Embed Size (px)
Citation preview

Journal of Clinical Immunology, Vol. 16, No. 4, 1996
Interleukin- 10 (IL- 10) Secretion in Systemic Lupus Erythematosus and Rheumatoid Arthritis: IL-10-Dependent CD4+CD45RO+ T Cell-B Cell Antibody Synthesis
MANSOUR AL-JANADI, t ABDULLAH AL-DALAAN, 2 SULIMAN AL-BALLA, t MOHAMMED AL-HUMAIDI, 1 and SYED RAZIUDDIN 2'3
Accepted. February 23, 1996
Interleukin-10 (IL-10) is a major immunoregulatory cytokine and has a multitude of immunomodulatory effects in the immune system. In this study, we have examined the secretion and in vitro function of IL-10 in B cell hyperactivity in antibody production in two common autoimmune diseases, systemic lupus erythematosus (SLE) and rheumatoid arthritis (RA). IL-10 was detectable in serum of all active SLE and serum and synovial fluid samples of all RA patients but in none of the normal controls. B cells and CD4+CD45RO+ "memo- ry" T cells secreted highly enhanced levels of IL-10 in SLE and RA versus normals. Increased IgM and IgG production by B cells-CD4+CD45RO+ T cells in SLE and RA was IL-10 dependent, since neutralization of IL- 10 cytokine by anti-IL- 10 antibody drastically reduced Ig synthesis in these coculture experiments. B cell hyperactivity in autoantibody production in SLE and RA may be a function of IL-10-dependent CD4+CD45RO+ Th2 cell activation. Therefore, IL-10 may play an important role in highly disturbed immune system and B cell-T cell function in these immune disorders.
KEY WORDS: Interleukin-10; CD4+CD45RO+ T cells; autoimmu- nity; systemic lupus erythematosus; rheumatoid arthritis.
INTRODUCTION
Interleukin-10 (IL-10) was initially identified as a T cell-derived cytokine and mainly a CD4+ Th2 cell response product that inhibited Thl proliferation by down regulating interferon-gamma (IFN-y) and IL-2 production (1). However, subsequent studies have shown
1Department of Internal Medicine (Rheumatology), King Saud Univer- sity, College of Medicine, Abha, Saudi Arabia.
2Tumor Immunology Unit, King Faisal Specialists Hospital & Re- search Center, Riyadh, Saudi Arabia.
3To whom correspondence should be addressed at Tumor Immunology Unit (MBC 03), King Faisal Specialists Hospital & Research Center, P.O. Box 3354, Riyadh 11211, Saudi Arabia.
that IL-10 is also produced by various other cells, including normal and malignant B cells (2), monocytes (3, 4), and mast cells (5) and that IL-10 activities are pleiotrophic (6). IL-10 has multiple biologic effects in the immune system and has been associated with suscepti- bility to certain microbial infections (7, 8). IL-10's inhibition of macrophage activation (9, 10) and T cell cytokine synthesis (IFN-3, and IL-2) suggests its use as an antigen nonspecific suppressor factor in certain T cell-mediated diseases. The known functions of IL-10 as a cytokine synthesis-inhibiting factor in various cell types (for review see Refs. 9 and 10) may indicate possible clinical applications in immune disorders.
Systemic lupus erythematosus (SLE) and rheumatoid arthritis (RA) are two well-known autoimmune diseases of unknown etiology (11, 12). B cell hyperactivity and autoantibody production are considered to be central to T cell abnormalities in the pathophysiological processes characteristic of SLE and RA. Moreover, cytokines determine whether T cells develop the capacity to pro- vide effective help for B cells to make antibody or to induce strong cell-mediated immunity. In view of the recognized functions of IL-10 in the immune system, and manifestations of the CD4+ Th2 cell response (9, 10), studies were undertaken to determine the secretion of IL-10 and to evaluate whether IL-10 was associated with B cell hyperactivity in antibody synthesis and hypergam- maglobulinemia in SLE and RA.
PATIENTS AND METHODS
198
Patients
All patients and controls were Saudi nationals. All patients were selected from those admitted to either
0271-9142/96/0700-0198509.50/0 �9 1996 Plenum Publishing Corporation

IL-10 SECRETION IN SLE AND RA 199
Rheumatology Ward, Assir Central Hospital, Abha, King Khalid University Hospital, Riyadh, or King Faisal Specialists Hospital & Research Center, Riyadh. Patients (26 women, 4 men; ages between 14 and 31 years), with active SLE, who fulfilled at least five of the American College of Rheumatology (ACR) criteria for SLE (13) were selected for these studies. All of these active SLE patients had anti-nuclear antibody (ANA) and anti- nDNA titers >-1:640, as determined by indirect immu- nofluorescence technique (Electronucleonics, Inc., Be- thesda, MD), as well as low serum levels of C3 (trace, 12 mg]dl) and C4 (trace, 10 mg/dl). Patients with RA (19 women, 11 men; ages between 31 and 52 years) fulfilled the ACR criteria for RA (14) and all of them were seropositive. Synovial fluid (SF) samples were obtained from the knee of these 21 patients with RA. Patients with Sjogren syndrome (SS) (6 women, 9 men; ages between 27 and 43 years) were diagnosed based on the presence of keratoconjunctivities sicca, xerostomia, a positive salivery gland biopsy showing mononuclear cell infil- trates, and the absence of an associated connective tissue disease. Patients with Behcet's disease (BD) (5 women, 10 men; ages between 23 and 36 years) had active signs and symptoms (painful oral ulcers, painful genital ulcers, some skin lesions such as erythema nodosum, folliculi- tis) and iritis (5 of them). All patients' serum and blood samples (10 cases each from active SLE and RA) were studied at diagnosis and before the start of any therapy. Patients with inactive SLE (13 women, 2 men; ages between 18 and 33 years) were those who were consid- ered in clinical remission and were taking low doses of prednisolone (5-10 mg/day). They had none of the clinical features of active SLE disease but had ANA and anti-nDNA titers of 1:20 to 1:80 and almost-normal levels of C3 (50-120 mg/dl) and C4 (20-72 mg/dl). Healthy controls (CO) were either laboratory workers or teaching staff members of the King Sand University, College of Medicine, Abha. All individuals tested were negative for human immunodeficiency virus.
Cell Preparation
Peripheral blood mononuclear cells (PBMC) were separated by the Histopaque-1077 (Sigma Chemical Co., St. Louis, MO) density-gradient centrifugation method described earlier (15).
Monocytes (adherent cells) were prepared by plastic adherence as follows: PBMC were layered onto plastic petri dishes (100 • 20 ram; Falcon Labware, Oxnard, CA) and incubated at 37~ for 60 min in culture medium [RPMI-1640 supplemented with L-glutamine (2 mM), penicillin (100 U/ml), streptomycin (100 /xg/ml), 5 X
10 5 ME, and 5% FCS (Flow Laboratories, Irvine, Scotland]. Nonadherent cells were removed by gently washing with warm medium. Adherent cells were then removed in cold medium. Monocytes were centrifuged at 1200g for 10 min at 4~ and were counted using ethidium bromide-acridine orange staining. The mono- cyte population consisted of 89-95% esterase-positive and OKMI/OKM5 (Ortho-Diagnostics, Raritan, NJ), mAb-positive cells.
PBMC were separated into T cells and non-T cells by the 2-aminoethylisothiouronium bromide-treated sheep erythrocyte (SRBC) rosetting technique described earlier (15, 16). Highly purified B cells were obtained from the unrosetted non-T cell fractions as follows: B cells were obtained by further depletion of T cells remaining in the non-T cell fractions by complement-mediated cell lysis with OKT3 mAb. This was followed by depletion of monocytes by removal of cells adhering (adherent) to plastic petri dishes and by complement-mediated cell lysis with OKMI/OKM5 mAb. The resultant B cell populations were 92-95% OKB2 (CD24+) and OKB7 (CD21+) mAb positive.
Twice-purified SRBC rosetting cells obtained from PBMC as described above were referred to as T cells. They were 91-96% CD3+ T cells (OKT3 mAb reac- tive). The CD3+ total T cells were separated into CD4+ and CD8 + T cell subsets by complement-mediated cell lysis with OKT8 and OKT4 mAbs, respectively (15, 16). The OKT4 mAb-treated cells contained 93-95% OKT8 mAb-reactive cells, thereafter called CD8+ T cells. The OKT8 mAb-treated cells yielded 92-96% OKT4 mAb- reactive ceils, thereafter called as CD4+ T cells.
The CD4+ T cells were further separated into C D 4 + C D 4 5 R O + (UCHLI mAb-react ive) and CD4+CD45RA+ (2H4 mAb-reactive) subpopulations by positive and negative selections using goat anti- mouse IgG (Ortho Diagnostics)-coated plastic plates (Fisher, Pittsburgh, PA) as described earlier (15). In brief, highly purified CD4+ T cells were reacted with saturating concentrations of either anti-2H4 or UCHLI mAbs. After washing to remove excess mAb, the cells were incubated on goat anti-mouse IgG (Genzyme, Boston, MA)-coated panning dishes for 60 rain at 4~ The adherent and nonadherent cell populations were separately collected and stained with 2H4 and 4B4/ UCHLI mAbs. This method yielded a purified CD4+CD45RO+ (91-96% UCHLI mAb-reactive) and CD4+CD45RA+ (89-94% 2H4 mAb-reactive) T-cell subpopulations. All mAbs used in these studies were obtained from Ortho Diagnostics (Raritan, NJ). The UCHLI mAb was a kind gift of Dr. Peter C. L. Beverley (University College Hospital, London, UK). A standard
Journal of Clinical Immunology, Vol. 16, No. 4, 1996

200 AL-JANADIETAL.
indirect immunofluorescence technique as described ear- lier was used to labels cells with a series of mAbs (16).
IL-IO Release
Purified mouocytes, B cells, and various T cell sub- populations were cultured in medium at a concentration of 2.5 X 105 cells/ml for a total period of 72 hr at 37~ and 5% CO 2 in 4-ml polystyrene tubes (Falcon, Lincoln Park, N J). The stimulants for monocytes and B cells was LPS (5/xg/ml Escherichia coli 0111 lipopolysaccharide; Difco, Detroit, MI), whereas T cells were stimulated with anti-CD3 (2/xg/ml OKT3 mAb). The supernatants were collected, centrifuged, aliquoted, and frozen at - 4 5 or -70~
In Vitro Ig Synthesis
Purified B cells (2.5 • 105 cells/ml) were cultured in complete RPMI-1640 medium with equal numbers of purified CD4+CD45RO+ T cell subsets with pokeweed mitogen (PWM; 10 /xg/ml) and with or without anti- IL-6, anti-IFN-7 (Genzyme, Boston, MA), or anti-IL-10 antibodies (PharMingen, San Diego, CA), each 10 /xg/ ml, in round-bottom microtiter plates (Costar, Cam- bridge, MA), for 7 days at 37~ and 5% CO 2. The cultured supernatants obtained were analyzed for IgM and IgG production by ELISA as described previously (16).
Detection of lL-lO by ELISA
IL-10 was determined using IL-10-specific mAbs in microtiter plates. In brief, 96-well microtiter plates (No. 25806 Coming, New York) were coated overnight at 4~ with 2/xg/well (100/xl) of anti-human IL-10 mAb (No. 18551D, PharMingen). Wells were next blocked with i% bovine serm albumin (BSA)/phosphate-buffered sa- line (PBS)-0.1% Tween, overnight at 4~ After five washes with PBS- 0.1% Tween, standard (human IL- 10) and serum samples were added to wells and allowed to incubate overnight at 4~ Plates were washed five times with PBS-0.1% Tween and secondary detection mAb, biotinylated anti-IL-10 (No. 18562D, PharMingen), 1 ~g]ml (100 ]xl) in 10% BSA/PBS, was added to each well. After 60 rain of incubation at room temperature, plates were washed five times (PBS-0.1% Tween) and 100 /xl of peroxidase-conjugated streptavidin, diluted 1:1000 in 10% BSA/PBS, was added to each well and allowed to incubate for an additional 60 rain at room temperature. Plates were washed five times (PBS-0.1% Tween), after which bound secondary mAb was allowed
to react with substrate (100 /M/well, 3,3', 5,5 '- tetramethylbenzadine; Sigma) for 10 min. Reaction was stopped by adding 2 M H2SO 4 (50/xl/well) and optical density was determined in a Flow Titertek microplate reader at 450 nm. The sensitivity of IL-10 in this ELISA was 40 pg/ml.
Statistical Analysis
Statistical significance was analyzed by Student's t test. P < 0.05 was considered statistically significant.
RESULTS
Serum IL-IO Levels in SLE and RA
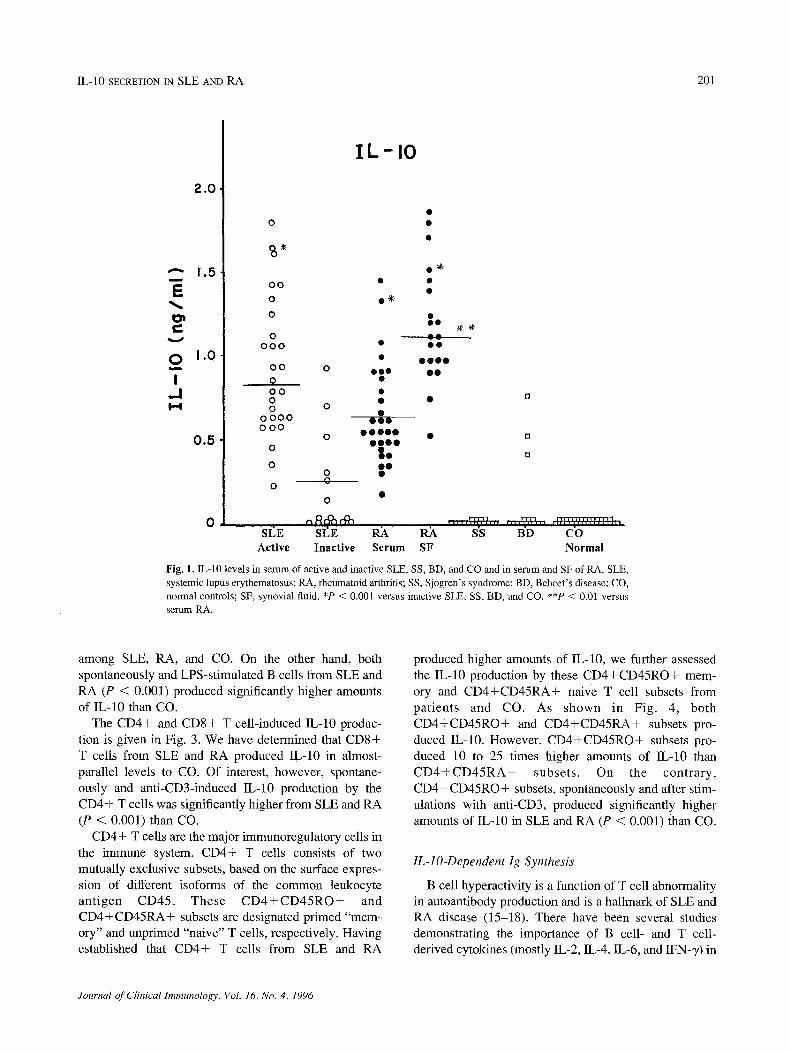
Serum IL-10 levels were measured in 30 patients with active SLE, 15 patients with inactive SLE, 30 patients with RA, 15 patients each with SS and BD, and 30 nornaal controls (CO). In addition, IL-10 was measured from SF samples of the 21 RA patients (Fig. 1). IL-10 was detectable in serum of all 30 active SLE, all 30 RA, 6 of the 15 inactive SLE, and 3 of the 15 BD patients, but none of the 15 SS and 30 CO. In RA, IL-10 levels were significantly higher in SF (P < 0.01) than serum. IL-10 levels were significantly elevated in active SLE and RA (serum and SF) (P < 0.001) compared to inactive SLE, SS, BD, and CO.
Serum IL-10 levels in active SLE were not correlated with any of the associated clinical signs such as nephritis, thrombocytopenia, and central nervous system disease or with complement C3 and C4 and anti-nDNA tilers (data not shown). Since ESR and serum CRP are frequently used to assess disease activity in RA, associations of serum and SF IL-10 with these variables were also evaluated in all 30 RA patients. However, no correlation was observed between IL-10 and any of these parameters in RA.
Source of lL-lO in SLE and RA
Increased concentrations of IL-10 in sera of all 30 active SLE and 30 RA patients prompted us to determine the source of IL-10 secretion in circulation of these patients. Therefore, we have determined the in vitro production of IL-10 by purified monocyte, B cell, and T cell populations from the circulation of 10 patients each with active SLE and RA and 10 CO (Figs. 2-4).
The data shown in Fig. 2 demonstrate that monocytes stimulated with LPS do produce some IL-10. But there was no major difference in IL-10 secretion by monocytes
Journal of Clinical Immunology, Vol. 16, No. 4, 1996
IL-10 SECRETION IN SLE AND RA 201
m
0 m
I ,..I H
2,0
1.5
1.0
0.5
IL-IO
0
S ~
O0
0
0
0 0 0 0
O 0 0
0
O 0 0 0 0
0 0 0 0 0 0 0
0
O 0
0
0
SI~E SEE Active Inactive
O ~
0 0 0
0 0 0 0 0 0 0 0
0 0 0 0
h $�9
O ~
O
O 0
O 0
0 0 0 0 O0
0 ~ ~ s's BD CO Serum SF Normal
Fig. 1. IL-10 levels in serum of active and inactive SLE, SS, BD, and CO and in serum and SF of RA. SLE, systemic lupus erythematosus; RA, rheumatoid arthritis; SS, Sjogren's syndrome; BD, Behcet's disease; CO, normal controls; SF, synovial fluid. *P < 0.001 versus inactive SLE, SS, BD, and CO. **P < 0.01 versus serum RA.
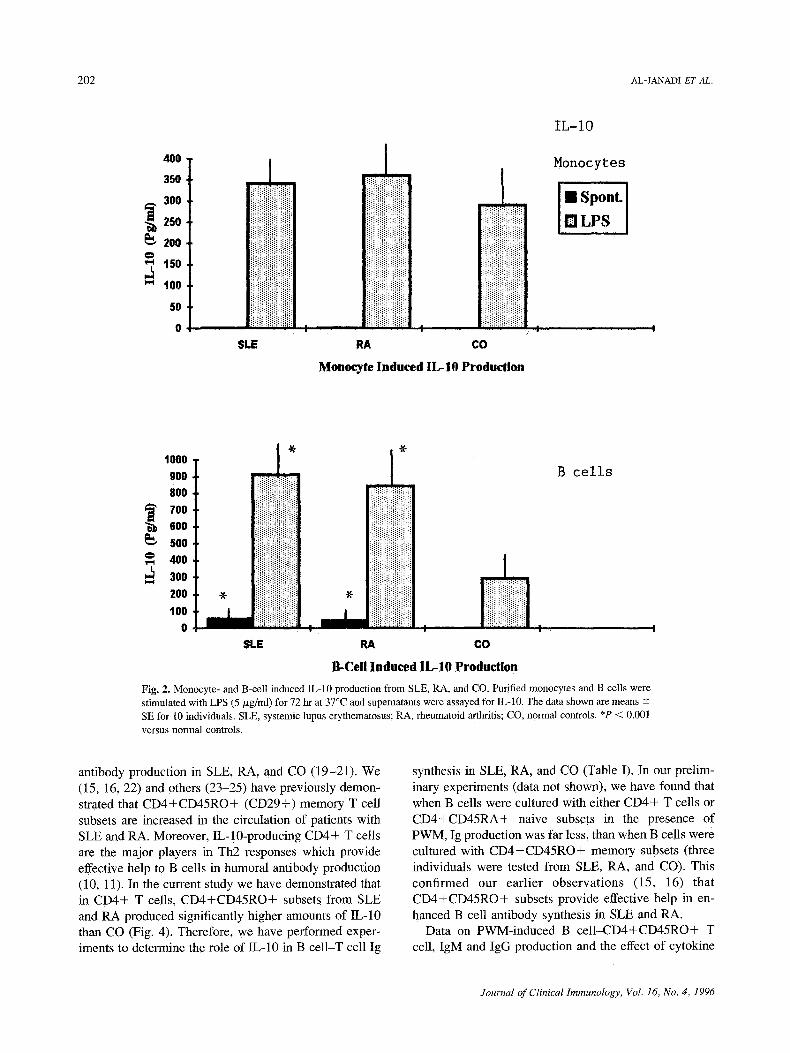
among SLE, RA, and CO. On the other hand, both spontaneously and LPS-stimulated B cells from SLE and RA (P < 0.001) produced significantly higher amounts of IL-10 than CO.
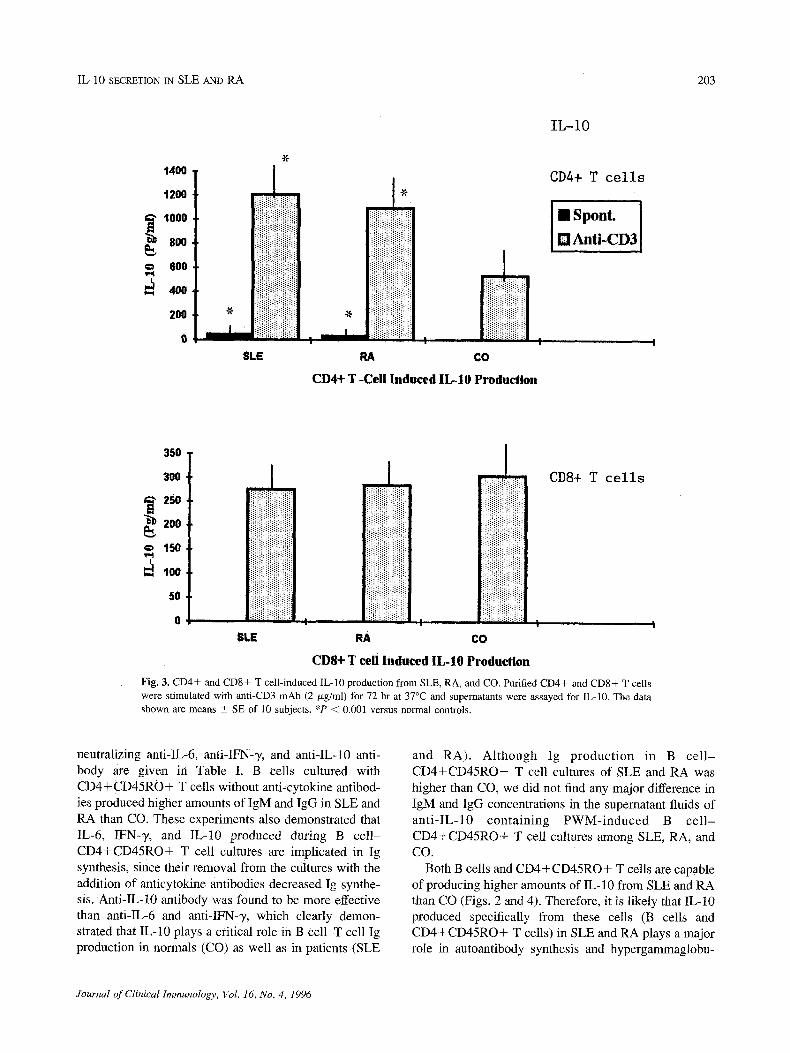
The CD4+ and CD8+ T cell-induced IL-10 produc- tion is given in Fig. 3. We have determined that CD8 + T cells from SLE and RA produced IL-10 in almost- parallel levels to CO. Of interest, however, spontane- ously and anti-CD3-induced IL-10 production by the CD4+ T cells was significantly higher from SLE and RA (P < 0.001) than CO.
CD4+ T cells are the major immunoregulatory cells in the immune system. CD4+ T cells consists of two mutually exclusive subsets, based on the surface expres- sion of different isoforms of the common leukocyte antigen CD45. These C D 4 + C D 4 5 R O + and CD4+CD45RA+ subsets are designated primed "mem- ory" and unprimed "naive" T cells, respectively. Having established that CD4+ T cells from SLE and RA
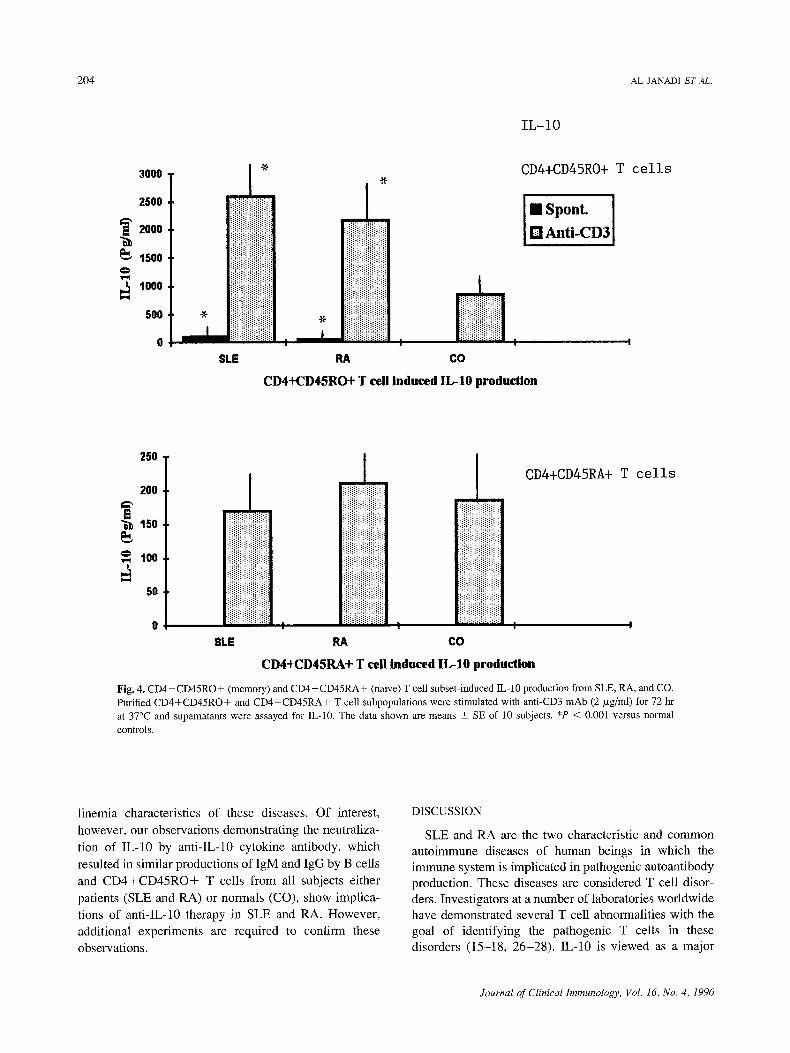
produced higher amounts of IL-10, we further assessed the IL-10 production by these CD4+CD45RO+ mem- ory and CD4+CD45RA+ naive T cell subsets from patients and CO. As shown in Fig. 4, both CD4+CD45RO+ and CD4+CD45RA+ subsets pro- duced IL-10. However, CD4+CD45RO+ subsets pro- duced 10 to 25 times higher amounts of IL-10 than C D 4 + C D 4 5 R A + subsets. On the contrary, CD4+CD45RO+ subsets, spontaneously and after stim- ulations with anti-CD3, produced significantly higher amounts of IL-10 in SLE and RA (P < 0.001) than CO.
IL-lO-Dependent Ig Synthesis
B cell hyperactivity is a function of T cell abnormality in autoantibody production and is a hallmark of SLE and RA disease (15-18). There have been several studies demonstrating the importance of B cell- and T cell- derived cytokines (mostly IL-2, IL-4, IL-6, and IFN-T) in
Journal of Clinical Immunology, Vol. 16, No. 4, 1996
202 AL-JANADI ET AL.
4OO
35O
300
-~ 250
2oo
" ~ t50
100
5O
0
i
iiiiiiiii iii iiiiiiii iiiiiil i
ii!ili ' iii! , liiiiiii!i[iiiiii ; ! i i iiiiiiiiiiii SLE RA CO
Monocyte Induced IL-IO Production
IL-IO
Monocytes
I Spont.
[] LPS
1000 900 800 700 600 5OO 400 3oo I,,,,,4
2O0 100
, ~/~
i!iiiiiii~iii!i~i~i iiii!iiii!!i!iiiiiii ii!iiii!;iiiii!ii~!
:iiiiiiiiiiililiiiii . . iiiiiiiiiii!iiiiiiii
i I ;iiiiiii!iiiiiFiil I , !
I~ c o
B-Cell Induced IL-IO Production
SLE I �9
B cells
Fig. 2. Monocyte- and B-cell induced IL-10 production from SLE, RA, and CO. Purified monocytes and B cells were stimulated with LPS (5 p~g/ml) for 72 hr at 37~ and supematants were assayed for IL-10. The data shown are means -+ SE for 10 individuals. SLE, systemic lupus erythematosus; RA, rheumatoid arthritis; CO normal controls. *P < 0.001 versus normal controls.
antibody production in SLE, RA, and CO (19-21). We (15, 16, 22) and others (23-25) have previously demon- strated that CD4+CD45RO+ (CD29+) memory T cell subsets are increased in the circulation of patients with SLE and RA. Moreover, IL-!0-producing CD4+ T cells are the major players in Th2 responses which provide effective help to B cells in humoral antibody production (10, 11). In the current study we have demonstrated that in CD4+ T cells, CD4+CD45RO+ subsets from SLE and RA produced significantly higher amounts of IL-10 than CO (Fig. 4). Therefore, we have performed exper- iments to determine the role of IL-10 in B cell-T cell Ig
synthesis in SLE, RA, and CO (Table I). In our prelim- inary experiments (data not shown), we have found that when B cells were cultured with either CD4+ T cells or CD4+CD45RA+ naive subsets in the presence of PWM, Ig production was far less, than when B cells were cultured with CD4+CD45RO+ memory subsets (three individuals were tested from SLE, RA, and CO). This confirmed our earlier observations (15, 16) that CD4+CD45RO+ subsets provide effective help in en- hanced B cell antibody synthesis in SLE and RA.
Data on PWM-induced B cell-CD4+CD45RO+ T cell, IgM and IgG production and the effect of cytokine
Journal of Clinical Immunology, Vol. 16, No. 4, 1996
IL-10 SECRETION IN SLE AND RA 203
1400
1200
.• 1000
800 600
200 ~F
I
$1-E
iil;i;i!i!ii! . i!iii!!i!ili!iiiill , iiiiiiiiiiiii!i~iii
RA CO
CD4+ T -Cell Induced [L-IO Production
IL-IO
CD4+ T cells
l~Anti-CD31
4~
4
350
300
250
2O0
150
IO0
50
0
i
i;ii!!iiii!iiii CD8+ T cells
D Ir i lii!~ii!!i~ii~!!i~i: |
Iiii!iiiiiiiii!iiiiil ", [ii!1! ! :: :::::: i ,
I
iii!iiiii!iiii!i!ii
$LE RA CO
CDS+ T cell Induced IL-10 Production
]Fig. 3. CD4+ and CD8+ T cell-induced IL-10 production from SLE, RA, and CO. Purified CD4+ and CD8+ T ceils were stimulated with anti-CD3 mAb (2/xg/ml) for 72 hr at 37~ and supernatants were assayed for IL-10. The data shown are means • SE of 10 subjects. *P < 0.001 versus normal Controls.
neutralizing anti-IL-6, anti-IFN-7, and anti-IL-10 anti- body are given in Table I. B cells cultured with CD4+CD45RO+ T cells without anti-cytokine antibod- ies produced higher amounts of IgM and IgG in SLE and RA than CO. These experiments also demonstrated that IL-6, IFN-7, and IL-10 produced during B cell- CD4+CD45RO+ T cell cultures are implicated in Ig synthesis, since their removal from the cultures with the addition of anticytokine antibodies decreased Ig synthe- sis. Anti-IL-10 antibody was found to be more effective than anti-IL-6 and anti-IFN-% which clearly demon- strated that IL-10 plays a critical role in B cell-T cell ig production in normals (CO) as well as in patients (SLE
and RA). Although Ig production in B ce l l - CD4+CD45RO+ T ceil cultures of SLE and RA was higher than CO, we did not find any major difference in IgM and IgG concentrations in the supernatant fluids of anti-IL-10 containing PWM-induced B ce l l - CD4+CD45RO+ T cell cultures among SLE, RA, and CO.
Both B cells and CD4+CD45RO+ T cells are capable of producing higher amounts of IL-10 from SLE and RA than CO (Figs. 2 and 4). Therefore, it is likely that IL-10 produced specifically from these cells (B cells and CD4+CD45RO+ T cells) in SLE and RA plays a major role in autoantibody synthesis and hypergammaglobu-
Journal of Clinical Immunology, Vol. 16, No. 4, 1996
204 AL-JANADI ET AL.
IL-IO
3000
2500
~,~2000
t5oo
4000
500 i
~.
!iii iiiiilUiiii!iill
SLE
n 1 I ..... ;q';'"";'; I I
RA CO
CD4+CD45RO+ T cell Induced IL-10 production
CD4+CD45RO+ T cells
[] Spont. [ M Anti-CD31
250
200
,.~ 100
50
' L I CD4+CD45RA+ T cells •
i:ifiJ:~:~:ifi:~:D ii!iiiiiiiii!iiiiii!
I I I I
SLE RA CO
CD4+CD45RA+ T cell induced IL-10 production
Fig. 4, CD4+CD45RO+ (memory) and CD4+CD45RA+ (naive) T cell subset-induced IL-10 production from SLE, RA, and CO. Purified CD4+CD45RO+ and CD4+CD45RA+ T cell subpopulations were stimulated with anti-CD3 mAb (2/xg/ml) for 72 hr at 37~ and supernatants were assayed for IL-10. The data shown are means _+ SE of l0 subjects. *P < 0.001 versus normal controls.
linemia characteristics of these diseases. Of interest, however, our observations demonstrating the neutraliza- tion of IL-10 by anti-IL-10 cytokine antibody, which resulted in similar productions of IgM and IgG by B cells and CD4+CD45RO+ T cells from all subjects either patients (SLE and RA) or normals (CO), show implica- tions of anti-IL-10 therapy in SLE and RA. However, additional experiments are required to confirm these observations.
DISCUSSION
SLE and RA are the two characteristic and common autoimmune diseases of human beings in which the immune system is implicated in pathogenic autoantibody production. These diseases are considered T cell disor- ders. Investigators at a number of laboratories worldwide have demonstrated several T cell abnormalities with the goal of identifying the pathogenic T cells in these disorders (15-18, 26-28). IL-10 is viewed as a major
Journal of Clinical Immunology, Vol. 16, No. 4, 1996

IL-10 SECRETION IN SLE AND RA 205
Table I. IL-10-Dependent Ig Production by B Cells-CD4+CD45RO+ T Cells a
Expt Cells Additions IgM IgG
1. SLE Nil Nil 0.2 -- 0.1 0.2 + 0.1 CD4+CD45RO+ Nil 14.8 -- 0.9 5.7 + 0.5 CD4+CD45RO+ Anti-IL-6 9.5 -+ 0.6 3.1 _+ 0.4 CD4+CD45RO+ Anti-IFN-y 10.8 +_ 0.8 4.3 _+ 0.4 CD4+CD45RO+ Anti-IL-10 1.7 _+ 0.2 0.6 + 0.1
2. RA Nil Nil 0.2 + 0.1 0 CD4+CD45RO+ Nil 10.6 _+ 0.7 3.8 _+ 0.4 CD4+CD45RO+ Anti-IL-6 8.1 _+ 0.5 2.9 _+ 0.4 CD4+CD45RO+ Anti-IFN-'y 8.8 -+ 0.5 3.6 _+ 0.5 CD4+CD45RO+ Anti-IL-10 1.5 + 0.3 0.8 _+ 0.1
3. CO (normal) Nil Nil 0.2 + 0.1 0 CD4+CD45RO+ Nil 7.9 + 0.4 3.2 _+ 0.4 CD4+CD45RO+ Anti-IL-6 5.8 -+ 0.4 2.5 +_ 0.3 CD4+CD45RO+ Anti-IFN-,y 6.3 -+ 0.5 1.9 +_ 0.2 CD4+CD45RO+ Anti-IL-10 1.5 _+ 0.2 0.7 +_ 0.1
a B cells (2.5 • 105 cells/ml) were cultured alone or with equal numbers of CD4+CD45RO+ T cells in the presence of PWM, and with or without anti-IL-6, anti-lFN-y, or anti-IL-10 for 7 days, and supernatants were assayed for IgM and IgG (p~g/ml). The data represent the mean _+ SE in the triplicate cultures. Similar results were obtained in additional experiments using B cells and CD4+CD45RO+ T cells from four patients each with SLE and RA and four CO.
immunoregulatory T cell Th2 response cytokine with a spectrum of pleiotrophic effects on various cells in the immune system (9, 10). In this report, we present the first documentation of the secretion of 1L-10 in serum of patients with active SLE and RA. Although IL-10 has been detected in serum samples of various other diseases including hematologic disorders (29-31) , normal serum samples do not contain IL-10. There is also scanty information available about the cytokine profile of monocytes and B cell and various T cell subsets in autoimmune diseases. Our study established that purified B cells and C D 4 + C D 4 5 R O + "memory" T cell sub- populations from blood produced higher amounts of IL-10 in SLE and RA relative to normal controls. Cytokines determine whether T cells develop the capac- ity to provide effective help for B cells to make antibody or to induce strong cel l-mediated immunity. Of interest, we demonstrate here that enhanced Ig production by B cells and C D 4 + C D 4 5 R O + T cells is a function of IL-10 cytokine in SLE and RA, since removal of IL-10 by anti-IL-10 antibody resulted in abrogation of this func- tion with a reduced production of Igs.
We demonstrate here that IL-10 levels were signifi- cantly higher in SF than serum samples of RA. A recent study (32) has also shown that SF of RA contains higher levels of IL-10. The synovium of RA patients is heavily infiltrated with T cells expressing some activation mark- ers (15). Of the various immunoregulatory functions of IL-10, prevention of the induction of apoptosis (9, 10) and differential expression of activation markers on T cells could explain some phenomena of chronic inflam- mation in RA. However, we could not study the cellular source or function of IL-10 from RA SF, because of the
small sample size. Al l our studies on secretion of IL-10 by monocytes, B cells, and T cells and IL-10 function in
B c e l l - C D 4 + C D 4 5 R O + T cell-induced Ig secretion were based on the cells isolated from blood. However, the increased secretion of IL-10 in SF of RA patients and the function of IL-10 in enhanced Ig synthesis by blood purified B and T cells may implicate this cytokine in the pathogenic autoantibody production in RA disease.
IL-10 is associated with the inhibition of C D 4 + Thl - type cytokines such as IFN-y and IL-2 (9, 10). The majority of direct effects of IL-10 on T cells described
has been inhibitory, in that IL-10 has been shown to inhibit T cell proliferation by reducing IL-2 production. In contrast to these reports, IL-10 has been shown to rescue human T cells from apoptotic cell death (33). In addition, IL-10 modulates activation marker expression, CD25 (IL-2 receptor), and HLA-DR on B cells and T cells (34) and is a cofactor for the proliferation of human peripheral T cells (35). IL-10 can have a multitude of effects on human T cells and should not be considered exclusively as a immunoinhibitory cytokine. High serum levels of IL-10 were found in various malignant diseases and correlates with poor survival in patients with inter- mediate and high-grade non-Hodgkin 's lymphomas (29, 30). High serum levels of IL-10 were related with malignant B cell proliferation and antibody synthesis in multiple myeloma (10). Therefore, IL- 10 is also a growth and differentiation factor for B cells. Enhanced secretion of IL-10 demonstrated in the serum of all active SLE, and serum and SF of RA, and IL-10-dependent aug- mented production of B ce l l -T cell Ig synthesis in these inflammatory disorders suggest that IL-10 also can act as an inflammatory cytokine.
Journal of Clinical Immunology, Vol. 16, No. 4, 1996

206 AL-JANADI ET AL.
B cells and CD4+CD45RO+ memory T cell subsets are the leading cells in the circulation of SLE and RA that were shown to be implicated in pathogenic autoan- tibody synthesis (15-19). These important components of the humoral immune system have been demonstrated to produce higher amounts of IL-10 both spontaneously and in response to stimulations with appropriate agents in these diseases. Previously, Emilie et al. (30) reported that in vitro stimulations of PBMC from AIDS patients also produced increased amounts of IL-10. The autocrine production of IL-10 may be a specific feature of B and T cells in certain autoimmune diseases and B cell malig- nancies with features of hypergammaglobulinemia. The spontaneously or antigen-induced IL-10 production by SLE and RA lymphocytes during B cell activation responses may amplify the differentiation of autoreactive B cells into autoantibody-secreting cells in an environ- ment where IL-2 and IFN-y is low. Indeed, studies of in
vitro antibody synthesis utilizing anti-lL-10 antibody indicated a clear role of IL-10 in B cell differentiations in patients and controls. An up-regulation of IL-10 produc- tion may play a role in B cell hyperactivation in SLE and RA. Therefore, a role of IL-10 in pathogenesis of these disorders cannot be excluded. Because all SLE and RA patients in the present study were believed to have very active disease, we were unable to determine whether a correlation does exist between IL-10 production and disease activity. Further studies with the aim of therapies directed at neutralizing the effects of IL-10 will hopefully lend additional insights into the role of this cytokine in disease mechanism. Antagonism of IL-10 may also be a desirable choice of therapy in SLE and RA. However, autocrine production should be taken into account when considering possible therapeutic approaches, as autocrine loops may be less sensitive than paracrine loops to the inhibitory effects of mAb directed toward cytokines.
In SLE, B cell hyperactivity is evident by almost all measures, but the extent of its dependence of Th (Thl/ Th2) cell activation and cytokines is unknown. Our studies provide some evidence that B cell hyperactivity in antibody product ion could depend on CD4+CD45RO+ Th2 cell activation (IL-10) parame- ters. In vitro, IL-10 augments proliferation of activated B cells and induces their differentiation into antibody- producing cells, with subsequent expression of large amounts of IgM and IgG antibodies. However, there is no conclusive evidence yet available to suggest that anti-IL-10 treatment in vivo also decreases Ig secretion by B cells-T cells in humans. A major point is to ascertain whether these in vitro properties accurately reflect the physiological functions of IL-10 in vivo.
CD5 + B cells are present in increased proportions in circulation of patients with SLE and RA, and they have been implicated in autoantibody synthesis (36, 37). Recent findings that CD5 + B cells are potent producers of IL-10 (38) indeed raises the possibility of a broader immunoregulatory role of IL-10 in autoantibody produc- tion. To date Ishida et al. (39) have conducted a series of extensive experiments to demonstrate the IL-10 function in vivo in mice. Anti-IL-10-treated mice from birth to 8 weeks of age appear to be essentially healthy and show no alteration of numerous parameters, such as total body weight, gross histology of liver, lungs, intestines, spleens, and thymocytes, hematocrit, and proportions of B cells, T cells, and monocytes (39). Furthermore, anti-IL-10-treated mice contained substantial levels of circulating IFN-y and frequently expressed IL-6 in their sera, two cytokines which were not found in the circu- lation of normal mice. In terms of serum Ig levels, anti-IL-10-treated mice were found to be drastically depleted of circulating IgM and IgA antibodies (39). The reduction of serum Igs can be explained by the selective elimination of a subpopulation of B cells designated CD5+ B cells from the anti-IL-10-treated mice. How- ever, more recent experiments by Ishida et al. (40) in lupus-prone NZB/W F 1 mice demonstrated that con- tinous treatment with anti-IL-10 antibodies substantially delays onset of autoimmunity. These data may suggest that IL-10 antagonists may be beneficial in the therapy of human SLE and RA. Based on these in vivo observations in mice, and on our own demonstrations of enhanced in
vitro secretion of IL-10 by SLE and RA B cells, it would be very interesting to evaluate the IL-10-dependent CD5+ B cell function in pathogenic autoantibody pro- duction. We hope that our future experimental approach will provide answers to some of these questions on the immunoregulatory role of IL-10 in autoimmune disease.
REFERENCES
1. Fiorentino DF, Bond MW, Mosmann TR: Two types of mouse helper T cell IV. Th2 clones secrete a factor that inhibits cytokine production by Thl clones. J Exp Med 170:2081-2089, 1989
2. O'Gara A, Stapleton G, Dhar V, Pearce M, Schumacher J, Rugo H, Barbis D, Stall A, Cupp J, Moore K, Vieira P, Mosmann T, Whitmore A, Arnold L, Haugton G, Howard M: Production of cytokines by mouse B cells: B lymphomas and normal B cells produce interleukin 10. Int Immunol 2:821-832, 1990
3. de Waal MR, Abrams J, Bennett B, Figdor C, de Vries JE: IL-10 inhibits cytokine synthesis by human monocytes. An autoregula- tory role of IL-10 produced by monocytes. J Exp Med 174:1209- 1220, 1991
4. Fiorentino DF, Zlotnik A, Mossmann TR, Howard M, O'Gara A: IL-10 inhibits cytokine production by activated macrophages. J Immunol 147:3815-3822, 1991
Journal of Clinical Immunology, Vol. 16, No. 4, 1996

IL-10 SECRETION IN SLE AND RA 207
5. Thompson-Snipes L, Dhar V, Bond MW, Mosmann TR, Moore KW, Rennick D: Interleukin-10: A novel stimulatory factor for mast cells and their progenitors. J Exp Med 173:507-510, 1991
6. Zlotnik A, Moore KW: Interleukin-10. Cytokines 3:366-371, 1991 7. Reed SG, Brownell CE, Russo DM, Silva JS, Grabstein KH,
Morrissey PJ. IL-10 mediates susceptibility to trypanosoma cruzi infection. J Immunol 153:3135-3140, 1994
8. Sher A, Coffman RL: Regulation of immunity to parasites by T cells and T cell derived cytokine. Annu Rev Immunol 10:385-399, 1992
9. Howard M, O'Garra A, Ishida H, de Waal MR, de Vries J: Biological properties of interleukin-10. J Clin Immunol 12:239- 247, 1992
10. Moore KW, O'Garra A, de Waal MR, Vieira P, Mosmann TR: Interleukin-10. Annu Rev Immunol 11:165-190, 1993
11. Hahn BH: An overview of the etiology and pathogenesis of systemic lupus erythematosus. In Systemic Lupus Erythematosus, 4th ed, Wallace, Hahn (eds). Philadelphia, Lea & Fibger, 1992, pp 65-70
12. Paget SA, Gibofsky A: Immunopathogenesis of rheumatoid arthri- tis. Am J Med 67:961-970, 1979
13. Tan EM, Cohen AS, Fries JF, Masi AT, McShane DJ, Rothfield NF, Schaller JG, Talal N, Winchester RJ: The 1982 revised criteria for the classification of systmic lupus erythematosus. Arth Rheum 25:1271-1277, 1982
14. Arnett FC, Edworthy SM, Bloch DA, McShane DJ, Fries JF, Cooper NS, Healey LA, Kaplan SR, Liang MH, Luthra HS, Medsger TA Jr, Mitchell DM, Neustadt DH, Pinals RS, Schaller JG, Sharp JT, Wilder RL. Hunder GG: The American Rheumatism Association 1987 revised criteria for the classification of rheuma- toid arthritis. Arth Rheum 31:315-324, 1988
15. A1-Janadi M, A1-Balla S, A1-Dalaan A, Raziuddin S: Cytokine production by helper T cell populations from the synovial fluid and blood in patients with rheumatoid arthritis. J Rheumatol 20:1647- 1653, 1993
16. A1-Janadi M, Raziuddin S: B cell hyperactivity is a function of T cell derived cytokines in systemic lupus erythematosus. J Rheu- matol 20:1885-1891, 1993
17. Klinman DM: B cells. In Steinberg AD, moderator. Systemic lupus erythematosus. Ann Intem Med 115:550-555, 1991
18. Ueda Y, Sakane T, Tsunematsu T: Hyperactivity of activated B cells to B cell growth factor in patints with systemic lupus erythematosus. J Immunol 143:3988-3993, 1989
19. Raziuddin S, A1-Janadi M, Al-Wabel AA: Soluble interleukin-2 receptor levels in serum and its relationship to T cell abnormality and clinical manifestation of the disease in patients with systemic lupus erythematosus. J Rheumatol 18:831-836, 1991
20. Linker-Israeli M, Deans RJ, Wallace DJ, Prehn J, Ozeri-Chen T, Klinberg JR: Elevated levels of endogeneous IL-6 in systemic lupus erythematosus. A putative role in pathogenesis. J Immunol 147:117-123, 1991
21. Hochberg MC: Systemic lupus erythematosus. Rheum Dis Clin North Am 16:613-639, 1990
22. Raziuddin S, Nur MA, A1-Wabel AA: Selective loss of the CD4+ inducers of T cell subsets (2H4) in active systemic lupus erythem- atosus. J Rheumatol 16:1315-1319, 1989
23. Morimoto C, Romain PL, Fox DA, Schlossman SF: Abnormalities in CD4+ T lymphocyte subsets in inflammatory rheumatic dis- eases. Am J Med 84:817-825, 1988
24. Emery P, Gentry KC, Makay IR, Muirden KD, Rowley M: Deficiency of the suppressor-inducer subsets of T lymphncytes in rheumatoid arthritis. Arth Rheum 30:849-856, 1987
25. Morimoto C, Steinberg AD, Letvin NL, Fox DA, Romain PL, Schlossman SF: A defect of immnnoregulatory T cell subsets in systemic lupus erythematosus patients dernonstrated by anti-2H4 antibody. J Clin Invest 79:762-768, 1987
26. Horwitz DA, Wang HA, Gray JD: Cytokine gene profile in circulating blood mononuclear cells from patients with systemic lupus erythematosus: Increased interleukin-2 but not interlenkin-4 mRNA. Lupus 3:423-428, 1994
27. Zagon G, Tumang JR, Li Y, Friedman ST, Crow MK: Increased frequency of Vbl7-positive T cells in patients with rheumatoid arthritis. Arth Rheum 37:1431-1440, 1994
28. Aringer M, Wintersberger W, Steiner CW, Kiener H, Presterl E, Smolen J, Graninger WB. High levels of bcl-2 protein in circulat- ing lymphocytes but not B lymphocytes of patients with systemic lupus erythematosus. Arth Rheum 37:1423-1430, 1994
29. Blay JY, Burdin N, Rousset F, Lenoir G, Biron P, Philip T, Bancherean J, Favrot MC: serum interleukin-10 in non-Hodgkin's lymphoma: A prognostic factor. Blood 82:2169-2174, 1993
30. Smilie D, Touitou R, Raphel M, Peuchmanr M, Devergnee O, Rea D, Galanaud P: In vivo production of interleukin-10 by malignant cells in AIDS lymphomas. Eur J Immunol 22:2937-2942
31. Baiocchi R, Ross ME, Tan JC, Chou CC, Sulivan L, Sklar J, Croce CM, Caliginri MA: Lymphomagenesis in the SCID-hu mouse involves abundent production of human interleukin-10. Blood 85:1063-1074, 1995
32. Katsikis PD, Chu CQ, Brenan FM, Mainin RN, Feldmann M. Immunoregulatory role in interleukin-10 in RA. J Exp Med 179:1517-1521, 1994
33. Taga K, Cheruay B, Tosato G: IL-10 inhibits apoptic cell death in humans T cells starved of IL-2. Int Immunol 5:1599-1604, 1993
34. Cohen SB, Katsikis PD, Feldmann M, Londei M: IL-10 enhance expression of the IL-2 receptor c~ chain on T cells. Immunology 83:329-332, 1994
35. MacNeil IA, Suda T, Moore KW, Mosmann TR, Zlotnik A. IL-10, a novel growth cofactor for mature and immature T ceils. J Im- mnnol 145:4167-4172, 1990
36. Via CS, Handwerger BS: T cell and B cell function in lupus. Curr Opinion Rhumatol 4:630-634, 1992
37. Murakami M, Tsubata T, Okamoto M, Shimizu A, Kumagai S, Imura H, Honjo T: Antigen-induced apoptic death of Ly 1 (CD5 +) B cells responsible for autoimmune disease in transgenic mice. Nature 357:77-80, 1992
38. O'Gara A, Chang R, Hastings R, Go N, Haughton G. howard M: LylB (B1) cells are the main source of B cell derived IL-10. Eur J Immunol 22:711-717, 1992
39. Ishida H, Hastings R, Kearuey J, Howard M: Continuous anti-IL-1 antibody administration depletes mice of Ly-1 B ceils but not conventional B cells. J Exp Med 175:1213-1220, 1992
40. Ishida H, Muchamuel T, Sakaguchi S, Andrade S, Menon S, Howard M: Continuous administration of anti-interleukin-10 anti- bodies delays onset of autoimmunity in NZB/W F1 mice. J Exp Med 179:305-310, 1994
Journal of Clinical Immunology, Vol. 16, No. 4, 1996





























