Embed Size (px)
Citation preview

The blood-brain barrier is disrupted in a mousemodel of infantile neuronal ceroid lipofuscinosis:amelioration by resveratrol
Arjun Saha1,{, Chinmoy Sarkar1,{, Satya P. Singh2,{, Zhongjian Zhang1, Jeeva Munasinghe3,
Shiyong Peng1, Goutam Chandra1, Eryan Kong1 and Anil B. Mukherjee1,∗
1Section on Developmental Genetics, Program on Endocrinology and Genetics, Eunice Kennedy Shriver National
Institute of Child Health and Human Development, 2Laboratory of Molecular Immunology, National Institute of Allergy
and Infectious Diseases and 3National Institute of Neurological Disorders and Stroke, In Vivo NMR Center (HNQ2-3),
National Institutes of Health, Bethesda, MD 20892-1830, USA
Received December 30, 2011; Revised and Accepted February 6, 2012
Disruption of the blood-brain barrier (BBB) is a serious complication frequently encountered in neurodegen-erative disorders. Infantile neuronal ceroid lipofuscinosis (INCL) is a devastating childhood neurodegenera-tive lysosomal storage disorder caused by palmitoyl-protein thioesterase-1 (PPT1) deficiency. It remainsunclear whether BBB is disrupted in INCL and if so, what might be the molecular mechanism(s) of this com-plication. We previously reported that the Ppt1-knockout (Ppt1-KO) mice that mimic INCL manifest high levelsof oxidative stress and neuroinflammation. Recently, it has been reported that CD41 T-helper 17 (TH17) lym-phocytes may mediate BBB disruption and neuroinflammation, although the precise molecular mechanism(s)remain unclear. We sought to determine: (i) whether the BBB is disrupted in Ppt1-KO mice, (ii) if so, do TH17-lymphocytes underlie this complication, and (iii) how might TH17 lymphocytes breach the BBB. Here, wereport that the BBB is disrupted in Ppt1-KO mice and that TH17 lymphocytes producing IL-17A mediate dis-ruption of the BBB by stimulating production of matrix metalloproteinases (MMPs), which degrade the tightjunction proteins essential for maintaining BBB integrity. Importantly, dietary supplementation of resveratrol(RSV), a naturally occurring antioxidant/anti-inflammatory polyphenol, markedly reduced the levels of TH17cells, IL-17A and MMPs, and elevated the levels of tight junction proteins, which improved the BBB integrityin Ppt1-KO mice. Intriguingly, we found that RSV suppressed the differentiation of CD41 T lymphocytes to IL-17A-positive TH17 cells. Our findings uncover a mechanism by which TH17 lymphocytes mediate BBB disrup-tion and suggest that small molecules such as RSV that suppress TH17 differentiation are therapeutic targetsfor neurodegenerative disorders such as INCL.
INTRODUCTION
The blood-brain barrier (BBB) is a selective permeabilitybarrier between the blood and the brain. It restricts the entryof plasma components, including red blood cells and leuko-cytes into the brain, which can damage neurons. In severalconditions, such as ischemic injury, trauma, cerebral hemor-rhage, neurodegeneration and neuroinflammation, the BBBmay be compromised. Thus, a breach of the BBB allows
immune cells to freely migrate to the brain damaging theneurons. Indeed, in chronic neurodegenerative diseases,disruption of the BBB is a frequent complication and thedevelopment of effective treatment remains challenging.The endothelial cells of the microvasculature and tight junc-tion proteins between adjacent endothelial cells are themajor constituents of the BBB (1). Previous work has shownthat CD4+ T-helper 17 (TH17) lymphocytes (2) expressing ret-inoic acid receptor-related orphan receptor gt (RORgt) (3)
†Equally contributed.
∗To whom correspondence should be addressed at: NIH, Building 10, Room 9D42, 10, Center Drive, Bethesda, MD 20892-1830, USA. Tel: +13014967213; Fax: +1 3014026632; Email: [email protected]
Published by Oxford University Press 2012.
Human Molecular Genetics, 2012, Vol. 21, No. 10 2233–2244doi:10.1093/hmg/dds038Advance Access published on February 13, 2012
Dow
nloaded from https://academ
ic.oup.com/hm
g/article-abstract/21/10/2233/2900646 by guest on 04 April 2019

mediate BBB disruption and neuroinflammation (4). However,it is unclear how these immune cells disrupt the BBB.
Lysosomal storage disorders (LSDs) affect 1 in 5000live-born infants and neurodegeneration is a devastatingmanifestation in the majority of the more than 50 LSDs (5).Neuronal Ceroid lipofuscinoses (NCLs) commonly known asBatten disease represent a group of common (1 in 12500births) neurodegenerative LSDs that mostly affect children(6,7). Mutations in at least eight different genes causevarious types of NCLs (8). The infantile form of NCL(INCL) (6,7) is a devastating childhood neurodegenerativestorage disease caused by inactivating mutations in thepalmitoyl-protein thioesterase-1 (PPT1) gene (9). ThePpt1-knockout (Ppt1-KO) mice (10) recapitulate virtually allclinical and pathological features of INCL (10,11), providinga reliable animal model of a neurodegenerative disease toexplore the mechanism(s) of BBB disruption and to developnovel therapeutic strategies.
In this study, we sought to determine whether BBB is dis-rupted in Ppt1-KO mice and if so what might be the mechan-ism of BBB disruption in this animal model of INCL. Ourresults showed that BBB is disrupted in Ppt1-KO mice andIL-17A, produced by TH17 lymphocytes mediate BBB disrup-tion by stimulating production of matrix metalloproteinases(MMPs), which degrade the tight junction proteins that are es-sential for maintaining BBB integrity. Most importantly, wefound that dietary supplementation of resveratrol (RSV), ananti-oxidant and anti-inflammatory polyphenol, prevented dis-ruption of the BBB by suppressing the differentiation of naıveCD4+ T cells to TH17 lymphocytes, which reduced productionof IL-17-mediated production of MMPs. We propose thatsmall molecules such as RSV that suppress TH17 differenti-ation are therapeutic targets for neurodegenerative disorders,such as INCL.
RESULTS
BBB integrity in Ppt1-KO mice evaluated bygadolinium-enhanced magnetic resonance imaging
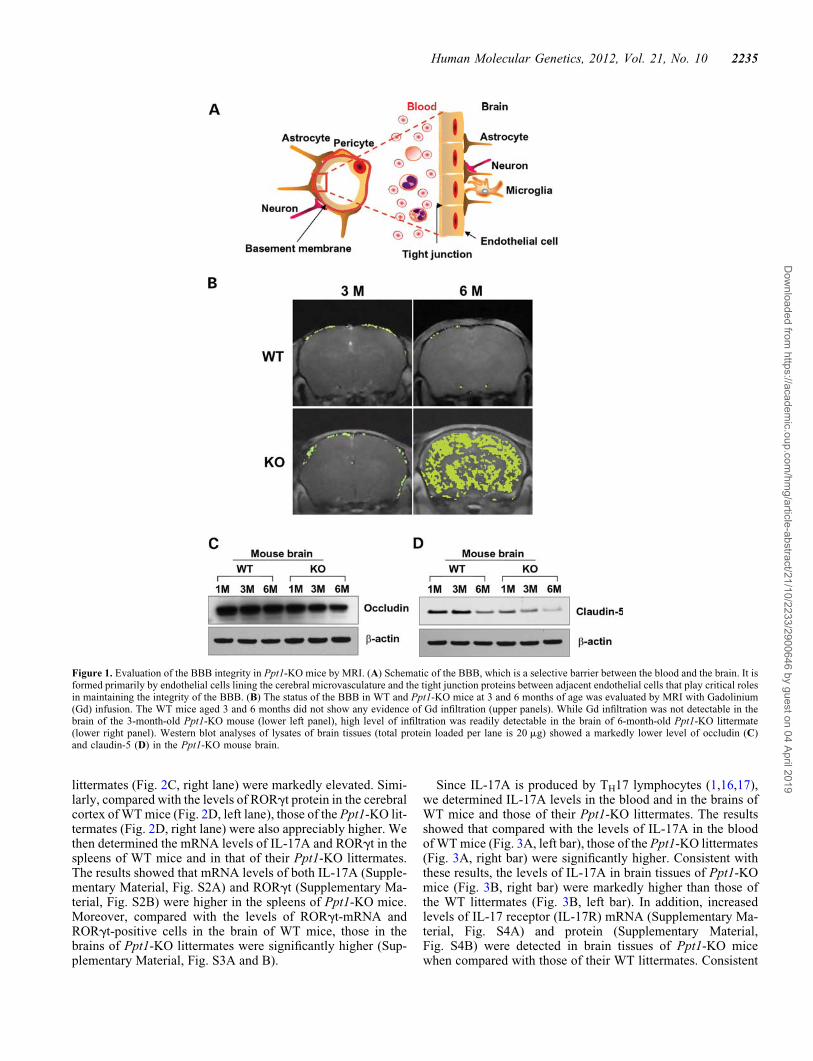
The endothelial cells lining the lumen of the cerebral microvas-culature and the tight junction proteins between adjacentendothelial cells maintain the integrity of the BBB (Fig. 1A).To determine whether the BBB is disrupted in Ppt1-KO mice,we performed brain magnetic resonance imaging (MRI) of 3-and 6-month-old Ppt1-KO mice and their wild-type (WT) litter-mates following intravenous infusion of gadolinium (Gd), acontrast medium, which does not infiltrate the normal brain par-enchyma due to an intact BBB (12). The results showed thatthere was no Gd infiltration in the brains of 3-month-old WTmice (Fig. 1B, upper left panel) as well as in those of theirPpt1-KO littermates (Fig. 1B, lower left panel). However,while the 6-month-old WT mice had no detectable Gd infiltra-tion (Fig. 1B, upper right panel), there was a massive infiltrationof Gd into the brain of their Ppt1-KO littermates (Fig. 1B, lowerright panel). Taken together, these results clearly demonstratedthat while the BBB was intact in WT mice and in their3-month-old Ppt1-KO littermates, it was breached in6-month-old Ppt1-KO mice. Remarkably, coinciding with
BBB disruption, the 6-month-old Ppt1-KO mice also mani-fested signs of neurological deterioration (10).
Reduced levels of tight junction proteins in the brain ofPpt1-KO mice
The tight junctions between adjacent endothelial cells provide ametabolic and physical barrier, which selectively allows move-ment of macromolecules between the blood and the brain(13,14) in order to maintain homeostasis. The tight junction pro-teins including occludin, claudin and junctional adhesionmolecule-1 (JAM1) play critical roles in maintaining the BBBintegrity, and degradation of the tight junction allows freemovement of molecules and immune cells from the blood tothe brain and vice versa. In addition, abnormal angiogenesis,regression of blood vessels, hypoperfusion of the brain andneuroinflammation may contribute to a ‘vicious circle’ thateventually leads to synaptic as well as neuronal dysfunctionand progression of neurodegeneration. We previously reportedthat Ppt1-KO mice suffer from neuroinflammation, which maycontribute to BBB disruption. Thus, we determined the levels oftight junction proteins, occludin and claudin-5, in corticaltissues from 1-, 3- and 6-month-old Ppt1-KO mice and thoseof their WT littermates by western blot analysis. The resultsshowed that the levels of occludin (Fig. 1C) as well asclaudin-5 (Fig. 1D) declined with increasing age of thePpt1-KO mice. Consistent with these findings, the levels ofother tight junction proteins such as JAM1 and claudin-1 werealso diminished in the brain of Ppt1-KO mice (SupplementaryMaterial, Fig. S1). These findings strongly suggested thatBBB integrity in Ppt1-KO mice is breached, at least in part,due to reduced levels of tight junction proteins.
Elevated levels of TH17 lymphocytes and IL-17A inPpt1-KO mice
While TH17 lymphocytes have been implicated in mediatingBBB disruption and neuroinflammation (4), the molecularmechanism(s), until now, remain unclear. To understand howTH17 lymphocytes may mediate BBB disruption, we soughtto determine whether the levels of these immune cells in lymph-oid organs and in the brains of Ppt1-KO mice are elevated com-pared with those of their WT littermates. Accordingly, weperformed fluorescence-activated cell sorting (FACS) analysisof IL-17A-positive TH17 lymphocytes in the spleen and in thebrain of WT mice and in those of their Ppt1-KO littermates.The results showed that compared with the levels ofIL-17A-positive cells in the spleens of WT mice (Fig. 2A, leftpanel), those in the spleens of Ppt1-KO littermates (Fig. 2A,right panel) were markedly elevated. Similarly, comparedwith the levels of IL-17A-positive cells in the brains of WTmice (Fig. 2B, left panel), those in the brains of their Ppt1-KOlittermates (Fig. 2B, right panel) were also significantly ele-vated. Since development and function IL-17A-producingTH17 lymphocytes (3) depend on RORgt (reviewed in 15), wefirst determined the levels of RORgt protein in the spleen andbrain tissues from perfused Ppt1-KO mice and those of theirWT littermates by western blot analysis. The results showedthat compared with the levels of RORgt protein in the spleensof WT mice (Fig. 2C, left lane), those of their Ppt1-KO
2234 Human Molecular Genetics, 2012, Vol. 21, No. 10
Dow
nloaded from https://academ
ic.oup.com/hm
g/article-abstract/21/10/2233/2900646 by guest on 04 April 2019
littermates (Fig. 2C, right lane) were markedly elevated. Simi-larly, compared with the levels of RORgt protein in the cerebralcortex of WT mice (Fig. 2D, left lane), those of the Ppt1-KO lit-termates (Fig. 2D, right lane) were also appreciably higher. Wethen determined the mRNA levels of IL-17A and RORgt in thespleens of WT mice and in that of their Ppt1-KO littermates.The results showed that mRNA levels of both IL-17A (Supple-mentary Material, Fig. S2A) and RORgt (Supplementary Ma-terial, Fig. S2B) were higher in the spleens of Ppt1-KO mice.Moreover, compared with the levels of RORgt-mRNA andRORgt-positive cells in the brain of WT mice, those in thebrains of Ppt1-KO littermates were significantly higher (Sup-plementary Material, Fig. S3A and B).
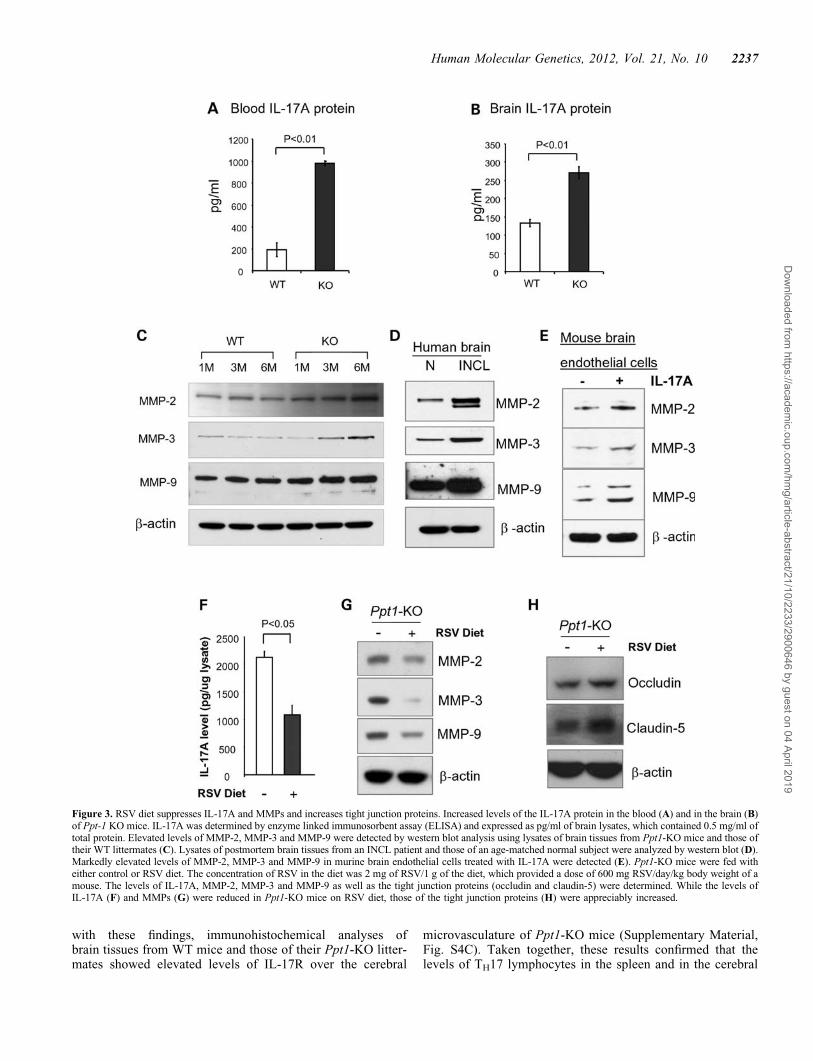
Since IL-17A is produced by TH17 lymphocytes (1,16,17),we determined IL-17A levels in the blood and in the brains ofWT mice and those of their Ppt1-KO littermates. The resultsshowed that compared with the levels of IL-17A in the bloodof WT mice (Fig. 3A, left bar), those of the Ppt1-KO littermates(Fig. 3A, right bar) were significantly higher. Consistent withthese results, the levels of IL-17A in brain tissues of Ppt1-KOmice (Fig. 3B, right bar) were markedly higher than those ofthe WT littermates (Fig. 3B, left bar). In addition, increasedlevels of IL-17 receptor (IL-17R) mRNA (Supplementary Ma-terial, Fig. S4A) and protein (Supplementary Material,Fig. S4B) were detected in brain tissues of Ppt1-KO micewhen compared with those of their WT littermates. Consistent
Figure 1. Evaluation of the BBB integrity in Ppt1-KO mice by MRI. (A) Schematic of the BBB, which is a selective barrier between the blood and the brain. It isformed primarily by endothelial cells lining the cerebral microvasculature and the tight junction proteins between adjacent endothelial cells that play critical rolesin maintaining the integrity of the BBB. (B) The status of the BBB in WT and Ppt1-KO mice at 3 and 6 months of age was evaluated by MRI with Gadolinium(Gd) infusion. The WT mice aged 3 and 6 months did not show any evidence of Gd infiltration (upper panels). While Gd infiltration was not detectable in thebrain of the 3-month-old Ppt1-KO mouse (lower left panel), high level of infiltration was readily detectable in the brain of 6-month-old Ppt1-KO littermate(lower right panel). Western blot analyses of lysates of brain tissues (total protein loaded per lane is 20 mg) showed a markedly lower level of occludin (C)and claudin-5 (D) in the Ppt1-KO mouse brain.
Human Molecular Genetics, 2012, Vol. 21, No. 10 2235
Dow
nloaded from https://academ
ic.oup.com/hm
g/article-abstract/21/10/2233/2900646 by guest on 04 April 2019
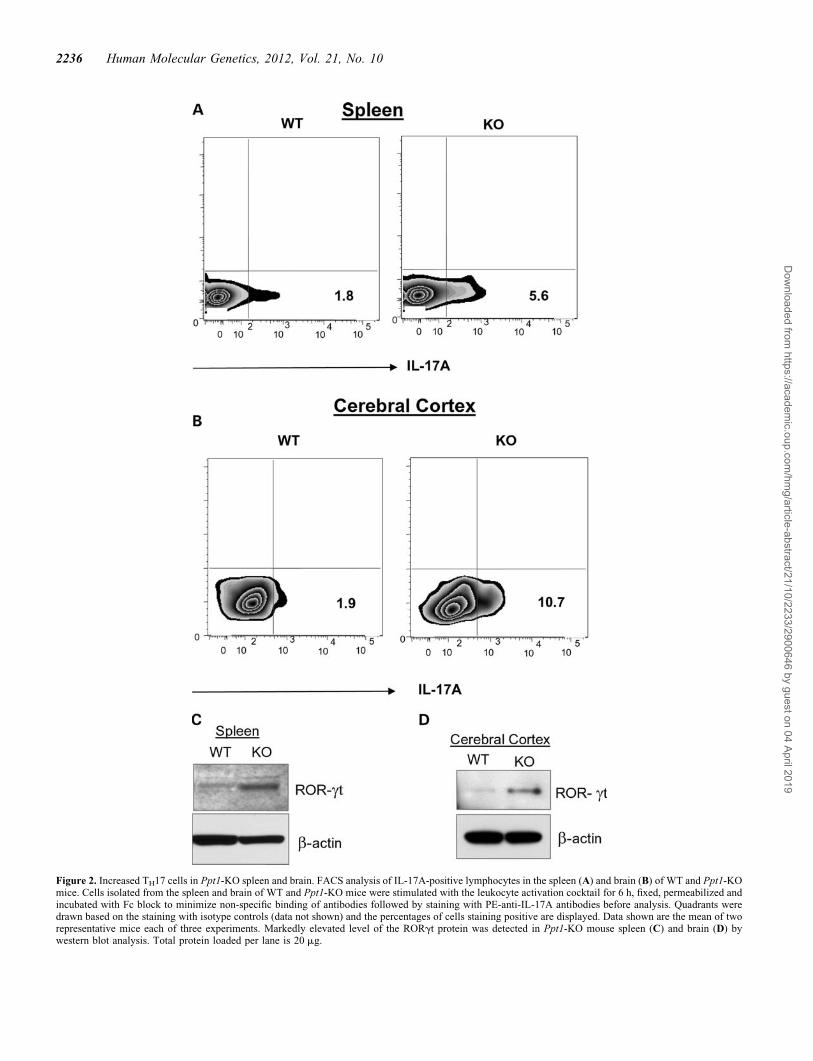
Figure 2. Increased TH17 cells in Ppt1-KO spleen and brain. FACS analysis of IL-17A-positive lymphocytes in the spleen (A) and brain (B) of WT and Ppt1-KOmice. Cells isolated from the spleen and brain of WT and Ppt1-KO mice were stimulated with the leukocyte activation cocktail for 6 h, fixed, permeabilized andincubated with Fc block to minimize non-specific binding of antibodies followed by staining with PE-anti-IL-17A antibodies before analysis. Quadrants weredrawn based on the staining with isotype controls (data not shown) and the percentages of cells staining positive are displayed. Data shown are the mean of tworepresentative mice each of three experiments. Markedly elevated level of the RORgt protein was detected in Ppt1-KO mouse spleen (C) and brain (D) bywestern blot analysis. Total protein loaded per lane is 20 mg.
2236 Human Molecular Genetics, 2012, Vol. 21, No. 10
Dow
nloaded from https://academ
ic.oup.com/hm
g/article-abstract/21/10/2233/2900646 by guest on 04 April 2019
with these findings, immunohistochemical analyses ofbrain tissues from WT mice and those of their Ppt1-KO litter-mates showed elevated levels of IL-17R over the cerebral
microvasculature of Ppt1-KO mice (Supplementary Material,Fig. S4C). Taken together, these results confirmed that thelevels of TH17 lymphocytes in the spleen and in the cerebral
Figure 3. RSV diet suppresses IL-17A and MMPs and increases tight junction proteins. Increased levels of the IL-17A protein in the blood (A) and in the brain (B)of Ppt-1 KO mice. IL-17A was determined by enzyme linked immunosorbent assay (ELISA) and expressed as pg/ml of brain lysates, which contained 0.5 mg/ml oftotal protein. Elevated levels of MMP-2, MMP-3 and MMP-9 were detected by western blot analysis using lysates of brain tissues from Ppt1-KO mice and those oftheir WT littermates (C). Lysates of postmortem brain tissues from an INCL patient and those of an age-matched normal subject were analyzed by western blot (D).Markedly elevated levels of MMP-2, MMP-3 and MMP-9 in murine brain endothelial cells treated with IL-17A were detected (E). Ppt1-KO mice were fed witheither control or RSV diet. The concentration of RSV in the diet was 2 mg of RSV/1 g of the diet, which provided a dose of 600 mg RSV/day/kg body weight of amouse. The levels of IL-17A, MMP-2, MMP-3 and MMP-9 as well as the tight junction proteins (occludin and claudin-5) were determined. While the levels ofIL-17A (F) and MMPs (G) were reduced in Ppt1-KO mice on RSV diet, those of the tight junction proteins (H) were appreciably increased.
Human Molecular Genetics, 2012, Vol. 21, No. 10 2237
Dow
nloaded from https://academ
ic.oup.com/hm
g/article-abstract/21/10/2233/2900646 by guest on 04 April 2019

cortex as well as IL-17A levels in the same organs of Ppt1-KOmice are significantly higher compared with those of their WTlittermates.
MMPs in the brain of Ppt1-KO mice and in that ofpostmortem INCL patient
How might an increase in TH17 lymphocytes mediate BBBdisruption? One possibility is that IL-17A produced by TH17lymphocytes (3) stimulates or activates proteases that catalyzethe degradation of tight junction proteins, essential for main-taining the integrity of the BBB. MMPs have been reportedto have multiple roles in a variety of physiological as wellas pathological conditions (18), including neurodegenerative(19) and neuroinflammatory diseases (20). Moreover, MMPshave been reported to degrade tight junction proteins such asoccludin (21). Therefore, we sought to test the hypothesisthat IL-17A produced by TH17 lymphocytes promotes the ex-pression of MMPs, which are reported to play critical roles inneurodegenerative and neuroinflammatory diseases (19–21).Accordingly, we performed western blot analysis of braintissue lysates from Ppt1-KO mice and those of their WT litter-mates using antibodies against MMP-2, MMP-3 and MMP-9.The results showed that protein levels of all three MMPs wereelevated in the brain of Ppt1-KO mice in an age-dependentmanner (Fig. 3C). We also determined the MMP-proteinlevels in postmortem brain tissues from an INCL patient andthose of an age-matched normal individual by western blotanalysis. The results showed that MMP-2-, MMP-3- andMMP-9-protein levels are markedly elevated in the braintissues of the INCL patient (Fig. 3D), although whetherincreased levels of MMPs in brain tissues of Ppt1-KO miceand in those of the INCL patient were mediated by IL-17Astill remained unclear.
IL-17A stimulates production of MMPs in cultured brainendothelial cells
To determine whether IL-17A stimulated the expression of theMMPs, we performed in vitro experiments in which culturedmurine brain endothelial cells were treated with IL-17A anddetermined the levels of MMPs by western blot analysis. Theuntreated endothelial cells served as controls. Our resultsshowed that the levels of MMP-2, MMP-3 and MMP-9 wereall elevated in IL-17A-treated cultured murine brain endothelialcells (Fig. 3E). Since the astrocyte population in the brain ofPpt1-KO mice increase with age contributing to neuroinflam-mation (22), we also sought to determine whether these cellsexpress IL-17A receptor (IL-17R) and are responsive toIL-17A treatment increasing the levels of MMPs. Accordingly,we determined the levels of IL-17R and MMP-2, MMP-3 andMMP-9 in untreated and IL-17A-treated cultured astrocytesfrom Ppt1-KO mice. The results showed that the culturedastrocytes from Ppt1-KO mice expressed IL-17R, which wasfurther augmented by IL-17A treatment (Supplementary Mater-ial, Fig. S5A). Moreover, treatment of these cells with IL-17Aalso stimulated the production of MMP-2, MMP-3 andMMP-9 (Supplementary Material, Fig. S5B). Although theseresults strongly suggest that TH17 lymphocytes mediate the dis-ruption of the BBB via IL-17A-stimulated production of MMPs
by brain endothelial cells and astrocytes, we cannot rule out thepossibility that other pro-inflammatory mediators causing neu-roinflammation (23,24) do not also contribute to the BBB dis-ruption in these mice.
RSV protects the BBB in Ppt1-KO mice
Previous reports have indicated that oxidative stress is prevalentin cultured cells as well as postmortem brain tissues from INCLpatients (25). MoreoverPpt1-KO mice (10) reliably recapitulateINCL phenotype (11) and manifest increased oxidative stress(25–27) as well as neuroinflammation (24). Furthermore, ithas been reported that both oxidative stress (28) and neuroin-flammation (22) contribute to BBB disruption (29). Additional-ly, we have reported that in Ppt1-KO mice, disruption ofadaptive energy metabolism contributed to neurodegenerationand dietary supplementation of RSV ameliorated this abnormal-ity (27). Recent reports also indicate that RSV improves mito-chondrial function (30), which is also abnormal in Ppt1-KOmice (27). Thus, we sought to determine whether compoundssuch as RSV with anti-oxidant and anti-inflammatory propertiesmay protect the BBB. Accordingly, we fed Ppt1-KO mice witheither control diet or RSV diet and determined the levels ofIL-17A, MMPs and tight junction proteins. Our resultsshowed that compared with Ppt1-KO mice on control diet,their Ppt1-KO littermates on RSV diet showed markedlydecreased levels of IL-17A (Fig. 3F) as well as MMP-2,MMP-3 and MMP-9 (Fig. 3G) and elevated the levels of occlu-din as well as claudin-5 (Fig. 3H). Taken together, these resultsstrongly suggested that RSV treatment has protective effects onthe BBB of Ppt1-KO mice.
RSV suppresses differentiation of TH17 cells from naıveCD41 T cells
How might RSV mediate improvement of the BBB in thePpt1-KO mice? Recent reports indicate that oxidative stresshas deleterious effects, which enhances permeability of theBBB (31). Moreover, disruption of the BBB and oxidativestress has been reported to be involved in the pathogenesis ofinflammatory brain disease (32). Further, TH17 lymphocyteshave been suggested to disrupt the BBB and cause neuroinflam-mation (4). We have previously reported that Ppt1-KO mice,which develop neuroinflammation and manifest high levels ofoxidative stress, when treated with RSV had improvement ofthese pathological conditions (27). Thus, we rationalized thatdue to its anti-oxidative and anti-inflammatory properties,RSV may also suppress the differentiation of TH17 cells fromnaıve CD4+ T cells and consequently, reduce the levels ofpro-inflammatory IL-17A levels in Ppt1-KO mice. We there-fore evaluated the effect of RSV on the differentiation ofTH17 phenotype from naıve (CD4+CD252CD62LhiCD44lo) Tcells. We isolated and cultured these naıve CD4+ T cells fromthe spleens of Ppt1-KO mice under TH17 polarizing conditionsand analyzed for IL-17A expression on day 5 of culture.The results showed that compared with the dimethylsulfoxide(DMSO)-treated controls, the treatment of naıve (CD4+CD252-
CD62LhiCD44lo) T cells with varying doses of RSV significant-ly inhibited TH17 differentiation (Fig. 4A–D). Moreover,compared with the level of TH17 cells in the spleen of WT
2238 Human Molecular Genetics, 2012, Vol. 21, No. 10
Dow
nloaded from https://academ
ic.oup.com/hm
g/article-abstract/21/10/2233/2900646 by guest on 04 April 2019
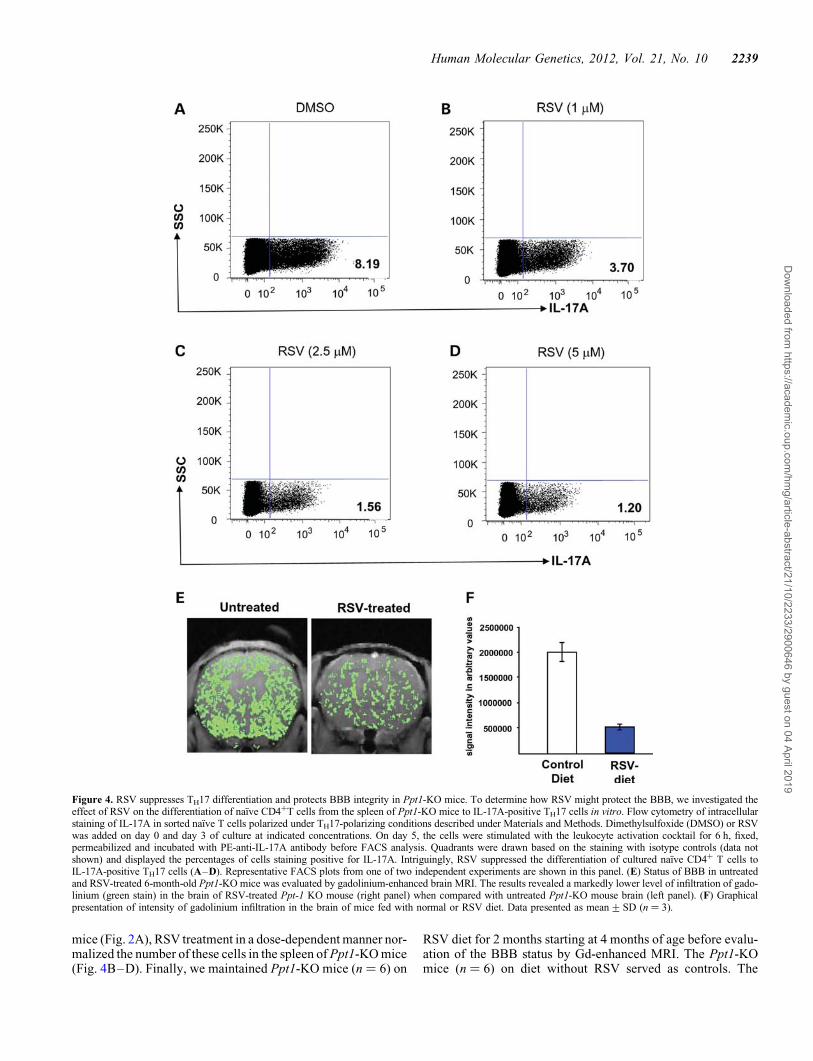
mice (Fig. 2A), RSV treatment in a dose-dependent manner nor-malized the number of these cells in the spleen of Ppt1-KO mice(Fig. 4B–D). Finally, we maintained Ppt1-KO mice (n ¼ 6) on
RSV diet for 2 months starting at 4 months of age before evalu-ation of the BBB status by Gd-enhanced MRI. The Ppt1-KOmice (n ¼ 6) on diet without RSV served as controls. The
Figure 4. RSV suppresses TH17 differentiation and protects BBB integrity in Ppt1-KO mice. To determine how RSV might protect the BBB, we investigated theeffect of RSV on the differentiation of naıve CD4+T cells from the spleen of Ppt1-KO mice to IL-17A-positive TH17 cells in vitro. Flow cytometry of intracellularstaining of IL-17A in sorted naıve T cells polarized under TH17-polarizing conditions described under Materials and Methods. Dimethylsulfoxide (DMSO) or RSVwas added on day 0 and day 3 of culture at indicated concentrations. On day 5, the cells were stimulated with the leukocyte activation cocktail for 6 h, fixed,permeabilized and incubated with PE-anti-IL-17A antibody before FACS analysis. Quadrants were drawn based on the staining with isotype controls (data notshown) and displayed the percentages of cells staining positive for IL-17A. Intriguingly, RSV suppressed the differentiation of cultured naıve CD4+ T cells toIL-17A-positive TH17 cells (A–D). Representative FACS plots from one of two independent experiments are shown in this panel. (E) Status of BBB in untreatedand RSV-treated 6-month-old Ppt1-KO mice was evaluated by gadolinium-enhanced brain MRI. The results revealed a markedly lower level of infiltration of gado-linium (green stain) in the brain of RSV-treated Ppt-1 KO mouse (right panel) when compared with untreated Ppt1-KO mouse brain (left panel). (F) Graphicalpresentation of intensity of gadolinium infiltration in the brain of mice fed with normal or RSV diet. Data presented as mean+SD (n ¼ 3).
Human Molecular Genetics, 2012, Vol. 21, No. 10 2239
Dow
nloaded from https://academ
ic.oup.com/hm
g/article-abstract/21/10/2233/2900646 by guest on 04 April 2019

results showed that compared with the Ppt1-KO mice on dietwithout RSV (Fig. 4E), the Ppt1-KO mice on RSV dietshowed significantly improved BBB status (Fig. 4F). Taken to-gether, these results showed that RSV protects the BBB in thePpt1-KO mice most likely through its anti-oxidative and anti-inflammatory properties as well as by its heretofore unknownproperty as suppressor of TH17 differentiation from naıveCD4+ T cells.
DISCUSSION
In this study, we have determined that BBB is disrupted in thePpt1-KO mice, a reliable animal model of INCL. Moreover,we show that elevated levels of RORgt-positive TH17 cellsare present in the brain as well as in the spleen of thesemice. We further demonstrate that IL-17A, produced byTH17 cells, stimulates the production of MMPs, which arereported to degrade tight junction proteins, which are essentialfor maintaining the integrity of the BBB. Importantly, wefound that Ppt1-KO mice put on a diet containing RSV, apolyphenol present in most plants including red grapes thathas anti-oxidant and anti-inflammatory properties, amelioratethe abnormalities that can disrupt the BBB. Most unexpected-ly, RSV was found to suppress TH17 differentiation, which atleast in part, one of its effects that may protect the BBB.
We previously reported that the Ppt1-KO mice suffer fromendoplasmic reticulum (33,34) as well as oxidative stress (25–27), which contribute to neuropathology in these mice. Wealso reported that the deficiency of Ppt1 in these mice mostlikely impairs dynamic palmitoylation of proteins that are crit-ical for synaptic vesicle recycling and regeneration (35),causing progressive decline in SV pool at the neuronal synap-ses (35,36). The results of our present experiments indicatethat oxidative stress caused by the deficiency of PPT1 maycontribute to many abnormalities, including disruption of theBBB. This assumption may be supported by the fact thatRSV appears to ameliorate this abnormality at least in partdue to its anti-oxidant property.
The disruption of the BBB is a serious complication innumerous neurodegenerative diseases, although to ourknowledge this complication in INCL has not been reportedpreviously. It allows blood components including immunecells to freely enter the brain. Some of these componentsmay cause neuronal death facilitating rapid progression ofthe disease. A clear understanding of the mechanism(s) bywhich the BBB is disrupted in a rare neurodegenerativedisease such as INCL may have implications for other morecommon neurodegenerative disorders, such as Alzheimer’sand Parkinson’s diseases. Such understanding may facilitatenot only the development of effective therapies to protectthe BBB but also the delivery of potentially therapeutic anddiagnostic compounds to specific areas of the brain (37). Onthe basis of our findings in this study, we present a model(Fig. 5) to explain a possible mechanism of BBB disruptionin neurodegenerative disorders, including INCL. In thismodel, we propose that increased IL-17A levels in the bloodof Ppt1-KO mice stimulate the endothelial cells of the brainmicrovasculature (constituent of the BBB) expressingIL-17R to produce high levels of MMPs. The MMPs then
mediate degradation of the tight junction proteins that are es-sential for maintaining the BBB integrity. Following thisevent, the blood components including IL-17A enter thebrain and stimulate the astrocytes that also express IL-17Rto produce more MMPs, which in turn further degrade thetight junction proteins disrupting the BBB. Despite the factthat disruption of the BBB is a serious complication frequentlyencountered in chronic neurodegenerative and neuroinflamma-tory diseases, the development of effective treatment remainschallenging. Our findings define a mechanism by which TH17lymphocytes via induction of MMPs mediate BBB disruptionand provide the proof of principle that small molecules such asRSV, which reduce oxidative stress and suppress TH17lymphocyte differentiation, may have therapeutic potentialfor protecting the BBB in neurodegenerative disorders, includ-ing INCL.
MATERIALS AND METHODS
Animals
Ppt1-KO mice (a generous gift from Dr S.L. Hofmann, Univer-sity of Texas Southwestern Medical Center, Dallas, TX, USA)were generated and characterized as previously described(10). These mice (generated using ES cells from 129 mousestrain) were backcrossed 10 generations with C57BL/6 strainin the laboratory of Dr M. S. Sands (Washington UniversitySchool of Medicine, St Louis, MO,USA) until congenicC57BL/6 genetic background was reached. We are grateful toDr Sands for providing a mating pair, which started ourmouse colony. Herterozygous mice (Ppt1+/2) were matedand the progeny were genotyped. All mice were maintainedand housed in a germ-free facility and animal procedureswere carried out in accordance with institutional guidelinesafter approval of an animal study protocol by the NICHDAnimal Care and Use Committee. The mice were fed standardnormal NIH-31 diet (Zeigler Brothers, Gardner, PA, USA) orNIH-31 diet containing resveratrol (RSV diet) as previouslydescribed (27,30). Briefly, 4 ml of RSV stock solution (50 mgRSV/ml of ethanol) was sprayed onto 100 g of pulverizeddiet. The final concentration of RSV in the diet was 2 mg ofRSV/gram of diet, which provided a dose of 600 mg of RSV/day/kg body weight.
Postmortem brain tissues from INCL patient and normalsubject
A clinical protocol (#01-CH-0086) was approved by the insti-tutional review board. Postmortem brain tissue samples wereobtained from the Human Brain and Spinal Fluid ResourceCenter, VA West Los Angeles Healthcare Center, LosAngeles, CA 90073, USA, which is sponsored by NINDS/NIMH, National Multiple Sciences Society and Departmentof Veterans Affairs. We also obtained control autopsy braintissue samples from the Brain and Tissue Bank for Develop-mental Disorders, University of Maryland, Baltimore, MD21201, USA.
2240 Human Molecular Genetics, 2012, Vol. 21, No. 10
Dow
nloaded from https://academ
ic.oup.com/hm
g/article-abstract/21/10/2233/2900646 by guest on 04 April 2019
Western blot analysis and cytokine ELISA
Western blot analyses of proteins from mouse brain tissueswere performed as previously described (25). The primaryantibodies used are: anti-b-actin (1:5000, US Biological),anti-Claudin-1 (1:1000, Cell signaling Technology),anti-Claudin-5 (1:500, Invitrogen), anti-IL17R (1:500, Santa-cruz Biotech), anti-JAM-1(1:1000, Abcam), anti-MMP2(1:1000, Abcam), anti-MMP3 (1:1000, Abcam), anti-MMP9(1:1000, Cell Signaling Technology), anti-Occludin (1:500,Invitrogen) and anti-RORgt (1:500, Abcam). The secondaryantibodies used are: goat anti-rabbit IgG, donkey anti-goatIgG and goat anti-mouse IgG (Santa Cruz Biotechnology).Chemiluminescent detection was performed by using Super-signal west pico luminol/enhancer solution (Thermo scientific)according to the manufacturer’s protocol. Amount ofproteins from brain lysates and sera from WT mice as wellas those of their Ppt1-KO littermates were adjusted to a
concentration of 0.5 mg/ml and used for enzyme linked im-munosorbent assay (ELISA) (Assaygate Inc.) for quantitatingthe IL-17A protein.
IL-17A treatment of cultured mouse brain endothelial cells
Immortalized mouse brain endothelial cells (bEnd.3) (Ameri-can Type Culture Collection) were cultured in Dulbecco’sminimal essential medium supplemented with 10% FBS at378C under humidified atmosphere containing 5% CO2. Daybefore treatment with IL-17A, cells were trypsinized andseeded in six-well plate at a density of 5 × 105 cells/well.Next day, cells were treated with or without 100 ng/ml of re-combinant IL-17A protein (Biolegend, USA) for 18 h. Proteinsamples were prepared from the treated as well as controluntreated endothelial cells and were used for western blotanalysis.
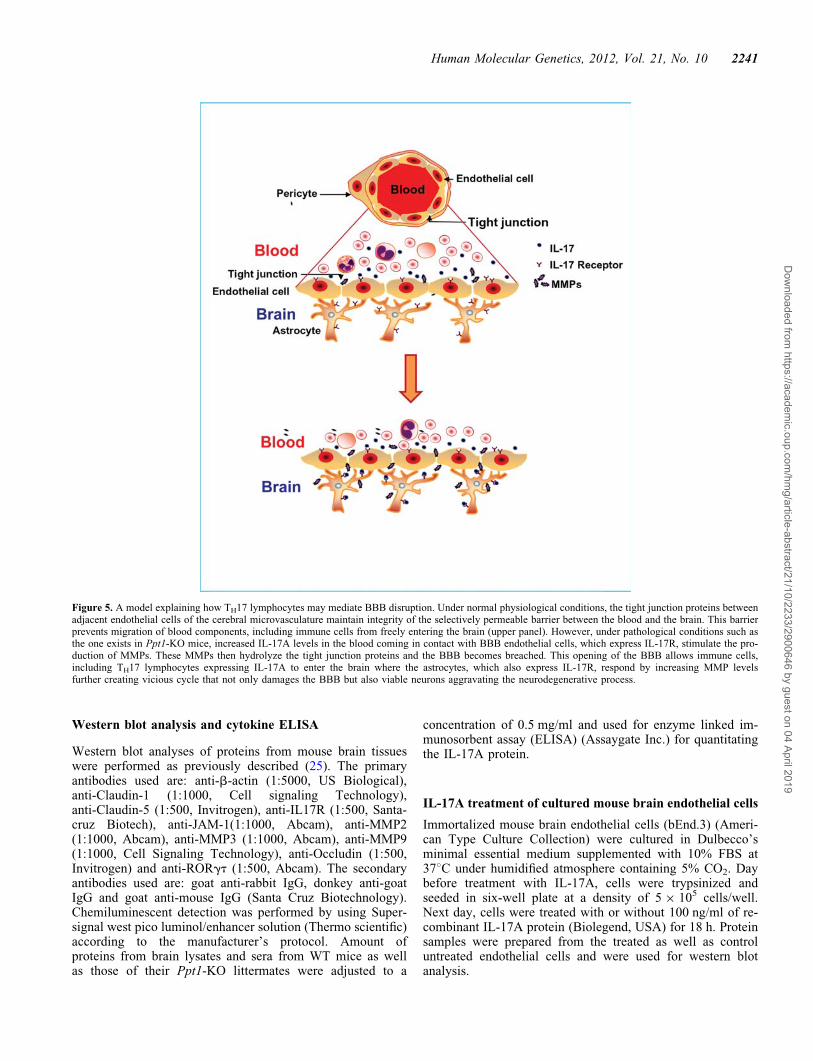
Figure 5. A model explaining how TH17 lymphocytes may mediate BBB disruption. Under normal physiological conditions, the tight junction proteins betweenadjacent endothelial cells of the cerebral microvasculature maintain integrity of the selectively permeable barrier between the blood and the brain. This barrierprevents migration of blood components, including immune cells from freely entering the brain (upper panel). However, under pathological conditions such asthe one exists in Ppt1-KO mice, increased IL-17A levels in the blood coming in contact with BBB endothelial cells, which express IL-17R, stimulate the pro-duction of MMPs. These MMPs then hydrolyze the tight junction proteins and the BBB becomes breached. This opening of the BBB allows immune cells,including TH17 lymphocytes expressing IL-17A to enter the brain where the astrocytes, which also express IL-17R, respond by increasing MMP levelsfurther creating vicious cycle that not only damages the BBB but also viable neurons aggravating the neurodegenerative process.
Human Molecular Genetics, 2012, Vol. 21, No. 10 2241
Dow
nloaded from https://academ
ic.oup.com/hm
g/article-abstract/21/10/2233/2900646 by guest on 04 April 2019

Evaluation of BBB status by gadolinium (Gd)-enhancedMRI
Age- and sex-matched Ppt1-KO mice and their WT litter-mates (n ¼ 5), at age 3 and 6 months were investigated forthe status of BBB Ppt1-KO mice. Mice were anesthetizedwith 1.5% isofluorane and a tail vein catheter was placed,for administering the contrast agent, Gd-DTPA (Bayer Phar-maceuticals Inc., NJ, USA) and positioned in a steriotaxicholder. The body core temperature was maintained at 378Cusing a circulating water pad. MRI was performed on a hori-zontal 7T Bruker (Bruker Biospin Inc., Bellerica, MA, USA)Avance scanner with the brain centered in a 72/25 mmtransmit/receive coil ensemble. T1-weighted axial images(matrix 256 × 256, number of averages ¼ 8, echo trainlength ¼ 8, repetition time (TR) ¼ 300 ms and echo time(TE) ¼ 6 ms, field of view ¼ 1.92 cm, slice thickness ¼1 mm), specified via a tri-axial pilot scan to encompass thewhole brain, were acquired using a gradient echo pulse se-quence. Post-contrast images, with identical parameters,were acquired using the same gradient echo sequence 5 minafter infusion of Gd DTPA (0.2 cm3/kg body weight). Theimages accentuating the relative contrast enhancement wereevaluated using MATLAB (Mathworks Inc., Natick, MA,USA) software.
Cell purification and flow cytometry
Mice were anesthetized and perfused with cold phosphate buf-fered saline (PBS) to remove leukocytes from the microvascu-lature of the brain (constituent of the BBB). Brain tissueswere manually minced in Hank’s balanced salt solution(HBSS) (Mediatech Inc.), supplemented with 5% FBS(HyClone) and filtered through a 70 mm nylon mesh cell strainerusing a rubber policeman. The resulting slurry was digested for30 min at 378C in HBSS supplemented with 2 mg/ml collage-nase type I (Sigma-Aldrich) and 5000 U/ml DNase I (Invitro-gen) to obtain a single-cell suspension. Cells wereresuspended in 37% percoll and gently laid over 70% percollin 15 ml tubes. Cells were centrifuged at 2400 rpm for 20 minin a swinging bucket rotor with centrifuge breaks turned off.After centrifugation, myelin debris was carefully aspiratedand the ring containing mononuclear cells at the interface wascollected, and washed extensively in HBSS. For analyzingIL-17A and transcription factor RORgt, cells were stimulatedwith Leukocyte Activation Cocktail, with GolgiPlusTM (BDPharmingen) for 6 h at 378C. Subsequently, cells were fixed,and permeabilized using Cytofix/CytoPerm Plus kit (BDPharmingen) according to the manufacturer’s protocol, andincubated with Fc block (BD Biosciences) to minimize non-specific antibodies binding followed by staining withPE-anti-IL-17A or PE-anti-RORgt (eBioscience). For isolationof splenocytes, spleen was harvested and perfused with PBS andcells were collected after centrifugation. RBCs were lysed withACK lysis buffer at room temperature. Single-cell splenocytesuspensions were incubated with Fc block (BD Biosciences)to minimize non-specific antibodies binding followed by stain-ing with FITC-anti-CD4 and Pacific blue-anti-CD3 for 30 minat room temperature. For analyzing IL-17A-producing cellsfrom the spleen, cells were activated, fixed, permeabilized and
stained with PE-anti-IL-17A antibody as stated above. Cellswere analyzed on a FACSCalibur cytometer with CellQuest(BD Biosciences), and data were analyzed using FlowJO(Tree Star). Control-included cells stained with directly conju-gated isotype control Abs to assess the degree of non-specificstaining.
Naıve CD41 T cell isolation and in vitro polarization
Naıve CD4+ T cells were isolated from mouse spleen usingCD4+ T cell isolation kit (Miltenyi Biotec) according to themanufacturer’s protocol. Cells were stained with APCcy7-anti-CD4, PE-anti-CD25, FITC-anti-CD62L (BD Pharmingen)and eFluor 605NC-anti-CD44 (eBioscience). Cell sorting wasperformed with a FacsAria cell sorter (Becton Dickinson) toobtain a naıve cell population of CD4+CD252CD62LhiCD44lo
T cells. Naıve T cells (0.5 × 106 cells/well) were cultured in48-well plate at 378C and 5% CO2 in T cell media:RPMI-1640 (Invitrogen) supplemented with 10% (vol/vol)heat-inactivated FCS, 1% L-glutamine, 1% penicillin/strepto-mycin and 50 mM 2-mercaptoethenol. Cells were stimulatedwith anti-CD3/CD28 coated Dynabeads (Invitrogen) at abead-to-cell ratio of 1:1. To polarize naıve cells to TH17 pheno-type, cells were cultured for 5 days in IL-6 (20 ng/ml), IL-1b(10 ng/ml), TNF-a (10 ng/ml), IL-23 (20 ng/ml), TGF-b 1.2(5 ng/ml), anti-IL-4 (0.4 mg/ml), anti-IL-12 (2 mg/ml) andanti-IFN-g (8 mg/ml). To study the effect of RSV on TH17 dif-ferentiation, DMSO (control) or varying concentrations of RSV(1, 2.5 and 5 mM) were added to the culture media at day 0 andsubsequently on alternate days. For IL-17A analysis, the cellswere stimulated with Leukocyte Activation Cocktail, withGolgiPlusTM for 6 h at 378C. Intracellular staining was per-formed according to manufacturer’s protocol using Cytofix/CytoPerm Plus kit (BD Pharmingen) with PE-anti-IL-17A(eBioscience). An LSR II (BD Bioscience) and FlowJo softwarewere used for flow cytometry and analysis.
Statistical analysis
Statistical analyses of the data were performed by Student’st-test using Excel Office 2000 (Microsoft) and a P-value of,0.05 was considered statistically significant.
SUPPLEMENTARY MATERIAL
Supplementary Material is available at HMG online.
AUTHOR CONTRIBUTIONS
A.S. and C.S. designed majority of the experiments. C.S. andS.P.S. designed and executed the FACS analyses for RORgtand TH17 lymphocytes. A.S., C.S. and S.P.S. analyzed thedata. J.M., G.C. and Z.Z. executed the MRI experiments andanalyzed the results. S.P., G.C. and E.K. performed the invivo RSV experiments and analyzed the results. A.B.M. con-ceived the project, provided guidance for experimentaldesign and analysis of data as well as wrote the manuscriptwith input from the co-authors.
2242 Human Molecular Genetics, 2012, Vol. 21, No. 10
Dow
nloaded from https://academ
ic.oup.com/hm
g/article-abstract/21/10/2233/2900646 by guest on 04 April 2019

ACKNOWLEDGEMENTS
We thank Dr S.L. Hofmann for the generous gift of thePpt1-KO mice (10) and Dr Mark Sands for providing amating pair of congenic C57BL/6 Ppt1-KO mice, whichwere used to establish our Ppt1-KO mouse colony. Wethank Dr J.M. Farber, Laboratory of Molecular Immunology,NIAID, NIH, for valuable discussions on the design of TH17-related experiments, helpful suggestions and critical review ofthe manuscript. We are also grateful to Drs S.W. Levin,I. Owens and J.Y. Chou for critical review of the manuscriptas well as helpful suggestions and A.M. Heffer for her helpin conducting the MRI experiments.
Conflict of Interest statement. None declared.
FUNDING
This research was supported in part by the Intramural Re-search Program of the Eunice Kennedy Shriver National Insti-tute of Child Health and Human Development [NICHD], andby Intramural Research Program of the National Institute ofAllergy and Infectious Diseases (NIAID), NIH.
REFERENCES
1. Zlokovic, B.V. (2008) The blood-brain barrier in health and chronicneurodegenerative disorders. Neuron 57, 178–201.
2. Wilson, N.J., Boniface, K., Chan, J.R., McKenzie, B.S., Blumenschein,W.M., Mattson, J.D., Basham, B., Smith, K., Chen, T., Morel, F. et al.(2007) Development, cytokine profile and function of human interleukin17-producing helper T cells. Nat. Immunol. 8, 950–957.
3. Ivanov, I.I., McKenzie, B.S., Zhou, L., Tadokoro, C.E., Lepelley, A.,Lafaille, J.J., Cua, D.J. and Littman, D.R. (2006) The orphan nuclearreceptor RORgamma directs the differentiation program ofproinflammatory IL-17+ T helper cells. Cell 126, 1121–1133.
4. Kebir, H., Kreymborg, K., Ifergan, I., Dodelet-Devillers, A., Cayrol, R.,Bernard, M., Giuliani, F., Arbour, N., Becher, B. and Prat, A. (2007)Human TH17 lymphocytes promote blood-brain barrier disruption andcentral nervous system inflammation. Nat Med. 13, 1173–1175.
5. Jeyakumar, M., Dwek, R.A., Butters, T.D. and Platt, F.M. (2005) Storagesolutions: treating lysosomal disorders of the brain. Nat Rev Neurosci. 6,713–725.
6. Goebel, H.H. and Wisniewski, K.E. (2004) Current state of clinical andmorphological features in human NCL. Brain Pathol. 14, 61–69.
7. Cooper, J.D., Russell, C. and Mitchison, H.M. (2006) Progress towardsunderstanding disease mechanisms in small vertebrate models of neuronalceroid lipofuscinoses. Biochim Biophys Acta. 1762, 873–889.
8. Jalanko, A. and Braulke, T. (2009) Neuronal ceroid lipofuscinoses.Biochim Biophys Acta. 1793, 697–709.
9. Vesa, J., Hellsten, E., Verkruyse, L.A., Camp, L.A., Rapola, J.,Santavuori, P., Hofmann, S.L. and Peltonen, L. (1995) Mutations in thepalmitoyl protein thioesterase gene causing infantile neuronal ceroidlipofuscinosis. Nature 376, 584–587.
10. Gupta, P., Soyombo, A.A., Atashband, A., Wisniewski, K.E., Shelton,J.M., Richardson, J.A., Hammer, R.E. and Hofmann, SL. (2001).Disruption of PPT1 or PPT2 causes neuronal ceroid lipofuscinosis inknockout mice. Proc. Natl Acad. Sci. USA 98, 13566–13571.
11. Bible, E., Gupta, P., Hofmann, S.L. and Cooper, J.D. (2004) Regional andcellular neuropathology in the palmitoyl protein thioesterase-1 null mutantmouse model of infantile neuronal ceroid lipofuscinosis. Neurobiol. Dis.16 346–359.
12. Grossman, R.I., Braffman, B.H., Brorson, J.R., Goldberg, H.I., Silberberg,D.H. and Gonzalez-Scarano, F. (1988) Multiple sclerosis serial study ofgadolinium-enhanced MR imaging. Radiology 169, 117–122.
13. Tsukita, S., Furuse, M. and Itoh, M. (2001) Multifunctional strands intight junctions. Nature Rev Mol Cell Biol 2, 285–293.
14. Persidsky, Y., Ramirez, S.H., Haorah, J. and Kanmogne, G.D. (2006)Blood-brain barrier: structural components and function under physiologicand pathologic conditions. J Neuroimmune Pharmacol. 1, 223–236.
15. Iwakura, Y., Ishigame, H., Saijo, S. and Nakae, S. (2011) Functionalspecialization of interleukin-17 family members. Immunity. 34, 149–162.
16. Dong, C. (2008) TH17 cells in development: an updated view oftheir molecular identity and genetic programming. Nat. Rev. Immunol. 8,335–348.
17. Zheng, Y., Josefowicz, S.Z., Kas, A., Chu, T.T. and Gavin, M.A. (2007)Genome-wide analysis of Foxo3 target genes in developing and matureregulatory T cells. Nature 445, 936–940.
18. Kaczmarek, L., Lapinska-Dzwonek, J. and Szymczak, S. (2002) Matrixmetalloproteinases in the adult brain physiology: a link between c-Fos,AO-1 and remodeling of neuronal connections? EMBO J. 21, 6643–6648.
19. Rosenberg, G.A. (2009) Matrix metalloproteinases and their multipleroles in neurodegenerative diseases. Lancet Neurol. 8, p205–216 (2009).
20. Rosenberg, G.A. (2002) Matrix metalloproteinases in neuroinflammation.Glia 39, 279–291.
21. Lischper, M., Beuck, S., Thanabalasundaram, G., Pieper, C. and Galla,H.J. (2010) Metalloproteinase mediated occludin cleavage in the cerebralmicrocapillary endothelium under pathological conditions. Brain Res.1326, 114–127.
22. Man, S., Ubogu, E.E. and Ransohoff, R.M. (2007) Inflammatory cellmigration into the central nervous system: a few new twists on an old tale.Brain Pathol. 17, 243–250.
23. Zhang, Z, Lee, YC, Kim, SJ, Choi, MS, Tsai, PC, Saha, A, Wei, H, Xu, Y,Xiao, YJ, Zhang, P, Heffer, A and Mukherjee, AB. (2007) Production oflysophosphatidylcholine by cPLA2 in the brain of mice lacking PPT1 is asignal for phagocyte infiltration. Hum Mol Genet. 16, 837–847.
24. Saha, A., Kim, S.J., Zhang, Z., Lee, Y.C., Sarkar, C., Tsai, P.C. andMukherjee, A.B. (2008) RAGE signaling contributes toneuroinflammation in infantile neuronal ceroid lipofuscinosis. FEBS Lett.582, 3823–3831.
25. Wei, H., Kim, S.J., Zhang, Z., Tsai, P.C., Wisniewski, K.E. andMukherjee, A.B. (2008) ER and oxidative stresses are common mediatorsof apoptosis in both neurodegenerative and non-neurodegenerativelysosomal storage disorders and are alleviated by chemical chaperones.Hum Mol Genet. 17, 469–477.
26. Kim, S.J., Zhang, Z., Lee, Y.C. and Mukherjee, A.B. (2006)Palmitoyl-protein thioesterase-1 deficiency leads to the activation ofcaspase-9 and contributes to rapid neurodegeneration in INCL. Hum Mol
Genet. 15, 1580–1586.27. Wei, H., Zhang, Z., Saha, A., Peng, S., Chandra, G., Quezado, Z. and
Mukherjee, A.B. (2011) Disruption of adaptive energy metabolism andelevated ribosomal p-S6K1 levels contribute to INCL pathogenesis:partial rescue by resveratrol. Hum Mol Genet. 20, 1111–1121
28. Chrissobolis, S. and Faraci, F.M. (2008) The role of oxidative stressand NADPH oxidase in cerebrovascular disease. Trends Mol Med. 14,495–502.
29. Lochhead, J.J., McCaffrey, G., Quigley, C.E., Finch, J., DeMarco, K.M.,Nametz, N. and Davis, T.P. (2010) Oxidative stress increases blood-brainbarrier permeability and induces alterations in occludin duringhypoxia-reoxygenation. J Cereb Blood Flow Metab. 30, 1625–1636.
30. Lagouge, M., Argmann, C., Gerhart-Hines, Z., Meziane, H., Lerin, C.,Daussin, F., Messadeq, N., Milne, J., Lambert, P., Elliott, P. et al. (2006)Resveratrol improves mitochondrial function and protects against metabolicdisease by activating SIRT1 and PGC-1alpha. Cell 127, 1109–1122.
31. Lagrange, P., Romero, I.A., Minn, A. and Revest, P.A. (1999)Transendothelial permeability changes induced by free radicals in an in vitromodel of the blood-brain barrier. Free Radic. Biol.Med. 27: 667–672.
32. Schreibelt, G., Musters, R.J., Reijerkerk, A., de Groot, L.R., van der Pol,S.M., Hendrikx, E.M., Dopp, E.D., Dijkstra, C.D., Drukarch, B. and deVries, H.E. (2006) Lipoic acid affects cellular migration into the centralnervous system and stabilizes blood-brain barrier integrity. J. Immunol.177, 2630–2637.
33. Zhang, Z., Lee, Y.C., Kim, S.J., Choi, M.S., Tsai, P.C., Xu, Y., Xiao, Y.J.,Zhang, P., Heffer, A. and Mukherjee, A.B. (2006) Palmitoyl-proteinthioesterase-1 deficiency mediates the activation of the unfolded proteinresponse and neuronal apoptosis in INCL. Hum Mol Genet. 15, 337–346.
34. Kim, S.J., Zhang, Z., Hitomi, E., Lee, Y.C. and Mukherjee, A.B.(2006) Endoplasmic reticulum stress-induced caspase-4 activationmediates apoptosis and neurodegeneration in INCL. Hum Mol Genet. 15,1826–1834.
Human Molecular Genetics, 2012, Vol. 21, No. 10 2243
Dow
nloaded from https://academ
ic.oup.com/hm
g/article-abstract/21/10/2233/2900646 by guest on 04 April 2019

35. Kim, S.J., Zhang, Z., Sarkar, C., Tsai, P.C., Lee, Y.C., Dye, L. andMukherjee, A.B. (2008) Palmitoyl protein thioesterase-1 deficiencyimpairs synaptic vesicle recycling at nerve terminals contributing toneuropathology in human and mice. J. Clin. Invest. 118, 3075–3086.
36. Virmani, T., Gupta, P., Liu, X., Kavalali, E.T. and Hofmann, S.L. (2005)Progressively reduced synaptic vesicle pool size in cultured neurons
derived from neuronal ceroid lipofuscinosis-1 knockout mice. Neurobiol.Dis. 20, 314–323.
37. Neuwelt, E.A., Bauer, B., Fahlke, C., Fricker, G., Iadecola, C., Janigro,D., Leybaert, L., Molnar, Z., O’Donnell, M.E., Povlishock, J.T. et al.(2011) Engaging neuroscience to advance translational research in brainbarrier biology. Nat Rev Neurosci. 12, 169–182.
2244 Human Molecular Genetics, 2012, Vol. 21, No. 10
Dow
nloaded from https://academ
ic.oup.com/hm
g/article-abstract/21/10/2233/2900646 by guest on 04 April 2019





























