Embed Size (px)
Citation preview

THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1987 by The American Society of Biological Chemists, Inc.
Vol. 262, No. 13, Issue of May 5, pp. 6318-6322, 1987 Printed in U.S.A.
A Fluorescent Analog of Colcemid, N-(7-Nitrobenz-2-oxa- 1,3-diazol- 4-yl)-colcemid, as a Probe for the Colcemid-binding Sites of Tubulin and Microtubules*
(Received for publication, December 1, 1986)
Toshiaki HiratsukaSg and Toyoki Katol[ From the $Department of Chemistry, Asahikawa Medical College, Asahikawa, Hokkaido 078 and liSugashima Marine Biological Laboratory, School of Science, Nagoya University, Sugashima, Toba, Mie 51 7, Japan
The synthesis and biological testing of the fluores- cent analog of colcemid, N-(7-nitrobenz-2-oxa-1,3- diazol-4-yl)-colcemid (NBD-colcemid), are here de- scribed. NBD-colcemid exhibited a visible absorption maximum at 465 nm and fluoresced in the range of 520-540 nm, highly in environments of low polarity, whereas only slightly in aqueous solution. The addition of NBD-colcemid to bovine brain tubulin was accom- panied by a striking enhancement of fluorescence. The fluorescent titration study suggested a stoichiometric binding of NBD-colcemid to tubulin. Assembled micro- tubules were directly visualized after mixing with NBD-colcemid using a fluorescence microscope. NBD- colcemid reversibly disrupted the metaphase spindles of sea urchin eggs as well as unlabeled colcemid. How- ever, even when the birefringence of spindles was mostly lost, self-quenching properties of the NBD flu- orescence allowed tubulin and its oligomers aggregated in higher concentrations in eggs to be visualized under a fluorescence microscope. The results suggest a wide applicability of NBD-colcemid as a fluorescent probe for studying the interactions of colcemid with tubulin and microtubules, as well as for localizing other col- cemid-binding structures within cells.
Essential to understanding the reaction of a ligand with a protein is knowledge of the properties and location of the ligand-binding sites on the protein. Such knowledge should also facilitate elucidation of the ligand-protein interaction. Biologically active fluorescent drugs and natural products are excellent as ligands in obtaining information of this sort because of the sensitivity to small changes in environment of the binding site as well as the low concentration sufficient to obtain meaningful results. Furthermore, the fluorescent prop- erties can be used to facilitate microscopic visualization of the ligand-binding protein in cells.
Although some drugs and natural products are intrinsically fluorescent, many biologically active fluorescent compounds have been made by the chemical modification of drugs and natural products to introduce fluorescent groups. As a part of our strategy for studying the ligand-protein interactions, we have synthesized fluorescent derivatives of ATP (1-3), GTP (3, 4), and CAMP and cGMP (5) with unmodified base moie-
* The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
5 To whom inquiries and reprint requests should be addressed Dept. of Chemistry, Asahikawa Medical College, Nishikagura 4-5, Asahikawa 078, Japan.
ties of nucleotides. These fluorescent nucleotides maintain specificity and biological activity of the unlabeled nucleotide. Because of the biological and spectroscopic properties, these fluorescent nucleotides have been widely used as substitutes for natural nucleotides in the study of various nucleotide- requiring systems (6-15).
It is well established that the action of the plant alkaloid colchicine on the mitotic spindle and, similarly, many other actions of colchicine on cellular processes are due to the disruption of MTs.’ A large number of investigators have demonstrated that colchicine can bind with high affinity to tubulin, the dimeric protein subunit of MTs, and that tubulin- colchicine complexes inhibit M T polymerization (see Refs. 16 and 17 for reviews). Colcemid (N-deacetyl-N-methylcolchi- cine), a structural analog of colchicine with a slightly modified B ring, also interferes with MT-dependent cell functions (18, 19). Both the drugs have played key roles as tools in the investigation of MT function.
In view of the high specificity of colcemid for tubulin and MTs (18, 19), we had hoped that fluorescence-labeled col- cemid would be useful for tubulin as a colcemid-binding site- directed probe. The present paper is a report of the synthesis and biological testing of the fluorescent derivative, N-(7- nitrobenz-2-oxa-1,3-diazol-4-yl)-colcemid (NBD-colcemid). The NBD fluorophore exhibits excitation and emission max- ima in the ranges of 460-480 and 520-550 nm, respectively (20). This excitation wavelength of the fluorophore is very desirable for design of fluorescence-labeled colcemid since colcemid decomposes upon irradiation with light at a wave- length of 300-400 nm, being converted to a mixture of biolog- ically inactive isomers of colcemid (lumicolcemids (21, 22). Because the NBD fluorophore has a favorably high quantum yield in environments of low polarity and a very low quantum yield in water (20), this fluorophore has long been used as a valuable fluorescent probe to follow the conformational changes of various proteins. In addition, it is smaller than most fluorophores, allowing only minor perturbations in var- ious biological systems to be produced.
Our results indicate that NBD-colcemid retains the speci- ficity and biological activity of unlabeled colcemid, fluorescing strongly when bound to tubulin with excitation and emission maxima at 465 and 530 nm, respectively. Therefore, NBD- colcemid is a very useful tool for studying the interaction between colcemid and tubulin, as well as for localizing other colcemid-binding structures within cells.
The abbreviations used are: MTs, microtubules; NBD, 7-nitro- benz-2-oxa-1,3-diazo1-4-yl; NBD-F, 4-fluoro-7-nitrobenz-2-oxa-1,3- diazole; PIPES, piperazine-N,N’-bis(2-ethanesulfonic acid); EGTA, [ethylenebis(oxyethylenenitrilo)]tetraa~etic acid; ASW, artificial sea- water.
6318
NBD-Colcernid: Fluorescent Probe for Tubulin 63 19
EXPERIMENTAL PROCEDURES
Materials-Colcemid was purchased from Wako Pure Chemicals. NBD-F was from Dojin Co. ATP was from Boehringer Mannehim. GTP was from Yamasa Shoyu Co. TLC (Silica Gel 60 F ~ M , 0.25 mm thick) and preparative layer chromatography (Silica Gel 60, 2 mm thick) plates were from Merck. Other reagents were of reagent or biological research grade.
Protein Preparations-MT proteins from bovine brains were ob- tained by two cycles of temperature-dependent polymerization by the
bly buffer solution (RAB) consisted of 0.1 M PIPES, 1 mM MgSO,, 1 method of Shelanski et al. (23) with some modifications. The reassem-
mM EGTA, and 1 mM GTP, pH 6.9. Tubulin was purified from the MT protein fraction by phosphocellulose column chromatography by the method of Weingarten et al. (24) with a minor modification. These proteins were stored at -80 "C until used.
For the measurements of self-quenching of the NBD fluorophore bound to tubulin, MT proteins (10 mg/ml) were polymerized in RAB at 37 "C for 30 min to form MTs. NBD-colcemid was conjugated to the assembled MT suspension in a 3:l molar ratio of NBD-colcemid to tubulin dimer at 37 'C for 10 min, and then centrifuged at 100,000 X g for 30 min at 30 "C. The resultant fluorescent MT pellets were homogenized with cold RAB without GTP and centrifuged at 100,000 x g for 20 min at 4 "C. The supernatant, MT proteins labeled with NBD-colcemid, was used for the experiments.
Preparation of Sea Urchin Eggs-Gammetes of the sea urchin, Hemicentrotus pulcherimus, were obtained by the injection of 0.5 M KC1 into the coelomic cavity. After insemination, eggs were developed in ASW (MBL formula V, Woods Hole, MA) at 15 "C. NBD-colcemid at a concentration of 6.5 I.LM in ASW was applied to the eggs.
Microscopy-Fluorescence observations were carried out using an Olympus BH-RFL epifluorescence microscope (Olympus Optical Co., Tokyo) equipped with a dichroic mirror (BH-DMV) and a 495-nm cut-off barrier filter. Excitation with violet light (BG.3 + IF405 filters) from a high pressure mercury lamp (Osrum HBO, 100 watts; Munich, West Germany). With this microscope, dark-field images in the same spots were also observed by changing the epiillumination to the transmitting illumination using a dark-field condenser (Olympus
A modified Leitz Ortholux Pol polarizing microscope (E. Leitz, Inc., Rockleigh, NJ) equipped with a set of rectified strain-free optics (Nippon Kogaku K. K., Tokyo) was used for the polarization obser- vations. Images were recorded on Fujichrome DX 400 D for fluores- cence and dark-field, and on Kodak plus-X pan for polarizing one using an Olympus PM-10 camera with an automatic control box (Olympus PM-CBA; Olympus Optical Co., Tokyo).
Spectral Measurements-Absorption spectra were measured with a Shimadzu double beam spectrophotometer Model UV-200.
Fluorescence emission spectra (uncorrected) were recorded with a Hitachi fluorescence spectrophotometer, Model MPF-2A or MPF- 4A, in a 0.5- or 1-cm light-path quartz cell(s). These instruments were operated in a ratio mode. Samples were placed in a temperature- controlled cell holder and maintained at 18 or 25 "C. Excitation wavelengths were 465 or 470 nm. The slit widths on the excitation and emission monochromators were 5-10 nm. Up to 10 p M NBD- colcemid there was less than 6% loss of fluorescence due to the inner filter effects. Excitation spectra (corrected) were measured with a Shimadzu Difference Spectrofluorophotometer RF-503A.
Fluorescence emission spectra of NBD-colcemid bound to tubulin were measured at 18 "C in RAB without GTP. The fluorescence enhancement for bound tubulin was measured at 530 nm by titrating a fixed amount of NBD-colcemid (10 p ~ ) with increasing amounts ot tubulin in a concentration range of 2-64 p ~ . The self-quenching of NBD fluorophore bound to tubulin was measured on solutions of MT proteins labeled with NBD-colcemid ranging in a concentration from 1 to 12 mg/ml.
Protein Determinotion-The concentration of proteins was deter- mined by the method of Lowry et al. (25) using bovine serum albumin as a standard. The molecular weight of tubulin was taken to be 100,000.
TLC-TLC was performed with chloroform/methanol/acetic acid (7686, v/v/v) (Solvent I) or henzene/methanol (3:1, v/v) (Solvent 11). Preparative layer chromatography was performed with Solvent I. Compounds were detected on chromatograms under a Camag Deluxe UV lamp.
Synthesis of NBD-colcemid-NBD-colcemid was synthesized by reaction of the methylamino group of colcemid with NBD-F the colcemid analog is now available from Wako Pure Chemicals Co.
DC, NA 1.33-1.2).
(Osaka, Japan). Colcemid (20 mg) was reacted with NBD-F (25 mg) in 1 ml of N,N-dimethylformamide. The solution was stirred at 30 "C in total darkness. The reaction was monitored with time by TLC, and the reaction was allowed to proceed until colcemid was undetect- able. After 1.5-2 h the mixture was purified by repeated preparative layer chromatography on Merck silica gel plates (20 X 20 cm; 2 mm thick) in Solvent I in a dark room. The product NBD-colcemid migrated as a single orange band (RF = 0.90), whereas unreacted NBD-F migrated more slowly (RF = 0.24). The band of NBD-colcemid was scraped off and eluted with Solvent I. The solid remaining after evaporation was dissolved in absolute ethanol and centrifuged to remove any remaining silica gel. After evaporation of ethanol, NBD- colcemid was stored as a powder in the dark at -20 "C.
NBD-colcemid was homogeneous and free from starting materials based on TLC on silica gel in Solvent I (RF values: NBD-colcemid, 0.90; NBD-F, 0.24; colcemid, 0.26) and in Solvent I1 (RF values: NBD- colcemid, 0.59; NBD-F, 0.17; colcemid, 0.43).
For routine purposes, the stock solutions of NBD-colcemid were prepared at concentrations of 0.5-1.5 mM in ethanol and stored at -20 "C. More dilute solutions in RAB and ASW were prepared just prior to use.
RESULTS AND DISCUSSION
Characterization of NBD-colcemid-The synthesis of NBD- colcemid is simple and straightforward. Following a one-step reaction of colcemid with NBD-F, the reaction mixture was separated by preparative layer chromatography. The product NBD-colcemid is chromatographically pure, as indicated by a single fluorescent spot, and free from starting materials.
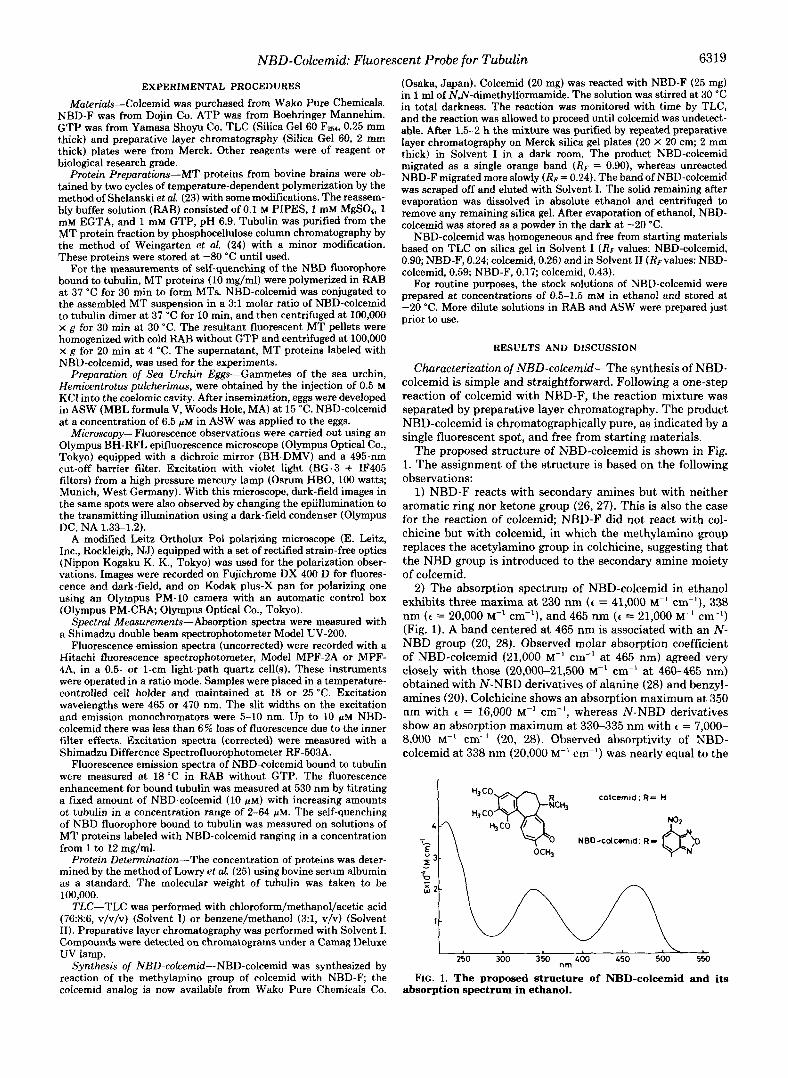
The proposed structure of NBD-colcemid is shown in Fig. 1. The assignment of the structure is based on the following observations:
1) NBD-F reacts with secondary amines but with neither aromatic ring nor ketone group (26, 27). This is also the case for the reaction of colcemid; NBD-F did not react with col- chicine but with colcemid, in which the methylamino group replaces the acetylamino group in colchicine, suggesting that the NBD group is introduced to the secondary amine moiety of colcemid.
2) The absorption spectrum of NBD-colcemid in ethanol exhibits three maxima at 230 nm (e = 41,000 "' cm"), 338 nm ( t = 20,000 M" cm"), and 465 nm ( t = 21,000 M" cm") (Fig. 1). A band centered at 465 nm is associated with an N- NBD group (20, 28). Observed molar absorption coefficient of NBD-colcemid (21,000 M" cm" at 465 nm) agreed very closely with those (20,000-21,500 "' cm" at 460-465 nm) obtained with N-NBD derivatives of alanine (28) and benzyl- amines (20). Colchicine shows an absorption maximum at 350 nm with e = 16,000 M" cm", whereas N-NBD derivatives show an absorption maximum at 330-335 nm with t = 7,000- 8,000 M-' cm" (20, 28). Observed absorptivity of NBD- colcemid at 338 nm (20,000 M" cm") was nearly equal to the
3 : c H J colccmid ; R = H
Y N ' \ OCn,
I . 250 300 350 nm 400 450 500 550
FIG. 1. The proposed structure of NBD-colcemid and its absorption spectrum in ethanol.
6320 NBD-Colcemid: Fluorescent Probe for Tubulin
sum of the values obtained with colchicine and N-NBD deriv- atives, being consistent with a conjugate containing 1 mol of colcemid and 1 mol of the NBD group.
3) There was no evidence from the absorption spectrum of the product for significant decomposition of the tropolone moiety; photo-degradation products of colcemid (lumicolcem- ids) exhibit an absorption maximum at 267 nm (c = 23,600 M" cm") (21).
4) NBD-colcemid fluoresced strongly in ethanol/water mix- tures in the range of 520-540 nm (see below). This is char- acteristic of an N-NBD derivative (20, 28).
500 550 nm
600
FIG. 2. Fluorescent properties of NBD-colcemid. The spectra of NBD-colcemid (10 p ~ ) were measured at 25 "C with 465 nm exciting light. A, emission spectra of NBD-colcemid in water/ethanol mixtures; the percentages of ethanol are indicated on the curves. B, the variation of fluorescence intensity (0) and the wavelength of maximum fluorescence emission (0) of NBD-colcemid in water/ ethanol mixtures.
I
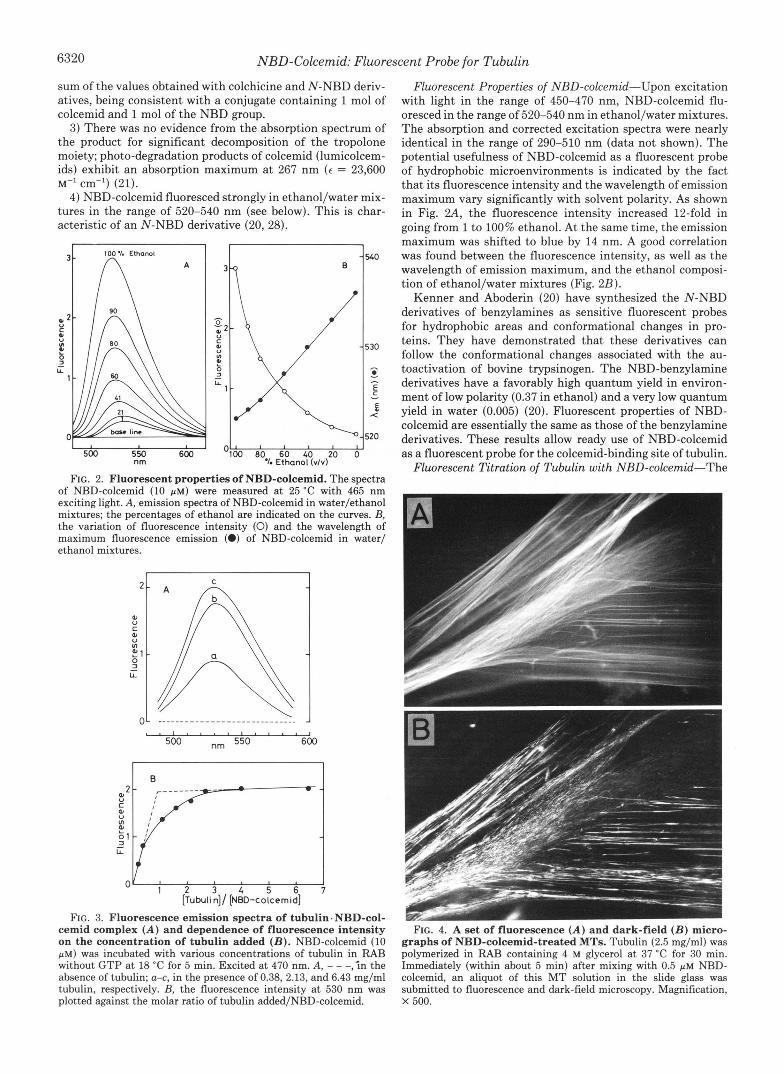
Fluorescent Properties of NBD-colcemid-Upon excitation with light in the range of 450-470 nm, NBD-colcemid flu- oresced in the range of 520-540 nm in ethanol/water mixtures. The absorption and corrected excitation spectra were nearly identical in the range of 290-510 nm (data not shown). The potential usefulness of NBD-colcemid as a fluorescent probe of hydrophobic microenvironments is indicated by the fact that its fluorescence intensity and the wavelength of emission maximum vary significantly with solvent polarity. As shown in Fig. 2 A , the fluorescence intensity increased 12-fold in going from 1 to 100% ethanol. At the same time, the emission maximum was shifted to blue by 14 nm. A good correlation was found between the fluorescence intensity, as well as the wavelength of emission maximum, and the ethanol composi- tion of ethanol/water mixtures (Fig. 2B).
Kenner and Aboderin (20) have synthesized the N-NBD derivatives of benzylamines as sensitive fluorescent probes for hydrophobic areas and conformational changes in pro- teins. They have demonstrated that these derivatives can follow the conformational changes associated with the au- toactivation of bovine trypsinogen. The NBD-benzylamine derivatives have a favorably high quantum yield in environ- ment of low polarity (0.37 in ethanol) and a very low quantum yield in water (0.005) (20). Fluorescent properties of NBD- colcemid are essentially the same as those of the benzylamine derivatives. These results allow ready use of NBD-colcemid as a fluorescent probe for the colcemid-binding site of tubulin.
Fluorescent Titration of Tubulin with NBD-cokemid-The
I . I I I , , , . , I
500 nm 550 600
I 0
[Tubulin]/ [NED-colcemid]
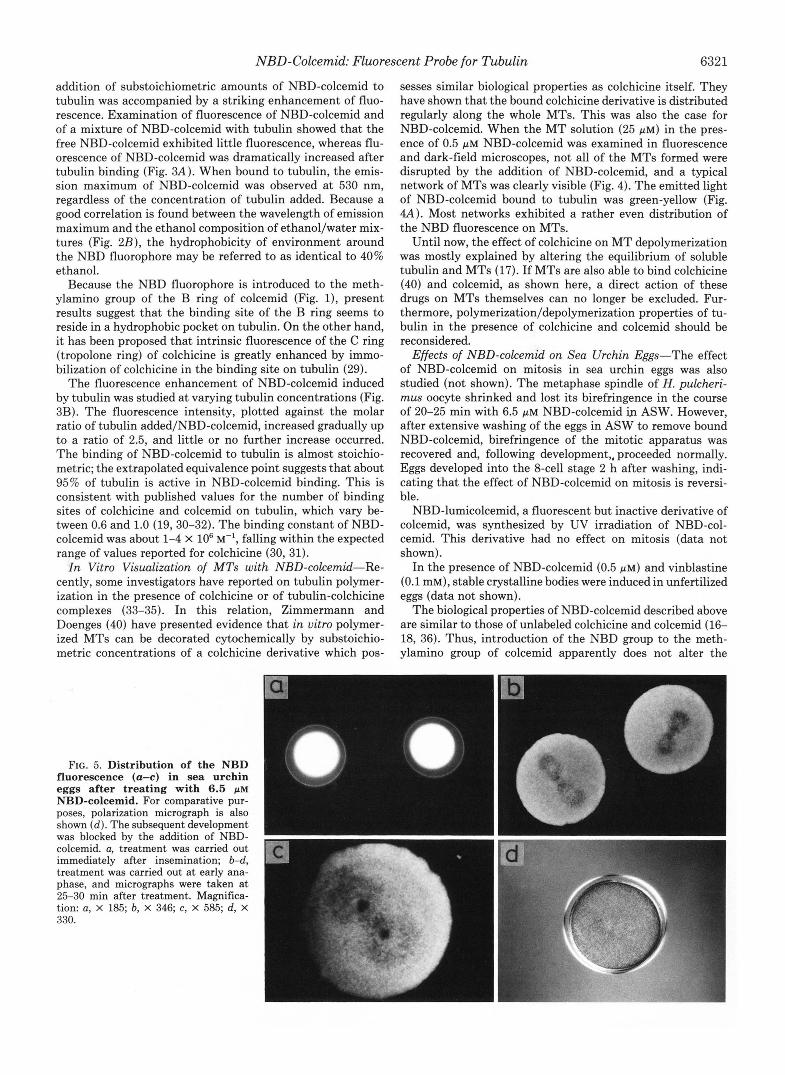
FIG. 3. Fluorescence emission spectra of tubulin.NBD-col- cemid complex ( A ) and dependence of fluorescence intensity on the concentration of tubulin added ( B ) . NBD-colcemid (10 p ~ ) was incubated with various concentrations of tubulin in RAB without GTP at 18 "C for 5 min. Excited at 470 nm. A, - - -,in the absence of tubulin; a-c, in the presence of 0.38, 2.13, and 6.43 mg/ml tubulin, respectively. B, the fluorescence intensity a t 530 nm was plotted against the molar ratio of tubulin added/NBD-colcemid.
FIG. 4. A set of fluorescence ( A ) and dark-field ( B ) micro- graphs of NBD-colcemid-treated MTs. Tubulin (2.5 mg/ml) was polymerized in RAB containing 4 M glycerol at 37 "C for 30 min. Immediately (within about 5 min) after mixing with 0.5 p~ NBD- colcemid, an aliquot of this MT solution in the slide glass was submitted to fluorescence and dark-field microscopy. Magnification, X 500.
NBD-Colcemid: Fluorescent Probe for Tubulin 6321
addition of substoichiometric amounts of NBD-colcemid to tubulin was accompanied by a striking enhancement of fluo- rescence. Examination of fluorescence of NBD-colcemid and of a mixture of NBD-colcemid with tubulin showed that the free NBD-colcemid exhibited little fluorescence, whereas flu- orescence of NBD-colcemid was dramatically increased after tubulin binding (Fig. 3A). When bound to tubulin, the emis- sion maximum of NBD-colcemid was observed a t 530 nm, regardless of the concentration of tubulin added. Because a good correlation is found between the wavelength of emission maximum and the ethanol composition of ethanol/water mix- tures (Fig. 2B), the hydrophobicity of environment around the NBD fluorophore may be referred to as identical to 40% ethanol.
Because the NBD fluorophore is introduced to the meth- ylamino group of the B ring of colcemid (Fig. l), present results suggest that the binding site of the B ring seems to reside in a hydrophobic pocket on tubulin. On the other hand, it has been proposed that intrinsic fluorescence of the C ring (tropolone ring) of colchicine is greatly enhanced by immo- bilization of colchicine in the binding site on tubulin (29).
The fluorescence enhancement of NBD-colcemid induced by tubulin was studied at varying tubulin concentrations (Fig. 3B). The fluorescence intensity, plotted against the molar ratio of tubulin added/NBD-colcemid, increased gradually up to a ratio of 2.5, and little or no further increase occurred. The binding of NBD-colcemid to tubulin is almost stoichio- metric; the extrapolated equivalence point suggests that about 95% of tubulin is active in NBD-colcemid binding. This is consistent with published values for the number of binding sites of colchicine and colcemid on tubulin, which vary be- tween 0.6 and 1.0 (19,30-32). The binding constant of NBD- colcemid was about 1-4 X lo6 M-', falling within the expected range of values reported for colchicine (30, 31).
In Vitro Visualization of MTs with NBD-colcemid-Re- cently, some investigators have reported on tubulin polymer- ization in the presence of colchicine or of tubulin-colchicine complexes (33-35). In this relation, Zimmermann and Doenges (40) have presented evidence that in vitro polymer- ized MTs can be decorated cytochemically by substoichio- metric concentrations of a colchicine derivative which pos-
sesses similar biological properties as colchicine itself. They have shown that the bound colchicine derivative is distributed regularly along the whole MTs. This was also the case for NBD-colcemid. When the MT solution (25 p ~ ) in the pres- ence of 0.5 p~ NBD-colcemid was examined in fluorescence and dark-field microscopes, not all of the MTs formed were disrupted by the addition of NBD-colcemid, and a typical network of MTs was clearly visible (Fig. 4). The emitted light of NBD-colcemid bound to tubulin was green-yellow (Fig. 4A). Most networks exhibited a rather even distribution of the NBD fluorescence on MTs.
Until now, the effect of colchicine on MT depolymerization was mostly explained by altering the equilibrium of soluble tubulin and MTs (17). If MTs are also able to bind colchicine (40) and colcemid, as shown here, a direct action of these drugs on MTs themselves can no longer be excluded. Fur- thermore, polymerization/depolymerization properties of tu- bulin in the presence of colchicine and colcemid should be reconsidered.
Effects of NBD-colcemid on Sea Urchin Eggs-The effect of NBD-colcemid on mitosis in sea urchin eggs was also studied (not shown). The metaphase spindle of H . pulcheri- mus oocyte shrinked and lost its birefringence in the course of 20-25 min with 6.5 p~ NBD-colcemid ip ASW. However, after extensive washing of the eggs in ASW to remove bound NBD-colcemid, birefringence of the mitotic apparatus was recovered and, following development,, proceeded normally. Eggs developed into the 8-cell stage 2 h after washing, indi- cating that the effect of NBD-colcemid on mitosis is reversi- ble.
NBD-lumicolcemid, a fluorescent but inactive derivative of colcemid, was synthesized by UV irradiation of NBD-col- cemid. This derivative had no effect on mitosis (data not shown).
In the presence of NBD-colcemid (0.5 p ~ ) and vinblastine (0.1 mM), stable crystalline bodies were induced in unfertilized eggs (data not shown).
The biological properties of NBD-colcemid described above are similar to those of unlabeled colchicine and colcemid (16- 18, 36). Thus, introduction of the NBD group to the meth- ylamino group of colcemid apparently does not alter the
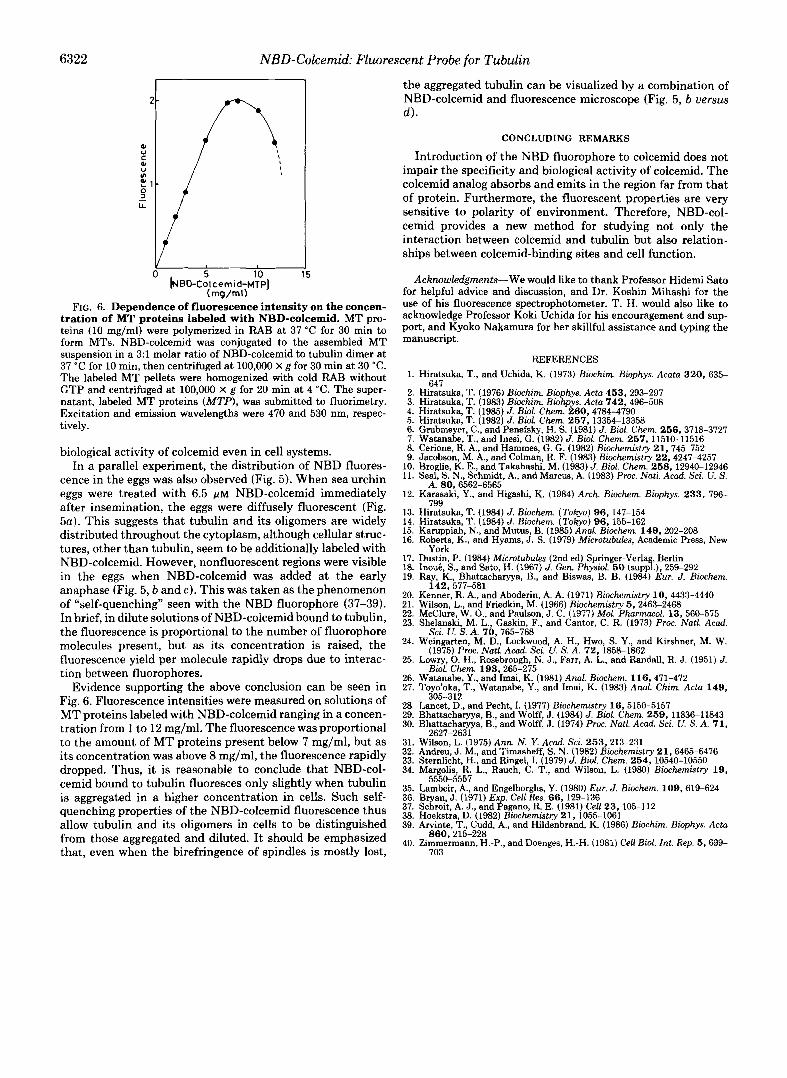
FIG. 5. Distribution of the NBD fluorescence (a-c) in sea urchin eggs after treating with 6.5 p~ NBD-colcemid. For comparative pur- poses, polarization micrograph is also shown (d ) . The subsequent development was blocked by the addition of NBD- colcemid. a, treatment was carried out immediately after insemination; b-d, treatment was carried out at early ana- phase, and micrographs were taken at 25-30 min after treatment. Magnifica- tion: a, X 185; b, X 346; c, X 585; d, X 330.
6322 NBD-Colcernid: Fluorescent Probe for Tubulin
- 4 ' 1 f LL
Y I 0 5 10 15
(mg/mlf FIG. 6. Dependence of fluorescence intensity on the concen-
tration of MT proteins labeled with NBD-colcemid. MT pro- teins (10 mg/ml) were polymerized in RAB at 37 "C for 30 min to form MTs. NBD-colcemid was conjugated to the assembled MT suspension in a 3:l molar ratio of NBD-colcemid to tubulin dimer at 37 "C for 10 min, then centrifuged at 100,000 X g for 30 min at 30 "C. The labeled MT pellets were homogenized with cold RAB without GTP and centrifuged at 100,000 X g for 20 min at 4 "C. The super- natant, labeled MT proteins (MTP), was submitted to fluorimetry. Excitation and emission wavelengths were 470 and 530 nm, respec- tively.
biological activity of colcemid even in cell systems. In a parallel experiment, the distribution of NBD fluores-
cence in the eggs was also observed (Fig. 5 ) . When sea urchin eggs were treated with 6.5 ~ L M NBD-colcemid immediately after insemination, the eggs were diffusely fluorescent (Fig. 5a). This suggests that tubulin and its oligomers are widely distributed throughout the cytoplasm, although cellular struc- tures, other than tubulin, seem to be additionally labeled with NBD-colcemid. However, nonfluorescent regions were visible in the eggs when NBD-colcemid was added at the early anaphase (Fig. 5 , b and c). This was taken as the phenomenon of "self-quenching'' seen with the NBD fluorophore (37-39). In brief, in dilute solutions of NBD-colcemid bound to tubulin, the fluorescence is proportional to the number of fluorophore molecules present, but as its concentration is raised, the fluorescence yield per molecule rapidly drops due to interac- tion between fluorophores.
Evidence supporting the above conclusion can be seen in Fig. 6. Fluorescence intensities were measured on solutions of MT proteins labeled with NBD-colcemid ranging in a concen- tration from 1 to 12 mg/ml. The fluorescence was proportional to the amount of MT proteins present below 7 mg/ml, but as its concentration was above 8 mg/ml, the fluorescence rapidly dropped. Thus, it is reasonable to conclude that NBD-col- cemid bound to tubulin fluoresces only slightly when tubulin is aggregated in a higher concentration in cells. Such self- quenching properties of the NBD-colcemid fluorescence thus allow tubulin and its oligomers in cells to be distinguished from those aggregated and diluted. I t should be emphasized that, even when the birefringence of spindles is mostly lost,
[NBD-Colcemid-MTP]
the aggregated tubulin can be visualized by a combination of NBD-colcemid and fluorescence microscope (Fig. 5 , b uers'sus 4 .
CONCLUDING REMARKS
Introduction of the NBD fluorophore to colcemid does not impair the specificity and biological activity of colcemid. The colcemid analog absorbs and emits in the region far from that of protein. Furthermore, the fluorescent properties are very sensitive to polarity of environment. Therefore, NBD-col- cemid provides a new method for studying not only the interaction between colcemid and tubulin but also relation- ships between colcemid-binding sites and cell function.
Acknowledgments-We would like to thank Professor Hidemi Sato for helpful advice and discussion, and Dr. Koshin Mihashi for the use of his fluorescence spectrophotometer. T. H. would also like to acknowledge Professor Koki Uchida for his encouragement and sup- port, and Kyoko Nakamura for her skillful assistance and typing the manuscript.
REFERENCES 1. Hiratsuka, T., and Uchida, K. (1973) Biochim. Biophys. A C Q ~ Q 320 , 635-
2. Hiratsuka, T. (1976) Biochim. Biophys. Acta 453 , 293-297
4. Hiratsuka, T. (1985) J. Biol. Chem. 260,4784-4790 3. Hiratsuka, T. (1983) Biochim. Biohpys. Acta 742,496-508
6. Grubmeyer, C., and Penefsky, H. S. (1981) J. Biol. Chem. 256,3718-3727 5. Hiratsuka, T. (1982) J. Biol. Chem. 257,13354-13358
7. Watanabe, T., and Inesi, G. (1982) J. Biol. Chem. 257,11510-11516 8. Cerione, R. A,, and Hammes, G. G. (1982) Biochemistry 21,745-752
10. Broglie, K. E., and Takahashi, M. (1983) J. Biol. Chem. 258,12940-12946 9. Jacobson, M. A,, and Colman, R. F. (1983) Biochemistry 22,4247-4257
11. Seal, S . N., Schmidt, A., and Marcus, A. (1983) Proc. NQtl. Acad. Sei. U. S.
12. Karasaki, Y., and Higashi, K. (1984) Arch. Biochem. Biophys. 2 3 3 , 796-
13. Hiratsuka, T. (1984) J. Biochem. (Tokyo) 9 6 , 147-154 14. Hiratsuka, T. (1984) J. Biochem. (Tokyo) 96,155-162 15. Karuppiah, N., and Mutus, B. (1985) A n d Biochem. 149,202-208 16. Roberts, K., and Hyams, J. S. (1979) Microtubules, Academic Press, New
17. Dustin, P. (1984) Microtubules (2nd ed) Springer-Verlag, Berlin 18. InouC., S., and Sato, H. (1967) J. Gen. Physiol. 50 (suppl.), 259-292 19. Ray, K., Bhattacharyya, B., and Biswas, B. B. (1984) Eur. J. Biochem.
20. Kenner, R. A,, and Aboderin, A. A. (1971) Biochemistry 10,4433-4440 21. Wilson, L., and Friedkin, M. (1966) Biochemistry 5,2463-2468 22. McClure, W. O., and Paulson, J. C. (1977) Mol. Phurmacol. 13,560-575 23. Shelanski. M. L.. Gaskin. F.. and Cantor. C. R. (1973) Proc. Natl. Acad.
647
A. 80,6562-6565
799
York
142,577-581
Sci. U. S . A . 70,765-768 '
(1975) Proc. Natl. Acad. Sci. U. S. A. 72.1858-1862
, . ~~
24. Weingarten, M. D., Lockwood, A. H., Hwo, S. Y., and Kirshner, M. W.
25. Lowry, 0. H., Rosebrough, N. J. , Farr, A. L., and Randall, R. J. (1951) J.
26. Watanabe, Y., and Imai, K. (1981) Anal. Biochem. 116,471-472 27. Toyo'oka, T., Watanabe, Y., and Imai, K. (1983) Anal. Chim. Acta 149 ,
Biol. Chem. 193,265-275
sn5-217 28. Lancet, D., and Pecht, I. (1977) Biochemistry 16,5150-5157 29. Bhattacharyya, B., and Wolff, J . (1984) J. Biol. Chem. 259,11836-11843 30. Bhattacharyya, B., and Wolff, J. (1974) Proc. Natl. Acad. Sci. U. S. A . 7 1 ,
31. Wilson, L. (1975) Ann. N. Y. Acad. Sci. 253,213-231 32. Andreu, 3. M., and Timasheff, S. N. (1982) Biochemistry 21,6465-6476
34. Margolis, R. L., Rauch, C. T., and Wilson, L. (1980) Biochemistry 19 , 33. Sternlicht, H., and Ringel, I. (1979) J. Biol. Chem. 254,10540-10550
-"" ""
2627-2631
35. Lambeir, A., and Engelborghs, Y. (1980) Eur. J . Biochem. 109,619-624 36. Bryan, J. (1971) Exp. Cell Res. 66,129-136 37. Schroit, A. J., and Pagano, R. E. (1981) Cell 23,105-112 38. Hoekstra, D. (1982) Biochemistry 21,1055-1061 39. Arvinte, T., Cudd, A,, and Hildenbrand, K. (1986) Biochim. Biophys. Acta
40. Zimmermann, H.-P., and Doenges, H.-H. (1981) Cell Biol. Int. Rep. 5,699-
5550-5557
860,215-228
703




![Nitro/Bromo Ketones to Access 2H-Pyrans Oxa-[3+3] Annulation … · 2020-05-19 · S1 Oxa-[3+3] Annulation of MBH-Carbonates of Propiolaldehydes with - Nitro/Bromo Ketones to Access](https://img.pdfslide.net/doc/110x75/5f1c5856ddd32b438e122c49/nitrobromo-ketones-to-access-2h-pyrans-oxa-33-annulation-2020-05-19-s1-oxa-33.jpg)























