Embed Size (px)
Citation preview

Brain Research 984 (2003) 63–75www.elsevier.com/ locate/brainres
Research report
T opographically graded postischemic presence of metalloproteinases isinhibited by hypothermia
a , a a a c*Simone Wagner , Simon Nagel , Britta Kluge , Stefan Schwab , Sabine Heiland ,b d aJames Koziol , Humphrey Gardner , Werner Hacke
aDepartment of Neurology University of Heidelberg Medical School, INF 400, 69120Heidelberg, GermanybScripps Research Institute, La Jolla, CA, USA
cDepartment of Neuroradiology University of Heidelberg Medical School, Heidelberg, GermanydBiogen, Cambridge, MA, USA
Accepted 28 May 2003
Abstract
To test the hypothesis that presence of metalloproteases (MMPs), their inhibitors (TIMPs) and their substrate laminin-5 differs betweenthe ischemic core and the surrounding tissue, we examined the impact of middle cerebral artery occlusion/ reperfusion (MCA:O/R) onthese molecules in different regions of the infarct. We also investigated the influence of hypothermia on the progression of the ischemiclesion and MMP activity. Brain sections from 64 Wistar rats subjected to MCA:O/R were examined by means of cytohistochemistry andzymography. The artery was occluded for 2 h followed by 3, 5, 8 and 12 h of reperfusion. Well characterized antibodies against laminin-5,MMPs and TIMP-2 were used. A total of 32 rats were treated with hypothermia. The presence of each antigen was related to thefollowing regions of interest: ischemic core with BBB breakdown (I ), surrounding ischemic tissue without BBB breakdown (I ), and thec r
contralateral non-ischemic region (N). Regions of interest were defined by MRI. The I increased over time at the cost of the I . BBBc r
breakdown occurred early in the ischemic core and increased over time. Hypothermia reduced the BBB breakdown at all time points. A2graded decreased presence of laminin-5 was observed with 16.563.7(N).1062.8(I ).461.4(I ) immunopositive microvessels /mm atr c
23 h of reperfusion. MMP-9 showed a reverse pattern with 0 (N),460.8(I ),1061.5(I ) immunopositive microvessels /mm .r c
Hypothermia decreased the MMP activity measured by zymography. Laminin-5 and MMP presence relate directly to the degree ofpostischemic injury. Hypothermia reduces the conversion from the I to ischemic core and the degree of BBB as well as MMP abundance.r
2003 Elsevier B.V. All rights reserved.
Theme: Disorders of the nervous system
Topic: Ischemia
Keywords: Cerebral ischemia; MRI; Hypothermia; Metalloproteinase
1 . Introduction
Middle cerebral artery occlusion (MCA:O) in the ratinduces changes in cerebral microvessels, which lead to
Abbreviations: BG, basal ganglia; BBB, blood–brain barrier; DWI, decreased microvascular integrity, oedema formation,diffusion weighted imaging; ECM, extracellular matrix; FITC, fluorescein
perivascular haemorrhage, and luminal obstruction in theisothiocyanate; GFAP, glial fibrillary protein; MCA:O/R, middle cerebralregion of striatal neurone injury[6].artery occlusion/ reperfusion; MMP, matrix–metalloproteinases; MRI,
magnetic resonance imaging; N, nucleus; NICU, neurologic intensive care Adhesion complexes consisting of extracellular matrixunit; PWI, perfusion weighted imaging; ROI, regions of interest; TIMP, molecules (ECMs) and integrins play an important role intissue inhibitor of metalloproteinases; TRITC, tetramethylrhodamine-iso- cell adhesion and cell–cell signalling[17]. Decreasedthiocyanate
expression of these molecules has been described in both*Corresponding author. Tel.:149-622-156-7504; fax:149-622-156-in vitro and in vivo models of cerebral ischemia[29,28].5461.Metalloproteases (MMPs), which degrade extracellularE-mail address: simone [email protected](S. Wagner).
]
0006-8993/03/$ – see front matter 2003 Elsevier B.V. All rights reserved.doi:10.1016/S0006-8993(03)03088-9

64 S. Wagner et al. / Brain Research 984 (2003) 63–75
matrix molecules e.g. laminins, are increased in ex- were removed from the skull, the brain was cut into fiveperimental cerebral ischemia[15]. They have also been coronal section of 2.5 mm thickness named A (rostral) to Eimplicated in blood brain barrier (BBB) damage[24]. (caudal). The caudal surface of block B is at the level ofHypothermia increases survival in patients with severe the bregma and level of the anterior commissure. ByMCA or hemispheric infarctions[26]. Many possible cutting block B rostrally for immuohistochemistry andmechanisms for hypothermia induced protection have been Block C for zymography, we attempted to stay well in theproposed: a decrease in metabolic requirement for oxygen territory of the middle cerebral artery.and decrease in cerebral blood flow, alterations in neuro-transmitter release, and suppression of the neutrophil-me-diated inflammatory response[12,18]. Given the tempera- 2 .2. Immunohistochemistry and zymographyture sensitivity of enzymes, the action of MMPs could wellbe inhibited by this therapeutic approach. The hypotheses Well-characterised anti-rat monoclonal antibodiestested herein are: (1) Changes in microvascular integrity (MoAb) identified antigens on MMP-2 (42-5D11, 1:50)during focal cerebral ischemia are graded in relation to and MMP-9 (VII C2, 1:100) (Oncogen). TIMP-2 wasischemic core and the surrounding tissue defined by MRI. stained with (MoAb, 674HII, 1:100, Oncogen). A rabbit(2) One important pathophysiological mechanism of mod- anti-rat polyclonal antibody (0668B, kind gift of V.erate hypothermia is the inhibition of MMPs leading to Quaranta, Scripps Research Institute, USA) as well as areduced BBB breakdown and slowed conversion of sur- mouse monoclonal (MIG-1 kind gift of V. Quaranta,rounding tissue to ischemic core. Scripps Research Institute, USA) against laminin-5 was
used to identify this MMP substrate. The antibody 0668Bis a polyclonal rabbit anti-rat and has been tested on
2 . Material and methods Western blot for its reactivity with rat astrocytes andendothelial cells[28]. CD 31 was used as a baseline
The experimental protocols used in this study were marker for endothelial cells (MoAb, TLD3A12, 1:200,approved by the National Ethics Committee for Animal Chemicon), GFAP (1:200, Dako) for astrocytes and NeuNResearch, Karlsruhe, Germany and adequate measures (MoAb, 1:100, Chemicon) for neurones. Immunohistoch-were taken to minimize pain or discomfort. emical procedures were performed as previously described
[28,29]. The cellular source of antigen presence was2 .1. Experimental stroke model in the rat investigated with immunofluorescence colocalization
studies with the use of FITC and TRITC-labelled sec-Occlusion of the middle cerebral artery for 2 h with ondary antibodies (Vector and Sigma). CD 31 (endothelial
subsequent reperfusion for 3, 5, 8 and 12 h (MCA:O/R) marker), GFAP (astrocytic marker) and NeuN (neuronalwas induced in 48 adult male Wistar rats (275 g610%) marker) were taken to identify the cellular origin of antigenaccording to a modification of the method ofZea longa presence. The number of cells or vessels identified byand has been described in detail elsewhere[8]. immunoperoxidase reactivity to antigens of MMP-2,
Eight animals underwent sham-operation involving fila- MMP-9, TIMP-2 and laminin-5 were measured afterment removal within 1 min, and eight subjects comprised scanning using NIH image software and expressed as
2the control group. A total of 32 rats were treated with absolute number /mm . Laminin-5 and MMP-9 positiveexperimental hypothermia. If animals died before the vessels were also expressed as percent of total vessel countcompletion of the experiment, they were excluded from on H&E sections (seeTable 1). Regions of interest (ROIs)analysis, and were replaced to ensure that six surviving namely centre of the infarct with BBB breakdown (I ),c
animals were in each group. Overall, five animals died area of risk (I ), and non ischemic left side (N) werer
under normothermic conditions, three died under hypo- identified on the slide by overlaying an image of the MRI.thermic conditions, five had no perfusion deficit and nine In the non-ischemic control subjects, ROIs were chosenanimals had to be excluded because of technical problems from the right hemisphere. Vessels were counted in theof the MR-Scanner. relevant regions.
Mild hypothermia (33C8) was induced by external body Zymography was performed as previously describedcooling with cool-packs after 1 h of occlusion and main- [22]. We used 2.5 mm of block C of each brain fortained for 4 h. The body temperature was continuously zymography. Gelatinases were bound to gelatine-coatedmonitored by a rectal thermometer and controlled with a Sephadex beads washed off with elution buffer (50 mMfeedback controlled heating pad (TKM-0902 temperature Tris, 1 M NaCl, 10 mM CaCl , 7.5% DMSO, 0.16%2
control module, Firma FMI, Bensheim-Jugendheim, Ger- Triton X-100) and subsequently loaded on the gel formany). A femoral artery was cannulated for the continuous zymography.monitoring of arterial blood pressure and for blood sam- Protease activity was visualised as clear bands. Gelspling. Arterial blood gases were measured. Tissue process- were scanned and bands quantitated by volume and densitying has been published in detail elsewhere[8]. After brains integration using the NIH Image program.

S. Wagner et al. / Brain Research 984 (2003) 63–75 65
T able 1Effect of focal ischemia and hypothermia on laminin and MMP-9
2Number of vessels /mm Laminin MMP-9(% of total vessel count)
Normothermia Hypothermia Normothermia Hypothermia
N 16.563.7 (2064.2) 15.963.0 (2063.5) 0 0L
Ir 2h/3h 1062.8 (1262.1) 11.262.7 (1561.2) 0 0Ic 2h/3h 461.4 (761.8) 3.861.2 (761.4) 960.8 (1663.5) 460.6 (762.2)Ir 2h/5h 10.362.7 (1461.9) 10.262.9 (1462) 460.8 (461.9) 3.160.9 (461.1)Ic 2h/5h 3.160.9 (662) 3.260.8 (561.7) 1061.5 (2065) 8.262.5 (1563.2)Ir 2h/8h 6.361.9 (1262.5) 9.460.8 (1562) 6.461.7 (1262.3) 4.061.2 (661.4)Ic 2h/8h 2.160.7 (561.2) 2.060.6 (561) 9.463.4 (2366.7) 8.064.0 (2365.4)Ir 2h/12h 3.7610 (761.4) 6.561.7 (1362.1) 6.462.0 (1363) 3.861.0 (762.1)Ic 2h/12h 1.860.8 (461) 1.960.5 (561.9) 10.361.4 (2566.9) 6.962.1 (2065.7)
N : non-ischemic left side of the animals undergoing MCA/O:R, Ir: periphery of the ischemic lesion, Ic: centre of the ischemic lesion.L
2 .3. MRI protocol antigens in each ROI. Mortality comparison was per-formed by Fisher’s exact test for 232 contingency tables.
The animals were examined in a 2.35 T scanner Multivariable analyses of variance with Bonferroni adjust-(Biospec 24/40, Bruker Medizintechnik, Ettlingen, Ger- ments for multiple comparisons were performed for allmany) using a home-built birdcage resonator. A first MR intergroup comparisons as well as for interantigen com-scan was done immediately after surgery to prove com- parisons and for the MRI data. Significance was set atplete occlusion of the MCA. For this purpose we used a 2P,0.05.perfusion-weighted imaging sequence (PWI). Serial MRexamination started after 3 h of reperfusion. Furtherexaminations were done after 5, 8, and 12 h of reperfusion.3 . ResultsIn each animal we performed diffusion-weighted MRimaging, T2-imaging, and T1-weighted MR imaging. T1 In all animals included in this study, PWI confirmedweighted MR imaging was performed before and immedi- successful occlusion and reperfusion of the territory ofately after injection of 0.5 mmol /kg Gd-DTPA middle cerebral artery (MCA).
(Magnevist , Schering, Berlin, Germany).From the T2-weighted MR-images we calculated the 3 .1. Physiology
volumes of infarction. From the diffusion-weighted imagesADC maps were generated to display the volume fraction pH,pO , pCO and mean arterial blood pressure were2 2
of different ADC values [13]. We also measured the within the expected physiological range for all groups.absolute volume of the lesion on diffusion weighted Body temperature was 37.160.88C in the normothermicimages and the volume of the area of contrast agent uptake group and 33.460.58C in the group treated with hypo-on T1-weighted images, displaying the area of BBB thermia.breakdown. We calculated the volume of infarction as thesum across slices of the area with infarction per slice 3 .2. Mortalitymultiplied by the thickness of the slice, i.e. 2 mm:V 5inf
o A (2 mm) ; A5area. Hemispheric lesion volume HLV Five of the 24 animals treated with normothermia ands dinf
is the ratio of V (V5volume of infarction) andV three of the 24 animals treated with hypothermia diedinf hi
(V 5volume of the ipsilateral hemisphere): HLV5V / before reaching the planned reperfusion time. This differ-hi inf
V . We calculated the difference between the two volumes ence was not statistically significant (Fisher’s exact test).hi
to quantify the ischemic area without visible signs of BBBbreakdown (I ). BBB breakdown only occurs in severely 3 .3. Results of the MRI studiesr
damaged tissue and is assumed to be part of the ischemiccore (I ). The third region of interest defined by MRI was T2 weighted images displayed a progressively increas-c
non-ischemic tissue in a corresponding area of the left ing lesion over time (Fig. 1). Hypothermia reduced thehemisphere. lesion sizes on T2 weighted and diffusion weighted
images. Diffusion weighted images showed an increase in2 .4. Statistical analysis lesion volume over time with significant differences in 3–8
h of reperfusion (P,0.05) (Fig. 2f). This increase wasAll data are presented as the mean6standard deviation found in both the normo- and the hypothermia groups, but
(S.D.). The distributions of relevant antigens are presented the volumes were smaller in the hypothermia comparedas numbers of cells or microvessels expressing the given with the normothermia groups: The mean lesion volumes

66 S. Wagner et al. / Brain Research 984 (2003) 63–75
Fig. 1. Example of infarct depiction on T2-weighted images.
after 5 and 12 h reperfusion were 192645 and 272652 left and right hemisphere. MMP-9 is not expressed in3mm , respectively, in the animals not treated with hypo- normal brain. Laminin-5 appeared on 1563.2 (N ) andL
3 2thermia, contrasted to 124643 and 181635 mm , respec- 17.564.1 (N ) microvessels /mm of the control animals.R
tively, in the hypothermia group.Fig. 2a,bshows the time The density of laminin-5 immunoreactive microvesselscourse of the lesion measured by serial MRI at 3 and 12 h was not different from the sham-operated subjects, or fromof reperfusion in an animal treated with hypothermia (Fig. the non ischemic hemisphere of subjects undergoing2b) and one animal not treated with hypothermia (Fig. 2a). MCA:OR. MMP-2 seemed to stain neurones and as-Postcontrast T1 weighted MRI demonstrating disruption of trocytes. However, because of the weak staining, the exactthe BBB in the ischemic area and diffusion weighted cellular origin could not be confirmed by immunofluores-images at 3 h and 12 h of reperfusion are shown inFig. cence. TIMP-2 was also expressed on neurones in the2a,b for representative animals from both groups. In hippocampal area as well as in the cortex. This wasanimals treated with hypothermia, the area of BBB break- confirmed by double labelling with the neuronal markerdown was significantly smaller than in the normothermia NeuN. In selected cases serial sections were stained for CDgroup (Fig. 2e). Moreover if we calculated the difference 31 adjacent to the sections stained for laminin-5 andbetween the lesion on the diffusion weighted image and MMP-9. In each case examined the laminin-5 and MMP-9the area of BBB breakdown of MRI, named I (ischemic positive structures were identified as vessels by eitherr
area without BBB breakdown), we found larger I s for all immunofluorescent double staining or staining of adjacentr
time points in rats treated with hypothermia (Fig. 2c,d). sections with CD 31.However the difference for I between these groups was After MCA:O/R 2/3 h, the number of microvesselsr
not statistically significant. expressing laminin-5 antigen decreased in the I and Ic r
regions compared to the contralateral non-ischemic zone2 23 .4. Abundance of laminin-5, MMPs and TIMP-2 during (N), such that N516.563.7/mm , I51062.8/mm ,L r
2MCA:O /R in the rat and I 5461.4/mm (2P,0.05) (Fig. 3a). Duringc
MCA:O/R, the absolute density of microvessels express-In non-ischemic controls, the abundance of MMP-2, ing laminin-5 antigen was dependent upon the region of
TIMP-2, and laminin-5 antigen was balanced between the neurone injury, displaying a graded significant decrease in

S. Wagner et al. / Brain Research 984 (2003) 63–75 67
Fig. 2. Results of the diffusion- and T1-weighted images. (a,b) Diffusion weighted (left side) and contrast enhanced T1 images (right side) after 3 h (top)and 12 h (bottom) of reperfusion in representative animals from the normothermic (a) and hypothermic groups (b). After 3 h of reperfusion, the lesion iseasily detectable on the diffusion weighted images. BBB breakdown is not detected. After 12 h of reperfusion, the area of BBB breakdown matches thelesion on the diffusion weighted image in the normothermic animal, whereas hypothermia leads to a reduction of lesion volume and area of BBBbreakdown preserving the area without BBB breakdown. DWI5diffusion weighted image; T1WI5T1 weighted image. (c,d) Flowchart of tworepresentative animals in the normo- and hypothermic groups. In the normothermic animal (c), the T1 lesion volume approaches the diffusion weightedlesion volume over time, and appears to meet at 12 h reperfusion, whereas under hypothermia (d) the 2 curves remain parallel. HLV5correctedhemispheric lesion volumes were calculated with the equation HLA (%)5 [LT 2 (RT2RI)] /LT 3 100, where LT is the area of the left hemisphere (in
2 2 2mm ), RT is the area of the right hemisphere (in mm ), and RI is the infarcted area (in mm ). (e,f) The bars indicate the volume of the lesion onpostcontrast T1 images over time in the normo- and hypothermic group (e). The volumes at 2/5 h, 2/8 h and 2/12 h are significantly smaller (P,0.05) inanimals treated with hypothermia. (f) Indicates the volume of the lesion on diffusion weighted images over time in the normo- and hypothermic group.There is a statistical significant difference between normo- and hypothermia at 2/12 h (P,0.05).

68 S. Wagner et al. / Brain Research 984 (2003) 63–75
Fig. 2. (continued)
the order N.I .I at 3, 5 and 8 h of reperfusion (2P, region displaying BBB breakdown (number of microves-r c20.05, seeTable 1). sels: 960.8/mm ). After 5 h of reperfusion MMP-9 was
2MMP-9 appeared after 3 h of reperfusion in the I expressed on significantly fewer vessels (460.8/mm ) inc

S. Wagner et al. / Brain Research 984 (2003) 63–75 69
Fig. 2. (continued)
2the I area, than in the I area (1061.5 mm ) (Table 1and on diffusion weighted images was significant: 181635r c3 3Fig. 3e). TIMP-2 appeared on few vessels after 5 h of mm in the hypothermia group versus 272652 mm in the
reperfusion in the I area. It was not expressed in the I normothermia group. Treatment with hypothermia signifi-r c
area. The neuronal TIMP presence disappeared at the cantly reduced the area of BBB breakdown in MRI after 5earliest time point after 3 h of reperfusion. We could never h of reperfusion. This effect was seen up to 12 h ofconvincingly detect MMP-2 on vessels. The abundance reperfusion (Fig. 2e). The I increased in both groups overc
increased after 5 h of reperfusion in comparison of the time, but more slowly under hypothermia. I grows at I ’sc r
control tissue on cells morphologically identified as as- expense, therefore I did not increase absolutely over timer
trocytes in the I and to a lesser extent in the I area. but the BBB (I ) is longer preserved by hypothermia.r c c
On zymography, MMP-2 was constitutively expressed Expressing the area of risk in percent of the infarct volumein normal brain and increased after 3 h of reperfusion. in diffusion weighted images, we find 17610% (nor-After the initial increase, levels of MMP-2 stayed constant mothermia) versus 59.8630.3% (hypothermia) at 5 h ofuntil 12 h of reperfusion, when a second marked increase reperfusion and 20613.3% (normothermia) versuswas observed (Fig. 3d). In concordance with immuno- 44.1625.3% (hypothermia) at 8 h of reperfusion.histochemistry, MMP-9 was not detected in normal brain. Hypothermia had a preservative effect on the numbersIt was detected after 3 h of reperfusion, and increased of laminin-5 positive vessels (Table 1). Hypothermia alsofurther with longer reperfusion times (Fig. 3c). In nor- decreased the abundance and activity in MMP-9 in the areamothermic animals, MMP-9 correlated with the extent of of BBB breakdown (Table 1). This was confirmed by athe lesion on diffusion weighted images (r50.69, P, decrease in optical density zymography bands of MMP-90.05) and with the area of BBB breakdown on postcontrast and MMP-2 in animals treated with hypothermia (Fig.T1 images (r50.93, y 5 0.18x 189.27,P,0.05). After 12 3b–d).h of reperfusion MMP-2 correlated significantly with thelesion on diffusion weighted images (r50.63, P,0.05).
4 . Discussion3 .5. Effect of hypothermia
This study was designed to test the hypothesis thatHypothermia significantly decreased the volume of the MMPs and their substrate laminin-5 are expressed in
ischemic region as well as the area of BBB breakdown different regions of ischemic tissue and are involved in(Fig. 2e,f). Even after 12 h of reperfusion, the difference BBB breakdown. For this purpose, serial MRIs were

70 S. Wagner et al. / Brain Research 984 (2003) 63–75
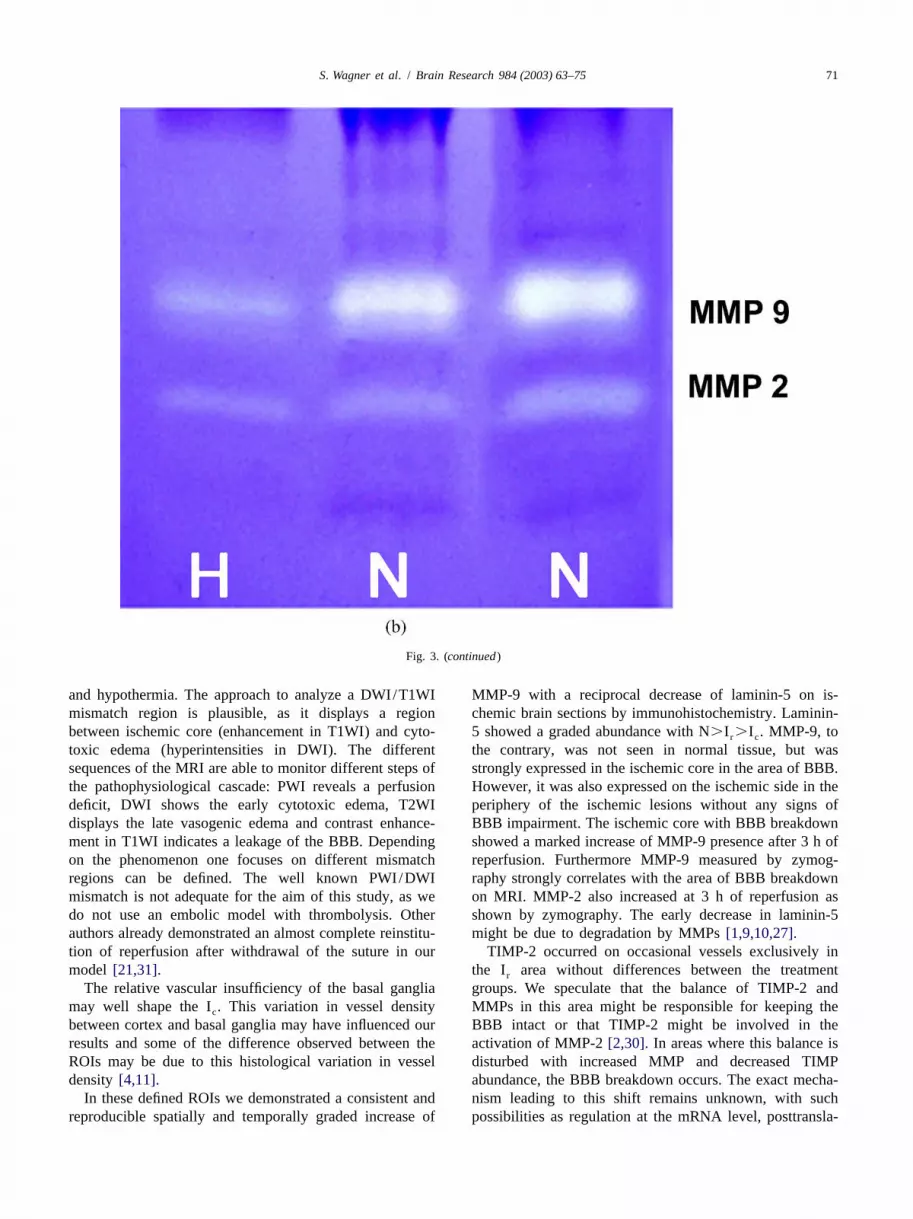
Fig. 3. Changes in laminin-5 and MMP abundance under focal cerebral ischemia: effect of hypothermia. (a) Effect of MCA:O/R on laminin-5-positivevessels. Already after 3 h of reperfusion, the gradation of N(A).I (B).I (C) is established. The separations in regional abundance for laminin-5 isr c
statistically significant (2P,0.05) (magnification340). (b) Zymogram of two normothermic and one hypothermic animal. Note the decrease in size andintensity of the bands of MMP-9 and MMP-2 under hypothermic conditions. N: normothermia; H: hypothermia. (c,d) Time course of MMP-9 (c) andMMP-2 (d) abundance after different reperfusion times (n56 in each group). The size of lysis zones is given as integrated density. MMP-2 increased after3 h of reperfusion and stayed constant to further increase after 12 h of reperfusion. MMP-9 gradually increased. Hypothermia reduced the amount of MMPabundance, especially MMP-9. * Indicates statistical significance (P,0.05). (e) Effect of MCA:O/R on MMP-9. Note the absence of MMP-9 positivevessels in normal tissue (A) and the graded increase from I (B) to I (C) at 5 h of reperfuson.(magnification340).r c
performed and the BBB breakdown was monitored in vivo reduced the size of the lesion on diffusion weighted imagesby contrast enhanced T1 imaging. We further set out to test and decreased the area of BBB breakdown and infarctthe hypothesis that hypothermia would have beneficial volume. In accord with previous experiments, we did noteffects on volume of infarct and on BBB disruption, ever observe recovery of ischemic regions on diffusionpossibly due to inhibitory effects on MMPs. weighted images after 2 h of occlusion[20]. The defined
We used MRI to define our regions of interest. One region I is an area which has signs of ischemia shown byr
could take regions defined by immunohistochemistry and abnormalities on the diffusion weighted image but displaystransfer those to the MRI scans. Changes depicted on the no visible BBB breakdown. The area of risk constitutes aMRI scans are, of course, secondary to different degrees of small volume resulting in small areas of this ROI ontissue damage. sections. This may have limited the accuracy with which
The lesion on MRI developed over time with a progres- the border to the adjacent ROI could be respected. We triedsive increase of lesion volume on T2 and diffusion to avoid this by choosing the ROIs of the area of riskweighted imaging. The lesion size derived from the directly adjacent to the ischemic core. This border isdifferent MR sequences is comparable with other inves- identifiable on H&E counterstained sections. The preserva-tigations of this filament MCAO model of the rat[16,18– tion of this area in most animals of the hypothermia group20]. The area of BBB breakdown detected on T1 weighted suggests that parts of it might be salvageable by neuro-images spread over time. In the animals in the normother- protective therapies. However, larger groups and longermia group, the area of BBB breakdown after 12 h of observation times are needed in future studies to follow thereperfusion covered large parts of the lesion on diffusion fate of this area of ischemic tissue. The authors are awareweighted images. In some cases, the areas were identical. that the penumbra, which is a term used to define potentialIn rats treated with hypothermia, this process was slowed salvageable tissue, is defined differently from and is notdown and even after 12 h of reperfusion a region within identical with, I[14,25]. The aim of this study was not tor
the lesion on diffusion weighted images without BBB examine the effect of ischemia on the penumbra; rather, webreakdown could be identified. Hypothermia significantly focused on the evaluation of BBB damage under normo-
S. Wagner et al. / Brain Research 984 (2003) 63–75 71
Fig. 3. (continued)
and hypothermia. The approach to analyze a DWI/T1WI MMP-9 with a reciprocal decrease of laminin-5 on is-mismatch region is plausible, as it displays a region chemic brain sections by immunohistochemistry. Laminin-between ischemic core (enhancement in T1WI) and cyto- 5 showed a graded abundance with N.I .I . MMP-9, tor c
toxic edema (hyperintensities in DWI). The different the contrary, was not seen in normal tissue, but wassequences of the MRI are able to monitor different steps of strongly expressed in the ischemic core in the area of BBB.the pathophysiological cascade: PWI reveals a perfusion However, it was also expressed on the ischemic side in thedeficit, DWI shows the early cytotoxic edema, T2WI periphery of the ischemic lesions without any signs ofdisplays the late vasogenic edema and contrast enhance- BBB impairment. The ischemic core with BBB breakdownment in T1WI indicates a leakage of the BBB. Depending showed a marked increase of MMP-9 presence after 3 h ofon the phenomenon one focuses on different mismatch reperfusion. Furthermore MMP-9 measured by zymog-regions can be defined. The well known PWI/DWI raphy strongly correlates with the area of BBB breakdownmismatch is not adequate for the aim of this study, as we on MRI. MMP-2 also increased at 3 h of reperfusion asdo not use an embolic model with thrombolysis. Other shown by zymography. The early decrease in laminin-5authors already demonstrated an almost complete reinstitu- might be due to degradation by MMPs[1,9,10,27].tion of reperfusion after withdrawal of the suture in our TIMP-2 occurred on occasional vessels exclusively inmodel [21,31]. the I area without differences between the treatmentr
The relative vascular insufficiency of the basal ganglia groups. We speculate that the balance of TIMP-2 andmay well shape the I . This variation in vessel density MMPs in this area might be responsible for keeping thec
between cortex and basal ganglia may have influenced our BBB intact or that TIMP-2 might be involved in theresults and some of the difference observed between the activation of MMP-2[2,30]. In areas where this balance isROIs may be due to this histological variation in vessel disturbed with increased MMP and decreased TIMPdensity [4,11]. abundance, the BBB breakdown occurs. The exact mecha-
In these defined ROIs we demonstrated a consistent and nism leading to this shift remains unknown, with suchreproducible spatially and temporally graded increase of possibilities as regulation at the mRNA level, posttransla-

72 S. Wagner et al. / Brain Research 984 (2003) 63–75
Fig. 3. (continued)
tional modification of TIMP, or simply binding to MMPs. using inhibitory MMP antibodies or synthetic MMP in-Presence of laminin may be taken as one sensitive in- hibitors have been undertaken by several groups. Blockingdicator of vascular matrix integrity and its loss is propor- of MMP led to decreased volume of the infarct measuredtional to the graded increase in MMP presence towards the on brain sections, and decreased indirect measures e.g.ischemic core of the lesion. Thus interventions which glucose uptake secondary to BBB breakdown[23]. Tomodulate the upregulation of MMPs or parallel downregu- date, a detailed monitoring of the effect of this therapy inlation of TIMP will reduce BBB breakdown. Attempts by vivo by e.g. MRI has not been shown. The presence of

S. Wagner et al. / Brain Research 984 (2003) 63–75 73
Fig. 3. (continued)
MMP-2 and MMP-9 has been reported before in different brain and disappeared after 3 h of reperfusion to sub-models, and their role in BBB breakdown has been implied sequently appear on vessels in the I region, where it isr
previously [15,23]. Rosenberg at al.[23] report a strong likely to counterbalance MMP activity.correlation of BBB breakdown measured by brain sucrose Neutrophils as a potential source of MMPs do not play auptake with MMP-2 and MMP-9 after 3 and 24 h, role in this study for two reasons: (1) our perfusionrespectively. As we were interested in early time points technique removes blood elements and plasma, and (2)and the immediate effect of hypothermia, we did not significant infiltration of leukocytes does not occur until 12extend our observation period beyond 12 h. A further h after MCA occlusion[7].increase of MMP-9 with a peak at 24 h is possible. We We studied the expression of laminin-5, which is ancould not detect an early peak of BBB breakdown at 3 h important substrate of gelatinases and a very dynamicwith MRI, this being the earliest time at which we component of the basement membrane. However, laminin-observed an increase in MMP-2. Depending on the model 5 is not the only ECM protein responsible for structuraland the reperfusion times, the time course of MMPs seems integrity: other ECM proteins could well maintain a certainto differ. Fujimura at al.[5] report an association of BBB level of integrity at a given time point. The failure to detectbreakdown with increased MMP-9 after 3 h. Hoe et al. laminin-5 by immunohistochemistry does not imply com-[15] on the other hand correlated MMPs with neurones plete absence of the antigen but reduction below a thres-displaying dUTP uptake. Their number correlated with hold level. Therefore we can indeed see vessels which areMMP-2 abundance. They also reported an increase of laminin-5 negative. There seem to be species differences inMMP-9 in a small subgroup of animals with haemorrhagic the expression of laminin-5 on cerebral vessels: for exam-transformation which, obviously, must have been preceded ple, it was below the threshold of detection in more vesselsby BBB breakdown. Despite certain differences, the results in the rat than in the non-human primate[29]. If thisof the studies including ours overlap sufficiently to infer difference in vessel count between the two species is duethat MMP-2 and MMP-9 are increased after focal cerebral to lack of detection in the rat brain or lack of specificity inischemia, and that they contribute to BBB breakdown and the non-human primate tissue is uncertain at this point.infarct progression. However a systematic error in laminin-5 positive vessels
MMPs have been described on astrocytes, neurones, due to low detection in rat tissue would affect all ROIsmicroglia, invading neutrophils and endothelial cells[3]. equally and this would not alter the finding of a gradedWe could identify MMP-9 on vessels and MMP-2 on loss.astrocytes. The absence of MMP-2 on the vessels pre- Hypothermia reduced MMP-9 presence as assessedvented us from quantitating it using the methods we used qualitatively on brain sections, and led to a significantfor MMP-9. However, by zymography MMP-2 does decrease in MMP-2 and MMP-9 activity as measured byindeed increase along with MMP-9. We therefore cannot zymography. This reduction was significant for MMP-9 atexclude the possibility that MMP-2 produced by neurons all time points. The effect outlasted the duration ofand astrocytes may contribute to basal lamina destruction hypothermia and even 12 h after reperfusion, 9 h afterin the vessels. TIMP-2 was detected on neurones in normal termination of hypothermia, the MMPs were significantly

74 S. Wagner et al. / Brain Research 984 (2003) 63–75
[5] M . Fujimura, Y. Gasche, Y. Morita-Fujimura, J. Massengale, M.less expressed than in the normothermia group. Hypo-Kawase, P.H. Chan, Early appearance of activated matrixthermia leads to decreased BBB breakdown and infarctmetalloproteinase-9 and blood–brain barrier disruption in mice after
volume on MRI. One pathophysiological aspect of this focal cerebral ischemia and reperfusion, Brain Res. 842 (1999)therapy is the inhibition of MMPs. We describe for the first 92–100.time this new pathophysiological aspect of mild hypo- [6] J .H. Garcia, K.F. Liu, J. Lian, J. Xu, Astrocytic and microvascular
responses to the occlusion of middle cerebral artery, J. Neuropathol.thermia. Other mechanisms for example reduced inflam-Exp. Neurol. 52 (1993) 288.matory response, decreased metabolic requirement for
[7] J .H. Garcia, K.F. Liu, Y. Yoshida, J. Lian, S. Chen, G.J. del Zoppo,oxygen, and alterations in neurotransmitter release have Influx of leukocytes and platelets in an evolving brain infarctbeen proposed. Kawai et al.[18] have recently shown that (Wistar rat), Am. J. Pathol. 144 (1994) 188–199.delayed postischemic hypothermia is as effective as intrais- [8] J .H. Garcia, S. Wagner, K.F. Liu, X.J. Hu, Neurological deficit and
extent of neuronal necrosis attributable to middle cerebral arterychemic hypothermia. This is very important with regard toocclusion in rats, Stroke 26 (1995) 627–635.treating patients on NICU, as treatment is often initialised
[9] G . Giannelli, J. Brassard, C. Foti, W.G. Stetler-Stevenson, J. Falk-after more than 6 h after the initial ischemic event[26].Marzillier, A. Zambonin-Zallone, O. Schiraldi, V. Quaranta, Altered
Kawai et al. [18] investigated the effect of intraischemic expression of basement membrane proteins and their integrinversus postischemic hypothermia after 2 h of MCA receptors in lichen planus: possible pathogenetic role of gelatinases
A and B, Lab. Invest. 74 (1996) 1091–1104.occlusion on neutrophil accumulation and ICAM-1 expres-[10] G . Giannelli, J. Falk-Marzillier, O. Schiraldi, W.G. Stetler-Steven-sion. Both forms of hypothermia proved to be equally
son, V. Quaranta, Induction of cell migration by matrixeffective. Neutrophils and monocytes are a source ofmetalloprotease-2 cleavage of laminin-5, Science 277 (1997) 225–
MMPs. Therefore, at later timepoints they are a likely 228.source of MMP release during degranulation[32]. As we [11] U . Gobel, H. Theilen, W. Kuschinsky, Congruence of total and
perfused capillary network in rat brains, Circ. Res. 66 (1990)examined at a time when neutrophil accumulation had not271–281.started yet, we postulate that downregulation of MMPs by
[12] M . Hagerdal, J. Harp, B.K. Siesjo, Effect of hypothermia uponhypothermia early (within 12 h) after MCAO is not due toorganic phosphates, glycolytic metabolites, citric acid cycle inter-
decreased neutrophil accumulation. One alternative is a mediates and associated amino acids in rat cerebral cortex, J.temperature dependent decrease of enzyme activity. Neurochem. 24 (1975) 743–748.
[13] S . Heiland, O. Dietrich, K. Sartor, Diffusion-weighted imaging ofIn conclusion, MMPs, especially MMP-9, are associatedthe brain: comparison of stimulated- and spin–echo echo–planarwith BBB breakdown. Increases in immunoreactive MMP-sequences, Neuroradiology 43 (2001) 442–447.9, together with reciprocal decreases in the levels of its
[14] W .D. Heiss, Ischemic penumbra: evidence from functional imagingsubstrate, laminin-5, correlate with a gradation. Hypo- in man, J. Cereb. Blood Flow Metab. 20 (2000) 1276–1293.thermia reduces infarct size and area of BBB breakdown [15] J .H. Heo, J. Lucero, T. Abumiya, J.A. Koziol, B.R. Copeland, G.J.
del Zoppo, Matrix metalloproteinases increase very early duringpossibly by inhibiting MMPs.experimental focal cerebral ischemia, J. Cereb. Blood Flow Metab.19 (1999) 624–633.
[16] P .W. Huh, L. Belayev, W. Zhao, S. Koch, R. Busto, M.D. Ginsberg,A cknowledgements Comparative neuroprotective efficacy of prolonged moderate intrais-
chemic and postischemic hypothermia in focal cerebral ischemia, J.Neurosurg. 92 (2000) 91–99.This work was funded by a grant (233/1999) of the
[17] R .O. Hynes, Integrins: versatility, modulation, and signaling in cellUniversity of Heidelberg. We are thankful to Professor V.adhesion, Cell 69 (1992) 11–25.Quaranta (Department of Cell Biology, Scripps Research
[18] N . Kawai, M. Okauchi, K. Morisaki, S. Nagao, Effects of delayedInstitute, La Jolla, USA) and Dr Clemens Sommer (De- intraischemic and postischemic hypothermia on a focal model ofpartment of Neuropathology, University of Heidelberg, transient cerebral ischemia in rats, Stroke 31 (2000) 1982–1989.
[19] R . Kollmar, W.R. Schabitz, S. Heiland, D. Georgiadis, P.D. Schell-Germany) for helpful discussions.inger, J. Bardutzky, S. Schwab, Neuroprotective effect of delayedmoderate hypothermia after focal cerebral ischemia: an MRI study,Stroke 33 (2002) 1899–1904.
R eferences [20] T . Neumann-Haefelin, A. Kastrup, A. de Crespigny, M.A. Yenari, T.Ringer, G.H. Sun, M.E. Moseley, Serial MRI after transient focalcerebral ischemia in rats: dynamics of tissue injury, blood–brain[1] S . Amano, N. Akutsu, Y. Matsunaga, T. Nishiyama, M.F. Cham-barrier damage, and edema formation, Stroke 31 (2000) 1965–1973.pliaud, R.E. Burgeson, E. Adachi, Importance of balance between
[21] K . Nishigaya, Y. Yoshida, M. Sasuga, H. Nukui, G. Ooneda, Effectextracellular matrix synthesis and degradation in basement mem-of recirculation on exacerbation of ischemic vascular lesions in ratbrane formation, Exp. Cell Res. 271 (2001) 249–262.brain, Stroke 22 (1991) 635–642.[2] W . Bode, C. Fernandez-Catalan, F. Grams, F.X. Gomis-Ruth, H.
[22] A . Pozzi, P.E. Moberg, L.A. Miles, S. Wagner, P. Soloway, H.A.Nagase, H. Tschesche, K. Maskos, Insights into MMP-TIMPGardner, Elevated matrix metalloprotease and angiostatin levels ininteractions, Ann. NY Acad. Sci. 878 (1999) 73–91.integrin alpha 1 knockout mice cause reduced tumor vascularization,[3] I .L. Campbell, A. Pagenstecher, Matrix metalloproteinases and theirProc. Natl. Acad. Sci. USA 97 (2000) 2202–2207.inhibitors in the nervous system: the good, the bad and the
enigmatic, Trends Neurosci. 22 (1999) 285–287. [23] G .A. Rosenberg, E.Y. Estrada, J.E. Dencoff, Matrix metalloprotein-[4] M . Cavaglia, S.M. Dombrowski, J. Drazba, A. Vasanji, P.M. ases and TIMPs are associated with blood–brain barrier opening
Bokesch, D. Janigro, Regional variation in brain capillary density after reperfusion in rat brain, Stroke 29 (1998) 2189–2195.and vascular response to ischemia, Brain Res. 910 (2001) 81–93. [24] G .A. Rosenberg, M. Navratil, F. Barone, G. Feuerstein, Proteolytic

S. Wagner et al. / Brain Research 984 (2003) 63–75 75
cascade enzymes increase in focal cerebral ischemia in rat, J. Cereb. [29] S . Wagner, M. Tagaya, J.A. Koziol, V. Quaranta, G.J. del Zoppo,Blood Flow Metab. 16 (1996) 360–366. Rapid disruption of an astrocyte interaction with the extracellular
[25] G . Schlaug, A. Benfield, A.E. Baird, B. Siewert, K.O. Lovblad, R.A. matrix mediated by integrin alpha 6 beta 4 during focal cerebralParker, R.R. Edelman, S. Warach, The ischemic penumbra: opera- ischemia/ reperfusion, Stroke 28 (1997) 858–865.tionally defined by diffusion and perfusion MRI, Neurology 53 [30] Z . Wang, R. Juttermann, P.D. Soloway, TIMP-2 is required for(1999) 1528–1537. efficient activation of proMMP-2 in vivo, J. Biol. Chem. 275 (2000)
[26] S . Schwab, S. Schwarz, M. Spranger, E. Keller, M. Bertram, W. 26411–26415.Hacke, Moderate hypothermia in the treatment of patients with [31] G .Y. Yang, A.L. Betz, Reperfusion-induced injury to the blood–brainsevere middle cerebral artery infarction, Stroke 29 (1998) 2461– barrier after middle cerebral artery occlusion in rats, Stroke 252466. (1994) 1658–1664, discussion 1664–1665.
[27] R .E. Seftor, E.A. Seftor, N. Koshikawa, P.S. Meltzer, L.M. Gardner, [32] S . Zucker, M. Hymowitz, C. Conner, H.M. Zarrabi, A.N. Hurewitz,M. Bilban, W.G. Stetler-Stevenson, V. Quaranta, M.J. Hendrix, L. Matrisian, D. Boyd, G. Nicolson, S. Montana, Measurement ofCooperative interactions of laminin 5 gamma2 chain, matrix matrix metalloproteinases and tissue inhibitors of metalloproteinasesmetalloproteinase-2, and membrane type-1-matrix /metalloproteinase in blood and tissues. Clinical and experimental applications, Ann.are required for mimicry of embryonic vasculogenesis by aggressive NY Acad. Sci. 878 (1999) 212–227.melanoma, Cancer Res. 61 (2001) 6322–6327.
[28] S . Wagner, H. Gardner, Modes of regulation of laminin-5 productionby rat astrocytes, Neurosci. Lett. 284 (2000) 105–108.





























