
doi:10.1016/j.jmb.2004.08.044 J. Mol. Biol. (2004) 343, 547–557
Conformational Changes Induced by NucleotideBinding in Cdc6/ORC From Aeropyrum pernix
Martin R. Singleton1†, Renaud Morales1†, Ian Grainge1†, Nicola Cook1
Michail N. Isupov2 and Dale B. Wigley1*
1Cancer Research UK ClareHall Laboratories, The LondonResearch Institute, Blanche LaneSouth Mimms, Potters BarHerts EN6 3LD, UK
2School of ChemistryUniversity of Exeter, StockerRoad, Exeter EX4 4QD, UK
0022-2836/$ - see front matter q 2004 E
†M R.S., R.M. & I.G. contributedAbbreviations used: pre-RC, pre-
MCM, mini chromosome maintenarecognition box; ORC, origin recognwinged helix.E-mail address of the correspond
Archaea contain one or more proteins with homology to eukaryoticORC/Cdc6 proteins. Sequence analysis suggests the existence of at leasttwo subfamilies of these proteins, for which we propose the nomenclatureORC1 and ORC2. We have determined crystal structures of the ORC2protein from the archaeon Aeropyrum pernix in complexes with ADP or anon-hydrolysable ATP analogue, ADPNP. Between two crystal forms, thereare three crystallographically independent views of the ADP complex andtwo of the ADPNP complex. The protein molecules in the three complexeswith ADP adopt very different conformations, while the two complexeswith ADPNP are the same. These structures indicate that there isconsiderable conformational flexibility in ORC2 but that ATP bindingstabilises a single conformation. We show that the ORC2 protein can bindDNA, and that this activity is associated with the C-terminal domain of theprotein. We present a model for the interaction of the winged helix (WH)domain of ORC2 with DNA that differs from that proposed previously forPyrobaculum aerophilum ORC/Cdc6.
q 2004 Elsevier Ltd. All rights reserved.
Keywords: Cdc6; ORC; replication origin; AAACproteins
*Corresponding authorIntroduction
In eukaryotes, replication occurs once, and onlyonce, per cell-cycle. It is clear that DNA replicationis a very tightly controlled process. Replicationbubbles emanate from distinct DNA regions and itis at these replication origins that all the factors forreplication are laid down in a temporally controlledway. The first step in activating an origin is a pro-cess called licensing, in which the pre-replicationcomplex (pre-RC) is localised to the origin. Thelicensing reaction culminates with the loading ofthe mini chromosome maintenance (MCM) com-plex, probably the replicative helicase, onto theorigin DNA but involves many other protein factorsto achieve this goal.1
In budding yeast, origins are relatively small andwell characterised. Throughout the cell-cycle, theorigin recognition complex (ORC) remains bound at
lsevier Ltd. All rights reserve
equally to this work.replication complex;nce; ORB, originition complex; WH,
ing author:
the origin and acts as a scaffold to recruit otherfactors during licensing. Two other proteins, Cdc6and Tah11 (Cdt1 in some other organisms), associ-ate with ORC and are required for the subsequentloading of the MCM complex.The AAAC family of proteins are involved in
many cellular processes, including DNA replica-tion.2 Although proteins of this family, whichincludes the Cdc6, ORC and MCM proteins, areall presumed ATPases on the basis of conservedsequence motifs, the role(s) of ATP binding and/orhydrolysis in these proteins is not always wellunderstood. For example, although the role of ATPhydrolysis in MCM is clear, namely to drive thehelicase reaction, the requirement for ATP by Cdc6and ORC proteins is less obvious. However, a likelyrole for ATP hydrolysis in these proteins is in thetemporal control of the initiation of replication,including the loading of other proteins at origins.3
In archaea, the replication, repair and transcrip-tional apparatus are related to eukaryotes moreclosely than to eubacteria. DNA replication appearsto be initiated from a single origin in Pyrococcusand Halobacterium species,4,5 but from at least twoorigins in Sulfolobus.6 Paradoxically, however, theorganisation of these origins appears to be morereminiscent of those in Eubacteria (e.g. oriC) than of
d.

Figure 1. Sequence alignment of archaeal ORC/Cdc6 proteins showing the division between class I and class IIproteins. The secondary structure of the winged helix region is shown at the top (helix as open rectangle, beta-strands asarrows). Conserved identical residues are highlighted in green and those conserved in themajority of proteins are shownin yellow. Archaeal species: Sso, Solfolobus solfataricus; Ape, Aeropyrum pernix; Pfu, Pyrococcus furiosus; Tac, Thermoplasmaacidophilum; Tvo, Thermoplasma volcanium; Mth, Methanobacterium thermoautotrophicum; Pae, Pyrobaculum aerophilum.
548 Nucleotide Binding in Aeropyrum pernix Cdc6/ORC
budding yeast, with several regions of invertedrepeat sequences located either side of an AT-richregion. With regard to proteins, in most archaeathere is a single gene encoding the MCM helicase,which then forms a homohexamer (or sometimes adouble hexamer) in solution,7–12 although a hepta-meric ring form has been reported for Methano-bacterium thermoautotrophicum MCM.13 For the ORCproteins and Cdc6, the situation is again simplifiedin most archaea, even to the extent that severalspecies encode a single gene with homology both toyeast Orc1 and Cdc6 proteins.14 However, in manyother species there are two genes, one with greatestsequence homology to the yeast Cdc6 gene and theother more like the Orc1 protein. The homologybetween these genes, however, makes ab initioassignment difficult. Furthermore, neither geneproduct has been characterised very well to date.To complicate the issue further, genome sequencingsuggests that other archaeal species appear to havethree or even as many as ten ORC/Cdc6 homol-ogues. Homologues of other proteins involved inpre-RC formation in eukaryotes appear to be absentfrom archaea.
Here, we present the crystal structures of Aero-pyrum pernix gene product APE0152, a member ofthe ORC/Cdc6 protein family. We have determinedstructures in two different crystal forms, one grownin the absence of added nucleotide and the othergrown in the presence of the non-hydrolysable ATPanalogue, ADPNP. Between these two crystalforms, we have five crystallographically indepen-dent views of the protein. Three of these have ADPbound to the protein, despite ADP not being added
to the crystallisation conditions. These three mol-ecules all show different conformations, which inturn are all different from the ADP complex of thePyrobaculum aerophilum ORC/Cdc6 protein.15 Theremaining two molecules have bound ADPNP.These molecules share a similar conformation, butone that is different from any of the ADP complexes.These structures reveal a conformational flexibilityof the ADP complex that is altered to a singleconformation when ATP replaces the ADP. Amodelfor DNA binding is presented that suggests amechanism for interaction between bound proteinmolecules and/or other components of the pre-replication complex.
Results and Discussion
Sequence homology analysis
A. pernix encodes two ORC/Cdc6 proteins (genesAPE0152 and APE0475). In order to classify theproteins, we aligned sequences of ORC/Cdc6proteins from a variety of archaeal species(Figure 1). These analyses revealed that two sub-families can be delineated, which we refer to asORC1 and ORC2. The basis for the division of thefamilies is a conserved motif that may be a DNArecognition helix (see below). This region is highlyconserved in ORC1 proteins but not in ORC2. Aspart of a study to understand the initiation of DNAreplication in archaea, we have overexpressed andpurified ORC1 and ORC2 from A. pernix forbiochemical analysis. We have also determined the
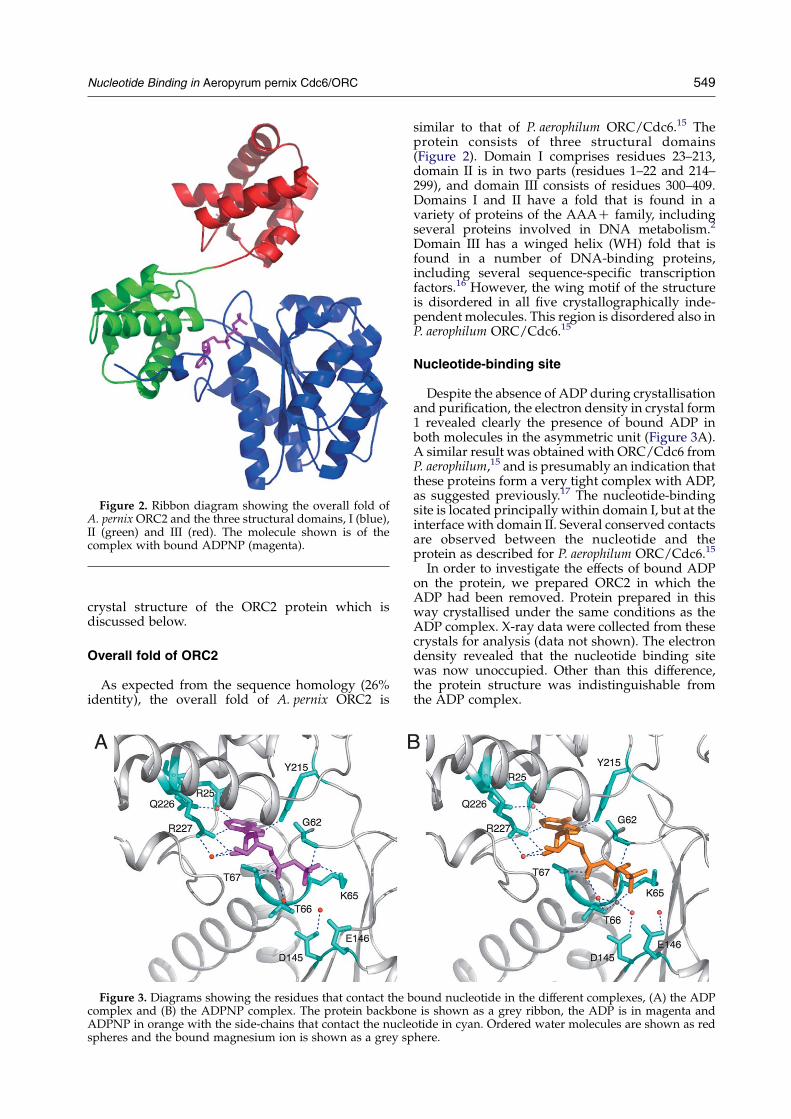
Figure 2. Ribbon diagram showing the overall fold ofA. pernix ORC2 and the three structural domains, I (blue),II (green) and III (red). The molecule shown is of thecomplex with bound ADPNP (magenta).
Nucleotide Binding in Aeropyrum pernix Cdc6/ORC 549
crystal structure of the ORC2 protein which isdiscussed below.
Overall fold of ORC2
As expected from the sequence homology (26%identity), the overall fold of A. pernix ORC2 is
Figure 3. Diagrams showing the residues that contact the bcomplex and (B) the ADPNP complex. The protein backbonADPNP in orange with the side-chains that contact the nuclespheres and the bound magnesium ion is shown as a grey sp
similar to that of P. aerophilum ORC/Cdc6.15 Theprotein consists of three structural domains(Figure 2). Domain I comprises residues 23–213,domain II is in two parts (residues 1–22 and 214–299), and domain III consists of residues 300–409.Domains I and II have a fold that is found in avariety of proteins of the AAAC family, includingseveral proteins involved in DNA metabolism.2
Domain III has a winged helix (WH) fold that isfound in a number of DNA-binding proteins,including several sequence-specific transcriptionfactors.16 However, the wing motif of the structureis disordered in all five crystallographically inde-pendent molecules. This region is disordered also inP. aerophilum ORC/Cdc6.15
Nucleotide-binding site
Despite the absence of ADP during crystallisationand purification, the electron density in crystal form1 revealed clearly the presence of bound ADP inboth molecules in the asymmetric unit (Figure 3A).A similar result was obtained with ORC/Cdc6 fromP. aerophilum,15 and is presumably an indication thatthese proteins form a very tight complex with ADP,as suggested previously.17 The nucleotide-bindingsite is located principally within domain I, but at theinterface with domain II. Several conserved contactsare observed between the nucleotide and theprotein as described for P. aerophilum ORC/Cdc6.15
In order to investigate the effects of bound ADPon the protein, we prepared ORC2 in which theADP had been removed. Protein prepared in thisway crystallised under the same conditions as theADP complex. X-ray data were collected from thesecrystals for analysis (data not shown). The electrondensity revealed that the nucleotide binding sitewas now unoccupied. Other than this difference,the protein structure was indistinguishable fromthe ADP complex.
ound nucleotide in the different complexes, (A) the ADPe is shown as a grey ribbon, the ADP is in magenta andotide in cyan. Ordered water molecules are shown as redhere.

550 Nucleotide Binding in Aeropyrum pernix Cdc6/ORC
For crystal form 2 (grown in the presence ofADPNP), we observe two molecules that arecomplexed with ADPNP (Figure 3B) but the thirdmolecule in the asymmetric unit contains a boundADP molecule. This suggests that ADPNP is notvery effective at replacing ADP in the ORC2structure, presumably due to a higher affinity ofORC2 for ADP compared to ADPNP, because theconcentration of ADPNP during crystallisation was1 mM while that of the ADP was no greater thanstoichiometric (!0.1 mM).
Contacts with the bound ADP (Figure 3A) aremuch the same as observed for P. aerophilum ORC/Cdc6.15 The ADPNP complex (Figure 3B) retainsthese contacts and has some additional contacts. Abound magnesium ion is coordinated by both the band g-phosphate groups. There is an additionalcontact with Lys65, which contacts both terminalphosphate moieties in the ADPNP complex. Someadditional ordered water molecules are present inthe ADPNP complex.
Although the reason why ORC2 binds to ADP sotightly compared with many other AAAC familyproteins is not evident from the crystal structure, itis likely that there is some biological significance tothis observation. We find that apo protein canundergo a single turnover of ATP hydrolysis andthen suffers from extreme product inhibition due tothe tight binding of ADP (data not shown) and nofurther ATPase activity can be detected. Similarly,protein isolated without the denaturation step isdevoid of ATPase activity due to ADP that is boundto the protein. One implication of this observation isthat ATP turnover controls the activity of ORC2 andthat some additional factor is required to facilitatethe release of ADP from the protein, presumably asa part of the process(es) controlling origin firing inarchaea.
It has been shown that ORC/Cdc6 homologuesfrom M. thermoautotrophicum, P. aerophilum and Sul-folobus solfataricus are capable of autophosphoryla-tion.17,18 We observe a low level ofautophosphorylation of A. pernix ORC2, althoughATPase assays reveal that greater than 95% of ATPhydrolysis is uncoupled from autophosphorylation,instead releasing ADP and inorganic phosphate(data not shown). The phosphorylation of M. ther-moautotrophicum ORC/Cdc6 was shown to be atserine residues, although the precise location of thephosphorylation site(s) is unknown. However, thecrystal structure of P. aerophilum ORC/Cdc6 did notshow any electron density that could be attributedto phosphorylation.15 Likewise, we do not observeelectron density at any serine residues of A. pernixORC2 that could be interpreted as phosphorylationin any of the five crystallographically independentmolecules.
Conformation of the protein
The structure of P. aerophilum ORC/Cdc6 con-tains bound ADP molecules in both complexes inthe asymmetric unit and the protein complexes are
very similar in each case.15 However, the twomolecules in crystal form 1 of the A. pernix ORC2reveal a rather different situation. Althoughdomains I and II can be superimposed, the relativedisposition of domain III is quite different in each ofthe two complexes and each is different again fromthe P. aerophilum ADP complex (Figure 4). Theseconformations of A. pernix ORC2 are observed inboth the presence and absence of bound ADP (datanot shown).
For crystal form 2, the situation is even moresurprising. Domains I and II of each of the threemolecules superimpose well with the structuresof all ORC2 molecules. However, the dispositionof domain III is more complex. For the moleculewith a bound ADP, the conformation of domainIII reveals yet another position for this domain,different from any of the other ADP complexes.Domain III for the two molecules that arecomplexed with ADPNP is in a similar locationin each case (0.35 A RMSD for Ca atoms), butdifferent again from all of the ADP complexes. Itis likely that this represents a preferred conformerof the protein when ADPNP and, by inference,when ATP, is bound to ORC2. By contrast, itwould appear that in the ADP complex of theprotein, domain III is extremely conformationallymobile (Figure 4). When domains I and II aresuperimposed, the positions of domain III in eachstructure fall on an arc between the extremes (inboth cases, complexes with ADP), suggesting thatthese conformations relate to a rigid-body rotationof domain III compared to the rest of the protein.If the binding of ATP does stabilise the confor-mation we observe for the ADPNP complexes onemight expect additional interactions betweendifferent domains of the protein and or inter-actions with the bound nucleoside triphosphatethat might explain this. However, we are unableto identify any additional interactions that mightexplain the stabilisation of the observed confor-mation of the ADPNP complex compared withany of the ADP complexes. It should be noted,however, that although the apo protein can becrystallised in crystal form 1, it cannot becrystallised in crystal form 2 even when ADPNPis added. A likely explanation is that if all threemolecules in the asymmetric unit are able to bindADPNP then the third molecule will be stabilisedin a conformation that is different from that seenin the crystals, presumably the same as in theother two ADPNP complexes. The conformationof the third molecule (containing ADP in crystalform 2) is disfavoured, thereby preventing crys-tallisation in form 2. That being said, the reasonfor the apparent stabilisation of one conformationover any other is unclear. It is unlikely that crystalpacking interactions account for the observedconformations, since they do not appear to beconstraining the position of domain 3, whichadopts the same position in molecules 1 and 2,despite their differing crystallographicenvironments.
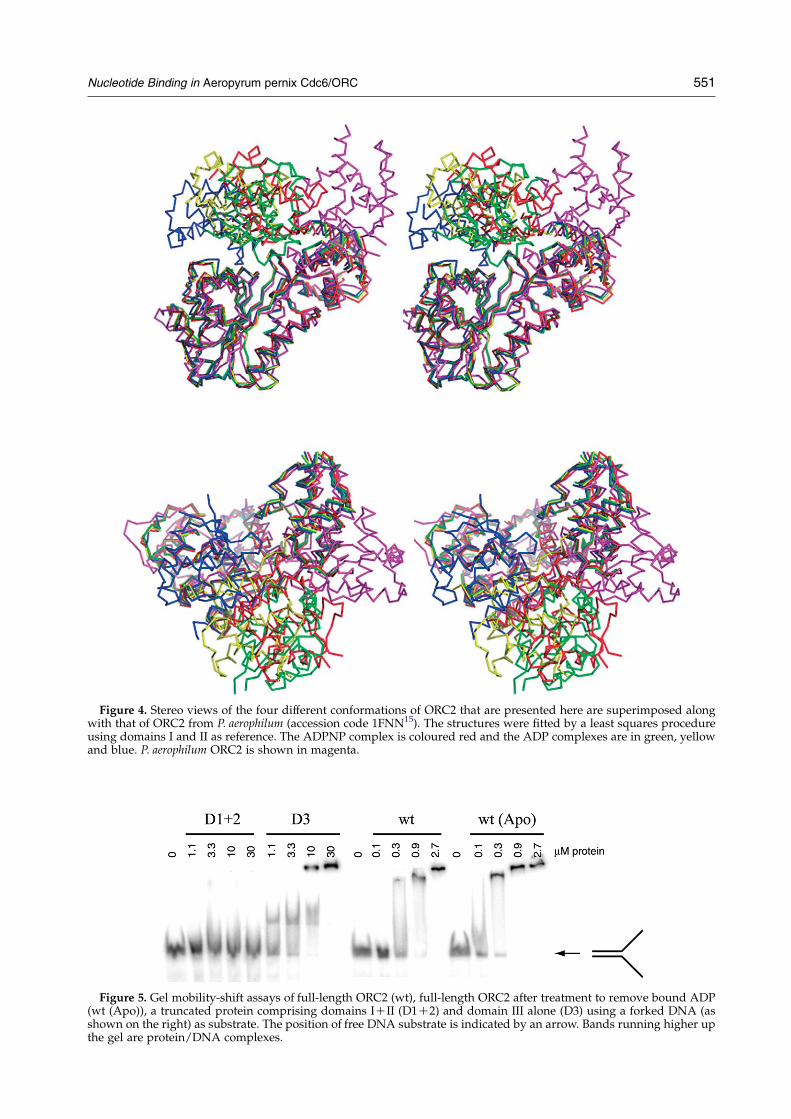
Figure 4. Stereo views of the four different conformations of ORC2 that are presented here are superimposed alongwith that of ORC2 from P. aerophilum (accession code 1FNN15). The structures were fitted by a least squares procedureusing domains I and II as reference. The ADPNP complex is coloured red and the ADP complexes are in green, yellowand blue. P. aerophilum ORC2 is shown in magenta.
Figure 5. Gel mobility-shift assays of full-length ORC2 (wt), full-length ORC2 after treatment to remove bound ADP(wt (Apo)), a truncated protein comprising domains ICII (D1C2) and domain III alone (D3) using a forked DNA (asshown on the right) as substrate. The position of free DNA substrate is indicated by an arrow. Bands running higher upthe gel are protein/DNA complexes.
Nucleotide Binding in Aeropyrum pernix Cdc6/ORC 551
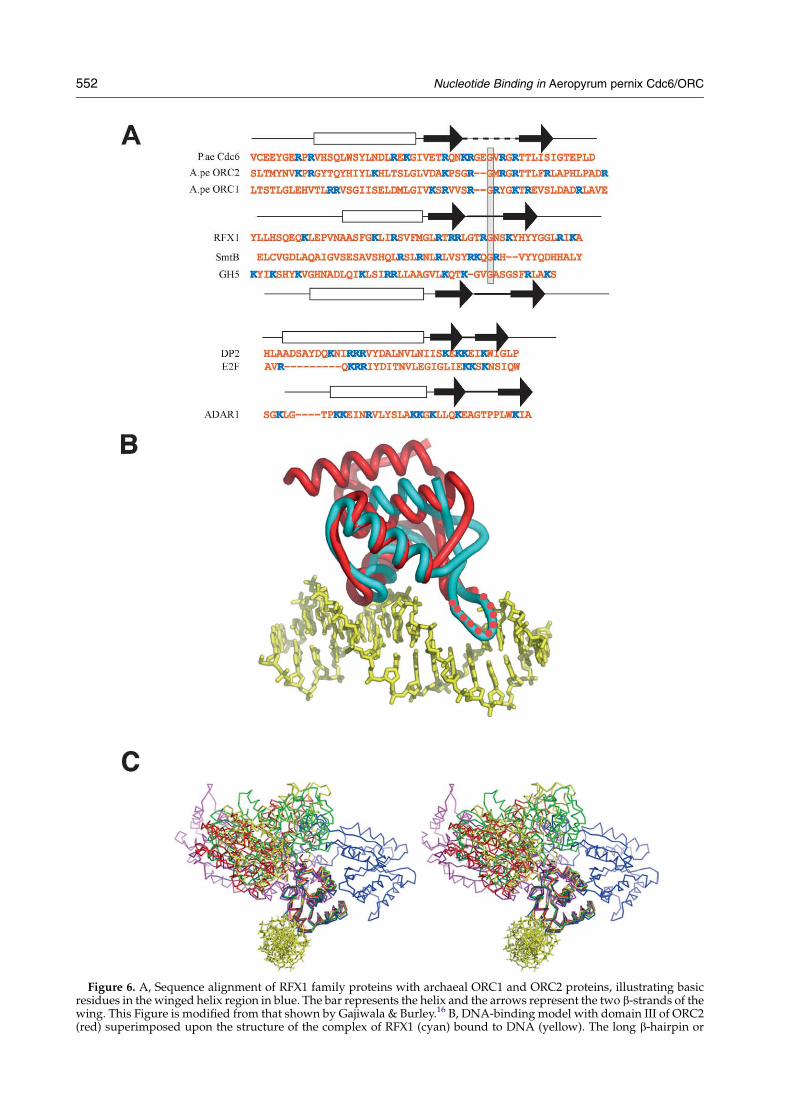
Figure 6. A, Sequence alignment of RFX1 family proteins with archaeal ORC1 and ORC2 proteins, illustrating basicresidues in the winged helix region in blue. The bar represents the helix and the arrows represent the two b-strands of thewing. This Figure is modified from that shown by Gajiwala & Burley.16 B, DNA-binding model with domain III of ORC2(red) superimposed upon the structure of the complex of RFX1 (cyan) bound to DNA (yellow). The long b-hairpin or
552 Nucleotide Binding in Aeropyrum pernix Cdc6/ORC

Nucleotide Binding in Aeropyrum pernix Cdc6/ORC 553
DNA-binding surface
When reporting the structure of P. aerophilumORC/Cdc6, Liu et al. noted a similarity betweendomain III and the DNA-binding domain ofwinged helix (WH) proteins, suggesting that theprotein would be able to bind DNA.15 Thissuggestion has been confirmed experimentally forORC1 of Archaeoglobus fulgidus,11 and for all threeORC/Cdc6 homologues in S. solfataricus.6,18 Wehave confirmed this activity for the A. pernixORC1 and ORC2 proteins using gel mobility-shiftassays (Figure 5 and our unpublished results).Furthermore, in order to determine which domainof ORC2 is able to interact with DNA, we madeconstructs to express domains I and II (residues1–297) and domain III (residues 298–412), separ-ately. These proteins were purified and tested in gelmobility-shift assays (Figure 5). We found that,although the fragment comprising domains I and IIwas unable to interact with DNA, domain III wasable to form a stable complex, albeit with loweraffinity than that of the full-length protein. Thisobservation is consistent with the proposal thatdomain III might be the DNA-binding site on thebasis of the structural similarity between this regionof ORC2 and other known DNA-binding proteinsthat contain the WH motif.15
There are two modes of interaction between WHdomains and DNA.16 The canonical WH proteins(e.g. HNF-3g, E2F) recognize DNA sequence in amanner similar to helix-turn-helix proteins; namely,by inserting the recognition helix into the majorgroove of DNA, while the b-hairpin “wing” inter-acts with the phosphodiester backbone. However,there is a subfamily of WH proteins that interactdifferently with DNA that was first illustrated bythe crystal structure of human RFX1 bound to itsDNA target.19 In the RFX sub-family it is the wingthat is inserted into the major groove, analogous tothe b-ribbon transcription factor family or TATA-binding protein, while the helix of the WH motifmakes a phosphate-backbone contact. Although it isnot straightforward to assign different WH proteinsto each of these families, Gajiwala & Burley domake some proposals.16 They observe that inter-action between WH proteins and their DNA targetsis via their most positively charged surfaces thatinteract with the phosphodiester backbone. Forcanonical WH proteins, this is the so-called recog-nition helix that precedes the wing in the structure,but for the RFX1 family it is the wing rather than thehelix that contains the positively charged residues.In addition, one interaction is observed between theend of the helix and the DNA backbone (Arg48 inRFX1). Therefore, examination of the amino acidsequence of WH domains in this regionmay give an
“wing” of RFX1 can be seen extending across the major grooP. aerophilumORC/Cdc6 and A. pernixORC2 crystal structureon domain III modelled bound to DNA as shown in B. The ADshown in green, yellow and blue. P. aerophilum ORC/Cdc6 is
idea of which mode of binding to DNA is adoptedby the WH protein of interest.With this in mind, we examined the sequences of
the Cdc6/ORC proteins from archaeal species.Several things were immediately apparent. Onesubgroup of proteins (class I) showed very goodsequence homology within the region correspond-ing to the recognition helix, while this sequence wasvery variable in another group of proteins (class II)(Figure 1). It has been reported recently that theS. solfataricus chromosome has at least two replica-tion origins.6 There are three ORC/Cdc6 homol-ogues in this organism and combinations of thesethree proteins bind at the origins. One of theproteins (Cdc6-1, a class I ORC) recognises a specificsequence referred to as an origin recognition box(ORB). Genome analysis revealed the presence ofORB sequences in several other archaeal species.Furthermore, the S. solfataricus class I protein is ableto bind to the ORB sequences at the Pyrococcusfuriosus replication origin.6 We note that there is anapparent correlation between the identification ofORB sequences in the genome and the presence ofan ORC1 protein with a conserved recognitionhelix. Organisms lacking an ORC1 protein (such asP. aerophilum) do not appear to have readily identi-fiable ORB sequences either. Furthermore, we haveshown that the A. pernix ORC1 protein interactswith the ORB sequences (data not shown) at aputative origin in this organism.6 By contrast, theORC2 protein from A. pernix does not bind to ORBsequences but does bind specifically to a differentDNA sequence (data not shown).If the model for DNA binding by P. aerophilum
ORC/Cdc6 proposed by Liu et al. is correct,15 thenrecognition of ORB sequences could be via theconserved recognition helix of ORC1 proteins in themanner they propose. However, since the ORBsequences are conserved, one might expect therecognition helix to be similarly conserved ifinteraction with the DNA were by the canonicalmechanism. Consequently, for the class II proteins,in which this helix is not conserved, an alternativemode of binding is a possibility. Consistent withthis proposal, the number of positively chargedresidues is greater in the wing region than in thehelix for class II proteins (Figure 6A). Theseobservations suggest that binding of these proteinscould be more similar to the RFX1 mode of bindingthan to the canonical mode of recognition of DNAbyWH proteins. This leads us to propose the modelfor DNA binding shown in Figure 6B. This modelwas prepared by superimposing domain III of eachORC2 molecule onto the structure of an RFX1monomer bound to DNA. The model shows howthe hairpin in ORC2 could fit into the major grooveof the DNA, although it is disordered in the crystalstructures in the absence of DNA. This model has
ve of the target DNA. The wing is disordered in both thes. C, Stereo view of all five ORC2 structures superimposedPNP complex is shown in red, and the ADP complexes arein magenta and DNA is shown in yellow.

Figure 7. Model of ORC2 binding to an ORB sequence.
554 Nucleotide Binding in Aeropyrum pernix Cdc6/ORC
implications for the conformations that we observefor ORC2, which can be regarded as rigid-bodymovements of domains I and II relative to domainIII when it is bound to DNA (Figure 6). None of theconformations of ORC2 appears to preclude bind-ing to duplex DNA, at least on steric grounds.Interestingly, a number of non-sequence-specificcontacts between the protein and the DNA back-bone are conserved or replaced by residues thatcould make similar contacts. By contrast, residuesthat make sequence-specific contacts in the majorgroove of the DNA in the RFX1/DNA complex arenot conserved in ORC2.
This model was derived by overlaying domain III of twoORC2 molecules upon the two molecules of RFX1 in thecrystal structure of the complex bound to its recognitionsequence, but with an additional base inserted in thecentral region between the inverted repeats. The confor-mation of ORC2 shown is that with ADPNP bound, butother conformations could be equally appropriate.
Potential for cooperative binding at ORB sites
The DNA-binding sites recognised by RFX1contain two inverted repeats of six bases each(half-sites) separated by a gap of two bases. Forexample, the oligonucleotide sequence used tocrystallise RFX1 was:
CGTTACCATGGTAACG
(half sites underlined).19 The consensus sequencefor ORBs determined recently for archaea alsocomprises two half-sites but with a three base-pairgap between them:6
ATTTTCCAGAGGAAACA
The spacings of the half-sites are similar in theORB and RFX consensus sequences and even thesequences themselves are remarkably similar. Wetherefore propose that both class I and class II Orcproteins may bind DNA containing two invertedhalf-sites. The RFX1/DNA complex appears, there-fore, to be a good model on which to base theinteraction between ORC proteins and DNA.
The RFX1 protein binds as a monomer to each ofthe half-sites in the binding site.19 Binding iscooperative despite a complete lack of contactsbetween the two protein monomers. It is proposedthat cooperativity arises from distortion of the DNAby one protein molecule such that binding of thesecond is favoured. By analogy, we would predictthat two ORC proteins could also bind to each ORBsite at an origin. Ourmodel of the binding of ORC toan ORB (Figure 7) raises some interesting conse-quences when the various conformations of theproteins are considered. As for the RFX1 structure,the DNA-binding regions of the ORC monomers inthe model do not contact each other. However, someof the different conformations of the protein that weobserve in our crystals place domains I and II of thetwo bound molecules in close proximity whenmodelled on an ORB site, while others place themfar apart. It is noteworthy that the closest proximityof these molecules arises with the conformationobserved for the ADPNP complexes (Figure 7). Ourpreliminary data suggest that binding of ORC2 tothe A. pernix origin is tighter to DNA containingmore than one ORB sequence than for isolated ORB
sequences, suggesting a higher order of coopera-tivity between proteins bound at separate ORBsequences (data not shown). How this might beaffected by the conformations that we describe hereis not clear. Furthermore, although it is also quiteplausible that these different conformations mayinfluence interactions with other proteins duringpre-RC assembly and/or origin firing, the details ofany such interactions are unclear at this time.
Experimental Procedures
Protein expression and purification
The gene APE0152 of A. pernix was amplified byPCR from genomic DNA using the oligonucleotideprimers:
5 0 GCATGCCATGGAAGTGCTGAGGCACGGCCTC 3 0
and:
5 0 AGCCCGGATCCTATTACTCGTAGCCTGAGGCCATTTTAGC 3 0
The PCR product was cut with NcoI and BamHI,and inserted into the same sites in pET28a. Theresultant plasmid was transformed into Escherichiacoli strain BL21(DE3) containing a plasmidencoding rare tRNAs (pSJS1240, a gift fromS. J. Sandler).23 Cells were grown in LB at 37 8C toan A600 of 0.5 and then induced with 1 mM IPTG at30 8C for four hours. Cells were harvested bycentrifugation and resuspended in 50 mM Tris(pH 7.5), 100 mM NaCl, 20% (w/v) sucrose, 1 mMEDTA, 140 ng/ml of pepstatin, 400 ng/ml ofleupeptin, 1 mM phenylmethanesulfonyl fluoride(PMSF), 1 mM dithtiothreitol (DTT). Cells weredisrupted by passaging through a homogeniser

Nucleotide Binding in Aeropyrum pernix Cdc6/ORC 555
(Stansted Fluid Instruments) at 15,000 psi(1 psiz6.9 kPa). The lysate was clarified by cen-trifugation at 20,000g for 20 minutes, followed byheat treatment at 55 8C for ten minutes, thenre-centrifugation. 0.4 g/ml of ammonium sulphatewas added to the resulting supernatant, followedby re-centrifugation. The pellet was then resus-pended in 20 mM Tris (pH 7.5), 1 mM EDTA, 1 mMDTT so that the conductivity was around15 mS/cm, and protein was loaded onto a Cibacronblue-Sepharose column equilibrated with buffer A(20 mM Tris (pH 7.5), 50 mM NaCl, 1 mM EDTA,1 mM DTT), washed with several column volumesof buffer and then eluted with buffer AC1 M NaCl.The resultant protein was diluted in buffer A to aconductivity of 20 mS/cm and loaded onto a 5 mlHiTrap heparin column (Pharmacia) equilibrated inbuffer A, washed with several column volumes ofbuffer A and then eluted with a linear gradient ofNaCl up to buffer AC1 M NaCl. The eluate waspassed through a HiLoad 16/60 Superdex 75 gel-filtration column (Pharmacia) equilibrated in20 mM Tris (pH 7.5), 500 mM NaCl, 1 mM EDTA,1 mM DTT. The yield was about 8 mg protein/l ofcells grown up. For concentration prior to crystal-lisation, protein was diluted with buffer until theconductivity was around 20 mS/cm and thenloaded onto a 1 ml Mono S column (Pharmacia) inbuffer A and eluted by a linear gradient of NaCl tobuffer AC1 M NaCl.
Purification of the apo-enzyme
Full-length protein was expressed and purified asdescribed above up to elution from the Cibacronblue column. Pooled fractions from elution wereconcentrated and dialysed overnight against 2 l of50 mM Tris (pH 7.5), 6 M guanidine hydrochloridecontaining activated charcoal. The dialysed proteinwas then diluted with an equal volume of low-saltbuffer (50 mM Tris (pH 7.5), 1 mM EDTA), followedby slow addition of ten volumes of 1 M MgCl2. Theresulting protein was concentrated then dilutedwith low-salt buffer to a conductivity of around20 mS/cm and loaded onto a 5 ml Hi-Trap heparincolumn in buffer A. Elution was with a lineargradient of NaCl to buffer AC1 M NaCl.
Cloning, expression and purification of ORC2domains
Domain ICII was amplified by PCR using thesame forward primer as with the wild-type enzymeand the reverse primer:
5 0 AGCCCGGATCCTTACTAACTGGCCGCCTCGTTCTCGG
The product was cloned and expressed asdescribed for the wild-type (see above). Purificationproceeded as for the wild-type up to resuspensionof the ammonium sulphate pellet. The protein wasthen loaded onto a higher substitution Cibacron
blue-Sepharose column, washed with severalcolumn volumes of buffer and then eluted withbuffer AC1 M NaCl. Saturated ammonium sul-phate solution was added to the protein to bring thefinal concentration of ammonium sulphate to 1 M.The protein was then loaded onto a low-substi-tution phenyl column (Pharmacia) in 50 mM Tris(pH 7.5), 1 mM EDTA, 1 mM DTT, 1 M ammoniumsulphate, and eluted with a linear gradient to bufferA. Eluate was run on a HiLoad 16/60 Superdex 75gel-filtration column (Pharmacia) equilibrated in20 mM Tris (pH7.5), 500 mM NaCl, 1 mM EDTA,1 mM DTT. The yield was about 25 mg of pureprotein per litre of cell culture.Domain III was amplified using the reverse
primer for the wild-type gene and:
5 0 GCATGCCATGGCAATACAAACCCACGAGCTTGAGG
as a forward primer. The product was cloned andexpressed as for the wild-type gene, and the earlystages of purification were identical up to resuspen-sion of the ammonium sulphate pellet. The resus-pended protein was loaded onto a 5 ml Hi-trapheparin column in buffer A and eluted with a lineargradient to buffer AC1 M NaCl. Eluate was dilutedwith low-salt buffer to conductivity w15 mS/cmand loaded onto a Source 30S column in buffer A,washed with several column volumes of buffer andeluted with a linear gradient of NaCl to buffer AC1 MNaCl. The yield was about 9 mg of pure proteinper litre of cell culture.
Gel mobility-shift assay
Oligonucleotide DNA (10 pmol) was labeledusing phage T4 PNK (New England BioLabs) and[g-32P]ATP. Labeled DNAwas annealed to a partlycomplementary oligonucleotide to produce a flayedduplex substrate.11 Binding reactions were set upusing 10 fmol of DNA and protein as indicated in abuffer containing 20 mM Hepes (pH 7.5), 10 mMMgCl2, 1 mM EDTA. After five minutes incuba-tion at room temperature, reactions were electro-phoresed through 6% (w/v) polyacrylamide gel inTBE buffer (90 mM Tris–borate, 1 mM EDTA), at60 V for 60 minutes. Gels were dried and visualisedon a phosphorimager.
Crystallisation, data collection and structuredetermination
Crystals of ADP-ORC2 (form 1) were grown bythe hanging-drop, vapour-diffusion method. ORC2protein was concentrated to 4 mg/ml in 10 mMTris–HCl (pH 7.5), 400 mM NaCl, 1 mM dithio-threitol and drops (2 ml protein:2 ml reservoir) wereequilibrated against a reservoir solution containing100 mM Tris–HCl (pH 7.0), 200 mM Li2SO4, 15%(w/v) PEG 5K MME. Small crystals appeared afterone day and were used as seeds to grow largerrectangular blocks of approximately 80 mm!
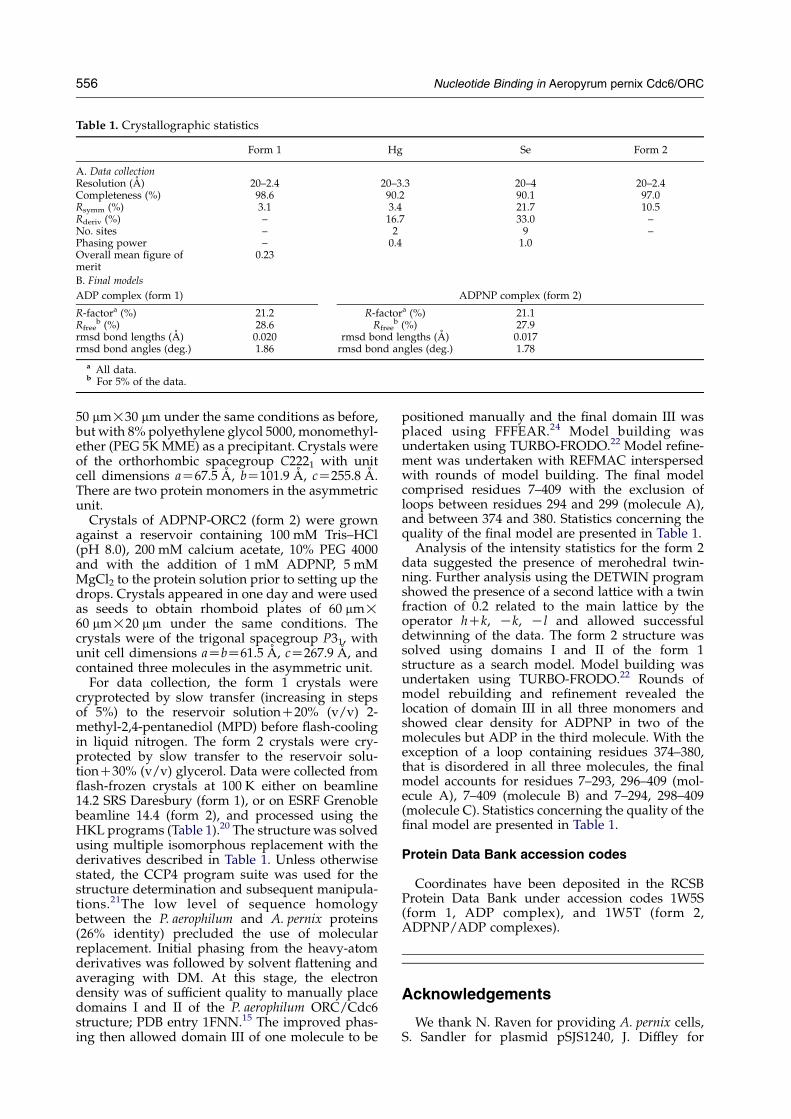
Table 1. Crystallographic statistics
Form 1 Hg Se Form 2
A. Data collectionResolution (A) 20–2.4 20–3.3 20–4 20–2.4Completeness (%) 98.6 90.2 90.1 97.0Rsymm (%) 3.1 3.4 21.7 10.5Rderiv (%) – 16.7 33.0 –No. sites – 2 9 –Phasing power – 0.4 1.0Overall mean figure ofmerit
0.23
B. Final models
ADP complex (form 1) ADPNP complex (form 2)
R-factora (%) 21.2 R-factora (%) 21.1Rfree
b (%) 28.6 Rfreeb (%) 27.9
rmsd bond lengths (A) 0.020 rmsd bond lengths (A) 0.017rmsd bond angles (deg.) 1.86 rmsd bond angles (deg.) 1.78
a All data.b For 5% of the data.
556 Nucleotide Binding in Aeropyrum pernix Cdc6/ORC
50 mm!30 mm under the same conditions as before,but with 8% polyethylene glycol 5000, monomethyl-ether (PEG 5KMME) as a precipitant. Crystals wereof the orthorhombic spacegroup C2221 with unitcell dimensions aZ67.5 A, bZ101.9 A, cZ255.8 A.There are two protein monomers in the asymmetricunit.
Crystals of ADPNP-ORC2 (form 2) were grownagainst a reservoir containing 100 mM Tris–HCl(pH 8.0), 200 mM calcium acetate, 10% PEG 4000and with the addition of 1 mM ADPNP, 5 mMMgCl2 to the protein solution prior to setting up thedrops. Crystals appeared in one day and were usedas seeds to obtain rhomboid plates of 60 mm!60 mm!20 mm under the same conditions. Thecrystals were of the trigonal spacegroup P31, withunit cell dimensions aZbZ61.5 A, cZ267.9 A, andcontained three molecules in the asymmetric unit.
For data collection, the form 1 crystals werecryprotected by slow transfer (increasing in stepsof 5%) to the reservoir solutionC20% (v/v) 2-methyl-2,4-pentanediol (MPD) before flash-coolingin liquid nitrogen. The form 2 crystals were cry-protected by slow transfer to the reservoir solu-tionC30% (v/v) glycerol. Data were collected fromflash-frozen crystals at 100 K either on beamline14.2 SRS Daresbury (form 1), or on ESRF Grenoblebeamline 14.4 (form 2), and processed using theHKL programs (Table 1).20 The structure was solvedusing multiple isomorphous replacement with thederivatives described in Table 1. Unless otherwisestated, the CCP4 program suite was used for thestructure determination and subsequent manipula-tions.21The low level of sequence homologybetween the P. aerophilum and A. pernix proteins(26% identity) precluded the use of molecularreplacement. Initial phasing from the heavy-atomderivatives was followed by solvent flattening andaveraging with DM. At this stage, the electrondensity was of sufficient quality to manually placedomains I and II of the P. aerophilum ORC/Cdc6structure; PDB entry 1FNN.15 The improved phas-ing then allowed domain III of one molecule to be
positioned manually and the final domain III wasplaced using FFFEAR.24 Model building wasundertaken using TURBO-FRODO.22 Model refine-ment was undertaken with REFMAC interspersedwith rounds of model building. The final modelcomprised residues 7–409 with the exclusion ofloops between residues 294 and 299 (molecule A),and between 374 and 380. Statistics concerning thequality of the final model are presented in Table 1.
Analysis of the intensity statistics for the form 2data suggested the presence of merohedral twin-ning. Further analysis using the DETWIN programshowed the presence of a second lattice with a twinfraction of 0.2 related to the main lattice by theoperator hCk, Kk, Kl and allowed successfuldetwinning of the data. The form 2 structure wassolved using domains I and II of the form 1structure as a search model. Model building wasundertaken using TURBO-FRODO.22 Rounds ofmodel rebuilding and refinement revealed thelocation of domain III in all three monomers andshowed clear density for ADPNP in two of themolecules but ADP in the third molecule. With theexception of a loop containing residues 374–380,that is disordered in all three molecules, the finalmodel accounts for residues 7–293, 296–409 (mol-ecule A), 7–409 (molecule B) and 7–294, 298–409(molecule C). Statistics concerning the quality of thefinal model are presented in Table 1.
Protein Data Bank accession codes
Coordinates have been deposited in the RCSBProtein Data Bank under accession codes 1W5S(form 1, ADP complex), and 1W5T (form 2,ADPNP/ADP complexes).
Acknowledgements
We thank N. Raven for providing A. pernix cells,S. Sandler for plasmid pSJS1240, J. Diffley for

Nucleotide Binding in Aeropyrum pernix Cdc6/ORC 557
helpful discussions, and the ESRF, Grenoble fordata collection facilities. This work was supportedby Cancer Research UK.
References
1. Diffley, J. F. X. (1998). Replication control: choreo-graphing replication origins. Curr. Biol. 8, R771–R773.
2. Neuwald, A. F., Aravind, L., Spouge, J. L. & Koonin,E. V. (1999). AAAC: a class of chaperone-like ATPasesassociated with the assembly, operation, and dis-assembly of protein complexes. Genome Res. 9, 27–43.
3. DeRyckere, D., Smith, C. L. &Martin, G. S. (1999). Therole of nucleotide binding and hydrolysis in thefunction of the fission yeast Cdc18(C) gene product.Genetics, 151, 1445–1457.
4. Myllykallio, H., Lopez, P., Lopez-Garcia, P., Heilig, R.,Saurin, W., Zivanovic, Y. et al. (2000). Bacterial modeof replication with eukaryotic-like machinery in ahyperthermophilic archaeon. Science, 288, 2212–2215.
5. Berquist, B. R. & DasSarma, S. (2003). An archaealchromosomal autonomously replicating sequenceelement from an extreme halophile, Halobacteriumsp. strain NRC-1. J. Bacteriol. 185, 5959–5966.
6. Robinson, N. P., Dionne, I., Lundgren, M., Marsh, V. L.,Bernander, R. & Bell, S. D. (2004). Identification of twoorigins of replication in the single chromosome of thearchaeon Sulfolobus solfataricus. Cell, 116, 25–38.
7. Kelman, Z., Lee, J. K. & Hurwitz, J. (1999). The singleminichromosome maintenance protein of Methano-bacterium thermoautotrophicum DeltaH contains DNAhelicase activity. Proc. Natl Acad. Sci. USA, 96,14783–14788.
8. Chong, J. P., Hayashi, M. K., Simon, M. N., Xu, R. M. &Stillman, B. (2000). A double-hexamer archaealminichromosome maintenance protein is an ATP-dependent DNA helicase. Proc. Natl Acad. Sci. USA,97, 1530–1535.
9. Schechter, D. F., Ying, C. Y. & Gautier, J. (2000). Theintrinsic DNA helicase activity of Methanobacteriumthermoautotrophicum delta H minichromosomemaintenance protein. J. Biol. Chem. 275, 15049–15059.
10. Carpentieri, F., De Felice, M., De Falco, M., Rossi, M. &Pisani, F. (2002). Physical and functional interactionbetween the mini-chromosome maintenance-likeDNA helicase and the single-stranded DNA bindingprotein from the crenarchaeon Sulfolobus solfataricus.J. Biol. Chem. 277, 12118–12127.
11. Grainge, I., Scaife, S. C. & Wigley, D. B. (2003).Biochemical analysis of components of the pre-replication complex of Archaeaoglobus fulgidus. Nucl.Acids Res. 31, 4888–4898.
12. Pape, T., Meka, H., Chen, S., Vicentini, G., van Heel,M. & Onesti, S. (2003). Hexameric ring structure of thefull-length archaeal MCM protein complex. EMBORep. 11, 1079–1083.
13. Yu, X., VanLoock, M. S., Poplawski, A., Kelman, Z.,Xiang, T., Tye, B. K. & Egelman, E. H. (2002). TheMethanobacterium thermoautotrophicum MCM proteincan form heptameric rings. EMBO Rep. 8, 792–797.
14. Grabowski, B. & Kelman, Z. (2003). Archaeal DNAreplication: eukaryal proteins in a bacterial context.Annu. Rev. Microbiol. 57, 487–516.
15. Liu, J., Smith, C. L., DeRyckere, D., DeAngelis, K.,Martin, G. S. & Berger, J. M. (2000). Structure andfunction of cdc6/Cdc18: implications for origin recog-nition and checkpoint control.Mol. Cell, 6, 637–648.
16. Gajiwala, K. S. & Burley, S. K. (2000). Winged helixproteins. Curr. Opin. Struct. Biol. 10, 110–116.
17. Grabowski, B. & Kelman, Z. (2001). Autophosphoryl-ation of archaeal Cdc6 homologues is regulated byDNA. J. Bacteriol. 183, 5459–5464.
18. De Felice, M., Esposito, L., Pucci, B., Carpentieri, F.,De Falco, M., Rossi, M. & Pisani, F. M. (2003).Biochemical characterisation of a CDC6-like proteinfrom the crenarchaeon Sulpholobus solfataricus. J. Biol.Chem. 278, 46424–46431.
19. Gajiwala, K. S., Chen, H., Cornille, F., Roques, B. P.,Reith, W., Mach, B. & Burley, S. K. (2000). Structure ofthe winged-helix protein hRFX1 reveals a new modeof DNA binding. Nature, 403, 916–921.
20. Otwinowski, Z. & Minor, W. (1997). Processing ofX-ray diffraction data collected in oscillation mode.Methods Enzymol. 276, 307–326.
21. Collaborative Computing Project Number 4. (1994).The CCP4 suite: programs for protein crystallo-graphy. Acta Crystallog. sect. D, 50, 760–763.
22. Roussel, A. & Cambillau, C. (1989). TURBO-FRODO.Silicon Graphics Geometry Partner Directory, SiliconGraphics, Mountain View, CA pp. 77–78.
23. DelTito,B. J., Jr,Ward, J.M.,Hodgson, J.,Gershater,C. J.,Edwards, H., Wysocki, L. A. et al. (1995). Effects of aminor isoleucyl tRNA on heterologous protein trans-lation in Escherichia coli. J. Bacteriol. 177, 7086–7091.
24. Cowtan, K. (1998). Modified phased translationfunctions and their application to molecular fragmentlocation. Acta Crystallog sect. D, 54, 750–756.
Edited by R. Huber
(Received 22 March 2004; received in revised form 12 August 2004; accepted 12 August 2004)
Recommended

















