Embed Size (px)
Citation preview

Dialyzability, Protein Binding, and RenalExcretion of Plasma Conjugated Bilirubin
Milford Fulop, … , John Sandson, Paul Brazeau
J Clin Invest. 1965;44(4):666-680. https://doi.org/10.1172/JCI105179.
Research Article
Find the latest version:
http://jci.me/105179-pdf

Journal of Clinical InvestigationVol. 44, No. 4, 1965
Dialyzability, Protein Binding, and Renal Excretion ofPlasma Conjugated Bilirubin *
MILFORD FULOP,t JOHN SANDSON,$ANDPAUL BRAZEAU(From the Departments of Medicine and Pharmacology, Albert Einstein College of Medicine
and the Bronx Municipal Hospital Center, New York, N. Y.)
Conjugated bilirubin is excreted in the urine,but unconjugated bilirubin is generally believednot to be excreted by the kidneys. The basis forthis difference has not been resolved by past stud-ies. Hoover and Blankenhorn reported that bili-rubin was dialyzable only from the plasma of pa-tients who had bilirubinuria, and that it was notdialyzable from the plasma of patients with acho-luric jaundice (3). Most subsequent investiga-tors, however, have maintained that neither con-jugated nor unconjugated bilirubin was dialyz-able or ultrafilterable from plasma (4-7). Thisled to the hypothesis that conjugated bilirubin wasprobably excreted in the urine by a process ofrenal tubular secretion (8).
Studies that were performed in jaundiced dogsin this laboratory gave results consistent with aprocess of glomerular filtration of conjugatedbilirubin (9). We therefore re-examined thequestion of the dialyzability of plasma bilirubin.A small fraction of plasma conjugated bilirubin injaundiced humans and dogs was found to be dia-lyzable, and hence available for urinary excretionby glomerular filtration. The studies indicatedthat this dialyzable fraction was bound to lowmolecular weight carrier(s), which migrated inthe alpha and beta globulin zones on paper electro-phoresis and which sedimented on ultracentri-fugation.
* Submitted for publication August 20, 1964; acceptedDecember 28, 1964.
Preliminary reports of portions of this work havebeen published (1, 2).
Supported by a grant from the U. S. Public HealthService (A-05503).
tAddress requests for reprints to: Dr. Milford Fulop,Albert Einstein College of Medicine, Eastchester Road &Morris Park Ave., New York 61, N. Y.
t Investigator of the Health Research Council of theCity of New York.
MethodsSubjects
The studies were performed in dogs and patients withobstructive jaundice. Jaundice was induced in the dogsby common bile duct ligation, and renal function stud-ies were performed as described previously (9). In twodogs the right ureter was ligated at the time of the com-
mon bile duct operation, and the left ureter was opera-
tively ligated about 10 days later. Both animals were
sacrificed 2 days after the second procedure.All of the patients had predominant conjugated hyper-
bilirubinemia (76 to 87% of total plasma bilirubin), as
judged by the method of Weber and Schalm (10), or
the 5-minute direct-reacting bilirubin. Jaundice was eitherwaxing or stable in each patient.
The diagnoses included extrahepatic biliary obstruc-
tion, hepatitis, and cirrhosis. Renal function studies were
performed after oral water loading, and urine was ob-
tained by spontaneous voiding. The clearance of en-
dogenous creatinine (Ccr) was used to estimate glomeru-lar filtration rate (GFR). Clearance values representan average of two consecutive 1-hour periods in most
patients. Plasma bilirubin studies were conducted seriallyin some patients who were followed through a phase ofrenal failure during their terminal illness.
Clearance studies were performed in patients withplasma bilirubin levels between 9 and 32 mg per 100 ml.The urinary excretion of nonbilirubin diazotizable sub-stances was equivalent to the excretion of an average of5.4 Asg of bilirubin per minute (range, 4.3 to 7.4) in sixnormal subjects. This represented less than 10% of thetotal urinary bilirubin excretion in almost all the jaun-diced patients, and no correction was made for this blank.The nonbilirubin diazo blank in normal dog urine was ofsimilar magnitude. Bilirubin clearance (CB) was cal-culated using total plasma bilirubin in the formula UV/P.None of the subjects received drugs known to interferewith the chemical methods or to affect the plasma proteinbinding of bilirubin. None of the subjects in whom clear-ance estimations were made, or whose urine was studiedby electrophoresis or ultracentrifugation, had pathologicproteinuria as judged by testing with sulfosalicylic acid.
Plasma specimens with high levels of unconjugatedbilirubin were prepared by dissolving bilirubin in 0.01 NNaOH, adding an equal volume of 0.15 Mphosphate buf-fer (pH 7.9), and then adding this bilirubin solution tonormal human or dog plasma (final pH 8.3).
666

PROTEIN BINDING OF CONJUGATEDBILIRUBIN
Analytical
A. Chemical. Bilirubin levels in plasma and urine wereestimated by the diazotization method of Malloy andEvelyn (11) adapted for the Coleman junior spectropho-tometer. Bilirubin in plasma dialyzates was estimatedfrom a) the OD in the range 433 to 450 m~umeasured ina Zeiss PMQII spectrophotometer in cuvettes of 5-cmlight path and b) the ODof the derivative violet azo pig-ment, measured in a Coleman senior or Zeiss spectro-photometer in cuvettes of 5-cm light path. In a few ofthe more recent studies (see Figure 3), diazotization wasaccomplished using 1 vol of dialyzate and 1 vol of re-agent without methanol. This method of performingthe diazo reaction yielded higher optical density val-ues than those obtained with the conventional propor-tions of 1: 6 or 1: 9 because of the smaller total volumecontaining the derivative azobilirubin. Bilirubin con-centrations in jaundiced urine and in jaundiced uremicplasma dialyzates were almost identical whether esti-mated this way or by using the standard method. Themodification was therefore used to calculate bilirubinconcentration in selected nonuremic plasma dialyzates.In six such dialyzates the bilirubin concentration esti-mated in this way averaged 69% (range, 62 to 81%) ofthat estimated from the absorbance at 433 to 450 miu.
Amino acid estimations were made using a Ninhydrinreagent (12). Serum urea nitrogen levels were esti-mated on an autoanalyzer.1 Hexuronic acid was deter-mined by the method of Dische (13). The other chemi-cal methods were described previously (9).
B. Ultrafiltration. Ultrafiltration of plasma was per-formed using the technique of Toribara, Terepka, andDewey (14). Plasma specimens in cellophane casing2were centrifuged for 3 to 4 hours at 2,000 rpm at roomtemperature, after gassing the apparatus with 5% CO-95%o N2. Similar results were obtained in several runsperformed at 40 C.
C. Dialysis. Cellophane casing was soaked in distilledwater for 1 to 2 days at 20° C and then wiped dry anddoubly knotted at one end. After introduction of plasma,the other end was also doubly knotted, leaving the tubinglax. The sacs were rinsed thoroughly with distilled wa-ter to remove traces of bilirubin from their outer sur-faces. When unsoaked fresh casing was used, the re-sulting pale yellow plasma dialyzates yielded spectral ab-sorption curves characteristic of bilirubin. The deriva-tive azo pigment, however, was brownish-violet instead ofviolet, although the OD at 540 to 560 m/u was the sameas when presoaked casing was used for simultaneous dialy-sis or ultrafiltration of paired specimens.
The dialysis sacs were placed in Erlenmeyer flaskswith phosphate-buffered saline,3 and the flasks weregassed with 5% C02-95% N2, stoppered, and covered
1 Technicon Instrument Corp., Chauncey, N. Y.2 19 mmdiameter, purchased from Will Scientific, Inc.,
Rochester 3, N. Y.3 0.313 g KH2PO4, 1.093 g Na2HPO4, 9.00 g NaCl per
L, pH 7.2.
with aluminum foil. Dialysis was carried out at 40 C ona mechanical shaker. In subjects with normal renalfunction plasma specimens of 5 to 7 ml were dialyzedagainst 5 to 7 ml of dialysis fluid for 21 to 26 hours.This volume relationship was selected in order to heightendialyzate bilirubin concentration. Optical density meas-urements were made on the native dialyzates between350 and 550 miA, and on the diazotized fluids between450 and 700 m/u, with dialyzates of normal plasma as theblanks. Plasma specimens from subjects with renal fail-ure were dialyzed for 48 hours, and the bilirubin concen-trations in these dialyzates were estimated by the diazomethod. When plasma specimens were dialyzed againstan equal volume of buffer, recovery of bilirubin after24 to 48 hours averaged 94%o (range, 91 to 101%o, 5 stud-ies). When large volumes of buffer were used, recoveryof bilirubin after 48 hours averaged 90%o (range, 87 to94%, 5 studies).
The integrity of the dialysis membranes was confirmedby immunoelectrophoresis (15) of plasma dialyzates, us-ing a potent antiserum to human serum. The methodcould readily detect human serum albumin at a level of25 to 50 ,g per ml. Before immunoelectrophoresis thedialyzates were concentrated fivefold by ultrafiltrationthrough collodion sacs.4 Seven 24-hour dialyzates oficteric nonuremic plasma and eight 48-hour dialyzates oficteric uremic plasma were examined. No serum pro-tein constituents were detectable in any of these speci-mens, indicating that dialyzate albumin concentrationwas less than 0.015 to 0.03%o of that in the originalplasma.
Fresh jaundiced urine was dialyzed on six occasions,and the conjugated bilirubin was found to be 76%o dialyz-able through cellophane (range, 66 to 83%o) after 48hours.
D. Ultracentrifugation. Ultracentrifugation of urine,urine and plasma dialyzates, and plasma ultrafiltrates wasusually carried out for 16 to 18 hours, and in one in-stance for 63 hours, at 80,000 X g in a Spinco model Lultracentrifuge using an angle rotor. Several ultracen-trifugations were also performed in a swinging bucketrotor for 60 to 65 hours at 100,000 X g.
E. Electrophoresis. Filter paper electrophoresis ofplasma and urine was performed in an aluminum foil-covered Beckman-Spinco model R electrophoretic ap-paratus using Whatman 3 MM paper. The use ofthick filter paper permitted the application of 150 to 300,ul of serum or plasma to each strip. Electrophoresis wasperformed in either barbital (r/2 0.075, pH 8.6), phos-phate (r/2 0.075, pH 7.4), or Tris-maleate (pH 8.6) (16)buffer at 4 v per cm. After a 16-hour run the paperstrips were dried for 60 to 90 minutes in a desiccator,which was evacuated to 1 mmHg and protected fromlight. The strips were stained for protein with brom-phenol blue or buffalo blue-black and for bilirubin byboth diazotization and oxidative conversion to biliverdin.The respective bilirubin staining baths consisted of a)
4Carl Schleicher & Schuell, Keene, N. H.
667
MILFORD FULOP, JOHN SANDSON,AND PAUL BRAZEAU
300 350 400 450 500 550X (mLu)
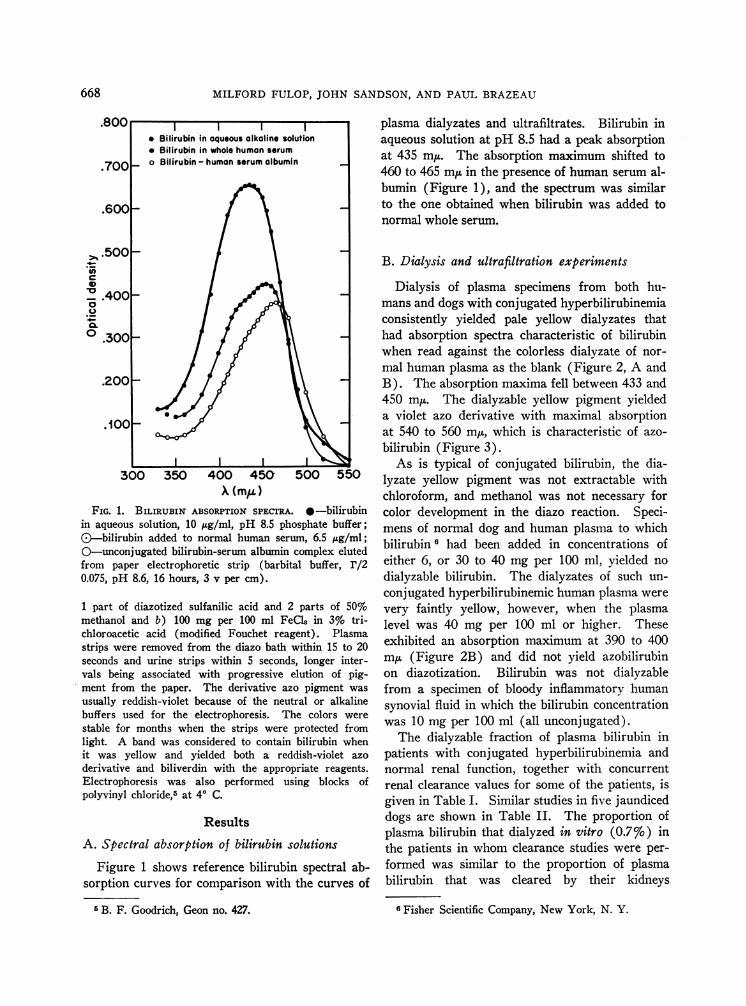
FIG. 1. BILIRUBIN ABSORPTION SPECTRA. 0-bilirubinin aqueous solution, 10 ,ug/ml, pH 8.5 phosphate buffer;&-bilirubin added to normal human serum, 6.5 ,ug/ml;O-unconjugated bilirubin-serum albumin complex elutedfrom paper electrophoretic strip (barbital buffer, r/20.075, pH 8.6, 16 hours, 3 v per cm).
1 part of diazotized sulfanilic acid and 2 parts of 50%methanol and b) 100 mg per 100 ml FeCls in 3% tri-chloroacetic acid (modified Fouchet reagent). Plasmastrips were removed from the diazo bath within 15 to 20seconds and urine strips within 5 seconds, longer inter-vals being associated with progressive elution of pig-ment from the paper. The derivative azo pigment was
usually reddish-violet because of the neutral or alkalinebuffers used for the electrophoresis. The colors were
stable for months when the strips were protected fromlight. A band was considered to contain bilirubin whenit was yellow and yielded both a reddish-violet azo
derivative and biliverdin with the appropriate reagents.Electrophoresis was also performed using blocks ofpolyvinyl chloride,5 at 40 C.
Results
A. Spectral absorption of bilirubin solutions
Figure 1 shows reference bilirubin spectral ab-sorption curves for comparison with the curves of
5 B. F. Goodrich, Geon no. 427.
plasma dialyzates and ultrafiltrates. Bilirubin inaqueous solution at pH 8.5 had a peak absorptionat 435 m~x. The absorption maximum shifted to460 to 465 muin the presence of human serum al-bumin (Figure 1), and the spectrum was similarto the one obtained when bilirubin was added tonormal whole serum.
B. Dialysis and ultrafiltration experiments
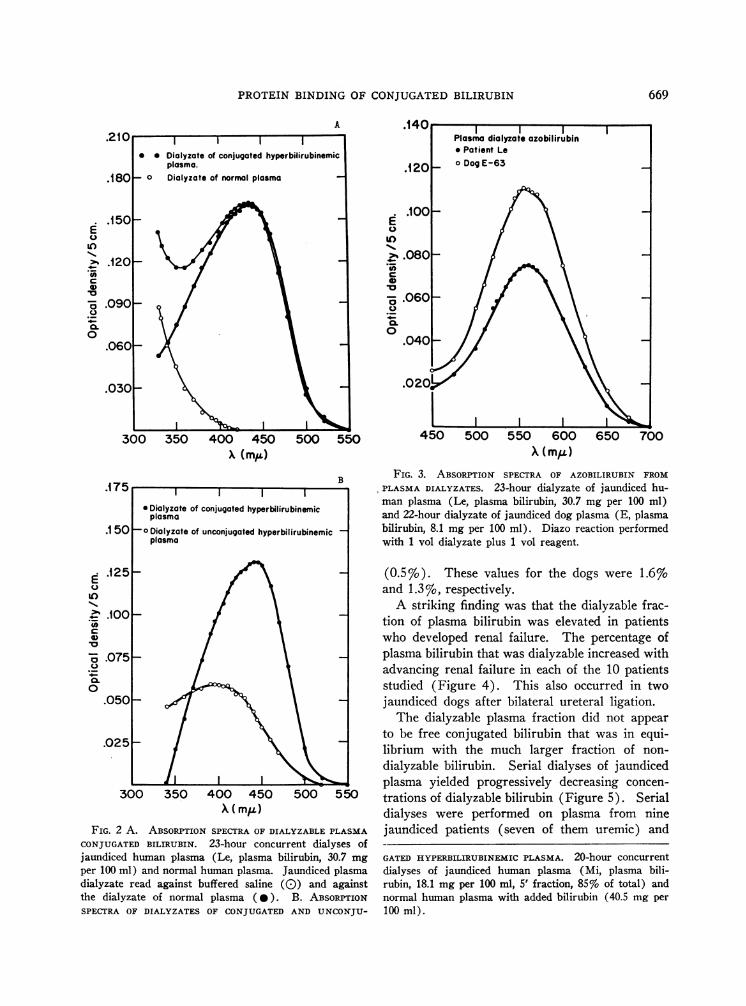
Dialysis of plasma specimens from both hu-mans and dogs with conjugated hyperbilirubinemiaconsistently yielded pale yellow dialyzates thathad absorption spectra characteristic of bilirubinwhen read against the colorless dialyzate of nor-mal human plasma as the blank (Figure 2, A andB). The absorption maxima fell between 433 and450 m~t. The dialyzable yellow pigment yieldeda violet azo derivative with maximal absorptionat 540 to 560 mu, which is characteristic of azo-bilirubin (Figure 3).
As is typical of conjugated bilirubin, the dia-lyzate yellow pigment was not extractable withchloroform, and methanol was not necessary forcolor development in the diazo reaction. Speci-mens of normal dog and human plasma to whichbilirubin 6 had been added in concentrations ofeither 6, or 30 to 40 mg per 100 ml, yielded nodialyzable bilirubin. The dialyzates of such un-conjugated hyperbilirubinemic human plasma werevery faintly yellow, however, when the plasmalevel was 40 mg per 100 ml or higher. Theseexhibited an absorption maximum at 390 to 400m'U (Figure 2B) and did not yield azobilirubinon diazotization. Bilirubin was not dialyzablefrom a specimen of bloody inflammatory humansynovial fluid in which the bilirubin concentrationwas 10 mg per 100 ml (all unconjugated).
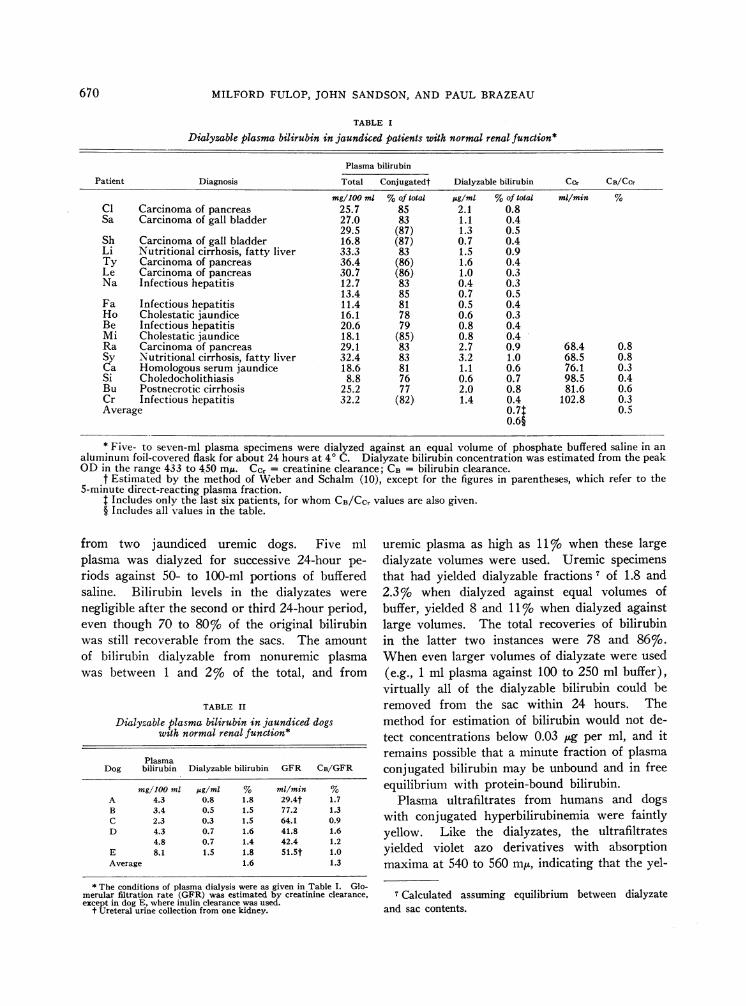
The dialyzable fraction of plasma bilirubin inpatients with conjugated hyperbilirubinemia andnormal renal function, together with concurrentrenal clearance values for some of the patients, isgiven in Table I. Similar studies in five jaundiceddogs are shown in Table II. The proportion ofplasma bilirubin that dialyzed in vitro (0.7%o) inthe patients in whom clearance studies were per-formed was similar to the proportion of plasmabilirubin that was cleared by their kidneys
6 Fisher Scientific Company, New York, N. Y.
668
PROTEIN BINDING OF CONJUGATEDBILIRUBIN
A
Ul
.120-
B.090l
0..060_
.030_
I1 'I I x300 350 400 450 500 550
X (maw)
757 5 B
* Dialyzate of conjugated hyperbilirubinemicplasma
.1 50- 0 Dialyzate of unconjugated hyperbilirubinemicplasma
125 -
.00 3 4
*~.075
0..050
.025-
300 350 400 450 500 550X(mMI)
FIG. 2 A. ABSORPTIONSPECTRAOF DIALYZABLE PLASMACONJUGATEDBILIRUBIN. 23-hour concurrent dialyses ofjaundiced human plasma (Le, plasma bilirubin, 30.7 mgper 100 ml) and normal human plasma. Jaundiced plasmadialyzate read against buffered saline ((0) and againstthe dialyzate of normal plasma (0). B. ABSORPTIONSPECTRA OF DIALYZATES OF CONJUGATEDAND UNCONJU-
X(mMs)
FIG. 3. ABSORPTION SPECTRA OF AZOBILIRUBIN FROM
PLASMA DIALYZATES. 23-hour dialyzate of jaundiced hu-man plasma (Le, plasma bilirubin, 30.7 mg per 100 ml)and 22-hour dialyzate of jaundiced dog plasma (E, plasmabilirubin, 8.1 mg per 100 ml). Diazo reaction performedwith 1 vol dialyzate plus 1 vol reagent.
(0.5%o). These values for the dogs were 1.6%oand 1.3%o, respectively.
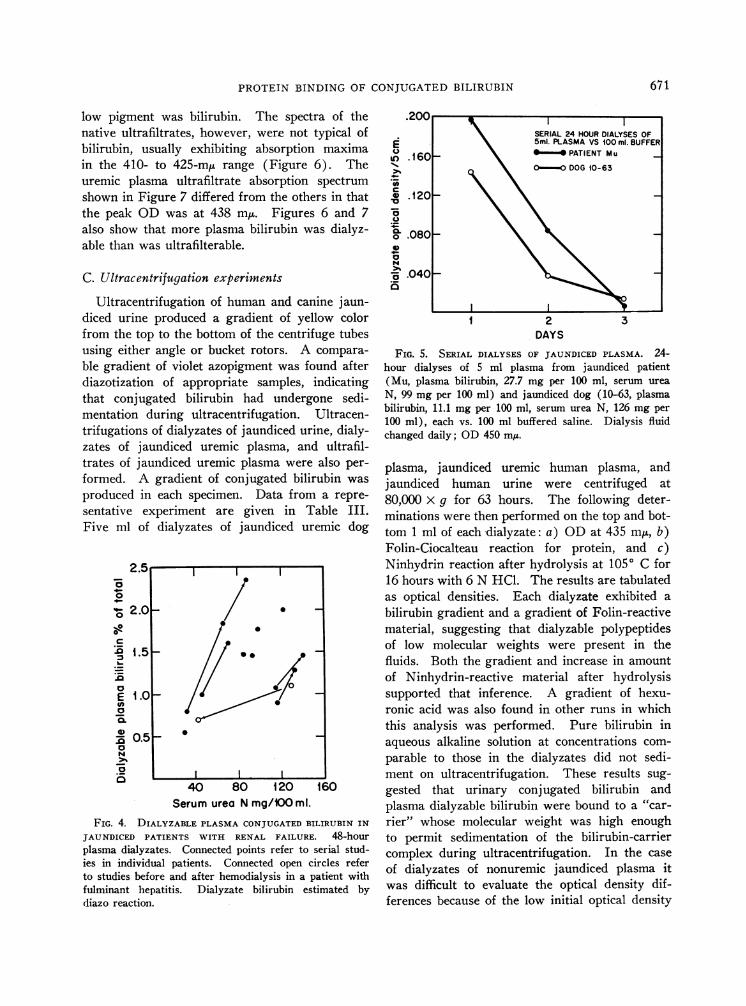
A striking finding was that the dialyzable frac-tion of plasma bilirubin was elevated in patientswho developed renal failure. The percentage ofplasma bilirubin that was dialyzable increased withadvancing renal failure in each of the 10 patientsstudied (Figure 4). This also occurred in twojaundiced dogs after bilateral ureteral ligation.
The dialyzable plasma fraction did not appearto be free conjugated bilirubin that was in equi-librium with the much larger fraction of non-dialyzable bilirubin. Serial dialyses of jaundicedplasma yielded progressively decreasing concen-trations of dialyzable bilirubin (Figure 5). Serialdialyses were performed on plasma from ninejaundiced patients (seven of them uremic) and
GATED HYPERBILIRUBINEMIC PLASMA. 20-hour concurrentdialyses of jaundiced human plasma (Mi, plasma bili-rubin, 18.1 mg per 100 ml, 5' fraction, 85% of total) andnormal human plasma with added bilirubin (40.5 mg per100 ml).
669
MILFORD FULOP, JOHN SANDSON,AND PAUL BRAZEAU
TABLE I
Dialyzable plasma bilirubin in jaundiced patients with normal renal function*
Plasma bilirubin
Patient Diagnosis Total Conjugatedt Dialyzable bilirubin Ccr CB/CCr
mg/100 ml %of total pg/ml %of total ml/min %Cl Carcinoma of pancreas 25.7 85 2.1 0.8Sa Carcinoma of gall bladder 27.0 83 1.1 0.4
29.5 (87) 1.3 0.5Sh Carcinoma of gall bladder 16.8 (87) 0.7 0.4Li Nutritional cirrhosis, fatty liver 33.3 83 1.5 0.9Ty Carcinoma of pancreas 36.4 (86) 1.6 0.4Le Carcinoma of pancreas 30.7 (86) 1.0 0.3Na Infectious hepatitis 12.7 83 0.4 0.3
13.4 85 0.7 0.5Fa Infectious hepatitis 11.4 81 0.5 0.4Ho Cholestatic jaundice 16.1 78 0.6 0.3Be Infectious hepatitis 20.6 79 0.8 0.4Mi Cholestatic jaundice 18.1 (85) 0.8 0.4Ra Carcinoma of pancreas 29.1 83 2.7 0.9 68.4 0.8Sy Nutritional cirrhosis, fatty liver 32.4 83 3.2 1.0 68.5 0.8Ca Homologous serum jaundice 18.6 81 1.1 0.6 76.1 0.3Si Choledocholithiasis 8.8 76 0.6 0.7 98.5 0.4Bu Postnecrotic cirrhosis 25.2 77 2.0 0.8 81.6 0.6Cr Infectious hepatitis 32.2 (82) 1.4 0.4 102.8 0.3Average 0.7t 0.5
0.6§
* Five- to seven-ml plasma specimens were dialyzed against an equal volume of phosphate buffered saline in analuminum foil-covered flask for about 24 hours at 4° C. Dialyzate bilirubin concentration was estimated from the peakOD in the range 433 to 450 mA. Ccr = creatinine clearance; CB = bilirubin clearance.
t Estimated by the method of Weber and Schalm (10), except for the figures in parentheses, which refer to the5-minute direct-reacting plasma fraction.
t Includes only the last six patients, for whomCB/CC, values are also given.§ Includes all values in the table.
from two jaundiced uremic dogs. Five mlplasma was dialyzed for successive 24-hour pe-riods against 50- to 100-ml portions of bufferedsaline. Bilirubin levels in the dialyzates werenegligible after the second or third 24-hour period,even though 70 to 80%o of the original bilirubinwas still recoverable from the sacs. The amountof bilirubin dialyzable from nonuremic plasmawas between 1 and 2%o of the total, and from
TABLE II
Dialyzable plasma bilirubin in jaundiced dogswith normal renal function*
PlasmaDog bilirubin Dialyzable bilirubin GFR CB/GFR
mg/100 ml ,g/ml % ml/min %A 4.3 0.8 1.8 29.4t 1.7B 3.4 0.5 1.5 77.2 1.3C 2.3 0.3 1.5 64.1 0.9D 4.3 0.7 1.6 41.8 1.6
4.8 0.7 1.4 42.4 1.2E 8.1 1.5 1.8 51.5t 1.0Average 1.6 1.3
* The conditions of plasma dialysis were as given in Table I. Glo-merular filtration rate (GFR) was estimated by creatinine clearance,except in dog E, where inulin clearance was used.
t Ureteral urine collection from one kidney.
uremic plasma as high as 11 o when these largedialyzate volumes were used. Uremic specimensthat had yielded dialyzable fractions 7 of 1.8 and2.3% when dialyzed against equal volumes ofbuffer, yielded 8 and 11% when dialyzed againstlarge volumes. The total recoveries of bilirubinin the latter two instances were 78 and 86%.When even larger volumes of dialyzate were used(e.g., 1 ml plasma against 100 to 250 ml buffer),virtually all of the dialyzable bilirubin could beremoved from the sac within 24 hours. Themethod for estimation of bilirubin would not de-tect concentrations below 0.03 ug per ml, and itremains possible that a minute fraction of plasmaconjugated bilirubin may be unbound and in freeequilibrium with protein-bound bilirubin.
Plasma ultrafiltrates from humans and dogswith conjugated hyperbilirubinemia were faintlyyellow. Like the dialyzates, the ultrafiltratesyielded violet azo derivatives with absorptionmaxima at 540 to 560 mpu, indicating that the yel-
7 Calculated assuming equilibrium between dialyzateand sac contents.
670
PROTEIN BINDING OF CONJUGATEDBILIRUBIN
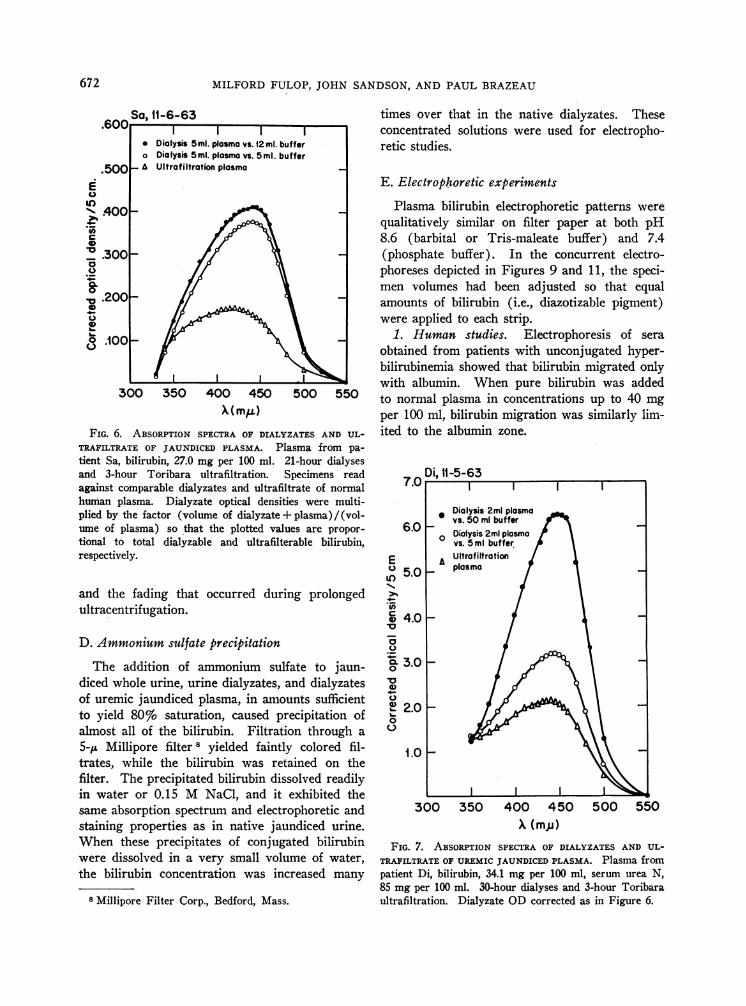
low pigment was bilirubin. The spectra of thenative ultrafiltrates, however, were not typical ofbilirubin, usually exhibiting absorption maximain the 410- to 425-mu range (Figure 6). Theuremic plasma ultrafiltrate absorption spectrumshown in Figure 7 differed from the others in thatthe peak OD was at 438 miu. Figures 6 and 7also show that more plasma bilirubin was dialyz-able than was ultrafilterable.
C. Ultracentrifugation experiments
Ultracentrifugation of human and canine jaun-diced urine produced a gradient of yellow colorfrom the top to the bottom of the centrifuge tubesusing either angle or bucket rotors. A compara-ble gradient of violet azopigment was found afterdiazotization of appropriate samples, indicatingthat conjugated bilirubin had undergone sedi-mentation during ultracentrifugation. Ultracen-trifugations of dialyzates of jaundiced urine, dialy-zates of jaundiced uremic plasma, and ultrafil-trates of jaundiced uremic plasma were also per-formed. A gradient of conjugated bilirubin wasproduced in each specimen. Data from a repre-sentative experiment are given in Table III.Five ml of dialyzates of jaundiced uremic dog
4-._
0
.0
0.
c
E0
4)
00
0
40 80 120Serum urea N mg/100 ml.
160
FIG. 4. DIALYZABLE PLASMACONJUGATEDBILIRUBIN IN
JAUNDICED PATIENTS WITH RENAL FAILURE. 48-hourplasma dialyzates. Connected points refer to serial stud-ies in individual patients. Connected open circles referto studies before and after hemodialysis in a patient withfulminant hepatitis. Dialyzate bilirubin estimated bydiazo reaction.
\ 5m1. PLASMAVS 100 ml. BUFFERn .160 _ \ *-* PATIENT Mu7%Q \ -00 DOG10-63
o .120 _\2
o~ .080-
-.040
1 2 3DAYS
FIG. 5. SERIAL DIALYSES OF JAUNDICED PLASMA. 24-hour dialyses of 5 ml plasma from jaundiced patient(Mu, plasma bilirubin, 27.7 mg per 100 ml, serum ureaN, 99 mg per 100 ml) and jaundiced dog (10-63, plasmabilirubin, 11.1 mg per 100 ml, serum urea N, 126 mg per100 ml), each vs. 100 ml buffered saline. Dialysis fluidchanged daily; OD450 mu.
plasma, jaundiced uremic human plasma, andjaundiced human urine were centrifuged at80,000 x 9 for 63 hours. The following deter-minations were then performed on the top and bot-tom 1 ml of each -dialyzate: a) ODat 435 miu, b)Folin-Ciocalteau reaction for protein, and c)Ninhydrin reaction after hydrolysis at 105° C for16 hours with 6 N HCl. The results are tabulatedas optical densities. Each dialyzate exhibited abilirubin gradient and a gradient of Folin-reactivematerial, suggesting that dialyzable polypeptidesof low molecular weights were present in thefluids. Both the gradient and increase in amountof Ninhydrin-reactive material after hydrolysissupported that inference. A gradient of hexu-ronic acid was also found in other runs in whichthis analysis was performed. Pure bilirubin inaqueous alkaline solution at concentrations com-parable to those in the dialyzates did not sedi-ment on ultracentrifugation. These results sug-gested that urinary conjugated bilirubin andplasma dialyzable bilirubin were bound to a "car-rier" whose molecular weight was high enoughto permit sedimentation of the bilirubin-carriercomplex during ultracentrifugation. In the caseof dialyzates of nonuremic jaundiced plasma itwas difficult to evaluate the optical density dif-ferences because of the low initial optical density
671
MILFORD FULOP, JOHN SANDSON,AND PAUL BRAZEAU
Sa, 11-6-63
300 350 400 450M(mM)
FIG. 6. ABSORPTION SPECTRA OF DIALYZ
TRAFILTRATE OF JAUNDICED PLASMA. Platient Sa, bilirubin, 27.0 mg per 100 ml. 2and 3-hour Toribara ultrafiltration. Sagainst comparable dialyzates and ultrafilthuman plasma. Dialyzate optical densiticplied by the factor (volume of dialyzate +ume of plasma) so that the plotted valuetional to total dialyzable and ultrafilterrespectively.
and the fading that occurred durirultracentrifugation.
D. Ammonium sulfate precipitation
The addition of ammonium sulfadiced whole urine, urine dialyzates, a
of uremic jaundiced plasma, in amou
to yield 80% saturation, caused proalmost all of the bilirubin. Filtratic5-,u Millipore filter s yielded faintlytrates, while the bilirubin was retafilter. The precipitated bilirubin dissin water or 0.15 M NaCl, and it (
same absorption spectrum and electrcstaining properties as in native jautWhen these precipitates of conjugawere dissolved in a very small voluithe bilirubin concentration was inc:
8 Millipore Filter Corp., Bedford, Mass.
times over that in the native dialyzates. Theseconcentrated solutions were used for electropho-
ffer retic studies.!fer
E. Electrophoretic experiments
Plasma bilirubin electrophoretic patterns werequalitatively similar on filter paper at both pH8.6 (barbital or Tris-maleate buffer) and 7.4(phosphate buffer). In the concurrent electro-phoreses depicted in Figures 9 and 11, the speci-men volumes had been adjusted so that equalamounts of bilirubin (i.e., diazotizable pigment)were applied to each strip.
1. Human studies. Electrophoresis of seraobtained from patients with unconjugated hyper-bilirubinemia showed that bilirubin migrated onlywith albumin. When pure bilirubin was added
500 550 to normal plasma in concentrations up to 40 mgper 100 ml, bilirubin migration was similarly lim-
'ATES AND UL- ited to the albumin zone.sma from pa-1-hour dialysespecimens read Di, 11i-5-63.rate of normal 70 I 1es were multi-,plasma) / (vol- * Dialysis 2m1 plasmaes are propor- 6.0 vs. 50 ml bufferYe are bilirub, 6.0 Dialysis 2ml plasmarable bilirubin, ° vs. 5ml buffer.
E A Ultrafiltrationo 5.0 plasma
ig prolonged .
4.0-
CL 3.0-ate to jaaun- 0nd dialyzates lfints sufficient i'ecipitation of ~ .0)n through a
colored fil- 1.lined on theolved readilyexhibited the)phoretic and 300 350 400 450 500 550ndiced urine. X (myu)Lted bilirubin FIG. 7. ABSORPTION SPECTRA OF DIALYZATES AND UL-me of water, TRAFILTRATE OF UREMICJAUNDICEDPLASMA. Plasma fromreased many patient Di, bilirubin, 34.1 mg per 100 ml, serum urea N,
85 mg per 100 ml. 30-hour dialyses and 3-hour Toribaraultrafiltration. Dialyzate ODcorrected as in Figure 6.
672
PROTEIN BINDING OF CONJUGATEDBILIRUBIN
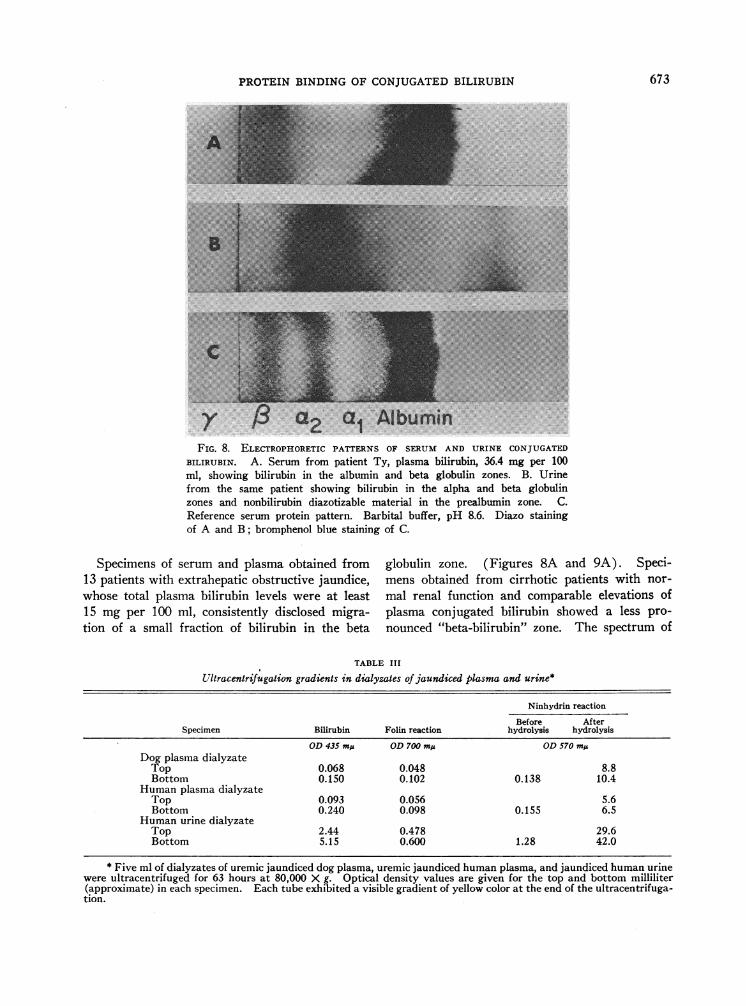
a2 a1 AlbuminFIG. 8. ELECTROPHORETICPATTERNS OF SERUM AND URINE CONJUGATED
BILIRUBIN. A. Serum from patient Ty, plasma bilirubin, 36.4 mg per 100ml, showing bilirubin in the albumin and beta globulin zones. B. Urinefrom the same patient showing bilirubin in the alpha and beta globulinzones and nonbilirubin diazotizable material in the prealbumin zone. C.Reference serum protein pattern. Barbital buffer, pH 8.6. Diazo stainingof A and B; bromphenol blue staining of C.
Specimens of serum and plasma obtained from13 patients with extrahepatic obstructive jaundice,whose total plasma bilirubin levels were at least15 mg per 100 ml, consistently disclosed migra-tion of a small fraction of bilirubin in the beta
globulin zone. (Figures 8A and 9A). Speci-mens obtained from cirrhotic patients with nor-
mal renal function and comparable elevations ofplasma conjugated bilirubin showed a less pro-nounced "beta-bilirubin" zone. The spectrum of
TABLE III
Ultracentrifugation gradients in dialyzates of jaundiced plasma and urine*
Ninhydrin reaction
Before AfterSpecimen Bilirubin Folin reaction hydrolysis hydrolysis
OD435 my OD700 my OD570 mewDog plasma dialyzate
Top 0.068 0.048 8.8Bottom 0.150 0.102 0.138 10.4
Human plasma dialyzateTop 0.093 0.056 5.6Bottom 0.240 0.098 0.155 6.5
Human urine dialyzateTop 2.44 0.478 29.6Bottom 5.15 0.600 1.28 42.0
* Five ml of dialyzates of uremic jaundiced dog plasma, uremic jaundiced human plasma, and jaundiced human urinewere ultracentrifuged for 63 hours at 80,000 X g. Optical density values are given for the top and bottom milliliter(approximate) in each specimen. Each tube exhibited a visible gradient of yellow color at the end of the ultracentrifuga-tion.
673

MILFORD FULOP, JOHN SANDSON,AND PAUL BRAZEAU
B j0ii
FIG. 9. ELECTROPHORETICPATTERNS OF PLASMA CON-JUGATED BILIRUBIN BEFORE AND AFTER UREMIA. Plasmafrom patient Co, plasma bilirubin, 32.9 mg per 100 ml.A. Preuremia. B. Serum urea N, 122 mg per 100 ml.C. Uremic plasma (B) after 48 hours of dialysis againstbuffered saline. Tris buffer, pH 8.6. Diazo staining.
the yellow pigment eluted from the beta globulinzone could not be characterized because the elu-ates were opalescent. Elution of this zone withdiazotized sulfanilic acid, however, yielded a vio-let pigment with the spectral absorption charac-teristics of azobilirubin. Staining of the paper
strips with diazo reagent and with Fouchet re-
agent yielded typical bilirubin derivatives in boththe albumin and beta globulin zones.
Paper electrophoresis of urine obtained fromeach of eight patients with obstructive jaundicedisclosed three zones of yellow diazotizable pig-
ment. Their mobilities corresponded with betaand alpha globulins and prealbumin (Figure 8B),although it was not possible to detect protein inany of these zones with either bromphenol blueor buffalo blue-black. The prealbumin pigmentzone was also present in normal urine, and inboth jaundiced and normal urine its derivativeazopigment was orange rather than violet, sug-
gesting that it was not bilirubin. The mobility ofurinary conjugated bilirubin on paper thereforecorresponded with the beta and alpha globulinzones. The relative proportions were variable,
and there was usually no sharp demarcation be-tween the alpha and beta zones.
Plasma electrophoretic studies were performedin two patients with extrahepatic obstructivejaundice before and after they developed renalfailure. The uremic specimens showed an in-creased proportion of total plasma bilirubin in thebeta globulin zone, and some appeared in thealpha globulin zone (Figure 9B). Dialysis ofthese uremic jaundiced plasma specimens causeda decrease of bilirubin in the beta globulin zone(and probably the alpha also) (Figure 9C). Intwo other jaundiced patients in whom preuremiaspecimens were not available, it was also possibleto demonstrate very prominent "beta- and alpha-bilirubin" zones in the uremic plasma specimensand a decrease in the intensity of those zones afterin vitro dialysis of plasma. The diazotized betaglobulin zone pigment in several of these uremicplasma specimens was eluted from both paperstrips and polyvinyl chloride and was found tohave an absorption spectrum characteristic of azo-bilirubin. Paper electrophoresis of jaundiced ure-mic plasma dialyzates in which the bilirubin hadfirst been concentrated by ammonium sulfateprecipitation disclosed bilirubin migration in thebeta-alpha globulin zone. This was similar to themigration of bilirubin in native jaundiced urineand urine dialyzates, as well as in these fluidsafter concentration by ammonium sulfate pre-cipitation.
On polyvinyl chloride at pH 7.4 (phosphate buf-fer) the pattern of plasma conjugated bilirubinmigration was similar to its pattern on paperelectrophoresis. The pattern in uremic jaundicedplasma was also similar to that on paper, exceptthat a faint prealbumin bilirubin zone was alsodiscernible. Electrophoresis of jaundiced urineand urine dialyzates on polyvinyl chloride showedmigration of most of the bilirubin in the prealbu-min zone, with broad trailing back to the origin.Analysis for Folin-reactive material disclosed sev-eral peaks, one of which coincided with the pre-albumin peak of bilirubin concentration. Thebilirubin concentration in native dialyzates ofjaundiced plasma was too low to permit satisfac-tory electrophoresis. Electrophoresis of uremicjaundiced plasma dialyzates in which the bilirubinhad been first concentrated by ammonium sulfateprecipitation disclosed a bilirubin migration pat-
674
PROTEIN BINDING OF CONJUGATEDBILIRUBIN
.375
t.3000
toIno .2250
za-
AC .150
ON .075
I~~~~~~
I ~~~~~~I \I ~~~~~~~~~I \/ ~~~~~~~~~~~~I
/~~~~~~~~~I
I.~~~~~~~~~~~~~~~~~~~~~~~/
.090
.075
.060I
I
I
z
0
Ull-
.015
15 14 13 12111 10 9 8 7 6 5 4 3 2 1
SEGMENTNUMBER
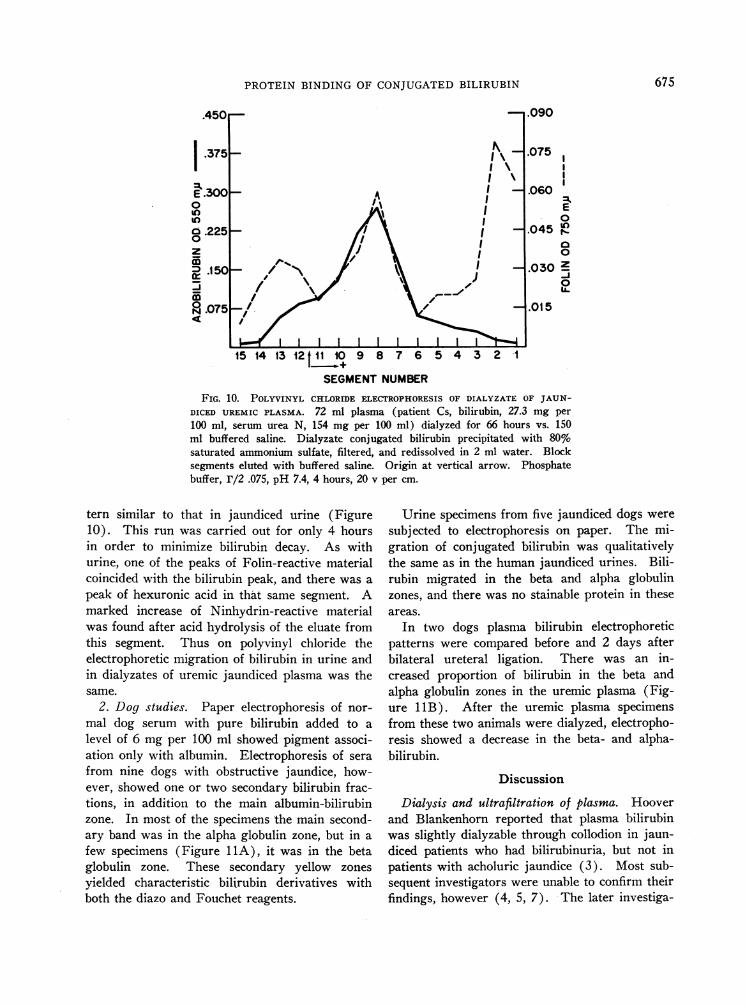
FIG. 10. POLYVINYL CHLORIDE ELECTROPHORESISOF DIALYZATE OF JAUN-DICED UREMIC PLASMA. 72 ml plasma (patient Cs, bilirubin, 27.3 mg per
100 ml, serum urea N, 154 mg per 100 ml) dialyzed for 66 hours vs. 150ml buffered saline. Dialyzate conjugated bilirubin precipitated with 80%saturated ammonium sulfate, filtered, and redissolved in 2 ml water. Blocksegments eluted with buffered saline. Origin at vertical arrow. Phosphatebuffer, r/2 .075, pH 7.4, 4 hours, 20 v per cm.
.045
.030
tern similar to that in jaundiced urine (Figure10). This run was carried out for only 4 hoursin order to minimize bilirubin decay. As withurine, one of the peaks of Folin-reactive materialcoincided with the bilirubin peak, and there was a
peak of hexuronic acid in that same segment. Amarked increase of Ninhydrin-reactive materialwas found after acid hydrolysis of the eluate fromthis segment. Thus on polyvinyl chloride theelectrophoretic migration of bilirubin in urine andin dialyzates of uremic jaundiced plasma was thesame.
2. Dog studies. Paper electrophoresis of nor-
mal dog serum with pure bilirubin added to a
level of 6 mg per 100 ml showed pigment associ-ation only with albumin. Electrophoresis of sera
from nine dogs with obstructive jaundice, how-ever, showed one or two secondary bilirubin frac-tions, in addition to the main albumin-bilirubinzone. In most of the specimens the main second-ary band was in the alpha globulin zone, but in a
few specimens (Figure 1lA), it was in the betaglobulin zone. These secondary yellow zones
yielded characteristic bilirubin derivatives withboth the diazo and Fouchet reagents.
Urine specimens from five jaundiced dogs were
subjected to electrophoresis on paper. The mi-gration of conjugated bilirubin was qualitativelythe same as in the human jaundiced urines. Bili-rubin migrated in the beta and alpha globulinzones, and there was no stainable protein in theseareas.
In two dogs plasma bilirubin electrophoreticpatterns were compared before and 2 days afterbilateral ureteral ligation. There was an in-creased proportion of bilirubin in the beta andalpha globulin zones in the uremic plasma (Fig-ure 11B). After the uremic plasma specimensfrom these two animals were dialyzed, electropho-resis showed a decrease in the beta- and alpha-bilirubin.
Discussion
Dialysis and ultrafiltration of plasma. Hooverand Blankenhorn reported that plasma bilirubinwas slightly dialyzable through collodion in jaun-diced patients who had bilirubinuria, but not inpatients with acholuric jaundice (3). Most sub-sequent investigators were unable to confirm theirfindings, however (4, 5, 7). The later investiga-
.450
I
675
)I

MILFORD FULOP, JOHN SANDSON,AND PAUL BRAZEAU
*:* . . .. . v...
A* .: .. .. .: ..*: :.* . ., . . . A . .
FIG. 11. ELECTROPHORETICPATTERNS OF PLASMA CONJUGATEDBILIRUBINBEFORE AND AFTER UREMIA. Plasma from jaundiced dog 10-63. A. Beforeuremia. B. 2 days after bilateral ureteral ligation. Tris buffer, pH 8.6.Diazo staining.
tors usually studied either normal plasma withadded bilirubin, or specimens obtained from pa-tients with predominant unconjugated hyperbili-rubinemia. The nondialyzability of plasma uncon-jugated bilirubin, recently confirmed in severalstudies as well as the present one, is consistentwith its lack of renal excretion (17-19).
The findings in our earlier renal function ex-periments in jaundiced dogs were consistent witha predominant process of glomerular filtration ofconjugated bilirubin (9). Collodion sac ultra-filtrations of jaundiced plasma failed to yield ul-trafilterable bilirubin in this laboratory. This maybe explained by the observation that urinary bili-rubin was adsorbed to the membrane when jaun-diced urine was ultrafiltered through these col-lodion sacs. The use of cellophane casingpermitted extensive dialysis of urinary bilirubinand also dialysis of a small fraction of plasma con-jugated bilirubin.
Dialyses of plasma from patients with conju-gated bilirubin levels below 9 mg per 100 ml arenot reported because the dialyzate optical densitymeasurements could not be translated accuratelyto concentrations. It has been possible, however,to verify the presence of bilirubin in dialyzatesfrom plasma specimens with conjugated bilirubinlevels as low as 5 mgper 100 ml by demonstratingabsorption maxima at 433 to 450 mu.
The proportion of plasma conjugated bilirubinthat was dialyzable in both humans and dogs was
large enough to account for urinary bilirubin ex-cretion by a process of glomerular filtration.9The precise proportion was difficult to define be-cause of the methods of dialysis and of bilirubinestimation. The typical absorption spectra of thenative pale yellow dialyzates and of their violetazo derivatives suggested that the dialyzableplasma pigment was bilirubin. The dialyzablepercentages of total plasma bilirubin given in Ta-bles I and II, however, are probably underesti-mates. Some methodologic factors resulted in un-derestimating the potential dialyzability, whereasothers might have resulted in overestimation.The most important among the former was theinefficiency of 24-hour dialyses in which plasmaspecimens were dialyzed against equal (small)volumes of dialyzing fluid. The use of largervolumes of bath fluid permitted increased dialysisof plasma bilirubin (Figures 6 and 7), althoughthe total amount dialyzable still remained a smallfraction of the total plasma bilirubin in nonuremicspecimens. The dialyzable fraction may also havebeen underestimated because of slight decay ofbilirubin during dialysis. The use of the peakabsorbance at 433 to 450 m/u probably did not in-troduce a major error because the conformation
9 The values of UBV determined from the diazo reac-tion in urine are overestimates, at least to the extent ofthe nonbilirubin diazo blank. The sufficiency of the dia-lyzable plasma fraction as the source of urinary bilirubinis actually enhanced by any overestimation of UBV.
676

PROTEIN BINDING OF CONJUGATEDBILIRUBIN
of the dialyzate spectral absorption curves (Fig-ure 2) suggested that bilirubin was the majorcontributor to the peak absorbances. It was oftennot possible to estimate dialyzate bilirubin con-
centrations from the concentration of derivativeazobilirubin in nonuremic specimens, becausethese optical densities were generally low.
The dialyzable plasma bilirubin fraction neednot be the same as CB/GFRin order to support theglomerular filtration hypothesis. It would befortuitous for an artificial membrane to have pore
and permeability characteristics identical withthose of the glomerular filter. Moreover, renalglomerular filtration involves ultrafiltration ofplasma rather than dialysis. The cellophane mem-
branes yielded less ultrafilterable than dialyzableplasma bilirubin'0 (Figures 6 and 7). Finally,it is possible that some of the conjugated bili-rubin which is filtered through the renal glomerulimay be reabsorbed, although our previous stud-ies in dogs did not suggest the presence of a ma-
jor tubular reabsorptive process.
The range of values for CB/GFR in the patientsin this report corresponds to that reported byWith (8) and by Wallace and Owen (20) inlarger groups of jaundiced patients. The valuesfor CB/GFR in the dogs (Table II) are in a range
similar to that previously reported from this lab-oratory in a larger series of animals (9).
Wallace and Owen found that CB/CCr was
higher in patients with reduced filtration ratesthan in those with normal Ccr (20). Wehavealso found that jaundiced patients and dogs withreduced GFR tend to have higher CB/GFR val-ues. The inclusion of several such patients in an
earlier publication (2) gave CB/GFR values up
to 0.9%o, and we have recently observed a jaun-diced patient with Ccr of 9 ml per minute whoseCB/CCr was 1.6%o. The conventional calculationof CB does not take into account the increasedpercentage of dialyzable plasma bilirubin that ispresent in patients with decreased GFR (Figure4). Although such subjects may have increasedglomerular permeability to a plasma bilirubin-protein complex, in our view the elevated CB/GFRdepends simply upon the use of either totalor direct-reacting plasma bilirubin as the de-
10 It was observed that urinary bilirubin was only about50% ultrafilterable through these cellophane membranes,whereas it was 70 to 80% dialyzable.
nominator in the clearance formula rather thandialyzable plasma bilirubin.
If the dialyzable plasma fraction was unboundconjugated bilirubin in free equilibrium with themuch larger nondialyzable fraction, repeated di-alyses of the same plasma specimen with serialchanges of bath fluid should yield, at equilibrium,similar bilirubin concentrations in successivedialyzates. The dialyzate bilirubin concentrationdecreased progressively until no more plasma bili-rubin could be dialyzed, even though only a frac-tion of the total bilirubin had been removed.When samples of the same plasma were dialyzedagainst different volumes of buffer, the dialyz-able bilirubin concentration was highest in thesmallest dialyzate, although the calculated amountof dialyzable bilirubin was highest in the largestdialyzate. These findings suggested that most orperhaps all of the dialyzable fraction was not freeconjugated bilirubin in equilibrium with the non-dialyzable plasma bilirubin.
Ultracentrifugation studies. Ultracentrifuga-tion of jaundiced human and dog urine resultedin sedimentation of bilirubin, and similar resultswere obtained with dialyzates of jaundiced urineand dialyzates of uremic jaundiced plasma (Ta-ble III). This suggested that the conjugatedbilirubin in these dialyzates was bound to a car-rier of high enough molecular weight so that thecomplex could sediment. The sedimentationgradients for Folin- and Ninhydrin-reactive ma-terial suggested that polypeptides were present inthe dialyzates of both urine and plasma. Thesegradients do not necessarily mean that bilirubinis bound to the sedimenting polypeptides, butthey are compatible with this hypothesis. It wasinferred, therefore, that urinary conjugated bili-rubin and the dialyzable fraction of plasma con-jugated bilirubin are both complexed with a dia-lyzable carrier molecule(s), possibly a large poly-peptide. The precipitation of bilirubin in dialy-zates of urine and uremic plasma by 80%o satu-rated ammonium sulfate also accords with thehypothesis that urinary bilirubin is bound to asmall protein or large polypeptide.
Electrophoretic studies. The electrophoreticstudies of jaundiced plasma and urine permittedfurther characterization of the bilirubin-carriercomplex. There was a bilirubin band in the betaglobulin zone in humans, in addition to the major
677

MILFORD FULOP, JOHN SANDSON,AND PAUL BRAZEAU
BILIRUBIN - PAPERELECTROPHORESIS
BILIRUBIN ADDEDTO NORMALPLASMA
i -lOBSTRUCTIVEJAUNDICE PLASMA
I '. UI1OBSTRUCTIVEJAUNDICE URINE
OBSTRUCTIVEJAUNDICE UREMIC PLASMA
ORIGIN ,8 a Alb
FIG. 12. ELECTROPHORETICPATTERNS OF PLASMA CON-JUGATED AND UNCONJUGATEDBILIRUBIN AND URINARY
BIImuBINz. A diagram showing the relative proportionsof bilirubin migrating in the albumin, alpha and betaglobulin zones. Note the small proportion of betamigrating conjugated bilirubin in nonuremic jaundicedplasma and the increase in beta and alpha migrating bili-rubin in uremic jaundiced plasma, paralleling the urinaryelectrophoretic migration of conjugated bilirubin.
one in the albumin zone, and in dogs in the alphaglobulin zone (sometimes alpha and beta).'
Earlier plasma bilirubin electrophoretic stud-ies concurred that most of the pigment migratedwith albumin. It was indicated in some reportsthat all the bilirubin was albumin-bound (4, 18,21-23), and in others it was suggested thatsmall amounts migrated in the alpha (19, 24-26)or beta globulin zones (26, 27). In most of thosestudies the electrophoreses were made using eithernormal plasma with added bilirubin or abnormalplasma with high levels of unconjugated biliru-bin. The present results accord with the earlierfindings that up to bilirubin levels of about 40 mg
per 100 ml, albumin seems to be the only plasmaprotein that binds unconjugated bilirubin in per-
ceptible amounts.The paper electrophoretic patterns of bilirubin
in jaundiced urine and uremic jaundiced plasmafrom humans and dogs were qualitatively similarand will be considered together. The diagrams inFigure 12 summarize the findings. Urinary bili-rubin migrated mainly in a broad zone of beta and
alpha globulin mobility, although there was nostainable protein on the paper strips. In somespecimens the color was more intense in the betazone (usually in dog urine), and in others in thealpha zone (usually in human urine), but therelative proportions were variable. The zone ofurinary beta bilirubin migration was slightlyfaster than the secondary beta bilirubin band pres-ent in nonuremic jaundiced human plasma. If thedialyzable plasma bilirubin fraction was the sourceof urinary bilirubin, it might be expected that itsincrement in the plasma in subjects with renalfailure could be appreciated electrophoretically inthe beta and alpha zones. The electrophoretic pat-terns of uremic jaundiced plasma in four patientsand two dogs bore out that prediction. This pat-tern was shown to differ from the preuremic pat-tern (Figures 9 and 11). In all of these speci-mens it was shown that at least some of the ex-cess uremic beta and alpha bilirubin could be re-moved by in vitro dialysis (Figure 9C). Theelectrophoretic migration of conjugated bilirubinin concentrated dialyzates of uremic jaundicedplasma was shown to conform with the resultsof the foregoing indirect studies.
These results indicated that the dialyzable frac-tion of plasma conjugated bilirubin was the sourceof urinary conjugated bilirubin and that it wascomplexed with a carrier molecule(s) which ex-hibited electrophoretic mobility on paper in thebeta and alpha globulin zones.
On polyvinyl chloride, as on cellulose acetateand agar gel (28), electrophoresis of jaundicedurine and urine dialyzates exhibited predominantprealbumin migration of conjugated bilirubin.Electrophoretic migration may vary in differentmedia (29), accounting for this difference fromthe pattern found on paper electrophoresis. Onpolyvinyl chloride the migration of plasma dia-lyzable bilirubin and urine bilirubin was con-cordant, both exhibiting prealbumin mobility.
The unavailability of pure bilirubin conjugateshas hampered efforts to elucidate the problem oftheir binding to plasma proteins. The presentstudies indicate the existence of qualitative dif-ferences in protein binding between conjugatedand unconjugated bilirubin. A solution of thequantitative aspects of the problem must awaitother approaches.
678

PROTEIN BINDING OF CONJUGATEDBILIRUBIN
Summary
Conjugated bilirubin was previously shown to
be excreted mainly by glomerular filtration in
jaundiced dogs. The present studies were under-taken to characterize the filtered urinary bilirubinin subjects with obstructive jaundice.
When plasma from subjects with obstructivejaundice was dialyzed in cellophane sacs againstan equal volume of buffer, a small fraction of theplasma conjugated bilirubin was found to be dia-lyzable. This dialyzable portion averaged 0.6%in humans and 1.6%o in dogs with normal renalfunction. In each species this was as large as, or
larger than, the fraction of plasma bilirubin clearedby the kidneys, suggesting that the dialyzableplasma fraction is filterable through the renal glo-meruli and that it is unnecessary to invoke tubu-lar secretion to explain renal excretion of conju-gated bilirubin. The dialyzable plasma bilirubinfraction increased in jaundiced subjects with renalinsufficiency.
Ultracentrifugation of dialyzates of jaundicedurine and jaundiced uremic plasma produced a
gradient of both conjugated bilirubin and poly-peptide. The conjugated bilirubin in these di-alyzates was precipitable with 80%o saturatedammonium sulfate, and these precipitates couldbe redissolved in water. It seems likely that thedialyzable conjugated bilirubin in both fluids iscombined with another- substance, possibly a
polypeptide.Paper electrophoresis of plasma from subjects
with obstructive jaundice showed migration ofbilirubin predominantly with albumin. There was
a minor secondary bilirubin band in the betaglobulin zone in humans and in the alpha, or oc-
casionally beta, globulin zone in dogs. Urinarybilirubin migrated in a broad region comprisingthe beta and alpha globulin zones. In subjectswith obstructive jaundice and renal failure, an
increased proportion of the plasma bilirubin mi-grated in the beta and alpha globulin zones. Bili-rubin appearing in the beta and alpha zones couldbe partially dialyzed from these plasma specimensin vitro. Electrophoresis of concentrated jaun-diced urine and plasma dialyzates prepared byammonium sulfate precipitation showed bilirubinmigration in the beta and alpha globulin zones.
On polyvinyl chloride electrophoresis, there was
predominant prealbumin migration of bilirubin injaundiced urine and in concentrated dialyzates ofuremic jaundiced plasma. These prealbuminzones contained hexuronic acid, and Folin- andNinhydrin-reactive material.
The basis for the difference in renal excretionbetween conjugated and unconjugated bilirubinresides in the dialyzability of a small fraction ofthe plasma conjugated bilirubin and the nondia-lyzability of plasma unconjugated bilirubin. Thisdifference is, in turn, attributable to the bindingof a small fraction of plasma conjugated bilirubinto a nonalbumin plasma constituent of low molecu-lar weight.
AcknowledgmentsWe are grateful to Alma Annexy, Luis Leon, and
Joseph DeVito for their capable technical assistance andto Dr. I. H. Scheinberg for the use of equipment in hislaboratory.
References1. Fulop, M., J. Sandson, and P. Brazeau. The mecha-
nism of excretion of bilirubin by the kidney (ab-stract). J. clin. Invest. 1964, 43, 1244.
2. Fulop, M., J. Sandson, and P. Brazeau. Dialysabilityof conjugated bilirubin from plasma of jaundiceddogs and patients (preliminary communication).Lancet 1964,1, 1017.
3. Hoover, C. F., and M. A. Blankenhorn. Dissociatedjaundice. Arch. intern. Med. 1916, 18, 289.
4. Snapper, I., and W. M. Bendien. On the physico-chemical condition of the bilirubin in the blood-serum and urine. Acta med. scand. 1938, 98, 77.
5. Barac, G. Cenapse albumino-bilirubinique et elimina-tion renale de la bilirubine. Bull. Soc. Chim. biol.(Paris) 1947, 29, 96.
6. Blondheim, S. H., D. Lathrop, and J. Zabriskie.Effect of light on the absorption spectrum ofjaundiced serum. J. Lab. clin. Med. 1962, 60, 31.
7. Gregory, R. L., and M. Andersch. The filtrability ofbilirubin in obstructive jaundice. J. Lab. clin. Med.1937, 22, 1111.
8. With, T. K. Bilirubin in urine and other secretionsapart from the bile and in the cerebrospinal and eyeliquors. Acta physiol. scand. 1945, 10, 355.
9. Fulop, M., and P. Brazeau. The renal excretion ofbilirubin in dogs with obstructive jaundice. J.clin. Invest. 1964, 43, 1192.
10. Weber, A. Ph., and L. Schalm. Quantitative sepa-ration and determination of bilirubin and con-jugated bilirubin in human serum. Clin. chim.Acta 1962, 7, 805.
11. Malloy, H. T., and K. A. Evelyn. The determina-tion of bilirubin with the photoelectric colorimeter.J. biol. Chem. 1937, 119, 481.
679

MILFORD FULOP, JOHN SANDSON,AND PAUL BRAZEAU
12. Moore, S., and W. H. Stein. Photometric Ninhydrinmethod for use in the chromatography of aminoacids. J. biol. Chem. 1948, 176, 367.
13. Dische, Z. A new specific color reaction of hexuronicacids. J. biol. Chem. 1947, 167, 189.
14. Toribara, T. Y., A. R. Terepka, and P. A. Dewey.The ultrafiltrable calcium of human serum. I.Ultrafiltration methods and normal values. J.clin. Invest. 1957, 36, 738.
15. Grabar, P., and C. A. Williams, Jr. Methode im-muno-electrophoretique d'analyse de melanges desubstances antigeniques. Biochim. biophys. Acta(Amst.) 1955, 17, 67.
16. Ingbar, S. H., and N. Freinkel. Regulation of theperipheral metabolism of the thyroid hormones.Recent Progr. Hormone Res. 1960, 16, p. 353.
17. Odell, G. B. Studies in kernicterus. I. The proteinbinding of bilirubin. J. clin. Invest. 1959, 38, 823.
18. Ostrow, J. D., and R. Schmid. The protein-bindingof C1'-bilirubin in human and murine serum. J.clin. Invest. 1963, 42, 1286.
19. Watson, D. The transport of bile pigments: the bind-ing of sodium bilirubinate to plasma protein. Clin.Sci. 1962, 22, 435.
20. Wallace, D. K., and E. E. Owen. An evaluation ofthe mechanism of bilirubin excretion by the humankidney. J. Lab. clin. Med. 1964, 64, 741.
21. Coolidge, T. B. Chemistry of the van den Berghreaction. J. biol. Chem. 1940, 132, 119.
22. Barac, G. Interactions of bilirubin with plasma pro-teins, and the state of bilirubin in normal humanplasma. Arch. int. Physiol. 1953, 61, 129.
23. Bourrillon, R. Isolement et proprietes du complexebilirubinserumalbumine du serum. Bull. Soc. Chim.biol. (Paris) 1957, 39, 385.
24. Gray, C. H., and R. A. Kekwick. Bilirubin-serumprotein complexes and the van den Bergh reaction.Nature (Lond.) 1948, 161, 274.
25. Martin, N. H. Preparation and properties of serumand plasma proteins. XXI. Interactions with bili-rubin. J. Amer. chem. Soc. 1949, 71, 1230.
26. Klatskin, G., and L. Bungards. Bilirubin-proteinlinkages in serum and their relationship to the vanden Bergh reaction. J. clin. Invest. 1956, 35, 537.
27. Westphal, U., H. Ott, and P. Gedigk. An welcheKomponenten der Serumproteine ist das Bilirubingebunden? Hoppe-Seylers Z. physiol. Chem. 1950,285, 200.
28. Sandson, J., and M. Fulop. Unpublished observa-tions.
29. Block, R. J., E. L. Durrum, and G. Zweig. A Manualof Paper Chromatography and Paper Electro-phoresis, 2nd ed. New York, Academic Press,1958, p. 501.
680





























