses synthesis ofurokinase-type plasminogen activator (uPA) but not its inhibitor PAI-1 Nucleic Acids Res 23 3887ndash3893
ROSETTE C and KARIN M (1996) Ultraviolet light and osmoticstress activation of the JNK cascade through multiple growth factorand cytokine receptors Science 274 1194ndash1197
SCHOUTEN GJ et al (1997) IkappaB alpha is a target for the mitogen-activated 90 kDa ribosomal S6 kinase EMBO J 163133ndash3144
SHIMADA T et al (1999) IKK-i a novel lipopolysaccharide-inducible kinase that is related to IkappaB kinases Int Immunol 111357ndash1362
SIMON M and GREEN H (1984) Participation of membrane-asso-ciated proteins in the formation of the cross-linked envelope of thekeratinocyte Cell 36 827ndash834
SIMON MM ARAGANE Y SCHWARZ A LUGER TA andSCHWARZ T (1994) UVB light induces nuclear factor kappa B(NF kappa B) activity independently from chromosomal DNA dam-age in cell-free cytosolic extracts J Invest Dermatol 102 422ndash427
TOBIN D NILSSON M and TOFTGARD R (1996) Ras-inde-pendent activation of Rel-family transcription factors by UVB andTPA in cultured keratinocytes Oncogene 12 785ndash793
TOMIC-CANIC M KOMINE M FREEDBERG IM and BLU-MENBERG M (1998) Epidermal signal transduction and tran-scription factor activation in activated keratinocytes J Dermatol Sci17 167ndash181
WILLIAMS BR (1999) PKR A sentinel kinase for cellular stressOncogene 18 6112ndash6120
WU Z WU J JACINTO E and KARIN M (1997) Molecularcloning and characterization of human JNKK2 a novel Jun NH2-terminal kinase-specific kinase Mol Cell Biol 17 7407ndash7416
YANG DD et al (1997) Absence of excitotoxicity-induced apopto-sis in the hippocampus of mice lacking the Jnk3 gene Nature 389865ndash870
YANKULOV K BLAU J PURTON T ROBERTS S and BENT-LEY DL (1994) Transcriptional elongation by RNA polymeraseII is stimulated by transactivators Cell 77 749ndash759
YIN MJ YAMAMOTO Y and GAYNOR RB (1998) The anti-inflammatory agents aspirin and salicylate inhibit the activity ofI(kappa)B kinase-beta Nature 396 77ndash80
YOSHIDA S et al (2001) The c-Jun NH2-terminal kinase3 (JNK3)gene Genomic structure chromosomal assignment and loss of ex-pression in brain tumors J Hum Genet 46 182ndash187
Address reprint requests toM Blumenberg PhD
The NYU Cancer InstituteNew York University School of Medicine
550 First AveNew York NY 10016
E-mail blumem01mednyuedu
Received for publication June 5 2003 received in revised formJuly 15 2003 accepted August 14 2003
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 677

nal However they do not formally exclude the possibility thatJNK3 and JNKK2 also play a role
To determine whether the Elk1 activation is specific for theUV responsive signals in keratinocytes we tested the effects ofEGF In other cell types EGF can act through pathways thatactivate both JNK and Elk1 However when we attempted toactivate Elk1 using EGF in keratinocytes we were unsuccess-ful (Fig 4b) This means that in keratinocytes the EGF and UVresponsive pathways do not overlap on the contrary UV specif-ically activates the JNK pathway This suggests that the epi-dermal keratinocytes exhibit distinctive specialized UV-responsive signal transduction pathways
Indeed when we compared the responses to UV in severalcell types we found that the response in keratinocytes is sig-nificantly more stringently controlled than in other cell typesFor example both UV and EGF activated the JNK pathway inHeLa cells (Fig 4c and d) demonstrating a lack of specificityMoreover targeting JNK1 and JNKK1 with antisense oligonu-cleotides only partially inhibited the UV effects in HeLa cells
implying that in these cells parallel pathways can respond toUV to activate Elk1 (Fig 4c) Suspecting that EGF activatesElk1 through both the JNK and the ERKMAPK pathways inHeLa we inhibited the latter with U0126 a specific MEK-12inhibitor We found that U0126 significantly blocks the acti-vation by EGF but not by UV (Fig 4e) again demonstratingrelaxed stringency in HeLa cells
To approach these phenomena from another angle we usedkinase assays and Western blots Consistent with the above re-sults we found that while in keratinocytes only UVB (andUVC) could activate the JNK pathway in HeLa cells (and fi-broblasts not shown) heat shock UVA and osmotic shockcould activate the pathway as well (Fig 5a) In HeLa cells andfibroblasts UVB could activate the ERK pathway (not shown)but in keratinocytes it could not (Fig 5b) We conclude fromthese results that keratinocytes being often exposed to ambi-ent UV have developed more stringent and specific responsesto UV than other cell types which are not commonly exposedto UV These data are summarized in Figure 5c
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 671
FIG 4 UV specifically activates JNK in keratinocytes but not in HeLa cells GAL4-Elk1 and GAL4CAT plasmids were co-transfected into keratinocytes (a and b) or HeLa cells (c d and e) The transfected cells were treated with either UVC (5 mJcm2)or EGF (100 ngml) and harvested 24 h later for determination of CAT activity U0126a specific inhibitor of the MAPK path-way blocks the effects of EGF but not of UV Open bars represent the untreated control cells closed bars the UV-treated onesand the shaded bars the EGF treated ones
While there are only two JNKKs kinases that directly phos-phorylate JNK there is a large family of JNKKKs kinases thatphosphorylate JNKKs In a manuscript in preparation we de-scribe our analysis of the kinases expressed by keratinocytes
To demonstrate that the JNK system is responsive in vivowe used a novel ex vivo approach Biopsies of human skin weretreated with as-JNKK1 oligonucleotide or left untreated as con-trol Then both the treated and untreated biopsies were illumi-nated with UVC and shortly thereafter processed for immuno-histochemical detection of activated JNK (Fig 6) We found inthese biopsies that UV caused activation and nuclear translo-cation of JNK Furthermore this activation was abrogated bypretreatment with as-JNKK1 the antisense oligonucleotide tar-geting JNKK1 again demonstrating that JNKK1 is essential foractivation of JNK by UV in epidermis These results mean that
the specific signal transduction pathways we found operationalin cultured keratinocytes are also functional and working thesame way in human skin in vivo
UVB signal transduction pathway part II activationof the NFkB pathway
Having established that keratinocytes possess a specific UV-responding pathway to activate JNK we wanted to examine thepossibility that the UV-specific signal transduction pathwaysactivating NFkB are also specific in these cells The canonicalactivator of NFkB is TNFa however recent evidence suggeststhat UV and TNFa may activate NFkB through different mech-anisms (N Li and Karin 1998) Demonstrating a UV-specificpathway would strengthen even more our hypothesis that thereis specificity in UV responsive signal transduction in humanepidermal keratinocytes Therefore we transfected into ker-atinocytes and into HeLa cells a reporter plasmid containingthree tandem NFkB sites linked to the TK promoter-drivenCAT We found that the exposure of transfected cells to TNFaUVB or UVC resulted in the induction of the NFkB responseelement (Fig 7) Generally at the optimal conditions the in-duction of NFkB by UV was weaker than the induction byTNFa On the other hand NFkB was not activated by UVA atall even at high doses (Fig 7)
To compare the mechanisms of NFkB activation by UV andTNFa we have used a battery of specific inhibitors of the NFkBpathway which inhibit the pathway at different points (Fig 8a)Namely Bay11-7085 inhibits upstream from IKK (Pierce et al1997) aspirin has recently been found to be a competitive in-hibitor of ATP binding to IKKb (Yin et al 1998) whereasparthenolide directly inhibits the degradation of IkBa (Pierceet al 1997) When we treated transfected keratinocytes or HeLacells with these three inhibitors we found that Bay11-7085 in-hibited the induction of NFkB by TNFa in a dose-dependentmanner while it has almost no effect on the induction of NFkBby UVC (Fig 8b) The same results were obtained by using as-pirin (Fig 8c) In contrast parthenolide inhibited both the UVCand the TNFa effects in parallel (Fig 8d) These results con-firm that TNFa and UVC use different pathways to activateNFkB in HeLa cells Specifically while both UVC and TNFaactivate NFkB the TNFa signal goes via IKKs but the UVCsignal does not From these data we conclude that the conver-gence of the UVB and TNFa signals occurs downstream fromIKKs that is at the modification of IkB
One caveat in the use of inhibitors of specific kinases is thepossibility that such inhibitors may affect additional pathwaysnot currently recognized as affected by the given inhibitor Toaddress such specificity concerns and to examine the involve-ment of IKK in the induction of NFkB by TNFa UVC andUVB we used antisense oligonucleotides specifically targetingNFkB and IKKb as-NFkB as-IKKb (Table 1) (Reuning etal 1995 Komine et al 2000) In parallel we added the senseoligonucleotides s-NFkB s-IKKb as controls The antisenseexperiments confirmed the fact that UVBUVC use a clearlydifferent pathway to activate NFkB than TNFa does The sup-pression of NFkB synthesis in cultured keratinocytes by addingas-NFkB resulted in inhibition of activation of NFkB by allthree agents UVC UVB and TNFa as expected (Fig 8b) Onthe other hand the suppression of IKKb synthesis by adding
ADACHI ET AL672
FIG 5 Specific activation of JNK in keratinocytes andpromiscuous activation of JNK in HeLa cells (a) JNK activityin keratinocytes and HeLa cells treated with UVC UVA heatshock or EGF Duplicate experiments were performed with allagents except EGF Each lane contains 10 mg of protein InHeLa cells (bottom panel) UVC and EGF strongly while UVAand heat detectably activate JNK Cells were harvested 30 minafter treatment (b) Lack of activation of ERK by UVB in ker-atinocytes The top panel shows a Western blot with the anti-body specific for the activated ERKs the bottom for totalERK1 as a control for loading and transfer (c) Specificity insignal transduction In keratinocytes UVB specifically acti-vates JNK and EGF specifically activates ERKs whereas inHeLa cells all three extracellular signals UVB EGF and heatactivate both JNK and ERK Fibroblasts respond similar toHeLa cells (data not shown)
as-IKKb only inhibited the activation by TNFa but had no ef-fect on the activation by either UVC or UVB (Fig 8b) In theseexperiments we used s-NFkB and s-IKKa as controls and con-firmed that they had no effect in this pathway (Fig 8b) Fur-thermore as-NIK targeting a kinase functioning upstream fromIKK also partially inhibited the effects of TNFa Therefore
the antisense oligonucleotide experiments confirm our conclu-sions that the activation of NFkB by TNFa depends on the IKKpathway whereas the response to UVC or UVB occurs througha mechanism independent of IKKs
Furthermore as an additional control we used oligonu-cleotides targeting the JNK pathway as-JNKK1 and s-JNKK1Although they affected the JNK pathway (see Figs 4 and 6)they did not affect the NFkB pathway (Fig 8b) which indi-cates that the activation of the JNK pathway and the NFkBpathway are independent in keratinocytes This means that epi-dermal keratinocytes contain multiple specific UV responsivemechanisms
DISCUSSION
Our data suggest that UVBUVC illumination in epidermalkeratinocytes is not a generic stress response inducer but a spe-cific extracellular agent that activates specific signal transduc-tion pathways In particular we show that in keratinocytes UVexclusively activates the JNK pathway while in HeLa cellsUV also activates the parallel ERK and p38 pathways Con-versely in keratinocytes exclusively UV activates the JNKpathway while in HeLa cells growth factors can do so as wellIn parallel in keratinocytes UV uses a distinct pathway to ac-tivate the NFkB transcription factor a pathway separate fromthe more common TNFa activated one (Fig 9)
The specific activation of JNK by UVBUVC is particularlyrelevant because the JNK pathway is in different cell types atarget of multiple stress agents The balance between apoptosisand proliferation is at times regulated by JNK and the tran-scription factors activated by JNK regulate the expression ofmany genes (Karin and Hunter 1995) It is uniquely importantin epidermis that JNK exclusively responds to UVBUVC be-cause epidermis is the natural target of UVBUVC and the JNK-
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 673
FIG 6 Activation of JNK in vivo and the effect of as-JNKK1 In the control sample (left) JNK is not activated as shown us-ing an antibody specific for the activated phosphorylated form of JNK Treatment with 100 mJcm2 causes strong activation ofJNK (middle) Pretreatment for 3 h with the antisense oligonucleotide targeting JNKK1 abolishes this activation (right)
FIG 7 TNFa UVB and UVC activate NFkB while UVAdoes not Reporter plasmid NFkB3-CAT (top) which containsthree tandem NFkB sites upstream from the TK promoter wastransfected into keratinocytes The transfected cells were eithertreated with 50 ngml of TNFa or illuminated with indicateddoses of UV harvested 16 h later and the CAT levels deter-mined Untreated cells served as a control The replicates dif-fered by 25 or less
ADACHI ET AL674
FIG 8 Specific inhibitors can differentiate between the TNFa-and UVB-dependent signal transduction pathways (a) The effectsof specific inhibitors on NFkB activation Cultured keratinocyteswere treated with either UVB (8 mJcm2) or TNFa (50 ngml) af-ter transfection of the NFkB response element Inhibitor Bay11-7085 specifically inhibits upstream kinases of IKK aspirin inhibitsIKKb parthenolide blocks IkBa degradation (b) the effects of an-tisense oligonucleotides targeting signaling cascades on NFkB ac-tivation in keratinocytes Oligonucleotide as-NFkB targeting thetranscription factor blocks the activation by UVB and UVC whileoligonucleotides targeting IKKb JNK1 and JNKK1 are withouteffect (top two panels) In contrast oligonucleotides targetingIKKb and NIK partially or completely block the effects of TNFa(bottom panel) Note that the sense oligonucleotides (s-NFkB s-IKKb) used as controls had no effect on NFkB activation (c)Schematics of NFkB pathways activated by UV light and TNFaIn keratinocytes the two extracellular stimuli work through inde-pendent pathways that merge at the NFkBIkB complex
regulated transcription factors mediate both photodamage andits alleviation (Fisher et al 1997)
In keratinocytes the EGF and UV responsive pathways donot overlap but UV specifically activates the JNK pathwayThis suggests that the epidermal keratinocytes exhibit distinc-tive specialized UV-responsive signal transduction pathwaysIndeed when we compared the responses to UV in several celltypes we found that the response in keratinocytes is signifi-cantly more stringently controlled than in other cell types Forexample HeLa cells demonstrated a lack of specificity Usinga specific MEK-12 inhibitor we again demonstrated relaxedstringency in HeLa cells We conclude from these results thatkeratinocytes being often exposed to ambient UV have de-veloped more stringent and specific responses to UV than othercell types which are not commonly exposed to UV
At present we do not understand the relevance of JNK3 ex-pression in keratinocytes This isoform it was reported isspecifically expressed in the brain (Yoshida et al 2001) Wespeculate that perhaps the sequestration of JNK1 by the speci-ficity of the UV response creates a need for JNK3 as the moregeneral less restrictive stress signal transducer
TNFa one of proinflammatory cytokines important in cuta-neous inflammation activates NFkB via IKKs kinases that canbe activated by MEKK or NIK (Hirano et al 1996 Lee et al1997) In HeLa cells the phosphorylation of IkBa at Ser-32and Ser-36 residues is essential for the IKK mediated IkBa
degradation caused by TNFa IL-1 or ionizing radiation Us-ing specific inhibitors and antisense oligonucleotides we foundthat the NFkB activation by UVBUVC does not involve IKKs
The Ser-32 and Ser-36 residues are not involved in the UV-induced IkBa degradation (Bender et al 1997 N Li and Karin1998) Because IKKb specifically phosphorylates IkB at bothSer-32 and Ser-36 of IkBa these facts support our results Re-cently it was suggested that UV activation of NFkB in HeLacells may involve the RasndashRaf pathway although not in ker-atinocytes (Tobin et al 1996) This leads to the question howUV activates NFkB in keratinocytes Two new kinases IKKlaquo
and pp90rsk have been found to phosphorylate IkBa (Ghodaet al 1997 Schouten et al 1997 Shimada et al 1999 Israel2000 Peters et al 2000) However IKKlaquo phosphorylates theSer-36 residue while pp90rsk phosphorylates the Ser-32residue only Therefore these kinases are probably not involvedin the UV activated pathway Amino acids between IkBa po-sition 277 and 297 (the PEST region) are apparently requiredfor the UV induced degradation of IkB (Ito et al 1995 Ben-der et al 1998) but the signaling events that effect this degra-dation in keratinocytes are still unknown The data suggest thatTNFa and UVC use different pathways to activate NFkB inHeLa cells Specifically while both UVC and TNFa activateNFkB the TNFa signal goes via IKKs but the UVC signaldoes not From these data it appears that the convergence ofthe UVB and TNFa signals occurs downstream from IKKs thatis at the modification of IkB Overall these results imply thatthe epidermis a natural target of UV light has an exceptionalmolecular UV defense system distinct from UV responsive sys-tems in other tissues
The responses to the three different wavelengths of UV areespecially strictly controlled in keratinocytes Although allwavelengths cause DNA damage and a release of cytokines wefound that the inherent molecular responses to UVA are com-pletely different from the responses to UVBUVC For exam-ple UVBUVC but not UVA activate JNK and NFkB in ker-atinocytes Furthermore UVA activates the JNK pathway inHeLa cells but not in keratinocytes while UVBUVC activatethis pathway in both cells (Rosette and Karin 1996) Similarlyin both HeLa cells and cultured keratinocytes UVBUVC ac-tivate NFkB via a kinase different from IKKs while UVA doesnot activate NFkB at all We conclude that UVBUVC activatea completely different system of signal transduction from UVA
The results suggesting specific signal transduction pathwaysfor UVBUVC in keratinocytes presented here agree well withour studies of UVB-regulated genes using DNA micro arrays(Li et al 2001) We found that in response to UVB ker-atinocytes not only induce the immediate early genes and stressresponse genes but also elaborate responses designed to pro-tect the organism underneath For example chemokines of theIL-8 family are induced which alert the surrounding tissue tothe UV damage synthesis of the cornified envelope compo-nents is enhanced which thickens the stratum corneum etc Weconclude that human epidermal keratinocytes being naturallyexposed to UV light have elaborated highly specific molecu-lar responses to UVBUVC at the signal transduction level aswell as the gene expression level
ACKNOWLEDGMENTS
Our research was supported by the National Institutes ofHealth Grants AR30682 AR39176 AR40522 and AR41850
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 675
FIG 9 Specificity in stress response Epidermal ker-atinocytes exhibit specialized UV-responsive signal transduc-tion pathways which apparently do not network and intersectwith other pathways such as those responsive to EGF or TNFaIn contrast in other cell types such as HeLa multiple path-ways crossover interact and overlap
and by the Anaderm Research Corporation Special thinks goto Dr M Simon for the generous gift of keratinocytes We alsothank members of our laboratory for advice reagents and en-couragement
REFERENCES
ALKALAY I YARON A HATZUBAI A ORIAN ACIECHANOVER A and BEN-NERIAH Y (1995) Stimulation-dependent I kappa B alpha phosphorylation marks the NF-kappa Binhibitor for degradation via the ubiquitin-proteasome pathway ProcNatl Acad Sci USA 92 10599ndash10603
BAEUERLE PA and HENKEL T (1994) Function and activationof NF-kappa B in the immune system Annu Rev Immunol 12141ndash179
BARNES PJ and KARIN M (1997) Nuclear factor-kappaB A piv-otal transcription factor in chronic inflammatory diseases N EnglJ Med 336 1066ndash1071
BEG AA FINCO TS NANTERMET PV and BALDWIN AJR (1993) Tumor necrosis factor and interleukin-1 lead to phos-phorylation and loss of I kappa B alpha A mechanism for NF-kappaB activation Mol Cell Biol 13 3301ndash3310
BENDER K BLATTNER C KNEBEL A IORDANOV M HER-RLICH P and RAHMSDORF HJ (1997) UV-induced signaltransduction J Photochem Photobiol B 37 1ndash17
BENDER K GOTTLICHER M WHITESIDE S RAHMSDORFHJ and HERRLICH P (1998) Sequential DNA damage-indepen-dent and -dependent activation of NF-kappaB by UV EMBO J 175170ndash5181
BROWN K GERSTBERGER S CARLSON L FRANZOSO Gand SIEBENLIST U (1995) Control of I kappa B-alpha proteoly-sis by site-specific signal-induced phosphorylation Science 2671485ndash1488
DELHASE M HAYAKAWA M CHEN Y and KARIN M(1999) Positive and negative regulation of IkappaB kinase activitythrough IKKbeta subunit phosphorylation Science 284 309ndash313
DEacuteRIJARD B et al (1994) JNK1 A protein kinase stimulated byUV light and Ha-Ras that binds and phosphorylates the c-Jun acti-vation domain Cell 76 1025ndash1037
DEVARY Y ROSETTE C DIDONATO JA and KARIN M(1993) NF-kappa B activation by ultraviolet light not dependent ona nuclear signal Science 261 1442ndash1445
DIDONATO JA HAYAKAWA M ROTHWARF DM ZANDIE and KARIN M (1997) A cytokine-responsive IkappaB kinasethat activates the transcription factor NF-kappaB Nature 388548ndash554
FANGER GR GERWINS P WIDMANN C JARPE MB andJOHNSON GL (1997) MEKKs GCKs MLKs PAKs TAKs andtpls Upstream regulators of the c-Jun amino-terminal kinases CurrOpin Genet Dev 7 67ndash74
FEIGE U MORIMOTO RI YAHARA I and POLLA BS eds(1996) Stress-Inducible Cellular Responses (Birkhauser VerlagBasel Switzerland)
FISHER GJ WANG ZQ DATTA SC VARANI J KANG Sand VOORHEES JJ (1997) Pathophysiology of premature skin ag-ing induced by ultraviolet light N Engl J Med 337 1419ndash1428
GHODA L LIN X and GREENE WC (1997) The 90-kDa ribo-somal S6 kinase (pp90rsk) phosphorylates the N-terminal regulatorydomain of IkappaBalpha and stimulates its degradation in vitro JBiol Chem 272 21281ndash21288
GILCHREST B (1995) Photodamage (Blackwell Scientific NewYork)
GRETHER-BECK S BUETTNER R and KRUTMANN J (1997)Ultraviolet A radiation-induced expression of human genes Molec-ular and photobiological mechanisms Biol Chem 378 1231ndash1236
GRILLI M PIZZI M MEMO M and SPANO P (1996) Neuro-protection by aspirin and sodium salicylate through blockade of NF-kappaB activation Science 274 1383ndash1385
GUPTA S et al (1996) Selective interaction of JNK protein kinaseisoforms with transcription factors EMBO J 15 2760ndash2770
HEHNER SP et al (1998) Sesquiterpene lactones specifically in-hibit activation of NF-kappa B by preventing the degradation of Ikappa B-alpha and I kappa B-beta J Biol Chem 273 1288ndash1297
HERSKOWITZ I (1995) MAP kinase pathways in yeast For matingand more Cell 80 187ndash197
HIBI M LIN A SMEAL T MINDEN A and KARIN M (1993)Identification of an oncoprotein- and UV-responsive protein kinasethat binds and potentiates the c-Jun activation domain Genes Dev7 2135ndash2148
HIRANO M et al (1996) MEK kinase is involved in tumor necro-sis factor alpha-induced NF-kappaB activation and degradation ofIkappaB-alpha J Biol Chem 271 13234ndash13238
HU Y et al (1999) Abnormal morphogenesis but intact IKK activa-tion in mice lacking the IKKalpha subunit of IkappaB kinase Sci-ence 284 316ndash320
IORDANOV M et al (1997) CREB is activated by UVC through ap38HOG-1-dependent protein kinase EMBO J 16 1009ndash1022
ISREAL A (2000) The IKK complex An integrator of all signals thatactivate NF-kappaB Trends Cell Biol 10 129ndash133
ITO CY ADEY N BAUTCH VL and BALDWIN AS (1995)Structure and evolution of the human IKBA gene Genomics 29490ndash495
JIANG CK CONNOLLY D and BLUMENBERG M (1991)Comparison of methods for transfection of human epidermal ker-atinocytes J Invest Dermatol 97 969ndash973
JIANG CK FLANAGAN S OHTSUKI M SHUAI K FREED-BERG IM and BLUMENBERG M (1994) Disease-activatedtranscription factor Allergic reactions in human skin cause nucleartranscription of STAT-91 and induce synthesis of keratin K17 MolCell Biol 14 4759ndash4769
JIANG CK MAGNALDO T OHTSUKI M FREEDBERG IMBERNERD F and BLUMENBERG M (1993) Epidermal growthfactor and transforming growth factor alpha specifically induce theactivation- and hyperproliferation-associated keratins 6 and 16 ProcNatl Acad Sci USA 90 6786ndash6790
KALLUNKI T DENG T HIBI M and KARIN M (1996) c-Juncan recruit JNK to phosphorylate dimerization partners via specificdocking interactions Cell 87 929ndash939
KARIN M (1995) The regulation of AP-1 activity by mitogen-acti-vated protein kinases J Biol Chem 270 16483ndash16486
KARIN M and HUNTER T (1995) Transcriptional control by pro-tein phosphorylation Signal transmission from the cell surface to thenucleus Curr Biol 5 747ndash757
KLAMPFER L LEE TH HSU W VILCEK J and CHEN-KIANG S (1994) NF-IL6 and AP-1 cooperatively modulate the ac-tivation of the TSG-6 gene by tumor necrosis factor alpha and in-terleukin-1 Mol Cell Biol 14 6561ndash6569
KOMINE M et al (2000) Inflammatory versus proliferative pro-cesses in epidermis Tumor necrosis factor alpha induces K6b ker-atin synthesis through a transcriptional complex containing NFkappaB and CEBPbeta J Biol Chem 275 32077ndash32088
KOPP E and GHOSH S (1994) Inhibition of NF-kappa B by so-dium salicylate and aspirin Science 265 956ndash959
KYRIAKIS JM et al (1994) The stress-activated protein kinase sub-family of c-Jun kinases Nature 369 156ndash160
LEE FS HAGLER J CHEN ZJ and MANIATIS T (1997) Ac-tivation of the IkappaB alpha kinase complex by MEKK1 a kinaseof the JNK pathway Cell 88 213ndash222
LEPPA S and BOHMANN D (1999) Diverse functions of JNK sig-naling and c-Jun in stress response and apoptosis Oncogene 186158ndash6162
ADACHI ET AL676
LI D TURI TG SCHUCK A FREEDBERG IM KHITROVG and BLUMENBERG M (2001) Rays and arrays The tran-scriptional program in the response of human epidermal keratinocytesto UVB illumination FASEB J 15 2533ndash2535
LI N and KARIN M (1998) Ionizing radiation and short wavelengthUV activate NF-kappaB through two distinct mechanisms ProcNatl Acad Sci USA 95 13012ndash13017
LIU ZG HSU H GOEDDEL DV and KARIN M (1996) Dis-section of TNF receptor 1 effector functions JNK activation is notlinked to apoptosis while NF-kappaB activation prevents cell deathCell 87 565ndash576
MERCURIO F and MANNING AM (1999) NF-kappaB as a pri-mary regulator of the stress response Oncogene 18 6163ndash6171
MIYAMOTO S and VERMA IM (1995) RelNF-kappa BI kappaB story Adv Cancer Res 66 255ndash292
NESTLE FO MITRA RS BENNETT CF CHAN H andNICKOLOFF BJ (1994) Cationic lipid is not required for uptakeand selective inhibitory activity of ICAM-1 phosphorothioate anti-sense oligonucleotides in keratinocytes J Invest Dermatol 103569ndash575
PETERS RT LIAO SM and MANIATIS T (2000) IKKepsilonis part of a novel PMA-inducible IkappaB kinase complex Mol Cell5 513ndash522
PIERCE JW READ MA DING H LUSCINSKAS FW andCOLLINS T (1996) Salicylates inhibit I kappa B-alpha phos-phorylation endothelial-leukocyte adhesion molecule expressionand neutrophil transmigration J Immunol 156 3961ndash3969
PIERCE JW et al (1997) Novel inhibitors of cytokine-induced Ikap-paBalpha phosphorylation and endothelial cell adhesion molecule ex-pression show anti-inflammatory effects in vivo J Biol Chem 27221096ndash21103
REUNING U et al (1995) Inhibition of NF-kappa B-Rel A expres-sion by antisense oligodeoxynucleotides suppresses synthesis ofurokinase-type plasminogen activator (uPA) but not its inhibitor PAI-1 Nucleic Acids Res 23 3887ndash3893
ROSETTE C and KARIN M (1996) Ultraviolet light and osmoticstress activation of the JNK cascade through multiple growth factorand cytokine receptors Science 274 1194ndash1197
SCHOUTEN GJ et al (1997) IkappaB alpha is a target for the mitogen-activated 90 kDa ribosomal S6 kinase EMBO J 163133ndash3144
SHIMADA T et al (1999) IKK-i a novel lipopolysaccharide-inducible kinase that is related to IkappaB kinases Int Immunol 111357ndash1362
SIMON M and GREEN H (1984) Participation of membrane-asso-ciated proteins in the formation of the cross-linked envelope of thekeratinocyte Cell 36 827ndash834
SIMON MM ARAGANE Y SCHWARZ A LUGER TA andSCHWARZ T (1994) UVB light induces nuclear factor kappa B(NF kappa B) activity independently from chromosomal DNA dam-age in cell-free cytosolic extracts J Invest Dermatol 102 422ndash427
TOBIN D NILSSON M and TOFTGARD R (1996) Ras-inde-pendent activation of Rel-family transcription factors by UVB andTPA in cultured keratinocytes Oncogene 12 785ndash793
TOMIC-CANIC M KOMINE M FREEDBERG IM and BLU-MENBERG M (1998) Epidermal signal transduction and tran-scription factor activation in activated keratinocytes J Dermatol Sci17 167ndash181
WILLIAMS BR (1999) PKR A sentinel kinase for cellular stressOncogene 18 6112ndash6120
WU Z WU J JACINTO E and KARIN M (1997) Molecularcloning and characterization of human JNKK2 a novel Jun NH2-terminal kinase-specific kinase Mol Cell Biol 17 7407ndash7416
YANG DD et al (1997) Absence of excitotoxicity-induced apopto-sis in the hippocampus of mice lacking the Jnk3 gene Nature 389865ndash870
YANKULOV K BLAU J PURTON T ROBERTS S and BENT-LEY DL (1994) Transcriptional elongation by RNA polymeraseII is stimulated by transactivators Cell 77 749ndash759
YIN MJ YAMAMOTO Y and GAYNOR RB (1998) The anti-inflammatory agents aspirin and salicylate inhibit the activity ofI(kappa)B kinase-beta Nature 396 77ndash80
YOSHIDA S et al (2001) The c-Jun NH2-terminal kinase3 (JNK3)gene Genomic structure chromosomal assignment and loss of ex-pression in brain tumors J Hum Genet 46 182ndash187
Address reprint requests toM Blumenberg PhD
The NYU Cancer InstituteNew York University School of Medicine
550 First AveNew York NY 10016
E-mail blumem01mednyuedu
Received for publication June 5 2003 received in revised formJuly 15 2003 accepted August 14 2003
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 677

While there are only two JNKKs kinases that directly phos-phorylate JNK there is a large family of JNKKKs kinases thatphosphorylate JNKKs In a manuscript in preparation we de-scribe our analysis of the kinases expressed by keratinocytes
To demonstrate that the JNK system is responsive in vivowe used a novel ex vivo approach Biopsies of human skin weretreated with as-JNKK1 oligonucleotide or left untreated as con-trol Then both the treated and untreated biopsies were illumi-nated with UVC and shortly thereafter processed for immuno-histochemical detection of activated JNK (Fig 6) We found inthese biopsies that UV caused activation and nuclear translo-cation of JNK Furthermore this activation was abrogated bypretreatment with as-JNKK1 the antisense oligonucleotide tar-geting JNKK1 again demonstrating that JNKK1 is essential foractivation of JNK by UV in epidermis These results mean that
the specific signal transduction pathways we found operationalin cultured keratinocytes are also functional and working thesame way in human skin in vivo
UVB signal transduction pathway part II activationof the NFkB pathway
Having established that keratinocytes possess a specific UV-responding pathway to activate JNK we wanted to examine thepossibility that the UV-specific signal transduction pathwaysactivating NFkB are also specific in these cells The canonicalactivator of NFkB is TNFa however recent evidence suggeststhat UV and TNFa may activate NFkB through different mech-anisms (N Li and Karin 1998) Demonstrating a UV-specificpathway would strengthen even more our hypothesis that thereis specificity in UV responsive signal transduction in humanepidermal keratinocytes Therefore we transfected into ker-atinocytes and into HeLa cells a reporter plasmid containingthree tandem NFkB sites linked to the TK promoter-drivenCAT We found that the exposure of transfected cells to TNFaUVB or UVC resulted in the induction of the NFkB responseelement (Fig 7) Generally at the optimal conditions the in-duction of NFkB by UV was weaker than the induction byTNFa On the other hand NFkB was not activated by UVA atall even at high doses (Fig 7)
To compare the mechanisms of NFkB activation by UV andTNFa we have used a battery of specific inhibitors of the NFkBpathway which inhibit the pathway at different points (Fig 8a)Namely Bay11-7085 inhibits upstream from IKK (Pierce et al1997) aspirin has recently been found to be a competitive in-hibitor of ATP binding to IKKb (Yin et al 1998) whereasparthenolide directly inhibits the degradation of IkBa (Pierceet al 1997) When we treated transfected keratinocytes or HeLacells with these three inhibitors we found that Bay11-7085 in-hibited the induction of NFkB by TNFa in a dose-dependentmanner while it has almost no effect on the induction of NFkBby UVC (Fig 8b) The same results were obtained by using as-pirin (Fig 8c) In contrast parthenolide inhibited both the UVCand the TNFa effects in parallel (Fig 8d) These results con-firm that TNFa and UVC use different pathways to activateNFkB in HeLa cells Specifically while both UVC and TNFaactivate NFkB the TNFa signal goes via IKKs but the UVCsignal does not From these data we conclude that the conver-gence of the UVB and TNFa signals occurs downstream fromIKKs that is at the modification of IkB
One caveat in the use of inhibitors of specific kinases is thepossibility that such inhibitors may affect additional pathwaysnot currently recognized as affected by the given inhibitor Toaddress such specificity concerns and to examine the involve-ment of IKK in the induction of NFkB by TNFa UVC andUVB we used antisense oligonucleotides specifically targetingNFkB and IKKb as-NFkB as-IKKb (Table 1) (Reuning etal 1995 Komine et al 2000) In parallel we added the senseoligonucleotides s-NFkB s-IKKb as controls The antisenseexperiments confirmed the fact that UVBUVC use a clearlydifferent pathway to activate NFkB than TNFa does The sup-pression of NFkB synthesis in cultured keratinocytes by addingas-NFkB resulted in inhibition of activation of NFkB by allthree agents UVC UVB and TNFa as expected (Fig 8b) Onthe other hand the suppression of IKKb synthesis by adding
ADACHI ET AL672
FIG 5 Specific activation of JNK in keratinocytes andpromiscuous activation of JNK in HeLa cells (a) JNK activityin keratinocytes and HeLa cells treated with UVC UVA heatshock or EGF Duplicate experiments were performed with allagents except EGF Each lane contains 10 mg of protein InHeLa cells (bottom panel) UVC and EGF strongly while UVAand heat detectably activate JNK Cells were harvested 30 minafter treatment (b) Lack of activation of ERK by UVB in ker-atinocytes The top panel shows a Western blot with the anti-body specific for the activated ERKs the bottom for totalERK1 as a control for loading and transfer (c) Specificity insignal transduction In keratinocytes UVB specifically acti-vates JNK and EGF specifically activates ERKs whereas inHeLa cells all three extracellular signals UVB EGF and heatactivate both JNK and ERK Fibroblasts respond similar toHeLa cells (data not shown)
as-IKKb only inhibited the activation by TNFa but had no ef-fect on the activation by either UVC or UVB (Fig 8b) In theseexperiments we used s-NFkB and s-IKKa as controls and con-firmed that they had no effect in this pathway (Fig 8b) Fur-thermore as-NIK targeting a kinase functioning upstream fromIKK also partially inhibited the effects of TNFa Therefore
the antisense oligonucleotide experiments confirm our conclu-sions that the activation of NFkB by TNFa depends on the IKKpathway whereas the response to UVC or UVB occurs througha mechanism independent of IKKs
Furthermore as an additional control we used oligonu-cleotides targeting the JNK pathway as-JNKK1 and s-JNKK1Although they affected the JNK pathway (see Figs 4 and 6)they did not affect the NFkB pathway (Fig 8b) which indi-cates that the activation of the JNK pathway and the NFkBpathway are independent in keratinocytes This means that epi-dermal keratinocytes contain multiple specific UV responsivemechanisms
DISCUSSION
Our data suggest that UVBUVC illumination in epidermalkeratinocytes is not a generic stress response inducer but a spe-cific extracellular agent that activates specific signal transduc-tion pathways In particular we show that in keratinocytes UVexclusively activates the JNK pathway while in HeLa cellsUV also activates the parallel ERK and p38 pathways Con-versely in keratinocytes exclusively UV activates the JNKpathway while in HeLa cells growth factors can do so as wellIn parallel in keratinocytes UV uses a distinct pathway to ac-tivate the NFkB transcription factor a pathway separate fromthe more common TNFa activated one (Fig 9)
The specific activation of JNK by UVBUVC is particularlyrelevant because the JNK pathway is in different cell types atarget of multiple stress agents The balance between apoptosisand proliferation is at times regulated by JNK and the tran-scription factors activated by JNK regulate the expression ofmany genes (Karin and Hunter 1995) It is uniquely importantin epidermis that JNK exclusively responds to UVBUVC be-cause epidermis is the natural target of UVBUVC and the JNK-
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 673
FIG 6 Activation of JNK in vivo and the effect of as-JNKK1 In the control sample (left) JNK is not activated as shown us-ing an antibody specific for the activated phosphorylated form of JNK Treatment with 100 mJcm2 causes strong activation ofJNK (middle) Pretreatment for 3 h with the antisense oligonucleotide targeting JNKK1 abolishes this activation (right)
FIG 7 TNFa UVB and UVC activate NFkB while UVAdoes not Reporter plasmid NFkB3-CAT (top) which containsthree tandem NFkB sites upstream from the TK promoter wastransfected into keratinocytes The transfected cells were eithertreated with 50 ngml of TNFa or illuminated with indicateddoses of UV harvested 16 h later and the CAT levels deter-mined Untreated cells served as a control The replicates dif-fered by 25 or less
ADACHI ET AL674
FIG 8 Specific inhibitors can differentiate between the TNFa-and UVB-dependent signal transduction pathways (a) The effectsof specific inhibitors on NFkB activation Cultured keratinocyteswere treated with either UVB (8 mJcm2) or TNFa (50 ngml) af-ter transfection of the NFkB response element Inhibitor Bay11-7085 specifically inhibits upstream kinases of IKK aspirin inhibitsIKKb parthenolide blocks IkBa degradation (b) the effects of an-tisense oligonucleotides targeting signaling cascades on NFkB ac-tivation in keratinocytes Oligonucleotide as-NFkB targeting thetranscription factor blocks the activation by UVB and UVC whileoligonucleotides targeting IKKb JNK1 and JNKK1 are withouteffect (top two panels) In contrast oligonucleotides targetingIKKb and NIK partially or completely block the effects of TNFa(bottom panel) Note that the sense oligonucleotides (s-NFkB s-IKKb) used as controls had no effect on NFkB activation (c)Schematics of NFkB pathways activated by UV light and TNFaIn keratinocytes the two extracellular stimuli work through inde-pendent pathways that merge at the NFkBIkB complex
regulated transcription factors mediate both photodamage andits alleviation (Fisher et al 1997)
In keratinocytes the EGF and UV responsive pathways donot overlap but UV specifically activates the JNK pathwayThis suggests that the epidermal keratinocytes exhibit distinc-tive specialized UV-responsive signal transduction pathwaysIndeed when we compared the responses to UV in several celltypes we found that the response in keratinocytes is signifi-cantly more stringently controlled than in other cell types Forexample HeLa cells demonstrated a lack of specificity Usinga specific MEK-12 inhibitor we again demonstrated relaxedstringency in HeLa cells We conclude from these results thatkeratinocytes being often exposed to ambient UV have de-veloped more stringent and specific responses to UV than othercell types which are not commonly exposed to UV
At present we do not understand the relevance of JNK3 ex-pression in keratinocytes This isoform it was reported isspecifically expressed in the brain (Yoshida et al 2001) Wespeculate that perhaps the sequestration of JNK1 by the speci-ficity of the UV response creates a need for JNK3 as the moregeneral less restrictive stress signal transducer
TNFa one of proinflammatory cytokines important in cuta-neous inflammation activates NFkB via IKKs kinases that canbe activated by MEKK or NIK (Hirano et al 1996 Lee et al1997) In HeLa cells the phosphorylation of IkBa at Ser-32and Ser-36 residues is essential for the IKK mediated IkBa
degradation caused by TNFa IL-1 or ionizing radiation Us-ing specific inhibitors and antisense oligonucleotides we foundthat the NFkB activation by UVBUVC does not involve IKKs
The Ser-32 and Ser-36 residues are not involved in the UV-induced IkBa degradation (Bender et al 1997 N Li and Karin1998) Because IKKb specifically phosphorylates IkB at bothSer-32 and Ser-36 of IkBa these facts support our results Re-cently it was suggested that UV activation of NFkB in HeLacells may involve the RasndashRaf pathway although not in ker-atinocytes (Tobin et al 1996) This leads to the question howUV activates NFkB in keratinocytes Two new kinases IKKlaquo
and pp90rsk have been found to phosphorylate IkBa (Ghodaet al 1997 Schouten et al 1997 Shimada et al 1999 Israel2000 Peters et al 2000) However IKKlaquo phosphorylates theSer-36 residue while pp90rsk phosphorylates the Ser-32residue only Therefore these kinases are probably not involvedin the UV activated pathway Amino acids between IkBa po-sition 277 and 297 (the PEST region) are apparently requiredfor the UV induced degradation of IkB (Ito et al 1995 Ben-der et al 1998) but the signaling events that effect this degra-dation in keratinocytes are still unknown The data suggest thatTNFa and UVC use different pathways to activate NFkB inHeLa cells Specifically while both UVC and TNFa activateNFkB the TNFa signal goes via IKKs but the UVC signaldoes not From these data it appears that the convergence ofthe UVB and TNFa signals occurs downstream from IKKs thatis at the modification of IkB Overall these results imply thatthe epidermis a natural target of UV light has an exceptionalmolecular UV defense system distinct from UV responsive sys-tems in other tissues
The responses to the three different wavelengths of UV areespecially strictly controlled in keratinocytes Although allwavelengths cause DNA damage and a release of cytokines wefound that the inherent molecular responses to UVA are com-pletely different from the responses to UVBUVC For exam-ple UVBUVC but not UVA activate JNK and NFkB in ker-atinocytes Furthermore UVA activates the JNK pathway inHeLa cells but not in keratinocytes while UVBUVC activatethis pathway in both cells (Rosette and Karin 1996) Similarlyin both HeLa cells and cultured keratinocytes UVBUVC ac-tivate NFkB via a kinase different from IKKs while UVA doesnot activate NFkB at all We conclude that UVBUVC activatea completely different system of signal transduction from UVA
The results suggesting specific signal transduction pathwaysfor UVBUVC in keratinocytes presented here agree well withour studies of UVB-regulated genes using DNA micro arrays(Li et al 2001) We found that in response to UVB ker-atinocytes not only induce the immediate early genes and stressresponse genes but also elaborate responses designed to pro-tect the organism underneath For example chemokines of theIL-8 family are induced which alert the surrounding tissue tothe UV damage synthesis of the cornified envelope compo-nents is enhanced which thickens the stratum corneum etc Weconclude that human epidermal keratinocytes being naturallyexposed to UV light have elaborated highly specific molecu-lar responses to UVBUVC at the signal transduction level aswell as the gene expression level
ACKNOWLEDGMENTS
Our research was supported by the National Institutes ofHealth Grants AR30682 AR39176 AR40522 and AR41850
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 675
FIG 9 Specificity in stress response Epidermal ker-atinocytes exhibit specialized UV-responsive signal transduc-tion pathways which apparently do not network and intersectwith other pathways such as those responsive to EGF or TNFaIn contrast in other cell types such as HeLa multiple path-ways crossover interact and overlap
and by the Anaderm Research Corporation Special thinks goto Dr M Simon for the generous gift of keratinocytes We alsothank members of our laboratory for advice reagents and en-couragement
REFERENCES
ALKALAY I YARON A HATZUBAI A ORIAN ACIECHANOVER A and BEN-NERIAH Y (1995) Stimulation-dependent I kappa B alpha phosphorylation marks the NF-kappa Binhibitor for degradation via the ubiquitin-proteasome pathway ProcNatl Acad Sci USA 92 10599ndash10603
BAEUERLE PA and HENKEL T (1994) Function and activationof NF-kappa B in the immune system Annu Rev Immunol 12141ndash179
BARNES PJ and KARIN M (1997) Nuclear factor-kappaB A piv-otal transcription factor in chronic inflammatory diseases N EnglJ Med 336 1066ndash1071
BEG AA FINCO TS NANTERMET PV and BALDWIN AJR (1993) Tumor necrosis factor and interleukin-1 lead to phos-phorylation and loss of I kappa B alpha A mechanism for NF-kappaB activation Mol Cell Biol 13 3301ndash3310
BENDER K BLATTNER C KNEBEL A IORDANOV M HER-RLICH P and RAHMSDORF HJ (1997) UV-induced signaltransduction J Photochem Photobiol B 37 1ndash17
BENDER K GOTTLICHER M WHITESIDE S RAHMSDORFHJ and HERRLICH P (1998) Sequential DNA damage-indepen-dent and -dependent activation of NF-kappaB by UV EMBO J 175170ndash5181
BROWN K GERSTBERGER S CARLSON L FRANZOSO Gand SIEBENLIST U (1995) Control of I kappa B-alpha proteoly-sis by site-specific signal-induced phosphorylation Science 2671485ndash1488
DELHASE M HAYAKAWA M CHEN Y and KARIN M(1999) Positive and negative regulation of IkappaB kinase activitythrough IKKbeta subunit phosphorylation Science 284 309ndash313
DEacuteRIJARD B et al (1994) JNK1 A protein kinase stimulated byUV light and Ha-Ras that binds and phosphorylates the c-Jun acti-vation domain Cell 76 1025ndash1037
DEVARY Y ROSETTE C DIDONATO JA and KARIN M(1993) NF-kappa B activation by ultraviolet light not dependent ona nuclear signal Science 261 1442ndash1445
DIDONATO JA HAYAKAWA M ROTHWARF DM ZANDIE and KARIN M (1997) A cytokine-responsive IkappaB kinasethat activates the transcription factor NF-kappaB Nature 388548ndash554
FANGER GR GERWINS P WIDMANN C JARPE MB andJOHNSON GL (1997) MEKKs GCKs MLKs PAKs TAKs andtpls Upstream regulators of the c-Jun amino-terminal kinases CurrOpin Genet Dev 7 67ndash74
FEIGE U MORIMOTO RI YAHARA I and POLLA BS eds(1996) Stress-Inducible Cellular Responses (Birkhauser VerlagBasel Switzerland)
FISHER GJ WANG ZQ DATTA SC VARANI J KANG Sand VOORHEES JJ (1997) Pathophysiology of premature skin ag-ing induced by ultraviolet light N Engl J Med 337 1419ndash1428
GHODA L LIN X and GREENE WC (1997) The 90-kDa ribo-somal S6 kinase (pp90rsk) phosphorylates the N-terminal regulatorydomain of IkappaBalpha and stimulates its degradation in vitro JBiol Chem 272 21281ndash21288
GILCHREST B (1995) Photodamage (Blackwell Scientific NewYork)
GRETHER-BECK S BUETTNER R and KRUTMANN J (1997)Ultraviolet A radiation-induced expression of human genes Molec-ular and photobiological mechanisms Biol Chem 378 1231ndash1236
GRILLI M PIZZI M MEMO M and SPANO P (1996) Neuro-protection by aspirin and sodium salicylate through blockade of NF-kappaB activation Science 274 1383ndash1385
GUPTA S et al (1996) Selective interaction of JNK protein kinaseisoforms with transcription factors EMBO J 15 2760ndash2770
HEHNER SP et al (1998) Sesquiterpene lactones specifically in-hibit activation of NF-kappa B by preventing the degradation of Ikappa B-alpha and I kappa B-beta J Biol Chem 273 1288ndash1297
HERSKOWITZ I (1995) MAP kinase pathways in yeast For matingand more Cell 80 187ndash197
HIBI M LIN A SMEAL T MINDEN A and KARIN M (1993)Identification of an oncoprotein- and UV-responsive protein kinasethat binds and potentiates the c-Jun activation domain Genes Dev7 2135ndash2148
HIRANO M et al (1996) MEK kinase is involved in tumor necro-sis factor alpha-induced NF-kappaB activation and degradation ofIkappaB-alpha J Biol Chem 271 13234ndash13238
HU Y et al (1999) Abnormal morphogenesis but intact IKK activa-tion in mice lacking the IKKalpha subunit of IkappaB kinase Sci-ence 284 316ndash320
IORDANOV M et al (1997) CREB is activated by UVC through ap38HOG-1-dependent protein kinase EMBO J 16 1009ndash1022
ISREAL A (2000) The IKK complex An integrator of all signals thatactivate NF-kappaB Trends Cell Biol 10 129ndash133
ITO CY ADEY N BAUTCH VL and BALDWIN AS (1995)Structure and evolution of the human IKBA gene Genomics 29490ndash495
JIANG CK CONNOLLY D and BLUMENBERG M (1991)Comparison of methods for transfection of human epidermal ker-atinocytes J Invest Dermatol 97 969ndash973
JIANG CK FLANAGAN S OHTSUKI M SHUAI K FREED-BERG IM and BLUMENBERG M (1994) Disease-activatedtranscription factor Allergic reactions in human skin cause nucleartranscription of STAT-91 and induce synthesis of keratin K17 MolCell Biol 14 4759ndash4769
JIANG CK MAGNALDO T OHTSUKI M FREEDBERG IMBERNERD F and BLUMENBERG M (1993) Epidermal growthfactor and transforming growth factor alpha specifically induce theactivation- and hyperproliferation-associated keratins 6 and 16 ProcNatl Acad Sci USA 90 6786ndash6790
KALLUNKI T DENG T HIBI M and KARIN M (1996) c-Juncan recruit JNK to phosphorylate dimerization partners via specificdocking interactions Cell 87 929ndash939
KARIN M (1995) The regulation of AP-1 activity by mitogen-acti-vated protein kinases J Biol Chem 270 16483ndash16486
KARIN M and HUNTER T (1995) Transcriptional control by pro-tein phosphorylation Signal transmission from the cell surface to thenucleus Curr Biol 5 747ndash757
KLAMPFER L LEE TH HSU W VILCEK J and CHEN-KIANG S (1994) NF-IL6 and AP-1 cooperatively modulate the ac-tivation of the TSG-6 gene by tumor necrosis factor alpha and in-terleukin-1 Mol Cell Biol 14 6561ndash6569
KOMINE M et al (2000) Inflammatory versus proliferative pro-cesses in epidermis Tumor necrosis factor alpha induces K6b ker-atin synthesis through a transcriptional complex containing NFkappaB and CEBPbeta J Biol Chem 275 32077ndash32088
KOPP E and GHOSH S (1994) Inhibition of NF-kappa B by so-dium salicylate and aspirin Science 265 956ndash959
KYRIAKIS JM et al (1994) The stress-activated protein kinase sub-family of c-Jun kinases Nature 369 156ndash160
LEE FS HAGLER J CHEN ZJ and MANIATIS T (1997) Ac-tivation of the IkappaB alpha kinase complex by MEKK1 a kinaseof the JNK pathway Cell 88 213ndash222
LEPPA S and BOHMANN D (1999) Diverse functions of JNK sig-naling and c-Jun in stress response and apoptosis Oncogene 186158ndash6162
ADACHI ET AL676
LI D TURI TG SCHUCK A FREEDBERG IM KHITROVG and BLUMENBERG M (2001) Rays and arrays The tran-scriptional program in the response of human epidermal keratinocytesto UVB illumination FASEB J 15 2533ndash2535
LI N and KARIN M (1998) Ionizing radiation and short wavelengthUV activate NF-kappaB through two distinct mechanisms ProcNatl Acad Sci USA 95 13012ndash13017
LIU ZG HSU H GOEDDEL DV and KARIN M (1996) Dis-section of TNF receptor 1 effector functions JNK activation is notlinked to apoptosis while NF-kappaB activation prevents cell deathCell 87 565ndash576
MERCURIO F and MANNING AM (1999) NF-kappaB as a pri-mary regulator of the stress response Oncogene 18 6163ndash6171
MIYAMOTO S and VERMA IM (1995) RelNF-kappa BI kappaB story Adv Cancer Res 66 255ndash292
NESTLE FO MITRA RS BENNETT CF CHAN H andNICKOLOFF BJ (1994) Cationic lipid is not required for uptakeand selective inhibitory activity of ICAM-1 phosphorothioate anti-sense oligonucleotides in keratinocytes J Invest Dermatol 103569ndash575
PETERS RT LIAO SM and MANIATIS T (2000) IKKepsilonis part of a novel PMA-inducible IkappaB kinase complex Mol Cell5 513ndash522
PIERCE JW READ MA DING H LUSCINSKAS FW andCOLLINS T (1996) Salicylates inhibit I kappa B-alpha phos-phorylation endothelial-leukocyte adhesion molecule expressionand neutrophil transmigration J Immunol 156 3961ndash3969
PIERCE JW et al (1997) Novel inhibitors of cytokine-induced Ikap-paBalpha phosphorylation and endothelial cell adhesion molecule ex-pression show anti-inflammatory effects in vivo J Biol Chem 27221096ndash21103
REUNING U et al (1995) Inhibition of NF-kappa B-Rel A expres-sion by antisense oligodeoxynucleotides suppresses synthesis ofurokinase-type plasminogen activator (uPA) but not its inhibitor PAI-1 Nucleic Acids Res 23 3887ndash3893
ROSETTE C and KARIN M (1996) Ultraviolet light and osmoticstress activation of the JNK cascade through multiple growth factorand cytokine receptors Science 274 1194ndash1197
SCHOUTEN GJ et al (1997) IkappaB alpha is a target for the mitogen-activated 90 kDa ribosomal S6 kinase EMBO J 163133ndash3144
SHIMADA T et al (1999) IKK-i a novel lipopolysaccharide-inducible kinase that is related to IkappaB kinases Int Immunol 111357ndash1362
SIMON M and GREEN H (1984) Participation of membrane-asso-ciated proteins in the formation of the cross-linked envelope of thekeratinocyte Cell 36 827ndash834
SIMON MM ARAGANE Y SCHWARZ A LUGER TA andSCHWARZ T (1994) UVB light induces nuclear factor kappa B(NF kappa B) activity independently from chromosomal DNA dam-age in cell-free cytosolic extracts J Invest Dermatol 102 422ndash427
TOBIN D NILSSON M and TOFTGARD R (1996) Ras-inde-pendent activation of Rel-family transcription factors by UVB andTPA in cultured keratinocytes Oncogene 12 785ndash793
TOMIC-CANIC M KOMINE M FREEDBERG IM and BLU-MENBERG M (1998) Epidermal signal transduction and tran-scription factor activation in activated keratinocytes J Dermatol Sci17 167ndash181
WILLIAMS BR (1999) PKR A sentinel kinase for cellular stressOncogene 18 6112ndash6120
WU Z WU J JACINTO E and KARIN M (1997) Molecularcloning and characterization of human JNKK2 a novel Jun NH2-terminal kinase-specific kinase Mol Cell Biol 17 7407ndash7416
YANG DD et al (1997) Absence of excitotoxicity-induced apopto-sis in the hippocampus of mice lacking the Jnk3 gene Nature 389865ndash870
YANKULOV K BLAU J PURTON T ROBERTS S and BENT-LEY DL (1994) Transcriptional elongation by RNA polymeraseII is stimulated by transactivators Cell 77 749ndash759
YIN MJ YAMAMOTO Y and GAYNOR RB (1998) The anti-inflammatory agents aspirin and salicylate inhibit the activity ofI(kappa)B kinase-beta Nature 396 77ndash80
YOSHIDA S et al (2001) The c-Jun NH2-terminal kinase3 (JNK3)gene Genomic structure chromosomal assignment and loss of ex-pression in brain tumors J Hum Genet 46 182ndash187
Address reprint requests toM Blumenberg PhD
The NYU Cancer InstituteNew York University School of Medicine
550 First AveNew York NY 10016
E-mail blumem01mednyuedu
Received for publication June 5 2003 received in revised formJuly 15 2003 accepted August 14 2003
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 677

as-IKKb only inhibited the activation by TNFa but had no ef-fect on the activation by either UVC or UVB (Fig 8b) In theseexperiments we used s-NFkB and s-IKKa as controls and con-firmed that they had no effect in this pathway (Fig 8b) Fur-thermore as-NIK targeting a kinase functioning upstream fromIKK also partially inhibited the effects of TNFa Therefore
the antisense oligonucleotide experiments confirm our conclu-sions that the activation of NFkB by TNFa depends on the IKKpathway whereas the response to UVC or UVB occurs througha mechanism independent of IKKs
Furthermore as an additional control we used oligonu-cleotides targeting the JNK pathway as-JNKK1 and s-JNKK1Although they affected the JNK pathway (see Figs 4 and 6)they did not affect the NFkB pathway (Fig 8b) which indi-cates that the activation of the JNK pathway and the NFkBpathway are independent in keratinocytes This means that epi-dermal keratinocytes contain multiple specific UV responsivemechanisms
DISCUSSION
Our data suggest that UVBUVC illumination in epidermalkeratinocytes is not a generic stress response inducer but a spe-cific extracellular agent that activates specific signal transduc-tion pathways In particular we show that in keratinocytes UVexclusively activates the JNK pathway while in HeLa cellsUV also activates the parallel ERK and p38 pathways Con-versely in keratinocytes exclusively UV activates the JNKpathway while in HeLa cells growth factors can do so as wellIn parallel in keratinocytes UV uses a distinct pathway to ac-tivate the NFkB transcription factor a pathway separate fromthe more common TNFa activated one (Fig 9)
The specific activation of JNK by UVBUVC is particularlyrelevant because the JNK pathway is in different cell types atarget of multiple stress agents The balance between apoptosisand proliferation is at times regulated by JNK and the tran-scription factors activated by JNK regulate the expression ofmany genes (Karin and Hunter 1995) It is uniquely importantin epidermis that JNK exclusively responds to UVBUVC be-cause epidermis is the natural target of UVBUVC and the JNK-
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 673
FIG 6 Activation of JNK in vivo and the effect of as-JNKK1 In the control sample (left) JNK is not activated as shown us-ing an antibody specific for the activated phosphorylated form of JNK Treatment with 100 mJcm2 causes strong activation ofJNK (middle) Pretreatment for 3 h with the antisense oligonucleotide targeting JNKK1 abolishes this activation (right)
FIG 7 TNFa UVB and UVC activate NFkB while UVAdoes not Reporter plasmid NFkB3-CAT (top) which containsthree tandem NFkB sites upstream from the TK promoter wastransfected into keratinocytes The transfected cells were eithertreated with 50 ngml of TNFa or illuminated with indicateddoses of UV harvested 16 h later and the CAT levels deter-mined Untreated cells served as a control The replicates dif-fered by 25 or less
ADACHI ET AL674
FIG 8 Specific inhibitors can differentiate between the TNFa-and UVB-dependent signal transduction pathways (a) The effectsof specific inhibitors on NFkB activation Cultured keratinocyteswere treated with either UVB (8 mJcm2) or TNFa (50 ngml) af-ter transfection of the NFkB response element Inhibitor Bay11-7085 specifically inhibits upstream kinases of IKK aspirin inhibitsIKKb parthenolide blocks IkBa degradation (b) the effects of an-tisense oligonucleotides targeting signaling cascades on NFkB ac-tivation in keratinocytes Oligonucleotide as-NFkB targeting thetranscription factor blocks the activation by UVB and UVC whileoligonucleotides targeting IKKb JNK1 and JNKK1 are withouteffect (top two panels) In contrast oligonucleotides targetingIKKb and NIK partially or completely block the effects of TNFa(bottom panel) Note that the sense oligonucleotides (s-NFkB s-IKKb) used as controls had no effect on NFkB activation (c)Schematics of NFkB pathways activated by UV light and TNFaIn keratinocytes the two extracellular stimuli work through inde-pendent pathways that merge at the NFkBIkB complex
regulated transcription factors mediate both photodamage andits alleviation (Fisher et al 1997)
In keratinocytes the EGF and UV responsive pathways donot overlap but UV specifically activates the JNK pathwayThis suggests that the epidermal keratinocytes exhibit distinc-tive specialized UV-responsive signal transduction pathwaysIndeed when we compared the responses to UV in several celltypes we found that the response in keratinocytes is signifi-cantly more stringently controlled than in other cell types Forexample HeLa cells demonstrated a lack of specificity Usinga specific MEK-12 inhibitor we again demonstrated relaxedstringency in HeLa cells We conclude from these results thatkeratinocytes being often exposed to ambient UV have de-veloped more stringent and specific responses to UV than othercell types which are not commonly exposed to UV
At present we do not understand the relevance of JNK3 ex-pression in keratinocytes This isoform it was reported isspecifically expressed in the brain (Yoshida et al 2001) Wespeculate that perhaps the sequestration of JNK1 by the speci-ficity of the UV response creates a need for JNK3 as the moregeneral less restrictive stress signal transducer
TNFa one of proinflammatory cytokines important in cuta-neous inflammation activates NFkB via IKKs kinases that canbe activated by MEKK or NIK (Hirano et al 1996 Lee et al1997) In HeLa cells the phosphorylation of IkBa at Ser-32and Ser-36 residues is essential for the IKK mediated IkBa
degradation caused by TNFa IL-1 or ionizing radiation Us-ing specific inhibitors and antisense oligonucleotides we foundthat the NFkB activation by UVBUVC does not involve IKKs
The Ser-32 and Ser-36 residues are not involved in the UV-induced IkBa degradation (Bender et al 1997 N Li and Karin1998) Because IKKb specifically phosphorylates IkB at bothSer-32 and Ser-36 of IkBa these facts support our results Re-cently it was suggested that UV activation of NFkB in HeLacells may involve the RasndashRaf pathway although not in ker-atinocytes (Tobin et al 1996) This leads to the question howUV activates NFkB in keratinocytes Two new kinases IKKlaquo
and pp90rsk have been found to phosphorylate IkBa (Ghodaet al 1997 Schouten et al 1997 Shimada et al 1999 Israel2000 Peters et al 2000) However IKKlaquo phosphorylates theSer-36 residue while pp90rsk phosphorylates the Ser-32residue only Therefore these kinases are probably not involvedin the UV activated pathway Amino acids between IkBa po-sition 277 and 297 (the PEST region) are apparently requiredfor the UV induced degradation of IkB (Ito et al 1995 Ben-der et al 1998) but the signaling events that effect this degra-dation in keratinocytes are still unknown The data suggest thatTNFa and UVC use different pathways to activate NFkB inHeLa cells Specifically while both UVC and TNFa activateNFkB the TNFa signal goes via IKKs but the UVC signaldoes not From these data it appears that the convergence ofthe UVB and TNFa signals occurs downstream from IKKs thatis at the modification of IkB Overall these results imply thatthe epidermis a natural target of UV light has an exceptionalmolecular UV defense system distinct from UV responsive sys-tems in other tissues
The responses to the three different wavelengths of UV areespecially strictly controlled in keratinocytes Although allwavelengths cause DNA damage and a release of cytokines wefound that the inherent molecular responses to UVA are com-pletely different from the responses to UVBUVC For exam-ple UVBUVC but not UVA activate JNK and NFkB in ker-atinocytes Furthermore UVA activates the JNK pathway inHeLa cells but not in keratinocytes while UVBUVC activatethis pathway in both cells (Rosette and Karin 1996) Similarlyin both HeLa cells and cultured keratinocytes UVBUVC ac-tivate NFkB via a kinase different from IKKs while UVA doesnot activate NFkB at all We conclude that UVBUVC activatea completely different system of signal transduction from UVA
The results suggesting specific signal transduction pathwaysfor UVBUVC in keratinocytes presented here agree well withour studies of UVB-regulated genes using DNA micro arrays(Li et al 2001) We found that in response to UVB ker-atinocytes not only induce the immediate early genes and stressresponse genes but also elaborate responses designed to pro-tect the organism underneath For example chemokines of theIL-8 family are induced which alert the surrounding tissue tothe UV damage synthesis of the cornified envelope compo-nents is enhanced which thickens the stratum corneum etc Weconclude that human epidermal keratinocytes being naturallyexposed to UV light have elaborated highly specific molecu-lar responses to UVBUVC at the signal transduction level aswell as the gene expression level
ACKNOWLEDGMENTS
Our research was supported by the National Institutes ofHealth Grants AR30682 AR39176 AR40522 and AR41850
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 675
FIG 9 Specificity in stress response Epidermal ker-atinocytes exhibit specialized UV-responsive signal transduc-tion pathways which apparently do not network and intersectwith other pathways such as those responsive to EGF or TNFaIn contrast in other cell types such as HeLa multiple path-ways crossover interact and overlap
and by the Anaderm Research Corporation Special thinks goto Dr M Simon for the generous gift of keratinocytes We alsothank members of our laboratory for advice reagents and en-couragement
REFERENCES
ALKALAY I YARON A HATZUBAI A ORIAN ACIECHANOVER A and BEN-NERIAH Y (1995) Stimulation-dependent I kappa B alpha phosphorylation marks the NF-kappa Binhibitor for degradation via the ubiquitin-proteasome pathway ProcNatl Acad Sci USA 92 10599ndash10603
BAEUERLE PA and HENKEL T (1994) Function and activationof NF-kappa B in the immune system Annu Rev Immunol 12141ndash179
BARNES PJ and KARIN M (1997) Nuclear factor-kappaB A piv-otal transcription factor in chronic inflammatory diseases N EnglJ Med 336 1066ndash1071
BEG AA FINCO TS NANTERMET PV and BALDWIN AJR (1993) Tumor necrosis factor and interleukin-1 lead to phos-phorylation and loss of I kappa B alpha A mechanism for NF-kappaB activation Mol Cell Biol 13 3301ndash3310
BENDER K BLATTNER C KNEBEL A IORDANOV M HER-RLICH P and RAHMSDORF HJ (1997) UV-induced signaltransduction J Photochem Photobiol B 37 1ndash17
BENDER K GOTTLICHER M WHITESIDE S RAHMSDORFHJ and HERRLICH P (1998) Sequential DNA damage-indepen-dent and -dependent activation of NF-kappaB by UV EMBO J 175170ndash5181
BROWN K GERSTBERGER S CARLSON L FRANZOSO Gand SIEBENLIST U (1995) Control of I kappa B-alpha proteoly-sis by site-specific signal-induced phosphorylation Science 2671485ndash1488
DELHASE M HAYAKAWA M CHEN Y and KARIN M(1999) Positive and negative regulation of IkappaB kinase activitythrough IKKbeta subunit phosphorylation Science 284 309ndash313
DEacuteRIJARD B et al (1994) JNK1 A protein kinase stimulated byUV light and Ha-Ras that binds and phosphorylates the c-Jun acti-vation domain Cell 76 1025ndash1037
DEVARY Y ROSETTE C DIDONATO JA and KARIN M(1993) NF-kappa B activation by ultraviolet light not dependent ona nuclear signal Science 261 1442ndash1445
DIDONATO JA HAYAKAWA M ROTHWARF DM ZANDIE and KARIN M (1997) A cytokine-responsive IkappaB kinasethat activates the transcription factor NF-kappaB Nature 388548ndash554
FANGER GR GERWINS P WIDMANN C JARPE MB andJOHNSON GL (1997) MEKKs GCKs MLKs PAKs TAKs andtpls Upstream regulators of the c-Jun amino-terminal kinases CurrOpin Genet Dev 7 67ndash74
FEIGE U MORIMOTO RI YAHARA I and POLLA BS eds(1996) Stress-Inducible Cellular Responses (Birkhauser VerlagBasel Switzerland)
FISHER GJ WANG ZQ DATTA SC VARANI J KANG Sand VOORHEES JJ (1997) Pathophysiology of premature skin ag-ing induced by ultraviolet light N Engl J Med 337 1419ndash1428
GHODA L LIN X and GREENE WC (1997) The 90-kDa ribo-somal S6 kinase (pp90rsk) phosphorylates the N-terminal regulatorydomain of IkappaBalpha and stimulates its degradation in vitro JBiol Chem 272 21281ndash21288
GILCHREST B (1995) Photodamage (Blackwell Scientific NewYork)
GRETHER-BECK S BUETTNER R and KRUTMANN J (1997)Ultraviolet A radiation-induced expression of human genes Molec-ular and photobiological mechanisms Biol Chem 378 1231ndash1236
GRILLI M PIZZI M MEMO M and SPANO P (1996) Neuro-protection by aspirin and sodium salicylate through blockade of NF-kappaB activation Science 274 1383ndash1385
GUPTA S et al (1996) Selective interaction of JNK protein kinaseisoforms with transcription factors EMBO J 15 2760ndash2770
HEHNER SP et al (1998) Sesquiterpene lactones specifically in-hibit activation of NF-kappa B by preventing the degradation of Ikappa B-alpha and I kappa B-beta J Biol Chem 273 1288ndash1297
HERSKOWITZ I (1995) MAP kinase pathways in yeast For matingand more Cell 80 187ndash197
HIBI M LIN A SMEAL T MINDEN A and KARIN M (1993)Identification of an oncoprotein- and UV-responsive protein kinasethat binds and potentiates the c-Jun activation domain Genes Dev7 2135ndash2148
HIRANO M et al (1996) MEK kinase is involved in tumor necro-sis factor alpha-induced NF-kappaB activation and degradation ofIkappaB-alpha J Biol Chem 271 13234ndash13238
HU Y et al (1999) Abnormal morphogenesis but intact IKK activa-tion in mice lacking the IKKalpha subunit of IkappaB kinase Sci-ence 284 316ndash320
IORDANOV M et al (1997) CREB is activated by UVC through ap38HOG-1-dependent protein kinase EMBO J 16 1009ndash1022
ISREAL A (2000) The IKK complex An integrator of all signals thatactivate NF-kappaB Trends Cell Biol 10 129ndash133
ITO CY ADEY N BAUTCH VL and BALDWIN AS (1995)Structure and evolution of the human IKBA gene Genomics 29490ndash495
JIANG CK CONNOLLY D and BLUMENBERG M (1991)Comparison of methods for transfection of human epidermal ker-atinocytes J Invest Dermatol 97 969ndash973
JIANG CK FLANAGAN S OHTSUKI M SHUAI K FREED-BERG IM and BLUMENBERG M (1994) Disease-activatedtranscription factor Allergic reactions in human skin cause nucleartranscription of STAT-91 and induce synthesis of keratin K17 MolCell Biol 14 4759ndash4769
JIANG CK MAGNALDO T OHTSUKI M FREEDBERG IMBERNERD F and BLUMENBERG M (1993) Epidermal growthfactor and transforming growth factor alpha specifically induce theactivation- and hyperproliferation-associated keratins 6 and 16 ProcNatl Acad Sci USA 90 6786ndash6790
KALLUNKI T DENG T HIBI M and KARIN M (1996) c-Juncan recruit JNK to phosphorylate dimerization partners via specificdocking interactions Cell 87 929ndash939
KARIN M (1995) The regulation of AP-1 activity by mitogen-acti-vated protein kinases J Biol Chem 270 16483ndash16486
KARIN M and HUNTER T (1995) Transcriptional control by pro-tein phosphorylation Signal transmission from the cell surface to thenucleus Curr Biol 5 747ndash757
KLAMPFER L LEE TH HSU W VILCEK J and CHEN-KIANG S (1994) NF-IL6 and AP-1 cooperatively modulate the ac-tivation of the TSG-6 gene by tumor necrosis factor alpha and in-terleukin-1 Mol Cell Biol 14 6561ndash6569
KOMINE M et al (2000) Inflammatory versus proliferative pro-cesses in epidermis Tumor necrosis factor alpha induces K6b ker-atin synthesis through a transcriptional complex containing NFkappaB and CEBPbeta J Biol Chem 275 32077ndash32088
KOPP E and GHOSH S (1994) Inhibition of NF-kappa B by so-dium salicylate and aspirin Science 265 956ndash959
KYRIAKIS JM et al (1994) The stress-activated protein kinase sub-family of c-Jun kinases Nature 369 156ndash160
LEE FS HAGLER J CHEN ZJ and MANIATIS T (1997) Ac-tivation of the IkappaB alpha kinase complex by MEKK1 a kinaseof the JNK pathway Cell 88 213ndash222
LEPPA S and BOHMANN D (1999) Diverse functions of JNK sig-naling and c-Jun in stress response and apoptosis Oncogene 186158ndash6162
ADACHI ET AL676
LI D TURI TG SCHUCK A FREEDBERG IM KHITROVG and BLUMENBERG M (2001) Rays and arrays The tran-scriptional program in the response of human epidermal keratinocytesto UVB illumination FASEB J 15 2533ndash2535
LI N and KARIN M (1998) Ionizing radiation and short wavelengthUV activate NF-kappaB through two distinct mechanisms ProcNatl Acad Sci USA 95 13012ndash13017
LIU ZG HSU H GOEDDEL DV and KARIN M (1996) Dis-section of TNF receptor 1 effector functions JNK activation is notlinked to apoptosis while NF-kappaB activation prevents cell deathCell 87 565ndash576
MERCURIO F and MANNING AM (1999) NF-kappaB as a pri-mary regulator of the stress response Oncogene 18 6163ndash6171
MIYAMOTO S and VERMA IM (1995) RelNF-kappa BI kappaB story Adv Cancer Res 66 255ndash292
NESTLE FO MITRA RS BENNETT CF CHAN H andNICKOLOFF BJ (1994) Cationic lipid is not required for uptakeand selective inhibitory activity of ICAM-1 phosphorothioate anti-sense oligonucleotides in keratinocytes J Invest Dermatol 103569ndash575
PETERS RT LIAO SM and MANIATIS T (2000) IKKepsilonis part of a novel PMA-inducible IkappaB kinase complex Mol Cell5 513ndash522
PIERCE JW READ MA DING H LUSCINSKAS FW andCOLLINS T (1996) Salicylates inhibit I kappa B-alpha phos-phorylation endothelial-leukocyte adhesion molecule expressionand neutrophil transmigration J Immunol 156 3961ndash3969
PIERCE JW et al (1997) Novel inhibitors of cytokine-induced Ikap-paBalpha phosphorylation and endothelial cell adhesion molecule ex-pression show anti-inflammatory effects in vivo J Biol Chem 27221096ndash21103
REUNING U et al (1995) Inhibition of NF-kappa B-Rel A expres-sion by antisense oligodeoxynucleotides suppresses synthesis ofurokinase-type plasminogen activator (uPA) but not its inhibitor PAI-1 Nucleic Acids Res 23 3887ndash3893
ROSETTE C and KARIN M (1996) Ultraviolet light and osmoticstress activation of the JNK cascade through multiple growth factorand cytokine receptors Science 274 1194ndash1197
SCHOUTEN GJ et al (1997) IkappaB alpha is a target for the mitogen-activated 90 kDa ribosomal S6 kinase EMBO J 163133ndash3144
SHIMADA T et al (1999) IKK-i a novel lipopolysaccharide-inducible kinase that is related to IkappaB kinases Int Immunol 111357ndash1362
SIMON M and GREEN H (1984) Participation of membrane-asso-ciated proteins in the formation of the cross-linked envelope of thekeratinocyte Cell 36 827ndash834
SIMON MM ARAGANE Y SCHWARZ A LUGER TA andSCHWARZ T (1994) UVB light induces nuclear factor kappa B(NF kappa B) activity independently from chromosomal DNA dam-age in cell-free cytosolic extracts J Invest Dermatol 102 422ndash427
TOBIN D NILSSON M and TOFTGARD R (1996) Ras-inde-pendent activation of Rel-family transcription factors by UVB andTPA in cultured keratinocytes Oncogene 12 785ndash793
TOMIC-CANIC M KOMINE M FREEDBERG IM and BLU-MENBERG M (1998) Epidermal signal transduction and tran-scription factor activation in activated keratinocytes J Dermatol Sci17 167ndash181
WILLIAMS BR (1999) PKR A sentinel kinase for cellular stressOncogene 18 6112ndash6120
WU Z WU J JACINTO E and KARIN M (1997) Molecularcloning and characterization of human JNKK2 a novel Jun NH2-terminal kinase-specific kinase Mol Cell Biol 17 7407ndash7416
YANG DD et al (1997) Absence of excitotoxicity-induced apopto-sis in the hippocampus of mice lacking the Jnk3 gene Nature 389865ndash870
YANKULOV K BLAU J PURTON T ROBERTS S and BENT-LEY DL (1994) Transcriptional elongation by RNA polymeraseII is stimulated by transactivators Cell 77 749ndash759
YIN MJ YAMAMOTO Y and GAYNOR RB (1998) The anti-inflammatory agents aspirin and salicylate inhibit the activity ofI(kappa)B kinase-beta Nature 396 77ndash80
YOSHIDA S et al (2001) The c-Jun NH2-terminal kinase3 (JNK3)gene Genomic structure chromosomal assignment and loss of ex-pression in brain tumors J Hum Genet 46 182ndash187
Address reprint requests toM Blumenberg PhD
The NYU Cancer InstituteNew York University School of Medicine
550 First AveNew York NY 10016
E-mail blumem01mednyuedu
Received for publication June 5 2003 received in revised formJuly 15 2003 accepted August 14 2003
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 677

ADACHI ET AL674
FIG 8 Specific inhibitors can differentiate between the TNFa-and UVB-dependent signal transduction pathways (a) The effectsof specific inhibitors on NFkB activation Cultured keratinocyteswere treated with either UVB (8 mJcm2) or TNFa (50 ngml) af-ter transfection of the NFkB response element Inhibitor Bay11-7085 specifically inhibits upstream kinases of IKK aspirin inhibitsIKKb parthenolide blocks IkBa degradation (b) the effects of an-tisense oligonucleotides targeting signaling cascades on NFkB ac-tivation in keratinocytes Oligonucleotide as-NFkB targeting thetranscription factor blocks the activation by UVB and UVC whileoligonucleotides targeting IKKb JNK1 and JNKK1 are withouteffect (top two panels) In contrast oligonucleotides targetingIKKb and NIK partially or completely block the effects of TNFa(bottom panel) Note that the sense oligonucleotides (s-NFkB s-IKKb) used as controls had no effect on NFkB activation (c)Schematics of NFkB pathways activated by UV light and TNFaIn keratinocytes the two extracellular stimuli work through inde-pendent pathways that merge at the NFkBIkB complex
regulated transcription factors mediate both photodamage andits alleviation (Fisher et al 1997)
In keratinocytes the EGF and UV responsive pathways donot overlap but UV specifically activates the JNK pathwayThis suggests that the epidermal keratinocytes exhibit distinc-tive specialized UV-responsive signal transduction pathwaysIndeed when we compared the responses to UV in several celltypes we found that the response in keratinocytes is signifi-cantly more stringently controlled than in other cell types Forexample HeLa cells demonstrated a lack of specificity Usinga specific MEK-12 inhibitor we again demonstrated relaxedstringency in HeLa cells We conclude from these results thatkeratinocytes being often exposed to ambient UV have de-veloped more stringent and specific responses to UV than othercell types which are not commonly exposed to UV
At present we do not understand the relevance of JNK3 ex-pression in keratinocytes This isoform it was reported isspecifically expressed in the brain (Yoshida et al 2001) Wespeculate that perhaps the sequestration of JNK1 by the speci-ficity of the UV response creates a need for JNK3 as the moregeneral less restrictive stress signal transducer
TNFa one of proinflammatory cytokines important in cuta-neous inflammation activates NFkB via IKKs kinases that canbe activated by MEKK or NIK (Hirano et al 1996 Lee et al1997) In HeLa cells the phosphorylation of IkBa at Ser-32and Ser-36 residues is essential for the IKK mediated IkBa
degradation caused by TNFa IL-1 or ionizing radiation Us-ing specific inhibitors and antisense oligonucleotides we foundthat the NFkB activation by UVBUVC does not involve IKKs
The Ser-32 and Ser-36 residues are not involved in the UV-induced IkBa degradation (Bender et al 1997 N Li and Karin1998) Because IKKb specifically phosphorylates IkB at bothSer-32 and Ser-36 of IkBa these facts support our results Re-cently it was suggested that UV activation of NFkB in HeLacells may involve the RasndashRaf pathway although not in ker-atinocytes (Tobin et al 1996) This leads to the question howUV activates NFkB in keratinocytes Two new kinases IKKlaquo
and pp90rsk have been found to phosphorylate IkBa (Ghodaet al 1997 Schouten et al 1997 Shimada et al 1999 Israel2000 Peters et al 2000) However IKKlaquo phosphorylates theSer-36 residue while pp90rsk phosphorylates the Ser-32residue only Therefore these kinases are probably not involvedin the UV activated pathway Amino acids between IkBa po-sition 277 and 297 (the PEST region) are apparently requiredfor the UV induced degradation of IkB (Ito et al 1995 Ben-der et al 1998) but the signaling events that effect this degra-dation in keratinocytes are still unknown The data suggest thatTNFa and UVC use different pathways to activate NFkB inHeLa cells Specifically while both UVC and TNFa activateNFkB the TNFa signal goes via IKKs but the UVC signaldoes not From these data it appears that the convergence ofthe UVB and TNFa signals occurs downstream from IKKs thatis at the modification of IkB Overall these results imply thatthe epidermis a natural target of UV light has an exceptionalmolecular UV defense system distinct from UV responsive sys-tems in other tissues
The responses to the three different wavelengths of UV areespecially strictly controlled in keratinocytes Although allwavelengths cause DNA damage and a release of cytokines wefound that the inherent molecular responses to UVA are com-pletely different from the responses to UVBUVC For exam-ple UVBUVC but not UVA activate JNK and NFkB in ker-atinocytes Furthermore UVA activates the JNK pathway inHeLa cells but not in keratinocytes while UVBUVC activatethis pathway in both cells (Rosette and Karin 1996) Similarlyin both HeLa cells and cultured keratinocytes UVBUVC ac-tivate NFkB via a kinase different from IKKs while UVA doesnot activate NFkB at all We conclude that UVBUVC activatea completely different system of signal transduction from UVA
The results suggesting specific signal transduction pathwaysfor UVBUVC in keratinocytes presented here agree well withour studies of UVB-regulated genes using DNA micro arrays(Li et al 2001) We found that in response to UVB ker-atinocytes not only induce the immediate early genes and stressresponse genes but also elaborate responses designed to pro-tect the organism underneath For example chemokines of theIL-8 family are induced which alert the surrounding tissue tothe UV damage synthesis of the cornified envelope compo-nents is enhanced which thickens the stratum corneum etc Weconclude that human epidermal keratinocytes being naturallyexposed to UV light have elaborated highly specific molecu-lar responses to UVBUVC at the signal transduction level aswell as the gene expression level
ACKNOWLEDGMENTS
Our research was supported by the National Institutes ofHealth Grants AR30682 AR39176 AR40522 and AR41850
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 675
FIG 9 Specificity in stress response Epidermal ker-atinocytes exhibit specialized UV-responsive signal transduc-tion pathways which apparently do not network and intersectwith other pathways such as those responsive to EGF or TNFaIn contrast in other cell types such as HeLa multiple path-ways crossover interact and overlap
and by the Anaderm Research Corporation Special thinks goto Dr M Simon for the generous gift of keratinocytes We alsothank members of our laboratory for advice reagents and en-couragement
REFERENCES
ALKALAY I YARON A HATZUBAI A ORIAN ACIECHANOVER A and BEN-NERIAH Y (1995) Stimulation-dependent I kappa B alpha phosphorylation marks the NF-kappa Binhibitor for degradation via the ubiquitin-proteasome pathway ProcNatl Acad Sci USA 92 10599ndash10603
BAEUERLE PA and HENKEL T (1994) Function and activationof NF-kappa B in the immune system Annu Rev Immunol 12141ndash179
BARNES PJ and KARIN M (1997) Nuclear factor-kappaB A piv-otal transcription factor in chronic inflammatory diseases N EnglJ Med 336 1066ndash1071
BEG AA FINCO TS NANTERMET PV and BALDWIN AJR (1993) Tumor necrosis factor and interleukin-1 lead to phos-phorylation and loss of I kappa B alpha A mechanism for NF-kappaB activation Mol Cell Biol 13 3301ndash3310
BENDER K BLATTNER C KNEBEL A IORDANOV M HER-RLICH P and RAHMSDORF HJ (1997) UV-induced signaltransduction J Photochem Photobiol B 37 1ndash17
BENDER K GOTTLICHER M WHITESIDE S RAHMSDORFHJ and HERRLICH P (1998) Sequential DNA damage-indepen-dent and -dependent activation of NF-kappaB by UV EMBO J 175170ndash5181
BROWN K GERSTBERGER S CARLSON L FRANZOSO Gand SIEBENLIST U (1995) Control of I kappa B-alpha proteoly-sis by site-specific signal-induced phosphorylation Science 2671485ndash1488
DELHASE M HAYAKAWA M CHEN Y and KARIN M(1999) Positive and negative regulation of IkappaB kinase activitythrough IKKbeta subunit phosphorylation Science 284 309ndash313
DEacuteRIJARD B et al (1994) JNK1 A protein kinase stimulated byUV light and Ha-Ras that binds and phosphorylates the c-Jun acti-vation domain Cell 76 1025ndash1037
DEVARY Y ROSETTE C DIDONATO JA and KARIN M(1993) NF-kappa B activation by ultraviolet light not dependent ona nuclear signal Science 261 1442ndash1445
DIDONATO JA HAYAKAWA M ROTHWARF DM ZANDIE and KARIN M (1997) A cytokine-responsive IkappaB kinasethat activates the transcription factor NF-kappaB Nature 388548ndash554
FANGER GR GERWINS P WIDMANN C JARPE MB andJOHNSON GL (1997) MEKKs GCKs MLKs PAKs TAKs andtpls Upstream regulators of the c-Jun amino-terminal kinases CurrOpin Genet Dev 7 67ndash74
FEIGE U MORIMOTO RI YAHARA I and POLLA BS eds(1996) Stress-Inducible Cellular Responses (Birkhauser VerlagBasel Switzerland)
FISHER GJ WANG ZQ DATTA SC VARANI J KANG Sand VOORHEES JJ (1997) Pathophysiology of premature skin ag-ing induced by ultraviolet light N Engl J Med 337 1419ndash1428
GHODA L LIN X and GREENE WC (1997) The 90-kDa ribo-somal S6 kinase (pp90rsk) phosphorylates the N-terminal regulatorydomain of IkappaBalpha and stimulates its degradation in vitro JBiol Chem 272 21281ndash21288
GILCHREST B (1995) Photodamage (Blackwell Scientific NewYork)
GRETHER-BECK S BUETTNER R and KRUTMANN J (1997)Ultraviolet A radiation-induced expression of human genes Molec-ular and photobiological mechanisms Biol Chem 378 1231ndash1236
GRILLI M PIZZI M MEMO M and SPANO P (1996) Neuro-protection by aspirin and sodium salicylate through blockade of NF-kappaB activation Science 274 1383ndash1385
GUPTA S et al (1996) Selective interaction of JNK protein kinaseisoforms with transcription factors EMBO J 15 2760ndash2770
HEHNER SP et al (1998) Sesquiterpene lactones specifically in-hibit activation of NF-kappa B by preventing the degradation of Ikappa B-alpha and I kappa B-beta J Biol Chem 273 1288ndash1297
HERSKOWITZ I (1995) MAP kinase pathways in yeast For matingand more Cell 80 187ndash197
HIBI M LIN A SMEAL T MINDEN A and KARIN M (1993)Identification of an oncoprotein- and UV-responsive protein kinasethat binds and potentiates the c-Jun activation domain Genes Dev7 2135ndash2148
HIRANO M et al (1996) MEK kinase is involved in tumor necro-sis factor alpha-induced NF-kappaB activation and degradation ofIkappaB-alpha J Biol Chem 271 13234ndash13238
HU Y et al (1999) Abnormal morphogenesis but intact IKK activa-tion in mice lacking the IKKalpha subunit of IkappaB kinase Sci-ence 284 316ndash320
IORDANOV M et al (1997) CREB is activated by UVC through ap38HOG-1-dependent protein kinase EMBO J 16 1009ndash1022
ISREAL A (2000) The IKK complex An integrator of all signals thatactivate NF-kappaB Trends Cell Biol 10 129ndash133
ITO CY ADEY N BAUTCH VL and BALDWIN AS (1995)Structure and evolution of the human IKBA gene Genomics 29490ndash495
JIANG CK CONNOLLY D and BLUMENBERG M (1991)Comparison of methods for transfection of human epidermal ker-atinocytes J Invest Dermatol 97 969ndash973
JIANG CK FLANAGAN S OHTSUKI M SHUAI K FREED-BERG IM and BLUMENBERG M (1994) Disease-activatedtranscription factor Allergic reactions in human skin cause nucleartranscription of STAT-91 and induce synthesis of keratin K17 MolCell Biol 14 4759ndash4769
JIANG CK MAGNALDO T OHTSUKI M FREEDBERG IMBERNERD F and BLUMENBERG M (1993) Epidermal growthfactor and transforming growth factor alpha specifically induce theactivation- and hyperproliferation-associated keratins 6 and 16 ProcNatl Acad Sci USA 90 6786ndash6790
KALLUNKI T DENG T HIBI M and KARIN M (1996) c-Juncan recruit JNK to phosphorylate dimerization partners via specificdocking interactions Cell 87 929ndash939
KARIN M (1995) The regulation of AP-1 activity by mitogen-acti-vated protein kinases J Biol Chem 270 16483ndash16486
KARIN M and HUNTER T (1995) Transcriptional control by pro-tein phosphorylation Signal transmission from the cell surface to thenucleus Curr Biol 5 747ndash757
KLAMPFER L LEE TH HSU W VILCEK J and CHEN-KIANG S (1994) NF-IL6 and AP-1 cooperatively modulate the ac-tivation of the TSG-6 gene by tumor necrosis factor alpha and in-terleukin-1 Mol Cell Biol 14 6561ndash6569
KOMINE M et al (2000) Inflammatory versus proliferative pro-cesses in epidermis Tumor necrosis factor alpha induces K6b ker-atin synthesis through a transcriptional complex containing NFkappaB and CEBPbeta J Biol Chem 275 32077ndash32088
KOPP E and GHOSH S (1994) Inhibition of NF-kappa B by so-dium salicylate and aspirin Science 265 956ndash959
KYRIAKIS JM et al (1994) The stress-activated protein kinase sub-family of c-Jun kinases Nature 369 156ndash160
LEE FS HAGLER J CHEN ZJ and MANIATIS T (1997) Ac-tivation of the IkappaB alpha kinase complex by MEKK1 a kinaseof the JNK pathway Cell 88 213ndash222
LEPPA S and BOHMANN D (1999) Diverse functions of JNK sig-naling and c-Jun in stress response and apoptosis Oncogene 186158ndash6162
ADACHI ET AL676
LI D TURI TG SCHUCK A FREEDBERG IM KHITROVG and BLUMENBERG M (2001) Rays and arrays The tran-scriptional program in the response of human epidermal keratinocytesto UVB illumination FASEB J 15 2533ndash2535
LI N and KARIN M (1998) Ionizing radiation and short wavelengthUV activate NF-kappaB through two distinct mechanisms ProcNatl Acad Sci USA 95 13012ndash13017
LIU ZG HSU H GOEDDEL DV and KARIN M (1996) Dis-section of TNF receptor 1 effector functions JNK activation is notlinked to apoptosis while NF-kappaB activation prevents cell deathCell 87 565ndash576
MERCURIO F and MANNING AM (1999) NF-kappaB as a pri-mary regulator of the stress response Oncogene 18 6163ndash6171
MIYAMOTO S and VERMA IM (1995) RelNF-kappa BI kappaB story Adv Cancer Res 66 255ndash292
NESTLE FO MITRA RS BENNETT CF CHAN H andNICKOLOFF BJ (1994) Cationic lipid is not required for uptakeand selective inhibitory activity of ICAM-1 phosphorothioate anti-sense oligonucleotides in keratinocytes J Invest Dermatol 103569ndash575
PETERS RT LIAO SM and MANIATIS T (2000) IKKepsilonis part of a novel PMA-inducible IkappaB kinase complex Mol Cell5 513ndash522
PIERCE JW READ MA DING H LUSCINSKAS FW andCOLLINS T (1996) Salicylates inhibit I kappa B-alpha phos-phorylation endothelial-leukocyte adhesion molecule expressionand neutrophil transmigration J Immunol 156 3961ndash3969
PIERCE JW et al (1997) Novel inhibitors of cytokine-induced Ikap-paBalpha phosphorylation and endothelial cell adhesion molecule ex-pression show anti-inflammatory effects in vivo J Biol Chem 27221096ndash21103
REUNING U et al (1995) Inhibition of NF-kappa B-Rel A expres-sion by antisense oligodeoxynucleotides suppresses synthesis ofurokinase-type plasminogen activator (uPA) but not its inhibitor PAI-1 Nucleic Acids Res 23 3887ndash3893
ROSETTE C and KARIN M (1996) Ultraviolet light and osmoticstress activation of the JNK cascade through multiple growth factorand cytokine receptors Science 274 1194ndash1197
SCHOUTEN GJ et al (1997) IkappaB alpha is a target for the mitogen-activated 90 kDa ribosomal S6 kinase EMBO J 163133ndash3144
SHIMADA T et al (1999) IKK-i a novel lipopolysaccharide-inducible kinase that is related to IkappaB kinases Int Immunol 111357ndash1362
SIMON M and GREEN H (1984) Participation of membrane-asso-ciated proteins in the formation of the cross-linked envelope of thekeratinocyte Cell 36 827ndash834
SIMON MM ARAGANE Y SCHWARZ A LUGER TA andSCHWARZ T (1994) UVB light induces nuclear factor kappa B(NF kappa B) activity independently from chromosomal DNA dam-age in cell-free cytosolic extracts J Invest Dermatol 102 422ndash427
TOBIN D NILSSON M and TOFTGARD R (1996) Ras-inde-pendent activation of Rel-family transcription factors by UVB andTPA in cultured keratinocytes Oncogene 12 785ndash793
TOMIC-CANIC M KOMINE M FREEDBERG IM and BLU-MENBERG M (1998) Epidermal signal transduction and tran-scription factor activation in activated keratinocytes J Dermatol Sci17 167ndash181
WILLIAMS BR (1999) PKR A sentinel kinase for cellular stressOncogene 18 6112ndash6120
WU Z WU J JACINTO E and KARIN M (1997) Molecularcloning and characterization of human JNKK2 a novel Jun NH2-terminal kinase-specific kinase Mol Cell Biol 17 7407ndash7416
YANG DD et al (1997) Absence of excitotoxicity-induced apopto-sis in the hippocampus of mice lacking the Jnk3 gene Nature 389865ndash870
YANKULOV K BLAU J PURTON T ROBERTS S and BENT-LEY DL (1994) Transcriptional elongation by RNA polymeraseII is stimulated by transactivators Cell 77 749ndash759
YIN MJ YAMAMOTO Y and GAYNOR RB (1998) The anti-inflammatory agents aspirin and salicylate inhibit the activity ofI(kappa)B kinase-beta Nature 396 77ndash80
YOSHIDA S et al (2001) The c-Jun NH2-terminal kinase3 (JNK3)gene Genomic structure chromosomal assignment and loss of ex-pression in brain tumors J Hum Genet 46 182ndash187
Address reprint requests toM Blumenberg PhD
The NYU Cancer InstituteNew York University School of Medicine
550 First AveNew York NY 10016
E-mail blumem01mednyuedu
Received for publication June 5 2003 received in revised formJuly 15 2003 accepted August 14 2003
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 677

regulated transcription factors mediate both photodamage andits alleviation (Fisher et al 1997)
In keratinocytes the EGF and UV responsive pathways donot overlap but UV specifically activates the JNK pathwayThis suggests that the epidermal keratinocytes exhibit distinc-tive specialized UV-responsive signal transduction pathwaysIndeed when we compared the responses to UV in several celltypes we found that the response in keratinocytes is signifi-cantly more stringently controlled than in other cell types Forexample HeLa cells demonstrated a lack of specificity Usinga specific MEK-12 inhibitor we again demonstrated relaxedstringency in HeLa cells We conclude from these results thatkeratinocytes being often exposed to ambient UV have de-veloped more stringent and specific responses to UV than othercell types which are not commonly exposed to UV
At present we do not understand the relevance of JNK3 ex-pression in keratinocytes This isoform it was reported isspecifically expressed in the brain (Yoshida et al 2001) Wespeculate that perhaps the sequestration of JNK1 by the speci-ficity of the UV response creates a need for JNK3 as the moregeneral less restrictive stress signal transducer
TNFa one of proinflammatory cytokines important in cuta-neous inflammation activates NFkB via IKKs kinases that canbe activated by MEKK or NIK (Hirano et al 1996 Lee et al1997) In HeLa cells the phosphorylation of IkBa at Ser-32and Ser-36 residues is essential for the IKK mediated IkBa
degradation caused by TNFa IL-1 or ionizing radiation Us-ing specific inhibitors and antisense oligonucleotides we foundthat the NFkB activation by UVBUVC does not involve IKKs
The Ser-32 and Ser-36 residues are not involved in the UV-induced IkBa degradation (Bender et al 1997 N Li and Karin1998) Because IKKb specifically phosphorylates IkB at bothSer-32 and Ser-36 of IkBa these facts support our results Re-cently it was suggested that UV activation of NFkB in HeLacells may involve the RasndashRaf pathway although not in ker-atinocytes (Tobin et al 1996) This leads to the question howUV activates NFkB in keratinocytes Two new kinases IKKlaquo
and pp90rsk have been found to phosphorylate IkBa (Ghodaet al 1997 Schouten et al 1997 Shimada et al 1999 Israel2000 Peters et al 2000) However IKKlaquo phosphorylates theSer-36 residue while pp90rsk phosphorylates the Ser-32residue only Therefore these kinases are probably not involvedin the UV activated pathway Amino acids between IkBa po-sition 277 and 297 (the PEST region) are apparently requiredfor the UV induced degradation of IkB (Ito et al 1995 Ben-der et al 1998) but the signaling events that effect this degra-dation in keratinocytes are still unknown The data suggest thatTNFa and UVC use different pathways to activate NFkB inHeLa cells Specifically while both UVC and TNFa activateNFkB the TNFa signal goes via IKKs but the UVC signaldoes not From these data it appears that the convergence ofthe UVB and TNFa signals occurs downstream from IKKs thatis at the modification of IkB Overall these results imply thatthe epidermis a natural target of UV light has an exceptionalmolecular UV defense system distinct from UV responsive sys-tems in other tissues
The responses to the three different wavelengths of UV areespecially strictly controlled in keratinocytes Although allwavelengths cause DNA damage and a release of cytokines wefound that the inherent molecular responses to UVA are com-pletely different from the responses to UVBUVC For exam-ple UVBUVC but not UVA activate JNK and NFkB in ker-atinocytes Furthermore UVA activates the JNK pathway inHeLa cells but not in keratinocytes while UVBUVC activatethis pathway in both cells (Rosette and Karin 1996) Similarlyin both HeLa cells and cultured keratinocytes UVBUVC ac-tivate NFkB via a kinase different from IKKs while UVA doesnot activate NFkB at all We conclude that UVBUVC activatea completely different system of signal transduction from UVA
The results suggesting specific signal transduction pathwaysfor UVBUVC in keratinocytes presented here agree well withour studies of UVB-regulated genes using DNA micro arrays(Li et al 2001) We found that in response to UVB ker-atinocytes not only induce the immediate early genes and stressresponse genes but also elaborate responses designed to pro-tect the organism underneath For example chemokines of theIL-8 family are induced which alert the surrounding tissue tothe UV damage synthesis of the cornified envelope compo-nents is enhanced which thickens the stratum corneum etc Weconclude that human epidermal keratinocytes being naturallyexposed to UV light have elaborated highly specific molecu-lar responses to UVBUVC at the signal transduction level aswell as the gene expression level
ACKNOWLEDGMENTS
Our research was supported by the National Institutes ofHealth Grants AR30682 AR39176 AR40522 and AR41850
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 675
FIG 9 Specificity in stress response Epidermal ker-atinocytes exhibit specialized UV-responsive signal transduc-tion pathways which apparently do not network and intersectwith other pathways such as those responsive to EGF or TNFaIn contrast in other cell types such as HeLa multiple path-ways crossover interact and overlap
and by the Anaderm Research Corporation Special thinks goto Dr M Simon for the generous gift of keratinocytes We alsothank members of our laboratory for advice reagents and en-couragement
REFERENCES
ALKALAY I YARON A HATZUBAI A ORIAN ACIECHANOVER A and BEN-NERIAH Y (1995) Stimulation-dependent I kappa B alpha phosphorylation marks the NF-kappa Binhibitor for degradation via the ubiquitin-proteasome pathway ProcNatl Acad Sci USA 92 10599ndash10603
BAEUERLE PA and HENKEL T (1994) Function and activationof NF-kappa B in the immune system Annu Rev Immunol 12141ndash179
BARNES PJ and KARIN M (1997) Nuclear factor-kappaB A piv-otal transcription factor in chronic inflammatory diseases N EnglJ Med 336 1066ndash1071
BEG AA FINCO TS NANTERMET PV and BALDWIN AJR (1993) Tumor necrosis factor and interleukin-1 lead to phos-phorylation and loss of I kappa B alpha A mechanism for NF-kappaB activation Mol Cell Biol 13 3301ndash3310
BENDER K BLATTNER C KNEBEL A IORDANOV M HER-RLICH P and RAHMSDORF HJ (1997) UV-induced signaltransduction J Photochem Photobiol B 37 1ndash17
BENDER K GOTTLICHER M WHITESIDE S RAHMSDORFHJ and HERRLICH P (1998) Sequential DNA damage-indepen-dent and -dependent activation of NF-kappaB by UV EMBO J 175170ndash5181
BROWN K GERSTBERGER S CARLSON L FRANZOSO Gand SIEBENLIST U (1995) Control of I kappa B-alpha proteoly-sis by site-specific signal-induced phosphorylation Science 2671485ndash1488
DELHASE M HAYAKAWA M CHEN Y and KARIN M(1999) Positive and negative regulation of IkappaB kinase activitythrough IKKbeta subunit phosphorylation Science 284 309ndash313
DEacuteRIJARD B et al (1994) JNK1 A protein kinase stimulated byUV light and Ha-Ras that binds and phosphorylates the c-Jun acti-vation domain Cell 76 1025ndash1037
DEVARY Y ROSETTE C DIDONATO JA and KARIN M(1993) NF-kappa B activation by ultraviolet light not dependent ona nuclear signal Science 261 1442ndash1445
DIDONATO JA HAYAKAWA M ROTHWARF DM ZANDIE and KARIN M (1997) A cytokine-responsive IkappaB kinasethat activates the transcription factor NF-kappaB Nature 388548ndash554
FANGER GR GERWINS P WIDMANN C JARPE MB andJOHNSON GL (1997) MEKKs GCKs MLKs PAKs TAKs andtpls Upstream regulators of the c-Jun amino-terminal kinases CurrOpin Genet Dev 7 67ndash74
FEIGE U MORIMOTO RI YAHARA I and POLLA BS eds(1996) Stress-Inducible Cellular Responses (Birkhauser VerlagBasel Switzerland)
FISHER GJ WANG ZQ DATTA SC VARANI J KANG Sand VOORHEES JJ (1997) Pathophysiology of premature skin ag-ing induced by ultraviolet light N Engl J Med 337 1419ndash1428
GHODA L LIN X and GREENE WC (1997) The 90-kDa ribo-somal S6 kinase (pp90rsk) phosphorylates the N-terminal regulatorydomain of IkappaBalpha and stimulates its degradation in vitro JBiol Chem 272 21281ndash21288
GILCHREST B (1995) Photodamage (Blackwell Scientific NewYork)
GRETHER-BECK S BUETTNER R and KRUTMANN J (1997)Ultraviolet A radiation-induced expression of human genes Molec-ular and photobiological mechanisms Biol Chem 378 1231ndash1236
GRILLI M PIZZI M MEMO M and SPANO P (1996) Neuro-protection by aspirin and sodium salicylate through blockade of NF-kappaB activation Science 274 1383ndash1385
GUPTA S et al (1996) Selective interaction of JNK protein kinaseisoforms with transcription factors EMBO J 15 2760ndash2770
HEHNER SP et al (1998) Sesquiterpene lactones specifically in-hibit activation of NF-kappa B by preventing the degradation of Ikappa B-alpha and I kappa B-beta J Biol Chem 273 1288ndash1297
HERSKOWITZ I (1995) MAP kinase pathways in yeast For matingand more Cell 80 187ndash197
HIBI M LIN A SMEAL T MINDEN A and KARIN M (1993)Identification of an oncoprotein- and UV-responsive protein kinasethat binds and potentiates the c-Jun activation domain Genes Dev7 2135ndash2148
HIRANO M et al (1996) MEK kinase is involved in tumor necro-sis factor alpha-induced NF-kappaB activation and degradation ofIkappaB-alpha J Biol Chem 271 13234ndash13238
HU Y et al (1999) Abnormal morphogenesis but intact IKK activa-tion in mice lacking the IKKalpha subunit of IkappaB kinase Sci-ence 284 316ndash320
IORDANOV M et al (1997) CREB is activated by UVC through ap38HOG-1-dependent protein kinase EMBO J 16 1009ndash1022
ISREAL A (2000) The IKK complex An integrator of all signals thatactivate NF-kappaB Trends Cell Biol 10 129ndash133
ITO CY ADEY N BAUTCH VL and BALDWIN AS (1995)Structure and evolution of the human IKBA gene Genomics 29490ndash495
JIANG CK CONNOLLY D and BLUMENBERG M (1991)Comparison of methods for transfection of human epidermal ker-atinocytes J Invest Dermatol 97 969ndash973
JIANG CK FLANAGAN S OHTSUKI M SHUAI K FREED-BERG IM and BLUMENBERG M (1994) Disease-activatedtranscription factor Allergic reactions in human skin cause nucleartranscription of STAT-91 and induce synthesis of keratin K17 MolCell Biol 14 4759ndash4769
JIANG CK MAGNALDO T OHTSUKI M FREEDBERG IMBERNERD F and BLUMENBERG M (1993) Epidermal growthfactor and transforming growth factor alpha specifically induce theactivation- and hyperproliferation-associated keratins 6 and 16 ProcNatl Acad Sci USA 90 6786ndash6790
KALLUNKI T DENG T HIBI M and KARIN M (1996) c-Juncan recruit JNK to phosphorylate dimerization partners via specificdocking interactions Cell 87 929ndash939
KARIN M (1995) The regulation of AP-1 activity by mitogen-acti-vated protein kinases J Biol Chem 270 16483ndash16486
KARIN M and HUNTER T (1995) Transcriptional control by pro-tein phosphorylation Signal transmission from the cell surface to thenucleus Curr Biol 5 747ndash757
KLAMPFER L LEE TH HSU W VILCEK J and CHEN-KIANG S (1994) NF-IL6 and AP-1 cooperatively modulate the ac-tivation of the TSG-6 gene by tumor necrosis factor alpha and in-terleukin-1 Mol Cell Biol 14 6561ndash6569
KOMINE M et al (2000) Inflammatory versus proliferative pro-cesses in epidermis Tumor necrosis factor alpha induces K6b ker-atin synthesis through a transcriptional complex containing NFkappaB and CEBPbeta J Biol Chem 275 32077ndash32088
KOPP E and GHOSH S (1994) Inhibition of NF-kappa B by so-dium salicylate and aspirin Science 265 956ndash959
KYRIAKIS JM et al (1994) The stress-activated protein kinase sub-family of c-Jun kinases Nature 369 156ndash160
LEE FS HAGLER J CHEN ZJ and MANIATIS T (1997) Ac-tivation of the IkappaB alpha kinase complex by MEKK1 a kinaseof the JNK pathway Cell 88 213ndash222
LEPPA S and BOHMANN D (1999) Diverse functions of JNK sig-naling and c-Jun in stress response and apoptosis Oncogene 186158ndash6162
ADACHI ET AL676
LI D TURI TG SCHUCK A FREEDBERG IM KHITROVG and BLUMENBERG M (2001) Rays and arrays The tran-scriptional program in the response of human epidermal keratinocytesto UVB illumination FASEB J 15 2533ndash2535
LI N and KARIN M (1998) Ionizing radiation and short wavelengthUV activate NF-kappaB through two distinct mechanisms ProcNatl Acad Sci USA 95 13012ndash13017
LIU ZG HSU H GOEDDEL DV and KARIN M (1996) Dis-section of TNF receptor 1 effector functions JNK activation is notlinked to apoptosis while NF-kappaB activation prevents cell deathCell 87 565ndash576
MERCURIO F and MANNING AM (1999) NF-kappaB as a pri-mary regulator of the stress response Oncogene 18 6163ndash6171
MIYAMOTO S and VERMA IM (1995) RelNF-kappa BI kappaB story Adv Cancer Res 66 255ndash292
NESTLE FO MITRA RS BENNETT CF CHAN H andNICKOLOFF BJ (1994) Cationic lipid is not required for uptakeand selective inhibitory activity of ICAM-1 phosphorothioate anti-sense oligonucleotides in keratinocytes J Invest Dermatol 103569ndash575
PETERS RT LIAO SM and MANIATIS T (2000) IKKepsilonis part of a novel PMA-inducible IkappaB kinase complex Mol Cell5 513ndash522
PIERCE JW READ MA DING H LUSCINSKAS FW andCOLLINS T (1996) Salicylates inhibit I kappa B-alpha phos-phorylation endothelial-leukocyte adhesion molecule expressionand neutrophil transmigration J Immunol 156 3961ndash3969
PIERCE JW et al (1997) Novel inhibitors of cytokine-induced Ikap-paBalpha phosphorylation and endothelial cell adhesion molecule ex-pression show anti-inflammatory effects in vivo J Biol Chem 27221096ndash21103
REUNING U et al (1995) Inhibition of NF-kappa B-Rel A expres-sion by antisense oligodeoxynucleotides suppresses synthesis ofurokinase-type plasminogen activator (uPA) but not its inhibitor PAI-1 Nucleic Acids Res 23 3887ndash3893
ROSETTE C and KARIN M (1996) Ultraviolet light and osmoticstress activation of the JNK cascade through multiple growth factorand cytokine receptors Science 274 1194ndash1197
SCHOUTEN GJ et al (1997) IkappaB alpha is a target for the mitogen-activated 90 kDa ribosomal S6 kinase EMBO J 163133ndash3144
SHIMADA T et al (1999) IKK-i a novel lipopolysaccharide-inducible kinase that is related to IkappaB kinases Int Immunol 111357ndash1362
SIMON M and GREEN H (1984) Participation of membrane-asso-ciated proteins in the formation of the cross-linked envelope of thekeratinocyte Cell 36 827ndash834
SIMON MM ARAGANE Y SCHWARZ A LUGER TA andSCHWARZ T (1994) UVB light induces nuclear factor kappa B(NF kappa B) activity independently from chromosomal DNA dam-age in cell-free cytosolic extracts J Invest Dermatol 102 422ndash427
TOBIN D NILSSON M and TOFTGARD R (1996) Ras-inde-pendent activation of Rel-family transcription factors by UVB andTPA in cultured keratinocytes Oncogene 12 785ndash793
TOMIC-CANIC M KOMINE M FREEDBERG IM and BLU-MENBERG M (1998) Epidermal signal transduction and tran-scription factor activation in activated keratinocytes J Dermatol Sci17 167ndash181
WILLIAMS BR (1999) PKR A sentinel kinase for cellular stressOncogene 18 6112ndash6120
WU Z WU J JACINTO E and KARIN M (1997) Molecularcloning and characterization of human JNKK2 a novel Jun NH2-terminal kinase-specific kinase Mol Cell Biol 17 7407ndash7416
YANG DD et al (1997) Absence of excitotoxicity-induced apopto-sis in the hippocampus of mice lacking the Jnk3 gene Nature 389865ndash870
YANKULOV K BLAU J PURTON T ROBERTS S and BENT-LEY DL (1994) Transcriptional elongation by RNA polymeraseII is stimulated by transactivators Cell 77 749ndash759
YIN MJ YAMAMOTO Y and GAYNOR RB (1998) The anti-inflammatory agents aspirin and salicylate inhibit the activity ofI(kappa)B kinase-beta Nature 396 77ndash80
YOSHIDA S et al (2001) The c-Jun NH2-terminal kinase3 (JNK3)gene Genomic structure chromosomal assignment and loss of ex-pression in brain tumors J Hum Genet 46 182ndash187
Address reprint requests toM Blumenberg PhD
The NYU Cancer InstituteNew York University School of Medicine
550 First AveNew York NY 10016
E-mail blumem01mednyuedu
Received for publication June 5 2003 received in revised formJuly 15 2003 accepted August 14 2003
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 677

and by the Anaderm Research Corporation Special thinks goto Dr M Simon for the generous gift of keratinocytes We alsothank members of our laboratory for advice reagents and en-couragement
REFERENCES
ALKALAY I YARON A HATZUBAI A ORIAN ACIECHANOVER A and BEN-NERIAH Y (1995) Stimulation-dependent I kappa B alpha phosphorylation marks the NF-kappa Binhibitor for degradation via the ubiquitin-proteasome pathway ProcNatl Acad Sci USA 92 10599ndash10603
BAEUERLE PA and HENKEL T (1994) Function and activationof NF-kappa B in the immune system Annu Rev Immunol 12141ndash179
BARNES PJ and KARIN M (1997) Nuclear factor-kappaB A piv-otal transcription factor in chronic inflammatory diseases N EnglJ Med 336 1066ndash1071
BEG AA FINCO TS NANTERMET PV and BALDWIN AJR (1993) Tumor necrosis factor and interleukin-1 lead to phos-phorylation and loss of I kappa B alpha A mechanism for NF-kappaB activation Mol Cell Biol 13 3301ndash3310
BENDER K BLATTNER C KNEBEL A IORDANOV M HER-RLICH P and RAHMSDORF HJ (1997) UV-induced signaltransduction J Photochem Photobiol B 37 1ndash17
BENDER K GOTTLICHER M WHITESIDE S RAHMSDORFHJ and HERRLICH P (1998) Sequential DNA damage-indepen-dent and -dependent activation of NF-kappaB by UV EMBO J 175170ndash5181
BROWN K GERSTBERGER S CARLSON L FRANZOSO Gand SIEBENLIST U (1995) Control of I kappa B-alpha proteoly-sis by site-specific signal-induced phosphorylation Science 2671485ndash1488
DELHASE M HAYAKAWA M CHEN Y and KARIN M(1999) Positive and negative regulation of IkappaB kinase activitythrough IKKbeta subunit phosphorylation Science 284 309ndash313
DEacuteRIJARD B et al (1994) JNK1 A protein kinase stimulated byUV light and Ha-Ras that binds and phosphorylates the c-Jun acti-vation domain Cell 76 1025ndash1037
DEVARY Y ROSETTE C DIDONATO JA and KARIN M(1993) NF-kappa B activation by ultraviolet light not dependent ona nuclear signal Science 261 1442ndash1445
DIDONATO JA HAYAKAWA M ROTHWARF DM ZANDIE and KARIN M (1997) A cytokine-responsive IkappaB kinasethat activates the transcription factor NF-kappaB Nature 388548ndash554
FANGER GR GERWINS P WIDMANN C JARPE MB andJOHNSON GL (1997) MEKKs GCKs MLKs PAKs TAKs andtpls Upstream regulators of the c-Jun amino-terminal kinases CurrOpin Genet Dev 7 67ndash74
FEIGE U MORIMOTO RI YAHARA I and POLLA BS eds(1996) Stress-Inducible Cellular Responses (Birkhauser VerlagBasel Switzerland)
FISHER GJ WANG ZQ DATTA SC VARANI J KANG Sand VOORHEES JJ (1997) Pathophysiology of premature skin ag-ing induced by ultraviolet light N Engl J Med 337 1419ndash1428
GHODA L LIN X and GREENE WC (1997) The 90-kDa ribo-somal S6 kinase (pp90rsk) phosphorylates the N-terminal regulatorydomain of IkappaBalpha and stimulates its degradation in vitro JBiol Chem 272 21281ndash21288
GILCHREST B (1995) Photodamage (Blackwell Scientific NewYork)
GRETHER-BECK S BUETTNER R and KRUTMANN J (1997)Ultraviolet A radiation-induced expression of human genes Molec-ular and photobiological mechanisms Biol Chem 378 1231ndash1236
GRILLI M PIZZI M MEMO M and SPANO P (1996) Neuro-protection by aspirin and sodium salicylate through blockade of NF-kappaB activation Science 274 1383ndash1385
GUPTA S et al (1996) Selective interaction of JNK protein kinaseisoforms with transcription factors EMBO J 15 2760ndash2770
HEHNER SP et al (1998) Sesquiterpene lactones specifically in-hibit activation of NF-kappa B by preventing the degradation of Ikappa B-alpha and I kappa B-beta J Biol Chem 273 1288ndash1297
HERSKOWITZ I (1995) MAP kinase pathways in yeast For matingand more Cell 80 187ndash197
HIBI M LIN A SMEAL T MINDEN A and KARIN M (1993)Identification of an oncoprotein- and UV-responsive protein kinasethat binds and potentiates the c-Jun activation domain Genes Dev7 2135ndash2148
HIRANO M et al (1996) MEK kinase is involved in tumor necro-sis factor alpha-induced NF-kappaB activation and degradation ofIkappaB-alpha J Biol Chem 271 13234ndash13238
HU Y et al (1999) Abnormal morphogenesis but intact IKK activa-tion in mice lacking the IKKalpha subunit of IkappaB kinase Sci-ence 284 316ndash320
IORDANOV M et al (1997) CREB is activated by UVC through ap38HOG-1-dependent protein kinase EMBO J 16 1009ndash1022
ISREAL A (2000) The IKK complex An integrator of all signals thatactivate NF-kappaB Trends Cell Biol 10 129ndash133
ITO CY ADEY N BAUTCH VL and BALDWIN AS (1995)Structure and evolution of the human IKBA gene Genomics 29490ndash495
JIANG CK CONNOLLY D and BLUMENBERG M (1991)Comparison of methods for transfection of human epidermal ker-atinocytes J Invest Dermatol 97 969ndash973
JIANG CK FLANAGAN S OHTSUKI M SHUAI K FREED-BERG IM and BLUMENBERG M (1994) Disease-activatedtranscription factor Allergic reactions in human skin cause nucleartranscription of STAT-91 and induce synthesis of keratin K17 MolCell Biol 14 4759ndash4769
JIANG CK MAGNALDO T OHTSUKI M FREEDBERG IMBERNERD F and BLUMENBERG M (1993) Epidermal growthfactor and transforming growth factor alpha specifically induce theactivation- and hyperproliferation-associated keratins 6 and 16 ProcNatl Acad Sci USA 90 6786ndash6790
KALLUNKI T DENG T HIBI M and KARIN M (1996) c-Juncan recruit JNK to phosphorylate dimerization partners via specificdocking interactions Cell 87 929ndash939
KARIN M (1995) The regulation of AP-1 activity by mitogen-acti-vated protein kinases J Biol Chem 270 16483ndash16486
KARIN M and HUNTER T (1995) Transcriptional control by pro-tein phosphorylation Signal transmission from the cell surface to thenucleus Curr Biol 5 747ndash757
KLAMPFER L LEE TH HSU W VILCEK J and CHEN-KIANG S (1994) NF-IL6 and AP-1 cooperatively modulate the ac-tivation of the TSG-6 gene by tumor necrosis factor alpha and in-terleukin-1 Mol Cell Biol 14 6561ndash6569
KOMINE M et al (2000) Inflammatory versus proliferative pro-cesses in epidermis Tumor necrosis factor alpha induces K6b ker-atin synthesis through a transcriptional complex containing NFkappaB and CEBPbeta J Biol Chem 275 32077ndash32088
KOPP E and GHOSH S (1994) Inhibition of NF-kappa B by so-dium salicylate and aspirin Science 265 956ndash959
KYRIAKIS JM et al (1994) The stress-activated protein kinase sub-family of c-Jun kinases Nature 369 156ndash160
LEE FS HAGLER J CHEN ZJ and MANIATIS T (1997) Ac-tivation of the IkappaB alpha kinase complex by MEKK1 a kinaseof the JNK pathway Cell 88 213ndash222
LEPPA S and BOHMANN D (1999) Diverse functions of JNK sig-naling and c-Jun in stress response and apoptosis Oncogene 186158ndash6162
ADACHI ET AL676
LI D TURI TG SCHUCK A FREEDBERG IM KHITROVG and BLUMENBERG M (2001) Rays and arrays The tran-scriptional program in the response of human epidermal keratinocytesto UVB illumination FASEB J 15 2533ndash2535
LI N and KARIN M (1998) Ionizing radiation and short wavelengthUV activate NF-kappaB through two distinct mechanisms ProcNatl Acad Sci USA 95 13012ndash13017
LIU ZG HSU H GOEDDEL DV and KARIN M (1996) Dis-section of TNF receptor 1 effector functions JNK activation is notlinked to apoptosis while NF-kappaB activation prevents cell deathCell 87 565ndash576
MERCURIO F and MANNING AM (1999) NF-kappaB as a pri-mary regulator of the stress response Oncogene 18 6163ndash6171
MIYAMOTO S and VERMA IM (1995) RelNF-kappa BI kappaB story Adv Cancer Res 66 255ndash292
NESTLE FO MITRA RS BENNETT CF CHAN H andNICKOLOFF BJ (1994) Cationic lipid is not required for uptakeand selective inhibitory activity of ICAM-1 phosphorothioate anti-sense oligonucleotides in keratinocytes J Invest Dermatol 103569ndash575
PETERS RT LIAO SM and MANIATIS T (2000) IKKepsilonis part of a novel PMA-inducible IkappaB kinase complex Mol Cell5 513ndash522
PIERCE JW READ MA DING H LUSCINSKAS FW andCOLLINS T (1996) Salicylates inhibit I kappa B-alpha phos-phorylation endothelial-leukocyte adhesion molecule expressionand neutrophil transmigration J Immunol 156 3961ndash3969
PIERCE JW et al (1997) Novel inhibitors of cytokine-induced Ikap-paBalpha phosphorylation and endothelial cell adhesion molecule ex-pression show anti-inflammatory effects in vivo J Biol Chem 27221096ndash21103
REUNING U et al (1995) Inhibition of NF-kappa B-Rel A expres-sion by antisense oligodeoxynucleotides suppresses synthesis ofurokinase-type plasminogen activator (uPA) but not its inhibitor PAI-1 Nucleic Acids Res 23 3887ndash3893
ROSETTE C and KARIN M (1996) Ultraviolet light and osmoticstress activation of the JNK cascade through multiple growth factorand cytokine receptors Science 274 1194ndash1197
SCHOUTEN GJ et al (1997) IkappaB alpha is a target for the mitogen-activated 90 kDa ribosomal S6 kinase EMBO J 163133ndash3144
SHIMADA T et al (1999) IKK-i a novel lipopolysaccharide-inducible kinase that is related to IkappaB kinases Int Immunol 111357ndash1362
SIMON M and GREEN H (1984) Participation of membrane-asso-ciated proteins in the formation of the cross-linked envelope of thekeratinocyte Cell 36 827ndash834
SIMON MM ARAGANE Y SCHWARZ A LUGER TA andSCHWARZ T (1994) UVB light induces nuclear factor kappa B(NF kappa B) activity independently from chromosomal DNA dam-age in cell-free cytosolic extracts J Invest Dermatol 102 422ndash427
TOBIN D NILSSON M and TOFTGARD R (1996) Ras-inde-pendent activation of Rel-family transcription factors by UVB andTPA in cultured keratinocytes Oncogene 12 785ndash793
TOMIC-CANIC M KOMINE M FREEDBERG IM and BLU-MENBERG M (1998) Epidermal signal transduction and tran-scription factor activation in activated keratinocytes J Dermatol Sci17 167ndash181
WILLIAMS BR (1999) PKR A sentinel kinase for cellular stressOncogene 18 6112ndash6120
WU Z WU J JACINTO E and KARIN M (1997) Molecularcloning and characterization of human JNKK2 a novel Jun NH2-terminal kinase-specific kinase Mol Cell Biol 17 7407ndash7416
YANG DD et al (1997) Absence of excitotoxicity-induced apopto-sis in the hippocampus of mice lacking the Jnk3 gene Nature 389865ndash870
YANKULOV K BLAU J PURTON T ROBERTS S and BENT-LEY DL (1994) Transcriptional elongation by RNA polymeraseII is stimulated by transactivators Cell 77 749ndash759
YIN MJ YAMAMOTO Y and GAYNOR RB (1998) The anti-inflammatory agents aspirin and salicylate inhibit the activity ofI(kappa)B kinase-beta Nature 396 77ndash80
YOSHIDA S et al (2001) The c-Jun NH2-terminal kinase3 (JNK3)gene Genomic structure chromosomal assignment and loss of ex-pression in brain tumors J Hum Genet 46 182ndash187
Address reprint requests toM Blumenberg PhD
The NYU Cancer InstituteNew York University School of Medicine
550 First AveNew York NY 10016
E-mail blumem01mednyuedu
Received for publication June 5 2003 received in revised formJuly 15 2003 accepted August 14 2003
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 677

LI D TURI TG SCHUCK A FREEDBERG IM KHITROVG and BLUMENBERG M (2001) Rays and arrays The tran-scriptional program in the response of human epidermal keratinocytesto UVB illumination FASEB J 15 2533ndash2535
LI N and KARIN M (1998) Ionizing radiation and short wavelengthUV activate NF-kappaB through two distinct mechanisms ProcNatl Acad Sci USA 95 13012ndash13017
LIU ZG HSU H GOEDDEL DV and KARIN M (1996) Dis-section of TNF receptor 1 effector functions JNK activation is notlinked to apoptosis while NF-kappaB activation prevents cell deathCell 87 565ndash576
MERCURIO F and MANNING AM (1999) NF-kappaB as a pri-mary regulator of the stress response Oncogene 18 6163ndash6171
MIYAMOTO S and VERMA IM (1995) RelNF-kappa BI kappaB story Adv Cancer Res 66 255ndash292
NESTLE FO MITRA RS BENNETT CF CHAN H andNICKOLOFF BJ (1994) Cationic lipid is not required for uptakeand selective inhibitory activity of ICAM-1 phosphorothioate anti-sense oligonucleotides in keratinocytes J Invest Dermatol 103569ndash575
PETERS RT LIAO SM and MANIATIS T (2000) IKKepsilonis part of a novel PMA-inducible IkappaB kinase complex Mol Cell5 513ndash522
PIERCE JW READ MA DING H LUSCINSKAS FW andCOLLINS T (1996) Salicylates inhibit I kappa B-alpha phos-phorylation endothelial-leukocyte adhesion molecule expressionand neutrophil transmigration J Immunol 156 3961ndash3969
PIERCE JW et al (1997) Novel inhibitors of cytokine-induced Ikap-paBalpha phosphorylation and endothelial cell adhesion molecule ex-pression show anti-inflammatory effects in vivo J Biol Chem 27221096ndash21103
REUNING U et al (1995) Inhibition of NF-kappa B-Rel A expres-sion by antisense oligodeoxynucleotides suppresses synthesis ofurokinase-type plasminogen activator (uPA) but not its inhibitor PAI-1 Nucleic Acids Res 23 3887ndash3893
ROSETTE C and KARIN M (1996) Ultraviolet light and osmoticstress activation of the JNK cascade through multiple growth factorand cytokine receptors Science 274 1194ndash1197
SCHOUTEN GJ et al (1997) IkappaB alpha is a target for the mitogen-activated 90 kDa ribosomal S6 kinase EMBO J 163133ndash3144
SHIMADA T et al (1999) IKK-i a novel lipopolysaccharide-inducible kinase that is related to IkappaB kinases Int Immunol 111357ndash1362
SIMON M and GREEN H (1984) Participation of membrane-asso-ciated proteins in the formation of the cross-linked envelope of thekeratinocyte Cell 36 827ndash834
SIMON MM ARAGANE Y SCHWARZ A LUGER TA andSCHWARZ T (1994) UVB light induces nuclear factor kappa B(NF kappa B) activity independently from chromosomal DNA dam-age in cell-free cytosolic extracts J Invest Dermatol 102 422ndash427
TOBIN D NILSSON M and TOFTGARD R (1996) Ras-inde-pendent activation of Rel-family transcription factors by UVB andTPA in cultured keratinocytes Oncogene 12 785ndash793
TOMIC-CANIC M KOMINE M FREEDBERG IM and BLU-MENBERG M (1998) Epidermal signal transduction and tran-scription factor activation in activated keratinocytes J Dermatol Sci17 167ndash181
WILLIAMS BR (1999) PKR A sentinel kinase for cellular stressOncogene 18 6112ndash6120
WU Z WU J JACINTO E and KARIN M (1997) Molecularcloning and characterization of human JNKK2 a novel Jun NH2-terminal kinase-specific kinase Mol Cell Biol 17 7407ndash7416
YANG DD et al (1997) Absence of excitotoxicity-induced apopto-sis in the hippocampus of mice lacking the Jnk3 gene Nature 389865ndash870
YANKULOV K BLAU J PURTON T ROBERTS S and BENT-LEY DL (1994) Transcriptional elongation by RNA polymeraseII is stimulated by transactivators Cell 77 749ndash759
YIN MJ YAMAMOTO Y and GAYNOR RB (1998) The anti-inflammatory agents aspirin and salicylate inhibit the activity ofI(kappa)B kinase-beta Nature 396 77ndash80
YOSHIDA S et al (2001) The c-Jun NH2-terminal kinase3 (JNK3)gene Genomic structure chromosomal assignment and loss of ex-pression in brain tumors J Hum Genet 46 182ndash187
Address reprint requests toM Blumenberg PhD
The NYU Cancer InstituteNew York University School of Medicine
550 First AveNew York NY 10016
E-mail blumem01mednyuedu
Received for publication June 5 2003 received in revised formJuly 15 2003 accepted August 14 2003
UV-SPECIFIC SIGNAL TRANSDUCTION IN KERATINOCYTES 677





























