Embed Size (px)
Citation preview


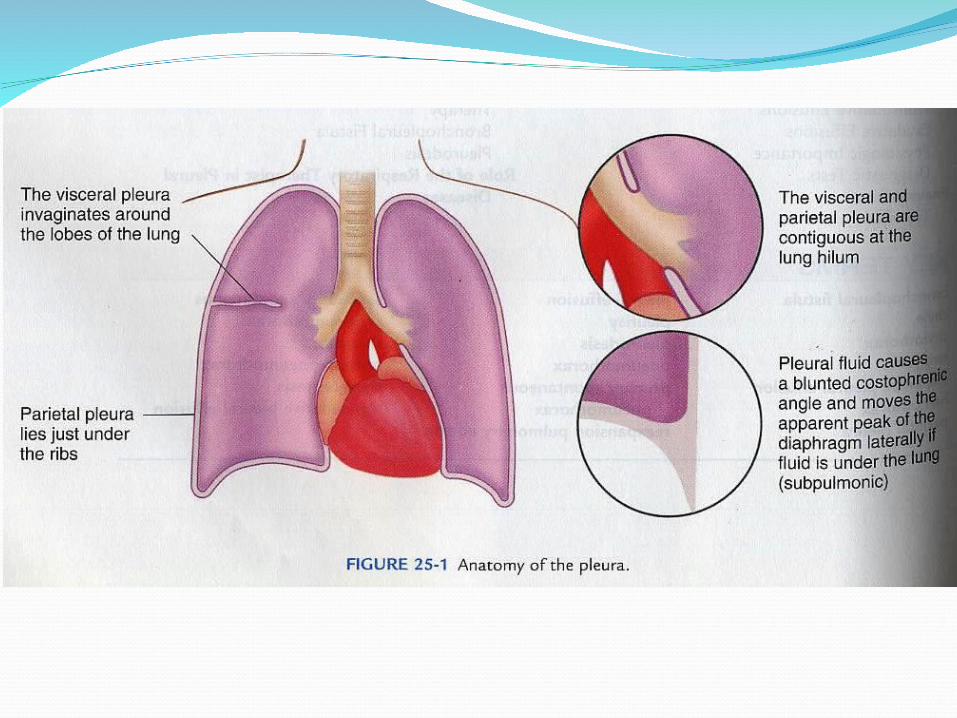

INTRODUCTIONThe pleural space is bounded by two
membranes, the visceral pleura covering
the lung and the parietal pleura covering
the chest wall and diaphragm. Into this
space, normal liquid and protein enter from
the systemic circulation and are removed
by the parietal pleural lymphatics.

Pleural pressure is subatmospheric and
ensures inflation of the lung. Because the
mesothelial boundaries are leaky, excess
liquid can move across into this lower-
pressure, high-capacitance space and collect
as a pleural effusion. Thus, pleural effusions
are common and of highly diverse etiologies.
These effusions can form based on disease
of the pleural membranes themselves or of
thoracic or abdominal organs.

Depending on the protein and lactate
dehydrogenase (LDH) concentrations of the
liquid and on the cellular composition, these
effusions can be categorized initially as
exudates or transudates. Exudative pleural
effusions meet at least one of the following
criteria, whereas transudative effusions meet
none (Light's criteria): pleural fluid protein–to–
serum protein ratio of more than 0.5, pleural
fluid LDH–to–serum LDH ratio of more than 0.6,
and pleural fluid LDH more than two thirds of
the upper normal limit for serum.

These two pleural membranes meet at the hilar
root of the lung. In the sheep, an animal with a
pleural anatomy similar to that of humans, the
surface area of the visceral pleura of one lung,
including that invaginating into the lung fissures,
is similar to that of the parietal pleura of one
hemithorax, approximately 1000 cm2.[1] The
normal pleural space is approximately 18 to 20 ým
in width, although it widens at its most dependent
areas. It has been shown that the pleural
membranes do not touch each other and that the
pleural space is a real, not a potential, space.
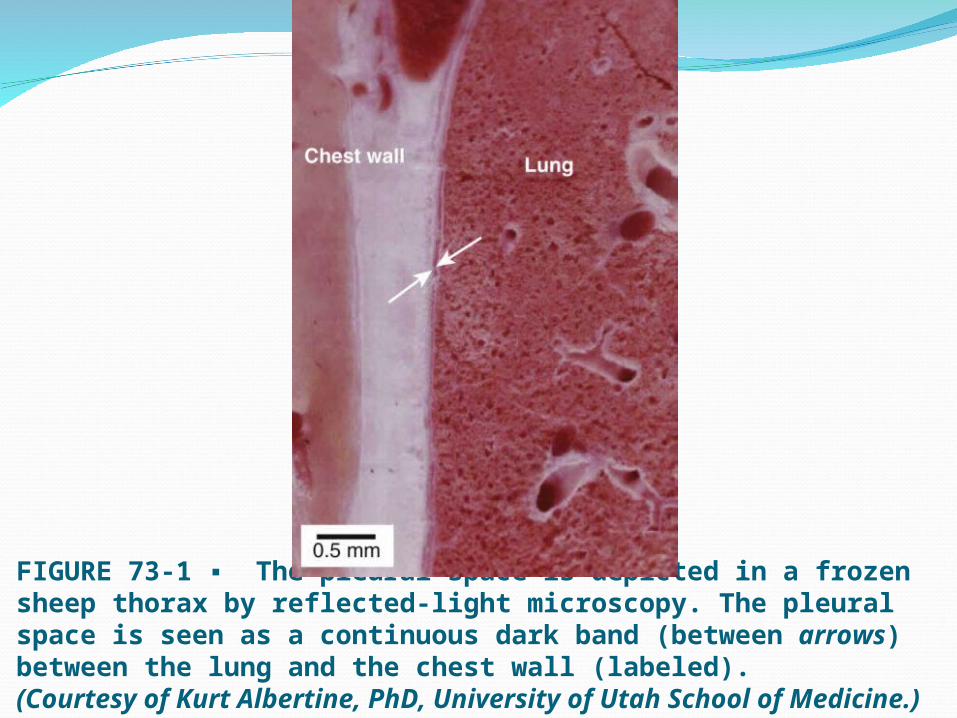
FIGURE 73-1 ▪ The pleural space is depicted in a frozen sheep thorax by reflected-light microscopy. The pleural space is seen as a continuous dark band (between arrows) between the lung and the chest wall (labeled).(Courtesy of Kurt Albertine, PhD, University of Utah School of Medicine.)

It is likely that the primary function of
the pleural membranes is to allow
extensive movement of the lung relative
to the chest wall. If the lung adhered
directly to the chest wall, its expansion
and deflation would be more limited.

The visceral pleura may also provide mechanical
support for the lung: contributing to the shape of
the lung, providing a limit to expansion, and
contributing to the work of deflation. Because the
submesothelial connective tissue is continuous with
the connective tissue of the lung parenchyma, the
visceral pleura may help to distribute the forces
produced by negative inflation pressures evenly over
the lung. In this way, overdistention of alveoli at the
pleural surface may be avoided, lessening the
chance of rupture and pneumothorax.

One more recently recognized function of the
pleural space is to provide a route by which edema
can escape the lung.[5] As has been shown in
several experimental studies of either hydrostatic
or increased permeability lung edema,[6],[7] the
pleural space can function as an additional safety
factor protecting against the development of
alveolar edema. The formation of transudative
effusions in patients with congestive heart failure
(CHF) apparently reflects the movement of edema
from the lung to a space where its effects on lung
function are relatively small.

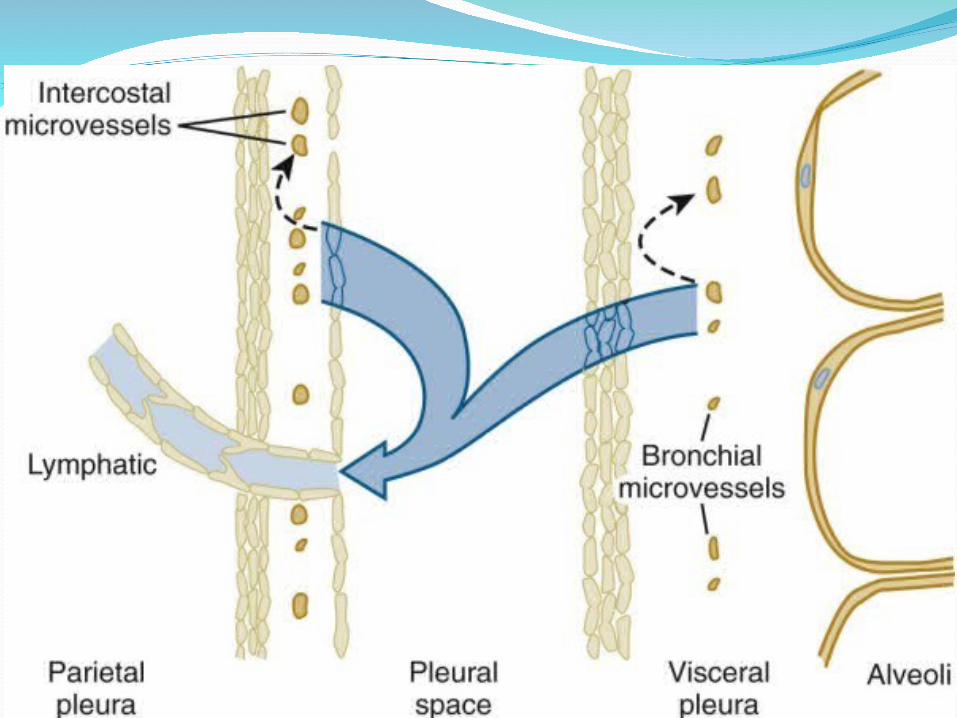
Blood SupplyThe parietal pleura is supplied by intercostal
arteries. The visceral pleura in humans, like that in sheep, is exclusively supplied by the bronchial circulation, which drains into pulmonary veins. The drainage route via pulmonary veins may have contributed to earlier confusion about whether the visceral pleural blood supply was from a systemic (bronchial) or a pulmonary circulation. Both pleurae in humans, therefore, have a systemic circulation, although the visceral pleural bronchial circulation may have a slightly lower perfusion pressure than the parietal pleural intercostal circulation because of its drainage into a lower-pressure venous system.
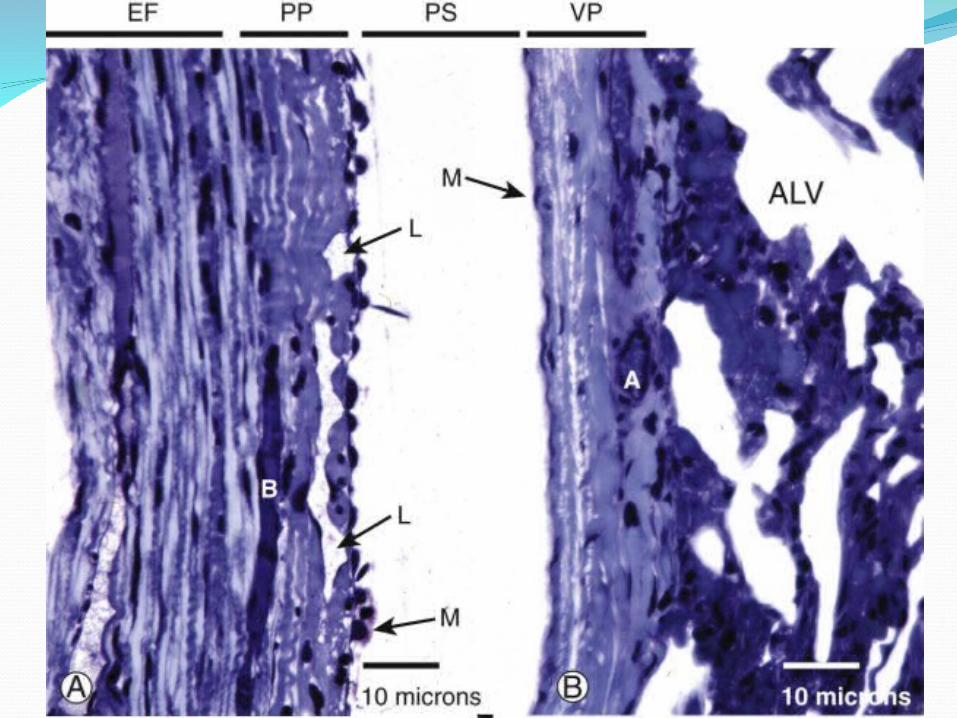

LymphaticsIf one injects carbon particles into the pleural
space as a visible marker of lymphatic drainage pathways, one later finds that the black carbon has been taken up into lymphatics on the parietal side, not the visceral side . The visceral pleura has extensive lymphatics, but they do not connect to the pleural space. Demonstrated in rabbits, sheep, and now in humans, the parietal pleural lymphatics connect to the pleural space via stomata, holes of 8 to 10 ým in diameter that are formed by discontinuities in the mesothelial layer where mesothelium joins to the underlying lymphatic endothelium.
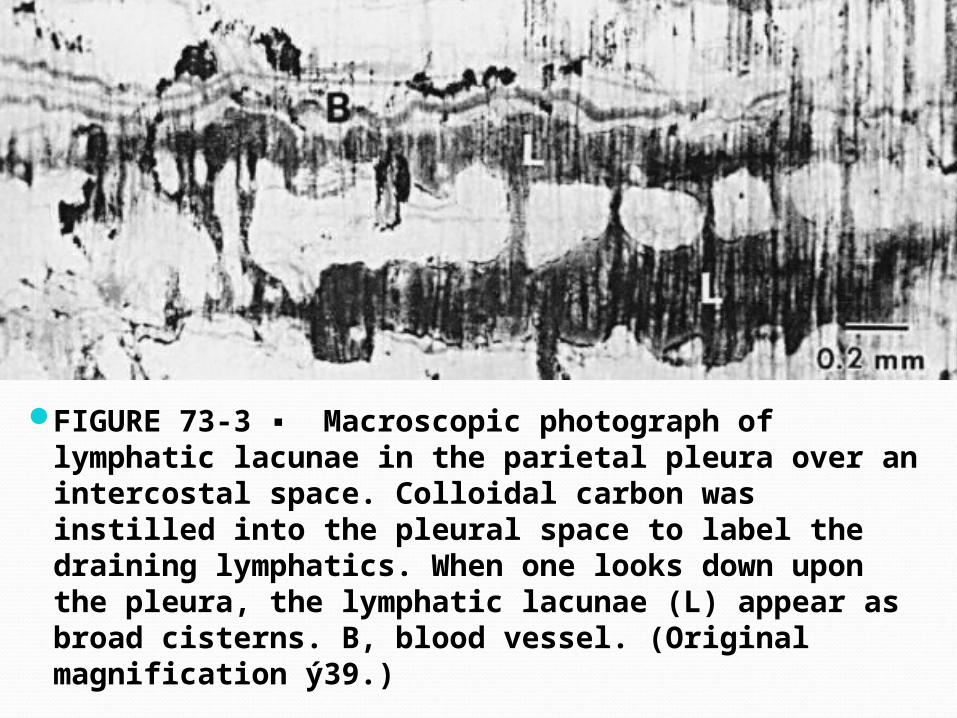
FIGURE 73-3 ▪ Macroscopic photograph of lymphatic lacunae in the parietal pleura over an intercostal space. Colloidal carbon was instilled into the pleural space to label the draining lymphatics. When one looks down upon the pleura, the lymphatic lacunae (L) appear as broad cisterns. B, blood vessel. (Original magnification ý39.)

Nerve SupplyThe parietal pleura contains sensory nerve
fibers, supplied by the intercostal and phrenic nerves, and has long been thought to be the major site of pain sensation in the pleura. The costal and peripheral diaphragmatic regions are innervated by the intercostal nerves, and pain from these regions is referred to the adjacent chest wall. The central diaphragmatic region is innervated by the phrenic nerve, and pain from this region is referred to the ipsilateral shoulder. The visceral pleura has more recently been shown to have sensory nerve fibers that may participate in pain or other sensations such as dyspnea.[19] In addition, pleural adhesions may contain pain fibers and contribute to post-thoracotomy or postpleurodesis pain.[20]

Normal Pleural Liquid and Protein Turnover
Since the 1980s, a consensus has developed
that normal pleural liquid arises from the
systemic pleural vessels in both pleurae,
flows across the leaky pleural membranes
into the pleural space, and exits the pleural
space via the parietal pleural lymphatics. In
this way, the pleural space is analogous to
other interstitial spaces of the body. There
are several lines of evidence for this view.

1. Intrapleural pressure is lower than
the interstitial pressure of either of the
pleural tissues. This pressure difference
constitutes a gradient for liquid
movement into but not out of the pleural
space.

2. The pleural membranes are leaky to
liquid and protein. Whether tested in
vitro[23],[24] or in situ,[25] the pleura offers
little resistance to liquid or protein
movement

3. Mesothelial cells express various
transporters and aquaporins but these have
not been shown to have a role in reabsorption
of effusions.[26] Although normal pleural liquid
has been reported to be alkaline with higher
bicarbonate than plasma, there is no evidence
for active mesothelial participation in
generating a bicarbonate gradient, which can
more likely be explained by a passive response
to protein gradients (Donnan equlibrium).[26a]

4. The entry of pleural liquid is slow and
compatible with known interstitial flow
rates. By noninvasive studies of the
equilibration of radiolabeled albumin, the
entry rate of pleural liquid is approximately
0.01 mL/kg/hr in a sheep, or about
0.5 mL/hr or 12 mL/day in an adult human.
[27] The halftime of pleural liquid turnover
in sheep and rabbits is 6 to 8 hours.[27],[28]

5. The protein concentration of normal pleural
liquid is low in sheep[27] and probably in humans,
which implies sieving of the protein across a high-
pressure gradient. The protein concentration of
sheep pleural liquid (10 g/L) and pleural–to–
plasma protein concentration ratio (0.15) are
similar to those of filtrates from high-pressure
systemic vessels. By comparison, a filtrate from
low-pressure pulmonary vessels has a higher
protein concentration (45 g/L) and ratio (lymph–
to–plasma protein concentration ratio 0.69).[29

6. The majority of liquid exits the pleural
space by bulk flow, not by diffusion or active
transport. This is evident because the protein
concentration of pleural effusions remains
constant as the effusion is absorbed, as is
expected with bulk flow. If liquid were absorbed
by diffusion or active transport, proteins would
diffuse at a slower rate, and the protein
concentration would progressively increase. In
addition, erythrocytes instilled into the pleural
space are absorbed intact and in almost the
same proportion as the liquid and protein.[18

] This indicates that the major route of exit is
via holes large enough to accommodate sheep
erythrocytes (6–8 ým diameter). The only
possible exit is via the parietal pleural stomata
(10–12 ým diameter) into the pleural
lymphatics. Of note, these lymphatics have a
large capacity for absorption. When artificial
effusions were instilled into the pleural space
of awake sheep, the exit rate (0.28 mL/kg/hr)
was nearly 30 times the baseline exit rate
(0.01 mL/kg/hr).[18]

Pleural PressureThe pleural pressure in humans is approximately
−5 cm H2O at midchest at functional residual
capacity and −30 cm H2O at total lung capacity.[30]
If the compliance of the lung decreased, pleural
pressures at the same lung volumes would be more
negative. In one study of patients undergoing
thoracentesis, those with more negative pleural
pressures had a smaller improvement in lung
volume than those with less negative pressures,
presumably reflecting the presence of underlying
diseased, noncompliant lung.[31]

Although the pleural space pressure is
subatmospheric, gases do not accumulate
there. The sum of all partial pressures of
gases in capillary blood is approximately
700 mm Hg, or 60 mm Hg below
atmospheric (water pressure [PH2O] = 47,
carbon dioxide pressure [PCO2] = 46,
partial pressure of nitrogen [PN2] = 570,
and oxygen pressure [PO2] = 40 mm Hg).

The subatmospheric pressure of
dissolved gases in capillary blood helps
to maintain the pleural space free of gas
and facilitates absorption of any gas that
does enter the pleural space. Of note, to
increase the gradient favoring absorption
of gas, one can lower the PN2 in the
blood by having a patient breathe
increased concentrations of inspired
oxygen.

The oxygen displaces alveolar nitrogen,
thereby lowering the PN2 in capillary
blood; because of the limited absorption
of oxygen due to the plateau of the
oxygen-hemoglobin dissociation curve,
the increase in inspired oxygen does not
add greatly to the PO2 in capillary blood.

PATHOPHYSIOLOGY OF THE PLEURAL SPACE
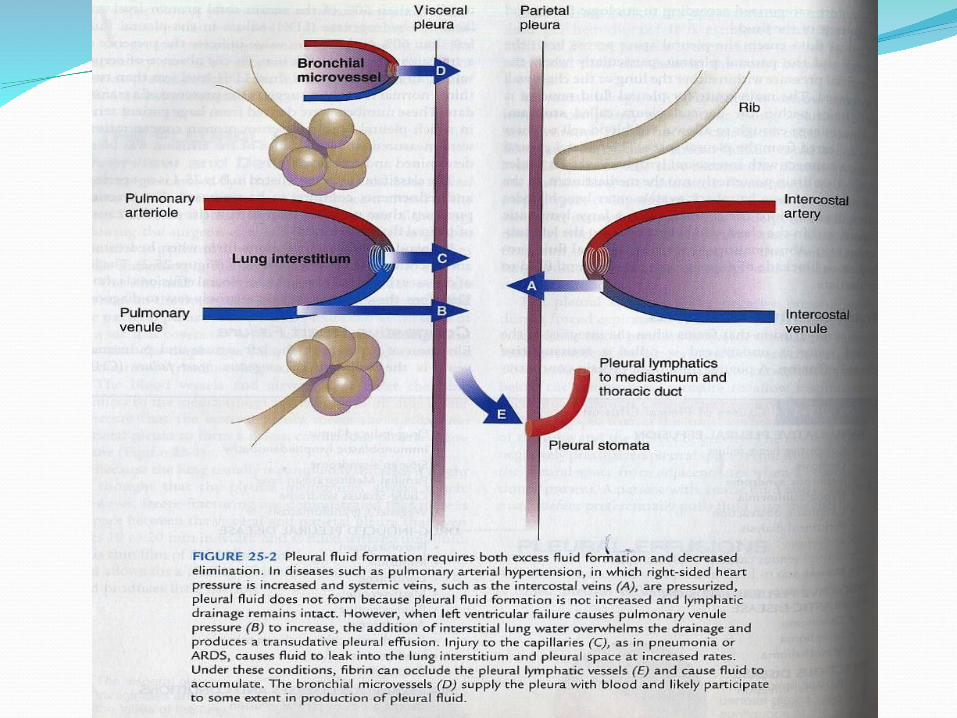
Pleural Effusions For pleural liquid to accumulate to form an
effusion, it is likely that both the entry rate of liquid must increase and the exit rate must decrease. If only the entry rate increased, it would require a sustained rate more than 30 times normal to exceed the reserve lymphatic removal capacity; if the exit rate decreased, it would take more than a month at the normal entry rate of 12 mL/day to produce an effusion detectable by chest radiograph.[22] Thus, in the clinical setting, it is most likely that excess pleural liquid accumulates due to changes in both entry and exit rates.

Increased entry of liquid may result from increased filtration across systemic or pulmonary capillaries or entry of another liqud (e.g., chyle, cerebrospinal fluid [CSF], urine, intravenous fluids). Decreased exit of liquid may result from interference with lymphatic function (e.g., obstruction of the parietal pleural stomata, inhibition of lymphatic contractility, infiltration of draining parasternal lymph nodes, or elevation of the systemic venous pressure into which the lymph drains).[22] There are few studies on the rate of removal of liquid in humans; however, decreases in lymphatic clearance have been confirmed in patients with tuberculous and malignant effusions[32] and in those with the yellow nail syndrome, a disease of lymphatic function.[33]

To determine the origin of effusions, a classic and useful distinction is between transudates and exudates (see later, “Separation of Exudates from Transudates”).[34] Transudates form by leakage of liquid across an intact capillary barrier owing to increases in hydrostatic pressures or decreases in osmotic pressures across that barrier. Transudates generally indicate that the pleural membranes are not themselves diseased. Exudates form from leakage of liquid and protein across an altered capillary barrier with increased permeability. The protein ratio, the LDH ratio, and the absolute pleural LDH concentration constitute Light's criteria.

Transudates include various low-protein liquids
that arise from noninjured capillary beds. The
majority of transudates are caused by CHF.
These transudates have been shown to form from
leakage of edema across normal pulmonary
capillaries into the pulmonary interstitium and
then across the leaky visceral pleura into the
pleural space.[6],[35] Other transudates, those
associated with the nephrotic syndrome or
atelectasis, may form because of altered
pressures (osmotic or hydrostatic) across the
pleural capillaries.

Some transudates, usually small, may develop
primarily because of an isolated decrease in
exit rate.[36] Hepatic hydrothorax and effusions
from peritoneal dialysis develop when liquid
flows from the peritoneal space into the lower
pressure pleural space across macroscopic
holes in the diaphragm. Finally, other very low
protein fluids such as urine or CSF or
intravenous liquids may find their way to the
pleural space if their normal course is
disrupted.

Exudates arise from injured capillary beds, in either the lung, the pleura, or adjacent tissues. Most exudates, such as those associated with pneumonia or pulmonary embolism, probably form following lung inflammation and injury when a high-protein lung edema leaks into the pleural space. Another large category of exudates arises from pleural injury due to inflammation, infection, or malignancy. Exudates can also form when exudative liquid in the mediastinum (esophageal rupture or chylothorax), retroperitoneum (pancreatic pseudocyst), or peritoneum (ascites with spontaneous bacterial peritonitis or Meigs’ syndrome) finds its way into the lower-pressure pleural space.

As stated, for either transudates or
exudates, lymphatic obstruction may
contribute to the accumulation of the
effusion. Nonetheless, because lymphatic
clearance does not alter the pleural fluid
protein concentration, the protein
concentration gives information about
the formation of the fluid, not its
removal.[22]

Effects of Pleural Effusions on Lung and Cardiac Function
In the presence of a space-occupying liquid in the pleural space, the lung recoils inward, the chest wall recoils outward, and the diaphragm is depressed inferiorly and is sometimes inverted.[37] If the lung and chest wall have normal compliances, the decrease in lung volume accounts for approximately a third of the volume of the pleural effusion, and the increase in the size of the hemithorax accounts for the remaining two thirds. As a result, lung volumes are reduced by less than the pleural effusion volume. If the lung is otherwise normal, there is no evidence that an effusion causes significant hypoxemia, presumably because ventilation and perfusion decrease similarly. In fact, in one study, hypoxemia was noted only after liquid was removed by thoracentesis,[38] when perfusion presumably was restored while ventilation remained inadequate.

In another study using multiple inert gas
techniques to quantify ventilation-perfusion
(V/Q) distributions, pleural effusion was
associated with a small intrapulmonary
perfusion shunt (6.9%) that did not change
significantly when measured again 30 minutes
after thoracentesis of approximately 700 mL
(6.1%).[39] It appears, therefore, that the effects
of pleural effusion and thoracentesis on
oxygenation are variable and may depend on
the underlying lung function.

Large pleural effusions may impair cardiac function, most likely by decreasing the distending pressures on the cardiac chambers and cardiac filling. In a study of 27 patients with large effusions occupying more than half the hemithorax, clinical and echocardiographic findings of cardiac tamponade were identified in most patients. These findings, including elevated jugular venous pressure, pulsus paradoxus, right ventricular diastolic collapse, or flow velocity paradoxus, resolved in all patients when studied again 24 hours after thoracentesis of more than 1.0 L.[40] Large pleural effusions, especially left-sided effusions, should be considered as potentially reversible causes of cardiac dysfunction.[41]

Common symptoms of patients with effusions
are pleuritic chest pain, cough, and dyspnea. It
appears that the three symptoms are due to
different causes. Pleuritic chest pain derives
from inflammation of the parietal pleura and
possibly the visceral pleura. Occasionally, this
symptom is accompanied by an audible or
palpable pleural rub, reflecting the movement
of abnormal pleural tissues. Cough may be due
to distortion of the lung, in the same way as
cough follows lung collapse from a
pneumothorax.

Dyspnea is most likely caused by the mechanical
inefficiency of the respiratory muscles that are
stretched by the outward displacement of the chest
wall and the downward displacement of the
diaphragm.[37] After the removal of large amounts of
pleural liquid, dyspnea is generally relieved
promptly, although the reduction in pleural liquid
volume is associated with only small increases in
lung volume and little improvement, or an actual
decrease, in PO2. In one study, nine patients
underwent removal of over 1800 mL of pleural
liquid, and despite increases in vital capacity of
only 300 mL, all patients experienced immediate
relief of dyspnea.[42]

Although the vital capacity changed little,
patients could generate a more negative
pleural pressure at the same lung volume
after thoracentesis than before, indicating an
improved efficiency of the respiratory muscles
following the return of the chest wall and
diaphragm to a more normal position after
thoracentesis. This improved efficiency of
respiration may explain the relief of dyspnea
after removal of pleural liquid.


Thank Thank youyou