Embed Size (px)
Citation preview

doi.org/10.26434/chemrxiv.11894904.v2
Triptolide Self-Assembling Nanoparticles Engineering with ModifiedErythrocyte Membranes for Targeting and Remodeling InflammatoryMicroenvironment in ArthritisJing Li, Sanpeng Li, Chunbin Li, Hongfeng Li, Chuangjun Liu, qi zhao, Pengfei Zhang, Ping Gong, Lintao Cai
Submitted date: 04/08/2020 • Posted date: 04/08/2020Licence: CC BY-NC-ND 4.0Citation information: Li, Jing; Li, Sanpeng; Li, Chunbin; Li, Hongfeng; Liu, Chuangjun; zhao, qi; et al. (2020):Triptolide Self-Assembling Nanoparticles Engineering with Modified Erythrocyte Membranes for Targetingand Remodeling Inflammatory Microenvironment in Arthritis. ChemRxiv. Preprint.https://doi.org/10.26434/chemrxiv.11894904.v2
Rheumatoid arthritis (RA) is an autoimmune disease causing severe joint damage, disability and decreasedquality of life. Pathologically, numerous blood-derived cells infiltrating in synovium and cytokine secretnecessitating formation of new blood vessels to generate pannus together form an inflammatorymicroenvironment. Triptolide with immunosuppressive activities is a potential drug to treat RA. However, it isstill lack of an effective targeting system to deliver triptolide to RA site safely. Herein an inflammatorymicroenvironment targeting and remodeling nanoplatform is developed to achieve significantly effective RAtreatment. In this system we synthesized a self-assembling triptolide nanoparticles (TPNs) mediated bydipeptide diphenylalanine which is the simplest self-assembly building block, then TPNs were entrapped bymannose-modified erythrocyte membranes to form engineering manRTPNs. For targeting, the immunologicalmolecule of erythrocytes was firstly introduced to target T cells by ligand binding of LFA-3/LFA-2, and thecoated mannose modified erythrocyte membrane also conferred the capacity of targeting to macrophages bymannose and its receptor CD206; for remodeling inflammatory microenvironment, TPNs could selectivelyexert its suppressive effects on different cells of RA including lymphocytes and synovial fibroblasts. Incollageninduced arthritis mice, manRTPNs showed excellent targeting effect and prolonged accumulation at inflamedjoint. After manRTPNs treatment, swollen paws of CIA considerably shrunk to normal, boss loss evenrecovered healthy level and cartilage preserved at synovium cavity, because of systemically conventionalcytokine reduction and expression shift of core genes in networks of RA microenvironment. Therefore, thiswell-defined manRTPNs might be a well promising systematic therapeutic agent for RA.
File list (2)
download fileview on ChemRxivmanuscript-update 20200804.docx (7.30 MiB)

download fileview on ChemRxivmanuscript-update 20200804.pdf (2.67 MiB)

Triptolide self-assembling nanoparticles engineering with
modified erythrocyte membranes for targeting and remodeling
inflammatory microenvironment in arthritis
Jing Li1, Sanpeng Li1, Chunbin Li1, Hongfeng Li1, Chuangjun Liu1, Qi Zhao3, Pengfei
Zhang1*, Ping Gong1,2*, Lintao Cai1*
1Guangdong Key Laboratory of Nanomedicine, CAS-HK Joint Lab for Biomaterials,
Shenzhen Institutes of Advanced Technology, Chinese Academy of Sciences, Shenzhen
518055, China
2Guangdong Key Laboratory for Research and Development of Natural Drugs,
Guangdong Medical University, Dongguan, 523808, China
3 Faculty of Health Sciences, University of Macau, Taipa, Macau, China.
*Corresponding author: Lintao Cai: E-mail: [email protected];
Ping Gong: E-mail: [email protected]
1
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
1

Abstract
Overview: Rheumatoid arthritis (RA) is an autoimmune disease characterized by persistent
synovitis, systemic inflammation and causing severe joint damage. Inflammation and influx of
immune cells is a hallmark of the disease. Triptolide with great immunosuppressive activities
is a potential drug to treat RA. However, its severe toxicity is still an unresolved bottleneck
and largely limits its clinical use. Recently, nanoparticle (NP)-based drug delivery systems
have been developed for detoxification of toxic drugs and diseases management.
Aim: To synthesize triptolide nanoparticles to decline its toxicity and improve water-solubility
and develop a dual targeted biomimicking delivery system which targets to inflammation
microenvironment. Finally, to achieve excellent therapeutic effect in rheumatoid arthritis.
Methods: We synthesized an amphiphilic molecule prodrug with the addition of
diphenylalanine peptide (FF) to triptolide. These prodrugs were self-assembled into
nanoparticles (TPNs) in water, then TPNs were coated with mannose-modified erythrocyte
membranes to form a dual targeting manRTPNs. Specifically, mannose and natural molecule
CD58 on RBC membrane guide it towards activated macrophages and T cells in inflamed
joint through innate and specific immune recognition, respectively. Different cell lines
including RAW264.7, CTLL-2, RSC364 and hFLS-RA and a collagen induced arthritis (CIA)
mice model were used to detect the targeting and therapeutic effect in vitro and vivo.
2
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
2

Results: manRTPNs showed an excellent dual targeting ability to activated macrophages and
T cells in vitro. We found that manRTPNs selectively induced anomalous activated cells death
and indeed altered genes expression and cytokines production of inflamed cells such as
macrophages, T cells and synoviocytes. In CIA mice, manRTPNs exhibited significantly
targeting effect and prolonged drug accumulation at inflamed joints, whereby increased drug
achieved significant therapeutic effect without adverse effect. In detail, manRTPNs
ameliorated joint destruction, swollen paws considerably shrunk to normal, bone and cartilage
destruction were suppressed and bone density was even recovered to healthy level. Basically,
several core genes like NF-κB, cox-2, rank, rankl, vegf, vegfr2, mmps, and cytokines such as
IL-6, TNF-α involved in inflammation, cartilage/bone destruction and angiogenesis were
regulated and altered expression and production. Thus, the inflammation microenvironment
was remodeled by TPNs.
Conclusion: This study demonstrated phenylalanine dipeptide mediating triptolide self-
assembled nanoparticles to solve current problems and the potential of engineered RBC
membranes for inflammation-targeted drug delivery. Therefore, our work is a good example
of triptolide reconstruction, and this agent based on immune recognition may provide an
immune cell targeted strategy for rheumatoid arthritis therapy.
Keywords: triptolide, nanodrug, dual targeting, inflammatory microenvironment, rheumatoid
arthritis
3
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
3
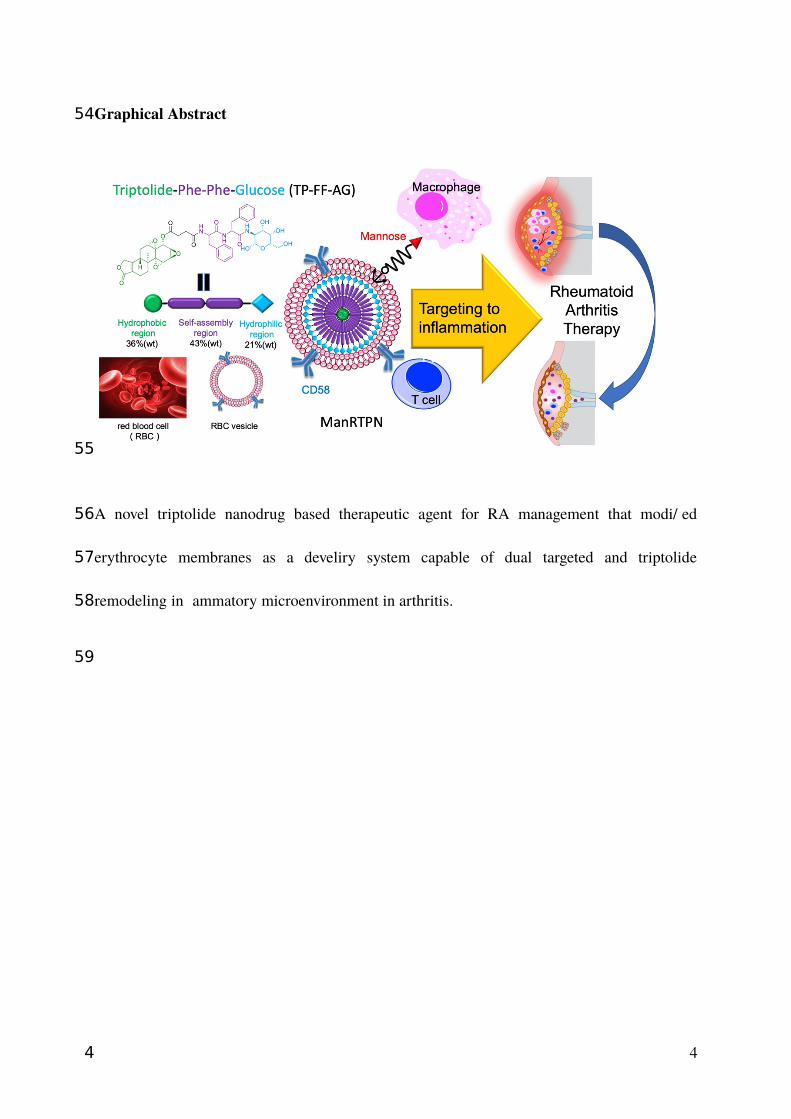
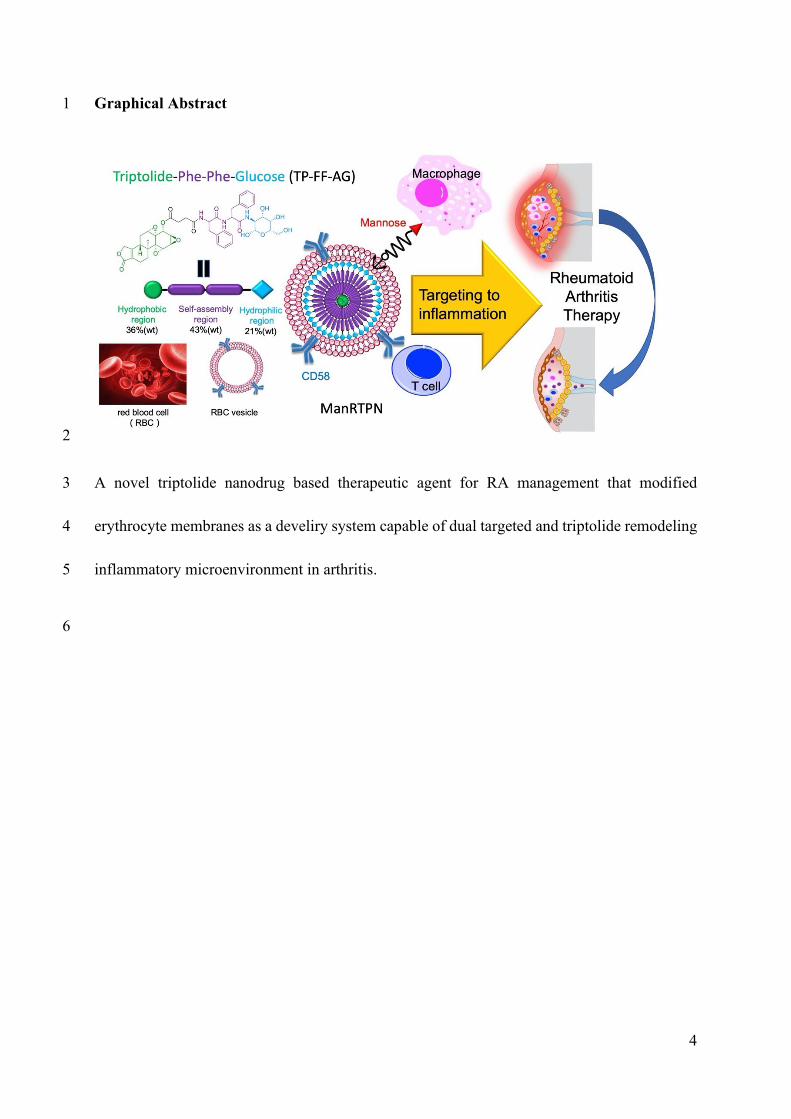
Graphical Abstract
A novel triptolide nanodrug based therapeutic agent for RA management that modified
erythrocyte membranes as a develiry system capable of dual targeted and triptolide
remodeling inflammatory microenvironment in arthritis.
4
54
55
56
57
58
59
4

Introduction
Rheumatoid arthritis (RA) is a serious long-term disease characterized by persistent
synovitis, systemic inflammation and autoantibodies and causing severe joint damage,
disability and decreased quality of life. The etiopathogenesis remains elusive, and viable and
effective treatment options are still limited[1]. Over the last two decades, long-term
management of RA has involved disease-modifying anti-rheumatic drugs (DMARDs),
biological agents such as tumor necrosis factor (TNF) inhibitors, and glucocorticoids[2].
Although the highest clinical remission rate achieved within 50% of control, not all patients
attain desirable levels of clinical remission[3] and clinical use of these therapies is limited
because of their high cost and frequency of adverse effects. The latter includes teratogenicity
and hepatotoxicity of DMARDs like methotrexate[4, 5], risks of infections such as
tuberculosis and osteoporosis caused by biological agents[6, 7] and long-term adverse
reactions to glucocorticoid therapy[8, 9]. Lately, new therapies based on inflammatory cells
such as macrophages and neutrophil or their products such as cell-free DNA in inflamed joint
have been reported[10-12], however inhibiting a single factor may not be enough to
sustainably halt or reverse disease progression due to RA’s complexity and heterogeneity.
Therefore, a new systemic design for drug candidate based on nano-medicine is urgently
needed.
Triptolide is a trace natural triepoxide diterpene from Tripterygium wilfordii (or Léi
Gōng Téng) and has been widely used to treat for RA patients since the 1970s in China[13].
Despite its various biological activities, low water solubility and severe toxicity, especially to
5
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
5

the liver and kidney, remarkably impedes the translation to clinical applications[14].
Considerable efforts in structural modification[15-18] and nanoparticle(NP) -based drug
delivery system [19-21] have been applied to reduce its toxicity. Unfortunately, common
practices that direct conjugations attached to the C14-hydroxyl group of triptolide via a
hydrolytic ester bond have proven biologically unstable; meanwhile, current targeted drug-
loading delivery systems are limited by drug loading or encapsulation ratio and they mostly
target to kidney or tumor[22]. Therefore, an effective and stable system that can deliver
triptolide to other organ or tissue such as joint is necessary. But until now, triptolide-formed
nanoparticles have been rarely reported before. From this point, inspired from
biomacromolecule self-assembly process, various peptide building blocks have been
performed for the creation of biomimetic or bioinspired nanostructured materials[23, 24].
Recently, diphenylalanine (FF) is the simplest peptide building block for self-assembly and
exhibits remarkable advantages including biocompatibility and non-immunogenicity and so
on. Many researchers have developed FF-based nanomaterials such as nanotubes, spherical
vesicles, nanofibrils and nanowires[25]. However, it is barely to link it with toxic drug for
nanodrug formation. Therefore, diphenylalanine (Phe-Phe, FF) mediating self-assembling
nanoparticles is a new strategy to reconstruct triptolide to be nanoparticles whose spherical
nanostructure was more stable and biocompatible than before.
In RA pathogenesis, numerous blood-derived cells infiltrate in synovium and cytokine
secret necessitates formation of new blood vessels to generate pannus, that all of these form an
inflammatory microenvironment[26]. Peripheral-blood monocytes are recruited to joint, and
functionally diverse macrophages distribute in the synovial sublining and lining layers. It is6
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
6

mentioned that Cluster of Differentiation 206 (CD206) on macrophages surface has been
induced to express more [27]. Besides, prominent T cells being activated, especially the CD4+
subset, also infiltrate the synovium. As reported, certain signal proteins on the T cell surface
such as lymphocyte function antigen 2 (LFA-2/CD2) have increased in both RA patients and
animal models[28, 29]. It was reported that CD2 can be effectively targeted by peptides
derived from LFA-3/CD58 as well as fusion protein constructs. Interestingly, the ligand of
CD2 namely CD58 widely distributes on erythrocyte membranes and CD58-CD2 interaction
have mediated process of T cells activation[30]. These immunological characteristics of
erythrocytes scarcely ever attracts attention in application, so it is a valuable attempt to apply
in drug targeting. Then, mannose, the ligand of CD206 was introduced in erythrocyte
membrane for ligand-binding targeting. In addition, the long circulation and flexible
functionalization of erythrocyte membrane also make it be a promising platform and have
wide application[31, 32]. In this study, we prepared mannose-labeled RBC membrane to
entrap newly synthesized triptolide nanoparticles (TPNs) for elevating its vast retention in
inflammation and achieving remarkably anti-inflammatory therapy. Triptolide, a hydrophobic
drug, was linked with diphenylalanine peptide to self-assembly into size-controlled
nanoparticles (TPNs) in water, and this reconstruction has successfully decreased its toxicity
and improved water-solubility. As shown in scheme 1, the nanodrug core was coated with
decorated RBC membranes to obtain mannose-modified RBC cell membrane-coated triptolide
nanoparticles (manRTPNs). By translocating RBC membranes to the nanoparticles, the
biological targeting properties of RBC cell can be conferred to manRTPNs as well, allowing
them to target activated T cells through the CD58-CD2 affinity. Further, the binding
7
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
7

interaction between the mannose and its receptor CD206 enhanced anchoring of manRTPNs
to activated macrophages. According to manRTPNs, the dual-targeting strategy facilitated the
accumulation of nanodrug in inflammation, and the accumulated TPNs efficiently achieved
systemic administration of rheumatoid arthritis by specifically inducing cell death and
remodeling the inflammatory microenvironment through affecting genes network,
downregulating cytokines and pro-angiogenic factors, regulating osteoclastogenesis, and
returning balance of metalloproteinases and their inhibitors. Overall, our study expanded the
RBC cell membrane biological application in nanocarrier for targeting inflammation through
innate and specific immune recognition, and the triptolide nanodrug showed high drug
content, low side effect, good biocompatibility which showed great potential for further
inflammation diseases therapy.
Results and Discussion
Preparation and characterization of manRTPNs
Triptolide is a diterpene triepoxide with several active sites that can be modified,
especially C14 position[33]. Because of its hydrophobicity, triptolide molecule acts as self-
assembly inducer with the addition of diphenylalanine peptide (FF), which is the key moiety
of the self-assembling peptide-drug conjugate. Initially, we added a glucosamine (AG) to the
FF terminal to lengthen the hydrophilic chain, and then linked triptolide to FF-AG by an ester
bond whose cleavage led to drug release in the cell; thus, the self-assembling monomer (TP-
FF-AG) as a triptolide prodrug was synthesized (Figure 1A). The chemical structures of these
compounds were confirmed via mass spectrometry (MS) and nuclear magnetic resonance
8
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
8

(NMR; Figure S1-S16). Due to hydrophobic interactions, triptolide nanoparticles (TPNs)
were formed via self-assembly of synthesized monomer namely TP-FF-AG, which is
uniformly easy to control TPNs size in water solution. The particles morphology was
observed via transmission electron microscopy (TEM; Figure 1B). To synthesize manRTPNs,
RBC membranes were extracted by hypotonic hemolysis[34], and mannose inserted by
PEG2000-DSPE. An extruder was using to camouflage mannose-modified erythrocyte
membranes onto TPNs then form a core-shell structure (Figure 1B). Dynamic light scattering
(DLS) measurements revealed that the TPNs were 135 nm in hydrodynamic diameter and
manRTPNs were about 10 nm larger with PDIs of 0.15-0.2. The surface zeta potential of the
shell was not as negative as that of the cores but was comparable to that of erythrocyte
membrane-derived vesicles (RBC vesicles) (Figure 1C). All these results demonstrated
successful cloaking of TPNs with erythrocyte membranes.
Next, TPNs and manRTPNs were stored in phosphate-buffered saline (PBS) for 1 month,
where they demonstrated a negligible change in size and showed an excellent stability (Figure
1D). Maintaining protein compounds of erythrocyte membranes on nanoparticles’ surfaces is
vital for harnessing their biofunctionality. Therefore, completeness of the membrane coating
was verified via sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE;
Figure S17), which indicated the manRTPNs maintained the whole effective biocomponent of
erythrocyte on the surface. Translocation of erythrocyte membranes might improve the
biocompatibility and circulating period of TPNs in vivo, which also sustains systemic delivery
and better targeting through both passive and active uptakes[35]. As reported, the elimination
T1/2 of triptolide molecules in rats following i.v. administration was 15-21 minutes, which
9
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
9

showed that drugs were rapidly eliminated[36]. However, TPNs and manRTPNs have
maintained higher plasma concentration after tail intravenous injection of mice in our work.
As the relative signal of drug down to 50%, TPNs lasted 4 h, and manRTPNs even had a
much longer elimination half-life of over 48 hrs (Figure 1E), which suggested that manRTPNs
significantly expanded blood circle compared to triptolide. Therefore, the self-assembling
TPNs nanostructure was more stable than original drug, and manRTPNs inherited biological
properties of erythrocyte membrane proteins for following efficacy.
Dual-Targeting capacity and immunosuppressive effect of manRTPNs
To determine each targeting capacity of manRTPNs to macrophage and T cell in vitro,
nanoparticles were Cy5.5-labelled and incubated with RAW264.7 and CTLL-2 treated with
IL-4 (100 ng/mL) or TNF-α (10 ng/mL), untreated cells and RSC364 as control. Flow
cytometry (FCM) analysis observed gradual increased uptake of nanoparticles by RAW264.7
and CTLL-2 for 6 hrs (Figure S18-20). An uptake curve was built using mean fluorescence
intensity (MFI) of cells at six time points, and cellular uptake rate was defined as k indicated
in figure 2A-C, which was the slope of the uptake curve. In detail, ks of nanoparticles in
activated RAW264.7 and CTLL-2 were significantly higher than that in untreated cells
(Figure 2A-C; Figure S21A & S22), and ks of manRTPNs were significantly higher than that
to TPNs both in RAW264.7 and CTLL-2 (Figure 2A-B), but no difference of ks existed
between TPNs and manRTPNs in RSC364 that no targeting ligands on it (Figure 2C). Further,
the k value in RAW264.7 was higher than that in CTLL-2 (Figure 2A-B), indicating the
affinity difference of interacted molecule pairs. To further confirm ligand-binding specificity,
10
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
10

Cy5.5-labelled TPNs (without mannose and CD58), RTPNs (with CD58) and manRTPNs
(with mannose and CD58) were incubated with treated macrophages and T cells for 3 hrs as
well, RSC364 and hFLS-RA as control. After treatment, CLSM images showed that CD206
and CD2 (Figure 2D-E, green) respectively overexpressed on surface of RAW264.7 and
CTLL-2. Significant red fluorescence was observed on RAW264.7 incubated with
manRTPNs, but not with TPNs and RTPNs (Figure 2D), while red fluorescence was observed
on CTLL-2 incubated with RTPNs and manRTPNs but not with TPNs (Figure 2E), which was
consistent with corresponding ligands distribution on the nanoparticles. When co-incubated
with RSC364, they also entered cells (Figure 2F; Figure S21b). Similar results were observed
in flow cytometry analysis (Figure 2G-I; Figure S21c). These results demonstrated the ability
of manRTPNs to target inflamed cells conferred by their mannose modified erythrocyte
membrane coating, which is probably attributed to specific affinity of mannose-CD206 and
CD58-CD2.
Except for migrated immune cells, synovial fibroblast is main resident effector of
inflammation. To determine immunosuppressive efficacy of manRTPNs in joint inflammatory
cells, LPS-treated RAW264.7, TNF-α-treated CTLL-2 and RSC364, and hFLS-RA were
incubated with manRTPNs in 96-well plates for 24 hrs. Firstly, cell viability was measured via
MTT assay. TPNs and manRTPNs didn’t significantly reduce RAW264.7 viability even at a
concentration of 500 nM (Figure 3A). However, at the same concentration CTLL-2, RSC364
and hFLS-RA showed an obvious cytotoxicity (Figure 3B-D). Specially, manRTPNs had an
IC50 (half maximal inhibitory concentration) values of 219 nM for activated CTLL-2
cytoxicity, 88.9 nM for activated RSC364 cytoxicity and 95.8 nM for hFLS-RA cytoxicity
11
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
11

(Figure 3B-D). The lower IC50 values of activated RSC364 and hFLS-RA indicated that
triptolide-induced cell death was mainly against anomalous proliferative synoviocytes in
inflamed joints, rather than lymphocytes like macrophages. Cells were following collected
and lysed for mRNA extraction to check certain inflammation associated genes variation by
qPCR. For LPS-treated RAW264.7, transcription factor NF-κB associated with most cytokine
transcription[37] and cyclooxygenase-2 (cox-2) were induced to decrease expression level, but
tissue inhibitor of metalloproteinase-1 (timp-1) showed opposite variation under manRTPNs
treatment. Receptor activator of NF-κB ligand (rankl) over-expressed on TNF-α-treated
CTLL-2 was also downregulated by manRTPNs. After activated with cytokines and
chemokines, synovial fibroblasts release matrix-degrading enzymes, including matrix
metalloproteinases (MMPs) and cathepsins causing cartilage and bone destruction[38]. In
RSC364, the transcripts of metalloproteases such as MMP-2 and MMP-9 and vascular
endothelial growth factor (VEGF) being critical for angiogenesis were lessen by manRTPNs
as well (Figure 3E). Thirdly, medium supernatants were collected to measured cytokines by
ELISA assay. supernatant IL-1β and IL-6 of LPS-treated RAW264.7 and supernatant IFN-� of
TNF-α-treated CTLL-2 lessened more quickly that treated with manRTPNs than TPNs
(Figure 3F-H), while supernatant IL-1β and IL-6 of TNF-α-treated RSC364 had a similar
decreasing curve between TPNs and manRTPNs groups (Figure 3I-J), which indicated that
manRTPNs had superior identification of immune cells than resident synoviocytes at least in
cytokine secretion. Consequently, manRTPNs selectively exerts its immunosuppressive
effects on selectively inducing lymphocytes and synovial fibroblasts cell death, but also
regulating production of proinflammatory cytokines, proinflammatory mediators and matrix
12
211
212
213
214
215
216
217
218
219
220
221
222
223
224
225
226
227
228
229
230
231
232
12

metalloproteinases, which suggested manRTPNs may have an ability to remodel
inflammatory microenvironment through its cell-specific anti-inflammation efficacy.
Targeting capacity and accumulation of manRTPNs in vivo
Upon the good capacity of manRTPNs to target activated macrophages and T cells in
vitro, we further verified their ability to target RA site in vivo. After CIA mouse model being
induced, all four paws swelled in varying degrees[39]. Hind paws were more heavily swollen,
and micro-computed tomography (Micro-CT) imaging showed distinct bone loss of joint
therein (Figure 4A). Then these mice received intravenous injections of free cyanine 5.5
(Cy5.5) solution and Cy5.5-labelled TPNs and manRTPNs for targeting imaging in vivo. As
shown in figure 4A, almost no fluorescence signal was detected in paws of the free-Cy5.5
group, indicating that the dye barely accumulated at local inflammatory sites. In the TPNs
group, we detected a weak signal, which peaked after 4 h (Figure 4A). While manRTPNs
obviously reached the RA sites and lasted there for ≥12 hrs, which would be attributed to their
long circulation and the targeting effect of the modified erythrocyte membranes. Thus,
significant fluorescence of manRTPNs lasted longer and showed greater accumulation than
that of TPNs. 24 hrs after intravenous injections, we dissected mouse and got froze sections of
left paws to observe nanoparticles’ retention in the inflamed joints. Contrary to free-Cy5.5 and
TPNs group, fluorescence microscopy indicated a very high fluorescence intensity of
manRTPNs in the synovial sublining layer, resulting in a reduced accumulation at the
inflamed joint due to the lack of permeability (Figure 4B). Tissue biodistribution further
revealed manRTPNs’ outstanding capacity to target the paws of RA mice, rather than other
13
233
234
235
236
237
238
239
240
241
242
243
244
245
246
247
248
249
250
251
252
253
13

groups (Figure 4C). Additionally, higher accumulation was observed in liver, suggesting that
this organ might be passive targeting of nanoparticles and the main organ metabolizing
triptolide (Figure 4C). The results confirmed that the effective accumulation of manRTPNs
provided a promising precondition of RA treatment.
Therapeutic efficacy of manRTPNs
The efficacy of ameliorating joint damage of manRTPNs was evaluated in a CIA mouse
model. Following arthritis induction, CIA mice presented symptoms of swelling in ankle and
paw joints and a clinical scoring was used to estimate the swelling degree (Figure S23).
According to protocol timeline, CIA mice were treated with intravenous injections of
triptolide at 15 mg/kg, TPNs and manRTPNs at doses of 25 mg/kg for 11 days (Figure 5A).
Surprisingly, swollen hindpaws and forepaws both shrunk considerably and even recovered to
the same extent as healthy ones with manRTPNs treatment rather than slightly shrunk in
TPNs and triptolide group, but swelling symptoms of other groups were increasingly severe
(Figure 5B), implying that inflammation process was exactly controlled by manRTPNs. As
paw swelling is only a surface phenomenon of RA, bone loss was further determined using
Micro-CT analysis. Reconstructed three dimensional (3D) bones images and calculated bone
mean density (BMD) showed that serious bone erosion was suppressed and bone density even
recovered to healthy mice level of manRTPNs treatment (Figure 5C-D), suggesting a
protective role of manRTPNs on cartilage and bone damage. Haemotoxylin and eosin (H&E)-
stained sections showed an intense anomalous proliferative cells infiltration in the joints and
synovium of control groups. By contrast, remarkable reduction of immune infiltration and
14
254
255
256
257
258
259
260
261
262
263
264
265
266
267
268
269
270
271
272
273
274
14

preservation of cartilage were observed in manRTPNs group (Figure 5E). Further, cytokines
including TNF-α and IL-6 are known to increase with the onset of arthritis and correlate
strongly with disease severity[40]. Immunohistochemical analysis showed that TNF-α and IL-
6 in synovium and articular cavity significantly declined in manRTPNs groups (Figure 7A-B)
and serum levels of them also observed to alleviate similarly (Figure 6C), indicating effective
reduction of arthritis at cytokines level. Moreover, gene heatmap showed that manRTPNs
suppressed several genes expression including rank, rankl, vegf, vegfr2, NF-κB, cox-2, IL-1β,
mmp-2, mmp-3, mmp-9 and mmp-13, that these transcripts are dramatically increased and
contribute in joints damage. Some negative regulators such as opg, timp1 and timp2 were
upregulated their gene expression by manRTPNs (Figure 6D), indicating manRTPNs preserve
appropriate and balanced concentration of cytokines and inflammatory factors through two-
way regulation. RANKL/RANK/ORG signal pathway is key of bone metabolism[41]. The
up-/down-regulation of manRTPNs also recovered the balance of RANKL/RANK/ORG and
metalloproteinases and their inhibitors (TIMP) to normal. Thus, manRTPNs ameliorated
destroyed through aiming to RA pathological factors including cytokines, extracellular matrix
(ECM), bone metabolism and angiogenesis.
Biosafety of manRTPNs nanoparticles
To investigate safety and biocompatibility of TPNs and manRTPNs in vivo, we
administered PBS, TPNs or manRTPNs at a single dose of 25 mg/kg to healthy mice via tail
vein, while to other mice we administered triptolide (original drug of TPNs and manRTPNs)
at 15 mg/kg by intraperitoneal injection. It is worth mentioning that most mice in the
15
275
276
277
278
279
280
281
282
283
284
285
286
287
288
289
290
291
292
293
294
295
15

triptolide group died soon (within 24 hrs) after 25 mg/kg administration, indicating the high
toxicity of the original drug (Figure S24). After 1-week treatment, mice treated with triptolide
showed a significant decrease in count of white blood cells (WBC), RBCs and platelets
(Figure 7A), suggesting the occurrence of bone marrow suppression, which is one of the main
side effects of triptolide. The main three type cells comprised WBC including neutrophils,
lymphocytes, and monocytes showed significant count differences between triptolide group
and other groups, which explained the WBC count variation conferred from triptolide (Figure
7B). The hemoglobin concentration did not differ significantly among all the groups in the
blood (HGB), mean corpuscular hemoglobin (MCHC) level, and hemoglobin distribution
width (HDW) (Figure 7C), indicating that triptolide didn’t influence the physiological
function of RBCs. Blood biochemistry analysis showed that blood urea nitrogen (BUN),
aspartate aminotransferase (AST), creatinine (Cr) and lactate dehydrogenase (LDH) levels of
mice treated with triptolide were significantly higher than those of mice in the TPNs and
manRTPNs groups (Figure 7D-G). These variations demonstrated that the hepatotoxicity and
nephrotoxicity of triptolide were greatly reduced in TPNs, with RBC membrane coating
yielding even better safety. Hematoxylin and eosin (H&E) staining of major organs showed
that triptolide treatment caused inflammatory infiltration of the liver and severe renal injury
characterized by loss of the brush border, tubular epithelial cells detachment from the
basement membrane and tubular obstruction. Meanwhile, the TPNs and manRTPNs groups
showed no significant variation in cellular morphology and negligible organ damage, in line
with the PBS group (Figure 7H). However, TPNs and manRTPNs significant reduced
triptolide toxicity.
16
296
297
298
299
300
301
302
303
304
305
306
307
308
309
310
311
312
313
314
315
316
317
16

Conclusion
Our study expanded the RBC cell membrane biological application in nanocarrier for
targeting inflammation through innate and specific immune recognition, and the triptolide
nanodrug showed high drug content, low side effect, good biocompatibility which showed
great potential for further inflammation diseases therapy. Generally, there are two
improvement of manRTPNs in cell-membrane-coated nanoparticle delivery system, namely
triptolide nanodrug core and RBC membrane shell. RBC membrane-camouflaging
nanoparticles has been extensive established platform, but there are still many deposits from
RBC cell itself to further develop disease-targeting and therapy platform except for tumor.
The natural immunological function of RBC cells has been always neglected in targeted drug
delivery construction. So, taking advantage of adhesion molecules affinity between RBC cells
and immunocytes in building nanocarrier for transporting drug to inflammation deeply
expands application of established RBC membrane platform. On the other side, triptolide has
excellent anti-inflammation and immunosuppressive effect and solving its low water-
solubility and toxicity is premise of using it in various diseases management. Our
reconstruction of triptolide isn’t content-limited loading in micelles, but drug-based self-
assembled size-controlled nanoparticles. This strategy takes advantages of drugs’
hydrophobicity to solve its bad water-solubility, which is magic of nanotechnology. While
inflammation is prevalent related with many diseases like cardiovascular diseases,
gastrointestinal tract, cancer and pathogen infection[42]. Therefore, manRTPNs as a drug
delivery system of inflammation targeting and treating may be a promising agent in other
inflammation-associated diseases therapy.
17
318
319
320
321
322
323
324
325
326
327
328
329
330
331
332
333
334
335
336
337
338
339
17

Materials and methods
Animal care
Mice were housed in an animal facility at Shenzhen Institutes of Advanced Technology,
Chinese Academy of Sciences (SIAT). All animal experiments were performed in accordance
with Guidance Suggestions for the Care and Use of Laboratory Animals and approved by
Sciences Animal Care and Use Committee of SIAT.
Erythrocyte membranes derivation
Erythrocyte membranes were collected according to a previously reported method with
modification. Briefly, fresh heparinized whole blood was collected from male mice (20-22 g)
and subsequently centrifuged at 2000 rpm for 15 min at 4 °C to remove the plasma and the
white buffy coat. The collected red blood cells (RBCs) were washed with 1× PBS three times
and suspended in 0.25× PBS for 2 hours at 4 °C, and then the hemoglobin was removed by
centrifugation at 9,000 rpm for 45 min. The resulting pink pellet was purified with 1× PBS,
and the collected erythrocyte membrane was suspended and stored in distilled water. To form
ligand-inserted erythrocyte ghosts, the light pink solution was incubated with DSPE-PEG-
mannose for 30 min, 37°C water bath. Erythrocyte membranes with PEG-mannose or not was
further prepared by extruding through a 200 nm polycarbonate porous membrane with an
Avanti mini-extruder (Avanti Polar Lipids).
Synthesis of TPNs
The TPNs were synthesized by a prodrug self-assembly method. Briefly, after dissolving
5mmol Boc-Phe-Phe-OH (BOC-FF, J&K scientific Ltd., China), 6 mmol 1-Ethyl-3-(3-
18
340
341
342
343
344
345
346
347
348
349
350
351
352
353
354
355
356
357
358
359
360
18

dimethylaminopropyl) carbodiimide (EDC, J&K scientific Ltd., China) and 6 mmol N-
Hydroxysuccinimide (NHS, J&K scientific Ltd.,China) in 3 mL anhydrous DMF, another
anhydrous DMF solution containing 5.5 mmol glucosamine (AG, Sigma-Aldrich, USA) and 5
mmol TEA (J&K scientific Ltd., China) was added drop-wise into it. The mixture was
evaporated under reduced pressure for 24 hours to get crude product BOC-FF-AG. Then,
crude BOC-FF-AG was dissolved in 30 mL equal volume mixture of anhydrous
dichloromethane and trifluoroacetic acid (TFA, J&K scientific Ltd., China) and stirred for 4
hours at room temperature. Afterward, pure NH2-FF-AG was obtained after evaporated under
reduced pressure and purified by silica gel column chromatography with dichloromethane and
methanol. Secondly, 1 mmol triptolide (TP, Shanghai Yuanye Bio-Technology Co., Ltd.,
China), 1.2 mmol succinic anhydride (SAA, Sigma-Aldrich, USA) and 0.2 mmol dimethyl
aminopyridine (DMAP, Sigma-Aldrich, USA) were dissolved in 3 mL pyridine and stirred
overnight. After then, the mixture was diluted with ethyl acetate and washed it with solution
containing saturated copper sulfate (J&K scientific Ltd.), water and brine. After reaction,
residue TP-COOH was obtained through drying with anhydrous Na2SO4 and concentration
with rotary evaporator. In the third step, TP-COOH, dicyclohexylcarbodiimide (DCC, Sigma-
Aldrich, USA) and 1-hydroxy-5-pyrrolidinedione (NHS, J&K scientific Ltd., China) of equal
molar ratio were dissolved into 3 mL anhydrous dichloromethane and stirred at room
temperature for overnight. The final product TP-FF-AG was gained by reaction of activated
TP-COOH and NH2-FF-AG for overnight at room temperature and purified by analytical RP-
HPLC and semi-preparative RP-HPLC with simultaneous detection at 265 nm. Analytical RP-
HPLC was performed at room temperature on the Shimadzu LC 20 with UV detector SPD-
19
361
362
363
364
365
366
367
368
369
370
371
372
373
374
375
376
377
378
379
380
381
382
19

20A using Inertsil ODS-SP column (4.6 x 250 mm, 5 µm, 100Å). The RP-HPLC gradient was
started at 10% of B (CH3CN, J&K scientific Ltd., China), then increased to 100% of B over
30 min (A: 0.1% TFA in water). Semi-preparative RP-HPLC was performed on the ULTIMAT
3000 Instrument (DIONEX). For fluorescence imaging experiments, cyanine5.5 (Cy5.5,
Lumiprobe, USA) was linked to intermediate product of NH2-FF-AG after esterification
reaction to obtain Cy5.5-FF-AG in the same way. Ultimately, nanoparticles were self-
assembled by added TP/Cy5.5-FF-AG dissolved in DMSO dropwise into PBS while vigorous
stirring and dialysis for 2 days. 1H NMR and 13C NMR spectra were recorded with a Bruker
VANCE III400 spectrometer (400 MHz). The high-resolution mass spectra (HR-MS) were
measured on a Bruker Micro TOF II 10257 instrument. For membrane coating, nanoparticles
were mixed with prepared membranes in ratio of 1:1. The resultant mixture was subsequently
extruded nine times through a 200 nm polycarbonate porous membrane using an Avanti mini-
extruder to yield the erythrocyte membranes-coating TPNs or mannose inserted erythrocyte
membrane-coating TPNs (namely RTPNs or manRTPNs).
Nanoparticles Characterization
The hydrodynamic diameter and zeta potential of NPs suspended in 1 × PBS were
measured by dynamic light scattering (DLS) (Zeta Plus, Brookhaven Instruments, USA). The
morphologies of TPNs and manRTPNs were observed by transmission electron microscope
(TEM, FEI spirit T12) at an accelerating voltage of 120 keV. The stability experiment of
TPNs and manRTPNs in PBS was monitored by DLS over five weeks and stored at 4°C. The
proteins retained on manRTPNs compared with those on the natural erythrocyte membrane
20
383
384
385
386
387
388
389
390
391
392
393
394
395
396
397
398
399
400
401
402
403
20

were observed by SDS-PAGE. Briefly, 2 mg of erythrocyte membrane vesicles and
manRTPNs were collected by centrifugation at 15,000g for 30 min, mixed in SDS sample
buffer (Invitrogen, USA), and heated at 90 °C for 5 min. Then, 20 μL of each sample was run
on a 10% SDS-polyacrylamide gel (Bio-Rad, SDS-PAGE Gel Preparation Kit, USA) at 120 V
for 1 h, followed by Coomassie blue staining and imaging.
Pharmacokinetics
To evaluate the circulation half-life of TPNs and manRTPNs in vivo, 150 μL Cy5.5-
labeled nanoparticles were injected into the tail vein of the mice (n = 3). Twenty microliters of
blood were collected at 1, 5, 15, 30 min, and 1, 2, 4, 8, 24, 48, and 72 h following the
injection. Each particle group contained 3-4 mice. The collected blood samples were diluted
with 30 μL PBS in a 96-well plate before fluorescence measurement. Pharmacokinetics
parameters were calculated to fit a two-compartment model and a one-way nonlinear model.
Drug toxicity analysis in vivo
Purchased from Vital River Laboratory Animal Technology Co. Ltd (Beijing, China),
males BALB/c mice (6∼8 weeks old) were randomly divided into four groups and
intravenously injected with PBS, TPNs and manRTPNs at a single dosage of 25 mg/kg and
triptolide of 15 mg/kg 4 times for eight days. A total of 7 days after last administration, all
mice were euthanized, and blood was collected for and blood routine examination and
biochemical parameters measurement, while main organs including heart, liver, spleen, lung,
and kidney were excised and stained by H&E for histological analysis.
Cell culture
21
404
405
406
407
408
409
410
411
412
413
414
415
416
417
418
419
420
421
422
423
424
21

RAW264.7 cells, CTLL-2, RSC364 and hFLS-RA were purchased from the American
Type Culture Collection (ATCC, USA) and cultured in Dulbecco’s minimum essential
medium (DMEM, Corning, USA) supplemented with 10% fetal bovine serum (FBS, Gibco,
USA), 1% penicillin (100 IU/mL, Corning, USA), and streptomycin (100 μg/mL, Corning,
USA) and placed at 37 °C in a 5% CO2 humidified atmosphere. Cell lines were purchased
with certification of authentication and free from Mycoplasma.
Quantification of cell targeting in vitro
A 6-well culture plate was prepared and RAW264.7, CTLL-2, RSC364 and hFLS-RA
cells were implanted at a density of 5 × 105 cells/well. 12-16 hrs later, 80% of cell coverage
was confirmed. 50 �L Cy5.5-labelled TPNs or manRTPNs was adding into prepared cells
every hour for six hours, incubated with covering foil paper and collected them in the sixth
hour. After digestion with trypsin (Gibco, USA), cells were put together and washed three
times with ice-cold PBS, then underwent flow cytometry by FACS Canto II (BD
Biosciences). The data was analyzed using FlowJo software. Mean fluorescence intensity
(MFI) reflected amount of intake nanoparticles amount. An uptake curve was constructed
with MFI and time, and a k value being the slope of uptake curve was defined as uptake rate
to measure targeting capacity of nanoparticles.
Immunofluorescence assay of macrophages and T cells with nanoparticles
Cells were cultured with medium added recombinant mouse IL-4 (100 ng/mL, R&D
System, USA) or recombinant TNF-α (10 ng/mL, R&D System, USA) for six hours and
incubated with TPNs, RTPNs or manRTPNs for 3 hours. Afterward, cells were fixed with 4%
22
425
426
427
428
429
430
431
432
433
434
435
436
437
438
439
440
441
442
443
444
445
22

paraformaldehyde (Sigma-Aldrich, USA) for 10 minutes, permeabilized with 0.1% Triton™
X-100 (Sigma-Aldrich, USA) for 10 minutes, blocked with 1% BSA for 1 hour and labeled
with 2 µg/mL anti-mannose receptor/AF488 conjugated antibody (ab195191, Abcam) and
rabbit anti-CD2/AF488 conjugated antibody (bs-2899R, Bioss) for 3 hours at room
temperature. Nuclei were stained with DAPI. All manipulation must be away from light. The
images were captured under confocal microscopy (Leica, TCS SP5).
Cellular cytotoxicity study
The cellular toxicities of TPNs and manRTPNs on macrophages, T cells, FLSs and
hFLSs were determined by MTT assay using CCK-8 Kit. Cells were implanted in a 96-well
culture plate at a density of 8 × 103 cells/well (5 replicates each treated group). After 12 hrs,
PBS, TPNs and manRTPNs with concentrations of 10 nM, 50 nM, 100 nM, 200 nM, 500 nM
were added into medium for further 24 hrs co-culture. Then the cells were mildly washed by
PBS before added CCK-8 mixed solution. Then, the optical density in 450 nm was measured
by Multiskan GO (Thermo Fisher Scientific).
Cytokines concentration analysis
The TNF-α, IL-6 and IFN-� concentrations of cells culture supernatants were determined
with ELISA Kit using mouse ELISA Kits (Biolegend, USA) as manipulation. Also, the TNF-α
and IL-6 concentrations of mouse serum samples were determined with TNF-α and IL-6
ELISA Kits (Biolegend, USA). Briefly, Serum samples of treated mice were collected at the
end of experiment and concentrations of TNF-α and IL-6 were quantified with ELISA kits.
Specifically, the whole blood of mice was collected by submandibular bleeding into
23
446
447
448
449
450
451
452
453
454
455
456
457
458
459
460
461
462
463
464
465
466
23

microtubes and allowed to clot at room temperature for 30 min. Samples were then
centrifuged at 2,000g for 6 min to collect supernatant serum. Serum samples were
immediately frozen at -20 °C until analysis by using mouse TNF-α and mouse IL-6 ELISA
kits (Biolegend) within 3 days of collection.
Genes expression profiles analysis of cells
After treated with nanoparticles of serious concentrations, cells were harvested to isolate
RNA. RNA was extracted with TRIzol reagent (Invitrogen, USA) according to the
manufacturer’s instructions. Complementary DNAs (cDNAs) were synthesized with
QuantiTect Reverse Transcription Kit (QIAGEN K.K., Tokyo, Japan) and the specific gene
transcripts were quantified by quantitative real-time PCR using QuantiTect SYBR Green PCR
Kit (QIAGEN K.K., Tokyo, Japan) and analyzed with ABI 7500 real-time PCR system
(Applied Biosystems, USA). The gene expression patterns along with concentration were
highlight with heatmap by using TBtools package.
Collagen induced arthritis (CIA) mice model induction
Seventy-two female DBA/1 mice (6∼8 weeks old) were purchased from Vital River
Laboratory Animal Technology Co. Ltd (Beijing, China). All mice were maintained in a SF
system which is room equipped with an air-filtering system, and the cages and water were
sterilized. CIA was induced as previously reported study[39]. Briefly, bovine type II collagen
(Chondrex, Redmond, WA, USA) was dissolved in 0.1M acetic acid overnight at 4°C. This
was emulsified in an equal volume of complete Freund’s adjuvant (Chondrex, Redmond, WA,
USA). The mice were immunized intradermal at the base of the tail with 100 �L of emulsion
24
467
468
469
470
471
472
473
474
475
476
477
478
479
480
481
482
483
484
485
486
487
24

containing 100 �g of type II collagen. On day 21, mice were boosted intraperitoneal with 100
�g type II collagen dissolved in phosphate buffered saline (PBS). Clinical scores of each
hindpaws and forepaws of mice were obtained following the standard evaluation process[39].
In detail, no evidence of erythema and swelling occurred is score 0, erythema and mild
swelling appeared is score 1, erythema and mild swelling extended from the ankle to the
tarsals is score 2, erythema and moderate swelling extended from the ankle to metatarsal
joints is score 3 and erythema and severe swelling encompassed the ankle, paws, and digits or
ankyloses of the limb is score 4. Then total clinical scores of hindpaws and forepaws were
calculated. At least six mice each group.
Nanoparticles targeting to inflamed joints
After CIA model was established, free Cy5.5, Cy5.5-labelled TPNs and manRTPNs were
intravenously injected into the CIA mice (20 mg/kg, n = 4). Fluorescence images were
obtained after 1, 2, 4, 8, 12, and 24 hours using an in vivo imaging system (IVIS, Caliper,
USA). Then, the mice were sacrificed for ex vivo tissue/organs distribution analysis at the end
of experiment. Meanwhile, the synovial tissue was dissected and dehydrated in 30% sucrose
solution overnight and then cry-sectioned at 8 �m. The sections were stained with DAPI and
washed twice. Fluorescence images were obtained using a confocal microscope (Leica, TCS
SP5). The fluorescence intensity of sections in different groups was calculated using software
Image J.
manRTPNs treatment assay
25
488
489
490
491
492
493
494
495
496
497
498
499
500
501
502
503
504
505
506
507
25

According to CIA mice model conduction, starting at day 21, the clinical scores were
calculated to make sure that CIA mice were almost equably and randomly divided into four
groups, while healthy DBA/1 mice were set as control (n = 5). Five different agents namely
PBS, RBC vesicle, triptolide, TPNs and manRTPNs were intravenously injected every two
days. The timeline of whole therapy experiment from model making to the end was according
to schematic diagram shown in figure 5A. The arthritis index of each group was recorded over
time to evaluate treatment efficacy. At the end, the mice were sacrificed for further analysis.
Micro-CT imaging of mice paws
The knee and ankle joints of each experimental mouse hindpaws were scanned using
micro-CT system (Bruker's SkyScan 1278, UK) to observe joint bone situation. Images were
acquired at 55 kV, 72 μA and 300 ms/frame, with 360 views. The 3D structures of knee joint
and ankle joint were reconstructed from 360 views and BDM was evaluated by corresponding
SkyScan NRecon package. Analysis typically took around 15 min per joint for an experienced
operator.
Histological analysis and immunohistochemical staining assay
At study endpoints, mice were euthanized and hind knee joints were collected for H&E
staining and immunohistochemical staining. Briefly, the ankle and knee joints of scarified
animals as well as their heart, liver, spleen, lung and kidney were fixed in 10% buffered
formalin and then joints were incubated in decalcifying solution (4% hydrochloric acid in 4%
formaldehyde) at room temperature for 7 days for decalcification. After paraffinization,
microtome (Leica) slices of 8 μm were prepared and stained with haematoxylin and eosin,
26
508
509
510
511
512
513
514
515
516
517
518
519
520
521
522
523
524
525
526
527
528
26

images were taken by using inverted microscope (Olympus, IX71, Japan) and the
inflammatory cell infiltration in synovial tissues, bone and cartilage was evaluated by Image J
software. After deparaffinization the slices were subjected to antigen recovery in 0.01 M
sodium citrate buffer at 125 °C for 30 s, followed by 10 s at 90 °C, and then subjected to the
endogenous peroxidase inactivation by covering tissue with 3% hydrogen peroxide for 5 min.
After blocking non-specific binding sites with 10% goat serum in PBS, the slices were
incubated with TNF-α/IL-6 monoclonal antibody (Invitrogen) with a dilution rate of 1:100,
respectively, at 4 °C for 24 h. Then the slices were incubated with horseradish peroxidase
(HRP)-conjugated secondary antibody with a dilution rate of 1:800 at 37 °C for 1 hour.
Sections were developed using the DAB substrate and then counterstained with haematoxylin.
The images were captured and analyzed by inverted microscope (Olympus, IX71, Japan) and
Image J software.
Gene expression profiles analysis of tissues
The paws and ankles were dissected from mice on day 22 of arthritis, snap-frozen in
liquid nitrogen, ground into powder, and homogenized. All procedure must be under RNase-
free conditions. The RNA isolation and real-time PCR assay were carried out following the
protocol following. Briefly, total RNA was extracted with TRIzol reagent (Invitrogen,
Carlsbad, CA, USA) from the tissue homogenates according to the manufacturer’s
instructions. The total RNA (1 �g) was reverse transcribed to cDNA using the QuantiTect
Reverse Transcription Kit (QIAGEN, Japan) and the specific gene transcripts were quantified
by quantitative real-time PCR using QuantiTect SYBR Green PCR Kit (QIAGEN K.K.,
27
529
530
531
532
533
534
535
536
537
538
539
540
541
542
543
544
545
546
547
548
549
27

Tokyo, Japan) and analyzed with ABI 7500 real-time PCR system (Applied Biosystems,
USA). PCR was performed as 40 cycles at 94°C for 15 s, 55°C for 30 s, and 72°C for 30 s.
The relative RNA expression was calculated with comparative �� method; �-actin as internal
ontrol. The gene expression patterns along with concentration were highlight with heatmap by
using TBtools package. Gene-specific primers were synthesized by Sangon Biotech
(Shanghai) Co., Ltd. and list in the TableS1.
Statistical Analysis
All the results are reported as mean ± SD. The differences among groups were analyzed
using one-way ANOVA analysis and Student’s t-test; *represents P < 0.05, ** represents P <
0.01.
Acknowledgments
This work was supported by National Natural Science Foundation of China (81801838,
31571013, 81701816 and 81601552), Guangdong Natural Science Foundation of Research
Team (2016A030312006), K.C. Wong Education Foundation (GJTD-2018-14), Natural
Science Foundation of Guangdong Province (2018A030313013), Shenzhen Science and
Technology Program (JCYJ20180302145912832, JCYJ20160429191503002,
JCYJ20170818162259843 and JCYJ20170818163739458). We thank Pro. Hongchang Li and
Dr. Yifan Ma of SIAT for their useful and meaningful recommendations to improve this paper,
and thank Mr. Danhui Quan for his help in some drawing.
Competing Interests
The authors have declared that no competing interest exists.
28
550
551
552
553
554
555
556
557
558
559
560
561
562
563
564
565
566
567
568
569
570
28

29
571
29
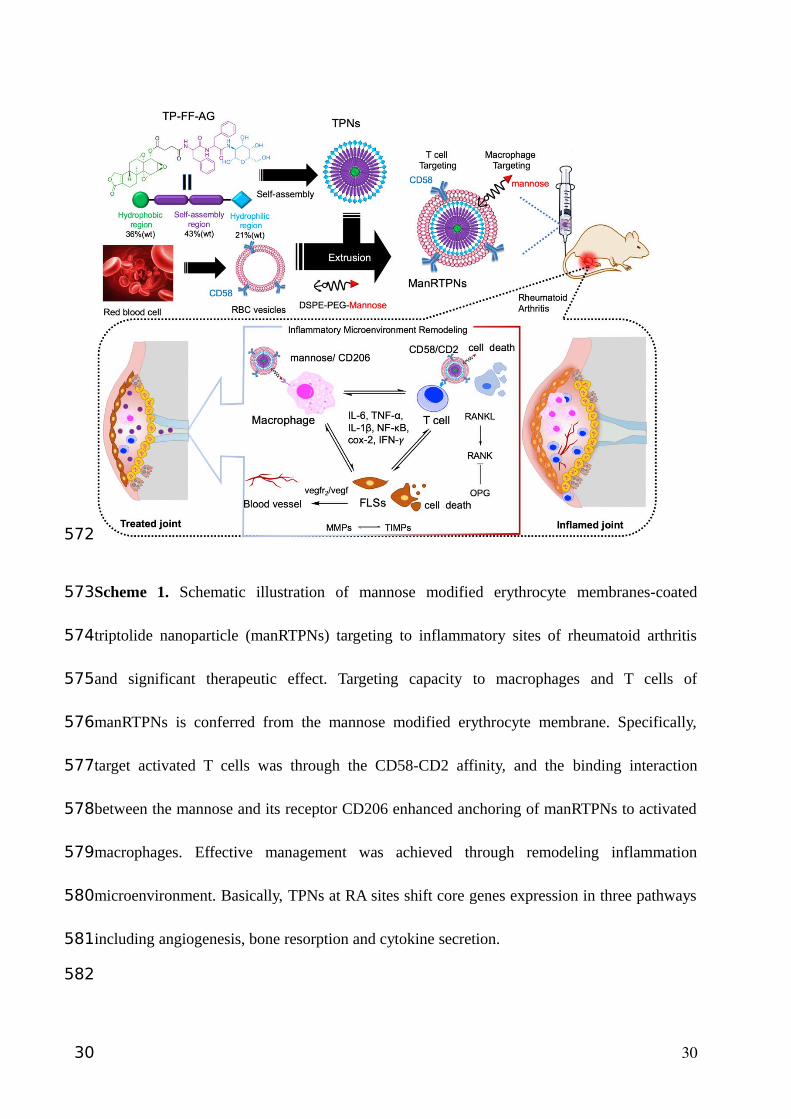
Scheme 1. Schematic illustration of mannose modified erythrocyte membranes-coated
triptolide nanoparticle (manRTPNs) targeting to inflammatory sites of rheumatoid arthritis
and significant therapeutic effect. Targeting capacity to macrophages and T cells of
manRTPNs is conferred from the mannose modified erythrocyte membrane. Specifically,
target activated T cells was through the CD58-CD2 affinity, and the binding interaction
between the mannose and its receptor CD206 enhanced anchoring of manRTPNs to activated
macrophages. Effective management was achieved through remodeling inflammation
microenvironment. Basically, TPNs at RA sites shift core genes expression in three pathways
including angiogenesis, bone resorption and cytokine secretion.
30
572
573
574
575
576
577
578
579
580
581
582
30
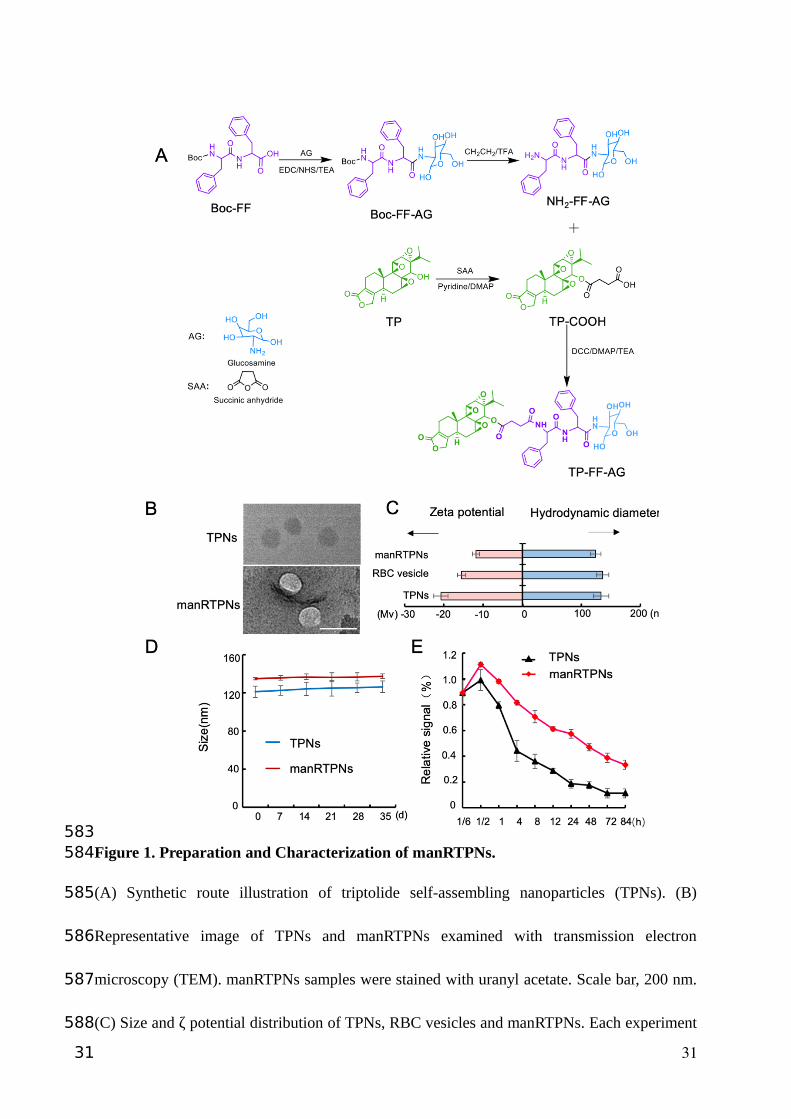
Figure 1. Preparation and Characterization of manRTPNs.
(A) Synthetic route illustration of triptolide self-assembling nanoparticles (TPNs). (B)
Representative image of TPNs and manRTPNs examined with transmission electron
microscopy (TEM). manRTPNs samples were stained with uranyl acetate. Scale bar, 200 nm.
(C) Size and ζ potential distribution of TPNs, RBC vesicles and manRTPNs. Each experiment
31
583584
585
586
587
588
31

had three replicates. (D) Stability of TPNs and manRTPNs in PBS over five weeks. Each
experiment had three replicates. (E) Pharmacokinetics of TPNs and manRTPNs in vivo. Each
experiment point had three replicates.
32
589
590
591
592
32
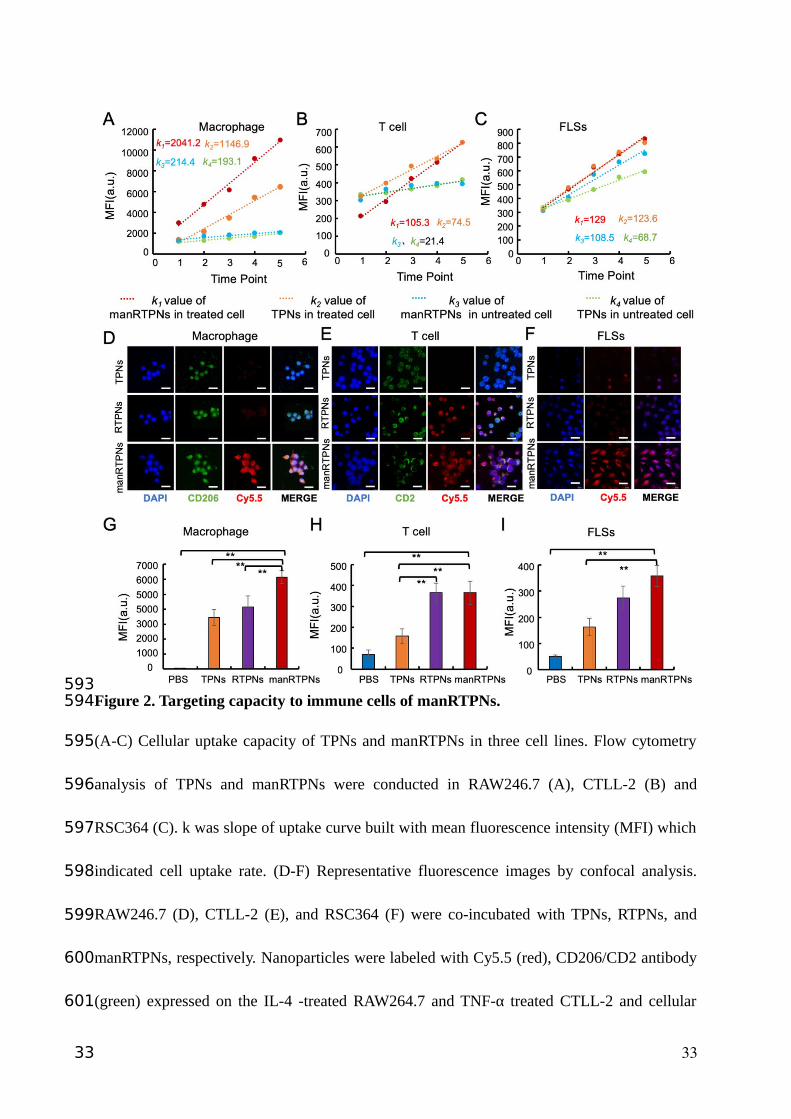
Figure 2. Targeting capacity to immune cells of manRTPNs.
(A-C) Cellular uptake capacity of TPNs and manRTPNs in three cell lines. Flow cytometry
analysis of TPNs and manRTPNs were conducted in RAW246.7 (A), CTLL-2 (B) and
RSC364 (C). k was slope of uptake curve built with mean fluorescence intensity (MFI) which
indicated cell uptake rate. (D-F) Representative fluorescence images by confocal analysis.
RAW246.7 (D), CTLL-2 (E), and RSC364 (F) were co-incubated with TPNs, RTPNs, and
manRTPNs, respectively. Nanoparticles were labeled with Cy5.5 (red), CD206/CD2 antibody
(green) expressed on the IL-4 -treated RAW264.7 and TNF-α treated CTLL-2 and cellular
33
593594
595
596
597
598
599
600
601
33

nuclear was labeled with DAPI (blue). Scale bars, 100 μm. (G-I) Flow cytometry analysis of
RAW246.7 (G), CTLL-2 (H) and RSC364 (I) incubated with various nanoparticles. **
indicated significant difference and p < 0.05.
34
602
603
604
605
34
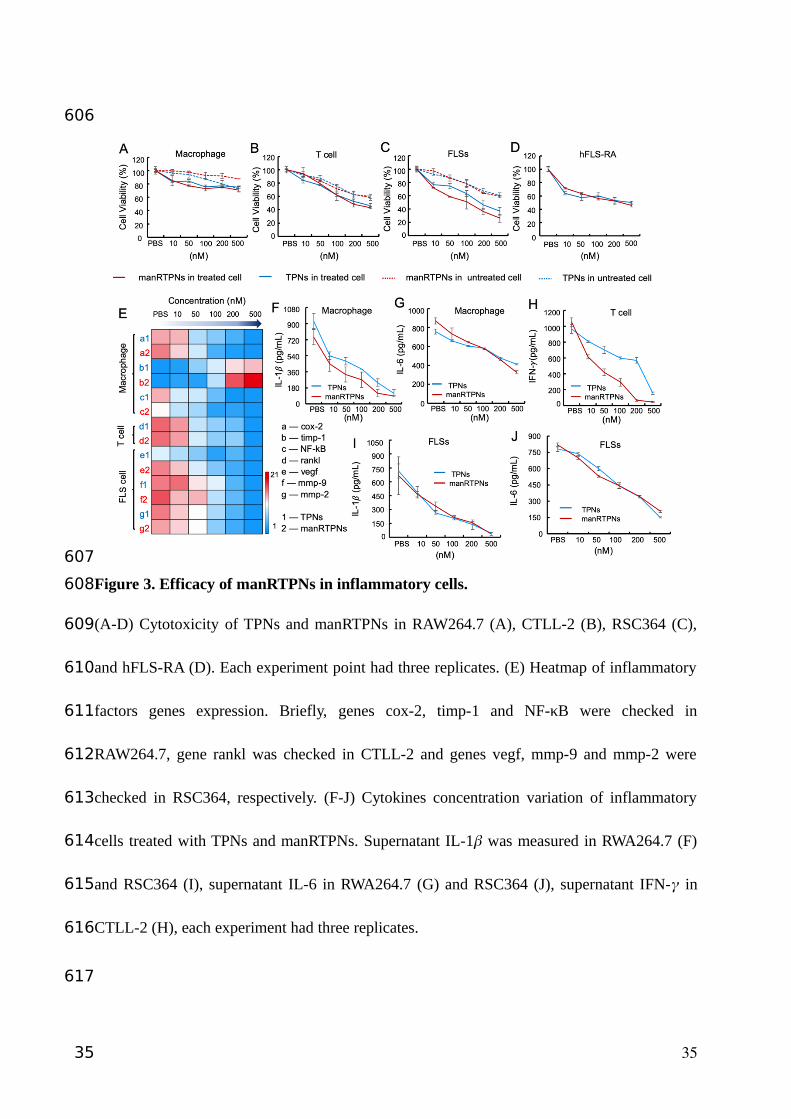
Figure 3. Efficacy of manRTPNs in inflammatory cells.
(A-D) Cytotoxicity of TPNs and manRTPNs in RAW264.7 (A), CTLL-2 (B), RSC364 (C),
and hFLS-RA (D). Each experiment point had three replicates. (E) Heatmap of inflammatory
factors genes expression. Briefly, genes cox-2, timp-1 and NF-κB were checked in
RAW264.7, gene rankl was checked in CTLL-2 and genes vegf, mmp-9 and mmp-2 were
checked in RSC364, respectively. (F-J) Cytokines concentration variation of inflammatory
cells treated with TPNs and manRTPNs. Supernatant IL-1� was measured in RWA264.7 (F)
and RSC364 (I), supernatant IL-6 in RWA264.7 (G) and RSC364 (J), supernatant IFN-� in
CTLL-2 (H), each experiment had three replicates.
35
606
607
608
609
610
611
612
613
614
615
616
617
35
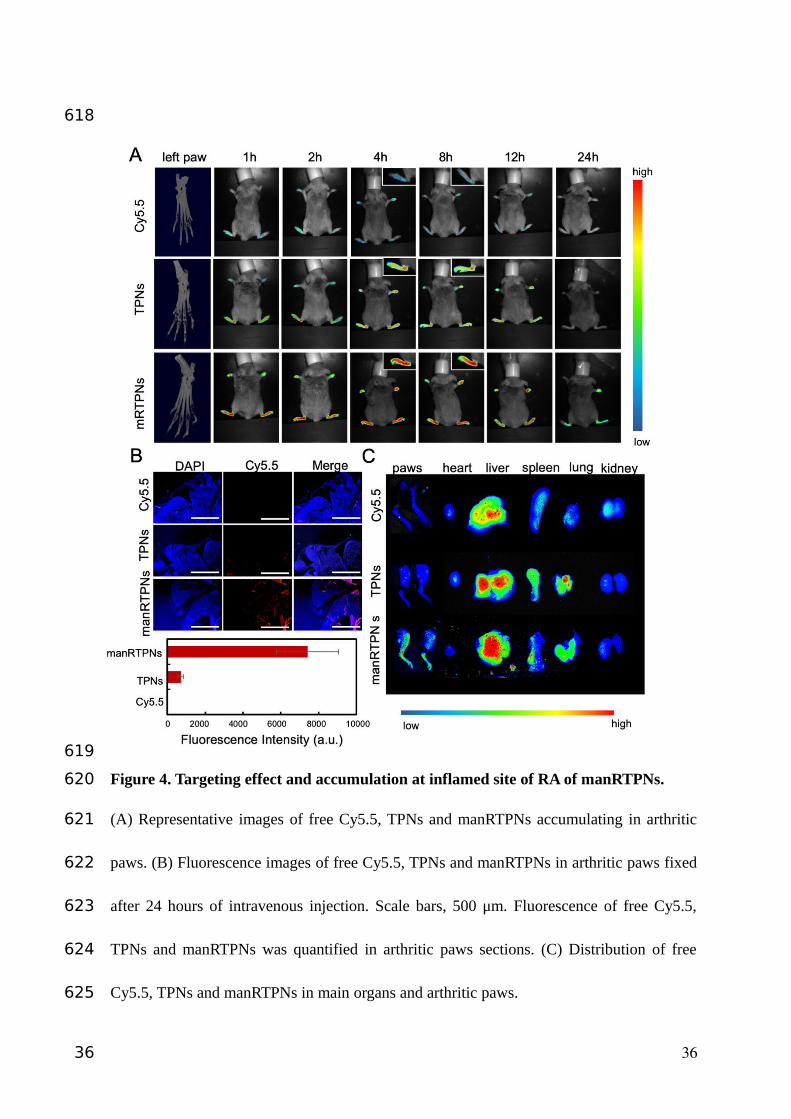
Figure 4. Targeting effect and accumulation at inflamed site of RA of manRTPNs.
(A) Representative images of free Cy5.5, TPNs and manRTPNs accumulating in arthritic
paws. (B) Fluorescence images of free Cy5.5, TPNs and manRTPNs in arthritic paws fixed
after 24 hours of intravenous injection. Scale bars, 500 μm. Fluorescence of free Cy5.5,
TPNs and manRTPNs was quantified in arthritic paws sections. (C) Distribution of free
Cy5.5, TPNs and manRTPNs in main organs and arthritic paws.
36
618
619
620
621
622
623
624
625
36

37
626
37
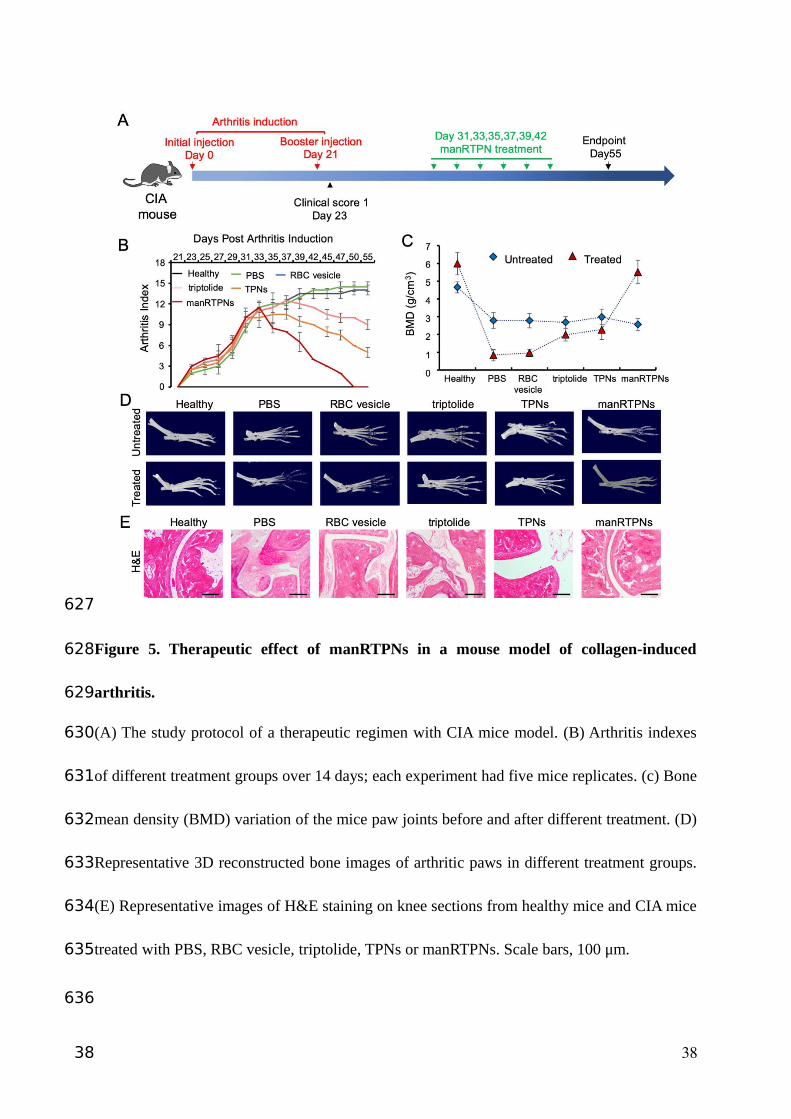
Figure 5. Therapeutic effect of manRTPNs in a mouse model of collagen-induced
arthritis.
(A) The study protocol of a therapeutic regimen with CIA mice model. (B) Arthritis indexes
of different treatment groups over 14 days; each experiment had five mice replicates. (c) Bone
mean density (BMD) variation of the mice paw joints before and after different treatment. (D)
Representative 3D reconstructed bone images of arthritic paws in different treatment groups.
(E) Representative images of H&E staining on knee sections from healthy mice and CIA mice
treated with PBS, RBC vesicle, triptolide, TPNs or manRTPNs. Scale bars, 100 μm.
38
627
628
629
630
631
632
633
634
635
636
38
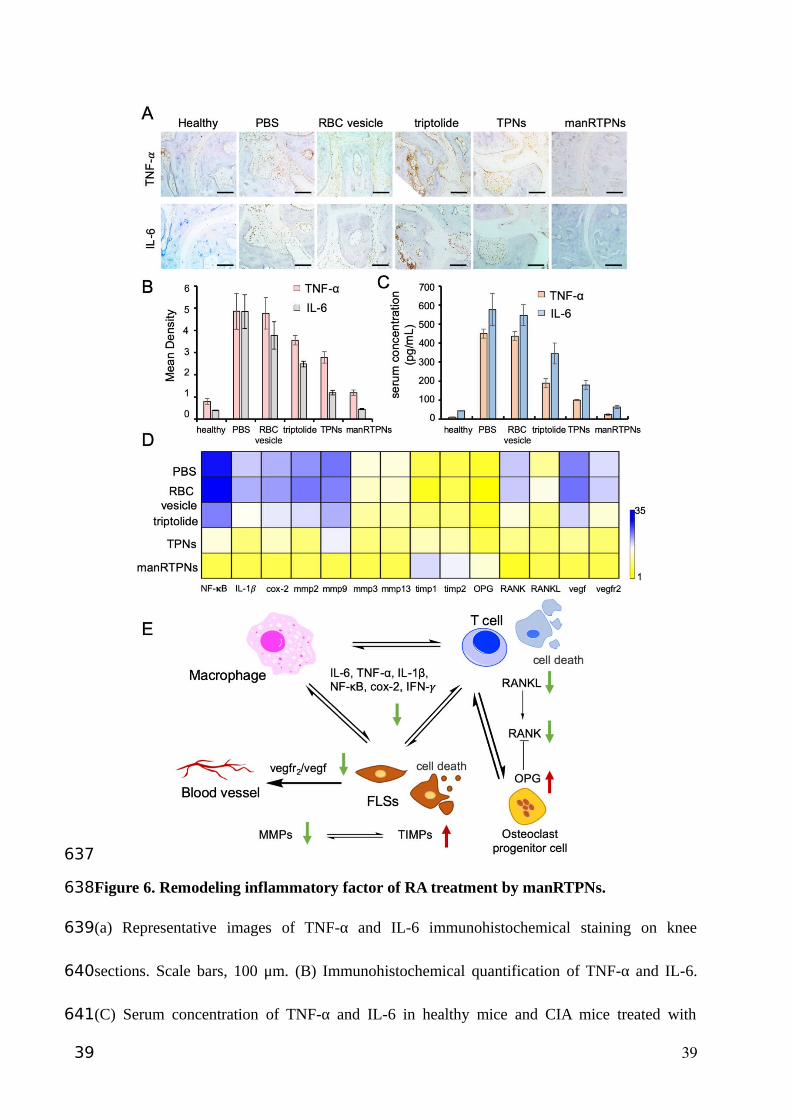
Figure 6. Remodeling inflammatory factor of RA treatment by manRTPNs.
(a) Representative images of TNF-α and IL-6 immunohistochemical staining on knee
sections. Scale bars, 100 μm. (B) Immunohistochemical quantification of TNF-α and IL-6.
(C) Serum concentration of TNF-α and IL-6 in healthy mice and CIA mice treated with
39
637
638
639
640
641
39

different groups. Each experiment point had three replicates. (D) Heatmap of genes
expression profiles in different treatment.
40
642
643
644
40
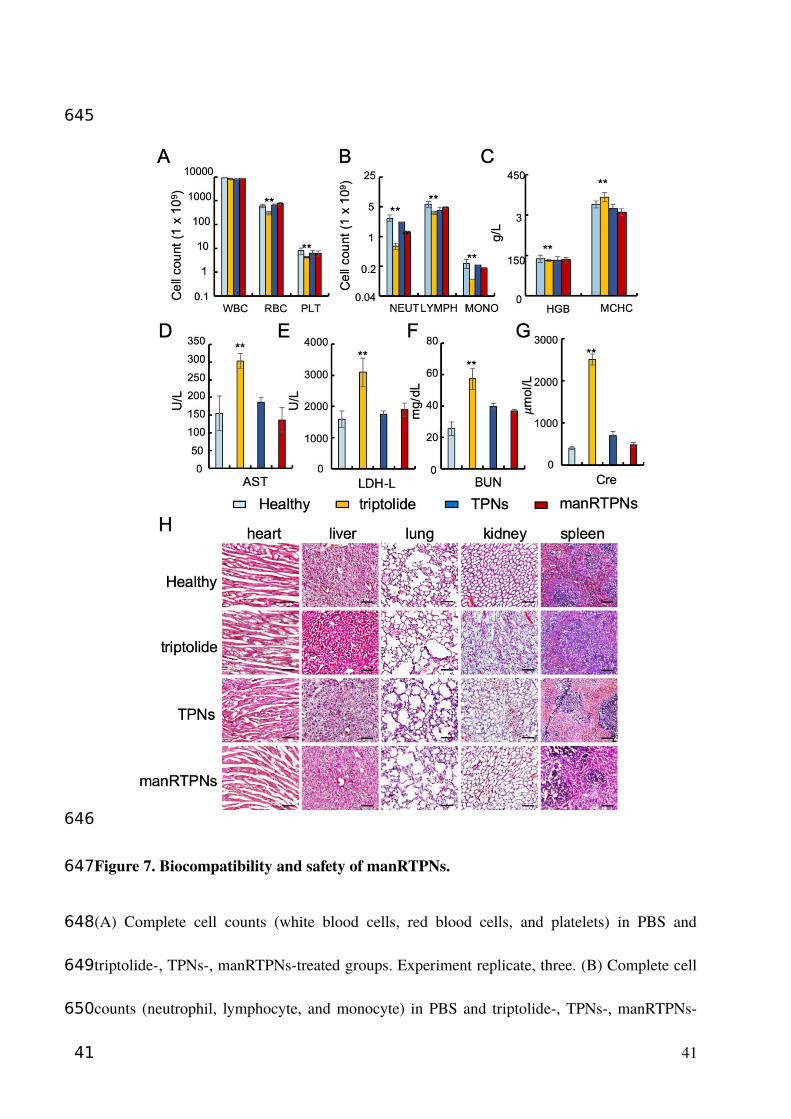
Figure 7. Biocompatibility and safety of manRTPNs.
(A) Complete cell counts (white blood cells, red blood cells, and platelets) in PBS and
triptolide-, TPNs-, manRTPNs-treated groups. Experiment replicate, three. (B) Complete cell
counts (neutrophil, lymphocyte, and monocyte) in PBS and triptolide-, TPNs-, manRTPNs-
41
645
646
647
648
649
650
41

treated groups. Experiment replicate, three. (C) Concentration of HGB and MCHC in four
groups indicated before. Experiment replicate, three. (D-G) Blood routine examination
parameters including AST(D), LDH-L(E), BUN(F) and Cre(G) after different treatment.
Experiment replicate, three. (H) H&E staining of main organs in four different groups. Scale
bars, 100 μm.
42
651
652
653
654
655
656
42

1. Scott DL, Wolfe F, Huizinga TWJ. Rheumatoid arthritis. Lancet. 2010; 376: 1094-1108.
2. Van Vollenhoven RF. Treatment of rheumatoid arthritis: state of the art 2009. Nat Rev
Rheumatol. 2009; 5: 531-541.
3. Bykerk V. Unmet needs in rheumatoid arthritis. J Rheumatol. 2009; 36: 42-46.
4. Salliot C, Van Der Heijde D. Long-term safety of methotrexate monotherapy in patients
with rheumatoid arthritis: a systematic literature research. Ann Rheum Dis. 2009; 68:
1100-1104.
5. Alcorn N, Saunders S, Madhok R. Benefit-risk assessment of leflunomide an appraisal of
leflunomide in rheumatoid arthritis 10 years after licensing. Drug Safety. 2009; 32: 1123-
1134.
6. Leombruno JP, Einarson TR, Keystone EC. The safety of anti-tumour necrosis factor
treatments in rheumatoid arthritis: meta and exposure-adjusted pooled analyses of serious
adverse events. Ann Rheum Dis. 2009; 68: 1136-1145.
7. Hyrich KL, Watson KD, Isenberg DA, Symmons DPM, Register BB. The british society
for rheumatology biologics register: 6 years on. Rheumatology. 2008; 47: 1441-1443.
8. Kirwan JR, Bijlsma JWJ, Boers M, Shea BJ. Effects of glucocorticoids on radiological
progression in rheumatoid arthritis. Cochrane Db Syst Rev. 2007.
9. Ravindran V, Rachapalli S, Choy EH. Safety of medium- to long-term glucocorticoid
therapy in rheumatoid arthritis: a meta-analysis. Rheumatology. 2009; 48: 807-811.
10. Zhang QZ, Dehaini D, Zhang Y, Zhou JL, Chen XY, Zhang LF, et al. Neutrophil
membrane-coated nanoparticles inhibit synovial inflammation and alleviate joint damage
in inflammatory arthritis. Nat Nanotechnol. 2018; 13: 1182-1190.
43
657
658
659
660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
678
43

11. Li RX, He YW, Zhu Y, Jiang LX, Zhang SY, Qin J, et al. Route to rheumatoid arthritis by
macrophage-derived microvesicle-coated nanoparticles. Nano Letters. 2019; 19: 124-134.
12. Liang HY, Peng B, Dong C, Liu LX, Mao JJ, Wei S, et al. Cationic nanoparticle as an
inhibitor of cell-free DNA-induced inflammation. Nat Commun. 2018; 9: 4291.
13. Tao X, Cush J, Garret M, Pe. L. A phase I study of ethylacetate extract of the Chinese
antirheumatic herb Tripterygium wilfordii Hook F in rheumatoid arthritis. J Rheumatol.
2001; 28: 2167.
14. Fan DP, Guo QQ, Shen JW, Zheng K, Lu C, Zhang G, et al. The effect of triptolide in
rheumatoid arthritis: from basic research towards clinical translation. Int J Mol Sci. 2018;
19.
15. Pao HP, Liao WI, Wu SY, Hung KY, Huang KL, Chu SJ. PG490-88, a derivative of
triptolide, suppresses ischemia/reperfusion-induced lung damage by maintaining tight
junction barriers and targeting multiple signaling pathways. Int Immunopharmacol. 2019;
68: 17-29.
16. Chen YK, Zhang L, Ni JS, Wang XY, Cheng J, Li YC, et al. LLDT-8 protects against
cerebral ischemia/reperfusion injury by suppressing post-stroke inflammation. J
Pharmacol Sci. 2016; 131: 131-137.
17. He QL, Minn I, Wang QL, Xu P, Head SA, Datan E, et al. Targeted delivery and sustained
antitumor activity of triptolide through glucose conjugation. Angew Chem Int Edit. 2016;
55: 12035-12039.
44
679
680
681
682
683
684
685
686
687
688
689
690
691
692
693
694
695
696
697
698
44

18. Yuan ZX, Wu XJ, Mo JX, Wang YL, Xu CQ, Lim LY. Renal targeted delivery of
triptolide by conjugation to the fragment peptide of human serum albumin. Eur J Pharm
Biopharm. 2015; 94: 363-371.
19. Mei ZN, Chen HB, Weng T, Yang YJ, Yang XL. Solid lipid nanoparticle and
microemulsion for topical delivery of triptolide. Eur J Pharm Biopharm. 2003; 56: 189-
196.
20. Huang CL, Zeng T, Li JW, Tan LS, Deng XL, Pan YC, et al. Folate receptor-mediated
renal-targeting nanoplatform for the specific delivery of triptolide to treat renal
ischemia/reperfusion injury. ACS Biomater Sci Eng. 2019; 5: 2877-2886.
21. Ling D, Xia H, Park W, Hackett MJ, Song C, Na K, et al. pH-sensitive nanoformulated
triptolide as a targeted therapeutic strategy for hepatocellular carcinoma. ACS Nano.
2014; 8: 8027-8039.
22. Xu HT, Liu B. Triptolide-targeted delivery methods. Eur J Med Chem. 2019; 164: 342-
351.
23. Abbas M, Zou QL, Li SK, Yan XH. Self-assembled peptide- and protein-based
nanomaterials for antitumor photodynamic and photothermal therapy. Adv Mater. 2017;
29.
24. Zhu PL, Yan XH, Su Y, Yang Y, Li JB. Solvent-induced structural transition of self-
assembled dipeptide: from organogels to microcrystals. Chem-Eur J. 2010; 16: 3176-
3183.
25. Yan XH, Zhu PL, Li JB. Self-assembly and application of diphenylalanine-based
nanostructures. Chem Soc Rev. 2010; 39: 1877-1890.
45
699
700
701
702
703
704
705
706
707
708
709
710
711
712
713
714
715
716
717
718
719
720
45

26. Elshabrawy HA, Chen ZL, Volin MV, Ravella S, Virupannavar S, Shahrara S. The
pathogenic role of angiogenesis in rheumatoid arthritis. Angiogenesis. 2015; 18: 433-
448.
27. Kinne RW, Brauer R, Stuhlmuller B, Palombo-Kinne E, Burmester GR. Macrophages in
rheumatoid arthritis. Arthritis Res. 2000; 2: 189-202.
28. Davis SJ, Ikemizu S, Evans EJ, Fugger L, Bakker TR, Van Der Merwe PA. The nature of
molecular recognition by T cells. Nat Immunol. 2003; 4: 217-224.
29. Raychaudhuri S, Thomson BP, Remmers EF, Eyre S, Hinks A, Guiducci C, et al. Genetic
variants at CD28, PRDM1 and CD2/CD58 are associated with rheumatoid arthritis risk.
Nat Genet. 2009; 41: 1313-1318.
30. Sable R, Durek T, Taneja V, Craik DJ, Pallerla S, Gauthier T, et al. Constrained cyclic
peptides as immunomodulatory inhibitors of the CD2:CD58 protein-protein interaction.
ACS Chemical Biology. 2016; 11: 2366-2374.
31. Hu CMJ, Fang RH, Copp J, Luk BT, Zhang LF. A biomimetic nanosponge that absorbs
pore-forming toxins. Nat Nanotechnol. 2013; 8: 336-340.
32. Rossi L, Fraternale A, Bianchi M, Magnani M. Red blood cell membrane processing for
biomedical applications. Front Physiol. 2019; 10.
33. He QL, Titov DV, Li J, Tan MJ, Ye ZH, Zhao YM, et al. Covalent modification of a
cysteine residue in the XPB subunit of the general transcription factor TFIIH through
single epoxide cleavage of the transcription inhibitor triptolide. Angew Chem Int Edit.
2015; 54: 1859-1863.
46
721
722
723
724
725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
46

34. Hu CMJ, Zhang L, Aryal S, Cheung C, Fang RH, Zhang LF. Erythrocyte membrane-
camouflaged polymeric nanoparticles as a biomimetic delivery platform. P Natl Acad Sci
USA. 2011; 108: 10980-10985.
35. Fang RNH, Hu CMJ, Chen KNH, Luk BT, Carpenter CW, Gao WW, et al. Lipid-insertion
enables targeting functionalization of erythrocyte membrane-cloaked nanoparticles.
Nanoscale. 2013; 5: 8884-8888.
36. Shao F, Wang GJ, Xie HT, Zhu XY, Sun JG, A JY. Pharmacokinetic study of triptolide, a
constituent of immunosuppressive chinese herb medicine, in rats. Biol Pharm Bull. 2007;
30: 702-707.
37. Ghosh S, Hayden MS. New regulators of NF-kappaB in inflammation. Nat Rev Immunol.
2008; 8: 837-848.
38. Niedermeier M PT, Korb A. Therapeutic opportunities in fibroblasts in inflammatory
arthritis. Best Pract Res Clin Rheumatol. 2010; 24: 527-540.
39. Brand DD, Latham KA, Rosloniec EF. Collagen-induced arthritis. Nat Protoc. 2007; 2:
1269-1275.
40. Noack M, Miossec P. Selected cytokine pathways in rheumatoid arthritis. Seminars in
Immunopathology. 2017; 39: 365-383.
41. Geusens P. The role of RANK ligand/osteoprotegerin in rheumatoid arthritis. Ther Adv
Musculoskelet Dis. 2012; 4: 225-233.
42. Chen L, Deng H, Cui H, Fang J, Zuo Z, Deng J, et al. Inflammatory responses and
inflammation-associated diseases in organs. Oncotarget. 2018; 9: 7204-7218.
47
742
743
744
745
746
747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
763
47

download fileview on ChemRxivmanuscript-update 20200804.docx (7.30 MiB)

1
Triptolide self-assembling nanoparticles engineering with 1
modified erythrocyte membranes for targeting and remodeling 2
inflammatory microenvironment in arthritis 3
Jing Li1, Sanpeng Li1, Chunbin Li1, Hongfeng Li1, Chuangjun Liu1, Qi Zhao3, Pengfei 4
Zhang1*, Ping Gong1,2*, Lintao Cai1* 5
1Guangdong Key Laboratory of Nanomedicine, CAS-HK Joint Lab for Biomaterials, 6
Shenzhen Institutes of Advanced Technology, Chinese Academy of Sciences, Shenzhen 7
518055, China 8
2Guangdong Key Laboratory for Research and Development of Natural Drugs, Guangdong 9
Medical University, Dongguan, 523808, China 10
3 Faculty of Health Sciences, University of Macau, Taipa, Macau, China. 11
12
*Corresponding author: Lintao Cai: E-mail: [email protected]; 13
Ping Gong: E-mail: [email protected] 14
15

2
Abstract 1
Overview: Rheumatoid arthritis (RA) is an autoimmune disease characterized by persistent 2
synovitis, systemic inflammation and causing severe joint damage. Inflammation and influx of 3
immune cells is a hallmark of the disease. Triptolide with great immunosuppressive activities 4
is a potential drug to treat RA. However, its severe toxicity is still an unresolved bottleneck and 5
largely limits its clinical use. Recently, nanoparticle (NP)-based drug delivery systems have 6
been developed for detoxification of toxic drugs and diseases management. 7
Aim: To synthesize triptolide nanoparticles to decline its toxicity and improve water-solubility 8
and develop a dual targeted biomimicking delivery system which targets to inflammation 9
microenvironment. Finally, to achieve excellent therapeutic effect in rheumatoid arthritis. 10
Methods: We synthesized an amphiphilic molecule prodrug with the addition of 11
diphenylalanine peptide (FF) to triptolide. These prodrugs were self-assembled into 12
nanoparticles (TPNs) in water, then TPNs were coated with mannose-modified erythrocyte 13
membranes to form a dual targeting manRTPNs. Specifically, mannose and natural molecule 14
CD58 on RBC membrane guide it towards activated macrophages and T cells in inflamed joint 15
through innate and specific immune recognition, respectively. Different cell lines including 16
RAW264.7, CTLL-2, RSC364 and hFLS-RA and a collagen induced arthritis (CIA) mice 17
model were used to detect the targeting and therapeutic effect in vitro and vivo. 18
Results: manRTPNs showed an excellent dual targeting ability to activated macrophages and 19
T cells in vitro. We found that manRTPNs selectively induced anomalous activated cells death 20
and indeed altered genes expression and cytokines production of inflamed cells such as 21

3
macrophages, T cells and synoviocytes. In CIA mice, manRTPNs exhibited significantly 1
targeting effect and prolonged drug accumulation at inflamed joints, whereby increased drug 2
achieved significant therapeutic effect without adverse effect. In detail, manRTPNs ameliorated 3
joint destruction, swollen paws considerably shrunk to normal, bone and cartilage destruction 4
were suppressed and bone density was even recovered to healthy level. Basically, several core 5
genes like NF-κB, cox-2, rank, rankl, vegf, vegfr2, mmps, and cytokines such as IL-6, TNF-α 6
involved in inflammation, cartilage/bone destruction and angiogenesis were regulated and 7
altered expression and production. Thus, the inflammation microenvironment was remodeled 8
by TPNs. 9
Conclusion: This study demonstrated phenylalanine dipeptide mediating triptolide self-10
assembled nanoparticles to solve current problems and the potential of engineered RBC 11
membranes for inflammation-targeted drug delivery. Therefore, our work is a good example of 12
triptolide reconstruction, and this agent based on immune recognition may provide an immune 13
cell targeted strategy for rheumatoid arthritis therapy. 14
Keywords: triptolide, nanodrug, dual targeting, inflammatory microenvironment, rheumatoid 15
arthritis 16
17
4
Graphical Abstract 1
2
A novel triptolide nanodrug based therapeutic agent for RA management that modified 3
erythrocyte membranes as a develiry system capable of dual targeted and triptolide remodeling 4
inflammatory microenvironment in arthritis. 5
6

5
Introduction 1
Rheumatoid arthritis (RA) is a serious long-term disease characterized by persistent 2
synovitis, systemic inflammation and autoantibodies and causing severe joint damage, 3
disability and decreased quality of life. The etiopathogenesis remains elusive, and viable and 4
effective treatment options are still limited[1]. Over the last two decades, long-term 5
management of RA has involved disease-modifying anti-rheumatic drugs (DMARDs), 6
biological agents such as tumor necrosis factor (TNF) inhibitors, and glucocorticoids[2]. 7
Although the highest clinical remission rate achieved within 50% of control, not all patients 8
attain desirable levels of clinical remission[3] and clinical use of these therapies is limited 9
because of their high cost and frequency of adverse effects. The latter includes teratogenicity 10
and hepatotoxicity of DMARDs like methotrexate[4, 5], risks of infections such as tuberculosis 11
and osteoporosis caused by biological agents[6, 7] and long-term adverse reactions to 12
glucocorticoid therapy[8, 9]. Lately, new therapies based on inflammatory cells such as 13
macrophages and neutrophil or their products such as cell-free DNA in inflamed joint have 14
been reported[10-12], however inhibiting a single factor may not be enough to sustainably halt 15
or reverse disease progression due to RA’s complexity and heterogeneity. Therefore, a new 16
systemic design for drug candidate based on nano-medicine is urgently needed. 17
Triptolide is a trace natural triepoxide diterpene from Tripterygium wilfordii (or Léi Gōng 18
Téng) and has been widely used to treat for RA patients since the 1970s in China[13]. Despite 19
its various biological activities, low water solubility and severe toxicity, especially to the liver 20
and kidney, remarkably impedes the translation to clinical applications[14]. Considerable 21

6
efforts in structural modification[15-18] and nanoparticle(NP) -based drug delivery system [19-1
21] have been applied to reduce its toxicity. Unfortunately, common practices that direct 2
conjugations attached to the C14-hydroxyl group of triptolide via a hydrolytic ester bond have 3
proven biologically unstable; meanwhile, current targeted drug-loading delivery systems are 4
limited by drug loading or encapsulation ratio and they mostly target to kidney or tumor[22]. 5
Therefore, an effective and stable system that can deliver triptolide to other organ or tissue such 6
as joint is necessary. But until now, triptolide-formed nanoparticles have been rarely reported 7
before. From this point, inspired from biomacromolecule self-assembly process, various 8
peptide building blocks have been performed for the creation of biomimetic or bioinspired 9
nanostructured materials[23, 24]. Recently, diphenylalanine (FF) is the simplest peptide 10
building block for self-assembly and exhibits remarkable advantages including 11
biocompatibility and non-immunogenicity and so on. Many researchers have developed FF-12
based nanomaterials such as nanotubes, spherical vesicles, nanofibrils and nanowires[25]. 13
However, it is barely to link it with toxic drug for nanodrug formation. Therefore, 14
diphenylalanine (Phe-Phe, FF) mediating self-assembling nanoparticles is a new strategy to 15
reconstruct triptolide to be nanoparticles whose spherical nanostructure was more stable and 16
biocompatible than before. 17
In RA pathogenesis, numerous blood-derived cells infiltrate in synovium and cytokine 18
secret necessitates formation of new blood vessels to generate pannus, that all of these form an 19
inflammatory microenvironment[26]. Peripheral-blood monocytes are recruited to joint, and 20
functionally diverse macrophages distribute in the synovial sublining and lining layers. It is 21

7
mentioned that Cluster of Differentiation 206 (CD206) on macrophages surface has been 1
induced to express more [27]. Besides, prominent T cells being activated, especially the CD4+ 2
subset, also infiltrate the synovium. As reported, certain signal proteins on the T cell surface 3
such as lymphocyte function antigen 2 (LFA-2/CD2) have increased in both RA patients and 4
animal models[28, 29]. It was reported that CD2 can be effectively targeted by peptides derived 5
from LFA-3/CD58 as well as fusion protein constructs. Interestingly, the ligand of CD2 namely 6
CD58 widely distributes on erythrocyte membranes and CD58-CD2 interaction have mediated 7
process of T cells activation[30]. These immunological characteristics of erythrocytes scarcely 8
ever attracts attention in application, so it is a valuable attempt to apply in drug targeting. Then, 9
mannose, the ligand of CD206 was introduced in erythrocyte membrane for ligand-binding 10
targeting. In addition, the long circulation and flexible functionalization of erythrocyte 11
membrane also make it be a promising platform and have wide application[31, 32]. In this 12
study, we prepared mannose-labeled RBC membrane to entrap newly synthesized triptolide 13
nanoparticles (TPNs) for elevating its vast retention in inflammation and achieving remarkably 14
anti-inflammatory therapy. Triptolide, a hydrophobic drug, was linked with diphenylalanine 15
peptide to self-assembly into size-controlled nanoparticles (TPNs) in water, and this 16
reconstruction has successfully decreased its toxicity and improved water-solubility. As shown 17
in scheme 1, the nanodrug core was coated with decorated RBC membranes to obtain mannose-18
modified RBC cell membrane-coated triptolide nanoparticles (manRTPNs). By translocating 19
RBC membranes to the nanoparticles, the biological targeting properties of RBC cell can be 20
conferred to manRTPNs as well, allowing them to target activated T cells through the CD58-21

8
CD2 affinity. Further, the binding interaction between the mannose and its receptor CD206 1
enhanced anchoring of manRTPNs to activated macrophages. According to manRTPNs, the 2
dual-targeting strategy facilitated the accumulation of nanodrug in inflammation, and the 3
accumulated TPNs efficiently achieved systemic administration of rheumatoid arthritis by 4
specifically inducing cell death and remodeling the inflammatory microenvironment through 5
affecting genes network, downregulating cytokines and pro-angiogenic factors, regulating 6
osteoclastogenesis, and returning balance of metalloproteinases and their inhibitors. Overall, 7
our study expanded the RBC cell membrane biological application in nanocarrier for targeting 8
inflammation through innate and specific immune recognition, and the triptolide nanodrug 9
showed high drug content, low side effect, good biocompatibility which showed great potential 10
for further inflammation diseases therapy. 11
Results and Discussion 12
Preparation and characterization of manRTPNs 13
Triptolide is a diterpene triepoxide with several active sites that can be modified, 14
especially C14 position[33]. Because of its hydrophobicity, triptolide molecule acts as self-15
assembly inducer with the addition of diphenylalanine peptide (FF), which is the key moiety of 16
the self-assembling peptide-drug conjugate. Initially, we added a glucosamine (AG) to the FF 17
terminal to lengthen the hydrophilic chain, and then linked triptolide to FF-AG by an ester bond 18
whose cleavage led to drug release in the cell; thus, the self-assembling monomer (TP-FF-AG) 19
as a triptolide prodrug was synthesized (Figure 1A). The chemical structures of these 20
compounds were confirmed via mass spectrometry (MS) and nuclear magnetic resonance 21

9
(NMR; Figure S1-S16). Due to hydrophobic interactions, triptolide nanoparticles (TPNs) were 1
formed via self-assembly of synthesized monomer namely TP-FF-AG, which is uniformly easy 2
to control TPNs size in water solution. The particles morphology was observed via transmission 3
electron microscopy (TEM; Figure 1B). To synthesize manRTPNs, RBC membranes were 4
extracted by hypotonic hemolysis[34], and mannose inserted by PEG2000-DSPE. An extruder 5
was using to camouflage mannose-modified erythrocyte membranes onto TPNs then form a 6
core-shell structure (Figure 1B). Dynamic light scattering (DLS) measurements revealed that 7
the TPNs were 135 nm in hydrodynamic diameter and manRTPNs were about 10 nm larger 8
with PDIs of 0.15-0.2. The surface zeta potential of the shell was not as negative as that of the 9
cores but was comparable to that of erythrocyte membrane-derived vesicles (RBC vesicles) 10
(Figure 1C). All these results demonstrated successful cloaking of TPNs with erythrocyte 11
membranes. 12
Next, TPNs and manRTPNs were stored in phosphate-buffered saline (PBS) for 1 month, 13
where they demonstrated a negligible change in size and showed an excellent stability (Figure 14
1D). Maintaining protein compounds of erythrocyte membranes on nanoparticles’ surfaces is 15
vital for harnessing their biofunctionality. Therefore, completeness of the membrane coating 16
was verified via sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE; 17
Figure S17), which indicated the manRTPNs maintained the whole effective biocomponent of 18
erythrocyte on the surface. Translocation of erythrocyte membranes might improve the 19
biocompatibility and circulating period of TPNs in vivo, which also sustains systemic delivery 20
and better targeting through both passive and active uptakes[35]. As reported, the elimination 21

10
T1/2 of triptolide molecules in rats following i.v. administration was 15-21 minutes, which 1
showed that drugs were rapidly eliminated[36]. However, TPNs and manRTPNs have 2
maintained higher plasma concentration after tail intravenous injection of mice in our work. As 3
the relative signal of drug down to 50%, TPNs lasted 4 h, and manRTPNs even had a much 4
longer elimination half-life of over 48 hrs (Figure 1E), which suggested that manRTPNs 5
significantly expanded blood circle compared to triptolide. Therefore, the self-assembling 6
TPNs nanostructure was more stable than original drug, and manRTPNs inherited biological 7
properties of erythrocyte membrane proteins for following efficacy. 8
Dual-Targeting capacity and immunosuppressive effect of manRTPNs 9
To determine each targeting capacity of manRTPNs to macrophage and T cell in vitro, 10
nanoparticles were Cy5.5-labelled and incubated with RAW264.7 and CTLL-2 treated with IL-11
4 (100 ng/mL) or TNF-α (10 ng/mL), untreated cells and RSC364 as control. Flow cytometry 12
(FCM) analysis observed gradual increased uptake of nanoparticles by RAW264.7 and CTLL-13
2 for 6 hrs (Figure S18-20). An uptake curve was built using mean fluorescence intensity (MFI) 14
of cells at six time points, and cellular uptake rate was defined as k indicated in figure 2A-C, 15
which was the slope of the uptake curve. In detail, ks of nanoparticles in activated RAW264.7 16
and CTLL-2 were significantly higher than that in untreated cells (Figure 2A-C; Figure S21A 17
& S22), and ks of manRTPNs were significantly higher than that to TPNs both in RAW264.7 18
and CTLL-2 (Figure 2A-B), but no difference of ks existed between TPNs and manRTPNs in 19
RSC364 that no targeting ligands on it (Figure 2C). Further, the k value in RAW264.7 was 20
higher than that in CTLL-2 (Figure 2A-B), indicating the affinity difference of interacted 21

11
molecule pairs. To further confirm ligand-binding specificity, Cy5.5-labelled TPNs (without 1
mannose and CD58), RTPNs (with CD58) and manRTPNs (with mannose and CD58) were 2
incubated with treated macrophages and T cells for 3 hrs as well, RSC364 and hFLS-RA as 3
control. After treatment, CLSM images showed that CD206 and CD2 (Figure 2D-E, green) 4
respectively overexpressed on surface of RAW264.7 and CTLL-2. Significant red fluorescence 5
was observed on RAW264.7 incubated with manRTPNs, but not with TPNs and RTPNs (Figure 6
2D), while red fluorescence was observed on CTLL-2 incubated with RTPNs and manRTPNs 7
but not with TPNs (Figure 2E), which was consistent with corresponding ligands distribution 8
on the nanoparticles. When co-incubated with RSC364, they also entered cells (Figure 2F; 9
Figure S21b). Similar results were observed in flow cytometry analysis (Figure 2G-I; Figure 10
S21c). These results demonstrated the ability of manRTPNs to target inflamed cells conferred 11
by their mannose modified erythrocyte membrane coating, which is probably attributed to 12
specific affinity of mannose-CD206 and CD58-CD2. 13
Except for migrated immune cells, synovial fibroblast is main resident effector of 14
inflammation. To determine immunosuppressive efficacy of manRTPNs in joint inflammatory 15
cells, LPS-treated RAW264.7, TNF-α-treated CTLL-2 and RSC364, and hFLS-RA were 16
incubated with manRTPNs in 96-well plates for 24 hrs. Firstly, cell viability was measured via 17
MTT assay. TPNs and manRTPNs didn’t significantly reduce RAW264.7 viability even at a 18
concentration of 500 nM (Figure 3A). However, at the same concentration CTLL-2, RSC364 19
and hFLS-RA showed an obvious cytotoxicity (Figure 3B-D). Specially, manRTPNs had an 20
IC50 (half maximal inhibitory concentration) values of 219 nM for activated CTLL-2 cytoxicity, 21

12
88.9 nM for activated RSC364 cytoxicity and 95.8 nM for hFLS-RA cytoxicity (Figure 3B-D). 1
The lower IC50 values of activated RSC364 and hFLS-RA indicated that triptolide-induced 2
cell death was mainly against anomalous proliferative synoviocytes in inflamed joints, rather 3
than lymphocytes like macrophages. Cells were following collected and lysed for mRNA 4
extraction to check certain inflammation associated genes variation by qPCR. For LPS-treated 5
RAW264.7, transcription factor NF-κB associated with most cytokine transcription[37] and 6
cyclooxygenase-2 (cox-2) were induced to decrease expression level, but tissue inhibitor of 7
metalloproteinase-1 (timp-1) showed opposite variation under manRTPNs treatment. Receptor 8
activator of NF-κB ligand (rankl) over-expressed on TNF-α-treated CTLL-2 was also 9
downregulated by manRTPNs. After activated with cytokines and chemokines, synovial 10
fibroblasts release matrix-degrading enzymes, including matrix metalloproteinases (MMPs) 11
and cathepsins causing cartilage and bone destruction[38]. In RSC364, the transcripts of 12
metalloproteases such as MMP-2 and MMP-9 and vascular endothelial growth factor (VEGF) 13
being critical for angiogenesis were lessen by manRTPNs as well (Figure 3E). Thirdly, medium 14
supernatants were collected to measured cytokines by ELISA assay. supernatant IL-1β and IL-15
6 of LPS-treated RAW264.7 and supernatant IFN-𝛾 of TNF-α-treated CTLL-2 lessened more 16
quickly that treated with manRTPNs than TPNs (Figure 3F-H), while supernatant IL-1β and 17
IL-6 of TNF-α-treated RSC364 had a similar decreasing curve between TPNs and manRTPNs 18
groups (Figure 3I-J), which indicated that manRTPNs had superior identification of immune 19
cells than resident synoviocytes at least in cytokine secretion. Consequently, manRTPNs 20
selectively exerts its immunosuppressive effects on selectively inducing lymphocytes and 21

13
synovial fibroblasts cell death, but also regulating production of proinflammatory cytokines, 1
proinflammatory mediators and matrix metalloproteinases, which suggested manRTPNs may 2
have an ability to remodel inflammatory microenvironment through its cell-specific anti-3
inflammation efficacy. 4
Targeting capacity and accumulation of manRTPNs in vivo 5
Upon the good capacity of manRTPNs to target activated macrophages and T cells in vitro, 6
we further verified their ability to target RA site in vivo. After CIA mouse model being induced, 7
all four paws swelled in varying degrees[39]. Hind paws were more heavily swollen, and micro-8
computed tomography (Micro-CT) imaging showed distinct bone loss of joint therein (Figure 9
4A). Then these mice received intravenous injections of free cyanine 5.5 (Cy5.5) solution and 10
Cy5.5-labelled TPNs and manRTPNs for targeting imaging in vivo. As shown in figure 4A, 11
almost no fluorescence signal was detected in paws of the free-Cy5.5 group, indicating that the 12
dye barely accumulated at local inflammatory sites. In the TPNs group, we detected a weak 13
signal, which peaked after 4 h (Figure 4A). While manRTPNs obviously reached the RA sites 14
and lasted there for ≥12 hrs, which would be attributed to their long circulation and the targeting 15
effect of the modified erythrocyte membranes. Thus, significant fluorescence of manRTPNs 16
lasted longer and showed greater accumulation than that of TPNs. 24 hrs after intravenous 17
injections, we dissected mouse and got froze sections of left paws to observe nanoparticles’ 18
retention in the inflamed joints. Contrary to free-Cy5.5 and TPNs group, fluorescence 19
microscopy indicated a very high fluorescence intensity of manRTPNs in the synovial sublining 20
layer, resulting in a reduced accumulation at the inflamed joint due to the lack of permeability 21

14
(Figure 4B). Tissue biodistribution further revealed manRTPNs’ outstanding capacity to target 1
the paws of RA mice, rather than other groups (Figure 4C). Additionally, higher accumulation 2
was observed in liver, suggesting that this organ might be passive targeting of nanoparticles 3
and the main organ metabolizing triptolide (Figure 4C). The results confirmed that the effective 4
accumulation of manRTPNs provided a promising precondition of RA treatment. 5
Therapeutic efficacy of manRTPNs 6
The efficacy of ameliorating joint damage of manRTPNs was evaluated in a CIA mouse 7
model. Following arthritis induction, CIA mice presented symptoms of swelling in ankle and 8
paw joints and a clinical scoring was used to estimate the swelling degree (Figure S23). 9
According to protocol timeline, CIA mice were treated with intravenous injections of triptolide 10
at 15 mg/kg, TPNs and manRTPNs at doses of 25 mg/kg for 11 days (Figure 5A). Surprisingly, 11
swollen hindpaws and forepaws both shrunk considerably and even recovered to the same 12
extent as healthy ones with manRTPNs treatment rather than slightly shrunk in TPNs and 13
triptolide group, but swelling symptoms of other groups were increasingly severe (Figure 5B), 14
implying that inflammation process was exactly controlled by manRTPNs. As paw swelling is 15
only a surface phenomenon of RA, bone loss was further determined using Micro-CT analysis. 16
Reconstructed three dimensional (3D) bones images and calculated bone mean density (BMD) 17
showed that serious bone erosion was suppressed and bone density even recovered to healthy 18
mice level of manRTPNs treatment (Figure 5C-D), suggesting a protective role of manRTPNs 19
on cartilage and bone damage. Haemotoxylin and eosin (H&E)-stained sections showed an 20
intense anomalous proliferative cells infiltration in the joints and synovium of control groups. 21

15
By contrast, remarkable reduction of immune infiltration and preservation of cartilage were 1
observed in manRTPNs group (Figure 5E). Further, cytokines including TNF-α and IL-6 are 2
known to increase with the onset of arthritis and correlate strongly with disease severity[40]. 3
Immunohistochemical analysis showed that TNF-α and IL-6 in synovium and articular cavity 4
significantly declined in manRTPNs groups (Figure 7A-B) and serum levels of them also 5
observed to alleviate similarly (Figure 6C), indicating effective reduction of arthritis at 6
cytokines level. Moreover, gene heatmap showed that manRTPNs suppressed several genes 7
expression including rank, rankl, vegf, vegfr2, NF-κB, cox-2, IL-1β, mmp-2, mmp-3, mmp-9 8
and mmp-13, that these transcripts are dramatically increased and contribute in joints damage. 9
Some negative regulators such as opg, timp1 and timp2 were upregulated their gene expression 10
by manRTPNs (Figure 6D), indicating manRTPNs preserve appropriate and balanced 11
concentration of cytokines and inflammatory factors through two-way regulation. 12
RANKL/RANK/ORG signal pathway is key of bone metabolism[41]. The up-/down-regulation 13
of manRTPNs also recovered the balance of RANKL/RANK/ORG and metalloproteinases and 14
their inhibitors (TIMP) to normal. Thus, manRTPNs ameliorated destroyed through aiming to 15
RA pathological factors including cytokines, extracellular matrix (ECM), bone metabolism and 16
angiogenesis. 17
Biosafety of manRTPNs nanoparticles 18
To investigate safety and biocompatibility of TPNs and manRTPNs in vivo, we 19
administered PBS, TPNs or manRTPNs at a single dose of 25 mg/kg to healthy mice via tail 20
vein, while to other mice we administered triptolide (original drug of TPNs and manRTPNs) at 21

16
15 mg/kg by intraperitoneal injection. It is worth mentioning that most mice in the triptolide 1
group died soon (within 24 hrs) after 25 mg/kg administration, indicating the high toxicity of 2
the original drug (Figure S24). After 1-week treatment, mice treated with triptolide showed a 3
significant decrease in count of white blood cells (WBC), RBCs and platelets (Figure 7A), 4
suggesting the occurrence of bone marrow suppression, which is one of the main side effects 5
of triptolide. The main three type cells comprised WBC including neutrophils, lymphocytes, 6
and monocytes showed significant count differences between triptolide group and other groups, 7
which explained the WBC count variation conferred from triptolide (Figure 7B). The 8
hemoglobin concentration did not differ significantly among all the groups in the blood (HGB), 9
mean corpuscular hemoglobin (MCHC) level, and hemoglobin distribution width (HDW) 10
(Figure 7C), indicating that triptolide didn’t influence the physiological function of RBCs. 11
Blood biochemistry analysis showed that blood urea nitrogen (BUN), aspartate 12
aminotransferase (AST), creatinine (Cr) and lactate dehydrogenase (LDH) levels of mice 13
treated with triptolide were significantly higher than those of mice in the TPNs and manRTPNs 14
groups (Figure 7D-G). These variations demonstrated that the hepatotoxicity and 15
nephrotoxicity of triptolide were greatly reduced in TPNs, with RBC membrane coating 16
yielding even better safety. Hematoxylin and eosin (H&E) staining of major organs showed 17
that triptolide treatment caused inflammatory infiltration of the liver and severe renal injury 18
characterized by loss of the brush border, tubular epithelial cells detachment from the basement 19
membrane and tubular obstruction. Meanwhile, the TPNs and manRTPNs groups showed no 20

17
significant variation in cellular morphology and negligible organ damage, in line with the PBS 1
group (Figure 7H). However, TPNs and manRTPNs significant reduced triptolide toxicity. 2
Conclusion 3
Our study expanded the RBC cell membrane biological application in nanocarrier for 4
targeting inflammation through innate and specific immune recognition, and the triptolide 5
nanodrug showed high drug content, low side effect, good biocompatibility which showed great 6
potential for further inflammation diseases therapy. Generally, there are two improvement of 7
manRTPNs in cell-membrane-coated nanoparticle delivery system, namely triptolide nanodrug 8
core and RBC membrane shell. RBC membrane-camouflaging nanoparticles has been 9
extensive established platform, but there are still many deposits from RBC cell itself to further 10
develop disease-targeting and therapy platform except for tumor. The natural immunological 11
function of RBC cells has been always neglected in targeted drug delivery construction. So, 12
taking advantage of adhesion molecules affinity between RBC cells and immunocytes in 13
building nanocarrier for transporting drug to inflammation deeply expands application of 14
established RBC membrane platform. On the other side, triptolide has excellent anti-15
inflammation and immunosuppressive effect and solving its low water-solubility and toxicity 16
is premise of using it in various diseases management. Our reconstruction of triptolide isn’t 17
content-limited loading in micelles, but drug-based self-assembled size-controlled 18
nanoparticles. This strategy takes advantages of drugs’ hydrophobicity to solve its bad water-19
solubility, which is magic of nanotechnology. While inflammation is prevalent related with 20
many diseases like cardiovascular diseases, gastrointestinal tract, cancer and pathogen 21

18
infection[42]. Therefore, manRTPNs as a drug delivery system of inflammation targeting and 1
treating may be a promising agent in other inflammation-associated diseases therapy. 2
Materials and methods 3
Animal care 4
Mice were housed in an animal facility at Shenzhen Institutes of Advanced Technology, 5
Chinese Academy of Sciences (SIAT). All animal experiments were performed in accordance 6
with Guidance Suggestions for the Care and Use of Laboratory Animals and approved by 7
Sciences Animal Care and Use Committee of SIAT. 8
Erythrocyte membranes derivation 9
Erythrocyte membranes were collected according to a previously reported method with 10
modification. Briefly, fresh heparinized whole blood was collected from male mice (20-22 g) 11
and subsequently centrifuged at 2000 rpm for 15 min at 4 °C to remove the plasma and the 12
white buffy coat. The collected red blood cells (RBCs) were washed with 1× PBS three times 13
and suspended in 0.25× PBS for 2 hours at 4 °C, and then the hemoglobin was removed by 14
centrifugation at 9,000 rpm for 45 min. The resulting pink pellet was purified with 1× PBS, and 15
the collected erythrocyte membrane was suspended and stored in distilled water. To form 16
ligand-inserted erythrocyte ghosts, the light pink solution was incubated with DSPE-PEG-17
mannose for 30 min, 37°C water bath. Erythrocyte membranes with PEG-mannose or not was 18
further prepared by extruding through a 200 nm polycarbonate porous membrane with an 19
Avanti mini-extruder (Avanti Polar Lipids). 20

19
Synthesis of TPNs 1
The TPNs were synthesized by a prodrug self-assembly method. Briefly, after dissolving 2
5mmol Boc-Phe-Phe-OH (BOC-FF, J&K scientific Ltd., China), 6 mmol 1-Ethyl-3-(3-3
dimethylaminopropyl) carbodiimide (EDC, J&K scientific Ltd., China) and 6 mmol N-4
Hydroxysuccinimide (NHS, J&K scientific Ltd.,China) in 3 mL anhydrous DMF, another 5
anhydrous DMF solution containing 5.5 mmol glucosamine (AG, Sigma-Aldrich, USA) and 5 6
mmol TEA (J&K scientific Ltd., China) was added drop-wise into it. The mixture was 7
evaporated under reduced pressure for 24 hours to get crude product BOC-FF-AG. Then, crude 8
BOC-FF-AG was dissolved in 30 mL equal volume mixture of anhydrous dichloromethane and 9
trifluoroacetic acid (TFA, J&K scientific Ltd., China) and stirred for 4 hours at room 10
temperature. Afterward, pure NH2-FF-AG was obtained after evaporated under reduced 11
pressure and purified by silica gel column chromatography with dichloromethane and 12
methanol. Secondly, 1 mmol triptolide (TP, Shanghai Yuanye Bio-Technology Co., Ltd., 13
China), 1.2 mmol succinic anhydride (SAA, Sigma-Aldrich, USA) and 0.2 mmol dimethyl 14
aminopyridine (DMAP, Sigma-Aldrich, USA) were dissolved in 3 mL pyridine and stirred 15
overnight. After then, the mixture was diluted with ethyl acetate and washed it with solution 16
containing saturated copper sulfate (J&K scientific Ltd.), water and brine. After reaction, 17
residue TP-COOH was obtained through drying with anhydrous Na2SO4 and concentration with 18
rotary evaporator. In the third step, TP-COOH, dicyclohexylcarbodiimide (DCC, Sigma-19
Aldrich, USA) and 1-hydroxy-5-pyrrolidinedione (NHS, J&K scientific Ltd., China) of equal 20
molar ratio were dissolved into 3 mL anhydrous dichloromethane and stirred at room 21

20
temperature for overnight. The final product TP-FF-AG was gained by reaction of activated 1
TP-COOH and NH2-FF-AG for overnight at room temperature and purified by analytical RP-2
HPLC and semi-preparative RP-HPLC with simultaneous detection at 265 nm. Analytical RP-3
HPLC was performed at room temperature on the Shimadzu LC 20 with UV detector SPD-20A 4
using Inertsil ODS-SP column (4.6 x 250 mm, 5 µm, 100Å). The RP-HPLC gradient was started 5
at 10% of B (CH3CN, J&K scientific Ltd., China), then increased to 100% of B over 30 min 6
(A: 0.1% TFA in water). Semi-preparative RP-HPLC was performed on the ULTIMAT 3000 7
Instrument (DIONEX). For fluorescence imaging experiments, cyanine5.5 (Cy5.5, Lumiprobe, 8
USA) was linked to intermediate product of NH2-FF-AG after esterification reaction to obtain 9
Cy5.5-FF-AG in the same way. Ultimately, nanoparticles were self-assembled by added 10
TP/Cy5.5-FF-AG dissolved in DMSO dropwise into PBS while vigorous stirring and dialysis 11
for 2 days. 1H NMR and 13C NMR spectra were recorded with a Bruker VANCE III400 12
spectrometer (400 MHz). The high-resolution mass spectra (HR-MS) were measured on a 13
Bruker Micro TOF II 10257 instrument. For membrane coating, nanoparticles were mixed with 14
prepared membranes in ratio of 1:1. The resultant mixture was subsequently extruded nine 15
times through a 200 nm polycarbonate porous membrane using an Avanti mini-extruder to yield 16
the erythrocyte membranes-coating TPNs or mannose inserted erythrocyte membrane-coating 17
TPNs (namely RTPNs or manRTPNs). 18
Nanoparticles Characterization 19
The hydrodynamic diameter and zeta potential of NPs suspended in 1 × PBS were 20
measured by dynamic light scattering (DLS) (Zeta Plus, Brookhaven Instruments, USA). The 21

21
morphologies of TPNs and manRTPNs were observed by transmission electron microscope 1
(TEM, FEI spirit T12) at an accelerating voltage of 120 keV. The stability experiment of TPNs 2
and manRTPNs in PBS was monitored by DLS over five weeks and stored at 4°C. The proteins 3
retained on manRTPNs compared with those on the natural erythrocyte membrane were 4
observed by SDS-PAGE. Briefly, 2 mg of erythrocyte membrane vesicles and manRTPNs were 5
collected by centrifugation at 15,000g for 30 min, mixed in SDS sample buffer (Invitrogen, 6
USA), and heated at 90 °C for 5 min. Then, 20 μL of each sample was run on a 10% SDS-7
polyacrylamide gel (Bio-Rad, SDS-PAGE Gel Preparation Kit, USA) at 120 V for 1 h, followed 8
by Coomassie blue staining and imaging. 9
Pharmacokinetics 10
To evaluate the circulation half-life of TPNs and manRTPNs in vivo, 150 μL Cy5.5-11
labeled nanoparticles were injected into the tail vein of the mice (n = 3). Twenty microliters of 12
blood were collected at 1, 5, 15, 30 min, and 1, 2, 4, 8, 24, 48, and 72 h following the injection. 13
Each particle group contained 3-4 mice. The collected blood samples were diluted with 30 μL 14
PBS in a 96-well plate before fluorescence measurement. Pharmacokinetics parameters were 15
calculated to fit a two-compartment model and a one-way nonlinear model. 16
Drug toxicity analysis in vivo 17
Purchased from Vital River Laboratory Animal Technology Co. Ltd (Beijing, China), 18
males BALB/c mice (6∼8 weeks old) were randomly divided into four groups and 19
intravenously injected with PBS, TPNs and manRTPNs at a single dosage of 25 mg/kg and 20
triptolide of 15 mg/kg 4 times for eight days. A total of 7 days after last administration, all mice 21

22
were euthanized, and blood was collected for and blood routine examination and biochemical 1
parameters measurement, while main organs including heart, liver, spleen, lung, and kidney 2
were excised and stained by H&E for histological analysis. 3
Cell culture 4
RAW264.7 cells, CTLL-2, RSC364 and hFLS-RA were purchased from the American 5
Type Culture Collection (ATCC, USA) and cultured in Dulbecco’s minimum essential medium 6
(DMEM, Corning, USA) supplemented with 10% fetal bovine serum (FBS, Gibco, USA), 1% 7
penicillin (100 IU/mL, Corning, USA), and streptomycin (100 μg/mL, Corning, USA) and 8
placed at 37 °C in a 5% CO2 humidified atmosphere. Cell lines were purchased with 9
certification of authentication and free from Mycoplasma. 10
Quantification of cell targeting in vitro 11
A 6-well culture plate was prepared and RAW264.7, CTLL-2, RSC364 and hFLS-RA 12
cells were implanted at a density of 5 × 105 cells/well. 12-16 hrs later, 80% of cell coverage was 13
confirmed. 50 𝜇L Cy5.5-labelled TPNs or manRTPNs was adding into prepared cells every 14
hour for six hours, incubated with covering foil paper and collected them in the sixth hour. 15
After digestion with trypsin (Gibco, USA), cells were put together and washed three times with 16
ice-cold PBS, then underwent flow cytometry by FACS Canto II (BD Biosciences). The data 17
was analyzed using FlowJo software. Mean fluorescence intensity (MFI) reflected amount of 18
intake nanoparticles amount. An uptake curve was constructed with MFI and time, and a k value 19
being the slope of uptake curve was defined as uptake rate to measure targeting capacity of 20
nanoparticles. 21

23
Immunofluorescence assay of macrophages and T cells with nanoparticles 1
Cells were cultured with medium added recombinant mouse IL-4 (100 ng/mL, R&D 2
System, USA) or recombinant TNF-α (10 ng/mL, R&D System, USA) for six hours and 3
incubated with TPNs, RTPNs or manRTPNs for 3 hours. Afterward, cells were fixed with 4% 4
paraformaldehyde (Sigma-Aldrich, USA) for 10 minutes, permeabilized with 0.1% Triton™ 5
X-100 (Sigma-Aldrich, USA) for 10 minutes, blocked with 1% BSA for 1 hour and labeled 6
with 2 µg/mL anti-mannose receptor/AF488 conjugated antibody (ab195191, Abcam) and 7
rabbit anti-CD2/AF488 conjugated antibody (bs-2899R, Bioss) for 3 hours at room 8
temperature. Nuclei were stained with DAPI. All manipulation must be away from light. The 9
images were captured under confocal microscopy (Leica, TCS SP5). 10
Cellular cytotoxicity study 11
The cellular toxicities of TPNs and manRTPNs on macrophages, T cells, FLSs and hFLSs 12
were determined by MTT assay using CCK-8 Kit. Cells were implanted in a 96-well culture 13
plate at a density of 8 × 103 cells/well (5 replicates each treated group). After 12 hrs, PBS, TPNs 14
and manRTPNs with concentrations of 10 nM, 50 nM, 100 nM, 200 nM, 500 nM were added 15
into medium for further 24 hrs co-culture. Then the cells were mildly washed by PBS before 16
added CCK-8 mixed solution. Then, the optical density in 450 nm was measured by Multiskan 17
GO (Thermo Fisher Scientific). 18
Cytokines concentration analysis 19
The TNF-α, IL-6 and IFN-𝛾 concentrations of cells culture supernatants were determined 20
with ELISA Kit using mouse ELISA Kits (Biolegend, USA) as manipulation. Also, the TNF-α 21

24
and IL-6 concentrations of mouse serum samples were determined with TNF-α and IL-6 ELISA 1
Kits (Biolegend, USA). Briefly, Serum samples of treated mice were collected at the end of 2
experiment and concentrations of TNF-α and IL-6 were quantified with ELISA kits. 3
Specifically, the whole blood of mice was collected by submandibular bleeding into microtubes 4
and allowed to clot at room temperature for 30 min. Samples were then centrifuged at 2,000g 5
for 6 min to collect supernatant serum. Serum samples were immediately frozen at -20 °C until 6
analysis by using mouse TNF-α and mouse IL-6 ELISA kits (Biolegend) within 3 days of 7
collection. 8
Genes expression profiles analysis of cells 9
After treated with nanoparticles of serious concentrations, cells were harvested to isolate 10
RNA. RNA was extracted with TRIzol reagent (Invitrogen, USA) according to the 11
manufacturer’s instructions. Complementary DNAs (cDNAs) were synthesized with 12
QuantiTect Reverse Transcription Kit (QIAGEN K.K., Tokyo, Japan) and the specific gene 13
transcripts were quantified by quantitative real-time PCR using QuantiTect SYBR Green PCR 14
Kit (QIAGEN K.K., Tokyo, Japan) and analyzed with ABI 7500 real-time PCR system 15
(Applied Biosystems, USA). The gene expression patterns along with concentration were 16
highlight with heatmap by using TBtools package. 17
Collagen induced arthritis (CIA) mice model induction 18
Seventy-two female DBA/1 mice (6∼8 weeks old) were purchased from Vital River 19
Laboratory Animal Technology Co. Ltd (Beijing, China). All mice were maintained in a SF 20
system which is room equipped with an air-filtering system, and the cages and water were 21

25
sterilized. CIA was induced as previously reported study[39]. Briefly, bovine type II collagen 1
(Chondrex, Redmond, WA, USA) was dissolved in 0.1M acetic acid overnight at 4°C. This was 2
emulsified in an equal volume of complete Freund’s adjuvant (Chondrex, Redmond, WA, 3
USA). The mice were immunized intradermal at the base of the tail with 100 𝜇L of emulsion 4
containing 100 𝜇g of type II collagen. On day 21, mice were boosted intraperitoneal with 100 5
𝜇g type II collagen dissolved in phosphate buffered saline (PBS). Clinical scores of each 6
hindpaws and forepaws of mice were obtained following the standard evaluation process[39]. 7
In detail, no evidence of erythema and swelling occurred is score 0, erythema and mild swelling 8
appeared is score 1, erythema and mild swelling extended from the ankle to the tarsals is score 9
2, erythema and moderate swelling extended from the ankle to metatarsal joints is score 3 and 10
erythema and severe swelling encompassed the ankle, paws, and digits or ankyloses of the limb 11
is score 4. Then total clinical scores of hindpaws and forepaws were calculated. At least six 12
mice each group. 13
Nanoparticles targeting to inflamed joints 14
After CIA model was established, free Cy5.5, Cy5.5-labelled TPNs and manRTPNs were 15
intravenously injected into the CIA mice (20 mg/kg, n = 4). Fluorescence images were obtained 16
after 1, 2, 4, 8, 12, and 24 hours using an in vivo imaging system (IVIS, Caliper, USA). Then, 17
the mice were sacrificed for ex vivo tissue/organs distribution analysis at the end of experiment. 18
Meanwhile, the synovial tissue was dissected and dehydrated in 30% sucrose solution overnight 19
and then cry-sectioned at 8 𝜇m. The sections were stained with DAPI and washed twice. 20

26
Fluorescence images were obtained using a confocal microscope (Leica, TCS SP5). The 1
fluorescence intensity of sections in different groups was calculated using software Image J. 2
manRTPNs treatment assay 3
According to CIA mice model conduction, starting at day 21, the clinical scores were 4
calculated to make sure that CIA mice were almost equably and randomly divided into four 5
groups, while healthy DBA/1 mice were set as control (n = 5). Five different agents namely 6
PBS, RBC vesicle, triptolide, TPNs and manRTPNs were intravenously injected every two 7
days. The timeline of whole therapy experiment from model making to the end was according 8
to schematic diagram shown in figure 5A. The arthritis index of each group was recorded over 9
time to evaluate treatment efficacy. At the end, the mice were sacrificed for further analysis. 10
Micro-CT imaging of mice paws 11
The knee and ankle joints of each experimental mouse hindpaws were scanned using 12
micro-CT system (Bruker's SkyScan 1278, UK) to observe joint bone situation. Images were 13
acquired at 55 kV, 72 μA and 300 ms/frame, with 360 views. The 3D structures of knee joint 14
and ankle joint were reconstructed from 360 views and BDM was evaluated by corresponding 15
SkyScan NRecon package. Analysis typically took around 15 min per joint for an experienced 16
operator. 17
Histological analysis and immunohistochemical staining assay 18
At study endpoints, mice were euthanized and hind knee joints were collected for H&E 19
staining and immunohistochemical staining. Briefly, the ankle and knee joints of scarified 20
animals as well as their heart, liver, spleen, lung and kidney were fixed in 10% buffered 21

27
formalin and then joints were incubated in decalcifying solution (4% hydrochloric acid in 4% 1
formaldehyde) at room temperature for 7 days for decalcification. After paraffinization, 2
microtome (Leica) slices of 8 μm were prepared and stained with haematoxylin and eosin, 3
images were taken by using inverted microscope (Olympus, IX71, Japan) and the inflammatory 4
cell infiltration in synovial tissues, bone and cartilage was evaluated by Image J software. After 5
deparaffinization the slices were subjected to antigen recovery in 0.01 M sodium citrate buffer 6
at 125 °C for 30 s, followed by 10 s at 90 °C, and then subjected to the endogenous peroxidase 7
inactivation by covering tissue with 3% hydrogen peroxide for 5 min. After blocking non-8
specific binding sites with 10% goat serum in PBS, the slices were incubated with TNF-α/IL-6 9
monoclonal antibody (Invitrogen) with a dilution rate of 1:100, respectively, at 4 °C for 24 h. 10
Then the slices were incubated with horseradish peroxidase (HRP)-conjugated secondary 11
antibody with a dilution rate of 1:800 at 37 °C for 1 hour. Sections were developed using the 12
DAB substrate and then counterstained with haematoxylin. The images were captured and 13
analyzed by inverted microscope (Olympus, IX71, Japan) and Image J software. 14
Gene expression profiles analysis of tissues 15
The paws and ankles were dissected from mice on day 22 of arthritis, snap-frozen in liquid 16
nitrogen, ground into powder, and homogenized. All procedure must be under RNase-free 17
conditions. The RNA isolation and real-time PCR assay were carried out following the protocol 18
following. Briefly, total RNA was extracted with TRIzol reagent (Invitrogen, Carlsbad, CA, 19
USA) from the tissue homogenates according to the manufacturer’s instructions. The total RNA 20
(1 𝜇g) was reverse transcribed to cDNA using the QuantiTect Reverse Transcription Kit 21

28
(QIAGEN, Japan) and the specific gene transcripts were quantified by quantitative real-time 1
PCR using QuantiTect SYBR Green PCR Kit (QIAGEN K.K., Tokyo, Japan) and analyzed 2
with ABI 7500 real-time PCR system (Applied Biosystems, USA). PCR was performed as 40 3
cycles at 94°C for 15 s, 55°C for 30 s, and 72°C for 30 s. The relative RNA expression was 4
calculated with comparative 𝐶𝑇 method; 𝛽-actin as internal ontrol. The gene expression 5
patterns along with concentration were highlight with heatmap by using TBtools package. 6
Gene-specific primers were synthesized by Sangon Biotech (Shanghai) Co., Ltd. and list in the 7
TableS1. 8
Statistical Analysis 9
All the results are reported as mean ± SD. The differences among groups were analyzed 10
using one-way ANOVA analysis and Student’s t-test; *represents P < 0.05, ** represents P < 11
0.01. 12
Acknowledgments 13
This work was supported by National Natural Science Foundation of China (81801838, 14
31571013, 81701816 and 81601552), Guangdong Natural Science Foundation of Research 15
Team (2016A030312006), K.C. Wong Education Foundation (GJTD-2018-14), Natural 16
Science Foundation of Guangdong Province (2018A030313013), Shenzhen Science and 17
Technology Program (JCYJ20180302145912832, JCYJ20160429191503002, 18
JCYJ20170818162259843 and JCYJ20170818163739458). We thank Pro. Hongchang Li and 19
Dr. Yifan Ma of SIAT for their useful and meaningful recommendations to improve this paper, 20
and thank Mr. Danhui Quan for his help in some drawing. 21

29
Competing Interests 1
The authors have declared that no competing interest exists. 2
3
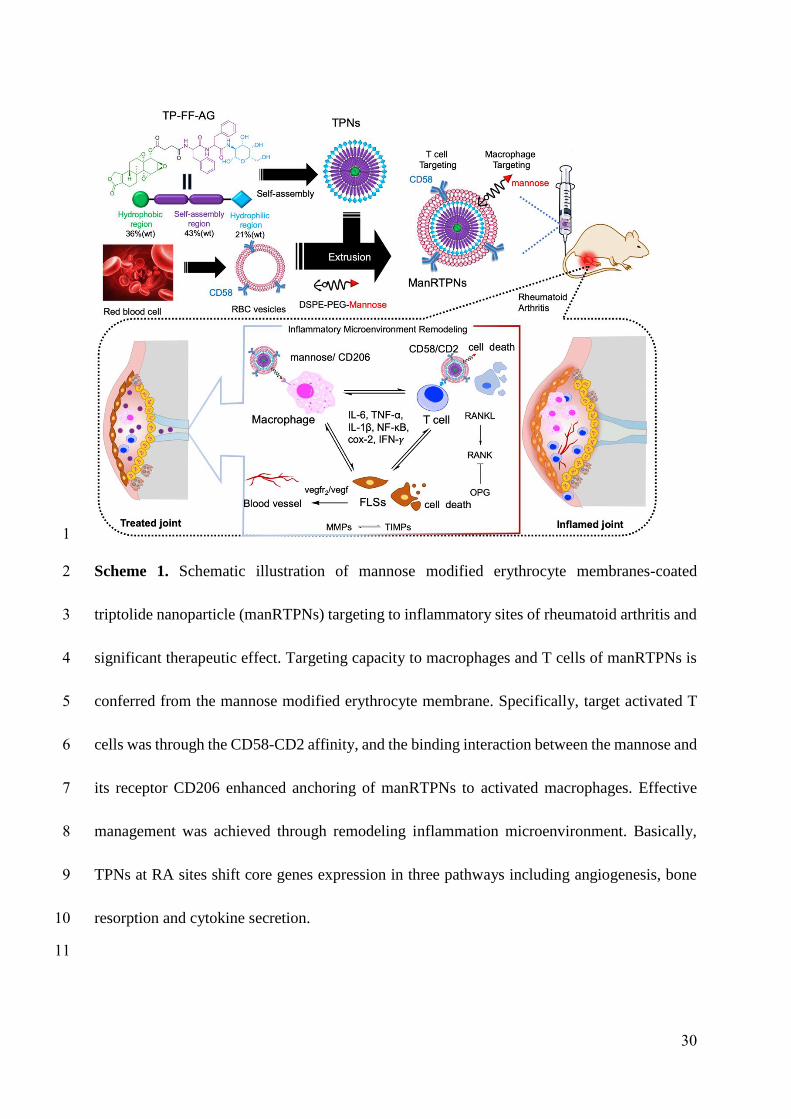
30
1
Scheme 1. Schematic illustration of mannose modified erythrocyte membranes-coated 2
triptolide nanoparticle (manRTPNs) targeting to inflammatory sites of rheumatoid arthritis and 3
significant therapeutic effect. Targeting capacity to macrophages and T cells of manRTPNs is 4
conferred from the mannose modified erythrocyte membrane. Specifically, target activated T 5
cells was through the CD58-CD2 affinity, and the binding interaction between the mannose and 6
its receptor CD206 enhanced anchoring of manRTPNs to activated macrophages. Effective 7
management was achieved through remodeling inflammation microenvironment. Basically, 8
TPNs at RA sites shift core genes expression in three pathways including angiogenesis, bone 9
resorption and cytokine secretion. 10
11
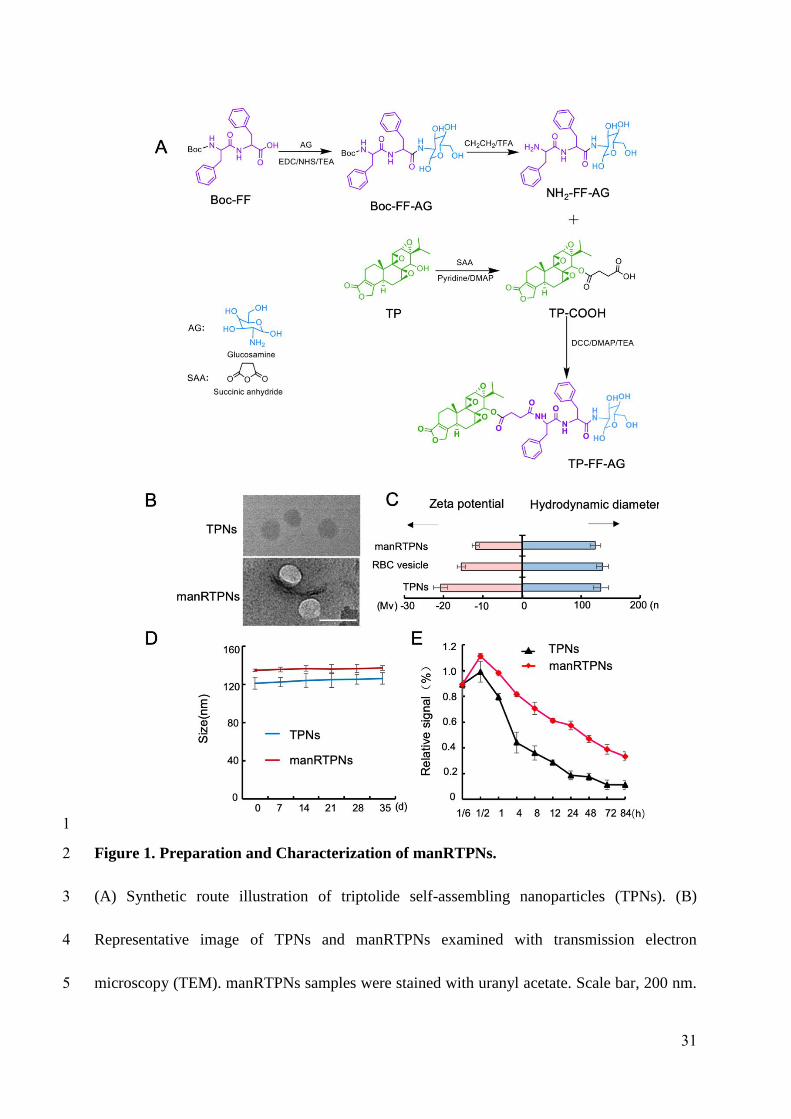
31
1
Figure 1. Preparation and Characterization of manRTPNs. 2
(A) Synthetic route illustration of triptolide self-assembling nanoparticles (TPNs). (B) 3
Representative image of TPNs and manRTPNs examined with transmission electron 4
microscopy (TEM). manRTPNs samples were stained with uranyl acetate. Scale bar, 200 nm. 5

32
(C) Size and ζ potential distribution of TPNs, RBC vesicles and manRTPNs. Each experiment 1
had three replicates. (D) Stability of TPNs and manRTPNs in PBS over five weeks. Each 2
experiment had three replicates. (E) Pharmacokinetics of TPNs and manRTPNs in vivo. Each 3
experiment point had three replicates. 4
5
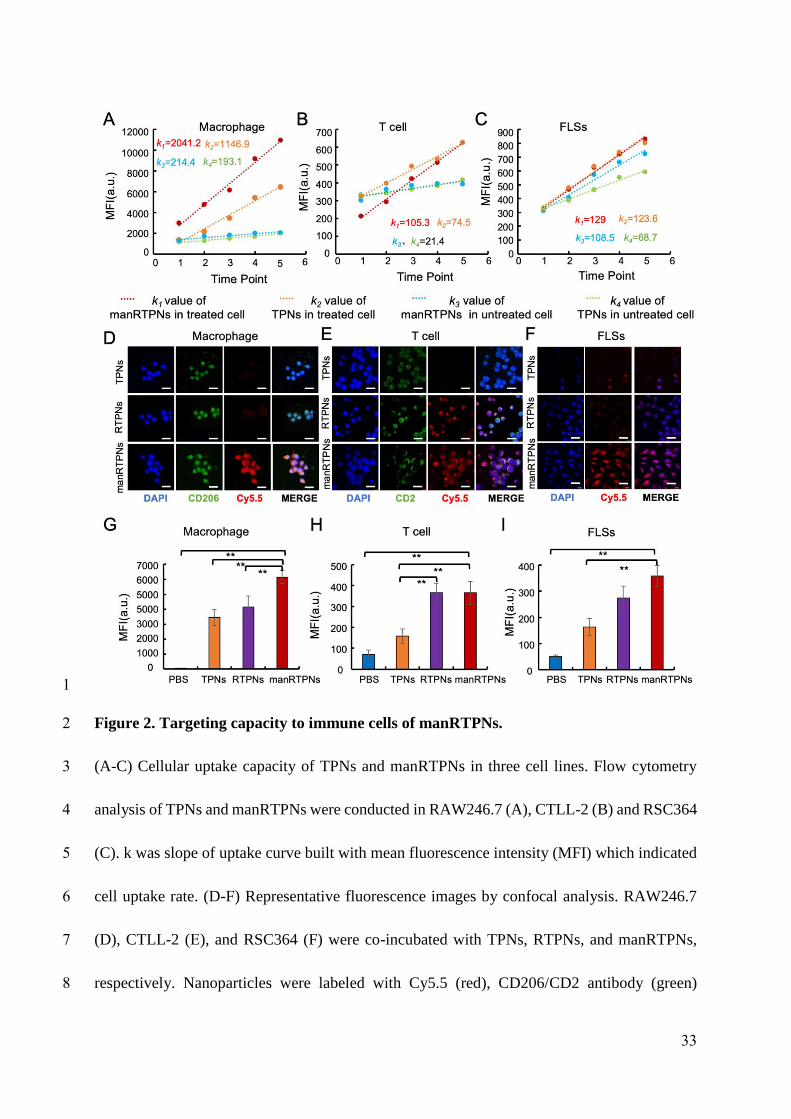
33
1
Figure 2. Targeting capacity to immune cells of manRTPNs. 2
(A-C) Cellular uptake capacity of TPNs and manRTPNs in three cell lines. Flow cytometry 3
analysis of TPNs and manRTPNs were conducted in RAW246.7 (A), CTLL-2 (B) and RSC364 4
(C). k was slope of uptake curve built with mean fluorescence intensity (MFI) which indicated 5
cell uptake rate. (D-F) Representative fluorescence images by confocal analysis. RAW246.7 6
(D), CTLL-2 (E), and RSC364 (F) were co-incubated with TPNs, RTPNs, and manRTPNs, 7
respectively. Nanoparticles were labeled with Cy5.5 (red), CD206/CD2 antibody (green) 8

34
expressed on the IL-4 -treated RAW264.7 and TNF-α treated CTLL-2 and cellular nuclear was 1
labeled with DAPI (blue). Scale bars, 100 μm. (G-I) Flow cytometry analysis of RAW246.7 2
(G), CTLL-2 (H) and RSC364 (I) incubated with various nanoparticles. ** indicated significant 3
difference and p < 0.05. 4
5
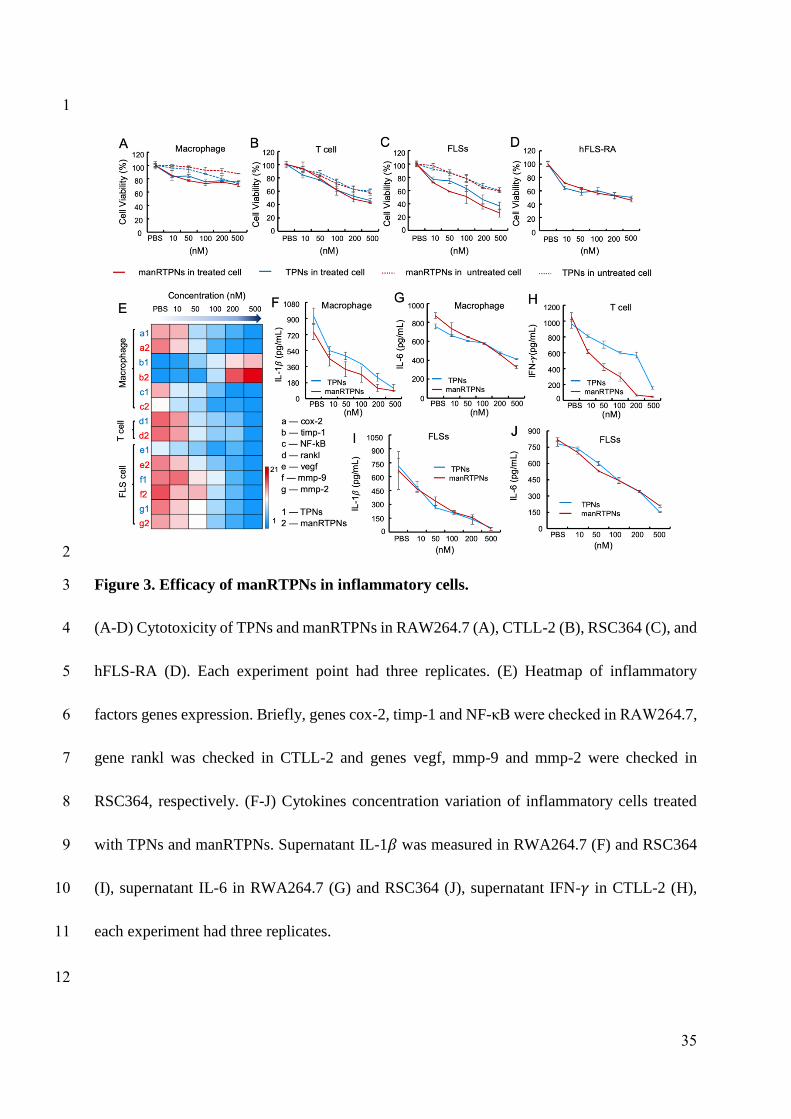
35
1
2
Figure 3. Efficacy of manRTPNs in inflammatory cells. 3
(A-D) Cytotoxicity of TPNs and manRTPNs in RAW264.7 (A), CTLL-2 (B), RSC364 (C), and 4
hFLS-RA (D). Each experiment point had three replicates. (E) Heatmap of inflammatory 5
factors genes expression. Briefly, genes cox-2, timp-1 and NF-κB were checked in RAW264.7, 6
gene rankl was checked in CTLL-2 and genes vegf, mmp-9 and mmp-2 were checked in 7
RSC364, respectively. (F-J) Cytokines concentration variation of inflammatory cells treated 8
with TPNs and manRTPNs. Supernatant IL-1𝛽 was measured in RWA264.7 (F) and RSC364 9
(I), supernatant IL-6 in RWA264.7 (G) and RSC364 (J), supernatant IFN-𝛾 in CTLL-2 (H), 10
each experiment had three replicates. 11
12
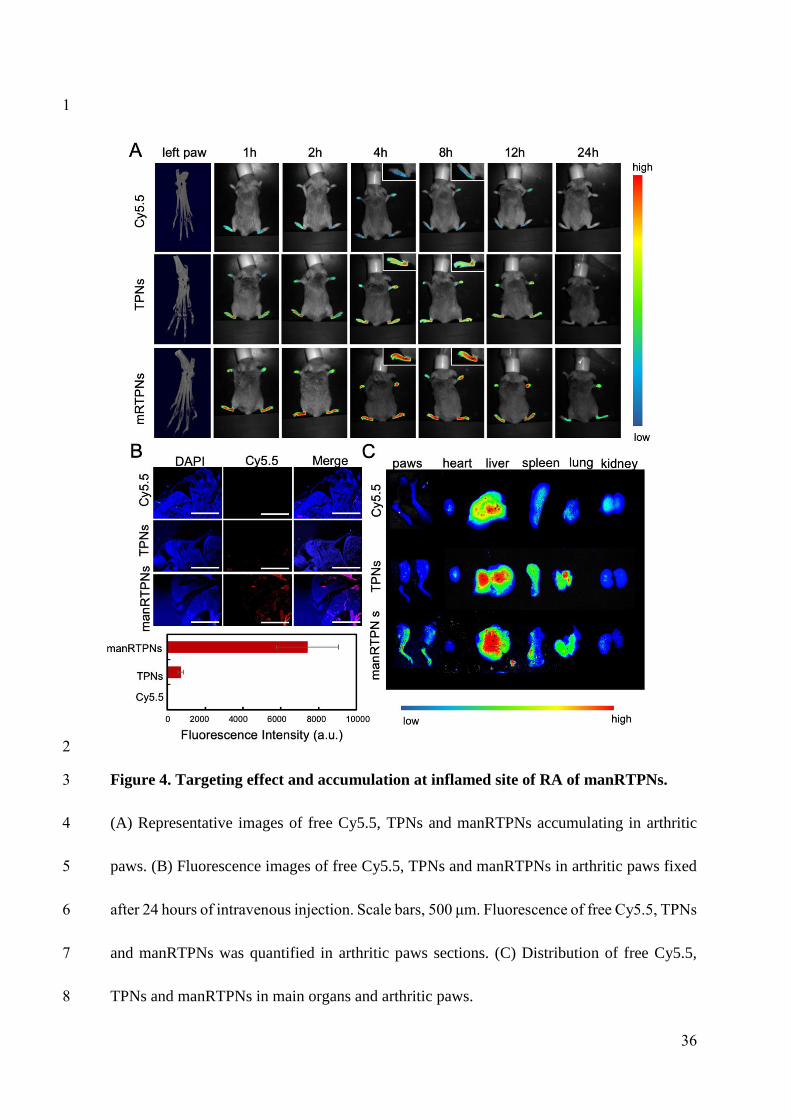
36
1
2
Figure 4. Targeting effect and accumulation at inflamed site of RA of manRTPNs. 3
(A) Representative images of free Cy5.5, TPNs and manRTPNs accumulating in arthritic 4
paws. (B) Fluorescence images of free Cy5.5, TPNs and manRTPNs in arthritic paws fixed 5
after 24 hours of intravenous injection. Scale bars, 500 μm. Fluorescence of free Cy5.5, TPNs 6
and manRTPNs was quantified in arthritic paws sections. (C) Distribution of free Cy5.5, 7
TPNs and manRTPNs in main organs and arthritic paws. 8

37
1
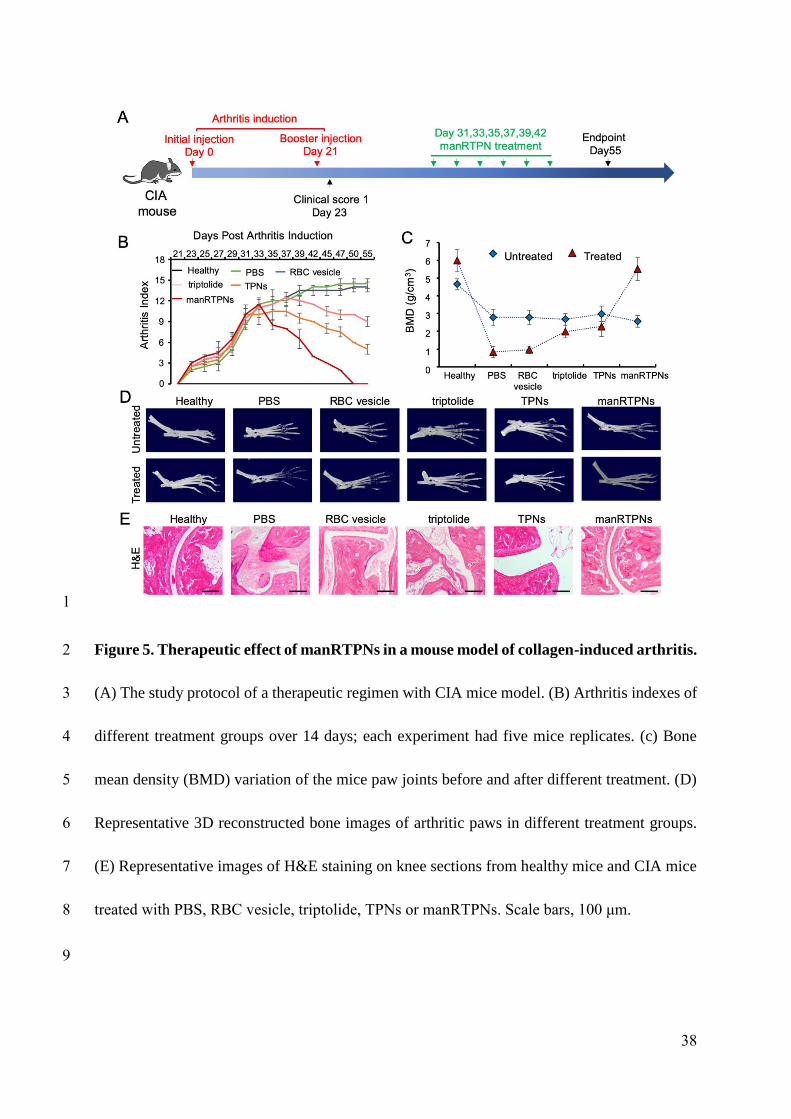
38
1
Figure 5. Therapeutic effect of manRTPNs in a mouse model of collagen-induced arthritis. 2
(A) The study protocol of a therapeutic regimen with CIA mice model. (B) Arthritis indexes of 3
different treatment groups over 14 days; each experiment had five mice replicates. (c) Bone 4
mean density (BMD) variation of the mice paw joints before and after different treatment. (D) 5
Representative 3D reconstructed bone images of arthritic paws in different treatment groups. 6
(E) Representative images of H&E staining on knee sections from healthy mice and CIA mice 7
treated with PBS, RBC vesicle, triptolide, TPNs or manRTPNs. Scale bars, 100 μm. 8
9
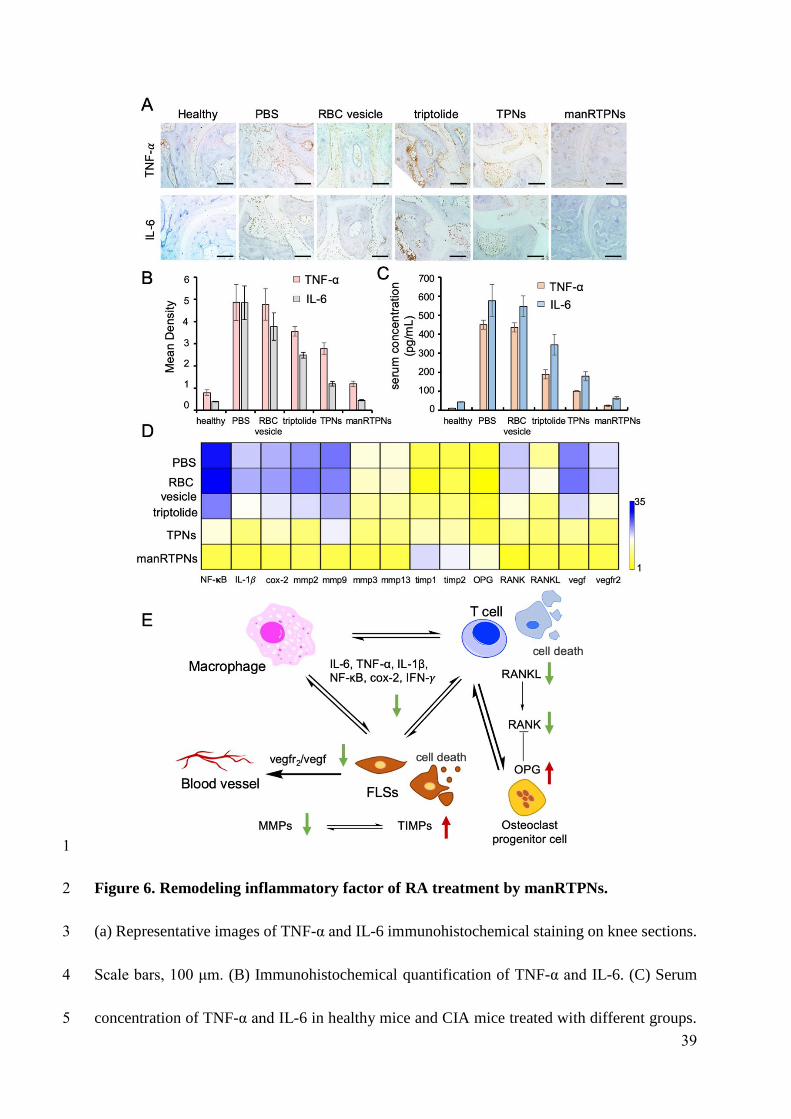
39
1
Figure 6. Remodeling inflammatory factor of RA treatment by manRTPNs. 2
(a) Representative images of TNF-α and IL-6 immunohistochemical staining on knee sections. 3
Scale bars, 100 μm. (B) Immunohistochemical quantification of TNF-α and IL-6. (C) Serum 4
concentration of TNF-α and IL-6 in healthy mice and CIA mice treated with different groups. 5

40
Each experiment point had three replicates. (D) Heatmap of genes expression profiles in 1
different treatment. 2
3
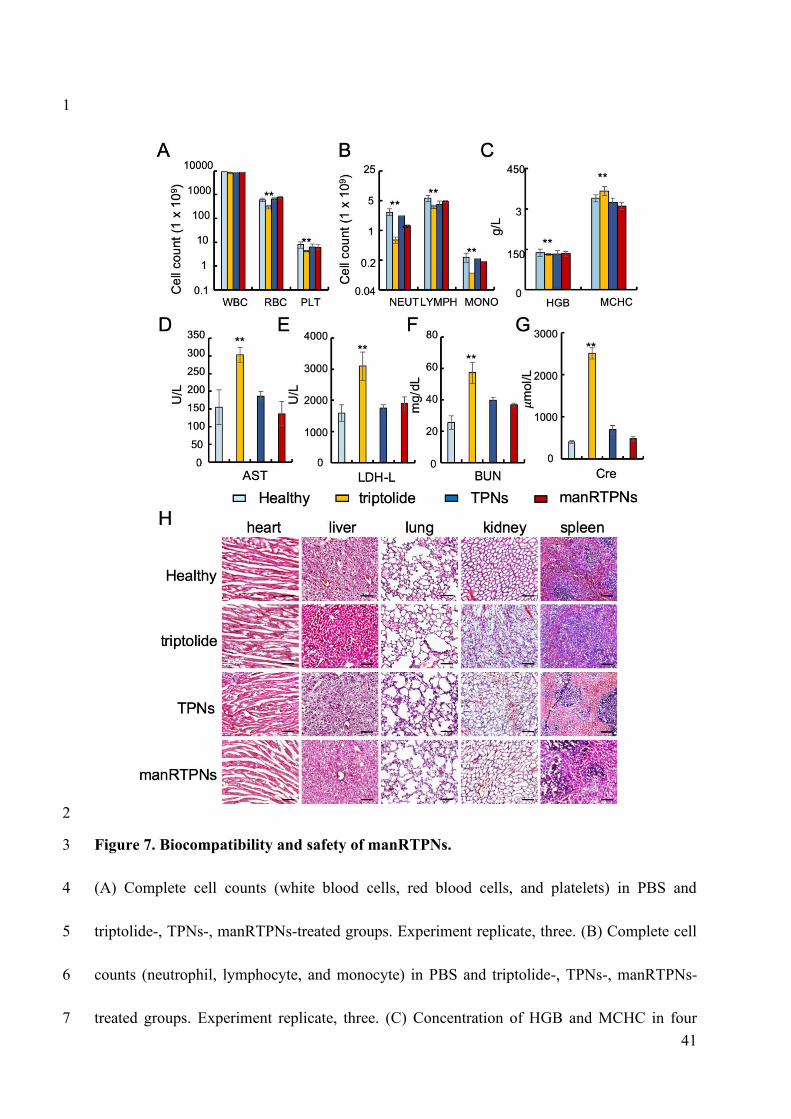
41
1
2
Figure 7. Biocompatibility and safety of manRTPNs. 3
(A) Complete cell counts (white blood cells, red blood cells, and platelets) in PBS and 4
triptolide-, TPNs-, manRTPNs-treated groups. Experiment replicate, three. (B) Complete cell 5
counts (neutrophil, lymphocyte, and monocyte) in PBS and triptolide-, TPNs-, manRTPNs-6
treated groups. Experiment replicate, three. (C) Concentration of HGB and MCHC in four 7

42
groups indicated before. Experiment replicate, three. (D-G) Blood routine examination 1
parameters including AST(D), LDH-L(E), BUN(F) and Cre(G) after different treatment. 2
Experiment replicate, three. (H) H&E staining of main organs in four different groups. Scale 3
bars, 100 μm. 4
5

43
1. Scott DL, Wolfe F, Huizinga TWJ. Rheumatoid arthritis. Lancet. 2010; 376: 1094-1108. 1
2. Van Vollenhoven RF. Treatment of rheumatoid arthritis: state of the art 2009. Nat Rev 2
Rheumatol. 2009; 5: 531-541. 3
3. Bykerk V. Unmet needs in rheumatoid arthritis. J Rheumatol. 2009; 36: 42-46. 4
4. Salliot C, Van Der Heijde D. Long-term safety of methotrexate monotherapy in patients 5
with rheumatoid arthritis: a systematic literature research. Ann Rheum Dis. 2009; 68: 1100-6
1104. 7
5. Alcorn N, Saunders S, Madhok R. Benefit-risk assessment of leflunomide an appraisal of 8
leflunomide in rheumatoid arthritis 10 years after licensing. Drug Safety. 2009; 32: 1123-9
1134. 10
6. Leombruno JP, Einarson TR, Keystone EC. The safety of anti-tumour necrosis factor 11
treatments in rheumatoid arthritis: meta and exposure-adjusted pooled analyses of serious 12
adverse events. Ann Rheum Dis. 2009; 68: 1136-1145. 13
7. Hyrich KL, Watson KD, Isenberg DA, Symmons DPM, Register BB. The british society 14
for rheumatology biologics register: 6 years on. Rheumatology. 2008; 47: 1441-1443. 15
8. Kirwan JR, Bijlsma JWJ, Boers M, Shea BJ. Effects of glucocorticoids on radiological 16
progression in rheumatoid arthritis. Cochrane Db Syst Rev. 2007. 17
9. Ravindran V, Rachapalli S, Choy EH. Safety of medium- to long-term glucocorticoid 18
therapy in rheumatoid arthritis: a meta-analysis. Rheumatology. 2009; 48: 807-811. 19

44
10. Zhang QZ, Dehaini D, Zhang Y, Zhou JL, Chen XY, Zhang LF, et al. Neutrophil 1
membrane-coated nanoparticles inhibit synovial inflammation and alleviate joint damage 2
in inflammatory arthritis. Nat Nanotechnol. 2018; 13: 1182-1190. 3
11. Li RX, He YW, Zhu Y, Jiang LX, Zhang SY, Qin J, et al. Route to rheumatoid arthritis by 4
macrophage-derived microvesicle-coated nanoparticles. Nano Letters. 2019; 19: 124-134. 5
12. Liang HY, Peng B, Dong C, Liu LX, Mao JJ, Wei S, et al. Cationic nanoparticle as an 6
inhibitor of cell-free DNA-induced inflammation. Nat Commun. 2018; 9: 4291. 7
13. Tao X, Cush J, Garret M, Pe. L. A phase I study of ethylacetate extract of the Chinese 8
antirheumatic herb Tripterygium wilfordii Hook F in rheumatoid arthritis. J Rheumatol. 9
2001; 28: 2167. 10
14. Fan DP, Guo QQ, Shen JW, Zheng K, Lu C, Zhang G, et al. The effect of triptolide in 11
rheumatoid arthritis: from basic research towards clinical translation. Int J Mol Sci. 2018; 12
19. 13
15. Pao HP, Liao WI, Wu SY, Hung KY, Huang KL, Chu SJ. PG490-88, a derivative of 14
triptolide, suppresses ischemia/reperfusion-induced lung damage by maintaining tight 15
junction barriers and targeting multiple signaling pathways. Int Immunopharmacol. 2019; 16
68: 17-29. 17
16. Chen YK, Zhang L, Ni JS, Wang XY, Cheng J, Li YC, et al. LLDT-8 protects against 18
cerebral ischemia/reperfusion injury by suppressing post-stroke inflammation. J Pharmacol 19
Sci. 2016; 131: 131-137. 20

45
17. He QL, Minn I, Wang QL, Xu P, Head SA, Datan E, et al. Targeted delivery and sustained 1
antitumor activity of triptolide through glucose conjugation. Angew Chem Int Edit. 2016; 2
55: 12035-12039. 3
18. Yuan ZX, Wu XJ, Mo JX, Wang YL, Xu CQ, Lim LY. Renal targeted delivery of triptolide 4
by conjugation to the fragment peptide of human serum albumin. Eur J Pharm Biopharm. 5
2015; 94: 363-371. 6
19. Mei ZN, Chen HB, Weng T, Yang YJ, Yang XL. Solid lipid nanoparticle and 7
microemulsion for topical delivery of triptolide. Eur J Pharm Biopharm. 2003; 56: 189-8
196. 9
20. Huang CL, Zeng T, Li JW, Tan LS, Deng XL, Pan YC, et al. Folate receptor-mediated 10
renal-targeting nanoplatform for the specific delivery of triptolide to treat renal 11
ischemia/reperfusion injury. ACS Biomater Sci Eng. 2019; 5: 2877-2886. 12
21. Ling D, Xia H, Park W, Hackett MJ, Song C, Na K, et al. pH-sensitive nanoformulated 13
triptolide as a targeted therapeutic strategy for hepatocellular carcinoma. ACS Nano. 2014; 14
8: 8027-8039. 15
22. Xu HT, Liu B. Triptolide-targeted delivery methods. Eur J Med Chem. 2019; 164: 342-16
351. 17
23. Abbas M, Zou QL, Li SK, Yan XH. Self-assembled peptide- and protein-based 18
nanomaterials for antitumor photodynamic and photothermal therapy. Adv Mater. 2017; 19
29. 20

46
24. Zhu PL, Yan XH, Su Y, Yang Y, Li JB. Solvent-induced structural transition of self-1
assembled dipeptide: from organogels to microcrystals. Chem-Eur J. 2010; 16: 3176-3183. 2
25. Yan XH, Zhu PL, Li JB. Self-assembly and application of diphenylalanine-based 3
nanostructures. Chem Soc Rev. 2010; 39: 1877-1890. 4
26. Elshabrawy HA, Chen ZL, Volin MV, Ravella S, Virupannavar S, Shahrara S. The 5
pathogenic role of angiogenesis in rheumatoid arthritis. Angiogenesis. 2015; 18: 433-448. 6
27. Kinne RW, Brauer R, Stuhlmuller B, Palombo-Kinne E, Burmester GR. Macrophages in 7
rheumatoid arthritis. Arthritis Res. 2000; 2: 189-202. 8
28. Davis SJ, Ikemizu S, Evans EJ, Fugger L, Bakker TR, Van Der Merwe PA. The nature of 9
molecular recognition by T cells. Nat Immunol. 2003; 4: 217-224. 10
29. Raychaudhuri S, Thomson BP, Remmers EF, Eyre S, Hinks A, Guiducci C, et al. Genetic 11
variants at CD28, PRDM1 and CD2/CD58 are associated with rheumatoid arthritis risk. 12
Nat Genet. 2009; 41: 1313-1318. 13
30. Sable R, Durek T, Taneja V, Craik DJ, Pallerla S, Gauthier T, et al. Constrained cyclic 14
peptides as immunomodulatory inhibitors of the CD2:CD58 protein-protein interaction. 15
ACS Chemical Biology. 2016; 11: 2366-2374. 16
31. Hu CMJ, Fang RH, Copp J, Luk BT, Zhang LF. A biomimetic nanosponge that absorbs 17
pore-forming toxins. Nat Nanotechnol. 2013; 8: 336-340. 18
32. Rossi L, Fraternale A, Bianchi M, Magnani M. Red blood cell membrane processing for 19
biomedical applications. Front Physiol. 2019; 10. 20

47
33. He QL, Titov DV, Li J, Tan MJ, Ye ZH, Zhao YM, et al. Covalent modification of a 1
cysteine residue in the XPB subunit of the general transcription factor TFIIH through single 2
epoxide cleavage of the transcription inhibitor triptolide. Angew Chem Int Edit. 2015; 54: 3
1859-1863. 4
34. Hu CMJ, Zhang L, Aryal S, Cheung C, Fang RH, Zhang LF. Erythrocyte membrane-5
camouflaged polymeric nanoparticles as a biomimetic delivery platform. P Natl Acad Sci 6
USA. 2011; 108: 10980-10985. 7
35. Fang RNH, Hu CMJ, Chen KNH, Luk BT, Carpenter CW, Gao WW, et al. Lipid-insertion 8
enables targeting functionalization of erythrocyte membrane-cloaked nanoparticles. 9
Nanoscale. 2013; 5: 8884-8888. 10
36. Shao F, Wang GJ, Xie HT, Zhu XY, Sun JG, A JY. Pharmacokinetic study of triptolide, a 11
constituent of immunosuppressive chinese herb medicine, in rats. Biol Pharm Bull. 2007; 12
30: 702-707. 13
37. Ghosh S, Hayden MS. New regulators of NF-kappaB in inflammation. Nat Rev Immunol. 14
2008; 8: 837-848. 15
38. Niedermeier M PT, Korb A. Therapeutic opportunities in fibroblasts in inflammatory 16
arthritis. Best Pract Res Clin Rheumatol. 2010; 24: 527-540. 17
39. Brand DD, Latham KA, Rosloniec EF. Collagen-induced arthritis. Nat Protoc. 2007; 2: 18
1269-1275. 19
40. Noack M, Miossec P. Selected cytokine pathways in rheumatoid arthritis. Seminars in 20
Immunopathology. 2017; 39: 365-383. 21

48
41. Geusens P. The role of RANK ligand/osteoprotegerin in rheumatoid arthritis. Ther Adv 1
Musculoskelet Dis. 2012; 4: 225-233. 2
42. Chen L, Deng H, Cui H, Fang J, Zuo Z, Deng J, et al. Inflammatory responses and 3
inflammation-associated diseases in organs. Oncotarget. 2018; 9: 7204-7218. 4
5

download fileview on ChemRxivmanuscript-update 20200804.pdf (2.67 MiB)


![Stimuli-responsive mesoporous silica nanoparticles for ... · polymers, lipids, self-assembling amphiphilic molecules, metals and other inorganic materials [14]. The raw material](https://img.pdfslide.net/doc/110x75/5fa6ffd31f655536fd2de42c/stimuli-responsive-mesoporous-silica-nanoparticles-for-polymers-lipids-self-assembling.jpg)


























