Embed Size (px)
Citation preview

Published by Durrell Conservation Academy

Solitaire No. 23 2
Welcome to Durrell Conservation Academy’s new-look Solitaire. In this edition we have begun to reshape the publication to achieve, ultimately, a number of objectives:
To produce a publication that challenges the way •we think about our conservation profession and how we can improve its effectivenessTo use • Solitaire as an outlet for scientifically robust research material as well as opinion pieces based on years of experience within the field To facilitate increased exchange of ideas and foster •partnerships between Durrell staff, Durrell associates and our Graduate Network.
There are still further changes to be made but we are keen to present you with our first step along this path and to ask you for your opinion – are we moving the publication in the right direction? What else would you recommend to help make the publication evolve into a ‘conservation professional magazine’, designed to challenge your thinking, provide diverse conservation-related short research articles and help build connections between Durrell staff, associates and the Graduate Network?
In this edition our own Chief Conservation Officer, Professor John Fa, kicks off with an opinion piece which sets out to challenge the role of zoos and encourage them to reflect on how they can make a genuine contribution to global conservation efforts. Another new feature sees the inclusion of several short research articles designed to test how best to manage particular primate species in captivity. From our Graduates we find out about securing watersheds for people and wildlife in Latin America, understanding the potential and limitations of habitat corridors as conservation tools in Sumatra and find out more about the evolution of zoo philosophy in India. In this issue we update you on courses we’ll be running in 2013 and funding opportunities to help kick-start your own conservation projects.
On a personal note it is with great excitement that I will be moving myself and my family to Mauritius to begin a three-year programme to establish Durrell’s first satellite training institution in Mauritius – Durrell
Conservation Academy: Indian Ocean Campus. We will begin in 2013 with a number of courses designed to draw on Durrell and Mauritian Wildlife Foundation projects that are ongoing in Mauritius, to provide first-hand experience for budding conservation leaders from around the world. In time we hope to begin building our own training campus which will run courses for an international group of students, with a particular focus on Africa, South-East Asia, China, Australia and New Zealand. I will maintain oversight of Durrell Conservation Academy in Jersey, but I am pleased to announce we will be leaving the day-to-day running of the academy in the capable hands of Dr Tim Wright, currently Deputy Head of the academy. Catherine Burrows will continue her invaluable role as Training Co-ordinator and Dr Eluned Price (editor of Solitaire!) will be focusing her time on developing on-site research capacity, which participants on our training courses will benefit from during their time with us in Jersey.
2012 has been an exciting year for the academy, with more than 450 individuals passing through our courses this year, our name change to Durrell Conservation Academy and the development of plans to establish ourselves in the Indian Ocean. 2013 will be an even more exciting year with our first Graduate Network meeting to be held in Colombia in February, co-hosting an international conference on capacity-building in Colombia in the same month, and a whole suite of training courses, internship opportunities and seed funding provision that will be up and running during the year. If you haven’t already done so please sign up to the Durrell Graduate Facebook page (http://www.facebook.com/#!/groups/199959050121085/) to help build the network and stay in touch.
From all of us here at Durrell Conservation Academy I would like to wish you a Happy New Year and a fruitful and exciting 2013!
Jamie CopseyHead of Learning and Development
Contents
2 Welcome to Solitaire
3 Corporate chimeras and zoos for conservation
6 Reciprocal agreements for watershed services in Latin America
7 Illegal wildlife trade threatens Asian bears
8 Enriched environments for callitrichids
13 The training and management of an insulin- dependent Sulawesi crested black macaque
16 The effect of a GnRH agonist on an aggressive adolescent male Macaca nigra: a case study
20 Dead end or thoroughfare? Habitat corridors and endangered mammals in Sumatra
22 Zoos in India: from menageries to naturalistic enclosures
26 Durrell Conservation Academy news
28 Congratulations and contact details

3 Solitaire No. 23
Corporate chimeras and zoos for conservation
. Zoos are chimeras*! They, like the mythological beasts, are metaphorically composed of the parts of three institution types: a business, a conservation body and a school. Zoos have to manage wild animals in captivity for exhibition pur-poses and to make money, they have to educate the large numbers of people that come to enjoy the spectacle of wild animals, but are also obliged (and often by law in some coun-tries) to contribute to safeguarding biodiversity. The latter is often achieved by inter-organisational endangered species breeding schemes and/or in-situ conser-vation programmes. However, unlike the chimeras of ancient Greece, zoos have a formidable challenge in balancing out the leanings of each of its three animal (insti-tutional!) body parts.
Zoos, like many other institutions involved in conservation, are shaken by non-science aspects, which reflect the pressure of multiple factors (e.g. societal values or even internal corporate biases). Possibly the strongest conditioner affecting zoos is related to anthropomorphic factors, such as the preference for the protection of more ‘charismatic’ taxa. Zoos are also affected by anthropocentric factors, for example the imperative of making money over funding conservation activities by the institution (Restani & Marzluff, 2002). As a result, zoos focus most exhibit, education and conservation attention on a select number of taxa, often the larger-bodied species (Pritchard et al., 2012). Such biases towards the ‘charismatics’ are not unusual in other conservation-related fields such as species conservation research. There is evidence from various countries that conservation research suffers from taxonomic bias, subjectivity in the choice of research topics, and even how resources are allocated (Clark & May, 2002). For example, conservation spending patterns and policy decisions are biased towards certain taxa, often revealing a strong connection between existing conservation policies and scientific information, research priorities and public concern (Martín-López et al., 2009). This effect is often brought about because the more threatened a taxon (at least for certain vertebrate groups), the more likely funds are to be channelled into research and conservation for that taxon. Garnett et al. (2003) presented data to suggest that the direct consequence of a species reaching critically endangered or endangered status is a need for a substantial increase in the funding levels for research on that species. Spending large amounts of money on
a particular priority species, however, is not the most efficient way to promote biodiversity conservation, because some of the most highly ranked species require huge conservation efforts with a small chance of success, whereas other, less threatened species might be successfully conserved for relatively little cost (Possingham et al., 2002).
There is clear evidence that zoos, in general, exhibit animal species that are not of conservation concern (Conde et al., 2011), and are often large-bodied taxa (Pritchard et al., 2011),
which are often more expensive to keep (Balmford et al., 1996; Fa, 2012). These taxonomic biases in zoos occur because species are often selected for exhibition (to which some conservation purpose may be attached later) based
on personal interests (not just of the zoo director, but also of the institution). In some cases, species selection may or may
not reflect certain actual or supposed values (Proença et al., 2008). Most in-
situ conservation work on endangered species undertaken by zoos, e.g. for WAZA projects, focuses on mammals, particularly on charismatic primates and carnivores, but other animal groups (e.g. amphibians and fishes) are significantly under-represented (Gusset & Dick, 2010). This is also the case for AZA and EAZA captive breeding programmes, for
John E. Fa
Durrell Wildlife Conservation Trust
* A chimera is an imaginary monster composed of incongruous parts.
‘There is clear evidence
that zoos, in general,
exhibit animal species
that are not of
conservation concern.’
Zoos often focus attention on large-bodied, charismatic species such as great apes.
Do
min
ic W
orm
ell
Guest essay

Solitaire No. 23 4
in-situ projects supported by European and North American zoos (EAZA, 2010; AZA, 2010), and also for Mexican zoos (Lascuráin et al., 2009).
A zoo’s organisational structure and management is directly relevant to achieving its conservation aims. In particular, it is not unusual within the zoo world in many parts of the world to encounter bureaucratic structures, sometimes highly specialised, with a strict hierarchy of authority that maintains impersonal, rigid rule systems. These structures are effective only when their tasks and their operating environments are simple and stable, but are totally ineffective in more complicated systems (Morgan, 1986; Blau & Myer, 1987). These highly centralised systems of control and their fixed rules, roles and regulations make them slow and ineffective in responding to changing circumstances. Flattening structures may be the way forward, but only as long as there is communication across. After all, it is no use having a horizontal structure that suffers the same ailments as a vertical one.
The unique challenges of conserving endangered species or shaping public understanding exceed current bureaucratic capabilities (Paehlke & Torgerson, 1990). Moreover, the influence of corporate management ethos, which permeates both public and private zoos in many parts of the world, has led to expanded senior management strata, heavily represented by business expertise (Mazur & Clark, 2001). This is always accompanied by the use of corporate tools (business strategies, product formats, and performance measures) to promote a paradigm of economic rationalism. Corporate management, and the economic rationalism it embodies, has been soundly criticised since the late 1980s by Considine (1988), Painter (1988) and Sinclair (1989), and more recently by Rees (1994) and Rhodes (1996). The main criticism is that corporate management narrowly construes economic efficiency as organisational effectiveness. Corporatisation has serious ramifications for all zoo policy. For example, animals may be managed as commodities and conservation may become a public relations ploy, all justified in terms of economic efficiency. Moreover, the mission of endangered species restoration and public education may easily fall prey to cost-benefit accounting (Adams, 1996). Consequently, animal welfare, wildlife conservation and public education may be overshadowed by a standard of economic efficiency common in corporate boardrooms.
Many zoos’ current market-oriented approaches and corporatised organisational practices result in an emphasis on maximising and stabilising revenues (Mazur, 1997). Official pressures and budget constraints balance conservation against corporate efficiency and other demands. Hence, conservation values translate into activities ‘not that the zoo must do – but that it must be able to afford to do’ (Mazur & Clark, 2001). Special tensions are created when conservation programmes and animals’ biological needs are balanced against zoos’ financial and public relations imperatives. Since zoos consider visitors’ recreational motives to be vital to their mission and survival (Chiszar et al., 1990; Bostock, 1993; Maple, 1995), it remains unclear what trade-offs are reasonable. Mazur & Clark (2001) suggest that these management structures are also promoted internally and externally, often by forceful
public relations and/or mechanisms that draw on ‘smoke and mirrors’ to promote something that is unreal. In other words, effectivenessis often measured by ‘how much’ is promoted as having been done (with phrases like ‘our zoo is here for conservation’) rather than measuring success and performance in more transparent and accountable ways.
So how do we move to zoos to become beacons of conservation? Perhaps the first barrier to overcome is to get zoos to integrate zoo conservation thinking within modern environmental problem solving. Some attempts have been made by the zoo community to give directions for zoological institutions to engage in conservation, but what may be lacking are clearer targets. Fa (2012) has suggested that zoos should focus on the more survival-compromised species, such as those listed in the Alliance for Zero Extinction (AZE, 2012). Perhaps it is easier (or economically safer) for zoos to argue that people visit to see the charismatic fauna in their collections (either because they are large and fierce as in the case of tigers and lions, or just show-off, send-ups of people as the
now ever-present meerkats) and that is what they should provide. The question is, are zoos unable to think ‘outside the box’ and
strive to change the way the public engage with any animal collection, or are there
unmovable visitor preferences to which the zoo has to conform to survive? Whatever
the case – and it should be resolved either way – the undeniable reality is that zoos
can be effective commercial enterprises. A possible option is for zoos to determine how
much emphasis (and funds) they are willing to commit to conservation. Once they do this they can then decide whether they merely exhibit animals for education and recreation (and make money), or really get involved in direct conservation. This could mean that the animal collections of ‘direct conservation’ zoos will look different from those of ‘indirect conservation’ zoos, e.g. they would develop coherent messages by exhibiting species that can change societal perceptions, and more importantly encourage adequate funding for those species needing conservation attention. But all zoos that want to achieve transparent and measureable conservation outcomes (ultimately meaning many more species conserved) will need to work together more effectively. If, like Durrell, other zoos are willing to invest in species conservation worldwide, educate the public and promote new conservation talent, the chimera that zoos are would be less an anomaly and more the norm.
ReferencesAdams, J (1996). Cost-benefit analysis: the problem, not the
solution. The Ecologist 26, 2–4.Alliance for Zero Extinction (2012). Alliance for Zero Extinction.
Available from: http://www.zeroextinction.org/AZA (2010). Association of Zoos and Aquariums. Animal
Programs. Available from: http://www.aza.org/animal-programs. Accessed 4 May 2010.
Balmford, A, Mace, GM & Leader-Williams, N (1996). Designing the ark: setting priorities for captive breeding. Conservation Biology 10, 719–727.
Blau, PM & Myer, MW (1987). Bureaucracy in Modern Society. New York: Random House.
Bostock, SC (1993). Zoos and Animal Rights: The Ethics of
‘Zoos need to integrate
conservation thinking
within modern
environmental problem-
solving.’
Guest essay

5 Solitaire No. 23
Keeping Animals. New York: Routledge.Chiszar, D, Murphy, JB & Iliff, W (1990). For zoos. The
Psychological Record 40, 3–13.Clark, JA & May, RM (2002). Taxonomic bias in conservation
research. Science 5579, 191–192.Conde, DA, Flesness, N, Colchero, F, Jones, OR & Scheuerlein,
A (2011). An emerging role of zoos to conserve biodiversity. Science 331: 1390-1391
Considine, M (1988). The corporate management framework as administrative science: a critique. Australian Journal of Public Administration 47, 4–18.
EAZA (2010). European Association of Zoos and Aquaria. EAZA collection planning. Available from: http://www.eaza.net/activities/cp/Pages/Collection%20Planning.aspx/
Fa, JE (2012). How to save more species: ‘zero tolerance’ conservation for zoos. WAZA News 2/2012, 2-4.
Garnett, S, Crowley, G & Balmford, A (2003). The costs and effectiveness of funding the conservation of Australian threatened birds. BioScience 7, 658–665.
Gusset, M & Dick, G (2010). ‘Building a Future for Wildlife’? Evaluating the contribution of the world zoo and aquarium community to in situ conservation. International Zoo Yearbook 44, 183–191.
Lascuráin, M, List, R, Barraza, L, Díaz Pardo, E, Gual Sill, F, Maunder, M, Dorantes, J & Luna, VE (2009). Conservación de especies ex situ. In Capital Natural de México, Vol. II: Estado de Conservación y Tendencias de Cambio,
517–544. Mexico City: CONABIO.Maple, T (1995). Towards a responsible zoo agenda. In Ethics
on the Ark: Zoos, Animal Welfare, and Wildlife Conservation, 20–30. Norton, BG, Hutchins, M, Stevens, EF & Maple, TL (eds). Washington, DC: Smithsonian Institution Press.
Martín-López, B, Montes, C, Ramírez, L & Benayas, J (2009). What drives policy decision-making related to species conservation? Biological Conservation 142, 1370–1380.
Mazur, NA (1997). Contextualising the Role of the Zoo in Conservation: An Australian Experience. PhD dissertation, University of Adelaide, South Australia.
Mazur, NA & Clark, TW (2001). Zoos and conservation: policy making and organizational challenges. In Species and Ecosystem Conservation: An Interdisciplinary Approach, Clark, TW, Stevenson, MJ, Ziegelmayer, K & Rutherford, MB (eds). Bulletin Series Yale School of Forestry & Environmental Studies, No. 105. New Haven: Yale University.
Morgan, G (1986). Images of Organization. Beverly Hills, California: Sage Publications.
Paehlke, R & Torgerson, D (eds.) (1990). Managing Leviathan: nvironmental Politics and the Administrative State, Peterborough, Ontario, Canada: Broadview Press.
Painter, M (1988). Public management: fad or fallacy. Australian Journal of Public Administration 47, 1–3.
Possingham, HP, Andelman, SJ, Burgman, MA, Medellín, RA, Master, LL & Keith, DA (2002). Limits to the use of threatened species lists. Trends in Ecology & Evolution 11, 503–507.
Pritchard, D, Fa, JE, Oldfield, S & Harrop, S (2011). Bring the captive closer to the wild: redefining the role of ex situ conservation. Oryx 46, 18–23.
Proença, VM, Pereira, HM & Vicente, L (2008). Organismal complexity is an indicator of species existence value. Frontiers in Ecology and Environment 6, 298–299.
Rees, S (1994). Economic rationalism: an ideology of exclusion. Australian Journal of Social Issues 29, 171–185.
Restani, M & Marzluff, JM (2002). Funding extinction? Biological needs and political realities in the allocation of resources to endangered species recovery. BioScience 2, 169–177.
Rhodes, R (1996). Looking beyond managerialism. Australian Journal of Public Administration 55, 106–109.
Sinclair, A (1989). Public sector culture: managerialism or multiculturalism. Australian Journal of Public Administration 48, 382–397.
John E. Fa is Durrell’s Chief Con-servation Officer, and leads the development of Durrell ’s long term strategy, focusing on ma-jor themes and emerging issues in conservation. John builds partnerships with conservation organisations and academic partners and has published
extensively on global biodiversity issues. In 2011 he co-authored a major text on zoo conservation biology. John is also Professor at Imperial College London.
E-mail: [email protected]
Is it just easier for zoos to argue that people visit in order to see popular species such as the ubiquitous meerkat?
Do
min
ic W
orm
ell
Zoos and conservation

Solitaire No. 23 6
Reciprocal agreements for watershed services in Latin AmericaSince 1980, Latin American countries have quadrupled the land area set aside in national, state or municipal protected areas. Public protected areas cannot fully provide the services communities need to survive, such as clean and consistent water supplies, biodiversity pro-tection, and areas for tourism and recreation. Areas of natural habitat outside protected areas, and located amidst largely agricultural landscapes, are needed for the survival of biodiversity, to provide services essential for the production of goods, and to buffer the effects of climate change on local communities.
In rural Latin America, the value of natural ecosystems has often been taken for granted. The last patches of forest and wetlands along streams and on steep slopes are being transformed by farmers into fields and pastures for agriculture and ranching. Small in-dividual benefits to local landowners prevail despite large social costs because institutions compensating landholders for stewardship of public goods are weak or nonexistent.
To address this problem, in the last decade Latin American countries have led institutional innovation in pioneering national programmes that compen-sate local landholders for specific, monitored actions to protect envi-ronmental assets producing public services on non-public land.
In 2009, RARE identified in Natura Bolivia’s ’Reciprocal Agreements for Watershed Services’ (Acuerdos Re-cipricos de Agua, or ARA) a bright spot for community watershed and biodiversity management, using a compensation system for forest conservation and water quality improvement.A first round of ARA campaigns was developed in the Andes region with the hypothesis that Pride Campaigns can become a catalyst for behavioural change to-
wards the adoption of reciprocal agreements and conservation of forests important for key biodiversity species and water production.
The primary objective of this cohort was to protect wa-ter-regulating habitats in the Tropical Montane Cloud Forest Ecosystem in order to provide clean water sup-
plies for human communities, and simultaneously avoid extinctions of AZE species, en-demic to specific ecosystems. In 2012, after evaluating the first ARA cohort, RARE has rep-licated this first experience and
identified 13 sites in Peru, Colom-bia, Ecuador and Mexico.
Since June 2012, 13 Campaign Managers from 13 different Latin American organizations or water en-terprises have begun training in Pride techniques to develop projects. During this process they establish their baseline, identify threats to be addressed, defin their conservation objectives at the local level, and develop their qualitative and quantitative research related to attitudes, barrier removal alternatives, etc. Campaign Managers will receive a University Certifica-tion after finishing the training process. All deliverables are evaluated as well as the effectiveness of their projects.
With this project RARE expects to conserve at least 15.000 ha of forest and paramo and their associated biodiversity in Ecuador, Peru, Colombia and Mexico.
‘Areas of natural habitat
outside protected areas are
needed for the survival of
biodiversity.’
Itala Yépez Zabala trained at Durrell in 1998 and also attended the ISLA course in 2006. She now works for RARE.
E-mail: [email protected]
Cantón Esíndola, Ecuador
A H
ass
e
A H
ass
e
Microcuenca de las Cruces
Conservation report

7 Solitaire No. 23
Illegal wildlife trade threatens Asian bearsThe real life counterpart of Rudyard Kipling’s Baloo faces severe pressure from illegal trade and strong enforcement efforts are needed to protect wild bears in Asia, delegates to an international conference on bear conservation were told recently.
More than 300 experts from 37 countries gathered in India from 26 to 30 November 2012 at the 21st International Conference on Bear Research and Management, organised jointly by the International Bear Association, Ministry of Environment and Forests, Central Zoo Authority of India, the IUCN SSC Bear Specialist Group, Wildlife Institute of India, Wildlife Trust of India, World Society for the Protection of Animals and the International Fund for Animal Welfare.
Asia’s bears are of special concern as demand for their parts for use in traditional medicines has increasingly become a serious threat to the conservation of these species. The illegal trade in bear bile and other bear products was highlighted, with the threats to Asia’s wild bear species being of extreme concern.
In 2011, TRAFFIC produced a report based on an investigation of the trade in bear parts across 13 countries and territories in South-East and East Asia. The report found evidence of widespread illegal sourcing and trade in wild bears and recommended increased efforts to enforce national laws, and better implement the Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES) in the region. However, as recently as 2010 CITES permitted more than 3000 kilos of bear galls to be exported from Russia to Korea. The conference
discussed the need to raise awareness of threats to bears around the world and to reduce demand for bear products.
The recent IUCN resolution concerning the threat to wild bear populations and their conservation was also presented at the conference. Among other things, the resolution calls for the closure of all illegal bear bile farms in Asia. ‘Efforts to shut down the illegal trade in
bear parts in Asia are essential’, Dr Chris R Shepherd, TRAFFIC’s Deputy Director in
South-East Asia, told conference delegates.
The conference also saw the official launch of the first National Bear Conservation and Welfare Action Plan by the Indian Ministry of Environment and Forests by the Environment Minister, Mrs Jayanthi Natarajan.
Conference delegates heard about the remarkable change in attitude towards bears in India, with the trade in sloth bears for roadside performances all but eliminated, for example. However, challenges still exist in India, including human–bear conflict, habitat loss and persistent poaching for illegal trade – all issues flagged as serious concerns in the National Bear Conservation and Welfare Action Plan. Participants highlighted the pressing need to tackle the ever growing cases of man–animal conflict in India as borne out by the attempted torching of a bear in Kashmir in November 2012.
Shubhobroto Ghosh is Senior Pro-gramme Officer at TRAFFIC India. He attended the Principles and Practice training course at Durrell Wildlife Conservation Trust in 1999.
E-mail: [email protected]
‘Efforts to shut down
the illegal trade in
bear parts in Asia are
essential.’
Bear parts are in demand for use in traditional medi-cines.
Asian bears are under severe threat (L); bear gallblad-der for sale in Malaysia (R).
Conservation report
TRAF
FIC
So
uthe
ast
Asia
TRAF
FIC
So
uthe
ast
Asia

Solitaire No. 23 8
IntroductionEnrichment can be described as any change in a captive animal’s life that has a stimulating and beneficial effect on psychological, physical and physiological well-being, and decreases the chance of abnormal behaviour developing (Shepherdson, 1998). The most appropriate way to do this is to maintain a behavioural repertoire in a given species that is as full as possible, within ethical constraints. Primates living in impoverished environments exhibit a lower proportion of normal behaviour in their repertoire, can have lower repro-ductive success, and may have increased cortisol levels – a measure of stress – that can lead to illness and behavioural problems such as stereotypies, aggression, self-harm and coprophagy (e.g. Johnson et al., 1991; Boinksi et al., 1999; Schoenfeld, 1989; Carlstead, 1996; Hosey & Skyner, 2007; Buchanan-Smith, 2010).
Appropriate enrichment can lead to substantial improvements in many of these indicators (e.g. Honess & Marin, 2006; Kitchen & Martin, 1996; Shyne, 2006; Roberts et al., 1999), and of all the primate groups that are kept in captivity, the callitrichids are perhaps the easiest to provide with a highly stimulating environment. No matter what the budget or the size of the enclosure, there can be no excuse for the available space and husbandry techniques failing to provide enrichment. Callitrichids can be very effective ‘flagship species’ (e.g. Dietz et al., 1994) and captive populations in enriched environments can act as a focus for increasing awareness and raising funds, as well as a resource for behavioural research (Kleiman, 1992) and a potential source of candidates for reintroduction programmes (Price et al., 2012). Enriched environments also allow infant callitrichids to develop play and exploratory skills sooner (e.g. Ventura & Buchanan-Smith, 2003).
Callitrichid ecology and enrichmentSubstratesThe evolution of the Callitrichidae (Callimico, marmosets and tamarins) involved reduction in body size (dwarfism) (Ford, 1980; Rosenberger, 1984). As a result, they can use many parts of the forest environment and a huge variety of substrates. They are not only able to move along very thin, flexible supports but also, because they have claws (instead of nails) on their digits, they can cling to large, rough tree trunks. Providing the opportunity to move around on all these different types of support and substrate is a basic requirement of enclosure design. This enables individuals to develop bal-ance and coordination, and the ability to judge distance, as
well as using a variety of different methods of locomotion, including climbing and leaping. Giving callitrichids the op-portunity to make extended leaps is important; animals that have been housed for long periods in small enclosures with no need to judge distances accurately are at a disadvantage if they are then given access to a large area, and may incur injuries from falls (E. Price, pers. obs.). However, they can learn these skills fairly rapidly when given appropriate hous-ing conditions.
Activity, diet and foraging techniquesFinding and processing food is central to the life of wild callitrichids, which spend up to half their day foraging (e.g. Veracini, 1998; Ferrari & Rylands, 1994; Ferrari & Digby, 1996; Digby & Barreto, 1996; Peres, 1989; Dietz et al., 1997; Albernaz, 1997; Rylands, 1989; Garber, 1993; Lopes & Ferrari, 1994; de la Torre et al., 1995; Savage, 1990; Egler, 1992). One of the primary needs in captivity is, therefore, to increase foraging behaviour and much enrichment is based on this. However, the various callitrichid genera have evolved to fit many different ecological niches, and even within a genus there are differences in foraging techniques. We need to consider the natural foraging styles of each species in de-veloping appropriate enrichment.
Insects are an important component of the diet of all callitrichids, and they forage for invertebrate prey at all levels, in foliage, tree crowns, leaf litter accumulations and very occasionally on the forest floor. Marmosets, however, have dentition that enables them to gouge holes in trees to extract gum, and up to 80% of the diet of wild marmosets may consists of exudates (see Huber & Lewis, 2011).
Lion tamarins have long hands that they can use to reach into holes and into the deep centres of plants, such as bromeliads (Rylands, 1989). This behaviour is frequently seen in captivity as lion tamarins probe every nook and cranny in their enclosure. Lion tamarins will automatically investigate a hole with their hands and frequently do so while looking in a different direction. Hanging plants with their associated gaps and crevices will stimulate this natural foraging behaviour. Other species, such as cotton-top tamarins Saguinus oedipus, may be much more reluctant to reach into holes when they cannot see what is in them (E. Price, pers. obs.).
The placement of enrichment within the enclosure can also be important. Marmosets are quicker to explore objects placed higher up and to spend more time investigating them (Majolo et al., 2003), while Hardie & Buchanan-Smith (2000) found differences between two species of Saguinus depending on
Enriched environments for callitrichids
Review article
Dominic Wormell1, Jenna Hunt1, Eric Bairrão Ruivo2 & Eluned Price1
1Durrell Wildlife Conservation Trust; 2ZooParc de Beauval, France
Abstract Environmental enrichment aims to promote well-being by providing opportunities for captive animals to exhibit a full behavioural repertoire. Callitrichids can easily be kept to extremely high standards in captivity, greatly enhancing their potential role as conservation ‘flagships’. Their particular characteristics mean that providing them with an environment that is naturally enriched, rather than supplied with artificial enrichment devices, is relatively easy and can be achieved at low cost. The key is to consider the ecology and social organization of marmosets and tamarins in the wild, and use this information to shape the design of accommodation, diets and social groupings, all of which can contribute to ‘enriching’ their environment.
Key words: callitrichid; conservation; ecology; enrichment; marmoset; social organization; tamarin

9 Solitaire No. 23
their ecological characteristics: S. labiatus, which uses higher levels of the forest than sympatric S. fuscicollis, approached novel objects located higher in the enclosure more quickly.
Enriched environments or environmental enrichment?
With a high-quality complex environment, even a small space can provide much of the enrichment a callitrichid re-quires without the need for artificial devices. Enrichment therefore starts with good enclosure design (Wormell & Brayshaw, 2000). Complex branching and rope systems give callitrichids the chance to display their full range of lo-comotor abilities, while planting is the easiest way to create a semi-natural environment where the animals can express their natural behavioural repertoire (Fig. 1). In tropical cli-mates this can be achieved easily with plants endemic to the region the species comes from. Small fruiting or flowering trees, while taking a while to establish, will provide sensory stimulation for the animals, not only by providing fruit/flow-ers but also by encouraging insects into the enclosure which the animals can then forage on naturally. Trees that produce edible exudates are ideal for marmosets and suitable species are available in temperate climates (Fig. 2).
For new enclosures in temperate climates the best plants are evergreen shrubs and trees and fast growing climbers; ivy Hedera spp is ideal. As well as providing substrates to forage in it also gives cover for these naturally shy animals, and in more tropical climates it also has the advantage of providing shade. A vine-covered log provides many microhabitats for invertebrates. Even a simple piece of rotting log will provide great enrichment as it is investigated and pulled apart.
If ground space is limited or cannot be planted, hanging baskets can be used, either suspended from the enclosure roof or attached to large branches/perching. Insect houses and logs covered with creepers can maintain self-sustaining insect populations, providing long-term enrichment with minimal cost in time or money.
Artificial devicesMany devices designed to provide stimulation for captive pri-mates have been developed with laboratory animals in mind (e.g. Buchanan-Smith, 2010). Zoo primates usually have
access to larger, and often more stimulating, environments than laboratory primates, and some measures of abnormal behaviour, such as self-injury, appear to be less of an issue in zoos than in laboratories (e.g. Hosey & Skyner, 2007). Nevertheless, problems may still arise if it is not possible to provide a sufficiently enriched environment simply through appropriate enclosure design and furnishing.
There may also be seasonal differences in the need to provide enrichment: in temperate climes where animals have outside as well as inside areas, winter has the obvious effect of confining animals indoors owing to poor weather, and there are few insects and fruits naturally available during the colder months. Social tension can also be exacerbated – colony housing increased aggression in Saguinus geoffroyi, and may have led to reduced infant rearing success (Kuhar et al., 2003), while aggression from parents to offspring increased in large groups of cotton-top tamarins S. oedipus under crowded conditions (Caperos et al., 2011). Inside areas also tend to provide a far less enriched environment and this, along with closer proximity, can lead to increased stress levels: a comparison of cortisol in three callitrichid species showed that baseline levels were higher in the winter than in the summer (McCallister, 2005).
Foraging devices can also be useful when it is unavoidable for a callitrichid to be temporarily housed alone, and they can increase foraging time much more in singly-housed callitrichids than in those living in groups (e.g. Chamove & Scott, 2005).
Any artificial enrichment device needs to be cheap to make, quick to use, easily serviced within the husbandry routine and, ideally, reusable. If too elaborate and time-consuming, they will not be used and will slip from the daily routine in a busy animal carer’s daily schedule.
The animals’ interest can also wane relatively quickly (e.g. Vigres et al., 2003). An unchanging environment, even if apparently relatively enriched, can soon decline in attractiveness and in its effect on behaviour. For example, Kitchen & Martin (1996) found that laboratory marmosets moved to an enriched but unchanging environment from a smaller cage initially increased their activity level, but it then declined again.
Fig. 2. Silvery marmoset at Durrell Wildlife Park gum-feeding on natural vegetation.
Callitrichid enrichment
Fig. 1. An outdoor enclosure for callitrichids at Durrell Wildlife Park with natural trees, climbing plants, ropes and rotting wood.

Solitaire No. 23 10
Providing several different enrichment items in enclosures allows members of the groups to forage independently and, therefore, relieves tension and prevents fights over food. Most methods are aimed at providing food in a more naturalistic way, increasing foraging and handling time (Fig. 3).
Simple foraging devices are often useful in indoor areas during periods of bad weather or in climates that are less than ideal. Puzzle feeders are attractive to callitrichids (e.g. Roberts et al., 1999; de Rosa et al., 2003), and foraging tasks tend to elicit stronger responses than devices that do not contain hidden food (Chamove & Scott, 2005; Majolo et al., 2003).
Gum-feeding devices such as the ‘gum-tree’ described by McGrew et al. (1986) can be constructed for marmosets, requiring them to gnaw holes into the wood before the gum can be extracted. Gum tends to be preferred in liquid rather than solid form (Herron et al., 2001). Surprisingly, however, gum feeders are not widely used in zoos (Huber & Lewis, 2011).
Insects or other treats can be given on hanging rotten logs or creeper-covered logs, in hanging baskets or plastic crates filled with substrates such as hay or wood wool, in insect houses or in logs or coconuts with large holes drilled into them, or in puzzle feeders. Cane or rope ‘kebabs’ with fruit attached means the animals have to jump onto a moving substrate in order to reach the food.
A variety of food items can be used in these devices but insects are ideal. However, mealworms Tenebrio molitor should not be used too much as they are relatively low in calcium, and a dietary calcium–phosphorous ratio between 1:1 and 2:1 is typically recommended (e.g. NRC, 2003). Care should also be taken not to use sweet foods, such as dried fruit or honey, as this can result in tooth abscesses (D. Wormell, pers. obs.), and the overall calorific content of the diet should not be increased to undesirable levels (Majolo et al., 2003).
Other forms of enrichmentA sprinkler system can mimic a rain shower and can encour-age invertebrates into the enclosure during periods of dry weather when the enclosure may otherwise become quite bare.
Simple platforms and shelters can have many benefits. Clumps of lianas are often used by callitrichids as safe roosting spots (Mittermeier & van Roosmalen, 1981), and
Leontopithecus spp use tree holes for daily roosting activities. Resting platforms should be placed high up so animals can relax with a feeling of security; enclosures should therefore be made as high as possible.
Conduit tubing cut into sections provides shelter and privacy for the animals at very little cost. It can diffuse tension within groups as it can provide a retreat during conflict, and gives nervous animals somewhere to hide when keepers are servicing the enclosure.
Callitrichids are social primates, living in families or extended groups with communal infant care, and complex vocal, visual and olfactory communication systems (Epple et al., 1993; Snowdon, 1993; Tardif et al., 1993). To stimulate olfaction, for example, occasional scent-based enrichment can be introduced; for example, by putting a branch marked by another group into the enclosure. Aromatic plant scents, such as lavender and scented mayweed, can be tried. However, its complex nature and the potential for introducing stressors into the environment mean that olfactory enrichment should be used with care (Clark & King, 2008). Callitrichids’ use of scent-marking to communicate means that they can, in a sense, enrich themselves, and the role of their carers is to facilitate this – for example, it is important to avoid over-cleaning so that scents communicating social information are not completely removed from the enclosure.
DiscussionA captive callitrichid can be said to have an enriched life when at any given point in the day it has a number of choic-es that it can make about what it will do next; enrichment is about empowering animals and increasing their ability to cope with challenges, and the provision of complexity, choice and control in captive environments is key to achiev-ing this (Buchanan-Smith, 2010; Markowitz & Aday, 1989).
This is where outdoor enclosures with natural vegetation that changes seasonally and houses an unending supply of insects etc have a clear advantage. Simply being outside can be a strong enough incentive for a callitrichid to use one area in preference to another – common marmosets given a choice between a smaller outdoor cage and a large, enriched indoor enclosure preferred to outdoor area, the indoor cage declining in attractiveness over time (Pines et al., 2007).
Fig. 3. Artificial enrichment devices for callitrichids. (L) Log drilled with holes; (R) insect foraging box.
Review article

11 Solitaire No. 23
It is important to consider that there may be forms of enrichment that provide stressful stimuli but that may in fact confer benefits, either immediately or in the future, as well as more closely mimicking the natural environment and prompting behavioural responses that would be normal in the wild. Some behaviours that occur in response to perceived threats, for example, may be necessary for mental fitness and, indeed, short acute stress may be needed to provide a healthy immune system. It can also promote positive social relationships (e.g. Chamove & Moodie, 1990).
Preparation for release back into the wild also means that behaviour that may only be rarely expressed in captivity needs to be stimulated. For example, captive tamarins have been taken by predators after release into the wild (e.g. Valladares-Padua et al., 2000) as they are much less reluctant than wild-born animals to descend to the ground, and may be unfamiliar with certain classes of predators as a result of lack of exposure. So it is important for a species to retain behavioural fitness, which may deteriorate with successive generations in captivity.
However, the aim is to mimic a natural existence to improve health and well-being and reduce stress caused by the captive environment; too much or too frequent alteration in their environment can in itself be detrimental. It is important to remember that the natural disposition of a callitrichid is to be alert to predators, as they have many in the wild (e.g. Stafford & Ferreira, 1995; Dunbar, 1995). The use of enrichment must not cause chronic stress.
Callitrichids, by their very nature, can take advantage of many different possible choices, and so offer tremendous opportunities for zoos to show themselves in the best possible light. A failure to enrich zoo environments will not only be detrimental to the well-being of the animals living in them but also will have a negative effect on the messages modern zoos should be seeking to convey and, more broadly, on conservation and research goals.
AcknowledgementsWe are grateful to the staff at Durrell Wildlife Conservation Trust and Beauval Zoo for their commitment to improving the management of callitrichids.
ReferencesAlbernaz, ALKM (1997). Home range size and habitat use
in the black lion tamarin (Leontopithecus chrysopygus). International Journal of Primatology 18, 877–887.
Boinksi, S, Swing, SP, Gross, TS & David, JK (1999). Environmental enrichment of brown capuchins (Cebus apella): behavioral and plasma and fecal cortisol measures of effectiveness. American Journal of Primatology 48, 49–68.
Bromm, DM & Zanella, AJ (2004). Brain measures which tell us about animal welfare. Animal Welfare 13, S41-45.
Buchanan-Smith, HM (2010). Environmental enrichment for primates in laboratories. Advances in Science and Research 5, 41-56.
Caperos, JM, Sánchez, S, Peláez, F, Fidalgo, A & Morcillo, A (2011). The effect of crowding on the social behavior of the cooperatively breeding cotton-top tamarin (Saguinus oedipus). International Journal of Primatology 32, 1179-1189.
Carlstead, K (1996). Effects of captivity on the behaviour of
wild mammals. In Wild Mammals in Captivity: Principles and Techniques, 317–333. Kleiman, DG, Allen, ME, Thompson, KV & Lumpkin, S (eds). Chicago: University of Chicago Press.
Chamove, AS & Moodie, EM (1990). Are alarming events good for captive monkeys? Applied Animal Behaviour Science 27, 169–176.
Chamove, AS & Scott, L (2005). Forage box as enrichment in single- and group-housed callitrichid monkeys. Laboratory Primate Newsletter 44(2), 13-17.
Clark, F & King, AJ (2008). A critical review of zoo-based olfactory enrichment. In Chemical Signals in Vertebrates 11, 391-398. Hurst, JL, Beynon, RJ, Roberts, SC & Wyatt, TD (eds). New York: Springer.
de Rosa, C, Vitale, A & Puopolo, M (2003). The puzzle-feeder as feeding enrichment for common marmosets (Callithrix jacchus): a pilot study. Laboratory Animals 37, 100-107.
Dietz, JM, Dietz, LA & Nagagata, EY (1994). The effective use of flagship species for conservation of biodiversity: the example of lion tamarins in Brazil. In Creative Conservation: Interactive Management of Wild and Captive Animals, 32–49. Olney, PJS, Mace, GM & Feistner, ATC (eds). London: Chapman & Hall.
Dietz, JM, Peres, CA & Pinder, L (1997). Foraging ecology and use of space in wild golden lion tamarins (Leontopithecus rosalia). American Journal of Primatology 41, 289–305.
Digby, LJ & Barreto, CE (1996). Activity and ranging patterns in common marmosets (Callithrix jacchus). Implications for reproductive strategies. In Adaptive Radiations of Neotropical Primates, 173–185. Norconk, MA, Rosenberger, AL & Garber, PA (eds). New York: Plenum Press.
Egler, SG (1992). Feeding ecology of Saguinus bicolor bicolor (Callitrichidae: Primates) in a relict forest in Manaus, Brazilian Amazonia. Folia Primatologica 59, 61–76.
Epple, G, Belcher, AM, Küderling, I, Zeller, U, Scolnick, L, Greenfield, KL & Smith III, AB (1993). Making sense out of scents: species differences in scent glands, scent-marking behaviour, and scent-mark composition in the Callitrichidae. In Marmosets and Tamarins: Systematics, Behaviour, and Ecology, 123–151. Rylands, AB (ed.). Oxford: Oxford University Press.
Ferrari, SF & Digby, LJ (1996). Wild Callithrix groups: stable extended families? American Journal of Primatology 38,19–27.
Ferrari, SF & Rylands, AB (1994). Activity budgets and differential visibility in field studies of three marmosets (Callithrix spp.). Folia Primatologica 63, 78–83.
Ford, SM (1980). Callitrichids as phyletic dwarfs, and the place of the Callitrichidae in Platyrrhini. Primates 21, 31–43.
Garber, PA (1993). Seasonal patterns of diet and ranging in two species of tamarin monkeys: stability versus variability. International Journal of Primatology 14, 145–166.
Hardie, SM & Buchanan-Smith, HM (2000). Responses of captive single- and mixed-species groups of Saguinus to novel nonthreatening objects. International Journal of Primatology 21, 629-648.
Herron, S, Price, EC & Wormell, D (2001). Feeding gum arabic to New World monkeys: species differences and palatability. Animal Welfare 10, 249-256.
Honess, PE & Marin, CM (2006). Enrichment and aggression in primates. Neuroscience and Bioehavioral Reviews 30,
Callitrichid enrichment

Solitaire No. 23 12
413-436.Hosey, GR & Skyner, LJ (2007). Self-injurious behaviour in zoo
primates. International Journal of Primatology 28, 1431–1437.
Huber, HF & Lewis, KP (2011). An assessment of gum-based environmental enrichment for captive gummivorous primates. Zoo Biology 30, 71-78.
Johnson, EO, Kamilaris, TC, Carter, S, Gold, PW & Chrousos, GP (1991). “Environmental stress” and reproductive success in the common marmoset (Callithrix jacchus). American Journal of Primatology 25, 191–201.
Kitchen, AM & Martin, AA (1996). The effects of cage size and complexity on the behaviour of captive common marmosets, Callithrix jacchus jacchus. Laboratory Animals 30, 317-326.
Kleiman, DG (1992). Behavior research in zoos: past, present, and future. Zoo Biology 11, 301–312.
Kuhar, CW, Bettinger, TL, Sironen, AL, Shaw, JH & Lasley, BL (2003). Factors affecting reproduction in zoo-housed Geoffroy’s tamarins (Saguinus geoffroyi). Zoo Biology 22, 545-559.
Lopes, MA & Ferrari, SF (1994). Foraging behavior of a tamarin group (Saguinus fuscicollis weddelli) and interactions with marmosets (Callithrix emiliae). International Journal of Primatology 15, 373–387.
Majolo, B, Buchanan-Smith, HM & Bell, J (2003). Response to novel objects and foraging tasks by common marmoset (Callithrix jacchus) female pairs. Lab Animal Europe 3(3), 25-32.
Markowitz, H & Aday, C (1998). Power for captive animals: contingencies and nature. In Second Nature: Environmental Enrichment for Captive Animals, 47–58. Shepherdson, D. J., Mellen, J. D. & Hutchins, M. (eds). Washington, DC: Smithsonian.
McCallister, JM (2005). Behavioural and Physiological Differences Between Callitrichid Primates. PhD dissertation, Queen’s University of Belfast.
McGrew, WC, Brennan, JA & Russell, J (1986). An artificial gum-tree for marmosets (Callithrix j. jacchus). Zoo Biology 5, 45–50.
Mittermeier, RA & van Roosmalen, MGM (1981). Preliminary observations on habitat utilization and diet in eight Surinam monkeys. Folia Primatologia 36, 1–39.
NRC (2003). Nutrient Requirements of Nonhuman Primates, 2nd edn. Washington, DC: National Academies Press.
Peres, CA (1989). Costs and benefits of territorial defense in wild golden lion tamarins, Leontopithecus rosalia. Behavioural Ecology and Sociobiology 25, 227–233.
Pines, M.K., G. Kaplan, and L.J. Rogers (2007). A note on indoor and outdoor housing preferences of common marmosets (Callithrix jacchus). Applied Animal Behaviour Science 108, 348-353.
Price, EC, Wormell, D, Brayshaw, M, Furrer, S, Heer, T & Steinmetz, HW (2012). Managing free-ranging callitrichids in zoos. International Zoo Yearbook 46, 123-136.
Roberts, RL, Roytburd, LA & Newman, JD (1999). Puzzle feeders and gum feeders as environmental enrichment for common marmosets. Contemporary Topics in Laboratory Animal Science 38(5), 27-31.
Rosenberger, AL (1984). Aspects of the systematics and evolution of the marmosets. In A Primatologia no Brasil, 159–180. de Mello, MT (ed.). Brasilia: Sociedade Brasileira
de Primatologia. Rylands, AB (1989). Sympatric Brazilian callitrichids: the black
tufted-ear marmoset, Callithrix kuhli, and the golden-headed lion tamarin, Leontopithecus chrysomelas. Journal of Human Evolution 18, 679–695.
Savage, A (1990). The Reproductive Biology of the Cotton-top Tamarin (Saguinus oedipus oedipus) in Colombia. PhD dissertation, University of Wisconsin-Madison.
Schoenfeld, D (1989). Effects of environmental impoverishment on the social behavior of marmosets (Callithrix jacchus). American Journal of Primatology Supplement 1, 45–51.
Shepherdson, DJ (1998). Tracing the path of environmental enrichment in zoos. In Second Nature: Environmental Enrichment for Captive Animals, 1–12.Shepherdson, DJ, Mellen, JD & Hutchins, M (eds). Washington, DC: Smithsonian.
Shyne, A (2006). Meta-analytic review of the effects of enrichment on stereotypic behavior in zoo mammals. Zoo Biology 25, 317-337.
Snowdon, CT (1993). A vocal taxonomy of the callitrichids. In Marmosets and Tamarins: Systematics, Behaviour, and Ecology, 78–94. Rylands, AB (ed.). Oxford: Oxford University Press.
Tardif, SD, Harrison, ML & Simek, MA (1993). Communal infant care in marmosets and tamarins: relation to energetics, ecology, and social organization. In Marmosets and Tamarins: Systematics, Behaviour, and Ecology, 220–234. Rylands, AB (ed.). Oxford: Oxford University Press.
de la Torre, S, Campos, F & de Vries, T (1995). Home range and birth seasonality of Saguinus nigricollis graellsi in Ecuadorian Amazonia. American Journal of Primatology 37, 39–56.
Valladares-Padua, C, Martins, CS, Wormell, D & Setz, EZF (2000). Preliminary evaluation of the reintroduction of a mixed wild-captive group of black lion tamarins (Leontopithecus chrysopygus). Dodo 36, 30-38.
Ventura, R & Buchanan-Smith, HM (2003). Physical environmental effects on infant care and development in captive Callithrix jacchus. International Journal of Primatology 24, 399-413.
Veracini, A (1998). Activity pattern in a wild group of Callithrix argentata. In XVIIth Congress of the International Primatological Society, Antananarivo, Madagascar, August 10-14, 1998. Antananarivo, Madagascar: University of Antananarivo. [Poster presentation.]
Vignes, S, Newman, JD & Roberts, RL (2001). Mealworm feeders as environmental enrichment for common marmosets. Contemporary Topics in Laboratory Animal Science 40(3), 26-29.
Wormell, D & Brayshaw, M (2000). The design and redevelopment of New World primate accommodation at Jersey Zoo: a naturalistic approach. Dodo 36, 9 –19.
Dominic Wormell is Head of Mammals and Jenna Hunt is a Senior Mammal Keeper at Durrell Wildlife Conservation Trust. Eric Bairrão Ruivo is Science and Conservation Director at ZooParc de Beauval, France, and Eluned Price is Wildlife Park Research Coordinator at Durrell.
E-mail: [email protected]
Review article

13 Solitaire No. 23
The training and management of an insulin-dependent Sulawesi crested black macaque
Louise Bestwick
Durrell Wildlife Conservation Trust
Abstract Diabetes mellitus occurs relatively frequently in captive Macaca nigra. A protocol is described that used positive reinforcement to train the dominant male in a captive group of M. nigra at Durrell Wildlife Park to allow daily unrestrained insulin injections to control diabetes mellitus.
Key words: diet, glycaemic index, injection, Macaca nigra, positive reinforcement, weight
IntroductionDiabetes mellitus is a metabolic disorder characterised by hyperglycaemia, and has been described in numerous non-human primates (Cheeke & Dierenfeld, 2010; Arslan et al., 2007). A particularly high incidence of diabetes has been recorded in captive Macaca nigra, and there is evidence to suggest it occurs in the wild population (Howard, 1974; Howard et al., 1999). There have been four confirmed cases of diabetes mellitus in this species at Durrell Wildlife Park.
In 2011 the fourteen year old dominant male Macaca nigra ‘Syfon’ was observed by keepers to have symptoms associated with diabetes: periodic lethargy, a swollen abdomen, polydipsia, polyuria, diarrhoea and weight loss. He was anaesthetised on 6 December 2011, and diabetes was confirmed with a blood glucose level of 15 and fructosamine level of 667.
In addition to changes in diet, a training plan was instigated immediately to allow monitoring and treatment with subcutaneous insulin. Positive reinforcement training is used throughout zoos in a variety of situations, and is a tool used to manage animals effectively and humanely, in a stress free environment (Laule et al., 2003). This paper discusses how he was treated from an animal manager’s perspective; it is not a veterinary management plan.
DietThe cause of diabetes in this species is not fully understood but is suspected to be a genetic predisposition exacerbated by captive diets and environments (Howard, 1974). Once Sy-fon was diagnosed with diabetes the macaque diet at Durrell Wildlife Park was reviewed. Diabetes is the only recorded dietary issue with this species so the diet was adjusted to keep it nutritionally similar whilst exchanging high glycae-mic index (GI) vegetables and fruit with low GI alternatives. The glycaemic index is a way of ranking foods according to their glycaemic effect – the rate at which carbohydrate is digested and absorbed. A low glycaemic response has been reported to facilitate blood glucose regulation and improve lipid metabolism in diabetes (Jarvi et al., 1999).
The basic source of nutrients in the diet came from a primate pellet, which was very high in sugar. As it took some time to match this pellet nutritionally, Syfon was initially separated from the group and given a lower sugar leaf-eater pellet, whilst the rest received primate pellet. Low GI fruits (apple and pear) were left in to be given at the morning feed,
and high-nutrient banana at the afternoon feed. These highly valued foods were used as a management tool for the whole group of macaques, and unless otherwise stated Syfon’s portion was cut up and used as the reward for training.
TrainingThere were two training sessions a day, one in the morning before he had any food, and the other in the afternoon, before the final feed.
Commands and rewards were initially given as a response to a desired behaviour, and once the association was made the command was given to promote the behaviour, and the reward to reinforce it.
Group separationTo allow for his different diet and training, Syfon needed to be separated from the group twice daily. Due to his condi-tion he was slower than the others, so they could be called into the adjacent cage (on the left in Fig. 1) from the inside and rewarded, and he could be left shut in and rewarded. It took three days before he purposely stayed behind. If any macaque stayed with him no macaque would get any food until they were in the correct place.
After a month Syfon started to move into the cage with the others, or would be waiting in the cage. He could then be encouraged into the tunnel between the cage and the inside area, for a reward, by calling his name, and then led inside. This became the standard routine.
The greatest complications throughout all his training came from separating Syfon from the group, even once he was regularly receiving his insulin. For example, after two weeks he stopped coming into the building for his training sessions, perhaps because he was being separated for approximately an hour to allow for his different food. As the diet had been changed by this point so the macaques all had the same pellet, Syfon only needed to be separated briefly for training, and so moved easily again.
On some occasions Syfon would not push past the other macaques blocking the tunnel, and so they were lightly sprayed with water to move them out of the way. Sometimes, he would not move at all from the middle of the cage. Feeding the others treats at the cage mesh encouraged him to come over, and he could then be led into the tunnel.
When the females were in oestrus he would not leave them, particularly as we had a boisterous juvenile male in the group.
Injection training for a male macaque

Solitaire No. 23 14
On those occasions it was necessary to separate the juvenile male off with some of the other young males or no oestrus females, and then Syfon could be separated.
There were different stages to his routine requiring him to move between rooms for weighing, urinating and insulin (see Fig. 1). Rather than being trained to move between rooms with further commands, he was instead led using a reward, and he quickly moved without reward, the trigger apparently being the opening of the slides between the rooms.
Urine training Urine sampling was necessary to monitor Syfon’s glucose and ketone levels. Peanuts were used as they are high value but low GI, and to avoid confusion, he was not given them at any other time. Initially he was given a peanut every time he urinated, and given the command ‘Wee wee Syfon’. How-ever, due to his condition he was urinating so frequently when keepers were not present that the command was not being sufficiently enforced. Therefore during his morning and afternoon separations he was encouraged into the small room with a reward and shut in until he urinated, then given a peanut and command, and let into the middle room for injec-tion training. It took three days, with two sessions per day, for him to either urinate or attempt to urinate immediately once shut in the room, and after three weeks he no longer needed a peanut reward.
Injection trainingA tunnel in the middle room (see Fig. 1) was used for his injection training so he could be injected through the mesh.
Step 1 – coming into the tunnel, 2 days. Syfon was led into the tunnel and rewarded for sitting down in a position he naturally chose. The trigger appeared to be placing a yellow cup with his rewards in by the tunnel, though the command ‘Syfon tunnel’ was used.
Step 2 – pen pressing, 5 days. The site on Syfon’s right leg that would be used for injection was touched through the mesh using a pen, with 3-5 presses per session and the com-mand ‘Syfon leg’. It took three days before he offered his leg against the mesh for pen pressing. If he pushed it away he was not rewarded. Pen pressing continued for two more days, increasing in force, until he showed no negative re-sponse.
Step 3 – syringe pressing and water squirting, 3 days. The same action, command and reward were used, except now with a 1 ml syringe. On some presses, at random, water was squirted out of the syringe when it was pressed into Syfon’s leg, with no negative response.
Step 4 – needle insertion, 2 days. The insulin needle was used for this, with the cap on or off, and the needle inserted into his leg. It was alternated at random to avoid sensitivity from overusing the needle. He showed no negative response.
Step 5- injection of fluid, 17 days. Syfon was initially inject-ed with 0.01 unit of saline solution, working up to 0.06 units (the amount of caninsulin he would be receiving), increasing in 0.01 ml increments and only when negative behaviours were minimal. He was only injected with the fluid once at each session, at random, and at other times pressed with a capped syringe or needle. He did initially respond negatively to the fluid (pulling his leg away, vocalising, grabbing the syringe), and so some sessions used just a capped syringe to ensure that he continued to come for reward and that there was not always a negative association. It was necessary to hold the needle in his leg for at least 5 secs before injecting the fluid to reduce negative responses. If he continued to react, the keeper turned his or her back to him, waited for him to come out of the tunnel, and began the session again. At 17 days we were confident we would succeed every time,
Technical report
Cage
Middle Room
Keeper area
Outside area
Small Room Large Room
KEY
Popholes
Walkthrough tunnels
=
Injection tunnel
Weighing scales
Urination site
Fig. 1. Diagram of macaque enclosure at Durrell Wildlife Park, showing access points and locations for various phases of training (not to scale).

15 Solitaire No. 23
so use of the actual medication could begin, and only the injection was given at the session, with no additional cap or needle pressing.
Initially Syfon’s insulin was given in the morning only, but in case the dose was increased to twice daily, afternoon training sessions continued with a capped syringe in order to reinforce the desired behaviours without causing unnecessary sensitivity.
After he had been on the insulin for about two weeks Syfon began to present his back, apparently because the site on his right leg was becoming sore. He was not rewarded for this, but he was rewarded for presenting his right leg and was given the command ‘Syfon thigh’. It took only a few days, following the training steps, for him to be injected in the leg, however he appeared to offer his legs by choice, rather then responding to alternate leg commands. Throughout his treatment he showed a preference for the right leg.
Scale trainingOnce he was fully trained to urinate, and was consistently doing so in the same place, scales were placed adjacent to this site in front of the mesh. After he had urinated a reward was placed in front of the scales, and he would climb onto the scales to get the reward, while the command ‘Syfon scales’ was given. In three days he responded to ‘Syfon scales’ and positioned himself correctly without being led.
ConclusionA key factor which contributed to the success of this train-ing was consistency; in the initial stages one keeper worked with Syfon all the time, with one other person covering days off – there was an existing good relationship between these keepers with a high level of communication. Even once he was fully trained, Syfon became confused at training session when more keepers became involved. A lot of time needed to be dedicated to training in the early stages, and so other members of staff were required to cover other aspects of the routine – this was important in ensuring that the keepers were never rushed, which could have led to frustration and lack of clarity for Syfon. The author has also observed a successful-ly trained diabetic lion-tailed macaque at Apenheul Primate Park, which provided additional motivation and encourage-ment to persevere with training.
This protocol would be used and recommended for managing diabetes in macaques in the future. Syfon responded very well to training; however, he did not respond as well to various type of insulin, so clearly more needs to be learnt here. He also suffered from pancreatitis and other health complications.
Although in the eight months following his diagnosis Syfon mated all three of the cycling females in his group, and fathered at least one offspring, due to a serious decline in health he was euthanased on 12 September 2012.
Future developmentsAs diabetes is a known condition in this species the aim is to train all macaques for urine sampling and weighing to monitor them regularly, permitting earlier diagnosis. Ideally they will also be injection-trained so that they can be given insulin if ever necessary, and to reduce the stress currently associated with using a dart gun when an animal needs to be anaesthetised for other reasons. It may also be possible to take
blood samples, by training the macaques to place an arm in a blood sleeve and remain stationary for venipuncture, as has been done in rhesus macaques (Coleman et al., 2008).
AcknowledgementsThanks to all members of Durrell’s macaque section and vet-erinary department at the time of this project for their help, to staff at Apenheul Primate Park for their advice, and to Sarah Fowkes and Jonny Poole for additional support.
ReferencesArslan, HH, Nisbet, C & Guvenc, T (2007). Spontaneous
amyloidosis and diabetes mellitus in a rhesus macaque (Macaca mulatta). Bulletin of the Veterinary Institute in Pulawy 51, 655-659.
Cheeke, PR & Dierenfeld, E (2010). Comparative animal nutrition and metabolism. CABI 16, 178-186.
Coleman, K, Pranger, L, Maier, A, Lambeth, SP, Perlman, JE, Thiele, E & Schapiro, SJ (2008). Training rhesus macaques for venipuncture using positive reinforcement techniques: a comparison with chimpanzees. Journal of the American Association for Laboratory Animal Science 47, 37–41.
Howard, CF (1974). Diabetes in Macaca nigra: metabolic and histologic changes. Diabetologia 10, 671-677.
Howard, CF, Fang, T-Y, Southwick, C, Erwin, J, Sugardjito, J, Supriatna, J, Kohlhaas, A & Lerche, N (1999). Islet cell antibodies in Sulawesi macaques. American Journal of Primatology 47, 223-229.
Jarvi, AE, Karlstrom, BE, Granfeldt, YE, Bjork, IE, Asp, N-GL & Vessby, BOH (1999). Improved glycemic control and lipid profile and normalized fibrinolytic activity on a low–glycemic index diet in type 2 diabetic patients. Diabetes Care 22, 10 -18
Laule, GE, Bloomsmith, MA & Schapiro, SJ (2003). The use of positive reinforcement training techniques to enhance the care, management and welfare of laboratory primates. Journal of Applied Animal Welfare Science 6, 163-173.
Louise Bestwick is a Mammal Keeper at Durrell Wild-life Park, specialising in the care of macaques and lemurs.
E-mail: [email protected]
Injection training for a male macaque
Fig. 2. Syfon in the injection tunnel, receiving a reward while having a syringe pressed against his leg.

Solitaire No. 23 16
IntroductionCrested black macaques (Macaca nigra; see Fig. 1) are the most endangered of the seven macaque species found on the Indonesian island of Sulawesi, and are critically endangered (Supriatna & Andayani, 2008). Captive breeding pro-grammes are vital for the conservation of such species, and fortunately there is a large captive population of M. nigra.
However, macaques have complex social structures, and live in mixed multi-male and multi-female groups (Reinhart, 2008). Males display a linear dominance hierarchy (Reed et al., 1997) with a dominant male in each group. Young males leave the group as they become sexually mature, but captivity restricts this natural process, making management of captive groups difficult. A key element in the development of young males is the need for a large social group to interact and learn social skills by playing (Poirier & Smith, 1974), but as male primates transition from childhood to adulthood, they undergo dramatic biological, physiological and morphological changes, known as adolescence (Richards et al., 2009). This stage of development may involve increased testosterone and aggression directed at others (Anestis, 2006). Rose et al. (1978) have also documented an increase in sexual behaviour in adolescent primates, thus increasing the risk of inbreeding with relatives in captivity, and creating competition for the dominant male, which could lead to severe and possibly fatal aggression.
Organisations that hold primates with complex social structures struggle to maintain control of aggression without removing individuals from groups, which is only possible if there are vacant enclosures. Exporting males to other breeding groups is ideal, but transport is expensive and the process can be time consuming.
Chemical castration is reversible and provides similar effects to that of surgical castration, such as reduced copulation and aggressive behaviour in mammals (Bertschinger et al., 2001; Asa & Porton, 2005). This could be a possible solution
for preventing inbreeding and aggression within social groups of Sulawesi crested black macaques.
An injectable hormone implant containing the active ingredient deslorelin acetate has been successfully used in many mammals, including some primate species, to reduce aggressive behaviour (De Nys et al., 2010; Wagner et al., 2005; Asa & Porton, 2005). It is a gonadotropin-releasing hormone (GnRH) agonist which triggers the release of follicle-stimulating hormone (FSH) and lutenising hormone (LH) from the anterior pituitary gland, both of which are responsible for the production of testosterone.
The objective of this project was to study play, aggression and copulation behaviour in male macaques of different ages before and during treatment of an adolescent M. nigra at Durrell Wildlife Conservation Trust with deslorelin acetate, to gain more information on methods to control aggression in this species.
The effect of a GnRH agonist on an aggressive adolescent male Macaca nigra: a case study
Stephanie Cox1, Sarah Fowkes2 & Eluned Price2
1Cardiff University, UK; 2Durrell Wildlife Conservation Trust
Abstract Adolescent male primates commonly exhibit aggressive behaviour, giving rise to captive management challenges. As a reversible alternative to surgical castration, an implant containing a gonadotropin-releasing hormone (GnRH) agonist (deslorelin) was tested in one adolescent male Sulawesi crested back macaque (Macaca nigra) at Durrell Wildlife Park. Play, aggression and copulation were recorded in all five males in the group for four weeks before the deslorelin implant was injected into the adolescent macaque and for 15 weeks during treatment. A strong inverse relationship between age and play behaviour was found, but no obvious relationship between age and aggression or copulation was observed. An initial increase in aggression was observed in the treated male after implantation, but overall there was no significant change in aggression. Copulation was observed frequently during treatment. These results suggest that the GnRH agonist had no effect on this individual, but further work on a larger sample is needed before this method of reducing aggression in adolescent male M. nigra can confidently be accepted or rejected.
Key words: aggression, captive management, copulation, play
Fig. 1. Members of the macaque group at Durrell Wildlife Park.
Research report
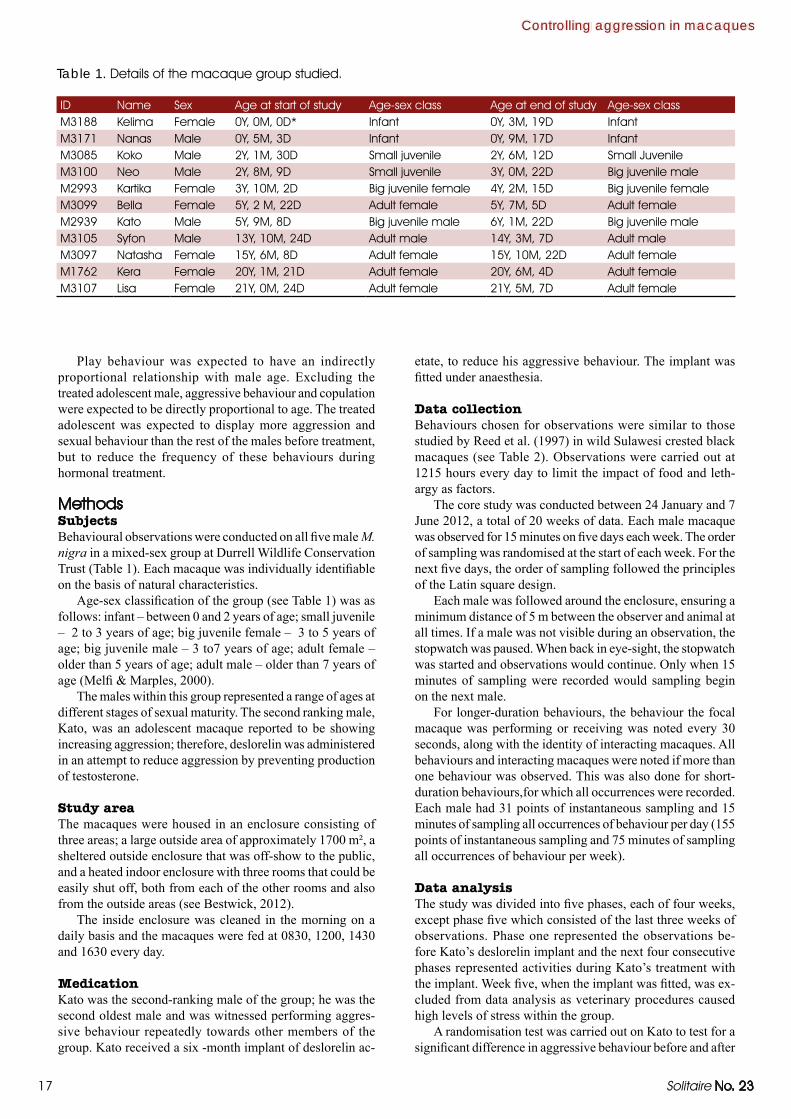
17 Solitaire No. 23
Play behaviour was expected to have an indirectly proportional relationship with male age. Excluding the treated adolescent male, aggressive behaviour and copulation were expected to be directly proportional to age. The treated adolescent was expected to display more aggression and sexual behaviour than the rest of the males before treatment, but to reduce the frequency of these behaviours during hormonal treatment.
MethodsSubjectsBehavioural observations were conducted on all five male M. nigra in a mixed-sex group at Durrell Wildlife Conservation Trust (Table 1). Each macaque was individually identifiable on the basis of natural characteristics.
Age-sex classification of the group (see Table 1) was as follows: infant – between 0 and 2 years of age; small juvenile – 2 to 3 years of age; big juvenile female – 3 to 5 years of age; big juvenile male – 3 to7 years of age; adult female – older than 5 years of age; adult male – older than 7 years of age (Melfi & Marples, 2000).
The males within this group represented a range of ages at different stages of sexual maturity. The second ranking male, Kato, was an adolescent macaque reported to be showing increasing aggression; therefore, deslorelin was administered in an attempt to reduce aggression by preventing production of testosterone.
Study areaThe macaques were housed in an enclosure consisting of three areas; a large outside area of approximately 1700 m², a sheltered outside enclosure that was off-show to the public, and a heated indoor enclosure with three rooms that could be easily shut off, both from each of the other rooms and also from the outside areas (see Bestwick, 2012).
The inside enclosure was cleaned in the morning on a daily basis and the macaques were fed at 0830, 1200, 1430 and 1630 every day.
MedicationKato was the second-ranking male of the group; he was the second oldest male and was witnessed performing aggres-sive behaviour repeatedly towards other members of the group. Kato received a six -month implant of deslorelin ac-
etate, to reduce his aggressive behaviour. The implant was fitted under anaesthesia.
Data collectionBehaviours chosen for observations were similar to those studied by Reed et al. (1997) in wild Sulawesi crested black macaques (see Table 2). Observations were carried out at 1215 hours every day to limit the impact of food and leth-argy as factors.
The core study was conducted between 24 January and 7 June 2012, a total of 20 weeks of data. Each male macaque was observed for 15 minutes on five days each week. The order of sampling was randomised at the start of each week. For the next five days, the order of sampling followed the principles of the Latin square design.
Each male was followed around the enclosure, ensuring a minimum distance of 5 m between the observer and animal at all times. If a male was not visible during an observation, the stopwatch was paused. When back in eye-sight, the stopwatch was started and observations would continue. Only when 15 minutes of sampling were recorded would sampling begin on the next male.
For longer-duration behaviours, the behaviour the focal macaque was performing or receiving was noted every 30 seconds, along with the identity of interacting macaques. All behaviours and interacting macaques were noted if more than one behaviour was observed. This was also done for short-duration behaviours,for which all occurrences were recorded. Each male had 31 points of instantaneous sampling and 15 minutes of sampling all occurrences of behaviour per day (155 points of instantaneous sampling and 75 minutes of sampling all occurrences of behaviour per week).
Data analysisThe study was divided into five phases, each of four weeks, except phase five which consisted of the last three weeks of observations. Phase one represented the observations be-fore Kato’s deslorelin implant and the next four consecutive phases represented activities during Kato’s treatment with the implant. Week five, when the implant was fitted, was ex-cluded from data analysis as veterinary procedures caused high levels of stress within the group.
A randomisation test was carried out on Kato to test for a significant difference in aggressive behaviour before and after
Table 1. Details of the macaque group studied.
Controlling aggression in macaques
ID Name Sex Age at start of study Age-sex class Age at end of study Age-sex classM3188 Kelima Female 0Y, 0M, 0D* Infant 0Y, 3M, 19D InfantM3171 Nanas Male 0Y, 5M, 3D Infant 0Y, 9M, 17D InfantM3085 Koko Male 2Y, 1M, 30D Small juvenile 2Y, 6M, 12D Small JuvenileM3100 Neo Male 2Y, 8M, 9D Small juvenile 3Y, 0M, 22D Big juvenile maleM2993 Kartika Female 3Y, 10M, 2D Big juvenile female 4Y, 2M, 15D Big juvenile femaleM3099 Bella Female 5Y, 2 M, 22D Adult female 5Y, 7M, 5D Adult femaleM2939 Kato Male 5Y, 9M, 8D Big juvenile male 6Y, 1M, 22D Big juvenile maleM3105 Syfon Male 13Y, 10M, 24D Adult male 14Y, 3M, 7D Adult maleM3097 Natasha Female 15Y, 6M, 8D Adult female 15Y, 10M, 22D Adult femaleM1762 Kera Female 20Y, 1M, 21D Adult female 20Y, 6M, 4D Adult femaleM3107 Lisa Female 21Y, 0M, 24D Adult female 21Y, 5M, 7D Adult female
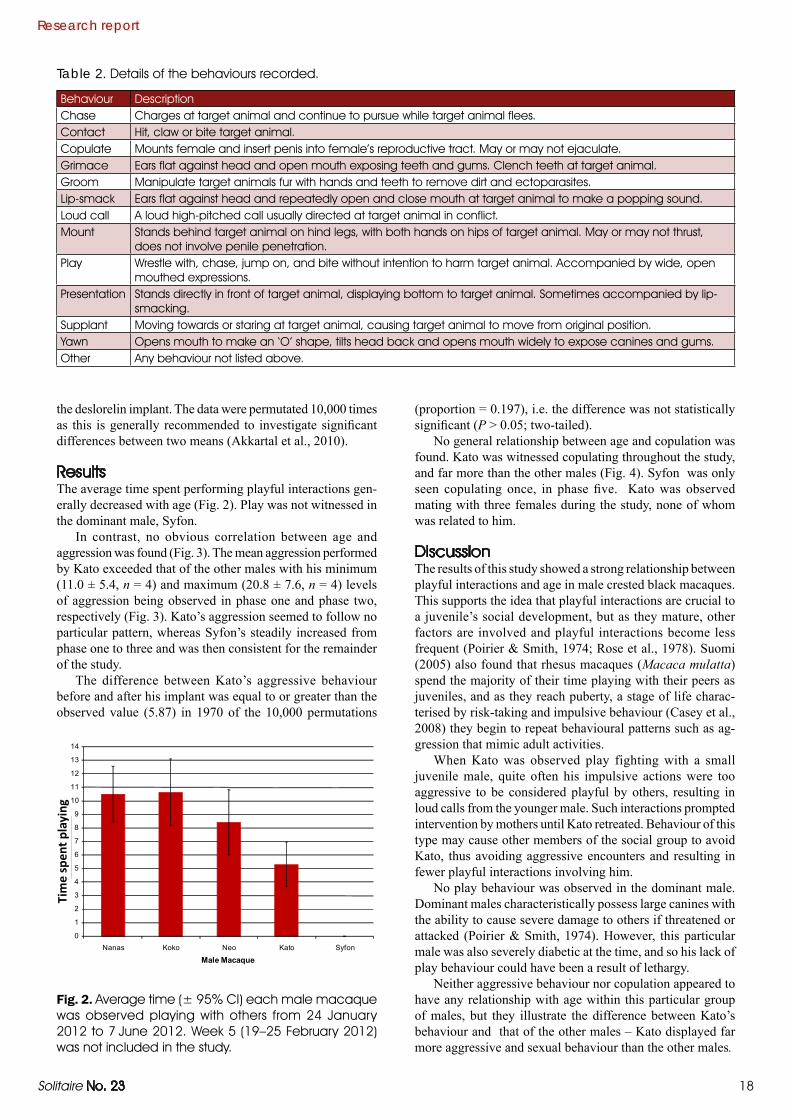
Solitaire No. 23 18
the deslorelin implant. The data were permutated 10,000 times as this is generally recommended to investigate significant differences between two means (Akkartal et al., 2010).
ResultsThe average time spent performing playful interactions gen-erally decreased with age (Fig. 2). Play was not witnessed in the dominant male, Syfon.
In contrast, no obvious correlation between age and aggression was found (Fig. 3). The mean aggression performed by Kato exceeded that of the other males with his minimum (11.0 ± 5.4, n = 4) and maximum (20.8 ± 7.6, n = 4) levels of aggression being observed in phase one and phase two, respectively (Fig. 3). Kato’s aggression seemed to follow no particular pattern, whereas Syfon’s steadily increased from phase one to three and was then consistent for the remainder of the study.
The difference between Kato’s aggressive behaviour before and after his implant was equal to or greater than the observed value (5.87) in 1970 of the 10,000 permutations
(proportion = 0.197), i.e. the difference was not statistically significant (P > 0.05; two-tailed).
No general relationship between age and copulation was found. Kato was witnessed copulating throughout the study, and far more than the other males (Fig. 4). Syfon was only seen copulating once, in phase five. Kato was observed mating with three females during the study, none of whom was related to him.
DiscussionThe results of this study showed a strong relationship between playful interactions and age in male crested black macaques. This supports the idea that playful interactions are crucial to a juvenile’s social development, but as they mature, other factors are involved and playful interactions become less frequent (Poirier & Smith, 1974; Rose et al., 1978). Suomi (2005) also found that rhesus macaques (Macaca mulatta) spend the majority of their time playing with their peers as juveniles, and as they reach puberty, a stage of life charac-terised by risk-taking and impulsive behaviour (Casey et al., 2008) they begin to repeat behavioural patterns such as ag-gression that mimic adult activities.
When Kato was observed play fighting with a small juvenile male, quite often his impulsive actions were too aggressive to be considered playful by others, resulting in loud calls from the younger male. Such interactions prompted intervention by mothers until Kato retreated. Behaviour of this type may cause other members of the social group to avoid Kato, thus avoiding aggressive encounters and resulting in fewer playful interactions involving him.
No play behaviour was observed in the dominant male. Dominant males characteristically possess large canines with the ability to cause severe damage to others if threatened or attacked (Poirier & Smith, 1974). However, this particular male was also severely diabetic at the time, and so his lack of play behaviour could have been a result of lethargy.
Neither aggressive behaviour nor copulation appeared to have any relationship with age within this particular group of males, but they illustrate the difference between Kato’s behaviour and that of the other males – Kato displayed far more aggressive and sexual behaviour than the other males.
0
1
2
3
4
5
6
7
8
9
10
11
12
13
14
Nanas Koko Neo Kato Syfon
Male Macaque
Time
playing
Tim
e sp
ent p
layi
ng
Fig. 2. Average time (± 95% CI) each male macaque was observed playing with others from 24 January 2012 to 7 June 2012. Week 5 (19–25 February 2012) was not included in the study.
Behaviour DescriptionChase Charges at target animal and continue to pursue while target animal flees.Contact Hit, claw or bite target animal.Copulate Mounts female and insert penis into female’s reproductive tract. May or may not ejaculate.Grimace Ears flat against head and open mouth exposing teeth and gums. Clench teeth at target animal.Groom Manipulate target animals fur with hands and teeth to remove dirt and ectoparasites.Lip-smack Ears flat against head and repeatedly open and close mouth at target animal to make a popping sound.Loud call A loud high-pitched call usually directed at target animal in conflict.Mount Stands behind target animal on hind legs, with both hands on hips of target animal. May or may not thrust,
does not involve penile penetration.Play Wrestle with, chase, jump on, and bite without intention to harm target animal. Accompanied by wide, open
mouthed expressions.Presentation Stands directly in front of target animal, displaying bottom to target animal. Sometimes accompanied by lip-
smacking.Supplant Moving towards or staring at target animal, causing target animal to move from original position.Yawn Opens mouth to make an ‘O’ shape, tilts head back and opens mouth widely to expose canines and gums.Other Any behaviour not listed above.
Table 2. Details of the behaviours recorded.
Research report
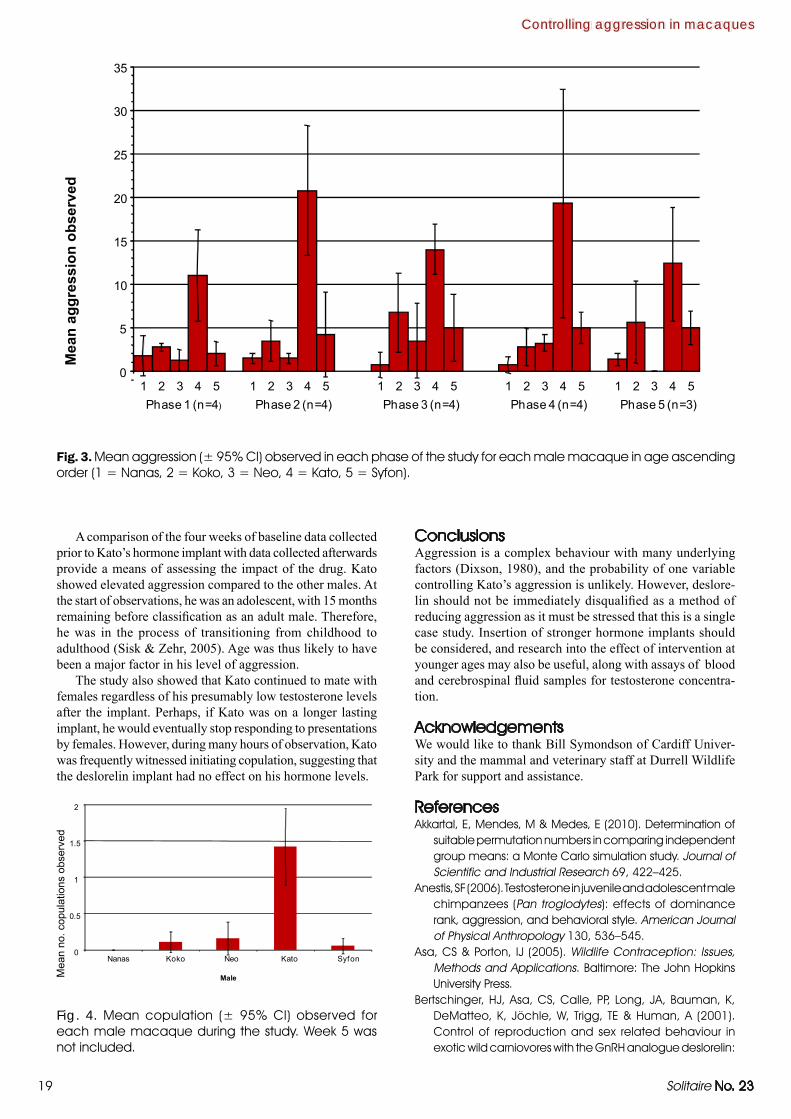
19 Solitaire No. 23
A comparison of the four weeks of baseline data collected prior to Kato’s hormone implant with data collected afterwards provide a means of assessing the impact of the drug. Kato showed elevated aggression compared to the other males. At the start of observations, he was an adolescent, with 15 months remaining before classification as an adult male. Therefore, he was in the process of transitioning from childhood to adulthood (Sisk & Zehr, 2005). Age was thus likely to have been a major factor in his level of aggression.
The study also showed that Kato continued to mate with females regardless of his presumably low testosterone levels after the implant. Perhaps, if Kato was on a longer lasting implant, he would eventually stop responding to presentations by females. However, during many hours of observation, Kato was frequently witnessed initiating copulation, suggesting that the deslorelin implant had no effect on his hormone levels.
ConclusionsAggression is a complex behaviour with many underlying factors (Dixson, 1980), and the probability of one variable controlling Kato’s aggression is unlikely. However, deslore-lin should not be immediately disqualified as a method of reducing aggression as it must be stressed that this is a single case study. Insertion of stronger hormone implants should be considered, and research into the effect of intervention at younger ages may also be useful, along with assays of blood and cerebrospinal fluid samples for testosterone concentra-tion.
AcknowledgementsWe would like to thank Bill Symondson of Cardiff Univer-sity and the mammal and veterinary staff at Durrell Wildlife Park for support and assistance.
ReferencesAkkartal, E, Mendes, M & Medes, E (2010). Determination of
suitable permutation numbers in comparing independent group means: a Monte Carlo simulation study. Journal of Scientific and Industrial Research 69, 422–425.
Anestis, SF (2006). Testosterone in juvenile and adolescent male chimpanzees (Pan troglodytes): effects of dominance rank, aggression, and behavioral style. American Journal of Physical Anthropology 130, 536–545.
Asa, CS & Porton, IJ (2005). Wildlife Contraception: Issues, Methods and Applications. Baltimore: The John Hopkins University Press.
Bertschinger, HJ, Asa, CS, Calle, PP, Long, JA, Bauman, K, DeMatteo, K, Jöchle, W, Trigg, TE & Human, A (2001). Control of reproduction and sex related behaviour in exotic wild carniovores with the GnRH analogue deslorelin:
Controlling aggression in macaques
0
5
10
15
20
25
30
35
1 2 3 4 5 1 2 3 4 5 1 2 3 4 5 1 2 3 4 5 1 2 3 4 5Phase 1 (n=4) Phase 2 (n=4) Phase 3 (n=4) Phase 4 (n=4) Phase 5 (n=3)
Mea
n ag
gres
sion
obs
erve
d
Fig. 3. Mean aggression (± 95% CI) observed in each phase of the study for each male macaque in age ascending order (1 = Nanas, 2 = Koko, 3 = Neo, 4 = Kato, 5 = Syfon).
0
0.5
1
1.5
2
Nanas Koko Neo Kato Syfon
MaleMea
n no
. cop
ulat
ions
obs
erve
d
Fig. 4. Mean copulation (± 95% CI) observed for each male macaque during the study. Week 5 was not included.

Solitaire No. 23 20
preliminary observations. Journal of Reproduction and Fertility Supplement 57, 275–283.
Bestwick, L (2012). The training and management of an insulin-dependent Sulawesi crested black macaque. Solitaire 23, 13–15.
Casey, BJ, Jones, RM & Hare, TA (2008). The adolescent brain. The Year in Cognitive Neuroscience 1124, 111–126.
De Nys, HM, Bertschinger, HJ, Turkstra, JA, Colenbrander, B, Palme, R & Human, AM (2010). Vaccination against GnRH may suppress aggressive behaviour and musth in African elephant (Loxodonta africana) bulls – a pilot study. Journal of the South African Veterinary Association 81, 8–15.
Dixson, AF (1980). Androgens and aggressive behavior in primates: A review. Aggressive Behavior 6, 37–67.
Melfi, VA & Marples, N (2000). How different captive environments affect the behaviour of Sulawesi crested black macaques (Macaca nigra). In Proceedings of the 2nd Annual Symposium on Zoo Research, Devon, UK, 6-7th July, 151–157. Plowman, AB (ed.). London: Federation of Zoological Gardens of Great Britain and Ireland.
Poirier, FE & Smith, EO (1974). Socializing functions of primate play. American Journal of Zoology 14, 275–287.
Reed, C, O’Brien, TG & Kinnaird, F (1997). Male social behavior and dominance hierarchy in the Sulawesi crested black macaque (Macaca nigra). International Journal of Primatology 18, 247–260.
Reinhart, CJ (2008). Cooperation and Competition during Play Fighting in Tonkean and Japanese Macaques: an Examination of Juvenile Behaviour within Egalitarian and Despotic Social Systems. PhD thesis, University of Lethbridge.
Richards, AB, Morris, RW, Ward, S, Schmitz, S, Rothmond, DA, Noble, PL, Woodward, RA, Winslow, JT & Weickert,
Stephanie Cox is an Ecology student at Cardiff University and undertook work experience at Durrell Wildlife Park in 2011–2012.
Sarah Fowkes is a Senior Mammal Keeper and Eluned Price the Wildlife Park Research Coordinator at Durrell.
E-mail: [email protected]
CS (2009). Gonadectomy negatively impacts social behavior of adolescent male primates. Hormones and Behavior 56, 140–148.
Rose, RM, Bernstein, IS, Gordon, TP & Lindsley, JG (1978). Changes in testosterone and behaviour during adolescence in the male rhesus monkey. Psychosomatic Medicine 40, 60–70.
Sisk, CL & Zehr, JL (2005). Pubertal hormones organize the adolescent brain and behaviour. Frontiers in Neuroendocrinology 26, 163–174.
Suomi, SJ (2005). Genetic and environmental factors influencing the expression of impulsive aggression and serotonergic functioning in rhesus monkeys. In Developmental Origins of Aggression, 63–82. Tremblay, RE, Hartup, WW & Archer, J (eds.). New York: Guilford.
Supriatna, J & Andayani, N (2008). Macaca nigra. In IUCN 2012. IUCN Red List of Threatened Species. Version 2012.1 [Online]. Available at: http://www.iucnredlist.org/details/12556/0 (accessed 10 September 2012).
Wagner, RA, Piché, CA, Jöchle, W & Oliver, JW (2005). Clinical and endocrine responses to treatment with deslorelin acetate implants in ferrets with adrenocortical disease. American Journal of Veterinary Research 66, 910–914.
Research report
Dead end or thoroughfare? Habitat corridors and endangered mammals in Sumatra
Wood-pulp plantations are growing dramatically in Indonesia and Malaysia. Each year, vast expanses of native forest are being cleared and fragmented for new plantations, especially in Borneo and Sumatra, potentially taking a serious toll on wildlife. In plantation landscapes in Sumatra, we have been studying whether and how narrow corridors of remnant native forest can facilitate the survival of larger mammals.
In fragmented landscapes, corridors may allow animals to move between surviving habitat patches and provide
additional habitat. When establishing their wood-pulp plantations, companies following the law and industry ‘best practice’ leave narrow strips of riparian forest along rivers. Are these corridors working? Will they help rare species such as tigers, elephants, tapirs and sun bears survive in plantation landscapes, or will they merely lure them from nearby protected areas, making them more vulnerable to hunting and other human persecution?
We recently completed fieldwork in Riau Province in central Sumatra, where forests are disappearing at an
Bets
y Ya
ap
Automatic camera photos of a sun bear, tiger and Malayan tapir.

21 Solitaire No. 23
Conservation report
William ‘Bill’ Laurance was one of the Trust’s first trainees in 1979. Today he is Distinguished Research Professor at James Cook University in Cairns, Australia. He recently won the Heineken Environment Prize for his long-term efforts to study and conserve tropical forests and biodiversity.
E-mail: [email protected]
Betsy Yaap is a PhD student under Bill’s supervision. She has lived in Indonesia for over seven years, work-ing with NGOs, government and the private sector on wildlife conservation issues.
E-mail: [email protected]
Betsy Yaap working with plantation workers in Sumatra.
Will
iam
Lau
ranc
e
Sumatran tigers are quite common near Tesso Nilo National Park.
Will
iam
Lau
ranc
e
alarming rate. We used automatic cameras to census mammal populations across a series of riparian ‘corridors’ (linear forest remnants) near Tesso Nilo National Park. The vanishing forests in this area are prime habitat for wildlife, including the critically endangered Sumatran tiger - of which fewer than 400 animals are thought to survive today.
The corr idors we studied were generally less than 200 metres in width. In many cases these remnants are actually ‘peninsulas’ rather than ‘corridors’ linking two separate forest blocks.
Very little is known about the degree to which such remnants are used by vulnerable wildlife, and
there are no guidelines for designing such linear remnants to increase their suitability for disturbance- and hunting-sensitive species.
We hope our work will influence the rapidly growing paper-pulp industries in Indonesia and Malaysia. Large expanses of forests
in the countries have been slated for industrial plantations, and our goal is
to minimize their impacts on imperiled wildlife.
With our fieldwork in Sumatra now completed, we are planning to conduct a similar study in the Osa Peninsula in Costa Rica. There, the landscape is different, with different land uses, but the fundamental question remains the same: will linear forest remnants help provide corridors for movement of jaguars and other forest-dependent wildlife species?
A riparian ‘corridor’ retained in an acacia plantation.
‘Will corridors help
rare species such
as Sumatran tigers,
or make them more
vulnerable to human
persecution?’
Will
iam
Lau
ranc
e

Solitaire No. 23 22
The first public zoo was established in India at Madras in 1855. Spurred on by the establishment of zoos by the British, the ruling kings of various Indian princely states set up zoos too. By the time India gained independence, there were 18 zoos in the country. As elsewhere around the globe, the objective of zoos in the pre-independence era was displaying wild animals in park-like settings and satisfying public curiosity by exhibiting as many species representing different continents as possible. As wildlife was plentiful and no restrictions were in place for the acquisition of wild animals from the natural ecosystems, the longevity and health of the animals displayed by zoos was not an area of concern.
Keeping up with global trends, soon after India became a Democratic Republic, the Government of India started making efforts to enhance the role played by zoos from mere entertainment to conservation. The Indian Board for Wildlife resolved in 1952 that the Central Government should set up modern zoos in the country’s major cities to eduate the people at large about the rich fauna of the country and the need to conserve it.
Construction work on Delhi Zoo was started in 1955. The government wanted this zoo to be really world class and therefore hired the services of the world’s most renowned zoo designer of that era, Carl Hagenbeck, owner of Hamburg Zoo, to prepare the master plan for Delhi Zoo. The services of the Director of Colombo Zoo were also obtained to assist Government officials in the construction phase of the zoo. The zoo was completed by 1959, when it was opened to the public. Further large, well planned zoos were set up by several State Governments soon afterwards.
The establishment of these zoos should have led to the beginning of a new era in the history of the development of zoos. However, the traditional perception of zoos as mere centres of recreation continued to prevail and State Governments and various other organizations continued to set up new menageries and deer parks, caring little for providing the financial and technical inputs required for management of these new, big, wild animal display facilities.
The Central Government, worried about the disturbing trend towards the unplanned establishment of new zoos, appointed a committee of experts in 1973 to study various zoos in the country and formulate norms on the basis of which zoos could be classified. The main aspects to be considered were the overall area of each zoo, the space available for animals in individual enclosures and the facilities available for feeding the animals. The Committee was also required to suggest measures for improving the efficiency of various zoo management operations. The Committee visited some of the important zoos in the country and held detailed discussions with forest officials, zoo officials and animal traders, to identify the constraints in the field of zoo management and to design a strategy for adressing these so that planned development of zoos could take place. The Committee’s report was accepted
by the Government of India and circulated to all concerned for implementation. The report not only highlighted the major problems that were plaguing the management of zoos in the country but also made very pertinent recommendations for improvements, some of which remain relevant even today, four decades later.
The report considered an area of less than 50 hectares to be inadequate to meet the requirements of zoo animals in accordance with modern concepts of zoo management. It also highlighted the need to give zoo animals sufficient space for movement and exercise and to meet their biological requirements. The report went further and suggested that the soil, water and vegetation in animal enclosures should be manipulated so as to make the animals at home. These recommendations, in conjunction with those that every species should be given an environment as close as possible to its natural habitat, resulted in quite comprehensive guidelines
about the designing of animal enclosures, as well as for feeding, upkeep and hygiene.
The report also highlighted the importance of breeding endangered species and recommended that enclosures should
be sufficiently large to give the fullest opportunity for breeding rituals, should
provide hiding places and privacy for the animals, and should simulate the natural habitat of the animal
housed as closely as possible.
Unplanned and substandard zoos mushroom
It is quite sad that the invaluable recommendations of the committee have been totally overlooked by the Government and zoo operators alike for decades, and the mushrooming of ill planned and inappropriately designed zoos continued for decades, making ‘zoo’ a derogatory word in the contempo-rary world. If the zoos have any respectability in this country today the credit goes to a few enthusiastic people in Govern-ment who ensured that modern zoos with adequate space and appropriately designed enclosures were established during
Zoos in India: from menageries to naturalistic enclosures
‘Ill planned and
inappropriately designed
zoos have made “zoo” a
derogatory word.’
Conservation report
Indira Gandhi Zoological Park, Vishakhapatnam
Total area provided in the paddock
Depth of the enclosure
No of feeding retiring cells available
3849 Sqmt 15 Mt Five
Sloth bear exhibit at Indira Gandhi Zoo.

23 Solitaire No. 23
the 1970s and 1980s. Examples include Indira Gandhi Zoo-logical Park, Visakhapatnam Zoo, Kanpur Zoo, Mahendra Chaudhary Zoological Park, Sanjay Gandhi Zoological Park, Arignar Anna Zoological Park, and Sri Venkateswara Zoo-logical Park.
These zoos had excellent layout plans and very well designed enclosures for various species of wild animals, and knowledgeable visitors were extremely pleased by them. However, State Governments and other zoo operators did not appear to be inspired to establish such modern zoos themselves, and their determination to have cramped menageries in every picturesque location or important town scaled new heights. The advent of the World Bank-funded Social Forestry Project and growing industrialisation gave a further boost to the tendency to set up new mini zoos and deer parks. Paucity of budgetary provisions became a chronic problem plaguing zoos all over the country. Even large and well managed zoos became victims of poor maintenance and sub-standard upkeep and healthcare of their animals.
The Central Government did extend a helping hand to State Governments to mitigate the deterioration in standards of zoo management and enhance their conservation role by launching new centrally sponsored schemes. However, as the funding under the schemes was on a 50:50 basis, states showed hardly any interest, and only the Chief Wildlife Wardens of
a few states took advantage of the schemes to improve the zoos under their charge.
Setting up of Central Zoo AuthorityBy the beginning of the 1990s it had become abundantly clear that no qualitative improvement of zoos in the country could be brought about without mandatory standards and norms
for regulating their management. The Central Government, therefore, through the amendment of the Wild Life (Pro-tection) Act in 1991, made a provision
that no zoo in the country could function without obtaining the prior recognition of the Central Zoo Authority. It was also provided in the Act that recognition would
be granted to zoos having due regard to the conservation of wildlife.
The Central Zoo Authority has the following functions:
to specify the minimum standards for housing, upkeep • and veterinary care of the animals kept in a zooto evaluate and assess the functioning of zoos with • respect to standards or norms prescribedto recognise or derecognise zoos• to identify endangered species of wild animals that • r equire captive breeding and assign responsibility for this to a zooto encourage zoos to exchange, loan, pool the animals •
Phayre’s leaf monkey.
Indian zoos
‘Zoos today must justify
their existence not by
the quantity of their
exhibits but by their
quality.’

Solitaire No. 23 24
available in the best interests of conservation to coordinate the acquisition, exchange and loan of • animals for breeding purposesto ensure proper genetic management and maintenance • of studbooks for endangered species bred in captivityto encourage and coordinate research in captive • breeding and educational programmesto provide technical and other assistance to zoos • for their proper management and development on scientific linesto upgrade the technical skills of zoo personnel • through training programmes and workshopsto help zoos identify priorities and themes for the • display of wild animals with the aim of enhancing the conservation value of zoos.
Exhibits for species survivalToday it has become mandatory for zoos to justify their ex-istence not by the quantity of exhibits but by their quality. Examples of ‘immersion’ exhibits in Indian zoos include:
Sepahijala Zoological Park, TripuraSepahijala Zoological Park in Tripura has provided immer-sion exhibits for many species. The zoo has access to areas of large trees which provide appropriate and adequate areas for animals to perform their natural behaviour.
The pig-tailed macaque enclosure is an example of this, the animals appearing to viewers to be living in wild habitat. Wild pig-tailed macaques do in fact live in areas adjoining the zoo.
Phayre’s leaf monkey is one of the rarest primates in India. Sepahijala Zoo has been selected as coordinating zoo for
Pig-tailed macaque at Sepahijala Zoo
Conservation report
Indira Gandhi Zoological Park, Vishakhapatnam
Total area provided in the paddock
Depth of the enclosure
No of feeding retiring cells available
3849 Sqmt 15 Mt Five
Nandankanan Zoo, Bhubaneshwar
Total area provided in the paddock
Area of the water body
Depth of the water body
3055 Sqmt 1040 sq mts. 1-12 mts
Sloth bear enclosure at Indira Gandhi Zoo. Gharial at Nandankanan Biological Park.

25 Solitaire No. 23
the species, and has good breeding success. The exhibit for this species provides opportunities for the monkeys to jump and climb from one tree to other, along with natural browse, protection during rains and places to hide if and when the dominant male chases other animals in the group.
Indira Gandhi Zoological Park, Vishakhap-atnamIndira Gandhi Zoological Park is one of the finest zoos in the country. Most of the zoo exhibits are based on the concept of ‘borrowing the landscape’.
The tiger exhibit provides natural landscaping with tall trees and thick bamboo groves, while the sloth bear enclosure has natural vegetation and rocks. The enclosure also provides ample opportunities for the bears to dig the soil, and large dead tree trunks to climb upon.
Nandankanan Biological Park, OdishaThe hippo enclosure at this zoo is world class, and is the largest exhib-it for this species anywhere. Visitors can see eight to ten hipps, and as the water body is attached to an ad-jacent lake, all waste flows from the exhibit to lake area. It has a huge sand bank and large trees to provide shade for the animals.
Brij Kishor Gupta graduated from the Durrell Conser-vation Academy in 1992 and has attended several other training courses. He works for India’s Central Zoo Authority.
E-mail: [email protected] enclosure at Indira Gandhi Zoo.
Indian zoos
The zoo is also known for breeding gharial and crocodile. The exhibit gives the impression of a vew of the animals in a large river.
The water is always clean, as it has a natural filtration system. The sandy banks
provide the animals with areas on which to to bask and lay eggs. The
green cover surrounding the exhibit provides a good buffer area.
ConclusionIn the 21st century, conservation efforts will need to preserve habitats with varying degrees of fragmentation and wildlife conservationists will need to be creative with broken sys-tems. Species survival will emerge as a clear separate agenda from habitat preservation, a global experiment in biospheric rescue where the lines between nature and captivity will be blurred.
Today, when wildlife habitats are under severe pressure and a large number of species of wild fauna have become endangered, zoos have not only to sustain their own populations but also augment the depleting populations of endangered species in the wild. This new role for the zoo has been acknowledged by the global conservation community.
The conservation breeding of identified endangered species has just begun, but exhibits for them need to be planned and designed in such a way that they meet the biological requirement of the animals housed there. The objective of keeping animals merely for entertainment is no longer the sole raison d’etre for Indian zoos, and it is high time for a more holistic development of zoos in the country to achieve the objective of making zoos into centres that can complement national effort for wildlife conservation in India. We need to infuse a more technical and scientific culture into the operation of our zoos and to change the general perception of zoos as mere picnic spots into one of scientific institutions.
‘We need to infuse a more
technical and scientific
culture into the operation of
our zoos.’
Nandankanan Zoo, Bhubaneshwar
Total area provided in the paddock
Feeding cubicle Area of krral
6240 Sqmt 77.4 sq mts. 801.3 sq mts.
Hippo exhibit at Nandankan Biological Park.

Solitaire No. 23 26
Durrell Conservation Award 2012: the winners are... Once again, the annual Durrell Conservation Awards have distributed around £3000 to successful applicants from three categories: Conservation Projects, Equipment, and Capacity Building. Many thanks to everyone who applied.
Academy news
The DCA Conservation Project grant was awarded to Camila Nali (DESMAN 2011) of Instituto de Pesquisas Ecológicas (IPE) towards a project to reconnect the now separate mainland and island populations of the critically endangered black-faced lion tamarin Leontopithecus caissara in the southeastern Atlantic rainforest of Brazil. The genetic isolation addressed by this project has been identified as a major threat to the survival of this species. This experimental metapopulation management project is part of IPE’s long-term programme to recover this species.
The DCA Equipment grant was awarded to Rosanna Carreras (ISLA 2011) of Grupo Jaragua towards field equipment costs for a study of the critically endangered Ricord’s iguana Cyclura ricordii in the Dominican Republic. As part of the study, Rosanna will be looking at genetic variability and relatedness among populations. This is just one component of Rosanna’s and Grupo Jaragua’s conservation work with this species.
The DCA Capacity Building grant was awarded to Marc Jean-Baptiste (DESMAN 2012) of Seychelles Islands Foundation towards the costs of participating in a three-week reptile survey on the island of Rodrigues with herpetologist Dr Chris Raxworthy. Through this, Marc will learn many valuable skills that he will then be able to apply to his work back in the Seychelles. The selection panel particularly liked the way in which Marc intends to transfer skills from one field project to another.
Durrell Conservation Learning Network
Calling all Durrell training graduates...
Please register with the Durrell Conservation Learning Network (DCLN).
By doing so, you will be able to:
get in touch with other graduates•search for graduates in particular countries•access useful information and post-course •materialbe eligible to apply for our annual conservation •grants.
Please sign up now – it doesn’t take long.
Visit www.durrell.org/graduate-registration
Once registered, you can access the DCLN at www.durrell.org/network
Please also join our active Facebook group by searching for ‘Durrell Graduate Network’ and clicking on the ‘Join’ button.

27 Solitaire No. 23
Academy news
Making a difference Eunice Ngwing Halle from Cameroon was a participant on the In-troduction to GIS for Conservation Managers course in 2012. She has sent us this message describing how the course has helped her in her conservation career.
‘The opportunity to attend this prestigious course organised by Durrell has been a turning point in my career. I was given a grant to attend the course by the Russell E. Train Education for Nature (EFN) Program, World Wildlife Fund WWF-USA. I learnt how to use GIS, and gained an understanding of how GIS data could be used to underpin conservation action.
One of the most enjoyable aspects of the course was the relaxed learning atmosphere, and I really felt very at home. I am gradually transferring the knowledge and ideas gained to my colleagues and to Masters students, and I will recommend the course to all aspiring conservationists as a stepping stone.’
Courses at Durrell Conservation Academy
Training courses available at Durrell Wildlife Conservation Trust include:
Conser vat ion pro ject management and •leadership
Conservation husbandry - primates, birds and •amphibians
Managing infectious disease in conservation •projects
Conservation education and interpretation •
Facilitation and communication skills•
Invasive species management•
GIS for conservation•
Endangered species recovery•
For full details of all our training, download our 2013 Training Prospectus from www.durrell.org/academy
Durrell Conservation Academy has 35 years experience of training the world’s conservationists. Drawing on Durrell Wildlife Conservation Trust’s more than 50 years of conservation experience, we have trained over 3300 people from 135 countries – over 350 during 2011 alone.
Over 40 experienced lecturers from some of the world’s leading conservation organisations help us provide unique conservation training that combines both theory and practice.

Congratulations Vikash!
Congratulations to Durrell training graduate Dr Vikash Tatayah on his recent promotion to Conservation Director within the Mauritian Wildlife Foundation (MWF).
Vikash, who attended the DESMAN 2000 course in Jersey, is responsible for overseeing MWF’s internationally renowned conservation and species recovery programmes for threatened fauna and flora on Mauritius and its offshore islands.
Recalling his time at Durrell, Vikash commented: ‘Training at Durrell has been a turning point in my career, so it means a lot to me.’
Vikash on Round Island
Contact us:Durrell Conservation Academy,Durrell Wildlife Conservation Trust,Trinity, Jersey, Channel Islands, JE3 5BP
E-mail: [email protected]: +44 (0) 1534 860037Fax: +44 (0) 1534 860002www.durrell.org
Solitaire is edited by Eluned PriceE-mail: [email protected]





























