Embed Size (px)
Citation preview

DEVELOPMENTAL BIOLOGY 32, 208-213 (1973)
Cell Surface Changes during Preimplantation Development
in the Mouse
PATRICIA G. CALARCO AND CHARLES J. EPSTEIN Departments of Anatomy, Pediatrics, and Biochemistry, University of California, San Francisco 94122
Accepted November 7, 1972
Scanning electron microscopy reveals microvilli on all preimplantation stages, indicates that their number and length may be dependent on embryo size, and provides examples of regional alterations in their number. Cellular adherence, as evidence by interactions of micro- villi, migration of cellular processes, and junctional complexes, increases during development and is accompanied by changes in the shapes of cells and embryos. Cell surfaces bordering the blastocoel differ markedly from the outer cell surfaces of the embryo.
INTRODUCTION
Changes in the cell surface and the de- velopment of cell junctions during pre- implantation development have been de- scribed at the fine structural level in sev- eral mammalian species (Enders and Schlafke, 1965; Mayer et al., 1967; Calarco and Brown, 1969; Enders, 1971). Studies utilizing the scanning electron microscope (SEM) have focused primarily on structures associated with implantation: surface microvilli, craters, and imprints on zona free blastocysts caused by the uterine epi- thelium in delayed implantation (Berg- strom and Nilsson, 1970, 1971; Berg- strom, 1972). However, the SEM is a powerful tool for investigating the number, size, and distribution of microvilli, the shapes of the blastomeres and of the em- bryo itself, and the region of cell-cell con- tact. We have, therefore, investigated the preimplantation stages of mouse develop- ment with the SEM and have observed characteristic changes of the cell surface and of the intracellular regions accompany- ing embryonic development.
MATERIALS AND METHODS
Swiss-Webster female mice were super- ovulated and mated, and eggs were col- lected at various times following fertiliza- tion. Primary oocytes obtained by punc- turing ovarian follicles, and unfertilized
208 Copyright 0 1973 by Academx Press, Inc. All rights of reproduction in any form reserved
superovulated eggs were also collected. For examination of the cell surface the zona pellucida surrounding each embryo was either removed enzymatically with pronase (Mintz, 1962) or manually dis- sected. Routine preparative procedures included primary fixation in 3% glutaral- dehyde in 0.1 M phosphate buffer, post- fixation in 2% osmium tetroxide in 0.1 M phosphate buffer, dehydration through increasing concentrations of ethanol, replacement of ethanol with amyl acetate, and critical point drying. During primary fixation the eggs were placed in empty ant pupae cases (Cooper and Ris, 1943; Mintz, 1971), which were tied shut with fine silk thread. After critical point dry- ing the cases were opened and the eggs removed and gold-coated for viewing on a Kent Cambridge S4 scanning electron microscope.
RESULTS
The acellular zona pellucida is relatively smooth and contains numerous small pits ranging from 70 to 220 nm in diameter at all stages examined. Primary (follicular) oocytes, secondary (metaphase II) oocytes and fertilized eggs possess microvilli evenly distributed on their surfaces. These microvilli are approximately 0.13 pm in diameter and vary from 0.1 pm to over 4 pm in length. While the first polar body is

BRIEF NOTES 209
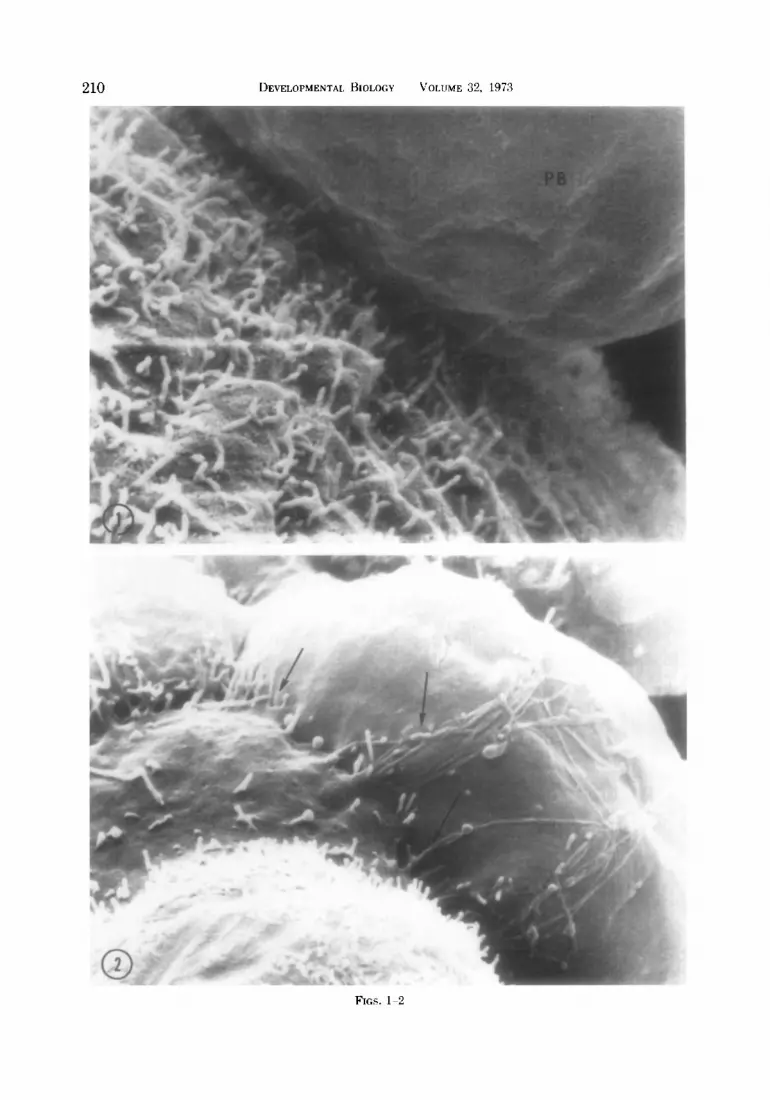
nonmicrovillous (Fig. 1) and is often near a smooth region on the egg’s surface, the second polar body is covered with micro- villi. The even distribution of microvilli persists through the 2-cell and 4-cell stages, and no specialized junctional re- gions are present. If the cells are separated, the area formerly between cells is revealed as somewhat flattened, with patches of smooth membrane alternating with micro- villous areas.
At the B-cell stage smooth portions of membrane are often seen near the regions of blastomere contact. Thin cell proc- esses occasionally extend from one cell across another (Fig. 21, possibly binding the cells more tightly together. Eight cell stages in vitro exhibit varying degrees of cell adhesion (Lewis and Wright, 1935) and examination of sectioned material shows that cell membrane contact becomes quite close at this stage (Calarco and Brown, 1969; Enders, 1971).
The micro,villi of morulae (16-cell em- bryos) are quite variable in number and distribution, and their number often ap- pears increased in the regions of cell con- tact. By transmission EM, terminal bars are known to be present apically between outer cells (presumptive trophoblast) at this time (Enders and Schlafke, 1965; Calarco and Brown, 1969), but they cannot yet be seen directly with the SEM. When a surface cell is dislodged during sample preparation the underlying cells are seen
to be bound together by thin cell proc- esses as described in the B-cell embryo.
During the transition from morula to blastocyst pn day 3 of development, the outer cells progress from a spherical to a flattened shape characteristic of tropho- blast cells, and the area of contact between cells increases strikingly. The surfaces of the day 3 blastocysts range from those which are very microvillous to those which exhibit fewer and shorter microvilli and are nearly smooth, the smoother blasto- cysts being larger in diameter than the microvillous ones. Possibly, blastocysts expand by utilizing the cell membrane present on the microvilli, which diminishes their number and length. On the smoother blastocyts, circular specializations (Fig. 3) are occasionally observed in the regions of cell contact. The spherical day 3 blasto- cysts progress to elongate and often pear- shaped blastocysts by day 4. The cells of 4-day blastocysts are not of uniform size and vary from microvillous to smooth. The significance of the smooth portions of mem- brane may be related to localized areas of membrane addition, mitosis, a randomly fluctuating population of microvilli, or to increased volume. Since microvilli fluctu- ate in number and length, estimates of the total surface area of these embryos have not been attempted.
When blastocysts spontaneously shed their zona pellucidas during day 4 of de- velopment, prominent smooth intercellu-
FIG. 1. Scanning electron micrograph (SEM) of a fertilized egg showing the nonmicrovillous first polar body (PB). x 15,680.
FIG. 2. SEM of portion of an S-cell embryo. Arrows indicate cell processes of one cell extending across another. x 6440.
FIG. 3. SEM of day 3 blastocyst. Note the circular specializations (arrows) between the 3 cells pictured. x 6000.
FIG. 4. SEM of 4-day blastocyst. Note the intercellular ridges. x 1530. FIG. 5. SEM of portion of 4-day blastocyst showing intercellular “junctional ridges” (R) and “Y” struc-
tures (arrows) described in the text. x 9190. FIG. 6. Transmission electron micrograph of a cellular junction from a 4-day blastocyst treated with
pronase, illustrating the apical terminal bar and a subjacent desmosome CD). The region delimited with arrows probably corresponds to the junctional ridges seen by SEM. x 25,670.
FIG. 7. SEM of the inner surface of several trophoblast cells from an opened 4-day blastocyst. Asterisk indicates a region where the cell processes of one cell lie across another. x 2300.
FIG. 8. SEM of the surface of several cells of the inner cell mass facing the blastocoel. Note the gaps between cells. Asterisk indicates where two microvilli of one cell extend across to another cell. x 5100.
210 DEVELOPMENTAL BIOLOGY VOLUME 32, 1973
FIGS. l-2

BRIEF NOTES 211
FIGS. 3-8

212 DEVELOPMENTAL BIOLOGY VOLUME 32, 1973
lar ridges averaging 0.6 pm in diameter are observed (Fig. 4). These ridges are also present between cells of blastocysts whose zonas are mechanically removed, but are not seen on blastocysts treated with pro- nase. (This is the only stage at which pronase has a detectable effect on the sur- face of the embryo.) By transmission EM, desmosomes are frequently found -be- tween the lateral walls of the trophoblast cells, but terminal bars still occupy the most apical position. A region averaging 0.6 pm extending 0.3 pm to either side of the terminal bar, and often projecting ex- ternally from the embryo, contains amor- phous material of medium density, from which cell organelles are excluded. After pronase treatment these 0.6 pm areas are still present at the bases of crypts formed by adjacent cells (Fig. 6). We be- lieve that the surface ridges seen with the SEM in nonpronase treated blastocysts correspond to these terminal bar regions, with the shrinkage which occurs in the preparation of eggs for SEM aiding in their visualization. Although the terminal bar regions are still present, pronase treat- ment appears to increase the convexity of the cells and thereby effectively ob- scures these ridges from view with the SEM. We have seen these “junctional ridges” on other epithelial cell types pre- pared for SEM, but have not found them described in the literature. Occasionally, very short microvilli protrude from a ridge itself. Wherever 3 or more cells come in contact, tripartite “Y” structures which are not continuous with the microvillous portions of the cell surface extend across the junctional ridges (Fig. 5). However, when four cells come in contact an “X” is not formed, but usually two adjacent Y’s (Fig. 5). In one instance, a Y structure was observed across the junctional ridge between 2 cells.
When blastocysts are dissected open, it is possible to compare the surface fea- tures of the two types of cells bordering
the blastocoel. The trophoblast cells are flat and often exhibit microvilli or larger cell processes extending from one cell across a neighboring cell. There are no gaps between trophoblast cells, and inter- cellular boundaries are not marked by surface ridges (Fig. 7). Cells of the inner cell mass, however, are rounder and show gaps between cells reflecting the absence of specialized attachment areas between these cells. Microvilli are occasionally seen crossing from one cell to another at cell-cell boundaries (Fig. 8). Both types of cells are markedly less microvillous than the outer surface of the embryo.
DISCUSSION
There appears to be a progressive series of changes affecting the adherence of embryonic cells to one another. In early stages adherence may be aided by the inter- digitation of microvilli, by the circular specializations seen on smoother surfaces, and by the spreading, which begins at the g-cell stage, of thin cellular processes over the surfaces of adjacent cells. The progres- sion of the outer round blastomeres of the morula to the flattened shape character- istic of trophoblast cells at the blastocyst stage begins at the time of terminal bar formation. In the late blastocysts inter- cellular junctional ridges are seen with the SEM and correspond to the protruding strands reported by Bergstrom (1972). The “Y” shapes crossing these ridges may represent a further specialized structure of cell adherence or communication, or, may only be an accumulation of material where intercellular ridges intersect.
This work was supported by a research grant (HD- 03132) from the National Institutes of Health, U. S. Public Health Service. Charles J. Epstein is the re- cipient of a Research Career Development Award from the National Institutes of Heath. We thank Maria Maglio for excellent technical assistance in the preparation and examination of specimens.

BRIEF NOTES 213
REFERENCES BERGSTR~~M, S. (1972). Delay of blastocysts implanta-
tion in the mouse by ovariectomy or lactation. A scanning electron microscope study. Fert. St&l. 23, 548-561.
BERGSTROM, S., and NILSSON, 0. (1970). Morphological changes of the trophoblast surface and implanta- tion in the mouse. J. Reprod. Fed. 23, 339-340.
BERGSTR~~M, S., and NILSSON, 0. (1971). Scanning electron microscopy of mouse blastocysts before and at implantation. In “Current Problems in Fertility” (A. Ingelman-Sundberg and N.-O. Lunell, eds.), pp. 118-23. Plenum Press, New York.
CALARCO, P. G., and BROWN, E. H. (19691. An ultra- structural and cytological study of preimplantation development of the mouse. J. Exp. Zool. 171, 253- 284.
COOPER, K. W., and Rrs, H. (19431. Handling small objects in bulk for cytological preparations. Stain Technol. 18, 175-6.
ENDERS, A. (1971). The fine structure of the blasto-
cysts. In “The Biology of the Blastocyst” (R. J. Blandau, ed.1, pp. 71-94. Univ. of Chicago Press, Chicago, Illinois.
ENDERS, A. C., and SCHLAFKE, S. J. (1965). In “Pre- implantation Stages of Pregnancy” (G. E. W. Wol- stenholme and M. O’Connor, eds.), pp. 29-54. Little, Brown, Boston, Massachusetts.
LEWIS, W. H., and WRIGHT, E. S. (1935). On the early development of the mouse egg. Carnegie Inst. Contrib. Embryol. 25, 115-143.
MAYER, G., NILSSON, O., and REINIUS, S. (1967). Cell membrane changes of uterine epithelium and trophoblasts during blastocyst attachment in rat. Z. Amt. Entwickhngsgesch. 126, 43.
MINTZ, B. (1962). Experimental study of the develop- ing mammalian egg; removal of the zona pellucida. Science 138, 594-5.
MINTZ, B. (1971). Allophenic mice of multi-embryo origin. In “Methods in Mammalian Embryology” (J. C. Daniel, Jr., ed.), pp. 186-214. Freeman, San Francisco, California.





























