Embed Size (px)
Citation preview

1989;49:1541-1547. Cancer Res Ritva P. Evarts, Peter Nagy, Harushige Nakatsukasa, et al.
Differentiation of Rat Liver Oval Cells into HepatocytesIn Vivo
Updated version
http://cancerres.aacrjournals.org/content/49/6/1541
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
Research. on September 2, 2014. © 1989 American Association for Cancercancerres.aacrjournals.org Downloaded from
Research. on September 2, 2014. © 1989 American Association for Cancercancerres.aacrjournals.org Downloaded from

[CANCER RESEARCH 49, 1541-1547, March 15, 1989]
In Vivo Differentiation of Rat Liver Oval Cells into HepatocytesRitva P. Evarts,1 Peter Nagy, Harushige Nakatsukasa, Elizabeth Marsden, and Snorri S. Thorgeirsson
Laboratory of Experimental Carcinogenesis, Division of Cancer Etiology, National Cancer Institute, Bethesda, Maryland 20892
ABSTRACT
The Solt-Farber protocol, in the absence of an initiating agent, wasused to examine the precursor-product relationship between oval cellsand hepatocytes in rat liver. The animals were administered 2-acetyla-minofluorene (AAF) by gavage for 2 wk combined with partial hepatec-
tomy 1 wk after administering AAF. Two dose levels of AAF were used:9- and 21-mg total dose for animals in Groups I and II, respectively. |'11|-
Thymidine was administered i.p. to one-half of the animals at Day 6post-partial hepatectomy. Animals were sacrificed 7, 9, 11, and 13 days
after surgery. Only oval cells became labeled on Day 7 in both groups.On Day 9 both labeled oval cells and labeled basophilic hepatocytes werepresent in Group I, whereas in Group II only oval cells remained labeled.On Days 11 and 13 both oval cells and basophilic hepatocytes werelabeled in both groups. The total amount of radioactivity in Group IIlivers remained the same on Day 9 when only labeled oval cells werepresent and on Days 11 and 13 when both labeled oval cells and labeledbasophilic hepatocytes were present. The calculated half-life for baso
philic hepatocytes was about 50 h. The differentiation of oval cells intobasophilic hepatocytes was delayed in Group II as compared to Group I,and the higher dose of AAF also induced the formation of both intestinalmetaplasia and bile duct formation. In situ hybridization with an a-
fetoprotein probe showed a strong expression in groups of typical ovalcells and in cells arranged in duct-like structures. In addition a transient
expression of AFP was also observed in the areas of basophilic hepatocytes 9 to 11 days after partial hepatectomy. Administration of AAFdecreased the level of albumin mRNA in preexisting hepatocytes andcaused a significant decrease of serum albumin. In contrast, oval cellsshowed a strong albumin expression, and basophilic hepatocytes formedislands of albumin-expressing cells.
Oval cells and the foci of early basophilic hepatocytes lacked glucose-6-phosphatase activity. At Day 13 significant numbers of basophilichepatocytes were positive for glucose-6-phosphatase. Oval cells werestrongly 7-glutamyltranspeptidase positive, whereas the foci of basophilichepatocytes were negative for •¿�y-glutamyltranspeptidase.Only occasionally were transiently -y-glutamyltranspeptidase-positive hepatocytes ob
served in basophilic foci. In summary our data indicate that oval cellscan differentiate to hepatocytes and may have an important physiologicalfunction as a source of major serum proteins when hepatocytes are unableto synthesize these proteins.
INTRODUCTIONThe emergence of "a new" cell population that has been
morphologically denned as oval cells is among the first cellularchanges induced by most hepatic carcinogens as well as byseveral toxic agents (1-5). Extensive analysis has suggested thatthe oval cell population is heterogeneous and constitutes sub-populations of epithelial cells having different developmentalpotential (6-8). The origin of oval cells, their physiologicalfunction, and ultimate fate have not been revealed. Although itis generally believed that oval cells and bile ductular cells arerelated, a possible precursor-product relationship with hepatocytes is undefined. Transformation of oval cells to transitionalcells in azo-dye carcinogenesis was the first evidence thattransition occurs between oval cells and hepatocytes (1, 2, 9-12). Several phenotypic markers are known to be common for
Received 8/12/88; revised 12/1/88: accepted 12/7/88.The costs of publication of this article were defrayed in part by the payment
of page charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
1To whom requests for reprints should be addressed, at the National Cancer
Institute, Bldg. 37. Room 3C28, Bethesda, MD 20892.
both oval cells and hepatocytes. Oval cells produce both albumin and AFP2 (13-15). In addition, fetal aldolases A and C
have been observed in oval cells and in transitional cells after3'-methyl-diaminoazobenzene administration as well as in liv
ers from rats fed choline-deficient, ethionine-supplemented diet(16, 17). The presence of these oncodevelopmental proteinsmay result from the proliferation of less-differentiated cellswhich normally express oncodevelopmental genes (17). A possible link between oval cells and immature hepatocytes has beensuggested and is based on antibody binding studies (18, 19).Furthermore, a number of investigators have been able toestablish phenotypically distinct nonparenchymal epithelial celllines from both neonatal and adult rat livers as well as fromrats fed a choline-deficient/DL-ethionine-supplemented dietthat show explicit phenotypic similarities to oval cells (3, 7,20).
We have recently demonstrated, using a combination of AAFand partial hepatectomy, that 3H-labeled thymidine was trans
ferred from oval cells to immature basophilic hepatocytes,suggesting a precursor-product relationship between these cells(21). Furthermore, Marceau and coworkers have recentlyshown that oval cells could be induced with dimethyl sulfoxideor sodium butyrate in vitro to express differentiated hepatocytictraits, such as albumin production and tyrosine aminotrans-ferase activity (22). In this paper we have extended our earlierobservations on the precursor-product relationship betweenoval cells and hepatocytes in the AAF/partial hepatectomyprotocol. By using this model in combination with thymidineincorporation and in situ hybridization, we now confirm thatoval cells give rise to basophilic hepatocytes.
MATERIALS AND METHODS
Treatment of Animals. Male Fischer rats (150 to 180 g) were usedthroughout the experiment. AAF was administered by gavage duringthe 2-wk period, and partial hepatectomy was performed at the midpoint of AAF treatment. The total dose of AAF administered was 9 mgfor Group 1and 21 mg for Group II. [Methyl- 3H]Thymidine (80.9 Ci/
mol; New England Nuclear, Boston, MA; 1 nCi/g of body weight) wasadministered i.p. to four animals in each group starting at Day 6 afterpartial hepatectomy at 8 a.m., 2 p.m., 8 p.m., and 2 a.m. Animals weresacrificed 7, 9, 11, and 13 days after partial hepatectomy (Fig. 1).
Histológica! Methods. Frozen serial sections of caudate lobe wereused for staining with hematoxylin and eosin and for the histochemicaldemonstration of GGT and GoPase. In addition, tissue samples fixedin Bouin's fixative was used for hematoxylin and eosin staining. GGT
staining was done according to the method of Rutenberg et al. (23),and GoPase according to the method of Wachstein and Meisel (24).
In Situ Hybridization. Frozen serial sections from the caudate lobewere used for in situ hybridization studies. Riboprobes for albumin andAFP were prepared as described earlier (25), but were labeled with 35Sto decrease the exposure time. Paraformaldehyde-fixed frozen sectionswere first incubated for 10 min in 5 mM MgCl2 in PBS, followed by a10-min incubation in 0.1 M glycine in 0.2 M Tris buffer (pH 7.4), asdescribed by Lawrence and Singer (26). The slides were then postfixedfor 20 min in 4% paraformaldehyde in PBS (pH 7-8). After rinsing inPBS the slides were dehydrated in ethanol. Prehybridization was per-
1The abbreviations used are: AFP, a-fetoprotein; AAF. 2-acetylaminofluorene;GoPase, glucose-6-phosphatase; GGT, 7-glutamyltranspeptidase; PBS, phosphate-buffered saline: SSC, standard saline citrate.
1541
Research. on September 2, 2014. © 1989 American Association for Cancercancerres.aacrjournals.org Downloaded from
DIFFERENTIATION OF OVAL CELLS
PH
O 9 11 13d
2-AAF 3H-TdR
Fig. 1. AAF was administered during the 2-wk period, and partial hepatectomywas performed at the midpoint of AAF treatment. Thymidine ('H-TdR) was
administered at Day 6 after partial hepatectomy. Animals were sacrificed at 7, 9,11, and 13 days after partial hepatectomy (I'll)
formed for 2 h at 47°C.The prehybridization mixture was similar to
that described by Shivers et al. (27). In order to reduce the nonspecificbinding, 100 nmol/ml of a-thiouridinetriphosphate (New England Nuclear) were added to the prehybridization mixture as described byBandtlow et al. (28). Hybridization with 35S-labeled probes was performed for 3 h at 48°Caccording to Shivers et al. (27).
RNase Treatment and Washings. The slides hybridized with 35S-labeled probes were washed for 20 min at S2*C in 1:1 dilution of
formamide and 2x SSC, rinsed 4 times in 2x SSC, air dried, andtreated with heat-denatured RNase A (Sigma Chemical Company, St.Louis, MO) (40 itg/ml) and RNase Ti (Boehringer-Mannheim, WestGermany) (250 units/ml) at 37°Cin humidified plastic boxes for 30
min. Slides were again washed with 1:1 dilution of formamide and 2x
SSC for 10 min at 52°C.The slides were then washed overnight with
0.5X SSC with two changes over 18-h period and with O.lx SSC for 1h at room temperature.
Autoradiography. After washing, the slides were dehydrated first with70% ethanol/0.3 M ammonium acetate and then in 95% ethanol/0.3 Mammonium acetate. The air-dried slides were immersed in Kodak NTB-2 emulsion diluted 1:1 with 0.6 M ammonium acetate at 45°Cand air
dried in the dark for 1 h. The slides were kept in dry light-protectedboxes for 6 to 14 days at 4"C. The slides were then developed in Kodak
D-19 developer for 4 min, submersed in water for 15s, and fixed in
Kodfix solution. The slides were counterstained with hematoxylin andeosin.
Automated Image Analysis. Magiscan image analysis system (Joyce-Lobel, England), equipped with in situ software, was used for thequantitative determination of silver grains after autoradiography ofslides probed for AFP. The number of silver grains on each slide wasdetermined over 15 fields (500 unr each) of basophilic and acidophilichepatocyte areas. The number of silver grains on the glass slide wasused as a background. The average size of the basophilic and acidophilichepatocytes on each slide was determined by the Magiscan imageanalysis system.
Determination of Liver Radioactivity. Silver grains in 200 nuclei were
y¡'¿*&?*&
SÃi/^iÃft*«:
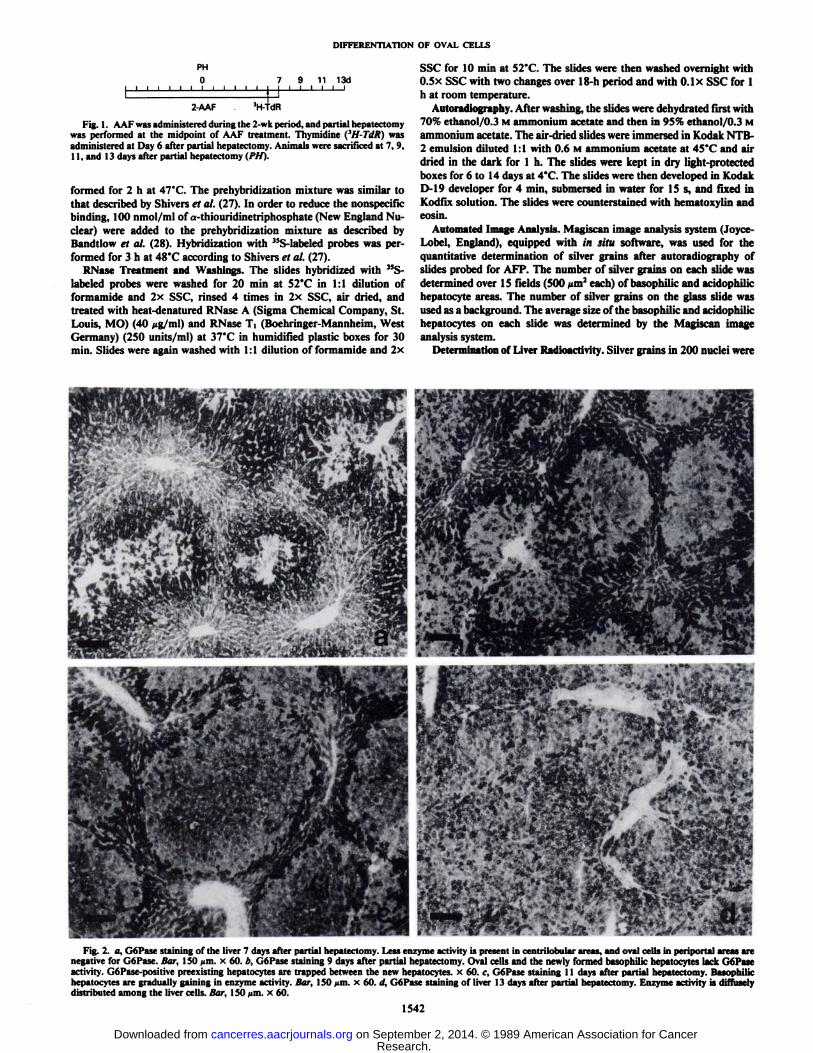
Fig. 2. a, G6Pase staining of the liver 7 days after partial hepatectomy. Less enzyme activity is present in centrilobular areas, and oval cells in periportal areas arenegative for GoPase. Hur. 150 urn. x 60. h. GoPase staining 9 days after partial hepatectomy. Oval cells and the newly formed basophilic hepatocytes lack GoPaseactivity. G6Pase-positive preexisting hepatocytes are trapped between the new hepatocytes. x 60. c, G6Pase staining 11 days after partial hepatectomy. Basophilichepatocytes are gradually gaining in enzyme activity. Bar, ISO uni- x 60. d, G6Pase staining of liver 13 days after partial hepatectomy. Enzyme activity is diffuselydistributed among the liver cells. Bar, ISO *<m.x 60.
1542
Research. on September 2, 2014. © 1989 American Association for Cancercancerres.aacrjournals.org Downloaded from
DIFFERENTIATION OF OVAL CELLS
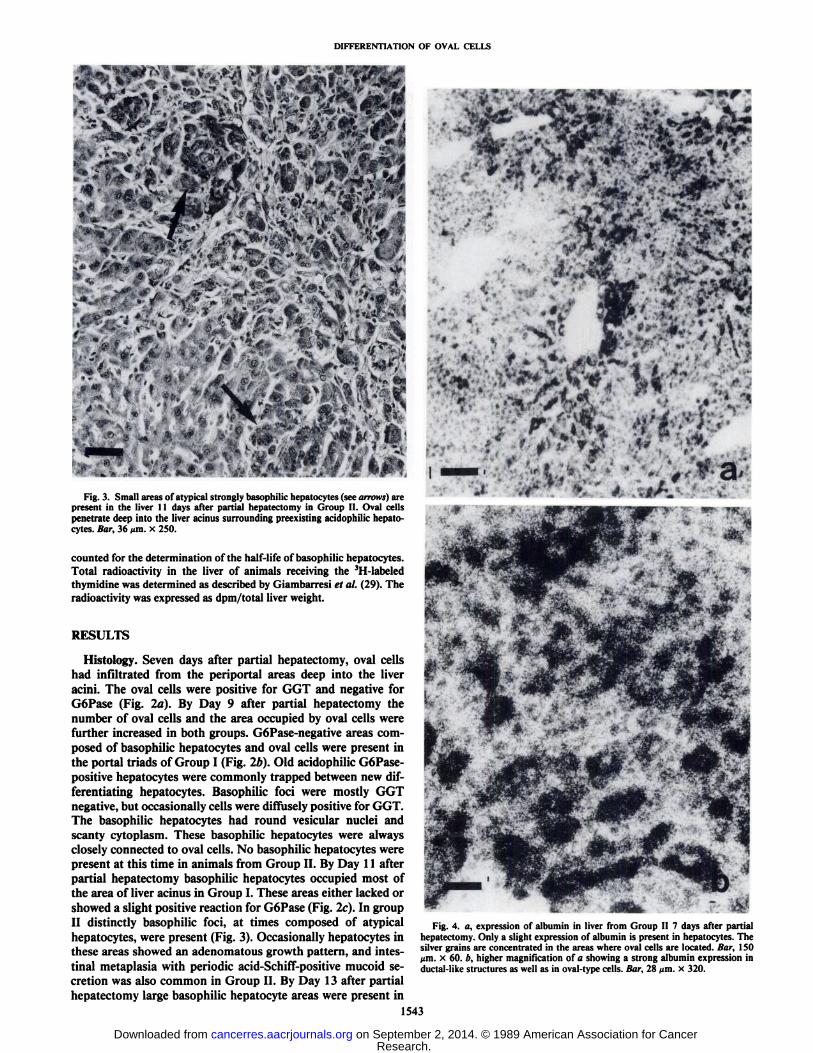
Fig. 3. Small areas of atypical strongly basophilic hepatocytes (see arrows) arepresent in the liver 11 days after partial hepatectomy in Group II. Oval cellspenetrate deep into the liver acinus surrounding preexisting acidophilic hepatocytes. Bar, 36 um. x 250.
counted for the determination of the half-life of basophilic hepatocytes.Total radioactivity in the liver of animals receiving the 3H-labeled
thymidine was determined as described by Giambarresi et al. (29). Theradioactivity was expressed as dpm/total liver weight.
RESULTS
Histology. Seven days after partial hepatectomy, oval cellshad infiltrated from the periportal areas deep into the liveracini. The oval cells were positive for GGT and negative forGoPase (Fig. la). By Day 9 after partial hepatectomy thenumber of oval cells and the area occupied by oval cells werefurther increased in both groups. GoPase-negative areas composed of basophilic hepatocytes and oval cells were present inthe portal triads of Group I (Fig. 2b). Old acidophilic GoPase-positive hepatocytes were commonly trapped between new differentiating hepatocytes. Basophilic foci were mostly GGTnegative, but occasionally cells were diffusely positive for GGT.The basophilic hepatocytes had round vesicular nuclei andscanty cytoplasm. These basophilic hepatocytes were alwaysclosely connected to oval cells. No basophilic hepatocytes werepresent at this time in animals from Group II. By Day 11 afterpartial hepatectomy basophilic hepatocytes occupied most ofthe area of liver acinus in Group I. These areas either lacked orshowed a slight positive reaction for GoPase (Fig. 2c). In groupII distinctly basophilic foci, at times composed of atypicalhepatocytes, were present (Fig. 3). Occasionally hepatocytes inthese areas showed an adenomatous growth pattern, and intestinal metaplasia with periodic acid-Schiff-positive mucoid secretion was also common in Group II. By Day 13 after partialhepatectomy large basophilic hepatocyte areas were present in
Fig. 4. a, expression of albumin in liver from Group II 7 days after partialhepatectomy. Only a slight expression of albumin is present in hepatocytes. Thesilver grains are concentrated in the areas where oval cells are located. Bar, 150/mi. x 60. b, higher magnification of a showing a strong albumin expression inductal-like structures as well as in oval-type cells. Bar, 28 ^m. x 320.
1543
Research. on September 2, 2014. © 1989 American Association for Cancercancerres.aacrjournals.org Downloaded from
DIFFERENTIATION OF OVAL CELLS
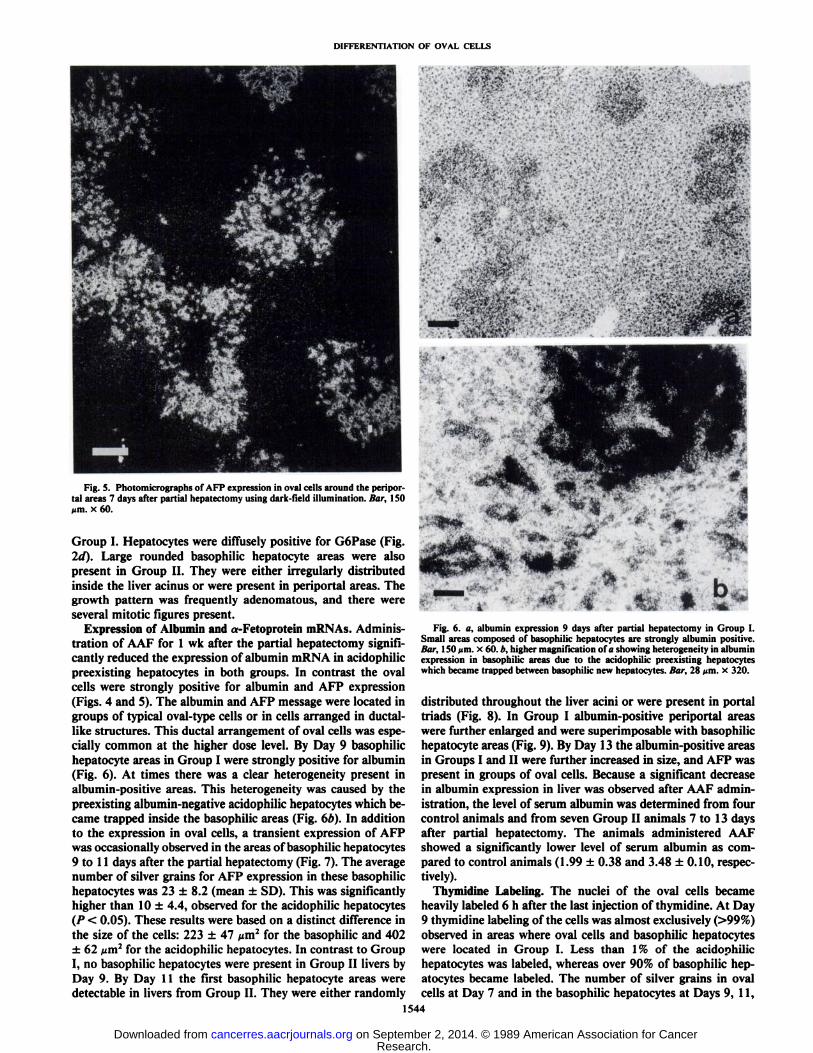
Fig. 5. Photomicrographs of AFP expression in oval cells around the peripor-tal areas 7 days after partial hepatectomy using dar k-field illumination. Bar, ISOMm. x 60.
Group I. Hepatocytes were diffusely positive for GoPase (Fig.2d). Large rounded basophilic hepatocyte areas were alsopresent in Group II. They were either irregularly distributedinside the liver acinus or were present in periportal areas. Thegrowth pattern was frequently adenomatous, and there wereseveral mitotic figures present.
Expression of Albumin and a-Fetoprotein mRNAs. Administration of AAF for 1 wk after the partial hepatectomy significantly reduced the expression of albumin mRNA in acidophilicpreexisting hepatocytes in both groups. In contrast the ovalcells were strongly positive for albumin and AFP expression(Figs. 4 and 5). The albumin and AFP message were located ingroups of typical oval-type cells or in cells arranged in ductal-like structures. This ductal arrangement of oval cells was especially common at the higher dose level. By Day 9 basophilichepatocyte areas in Group I were strongly positive for albumin(Fig. 6). At times there was a clear heterogeneity present inalbumin-positive areas. This heterogeneity was caused by thepreexisting albumin-negative acidophilic hepatocytes which became trapped inside the basophilic areas (Fig. 6/>). In additionto the expression in oval cells, a transient expression of AFPwas occasionally observed in the areas of basophilic hepatocytes9 to 11 days after the partial hepatectomy (Fig. 7). The averagenumber of silver grains for AFP expression in these basophilichepatocytes was 23 ±8.2 (mean ±SD). This was significantlyhigher than 10 ±4.4, observed for the acidophilic hepatocytes(P < 0.05). These results were based on a distinct difference inthe size of the cells: 223 ±47 ^m2 for the basophilic and 402±62 /urn2for the acidophilic hepatocytes. In contrast to Group
I, no basophilic hepatocytes were present in Group II livers byDay 9. By Day 11 the first basophilic hepatocyte areas weredetectable in livers from Group II. They were either randomly
tFig. 6. a, albumin expression 9 days after partial hepatectomy in Group I.
Small areas composed of basophilic hepatocytes are strongly albumin positive.liar. 1SO/mi. x 60. b, higher magnification of a showing heterogeneity in albuminexpression in basophilic areas due to the acidophilic preexisting hepatocyteswhich became trapped between basophilic new hepatocytes. Bar, 28 /¡m.x 320.
distributed throughout the liver acini or were present in portaltriads (Fig. 8). In Group I albumin-positive periportal areaswere further enlarged and were superimposable with basophilichepatocyte areas (Fig. 9). By Day 13 the albumin-positive areasin Groups I and II were further increased in size, and AFP waspresent in groups of oval cells. Because a significant decreasein albumin expression in liver was observed after AAF administration, the level of serum albumin was determined from fourcontrol animals and from seven Group II animals 7 to 13 daysafter partial hepatectomy. The animals administered AAFshowed a significantly lower level of serum albumin as compared to control animals (1.99 ±0.38 and 3.48 ±0.10, respectively).
Thymidine Labeling. The nuclei of the oval cells becameheavily labeled 6 h after the last injection of thymidine. At Day9 thymidine labeling of the cells was almost exclusively (>99%)observed in areas where oval cells and basophilic hepatocyteswere located in Group I. Less than 1% of the acidophilichepatocytes was labeled, whereas over 90% of basophilic hepatocytes became labeled. The number of silver grains in ovalcells at Day 7 and in the basophilic hepatocytes at Days 9, 11,
1544
Research. on September 2, 2014. © 1989 American Association for Cancercancerres.aacrjournals.org Downloaded from

DIFFERENTIATION OF OVAL CELLS
Fig. 7. Dark field photomicrograph of basophilic area (delineated by arrows)hybridized with a 35S-labeled riboprobe for AFP. The number of silver grains in
basophilic small hepatocytes is significantly higher than in the surroundinghepatocytes. Oval cells inside the basophilic area show a strong AFP expression.Bar, 36 ¡im.X 250.
,'«•..•¿�•¿� ' •¿�;;;.' ' •¿�,
fei
Fig. 9. Expression of albumin 11 days after partial hepatectomy in Group I.Albumin-positive areas are superimposable with basophilic hepatocytes. liar, 150¿im.X 60.
200 -
C/3=> 100
iZ¡fiZ
OILOocHIffl2
10
t V, = 50 hr
9 11
DAYS AFTER PH
13
Fig. 10. | 'l l [Umiliili ne was administered to 16 animals on Day 6 after partial
hepatectomy, and the animals were sacrificed at Days 7, 9, 11, and 13 after theoperation. The number of silver grains in oval cells at Day 7 and in the basophilichepatocytes at Days 9, II, and 13 was counted. The half-life obtained forbasophilic hepatocytes is 50 h.
Fig. 8. Expression of albumin 11 days after partial hepatectomy in Group II.Few albumin-positive small areas are distributed throughout the liver acini. Bar,ISO^m. x60.
and 13 was used for estimation of half-life. A half-life of 50 hwas obtained for this pool of dividing hepatocytes (Fig. 10). InGroup II only oval cells were labeled at Day 9. However, atDays 11 and 13 most basophilic hepatocytes in this groupbecame heavily labeled with thymidine (Fig. 11). However, therewere always a significant number of labeled oval cells presentat later time points in both groups.
Total Radioactivity of the Liver. In order to exclude the
possibility of the reutilization of 3H-labeled thymidine from
dying labeled cells by the newly formed basophilic hepatocytes,the total liver radioactivity in Group II was determined at Day9, when only oval cells were present and at Days 11 and 13,when large basophilic areas could be seen. As shown in Fig. 12,total radioactivity remained constant.
DISCUSSION
In the "resistant hepatocyte model" for hepatocarcinogenesis,
administration of AAF combined with partial hepatectomy isused to promote growth of early preneoplastic and neoplasticcell populations (30). In the absence of promotion only a fewpreneoplastic and neoplastic lesions are seen (31). Thereforethe promoting regimen plays a significant part in the eventsleading to neoplastic tumor formation. Important features of
1545
Research. on September 2, 2014. © 1989 American Association for Cancercancerres.aacrjournals.org Downloaded from
DIFFERENTIATION OF OVAL CELLS
-
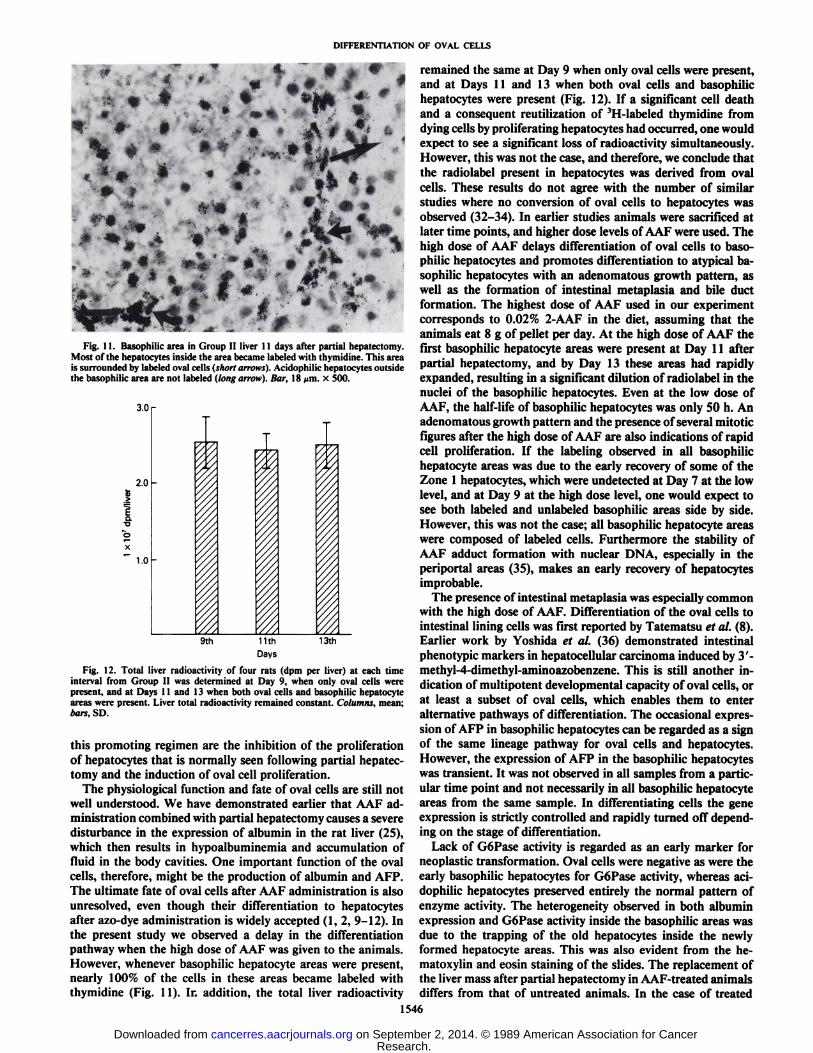
Fig. 11. Basophilic area in Group II liver 11 days after partial hepatectomy.Most of the hepatocytes inside the area became labeled with thymidine. This areais surrounded by labeled oval cells (short arrows). Acidophilic hepatocytes outsidethe basophilic area are not labeled (long arrow). Bar, 18 >im. x 500.
3.0
2.0
•¿�o'b
1.0
9th 11th
Days
13th
Fig. 12. Total liver radioactivity of four rats (dpm per liver) at each timeinterval from Group II was determined at Day 9, when only oval cells werepresent, and at Days 11 and 13 when both oval cells and basophilic hepatocyteareas were present. Liver total radioactivity remained constant. Columns, mean;bars, SD.
this promoting regimen are the inhibition of the proliferationof hepatocytes that is normally seen following partial hepatectomy and the induction of oval cell proliferation.
The physiological function and fate of oval cells are still notwell understood. We have demonstrated earlier that AAF administration combined with partial hepatectomy causes a severedisturbance in the expression of albumin in the rat liver (25),which then results in hypoalbuminemia and accumulation offluid in the body cavities. One important function of the ovalcells, therefore, might be the production of albumin and AFP.The ultimate fate of oval cells after AAF administration is alsounresolved, even though their differentiation to hepatocytesafter azo-dye administration is widely accepted (1,2, 9-12). Inthe present study we observed a delay in the differentiationpathway when the high dose of AAF was given to the animals.However, whenever basophilic hepatocyte areas were present,nearly 100% of the cells in these areas became labeled withthymidine (Fig. 11). In addition, the total liver radioactivity
remained the same at Day 9 when only oval cells were present,and at Days 11 and 13 when both oval cells and basophilichepatocytes were present (Fig. 12). If a significant cell deathand a consequent reutilization of '11-laheled thymidine from
dying cells by proliferating hepatocytes had occurred, one wouldexpect to see a significant loss of radioactivity simultaneously.However, this was not the case, and therefore, we conclude thatthe radiolabel present in hepatocytes was derived from ovalcells. These results do not agree with the number of similarstudies where no conversion of oval cells to hepatocytes wasobserved (32-34). In earlier studies animals were sacrificed atlater time points, and higher dose levels of AAF were used. Thehigh dose of AAF delays differentiation of oval cells to basophilic hepatocytes and promotes differentiation to atypical basophilic hepatocytes with an adenomatous growth pattern, aswell as the formation of intestinal metaplasia and bile ductformation. The highest dose of AAF used in our experimentcorresponds to 0.02% 2-AAF in the diet, assuming that theanimals eat 8 g of pellet per day. At the high dose of AAF thefirst basophilic hepatocyte areas were present at Day 11 afterpartial hepatectomy, and by Day 13 these areas had rapidlyexpanded, resulting in a significant dilution of radiolabel in thenuclei of the basophilic hepatocytes. Even at the low dose ofAAF, the half-life of basophilic hepatocytes was only 50 h. Anadenomatous growth pattern and the presence of several mitoticfigures after the high dose of AAF are also indications of rapidcell proliferation. If the labeling observed in all basophilichepatocyte areas was due to the early recovery of some of theZone 1 hepatocytes, which were undetected at Day 7 at the lowlevel, and at Day 9 at the high dose level, one would expect tosee both labeled and unlabeled basophilic areas side by side.However, this was not the case; all basophilic hepatocyte areaswere composed of labeled cells. Furthermore the stability ofAAF adduct formation with nuclear DNA, especially in theperiportal areas (35), makes an early recovery of hepatocytesimprobable.
The presence of intestinal metaplasia was especially commonwith the high dose of AAF. Differentiation of the oval cells tointestinal lining cells was first reported by Tatematsu et al. (8).Earlier work by Yoshida et al. (36) demonstrated intestinalphenotypic markers in hepatocellular carcinoma induced by 3'-methyl-4-dimethyl-aminoazobenzene. This is still another indication of multipotent developmental capacity of oval cells, orat least a subset of oval cells, which enables them to enteralternative pathways of differentiation. The occasional expression of AFP in basophilic hepatocytes can be regarded as a signof the same lineage pathway for oval cells and hepatocytes.However, the expression of AFP in the basophilic hepatocyteswas transient. It was not observed in all samples from a particular time point and not necessarily in all basophilic hepatocyteareas from the same sample. In differentiating cells the geneexpression is strictly controlled and rapidly turned off depending on the stage of differentiation.
Lack of GoPase activity is regarded as an early marker forneoplastic transformation. Oval cells were negative as were theearly basophilic hepatocytes for GoPase activity, whereas aci-dophilic hepatocytes preserved entirely the normal pattern ofenzyme activity. The heterogeneity observed in both albuminexpression and GoPase activity inside the basophilic areas wasdue to the trapping of the old hepatocytes inside the newlyformed hepatocyte areas. This was also evident from the he-matoxylin and eosin staining of the slides. The replacement ofthe liver mass after partial hepatectomy in AAF-treated animalsdiffers from that of untreated animals. In the case of treated
1546
Research. on September 2, 2014. © 1989 American Association for Cancercancerres.aacrjournals.org Downloaded from

DIFFERENTIATION OF OVAL CELLS
animals, the liver has to rely on a more primitive cell populationto provide functional hepatocytes. After partial hepatectomy ofthe untreated animals no change in albumin expression wasobserved at any time after the operation (data not shown).When partial hepatectomy is performed on AAF-treated animals the replacement of liver is similar to that seen during fetallife; both AFP and albumin are present. The expression of AFPdisappears, while that of albumin remains. The uninitiatedversion of the Solt-Farber model offers many opportunities forstudying the gene expression in the proliferating and differentiating stem cell population. Furthermore, using the completeSolt-Farber protocol, exploration of gene expression, in bothinitiated and uninitiated hepatocytes, is now feasible by usingthe in situ hybridization technique.
REFERENCES
1. Price, J. M, Harman, J. W., Miller, E. C, and Miller, J. A. Progressivemicroscopic alterations in the livers of rats fed hepatic carcinogens 3'-methyl-4-dimethylaminoazobenzene and 4'-fluoro-4-dimethylaminoazobenzene.Cancer Res., 12:192-200, 1952.
2. Farber, E. Similarities in the sequence of early histological changes inducedin the liver of the rat by ethionine, 2-acetylaminofluorene, and 3' -methyl-4-dimethylaminoazobenzene. Cancer Res., 16: 142-148, 1956.
3. Sell, S., and I .effort, H. L. An evaluation of cellular lineages in the pathogen-esis of experimental hepatocellular carcinoma. Hepatology, 2: 77-86, 1982.
4. Wilson, J. W., and I-educe. E. H. Role of cholangioles in restoration of theliver of the mouse after dietary injury. J. Pathol. Bacterio!., 76: 441-449,1958.
5. Lopez, M., and Mazzanti, L. Experimental investigation on a-naphthyliso-thiocyanate as a hyperplastic agent of the biliary ducts in the rat. J. Pathol.Bacterio!., 69: 243-250, 1955.
6. Dunsford, H. A., Naret, R., and Sell, S. Connection of chemical hepatocar-cinogen-induced oval cell derived duct-like structures to bile ducts in rat liverdemonstrated by injection of bile ducts with a pigmented gelatin medium.Proc. Am. Assoc. Cancer Res., 25:77, 1984.
7. Fausto, N., Thompson, N. L., and Braun, L. Purification and culture of ovalcells from rat liver. Cell Separation Methods, 4:45-77, 1987.
8. Tatematsu, M., Kaku. T., Mediine, A., and Farber, E. Intestinal metaplasiaas a common option of oval cells in relation to cholangiofibrosis in liver ofrats exposed to 2-acetylaminofluorene. Lab. Invest., 52: 354-362, 1985.
9. lanaoka, Y. Significance of the so-called oval cell proliferation during azo-dye hepatocarcinogenesis. Gann, 58: 355-366, 1967.
10. Ogawa, K., Minase, T., and Onore, T. Demonstration of glucose-6-phospha-tase activity in the oval cells of the rat liver and the significance of the ovalcells in azo dye carcinogenesis. Cancer Res., 34: 3379-3386,1974.
11. Onda, H. Immunohistochemical studies in alpha-fetiiprotcin and alpha ¡-acidglycoprotein during azodye hepatocarcinogenesis in rats. Gann, 67:253-262,1976.
12. Dempo, K., Chisaka, N., Yoshida, Y., Kaneko, A., and Onoe, T. Immunoflu-orescent study on alpha-fetoprotein-producing cells in the early stage of 3'-methyl-4-dimethylaminoazobenzene carcinogenesis. Cancer Res., 35: 1282-1287, 1975.
13. Sell, S. Distribution of a-fetoprotein- and albumin-containing cells in thelivers of Fischer rats fed four cycles on A'-2-fluorenylacetamide. Cancer Res.,J«:3107-3113, 1978.
14. Sell, S., and Salman, J. Light- and electron-microscopic autoradiographicanalysis of proliferating cells during the early stages of chemical hepatocarcinogenesis in the rat induced by feeding .V-2-fluoren\ lacciamide in a choline-deficient diet. Am. J. Pathol., 114: 289-300, 1984.
15. Yaswen, P., Hayner, N. T., and Fausto, N. Isolation of oval cells by centrif
ugal elutriation and comparison with other cell types purified from normaland preneoplastic livers. Cancer Res., 44: 324-331, 1984.
16. Weber, A., Le Provost, E., Boisnard-Rissel, M., Berges, J., Schapira, F., andGuillouzo, A. Localization of fetal aldolase during early stages of azo-dyehepatocarcinogenesis in rat. Biochem. Biophys. Res. Commun., 92: 591-597, 1980.
17. Hayner, N. T., Braun, L., Yaswen, P., Brooks, M., and Fausto, N. Isozymeprofiles of oval cells, parenchyma! cells, and biliary cells isolated by centrifugal elutriation from normal and preneoplastic livers. Cancer Res., 44: 332-338, 1984.
18. Hixon, D. C., and Allison, J. P. Monoclonal antibodies recognizing oval cellsinduced in the liver of rats by W-2-fluorenylacetamide or ethionine in acholine-deficient diet. Cancer Res., 45: 3750-3760, 1985.
19. Germain, L., Goyette, R., and Marceau, N. Differential cytoken«in and a-fetoprotein expression in morphologically distinct epithelial cells emergingat the early stage of rat hepatocarcinogenesis. Cancer Res., 45: 673-681,1985.
20. Grisham, J. W. Cell types in long-term propagable cultures of rat liver. Ann.NY Acad. Sci., 349:128-137, 1980.
21. Evarts, R. P., Nagy, P., Marsden, E. R., and Thorgeirsson, S. S. A precursor-product relationship exists between oval cells and hepatocytes in rat liver.Carcinogenesis (Lond.), «.-1737-1740, 1987.
22. Germain, L., Noel, M., Gourdeau, H., and Marceau, N. Promotion of growthand differentiation of rat ductular oval cells in primary culture. Cancer Res.,¥«.-368-378,1988.
23. Rutenberg, A. M., Kim, H., Fishbein, J. W., Hanker, J. S., Waserkrug, H.L., and Seligman, A. M. Histochemical and ultrastructural demonstration off-glutamyltranspeptidase activity. J. Histochem. Cytochem., 17: 517-526,1969.
24. Wachstein, M., and Meisel, E. On the histochemical demonstration ofglucose-6-phosphatase. J. Histochem. Cytochem., 4: 592, 1956.
25. Evarts, R. P., Nagy, P., Marsden, E. R., and Thorgeirsson, S. S. In situhybridization studies on expression of albumin and a-fetoprotein during earlystage of neoplastic transformation in rat liver. Cancer Res., 47: 5469-5475,1987.
26. Lawrence, J. B., and Singer, R. H. Quantitative analysis of in situ hybridization methods for the detection of actin gene expression. Nucleic AcidsRes., 13:1777'-1799, 1985.
27. Shivers, B. D., Schachter, B. S., and Pfaff, D. W. In situ hybridization forthe study of gene expression in the brain. Methods Enzymol., 124:497-510,1986.
28. Bandtlow, C. E., Heimann, R., Schwab, M. E., and Thoenen, H. Cellularlocalization of nerve growth factor synthesis by in situ hybridization. EMBOJ., 0:891-899, 1987.
29. Giambarresi, L. I., Katyal, S. L., and Lombardi, B. Promotion of livercarcinogenesis in the rat by a choline-devoid diet: role of liver cell necrosisand regeneration. Br. J. Cancer, 46: 825-829, 1982.
30. Solt, D., and Farber, E. New principle for the analysis of chemical carcinogenesis. Nature (Lond.), 263: 701-703, 1976.
31. Farber, E. Response of liver to carcinogens: a new analytical approach. In:E. Farber and M. M. Fisher (eds.), Toxic Liver Injury, pp. 445-467. NewYork: Marcel Dekker, 1979.
32. Tatematsu, M., Kaku, T., Ekem, J. K., and Farber, E. Studies on theproliferation and fate of oval cells in the liver of rats treated with 2-acetylaminofluorene and partial hepatectomy. Am. J. Pathol., 114:418-430,1984.
33. Grisham, J. W., and Porta, E. A. Origin and fate of proliferated hepaticductal cells in the rat: electron microscopic and autoradiographic studies.Exp. Mol. Pathol., 3: 242-261, 1964.
34. Rubin, E. The origin and fate of proliferated bile ductular cells. Exp. Mol.Pathol., 3:279-286, 1986.
35. Huitfeldt, H. S., Spangler, E. F., Hunt, J. M., and Poirier, M. C. Immunohistochemical localization of DNA adducts in rat liver tissue and phenotyp-ically altered foci during oral administration of 2-acetyl-aminofluorene. Carcinogenesis (Lond.), 7:123-129, 1986.
36. Yoshida, Y., Kaneko, A., Chisaka, N., and Onoe, T. Appearance of intestinaltype of tumor cells in hepatoma tissue induced by 3'-methyl-4-dimethyla-minoazobenzene. Cancer Res., 38: 2753-2758, 1978.
1547
Research. on September 2, 2014. © 1989 American Association for Cancercancerres.aacrjournals.org Downloaded from





























