Embed Size (px)
Citation preview

The stimulatory effect of PYY3-36 on gonadotropin secretion is potentiated
in fasted rats
L. Pinilla1, R. Fernández-Fernández1, E. Vigo, V.M. Navarro, J. Roa, J.M. Castellano, R.
Pineda, M. Tena-Sempere and E. Aguilar
Department of Cell Biology, Physiology and Immunology. University of Córdoba, 14004.
Córdoba, Spain
Running Head: PYY3-36 and gonadotropins in adult rats
Key words: gonadotropin-releasing hormone, luteinizing hormone, follicle-stimulating
hormone, fasting, pituitary
Corresponding author: Enrique Aguilar
Physiology Section. Department of Cell Biology, Physiology and
Immunology. Faculty of Medicine, University of Córdoba
Avda. Menéndez Pidal s/n
14004 Córdoba, SPAIN
FAX: +34-957-218288
E-mail: [email protected]
1 Equally contributed to this work and should be considered as joint first authors
* This work was supported by grants BFI 2000-0419-CO3-03 and BFI 2002-00176 from
DGESIC (Ministerio de Ciencia y Tecnología, Spain)
Articles in PresS. Am J Physiol Endocrinol Metab (January 3, 2006). doi:10.1152/ajpendo.00469.2005
Copyright © 2006 by the American Physiological Society.

2
ABSTRACT
Development and normal function of the reproductive axis requires a precise degree of
body energy stores. Polypeptide YY3-36 (PYY3-36) is a gastrointestinal secreted molecule recently
shown to be involved in the control of food intake with agonistic activity upon neuropeptide Y
(NPY) receptor subtypes Y2 and Y5. Notably, PYY3-36 has been recently demonstrated as
putative regulator of gonadotropin secretion in the rat. However, the “reproductive” facet of this
factor remains to be fully elucidated. In this context, we report herein our analyses of the
influence of the nutritional status on the effects of PYY3-36 upon GnRH and gonadotropin
secretion. The major findings of our study are: (1) the stimulatory effect of central administration
of PYY3-36 on LH secretion was significantly enhanced after fasting, and blocked by a GnRH
antagonist; (2) besides central effects, PYY3-36 elicited LH and FSH secretion directly at the
pituitary level; a response which is also augmented by fasting; (3) PYY3-36 inhibited GnRH
secretion by hypothalamic fragments from male rats fed ad libitum, whereas a significant
stimulatory effect was observed after fasting; (4) the increase in the gonadotropin
responsiveness to PYY3-36 in fasting was not associated with changes in the expression of Y2
and Y5 receptor genes at hypothalamus and/or pituitary. In conclusion, our study extends our
previous observations suggesting a relevant, mostly stimulatory, role of PYY3-36 in the control of
gonadotropin secretion. Strikingly, such an effect was significantly enhanced by fasting.
Considering the proposed decrease in PYY3-36 levels after fasting, the possibility that reduced
PYY3-36 secretion might contribute to defective function of the gonadotropic axis after food
deprivation merits further investigation.

3
INTRODUCTION
Although it is known that conditions of negative energy balance are frequently linked to
lack of puberty onset and reproductive failure, at the moment the mechanisms involved in fitting
the reproductive function to body energy stores remain incompletely defined. Compelling
evidence has recently demonstrated that central and peripheral endocrine signals governing
energy homeostasis, such as the adipocyte-derived hormone leptin, the stomach-derived
hormone ghrelin, orexins and neuropeptide Y (NPY), are also involved in the control of
reproductive function by acting at different levels of hypothalamic-pituitary-gonadal axis.
In this context it has been demonstrated that NPY, a member of the pancreatic
polypeptide family (54), is involved in the control of food intake, reproductive function and
pituitary secretion (26, 27, 30, 33). In rats, central administration of NPY advances puberty (43),
while immunoneutralization of NPY reduced the magnitude of the LH surge during the afternoon
of first proestrus (42). A facilitatory role of NPY on the onset of puberty has been also reported
in the female rhesus monkey (23) and in chicks (18). Secretion of NPY to portal vasculature is
increased on the afternoon of proestrus and served to amplify the actions of GnRH in initiating
the preovulatory surges of LH and probably FSH (53). Acute intracerebroventricular (i.c.v.)
administration of NPY stimulated LH release in ovariectomized rats primed with ovarian steroids
(4). These excitatory effects have been shown to be the consequence of an increase in
secretion and effectiveness of GnRH (5, 14). In contrast, chronic treatment with the peptide
decreased FSH, LH and testosterone secretion (47, 9). NPY hypersecretion is observed in
genetically obese and sterile hyperphagic rodents, which demonstrated the inverse relationship
between chronic NPY secretion and reproductive function (15).
NPY exerts its actions throughout at least five receptor subtypes (16). Development of
selective agonists/antagonists for different receptors and utilization of knock-out animals have
improved our understanding about the role of different receptor subtypes for NPY in the control
of reproductive axis, although at the moment the characterization of the specific role of the
different receptor subtypes is scarce. Recent experiments have shown that the NPY Y1 receptor
inhibits the gonadotrope axis (22) and that its blockade accelerates the onset of puberty (49). In
addition, NPY Y4 receptors have been involved in the NPY effects on LH release (25) and

4
experimental evidence suggests that the inhibition of LH secretion exerted by central
administration of NPY in the rats is predominantly mediated by Y5 receptors (51).
Polypeptide YY 3-36 (PYY3-36), a hormone from gastrointestinal origin structurally
related with NPY and agonist of receptor subtypes Y2 and Y5 (36), has been recently proposed
as a putative anorexigenic signal involved in the control of food intake (3, 24, 50). Conflicting
results on the repeatability of the effects of PYY3-36 in terms of body weight control have been
also published (13, 60). Although the potential role of PYY3-36 in fitting the reproductive function
to body energy stores is still poorly characterized, recent data suggested its involvement in the
control of reproductive axis. Thus, in addition to the reported presence of PYY3-36 in placenta
(62), recent data from our laboratory indicated that PYY3-36 stimulates in vitro FSH and LH
secretion by pituitaries from prepubertal female and male rats and inhibited in vitro GnRH
secretion selectively in males (17). In addition, infusion of PYY3-36 into the lateral ventricle
rapidly inhibited estrous behavior in ovariectomized steroid-primed hamsters (35), and
decreased LH secretion in prepubertal male rats (17).
It is well known that fasting strongly inhibits gonadotropin secretion (6, 7). Considering
that PYY3-36 stimulates in vitro gonadotropin secretion (17), and that PYY3-36 secretion is
depressed during fasting (59), we hypothesized that decreased secretion of PYY3-36 during food
deprivation might be involved in the suppressive effect on pituitary-gonadal function. To test this
hypothesis, we have analyzed the in vivo and in vitro effects of PYY3-36 on GnRH and
gonadotropin secretion after fasting.

5
MATERIAL AND METHODS
Animals and drugs
Wistar rats born in our laboratory were kept under controlled conditions of light (12 h
light: 12 h darkness, lights on at 07.00 h) and temperature (22º C) with free access to pelleted
food (Pacsa Sanders, Seville, Spain) and tap water. Experiments were carried out in adult (90-
100 days) animals. The vaginal smears of adult females were monitored daily and only those
rats exhibiting two or more regular cycles were used. PYY3-36 was purchased from Bachem
(Barcelona, Spain). GnRH antagonist (GnRH-ANT) was ORG.30276 (Ac-D-p-Cl-Phe-D-p-Cl-
Phe-D-Trp-Ser-Tyr-D-Arg-Leu-Arg-Prol-D-Ala-NH2 CH3·COOH) and was purchased from
Organon International (Oss, The Netherlands). For in vivo experiments, PYY3-36 and GnRH-ANT
were dissolved in saline immediately before use, whereas for in vitro experiments PYY3-36, was
dissolved in Dulbecco’s Modified Eagle’s Medium (DMEM) (BioWhittaker; Verviers, Belgium)
immediately before use. Doses of drugs were selected on the basis of previous studies (17, 57).
Experimental designs
Experimental procedures were approved by the Córdoba University Ethical Committee
for animal experimentation and were conducted in accordance with the European Union
normative for care and use of experimental animals. The number of animals per experimental
group is provided in the figure legends. Experiments were carried out between 10.00 and 12.00
h. Special caution was taken to avoid any stressing influences upon the experimental animals
(all the animals were handled daily for a week before the experiment and humanely killed by the
same person and the different drugs were injected at random). The following experiments were
conducted:
In vivo experiments
In the first set of experiments, and to characterize the possible effects of fasting in the
effects of PYY3-36 on gonadotropin secretion, adult (90 days) males were submitted to a 4-days-
period of absolute restriction of food. Control animals were fed ad libitum. The animals were
i.c.v. injected with 3 nmol/rat of PYY3-36 or vehicle. The procedure of i.c.v. injection was as
previously described (48). Briefly, animals were implanted, two days before PYY3-36
administration, with i.c.v. cannulae under light ether anesthesia. To allow delivery of PYY3-36 into
the lateral cerebral ventricle, the cannulae were lowered to a depth of 4 mm beneath the

6
surface of the skull; the insert point was 1 mm posterior and 1.2 mm lateral to bregma. Animals
were humanely killed 15, 30 and 60 min. after injection and trunk blood samples were collected.
To analyze whether the stimulatory effect of PYY3-36 on gonadotropin secretion was
exerted throughout an increase in GnRH release, adult male rats submitted to a 4-days period
of absolute restriction of food and corresponding control groups were subcutaneously (s.c.)
injected with GnRH-ANT (5 mg/kg/rat) 48 and 24 h before i.c.v. administration of PYY3-36 (3
nmol/rat). Blood samples were obtained by decapitation 15, 30 and 60 min. after PYY3-36
injection.
In vitro experiments
Adult cyclic female and male rats were submitted to a 4-day period of absolute
restriction of food. Control animals received food ad libitum. Thereafter, the animals were
humanely killed (the control females being in metestrus) and hypothalami and pituitaries
removed to analyze the effects of PYY3-36 on GnRH and gonadotropin secretion. Hypothalamic
samples were dissected out as described in detail elsewhere (45), by a horizontal cut of about 2
mm depth with the following limits: 1 mm anteriorly from the optic chiasma, the posterior border
of mamillary bodies and the hypothalamic fisures. Hypothalamus were placed in scintillation
vials and incubated in 500 µl of DMEM in a Dubnoff shaker incubator under an atmosphere of
95% O2 and 5% CO2 at 37.5º C. After a 30-min. preincubation, the medium was removed and
hypothalamic fragments were challenged for 45 min with PYY3-36 (10-6 M) or DMEM alone. At
the end of the incubation period, medium samples were boiled for 30 min. to inactivate
endogenous protease activity and stored at -80º C until used for GnRH determinations. Anterior
pituitaries were halved and placed in scintillation vials. After 1 h. of preincubation, the medium
was replaced by fresh medium alone or medium containing PYY3-36 (10-8 and 10-6 M). Samples
were collected at 60 and 120 min. of the incubation period for LH and FSH determinations.
Finally, to analyze whether the increase in the responsiveness to gonadotropin at PYY3-
36 in fasted rats can be mediated by an increase in the NPY Y2 and Y5 receptor subtypes, adult
male were submitted to a 4-days period of absolute restriction of food. Control animals were fed
ab libitum. Thereafter, animals (9-12 animals/group) were humanely killed by decapitation and
pituitary and hypothalamus were immediately dissected (as above described), snap frozen in
liquid nitrogen and stored at -80º C until use for RNA isolation and analysis.

7
LH, FSH, GnRH and leptin measurements by specific RIAs
After centrifugation (1600 g at 4º C for 20 min.), serum was collected, frozen and stored
at –20º C until use. The concentrations of LH and FSH were measured in 5-50 µL by a double-
antibody method using RIA kits supplied by National Institute of Diabetes and Digestive and
Kidney Diseases (Bethesda, MD.USA). Rat-LH-I-10 and FSH-I-9 was labeled with 125I by the
chloramine T method and hormone concentrations were expressed using a reference
preparation LH-RP3 and FSH-RP2 as standard. Intra- and inter-assay variations were,
respectively, 8 and 10%, for LH and 6 and 9% for FSH. The sensitivities of the assay were 75
and 400 pg/mL for LH and FSH, respectively. In addition, GnRH concentrations in the
incubation media from hypothalamic explants were measured in 100-µL aliquots using a
commercial RIA kit purchased from Peninsula Laboratories Inc (Bachem group, San Carlos,
CA) following the instructions of the manufacturer. The sensitivity of the assay was 1 pg/tube.
All samples of each experiment were measured in the same assay. Serum leptin concentrations
were measured in control and fasted rats using a commercial kit from LINCO Research (St.
Charles, MI, USA), following the instructions of the manufacturer. The sensitivity of the assay
was 0.05 ng/tube, while the intra-assay coefficient of variation was below 5%.
RNA Analysis by RT-PCR
Hypothalamic and pituitary expression of NPY Y2 and NPY-Y5 receptor mRNAs was
assessed by semi-quantitative RT-PCR. Total mRNA was isolated from tissue samples using
the single-step, acid guanidinum thiocyanate-phenol-chloroform extraction method, followed by
DNase I treatment (12). For amplification of the target genes, the following primer pairs were
used: NPY-Y2-sense (nt 375-398; 5′-GGT GCC CTA TGC CCA GGG TCT GGC-3′) and NPY-
Y2-antisense (nt 530-509; 5′-GCG CTG ACA CCC CAC GCC AGG C-3′) for amplification of a
156-bp fragment of rat NPY-Y2 receptor cDNA; and NPY Y5-sense (nt 131-153; 5′-GGT CCT
GCT CCT GCC GCC ACC GC-3′) and NPY Y5-antisense (nt 274-253; 5′-CTT GTT AAA ATG
CTC CTC AAG C-3′) for amplification of a 144-bp fragment of rat NPY-Y5 receptor cDNA.
These oligo-primers were synthesized according to the published rat cDNA sequences of NPY-
Y2 and NPY-Y5 (GenBank accession nº NM-023968 and NM-012869, respectively). In addition,
to provide an appropriate internal control, parallel amplification of a 241-bp fragment of S11
ribosomal protein mRNA was carried out in each sample, using the primer pair S11-sense (nt

8
11/32; 5′-CAT TCA GAC GGA GCG TG TTA C-3′) and S11-antisense (nt 231/250; 5′-TGC ATC
TTC ATC TTC GTC AC-3′).
For amplifications of the targets, RT and PCR were run in two separate steps.
Furthermore, to enable appropriate amplification in the exponential phase for each target, PCR
amplification of specific signal and S11 ribosomal protein transcripts was carried out in separate
reactions with different number of cycles (see below), but using similar amounts of the
corresponding cDNA templates, generated in single RT reactions, as previously described (55,
58). Briefly, equal amounts of total RNA (2 µg) were heat denatured and reverse transcribed by
incubation at 42º C for 90 min with 12.5 U avian myeloblastosis virus (AMV) RT (Promega,
Madison, WI, USA), 20 U ribonuclease inhibitor RNasin (Promega), 200 mM deoxy-NTP
mixture, and 1 nM specific and internal control antisense primers in a final volume of 30 µL of 1x
AMV-RT buffer. The reactions were terminated by heating at 97º C for 5 min. and cooling on
ice, followed by dilution of the RT cDNA samples with nuclease-free H2O (final volume, 60 µL).
For semi-quantitative PCR, 10 µL-aliquots of the cDNA samples (equivalent to 650 ng total RNA
input) were amplified in 50 µL of 1x PCR buffer in the presence of 2.5 U Taq DNA polymerase
(Promega), 200 nM deoxy-NTP mixture, and the appropriate primer pairs (1 nM of each primer).
PCR reactions consisted in a first denaturing cycle at 97º C for 5 min., followed by a variable
number of cycles of amplification (n= 36 cycles for NPY-Y2 and –Y5; n= 26 cycles for RP-S11)
defined by denaturation at 96º C for 1.5 min., annealing for 1.5 min, and extension at 72º C for 3
min. A final extension cycle of 72º C for 15 min. was included. Annealing temperature was
adjusted for each target: 58º C for NPY-Y2 receptor and S11, and 61.5º C for NPY-Y5 receptor.
Different numbers of cycles were tested to optimize amplification in the exponential phase of
PCR. On this basis, the numbers of PCR cycles indicated above were chosen for further semi-
quantitative analysis targets and RP-S11 internal control.
PCR-generated DNA fragments were resolved in Tris-borate buffered 1.5% agarose
gels and visualized by ethidium bromide staining. Specificity of PCR products was confirmed by
direct sequencing (Central Sequencing Service, University of Córdoba). Quantification of
intensity of RT-PCR signals was carried out by densitometric scanning and values of the
specific targets were normalized to those of internal control to express arbitrary units of relative

9
expression. In all assays, liquid controls and reactions without RT were included, yielding
negative amplification.
Presentation of data and statistics
Values are expressed as means ± SEM. When relevant, integrated LH and FSH
secretory responses were calculated as the area under the curve (AUC), obtained following the
trapezoidal rule, over the 60-min period after administration of PYY3-36. Differences between
groups were analyzed using Student t test or two-way ANOVA followed by Student-Newman-
Keuls multiple range test.
RESULTS
Effects of PYY3-36 on in vivo gonadotropin secretion
In adult males, a 4-day period of absolute restriction of food induced a significant
decrease in body weight (282 ± 10 g vs 325 ± 5 g in controls) and in serum LH and leptin
concentrations (Table 1). I.c.v. administration of PYY3-36 (3 nmol/rat) significantly stimulated LH
and FSH secretion in controls fed ad libitum and in fasted animals (Table 1), without affecting
serum leptin concentrations. Interestingly, the LH response in fasted animals was higher than in
the control group, either when response was estimated by absolute LH levels reached after
PYY3-36 administration or by the fold-increase in serum LH concentrations (Table 1). To obtain
information about the time-course of the stimulatory effect of PYY3-36 on gonadotropin secretion,
blood samples were obtained 15, 30 and 60 min. after peptide administration in adult controls
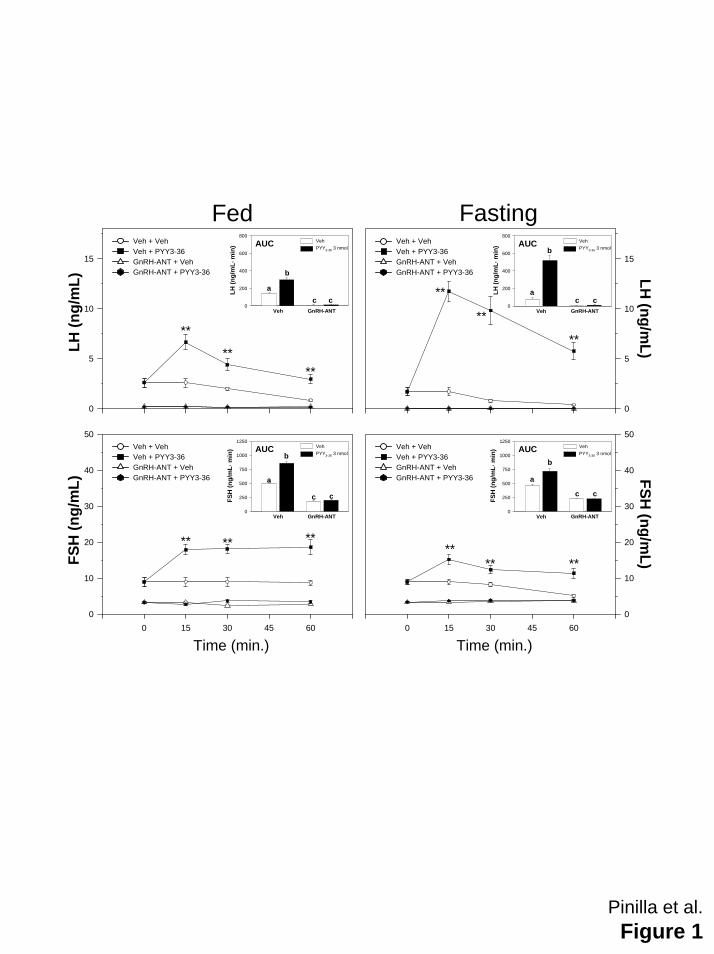
and fasted male rats. Results obtained showed that the stimulatory effect on gonadotropin
secretion remain evident, at least, 60 min after i.c.v. administration of PYY3-36 (Figure 1).
Treatment with a GnRH-ANT abolished completely the stimulatory effect of PYY3-36 on LH and
FSH secretion in controls and fasted rats (Figure 1).
Effects of PYY3-36 on in vitro GnRH secretion in adult male rats
PYY3-36 (10-6 M) significantly inhibited GnRH secretion by hypothalamic fragments from
control males fed ad libitum (Figure 2). Food deprivation during 4 days reduced, in the limits of
statistical significance, basal GnRH secretion (11.07 ± 2.21 pg/fragment vs 16.42 ± 1.40). In
contrast with the inhibitory effect in ad libitum control animals, PYY3-36 significantly stimulated
GnRH by hypothalamic samples from fasted animals (Figure 2).

10
Effects of PYY3-36 on in vitro gonadotropin secretion
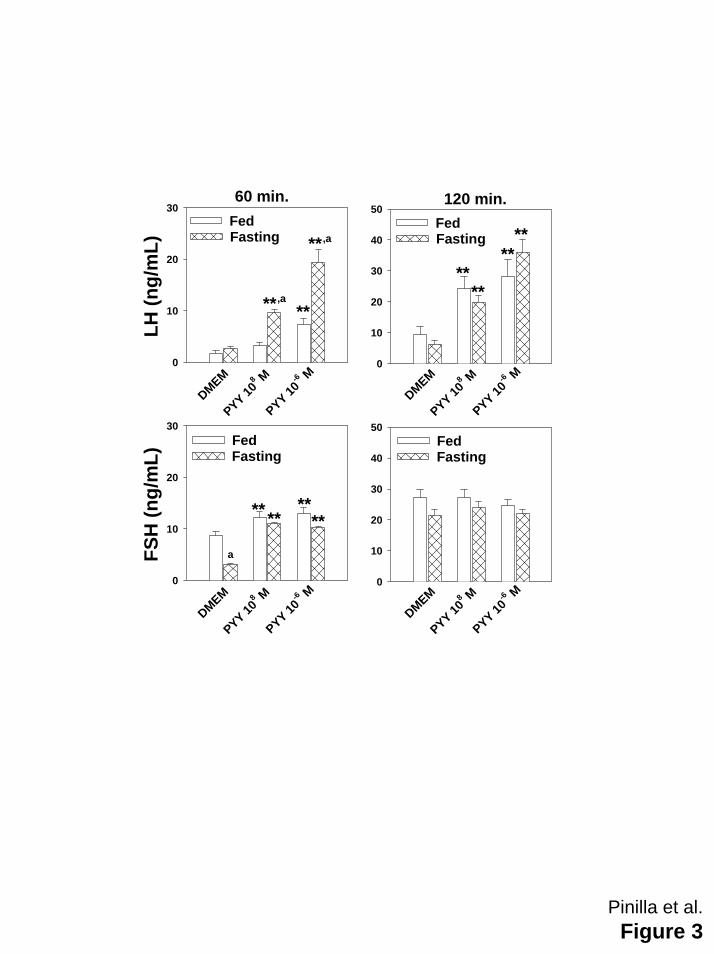
Males: In hemi-pituitaries from control rats fed ad libitum, 10-6 M PYY3-36 significantly increased
LH secretion at 60 and 120 min. of incubation period, whereas the dose of 10-8M only increased
LH secretion at 120 min. (Figure 3). After a 4-day period of absolute food restriction, both doses
of PYY3-36 significantly increased LH secretion at 60 and 120 min. of incubation period, the
responses being significantly greater than in control group (Figure 3). FSH secretion was
similarly stimulated at 60 min. of the incubation period in controls fed ad libitum and fasted rats
by both doses of PYY3-36, and the effect disappeared at 120 min of the incubation period
(Figure 3).
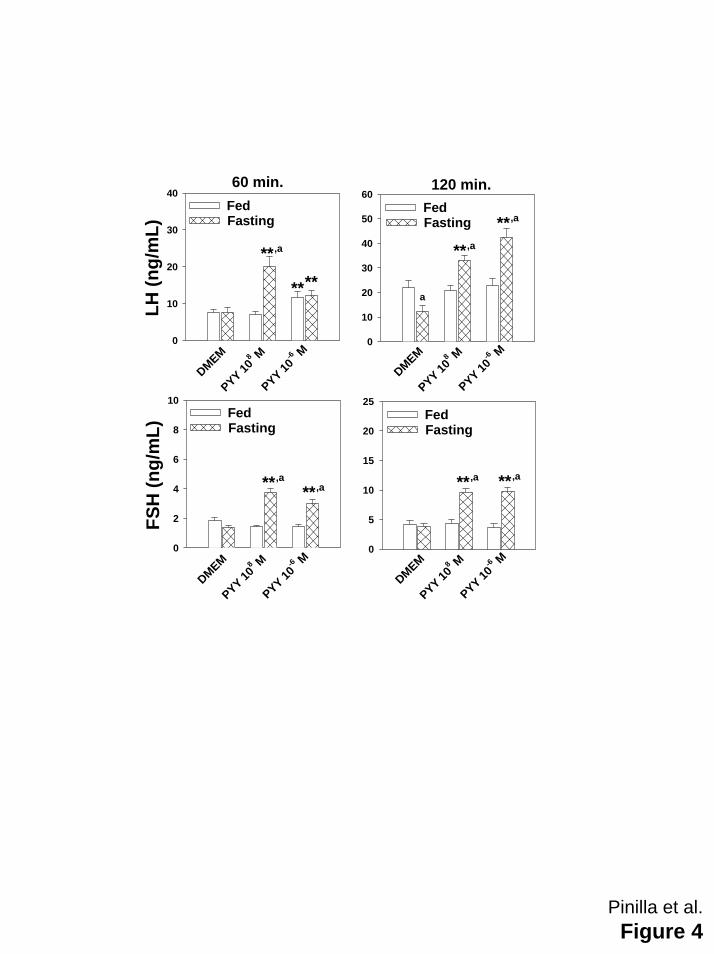
Females: In hemi-pituitaries from control metestrous females fed ad libitum, PYY3-36 was unable
to stimulate LH or FSH secretion, except for a significant increase in LH observed at 60 min of
the incubation period in presence of higher dose of PYY3-36 (Figure 4). However, after a 4-day
period of absolute food restriction, LH and FSH secretion was significantly stimulated by both
doses of PYY3-36 (Figure 4).
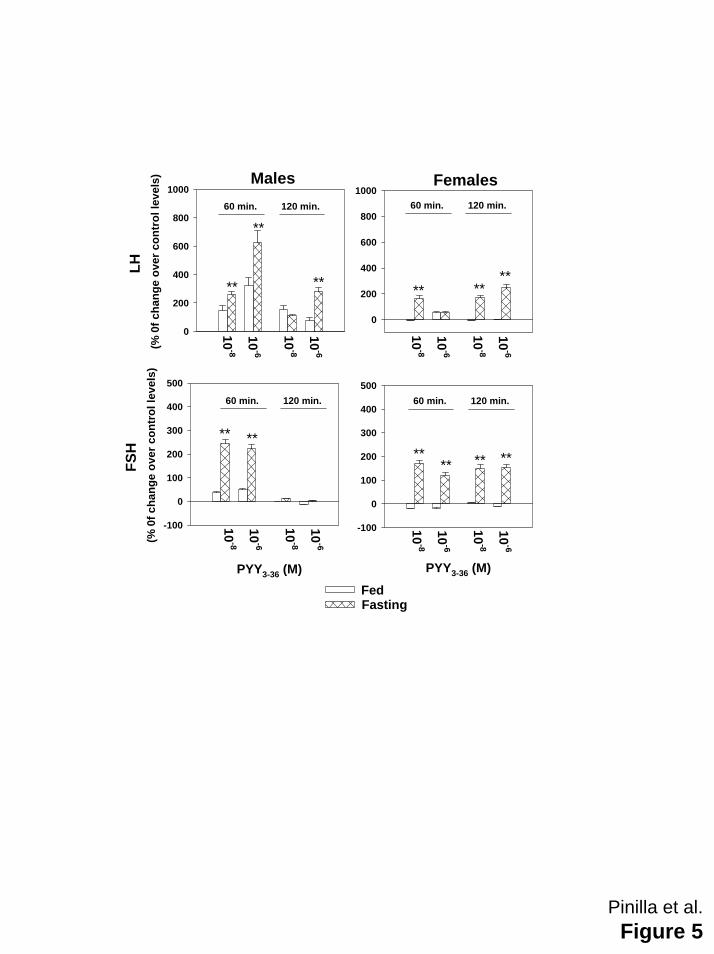
In order to compare the effectiveness of PYY3-36 in controls fed ad libitum and fasted rats,
results were expressed as changes over the concentrations measured in absence of PYY3-36.
Figure 5 clearly evidences that the overall response was greater in fasted animals than in ad
libitum control animals.
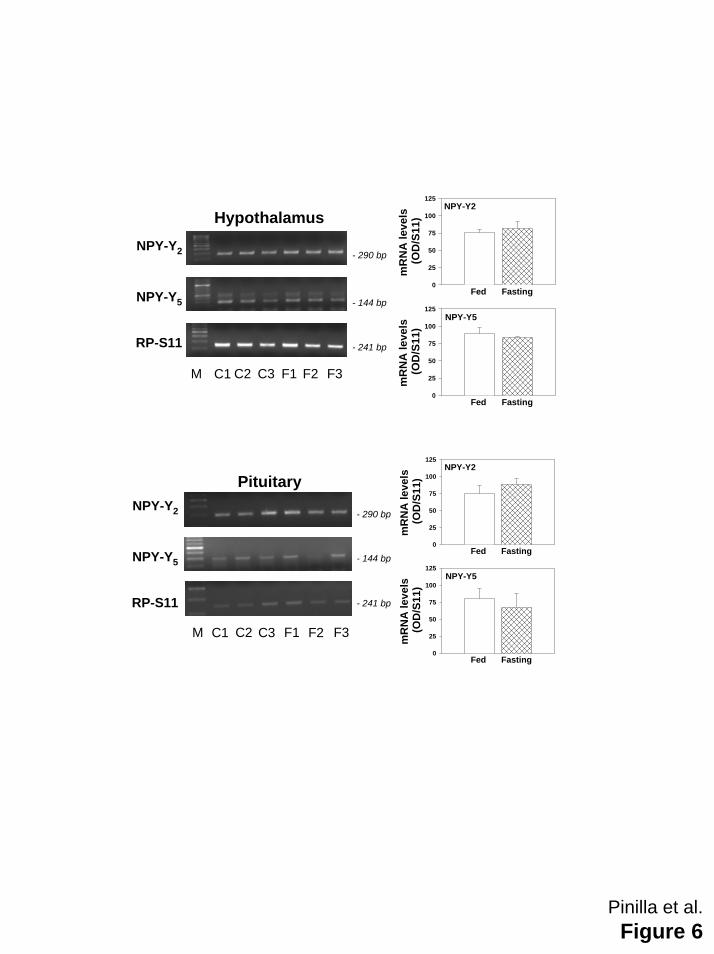
Hypothalamic and pituitary mRNA expression of NPY Y2 and Y5 receptors in fed and fasted rats
RT-PCR analysis using specific primer pairs demonstrated that the expression of the
genes encoding NPY receptors Y2 and Y5 at the hypothalamus and pituitary of adult rats was
similar in ad libitum control and fasted rats (Figure 6).

11
DISCUSSION
The relationship between energy balance, body weight and reproductive function is well
known after the pioneer works of Frish group (20, 21). Different hormonal signals such as leptin,
ghrelin and orexins seem to be involved in the fine coordination between body weight and
reproductive function (2, 52, 56).
The key signal in the regulation of reproductive function is the pulsatile secretion of
GnRH which controls gonadotropin release. GnRH release is primarily controlled by central and
peripheral signals (28, 29, 41). NPY, secreted from hypothalamus to hypothalamic-pituitary
portal system, was characterized as “an unique member of the family of gonadotropic releasing
hormones“ (19), due to their multiple effects on hypothalamic GnRH secretion and in the
pituitary responsiveness to GnRH. Selective agonists and antagonists for the different NPY
receptors and different knock-out animals have been used to elucidate the role of different NPY
receptors in the control of feeding behavior and reproductive function. Recently, it has been
demonstrated that PYY3-36, agonist of NPY Y2 and Y5 receptors, is secreted from
gastrointestinal tract depending on the nutritional status of the animal. A regulatory loop
involving this peptide has been proposed: after food intake, PYY3-36 is secreted and inhibits
feeding behavior, whereas its release is depressed in underfeeding conditions (3, 59).
In previous experiments, we demonstrated the presence of NPY Y2 and Y5 receptor
subtypes in hypothalamus and pituitary and the complex actions of PYY3-36 at hypothalamic and
pituitary level (17). PYY3-36 directly stimulated gonadotropin secretion by prepubertal pituitaries
and significantly inhibited GnRH secretion by hypothalamic explants in males. The systemic and
central administration of the peptide exerted changes in serum concentrations of gonadotropins,
the effects being sexually dimorphic (17). The present studies extend and reinforce our previous
hypothesis that PYY3-36 is involved in the control of gonadotropin secretion. Specifically, herein
we have analyzed the effects of PYY3-36 in the control of gonadotropin and GnRH secretion
during fasting.
Periods of chronic undernutrition, as well as short periods of fasting, have been shown
to adversely impact mammalian reproduction in various ways. Fasting suppresses GnRH
release and hence pituitary LH, FSH and testosterone secretion, which are reversed by pulsatile
GnRH substitution (6, 7, 31, 61). The neural pathways that relay information on insufficient

12
energy stores to GnRH neurons included many signals. Different neuropeptides and
neurotransmitters [such as NPY, adrenaline via alpha 2-adrenergic receptors, Corticotropin
releasing hormone (CRH), leptin, ciliary neurotropic factor (CNTF), gamma-aminobutyric acid
(GABA) and opioids] have been implicated in the effects of fasting on pituitary secretion (8, 32,
38, 39, 40, 44).
Acute administration of NPY stimulated GnRH/LH release (4, 5, 14), whereas
continuous NPY receptor activation results in suppression of gonadotropin secretion (9, 15, 47),
which explains that in experimental conditions that up-regulate NPY synthesis and release,
such as diabetes, the reproductive function is also impaired (31). Acute and chronic food
deprivation and undernutrition stimulate hypothalamic NPY synthesis, storage and release in
the hypothalamus (31, 46, 61). Consequently, fasting-induced up-regulation of NPY secretion
may diminish gonadotropin secretion. This hypothesis is sustained by the finding that
counteracting the fasting-induced NPY up-regulation experimentally with those naturally-
occurring compounds that inhibit the hypothalamic NPY system, such as CNTF, it is possible to
attenuate the fasting-induced suppression of pituitary LH secretion (32). In summary,
undernutrition induces an up-regulation of NPY system in arcuate nucleus which, in turn,
suppresses GnRH and gonadotropin secretion.
In addition to such phenomenon, we hypothesized that changes in PYY3-36 secretion
should also be involved in the adverse effects of fasting on reproductive function. PYY3-36
release is regulated by food intake, increasing after meal intake and decreasing during fasting
(3, 59). On the basis of this secretory profile, reduction of gonadotropin secretion during fasting
would be due, at least in part, to the decrease in PYY3-36 release. To confirm this hypothesis, we
have analyzed the effects in vivo and in vitro of PYY3-36 on gonadotropin secretion in controls
fed ad libitum and fasted rats. In adult male rats, deprivation of food during 4 days significantly
decreased serum LH concentrations (Table 1). I.c.v. administration of PYY3-36 stimulated more
efficiently LH secretion in fasted than in control animals fed ad libitum, and serum LH
concentrations reached levels higher than obtained in control males. The increased in vivo
efficiency of PYY3-36 in fasted animals was confirmed in vitro. These results support our
hypothesis that depression of gonadotropin secretion during fasting in adult animals may be

13
due, at least partially, to the decrease in the stimulatory effect carried out by PYY3-36 and that
the efficiency of this signal was increased during food deprivation.
To analyze the participation of GnRH on the stimulatory effect of i.c.v. administration of
PYY3-36 on gonadotropin release in adult fasted rats, we analyzed the effects of PYY3-36 after
pretreatment with a GnRH-ANT. Results obtained demonstrated that the blockade of GnRH
action abolished the stimulatory effect of PYY3-36 on gonadotropin secretion. In addition, we
analyzed in vitro the GnRH release in presence of PYY3-36. Two important findings were
observed: PYY 3-36 inhibited GnRH release by hypothalamus obtained from males fed ad
libitum, which agrees with data obtained in prepubertal males (17), whereas a clear stimulatory
effect was observed after fasting. The reasons for the switch from an inhibitory action of PYY3-36
on GnRH release in control males to the stimulatory effect observed after fasting are unknown
at the present. However, this finding is in strong agreement with the potentiation of PYY3-36
effectiveness on gonadotropin secretion after fasting. Overall it is evident that the control of
GnRH /gonadotropin release by PYY3-36 is critically dependent of the nutritional status. Since
the effects of PYY3-36 on GnRH are opposite in fasted and ad lib fed rats, the stimulatory effect
on LH observed in both groups of animals suggests either that PYY3-36 modulated the
hypothalamic release of different signals other than GnRH involved in the control of LH, or a
possible direct pituitary effect (via hypothalamic-pituitary portal system) of the peptide icv
delivered.
The present data also indicated that PYY3-36 directly increases in both sexes LH and
FSH secretion at the pituitary level, in accordance with data obtained in prepubertal rats (17).
The stimulatory effect is enhanced after fasting. The mechanisms of the stimulatory action of
PYY3-36 at the pituitary level and the reasons for its increase effectiveness during fasting are
unknown. It has been previously shown that the NPY Y2 and Y5 receptor mRNA expression
patterns in hypothalamus do not change during fasting, in contrast with the increase observed
for Y1 mRNA expression (11, 63), but the expression in pituitary have not been studied. To
analyze whether, in our experimental paradigm, fasting increased the expression of genes
encoding Y2 and Y5 receptors, we studied their expression levels in hypothalamus and pituitary.
The fact that similar levels of mRNA were detected after fasting argues against the possibility
that an up-regulation of NPY Y2 and Y5 receptors in hypothalamus and/or pituitary might explain

14
the increase in the effectiveness of PYY3-36 in fasted rats. Nonetheless, the possibility that
fasting-induced changes in receptor number of signaling might take place at a post-
trascriptional level cannot be ruled out on the basis of our present data.
Leptin stimulates GnRH and LH secretion (64), and serum leptin levels decreased
during fasting (1). Different experimental approaches demonstrated the cross-talk between
leptin and NPY in the control of reproductive function. For example: the inhibition of
gonadotropic axis in leptin-deficient mice is attenuated by removal of NPY Y1 receptor and
acceleration of puberty by leptin is largely facilitated in mice deficient for NPY Y1 receptors (22).
It is conceivable that suppression of leptin input to GnRH neurons and pituitary gonadotrops in
fasted rats may enhance the responsiveness to other stimulating signals such as PYY 3-36. If this
hypothesis is correct, the increased hypothalamic/pituitary responsiveness to PYY 3-36 after
fasting could be secondary to the decrease in serum leptin levels. Since underrnutrition induces,
in addition to changes observed in serum leptin concentrations, a decrease in serum levels of
insulin and an increase in those of corticosterone, and both hormones are involved in the
control of LH secretion (10, 34, 37), the possibility that these changes could be involved in the
potentiation of stimulatory effect of PYY 3-36 after fasting merits further investigation
In conclusion, present experiments demonstrated that the stimulatory effect of PYY3-36
on gonadotropin secretion is enhanced after fasting. This phenomenon included an increase in
gonadotropin responsiveness to PYY3-36 in vivo and in vitro, as well as a clear stimulatory effect
on GnRH release. We proposed that the inhibition of pituitary gonadotropin secretion which
occurs in undernutrition could be mediated, at least in part, by the decrease in PYY 3-36
secretion and is reversed by exogenous administration of the peptide.
ACKNOWLEDGMENTS
This work was supported by grants BFI 2000-0419-CO3 and BFI 2002-00176 from
DGESIC (Ministerio de Ciencia y Tecnología, Spain). The collaboration of A. Mayen is
recognized.

15
REFERENCES
1. Aubert ML, Pierroz DD, Gruaz NM, d'Alleves V, Vuagnat BA, Pralong FP, Blum WF, Sizonenko
PC. Metabolic control of sexual function and growth: role of neuropeptide Y and leptin. Mol Cell Endocrinol
140:107-113, 1998.
2. Barreiro ML, Tena-Sempere M. Ghrelin and reproduction: a novel signal linking energy status and
fertility? Mol Cell Endocrinol 226:1-9, 2004.
3. Batterham RL, Cowley MA, Small CJ, Herzog H, Cohen MA, Dakin CL, Wren AM, Brynes AE,
Low MJ, Ghatei MA, Cone RD, Bloom SR. Gut hormone PYY 3-36 physiologically inhibits food intake.
Nature 418:650-654, 2002.
4. Bauer-Dantoin AC, McDonald JK, Levine JE. Neuropeptide Y potentiates LHRH-induced LH
secretion only under conditions leading to preovulatory LH surges. Endocrinology 131:2946-2952
5. Bauer-Dantoin AC, Tabesh B, Norgle JR, Levine JE. RU486 administration blocks neuropeptide Y
potentiation of luteinizing-hormone (LH)-releasing hormone-induced LH surges in proestrous rats.
Endocrinology 133:2418-2423, 1993.
6. Bergendahl M, Perheentupa A, Huhtaniemi I. Effect of short-term starvation on reproductive
hormone gene expression, secretion and receptor levels in male rats. J Endocrinol 121:409-417, 1989.
7. Bergendahl M, Perheentupa A, Huhtaniemi I. Starvation-induced suppression of pituitary-testicluar
function in rats is reversed by pulsatile gonadotropin-releasing hormone. Biol Reprod 44:413-419, 1991.
8. Cagampang FR, Ohjura S, Tsukamura H, Coen CW, Ota K, Maeda K. Alpha 2-adrenegic
receptors are involved in the suppression of luteinizing hormone release during acute fasting in the
ovariectomized estradiol-primed rats. Neuroendocrinology 56:724-728, 1992.
9. Catzeflis C, Pierroz DD, Rohner-Jeanrenaud F, Rivier JE, Sizonenko PC, Aubert ML.
Neuropeptide Y administered chronically into the lateral ventricle profoundly inhibits both the gonadotropic
and the somatotropic axis in intact adult female rats. Endocrinology 132:224-234, 1993.
10. Chang LL, Kau MM, Wum WS, Ho LT, Wang PS. Effects of fasting on corticosterone production by
zona fasciculate-reticularis cells in ovariectomized rats. J Invest Med 50:86-94, 2002.
11. Cheng X, Broberger C, Tong Y, Yongtao X, Yu G, Zhang X, Hokfelt T. Regulation of expression
of neuropeptide Y Y1 and Y2 receptors in the arcuate nucleus of fasted rats. Brain Res 792:89-96, 1998.
12. Chomczynski P, Sacchi N. Single-step method of RNA isolation by acid guanidinium thiocianate-
phenol-chloroform extraction. Anal Biochem 162:156-159, 1987.
13. Coll AP, Challis BG, O´Rahilly S. Peptide YY 3-36 and satiety: clarity or confusion? Endocrinology
145:2582-2584, 2004.

16
14. Crowley WR, Shah GV, Carroll BL, Kennedy D, Dockter ME, Kalra SP. Neuropeptide-Y enhances
luteinizing hormone (LH)-releasing hormone-induced LH release and elevations in cytosolic Ca2+ in rat
anterior pituitary cells: evidence for involvement of extracellular Ca2+ influx through voltage-sensitive
channels. Endocrinology 127:1487-1494, 1990.
15. Dryden S, Pickavana L, Frankish HM, Williams G. Increased neuropeptide Y secretion in the
hypothalamic paraventricular nucleus of obese (fa/fa) Zucker rats. Brain Res 690:185-188, 1995.
16. Dumont Y, Jacques D, Bouchard P, Quirion R. Species differences in the expression and
distribution of the neuropeptide Y Y1, Y2, Y4 and Y5 receptors in rodents, guinea pig, and primate brains.
J Comp Neurol 402:372-384, 1998.
17. Fernández-Fernández R, Aguilar E, Tena-Sempere M, Pinilla L. Effects of polypeptide YY3-36
upon luteinizing hormone-releasing hormone and gonadotropin secretion in prepubertal rats: in vivo and in
vitro studies. Endocrinology 146:1403-1410, 2005.
18. Fraley GS, Kuenzel WJ. Precocious puberty in chicks (Gallus domesticus) induced by central
injections of neuropeptide Y. Life Sci 52:1649-1656, 1993.
19. Freeman ME. Editorial: Neuropeptide Y: A unique member of the constellations of gonadotropin-
releasing hormones. Endocrinology 133:2411-2412, 1993.
20. Frisch R, McArthur J. Menstrual cycles: fatness as a determinant of minimum weight for height
necessary for their maintenance or onset. Science 169:397-399, 1974.
21. Frisch R, Revelle R. Height and weight at menarche: a hypothesis of critical body weights and
adolescent events. Science 169:397-399, 1970.
22. Gonzales C, Voirol MJ, Giacomini M, Gaillard RC, Pedrazzini T, Pralong FP. The neuropeptide Y
Y1 receptor mediates NPY-induced inhibition of the gonadotrope axis under poor metabolic conditions.
FASEB J 18:137-139, 2004.
23. Gore AC, Mitsushima D, Teresawa E. A possible role of neuropeptide Y in the control of the onset
puberty in female rhesus monkey. Neuroendocrinology 58:23-34, 1993.
24. Halatchev IG, Ellacott KLJ, Fan W, Cone RD. Peptide YY 3-36 inhibits food intake in mice through
a melanocortin-4-receptor-independent mechanism. Endocrinology 145:2585-2590, 2004.
25. Jain MR, Pu S, Kalra PS, Kalra SP. Evidence that stimulation of two modalities of pituitary
luteinizing hormone release in ovarian steroid-primed ovariectomized rats may involve neuropeptide Y Y1
and Y4 receptors. Endocrinology 140:5171-5177, 1999.
26. Kalra SP, Crowley WR. Neuropeptide Y: a novel neuroendocrine peptide in the control of pituitary
hormone secretion and its relation to luteinizing hormone. Front Neuroendocrinol 13:1-46, 1992.

17
27. Kalra SP, Dube MG, Pu S, Xu B, Horvath TL, Kalra PS. Interacting appetite-regulating pathways in
the hypothalamic regulation of body weight. Endocr Rev 20:68-100, 1999.
28. Kalra SP, Kalra PS. Do testosterone and estradiol-17 beta enforce inhibition or stimulation of
luteinizing hormone-releasing hormone secretion? Biol Reprod 41:559-570, 1989.
29. Kalra SP, Kalra PS. Neural regulation of luteinizing hormone secretion in the rat. Endocr Rev 4:311-
351, 1983.
30. Kalra SP, Kalra PS. NPY an endearting journey in search of a neurochemical on/off switch for
appetite, sex and reproduction. Peptides 25:465-471, 2004.
31. Kalra SP, Kalra PS. Nutritional infertility in the role of the interconnected hypothalamic neuropeptide
Y-galanin-opioid network. Front Neuroendocrinol 17:371-401, 1996..
32. Kalra SP, Xu B, Dube MG, Moldawer LL, Martin D, Kalra PS. Leptin and ciliary neurotropic factor
(CNTF) inhibit fasting-induced suppression of luteinizing hormone release in rats: role of neuropeptide Y.
Neurosci Lett 240:45-49, 1998.
33. Kalra SP. Appetite and body weight regulation: is all in the brain? Neuron 19:227-230, 1997.
34. Kamel F, Kubajak CL. Modulation of gonadotropin secretion by corticosterone: interaction with
gonadal steroids and mechanism of action. Endocrinology 121:561-568, 1987.
35. Keene AC, Jones JE, Wade GN, Corp ES. Forebrain sites of NPY action on estrous behavior in
Syrian hamsters. Physiol Behav 78:711-716, 2003.
36. Keire DA, Mannon P, Kobayashi M, Walsh JH, Solomon TE, Reeve JR Jr. Primary structures of
PYY, [Pro(34)] PYY, and PYY 3-36 confer different conformations and receptor selectivity. Am J Physiol
Gastrointest and Liver Physiol 279:G126-131, 2000.
37. Kovacs P, Parlow AF, Karkanias GB. Effect of centrally administered insulin on gonadotropin-
releasing hormone neuron activity and luteinizing hormone surge in the diabetic female rat.
Neuroendocrinology 76:357-365, 2002.
38. Leonhardt S, Shahab M, Luft H, Wuttke W, Jarry H. Reduction of luteinizing hormone secretion
induced by long-term feed restriction in male rats is associated with increased expression of GABA-
synthesizing enzymes without alterations of GnRH expression. J Neuroendocrinol 11:613-619, 1999.
39. Maeda K, Cagampang FR, Coen CW, Tsukamura H. Involvement of the catecholaminergic input to
the paraventricular nucleus and corticotrophin-releasing hormone in the fasting-induced suppression of
luteinizing hormone release in female rats. Endocrinology 134:1718-1722, 1994.
40. Maeda K, Tsukamura H. Neuroendocrine mechanism mediated fasting-induced suppression of
luteinizing hormone secretion in female rats. Acta Neurobiol Exp (Wars) 56:787-796, 1996.

18
41. McCann SM, Krulich L, Cooper KJ, Kalra PS, Kalra SP, Libertun C, Negro-Vilar A, Orias R,
Ronnekleiv O, Fawcett CP. Hypothalamic control of gonadotrophin and prolactin secretion, implications
for fertility control. J Reprod Fertil Suppl 20:43-59, 1973.
42. Minami S, Frautschy SA, Plotsky PM, Sutton SW, Sarkar DK. Facilitatory role of neuropeptide Y
on the onset of puberty: effect of immunoneutralization of neuropeptide Y on the release of luteinizing
hormone and luteinizing-hormone releasing hormone. Neuroendocrinology 52:112-115, 1990.
43. Minami S, Sarkar DK. Central administration of neuropeptide Y induces precocious puberty in
female rats. Neuroendocrinology 56:930-934, 1992.
44. Nagatani S, Guthikonda P, Thompson RC, Tsukamura H, Maeda KI, Foster DL. Evidence for
GnRH by leptin: leptin administration prevents reduced pulsatile LH secretion during fasting.
Neuroendocrinology 67:370-376, 1998.
45. Navarro VM, Fernández-Fernández R, Castellano JM, Roa J, Mayén A, Barreiro ML, Gaytán F,
Aguilar E, Pinilla L, Diéguez C, Tena-Sempere M. Advanced vaginal opening and precocious activation
of the reproductive axis by kiss-1 peptide, the endogenous ligand of GPR54. J Physiol 561:379-386, 2004.
46. Pelleymounter MA, Cullen MJ, Baker MB, Hecht R, Winters D, Boone T, Collins F. Effects of
the obese gene product on the body weight regulation in ob/ob mice. Science 269:540-543, 1995.
47. Pierroz DD, Catzeflis C, Aebi AC, Rivier JE, Aubert ML. Chronic administration of neuropeptide Y
into the lateral ventricle inhibits both the pituitary-testicular axis and growth hormone and insuline-like
growth factor I secretion in intact adult male rats. Endocrinology 137:3-12, 1996.
48. Pinilla L, González L, Tena Sempere M, Aguilar E. Activation of AMPA receptors inhibits prolactin
and estradiol secretion and delays the onset of puberty in female rats. J Steroid Biochem Mol Biol 75:277-
281, 2000.
49. Pralong FP, Voirol MJ, Giacomini M, Gaillard RC, Grouzmann E. Acceleration of pubertal
development following central blockade of the Y1 subtype of neuropeptide Y receptors. Regul Pept 95:47-
52, 2000.
50. Prasanth KC, Haver AC, Reidelberger RD. Intravenous infusion of peptide YY (3-36) potently
inhibits food intake in rats. Endocrinology 146:1879-1888, 2005.
51. Raposinho PD, Broqua P, Pierroz DD, Hayward A, Dumont Y, Quirion R, Junien JL, Aubert ML.
Evidence that the inhibition of luteinizing hormone secretion exerted by central administration of
neuropeptide Y (NPY) in the rat is predominantly mediated by the NPY Y5 receptor subtype.
Endocrinology 140:4046-4055, 1999.

19
52. Russell SH, Small CJ, Kennedy AR, Stanley SA, Seth A, Murphy KG, Taheri S, Ghatei MA,
Bloom SR. Orexin A interactions in the hypothalamo-pituitary gonadal axis. Endocrinology 142:5294-
5302, 2001.
53. Sutton SW, Toyama TT, Otto S, Plotsky PM. Evidence that neuropeptide Y (NPY) released into the
hypophysial portal circulation participates in priming gonadotropes to the effects of gonadotropin releasing
hormone (GnRH). Endocrinology 123:1208-1210, 1988.
54. Tatemoto K. Neuropeptide Y: complete amino acid sequence of the brain peptide. Proc Natl Acad
Sci USA 79:5485-5489, 1982.
55. Tena-Sempere M, Barreiro ML, González LC, Gaytán F, Zhang FP, Caminos JE, Pinilla L,
Casanueva FF, Diéguez C, Aguilar E. Novel expression and functional role of ghrelin in rat testis.
Endocrinology 143:717-725, 2002.
56. Tena-Sempere M, Barreiro ML. Leptin in male reproduction: the testis paradigm. Mol & Cell
Endocrinol 188:9-13, 2002.
57. Tena-Sempere M, Pinilla L, Aguilar E. Orchidectomy selectively increases follicle-stimulating
hormone secretion in gonadotropin-releasing hormone antagonist-treated rats. Eur J Endocrinol 132:357-
362, 1995.
58. Tena-Sempere M, Pinilla L, Zhang FP, González LC, Huhtaniemi I, Casanueva FF, Diéguez C,
Aguilar E. Developmental and hormonal regulation of leptin receptor (Ob-R) messenger ribonucleic acid
expression in the rat testis. Biol Reprod 64:634-643, 2001.
59. Tovar S, Seoane LM, Caminos JE, Nogueiras R, Casanueva FF, Dieguez C. Regulation of
peptide YY levels by age, hormonal and nutritional status. Obes Res 12:1944-1952, 2004.
60. Tschop M, Castaneda TR, Joost HG, Thone-Reineke C, Ortmann S, Klaus S, Hagan MM,
Chandler PC, Oswald KD, Benoit SC, Seeley RJ, Kinzig KP, Moran TH, Beck-sickinger AG, Koglin N,
Rodgers RJ, Blundell JE, Ishii Y, Beattie AH, Holch P, Allison DB, Raun K, Madsen K, Wulff BS,
Stidsen CE, Birringer M, Kreuzer OJ, Schindler M, Arndt K, Rudolf K, Mark M, Deng XY, Withcomb
DC, Halem H, Taylor J, Dong J, Datta R, Culler M, Craney S, Flora D, Smiley D, Heiman ML. Does gut
hormone PYY 3-36 decrease food intake in rodents? Nature 431:430-431, 2004.
61. Wade GN, Schneider JE. Metabolic levels and reproduction in female mammals. Neurosci Biobehav
Rev 16:235-272, 1992.
62. Xiao Q, Han X, Arany E, Hill D, Challis JR, McDonald TJ. Human placenta and fetal membranes
contain peptide YY1-36 and peptide YY3-36. J Endocrinol 156:485-492, 1998.

20
63. Xu B, Kalra PS, Moldawer LL, Kalra SP. Increased appetite augments hypothalamic NPY Y1
receptor gene expression: effects of anorexigenic ciliary neurotropic factor. Regul Pept 75-76:391-395,
1998.
64. Yu WH, Kimura M, Walczewska A, Karanth S, McCann SM. Role of leptin in hypothalamic-pituitary
function. Proc Natl Acad Sci USA 94:1023-1028, 1997.

21
LEGEND TO FIGURES
Figure 1: Serum concentrations of LH (upper panels) and FSH (lower panels) in male rats,
either fed ad libitum or at fasting, which were s.c. injected with GnRH-ANT (5 mg/kg) or vehicle
48 and 24 h before i.c.v.injection with vehicle or 3 nmol/rat of PYY3-36. Food deprivation was
maintained during 4 days. Values are expressed as means ± SEM (10-12 animals/group). **p≤
0.01 vehicle plus PYY3-36 vs other groups plus PYY3-36. (ANOVA followed by Student -Newman-
Keuls test). In the inside panels, the integrated gonadotropin secretory responses following
central administration of PYY3-36 and peripheral administration of GnRH-ANT (calculated as the
area under the curve-AUC- during the 60-min study-period) in control and fasted rats are
shown. Bars with different superscript letter were statistically differents (ANOVA followed by
Student -Newman-Keuls test)
Figure 2: GnRH released (pg/hypothalamus/45 min.) by hypothalami from adult male rats,
either fed ad libitum or at fasting, incubated in the presence of PYY3-36 (10-6M) or medium
(DMEM) alone. Food deprivation was maintained during 4 days before incubation of
hypothalami. Values are expressed as means ± SEM (10-12 samples/group). **p≤ 0.01 and *p≤
0.05 vs corresponding control (DMEM) group (ANOVA followed by Student-Newman-Keuls
test).
Figure 3: Effects of PYY3-36 (10-8 and 10-6 M) on LH (upper panels) and FSH (lower panels)
secreted by hemi-pituitaries obtained from adult controls fed ad libitum (open bars) or fasting
males (hatched bars). Food deprivation was maintained during 4 days before incubation of
hemi-pituitaries. Values are expressed as means ± SEM (10-12 samples/group). **p≤ 0.01 vs
hemi-pituitaries incubated in absence of PYY3-36; ap≤ 0.01 vs corresponding fed ad libitum
control group (ANOVA followed by Student-Newman-Keuls test).
Figure 4: Effects of PYY3-36 (10-8 and 10-6 M) on LH (upper panels) and FSH (lower panels)
secreted by hemi-pituitaries obtained from adult controls fed ad libitum (open bars) or fasting
female rats (hatched bars). Control females were studied in metestrus. Food deprivation was
maintained during 4 days before incubation of hemi-pituitaries. Values are expressed as means
± SEM (10 samples/group). **p≤ 0.01 vs hemi-pituitaries incubated in absence of PYY3-36; ap≤

22
0.01 vs corresponding fed ad libitum control group (ANOVA followed by Student-Newman-Keuls
test).
Figure 5: Comparison between the effects of PYY3-36 on LH (upper panels) and FSH (lower
panels) secreted by hemi-pituitaries obtained from adult fed ad libitum control (open bars) and
fasting (hatched bars) male and female rats. Results are presented as changes over levels
obtained in the absence of PYY3-36. **p≤ 0.01 vs corresponding fed ad libitum control group
(ANOVA followed by Student-Newman-Keuls test).
Figure 6: Expression of the genes encoding NPY receptors Y2 and Y5 at the hypothalamus
(upper panels) and pituitary (lower panels) from adult male rats fed ad libitum (open bars) or
fasting males (hatched bars). Food deprivation was maintained during 4 days before study.
Representative images of ethidium bromide-stained gel electrophoresis of the specific
amplicons are presented. Three independent samples from controls fed ad libitum (C1-C3) and
fasting rats (F1-F3) are shown. Amplification of a fragment of RP-S11 mRNA served as internal
control. Semi-quantitative values of gene expression levels in the experimental groups are
shown, which are the mean ± SEM of at least three independent determinations. Negative
controls were run in parallel with specific RT-PCR assays and yielded negative amplification
(data not shown).
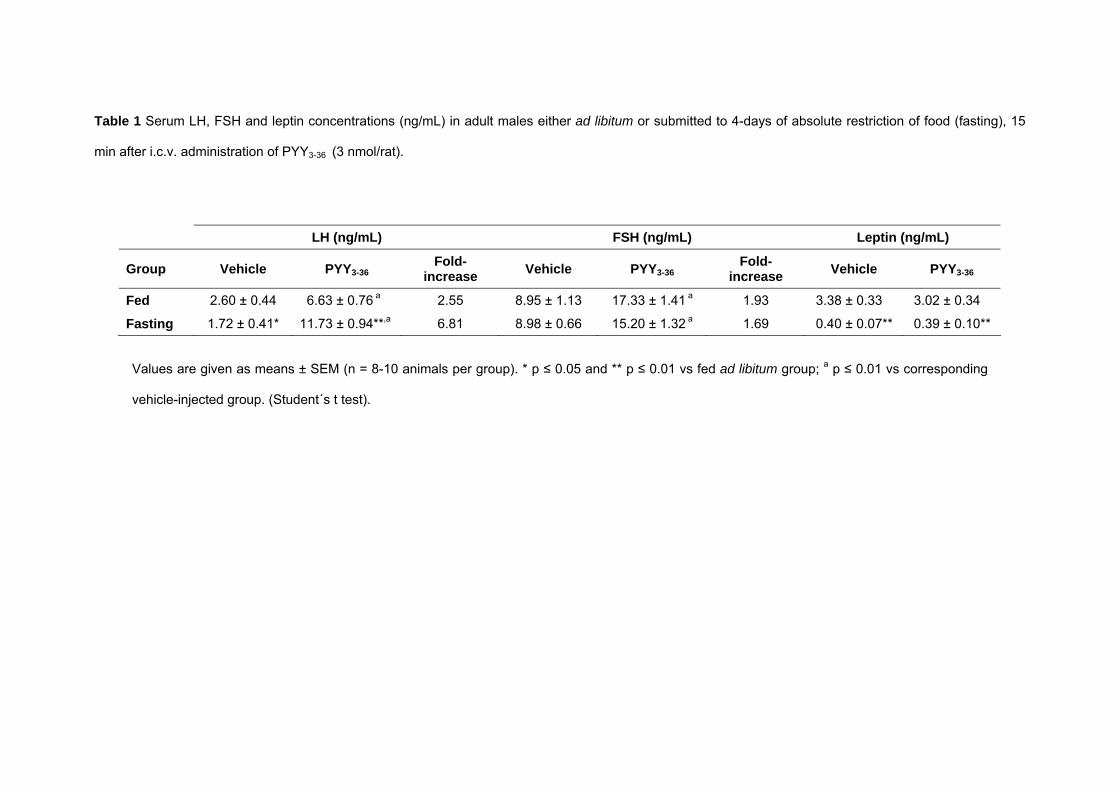
Table 1 Serum LH, FSH and leptin concentrations (ng/mL) in adult males either ad libitum or submitted to 4-days of absolute restriction of food (fasting), 15
min after i.c.v. administration of PYY3-36 (3 nmol/rat).
LH (ng/mL) FSH (ng/mL) Leptin (ng/mL)
Group Vehicle PYY3-36Fold-
increase Vehicle PYY3-36Fold-
increase Vehicle PYY3-36
Fed 2.60 ± 0.44 6.63 ± 0.76 a 2.55 8.95 ± 1.13 17.33 ± 1.41 a 1.93 3.38 ± 0.33 3.02 ± 0.34
Fasting 1.72 ± 0.41* 11.73 ± 0.94**,a 6.81 8.98 ± 0.66 15.20 ± 1.32 a 1.69 0.40 ± 0.07** 0.39 ± 0.10**
Values are given as means ± SEM (n = 8-10 animals per group). * p ≤ 0.05 and ** p ≤ 0.01 vs fed ad libitum group; a p ≤ 0.01 vs corresponding
vehicle-injected group. (Student´s t test).
LH (n
g/m
L)
0
5
10
15
AUCLH
(ng/
mL.
min
)
0
200
400
600
800
b
ac c
Veh GnRH-ANT
VehPYY3-36 3 nmol
**
****
Veh + VehVeh + PYY3-36GnRH-ANT + VehGnRH-ANT + PYY3-36 LH
(ng/mL)
0
5
10
15
AUC
LH (n
g/m
L. m
in)
0
200
400
600
800
b
ac c
Veh GnRH-ANT
VehPYY3-36 3 nmol
**
**
**
Veh + VehVeh + PYY3-36GnRH-ANT + VehGnRH-ANT + PYY3-36
0 15 30 45 60
FSH
(ng/
mL)
0
10
20
30
40
50
AUC
FSH
(ng/
mL.
min
)
0
250
500
750
1000
1250
b
Time (min.)
a
c c
Veh GnRH-ANT
VehPYY3-36 3 nmol
** ** **
Veh + VehVeh + PYY3-36GnRH-ANT + VehGnRH-ANT + PYY3-36
0 15 30 45 60
FSH (ng/m
L)
0
10
20
30
40
50
AUC
FSH
(ng/
mL.
min
)
0
250
500
750
1000
1250
b
Time (min.)
a
c c
Veh GnRH-ANT
VehPYY3-36 3 nmol
**** **
Veh + VehVeh + PYY3-36GnRH-ANT + VehGnRH-ANT + PYY3-36
Fed Fasting
Pinilla et al.Figure 1

GnR
H
(pg/
hypo
thal
amus
/45
min
)
0
10
20
30DMEMPYY3-36 10-6 M
**
*
Fed Fasting
Pinilla et al.Figure 2
0
10
20
30
40
50
DMEM
****
PYY 108 M
PYY 10-6 M
DMEM
PYY 108 M
PYY 10-6 M
** **
FSH
(ng/
mL)
0
10
20
30Fed Fasting
Fed Fasting
** ****
**
a
LH (n
g/m
L)
0
10
20
30
0
10
20
30
40
5060 min.
**
DMEM
**
120 min.
PYY 108 M
PYY 10-6 M
DMEM
PYY 108 M
PYY 10-6 M
Fed Fasting
Fed Fasting
**
****
,a**
,a**
Pinilla et al.Figure 3
FSH
(ng/
mL)
0
2
4
6
8
10
0
5
10
15
20
25
DMEM
PYY 108 M
PYY 10-6 M
DMEM
PYY 108 M
PYY 10-6 M
Fed Fasting
Fed Fasting
,a** ,a**,a** ,a**
LH (n
g/m
L)
0
10
20
30
40
0
10
20
30
40
50
6060 min.
DMEM
**
120 min.
PYY 108 M
PYY 10-6 M
DMEM
PYY 108 M
PYY 10-6 M
Fed Fasting
Fed Fasting
** a
,a** ,a**
,a**
Pinilla et al.Figure 4
0
200
400
600
800
1000
LH
(% 0
f cha
nge
over
con
trol
leve
ls)
0
200
400
600
800
1000Males Females
Fed Fasting
FSH
(%
0f c
hang
e ov
er c
ontr
ol le
vels
)
-100
0
100
200
300
400
500
-100
0
100
200
300
400
500
PYY3-36 (M) PYY3-36 (M)
10-8
10-6
10-8
10-6
10-8
10-6
10-8
10-6
10-8
10-6
10-8
10-6
10-8
10-6
10-8
10-6
60 min. 120 min.
60 min. 120 min.
60 min. 120 min.
60 min. 120 min.
**
**
****
**** ** **
**
**
** **
Pinilla et al.Figure 5
NPY-Y2
NPY-Y5
RP-S11
- 290 bp
- 144 bp
- 241 bp
Hypothalamus
C1 F2C3C2 F1M F3
mR
NA
leve
ls (O
D/S
11)
0
25
50
75
100
125NPY-Y2
mR
NA
leve
ls (O
D/S
11)
0
25
50
75
100
125
Fed Fasting
NPY-Y5
Fed Fasting
- 290 bp
- 144 bp
- 241 bp
Pituitary
mR
NA
leve
ls (O
D/S
11)
0
25
50
75
100
125NPY-Y2
mR
NA
leve
ls (O
D/S
11)
0
25
50
75
100
125
Fed Fasting
NPY-Y5
Fed Fasting
NPY-Y2
NPY-Y5
RP-S11
C2 F1M F3C1 C3 F2
Pinilla et al.Figure 6
























