Embed Size (px)
Citation preview

Molecular Cell
Short Article
E3-Independent Monoubiquitinationof Ubiquitin-Binding ProteinsDaniela Hoeller,1,3 Christina-Maria Hecker,1,3 Sebastian Wagner,1 Vladimir Rogov,2 Volker Dotsch,2
and Ivan Dikic1,*1 Institute for Biochemistry II, Goethe University Medical School, Theodor-Stern-Kai 7, 60590 Frankfurt am Main, Germany2 Institut fur Biophysikalische Chemie, Goethe University, Biozentrum, Max-von-Laue-Strasse 9, 60438 Frankfurt am Main,Germany3These authors contributed equally to this work.
*Correspondence: [email protected]
DOI 10.1016/j.molcel.2007.05.014
SUMMARY
Ubiquitin (Ub)-binding domains (UBDs) are keyelements in conveying Ub-based cellular sig-nals. UBD-containing proteins interact withubiquitinated targets and control numerousbiological processes. They themselves undergoUBD-dependent monoubiquitination, whichpromotes intramolecular binding of the UBD tothe attached Ub and leads to their inactivation.Here, we report that, in contrast to the estab-lished ubiquitination pathway, the presence ofUBDs allows the ubiquitination of host proteinsindependently of E3 ligases. UBDs of differenttypes, including UBA, UIM, UBM, NFZ, andUBZ, can directly cooperate with Ub-chargedE2 enzymes to promote monoubiquitination.Using FRET and siRNA technologies, we verifythat Ub-loaded E2 and substrates interact incells and that E2 enzymes are essential for theirmonoubiquitination in vivo. This modification ismechanistically and functionally distinct fromE3-mediated and growth factor-dependentmonoubiquitination.
INTRODUCTION
Ubiquitination is a highly regulated process involving the
action of three enzymes. In the first step, E1 activates
Ub, which is then transferred on an E2 Ub-conjugating
enzyme that finally cooperates with an E3 Ub ligase to
attach Ub to a lysine residue on the substrate (Hershko
and Ciechanover, 1998; Pickart and Eddins, 2004). Differ-
ent types of Ub modifications have distinct cellular
functions: polyUb (linked via Lys-48: proteasomal degra-
dation; linked via Lys-63: endocytosis, DNA repair),
monoUb (signaling, endocytosis, DNA repair), or multiple
monoUb (endosomal sorting and lysosomal degradation)
(reviewed in Haglund and Dikic [2005], Harper and Schul-
man [2006], and Pickart and Fushman [2004]). UBDs are
crucial mediators of Ub-based signals, as they are able
to recognize proteins carrying different types of Ub
Mole
modifications and translate them into the appropriate
cellular response (Hicke et al., 2005; Hurley et al., 2006).
Notably, most of the proteins that are able to bind ubiquitin
noncovalently can be monoubiquitinated themselves (Di
Fiore et al., 2003). This process is called ‘‘coupled mono-
ubiquitination’’ because a functional UBD is a prerequisite
for their self-ubiquitination (Hoeller et al., 2006; Miller et al.,
2004; Polo et al., 2002). Monoubiquitination of UBD-
containing proteins imposes an autoinhibitory conforma-
tion rendering them unable to bind in trans to ubiquitinated
targets, thus providing an intrinsic switch-off mechanism
for Ub-binding proteins (Hoeller et al., 2006).
Some insights into the mechanism of coupled monoubi-
quitination have been gained recently. Two different E3
ubiquitin ligases, the RING-type E3 ubiquitin ligase Parkin
and the HECT-type E3 ligase Nedd4, have been shown
to mediate monoubiquitination of the endocytic protein
Eps15 by mechanisms that require an intact UIM of
Eps15 (Fallon et al., 2006). In the case of Parkin, the UIM
of Eps15 interacts with the ubiquitin-like (Ubl) domain of
Parkin, which in turn mediates the Eps15 monoubiquitina-
tion. In the other case, the Eps15 UIM binds monoubiquitin
attached to Nedd4 via an isopeptide bond (Woelk et al.,
2006). Nedd4 then transfers thiolester-bound ubiquitin
to Eps15. It remains to be determined whether similar
mechanisms apply for coupled monoubiquitination of
UIM-containing proteins other than Eps15, and for mono-
ubiquitination of UBD-containing proteins in general.
In order to further investigate the molecular principles
underlying monoubiquitination of UBD proteins, we have
employed in vitro ubiquitination assays and fluores-
cence-resonance energy transfer (FRET)-based protein
interaction studies in cells. We demonstrate that multiple
UBD-containing proteins can be monoubiquitinated
in vitro independently of the activity of E3 ligases. We
also show that UBDs can directly recruit Ub-loaded E2
enzymes in cells and that E2, but not E3, enzymes
are essential for efficient monoubiquitination of UBD-
containing proteins in vivo.
RESULTS
Monoubiquitination of several UBD-containing proteins is
induced upon overexpression of Ub in different cell types
cular Cell 26, 891–898, June 22, 2007 ª2007 Elsevier Inc. 891
Molecular Cell
Self-Monoubiquitination of Ub-Binding Proteins
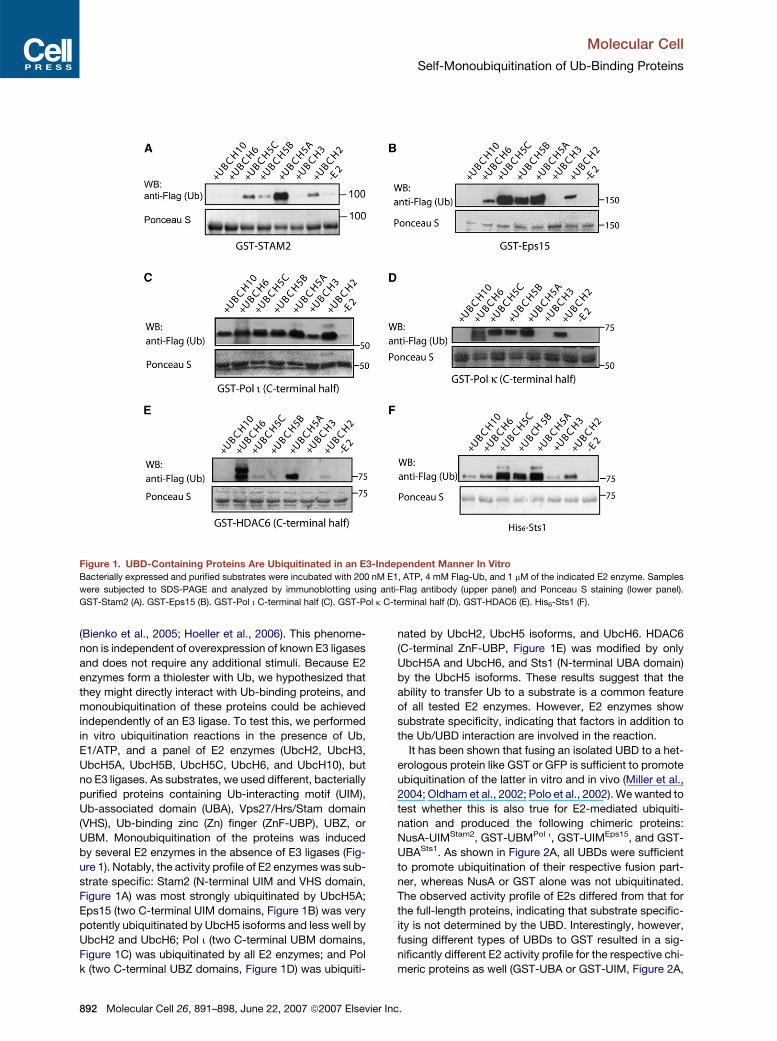
Figure 1. UBD-Containing Proteins Are Ubiquitinated in an E3-Independent Manner In Vitro
Bacterially expressed and purified substrates were incubated with 200 nM E1, ATP, 4 mM Flag-Ub, and 1 mM of the indicated E2 enzyme. Samples
were subjected to SDS-PAGE and analyzed by immunoblotting using anti-Flag antibody (upper panel) and Ponceau S staining (lower panel).
GST-Stam2 (A). GST-Eps15 (B). GST-Pol i C-terminal half (C). GST-Pol k C-terminal half (D). GST-HDAC6 (E). His6-Sts1 (F).
(Bienko et al., 2005; Hoeller et al., 2006). This phenome-
non is independent of overexpression of known E3 ligases
and does not require any additional stimuli. Because E2
enzymes form a thiolester with Ub, we hypothesized that
they might directly interact with Ub-binding proteins, and
monoubiquitination of these proteins could be achieved
independently of an E3 ligase. To test this, we performed
in vitro ubiquitination reactions in the presence of Ub,
E1/ATP, and a panel of E2 enzymes (UbcH2, UbcH3,
UbcH5A, UbcH5B, UbcH5C, UbcH6, and UbcH10), but
no E3 ligases. As substrates, we used different, bacterially
purified proteins containing Ub-interacting motif (UIM),
Ub-associated domain (UBA), Vps27/Hrs/Stam domain
(VHS), Ub-binding zinc (Zn) finger (ZnF-UBP), UBZ, or
UBM. Monoubiquitination of the proteins was induced
by several E2 enzymes in the absence of E3 ligases (Fig-
ure 1). Notably, the activity profile of E2 enzymes was sub-
strate specific: Stam2 (N-terminal UIM and VHS domain,
Figure 1A) was most strongly ubiquitinated by UbcH5A;
Eps15 (two C-terminal UIM domains, Figure 1B) was very
potently ubiquitinated by UbcH5 isoforms and less well by
UbcH2 and UbcH6; Pol i (two C-terminal UBM domains,
Figure 1C) was ubiquitinated by all E2 enzymes; and Pol
k (two C-terminal UBZ domains, Figure 1D) was ubiquiti-
892 Molecular Cell 26, 891–898, June 22, 2007 ª2007 Elsevier
nated by UbcH2, UbcH5 isoforms, and UbcH6. HDAC6
(C-terminal ZnF-UBP, Figure 1E) was modified by only
UbcH5A and UbcH6, and Sts1 (N-terminal UBA domain)
by the UbcH5 isoforms. These results suggest that the
ability to transfer Ub to a substrate is a common feature
of all tested E2 enzymes. However, E2 enzymes show
substrate specificity, indicating that factors in addition to
the Ub/UBD interaction are involved in the reaction.
It has been shown that fusing an isolated UBD to a het-
erologous protein like GST or GFP is sufficient to promote
ubiquitination of the latter in vitro and in vivo (Miller et al.,
2004; Oldham et al., 2002; Polo et al., 2002). We wanted to
test whether this is also true for E2-mediated ubiquiti-
nation and produced the following chimeric proteins:
NusA-UIMStam2, GST-UBMPol i, GST-UIMEps15, and GST-
UBASts1. As shown in Figure 2A, all UBDs were sufficient
to promote ubiquitination of their respective fusion part-
ner, whereas NusA or GST alone was not ubiquitinated.
The observed activity profile of E2s differed from that for
the full-length proteins, indicating that substrate specific-
ity is not determined by the UBD. Interestingly, however,
fusing different types of UBDs to GST resulted in a sig-
nificantly different E2 activity profile for the respective chi-
meric proteins as well (GST-UBA or GST-UIM, Figure 2A,
Inc.
Molecular Cell
Self-Monoubiquitination of Ub-Binding Proteins
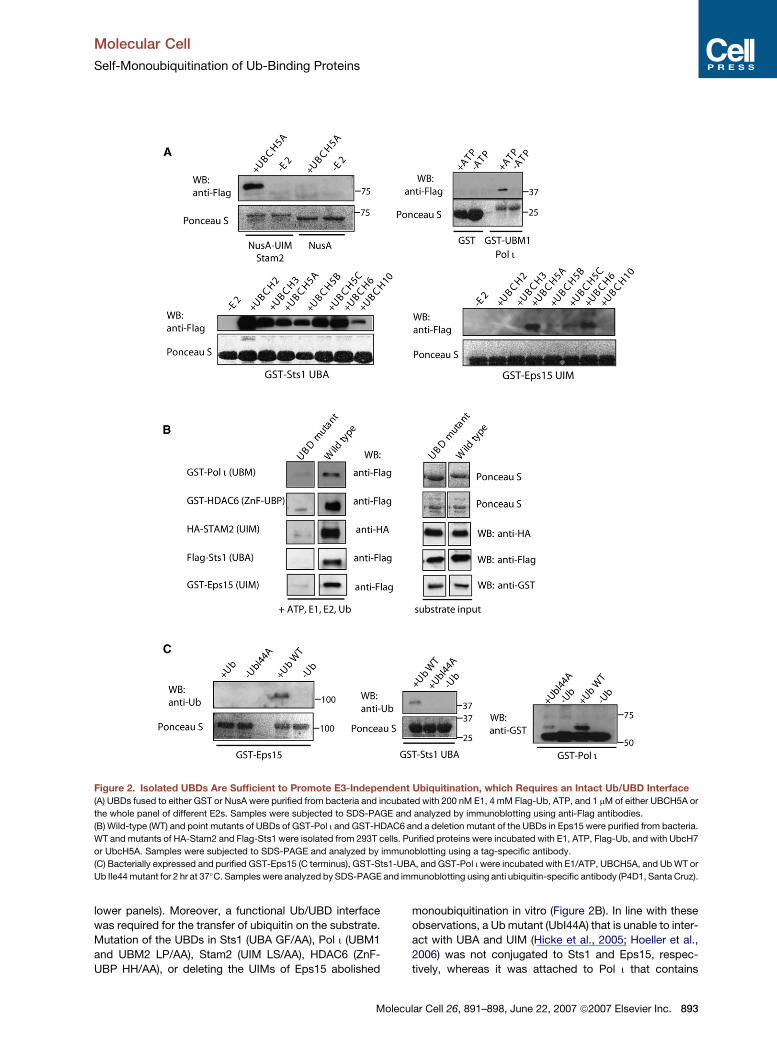
Figure 2. Isolated UBDs Are Sufficient to Promote E3-Independent Ubiquitination, which Requires an Intact Ub/UBD Interface
(A) UBDs fused to either GST or NusA were purified from bacteria and incubated with 200 nM E1, 4 mM Flag-Ub, ATP, and 1 mM of either UBCH5A or
the whole panel of different E2s. Samples were subjected to SDS-PAGE and analyzed by immunoblotting using anti-Flag antibodies.
(B) Wild-type (WT) and point mutants of UBDs of GST-Pol i and GST-HDAC6 and a deletion mutant of the UBDs in Eps15 were purified from bacteria.
WT and mutants of HA-Stam2 and Flag-Sts1 were isolated from 293T cells. Purified proteins were incubated with E1, ATP, Flag-Ub, and with UbcH7
or UbcH5A. Samples were subjected to SDS-PAGE and analyzed by immunoblotting using a tag-specific antibody.
(C) Bacterially expressed and purified GST-Eps15 (C terminus), GST-Sts1-UBA, and GST-Pol i were incubated with E1/ATP, UBCH5A, and Ub WT or
Ub Ile44 mutant for 2 hr at 37�C. Samples were analyzed by SDS-PAGE and immunoblotting using anti ubiquitin-specific antibody (P4D1, Santa Cruz).
lower panels). Moreover, a functional Ub/UBD interface
was required for the transfer of ubiquitin on the substrate.
Mutation of the UBDs in Sts1 (UBA GF/AA), Pol i (UBM1
and UBM2 LP/AA), Stam2 (UIM LS/AA), HDAC6 (ZnF-
UBP HH/AA), or deleting the UIMs of Eps15 abolished
Mo
monoubiquitination in vitro (Figure 2B). In line with these
observations, a Ub mutant (UbI44A) that is unable to inter-
act with UBA and UIM (Hicke et al., 2005; Hoeller et al.,
2006) was not conjugated to Sts1 and Eps15, respec-
tively, whereas it was attached to Pol i that contains
lecular Cell 26, 891–898, June 22, 2007 ª2007 Elsevier Inc. 893
Molecular Cell
Self-Monoubiquitination of Ub-Binding Proteins
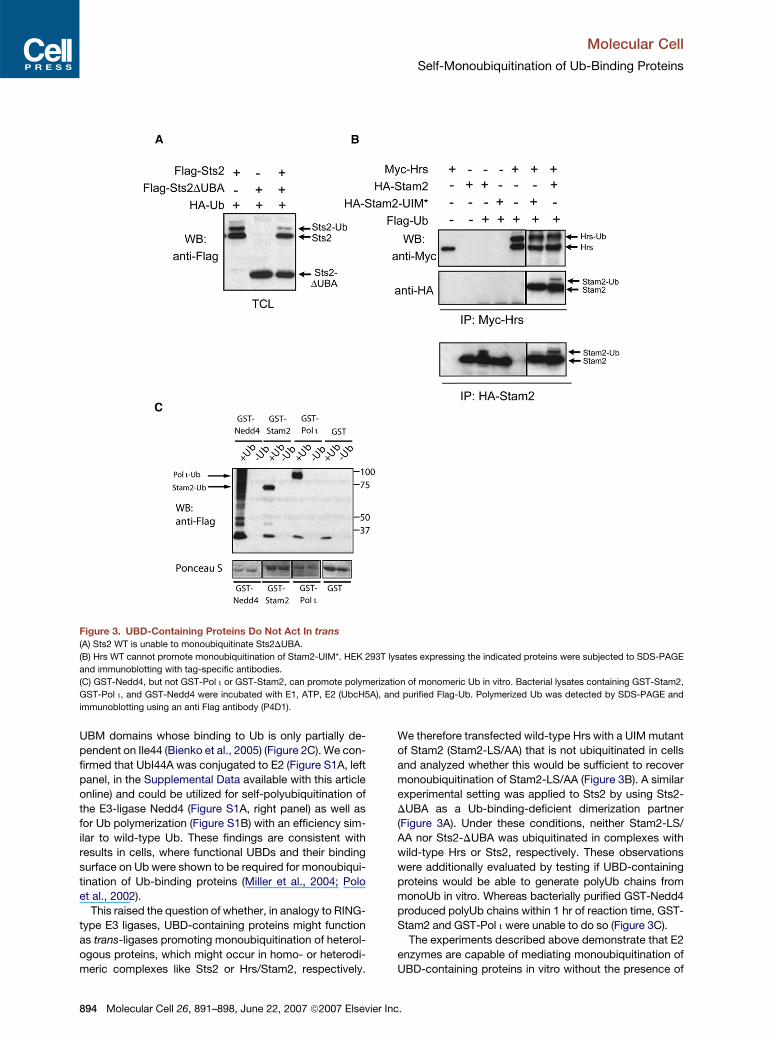
Figure 3. UBD-Containing Proteins Do Not Act In trans
(A) Sts2 WT is unable to monoubiquitinate Sts2DUBA.
(B) Hrs WT cannot promote monoubiquitination of Stam2-UIM*. HEK 293T lysates expressing the indicated proteins were subjected to SDS-PAGE
and immunoblotting with tag-specific antibodies.
(C) GST-Nedd4, but not GST-Pol i or GST-Stam2, can promote polymerization of monomeric Ub in vitro. Bacterial lysates containing GST-Stam2,
GST-Pol i, and GST-Nedd4 were incubated with E1, ATP, E2 (UbcH5A), and purified Flag-Ub. Polymerized Ub was detected by SDS-PAGE and
immunoblotting using an anti Flag antibody (P4D1).
UBM domains whose binding to Ub is only partially de-
pendent on Ile44 (Bienko et al., 2005) (Figure 2C). We con-
firmed that UbI44A was conjugated to E2 (Figure S1A, left
panel, in the Supplemental Data available with this article
online) and could be utilized for self-polyubiquitination of
the E3-ligase Nedd4 (Figure S1A, right panel) as well as
for Ub polymerization (Figure S1B) with an efficiency sim-
ilar to wild-type Ub. These findings are consistent with
results in cells, where functional UBDs and their binding
surface on Ub were shown to be required for monoubiqui-
tination of Ub-binding proteins (Miller et al., 2004; Polo
et al., 2002).
This raised the question of whether, in analogy to RING-
type E3 ligases, UBD-containing proteins might function
as trans-ligases promoting monoubiquitination of heterol-
ogous proteins, which might occur in homo- or heterodi-
meric complexes like Sts2 or Hrs/Stam2, respectively.
894 Molecular Cell 26, 891–898, June 22, 2007 ª2007 Elsevie
We therefore transfected wild-type Hrs with a UIM mutant
of Stam2 (Stam2-LS/AA) that is not ubiquitinated in cells
and analyzed whether this would be sufficient to recover
monoubiquitination of Stam2-LS/AA (Figure 3B). A similar
experimental setting was applied to Sts2 by using Sts2-
DUBA as a Ub-binding-deficient dimerization partner
(Figure 3A). Under these conditions, neither Stam2-LS/
AA nor Sts2-DUBA was ubiquitinated in complexes with
wild-type Hrs or Sts2, respectively. These observations
were additionally evaluated by testing if UBD-containing
proteins would be able to generate polyUb chains from
monoUb in vitro. Whereas bacterially purified GST-Nedd4
produced polyUb chains within 1 hr of reaction time, GST-
Stam2 and GST-Pol i were unable to do so (Figure 3C).
The experiments described above demonstrate that E2
enzymes are capable of mediating monoubiquitination of
UBD-containing proteins in vitro without the presence of
r Inc.

Molecular Cell
Self-Monoubiquitination of Ub-Binding Proteins
E3 enzymes. We next tested if the E3-independent mono-
ubiquitination of Ub-binding proteins also takes place in
cells. Nedd4 and Parkin have been described as E3
ligases responsible for growth factor-induced monoubi-
quitination of Eps15 and Hrs (Fallon et al., 2006; Katz
et al., 2002; Woelk et al., 2006). In order to test if these
ligases are generally required for monoubiquitination of
UBD-containing proteins, we used siRNA against Nedd4
as well as Aip4, a closely related HECT-type ligase, to
knock down expression of these ligases in HeLa cells
that do not possess endogenous Parkin (Fallon et al.,
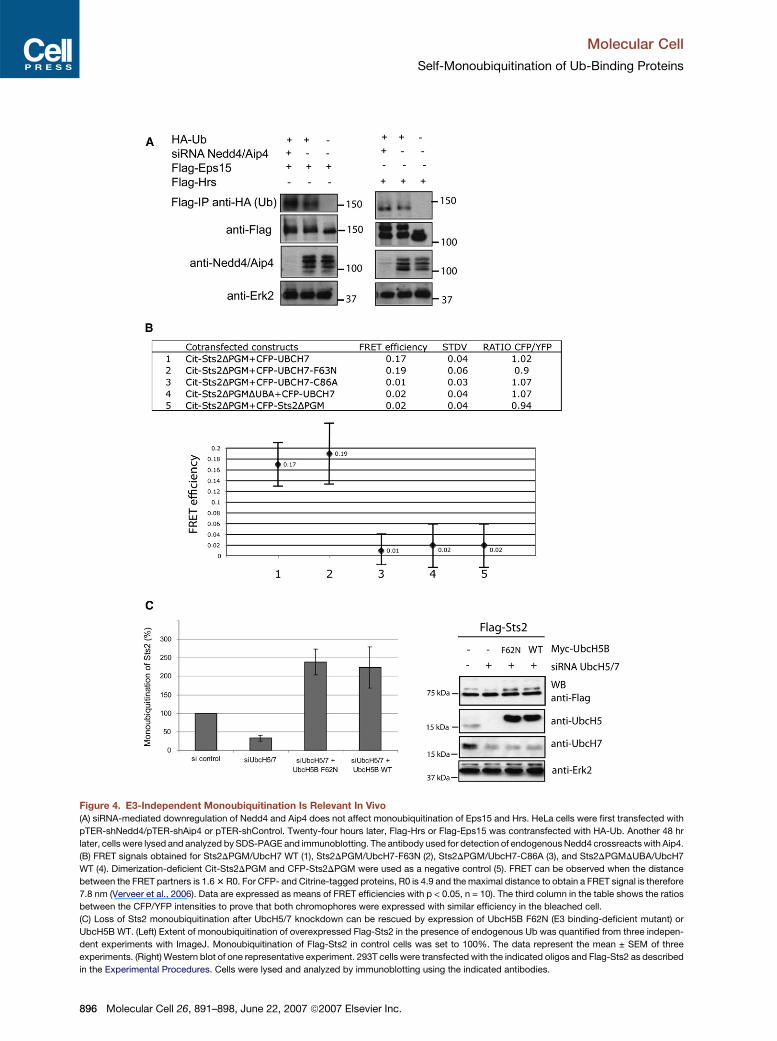
2006). Under these conditions, monoubiquitination of
Eps15, Hrs, and Sts2 was not affected (Figure 4A). We
cannot rule out that a number of so far unidentified ligases
contribute to ubiquitination of these substrates in vivo.
However, the results show that their constitutive monoubi-
quitination is not dependent on the known E3 ligases.
In order to further analyze the interaction between E2
and substrate in vivo, we used FRET after acceptor photo-
bleaching in intact cells. UbcH7 was fused to the FRET do-
nor CFP (CFP-UbcH7), and Citrine-tagged Sts2DPGM (a
Sts2 mutant lacking the dimerization domain) was used
as substrate. These constructs were cotransfected in
HeLa cells, and after photobleaching of the acceptor, an
increase in intensity of donor fluorescence was observed
(Figures S2A–S2C). To evaluate the FRET efficiency, we
calculated the increase of the CFP intensity according to
Bastiaens et al. (1996) for each pair of transfected proteins
(Figure 4B). As a negative control, we used CFP-
Sts2DPGM and Cit-Sts2DPGM, which colocalize but do
not interact with each other due to the deletion of the di-
merization domain (Hoeller et al., 2006). To exclude the
possibility that E3 ligases (such as Cbl) function as a link
between UbcH7 and Sts2, we mutated Phe63 of UbcH7
(CFP-UbcH7-F63N), which is unable to bind to E3 ligases
(Huang et al., 1999; Zheng et al., 2000). Coexpression of
CFP-UbcH7-F63N and Cit-Sts2DPGM led to a similar
FRET signal as compared to the wild-type UbcH7,
whereas the FRET signal between Sts2DPGM and mutant
UbcH7 that cannot be loaded with Ub (UbcH7 C86A). or
Ub-binding-deficient Sts2DPGMDUBA and wild-type
UbcH7 was reduced to background level (Figure 4B).
The FRET experiments thus show that substrate and
Ub-loaded E2 enzyme can directly interact with each
other in cells and that this interaction is dependent on
a functional Ub/UBD interface.
Finally, it was important to determine the significance of
E2-mediated monoubiquitination of UBD proteins in vivo.
We thus tested whether targeted knockdown of selected
E2 enzymes would negatively affect the extent of monou-
biquitination of Sts2 and whether this could be rescued by
expressing an E3 binding-deficient E2 mutant. Among the
E2s tested in the ubiquitination assays, the UbcH5 iso-
forms were the most potent ones for several substrates
(Figure 1). By selectively knocking down the isoforms in
several cell lines, we found that the isoforms UbcH5B
and UbcH5C are expressed in HEK 293T cells (Figure 4C,
right panels). HEK 293T cells also showed substantial ex-
Mol
pression of UbcH7 that interacts with Sts2 in vivo (Fig-
ure 4B) and strongly ubiquitinates Sts2 in in vitro assays
(Figure S1C). We thus targeted these E2 enzymes and
quantified the extent of Sts2 monoubiquitination in
HEK293T cells transfected with Flag-Sts2 in the presence
of endogenous Ub. Downregulation of UbcH5B/C and
UbcH7 led to a marked decrease in Sts2 ubiquitination
compared to control cells (Figure 4C). This effect could be
rescued by transfection of siRNA-resistant Myc-UbcH5B
WT or Myc-UbcH5B F62N, respectively. In fact, overex-
pression of these E2 enzymes led to a more than 2-fold
increase in Sts2 monoubiquitination compared to control
cells (Figure 4C). Importantly, UbcH5B F62N, the E3 bind-
ing uncoupled E2 enzyme (Nuber and Scheffner, 1999),
promoted Sts2 monoubiquitination with a similar effi-
ciency as UbcH5 WT. We would like to note that we can-
not be entirely sure that the F62N (UbcH5)/F63N (UbcH7)
mutation abrogates binding to all classes of E3 ligases.
DISCUSSION
In this report we demonstrate that Ub-binding domains
can confer cis-, but not trans-, ligase activity on host
proteins in vitro and in vivo. This is accomplished by the
direct recruitment of Ub-loaded E2 enzymes into a com-
plex with UBD-containing substrates, a process that is
independent of E3 Ub ligases. Notably, all tested E2
enzymes possessed the ability to participate in the reac-
tion with a different degree of efficiency and substrate
specificity (Figure 1). This specificity could also be
observed with nonphysiological substrates. The type of
UBD (Figure 2A) and residues located outside the Ub/
UBD interface (not all E2s can ubiquitinate a given sub-
strate, Figure 1) appear to be important in defining speci-
ficity of this interaction. The impact of the UBD might be
based on the different Ub-binding modes. In other words,
the Ub-loaded E2 is oriented differently on the substrate
depending on the UBD that recruits it. Efficient substrate
ubiquitination by E2 requires Ub/UBD interactions and
probably additional contact sites that contribute to
substrate positioning. Our preliminary NMR studies do
not imply any extended protein-protein interaction inter-
face between the E2 and (nonphysiological) substrates
but rather suggest a model in which specificity is created
by the positioning of a reactive lysine of the substrate pro-
tein with respect to the conjugated ubiquitin. This might be
defined by steric and electrostatic constraints that are
characteristic for a specific E2-substrate pair.
By downregulating selected E2 enzymes in cells, a
significant reduction of Sts2 monoubiquitination was
achieved in vivo (Figure 4C) that was rescued by an E3
binding-deficient UbcH5B enzyme, indicating that E2-
mediated but E3-independent ubiquitination plays an
important role in modifying UBD-containing proteins
in vivo. The data presented here do not exclude the pos-
sibility that E3 ligases can promote monoubiquitination
of UBD-containing proteins but rather suggest that there
ecular Cell 26, 891–898, June 22, 2007 ª2007 Elsevier Inc. 895
Molecular Cell
Self-Monoubiquitination of Ub-Binding Proteins
Figure 4. E3-Independent Monoubiquitination Is Relevant In Vivo
(A) siRNA-mediated downregulation of Nedd4 and Aip4 does not affect monoubiquitination of Eps15 and Hrs. HeLa cells were first transfected with
pTER-shNedd4/pTER-shAip4 or pTER-shControl. Twenty-four hours later, Flag-Hrs or Flag-Eps15 was contransfected with HA-Ub. Another 48 hr
later, cells were lysed and analyzed by SDS-PAGE and immunoblotting. The antibody used for detection of endogenous Nedd4 crossreacts with Aip4.
(B) FRET signals obtained for Sts2DPGM/UbcH7 WT (1), Sts2DPGM/UbcH7-F63N (2), Sts2DPGM/UbcH7-C86A (3), and Sts2DPGMDUBA/UbcH7
WT (4). Dimerization-deficient Cit-Sts2DPGM and CFP-Sts2DPGM were used as a negative control (5). FRET can be observed when the distance
between the FRET partners is 1.6 3 R0. For CFP- and Citrine-tagged proteins, R0 is 4.9 and the maximal distance to obtain a FRET signal is therefore
7.8 nm (Verveer et al., 2006). Data are expressed as means of FRET efficiencies with p < 0.05, n = 10). The third column in the table shows the ratios
between the CFP/YFP intensities to prove that both chromophores were expressed with similar efficiency in the bleached cell.
(C) Loss of Sts2 monoubiquitination after UbcH5/7 knockdown can be rescued by expression of UbcH5B F62N (E3 binding-deficient mutant) or
UbcH5B WT. (Left) Extent of monoubiquitination of overexpressed Flag-Sts2 in the presence of endogenous Ub was quantified from three indepen-
dent experiments with ImageJ. Monoubiquitination of Flag-Sts2 in control cells was set to 100%. The data represent the mean ± SEM of three
experiments. (Right) Western blot of one representative experiment. 293T cells were transfected with the indicated oligos and Flag-Sts2 as described
in the Experimental Procedures. Cells were lysed and analyzed by immunoblotting using the indicated antibodies.
896 Molecular Cell 26, 891–898, June 22, 2007 ª2007 Elsevier Inc.

Molecular Cell
Self-Monoubiquitination of Ub-Binding Proteins
are multiple ways by which UBD-containing proteins are
regulated in cells. It is likely that the E3-dependent mech-
anism is specific for inducible monoubiquitination after
cell stimulation because E3 ligases are commonly re-
cruited to substrates after their inducible posttranslational
modifications. In accordance, the E3 ligases Nedd4 and
Parkin can facilitate monoubiquitination of Eps15 and
Hrs upon growth factor stimulation (Fallon et al., 2006;
Katz et al., 2002; Polo et al., 2002).
Given the fact that monoubiquitination of most UBD-
containing proteins can occur independently of cell
stimulation (Bienko et al., 2005; Hoeller et al., 2006; Miller
et al., 2004; Polo et al., 2002), cis- ubiquitination seems to
be a constitutively active homeostatic process, which
determines the amount of Ub-binding-competent effector
proteins being available for promoting Ub-dependent
signals. The cellular pool of autoinhibited, monoubiquiti-
nated Ub-binding proteins could be rapidly activated by
deubiquitinating enzymes (DUBs). In this way, DUBs might
be capable of activating/promoting cellular processes
controlled by monoubiquitination such as receptor endo-
cytosis, DNA repair, translesion DNA synthesis, or protein
trafficking. In conclusion, we propose that E2- and UBD-
dependent cis ubiquitination of host proteins represents
an active homeostatic mechanism, which regulates local-
ization, activity, and signaling competence of UBD-
containing proteins in a given physiological state.
EXPERIMENTAL PROCEDURES
Reagents, Cells, Plasmids, and Antibodies
Constructs for Flag-tagged Sts1 and Sts2 have been described
recently (Hoeller et al., 2006). Mouse anti-HA (12CA5) was purchased
from Roche, anti-FLAG (M2 and M5) antibodies from Sigma, and anti-
GST antibodies from Santa Cruz. Affinity-purified rabbit antibodies
against recombinant Hrs have been described previously (Raiborg
et al., 2001).
For overexpression experiments, HEK293T cells were transfected
with Lipofectamine Reagent (Invitrogen) and HeLa cells with Effectene
(Qiagen) according to the manufacturer’s instructions. Cells were
lysed 24–36 hr after transfections.
The construct for bacterial expression of GST-Eps15 was kindly
provided by P.P. Di Fiore (IFOM, Milan, Italy). GST-HDAC6 and GST-
HDAC6 BUZ mutant (H1094 and H1098 changed to A) were described
recently (Boyault et al., 2006) and kindly provided by Cyril Boylault.
Constructs for GST-Pol i (C-terminal half) and GST-UBM1 were de-
scribed recently (Bienko et al., 2005). The GST-Pol i UBM mutant
was created by mutating Leu508/Pro509 (UBM1) and Leu693/
Pro694 (UBM2) of GST-Pol i to alanines by site-directed mutagenesis
(Stratagene). GST-Pol k was created by subcloning both UBZs in
pGEX4T1 using BamH1 and XhoI restriction sites. NusA-UIM of
STAM2 was kindly provided by Dr. Silke Wiesner. GST-UBA of Sts1
was subcloned into pGEX4T1 by using XhoI restriction site. CFP-
Sts2DPGM and Cit-Sts2DPGM were described previously (Hoeller
et al., 2006). CFP-UbcH7 and Cit-UbcH7 were generated by subclon-
ing of full-length UbcH7 into pcDNA3-CFP or pcDNA3-Cit using HindIII
and BamHI restriction sites. CFP-UbcH7-F63, CFP-UbcH7-C68A, and
CFP-Sts2DPGMDUBA were created by site-directed mutagenesis.
pcDNA3-myc-Hrs was kindly provided by H. Stenmark (The Norwe-
gian Radium Hospital, Oslo, Norway). HA-tagged Stam2 for mamma-
lian expression and GST-Stam2 for bacterial expression were kindly
provided by S. Urbe (University of Liverpool, UK). Constructs for mam-
Mole
malian and bacterial expression of Sts1 have been described recently
(Hoeller et al., 2006).
In Vitro Ubiquitination Assays
Reaction mixtures containing 5 mM Ub or UbI44A (Boston Biochem,
Boston, USA), 200 nM E1 (Boston Biochem, Boston, USA), 100 nM–
5 mM E2 (Boston Biochem), and 4 mM ATP in reaction buffer (50 mM
Tris-HCl [pH 7.5], 100 mM NaCl, 2.5 mM MgCl2, and 1 mM DTT) and
the indicated substrate (5–10 mM) were incubated in the absence of
E3 enzyme for 1 hr at 37�C. The reaction was stopped by adding
Laemmli buffer and boiling for 5 min. For preserving thiolester bounds
in E2 enzymes under nonreducing conditions, Laemmli buffer without
b-mercaptoethanol was used.
Immunoprecipitation
HEK 293T or HeLa cells were transfected with the indicated con-
structs, lysed for 10 min on ice in lysis buffer (50 mM HEPES, 150
mM NaCl, 1 mM EDTA, 1 mM EGTA, 10% glycerol, 1% Triton X-100,
25 mM NaF, and 10 mM ZnCl2 [pH 7.5]) containing protease inhibitors
(aprotinin, leupeptin, and PMSF). Cell lysates were collected, centri-
fuged for 15 min (13,000 rpm) to remove the Triton X-100-insoluble
fraction, and incubated with the indicated antibody and Immunosorb
Protein A beads (Sweden) for >3 hr at 4�C. After incubation, the Se-
pharose matrix was washed three times with lysis buffer, and bound
proteins were eluted by boiling the samples for 5 min with Laemmli
buffer containing 5% b-mercaptoethanol. For preserving thiolester
bounds in E2 enzymes, elution was done under nonreducing condi-
tions by using Laemmli buffer without b-mercaptoethanol and boiling
for only 2 min.
siRNA Experiments
Predesigned siRNAs directed against UbcH5A, UbcH5B, UbcH5C,
and UbcH7 were obtained from Perbio (Darmacon, Germany). We
used smartpool oligos (pool of four oligos) for knockdown of UbcH5A,
UbcH5C, and UbcH7. UbcH5B was targeted with two different oligos
binding to the 30UTR to allow for a rescue with UbcH5B F62N and WT,
respectively. To achieve a sufficient downmodulation of the UbcH5
isoforms, 293T cells were transfected with siRNAs using Lipofect-
amine2000 (Invitrogen) 72 hr prior to cell lysis according to the manu-
facturer’s protocol. UbcH7 smartpool oligos were cotransfected with
Flag-Sts2 using Lipofectamine (Invitrogen) 24 hr after transfection of
UbcH5 siRNA. Control cells were treated similarly with scrambled
siRNA. To determine the knockdown efficiency, the cell lysates were
subjected to SDS-PAGE and analyzed by immunoblotting using
a UbcH7-specific antibody (BD transduction labs) and a UbcH5 anti-
body recognizing all three isoforms (BD transduction labs). Knock-
down efficiencies were quantified with ImageJ software. For quan-
tification of Flag-Sts2 monoubiquitination, the ratio between
monoubiquitinated Flag-Sts2 and unmodified Flag-Sts2 was deter-
mined for three independent experiments with ImageJ. The value
obtained in the control cells was set to 100%.
Nedd4 and Aip4 were downmodulated by using a vector-based
approach. shRNA oligos specific for human Nedd4 and human Aip4
were cloned in pTER as described recently (van de Wetering et al.,
2003). HeLa cells were first cotransfected with pTER-Nedd4 and
pTER-Aip4. Twenty-four hours after transfection, cells were split into
6-well cell-culture dishes and transfected the next day with Ub-HA
and the substrate (Flag-Hrs, Flag-Sts1/2, or Flag-Eps15). After an
additional 24 hr, cells were lysed and analyzed by SDS-PAGE and
immunoblotting. The antibody recognizing endogenous Nedd4 cross-
reacts with Aip4 and was purchased from Santa Cruz.
Confocal Microscopy and FRET Analysis
Cells were plated on glass coverslips and transfected with Effectene
Reagent (Qiagen). Twenty-four to thirty-six hours after transfection,
the cells were fixed with 4% paraformaldehyde. All experiments
cular Cell 26, 891–898, June 22, 2007 ª2007 Elsevier Inc. 897

Molecular Cell
Self-Monoubiquitination of Ub-Binding Proteins
were done with the confocal laser-scanning LSM 510 meta micro-
scope from Zeiss.
For FRET analysis, LSM 510 meta software and the following set-
tings were applied: Cyanfluorescent protein (CFP) was measured by
using argon laser 458 nm excitation and 470–500 band-pass emission
filter, whereas Citrine was detected by using argon laser 514 nm
excitation and band pass 530–600. To permit this dual excitation,
the microscope was configured with a 458/514 nm main dichroic filter
and with 515 nm secondary dichroic filter. Photobleaching of Citrine
fluorescence was achieved by irradiation of a 10 3 10 mm big square
with the 514 nm excitation filter at maximum intensity. FRET efficiency
was calculated by normalizing the difference of the donor post- and
prebleach intensity by the postbleach intensity according to E =
(Ipostbleach � Iprebleach)/Ipostbleach (Bastiaens et al. 1996).
Supplemental Data
Supplemental Data include two figures and can be found with this ar-
ticle online at http://www.molecule.org/cgi/content/full/26/6/891/
DC1/.
ACKNOWLEDGMENTS
We thank Silke Wiesner, Sylvie Urbe, Yosef Yarden, Pier Paolo Di
Fiore, and Harald Stenmark for reagents used in these studies. We
are grateful to Harald Stenmark and Rainer Breitling for critical com-
ments on the manuscript. C.M.H is a fellow of the Hessian ministry
of arts and science. This work was supported by grants from the Deut-
sche Forschungsgemeinschaft (DI 931/1-1) and the German-Israeli
Foundation to I.D.
Received: November 30, 2006
Revised: March 15, 2007
Accepted: May 16, 2007
Published: June 21, 2007
REFERENCES
Bastiaens, P.I., Majoul, I.V., Verveer, P.J., Soling, H.D., and Jovin, T.M.
(1996). Imaging the intracellular trafficking and state of the AB5 quater-
nary structure of cholera toxin. EMBO J. 15, 4246–4253.
Bienko, M., Green, C.M., Crosetto, N., Rudolf, F., Zapart, G., Coull, B.,
Kannouche, P., Wider, G., Peter, M., Lehmann, A.R., et al. (2005).
Ubiquitin-binding domains in Y-family polymerases regulate transle-
sion synthesis. Science 310, 1821–1824.
Boyault, C., Gilquin, B., Zhang, Y., Rybin, V., Garman, E., Meyer-
Klaucke, W., Matthias, P., Muller, C.W., and Khochbin, S. (2006).
HDAC6-p97/VCP controlled polyubiquitin chain turnover. EMBO J.
25, 3357–3366.
Di Fiore, P.P., Polo, S., and Hofmann, K. (2003). When ubiquitin meets
ubiquitin receptors: a signalling connection. Nat. Rev. Mol. Cell Biol. 4,
491–497.
Fallon, L., Belanger, C.M., Corera, A.T., Kontogiannea, M., Regan-
Klapisz, E., Moreau, F., Voortman, J., Haber, M., Rouleau, G., Thorar-
insdottir, T., et al. (2006). A regulated interaction with the UIM protein
Eps15 implicates parkin in EGF receptor trafficking and PI(3)K-Akt
signalling. Nat. Cell Biol. 8, 834–842.
Haglund, K., and Dikic, I. (2005). Ubiquitylation and cell signaling.
EMBO J. 24, 3353–3359.
898 Molecular Cell 26, 891–898, June 22, 2007 ª2007 Elsevier
Harper, W., and Schulman, B.A. (2006). Structural complexity in ubiq-
uitin recognition. Cell 124, 1133–1136.
Hershko, A., and Ciechanover, A. (1998). The ubiquitin system. Annu.
Rev. Biochem. 67, 425–479.
Hicke, L., Schubert, H.L., and Hill, C.P. (2005). Ubiquitin-binding
domains. Nat. Rev. Mol. Cell Biol. 6, 610–621.
Hoeller, D., Crosetto, N., Blagoev, B., Raiborg, C., Tikkanen, R.,
Wagner, S., Kowanetz, K., Breitling, R., Mann, M., Stenmark, H., and
Dikic, I. (2006). Regulation of ubiquitin-binding proteins by monoubi-
quitination. Nat. Cell Biol. 8, 163–169.
Huang, L., Kinnucan, E., Wang, G., Beaudenon, S., Howley, P.M., Hui-
bregtse, J.M., and Pavletich, N.P. (1999). Structure of an E6AP-UbcH7
complex: insights into ubiquitination by the E2-E3 enzyme cascade.
Science 286, 1321–1326.
Hurley, J.H., Lee, S., and Prag, G. (2006). Ubiquitin-binding domains.
Biochem. J. 399, 361–372.
Katz, M., Shtiegman, K., Tal-Or, P., Yakir, L., Mosesson, Y., Harari, D.,
Machluf, Y., Asao, H., Jovin, T., Sugamura, K., and Yarden, Y. (2002).
Ligand-independent degradation of epidermal growth factor receptor
involves receptor ubiquitylation and Hgs, an adaptor whose ubiquitin-
interacting motif targets ubiquitylation by Nedd4. Traffic 3, 740–751.
Miller, S.L., Malotky, E., and O’Bryan, J.P. (2004). Analysis of the role of
ubiquitin-interacting motifs in ubiquitin binding and ubiquitylation. J.
Biol. Chem. 279, 33528–33537.
Nuber, U., and Scheffner, M. (1999). Identification of determinants in
E2 ubiquitin-conjugating enzymes required for hect E3 ubiquitin-
protein ligase interaction. J. Biol. Chem. 274, 7576–7582.
Oldham, C.E., Mohney, R.P., Miller, S.L., Hanes, R.N., and O’Bryan,
J.P. (2002). The ubiquitin-interacting motifs target the endocytic adap-
tor protein epsin for ubiquitination. Curr. Biol. 12, 1112–1116.
Pickart, C.M., and Eddins, M.J. (2004). Ubiquitin: structures, functions,
mechanisms. Biochim. Biophys. Acta 1695, 55–72.
Pickart, C.M., and Fushman, D. (2004). Polyubiquitin chains: polymeric
protein signals. Curr. Opin. Chem. Biol. 8, 610–616.
Polo, S., Sigismund, S., Faretta, M., Guidi, M., Capua, M.R., Bossi, G.,
Chen, H., De Camilli, P., and Di Fiore, P.P. (2002). A single motif
responsible for ubiquitin recognition and monoubiquitination in endo-
cytic proteins. Nature 416, 451–455.
Raiborg, C., Bache, K.G., Mehlum, A., Stang, E., and Stenmark, H.
(2001). Hrs recruits clathrin to early endosomes. EMBO J. 20, 5008–
5021.
van de Wetering, M., Oving, I., Muncan, V., Pon Fong, M.T., Brantjes,
H., van Leenen, D., Holstege, F.C., Brummelkamp, T.R., Agami, R.,
and Clevers, H. (2003). Specific inhibition of gene expression using
a stably integrated, inducible small-interfering-RNA vector. EMBO
Rep. 4, 609–615.
Verveer, P.J., Rocks, O., Harpur, A.G., and Bastiaens, P.I.H. (2006).
Imaging protein interactions by FRET Microscopy. Cold Spring Harbor
Protocols. 10.1101/pdb.ip15.
Woelk, T., Oldrini, B., Maspero, E., Confalonieri, S., Cavallaro, E.,
Di Fiore, P.P., and Polo, S. (2006). Molecular mechanisms of coupled
monoubiquitination. Nat. Cell Biol. 8, 1246–1254.
Zheng, N., Wang, P., Jeffrey, P.D., and Pavletich, N.P. (2000). Struc-
ture of a c-Cbl-UbcH7 complex: RING domain function in ubiquitin-
protein ligases. Cell 102, 533–539.
Inc.





























